Embed Size (px)
Citation preview

NATURALIZED POPULATIONS OF OYSTERS, CRASSOSTREA GIGAS ALONG THE SOUTHAFRICAN COAST: DISTRIBUTION, ABUNDANCE AND POPULATION STRUCTURE
T. B. ROBINSON,1 C. L. GRIFFITHS,1 A. TONIN,2 P. BLOOMER3 AND M. P. HARE4
1Centre of Invasion Biology, Zoology Department and Marine Biology Research Institute, University ofCape Town, Rondebosch 7700, South Africa; 2Mariculture Development Services, PO Box 200,Alexander Bay 8290; 3Molecular Ecology and Evolution Programme, Department of Genetics,University of Pretoria, 0002, South Africa; 4Biology Department, University of Maryland,College Park, Maryland 20742
ABSTRACT The Japanese oyster Crassostrea gigas forms the mainstay of the South African oyster farming industry. Despite havingbeen cultured along the South African coast for 30 years, this is the first record of naturalized populations of this well known globalinvader from the region. DNA sequence data confirmed the presence of C. gigas in the Breede and Goukou estuaries. Populations of184,206 ± 21 058.9 (SE), 876 ± 604.2 (SE) and 1 228 ± 841.8 (S.E.) individuals were recorded in the Breede, Goukou and Knysnaestuaries respectively. No C. gigas were recorded from the open coast. The condition index of oysters in the Goukou and Knysnaestuaries differed significantly from those in the Breede Estuary (Kruskal-Wallis ANOVA H � 48.249, P < 0.01) but no differencewas found between the Goukou and Knysna estuaries (Kruskal-Wallis ANOVA H � 48.249, P > 0.05). The lower condition indexrecorded in the Breede Estuary is believed to reflect decreased food supply as a result of the large C. gigas population present in theestuary.
KEY WORDS: Crassostrea gigas, oyster, South Africa, introduced species, diagnostic DNA identification
INTRODUCTION
Over the last century, oysters have been extensively movedaround the globe as highly successful aquaculture species (An-drews 1980). As the earliest of these translocations took placebefore the implications of introducing non-native species wererealized or understood, they very often lead to the establishment oflarge invasive populations. Despite a more responsible approach totranslocations of nonindigenous species in recent times (Shatkin etal. 1997), modern introductions have none-the-less resulted innaturalized populations establishing. Of all oysters used in aqua-culture, the Japanese oyster Crassostrea gigas Thunberg is argu-ably the most aggressive invader. Although originating in the northwest Pacific, C. gigas presently has established naturalized popu-lations on all major coastlines of the Northern Hemisphere (withthe exception of the Atlantic coast of North America) (Andrews1980, Carlton et al.1990, Minchin 1996, Shatkin et al. 1997, Wehr-mann et al. 2000), as well as on the coasts of Tasmania, southernand eastern Australia (Thomson 1959, Shatkin et al. 1997) andNew Zealand (Dinamani 1971). It is however unclear if C. gigasexhibits heightened invasiveness in comparison to other commonaquaculture species, or if the high frequency of transport of thisspecies is responsible for the elevated incidence of naturalizedpopulations.
Following global trends, the South African oyster industry isbased on C. gigas. This species was first imported to KnysnaEstuary in the 1950s (Korringa 1956), where it is still farmed.Presently two other South African estuaries (Kowie & SwartkopsEstuaries) support oyster farms, whereas a further three fully ma-rine farms are based in Algoa Bay, Saldanha Bay and AlexanderBay (Fig. 1). Due to difficulties in inducing predictable spawningand subsequent settling under South African conditions, the entireindustry has been based on spat imported from Chile, the UnitedKingdom and France. As C. gigas has appeared unable to completeits life cycle under local environmental conditions, this species hasnot previously been considered to pose a threat as an invader alongSouth African shores (Griffiths et al. 1992, C. Smith pers comm.).
This article documents the first record of naturalized popula-
tions of C. gigas along the coast of South Africa, and establishesthe present distribution and status of this species.
METHODS
In 2001, 20 estuaries along the southern and eastern coasts ofSouth Africa were surveyed for the presence of oysters. In in-stances where the identification of individuals was not possibleusing morphometric characters, tissue samples were collected andlater used in genetic analyses to obtain confirmation of speciesidentification.
Several recent studies have used DNA sequences of either mi-tochondrial protein coding, ribosomal genes (rDNA) or nuclearrDNA genes for studying phylogenetic relationships among oys-ters (Jozefowicz and Foighil 1998, Lapegue et al. 2002, Matsu-moto 2003) as well as for species identification purposes (Banks etal. 1993). In this study C. gigas (n � 18) from 3 oyster farms(Walvis Bay and Swakopmund in Namibia and Saldanha Bay inSouth Africa) were used to compare with the indigenous Cape rockoyster Striostrea margaritacea Lamarck (n � 9), collected fromthe Breede (samples 1–6) and Goukou (samples 7–9) Estuaries.Diagnostic genetic differences were used to determine the identityof 12 C. gigas specimens collected from the Breede Estuary. Sev-eral published sequences were used for comparative purposes(numbers in parentheses refer to sequence accession numbers in arange of international data bases such as GenBank): C. gigas (COI,GB AF152565; 18S rDNA DBJ AB064942.1), Crassostrea vir-ginica Gmelin (COI, GB AF152566; 18S rDNA EMB X60315.1),Crassostrea angulata Lamarck (COI, GB AF152567) and Saccos-trea cuccullata Born (18S rDNA, EMB AJ389634.1). Ostrea chil-ensis_Philippi (COI, GB AF112289) or O. edulis (18S rDNA, GBU88709.1) was used for outgroup rooting of phylogenetic trees.
DNA was extracted from 2 mg of gill tissue using the DNeasy96 Tissue Kit (Qiagen) according to the manufacturer’s instruc-tions. Two DNA regions were amplified via the polymerase chainreaction (PCR; Saiki et al. 1988): A 700 base pair (bp) fragment ofthe mitochondrial DNA (mtDNA) cytochrome oxidase subunit I(COI) gene (Folmer et al. 1994) and a 520 bp fragment of the 18S
Journal of Shellfish Research, Vol. 24, No. 2, 443–450, 2005.
443

nuclear rDNA gene (Lane et al. 1985, Diez et al. 2001). Amplifi-cations were performed in a total volume of 25 �l containing 0.5units Taq DNA polymerase (Invitrogen). For COI 2.5 mM MgCl2was used at an annealing temperature of 45°C, while 18S wasannealed at 50°C in reactions containing 1.5 mM MgCl2. Suc-cessful PCR reactions were purified using the SAP/Exo systemaccording to the manufacturer’s protocol (http://www.pebiodocs.com/pebiodocs/04305080.pdf). Successful PCR reactions were se-quenced in the forward and reverse directions using dye-terminatorcycle sequencing (Big Dye DNA sequencing kit, Applied Biosys-tems) with enzymatic template purification according to the manu-facturer’s recommendations; sequences were electrophoresed onan ABI 3100 automated capillary DNA sequencer (Applied Bio-systems). Sequences were deposited in GenBank.
Consensus sequences of each specimen were aligned to eachother in ClustalX (Thompson et al. 1997). Aligned sequences werecompared in MEGA version 2.1 (Kumar et al. 2001) where geneticdistances were calculated according to the Kimura-two-parametermodel (Kimura 1980) and sequences were clustered using theneighbor-joining algorithm (Saitou & Nei 1987). Confidence innodes was assessed using bootstrapping (Felsenstein 1985). Se-quence statistics such as nucleotide diversity (Saitou & Nei 1987)was also calculated in MEGA. A diagnostic restriction enzyme testwas developed based on observed sequence differences.
Following genetic confirmation of the presence of C. gigaswithin South African waters, the six estuaries in which this specieswas originally found were extensively resampled in 2003. Duringresampling, the morphologic characteristics detailed in Table 1were used to distinguish C. gigas from indigenous oyster species.In each estuary, all rocky shores and artificial structures offeringsuitable habitat for C. gigas were visited, and at least four intertidaltransects were surveyed for every 200 m of suitable habitat. Ateach transect site the intertidal was divided into three tidal zones
(i.e., high-, mid- and low-shore zones). Using a 0.5 m2 quadrat,which was rolled up the shore from Mean Low Water Spring toMean High Water Spring, the number of C. gigas per quadrat, ineach tidal zone, was recorded. This was used to calculate the meannumber of C. gigas individuals supported per meter square in thethree tidal zones as well as the total population present in eachestuary. Due to the eco-morphic nature of oysters, the capacity ofthe inner side of the shell was used as a measure of size of indi-viduals. Sample sizes varied between the estuaries depending onthe size of the population. Individuals were randomly selectedfrom various tidal heights and at varying distances from the mouthof the estuary. Each individual was weighed whole to the nearest0.01g. The shell and soft tissue were then separated. Shells weredried at room temperature for 24 h and then weighed again,whereas the soft tissues were oven dried at 105°C for 24 h andweighed. The following equation was used when calculating innershell capacity (Cosby & Gale 1990):
Capacity of the inner side of the shell � Whole wet weight (g)– Dry shell weight (g)
Condition index (CI) was calculated on a gravimetric basis usingthe following equation (Rainer & Mann 1992):
CI � Soft tissue dry weight (g) × 1,000Capacity of the inner shell (g)
Besides sampling the estuaries in which C. gigas was firstrecorded in 2001, three sites along the open coast between each ofthe estuaries were surveyed for the presence of C. gigas during2003 (Fig. 1).
Statistical Analyses
The sizes of C. gigas individuals occurring in the three estu-aries were compared using the Kruskal-Wallis ANOVA for mul-
Figure 1. Map of the South Africa showing the location of oyster farms and naturalized populations of C. gigas.
ROBINSON ET AL.444

tiple comparisons as non-normal distributions were recorded(Kolmogorov-Smirnov one-sample test). Because the condition in-dices of individuals were found to have heteroskedastic distribu-tions (Kolmogorov-Smirnov one-sample test), they were compared
using the Kruskal-Wallis ANOVA, followed by nonparametricmultiple comparisons. All statistical analyses were conductedusing STATISTICA for Windows (Version 6), StatSoft Inc.(2001).
TABLE 2.
A list of all estuaries searched for natural populations of oysters in 2001. The presence of C. gigas populations are indicated in bold.
EstuaryGeographical
Position
Oyster Species Present
Crassostrea gigas Striostrea margaritacea Saccostrea cuccullata
1. Breede 34°26�S 20°52�E � �
2. Duiwenhoks 34°24�S 21°00�E � �
3. Goukou 34°26�S 21°25�E � �
4. Gourits 34°24�S 22°82�E �
5. Knysna 34°09�S 23°04�E � �
6. Keurbooms 34°01�S 23°23�E7. Kromme 34°10�S 24°53�E � �
8. Swartkops 33°56�S 25°38�E �
9. Sundays 33°45�S 25°52�E �
10. Bushmans 33°44�S 26°40�E �
11. Kariega 33°43�S 26°41�E �
12 Kowie 33°37�S 36°55�E �
13. Keiskamma 33°20�S 27°29�E � � �
14. Tyolomnqa 33°16�S 27°33�E � �
15. Kwelera 32°59�S 28°03�E � �
16. Qora 32°31�S 28°41�E � �
17. Mbashe 32°17�S 28°54�E � �
18. Xora 32°11�S 29°00�E � �
19. Umtata 31°56�S 29°11�E � �
20. Umdumbe 31°55�S 29°13�E � �
TABLE 1.
Characterising features of the three dominant intertidal oyster species found along the South African coast.
Crassostrea gigas Striostrea margaritacea Saccostrea cuccullata
Radial threads No radical threads Fine radial threads on the surface of thetop valve
No radial threads
Shell interior Non-iridescent: White to off-white, mayhave mauve flecks
Iridescent: Mother-of-pearl to gold incolor
Non-iridescent: White to grey
Adductor scar At least one adductor scar purple in color Most scars have no distinct color, somemay be tinged pink-purple
Adductor scar on top valve always blackin color, lower scar not coloured
Valve margins Generally show no coloration but fewmay be mauve−black. Margins mostoften undulating
Margins smooth with no distinctcoloration
Margins of both valves generally blackwith undulating folds
Max. size 200 mm 180 mm 70 mmLocation Select estuaries on the south coast. Not
known from the open coast along SAshores
Mozambique to False Bay. Predominantlyon the open coast, but penetrate themouths of estuaries
Indo-West-Pacific to Algoa Bay. Foundmainly on the open coast but mayoccur in estuaries
Vertical positionon shore
Low intertidal zone to a depth of 1 m Extreme low tide to a depth of 5 m Upper mid-intertidal zone
Diagram
NATURALIZED CRASSOSTREA GIGAS IN SOUTH AFRICA 445

RESULTS
Indigenous or introduced oyster populations were recorded inall but one of the estuaries (Keurbooms Estuary) surveyed during2001 (Table 2). Of the indigenous species S. margaritacea wasfound throughout the survey region, whereas estuaries to the eastof Keiskamma also harbored populations of the Natal rock oysterS. cucculata. Natural populations of oysters appearing unlike ei-ther known indigenous oyster species and morphologically resem-bling C. gigas were discovered in six estuaries in the region (Table2). This identification was subsequently genetically confirmed.
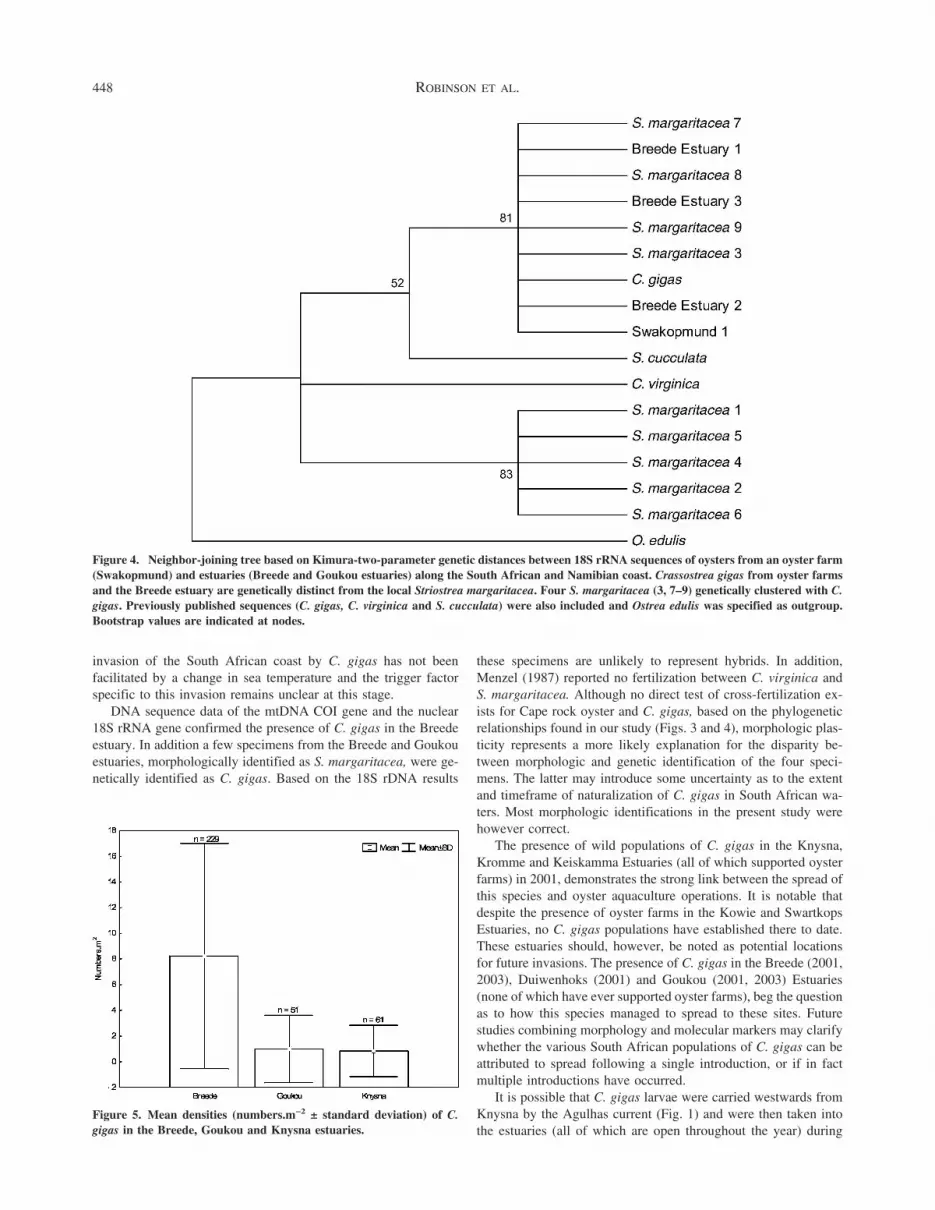
DNA sequences of the mtDNA COI gene showed substantialdifferences between C. gigas and S. margaritacea (Fig. 2a). Thesesequences in comparison with other published oyster sequencesalso revealed very high levels of sequence divergence betweenthese taxa and S. margaritacea (109-140 nucleotide differences or27% to 36% nucleotide sequence divergence and 21-30 amino aciddifferences). When compared with other species based on nucle-otide divergence, Crassostrea species grouped with C. gigas to theexclusion of S. margaritacea (Fig. 3; S. cucculata COI sequencewas unavailable). However, four exceptional S. margaritacea
samples (samples 3, 7-9) grouped with C. gigas suggesting eithermisidentification or hybridization. Mitochondrial DNA is haploidand therefore cannot be used to test for hybridization. Multicopynuclear DNA loci such as rRNA are expected to show, within anindividual, a mixture of polymorphism from two species that re-cently hybridized. Nuclear 18S sequences showed a large diver-gence between C. gigas and S. margaritacea (Fig. 2b) and groupedS. margaritacea separately from Crassostrea and Saccostrea spe-cies (Fig. 4). The four exceptional S. margaritacea samples withC. gigas mtDNA, had 18S polymorphisms characteristic of C.gigas and there is thus no genetic evidence of hybridization.
A diagnostic restriction pattern was found between C. gigasand S. margaritacea COI using the HaeIII restriction, enzyme thatrecognizes the DNA sequence 5� GG↓CC 3� (Fig. 2a). To confirmidentifications, all amplified COI products (n � 39) were subse-quently digested with HaeIII to identify S. margaritacea based onthe absence of a cut and C. gigas based on a two-fragment elec-trophoretic profile. All C. gigas from the oyster farms showed theC. gigas restriction profile. All S. margaritacea restriction patternsconfirmed the earlier sequencing results as outlined earlier. The 12specimens from the Breede estuary that were morphologically
Figure 2. Alignment of mitochondrial DNA cytochrome oxidase c subunit I (COI)(a) and nuclear 18S rDNA (b) sequences of a Cape rock oysterfrom the Breede River estuary (S. margaritacea), an oyster morphologically identified as a naturalized Crassostrea gigas (“C. gigas”) from thesame estuary and a C. gigas sequence from GenBank. The two C. gigas sets of sequences are identical. Nucleotide differences between the twogenera are highlighted in bold face. The diagnostic restriction enzyme site (HaeIII) is shaded in the COI sequence.
ROBINSON ET AL.446

identified as C. gigas, showed the C. gigas restriction profile(seven of the samples were also sequenced, see Fig. 3). Overall C.gigas from the oyster farms and from the estuaries showed verylow levels of nucleotide diversity (� � 0.001 or 0.1%). The localS. margaritacea showed slightly higher levels of nucleotide diver-sity (� � 0.003 or 0.3%) although this estimate should be recal-culated based on a larger number of individuals.
Crassostrea gigas populations have probably not been recog-nized along the South African coast to date because they are soeasily confused with the two indigenous species found in the re-gion. To simplify field identification, the physical features, whichcan be used to distinguish C. gigas from the two dominant indig-enous intertidal oysters found along the South African coast aresummarized in Table 1. No single feature provides unequivocalidentification of these oysters, because natural variability is great.Rather, a combination of features is recommended to ensure ac-curate identification of all size classes. The most discerning featureof C. gigas is the presence of at least one purple adductor scar ona non-iridescent white shell interior. The vertical range of thisspecies, from the low-shore zone to a depth of 1 m, is also animportant field characteristic. The Cape rock oyster S. margarita-cea is most reliably distinguished by the presence of fine radial
threads on the surface of the top valve. In addition, an iridescentmother-of-pearl shell interior and smooth margins on both valvesare useful identifying features. The natal rock oyster S. cuccullatais much smaller than either C. gigas or S. margaritacea, and iseasily distinguished by the presence of a black adductor scar on thetop valve, while no scar is visible on the lower valve. Blackenedvalve margins with undulating folds are also present on most in-dividuals.
Although C. gigas was recorded in six estuaries along thesouthern and eastern coasts in 2001 (Table 2), wild populationswere only present in three of these estuaries in 2003. These werethe Breede, Goukou and Knysna Estuaries. No C. gigas wererecorded on the open coast. In all three of these estuaries the entirepopulation was restricted to the low-shore zone. Breede Estuarysupported by far the highest density of individuals per metersquare (i.e., 8.3 ± 8.8 [SD].m−2) while the Goukou and KnysnaEstuaries supported 1.0 ± 2.6 (SD) and 0.9 ± 2.1 (SD) individu-als.m−2 respectively (Fig. 5). Total population size within theBreede Estuary was 184 206 ± 21 058.9 (SE) individuals (95%confidence limits � 0–496 363), compared with only 876 ± 604.2(SE) individuals (95% confidence limits � 0–5 482) and 1 228 ±841.8 (SE) individuals (95% confidence limits � 0–6 614) in theGoukou and Knysna Estuaries respectively. Thus the Breede Es-tuary population is more than two orders of magnitude larger thanthose in either of the other two estuaries.
No significant difference was recorded in the size of individu-als from the various estuaries (Kruskal-Wallis ANOVA H �4.547, P > 0.05). The Goukou Estuary supported the largest indi-viduals (40.95 ± 57.84 SD), followed by the Breede (37.02 ±26.08) and Knysna (24.47 ± 19.34) Estuaries.
Although the smallest population of C. gigas was recorded inthe Goukou Estuary, these oysters had a higher mean CI than thosein the other estuaries (Fig. 6). The CI of C. gigas differed signifi-cantly between estuaries in the Goukou and Knysna Estuaries,from those in the Breede Estuary (Kruskal-Wallis ANOVA H �48.249, P < 0.01), but no difference was found between theGoukou and Knysna Estuaries (Kruskal-Wallis ANOVA H �48.249, P > 0.05).
DISCUSSION
There are numerous records from around the world of C. gigasbeing introduced as an aquaculture species, and remaining con-fined to these operations for many years; however, unpredictablerecruitment and subsequent establishment of natural populationshas eventually occurred in nearly all cases (Shatkin et al. 1997,Wehrmann et al. 2000). Examples of such “dormancy” have beenshown in the Netherlands and in the United Kingdom, where C.gigas was first imported in 1964 and 1965, but only establishedwild populations in 1976 and 1991 respectively (Mann 1983, Shat-kin et al. 1997). In both cases, unpredicted increases in sea tem-perature probably enabled the establishment of naturalized stocks.The history of C. gigas along South African shores has shown asimilar pattern. The species was first introduced in the 1950s (Kor-ringa 1956) and wild populations were first recorded only in 2001.The difference in the South African situation is that the southernand eastern coasts offer a coastal environment with sea tempera-tures varying between 10°C and 21°C (Lutjeharms 1998)—wellwithin the range tolerated by C. gigas during the completion of itslifecycle (i.e., 4°C–24°C) (Shatkin et al. 1997). Thus, the delayed
Figure 3. Neighbor-joining phylogram based on Kimura-two-parameter genetic distances between mitochondrial DNA cytochromeoxidase C subunit I (COI) sequences of oysters from oyster farms(Swakopmund, Walvis Bay and Saldanha Bay) and estuaries (Breedeand Goukou estuaries) along the South African and Namibian coast.Crassostrea gigas from oyster farms and the Breede estuary are ge-netically distinct from the local Striostrea margaritacea. Four S. mar-garitacea (3, 7–9) genetically clustered with C. gigas. Previously pub-lished sequences (C. gigas, C. angulata, C. virginica) were also includedand Ostrea chilensis was specified as outgroup. Bootstrap values areindicated at nodes.
NATURALIZED CRASSOSTREA GIGAS IN SOUTH AFRICA 447
invasion of the South African coast by C. gigas has not beenfacilitated by a change in sea temperature and the trigger factorspecific to this invasion remains unclear at this stage.
DNA sequence data of the mtDNA COI gene and the nuclear18S rRNA gene confirmed the presence of C. gigas in the Breedeestuary. In addition a few specimens from the Breede and Goukouestuaries, morphologically identified as S. margaritacea, were ge-netically identified as C. gigas. Based on the 18S rDNA results
these specimens are unlikely to represent hybrids. In addition,Menzel (1987) reported no fertilization between C. virginica andS. margaritacea. Although no direct test of cross-fertilization ex-ists for Cape rock oyster and C. gigas, based on the phylogeneticrelationships found in our study (Figs. 3 and 4), morphologic plas-ticity represents a more likely explanation for the disparity be-tween morphologic and genetic identification of the four speci-mens. The latter may introduce some uncertainty as to the extentand timeframe of naturalization of C. gigas in South African wa-ters. Most morphologic identifications in the present study werehowever correct.
The presence of wild populations of C. gigas in the Knysna,Kromme and Keiskamma Estuaries (all of which supported oysterfarms) in 2001, demonstrates the strong link between the spread ofthis species and oyster aquaculture operations. It is notable thatdespite the presence of oyster farms in the Kowie and SwartkopsEstuaries, no C. gigas populations have established there to date.These estuaries should, however, be noted as potential locationsfor future invasions. The presence of C. gigas in the Breede (2001,2003), Duiwenhoks (2001) and Goukou (2001, 2003) Estuaries(none of which have ever supported oyster farms), beg the questionas to how this species managed to spread to these sites. Futurestudies combining morphology and molecular markers may clarifywhether the various South African populations of C. gigas can beattributed to spread following a single introduction, or if in factmultiple introductions have occurred.
It is possible that C. gigas larvae were carried westwards fromKnysna by the Agulhas current (Fig. 1) and were then taken intothe estuaries (all of which are open throughout the year) during
Figure 5. Mean densities (numbers.m−2 ± standard deviation) of C.gigas in the Breede, Goukou and Knysna estuaries.
Figure 4. Neighbor-joining tree based on Kimura-two-parameter genetic distances between 18S rRNA sequences of oysters from an oyster farm(Swakopmund) and estuaries (Breede and Goukou estuaries) along the South African and Namibian coast. Crassostrea gigas from oyster farmsand the Breede estuary are genetically distinct from the local Striostrea margaritacea. Four S. margaritacea (3, 7–9) genetically clustered with C.gigas. Previously published sequences (C. gigas, C. virginica and S. cucculata) were also included and Ostrea edulis was specified as outgroup.Bootstrap values are indicated at nodes.
ROBINSON ET AL.448

tidal exchange. Live adult oysters bought for personal consump-tion, may also have been deliberately, but illegally, introduced bywaterfront property owners. A third possibility is that oysters pur-chased for personal consumption may have been stored in theestuaries for short periods prior to consumption and have spawnedover this time. Holidaymakers frequent these areas particularlyduring the December and April holidays and C. gigas from Knysna(the most likely source of oysters for holidaymakers in this region)are known to spawn between September and April (Tonin pers.ob.). This makes it quite possible that spawning of farmed indi-viduals temporarily held in the estuaries could have resulted in theestablishment of the naturalized populations.
Knysna Estuary is the largest estuary along the southern coast.This, combined with the fact that it supports a large oyster farm,might lead one to expect that the largest wild population of C.gigas would occur within this estuary. In fact, Knysna Estuarysupported a population of only 1 228 ± 841.8 (SE) Japanese Oys-ters (95% confidence limits � 0–6 614), while by far the largestpopulation (184 206 ± 21 058.9 (SE) individuals (95% confidencelimits � 0–496 363)) was recorded in the Breede Estuary. Thereason for the observed pattern may relate to flooding of the riversfeeding the respective estuaries. Knysna Estuary experienced ex-tensive and prolonged flooding in 1998, which had severe effectson many intertidal invertebrates (P. Joubert pers comm.). In par-ticular, it was noticed by South African National Parks officialsthat the number of oysters in the estuary declined dramatically
after flooding subsided. Thus, the present size of the C. gigaspopulation in the Knysna Estuary may reflect a population yet torecover from the effects of flooding.
The significantly higher CI of C. gigas individuals from theGoukou and Knysna estuaries, in comparison to the Breede Estu-ary, may reflect a limited food supply available to Breede indi-viduals as a result of the dramatically larger population in thisestuary.
The absence of C. gigas from open coasts adjoining invadedestuaries may reflect the mediating effect of wave action on inva-sive species recorded by other authors along the South Africancoast. The density, recruitment and growth of the Mediterraneanmussel Mytilus galloprovincialis Lamarck along the west coast ofSouth Africa, has been found to peak under conditions of highwave exposure while decreasing towards the two extremes waveenergy (Steffani & Branch 2003a, 2003b, 2003c). Similarly, theEuropean shore crab Carcinus maens Linnaeus has been shown tobe restricted in abundance along open coast wave exposed shoreswhile flourishing in areas of shelter (Le Roux et al. 1990, Robin-son in prep). Should the high energy of the South African coastprove limiting to the distribution of C. gigas, the yet undeterminedimpact of this species may be focused on estuarine habitats and noton the coastal intertidal as has been reported globally (Shatkin etal. 1997).
As the presence of naturalized C. gigas along the South Africancoast has only just been discovered, the rate of spread of thisspecies remains unknown, as do the ecologic implications of thisinvasion. Internationally, impacts resulting from the invasion of C.gigas include the simultaneous introduction of associated fauna(Critchley et al. 1983, Kaiser et al. 1998), the introduction ofdisease organisms (Ford 1992), genetic pollution of local oysterspecies (Gaffney and Allen 1992, Gaffney and Allen 1993) and thereduction of indigenous oyster populations to threatened levels(Williams et al. 1988, Mann et al.1991). It is thus clear, that theinvasion of the South African coast by C. gigas requires urgentscientific attention if its environmental impacts are to be managedresponsibly.
ACKNOWLEDGMENTS
The authors thank South African National Parks for facilitatingsampling within the Knysna Estuary and Dr. Anesh Govender forproviding valuable statistical assistance. Funding for this study wasprovided by the National Research Foundation and Marine andCoastal Management, through a grant to C L Griffiths from the Seaand the Coast 2 Programme. The genetic analyses were funded by aNational Research Foundation and Marine and Coastal Managementsabbatical grant to P Bloomer and a SeaGrant to M P Hare.
LITERATURE CITED
Andrews, J. 1980. A review of introductions of exotic oysters and biologi-cal planning for new importations. Mar. Fish. Rev. 42:1–11.
Banks, M. A., D. Hedgecock & C. Waters. 1993. Discrimination betweenclosely related Pacific oyster spp. (Crassostrea) via mitochondrialDNA sequences encoding for large subunit rRNA. Mol. Mar. Biol.Biotechnol 2:129–136.
Carlton, J. T., J. K. Thompson, L. E. Schemel & F. H. Nichols. 1990. Re-markable invasion of San Francisco Bay (California, USA) by theAsian clam Potamocorbula amurensis. I. Introduction and dispersal.Mar. Ecol. Prog. Ser. 66:81–94.
Cosby, M. P. & L. D. Gale. 1990. A review and evaluation of bivalve
condition index methodologies with a suggested standard method. J.Shellfish Res. 9:233–237.
Critchley, A. T., W. F. Farnham & S. L. Morrell. 1983. A chronology ofnew European sites of attachment for the invasive brown algae Sar-gassum muticum, 1973–1981. J. Mar. Biol. Assoc. UK. 63:799–811.
Diez, B., C. Pedros-Alio, T. L. Marsh & R. Massana. 2001. Application ofdenaturing gradient gel electrophoresis (DGGE) to study the diversityof marine picoeukaryotic assemblages and comparison of DGGE withother molecular techniques. Appl. Env. Microbiol. 67:2942–2951.
Dinamani, P. 1971. Occurrence of the Japanese oyster, Crassostrea gigas(Thunberg), in Northland, New Zealand. New Zealand J. Mar. Fresh-water Res. 5:352–357.
Figure 6. Mean condition indices (±standard deviation) of C. gigas theBreede, Goukou and Knysna estuaries.
NATURALIZED CRASSOSTREA GIGAS IN SOUTH AFRICA 449

Felsenstein, J. 1985. Confidence limits on phylogenies: an approach usingthe bootstrap. Evolution 39:783–791.
Ford, S. E. 1992. Avoiding the transmission of disease in commercialculture of mollusks, with special reference to Perkinsus marinus(Dermo) and Haplosporidium nelsoni (MSX). J. Shellfish Res. 11:539–546.
Folmer, O., M. Black, W. Hoeh, R. Lutz & R. Vrijenhoek. 1994. DNAprimers for amplification of mitochondrial cytochrome c oxidase sub-unit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotech.3:294–299.
Gaffney, P. M. & S. K. Allen. 1992. Genetic effects of introduction andtransfer of mollusks. J. Shellfish Res. 11:535–538.
Gaffney, P. M. & S. K. Allen. 1993. Hybridization among Crassostreaspecies: a review. Aquaculture 116:1–13.
Griffiths, C. L., P. A. R. Hockey, C. Van Erkom Schurink, & P. J. LeRoux. 1992. Marine invasive aliens on South African shores: Implica-tions for community structure and trophic functioning. S. Afr. J. MarSci 12:713–722.
Jozefowicz, C. J. & D. O. Foighil. 1998. Phylogenetic analysis of SouthernHemisphere flat oysters based on partial mitochondrial 16S rDNA genesequences. Mol. Phylogenet. Evol. 10:426–435.
Kaiser, M. J., I. Laing, D. S. Utting & G. M. Burnell. 1998. Environmentalimpacts of bivalve mariculture. J. Shellfish Res. 17:59–66.
Kimura, M. 1980. A simple method for estimating evolutionary rates ofbase substitutions through comparative studies of nucleotide se-quences. J. Mol. Evol. 16:111–120.
Korringa, P. 1956. Oyster culture in South Africa. Hydrographical, bio-logical and ostreological observations in the Knysna Lagoon, withnotes on conditions in other South African waters. Invest. Rep. Div.Fish. S. Afr. 20:84.
Kumar, T., K. Tamura, I. Jakobsen, & M. Nei. 2001. MEGA2: MolecularEvolutionary Genetic Analysis software. Bioinformatics 17:1244–1245.
Lane, D. J., B. Pace, G. J. Olsen, D. A. Stahl, M. L. Sogin & N. R. Pace.1985. Rapid determination of 16S ribosomal RNA sequences for phy-logenetic analyses. Proc. Natl. Acad. Sci. USA 82:6955–6959.
Lapegue, S., I. Boutet, A. Leitao, S. Heurtebise, P. Garcia, C. Thiriot-Quievreux & P. Boudry. 2002. Trans-Atlantic distribution of a man-grove oyster species revealed by 16S mtDNA and karyological analy-ses. Biol. Bull. 202:232–242.
Le Roux, P. J., G. M. Branch & M. A. P. Joska. 1990. On the distribution,diet and possible impact of the invasive European shore crab Carcinusmaens (L.) along the South African Coast. S. Afr. J. Mar. Sci. 9:85–93.
Lutjeharms, J. R. E. 1998. Coastal hydrography. In: R. Lubke & I. Moor,editors. Field guide to the Eastern and Southern Cape coasts. CapeTown, South Africa: University of Cape Town Press. pp. 50–61.
Mann, R., E. Burreson & P. Baker. 1991. The decline of the Virginia oysterfishery in Chesapeake Bay: considerations for introduction of a non-endemic species, Crassostrea gigas (Thunberg 1793). J. Shellfish Res.10:379–388.
Mann, R. 1983. The role of introduced bivalve mollusk species in mari-culture. J. World Mariculture Soc. 14:546–559.
Matsumoto, M. 2003. Phylogenetic analysis of the subclass Pteriomorpha(Bivalvia) from mtDNA COI sequences. Mol. Phylogenet. Evol. 27:429–440.
Menzel, W. 1987. Hybridization of oysters and clams. In: K. Tiews, editor.Selection, hybridization, and genetic engineering in Aquaculture. Vol.2. Berlin:Heeneman. pp. 47–59.
Minchin, D. 1996. Management of the introduction and transfer of marinemollusks. Aquatic Conservation. Mar. Freshwater Eco. 6:229–244.
Rainer, J. S. & R. Mann. 1992. A comparison of methods for calculatingcondition index in Eastern Oysters, Crassostrea virginica (Gmelin,1791). J. Shellfish Res. 11:55–58.
Robinson, T. B., C. L. Griffiths, C. D. McQuaid, & M. Ruis. In Press.Marine alien species of South Africa—status and impacts. Afr. J. MarSci.
Saiki, R. K., D. H. Gelfand, S. Stoffel, S. J. Scharf, R. Higuchi, G. T. Horn,K. B. Mullis & H. A. Erlich. 1988. Primer-directed enzymatic ampli-fication of DNA with a thermostabile DNA polymerase. Science 239:487–491.
Saitou, N. & M. Nei. 1987. The neighbor-joining method: a new methodfor reconstructing phylogenetic trees. Mol. Biol. Evol. 4:406–425.
Shatkin, G., S. E. Shumway & R. Hawes. 1997. Considerations regardingthe possible introduction of the Pacific oyster (Crassostrea gigas) tothe gulf of Maine: A review of global experience. J. Shellfish Res.16:463–477.
Steffani, C. N. & G. M. Branch. 2003a. Growth rate, condition, and shellshape of Mytilus galloprovincialis: responses to wave exposure. Mar.Ecol. Prog. Ser. 246:197–209.
Steffani, C. N. & G. M. Branch. 2003b. Spatial comparisons of populationsof an indigenous limpet Scutellastra argenvillei and the alien musselMytilus galloprovincialis along a gradient of wave energy. S. Afr. J.Mar. Sci. 25:195–212.
Steffani, C. N. & G. M. Branch. 2003c. Temporal changes in an interactionbetween an indigenous limpet Scutellastra argenvillei and an alienmussel Mytilus galloprovincialis: effects of wave exposure. S. Afr. J.Mar. Sci. 25:213–229.
Thompson, J. D., T. J. Gibson, F. Plewniak, F. Jeanmougin & D. G. Hig-gins. 1997. The ClustalX windows interface: flexible strategies formultiple sequence alignment aided by quality analysis tools. Nucl. Ac-ids Res. 24:4876–4882.
Thomson, J. 1959. The naturalization of the Pacific oyster in Australia.Aust. J. Mar. Freshwater Res. 10:144–149.
Wehrmann, A., M. Herlyn, F. Bungenstock, G. Hertweck & G. Millat.2000. The distribution gap is closed—first record of naturally settledPacific oysters Crassostrea gigas in the east Frisian Wadden Sea, NorthSea. Senckenbergiana Marit. 30:153–160.
Williams, R. J., F. B. Griffiths, E. J. Van Der Wal, & J. Kelly. 1988. Cargovessel ballast water as a vector for the transport of non-indigenousmarine species. Estuarine, Coastal and Shelf Science 27:409–420.
ROBINSON ET AL.450





























