Embed Size (px)
Citation preview

© 2002 Blackwell Science Ltd
479
Parasite Immunology
,
2002,
24
, 479–488
Blackwell Publishing Ltd.
ORIGINAL ARTICLE
NEUTROPENIA IN
S. JAPONICUM
EGG GRANULOMA FORMATION
Neutropenia augments experimentally induced
Schistosoma japonicum
egg granuloma formation in CBA mice, but not in
C57BL/6 mice
MIZUKI HIRATA
1
, TATSURU HARA
1
, MASAYOSHI KAGE
2
, TOSHIHIDE FUKUMA
1
& FUJIRO SENDO
3
1
Department of Parasitology and
2
Department of Pathology, Kurume University School of Medicine, Kurume, Fukuoka, Japan and
3
Department of Immunology and Parasitology, Yamagata University School of Medicine, Yamagata, Japan
SUMMARY
The present study was designed to investigate the role of neu-trophils during the development of
Schistosoma japonicum
egg granulomas, in C57BL/6 and CBA mice. Laid eggs wereimplanted into the liver and monoclonal antibody, RB6-8C5,was used to eliminate neutrophils. After daily antibody treat-ment between days 9 and 13 of egg implantation, both strainsof mice showed a marked decrease in neutrophil infiltrationand coagulative hepatocyte necrosis at 2 weeks. At 4 weeks,after antibody administration every other day between days16 and 26, granuloma formation in C57BL/6 mice was notaffected by the treatment, whereas CBA mice exhibited a sig-nificant increase of reactions. Neutropenia augmented the Th2cytokine response (IL-4, IL-13 and IL-5), but not for IFN-
γ
at any time point examined and in either strain of mice.Higher levels of IL-4 and IL-13 were noted in CBA mice atearly and late stages of granuloma formation, compared toC57BL/6 mice. There was also a striking difference in IL-13production between the two strains. Our results indicate thatneutropenia is associated with a significant augmentation of
S. japonicum
egg-induced granuloma formation in CBA mice,probably through increase in Th2 cytokines, however, theeffects differ between early and late stages and between highand low responders.
Keywords
neutrophils
, Schistosoma japonicum,
granulomaformation, Th2 cytokines, CBA mouse, C57BL/6 mouse
INTRODUCTION
Schistosoma japonicum
is prevalent in South-east Asia andis one of the major schistosome species world-wide, causingsevere intestinal and hepatosplenic disease. Schistosome wormsthat settle in the portal vein continually lay eggs that aredeposited in tissues of the intestine and liver, and provoke aserious granulomatous hypersensitivity reaction. Studieshave indicated that the reactions are mediated by CD4
+
Thelper cells (1) and Th2 type cytokines, such as interleukin(IL)-4 and IL-13 (2). Using the hepatic model of granulomaformation, which is experimentally induced by the implanta-tion of
in vitro
laid eggs, we have previously shown that theeggs have a strong granuloma-evoking capacity relative tolive eggs extracted from tissues and that marked neutrophilinfiltration occurs between 8 and 14 days after egg implanta-tion (stages of abscess formation), corresponding to the timeof egg maturation (3). With time, neutrophil infiltrationdiminishes and eosinophils and mononuclear cells becomepredominant. At 4 weeks, development of fibrosis becomesobvious (stages of granuloma formation), although delayedtype hypersensitivity (DTH) response and the magnitude oftissue reactivity differ considerably between high (CBAmice) and low responders (C57BL/6 mice) (4).
Neutrophils have been reported to play a crucial role inthe defence against parasitic infections, such as those causedby
Entamoeba histolytica
(5,6),
Leishmania major
(7), and
Leishmania donovani
(8) and to be cytotoxic to
Schistosomamansoni
eggs (9,10), as well as bacterial infection. They alsoparticipate in the pathogenesis of various infectious diseasesor physiological disorders, as emphasized by recent resultsthat neutrophils can produce TNF-
α
, IL-1
α
, IL-10 and IL-12 (11,12), a crucial inducer of IFN-
γ
, and induce MCP-1(13). These findings strongly suggest that neutrophils play arole during the course of schistosome egg-induced granulomaformation; however, little is known about how neutrophilsparticipate in this process.
Correspondence
: Mizuki Hirata, Ph.D., Department of Parasitology, Kurume University School of Medicine, Kurume, Fukuoka 830-0011, Japan (e-mail: [email protected]).
Received
: 11 October 2002
Accepted for publication
: 22 November 2002

480
© 2002 Blackwell Science Ltd,
Parasite Immunology
,
24
, 479–488
M. Hirata
et al
. Parasite Immunology
In order to elucidate the role of neutrophils during
S.japonicum
egg granuloma formation, a mouse hepatic modelthat was induced by implantation of
in vitro
laid eggs wasemployed. We used a monoclonal antibody (RB6-8C5) toeliminate neutrophils and analysed tissue reactions (cell-ularity and magnitude). Studies were performed at early andlate stages of egg implantation when different cellularity(abscess and granuloma formation) was observed. We usedtwo mouse strains, C57BL/6 and CBA, because they displaydifferent DTH responses (4). Particular attention was paidto the effect of neutrophil depletion on Th2 cytokines, IL-4and IL-13. Our results demonstrate that neutrophil depletionsignificantly alters granulomatous reactions and augmentsthe Th2 cytokine response. However, the effect differs consider-ably between different stages and between mouse strains.
MATERIALS AND METHODS
Animals and parasites
The animals were all female CBA and C57BL/6 mice, aged9–12 weeks. They were purchased from the Shizuoka Labo-ratory Animal Centre, Japan, and were bred and maintainedusing standard laboratory care procedures. A Japanesestrain of
S. japonicum
was maintained in our laboratory bypassage through
Oncomelania hupensis nosophora
and rab-bits for 25 years. The experimental protocol was approvedby the Ethics Review Committee for Animal Experimenta-tion of Kurume University School of Medicine.
Induction of granuloma formation
Hepatic granulomas were induced by implantation of eggs(3). Freshly laid
S. japonicum
eggs were obtained frommated worms taken from infected rabbits and were culturedfor 2 days in RPMI1640 medium containing 10% foetalbovine serum (FBS). After washing with serum-freeRPMI1640, 1000 eggs were implanted into the livers of micethrough the caecal vein.
Depletion of neutrophils
The hybridoma was a kind gift from Dr R. Coffman(DNAX Research Institute, Palo Alto, CA). RB6-8C5 mon-oclonal antibody was obtained from the culture supernatant(ASF-103, Ajinomoto, Japan) and purified using a methoddescribed previously by Chen
et al
. (14) and stored at
−
80
°
C. The monoclonal antibody was intra-peritoneallyadministered at an initial dose of 0·1 mg/mouse, and laterthe dose was increased to 0·2 mg/mouse. In control mice, thesame concentration of normal rat IgG (Caltag Laboratories,Burlingame, CA) was administered. Each group of mice
consisted of 3–4 animals. To examine the effect of antibodytreatment, a peripheral blood smear was prepared the dayfollowing the last injection and stained with Giemsa. Experi-ments were repeated twice or thrice.
Histopathological examination
For histological examination, the liver was fixed in 10%neutral buffered formalin, embedded in paraffin wax, andstained with haematoxylin and eosin. To assess fibrosis,sections were stained with Sirius red (0·1% Sirius red F3BAcolour index (CI) = 35780) (15). For evaluation of tissuereactivity, lesions larger than 100
µ
m in diameter wereselected, as disintegrated eggs induced only minimal reac-tions. Diameters (mean width and length of the lesion) weremeasured using a micrometer. In each specimen, 25–30 lesionswere measured and the mean values were determined.
Cytokine measurement
Cytokine production in spleen cells was examined asdescribed previously (2). Briefly, 5
×
10
6
nucleated cells /wellwere plated in 24-well plates and challenged with 2·5
µ
g /mLsoluble egg antigen. After the total volume was adjusted to2 mL, plates were incubated for 48 h at 37
°
C in a humidifiedCO
2
incubator. Supernatant fluids were harvested aftercentrifugation, aliquoted and stored at
−
80
°
C until use.Cytokines and chemokines investigated included IL-4, IL-5,IL-13, and IFN-
γ
, and their levels were determined by sand-wich enzyme-linked immunosorbent assay (ELISA) usingcommercially available kits. The kit used for determinationof IL-5 was obtained from Genzyme (Cambridge, MA),while reagents for the other cytokines were from R & DSystems (Minneapolis, MN). The samples were diluted 5- or10-fold when values reached the upper limits of the assaysystem. Five to six samples obtained from separate experi-ments were collectively tested for each time point.
Statistical analysis
All data were expressed as mean
±
SD. Differences betweengroups were examined for statistical significance using theMann–Whitney
U
-test. A
P
-value less than 0·05 denoted thepresence of a statistically significant difference.
RESULTS
Effect on abscess formation
Marked neutrophil infiltration around eggs was commonlyobserved during the early stage of
S. japonicum
infection.This histological feature was most marked at 2 weeks in
© 2002 Blackwell Science Ltd,
Parasite Immunology
,
24
, 479–488
481
Volume 24, Number 9/10, September/October 2002 Neutropenia in
S. japonicum
egg granuloma formation
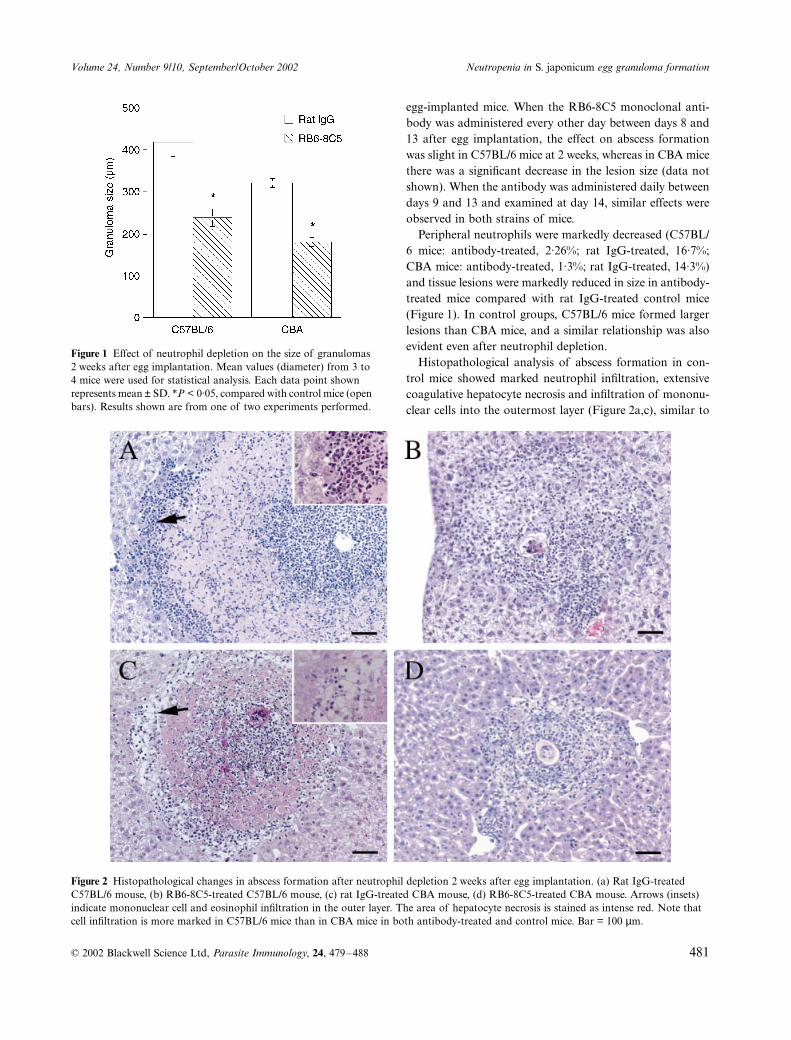
egg-implanted mice. When the RB6-8C5 monoclonal anti-body was administered every other day between days 8 and13 after egg implantation, the effect on abscess formationwas slight in C57BL/6 mice at 2 weeks, whereas in CBA micethere was a significant decrease in the lesion size (data notshown). When the antibody was administered daily betweendays 9 and 13 and examined at day 14, similar effects wereobserved in both strains of mice.
Peripheral neutrophils were markedly decreased (C57BL/6 mice: antibody-treated, 2·26%; rat IgG-treated, 16·7%;CBA mice: antibody-treated, 1·3%; rat IgG-treated, 14·3%)and tissue lesions were markedly reduced in size in antibody-treated mice compared with rat IgG-treated control mice(Figure 1). In control groups, C57BL/6 mice formed largerlesions than CBA mice, and a similar relationship was alsoevident even after neutrophil depletion.
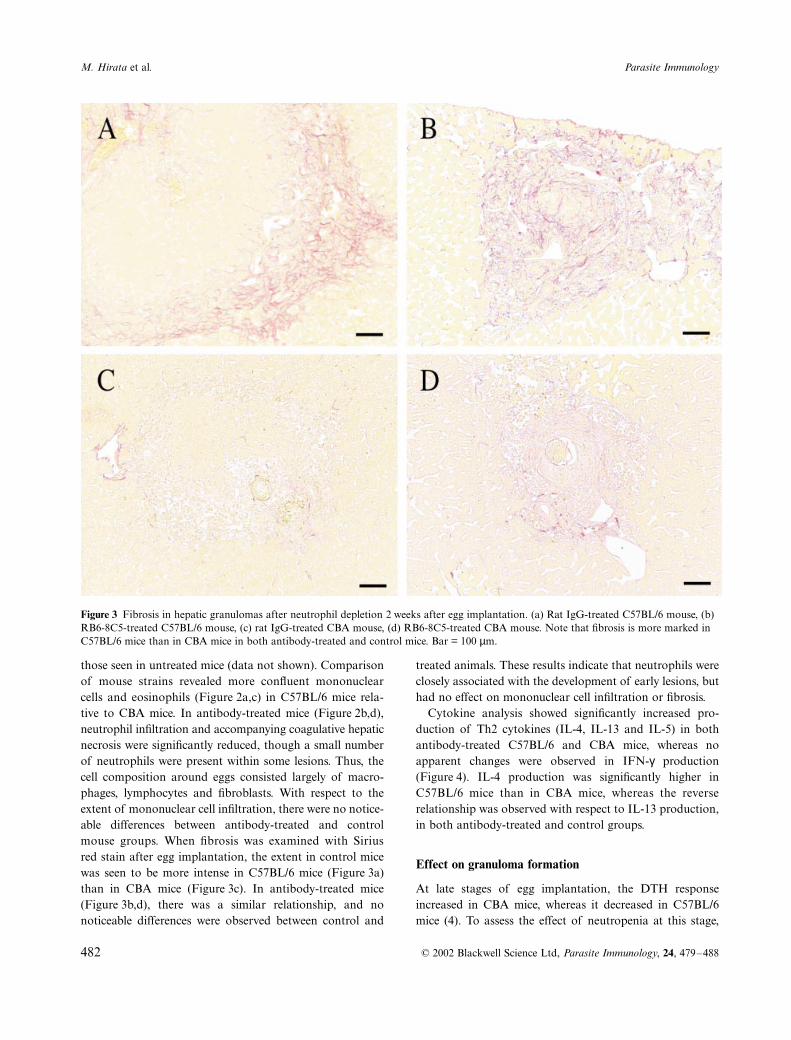
Histopathological analysis of abscess formation in con-trol mice showed marked neutrophil infiltration, extensivecoagulative hepatocyte necrosis and infiltration of mononu-clear cells into the outermost layer (Figure 2a,c), similar to
Figure 2 Histopathological changes in abscess formation after neutrophil depletion 2 weeks after egg implantation. (a) Rat IgG-treated C57BL/6 mouse, (b) RB6-8C5-treated C57BL/6 mouse, (c) rat IgG-treated CBA mouse, (d) RB6-8C5-treated CBA mouse. Arrows (insets) indicate mononuclear cell and eosinophil infiltration in the outer layer. The area of hepatocyte necrosis is stained as intense red. Note that cell infiltration is more marked in C57BL/6 mice than in CBA mice in both antibody-treated and control mice. Bar = 100 µm.
Figure 1 Effect of neutrophil depletion on the size of granulomas 2 weeks after egg implantation. Mean values (diameter) from 3 to 4 mice were used for statistical analysis. Each data point shown represents mean ± SD. *P < 0·05, compared with control mice (open bars). Results shown are from one of two experiments performed.
482
© 2002 Blackwell Science Ltd,
Parasite Immunology
,
24
, 479–488
M. Hirata
et al
. Parasite Immunology
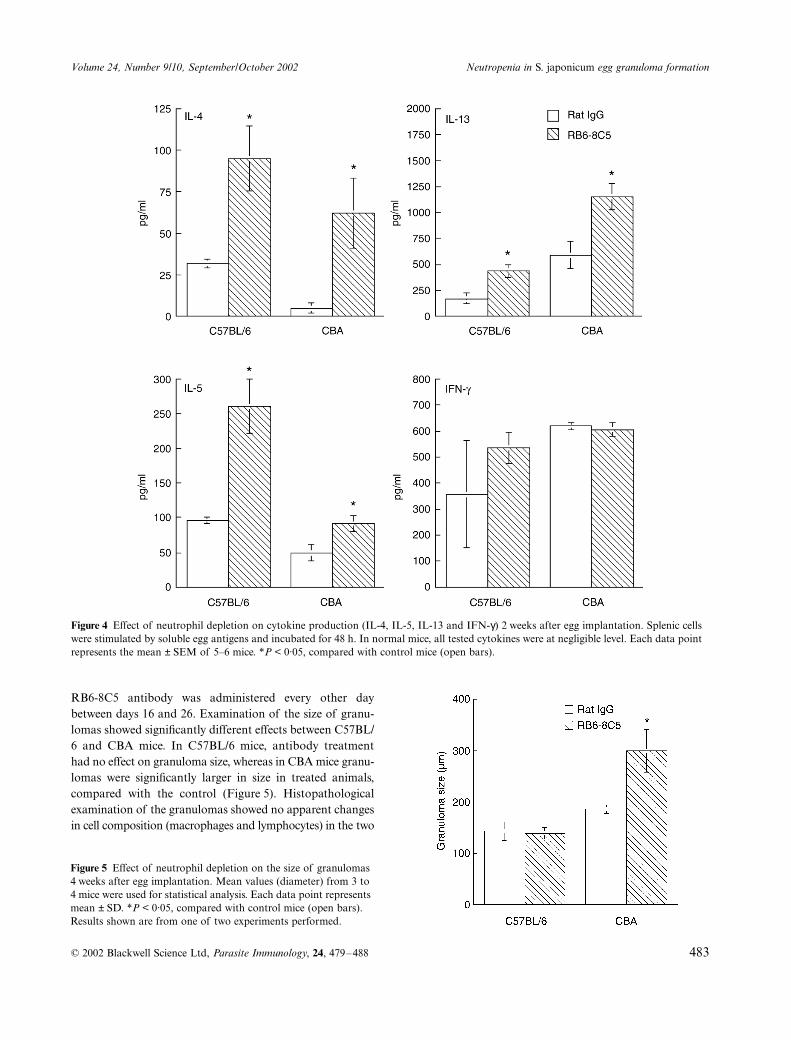
those seen in untreated mice (data not shown). Comparisonof mouse strains revealed more confluent mononuclearcells and eosinophils (Figure 2a,c) in C57BL/6 mice rela-tive to CBA mice. In antibody-treated mice (Figure 2b,d),neutrophil infiltration and accompanying coagulative hepaticnecrosis were significantly reduced, though a small numberof neutrophils were present within some lesions. Thus, thecell composition around eggs consisted largely of macro-phages, lymphocytes and fibroblasts. With respect to theextent of mononuclear cell infiltration, there were no notice-able differences between antibody-treated and controlmouse groups. When fibrosis was examined with Siriusred stain after egg implantation, the extent in control micewas seen to be more intense in C57BL/6 mice (Figure 3a)than in CBA mice (Figure 3c). In antibody-treated mice(Figure 3b,d), there was a similar relationship, and nonoticeable differences were observed between control and
treated animals. These results indicate that neutrophils wereclosely associated with the development of early lesions, buthad no effect on mononuclear cell infiltration or fibrosis.
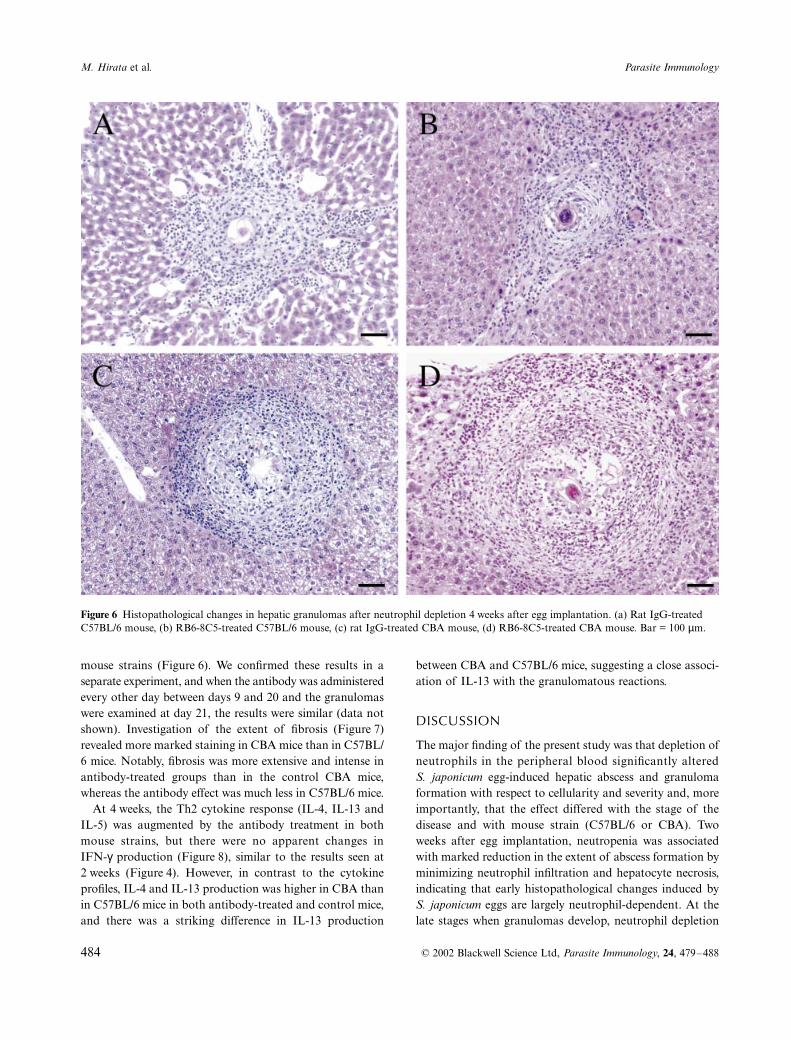
Cytokine analysis showed significantly increased pro-duction of Th2 cytokines (IL-4, IL-13 and IL-5) in bothantibody-treated C57BL/6 and CBA mice, whereas noapparent changes were observed in IFN-
γ
production(Figure 4). IL-4 production was significantly higher inC57BL/6 mice than in CBA mice, whereas the reverserelationship was observed with respect to IL-13 production,in both antibody-treated and control groups.
Effect on granuloma formation
At late stages of egg implantation, the DTH responseincreased in CBA mice, whereas it decreased in C57BL/6mice (4). To assess the effect of neutropenia at this stage,
Figure 3 Fibrosis in hepatic granulomas after neutrophil depletion 2 weeks after egg implantation. (a) Rat IgG-treated C57BL/6 mouse, (b) RB6-8C5-treated C57BL/6 mouse, (c) rat IgG-treated CBA mouse, (d) RB6-8C5-treated CBA mouse. Note that fibrosis is more marked in C57BL/6 mice than in CBA mice in both antibody-treated and control mice. Bar = 100 µm.
© 2002 Blackwell Science Ltd,
Parasite Immunology
,
24
, 479–488
483
Volume 24, Number 9/10, September/October 2002 Neutropenia in
S. japonicum
egg granuloma formation
RB6-8C5 antibody was administered every other daybetween days 16 and 26. Examination of the size of granu-lomas showed significantly different effects between C57BL/6 and CBA mice. In C57BL/6 mice, antibody treatmenthad no effect on granuloma size, whereas in CBA mice granu-lomas were significantly larger in size in treated animals,compared with the control (Figure 5). Histopathologicalexamination of the granulomas showed no apparent changesin cell composition (macrophages and lymphocytes) in the two
Figure 4 Effect of neutrophil depletion on cytokine production (IL-4, IL-5, IL-13 and IFN-γ) 2 weeks after egg implantation. Splenic cells were stimulated by soluble egg antigens and incubated for 48 h. In normal mice, all tested cytokines were at negligible level. Each data point represents the mean ± SEM of 5–6 mice. *P < 0·05, compared with control mice (open bars).
Figure 5
Effect of neutrophil depletion on the size of granulomas 4 weeks after egg implantation. Mean values (diameter) from 3 to 4 mice were used for statistical analysis. Each data point represents mean
±
SD. *
P
< 0·05, compared with control mice (open bars). Results shown are from one of two experiments performed.
484
© 2002 Blackwell Science Ltd,
Parasite Immunology
,
24
, 479–488
M. Hirata
et al
. Parasite Immunology
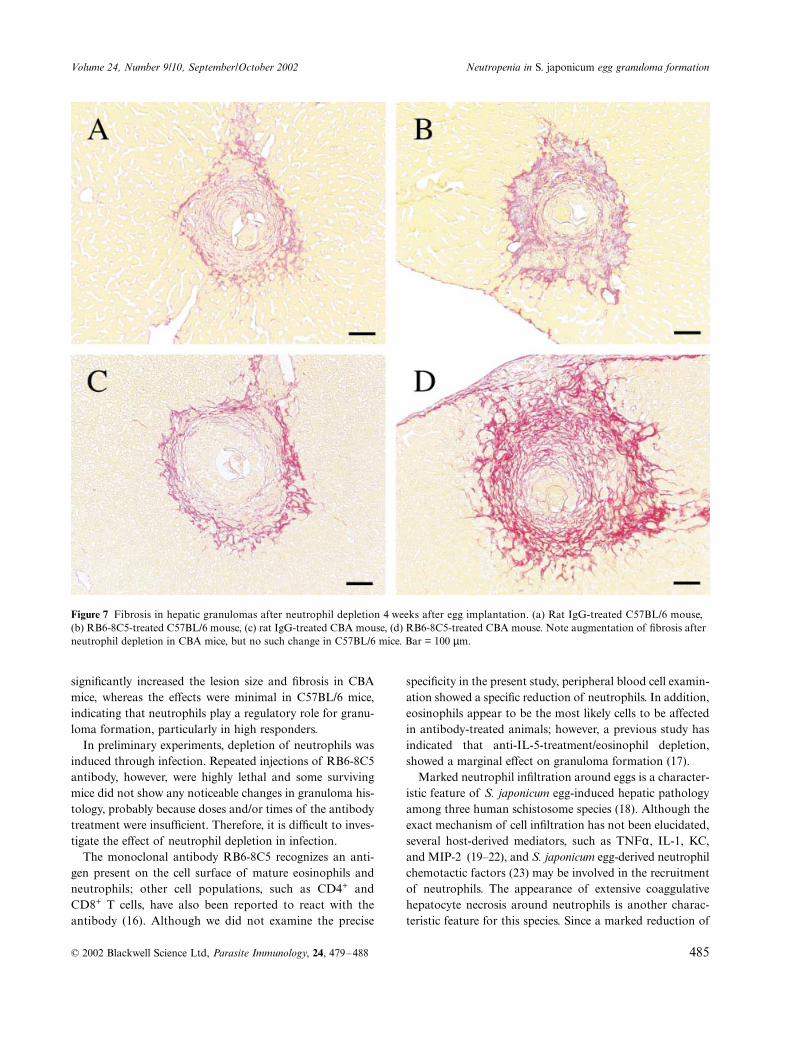
mouse strains (Figure 6). We confirmed these results in aseparate experiment, and when the antibody was administeredevery other day between days 9 and 20 and the granulomaswere examined at day 21, the results were similar (data notshown). Investigation of the extent of fibrosis (Figure 7)revealed more marked staining in CBA mice than in C57BL/6 mice. Notably, fibrosis was more extensive and intense inantibody-treated groups than in the control CBA mice,whereas the antibody effect was much less in C57BL/6 mice.
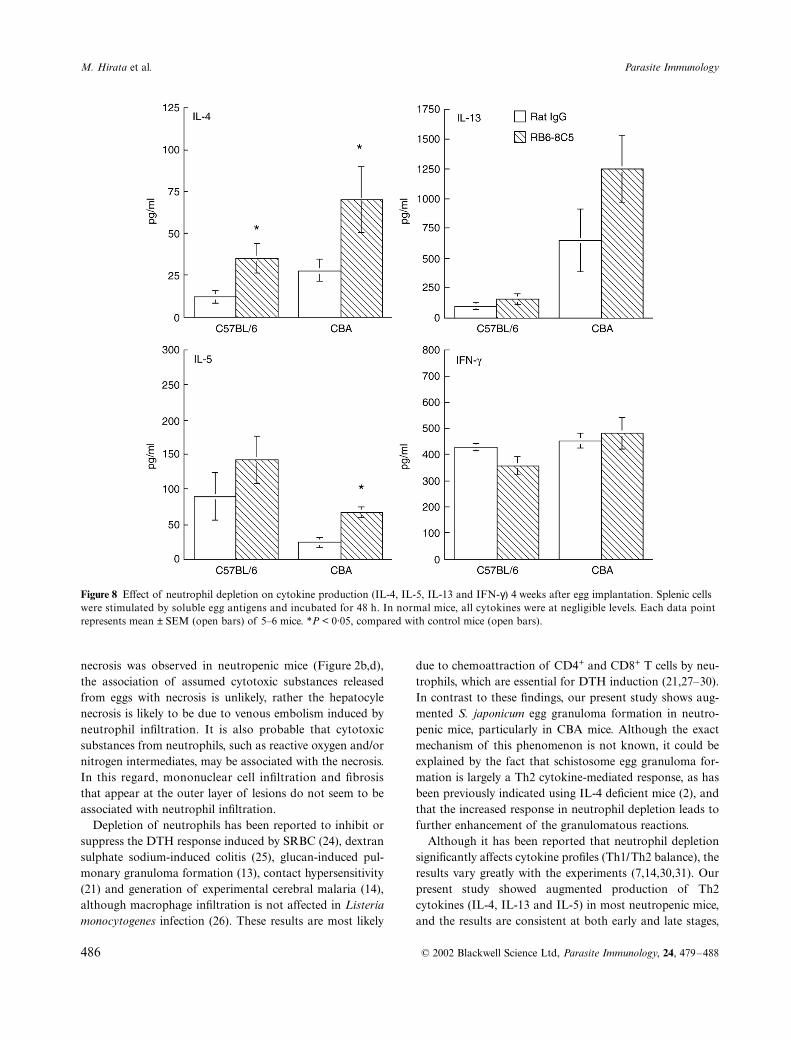
At 4 weeks, the Th2 cytokine response (IL-4, IL-13 andIL-5) was augmented by the antibody treatment in bothmouse strains, but there were no apparent changes inIFN-
γ
production (Figure 8), similar to the results seen at2 weeks (Figure 4). However, in contrast to the cytokineprofiles, IL-4 and IL-13 production was higher in CBA thanin C57BL/6 mice in both antibody-treated and control mice,and there was a striking difference in IL-13 production
between CBA and C57BL/6 mice, suggesting a close associ-ation of IL-13 with the granulomatous reactions.
DISCUSSION
The major finding of the present study was that depletion ofneutrophils in the peripheral blood significantly altered
S. japonicum
egg-induced hepatic abscess and granulomaformation with respect to cellularity and severity and, moreimportantly, that the effect differed with the stage of thedisease and with mouse strain (C57BL/6 or CBA). Twoweeks after egg implantation, neutropenia was associatedwith marked reduction in the extent of abscess formation byminimizing neutrophil infiltration and hepatocyte necrosis,indicating that early histopathological changes induced by
S. japonicum
eggs are largely neutrophil-dependent. At thelate stages when granulomas develop, neutrophil depletion
Figure 6 Histopathological changes in hepatic granulomas after neutrophil depletion 4 weeks after egg implantation. (a) Rat IgG-treated C57BL/6 mouse, (b) RB6-8C5-treated C57BL/6 mouse, (c) rat IgG-treated CBA mouse, (d) RB6-8C5-treated CBA mouse. Bar = 100 µm.
© 2002 Blackwell Science Ltd,
Parasite Immunology
,
24
, 479–488
485
Volume 24, Number 9/10, September/October 2002 Neutropenia in
S. japonicum
egg granuloma formation
significantly increased the lesion size and fibrosis in CBAmice, whereas the effects were minimal in C57BL/6 mice,indicating that neutrophils play a regulatory role for granu-loma formation, particularly in high responders.
In preliminary experiments, depletion of neutrophils wasinduced through infection. Repeated injections of RB6-8C5antibody, however, were highly lethal and some survivingmice did not show any noticeable changes in granuloma his-tology, probably because doses and/or times of the antibodytreatment were insufficient. Therefore, it is difficult to inves-tigate the effect of neutrophil depletion in infection.
The monoclonal antibody RB6-8C5 recognizes an anti-gen present on the cell surface of mature eosinophils andneutrophils; other cell populations, such as CD4
+
andCD8
+
T cells, have also been reported to react with theantibody (16). Although we did not examine the precise
specificity in the present study, peripheral blood cell examin-ation showed a specific reduction of neutrophils. In addition,eosinophils appear to be the most likely cells to be affectedin antibody-treated animals; however, a previous study hasindicated that anti-IL-5-treatment/eosinophil depletion,showed a marginal effect on granuloma formation (17).
Marked neutrophil infiltration around eggs is a character-istic feature of
S. japonicum
egg-induced hepatic pathologyamong three human schistosome species (18). Although theexact mechanism of cell infiltration has not been elucidated,several host-derived mediators, such as TNF
α
, IL-1, KC,and MIP-2 (19–22), and
S. japonicum
egg-derived neutrophilchemotactic factors (23) may be involved in the recruitmentof neutrophils. The appearance of extensive coaggulativehepatocyte necrosis around neutrophils is another charac-teristic feature for this species. Since a marked reduction of
Figure 7 Fibrosis in hepatic granulomas after neutrophil depletion 4 weeks after egg implantation. (a) Rat IgG-treated C57BL/6 mouse, (b) RB6-8C5-treated C57BL/6 mouse, (c) rat IgG-treated CBA mouse, (d) RB6-8C5-treated CBA mouse. Note augmentation of fibrosis after neutrophil depletion in CBA mice, but no such change in C57BL/6 mice. Bar = 100 µm.
486
© 2002 Blackwell Science Ltd,
Parasite Immunology
,
24
, 479–488
M. Hirata
et al
. Parasite Immunology
necrosis was observed in neutropenic mice (Figure 2b,d),the association of assumed cytotoxic substances releasedfrom eggs with necrosis is unlikely, rather the hepatocylenecrosis is likely to be due to venous embolism induced byneutrophil infiltration. It is also probable that cytotoxicsubstances from neutrophils, such as reactive oxygen and/ornitrogen intermediates, may be associated with the necrosis.In this regard, mononuclear cell infiltration and fibrosisthat appear at the outer layer of lesions do not seem to beassociated with neutrophil infiltration.
Depletion of neutrophils has been reported to inhibit orsuppress the DTH response induced by SRBC (24), dextransulphate sodium-induced colitis (25), glucan-induced pul-monary granuloma formation (13), contact hypersensitivity(21) and generation of experimental cerebral malaria (14),although macrophage infiltration is not affected in
Listeriamonocytogenes
infection (26). These results are most likely
due to chemoattraction of CD4
+
and CD8
+
T cells by neu-trophils, which are essential for DTH induction (21,27–30).In contrast to these findings, our present study shows aug-mented
S. japonicum
egg granuloma formation in neutro-penic mice, particularly in CBA mice. Although the exactmechanism of this phenomenon is not known, it could beexplained by the fact that schistosome egg granuloma for-mation is largely a Th2 cytokine-mediated response, as hasbeen previously indicated using IL-4 deficient mice (2), andthat the increased response in neutrophil depletion leads tofurther enhancement of the granulomatous reactions.
Although it has been reported that neutrophil depletionsignificantly affects cytokine profiles (Th1/Th2 balance), theresults vary greatly with the experiments (7,14,30,31). Ourpresent study showed augmented production of Th2cytokines (IL-4, IL-13 and IL-5) in most neutropenic mice,and the results are consistent at both early and late stages,
Figure 8 Effect of neutrophil depletion on cytokine production (IL-4, IL-5, IL-13 and IFN-γ) 4 weeks after egg implantation. Splenic cells were stimulated by soluble egg antigens and incubated for 48 h. In normal mice, all cytokines were at negligible levels. Each data point represents mean ± SEM (open bars) of 5–6 mice. *P < 0·05, compared with control mice (open bars).

© 2002 Blackwell Science Ltd,
Parasite Immunology
,
24
, 479–488
487
Volume 24, Number 9/10, September/October 2002 Neutropenia in
S. japonicum
egg granuloma formation
and in C57BL/6 and CBA mice. These different observa-tions may be attributable to differences in stimulatoryagents or cells and organs studied, or types of inflammation.With respect to the potential association of neutrophils withTh1/Th2 balance, neutrophils are an important source ofIL-12 and also of other cytokines (11,31). It has been shownthat neutrophil depletion in Legionella pneumophila infectedmice results in a decrease of IFN-γ production and IL-12,and increased Th2 cytokines (31). In our study, however,no noticeable changes were observed in IFN-γ production.Further studies are needed to investigate the association ofneutrophils with the Th2 cytokine response.
Although IL-4 and IL-13 have many similar biologicalfunctions (32), recent studies have indicated distinct func-tions for each cytokine. In studies of S. mansoni-inducedgranulomatous inflammation, IL-13 is associated with thedevelopment of fibrosis in IL-13 knockout mice or sIL-13Rα2-Fc-treated animals (33–35). In the present study, weobserved a striking difference in IL-13 production betweenC57BL/6 and CBA mice, particularly at stages of granulomaformation. Given that granulomas are significantly largerand that there is intense fibrosis in CBA mice, our resultsappear to coincide with the reported profibrogenic roleof IL-13. Recent studies reported that IL-13 induces TGF-β, a major fibrosis inducer (36). In contrast, no associationbetween IL-13 and fibrosis was observed at stages of abscessformation, as CBA mice showed less fibrosis and a higherIL-13 expression, compared to C57BL/6 mice (Figures 3and 4). The fact that mononuclear cell infiltration wasapparently less in CBA mice than C57BL/6 mice may sug-gest that local cytokine production is more crucial thansplenic cell production presently examined, though there issome difficulty in collecting sufficient number of granulomacells in egg-implanted mice, because granulomas induced byegg implantation are relatively small in number. Furtherstudies using IL-13 deficient mice may clarify this point.
Finally, Cheever et al. (37) reported a strain differencein S. japonicum egg granuloma formation. In our prev-ious comparative study, CBA mice exhibited significantlylarger granulomas and a higher DTH response at stages ofgranuloma formation, compared to C57BL/6 mice (4). Thepresent study confirms our early results and further demon-strates that neutropenia does not affect the relationship,but rather amplifies it. Our results showed that the effect ofneutropenia differed in the two mouse strains, CBA andC57BL/6 mice. Granuloma-associated cytokines, IL-4and IL-13, increased in both strains after neutrophil deple-tion, but the increase was more marked in CBA mice(Figure 8). Our results also showed a marked differencein the production of a reportedly profibrogenic cytokine,IL-13, between the strains, particularly at stages of granu-loma formation.
ACKNOWLEDGEMENTS
We thank Drs R. Coffman (DNAX Research Institute,Palo Alto, CA), M. Suzuki (Gumma University, Japan), andR. L. Tarleton (University of Georgia, Athens, GA) forgenerously providing RB6-8C5 hybridoma. The authorsthank Manami Ohba for the excellent technical assistance.This work was supported in part by a Grant-in-Aid(14570227) from the Ministry of Education, Science andCulture, Japan.
REFERENCES
1 Cheever AW, Deb S, Duvall RH. Granuloma formation inSchistosoma japonicum infected nude mice: the effects of recon-stitution with L3T4+ or Lyt2+ splenic cells. Am J Trop Med Hyg1989; 40: 66–71.
2 Hirata M, Hirata K, Kage M, Zhang M, Hara T, Fukuma T.Schistosoma japonicum egg granuloma formation in the interleukin-4 or interferon-γ deficient host. Parasite Immunol 2001; 23: 271–280.
3 Hirata M, Takushima M, Kage M, Fukuma T. Induction ofexperimental murine granuloma formation against Schistosomajaponicum eggs produced by in vitro ova deposition, in vitro tissueextraction, or lyophilization. Parasitol Res 1991; 77: 315–319.
4 Hirata M, Kage M, Takushima M, Fukuma T. Different coursesof granulomatous reactions around Schistosoma japonicum eggsin three strains of mice. J Parasitol 1993; 79: 266–273.
5 Seydel KB, Zhang T, Stanley SL Jr. Neutrophils play a criticalrole in early resistance to amebic liver abscesses in severe com-bined immunodeficient mice. Infect Immun 1997; 65: 3951–3953.
6 Velazquez C, Shibayama-Salas M, Aguirre-Garcia J, Tsutsumi V,Calderon J. Role of neutrophils in innate resistance to Enta-moeba histolytica liver infection in mice. Parasite Immunol 1998;20: 255–262.
7 Tacchini-Cottier F, Zweifel C, Belkaid Y et al. An immu-nomodulatory function for neutrophils during the induction ofa CD4+ Th2 response in BALB/c mice infected with Leishmaniamajor. J Immunol 2000; 165: 2628–2636.
8 Smelt SC, Cotterell SE, Engwerda CR, Kaye PM. B cell-deficientmice are highly resistant to Leishmania donovani infection, butdevelop neutrophil-mediated tissue pathology. J Immunol 2000;164: 3681–3688.
9 De Brito PA, Kazura JW, Mahmoud AA. Host granulomat-ous response in schistosomiasis mansoni. Antibody and cell-mediated damage of parasite eggs in vitro. J Clin Invest 1984; 74:1715–1723.
10 Kazura JW, de Brito P, Rabbege J, Aikawa M. Role of granu-locyte oxygen products in damage of Schistosoma mansoni eggsin vitro. J Clin Invest 1985; 75: 1297–1307.
11 Romani L, Mencacci A, Cenci E et al. Neutrophil production ofIL-12 and IL-10 in candidiasis and efficacy of IL-12 therapy inneutropenic mice. J Immunol 1997; 158: 5349–5356.
12 Bliss SK, Butcher BA, Denkers EY. Rapid recruitment of neu-trophils containing prestored IL-12 during microbial infection.J Immunol 2000; 165: 4515–4521.
13 Kilgore KS, Powers KL, Imlay MM et al. The carbohydratesialyl Lewisx (sLex) sulfated glycomimetic GM2941 attenuatesglucan-induced pulmonary granulomatous vasculitis in the rat.J Pharmacol Exp Ther 1998; 286: 439– 446.

488 © 2002 Blackwell Science Ltd, Parasite Immunology, 24, 479–488
M. Hirata et al. Parasite Immunology
14 Chen L, Zhang Z, Sendo F. Neutrophils play a critical role inthe pathogenesis of experimental cerebral malaria. Clin ExpImmunol 2000; 120: 125–133.
15 Sweat F, Puchtler H, Rosenthal SI. Sirius red F3BA as a stainfor connective tissue. Arch Pathol 1964; 78: 69–72.
16 Goossens PL, Jouin H, Milon G. Dynamics of lymphocytes andinflammatory cells recruited in liver during murine listeriosis. Acytofluorimetric study. J Immunol 1991; 147: 3514–3520.
17 Sher A, Coffman RL, Hieny S, Scott P, Cheever AW. Inter-leukin 5 is required for the blood and tissue eosinophilia but notgranuloma formation induced by infection with Schistosomamansoni. Proc Natl Acad Sci USA 1990; 87: 61–65.
18 Von Lichtenberg F, Erickson DG, Sadun EH. Comparativehistopathology of schistosome granulomas in the hamster. AmJ Pathol 1973; 72: 149–178.
19 Butcher EC, Picke LJ. Lymphocyte homing and homeostasis.Science 1996; 272: 60–66.
20 Baggiolini M. Chemokines and leukocyte traffic. Nature 1998;392: 565–568.
21 Dilulio NA, Engeman T, Armstrong D, Tannenbaum C,Hamilton TA, Fairchild RL. Groalpha-mediated recruitment ofneutrophils is required for elicitation of contact hypersensitivity.Eur J Immunol 1999; 29: 3485–3495.
22 Zlotnik A, Yoshie O. Chemokines: a new classification systemand their role in immunity. Immunity 2000; 12: 121–127.
23 Owhashi M, Horii Y, Ishii A. Schistosoma japonicum:identification and characterization of neutrophil chemotacticfactors from egg antigen. Exp Parasitol 1985; 60: 229–238.
24 Kudo C, Yamashita T, Araki A et al. Modulation of in vivoimmune response by selective depletion of neutrophils using amonoclonal antibody, RP-3. I. Inhibition by RP-3 treatment ofthe priming and effector phases of delayed type hypersensitivityto sheep red blood cells in rats. J Immunol 1993; 150: 3728–3738.
25 Natsui M, Kawasaki K, Takizawa H et al. Selective depletionof neutrophils by a monoclonal antibody, RP-3, suppressesdextran sulphate sodium-induced colitis in rats. J GastroenterolHepatol 1997; 12: 801–808.
26 Rogers HW, Unanue ER. Neutrophils are involved in acute,nonspecific resistance to Listeria monocytogenes in mice. InfectImmun 1993; 61: 5090–5096.
27 Kudo C, Araki A, Matsushima K, Sendo F. Inhibition of IL-8-induced W3/25+ (CD4+) T lymphocyte recruitment intosubcutaneous tissues of rats by selective depletion of in vivoneutrophils with a monoclonal antibody. J Immunol 1991; 147:2196–2201.
29 Tanaka E, Sendo F. Abrogation of tumor-inhibitory MRC-OX8+ (CD8+) effector T-cell generation in rats by selectivedepletion of neutrophils in vivo using a monoclonal antibody.Int J Cancer 1993; 54: 131–136.
30 De Oca RM, De Buendia AJI, Rio L, Sanchez J, Salinas J,Navarro JA. Polymorphonuclear neutrophils are necessary forthe recruitment of CD8+ T cells in the liver in a pregnant mousemodel of Chlamydophila abortus (Chlamydia psittaci serotype 1)infection. Infect Immun 2000; 68: 1746–1751.
31 Tateda K, Moore TA, Deng JC et al. Early recruitment ofneutrophils determines subsequent T1/T2 host responses in amurine model of Legionella pneumophila pneumonia. J Immunol2001; 166: 3355–3361.
32 Zurawski G, de Vries JE. Interleukin 13, an interleukin 4-likecytokine that acts on monocytes and B cells, but not on T cells.Immunol Today 1994; 15: 19–26.
33 Chiaramonte MG, Donaldson DD, Cheever AW, Wynn TA.An IL-13 inhibitor blocks the development of hepatic fibrosisduring a T-helper type 2-dominated inflammatory response. JClin Invest 1999; 104: 777–785.
34 Fallon PG, Richardson EJ, McKenzie GJ, McKenzie AN.Schistosome infection of transgenic mice defines distinct andcontrasting pathogenic roles for IL-4 and IL-13: IL-13 is a profi-brotic agent. J Immunol 2000; 164: 2585–2591.
35 Chiaramonte MG, Cheever AW, Malley JD, Donaldson DD,Wynn TA. Studies of murine schistosomiasis reveal interleukin-13 blockade as a treatment for established and progressive liverfibrosis. Hepatology 2001; 34: 273–282.
36 Lee CG, Homer RJ, Zhu Z, Lanone S et al. Interleukin-13induces tissue fibrosis by selectively stimulating and activatingtransforming growth factor β1. J Exp Med 2001; 194: 809–821.
37 Cheever AW, Duvall RH, Hallack TA. Differences in hepaticfibrosis and granuloma size in several strains of mice infectedwith Schistosoma japonicum. Am J Trop Med Hyg 1984; 33:602–607.





























