Embed Size (px)
Citation preview

ANNALS OF ANATOMY
Normalization of keratinocyte-type integrins during the establishment of the oral mucosa phenotype
in vitro
Pascal Tomakidi, Dirk Breitkreutz 1, Annette Kohl and G. Komposch
Policlinic of Orthodontics, Im Neuenheimer Feld 400, and 1 German Cancer Research Center, Division of Differentiation and Carcinogenesis, Im Neuenheimer Feld 280,
D-69120 Heidelberg, Germany
Summary. In stratified epithelia, integrins play a funda- mental role in mediating basal cell attachment to a vari- ety of extracellular matrix molecules. To assess whether keratinocyte-specific integrins are expressed in a similar way as in the normal situation also under in vivo related conditions, we processed oral mucosa equivalents consist- ing of keratinocytes and fibroblasts from non-cornified gingiva. In this model histomorphology, the expression of differentiation-specific keratins and keratinocyte-type in- tegrins exhibited similarity to the tissue of origin. The stages of tissue normalization were assessed on frozen sections by indirect immunofluorescence. The initial activ- ated stage (1 week) was characterized by (i) incomplete epithelial organization and a weak presence of the supra- basal mucosa type keratin K4, (ii) diffuse expression of the integrin chains fll and a 6 and (iii) abundance of the wound healing-associated integrin a v throughout the whole epithelium. After 2 weeks, the increase in epithe- lial organization was characterized by (i) the presence of a basal and suprabasal cell compartment, (ii) extension of K4 in the suprabasal compartment, (iii) extended expres- sion of the keratinocyte integrins fll and a 6 and (iv) con- centration of a v integrin underneath basal cells. Further normalization of tissue architecture was indicated by (i) a slight increase in K4 extension, (ii) appearance of kerati- nocyte integrins fll and a 6 in basal and parabasal cells and (iii) interruption of the band-like a v integrin immu- nolocalization at the subepithelial site. The findings in the in vitro model system indicate that these oral mucosa equivalents exhibit similarities to the in vivo situation of non-cornified gingiva, thus rendering them a suitable model for the assessment of stages during epithelial re- construction or in vivo relevant studies on material ef- fects.
Correspondence to: R Tomakidi
Key words: Organotypic cocultures - Oral mucosa - Cell matrix interactions - Integrins
Introduction
Integrins are heterodimeric transmembrane molecules consisting of an a- and fl- subunit, which mediate cell-cell and cell-matrix interactions. Hence they are essential for cell adhesion, proliferation and migration (Hynes et al. 1992), and are basically involved in the maintenance of tissue integrity. These functions become evident in nor- mal as well as pathological processes such as deve- lopment, wound healing, inflammation and tumor progression (for review see Thomas et al. 1997). Further- more, integrins are also receptors for intracellular signal- ling which regulate cell function (Clark and Brugge 1995). Those integrins, which are mainly expressed in stratified squamous epithelia like epidermis or oral muco- sa, are members of the fl! family, as well as a @4, and a v integrins (Watt et al. 1994). While the integrin chains av and a 5 in epidermis are only expressed during keratino- cyte growth or migration (av, Marchisio et al. 1991), wound healing (Cavani et al. 1993) or hyperproliferation (Gianelli et al. 1994), respectively (a 5), both chains are expressed in oral mucosa, exhibiting a variable distribu- tion (Jones et al. 1993). In the oral cavity the fll subunit generally exhibits a pericellular distribution whereas a 6, which is localized in the hemidesmosome together with f14, is concentrated at the basal aspect of the basal cells. Unlike skin, site-specific variations from this general si- tuation, including extensive suprabasal staining, are not unusual and described for the floor of the mouth and the lateral border of the tongue, possibly linked to an in-
Ann Anat (1999) 181:127-132 © Urban & Fischer Verlag
0940-9602/99/181/1-127512.00/0

creased turnover rate in oral epithelium (see Jones et al. 1993). Another prerequisite for tissue integrity is cell dif- ferentiation. In squamous epithelia, differentiation is not only linked to morphological cell changes, e.g. flattening of cells in the top layers, but also to the synthesis of bio- chemical markers which, in terms of non-cornified epithe- lia, are the keratins K4 and K13. In the oral cavity, these indicators of early differentiation are mainly expressed in the suprabasal cell compartment of non-keratinizing epithelia, such as non-cornified gingiva, the alveolar mu- cosa, the buccal mucosa, the floor of the mouth and the ventral part of the tongue (Mackenzie et al. 1991; Fegha- li-Assaly et al. 1994; Gao et al. 1996; Sch6n et al. 1996).
To investigate a possible correlation of the keratinocyte integrins of oral mucosa with histodifferentiation under in vitro conditions, we analyzed organotypic cocultures from non-cornified gingiva tissue at different growth states. The oral mucosa equivalents reached a stage exhibiting parallels to the normal situation, thus rendering them candidates for studying (i) processes during epithelial re- construction or (if) material effects under in vivo relevant conditions.
Material and methods
Cell Culture and Organotypic Coculture
Primary human keratinocytes and fibroblasts from non-cornified gingiva were isolated from tissue obtained after third molar ex- traction as described previously (Tomakidi et al. 1997). In brief, epithelial tissue fragments and keratinocytes from dispase treat- ment were seeded and grown in KGM medium with bovine pi- tuitary extract. Connective tissue fibroblasts were obtained from explants of the same donor tissue using DMEM medium contain- ing 10% fetal calf serum (FCS). For the generation of organoty- pic cocultures both keratinoeytes and fibroblasts were used at the first passage. The production of fibroblast-containing col- lagen lattices is published in detail elsewhere (Tomakidi et al. 1998). Briefly, 5 x 10 s keratinocytes were seeded per gel and or- ganotypic cocultures were cultured for 7, 14 and 21 days in FAD medium with 5% FCS (a schematic overview of the organotypic coculture system is given in Fig. 1 c). After one day, the collagen gels were lifted to the air-liquid interface and covered with a slight medium overlay to allow epithelial growth similar to oral cavity conditions.
Indirect Immunofluorescence
After specimens of oral mucosa equivalents were embedded and frozen in liquid nitrogen, vapor serial cryostat sections (8 gin) were fixed in acetone (5 rain; -20°C). Incubation of the primary antibodies overnight at 4°C in a humid chamber was followed by washing (3 x ) in phosphate-buffered solution (PBS) and incuba- tion of secondary fluorochrome conjugated antibodies (dilution 1:50-1:100) at room temperature for 45 minutes. After addi- tional PBS washing, samples were embedded in a mounting medium (Vectashield, Camon, Wiesbaden, Germany) and photo-
graphed with an inverted microscope (Zeiss Axiovert 35, Ober- kochen, Germany). Monoclonal antibodies from mouse against keratin K4 were purchased from Progen (Heidelberg, Germany), and those against integrin chains fll and av (Klein et al. 1990) were a generous gift from Eberhard Klein and Eva Br6cker (Wtirzburg, Germany). Rat monoclonal antibody against the in- tegrin chain a6 (Sonnenberg et al. 1991) was kindly provided by Arnoud Sonnenberg (Amsterdam, The Netherlands).
Results
Cytomorphology of Primary Gingival Cells and Progres- sion of Histodifferentiation in Organotypic Cocultures
Frequently, growth of keratinocytes from tissue fragments or single cells was observed 7-10 days after plating. Both keratinocytes as well as connective tissue fibroblasts reached confluency after 2-3 weeks. While confluent pri- mary keratinocytes exhibited the typical cobble stone mor- phology (Fig. 1 a), primary fibroblasts were of a spindle shape after reaching confluency (Fig. l b). Separated growth of the isolated cell types under physiological con- ditions, i.e. separated by an interposed collagen type I cell culture matrix, led to the formation of a multilayered but less organized epithelium (not shown). The poor mor- phological organization was reflected by the immunoloca- lization of the early differentiation marker of non- cornified epithelia, keratin K4, which was only detectable in the uppermost cell layers (Fig. 1 d). After 2 weeks, epithelial morphology revealed a much better organiza- tion substantiated by a distinct basal cell compartment harbouring cuboidal cells and a suprabasal cell compart- ment consisting of flattened cells (Fig. I e 1). This gain of morphological differentiation was concomitantly asso- ciated with an extension of K4 immunolocalization within the suprabasal cell compartment (Fig. i e 2). A further progression of histodifferentiation in these oral mucosa equivalents was documented by a slight increase in the extension of K4 expression, already detectable in some of the basal cells (Fig. 1 f).
Normalization of Keratinocyte Integrins
Based on the integrin function as key cell surface mole- cules in cell adhesion and maintenance of epithelial archi- tecture, we analyzed the expression patterns of integrin chains a 6, fll and a v as representatives for stable (a 6, component of the hemidesmosome, see Sonnenberg et al. 1991) as well as focal contacts (ill, Carter et al. 1990) and hyperproliferative tissue conditions such as wounding (a v, see Thomas et al. 1997) during epithelial develop- ment in vitro. While there was variable staining of the in- tegrin subunit a 6 at day 7, visible at the basal pole of basal cells but also pericellularly (Fig. 2 a), the fll chain, appearing pericellularly, was mostly detectable in the ba-
128

sal layer (Fig. 2 b). In this early stage the av chain per- sisted throughout the whole epithelium also showing a pericellular localization (Fig. 2 c). After 2 weeks the integ- rin chains a6 and //1 showed a remarkable increase in abundance now decorating basal and parabasal cells peri- cellularly, this being much more pronounced for //1 (Figs. 2d, e). Furthermore, a6 immunolocalization ap- peared close to the regular pattern due to the band-like subepithelial staining, and//1 also gradually appeared to be slightly more concentrated in the basal cell layer (see Figs. 2 d, e). In accordance with this approach to the nor- mal tissue situation, av also became gradiently localized
in the basal as well as suprabasal cell compartment and partially showed a polarized, band-like appearance at the subbasal pole of basal cells (Fig. 2f). Finally, after 3 weeks, the presence of a6 mainly at the basal pole of ba- sal keratinocytes (Fig. 2g) and ill, with a pericellular staining of basal and focally parabasal as well as supraba- sal cells (Fig. 2h), demonstrated a further normalization of these keratinocyte-type integrins. The av chain was now present in all epithelial cell layers showing an in- crease in the discontinuity of the polarized band-like im- munolocalization at the epithelium-matrix border (Fig. 2 i).
Fig. 1. Cytomorphology and Progressive Epithelial Histodifferentiation. Phase contrast microscopy revealed characteristic cell morphology in confluent primary cultures of keratinocytes exhibiting cobble stone morphology (a) and fibroblasts showing spindle shape (b). The concept of co-culture of these primary cells in the organotypic system is schematically exemplified in (e), demonstrating K4 protein expression by IIF. On day 7, K4 expression is limited to the sur- face epithelial layers (d), extending within the suprabasal compartment after 14 days (e2) and showing a slight progression of exten- sion to some basal cells (small arrowheads) at day 21 (f). Concomitant with suprabasal K4 expression on day 14, histoarchitecture is characterized by a distinct basal and suprabasal cell compartment (el). Scale Bars 90 tim (a, b) and 55 gm (d, el, e2 and f).
129
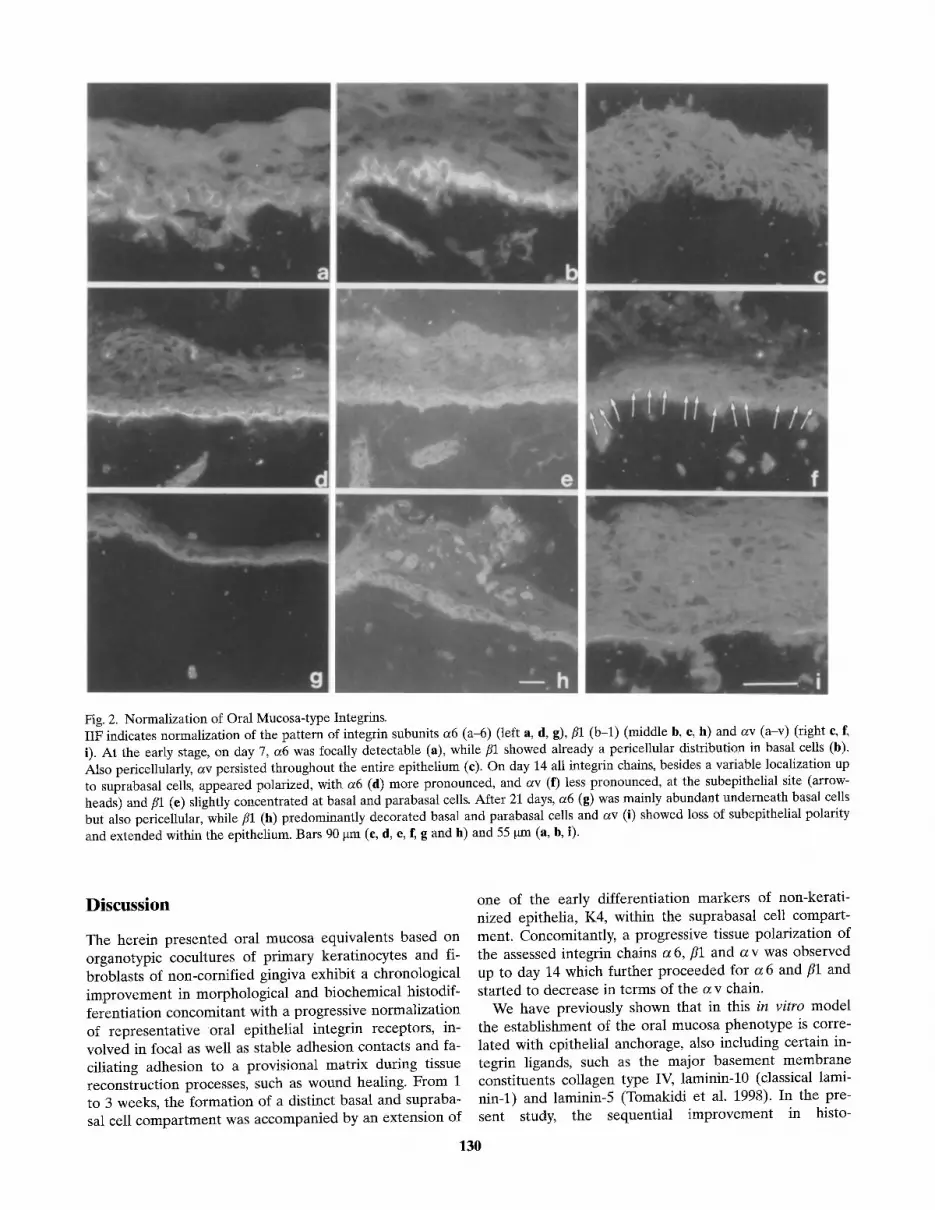
Fig. 2. Normalization of Oral Mucosa-type Integrins. IIF indicates normalization of the pattern of integrin subunits a6 (a-6) (left a, d, g), fll (b-l) (middle b, e, h) and av (a-v) (right e, f~ i). At the early stage, on day 7, a6 was focally detectable (a), while fll showed already a pericellular distribution in basal cells (b). Also pericellularly, av persisted throughout the entire epithelium (c). On day 14 all integrin chains, besides a variable localization up to suprabasal cells, appeared polarized, with a6 (d) more pronounced, and av (f) less pronounced, at the subepithelial site (arrow- heads) and/~1 (e) slightly concentrated at basal and parabasal cells. After 21 days, a6 (g) was mainly abundant underneath basal cells but also pericellular, while fll (h) predominantly decorated basal and parabasal cells and av (i) showed loss of subepithelial polarity and extended within the epithelium. Bars 90 gm (e, d, e, f, g and h) and 55 gm (a, b, i).
Discussion
The herein presented oral mucosa equivalents based on organotypic cocultures of pr imary kerat inocytes and fi- broblasts of non-cornif ied gingiva exhibit a chronological improvemen t in morphological and biochemical histodif- ferentiat ion concomitant with a progressive normalizat ion of representat ive oral epithelial integrin receptors, in- volved in focal as well as stable adhesion contacts and fa- ciliating adhesion to a provisional matr ix during tissue reconstruct ion processes, such as wound healing. From 1 to 3 weeks, the format ion of a distinct basal and supraba- sal cell compar tmen t was accompanied by an extension of
one of the early differentiation markers of non-kerat i- nized epithelia, K4, within the suprabasal cell compar t - ment . Concomitantly, a progressive tissue polarizat ion of the assessed integrin chains a 6,/ /1 and a v was observed up to day 14 which fur ther p roceeded for a 6 and fll and started to decrease in terms of the a v chain.
We have previously shown that in this in vitro model the establ ishment of the oral mucosa phenotype is corre- lated with epithelial anchorage, also including certain in- tegrin ligands, such as the major basement m e m b r a n e constituents collagen type IV, laminin-10 (classical lami- nin-1) and laminin-5 (Tomakidi et al. 1998). In the pre- sent study, the sequential improvement in histo-
130

differentiation and the normalization in the major oral epithelial integrin receptors seems to be correlated with a time-dependent modulation of such integrins which char- acterize strongly activated tissue conditions. The persis- tence of av integrin throughout the whole epithelium after 1 week, together with a single focal presence of a6 and a limited K4 expression at this time point suggests an activated immature tissue state. This is emphasized by the already clearly pericellular abundance of the fll chain and an also pronounced a5 protein expression which already shows a slight focus to basal cells (not shown). Since aSfll and the a v chain associated with r5 are described as the major wound integrins in the epidermis and the oral cav- ity epithelia (Cavani et al. 1993; Thomas et al. 1997), as also in our system, fll might not only form complexes with a3, which is also shown to be weakly expressed in the system at this early timepoint (see Tomakidi et al. 1998), but also with a5 and av. During the culture period the improvement in histodifferentiation generally corre- lated with a polarization in the immunolocalization of in- tegrins a6 and fll subunits and a tissue distribution which is also described for normal oral epithelia of distinct re- gions (see Thomas et al. 1997). The discrepancy between the epidermis and the oral situation is not only limited to a6f14 and a3fll expression which is, as regards its topogra- phy, less restrictive compared to epidermis but also in- cludes the presence of av and a5 which is likely to be more frequently observed in the oral epithelia. This is as- sumed to be linked to the higher rate of cell turnover in the oral cavity (see Jones et al. 1993; Thomas et al. 1997). Although our data revealed the presence of a5 and av, they demonstrate a time-dependent modulation in tissue abundance, possibly correlated to progressive tissue differ- entiation. Since av persists at the early stage it becomes predominantly focused, besides a localization in the su- prabasal compartment, underneath basal cell keratino- cytes, similar to a6. This indicates a more balanced tissue state after 2 weeks, favorizing such contacts which me- diate focal and, what is more important, stable basal cell adhesion. This turn to a more differentiated phenotype is supported by the progressively regular expression of the f14- and a3- chain and the regular deposition of the re- spective integrin ligands (see Tomakidi et al. 1998). This development is in accordance with findings on a6f14 ex- pression and its binding to laminin which is associated with t h e initial steps in keratinocyte maturation (Ten- nenbaum et al. 1996). Whil~ a6 and fll become continu- ously normalized by slightly progressive polarization, the av chain may Start to become depolarized as indicated by the decrease in Subepithelial localization and the pre- sence of panepithelial staining. This av depolarization, which is even more pronounced for a5 (not shown), may suggest a persistence! of the protein which is still detected by the antibody but a downregulation in the synthesis of integrin receptors reflecting an activated tissue state. This may be due to the concomitant increase in histodifferen- tiation, as demonstrated by K4 extension, by culture time. Taken together, the observed stages in this in vitro model
of non-keratinized oral mucosa provide evidence that the expression and localization of cell-matrix interaction re- ceptors is correlated with histodifferentiation. The pro- gressive polarization of integrin chains mediating focal and stable contacts and the vice versa depolarization of integrin subunits of provisorial matrix contacts may lead to the sequential predominance of differentiation. This modulation of integrin expression makes this model a sui- table system to assess events occuring during epithelial reconstruction.
Acknowledgements. We would like to thank Martina Kegel for stylistic improvements and the photographic department of the German Cancer Research Center for excellent collaboration. This work was supported by a grant (to P.T.) from the Deutsche Forschungsgemeinschaft (DFG, Bonn, Germany).
References
Cavani A, Zambruno G, Marcono A (1993) Distinctive integrin expression in the newly forming epidermis during wound heal- ing in humans. J Invest Dermatol 101:600-604
Carter WG, Kaur P, Gil SG, Gahr PJ, Wayner EA (1990) Distinct functions for integrins a3fll in focal adhesions and a6fl4/bul- lous pemphigoid antigen in a new stable anchoring contact (SAC) of keratinocytes: relation to hemidesmosomes. J Cell Biol 111:3141-3154
Clark E, Brugge JS (1995) Integrins and signal transduction path- ways: the road taken. Science 268:233-239
Feghali-Assaly M, Sawaf MH, Serres G, Forest N, Ouhayoun JP (1994) Cytokeratin profile of the junctional epithelium in par- tially erupted teeth. J Periodontal Res 29:185-195
Gao Z, Mackenzie IC (1996) Influence of retinoic acid on the ex- pression of cytokeratins, vimentin and ICAM-1 in human gin- gival epithelia in vitro. J Periodontal Res 31:81-89
Gianelli G, Savoia P, Schiraldi M (1994) Psoriatic lesions in pa- tients with chronic liver disease are distinct from psoriasis vul- garis lesions as judged on the basis of integrin adhesion receptors. Hepatology 20:56-455
Hynes RO (1991) Integrins: versatility, modulation and signalling in cell adhesion. Cell 69:11-25
Jones J, Sugiyama M, Watt FM, Speight PM (1993) Integrin ex- pression in normal hyperplastic, dysplastic and malignant oral epithelium. J Pathol 169:235-243
Klein EC, Steinmayer T, Mattes JM, Kaufmann D, Weber L (1990) Integrins of normal human epidermis: differential ex- pression, synthesis and molecular structure. Br J Dermatol 123:171-178
Mackenzie IC, Rittman G, Gao Z, Leigh I, Lane B (1991) Pat- terns of cytokeratin expression in human gingival epithelia. J Periodontal Res 26:468-478
MarchisioPC, Bondanza S, Cremona O, ConceddaR, De Lu- ca M (1991) Polarized expression of integrin receptors (a6p4, a2fll, a3fll and avfl5) and their relationship with the cytoske- leton and basement membrane matrix in cultured human ker- atinocytes. J Cell Biol 2:761-773
SchOn M, Rheinwald JG (1996) A limited role for retinoic acid and retinoic acid receptors RARa and RARfl in regulating keratin 19 expression and keratinization in oral and epidermal keratinocytes. J Invest Dermatol 107:428-438
131

Sonnenberg A, Calafat J, Janssen H, Daams H, Van der Raaij- Helmer LMH, Falcioni R, Kennel SJ, Aplin JD, Baker J, Loizi- dou M, Garrod D (1991) Integrin a6f14 complex is located in hemidesmosomes, suggesting a major role in epidermal cell- basement membrane adhesion. J Cell Biol 113:907-917
Tennenbaum T, Li L, Belanger AJ, De Luca LM, Yuspa SH (1996) Selective changes in laminin adhesion and a6f14 in- tegrin regulation are associated with the initial steps in kera- tinocyte maturation. Cell Growth Differ 7:615-628
Thomas GJ, Jones J, Speight PM (1997) Integrins and oral cancer. Oral Oncology 33:381-388
Tomakidi P, Breitkreutz D, Fusenig NE, ZOller J, Kohl A, Kom- posch G (1998) Establishment of oral mucosa phenotype in vi- tro in correlation to epithelial anchorage. Cell Tissue Res 292: 355-366
Watt FM, Hertle MD (1994) Keratinocyte integrins. In: Leigh IR, Lane EB, Watt FM (Eds) The Keratinocyte Handbook. Cam- bridge University Press, Cambridge, pp 153-164
132





























