Embed Size (px)
Citation preview

Elsevier Editorial System(tm) for Antiviral Research Manuscript Draft Manuscript Number: Title: NOVEL HBSAG MARKERS TIGHTLY CORRELATE WITH OCCULT HBV INFECTION AND STRONGLY AFFECT HBSAG DETECTION Article Type: Research Paper Section/Category: Host - Virus Interactions Keywords: HBV, Occult infection, HBsAg structure, HBsAg detection Corresponding Author: Dr. C.F. Perno, Corresponding Author's Institution: Universita' degli Studi di Roma First Author: Valentina Svicher Order of Authors: Valentina Svicher; Valeria Cento; Martina Bernassola; Maria Neumann-Fraune; Formijn Van Hemert; Mengjie Chen; Romina Salpini; Chang Liu; Roberta Longo; Michela Visca; Sara Romano; Valeria Micheli; Ada Bertoli; Caterina Gori; Francesca Ceccherini-Silberstein; Cesare Sarrecchia; Massimo Andreoni; Mario Angelico; Antonella Ursitti; Alberto Spanò; Jing Maria Zhang; Jens Verheyen; Giuseppina Cappiello; C.F. Perno Manuscript Region of Origin: ITALY Abstract: Occult HBV infection (OBI) is a threat for the safety of blood-supply, and has been associated with the onset of HBV-related hepatocellular carcinoma and lymphomagenesis. Nevertheless, genetic markers in HBsAg (particularly in D-genotype, the most common in Europe) significantly associated with OBI in vivo are missing. Thus, the goal of this study is to define: i) prevalence and clinical profile of OBI among blood-donors; ii) HBsAg-mutations associated with OBI; iii) their impact on HBsAg-detection. OBI was searched among 422,278 blood-donors screened by Nucleic-Acid-Testing. Following Taormina-OBI-definition, 26 (0.006%) OBI-patients were identified. Despite viremia<50IU/ml, HBsAg-sequences were obtained for 25/26 patients (24/25 Genotype-D). OBI-associated mutations were identified by comparing OBI-HBsAg with that of 82 chronically-infected (genotype-D) patients as control. Twenty HBsAg-mutations significantly correlated for the first time with OBI. By structural analysis, they localized in the major HBV B-cell-epitope, and in HBsAg-capsid interaction region. 14/24 OBI-patients (58.8%) carried in median 3 such mutations (IQR:2.0-6.0) against 0 in chronically-infected patients. By co-variation analysis, correlations were observed for R122P+S167L (phi=0.68,P=0.01), T116N+S143L (phi=0.53,P=0.03), and Y100S+S143L (phi=0.67,P<0.001). Mutants (obtained by site-directed mutagenesis) carrying T116N, T116N+S143L, R122P, R122P+Q101R, or R122P+S167L strongly decreased HBsAg-reactivity (54.9±22.6S/CO, 31.2±12.0S/CO, 6.1±2.4S/CO, 3.0±1.0S/CO and 3.9±1.3S/CO, respectively) compared to wild-type (306.8±64.1S/CO). Even more, Y100S and Y100S+S143L supernatants show no detectable-HBsAg (experiments in quadruplicate). In conclusions, unique HBsAg-mutations in genotype-D, different than those described in genotypes B/C (rarely found in western countries), tightly correlate with OBI, and strongly affect HBsAg-

detection. By altering HBV-antigenicity and/or viral-particle maturation, they may affect full-reliability of universal diagnostic-assays for HBsAg-detection. Suggested Reviewers: Vincent Calvez Department of Virology, Hospital Pitiè-Salpetriere, Paris, France [email protected] Charles Boucher Department of Virology, Erasmus University Rotterdam, the Netherlands [email protected] Hans Zaaijer Department of Clinical Virology, University of Amsterdam [email protected] Daniel Douek Department of Immunology, Vaccine Research Center, Bethesda [email protected]

Prof. Carlo-Federico Perno
Dr. Valentina Svicher
University of Rome Tor Vergata
e-mail: [email protected]
Dr Erik De Clercq
Editor, Antiviral Research
August 18th, 2011
Dear Erik,
On behalf of our co-authors, we submit the enclosed manuscript entitled “Novel HBsAg
markers tightly correlate with occult HBV infection and strongly affect HBsAg detection"
by V Svicher, V Cento, M Bernassola, M Neumann-Fraune, F Van Hemert, M Chen, R
Salpini, C Liu, R Longo, M Visca, S Romano, V Micheli, G Gubertini, A Bertoli, N
Marino, F Mazzotta, C Gori, V Micheli, C Sarrecchia, M Andreoni, M Angelico, A Ursitti,
A Spanò, F Ceccherini-Silberstein, JM Zhang, J Verheyen, G Cappiello, CF Perno for
potential publication in Antiviral Research.
The manuscript has not been published in this or a substantially similar form (in print or
electronically, including on a web site), nor accepted for publication elsewhere, nor it is
under consideration by another publication.
The philosophy of this study is based on a strong interdisciplinary approach joining
together a vertical network of researchers with particular and diverse skills, including
virologists, molecular biologists, biochemists, clinicians, and mathematicians. Therefore,
all people listed as authors actively participated in this study and each gave substantial
contribution at different levels in the conception, design and performance of the work.
Occult HBV infection (characterized by the persistence of HBV-DNA in the serum or liver
tissu in absence of HBsAg) is so far of clinical and pathological importance due to i) its
association with the progression of hepatitis B towards cirrhosis and hepatocellular
carcinoma despite the apparently low HBV-replication rate, ii) HBV-related
lymphomagenesis, iii) its potential risk of transmission in cases of transfusion of blood
products, and of orthotopic liver transplantation.
Several host and viral factors have been hypothesized for the OBI pathogenesis.
Nevertheless, conclusive studies on HBV genetic markers (in particular in the HBsAg)
responsible for Occult HBV infection in vivo are lacking.
In this light, this study is aimed at:
1. Defining the prevalence of occult HBV infection in a large cohort (N=422,278) of
blood donors tested by Nucleic Acid Testing procedure between 2004 and 2008 in
an important reference center for blood transfusion in Central Italy.
2. Providing a systematic and comprehensive characterization of HBsAg genetic
markers underlying occult D-genotype infection in-vivo by using an integrated
clinical, structural and mathematical approach in one of the largest cohort of
patients with OBI so far analyzed for this purpose. Twenty unique HBsAg-
UNIVERSITA' DEGLI STUDI DI ROMA "TOR VERGATA"
DIPARTIMENTO DI MEDICINA SPERIMENTALE E SCIENZE BIOCHIMICHE
Cover Letter

mutations in HBV D genotype were found for the first time significantly correlated
with OBI in-vivo. This is the first study showing such correlation of OBI with the
HBV genotype most commonly found in western countries. Of note, these HBsAg
genetic markers are different from those recently identified in HBV B and C
genotype in far East Asia (Yuan et al., 2010), thus supporting the concept that the
natural variability of HBV genotypes makes difficult the definition of universal
diagnostic markers of HBV.
3. Investigating the impact of such markers on the detection of HBsAg in the
supernatants of cell culture experiments, by means of a clinically-validated test
with a known sensitivity >99%. These in vitro experiments showed that these OBI-
related HBsAg mutations strongly decrease (up to abrogate) the amount of detected
HBsAg in the supernatants, thus potentially altering the full reliability of
diagnostic-assays for HBsAg-detection in blood-samples.
Overall, this information can be used for planning and implementation of screening of this
disease in transfusion services, and to further improve the rate of true-positive results by
standard assay for HBsAg detection in diagnostic practice
These data have been presented in part at the 46th
Annual Meeting of the European
Association for the Study of the Liver (EASL), held in Berlin, Germany, March 30-April 3,
2011, where the abstract was scored among the TOP 10%
For all these reasons we propose our paper to the attention of Antiviral Research.
Among scientists renowned for their expertise in the field, may we suggest the following
names:
Dr Vincent Calvez
Department of Virology,
Hospital Pitiè-Salpetriere, Paris, France
83 Boulevard de l’Ospital, F-75013 Paris
Tel: +33-1-42177416
E-mail: [email protected]
Dr Charles Boucher
Department of Virology,
Erasmus University Rotterdam, the Netherlands
Tel: +30 250 6526
E-mail: [email protected]
Dr Hans L Zaaijer
Department of Clinical Virology,
Academic Medical Center (AMC), Clinical Virology (L1-104),
University of Amsterdam, P.O.Box 22660, 1100DD Amsterdam, The Netherlands
Tel: +31-20-5669111
E-mail: [email protected]
Dr Daniel Douek,
Vaccine Research Center, Bethesda
Bldg 40
Room 3504
MSC 3022
Bethesda, MD 20892-3005

Tel: +1 301-594-8484
E-mail: [email protected]
Looking forward to hearing from you.
Yours sincerely,
Prof. Carlo Federico Perno, MD, PhD
and Dr. Valentina Svicher, PhD
Department of Experimental Medicine
University of Rome Tor Vergata
Via Montpellier 1, 00133 Rome, Italy
Tel: +39-06-72596566, 06-72596551
Fax: +39-06-72596039
E-mail: [email protected],

Highlights
HBsAg genetic markers correlated with occult HBV D genotype infection in vivo were
studied
Twenty HBsAg-mutations significantly correlated for the first time with OBI were found.
They localized in the major HBV B-cell-epitope, and in HBsAg-capsid interaction region.
They strongly affect, up to abrogate, HBsAg detection,
Thus, they can affect full-reliability of diagnostic-assays for HBsAg-detection.
*Highlights

1
NOVEL HBSAG MARKERS TIGHTLY CORRELATE WITH OCCULT HBV 1
INFECTION AND STRONGLY AFFECT HBSAG DETECTION 2
Running Title: HBsAg Markers in Occult HBV infection 3
Valentina Svicher1#
, Valeria Cento1#
, Martina Bernassola2, Maria Neumann-Fraune
3, 4
Formijn Van Hemert4, Mengjie Chen
5, Romina Salpini
1, Chang Liu
5, Roberta Longo
2, 5
Michela Visca2, Sara Romano
2, Valeria Micheli
6, Ada Bertoli
7, Caterina Gori
8, 6
Francesca Ceccherini-Silberstein1, Cesare Sarrecchia
7, Massimo Andreoni
7, Mario 7
Angelico7, Antonella Ursitti
2, Alberto Spanò
2, Jing Maria Zhang
5, Jens Verheyen
3, 8
Giuseppina Cappiello2, Carlo Federico Perno
1,7* 9
1 University of Rome “Tor Vergata” Rome, Italy; 2 “S. Pertini” Hospital, Rome, Italy; 10
3 University of Cologne, Cologne, Germany; 4 Center for Infection and Immunity 11
Amsterdam (CINIMA), Academic Medical Center, University of Amsterdam, 12
Amsterdam, the Netherlands; 5 Yale University, USA; 6 “L. Sacco” Hospital, Milan, 13
Italy; 7 University Hospital of Rome “Tor Vergata” Rome, Italy; 8 “L. Spallanzani” 14
Hospital, Rome, Italy 15
# equally contributed to the manuscript 16
Corresponds to: Carlo-Federico Perno, PhD, MD; University of Rome Tor Vergata; 17
Via Montpellier 1, 00133 Rome, Italy. Tel: +39 06-72596551; Fax: +39 06-72596039. 18
E-mail: [email protected]. 19
20
These data have been presented in part at the 46th Annual Meeting of the European 21
Association for the Study of the Liver (EASL), held in Berlin, Germany, March 30-April 22
3, 2011 where the abstract was scored among the 10%, and at the 2nd
International HIV 23
and Viral Hepatitis Drug Resistance Workshop held in Los Cabos, Mexico, June 8-11, 24
2011. 25
26
27
*ManuscriptClick here to view linked References

2
ABSTRACT 28
Occult HBV infection (OBI) is a threat for the safety of blood-supply, and has been 29
associated with the onset of HBV-related hepatocellular carcinoma and 30
lymphomagenesis. Nevertheless, genetic markers in HBsAg (particularly in D-31
genotype, the most common in Europe) significantly associated with OBI in vivo are 32
missing. Thus, the goal of this study is to define: i) prevalence and clinical profile of 33
OBI among blood-donors; ii) HBsAg-mutations associated with OBI; iii) their impact 34
on HBsAg-detection. OBI was searched among 422,278 blood-donors screened by 35
Nucleic-Acid-Testing. Following Taormina-OBI-definition, 26 (0.006%) OBI-patients 36
were identified. Despite viremia<50IU/ml, HBsAg-sequences were obtained for 25/26 37
patients (24/25 Genotype-D). OBI-associated mutations were identified by comparing 38
OBI-HBsAg with that of 82 chronically-infected (genotype-D) patients as control. 39
Twenty HBsAg-mutations significantly correlated for the first time with OBI. By 40
structural analysis, they localized in the major HBV B-cell-epitope, and in HBsAg-41
capsid interaction region. 14/24 OBI-patients (58.8%) carried in median 3 such 42
mutations (IQR:2.0-6.0) against 0 in chronically-infected patients. By co-variation 43
analysis, correlations were observed for R122P+S167L (phi=0.68,P=0.01), 44
T116N+S143L (phi=0.53,P=0.03), and Y100S+S143L (phi=0.67,P<0.001). 45
Mutants (obtained by site-directed mutagenesis) carrying T116N, T116N+S143L, 46
R122P, R122P+Q101R, or R122P+S167L strongly decreased HBsAg-reactivity 47
(54.9±22.6S/CO, 31.2±12.0S/CO, 6.1±2.4S/CO, 3.0±1.0S/CO and 3.9±1.3S/CO, 48
respectively) compared to wild-type (306.8±64.1S/CO). Even more, Y100S and 49
Y100S+S143L supernatants show no detectable-HBsAg (experiments in quadruplicate). 50

3
In conclusions, unique HBsAg-mutations in genotype-D, different than those described 51
in genotypes B/C (rarely found in western countries), tightly correlate with OBI, and 52
strongly affect HBsAg-detection. By altering HBV-antigenicity and/or viral-particle 53
maturation, they may affect full-reliability of universal diagnostic-assays for HBsAg-54
detection. 55
Keywords: HBV, Occult infection, HBsAg structure, HBsAg detection 56
1. INTRODUCTION 57
Chronic overt hepatitis B is characterized by detectable hepatitis B surface 58
antigen (HBsAg) in the serum, while occult HBV infection (OBI) is defined by the 59
persistence of HBV-DNA in the liver tissue (and in some cases also in the serum) in 60
absence of HBsAg (Raimondo et al., 2010). OBI is one of the most challenging topics 61
in the field of viral hepatitis. Indeed, OBI can be transmitted in cases of transfusion of 62
blood products, as well as during orthotopic liver transplantation, and it has been 63
associated with the progression of chronic liver damage towards cirrhosis and 64
hepatocellular carcinoma despite the apparently low HBV-replication rate (Raimondo et 65
al., 2008). Indeed, OBI has been shown to maintain the pro-oncogenic properties typical 66
of the chronic infection, and has been identified in up to 70.4% of HBsAg-negative 67
patients with hepatocellular carcinoma (Fang et al., 2009). In addition, it has been 68
shown that lymphotropism is a major feature of OBI, and that a relationship may exist 69
between OBI and lymphomagenesis (Engels et al., 2010). Finally, patients with OBI are 70
at potential risk of HBV reactivation during immune-suppressive therapy in the future, 71
especially in regimens containing rituximab (Hoofnagle et al., 2009). This supports the 72
need to set up effort for a correct detection of OBI. 73

4
OBI is also a potential threat for the safety of blood supply. The prevalence of OBI in 74
blood donors has been reported in some European Countries such as Poland, Germany, 75
and France (Broyer et al., 2006; Katsoulidou et al., 2009; Levast et al., 2010), while in 76
others, such Italy, recent data are limited (Velati et al., 2008; Dettori et al., 2009). This 77
information is critical in order to set up optimized transfusion services. 78
Several host and viral factors have been hypothesized for the OBI pathogenesis 79
(Blum et al., 1991; Chaudhuri et al., 2004; Zaaijer et al., 2008; Vivekanandan et al., 2008; 80
El Chaar et al., 2010). Nevertheless, conclusive studies on HBV genetic markers (in 81
particular in the HBsAg) responsible for OBI in vivo are lacking. This can be due to a 82
number of factors: 1. The number and type of patients studied: the majority of reports 83
included a limited number of patients infected with different HBV genotypes, thus 84
making difficult to extrapolate the effect of mutations from the different genetic 85
background; 2. The technology used for viral genome sequencing: not widely available 86
for low level viremia; 3. The analysis of different fragments of HBV genome and the 87
methodologies used to characterize HBV genetic variability: the majority of studies 88
reported single mutations correlated with occult HBV infection without providing a 89
more complex level of information regarding the cluster of mutations underlying occult 90
HBV infection in-vivo. 91
Thus, and despite its clinical relevance, the genetic characteristics in HBsAg 92
from patients with occult HBV is still poorly known. This is particularly true for HBV, 93
D-genotype infection, that is the most prevalent in Mediterranean Countries including 94
Southern Europe. 95
In this light, using extensive sequence analyses, unique computational methods, 96
and phenotypic characterization, this study is aimed i) at defining the extent of occult 97

5
HBV infection in asymptomatic blood donors in a Country with low HBV endemicity, 98
ii) at identifying HBsAg-genetic markers specifying occult HBV infection in-vivo in 99
one of the largest homogenous cohort of OBI-patients so far analyzed, iii) as proof of 100
concept, at investigating the impact of such markers on the detection of HBsAg in the 101
supernatants of cell culture experiments. 102
Overall, this information can provide new insights regarding the pathogenesis of OBI, 103
and can be used to further improve the rate of true-positive results by standard assay for 104
HBsAg detection in diagnostic practice. 105
106
2. METHODS 107
2.1 Patients. The present study was carried out in two cohorts involving a total of 108
422,278 regular and occasional blood donors, screened for HCV, HIV-1/2 and HBV 109
infection by Nucleic Acid Testing (NAT) at a reference center for transfusion security in 110
Lazio region. The first cohort involved blood donors who donated blood between 111
December 2004 and April 2007 These samples were tested using polymerase chain 112
reaction (PCR)-based HIV-1/HCV/HBV test (COBAS Ampliscreen, Roche 113
Diagnostics, Branchburg, NJ; lower limit of detection [LOD]: 5 IU/mL, 95% 114
confidence interval: 3-13 IU/mL) in pools of 20 samples, generated with Tecan Genesis 115
pipetting machine (Tecan Schweiz AG, Männedorf, Switzerland). HBV-DNA reactive 116
pools were resolved by minipools of 5 samples testing. Donations of minipool, found to 117
be NAT-reactive, were tested individually to identify the positive unit. 118
The second cohort of donors, who donated blood between March 2007 and December 119
2008, were screened in minipools of 6 samples using the COBAS Taqscreen MPX 120
(Roche Molecular Systems, Branchburg, NJ; LOD: 3.1 IU/mL, 95% confidence 121

6
interval: 1.7-7.8 IU/mL) test on the COBAS s201 system for the direct detection of 122
HIV1-2/HCV/HBV (Roche Instrument Center, Rotkreuz, Switzerland). When a 123
minipool was reported as reactive, the 6 donation were individually tested with the 124
COBAS Taqscreen MPX test on the COBAS s201 system as a part of the resolution 125
process to identify the reactive and non-reactive donor sample. 126
Any individual specimens found to be reactive was tested using HIV1/HCV/HBV 127
COBAS Ampliscreen (Roche Diagnostics, Branchburg, NJ) for viral discrimination. 128
HBV-DNA was quantified by quantitative PCR COBAS Taqman High Pure System 129
HBV (Roche Molecular Systems, Branchburg, NJ; LOD: 6 IU/mL) between December 130
2004 and December 2005 and by COBAS Ampliprep/COBAS Taqman HBV test 131
(Roche Molecular Systems, Branchburg, NJ; LOD: 12 IU/mL) between November 2005 132
and December 2008. 133
Confirmed HBV-DNA positive samples were retested for serological markers of HBV 134
infection (HBsAg, anti-HBc, anti-HBc-IgM, HBeAg, anti-HBe, anti-HBs) using 135
specific tests on the Roche Elecsys 2010 immunoassay analyzer (Roche Diagnostics, 136
Branchburg, NJ). 137
2.2 HBsAg-DNA sequencing. HBsAg-DNA sequencing was performed following the 138
methodology reported in (Weinberger et al., 1999). Specific primers and amplification 139
protocols are reported in detail in the supplementary text 1. Translated HBsAg-140
sequences were generated and aligned using Bioedit 7.0 software and CLUSTALW1.8. 141
All sequences are in process to be submitted to GenBank. A phylogenetic analysis was 142
performed to exclude possible contaminations. 143
2.3 Genetic variability analysis. 144

7
Shannon entropy (Sn = -Σi(pi lnpi)/lnN) was calculated, where pi was the frequency of 145
each distinct nucleotide sequence and N was the total number of sequences analyzed. 146
Statistical significance was assessed trough Wilcoxon rank test. Evolutionary 147
divergence of HBsAg-sequences was estimated as the extent of nucleotide substitutions 148
per site determined by the HKY model of MEGAv5.0 (Tamura et al., 2011), applying a 149
gamma distribution with shape parameter = 0.8. The nested codon models 7 and 8 of 150
PAML 3.15 (Yang et al., 2007) were used to calculate dN/dS ratio values. The nwk user 151
tree was generated in MEGA5 (amino acid alignment, JTT replacement model), 152
CodonFreq F3x4 was chosen and gaps were treated as “missing data” (cleandata=0). 153
Likelihood Ratio Tests were performed to verify a better fit of the data with model 8 154
(extra size class (dN/dS >1) with respect to the null-hypothesis (model 7, 10 size 155
classes, dN/dS =<1). 156
2.4 Statistical analysis. 157
(i) Mutation prevalence. To assess the association of HBsAg-mutations with occult 158
infection, we calculated the prevalence of all amino acids (including the wild-type 159
amino acids) found at each HBsAg-position in the 24 sequences from occult infected 160
patients, and 82 sequences from chronically infected patients. We then performed chi-161
squared tests of independence with a Benjamini-Hochberg correction (false discovery 162
rate, FDR=0.05) to identify statistically significant differences in frequency between the 163
two groups of patients. 164
(ii) Mutation covariation. Binomial-correlation coefficient (phi) was calculated to 165
assess covariation among mutations associated with occult infection (Svicher et al., 166
2009). Statistically significant pairs of mutations were identified by Fisher’s exact test, 167
corrected for multiple-testing by Benjamini-Hochberg method (FDR=0.05). 168

8
(iii) Cluster analysis. In order to identify and summarize higher-order interactions of 169
mutations, we transformed the pairwise phi correlation coefficients into dissimilarity 170
values. A dendrogram was then computed by hierarchical clustering, and its stability 171
was assessed from 100 bootstrap replicates. The details of this explorative data analysis 172
procedure have been described elsewhere (Svicher et al., 2009). 173
2.5 Bayesian Variable Partition Model and Recursive Model Selection. Bayesian 174
Variable Partition Model and Recursive Model Selection were carried out according to 175
the methodology described in (Zhang et al., 2010). A detailed description of these 176
models is reported in supplementary text 2. 177
2.6 HBsAg three-dimensional structure prediction. An in silico generated tertiary 178
structure of HBV S-protein was available including the amino acid residues comprising 179
the interface composition of a surface-capsid complex (Roy et al., 2010). The amino 180
acid replacements described to be associated with occult HBV infection (this paper) 181
were manually introduced into the sequence. To generate a 3D model of surface protein 182
carrying these mutations, the file was submitted to I_TASSER for homology modeling 183
with the original "D-consensus" structure as a custom modeling template. Residues are 184
colored according to membrane topology, A-determinant size and surface-capsid 185
interface composition as described in (Roy et al., 2010). Hydrophobicity profiles were 186
constructed using values normalized between 0 and 1 (Black et al., 1991). A sliding 187
window of 5 residues with a step size of 1 was applied. 188
2.7 Site-specific mutagenesis and HBsAg detection assay. The expression vector 189
pCHsAg (genotype D) under the control of cytomegalovirus immediate early promoter-190
enhancer was used as wild type control and as backbone vector for the expression of 191
HBsAg. Single or double HBsAg-mutations were introduced by site directed 192

9
mutagenesis in the expression vector and confirmed by sequence reactions. Wild type 193
and HBsAg mutants were tested by transfection of human HuH7 cells with pCHsAg 194
plasmids using FuGene6 (Roche diagnostics GmbH, Mannheim, Germany). The 195
supernatant was collected after 72 hours and tested using the ABBOTT Architect 196
HBsAg (Qual) assay. HBsAg was tested in cell-supernatant obtained from four 197
independent experiments for each mutant and wild type. These results were used to 198
calculate the mean value and standard deviation for each HBsAg mutant. 199
200
3. RESULTS 201
3.1 Prevalence of Occult infection and Description of the Study Population 202
The frequency of HBV-DNA positive/HBsAg negative donations was assessed in a 203
large cohort of blood donors recruited in an important reference center for transfusion 204
security in Central Italy. 205
Among 422,278 blood donors tested, 269 (0.06%) were positive for serum HBV-DNA 206
by NAT procedure. After HBsAg testing, 243/269 (90.3%) patients were HBV-DNA-207
positive and HBsAg-positive (thus with diagnosis of overt chronic HBV hepatitis), 208
while the other 26 patients (0.006%) were HBV-DNA-positive and HBsAg-negative. 209
No significant difference in the prevalence of HBV-DNA-positive and HBsAg-negative 210
patients was observed in the two cohorts of blood donors (0.005% in the first cohort 211
screened between 2004-2007 versus 0.008% in the second cohort screened between 212
2007-2008). Following the position paper issued by the recent meeting of experts in 213
Taormina (Raimondo et al., 2008), OBI is associated with undetectable or low level 214
HBV-DNA (<200IU/ml) in the serum of HBsAg negative individuals. For 22/26 215
patients (84.6%), serum HBV-DNA quantification was <12IU/ml, while for the 216

10
remaining 4 patients, viremia ranged from 30 to 50IU/ml (Table 1). ALT values were in 217
the normal range. Therefore, these 26 HBV-DNA-positive and HBsAg-negative patients 218
were considered as occult HBV-infected patients. 219
Demographic and serological characteristics of the 26 blood donors with OBI are 220
summarized in Table 1. The majority were Italian (25/26, 96.2%), and male (22/26, 221
84.6%). All blood donors with OBI were anti-HBc positive, and anti-HBc IgM 222
negative, thus indicating that they were not in the window phase of acute hepatitis B 223
infection. Eleven out of 26 (42.3%) patients also showed low level of protective anti-224
HBs (>10mUI/ml) (Median [IQR]= 28.0 [20.0-54.0]). 225
Despite HBV-DNA<50IU/ml, HBsAg-DNA sequences were obtained for 25/26 (96%). 226
Direct HBsAg sequencing showed that genotype-D was dominant (24/25, 96.0%), 227
while genotype-A was present only in 1 patient. 228
3.2 HBsAg genetic markers underlying OBI in-vivo. 229
HBsAg genetic characteristics underlying OBI in-vivo were investigated. In order to 230
make data more clear and readable, the analysis was focused on 24 patients with occult 231
HBV D-genotype infection (thus excluding the only genotype-A patient). A group of 82 232
randomly-selected, chronically HBV D-genotype infected patients was used as control. 233
All chronically infected patients were HBsAg-positive, anti-HBc positive with a median 234
(IQR) serum HBV-DNA of 5.9 (3.3-6.2) logIU/ml (Table S1). All of them were naïve 235
to any anti-HBV drug-treatment. 236
A higher degree of genetic variability was found in HBsAg-sequences from OBI 237
compared to patients with chronic HBV infection. Indeed, both the median Shannon 238
Entropy, the median evolutionary rate, and the dN/dS ratio were remarkably higher in 239
patients with OBI than in patients with chronic HBV infection (median Shannon 240

11
Entropy (IQR): 0.29 (0.00-0.54) versus 0.07 (0.00-0.18), P=0.000068; median 241
evolutionary divergence (IQR): 0.06 (0.03-0.08) versus 0.02 (0.02-0.04), P=0.0003; 242
dN/dS: 0.84 (0.74-2.36) versus 0.32 (0.15-0.59), P=4.863∙10-29
) (Fig.1). 243
This result prompted us to investigate the existence of HBsAg markers specifying OBI 244
in-vivo. 245
Overall, 20 HBsAg-mutations were found for the first time significantly correlated with 246
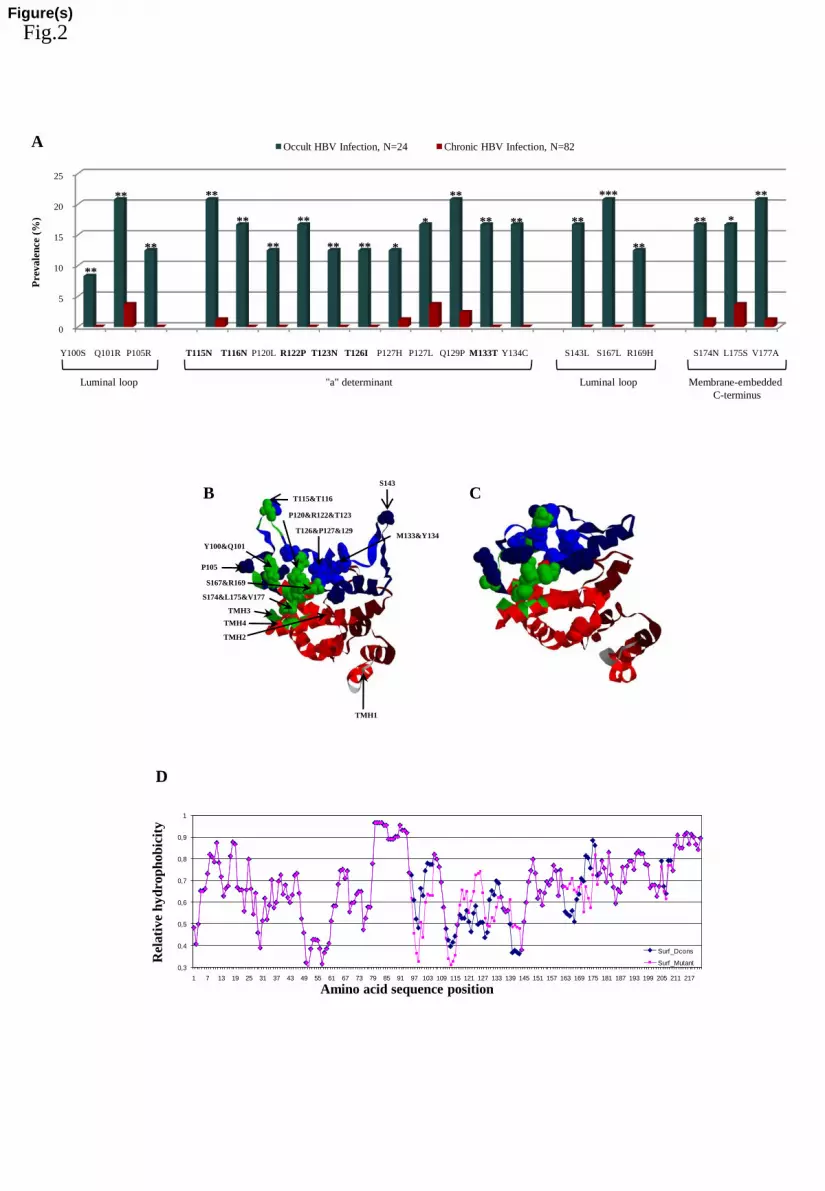
occult HBV D-genotype infection in-vivo (Fig.2A) (P-values from 10-2
to 10-4
after 247
correction for multiple comparison). Fourteen out of 24 (58.8%) patients with OBI (vs. 248
only 1/82 chronic infected patients) carried >1 of these mutations (median number 249
[IQR]: 3.0 [2.0-6.0]). Conversely, in our cohort of 82 chronically-infected patients, such 250
mutations were completely absent (0/82 for Y100S, P105R, T116N, P120L, R122P, 251
T126I, Y134C, S143L, S167L, R169H) or nearly absent (1/82 for T115N, P127H, 252
S174N, V177A, 2/82 for Q129P, and 3/82 for Q101R, P127L, L175S, one mutation per 253
patient). Their prevalence ranged from 8.3 to 20.8% in OBI patients, compared to 0-254
3.7% in the 82 chronically-infected patients. 255
A Bayesian variable partition model (Zhang et al., 2010) was then applied to further 256
confirm whether HBsAg-positions are associated with OBI. All HBsAg-mutations 257
identified in the above-mentioned analysis were fully confirmed by this model. Indeed, 258
all mutations showed a posterior probability ranging from 0.75 to 1, indicating a strong 259
association with occult infection (data not shown). 260
3.3 Localization of HBsAg residues in the predicted HBsAg three-dimensional 261
structure. 262
To gain insight into effects of OBI-associated amino acid replacements on HBsAg, we 263
introduced these replacements in the sequence and built a 3D-model of mutant HBsAg 264

12
for comparison with D-consensus HBsAg. Fig.2B shows that OBI-associated residues 265
are mainly located at positions involved in antibody recognition and capsid interaction. 266
In the 3D-structure of mutant HBsAg (Fig.2C), the luminal loop including the A-267
determinant region displays a "collapse" towards the hydrophobic center of the 268
molecule. The architecture at the capsid-interaction regions is less modified. A similar 269
view is presented by the hydrophobicity profiles (Fig.2D) showing a replacement of 270
hydrophilic into more hydrophobic residues particularly at the A-determinant region 271
(110-140). 272
3.4 Specific associations among HBsAg markers. 273
A further step of this study was to investigate the complex interaction patterns of 274
HBsAg-mutations associated with OBI in-vivo. This is the first study addressing this 275
point. 276
Associations among HBsAg-mutations. To identify significant patterns of pairwise 277
correlations between mutations at the above mentioned positions, we calculated the 278
binomial correlation coefficient (phi) and its statistical significance for each pair of 279
mutations. A positive and statistically significant correlation between mutations at two 280
specific positions (0<phi<1, P< 0.05) indicates that these two positions co-evolve; thus 281
it indicates that the co-occurrence of mutations is not due by chance. A strong and 282
significant positive association of S143L with either T116N (phi=0.68) or Y100S 283
(phi=0.66), and of R122P with either T127S (phi=0.84), Q101R (phi=0.56), or S167L 284
(phi=0.56) was found. A significant correlation was also observed between L175S and 285
V177A, both localized in the HBsAg hydrophobic C-terminus (phi=0.56). 286
Clusters of correlated mutations. Because pairwise analysis suggested that some novel 287
mutations are associated with specific evolutionary pathways, an average linkage 288

13
hierarchical agglomerative cluster analysis was performed to investigate this hypothesis 289
in more detail. 290
The topology of the dendrogram (Fig.S1) highlighted the existence of two distinct 291
clusters of HBsAg-mutations in OBI. The first cluster included the mutations T116N 292
and S143L (bootstrap value=0.51), that were linked to Y100S (bootstrap=0.54). The 293
second pattern included the mutations S167L, and Q101R that were linked to T127S 294
and R122P (bootstrap value=0.50). 295
3.5 Phenotypic Analysis of HBsAg-mutation’s Patterns in Occult Infection 296
Cluster analysis highlighted the existence of specific cluster underlying OBI in-vivo. As 297
proof of concept, the ability of these mutation clusters to hamper HBsAg detection was 298
investigated in cell-culture using the ABBOTT Architect HBsAg (Qual) assay. 299
The detection rates of expressed HBsAg mutants significantly differed according to the 300
pattern of mutations analyzed (Fig. 3). The supernatants of mutants Y100S and 301
Y100S+S143L showed no detectable HBsAg (<1 sample/cutoff-ratio [S/CO]). A strong 302
decrease in the detection was also observed for HBsAg-mutants carrying T116N, 303
T116N+S143L, R122P, R122P+Q101R, or R122P+S167L. These mutations were 304
associated with remarkable decrease of HBsAg-reactivity from 306.8±64.1S/CO for 305
wild-type to 54.9±22.6S/CO, 31.2±12.0S/CO, 6.1±2.4S/CO, 3.0±1.0S/CO and 306
3.9±1.3S/CO, respectively (p<0.01 for all tests). By contrast, HBsAg-results for Q101R, 307
S167L and S143L were comparable to wild-type. 308
309
4. DISCUSSION 310
This study documented a prevalence of 0.057% of HBsAg-positive HBV-DNA 311
positive patients, and of 0.006% of OBI in a large cohort of blood donors tested by 312

14
NAT procedure, resolved by minipools of 5/6 samples, between 2004 and 2008. This is 313
in the range of prevalence of OBI estimated in Italy in previous studies (0.013%-314
0.006%) (Dettori et al., 2009; Velati et al., 2008) and in other European Countries, such 315
as Greece (0.0002%) and Poland (0.084%) (Brojer et al., 2006; Katsoulidou et al, 316
2009). 317
Several possible mechanisms have been hypothesized for the pathogenesis of 318
OBI and the condition is probably multi-factorial (Raimondo et al., 2008). Due to the 319
very small group of OBIs so far analyzed, HBsAg genetic markers in D-genotype 320
significantly associated with OBI in vivo are still missing. 321
By an integrated clinical, structural and mathematical approach, 20 mutations 322
(18 not previously correlated, and two previously associated with HBV immune-escape, 323
T123N and M133T) (Hou et al., 2001; Torresi et al., 2002) were found to be 324
significantly associated with occult HBV D-genotype infection in vivo, mainly located 325
in the “a-determinant” region. This is the first study showing such correlation of OBI 326
with an HBV genotype most commonly found in western countries. 327
The majority of these OBI-associated HBsAg genetic-markers were completely 328
absent in patients with chronic hepatitis. In addition, they occurred with a median 329
number [IQR] of 3.0 [2.0-6.0] in patients with OBI. The route towards the occult state 330
may be therefore characterized by an accumulation of variants, followed by a step-by-331
step loss of antigen-antibody reactivity (that is the basis of HBsAg detection test in 332
clinical practice). 333
In-vitro experiments showed that specific patterns of HBsAg-mutations (Y100S; 334
Y100S+S143L; T116N+S143L; R122P; R122P+Q101R and R122P+167L) strongly 335

15
decreased the amount of detected HBsAg in the supernatants, suggesting an impairment 336
in HBsAg-detection and/or changes in the antigenicity. 337
The impact of such mutation-patterns on HBsAg detection has not yet been 338
evaluated before. An important HBsAg function is the anchorage in the endoplasmatic-339
reticulum followed by assembly and secretion of mature virions (also HBsAg-mediated 340
processes). Notably, transmembrane helices and the hydrophobic C-terminus are largely 341
saved from extensive mutation in OBI. Conversely, we observed a linkage between 342
amino acid replacement near or at positions involved in capsid interaction (100, 101, 343
116, 122, 169, 174, 175, 177), suggesting a potential ability to impair viral particle 344
maturation and/or to hamper viral infectivity (van Hemert et al., 2008). Consistent with 345
this hypothesis, a recent in-vitro study showed the ability of mutations at position 169 346
(such as the R169H) to drastically hamper virion secretion, and to have a dominant 347
negative effect when it is co-expressed with wild-type envelope proteins (Ito et al., 348
2010). 349
For mutations T115N , T116N and T123N, the correlation with OBI can be 350
explained by the fact that these mutations introduce a novel N-liked glycosylation site, 351
that may impair virion secretion, along with reduction of HBsAg immunogenicity; 352
indeed T123N has been recently shown to strongly reduce virion assembly by leading to 353
the production of a new N-glycosylated form of HBsAg (Wu et al., 2010). 354
Residues associated with occult HBV D-genotype infection at positions 105, 355
120, 122, and 123 have been recently shown to be critical conformation-dependent 356
features for HBV entry into the target cells (Salisse et al., 2009). Mutations at these 357
residues have been found to drastically hamper (or even abrogate) in-vitro viral 358
infectivity (Salisse et al., 2009), thus supporting their association with OBI in-vivo. 359

16
Notably, while mutations in the “a-determinant” often resulted in non-360
synonymous mutations in the reverse transcriptase (RT) protein, the majority of the 361
amino acid substitutions involving capsid-interacting residues were silent in the RT. 362
From the evolutionary point of view, this observation points towards an accumulation of 363
mutations due to a relaxation of selective constraints and may indicate a contribution of 364
these regions to the evolutionary transition from a chronic into an OBI, observation also 365
supported by previous studies (van Hemert et al., 2008) and by the higher values of 366
evolutionary divergence and dN/dS found in occult infection respecting to chronic. 367
Functionally, a diminished interaction between core and surface proteins due to the 368
mutations introduced at these regions may support this process, rendering the transition 369
irreversible. 370
A recent study showed an association between OBI and immune-escape (El 371
Chaar et al., 2010), postulating that patients with OBI carrying both anti-HBc and anti-372
HBs, are under strong humoral immune-pressure. This selecting-pressure may lead to 373
the selection of viral strains with highly mutated HBsAg, in particular in the HBsAg 374
regions 125-131 and 158-169, crucial for HBsAg recognition and to reduce the 375
exposure of HBsAg immunogenic surface (Wu et al., 2010). These mutations may be 376
responsible for HBV evasion from the immune-system, and, at the same time, may 377
hamper viral replication, justifying the very low serum HBV-DNA levels classically 378
observed in OBI. Due to the quasispecies nature of HBV, the reactivation of viral 379
replication under immune-suppression may be mainly sustained by the re-emergence of 380
the wild-type strain. 381
OBI has been associated with the onset of HBV-related hepatocarcinogenesis 382
and lymphomagenesis (Fang et al., 2009; Engels et al., 2010). Further studies are thus 383

17
necessary to verify the ability of these mutations to modulate HBV oncogenic potential. 384
In addition, recent studies have highlighted new insights regarding HBV pathogenesis 385
(Neumann et al., 2010; Tian et al., 2010). In particular, a study shows that expression 386
and secretion of HBsAg is a novel mode of inducing secretion of Cyclophilin A 387
(CypA), that has been involved in the pathogenesis of HBV infection (Tian et al., 2010).
388
Another study identifies a novel antiviral mechanism of antibodies to HBsAg involving 389
prolonged blocking of the HBV and HBsAg subviral particles release from infected 390
cells (Neumann et al., 2010). The impact of HBsAg mutations correlated with OBI on 391
these two novel mechanisms underlying HBV pathogenesis should be matter of further 392
investigation. 393
A new Abbott ARCHITECT® assay for the detection of HBsAg has been 394
recently proposed to specifically enhance the detection of some HBsAg mutants at 395
position 122 and 123 (Lou et al., 2011). The impact of the other genetic markers we 396
identified on the performance of this new assay has not yet been defined. 397
It should be noted that such study has been conducted in OBIs all carrying D-398
genotype HBV. Therefore, the conclusions driven by these results may not be 399
applicable to other HBV-genotypes. Consistent with this hypothesis, a recent study, led 400
in China, identified specific HBsAg mutations (in the regions from amino acids 117 to 401
121 and amino acids 144 to 147) potentially associated with OBI-infection in HBV B- 402
and C-genotype. These mutations are completely different from those we identified in 403
HBV D-genotype (none among those described in genotype D are reported in genotype 404
B and C, and viceversa) (Yuan et al., 2010), thus supporting the concept that immune 405
escape and OBI can be driven by HBV-associated mutations different for each HBV 406

18
genotype. As a consequence, the natural variability of HBV genotypes may make 407
difficult the definition of universal diagnostic markers of HBV. 408
In conclusions, unique HBsAg-mutations in genotype-D, different than those 409
described in genotypes B/C (rarely found in Western-Countries), tightly correlate with 410
OBI, and strongly affect HBsAg-detection. These mutations can alter HBV-antigenicity 411
and/or viral-particle maturation, and can affect full-reliability of diagnostic-assays for 412
HBsAg-detection. Thus, the incorporation of these mutants in reagent development can 413
further improve the rate of true-positive results. 414
415
416
417
418
419
420
421
422
423
424
425
426

19
REFERENCES 427
1. Black, S.D., Mould, D.R., 1991. Development of hydrophobicity parameters to 428
analyze proteins which bear post- or cotranslational modifications. Anal. Biochem. 429
193, 72-82. 430
2. Blum, H.E., Galun, E., Liang, T.J., Wands, J.R., 1991. Naturally occurring missense 431
mutation in the polymerase gene terminating hepatitis B virus replication. J. Virol. 432
65, 1836-1842. 433
3. Brojer, E., Grabarczyk, P., Liszewski, G., Mikulska, M., Allain, J.P., Letowska, M., 434
2006. Characterization of HBV DNA+/HBsAg- blood donors in Poland identified by 435
triplex NAT. Hepatology 44, 1666-1674. 436
4. Chaudhuri, V., Tayal, R., Nayak, B., Acharya, S.K., Panda, S.K., 2004. Occult 437
hepatitis B virus infection in chronic liver disease: full-length genome and analysis 438
of mutant surface promoter. Gastroenterology 127, 1356-1371. 439
5. Dettori, S., Candido, A., Kondili, L.A., Chionne, P., Taffon, S., Genovese, D., 440
Iudicone, P., Miceli, M., Rapicetta, M., 2009. Identification of low HBV-DNA levels 441
by nucleic acid amplification test (NAT) in blood donors. J. Infect. 59, 128-133. 442
6. El Chaar, Candotti, D., Crowther, R.A., Allain, J.P., 2005. Impact of hepatitis B virus 443
surface protein mutations on the diagnosis of occult hepatitis B virus infection. 444
Hepatology 52, 1600-1610. 445
7. Engels, E.A., Cho, E.R., Jee, S.H., 2010. Hepatitis B virus infection and risk of non-446
Hodgkin lymphoma in South Korea: a cohort study. Lancet Oncol. 11, 827-834. 447

20
8. Fang, Y., Shang, Q.L., Liu, J.Y., Li, D., Xu, W.Z., Teng, X., Zhao, H.W., Fu, L.J., 448
Zhang, F.M., Gu, H.X., 2009. Prevalence of occult hepatitis B virus infection 449
among hepatopathy patients and healthy people in China. J. Infect. 58, 383-388. 450
9. Hoofnagle, J.H., 2009. Reactivation of hepatitis B. Hepatology 49, S156-S165 451
10. Hou, J., Wang, Z., Cheng, J., Lin, Y., Lau, G.K., Sun, J., Zhou, F., Waters, J., 452
Karayiannis, P., Luo, K., 2001. Prevalence of naturally occurring surface gene 453
variants of hepatitis B virus in nonimmunized surface antigen-negative Chinese 454
carriers. Hepatology 34, 1027-1034. 455
11. Ito, K., Qin, Y., Guarnieri, M., Garcia, T., Kwei, K., Mizokami, M., Zhang, J., Li, J., 456
Wands, J.R., Tong, S., 2010. Impairment of hepatitis B virus virion secretion by 457
single-amino-acid substitutions in the small envelope protein and rescue by a novel 458
glycosylation site. J. Virol. 84, 12850-12861. 459
12. Katsoulidou, A., Paraskevis, D., Magiorkinis, E., Moschidis, Z., Haida, C., 460
Hatzitheodorou, E., Varaklioti, A., Karafoulidou, A., Hatzitaki, M., Kavallierou, L., 461
Mouzaki, A., Andrioti, E., Veneti, C., Kaperoni, A., Zervou, E., Politis, C., 462
Hatzakis, A., 2009. Molecular characterization of occult hepatitis B cases in Greek 463
blood donors. J. Med. Virol. 81, 815-825. 464
13. Levast, M., Larrat, S., Thelu, M.A., Nicod, S., Plages, A., Cheveau, A., Zarski, J.P., 465
Seigneurin, J.M., Morand, P., Leroy, V., 2010. Prevalence and impact of occult 466
hepatitis B infection in chronic hepatitis C patients treated with pegylated interferon 467
and ribavirin. J. Med. Virol. 82, 747-754. 468

21
14. Lou, S.C., Pearce, S.K., Lukaszewska, T.X., Taylor, R.E., Williams, G.T., Leary, 469
T.P., 2011. An improved Abbott ARCHITECT((R)) assay for the detection of 470
hepatitis B virus surface antigen (HBsAg). J. Clin. Virol. 51, 59-63. 471
15. Neumann, A.U., Phillips, S., Levine, I., Ijaz, S., Dahari, H., Eren, R., Dagan, S., 472
Naoumov, N.V., 2010. Novel mechanism of antibodies to hepatitis B virus in 473
blocking viral particle release from cells. Hepatology 52, 875-885. 474
16. Raimondo, G., Pollicino, T., Romano, L., Zanetti, A.R., 2010. A 2010 update on 475
occult hepatitis B infection. Pathol. Biol. (Paris) 58, 254-257. 476
17. Raimondo, G., Allain, J.P., Brunetto, M.R., Buendia, M.A., Chen, D.S., Colombo, 477
M., Craxì, A., Donato, F., Ferrari, C., Gaeta, G.B., Gerlich, W.H., Levrero, M., 478
Locarnini, S., Michalak, T., Mondelli, M.U., Pawlotsky, J.M., Pollicino, T., Prati, 479
D., Puoti, M., Samuel, D., Shouval, D., Smedile, A., Squadrito, G., Trépo, C., Villa, 480
E., Will, H., Zanetti, A.R., Zoulim, F., 2008. Statements from the Taormina expert 481
meeting on occult hepatitis B virus infection. J. Hepatol. 49, 652-657. 482
18. Salisse, J., Sureau, C., 2009. A function essential to viral entry underlies the 483
hepatitis B virus "a" determinant. J. Virol. 83, 9321-9328. 484
19. Roy, A., Kucukural, A., Zhang, Y. I-TASSER, 2010. A unified platform for 485
automated protein structure and function prediction. Nat. Protoc. 5, 725-738. 486
20. Svicher, V., Gori, C., Trignetti, M., Visca, M., Micheli, V., Bernassola, M., Salpini, 487
R., Gubertini, G., Longo, R., Niero, F., Ceccherini-Silberstein, F., De Sanctis, 488
G.M., Spanò, A., Cappiello, G., Perno, C.F., 2009. The profile of mutational 489

22
clusters associated with lamivudine resistance can be constrained by HBV 490
genotypes. J. Hepatol. 50, 461-470. 491
21. Tamura, K,, Peterson, D,, Peterson, N., Stecher, G., Nei, M., Kumar, S., 2011. 492
MEGA5 2011: Molecular Evolutionary Genetics Analysis using Maximum 493
Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Molecular 494
Biology and Evolution. 495
22. Tian, X., Zhao, C., Zhu, H., She, W., Zhang, J., Liu, J., Li, L., Zheng, S., Wen, 496
Y.M., Xie, Y., 2010. Hepatitis B virus (HBV) surface antigen interacts with and 497
promotes cyclophilin a secretion: possible link to pathogenesis of HBV infection. J. 498
Virol. 84, 3373-3381. 499
23. Torresi, J., 2002. The virological and clinical significance of mutations in the 500
overlapping envelope and polymerase genes of hepatitis B virus. J. Clin. Virol. 25, 501
97-106. 502
24. van Hemert, F.J., Zaaijer, H.L., Berkhout, B., Lukashov, V.V., 2008. Occult 503
hepatitis B infection: an evolutionary scenario. Virol. J. 5, 146. 1743-1746. 504
25. Velati, C., Romano, L., Fomiatti, L., Baruffi, L., Zanetti, A.R., 2008. Impact of 505
nucleic acid testing for hepatitis B virus, hepatitis C virus, and human 506
immunodeficiency virus on the safety of blood supply in Italy: a 6-year survey. 507
Transfusion 48, 2205-2213. 508
26. Wu, C., Zhang, X., Tian, Y., Song, J., Yang, D., Roggendorf, M., Lu, M., Chen, X., 509
2010. Biological significance of amino acid substitutions in hepatitis B surface 510

23
antigen (HBsAg) for glycosylation, secretion, antigenicity and immunogenicity of 511
HBsAg and hepatitis B virus replication. J. Gen. Virol. 91, 483-492. 512
27. Weinberger, K.M., Zoulek, G., Bauer, T., Bohm, S., Jilg, W., 1999. A novel 513
deletion mutant of hepatitis B virus surface antigen. J. Med. Virol. 58, 105-110. 514
28. Yang, Z., 2007. PAML 4: phylogenetic analysis by maximum likelihood. Mol. Biol. 515
Evol. 24, 1586-1591. 516
29. Yuan, Q., Ou, S.H., Chen, C.R., Ge, S.X., Pei, B., Chen, Q.R., Yan, Q., Lin, Y.C., 517
Ni, H.Y., Huang, C.H., Yeo, A.E., Shih, J.W., Zhang, J., Xia, N.S., 2010. 518
Molecular characteristics of occult hepatitis B virus from blood donors in southeast 519
China. J. Clin. Microbiol. 48, 357-62. 520
30. Zaaijer, H.L., Torres, P., Ontañón, A., Ponte, L.G., Koppelman, M.H., Lelie, P.N., 521
van Hemert, F.J., Boot, H.J., 2008. Multiple surface antigen mutations in five blood 522
donors with occult hepatitis B virus infection. J. Med. Virol. 80, 1344-1349. 523
31. Zhang, J., Hou, T., Wang, W., Liu, J.S., 2010. Detecting and understanding 524
combinatorial mutation patterns responsible for HIV drug resistance. Proc. Natl. 525
Acad. Sci. U. S. A. 107, 1321-1326 526
527
528
529
530
531
532

24
LEGENDS TO FIGURES 533
Figure 1. Nucleotidic and amino acidic variability in HBsAg-gene and protein. The 534
three box-plots represent Shannon Entropy (A), Evolutionary Divergence (B) and the 535
ratio of non-synonymous (dN) and synonymous (dS) substitutions rates (C) among 536
patients with chronic infection and OBI. P-Values were calculated by Mann-Whitney 537
Test. 538
Figure 2. Amino acidic HBsAg-protein mutations associated with OBI. (A) prevalence 539
of HBsAg-mutations associated with OBI. Mutations reported in bold can introduce a 540
new N-linked glycosylation site in HBsAg-protein. P-values were determined by Fisher 541
exact test applying a Benjamini-Hochberg correction for multiple comparison at a false 542
positive rate of 0.05. * p<0.05; ** p<0.01; *** p<0.001. (B) Localization of amino acid 543
residues associated with HBV occult infection in 3D models of "D-consensus" and 544
mutant HBsAg-protein. Functional HBsAg domains are color-coded in the "D-545
consensus" protein: amino acids not assigned (1-7, gray), transmembrane signals (8-22, 546
80-98, red) and membrane-embedded C-terminus (169-226, red), cytosolic loop (23-79, 547
dark-red), luminal loop (99-169, dark-blue), the a-determinant region (110-140, blue) 548
and capsid interaction (96, 100, 103, 104, 113, 116, 117, 118, 121, 122, 169, 173, 174, 549
175, 177, 181, 182, 191, 192, 195, green). OBI-related residues are marked by 550
spacefilling in the "D-consensus" 3D structure. (C) The 3D-modeled structure of mutant 551
HBsAg with similar color coded and spacefilled residues shows to what extent the 552
HBsAg architecture becomes affected by amino acid replacements associated with 553
occult HBV infection. (D) Superposition of normalized hydrophobicity profiles of D-554
consensus (blue) and mutant (red) HBsAg-proteins (window 5 and step-size 1 residues) 555

25
displays these alterations particularly in the A-determinant region (110-140) and also at 556
capsid interacting residues (97-105, 160-177). 557
Figure 3. Impact of HBsAg-mutations associated with OBI on HBsAg reactivity. The 558
graph reports the mean of 4 independent experiments of the HBsAg amount detected in 559
presence of the with wild-type and mutated HBV clones. HBsAg has been quantified by 560
the Abbott Architect HBsAg assay. Specimens <1.0 S/CO are considered not reactive. 561
562
563
564
565
566
567
568
569
570
571
572
573
574
575
576
577
578
579

26
ACKNOWLEDGMENTS 580
We thank Laura Scipioni, and Andrea Biddittu for data-management.This work was 581
financially supported by grants from the Italian National Institute of Health, the 582
Ministry of University and Scientific Research, Current and Finalized Research of the 583
Italian Ministry of Health. The funders had no role in study design, data collection and 584
analysis, decision to publish, or preparation of the manuscript 585
586
587
TRANSPARENCY DECLARATION 588
C.-F. Perno have received funds for attending symposia, speaking, organizing 589
educational activities, grant research support, consultancy and advisory board 590
membership, from Abbott, Boehringer Ingelheim, Bristol Myers Squibb, Gilead, Merck 591
Sharp & Dohme, Janssen Cilag, Pfizer, Tibotec, Roche, and ViiV. F. Ceccherini-592
Silberstein has received funds for attending symposia, speaking and organizing 593
educational activities from Abbott, Merck Sharp & Dohme, Janssen Cilag, and Virco. V 594
Svicher has received funds for attending symposia, speaking and organizing educational 595
activities from VIIV. The other authors declare that no competing interests exist. 596
597
598
599
600
601
602
603
604

27
Table 1. Demographic and serological characteristics of the study population. 605
Characteristic Patients with Occult HBV infection
Patients, N 26
Age (years), Median (IQR) 56.0 (51.1-62.6)
Male, N (%) 22 (84.6)
Italian Nationality, N (%) 25 (96.2)
HBV Genotype
D 24 (92.3)
A 1 (3.8)
Undetermined 1 (3.8)
Type of Blood Donation
First time donors 10 (38.5)
Habitual donors 16 (61.5)
N (%) of patients with serum HBV-DNA
< 12 UI/ml 22 (84.6)
> 12 UI/ml 4 (15.4)b
ALT (IU/ml), Median (IQR) 21.5 (19.0-29.3)
Serological characteristics of patients
HBcAb IgG+, N (%) 26 (100)
HBcAb IgM+, N (%) 0
HBeAg+, N (%) 0
HBeAb+, N (%) 19 (73.1)
HBsAg (S/CO), Median, (IQR) 0.46 (0.38-0.59)
HBsAb+, N (%) 11 (42.3)
HBsAb (mIU/ml), Median, (IQR)a 28.0 (20.0-54.0)
a Serum HBV-DNA was determined by COBAS TaqMan High Pure System and by COBAS AmpliPrep/TaqMan 606
HBV. 607
b Viremia ranges from 30IU/ml to 50IU/ml. 608
IQR, interquartile range. 609
610
611
612

28
613

10
8
6
4
2
0
Patients with chronic HBV
infection (N=82)
Patients with occult HBV
infection (N=24)
dN
/dS
2.0
1.5
1.0
0.5
0.0
Patients with chronic HBV
infection (N=82)
Patients with occult HBV
infection (N=24)
Sh
ann
on
En
trop
y
A
P = 6.855e-05 0.2
0.15
0.1
0.05
0.0
Patients with chronic HBV
infection (N=82)
Patients with occult HBV
infection (N=24)
Aver
age
Evolu
tion
ary D
iver
gen
ce (
Nsu
bs/
Sit
e)
B
P = 0.0003
C
P = 4.863e-29
Fig.1
Figure(s)
0
5
10
15
20
25
Prevale
nce (
%)
Occult HBV Infection, N=24 Chronic HBV Infection, N=82
*
Y100S Q101R P105R T115N T116N P120L R122P T123N T126I P127H P127L Q129P M133T Y134C S143L S167L R169H S174N L175S V177A
*
*
**
**
**
**
**
**
**
** **
**
** ** **
***
**
**
**
Luminal loop "a" determinant Luminal loop Membrane-embedded
C-terminus
A
S143
TMH1
TMH2
TMH4
TMH3
S174&L175&V177
S167&R169
Y100&Q101
P105
T115&T116
P120&R122&T123
T126&P127&129M133&Y134
CB
Rel
ati
ve
hy
dro
ph
ob
icit
y
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1
1 7 13 19 25 31 37 43 49 55 61 67 73 79 85 91 97 103 109 115 121 127 133 139 145 151 157 163 169 175 181 187 193 199 205 211 217
Surf_Dcons
Surf_Mutant
Amino acid sequence position
D
Fig.2Figure(s)

S/CO WT 100S 116N 143L100S,
143L
116N,
143L122P 101R 167L
122P,
167L
101R,
122P
Mean 306.86 0.55 54.91 229.97 0.47 31.20 6.12 169.04 303.42 3.89 3.02
SE 64.10 0.05 22.57 90.30 0.02 12.05 2.44 91.27 74.71 1.33 1.00
0
50
100
150
200
250
300
350
400
1 S/CO
Am
ou
nt
of
HB
sAg
det
ecte
d (
S/C
O)
Fig.3
Figure(s)

e-componentsClick here to download e-components: SI_text for AVR.doc

e-componentsClick here to download e-components: Table S1.docx

e-componentsClick here to download e-components: Figura S1.ppt





























