Embed Size (px)
Citation preview

J. Evol. Biol. 8: 355-367 (1995) 1010-061X/95/030355-13 $ 1.50 +0.20/O 0 1995 Birkhluser Verlag, Base1
Nucleotide Polymorphism at the rp49 region of Drosophila subobscura: Lack of geographic subdivision within chromosomal arrangements in Europe
J. Rozas*, C. Segarra*, C. Zapatat, G. Alvarez7 and M. Aguade*
*Departament de Genetica, Facultat de Biologia, Universitat de Barcelona, Barcelona, Spain; TDepartamento de Biologia Fundamental, Fact&ad de Biologia, Universidad de Santiago de Compostela, Santiago de Compostela, Spain.
Key words: rp49 region; restriction-map variation; Drosophila subobscura; chromo- somal polymorphism.
Abstract
Restriction-site polymorphism at the rp49 gene region has been studied in 234 lines of Drosophila subobscura representing different gene arrangements for the 0 chromosome. The population surveyed (El Pedroso, Spain) was sampled four times in each of two consecutive years. The data indicate that the two chromosomal classes studied, OtSTI and Ot3++ are genetically differentiated in El Pedroso. Comparison of the present results with those previously obtained for two other populations further supports that, for a given chromosomal class, European populations are not genetically differentiated. This lack of differentiation at the rp49 region within OtSTI and within O,, +41 stands in contrast to the clear latitudinal clines found in Europe for these arrangements.
Introduction
Drosophila subobscura, a typically Palearctic species, has been extensively studied at the chromosomal and allozyme levels (Loukas et al., 1979; Krimbas and Loukas, 1980; Prevosti et al., 1983; Zapata and Alvarez, 1987; Krimbas, 1992). Data on nuclear DNA variation in this species are still scarce. Only variation at the region encompassing the rp49 gene, which codes for the ribosomal protein 49, has been analyzed in natural populations (Rozas and Aguade, 1990, 1991a, b, 1993, 1994). In D. subobscura, the rp49 region is located very close to one of the breakpoints of
355

356 Rozas et al.
the O3 inversion, which, together with the overlapping inversion 04, constitutes the 0 3+4 chromosomal arrangement that defines segment I (distal part) of the 0 chromosome. Chromosomal gene arrangements Os, and O,,, show clear latitudi- nal clines in Europe (Krimbas and Loukas, 1980; Krimbas, 1992) and in the recently colonized areas of America (Prevosti et al., 1988). The close proximity of the rp49 gene to the 0, inversion breakpoint makes this gene an appropriate marker for the study of the evolution of segment I of the 0 chromosome.
Nucleotide variation at the rp49 gene region has been studied in two European populations: Barcelona and Ter Ape1 (Rozas and Aguade, 1990). This study indicated that for a given chromosomal class these populations were not genetically differentiated. On the other hand, significant heterogeneity was detected between
OL3 + 4l and OfSTl chromosomal classes, and a population from the Canary Islands (monomorphic for the 0, + 4 chromosomal arrangement) proved to be genetically differentiated from both European populations (Rozas and Aguade, 1991a).
The lack of geographic subdivision within chromosomal classes detected in these European populations stands in contrast to the latitudinal clines for the frequencies
of 03+4 and Os, arrangements. However, due to the location of these populations in the latitudinal cline for inversion polymorphism the sample size was rather small
for 03+, in Ter Apel, and for Os, in Barcelona (Rozas and Aguadt, 1990). This lack of differentiation needed therefore to be confirmed. In order to corroborate these results and to obtain additional information on nucleotide variation in Europe, we studied a large sample for each chromosomal gene arrangement from a third population (El Pedroso, Spain). Previous data on allozyme and chromosomal polymorphism showed some seasonal changes in this population (Fontdevila et al., 1983), which motivated our seasonal sampling strategy,
Materials and methods
Fly samples
A total of 379 lines of Drosophila subobscura were used (Tab. 1). Lines from Barcelona (Spain), Ter Ape1 (Holland) and Tenerife (Canary Islands, Spain) have been previously analyzed (Rozas and Aguade, 1990, 1991a).
The El Pedroso population (Spain) was sampled in four different periods of the year (spring, early summer, late summer, autumn) over two consecutive years ( 1989 and 1990). We used the Va/Ba balancer stock (Sperlich et al., 1977) to obtain either isochromosomal lines or heterozygotes of the wild chromosome over one balancer- stock chromosome, as described in Rozas and Aguade (1990). Two hundred and thirty four lines were established and the gene arrangement for the 0 chromosome was determined for each line. These lines are not a random sample of the population. In fact, we studied 94 OtsT1 chromosomes, while the frequency of this arrangement in the population is 0.13 (Tab. 1); however, for each chromosomal class, the lines were chosen at random.
rp49 restriction-site variation
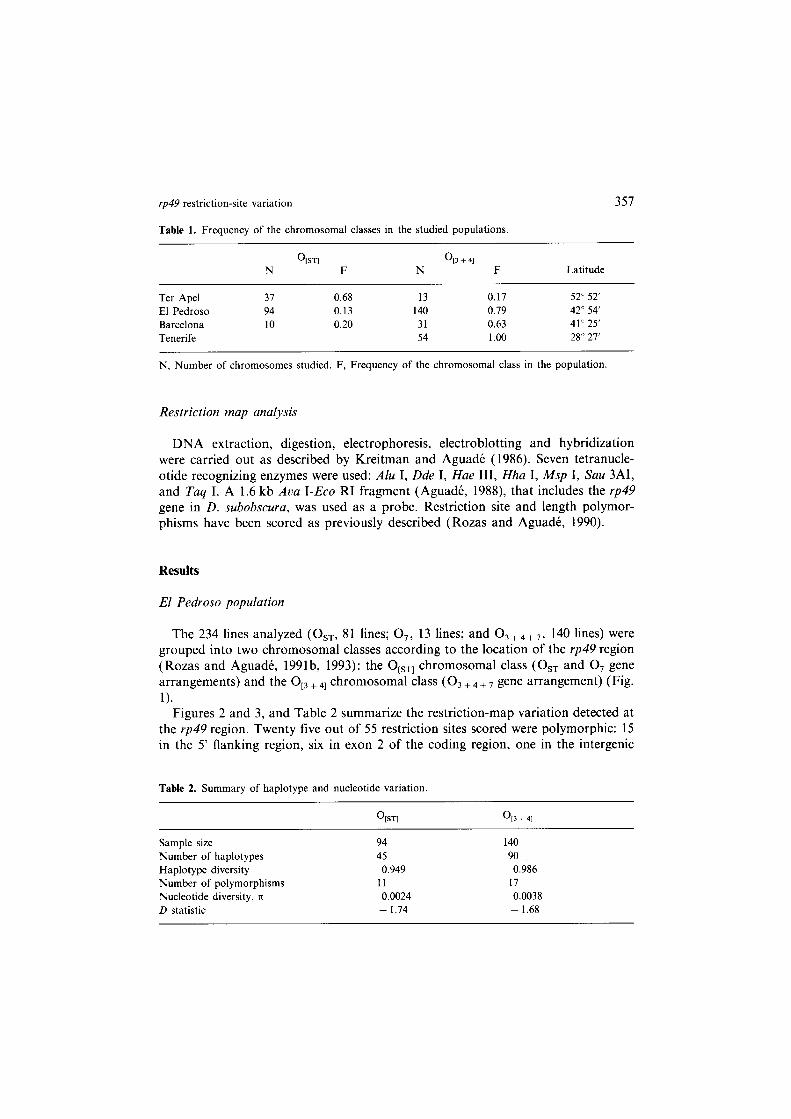
Table 1. Frequency of the chromosomal classes in the studied populations.
357
%I 0 13 + 41 N F N F Latitude
Ter Ape1 37 0.68 13 0.17 52” 52’ El Pedroso 94 0.13 140 0.79 42” 54’ Barcelona 10 0.20 31 0.63 41” 25’ Tenerife 54 1.00 28” 27’
N, Number of chromosomes studied. F, Frequency of the chromosomal class in the population.
Restriction map analysis
DNA extraction, digestion, electrophoresis, electroblotting and hybridization were carried out as described by Kreitman and Aguade (1986). Seven tetranucle- otide recognizing enzymes were used: Alu I, Dde I, Hae III, Hha I, Msp I, Sau 3A1, and Taq I. A 1.6 kb Ava I-&o RI fragment (Aguade, 1988), that includes the rp49 gene in D. subobscura, was used as a probe. Restriction site and length polymor- phisms have been scored as previously described (Rozas and Aguade, 1990).
Results
El Pedroso population
The 234 lines analyzed (O,,, 81 lines; 0,) 13 lines; and 0, + 4 + ,, 140 lines) were grouped into two chromosomal classes according to the location of the rp49 region (Rozas and Aguadt, 1991b, 1993): the OtST1 chromosomal class (O,, and 0, gene arrangements) and the O,, + 4l chromosomal class (0, + 4 + 7 gene arrangement) (Fig.
1). Figures 2 and 3, and Table 2 summarize the restriction-map variation detected at
the rp49 region. Twenty five out of 55 restriction sites scored were polymorphic: 15 in the 5’ flanking region, six in exon 2 of the coding region, one in the intergenic
Table 2. Summary of haplotype and nucleotide variation.
Sample size 94 140 Number of haplotypes 45 90 Haplotype diversity 0.949 0.986 Number of polymorphisms 11 17 Nucleotide diversity, n 0.0024 0.0038 D statistic - 1.74 - 1.68

358 Rozas et al.
Fig. 1. Location of the rp49 region in the three chromosomal gene arrangements studied. Within any chromosomal class the rp49 region has the same location, and consequently crossing over should not be prevented in homokaryotypes for segment I.
region between rp49 and 6-sry genes, and three in the coding region of the b-sry gene.
Eight length polymorphisms (insertions-deletions ranging from 1 to 12 bp in length) were also detected, all of them located in noncoding regions (either in the flanking regions or in the intron of the rp49 gene). Four of these length polymor- phisms (I/D 2, I/D 4, I/D 6, I/D 8) show more than two variants. Some of these polymorphisms are located in regions with tandemly repeated motifs of a short number of nucleotides, and they represent variation in the number of repeats
Fig. 2. Summary of restriction map variation in the rp49 region of D. subobscura for the El Pedroso population. Polymorphisms are indicated above the bar: restriction-site and length polymorphisms are
shown by vertical bars and triangles, respectively. Monomorphic restriction sites are indicated below the bar. A, Alu I; D, Dde I; E, Hae III; H, Hha I; M, Msp I; S, Sau 3AI; T, Taq I. Boxes indicate the rp49 and the S-sry (partial) coding regions. Coordinates, in base pairs, correspond to those of Aguadt (1988). Arrows indicate the direction of transcription.

rp49 restriction-site variation
Fig. 3.
359
(Continued ouerleaf )

360 Rozas et al.
Fig. 3. (Continued). Haplotypes (Hap.) identified in El Pedroso. A plus sign ( + ) and a minus sign ( - ) denote the presence and absence of a restriction site or of an insertion/deletion. I/D = insertion/deletion. Length polymorphisms with more than two variants are indicated by numbers according to Rozas and Aguadt (1990), and represent insertions or deletions from I to 12 bp. S, spring; ES, early summer; LS,
late summer; AU, autumn.

rp49 restriction-site variation 361
(Rozas and Aguade, 1993). When both restriction site and length polymorphisms are considered, 123 different haplotypes were identified (Fig. 3).
Genetic differentiation between subsamples was examined by Hudson et al.‘s (1992) method. The statistic used was KST, defined as
f&T = 1 -V&/K,)
where KT is the average number of differences between two sequences of the total population, and KS is a weighted average of the average number of differences between sequences within subsamples. The statistical significance of the observed KST values was determined by Monte Carlo simulations (1000 iterations). Since KT is constant over iterations, the P value (the probability of obtaining the observed or a more extreme value of the statistic) of KST is identical to the P value of KS, and therefore KS is equivalent to KST as a test statistic. For a given chromosomal class and season, no significant heterogeneity in the distribution of haplotypes could in general be detected between consecutive years (only 1 out of the 8 tests performed was significant, P = 0.01 I). These results allowed us to pool the data of both years and to test for putative heterogeneity among seasons. No significant heterogeneity could be detected among seasons (each chromosomal class was tested separately). This lack of heterogeneity allowed the pooling of all data from a given chromoso- ma1 class. In the pooled data significant differentiation was detected between OLsrl
and 013 + 4l (P = O.OOO), despite the existence of some polymorphisms segregating in both chromosomal classes (Fig. 4).
Nucleotide variation was estimated as haplotype and nucleotide diversity (Nei and Tajima, 1981) (Tab. 2). Haplotype and nucleotide diversity were lower among O,sTl chromosomes than among O,, + 4l ones. Departure from a neutral expectation was tested using Tajima’s method (Tajima, 1989). The method compares two estimates of 4Nu, where N is the effective population size and u is the mutation rate per DNA sequence per generation. One of these estimates is based on the average number of pairwise nucleotide differences among the DNA sequences, and the other on the number of segregating sites. Under a strictly neutral model the statistic proposed by
Fig. 4. Frequency spectrum of restriction-site polymorphisms within chromosomal classes. Nine out of I I restriction-site polymorphisms present in O,srl and nine out of 17 in O,, + 4l are present only once in the sample. Polymorphisms segregating in both chromosomal classes (I, Dde I ~ 136; 14, Dde I 716; 25, Hue III 1993) are indicated.

362 Kozas et al.
Tajima, Tajima’s D, should not deviate significantly from zero. Although D estimates were not significant, they were very close to the critical value (Tab. 2).
Genetic variation between populations
Table 1 shows the frequency of the chromosomal classes in the surveyed populations. Like in previous studies in this species (see Krimbas and Loukas, 1980; Prevosti et al., 1988; Krimbas, 1992), the frequency of the OtST1 is highest in the most northern population, while O,, + 4l shows the lowest frequency in that population.
We have estimated the average level of gene flow between populations by Nm. This parameter was estimated from the proportion of nucleotide diversity that is attributable to variation between populations, ysT, assuming that populations are structured in an island model and that they are in a genetic equilibrium between migration and genetic drift (Wright, 1951; Nei, 1982). It is not clear that our populations conform to these assumptions. In addition, the variance of ysT due to the sampling of individuals, populations and nucleotides might be very large. In our case, the variance due to the sampling of nucleotides could be important due to the rather small number of restriction sites analyzed. Nevertheless, the Nm estimates for the three OtST1 and the three O,, +41 mainland subpopulations ( 17.8 and 19.9, respectively) are higher than the Nm estimate (5.8) obtained for the O,, + 41 when the Tenerife population is also included.
We used Hudson et al.‘s (1992) method to detect whether populations are genetically differentiated. Table 3 shows the values of the test statistic Ks for all pairwise comparisons. For the European populations, all pairwise comparisons between OtST1 and O,, +41 were significant; moreover all pairwise comparisons including Tenerife were also significant. For a given chromosomal class, none of the pairwise comparisons between European populations were significant.
Table 3. KS values between populations.
El Pedroso Barcelona Ter Ape1 El Pedroso Barcelona Ter Ape1
%I Barcelona
Ter Ape1
2.213
2.344 2.671
0 I3 + 41 El Pedroso Barcelona Ter Ape1
Tenerife
2.879*** 3.283* 3.217***
2.620*** 3.5W 3.282;‘; 3.449 2.431*** 3.454** 3.128*** 3.419 4.018
2.168*** 2.180*** 2.319*** 3.006*** 2.787*** 2.539***
* P < 0.05; ** P < 0.01; *** P < 0.001
rp49 restriction-site variation
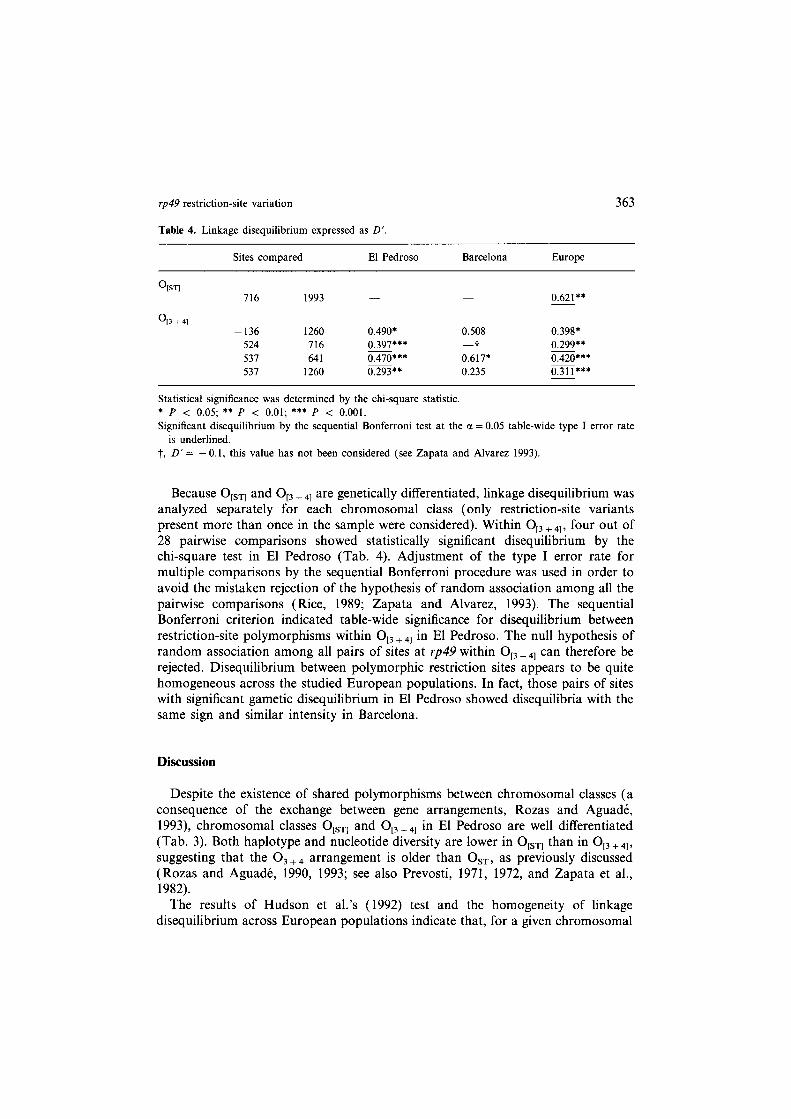
Table 4. Linkage disequilibrium expressed as D’.
363
Sites compared El Pedroso Barcelona Europe
%I 716 1993 0.621**
0,3 + 41 -136 1260 0.490* 0.508 0.398*
524 716 0.397*** -t 0.299** 537 641 0.470*** 0.617* 0.420*** 537 1260 0.293** 0.235 0.311***
Statistical significance was determined by the chi-square statistic.
* P < 0.05; ** P < 0.01; *** P < 0.001. Significant disequilibrium by the sequential Bonferroni test at the a = 0.05 table-wide type I error rate
is underlined. t, D’ = - 0.1, this value has not been considered (see Zapata and Alvarez 1993).
Because OtsTl and O,, + 41 are genetically differentiated, linkage disequilibrium was analyzed separately for each chromosomal class (only restriction-site variants present more than once in the sample were considered). Within O,, + 41, four out of 28 pairwise comparisons showed statistically significant disequilibrium by the chi-square test in El Pedroso (Tab. 4). Adjustment of the type I error rate for multiple comparisons by the sequential Bonferroni procedure was used in order to avoid the mistaken rejection of the hypothesis of random association among all the pairwise comparisons (Rice, 1989; Zapata and Alvarez, 1993). The sequential Bonferroni criterion indicated table-wide significance for disequilibrium between restriction-site polymorphisms within O,, + 41 in El Pedroso. The null hypothesis of random association among all pairs of sites at rp49 within O,, + 41 can therefore be rejected. Disequilibrium between polymorphic restriction sites appears to be quite homogeneous across the studied European populations. In fact, those pairs of sites with significant gametic disequilibrium in El Pedroso showed disequilibria with the same sign and similar intensity in Barcelona.
Discussion
Despite the existence of shared polymorphisms between chromosomal classes (a consequence of the exchange between gene arrangements, Rozas and Aguadt, 1993), chromosomal classes OtST1 and O,, + 41 in El Pedroso are well differentiated (Tab. 3). Both haplotype and nucleotide diversity are lower in OtsT1 than in O,, +41, suggesting that the 0, +4 arrangement is older than O,,, as previously discussed (Rozas and Aguadt, 1990, 1993; see also Prevosti, 1971, 1972, and Zapata et al., 1982).
The results of Hudson et al.‘s (1992) test and the homogeneity of linkage disequilibrium across European populations indicate that, for a given chromosomal

364 Rozas et al.
class, there is no genetic differentiation among these European populations. How- ever, these populations are genetically differentiated from Tenerife as indicated both by Hudson et al.‘s (1992) test and by the relative Nm values (Fig. 5).
The rate of migration in Europe is therefore high enough to homogenize the genetic content of a given chromosomal class in these populations. However, O,sTl and O,, + 4l are genetically differentiated. This could be explained either by selection, or by the low relative rates of exchange between chromosomal arrangements. In the case of the rp49 region, positive selection does not seem to be playing an important role: ribosomal protein 49 is monomorphic in natural populations of D. subobscura (Rozas and Aguadt, 1993, 1994) and data on intra and interspecific variation at the rp49 region show no departure from neutrality using the HKA (Hudson et al., 1987) test (Rozas and Aguade, 1994). This would suggest that variation at this region is neutral, and that the rp49 region is not tightly linked to any region with a balanced polymorphism (Hudson and Kaplan, 1988). We conclude therefore that the genetic differentiation between OtST1 and O,, + 4l could be explained by the initial association due to the unique origin of the inversions (see Rozas and Aguade 1993). In this case, and despite the gene flow between gene arrangements (Rozas and Aguade, 1990, 1993), the elapsed time since the origin of inversions would not have been long enough to allow the decay of the initial association. This conclusion might also be supported by the negative values of Tajima’s D. In fact, the unique origin of inversions involves an extreme bottleneck, and in this situation negative values of D would be expected.
The co-existence of latitudinal clines for inversion polymorphism in Europe with high rates of migration among populations confirms that selection is responsible for maintaining the inversion polymorphism in D. subobscura. If gene flow among populations is high enough to homogenize the genetic content of a given chromoso- ma1 class then, in the absence of selection, it should also homogenize the frequen-
Fig. 5. A) Map showing the geographical location of the D. subobscura populations studied. B) Two-
dimensional map obtained by the multidimensional scaling method (Rohlf 1990) based on the number of net nucleotide substitutions between populations (6) (Nei and Li, 1979). Populations with no evidence of genetic differentiation are enclosed in an oval figure. B, Barcelona; T, Ter Apel; P, El Pedroso; Tf, Tenerife. Shaded ( W) and non-shaded symbols ( U) represent the O,, + 4I and O,,,,chromosomal classes,
respectively.

rp49 restriction-site variation 365
ties of the different gene arrangements among populations. This selective explana- tion of the clines was previously reported by Prevosti et al. (1988) based on the observation of similar latitudinal clines in North and South America that were established after the recent colonization of these areas by D. subobscura.
According to the adaptive character of the inversion polymorphism, one would expect that some loci within the inverted region (other than rp49) were under selection. Results of electrophoretic studies of loci within the inversions have been equivocal: some studies (Pinsker and Sperlich, 1981; Larruga and Pinsker, 1984; Pinsker and Biihm, 1989) did not detect genetic differentiation among populations at the Lap, Pept-1 and Acph loci (located in segment I of the 0 chromosome) within a given chromosomal arrangement. Prevosti et al. (1983) showed that the frequency of allele 0.40 of the Pept- 1 locus within the 0, + 4 gene arrangement is negatively correlated with latitude. In addition, Fontdevila et al. (1983) and Rodriguez-Trelles ( 1993) detected seasonal changes repeated over a period of fifteen years for Lap and Pept-I within the OsT arrangement, suggesting that selection acts to maintain the observed patterns of variation for Lap and Pept-1. Moreover, analysis of disequilibrium between Lap and Pept-I for a set of European popula- tions of D. subobscura showed that the observed deviations from random associa- tion are inconsistent with sampling error (Zapata and Alvarez, 1992). Therefore, the study of DNA sequence variation at these loci would be worthwhile: the detection of balancing selection at sites in linkage disequilibrium with the inversions would explain the selective maintenance of the inversion polymorphism.
Acknowledgements
This work was supported by grant PB88-0196 from the Dire&on General de Investigation Cientilica
y Tecnica (Spain) to M. A. and to C. Z.
References
Aguade, M. 1988. Nucleotide sequence comparison of the rp49 gene region between Drosophila subobscura and D. melanogaster. Mol. Biol. Evol. 5: 433-441.
Fontdevila, A., C. Zapata, G. Alvarez, L. Sanchez, J. Mendez and I. Enriquez. 1983. Genetic coadaptation in the chromosomal polymorphism of Drosophila subobscura. I. Seasonal changes of
gametic disequilibrium in a natural population. Genetics 105: 935955. Hudson, R. R., D. D. Boos and N. L. Kaplan. 1992. A statistical test for detecting population
subdivision. Mol. Biol. Evol. 9: 138-151.
Hudson, R. R. and N. L. Kaplan. 1988. The coalescent process in models with selection and
recombination. Genetics 120: 831-840. Hudson, R. R., M. Kreitman, and M. Aguadt. 1987. A test of neutral molecular evolution based on
nucleotide data. Genetics 116: 153-159.
Kreitman, M. and M. Aguadt. 1986. Genetic unifomity in two populations of Drosophila melunogaster as revealed by filter hybridization of four-nucleotide-recognizing restriction enzyme digests. Proc.
Natl. Acad. Sci. USA 83: 3562-3566. Krimbas, C. B. 1992. The inversion polymorphism of Drosophila subobscura. pp. 127-220. In C. B.

366 Rozas et al.
Krimbas and J. R. Powell (eds.), Drosophila Inversion Polymorphism. CRC Press Inc., Boca Raton, Florida.
Krimbas, C. B. and M. Loukas. 1980. The inversion polymorphism of Drosophila subobscura. Evolution- ary Biology 12: 163-234.
Larruga, J. M. and W. Pinsker. 1984. The importance of geographic isolation, structural rearrangement and speciation for the genetic divergence of chromosome 0 in Drosophila subobscura, Drosophila guanche, and Drosophila madeirensis. Z. zool. Syst. Evolut.-forsch. 22: 103- 113.
Loukas, M., C. B. Krimbas, P. Mavragani-Tsipidou and C. D. Kastritsis. 1979. Genetics of Drosophila subobscura populations. VIII. Allozyme loci and their chromosome maps. J. Hered. 70: 17726.
Nei, M. 1982. Evolution of human races at the gene level, pp. 167-181. In B. Bonne-Tamir, T. Cohen, and R. M. Goodman (eds.), Human Genetics, Part A: The Unfolding Genome. Alan R. Liss, New York.
Nei, M. and W.-H. Li. 1979. Mathematical model for studying genetic variations in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA. 76: 526995273.
Nei, M. and F. Tajima. 1981. DNA polymorphism detectable by restriction endonucleases. Genetics 97:
145-163. Pinsker, W. and I. Biihm. 1989. Allozyme divergence between different gene arrangements in Drosophila
subobscura. J. Evol. Biol. 2: 353-366. Pinsker, W. and D. Sperlich. 1981. Geographic pattern of allozyme and inversion polymorphism on
chromosome 0 of Drosophila subobscura and its evolutionary origin. Genetica 57: 51-64. Prevosti, A. 1971. Chromosomal polymorphism in Drosophila subobscura Coll. populations from the
Canary islands. Get&. Ib&r. 23: 69-84. Prevosti, A. 1972. Chromosomal polymorphism in Drosophila subobscura populations from the Madeira
island. Gentt. Iber. 24: 11-21. Prevosti, A., M. P. Garcia, L. Serra, M. Aguade, G. Ribb and E. Sagarra. 1983. Association between
isozyme alleles and chromosomal arrangements in European populations and Chilean colonizers of
Drosophila subobscura, pp. 171-191. In M. C. Ratazzi, J. G. Scandalios and G. S. Whitt (eds.), Isozymes: Current Topics in Biological and Medical Research. Vol. 10: Genetics and Evolution. Alan R. Liss, New York.
Prevosti, A., G. Rib& L. Serra, M. Aguadt, J. Balafia, M. Moncllis and F. Mestres. 1988. Colonization of America by Drosophila subobscuru: Experiment in natural populations that supports the adaptive role of chromosomal-inversion polymorphism. Proc. Natl. Acad. Sci. USA 85: 5597-
5600. Rice, W. R. 1989. Analyzing tables of statistical test. Evolution 43: 2233225. Rodriguez-Trelles, F. 1993. Analisis estacional de sistemas geneticos multiloci y estructura genetica en
una poblacion natural de Drosophila subobscura. Ph. D. thesis, Universidad de Santiago de Compostela.
Rohlf, F. J. 1990. Ntsys-pc. Numerical taxonomy and multivariate analysis system. Version 1.60. Exeter Software, Setauket.
Rozas, J. and M. Aguade. 1990. Evidence of extensive genetic exchange in the rp49 region among polymorphic chromosome inversions in Drosophila subobscuru. Genetics 126: 4177426.
Rozas, J. and M. Aguade. 1991a. Study of an isolated population at the nucleotide level: rp49 region of a Canarian population of Drosophila subobscura. Mol. Biol. Evol. 8: 202-211.
Rozas, J. and M. Aguade. 199lb. Using restiction-map analysis to characterize the colonization process of Drosophila subobscura on the American continent. I. rp49 region. Mol. Biol. Evol. 8: 4477457.
Rozas, J. and M. Aguade. 1993. Transfer of genetic information in the rp49 region of Drosophila subobscuru between different chromosomal gene arrangements. Proc. Natl. Acad. Sci. USA. 90: 8083-8087.
Rozas, J. and M. Aguadt. 1994. Gene conversion is involved in the transfer of genetic information between naturally occurring inversions of Drosophila Proc. Natl. Acad. Sci. USA. 91: 11517-l 1521
Sperlich, D., H. Feuerbach-Mravlag, P. Lange, A. Michaelidis and A. Pentzos-Daponte. 1977. Genetic load and viability distribution in central and marginal populations of Drosophila subobscura. Genetics 86: 8355848.

rp49 restriction-site variation 367
Tajima, F. 1989. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 123: 585-595.
Wright, S. 1951. The genetical structure of populations. Ann. Eugenics 15: 323-354. Zapata, C. and G. Alvarez. 1987. Gametic disequilibrium in populations of Drosophila subobscura: A
review ‘of experimental evidence. Genet. Iber. 39: 5933616. Zapata, C. and G. Alvarez. 1992. The detection of gametic disequilibrium between allozyme loci in
natural populations of Drosophila. Evolution 46: 1900-1917.
Zapata, C. and G. Alvarez. 1993. On the detection of nonrandom associations between DNA polymor- phisms in natural populations of Drosophila. Mol. Biol. Evol. 10: 8233841.
Zapata, C., Santos, M. and G. Alvarez. 1982. Origin of inversions and Wallace’s rule of triads. Evolution 36: 407-409.
Received 24 March, 1994;
accepted 8 September, 1994. Corresponding Editor: 0. Savolainen





























