Embed Size (px)
Citation preview

Parasitology International 52(2003) 259–268
1383-5769/03/$ - see front matter� 2003 Elsevier Ireland Ltd. All rights reserved.doi:10.1016/S1383-5769(03)00029-1
Onset of hepatic erythropoiesis after malarial infection in mice
Ramesh C. Halder , Tetsuya Abe , M. Kaiissar Mannoor , Sufi Reza M. Morshed ,a a a a
Anoja Ariyasinghe , Hisami Watanabe , Hiroki Kawamura , Hiroho Sekikawa ,a a a a
Hiromasa Hamada , Yasuhiro Nishiyama , Hiromichi Ishikawa , Ken Toba , Toru Abo *b b b c a,
Department of Immunology, Niigata University School of Medicine, Niigata 951-8510, Japana
Department of Microbiology, Keio University School of Medicine, Tokyo 160-8582, Japanb
First Department of Internal Medicine, Niigata University School of Medicine, Niigata 951-8510, Japanc
Received 10 June 2002; accepted 16 July 2003
Abstract
Plasmodium yoelii-infected erythrocytes were injected into mice with or without 6.5 Gy irradiation. This irradiationsuppressed erythropoiesis and induced severe immunosuppression. However, these mice showed a rather delayedinfection, suggesting that fresh erythrocytes may become malarial targets. In other words, malarial infection did notpersist without newly generated erythrocytes in mice. We then examined erythropoiesis in the liver and bone marrowof mice with malaria. Surprisingly, erythropoiesis began in the liver. At this time, the serum level of erythropoietin(EPO) was prominently elevated and the EPO mRNA also became detectable in the kidney. Many clusters of redblood cells appeared de novo in the parenchymal space of the liver. These results revealed that malarial infection hada potential to induce the onset of hepatic erythropoiesis in mice.� 2003 Elsevier Ireland Ltd. All rights reserved.
Keywords: Plasmodium yoelii; Liver; Erythropoiesis; Merozoites
1. Introduction
We previously reported that intermediate TCRcells (TCR cells), mainly a NK1.1 subset,int y
expanded in the liver of mice that were injectedwith 10 Plasmodium (P.) yoelii 17XNL-infected4
Abbreviations: P. yoelii, Plasmodium yoelii; NKT cells,natural killer T cells; B6, C57BLy6; MNC, mononuclear cells;RT-PCR, reverse transcription-polymerase chain reaction;EPO, erythropoietin.*Corresponding author. Tel.:q81-25-227-2133; fax:q81-
25-227-0766.E-mail address: [email protected](T. Abo).
erythrocytesw1,2x. Other investigators have alsofocused attention on TCR cells and natural killerint
T (NKT) cells in the liver of mice with malarialinfection w3,4x. TCR cells comprise both aint
NK1.1 subset(i.e. NKT cells) and a NK1.1q y
subset at a ratio of 1:1w5x. NKT cells are generatedby an alternative intrathymic pathway and someof them home to the liver w6–8x, whereasNK1.1 TCR cells are generated extrathymicallyy int
in the liver w9x. All T cells seen in athymic nudemice are classified as NK1.1 TCR cellsw10,11x.y int
The liver is also known to be the site whereactivated T cells are removedw12x. In a previous

260 R.C. Halder et al. / Parasitology International 52 (2003) 259–268
study, TCR cells were isolated from the liver ofint
mice that had recovered fromP. yoelii 17XNLinfection and were transferred into 6.5 Gy-irradi-ated micew1x. These mice did not succumb tomalaria and showed no parasitemia.In light of these findings, we then investigated
how such liver lymphocytes interact withPlas-modium-infected erythrocytes in the liver. Persist-ence of malaria occurs by a transition from theinitial liver stage to the blood stage ofPlasmodiuminfection w13–15x. Some investigators havebelieved that fresh erythrocytes become targets formerozoitesw16,17x. In the present study, we presentevidence that erythropoiesis consistently occurredin the liver after malarial infection. It raises thepossibility that erythropoiesis in the liver mightplay some roles for the persistence of malarialinfection.
2. Materials and methods
2.1. Mice and malarial infection
C57BLy6 (B6) mice were used at the age of8–15 weeks. All mice were fed under specificpathogen-free conditions. Some mice were usedafter 6.5 Gy irradiation(sublethal dose ofg-rayirradiation). Control and irradiated mice wereinfected with 10 P. yoelii 17XNL (non-lethal4
strain)-infected erythrocytesw1x. Parasitemia wasestimated by a cell smear of the blood at theindicated points of time after malarial infection. Insome experiments, erythropoietin(EPO) (200 IUymouse, Chugai Pharmaceuticals Co., Tokyo,Japan) was injected into irradiated malaria-infectedmice to activate erythropoiesisw18x.
2.2. Cell preparation
Mononuclear cells(MNC) were isolated fromthe liver. Hepatic MNC were isolated by a methoddescribed previouslyw19x. Briefly, the liver wasremoved, pressed through 200-gauge stainless steelmesh and suspended in Eagle’s MEM medium(Nissui Pharmaceutical Co., Tokyo, Japan) supple-mented with 5 mM Hepes and 2% heat-inactivatednewborn calf serum. After being washed once withmedium, the cells were fractionated by centrifu-
gation in 15 ml of 35% Percoll solution(Pharma-cia Fine Chemicals, Piscataway, NJ) for 15 min at2000 rpm. The pellet was resuspended in erythro-cyte lysing solution(155 mM MH Cl, 10 mM4
KHCO , 1 mM EDTA–Na, 170 mM Tris, pH 7.3).3
The splenocytes were obtained by forcing thespleen, through stainless steel mesh. Splenocyteswere used after erythrocyte lysis. Bone marrowcells were obtained by flushing the femurs withthe medium followed by erythrocyte lysis.
2.3. Immunofluorescence tests
The surface phenotype of cells was identifiedby two-color immunofluorescence testsw5x.FITC-, PE- or biotin-conjugated mAbs were usedand biotin-conjugated reagents were developedwith TRIcolor-conjugated streptavidin(CaltagLab., San Francisco, CA). mAbs used here includ-ed anti-CD3(145-2C11), anti-IL-2Rb (TM-b1),anti-NK1.1 (PK136), anti-I-A , anti-K and anti-b b
erythrocyte(TER119) antibody(PharMingen Co.,San Diego, CA). Cells were analyzed by FACScan(Becton Dickinson Co., Mountain View, CA). Toprevent nonspecific binding of mAb, CD16y32(2.4G2) was added before staining with labeledmAb. Dead cells were excluded by forward scatter,side scatter and propidium iodide gating. Thefluorescence-positive cells were analyzed withFACScan using LYSIS II software (BectonDickinson).Since mature erythrocytes were ruptured by a
NH Cl-lysing buffer, all remaining TER119 cellsq4
were nucleated erythrocytes.
2.4. Cell sorting
CD3 TER119 cells were purified by a celly q
sorter (FACS Vantage, Becton Dickinson), afterimmunofluorescence staining.
2.5. Serum titer of erythropoietin
Serum titers of EPO were measured by radio-immunoassay(SRL, Tokyo, Japan).
2.6. Detection of mRNA of erythropoietin
Total RNA was extracted from the kidney andliver before and after malarial infection(day 21)
261R.C. Halder et al. / Parasitology International 52 (2003) 259–268
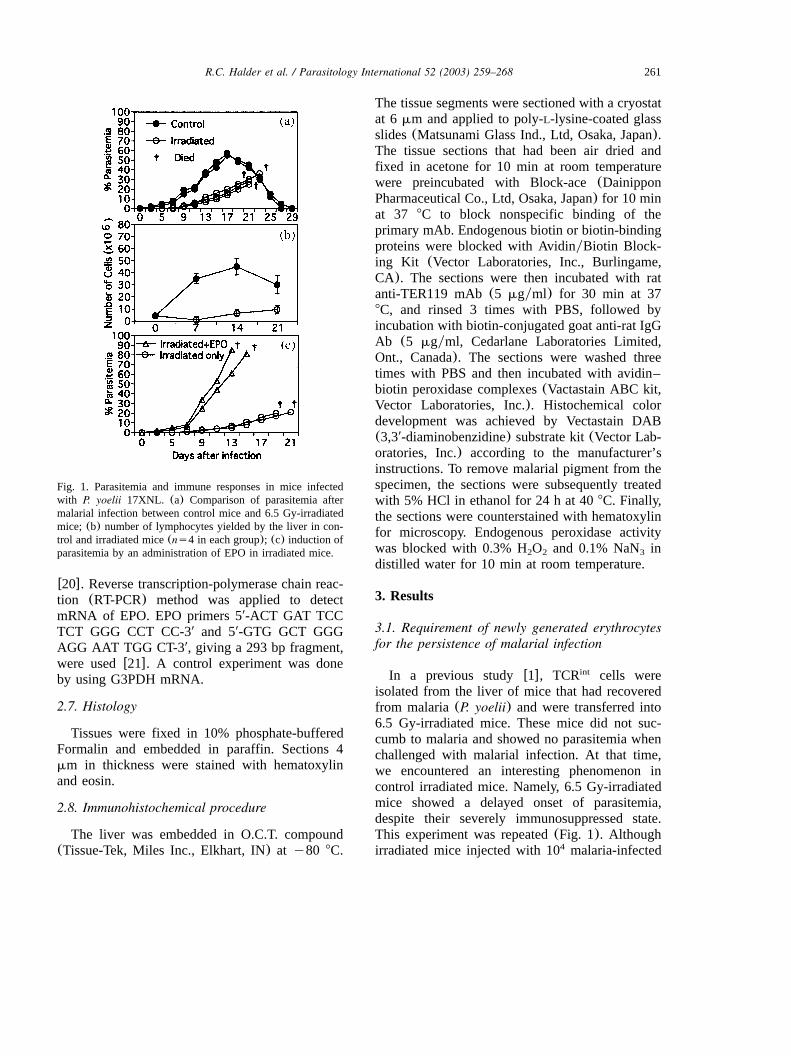
Fig. 1. Parasitemia and immune responses in mice infectedwith P. yoelii 17XNL. (a) Comparison of parasitemia aftermalarial infection between control mice and 6.5 Gy-irradiatedmice; (b) number of lymphocytes yielded by the liver in con-trol and irradiated mice(ns4 in each group); (c) induction ofparasitemia by an administration of EPO in irradiated mice.
w20x. Reverse transcription-polymerase chain reac-tion (RT-PCR) method was applied to detectmRNA of EPO. EPO primers 59-ACT GAT TCCTCT GGG CCT CC-39 and 59-GTG GCT GGGAGG AAT TGG CT-39, giving a 293 bp fragment,were usedw21x. A control experiment was doneby using G3PDH mRNA.
2.7. Histology
Tissues were fixed in 10% phosphate-bufferedFormalin and embedded in paraffin. Sections 4mm in thickness were stained with hematoxylinand eosin.
2.8. Immunohistochemical procedure
The liver was embedded in O.C.T. compound(Tissue-Tek, Miles Inc., Elkhart, IN) at y80 8C.
The tissue segments were sectioned with a cryostatat 6mm and applied to poly-L-lysine-coated glassslides(Matsunami Glass Ind., Ltd, Osaka, Japan).The tissue sections that had been air dried andfixed in acetone for 10 min at room temperaturewere preincubated with Block-ace(DainipponPharmaceutical Co., Ltd, Osaka, Japan) for 10 minat 37 8C to block nonspecific binding of theprimary mAb. Endogenous biotin or biotin-bindingproteins were blocked with AvidinyBiotin Block-ing Kit (Vector Laboratories, Inc., Burlingame,CA). The sections were then incubated with ratanti-TER119 mAb(5 mgyml) for 30 min at 378C, and rinsed 3 times with PBS, followed byincubation with biotin-conjugated goat anti-rat IgGAb (5 mgyml, Cedarlane Laboratories Limited,Ont., Canada). The sections were washed threetimes with PBS and then incubated with avidin–biotin peroxidase complexes(Vactastain ABC kit,Vector Laboratories, Inc.). Histochemical colordevelopment was achieved by Vectastain DAB(3,39-diaminobenzidine) substrate kit(Vector Lab-oratories, Inc.) according to the manufacturer’sinstructions. To remove malarial pigment from thespecimen, the sections were subsequently treatedwith 5% HCl in ethanol for 24 h at 408C. Finally,the sections were counterstained with hematoxylinfor microscopy. Endogenous peroxidase activitywas blocked with 0.3% H O and 0.1% NaN in2 2 3
distilled water for 10 min at room temperature.
3. Results
3.1. Requirement of newly generated erythrocytesfor the persistence of malarial infection
In a previous studyw1x, TCR cells wereint
isolated from the liver of mice that had recoveredfrom malaria(P. yoelii) and were transferred into6.5 Gy-irradiated mice. These mice did not suc-cumb to malaria and showed no parasitemia whenchallenged with malarial infection. At that time,we encountered an interesting phenomenon incontrol irradiated mice. Namely, 6.5 Gy-irradiatedmice showed a delayed onset of parasitemia,despite their severely immunosuppressed state.This experiment was repeated(Fig. 1). Althoughirradiated mice injected with 10 malaria-infected4

262 R.C. Halder et al. / Parasitology International 52 (2003) 259–268
erythrocytes finally died from malaria, theyshowed a delayed onset of parasitemia(Fig. 1a).There was no expansion of lymphocytes in theliver of these irradiated mice even after malariainfection (Fig. 1b). These results suggested thepossibility that newly generated erythrocytes arerequired for the parasite infection in vivo. Tofurther examine this possibility, we injected EPOinto these irradiated malaria-infected mice on day7 (Fig. 1c). In parallel with a new generation oferythrocytes, parasitemia appeared in the bloodand these mice quickly died due to their immu-nosuppressed state.
3.2. Onset of erythropoiesis in the liver
Although the importance of newly generatederythrocytes for the targets of parasites wasrevealed, malarial inflammation is always moreprominent in the liver than in the bone marrow.Since c-kit stem cells are abundant even in theq
adult liver w22x, we examined the possibility thatsuch stem cells give rise to erythrocytes in theliver before and after malarial infection(Fig. 2).Two-color staining for CD3 and IL-2Rb and thatfor CD3 and TER119(an erythroid marker) wereconducted in the liver(Fig. 2A). As shown pre-viously w1x, unconventional T cells(i.e. CD3 IL-int
2Rb ) expanded in the liver from day 7 afterq
malarial infection. Initially, CD3 TER119 cellsy q
(erythrocytes) were almost absent in the liver(-1%), but they appeared from day 14 after theinfection (indicated by an arrowhead). In thisidentification, only nucleated cells were isolatedfrom the liver and denucleated erythrocytes weredestroyed by a lysing buffer. Therefore, allCD3 TER119 cells were immature nucleatedy q
erythrocytes. In other words, 20%(Fig. 2A) of3=10 nucleated cellsymouse (Fig. 1B) were7
estimated to be such immature erythrocytes in theliver on day 21 after malarial infection. Theseerythrocytes still expressed MHC class I antigens,but not class II antigens, on the surface(Fig. 2B).Almost all erythrocytes in the peripheral bloodwere denucleated and had lost MHC class I anti-gens(data not shown).
To determine how erythropoiesis occurred in theliver after malarial infection, serum titers of EPOwere measured by radioimmunoassay(Fig. 2C).In control mice, serum titers of EPO ranged from30 to 50 mUyml. Prominent increase in the titerwas observed after malarial infection as early asday 7. To detect the actual site for the productionof EPO, total RNA was extracted from both thekidney and the liver before and after malarialinfection (day 14). The presence of EPO mRNAwas tested for by RT-PCR method. Only in thekidney was a prominent signal for EPO mRNAdetected after malarial infection(Fig. 2D). Such asignal was not detected in the liver even aftermalarial infection(data not shown).
3.3. Histology of infected liver
A liver section (on day 21 after infection)stained by hematoxylin–eosin was observed bylight microscopy(Fig. 3). Many massive clustersof nucleated erythroid and lymphoid cells werefound in the parenchymal space of the liver. Thissituation resembles the erythropoiesis seen in thefetal liver, although the demarcation of erythropoi-esis area in the liver was more prominent inmalarial infection than that in fetal livererythropoiesis.
3.4. Immunohistochemical staining of nucleatederythrocytes in the liver
To directly detect nucleated erythrocytes in theliver of mice with malarial infection, we conductedan immunohistochemical staining of the liver byusing anti-erythrocyte mAb(TER119) (Fig. 4).When the liver was isolated from control B6 mice,TER119 cells were present in the vessels(=40).q
This was also true in a high magnification(=200).Then, the liver was isolated from mice infectedwith P. yoelii (day 21 after infection). Since muchmalarial pigments were distributed in the liver ofthese mice, liver specimens with and without thetreatment to eliminate malarial pigment were rep-resented. With or without the elimination proce-dure, many nucleated TER119 cells were foundq
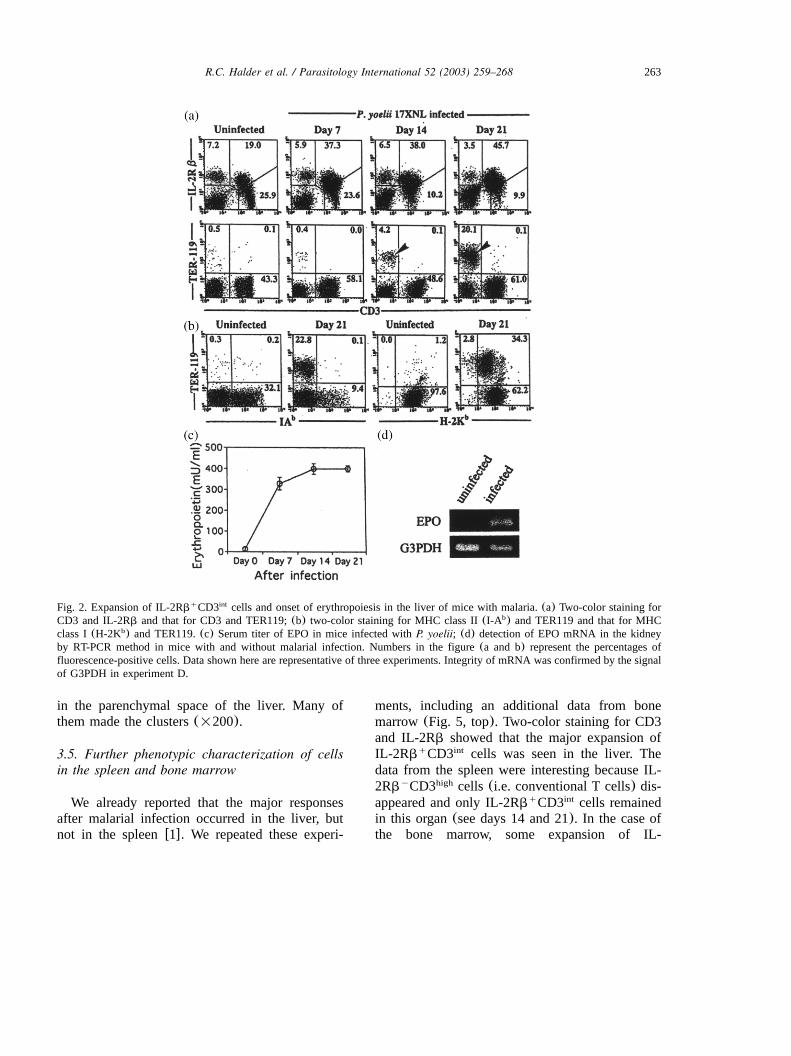
263R.C. Halder et al. / Parasitology International 52 (2003) 259–268
Fig. 2. Expansion of IL-2Rb CD3 cells and onset of erythropoiesis in the liver of mice with malaria.(a) Two-color staining forq int
CD3 and IL-2Rb and that for CD3 and TER119;(b) two-color staining for MHC class II(I-A ) and TER119 and that for MHCb
class I(H-2K ) and TER119.(c) Serum titer of EPO in mice infected withP. yoelii; (d) detection of EPO mRNA in the kidneyb
by RT-PCR method in mice with and without malarial infection. Numbers in the figure(a and b) represent the percentages offluorescence-positive cells. Data shown here are representative of three experiments. Integrity of mRNA was confirmed by the signalof G3PDH in experiment D.
in the parenchymal space of the liver. Many ofthem made the clusters(=200).
3.5. Further phenotypic characterization of cellsin the spleen and bone marrow
We already reported that the major responsesafter malarial infection occurred in the liver, butnot in the spleenw1x. We repeated these experi-
ments, including an additional data from bonemarrow (Fig. 5, top). Two-color staining for CD3and IL-2Rb showed that the major expansion ofIL-2Rb CD3 cells was seen in the liver. Theq int
data from the spleen were interesting because IL-2Rb CD3 cells(i.e. conventional T cells) dis-y high
appeared and only IL-2Rb CD3 cells remainedq int
in this organ(see days 14 and 21). In the case ofthe bone marrow, some expansion of IL-
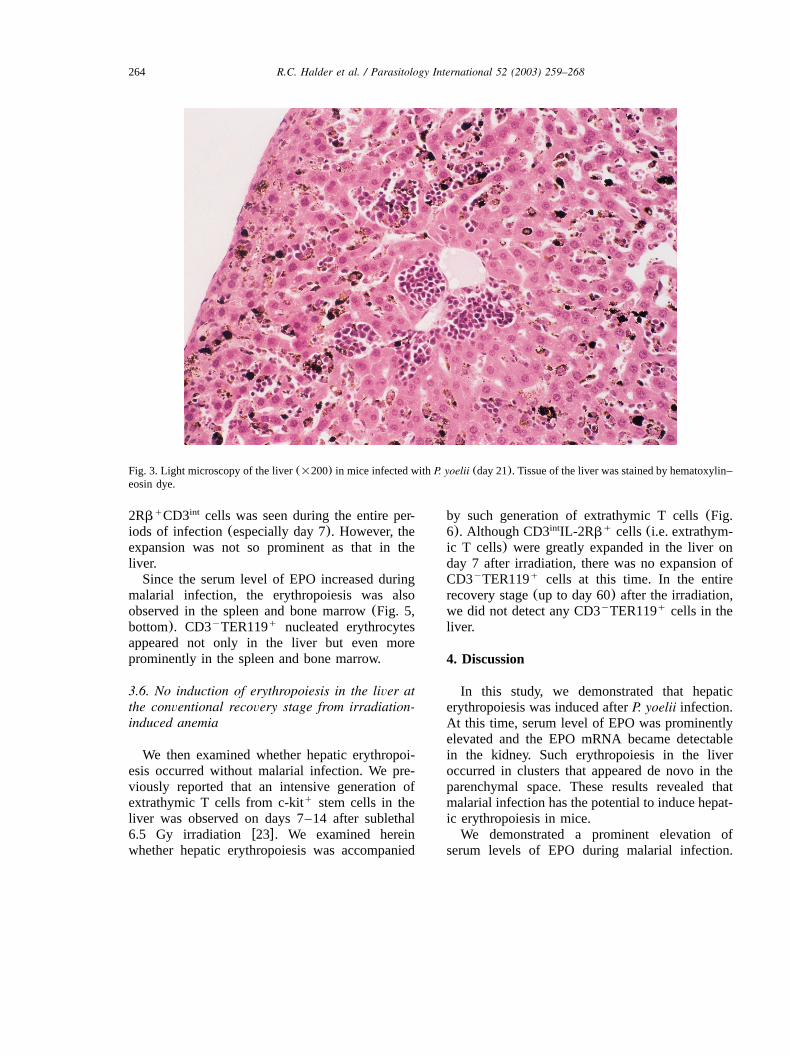
264 R.C. Halder et al. / Parasitology International 52 (2003) 259–268
Fig. 3. Light microscopy of the liver(=200) in mice infected withP. yoelii (day 21). Tissue of the liver was stained by hematoxylin–eosin dye.
2Rb CD3 cells was seen during the entire per-q int
iods of infection(especially day 7). However, theexpansion was not so prominent as that in theliver.Since the serum level of EPO increased during
malarial infection, the erythropoiesis was alsoobserved in the spleen and bone marrow(Fig. 5,bottom). CD3 TER119 nucleated erythrocytesy q
appeared not only in the liver but even moreprominently in the spleen and bone marrow.
3.6. No induction of erythropoiesis in the liver atthe conventional recovery stage from irradiation-induced anemia
We then examined whether hepatic erythropoi-esis occurred without malarial infection. We pre-viously reported that an intensive generation ofextrathymic T cells from c-kit stem cells in theq
liver was observed on days 7–14 after sublethal6.5 Gy irradiation w23x. We examined hereinwhether hepatic erythropoiesis was accompanied
by such generation of extrathymic T cells(Fig.6). Although CD3 IL-2Rb cells(i.e. extrathym-int q
ic T cells) were greatly expanded in the liver onday 7 after irradiation, there was no expansion ofCD3 TER119 cells at this time. In the entirey q
recovery stage(up to day 60) after the irradiation,we did not detect any CD3 TER119 cells in they q
liver.
4. Discussion
In this study, we demonstrated that hepaticerythropoiesis was induced afterP. yoelii infection.At this time, serum level of EPO was prominentlyelevated and the EPO mRNA became detectablein the kidney. Such erythropoiesis in the liveroccurred in clusters that appeared de novo in theparenchymal space. These results revealed thatmalarial infection has the potential to induce hepat-ic erythropoiesis in mice.We demonstrated a prominent elevation of
serum levels of EPO during malarial infection.
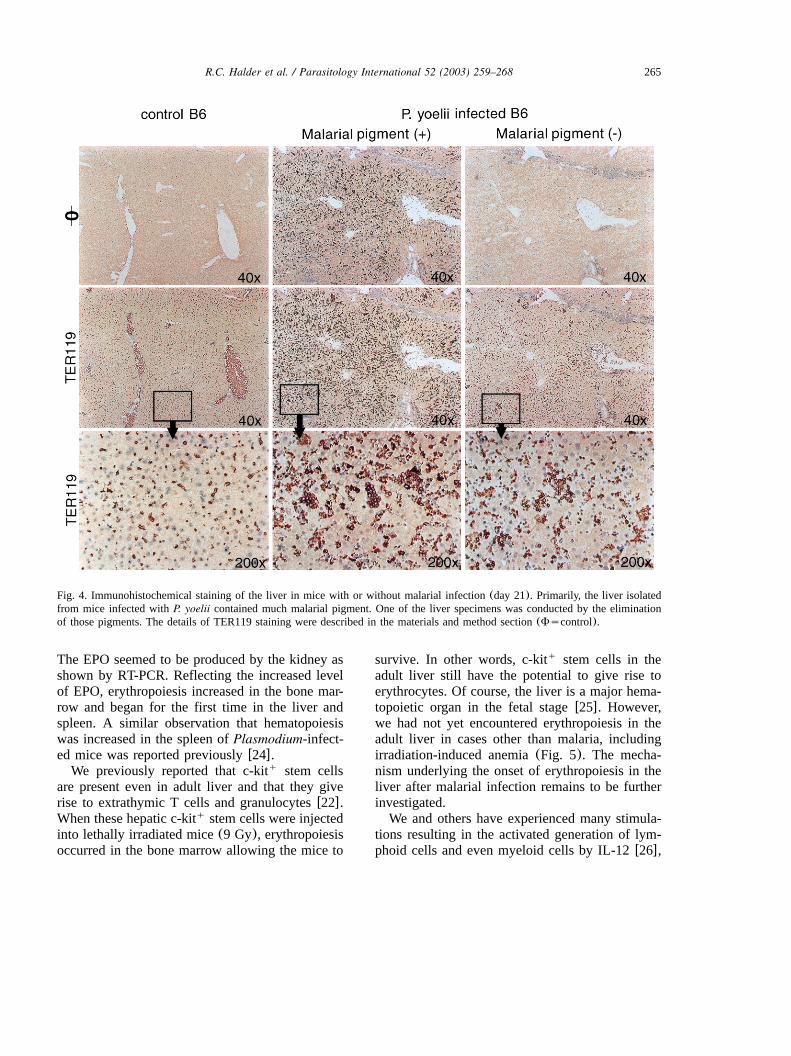
265R.C. Halder et al. / Parasitology International 52 (2003) 259–268
Fig. 4. Immunohistochemical staining of the liver in mice with or without malarial infection(day 21). Primarily, the liver isolatedfrom mice infected withP. yoelii contained much malarial pigment. One of the liver specimens was conducted by the eliminationof those pigments. The details of TER119 staining were described in the materials and method section(Fscontrol).
The EPO seemed to be produced by the kidney asshown by RT-PCR. Reflecting the increased levelof EPO, erythropoiesis increased in the bone mar-row and began for the first time in the liver andspleen. A similar observation that hematopoiesiswas increased in the spleen ofPlasmodium-infect-ed mice was reported previouslyw24x.We previously reported that c-kit stem cellsq
are present even in adult liver and that they giverise to extrathymic T cells and granulocytesw22x.When these hepatic c-kit stem cells were injectedq
into lethally irradiated mice(9 Gy), erythropoiesisoccurred in the bone marrow allowing the mice to
survive. In other words, c-kit stem cells in theq
adult liver still have the potential to give rise toerythrocytes. Of course, the liver is a major hema-topoietic organ in the fetal stagew25x. However,we had not yet encountered erythropoiesis in theadult liver in cases other than malaria, includingirradiation-induced anemia(Fig. 5). The mecha-nism underlying the onset of erythropoiesis in theliver after malarial infection remains to be furtherinvestigated.We and others have experienced many stimula-
tions resulting in the activated generation of lym-phoid cells and even myeloid cells by IL-12w26x,

266 R.C. Halder et al. / Parasitology International 52 (2003) 259–268
Fig. 5. Further phenotypic characterization of cells in the liver, spleen and bone marrow after malarial infection. Two-color stainingfor CD3 and IL-2Rb and that for CD3 and TER119 were conducted at the indicated points of time. Numbers in the figure representthe percentages of fluorescence-positive cells(BMsbone marrow).
estrogenw27x, catecholaminesw28x, glucocorticoidsw29x and others. However, we did not detect thegeneration of nucleated erythrocytes in the liverunder these conditions(data not shown).We should mention the nature of c-kit stemq
cells in the liver. When parabiosis experimentswere performed in mice(Ly5.1 and Ly5.2 mice),extrathymic T cells in the liver showed difficultiesin mixing with each otherw30x. On the other hand,conventional T and B cells in the periphery wereable to mix well with partner cells by 14 days
after parabiosis. This was due to the fact thatperipheral T and B cells were supplied by otherorgans. In contrast, many extrathymic T cells aswell as their precursors(including c-kit stemq
cells) did not mix with partner cells. It is thereforeconceivable that newly generated erythrocytes inthe liver after malarial infection might be derivedfrom stem cells existing in the liver itself ratherthan in the bone marrow. In any case, the onset ofhepatic erythropoiesis is an important physiologi-cal response seen in malaria.

267R.C. Halder et al. / Parasitology International 52 (2003) 259–268
Fig. 6. A limited expansion of extrathymic T cells in the liver of 6.5 Gy-irradiated mice without malarial infection. MNC wereisolated from the liver at the indicated days after sublethal irradiation. Two-color staining for CD3 and TER119, that for CD3 andIL-2Rb and that for CD3 and NK1.1 were conducted. Date shown here are representative of three experiments.
In the case of human malaria, CD36 antigens(sequestrin) on erythrocytes are known to mediateadherence and subsequent infection ofP. falcipa-rum w31,32x. These CD36 antigens are expressedon immature erythrocytes. Newly generated eryth-rocytes may be critically important in humanmalarial infections as well as in rodent malaria. Astudy on the expression of CD36 antigens onnucleated erythrocytes in the liver and other organsof mice with or without malarial infection isongoing.
Acknowledgments
This work was supported by a Grant-in-Aid forScientific Research on Priority Areas 08281103from the Ministry of Education, Science, Cultureand Sports of Japan. We thank Dr Hiroki Maruy-ama(the Second Department of Internal Medicine,Niigata University School of Medicine, Niigata,Japan) for helpful suggestions and Mrs YukoKaneko for preparation of the manuscript.
References
w1x Weerasinghe A, Sekikawa H, Watanabe H, et al. Asso-ciation of intermediate T cell receptor cells, mainly theirNK1.1 subset, with protection from malaria. Celly
Immunol 2001;207:28–35.w2x Mannoor MK, Halder RC, Morshed SRM, et al. Essen-
tial role of extrathymic T cells in protection againstmalaria. J Immunol 2002;169:301–6.
w3x Pied S, Roland J, Louise A, et al. LiverCD4 CD8 NK1.1 TCRab intermediate cells increasey y q
during experimental malaria infection and are able toexhibit inhibitory activity against the parasite liver stagein vitro. J Immunol 2000;164:1463–9.
w4x Molano A, Park SH, Chiu YH, Nosseir S, Bendelac A,Tsuji M. The IgG response to the circumsporozoiteprotein is MHC class II-dependent and CD1d-independ-ent: exploring the role of GPIs in NK T cell activationand antimalarial responses. J Immunol 2000;164:5005–9.
w5x Watanabe H, Miyaji C, Kawachi Y, et al. Relationshipsbetween intermediate TCR cells and NK1.1 T cells inq
various immune organs. NK1.1 T cells are presentq
within a population of intermediate TCR cells. J Immu-nol 1995;155:2972–83.
w6x Schulz RJ, Parkes A, Mizoguchi E, Bhan AK, KoyasuS. Development of CD4 CD8 abTCR NK1.1 Ty y q q

268 R.C. Halder et al. / Parasitology International 52 (2003) 259–268
lymphocytes: thymic selection by self antigen. J Immu-nol 1996;157:4379–89.
w7x Coles MC, Raulet DH. NK1.1 T cells in the liverq
arise in the thymus and are selected by interactions withclass I molecules on CD4 CD8 cells. J Immunolq q
2000;164:2412–8.w8x Emoto M, Miyamoto M, Namba K, et al. Participation
of leukocyte function-associated antigen-1 and NK cellsin the homing of thymic CD8 NKT cells to the liver.q
Eur J Immunol 2000;30:3049–56.w9x Abo T, Kawamura T, Watanabe H. Physiological
responses of extrathymic T cells in the liver. ImmunolRev 2000;174:135–49.
w10x Emoto M, Emoto Y, Kaufmann SHE. CD8abq
TCRab lymphocytes expressing skewedintermediate
TCRVb repertoire in the liver of aged athymicnuynumice. J Immunol 1997;158:1041–50.
w11x Halder RC, Kawamura T, Bannai M, et al. Intensivegeneration of NK1.1 extrathymic T cells in the livery
by injection of bone marrow cells isolated from micewith a mutation of polymorphic major histocompatibil-ity complex antigens. Immunology 2001;102:450–9.
w12x Crispe IN, Mehal WZ. Strange brew: T cells in theliver. Immunol Today 1996;17:522–5.
w13x Plebanski M, Hill AV. The immunology of malariainfection. Curr Opin Immunol 2000;12:437–41.
w14x Krzych U, Schwenk R, Guebre-Xabier M, et al. Therole of intrahepatic lymphocytes in mediating protectiveimmunity induced by attenuatedPlasmodium bergheisporozoites. Immunol Rev 2000;174:123–34.
w15x Taylor-Robinson AW, Phillips RS. Infective dose mod-ulates the balance between Th1- and Th2-regulatedimmune responses during blood-stage malaria infection.Scand J Immunol 1998;48:527–34.
w16x Aikawa M, Atkinson CT. Immunoelectron microscopyof parasites. Adv Parasitol 1990;29:151–214.
w17x Chitnis CE, Miller LH. Identification of the erythrocytebinding domains ofPlasmodium vivax andPlasmodiumknowlesi proteins involved in erythrocyte invasion. JExp Med 1994;180:497–506.
w18x Koya T, Narita J, Honda S, Watanabe H, Arakawa M,Abo T. Erythropoietin induces the expansion of c-kitq
progenitors for myeloid and erythroid cells, but not forlymphoid cells, in the bone marrow and liver. Eur JHaematol 1999;63:306–12.
w19x Kawamura H, Kawamura T, Kokai Y, et al. Expansionof extrathymic T cells as well as granulocytes in theliver and other organs of G-CSF transgenic mice: whythey lost the ability of hybrid resistance. J Immunol1999;16:5957–64.
w20x Narita J, Kawamura T, Miyaji C, et al. Abundance ofNKT cells in the salivary glands but absence thereof inthe liver and thymus ofalyyaly mice with Sjogren¨syndrome. Cell Immunol 1999;192:149–58.
w21x Shoemaker CB, Mitsock LD. Murine erythropoietingene: cloning, expression and human gene homology.Mol Cell Biol 1986;6:849–58.
w22x Watanabe H, Miyaji C, Seki S, Abo T. c-kit stem cellsq
and thymocyte precursors in the livers of adult mice. JExp Med 1996;184:687–93.
w23x Shimizu T, Sugahara S, Oya H, et al. The majority oflymphocytes in the bone marrow, thymus and extracym-ic T cells in the liver are generatedin situ from theirown preexisting precursors. Microbiol Immunol1999;43:596–608.
w24x Asami M, Owhashi M, Abe T, Nawa Y. A comparativestudy of the kinetic changes of hemopoietic stem cellsin mice infected with lethal and non-lethal malaria. IntJ Parasitol 1992;22:43–7.
w25x Kawamura T, Toyabe S, Moroda T, et al. Neonatalgranulocytosis is a postpartum event which is seen inthe liver as well as in the blood. Hepatology1997;26:1567–72.
w26x Eberl G, MacDonald HR. Rapid death and regenerationof NKT cells in anti-CD3́ - or IL-12-treated mice: amajor role for bone marrow in NKT cell homeostasis.Immunity 1998;9:345–53.
w27x Narita J, Miyaji C, Watanabe H, et al. Differentiationof forbidden T cell clones and granulocytes in theparenchymal space of the liver in mice treated withestrogen. Cell Immunol 1998;185:1–13.
w28x Minagawa M, Oya H, Yamamoto S, et al. Intensiveexpansion of natural killer T cells in the early phase ofhepatocyte regeneration after partial hepatectomy inmice and its association with sympathetic nerve activa-tion. Hepatology 2000;31:907–15.
w29x Shimizu T, Kawamura T, Miyaji C, et al. Resistance ofextrathymic T cells to stress and the role of endogenousglucocorticoids in stress-associated immuno suppres-sion. Scand J Immunol 2000;51:285–92.
w30x Suzuki S, Sugahara S, Shimizu T, et al. Low level ofmixing of partner cells seen in extrathymic T cells inthe liver and intestine of parabiotic mice: its biologicalimplication. Eur J Immunol 1998;28:3719–29.
w31x Oquendo P, Hundt E, Lawler J, Seed B. CD36 directlymediated cytoadherence ofPlasmodium falciparum par-asitized erythrocytes. Cell 1989;58:95–101.
w32x Ockenhouse CF, Klotz FW, Tandon NN, Jamieson GA.Sequestrin, a CD36 recognition protein onPlasmodiumfalciparum malaria-infected erythrocytes identified byanti-idiotype antibodies. Proc Natl Acad Sci USA1991;88:3175–9.





























