Embed Size (px)
Citation preview

&p.1:Abstract Certain components of the jaw apparatus, orlantern, of regular sea-urchins form a ‘compass system’the function of which has still to be established. Thissystem includes ten compass depressors (CDs) whichconnect the compass ossicles to the inner edge of thetest. Previous studies focused on the CDs of euechinoidsea-urchins. This paper provides the first detailed ac-count of the organization and mechanical behaviour ofthe CDs of a cidaroid sea-urchin, Stylocidaris affinis.The outermost layer of its CDs is a coelothelium com-prising apical peritoneocytes, sparse subapical myo-cytes and two types of granule-containing cells. Thecentral core of the CD is dominated by longitudinallyorientated collagen fibrils arranged in bundles surround-ed by networks of beaded microfibrils. Myocytes arescattered throughout the core and are always surround-ed by a basal lamina which usually encloses the sametwo types of granule-containing cells observed in thecoelothelium. Each CD is attached to the outer coelo-mic septum by a mesentery, both the connective tissueand coelothelia of which lack myocytes and granule-containing cells. From data on the relationship betweenthe vertical position of the lantern and the passive andactive forces developed by intact CD sets, it appearsthat the CDs resist elevation of the lantern above its‘resting position’ and that they develop a maximal con-tractile force when their length corresponds to that atthe resting position. The connective tissue of the CDshas the capacity to undergo nervously mediated changesin stretch resistance, although this is expressed moreweakly than in the euechinoid CDs. These results are
related to current views on the physiological role of thecompass system.&bdy:
A. Introduction
An important feature of animal design is the ubiquitousoccurrence of close anatomical and functional associa-tions between muscle and connective tissue. Vertebrateskeletal muscles provide an obvious illustration of this,where tendons transmit force to the rigid skeleton, andthe hierarchy of intramuscular connective tissue, endo-mysium, perimysium and epimysium, both transmitsforce and maintains the alignment of the muscle fibres(Herzog and Loitz 1994; Nishimura et al. 1994; Purslow1994). Other well-known examples are certain molluscmuscles that are permeated by connective tissue net-works (Shadwick and Gosline 1985; Olaechea et al.1993) and the various patterns of smooth muscle and ex-tracellular matrix components in the walls of blood ves-sels and other tubular organs (Davison et al. 1995). Inthe Echinodermata there is a wide diversity of such com-binations, ranging from associations between discretebut contiguous muscles and ligaments (e.g. the ophiuroidintervertebral ligament and muscles; see Wilkie 1988) tointricate admixtures of myocytes and collagen fibressuch as occur in the starfish lower transverse ambulacralmuscle (Eylers 1976), the sea-urchin spine ligament(Smith et al. 1981) and, the subject of this paper, thecompass depressors (CDs) of the sea-urchin lantern.
The sea-urchin lantern is first and foremost a foodcollecting and processing apparatus. However, in all reg-ular sea-urchins it includes a set of structural compo-nents comprising the ‘compass system’ which do notcontribute directly to food handling. The compasssystem consists of: (1) five skeletal elements, the com-passes, lying on top of the lantern and hinged at their in-ner ends to the underlying rotular ossicles, (2) five com-pass elevator muscles which link adjacent compasses andwhose contraction causes the compasses to rotate up-wards and (3) the CDs themselves: ten straps of soft tis-
I.C. Wilkie (✉)Department of Biological Sciences,Glasgow Caledonian University,70 Cowcaddens Road, Glasgow G4 0BA, UKe-mail: [email protected],Tel.: 0141 331 3207, Fax: 0141 331 3208
M.D. Candia Carnevali · F. BonasoroDipartimento di Biologia “Luigi Gorini”,Università degli Studi di Milano, Via Celoria 26,I-20133 Milan, Italy&/fn-block:
Zoomorphology (1998) 118:87–101 © Springer-Verlag 1998
O R I G I N A L A RT I C L E
&roles:Iain C. Wilkie · M. Daniela Candia CarnevaliFrancesco Bonasoro
Organization and mechanical behaviour of myocyte–ligament compositesin a sea-urchin lantern: the compass depressors of Stylocidaris affinis(Echinodermata, Echinoida)
&misc:Accepted: 15 March 1998

sue which run from the forked outer ends of the com-passes to the edge of the sea-urchin’s test (perignathicgirdle) bordering the mouth area (Fig. 1). The compassossicles are attached to membranous structures, the exte-rior coelomic septa, that form fluid-filled pouches sepa-rating the lantern and its muscles from the main bodycavity. These pouches are connected to external or inter-nal, gill-like appendages, and it has been suggested thatthe compass system functions as a respiratory pump thatdelivers oxygen to the lantern muscles (von Uexküll1896) and/or as a pressure-compensating device that sta-bilizes pressure within the rigid test whilst the lantern isbeing protracted and retracted during feeding activity(Kier 1974).
Although the CDs are contractile and were consideredformerly to be muscles, the ultrastructural investigationsof Saita (1969), Lanzavecchia et al. (1988) and Wilkie etal. (1992) established that those of Paracentrotus lividus(Lamarck, 1816) (Euechinoida) are essentially ligamentsassociated with a myoepithelium which representsaround 8% of the total cross-sectional area. Wilkie et al.(1992) also provided information on the contractile be-haviour of the CD and demonstrated that its connectivetissue component consists of mutable collagenous tissue,
i.e. its tensile stiffness is variable and under direct ner-vous control (see Wilkie 1996).
This paper deals with the CDs of Stylocidaris affinis(Philippi, 1845) (Cidaroida). Comparative investigationshave identified significant differences between the lan-terns of cidaroid and euechinoid sea-urchins regardingboth their structure and functioning (Märkel 1979; Can-dia Carnevali et al. 1993; Andrietti et al. 1990, 1993;Wilkie et al. 1993, 1996, 1998). Our primary aim in thepresent study was to determine whether these differencesextend to the CDs by describing the ultrastructural orga-nization of these structures in S. affinisand by character-izing their contractile behaviour and passive mechanicalproperties.
B. Materials and methods
Specimens of S. affiniswith a test diameter (TD) of 28–45 mmwere collected by scuba divers from depths of 60–70 m in the Gulfof Taranto, Italy. They were transported to the University of Milanand maintained in aquaria at 14–16°C.
I. Microscopy
1. Transmission electron microscopy
Specimens were fixed in 2% glutaraldehyde in 0.1 M cacodylatebuffer at pH 7.2, postfixed in 1% osmium tetraoxide in the samebuffer, dehydrated in an alcohol series and embedded in an Epon812–Araldite mixture. Semithin sections (1 µm) were stained withcrystal violet and basic fuchsin, and observed in a Jenaval lightmicroscope. Ultrathin sections (50 nm) were stained with uranylacetate and lead citrate and observed in a Jeol 100 SX electron mi-croscope.
2. Histometry
The proportion of the CD cross-sectional area occupied by myo-cytes was estimated from semithin transverse sections by a point-counting method. Sections were examined at a final magnificationof ×1000 using an eyepiece graticule with a squared grid, and foreach field of view the number of grid intersections overlying myo-cytes was expressed as a percentage of the total number of inter-sections overlying the whole CD. Counts were made from threedifferent sections at each of four different vertical levels in theCD, and from two or three non-overlapping fields in each section.
II. Mechanical tests
1. Lantern preparation
Animals were prepared for experimentation by removing the toptwo-thirds of the test. The remainder, consisting of the lower one-third of the test and the intact lantern complex, was immersed in ananaesthetic solution of 0.1% propylene phenoxetol in seawater forat least 30 min, during which time the maximum TD, perignathicgirdle diameter (see Fig. 2a) and length of the CDs were measured.The coelomic septa and all the compass elevator, protractor and re-tractor muscles were transected, and the peristomial membrane wasexcised completely, thus leaving the lantern connected to the testby only a full set of CDs. Taking every precaution to avoid over-stretching the CDs, an aluminium rod (diameter 1.5 mm) was in-serted down the central axis of the lantern until its tip projected be-yond the teeth by a few mm, and the tip was bent into a right angle.
88
Fig. 1a, b Lantern of Stylocidaris affinis. a Drawing of the wholelantern system, on the leftshowing the skeleton only and on theright with the main muscles and ligaments added. a Apophysis, amambulacrum, cd compass depressor, cemcompass elevator muscle,cp compass, inm interpyramidal muscle, j jaw, pm peristomialmembrane, prm protractor muscle, rem retractor muscle, r rotula.b Diagrammatic vertical section through the lantern system (not toscale). Arrowsexterior coelomic septum, ap ampullae of tube-feet,oeoesophagus, sStewart’s organs, tf tube-feet, to tooth&/fig.c:
The whole preparation was placed in a large volume of pure seawa-ter for at least 20 min to recover from the anaesthetic.
The lower one-third of the test was clamped horizontally, andthe upper end of the aluminium rod was attached to an isometricforce transducer which was fixed rigidly to a manipulator(Fig. 2a). Adjustment of the manipulator enabled the vertical posi-tion of the transducer to be altered by known distances. Elevationof the transducer pulled the lantern upwards and thus stretched theCDs connecting the lantern to the clamped test. All experimentswere conducted with the preparations held in air at room tempera-ture (23–26°C), but kept moist by frequent additions of seawateror test solution.
At the start of each experiment, the height of the lantern withrespect to the test was adjusted until it corresponded to the normalresting position in the living animal, i.e. where it is protracted byabout 1.5 mm below the level at which the peristomial membraneis horizontal (Andrietti et al. 1993). Hereafter, the location of thelantern is quantified as mm ‘retraction’ (displacement above thisresting position).
2. Dose-response experiments
The contractile properties of the CDs were examined by stimulat-ing them with either elevated K+ concentrations or acetylcholine.Initial experiments were conducted to determine the optimum dos-es of these agents. Preparations were set up as in Fig. 2a and thelanterns were held at 0.5 mm retraction to pre-stretch the tissues.Each preparation was stimulated using either mixtures of 0.56 M
potassium chloride solution and seawater giving a range of K+
concentrations (20–100 mM), or a series of dilutions of acetylcho-line chloride (10–9–10–3 M) in seawater. For each concentration ofK+ ions or acetylcholine all the CDs of the preparations wereflooded with solution until the contractile force stopped rising.The preparation was then drenched with seawater until relaxationwas complete, and the next concentration was then applied.
3. Characterization of contractile properties
The protocol for these experiments is illustrated in Fig. 2b. Withthe lantern 0.5 mm below the resting position, the preparation wasflooded with the optimum concentration of K+ ions or acetylcho-line until the contractile force maximized, at which point the prep-aration was washed with seawater. Once relaxation was complete,the transducer was elevated by 0.5 mm, thus increasing passivetension on the CDs. Once the passive tension had stabilized, thepreparation was stimulated and washed as before. This sequencewas repeated as the lantern was elevated by further 0.5 mm incre-ments up to a maximum of 2 mm beyond the resting position.
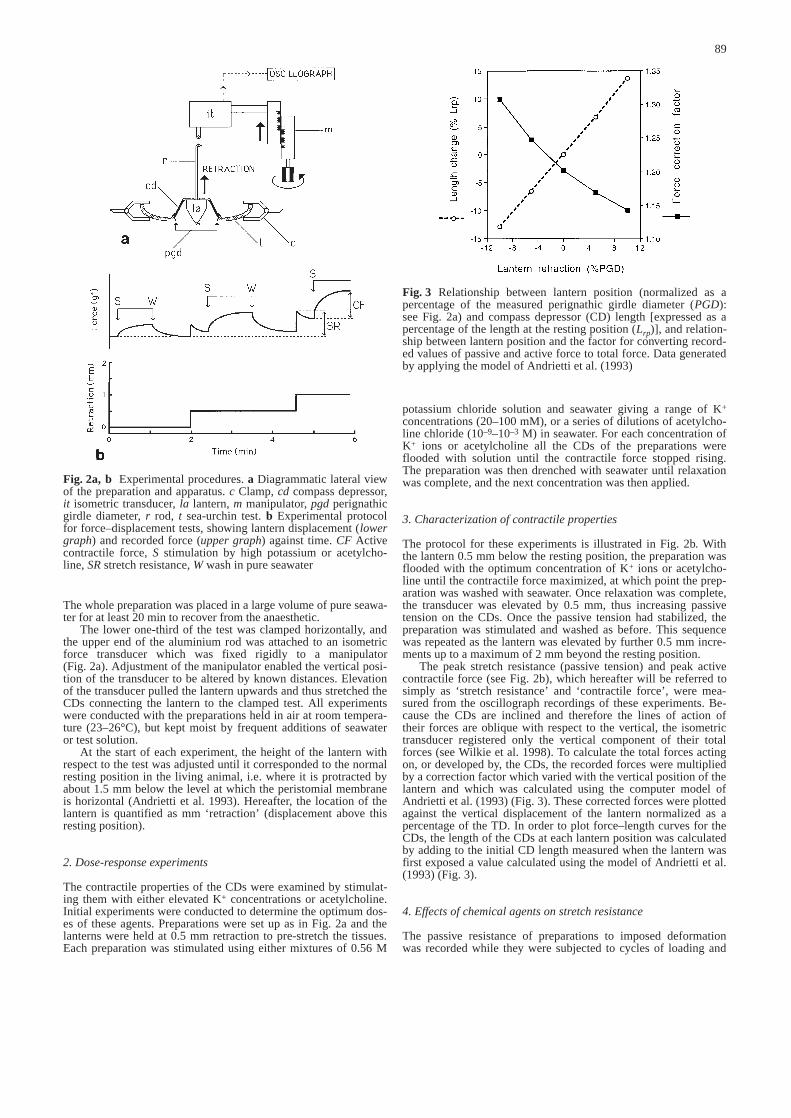
The peak stretch resistance (passive tension) and peak activecontractile force (see Fig. 2b), which hereafter will be referred tosimply as ‘stretch resistance’ and ‘contractile force’, were mea-sured from the oscillograph recordings of these experiments. Be-cause the CDs are inclined and therefore the lines of action oftheir forces are oblique with respect to the vertical, the isometrictransducer registered only the vertical component of their totalforces (see Wilkie et al. 1998). To calculate the total forces actingon, or developed by, the CDs, the recorded forces were multipliedby a correction factor which varied with the vertical position of thelantern and which was calculated using the computer model ofAndrietti et al. (1993) (Fig. 3). These corrected forces were plottedagainst the vertical displacement of the lantern normalized as apercentage of the TD. In order to plot force–length curves for theCDs, the length of the CDs at each lantern position was calculatedby adding to the initial CD length measured when the lantern wasfirst exposed a value calculated using the model of Andrietti et al.(1993) (Fig. 3).
4. Effects of chemical agents on stretch resistance
The passive resistance of preparations to imposed deformationwas recorded while they were subjected to cycles of loading and
89
Fig. 2a, b Experimental procedures. a Diagrammatic lateral viewof the preparation and apparatus. c Clamp, cd compass depressor,it isometric transducer, la lantern, m manipulator, pgd perignathicgirdle diameter, r rod, t sea-urchin test. b Experimental protocolfor force–displacement tests, showing lantern displacement (lowergraph) and recorded force (upper graph) against time. CF Activecontractile force, S stimulation by high potassium or acetylcho-line, SRstretch resistance, W wash in pure seawater&/fig.c:
Fig. 3 Relationship between lantern position (normalized as apercentage of the measured perignathic girdle diameter (PGD):see Fig. 2a) and compass depressor (CD) length [expressed as apercentage of the length at the resting position (Lrp)], and relation-ship between lantern position and the factor for converting record-ed values of passive and active force to total force. Data generatedby applying the model of Andrietti et al. (1993)&/fig.c:

unloading. The transducer was raised 0.5 mm every 30 s until thedesired maximum deformation had been attained, and then low-ered back to the starting position in 0.5 mm/30 s steps. This cyclewas repeated after the preparation had been treated with seawatercontaining either 100 mM K+ or 10–3 M acetylcholine, and thenagain after the preparation had been washed with pure seawater.
C. Results
I. Gross structure
The gross structure of the whole lantern of S. affinishasbeen described by Candia Carnevali et al. (1995). TheCDs connect the compass ossicles to the edge of the testat its junction with the peristomial membrane. Two CDsdiverge from the bilobed outer tip of each compass andrun diagonally to the central notch of two adjacentapophyses, the skeletal projections of the perignathic gir-dle to which the retractor and protractor muscles are alsoattached (Fig. 1). Near each apophysis, the two CDsfrom adjacent compasses fuse together. The exterior coe-lomic septum separates the whole ‘lantern complex’ (i.e.the lantern itself, retractor and protractor muscles, peri-stomial membrane and compass system) from the mainbody cavity of the sea-urchin, the perivisceral coelom(Fig. 1b). This septum forms the outer wall of a set of tenfluid-filled compartments, each of which encloses oneCD (see Stauber 1993; Candia Carnevali et al. 1995 forfull descriptions), and is itself contractile. Its organiza-tion and mechanical behaviour are the subject of a sepa-rate paper (in preparation). The CD is attached to theseptum along its full length by a narrow sheet of tissue(referred to hereafter as the ‘attachment sheet’), and con-sists of a core of longitudinally disposed myocytes andconnective tissue fibres covered by a coelomic epitheli-um (‘coelothelium’; Fig. 4).
II. Ultrastructure
1. Coelothelium
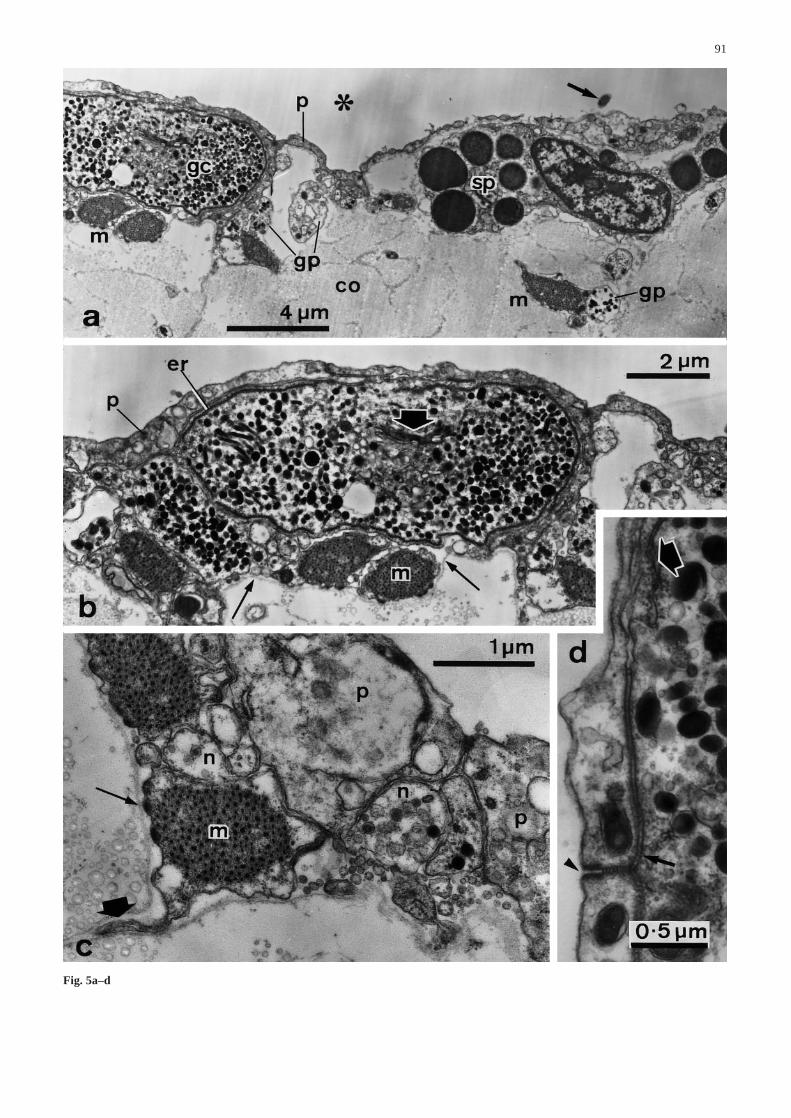
The outermost layer of the CD is a coelothelium which iscontinuous with that of the attachment sheet and with theinner coelothelium of the exterior coelomic septum. Thecoelothelium is a bipartite pseudostratified myoepitheliumdominated by apical peritoneocytes and subapical myo-cytes. The peritoneocytes have T-shaped profiles and ex-tend from the lumen, where their lateral extensions form athin pavement overlying other cell types, to the basal lami-na. The peritoneocytes are connected to each other byjunctional complexes consisting of apical zonulae adhae-rentes and septate belt junctions and to the basal lamina byhemidesmosomes. They are monociliated and their cyto-plasm contains mitochondria, a small amount of rough en-doplasmic reticulum and a variety of small vesicles withcontents of medium to low electron density (Fig. 5a–d).
The myocytes of the coelothelium do not form a con-tinuous layer. They are scattered very sparsely next to
the basal lamina to which they are attached by hemides-mosomes (Fig. 5a–c). In transverse sections of the CDthey are seen to have a well-defined contractile apparatusof regularly arranged thick and thin filaments. The sur-face of the myocytes is formed into longitudinal ribs. Atthe basal side these can be attenuated into narrow sheetswhich, with the basal lamina, penetrate between fibres ofthe underlying connective tissue (Fig. 5c). No special-ized junctions between the myocytes and any other celltype were found.
The coelothelium encloses neural elements. Small di-ameter axon-like processes containing spherical vesicleswith a diameter of up to 140 nm and with a core of lightto medium electron density form a basiepithelial plexus
90
Fig. 4 Semithin transverse section of the upper region of a CD(cd) and adjacent coelomic septa. At this vertical level the coelo-mic cavity of the CD (asterisk) is very narrow. Fat arrowscoelo-thelium, thin arrow attachment sheet, ecsexterior coelomic sep-tum, m myocyte, pccperivisceral coelomic cavity&/fig.c:
Fig. 5a–d Ultrastructure of the coelothelium. a Low magnifica-tion view of the coelothelium and edge of the core (transverse sec-tion). Arrow Cilium of a peritoneocyte, asteriskcoelomic cavity ofCD, co collagen fibrils of core, gc large granule-containing cellbody, gp large granule-containing cell process, p peritoneocyte, spspherulocyte. b Higher magnification view of large granule-con-taining cell body in a. Thin arrowsbasal lamina, fat arrow Golgiapparatus, er subplasmalemmal rough endoplasmic reticulum. cPeritoneocytes (p), neural processes containing dense core vesicles(n) and myocyte (m). Note the prominent projections on the rightsideof the last cell, interpreted as longitudinal ribs. Myocytes areattached to the basal lamina by hemidesmosomes (thin arrow) andhave processes which run into the core (fat arrow). d Junctionalcomplex (arrowhead) between adjacent peritoneocytes which arelinked to the underlying large granule-containing cell by septatejunctions (thin arrow). Part of the subplasmalemmal endoplasmicreticulum in the latter cell is also visible (fat arrow) &/fig.c:
▲
91
Fig. 5a–d
92
Fig. 6a–f

(Fig. 5c). There are also cell bodies and processes con-taining granules of medium to high electron density, cir-cular to elongate-ellipsoid profile and largest dimensionup to 380 nm (Fig. 5a,b,d). The perikarya of the lattercells contain a prominent Golgi apparatus from whichthe granules are derived (Fig. 6a) and are characterizedby the presence of a single subplasmalemmal cisterna ofrough endoplasmic reticulum which can almost encirclethe rest of the cell contents (Fig. 5b). These cells can beattached to overlying peritoneocytes by septate junctions(Fig. 5d). Semithin sections showed that each has a sin-gle prominent process which passes into the CD core andcan often be seen to contact myocyte profiles.
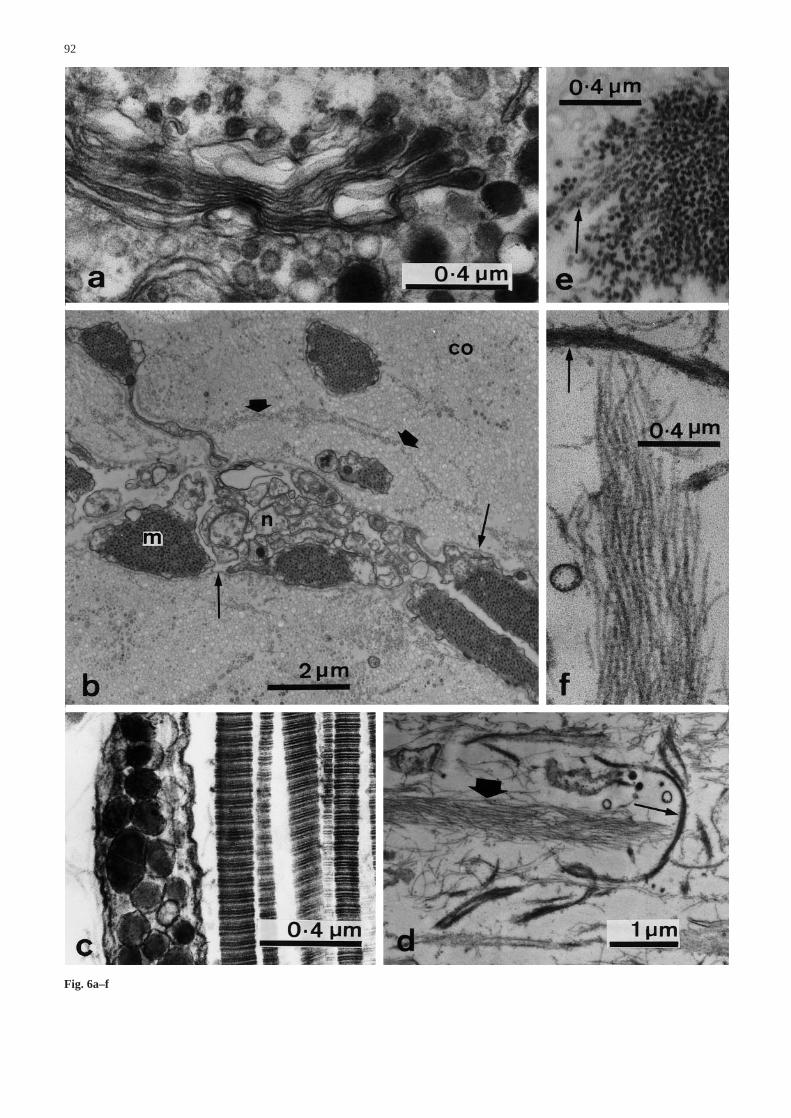
2. Core
The subcoelothelial core of the CD consists mainly ofcross-striated collagen fibrils with a periodicity ofaround 60 nm and a maximum diameter of 280 nm(Fig. 6b,c). The fibrils are mostly parallel to the longitu-dinal axis of the CD and grouped in bundles (i.e. ‘fi-bres’) which are delimited and interconnected by loosenetworks of hollow, beaded microfibrils with a diameterof around 15 nm and with a periodicity of about 20 nm(Fig. 6b,d,e). Near the insertion into the perignathic gir-dle, the microfibrils are much more abundant and aggre-gate to form discrete, longitudinal fibres which are inter-woven with other fibres of basal lamina-like material(Fig. 6d,f).
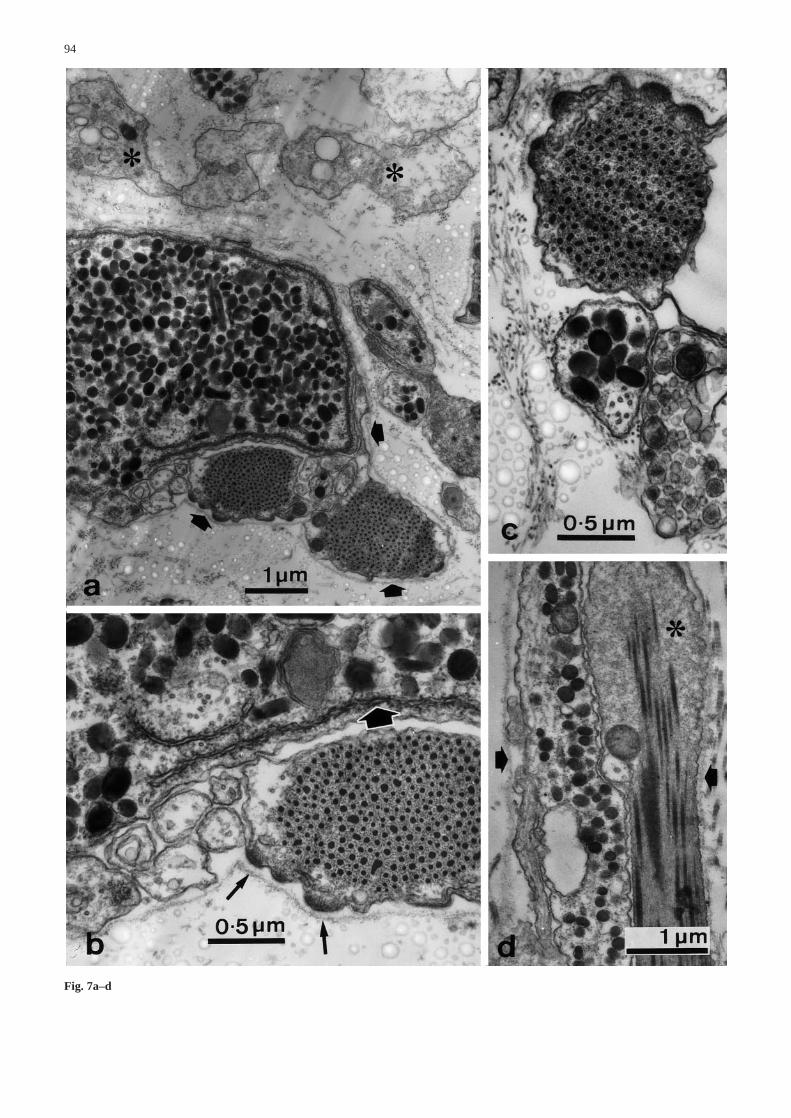
The dominant cells of the core are extremely elongat-ed myocytes which lie parallel to the collagen fibres(Fig. 6b). Myocytes are scattered throughout the core,but there is an outer band below the coelothelium wherethey are sparser than in more central parts. Their generaldistribution is irregular but clustered. In transverse sec-tions, myocyte profiles often form linear arrays betweenlarger bundles of collagen fibrils, although isolated myo-cytes can also be found inside such bundles (Fig. 6b).They are always separated from the surrounding extra-cellular matrix by a basal lamina. They are attached tothe latter by abundant hemidesmosomes (Fig. 7a,b) and
to each other by spot desmosomes (not illustrated).Whilst their general cytology is identical to that of thecoelothelial myocytes, their normally cylindrical shapeshows occasional dilatations which lack thick filamentsbut may include mitochondria and/or electron-lucentvesicles (Fig. 7d). The myocytes may possess narrow,sometimes very elongated, extensions which run towardsor into aggregations of neural elements (Fig. 6b).
The basal lamina surrounding each myocyte profileusually encloses one or more neural elements of thesame two types observed in the coelothelium, i.e. con-taining either small dense-core vesicles or large electron-dense granules (Figs. 6b, 7a,c,d). Of 114 myocyte pro-files which were observed, 70.2% were in direct contactwith at least one such neural element. Profiles of gran-ule-containing cell processes that are not near, or in con-tact with, myocytes are very rare and we were able toidentify with certainty very few granule-containing pro-files which were not surrounded by a basal lamina. Gran-ule-containing cell bodies with their prominent sub-plasmalemmal RER cisternae are also present and are al-ways surrounded by a basal lamina (Fig. 7a,b). Near thejunction with the perignathic girdle there are many vari-cose cell processes containing large, electron-densegranules (Fig. 8a).
Two other cell types were seen in the core: the firstcontains many vesicles which are heterogeneous in size,contents and electron density (Fig. 7a), and the secondare ‘spherulocytes’ which have very large, spherical, uni-formly electron dense granules around 2 µm in diameterand which are often close to the coelothelium (Fig. 5a).Using the point-counting method, it was calculated thatmyocytes occupy 12.8±3.85% of the total cross-sectionalarea of the CD (coelothelium and core).
3. Attachment sheet
The attachment sheet consists of a layer of connectivetissue sandwiched between two coelothelia. The coelo-thelia resemble that of the CD proper except that myo-cytes and large granule-containing cells (LGCs) or pro-cesses are extremely rare (Fig. 8b–d). The central layerof connective tissue is a dense aggregation of collagen fi-brils which are parallel to the longitudinal axis of theCD. These do not appear to be grouped into fibres sur-
93
Fig. 6a–f Ultrastructure of the coelothelium and core. a Coelothe-lium. Higher magnification view of the large granule-containingcell body in Fig. 5b, showing the elaboration of electron-densegranules by the Golgi apparatus. b–f Core. b Low magnificationview (transverse section), showing cross-sections of collagen fi-brils (co) forming bundles surrounded by microfibrils (fat arrows),myocytes (m) and neural elements (n). The cellular elements areenclosed within basal laminae (thin arrows). Note the long processextending from the myocyte at the top left. c Longitudinal sectionshowing collagen fibrils and a cell process containing large gran-ules and microtubules. d Longitudinal section from region near theperignathic girdle, where beaded microfibrils are aggregated intofibres (fat arrow) which are interwoven with other fibres that con-sist of basal lamina-like material (thin arrow). e Transverse sec-tion of a bundle of microfibrils, illustrating their hollow appear-ance (arrow). f High magnification view of part of d illustratingthe beaded appearance of the microfibrils and the basal lamina-like appearance of the other fibre type (arrow) at this location&/fig.c:
Fig. 7a–d Ultrastructure of the core. a–c Transverse sections. aAggregations of myocytes and large granule-containing cell bod-ies and processes are enclosed within basal laminae (arrows). Oth-er cells contain heterogeneous vacuoles (asterisks). b High magni-fication view of part of a. The large granule-containing cell bodyhas a prominent subplasmalemmal vesicle of rough endoplasmicreticulum (fat arrow). Myocytes are attached to the basal laminaby hemidesmosomes (thin arrows). c Myocytes (top) are usuallyin close contact with large granule-containing cell processes (mid-dle) and other processes containing dense-core vesicles (bottom).d Longitudinal section showing a myocyte with a dilated region(asterisk) and a large granule-containing process, the two cells be-ing enclosed within a basal lamina (arrows) &/fig.c:
▲▲
94
Fig. 7a–d
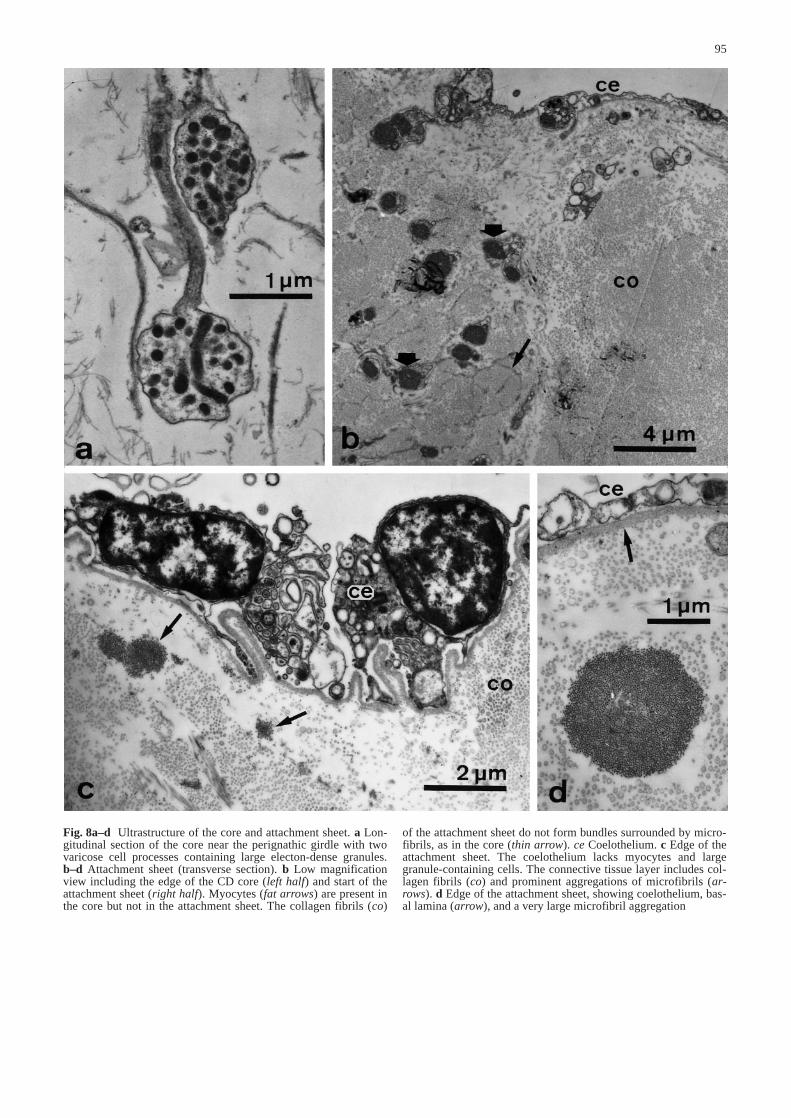
95
Fig. 8a–d Ultrastructure of the core and attachment sheet. a Lon-gitudinal section of the core near the perignathic girdle with twovaricose cell processes containing large electon-dense granules.b–d Attachment sheet (transverse section). b Low magnificationview including the edge of the CD core (left half) and start of theattachment sheet (right half). Myocytes (fat arrows) are present inthe core but not in the attachment sheet. The collagen fibrils (co)
of the attachment sheet do not form bundles surrounded by micro-fibrils, as in the core (thin arrow). ce Coelothelium. c Edge of theattachment sheet. The coelothelium lacks myocytes and largegranule-containing cells. The connective tissue layer includes col-lagen fibrils (co) and prominent aggregations of microfibrils (ar-rows). d Edge of the attachment sheet, showing coelothelium, bas-al lamina (arrow), and a very large microfibril aggregation&/fig.c:
rounded by microfibrillar networks as in the CD core(Fig. 8b), although microfibrils occur in small groupsscattered amongst the collagen fibrils and in very large,longitudinally aligned aggregations which can be over2 µm in diameter (Fig. 8c,d). No cellular elements werefound in the connective tissue layer.
III. Mechanical behaviour
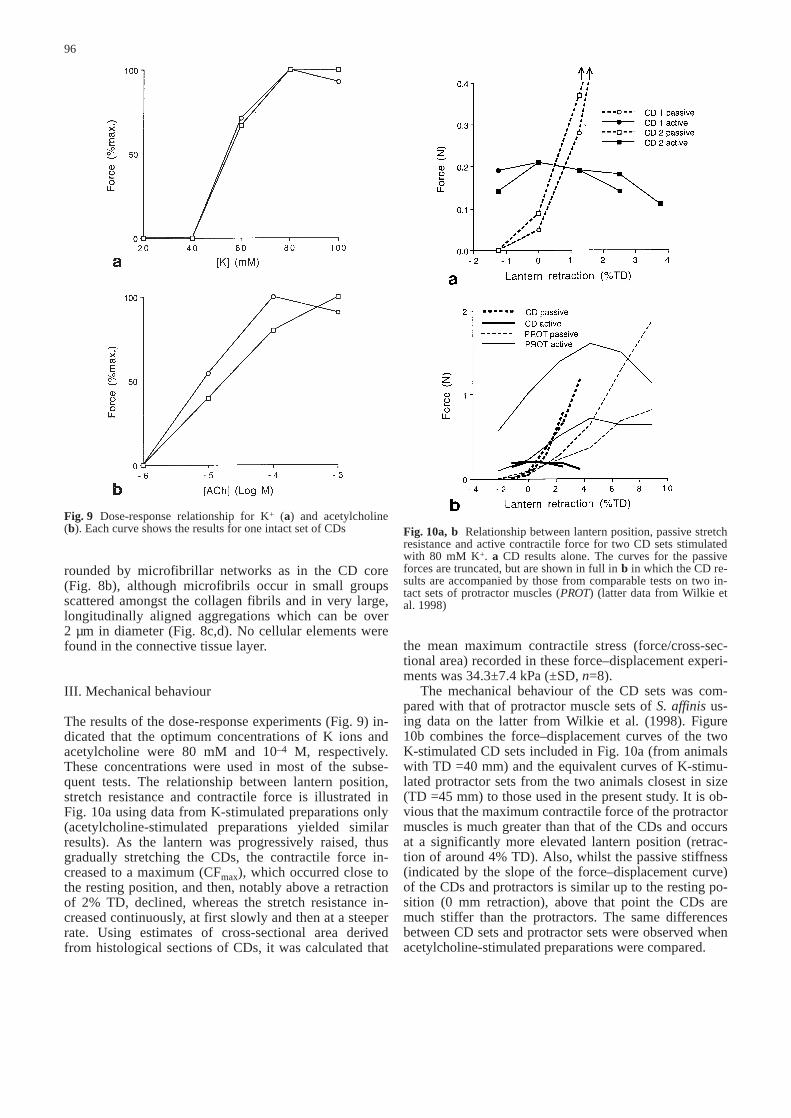
The results of the dose-response experiments (Fig. 9) in-dicated that the optimum concentrations of K ions andacetylcholine were 80 mM and 10–4 M, respectively.These concentrations were used in most of the subse-quent tests. The relationship between lantern position,stretch resistance and contractile force is illustrated inFig. 10a using data from K-stimulated preparations only(acetylcholine-stimulated preparations yielded similarresults). As the lantern was progressively raised, thusgradually stretching the CDs, the contractile force in-creased to a maximum (CFmax), which occurred close tothe resting position, and then, notably above a retractionof 2% TD, declined, whereas the stretch resistance in-creased continuously, at first slowly and then at a steeperrate. Using estimates of cross-sectional area derivedfrom histological sections of CDs, it was calculated that
the mean maximum contractile stress (force/cross-sec-tional area) recorded in these force–displacement experi-ments was 34.3±7.4 kPa (±SD, n=8).
The mechanical behaviour of the CD sets was com-pared with that of protractor muscle sets of S. affinisus-ing data on the latter from Wilkie et al. (1998). Figure10b combines the force–displacement curves of the twoK-stimulated CD sets included in Fig. 10a (from animalswith TD =40 mm) and the equivalent curves of K-stimu-lated protractor sets from the two animals closest in size(TD =45 mm) to those used in the present study. It is ob-vious that the maximum contractile force of the protractormuscles is much greater than that of the CDs and occursat a significantly more elevated lantern position (retrac-tion of around 4% TD). Also, whilst the passive stiffness(indicated by the slope of the force–displacement curve)of the CDs and protractors is similar up to the resting po-sition (0 mm retraction), above that point the CDs aremuch stiffer than the protractors. The same differencesbetween CD sets and protractor sets were observed whenacetylcholine-stimulated preparations were compared.
96
Fig. 9 Dose-response relationship for K+ (a) and acetylcholine(b). Each curve shows the results for one intact set of CDs&/fig.c: Fig. 10a, b Relationship between lantern position, passive stretch
resistance and active contractile force for two CD sets stimulatedwith 80 mM K+. a CD results alone. The curves for the passiveforces are truncated, but are shown in full in b in which the CD re-sults are accompanied by those from comparable tests on two in-tact sets of protractor muscles (PROT) (latter data from Wilkie etal. 1998)&/fig.c:
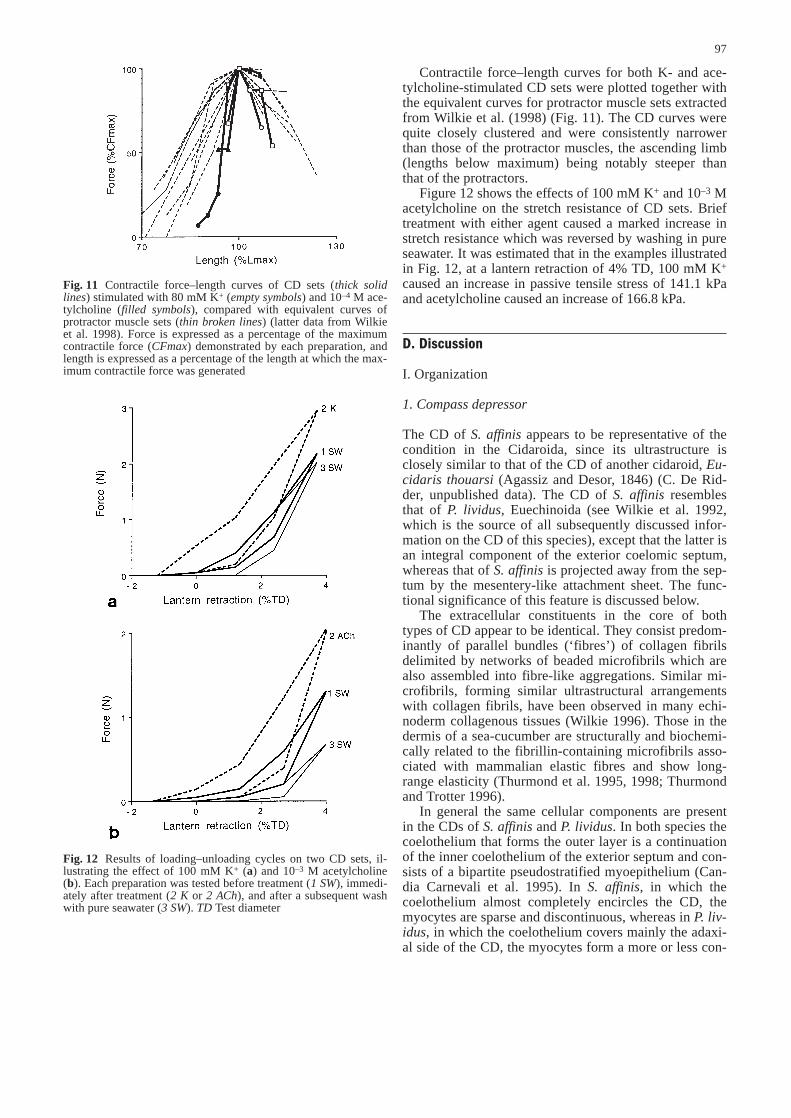
Contractile force–length curves for both K- and ace-tylcholine-stimulated CD sets were plotted together withthe equivalent curves for protractor muscle sets extractedfrom Wilkie et al. (1998) (Fig. 11). The CD curves werequite closely clustered and were consistently narrowerthan those of the protractor muscles, the ascending limb(lengths below maximum) being notably steeper thanthat of the protractors.
Figure 12 shows the effects of 100 mM K+ and 10–3 Macetylcholine on the stretch resistance of CD sets. Brieftreatment with either agent caused a marked increase instretch resistance which was reversed by washing in pureseawater. It was estimated that in the examples illustratedin Fig. 12, at a lantern retraction of 4% TD, 100 mM K+
caused an increase in passive tensile stress of 141.1 kPaand acetylcholine caused an increase of 166.8 kPa.
D. Discussion
I. Organization
1. Compass depressor
The CD of S. affinisappears to be representative of thecondition in the Cidaroida, since its ultrastructure isclosely similar to that of the CD of another cidaroid, Eu-cidaris thouarsi(Agassiz and Desor, 1846) (C. De Rid-der, unpublished data). The CD of S. affinisresemblesthat of P. lividus, Euechinoida (see Wilkie et al. 1992,which is the source of all subsequently discussed infor-mation on the CD of this species), except that the latter isan integral component of the exterior coelomic septum,whereas that of S. affinisis projected away from the sep-tum by the mesentery-like attachment sheet. The func-tional significance of this feature is discussed below.
The extracellular constituents in the core of bothtypes of CD appear to be identical. They consist predom-inantly of parallel bundles (‘fibres’) of collagen fibrilsdelimited by networks of beaded microfibrils which arealso assembled into fibre-like aggregations. Similar mi-crofibrils, forming similar ultrastructural arrangementswith collagen fibrils, have been observed in many echi-noderm collagenous tissues (Wilkie 1996). Those in thedermis of a sea-cucumber are structurally and biochemi-cally related to the fibrillin-containing microfibrils asso-ciated with mammalian elastic fibres and show long-range elasticity (Thurmond et al. 1995, 1998; Thurmondand Trotter 1996).
In general the same cellular components are presentin the CDs of S. affinisand P. lividus. In both species thecoelothelium that forms the outer layer is a continuationof the inner coelothelium of the exterior septum and con-sists of a bipartite pseudostratified myoepithelium (Can-dia Carnevali et al. 1995). In S. affinis, in which thecoelothelium almost completely encircles the CD, themyocytes are sparse and discontinuous, whereas in P. liv-idus, in which the coelothelium covers mainly the adaxi-al side of the CD, the myocytes form a more or less con-
97
Fig. 11 Contractile force–length curves of CD sets (thick solidlines) stimulated with 80 mM K+ (empty symbols) and 10–4 M ace-tylcholine (filled symbols), compared with equivalent curves ofprotractor muscle sets (thin broken lines) (latter data from Wilkieet al. 1998). Force is expressed as a percentage of the maximumcontractile force (CFmax) demonstrated by each preparation, andlength is expressed as a percentage of the length at which the max-imum contractile force was generated&/fig.c:
Fig. 12 Results of loading–unloading cycles on two CD sets, il-lustrating the effect of 100 mM K+ (a) and 10–3 M acetylcholine(b). Each preparation was tested before treatment (1 SW), immedi-ately after treatment (2 K or 2 ACh), and after a subsequent washwith pure seawater (3 SW). TD Test diameter&/fig.c:

tinuous layer one to a few cells thick. The coelothelia ofboth species include axon-like processes with dense-corevesicles of the same size and cells containing large gran-ules with circular to ovoid profiles 370–380 nm long.
The CD core of S. affiniscontains many myocytes,whereas none are present in that of P. lividus. CandiaCarnevali et al. (1995) argued that the myocytes in thecore of S. affinisare derived from the coelothelium, sincethey are always separated from the surrounding connec-tive tissue by a basal lamina, and this is confirmed by thepresent observation that they are usually accompanied bythe same two granule-containing cell types as occur inthe coelothelium. The different myocyte distributions inthe CDs of S. affinisand P. lividus imply that the role ofthe CD as an actively contracting muscle, rather than apassive ligament, is more important in S. affinisthan inP. lividus, a view supported by the greater proportion ofmyocytes in the CD of S. affinis(ca 13% of the cross-sectional area) than in that of P. lividus(ca 8%). Anotherfunctional consequence may arise from the fact that CDshortening necessitates the compression of the connec-tive tissue component. Evidence from other echinodermmyocyte–ligament composites, principally the sea-urchincatch apparatus (Hidaka and Takahashi 1983; Smith etal. 1981; Perez-Acevedo et al. 1998), indicates that sig-nificant changes in length are achieved in vivo throughslippage between collagen fibrils or fibril bundles ratherthan by the crimping and decrimping of fibres which al-lows only limited length changes in mammalian tendonand ligament (Frank and Shrive 1994; Herzog and Loitz1994). If it is assumed that the sea-urchin CD conformsto the echinoderm norm, it is plausible that the interfibreshear that accompanies shortening of the CD can be ac-complished more efficiently by a dispersed myocyte ar-rangement than by a concentration of myocytes at oneside of the ligament.
In S. affinis, cells containing large granules are codis-tributed with myocytes in the CD coelothelium and core,whereas both cell types are absent from the coelotheliumand connective tissue layer of the attachment sheet. Iden-tical LGCs are present in the CD myoepithelium of P. liv-idus, and LGC–myocyte associations are common in thecoelothelia of echinoderm tube-feet (e.g. Rieger andLombardi 1987), gonads (e.g. Byrne 1989) and digestivetracts (e.g. Chia and Koss 1994). Cobb (1987) noted themorphological similarity between LGCs and the ‘juxta-ligamental cells’ that have been found in all investigatedexamples of echinoderm mutable collagenous tissues andare likely to be the effector cells modulating directly thepassive mechanical properties of the surrounding extra-cellular matrix (Trotter and Koob 1995; Wilkie 1996).Cobb (1987) speculated that coelothelial LGCs mighthave an analogous function in controlling cohesion be-tween myocytes and the extensibility of myocyte bundles.
Though lacking subcoelothelial myocytes, the CDcore of P. lividus is densely permeated by granule-con-taining cell bodies and processes interpreted by Wilkie etal. (1992) as being juxtaligamental cells. Their appear-ance is remarkably similar to that of the myocyte-associ-
ated LGCs in the core of S. affinis(and in the coelotheliaof both species). The cells in the CD core of P. lividusare not, however, separated from the extracellular matrixby a basal lamina and their processes make frequent con-tact with what appear to be cholinergic axons. In the CDcore of S. affinis, only a few granule-containing process-es are located outwith the basal laminae of the myocytecomplexes and there are no putative cholinergic axons.Recent evidence indicates that juxtaligamental cells in-fluence the tensility of connective tissue by secreting ef-fector molecules that interact directly with componentsof the extracellular matrix (Szulgit and Shadwick 1994;Trotter and Koob 1995). The separation of myocyte-as-sociated LGCs from the extracellular matrix by basallaminae makes it unlikely that these cells serve the samefunction.
2. Attachment sheet
Since neither myocytes nor any type of granule-contain-ing cell are present in the connective tissue layer of theattachment sheet and both cell types are very rare in itscoelothelia, it is highly improbable that this structure isactively contractile or that its connective tissue is mut-able. Moreover, the connective tissue layer of the attach-ment sheet differs from the extracellular compartment ofthe CD core in that its densely packed collagen fibrils arenot organized into bundles delimited by microfibrillarnetworks. If, as discussed above, the latter organizationis the morphological correlate of tissue extensibility andcompressibility, its absence from the connective tissue ofthe attachment sheet implies that this structure is inex-tensible and that under compressive forces it will shortenby buckling rather than by interfibre shear. If this is thecase, then the role of the prominent, longitudinallyaligned (and probably elastic) microfibril aggregationsmay be to promote unfolding and straightening of the at-tachment sheet after it has been distorted by contractionof the CD itself.
The presence of the attachment sheet in S. affinisbutnot P. lividus is probably correlated with the structure ofthe external coelomic septa. In P. lividusand related Euec-hinoida the latter are very thin, delicate membraneswhich are unlikely to interfere with the mechanical func-tioning of the CDs. In cidaroids they are much more sub-stantial structures with a thick central layer of connectivetissue reinforced by calcite spicules (I.C. Wilkie et al.unpublished data). The attachment sheet may allow thecidaroid CDs to shorten and re-extend independently of,and unhindered by, the coelomic septa.
II. Mechanical behaviour
1. Conventional mechanical behaviour
As in typical muscles, when the CDs of S. affinisaregradually elongated, their contractile force increases to
98

a maximum and then decreases. It is widely recognizedthat the mechanical properties of muscles, includingtheir force–length curves, are optimized to match theirnormal working conditions (Herzog 1994). In this inves-tigation the behaviour of the CDs was compared withthat of the lantern protractor muscles of S. affinis. Theforce–displacement experiments showed that the CDsdevelop a maximum contractile force when the lanternis around the resting position, whereas the protractorsdevelop maximum contractile force when the lantern isretracted above the resting position by around 4% TD.In addition, the force–length curves of the CDs are nar-rower than those of the protractors, which themselvesare close to the curves of striated muscles such as thefrog sartorius and crab scaphognathite elevator (Wilkieet al. 1998). The CDs of S. affinis thus appear to beadapted to contract most forcefully when their startinglength is at or close to their resting length, i.e. the lengththey take up when the compasses are fully depressedand the lantern is in the resting position. Moreover, thepassive stretch resistance of the CDs increases greatlywhen they are stretched beyond their resting length. TheJ-shaped passive force–length curves of the CD resem-ble those of other collagenous structures in which theinflection between the low resistance and high resis-tance phases occurs at around the maximum length towhich the structure is stretched in vivo (Frank andShrive 1994; Herzog and Loitz 1994). It seems then thatthe connective tissue component of the CD is designedto resist stretching beyond the resting length, the opti-mal length for force development by its contractile com-ponent.
This has interesting implications for the relationshipbetween lantern position and the functioning of the com-pass system in vivo. The two physiological roles thathave been attributed to the compass system depend onthe compass ossicles being able to swing up and down asa result of the reciprocal contraction of the compass ele-vator muscles and compass depressors. Von Uexküll(1896) proposed that the system is a respiratory pumpwhich by raising and lowering the compasses circulatesfluid through the peripharyngeal coelomic pouches andboosts the oxygenation of the lantern muscles. Kier(1974) argued that it is a pressure-compensating device:as the lantern is protracted, a pressure drop within thetest is prevented by the raising of the compasses whichexpands the peripharyngeal pouches by ‘sucking’ in flu-id from external peristomial ‘gills’ (present only in someeuechinoids), the opposite happening when the lantern isretracted again. For either of these physiological roles tobe fulfilled, the lantern must be at a vertical level thatpermits compass elevation and depression. When the lan-tern of S. affinisis at the resting position, the CDs are attheir resting length and compass elevation will be pre-vented by the stretch resistance of the CDs. This impliesthat compass elevation takes place only when the lanternis protracted below the resting position. However, lanternprotraction in S. affinis is constrained by the unfavour-able working length of the protractor muscles (Andrietti
et al. 1993) and by the stretch resistance of the peristo-mial membrane whose passive force–displacement curveshows an inflection between low and high resistancephases at a protraction of at most 3% TD (equivalent tothe maximum value of 6% perignathic girdle diametercited by Wilkie et al. 1996). Accordingly, the range ofvertical movement permitted to the compasses is also atmost 3% TD, which in an animal with a test diameter of40 mm would be only 1.2 mm. The restricted range ofvertical movement permissable to the compasses of S. af-finis seems more compatible with its compass system be-ing a respiratory pump, the efficiency of which dependson both the amplitude of the compass oscillations andtheir frequency, rather than a pressure-compensator, forwhich there would seem to be little need when the wholelantern has such limited vertical mobility. The case forthe compass system in the Cidaroida being first and fore-most a respiratory pump is strengthened by the presenceof Stewart’s organs (see Fig. 1b) which are outgrowths ofthe peripharyngeal pouches. These have a lobate lowerwall which lies above, and in some cidaroid speciesagainst, the tube-foot ampullae and has an ultrastructureconsistent with its being adapted for gaseous exchange(De Ridder 1988).
In terms of the range of vertical, lateral and tiltingmovements of which it is capable, the lantern of P. liv-idus is considerably more mobile than that of S. affinis(Andrietti et al. 1993). This is matched by the behaviourof the compass system. For example, direct observationof the intact lantern of P. lividus at the resting positionrevealed that compass elevation can elongate the CDs byover 40% (Wilkie et al. 1992), whereas we argue abovethat the CDs of S. affinisare unlikely to be stretched be-yond their resting length.
2. Variable tensility
A notable feature of the CDs of P. lividusand other Euec-hinoida is that the core consists of mutable collagenoustissue which undergoes reversible, nervously mediatedchanges in stiffness. Wilkie et al. (1992) provided evi-dence for this by showing that treatment with either ace-tylcholine or a high K+ concentration causes an increasein stretch resistance too great to be due to contraction ofthe myocytes alone. The same treatments increased thestretch resistance of S. affinisCDs. In the illustrated ex-amples (Fig. 12), high K+ and acetylcholine caused anincrease in passive tensile stress of 141.1 kPa and166.8 kPa, respectively. Myocytes occupy 12.8% of thecross-sectional area of the CDs of this species. If it is as-sumed that the myocytes alone were responsible forthese changes in passive stress, i.e. by developing an iso-metric tension that resisted the imposed stretch, theywould have to have generated a contractile stress of1.1 MPa and 1.3 MPa, respectively. However, this is un-likely, because the maximum contractile stress producedby whole CDs in the dose-response tests and force–dis-placement experiments was estimated to be around
99

34 kPa, which implies that the myocytes themselves cangenerate a maximum stress of only 0.27 MPa.
Whilst this is evidence that connective tissue stiffen-ing contributes to the response induced by high K+ oracetylcholine, the CDs of S. affinisshow much smallerchanges in stretch resistance than those of P. lividus: Wi-lkie et al. (1992) found that the same agents increasedthe stretch resistance of the latter by up to 200%, where-as the maximum increase recorded in the CDs of S. affi-nis was 56%. This finding is complemented by data onthe physicochemical properties of CDs from a range ofsea-urchins (I.C. Wilkie et al. unpublished data) whichindicate that the CDs of S. affinis, in comparison withthose of P. lividus and three other Euechinoida species,express only weakly the response to distilled water thatis characteristic of mutable collagenous tissues (see e.g.Trotter and Koob 1995; Wilkie et al. 1995). The observa-tion that almost all the LGCs in the CD core are associat-ed with myocytes and very few, if any, ‘target’ the extra-cellular matrix (i.e. could be juxtaligamental cell equiva-lents) matches these other indications that the CD corehas a limited capacity for variable tensility. This is notunexpected, since the peristomial membrane of S. affinisprovides little evidence for the presence of mutable col-lagenous tissue (Wilkie et al. 1996).
III. Conclusions
The CDs of S. affiniscombine the function of inextensi-ble ligaments, designed to resist stretching beyond theirlength when the lantern and compass system are at theresting position, with that of muscles which contract mostforcefully at or close to this same length. The disperseddistribution and greater abundance of their myocytes sug-gests that the contractile function of these CDs is moreimportant than in euechinoid CDs. The significance ofthis is not clear at present. It could be related to the con-jectured role of the cidaroid compass system as a respira-tory pump in which the CDs and compass elevator mus-cles bring about low amplitude, but perhaps rapid, rhyth-mical movements of the compasses while the lantern isslightly protracted. In contrast to that of euechinoid CDs,the connective tissue in the CDs of S. affinisshows a lim-ited capacity for variable tensility. In the CDs of Cidaro-ida connective tissue mutability may be largely redundantbecause of the restricted mobility of the lantern and be-cause lantern stabilization in these sea-urchins, which inEuechinoida depends partly on reversible stiffening of theCDs and peristomial membrane (Andrietti et al. 1990), isan important function of the retractor and protractor mus-cles (Andrietti et al. 1993; Wilkie et al. 1998).
&p.2:Acknowledgements We are grateful to Dr. C. De Ridder, Univer-sité Libre de Bruxelles, for providing unpublished electron micro-graphs of Eucidaris thouarsi. This project received financial sup-port from the Consiglio Nazionale delle Ricerche, Rome, the Roy-al Society, London, Glasgow Caledonian University and the Car-negie Trust for the Universities of Scotland.
References
Andrietti F, Candia Carnevali MD, Wilkie IC, Lanzavecchia G,Melone G, Celentano FC (1990) Mechanical analysis of thesea-urchin lantern: the overall system in Paracentrotus lividus.J Zool Lond 220:345–366
Andrietti F, Candia Carnevali MD, Wilkie IC (1993) A biome-chanical comparison of the lantern of the cidarid sea-urchinStylocidaris affiniswith the typical camarodont lantern. J ZoolLond 231:595–610
Byrne M (1989) Ultrastructure of the ovary and oogenesis in theovoviviparous ophiuroid Ophiolepis paucispina(Echinoder-mata). Biol Bull 176:79–95
Candia Carnevali MD, Wilkie IC, Lucca E, Andrietti F, Melone G(1993) The Aristotle’s lantern of the sea-urchin Stylocidarisaffinis (Echinoida, Cidaridae): functional morphology of themusculo-skeletal system. Zoomorphology 113:173–189
Candia Carnevali, MD, Bonasoro F, Wilkie IC (1995) Coelom and“tinkering” in echinoids: morpho-functional adaptations of thelantern coelom. In: Lanzavecchia G, Valvassori R, CandiaCarnevali MD (eds) Body cavities: function and phylogeny.Mucchi, Modena, pp 135–165
Chia FS, Koss R (1994) Asteroidea. In: Harrison FW, Chia FS(eds) Microscopic anatomy of invertebrates, vol 14: Echino-dermata. Wiley-Liss, New York, pp 169–245
Cobb JLS (1987) Neurobiology of the Echinodermata. In: Ali MA(ed) Nervous systems in invertebrates. Plenum Press, NewYork, pp 483–525
Davison IG, Wright GM, De Mont ME (1995) The structure andphysical properties of invertebrate and primitive vertebrate ar-teries. J Exp Biol 198:2185–2196
De Ridder C (1988) Could the Stewart’s organs of cidaroidechinoids be internal gills? In: Burke RD, Mladenov PV, Lam-bert P, Parsley RL (eds) Echinoderm biology. Balkema, Rotter-dam, pp 675–681
Eylers JP (1976) Aspects of skeletal mechanics of the starfish As-terias forbesii. J Morphol 149:353–368
Frank CB, Shrive NG (1994) Ligament. In: Nigg BM, Herzog W(eds) Biomechanics of the musculo-skeletal system. Wiley,Chichester, pp 106–131
Herzog W (1994) Muscle. In: Nigg BM, Herzog W (eds) Bio-mechanics of the musculo-skeletal system. Wiley, Chichester,pp 154–190
Herzog W, Loitz B (1994) Tendon. In: Nigg BM, Herzog W (eds)Biomechanics of the musculo-skeletal system. Wiley, Chiches-ter, pp 154–190
Hidaka M, Takahashi K (1983) Fine structure and mechanicalproperties of the catch apparatus of the sea-urchin spine, a col-lagenous connective tissue with muscle-like holding activity. JExp Biol 103:1–14
Kier PM (1974) Evolutionary trends and their functional signifi-cance in the post-paleozoic echinoids. J Paleontol 48 (suppl 3,Paleontol Soc Mem 5):1–95
Lanzavecchia G, Candia Carnevali MD, Melone G, Celentano FC,Andrietti F (1988) Aristotle’s lantern in the regular sea-urchinParacentrotus lividus.I. Functional morphology and signifi-cance of bones, muscles and ligaments. In: Burke RD, Mlade-nov PV, Lambert P, Parsley RL (eds) Echinoderm biology.Balkema, Rotterdam, pp 649–662
Märkel K (1979) Structure and growth of the cidaroid socket-jointlantern of Aristotle compared to the hinge-joint lanterns ofnon-cidaroid regular echinoids (Echinodermata, Echinoidea).Zoomorphologie 94:1–32
Nishimura T, Hattori A, Takahashi K (1994) Ultrastructure of theintramuscular connective tissue in bovine skeletal muscle. Ademonstration using the cell-maceration/scanning electron mi-croscope method. Acta Anat 151:250–257
Olaechea RP, Ushio H, Watabe S, Takada K, Hatae K (1993)Toughness and collagen content of abalone muscles. BiosciBiotech Biochem 57:6–11
100

Perez-Acevedo NL, Marrero H, Castillo J del, Smith DS (1998)Transient wrinkles in a variable length tendon. In: Mooi R,Telford M (eds) Echinoderms: San Francisco. Proc Ninth IntEchinoderm Conference, San Francisco, California 1996.Balkema, Rotterdam, pp 183–190
Purslow PP (1994) The morphology and mechanical properties ofendomysium in series-fibred muscles: variations with musclelength. J Muscle Res Cell Motil 15:299–308
Rieger RM, Lombardi J (1987) Ultrastructure of coelomic liningin echinoderm podia: significance for concepts in the evolu-tion of muscle and peritoneal cells. Zoomorphology 107:191–208
Saita A (1969) La morfologia ultrastrutturale dei muscoli della“Lanterna di Aristotele” di alcuni echinoidi. Ist Lomb AccadSci Lett Rend Sci B 103:297–313
Shadwick RE, Gosline JM (1985) The role of collagen in the me-chanical design of squid mantle. In: Bairati A, Garrone R (eds)Biology of invertebrate and lower vertebrate collagen. PlenumPress, New York, pp 299–307
Smith DS, Wainwright SA, Baker J, Cayer ML (1981) Structuralfeatures associated with movement and ‘catch’ of sea-urchinspines. Tissue Cell 13:299–320
Stauber M (1993) The lantern of Aristotle: organization of its coe-lom and origin of its muscles (Echinodermata, Echinoida).Zoomorphology 113:137–151
Szulgit GK, Shadwick RE (1994) The effects of calcium chelationand cell perforation on the mechanical properties of sea urchinligaments. In: David B, Guille A, Féral JP, Roux M (eds)Echinoderms through time. Balkema, Rotterdam, pp 887–892
Thurmond FA,Trotter JA (1996) Morphology and biomechanics ofthe microfibrillar network of sea cucumber dermis. J Exp Biol199:1817–1828
Thurmond FA, Koob TJ, Trotter JA (1995) The microfibrils of seacucumber dermis are similar to fibrillin microfibrils and forma network that has long-range elasticity. Mol Biol Cell 6:2210
Thurmond FA, Trotter JA, Koob TJ, Bowness JM (1998) Microfi-brils from sea cucumber dermis belong to the fibrillin family,and their long-range elasticity is a crucial component of mut-
able collagenous tissues. In: Mooi R, Telford M (eds) Echino-derms: San Francisco. Proc Ninth Int Echinoderm Conference,San Francisco, California 1996. Balkema, Rotterdam, p 528
Trotter JA, Koob TJ (1995) Evidence that calcium-dependent cel-lular processes are involved in the stiffening response of ho-lothurian dermis and that dermal cells contain an organic stiff-ening factor. J Exp Biol 198:1951–1961
Uexküll J von (1896) Über die Funktion der Polischen Blasen amKauapparat der regulären Seeigel. Mitt Zool Staz Neapel12:463–476
Wilkie IC (1988) Design for disaster: the ophiuroid intervertebralligament as a typical mutable collagenous structure. In: BurkeRD, Mladenov PV, Lambert P, Parsley RL (eds) Echinodermbiology. Balkema, Rotterdam, pp 25–38
Wilkie IC (1996) Mutable collagenous tissues: extracellular matrixas mechano-effector. In: Jangoux M, Lawrence JM (eds) Echi-noderm studies, vol 5. Balkema, Rotterdam, pp 61–102
Wilkie IC, Candia Carnevali MD, Bonasoro F (1992) The compassdepressors of Paracentrotus lividus(Echinodermata, Echino-ida): ultrastructural and mechanical aspects of their variabletensility and contractility. Zoomorphology 112:143–153
Wilkie IC, Candia Carnevali MD, Andrietti F (1993) Variabletensility of the peristomial membrane of the cidaroid sea-ur-chin Paracentrotus lividus(Lamarck). Comp Biochem Physiol(A) 105:493–501
Wilkie IC, McKew M, Candia Carnevali MD (1995) Anomalousphysico-chemical properties of the compass-rotular ligamentsin two species of sea-urchins: preliminary observations. In:Emson RH, Smith AB, Campbell AC (eds) Echinoderm re-search 1995. Balkema, Rotterdam, pp 147–152
Wilkie IC, Candia Carnevali MD, Andrietti F (1996) Mechanicalproperties of the peristomial membrane of the cidaroid sea-ur-chin Stylocidaris affinis. J Zool Lond 238:557–569
Wilkie IC, Candia Carnevali MD, Andrietti F (1998) Mechanicalproperties of sea-urchin lantern muscles: a comparative inves-tigation of intact muscle groups in Paracentrotus lividus(Lam.) and Stylocidaris affinis (Phil.) (Echinodermata,Echinoidea). J Comp Physiol B (in press)
101





























