Embed Size (px)
Citation preview

Seediscussions,stats,andauthorprofilesforthispublicationat:http://www.researchgate.net/publication/11407952
Paenibacilluspolymyxaproducesfusaricidin‐typeantifungalantibioticsactiveagainstLeptosphaeriamaculans,thecausativeagentofblacklegdiseaseofcanola
ARTICLEinCANADIANJOURNALOFMICROBIOLOGY·FEBRUARY2002
ImpactFactor:1.18·DOI:10.1139/w02-002·Source:PubMed
CITATIONS
84
DOWNLOADS
108
VIEWS
157
2AUTHORS,INCLUDING:
PerrinHudsonBeatty
UniversityofAlberta
34PUBLICATIONS601CITATIONS
SEEPROFILE
Availablefrom:PerrinHudsonBeatty
Retrievedon:31July2015

Paenibacillus polymyxa produces fusaricidin-typeantifungal antibiotics active against Leptosphaeriamaculans, the causative agent of blackleg diseaseof canola
Perrin H. Beatty and Susan E. Jensen
Abstract: A bacterial isolate capable of inhibiting the growth ofLeptosphaeria maculans(Desmaz.) Ces. & De Not.,the causative agent of blackleg disease of canola (Brassica napusL. and Brassica rapaL.), was identified as a poten-tial biological control agent. This environmental isolate was determined to bePaenibacillus polymyxabased on its(i) biochemical and growth characteristics and (ii ) 16S rRNA sequence similarity, and was given the strain designationPKB1. Antifungal peptides were produced byP. polymyxaPKB1 around the onset of sporulation, with optimal produc-tion on potato dextrose broth. The antifungal peptides were extracted fromP. polymyxaPKB1 cells and (or) spores us-ing methanol and were purified using size exclusion and reverse-phase chromatography. Characterization of theantifungal peptides was done using amino acid compositional analysis, Edman degradation sequencing from partiallyhydrolyzed material, and a variety of mass spectrometric methods. The purified antifungal material was found to be amixture of related peptides of molecular masses 883, 897, 948, and 961 Da, with the most likely structure of the 897-Da component determined to be a cyclic depsipeptide with an unusual 15-guanidino-3-hydroxypentadecanoic acid moi-ety bound to a free amino group. These compounds are therefore members of the fusaricidin group of cyclicdepsipeptides.
Key words: cyclic depsipeptide, biocontrol agent, allo-threonine.
Beatty and JensenRésumé: Un isolat bactérien capable d’inhiber la croissance deLeptosphaeria maculans(Desmaz.) Ces. & De Not.,l’agent causal de la jambe noire du canola (Brassica napusL. and Brassica rapaL.), a été jugé comme un agentpotentiellement utile dans la lutte biologique. Cet isolat environnemental a été identifiéPaenibacillus polymyxad’après(i) ses caractéristiques biochimiques et de croissance et (ii ) la similitude de la séquence de l’ARNr 16S et il a étédésigné souche PKB1. Les peptides antifongiques deP. polymyxaPKB1 sont produits au début de la phase desporulation et leur production est optimale dans un bouillon pomme de terre-dextrose. Ces peptides antifongiques ontété extraits de cellules et (ou) spores deP. polymyxaPKB1 par un procédé au méthanol et purifiés par tamisagemoléculaire et chromatographie de phase inversée. La caractérisation de ces peptides antifongiques a été obtenue parune analyse de la composition en acides aminés, séquençage après dégradation d’Edman sur du matériel partiellementhydrolysé et une variété de méthodes de spectrométrie de masse. Le matériel antifongique purifié s’est révélé commeétant un mélange de peptides apparentés de poids moléculaires de 883, 897, 948 et 961 Da et il semble que la struc-ture la plus vraisemblable du composé de 897 Da soit un depsipeptide cyclique possédant une moitié peu fréquented’acide 15-guanidino-3-hydroxypentadécanoïque liée à un groupement aminé libre. Ces composés sont donc desmembres du groupe de la fusaricidine des depsipepsides cycliques.
Mots clés: depsipepside cyclique, agent de contrôle biologique, allo-thréonine.
[Traduit par la Rédaction] 169
Introduction
Significant losses in canola (Brassica napusL. and Bras-sica rapaL.) seed quality and yield are seen every year be-cause of fungal diseases, such as blackleg, caused by the
soilborne fungusLeptosphaeria maculans(Desmaz.) Ces. &De Not. Other pathogenic fungi, includingSclerotiniasclerotiorum (Lib.) de Bary, Rhizoctonia solani Kühn,Fusarium avenaceum(Corda:Fr.) Sacc., andAlternariabrassicae(Berk.) Sacc., cause additional losses. Blackleg isconsidered an endemic disease in most regions of the worldwhere these crops are grown. It has been a serious impedi-ment to rapeseed and canola cultivation in Europe since1950, and the disease virtually wiped out the rapeseed andcanola industry in western Australia in 1972. In Canada,economic losses due to blackleg are estimated to be nearly50 million dollars annually (Kharbanda 1994).
Methods used to date to slow or stop the spread and sever-ity of blackleg have included the use of fungicides, the devel-
Can. J. Microbiol.48: 159–169 (2002) DOI: 10.1139/W02-002 © 2002 NRC Canada
159
Received 6 September 2001. Revision received 14 December2001. Accepted 17 December 2001. Published on the NRCResearch Press Web site at http://cjm.nrc.ca on 3 March 2002.
P.H. Beatty and S.E. Jensen.1 Department of BiologicalSciences, University of Alberta, Edmonton, AB T6G 2E9,Canada.
1Corresponding author (e-mail: [email protected]).
I:\cjm\cjm48\4802\W02-002.vp27-Feb-2002 10:08:19 AM
Color profile: DisabledComposite Default screen

opment of blackleg-resistant canola cultivars, and the use ofcultural practices that limit the spread of disease (Kharbanda1993). In the past decade, research has been conducted to finda biocontrol agent active againstL. maculans(Kharbanda andDahiya 1990; Chakraborty et al. 1994), but no effective agentis available for use at present. However, interest in the devel-opment of biocontrol agents active against agricultural pestscontinues to be strong because of public pressure on the agri-cultural industry to reduce the use of chemicals.
Biocontrol agents in use or under investigation for thecontrol of agricultural diseases and pests include many gen-era of bacteria and filamentous fungi, but not all organismsare equally well suited for such applications. In addition tothe need to provide effective control of the disease or pest, asuccessful biocontrol agent must be robust enough to survivethe processing and storage conditions involved in its formu-lation and use. For this reason, endospore-forming bacteria,in particular Bacillus spp. and related genera, have espe-cially desirable properties. These microbes are well-knownas producers of a broad range of peptide metabolites withantibacterial and (or) antifungal activities, and in addition,they form endospores that are highly resistant to both chemi-cal and physical stresses. These properties combined withtheir natural ability to flourish in the soil environment makethem well suited for biocontrol applications.
In this study, we report the identification of a soil bacte-rium able to inhibit the growth ofL. maculans(Kharbandaet al. 1998) as a strain ofPaenibacillus polymyxa(formerlyBacillus polymyxa; reclassified by Ash et al. 1993). Strainsof this species have been isolated previously from therhizosphere or mycorrhizosphere of white clover, perennialryegrass, crested wheatgrass (Holl et al. 1988), lodgepolepine (Holl and Chanway 1992), wheat (Heulin et al. 1994),Douglas fir (Shishido et al. 1996), green bean (Petersen etal. 1996), and garlic (Kajimura and Kaneda 1996, 1997), andsome of these strains exhibit plant growth-promoting proper-ties, such as nitrogen fixation and phosphate solubilization,while others produce various peptide antibiotics. A majorfamily of peptide antibiotics produced by some but not allstrains of this species is the polymyxin–colistin–circulinfamily (Umezawa et al. 1978; Shoji et al. 1977). Otherstrains of the same species fail to produce the polymyxin–colistin–circulin classes of compounds and instead producedifferent peptides, including polypeptins (Doi andMcGloughlin 1992); jolipeptin (Ito and Koyama 1972a,1972b); gatavalin (Nakajima et al. 1972, 1975); gavaserin;saltavidin (Pichard et al. 1995); antifungal compounds,named as LI-F03, -F04, -F05, -F07, and -F08 (Kurusu et al.
1987); fusaricidins (Kajimura and Kaneda 1996, 1997); andpolyxin (Piuri et al. 1998). In addition, there are many re-ports in the literature of antimicrobial and antifungal proper-ties of P. polymyxa isolates where the nature of theinhibitory agent is undefined (Rosado and Seldin 1993;Smid et al. 1993; Oedjijono et al. 1993; Mavingui andHeulin 1994; Liang et al. 1996; Dijksterhuis et al. 1999;Seldin et al. 1999). Therefore, antibiotic production is a fre-quently encountered but nonuniform characteristic ofP. polymyxastrains.
In this study, we report the purification of the antifungalmaterial active againstL. maculansproduced by the newlyisolated strain ofP. polymyxaand its identification as repre-sentative of the fusaricidin family of peptides.
Methods
Bacterial and fungal strains, reagents, and suppliesPaenibacillus polymyxaPKB1 (= ATCC 202127) andL.
maculanswere obtained from P. Kharbanda at the Alberta Re-search Council.Bacillus polymyxa ssp. colistinus (ATCC21830) was obtainedfrom the American Type Culture Collec-tion. Most premixed solid and liquid media were purchased fromDifco (Detroit, Mich.) except for trypticase soy broth, which wasfrom Becton Dickinson and Company (Cockeysville, Md.).Trypticase soy with starch medium (TCSS), used for starter cul-tures ofP. polymyxa, contained 3% trypticase soy broth and 1%soluble starch. Potato dextrose broth (PDB) was used for produc-tion cultures ofP. polymyxa, and potato dextrose agar (PDA) wasused for plate cultures of bothP. polymyxaandL. maculans.
Culture and storage of microorganismsPaenibacillus polymyxastrains were maintained as spore
stocks in 20% glycerol stored at –70°C or as lyophilizedspores stored at –20°C. Standard growth conditions for thevarious strains ofP. polymyxainvolved the preparation of astarter culture by inoculation of TCSS medium to 1% (v/v)with a glycerol spore stock and then incubation at 28°C and200 rpm for 24 h. Production cultures were inoculated to 1%(v/v) with starter culture and then incubated as describedabove. When PKB1 biomass was required for purification ofthe antifungal material, 14-L stirred tank fermenters (NewBrunswick Scientific, Edison, N.J.) containing 10 L of PDB(with 1% polypropylene glycol as an antifoam agent) wereinoculated to 1% (v/v) with a TCSS starter culture. Thefermenters were stirred at 200 rpm, and airflow was set to15 L/min. After 72 h of incubation (or when sporulation wascomplete) at 25°C, spores were harvested by centrifugation
© 2002 NRC Canada
160 Can. J. Microbiol. Vol. 48, 2002
Primername Primer sequence* Application
8F 5′ GAGCTCTAGAATTCAGAGTTTGATCATGGCTCAG 3′ Amplification; sequencing704F 5′ TGTGTAGCGGTGAAATGCGTAGA 3′ Sequencing1176F 5′ AGGAAGGGGGGGAGGAGGT 3′ Sequencing358R 5′ CCCACTGGTGCCTCCCGTA 3′ Sequencing1106R 5′ CGCCCTTTTCGGGACTTAACCC 3′ Sequencing1403R 5′ TCGAGCTCTAGAATTCGGGCGGTGTGTACAAGGC 3′ Amplification; sequencing
*Primers were based on similar sequences reported by Hauben et al. (1997).
Table 1. Amplification and sequencing primers used to determine the 16S rRNA sequence ofPaenibacillus polymyxaPKB1.
I:\cjm\cjm48\4802\W02-002.vp27-Feb-2002 10:08:19 AM
Color profile: DisabledComposite Default screen

at 15 300 ×g for 10 min, resuspended in distilled water toform a thick slurry, and lyophilized.
Leptosphaeria maculanswas stored at –70°C as a 20%glycerol stock of pycnidiospores at 2 × 107 spores/mL.
Biochemical tests and growth on selective anddifferential media
Biochemical tests were conducted using API 20E and API20NE identification strips (bioMérieux, Marcy-l’Etoile,France). Additional selective and differential media wereprepared according to theDifco Manual(1984). Compoundstested as sole carbon sources forP. polymyxaPKB1 wereadded to B+N8P agar medium (Kropp et al. 1994). Chitin-degrading ability was assessed using the assay procedureoutlined by O’Brien and Colwell (1987), using 4-methyl-umbelliferyl-N-acetyl-β-D-glucosaminide (Sigma ChemicalCorp., St. Louis, Mo.) as substrate.
16S rDNA analysisPolymerase chain reaction (PCR) primers used to amplify
and sequence a 1390-bp region of the 16S rRNA gene ofP. polymyxaPKB1 are listed in Table 1. The PCR protocolused to amplify the gene fragment involved an initial dena-turation step at 94°C for 5 min, followed by 30 cycles of94°C for 30 s, 55°C for 30 s, and 72°C for 1.5 min, and a fi-nal extension step of 72°C for 5 min. PCR reaction mixturesfor sequencing contained the following: 1.2µL ofP. polymyxaPKB1 genomic DNA (25 ng/µL), 0.67µL eachof reverse and forward primers (30 pmol/µL), 0.4 µL ofdNTP mix (25 mM each of dATP, dCTP, dGTP, and dTTP;Boehringer-Mannheim, Indianapolis, Ind.), 5µL of 10× PCRbuffer (0.5 g KCl, 8.48 g Tris–HCl, 0.01 mL of Triton X-100/100 mL of dH2O, pH 8.8), 0.75µL of 0.1 M MgCl2,28.25µL of dH2O, and 1µL of Taq DNA polymerase (5 U/
µL). The Taq DNA polymerase was added to the reactionmixtures after they had been heated to 94°C for 2 min. DNAsequence analysis was conducted by the Biological SciencesMolecular Biology Service Unit, University of Alberta usingthe oligonucleotide primers listed in Table 1 together with aThermosequenase II kit (Amersham Pharmacia, Baie d’Urfe,Que.). Reaction mixtures were analyzed on a AppliedBiosystems model 373A sequencer with Stretch Upgrade.
Computer analysis of DNA sequencesBLASTn (National Center for Biotechnology Informa-
tion) was used as a search tool to examine DNA sequencesin the public databases (http://www.ncbi.nlm.nih.gov;Altschul et al. 1997), and an advanced ungapped BLASTsearch of the nucleic acid databases was also performed. Se-quence alignment was performed from the Ribosomal Data-base Project Web site (http://rdpwww.life.uiuc.edu; Maidaket al. 1997).
Small scale extraction and bioassay of antifungalantibiotic
Samples ofP. polymyxacultures were harvested after vari-ous incubation periods by centrifugation at 16 000 ×g for5 min. Cell pellets were resuspended in methanol(200 µL/1.5 mL of culture), incubated at 21°C for 20 min,and then centrifuged as above. Seventy-five microlitreamounts of the methanol extracts were applied to PDA plates,each consisting of 100 mL of medium in a 245 × 245 mmplate (Nalge Nunc International, Rochester, N.Y.), that hadpreviously been spread with 500–800µL of L. maculansglyc-erol spore stock. Sample wells were punched out of the agarwith the large end of a sterile 1-mL pipette tip. Plates wereincubated at 21°C, and zones of inhibition (ZOI) were mea-sured after 48–72 h.
© 2002 NRC Canada
Beatty and Jensen 161
Characteristic or test* Isolate PKB1 P. polymyxa†
Voges-Proskauer test + +, 90%Catalase producer + +, 90%Gelatin hydrolysis + +, 90%H2S from thiosulfate – –, 90%Tryptophan deaminase – –, 90%Urease; release of NH4
+ – –, 90%Citrate; C source – –, 90%Chitinase test + +, 90%Glucose fermentation – +, 90%Nitrate reduction + +, 90%Arabinose; C source + +, 90%Mannitol; C source + +, 90%Blood agar +, clear colourless zones +Acid from glucose + +, 90%Starch agar +, clear zones +, 90%Triple sugar iron slant
(glucose, lactose, sucrose)Growth on slope and inside
agar, gas produced+, 90% anaerobic growth;
+, 90% gas from glucose
Note: A positive (+) result for a substrate indicates utilization or production of the compound within 48h of inoculation in the test strip.
*Where only a substrate name is indicated, agar plates consisted of B+N8P medium with the substrate asthe sole carbon source and bromthymol blue as a pH indicator.
†The results of these tests on knownP. polymyxastrains are from Parry et al. (1983) and Holt et al.(1994).
Table 2. Biochemical and substrate utilization profile ofPaenibacillus polymyxaPKB1 com-pared withP. polymyxatype strains.
I:\cjm\cjm48\4802\W02-002.vp27-Feb-2002 10:08:20 AM
Color profile: DisabledComposite Default screen
This same bioassay procedure was used to determine thespectrum of inhibition of the PKB1 antifungal antibioticexcept that other indicator organisms were used in placeof L. maculans. In the case ofMicrococcus luteus, whole bac-terial culture was added to cooled molten agar medium usedto form the bioassay plate. ForNeurospora crassaShear &Dodge, Penicillium roquefortii Thom, and Streptomycesclavuligerus, spore suspensions were spread on the surface ofthe bioassay plates, just as described forL. maculans.
Similarly, the bioassay procedure withL. maculansas in-dicator organism was used to monitor antifungal activity ofvarious fractions throughout the purification procedure andresidual antifungal activity of samples after various physical,chemical, and enzymatic treatments.
Purification of antifungal antibioticLyophilizedP. polymyxaspores (1 g) were washed by sus-
pending in 100 mL of distilled water and harvested bycentrifugation at 15 300 ×g for 20 min. The wash procedurewas repeated once. Antifungal antibiotic was extracted fromthe washed spores by resuspending in 60 mL of methanol,incubating for 1 h at21°C, and harvesting by centrifugationat 10 000 ×g for 15 min. The methanol extraction was re-peated once, and the pooled methanol extracts were reducedto 10 mL by rotary evaporation. After centrifugation at 890× g for 12 min to sediment particulate matter, the crudemethanol extract was further reduced to 6 mL by rotaryevaporation.
Sephadex LH-20 size exclusion chromatographyThe crude methanol extract was applied in 3-mL aliquots
to a Sephadex LH-20 (Amersham Pharmacia) column (2 ×24 cm, equilibrated in methanol) and eluted with methanolat a flow rate of 0.2 mL·min–1. Two-millilitre fractions werecollected, and antifungal activity was located by bioassay.Fractions containing antifungal material were pooled and re-duced to 4 mL under a stream of nitrogen. These samplesare referred to as LH-20 purified material.
Reverse-phase (RP) C18 chromatographyThe LH-20 purified material was diluted twofold with wa-
ter to give 50% methanol and purified further by passagethrough a C18 Sep-pak Classic Short Body column (WatersAssociates, Milford, Mass.), following the manufacturer’sprotocol. The active component, located by bioassay, waseluted from the column in 100% methanol. Active fractions
were pooled and dried under a stream of nitrogen and werereferred to as Sep-pak purified material.
Reverse-phase high-performance liquid chromatography(RP-HPLC)
Chromatography on a C18 µBondapak™ radially com-pressed 8 × 100 mm RP-HPLC column (Waters Associates)was used as a final purification step. Sep-pak purified mate-rial was dried and resuspended in 1 mL of the mobile phaseused for RP-HPLC. Samples (200µL) were analyzed using abinary gradient mobile phase at a flow rate of 1 mL·min–1.Mobile phase composition was as follows: 100% A goes to40% A over 2.5 min; 40% A for 7.5 min; 40% A goes to 0%A over 40 min; 0% A goes to 100% A over 5 min; 100% Afor 10 min, where solvent A was 0.1% (w/v) disodiumorthophosphate and solvent B was acetonitrile. Eluate wasmonitored at 214 nm using a Lambda Max model 480 LCUV-Spectrophotometer (Waters Associates), and 1.0-mLfractions were collected. Active fractions were located bybioassay, pooled, and dried under a stream of nitrogen andare referred to as pure antifungal material.
Enzyme digestion
Pronase digestionPronase is a crude mixture of enzymes that includes car-
boxypeptidases and aminopeptidases as well as at least fiveserine-type proteases and two zinc endopeptidases (Hedigeret al. 1973). One millilitre of pronase E (Sigma) solution at0.3, 0.5, 0.6, or 0.7 mg·mL–1 in 0.01 M Tris–HCl (pH 7.8),0.01 M EDTA, and 0.5% (w/v) SDS buffer was added to100-µL samples of crude methanol extract, which had beenreduced to dryness under a stream of nitrogen. Reactionmixtures were incubated at room temperature for 2 h, driedusing a SpeedVac (Savant Instruments Inc., Farmingdale,
© 2002 NRC Canada
162 Can. J. Microbiol. Vol. 48, 2002
Inhibition against fungi Inhibition against bacteria
Alternaria brassicae* Micrococcus luteusFusarium avenaceum* Streptomyces clavuligerusLeptosphaeria maculansMarasmius oreadesFries*Neurospora crassaPenicillium roquefortiiRhizoctonia solani*Sclerotinia sclerotiorum*
*Kharbanda 1999.
Table 3. Spectrum of inhibition for thePaenibacilluspolymyxaPKB1 antifungal antibiotic.
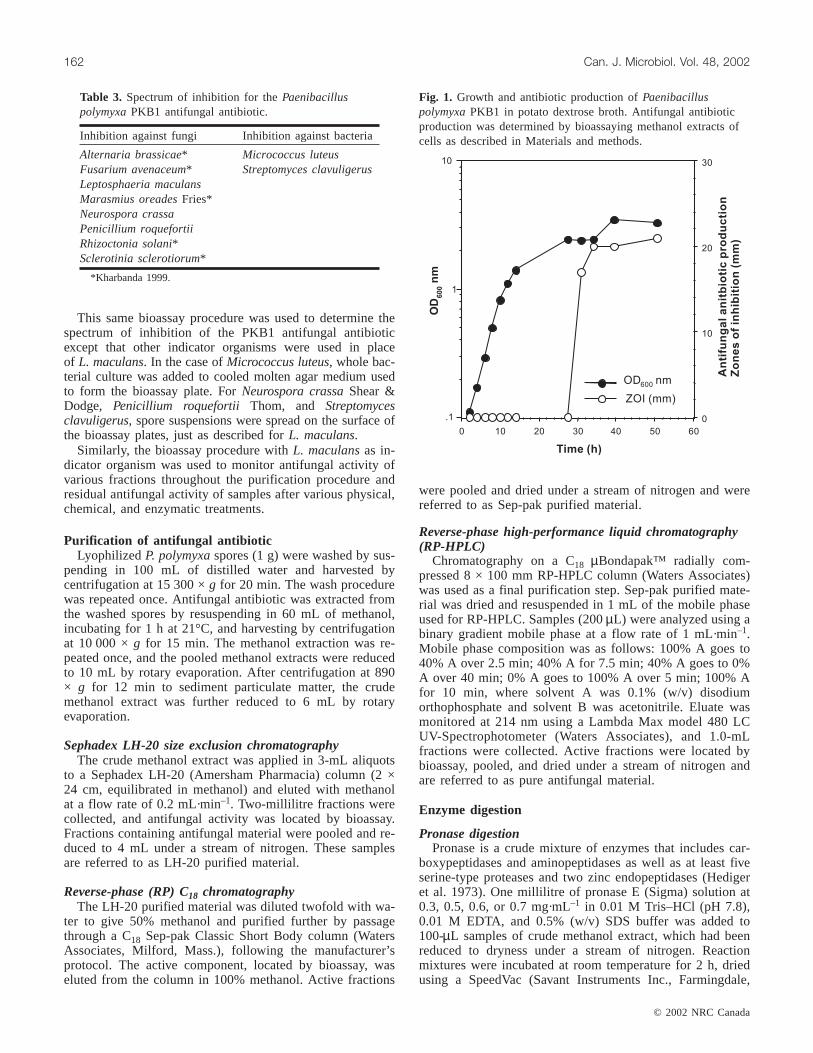
Fig. 1. Growth and antibiotic production ofPaenibacilluspolymyxaPKB1 in potato dextrose broth. Antifungal antibioticproduction was determined by bioassaying methanol extracts ofcells as described in Materials and methods.
I:\cjm\cjm48\4802\W02-002.vp27-Feb-2002 10:08:20 AM
Color profile: DisabledComposite Default screen
N.Y.), and the residue was resuspended in 100µL of metha-nol and assayed for antifungal activity by bioassay.
Trypsin and carboxypeptidase digestionOne hundred microlitres of Sep-pak purified material (re-
duced to dryness) was subjected to digestion with either 150µg trypsin (Boehringer-Mannheim), 40µg of carboxy-peptidase (Boehringer-Mannheim), or both of the proteases.The reactions were carried out in a final volume of 1 mL of0.1 M ammonium bicarbonate and incubated at room tem-perature for 30 min. When the antifungal material was sub-jected to digestion by both proteases, carboxypeptidase wasadded first and incubated for 30 min, followed by trypsin,with a further 30 min of incubation before the enzymes wereheat inactivated at 70°C for 10 min. Samples were analyzedby C18 RP-HPLC, and the resulting HPLC fractions weremonitored by bioassay.
Asp-N endoproteinase digestionThe endoproteinase Asp-N can cleave amide bonds on the
carboxyl side of aspartic acid residues (rapid) and on thecarboxyl side of glutamic acid residues (slow), depending onthe reaction conditions used. Pure antifungal material wassubjected to digestion with endoproteinase Asp-N(Boehringer-Mannheim) according to the manufacturer’s in-structions. One series of reactions was carried out underconditions designed to cleave at aspartic acid residues. Pureantifungal material, dried under a stream of nitrogen, wassuspended in 200µL of 50 mM sodium phosphate buffer(pH 8.0) to a concentration of 0.036 mg·mL–1 as determinedby the bicinchoninic acid assay (Pierce Chemicals) using bo-vine serum albumin as standard. Aliquots (50µL) of thismaterial were mixed with 0.0015–1.5 U of Asp-Nendoproteinase. Reaction mixtures were incubated for 3 h at37°C, then the reactions were terminated by heat inactiva-tion at 80°C for 10 min. A second series of reactions wasperformed under conditions designed to cleave at glutamicacid residues. Pure antifungal material was dried and sus-pended into 150µL of 50 mM sodium phosphate (pH 8.0),containing 10% acetonitrile, to a concentration of
0.048 mg·mL–1 as determined by the bicinchoninic acid as-say. Aliquots (50µL) of this sample were mixed with 0.24–2.4 mU of Asp-N endoproteinase. Reaction mixtures wereincubated at 37°C for 19 h, and then the reactions were ter-minated by heat inactivation at 80°C for 10 min. Samples ofeach reaction mixture (4µL in the first series and 10µL inthe second series) were tested for antifungal activity bybioassay. The remainder of the mixture was subjected to N-terminal amino acid sequence analysis (Micro-Sequencinglaboratory, University of Victoria, B.C.).
Esterase digestionSep-pak purified material (5 mL) was dried under a stream
of nitrogen and treated with 100 U of porcine liver esterase(Boehringer-Mannheim) in 1 mL of 50 mM sodium phos-phate buffer, pH 8.0. Samples (0.33 mL) were taken at 30, 60,and 90 min and heat inactivated at 70°C for 10 min. The sam-ples were dried under a stream of nitrogen and resuspended inacetonitrile–H2O (2:1) containing 0.1% trifluoroacetic acid,before analysis by C18 RP-HPLC. Fractions were monitoredby bioassay.
Mass spectrometryPure antifungal material was evaporated to dryness and
subjected to mass spectroscopic analyses by a variety of pro-cedures. The antifungal material was dissolved in a glyc-erol–thioglycerol matrix and analyzed by fast atombombardment (FAB) and metastable ion analysis (Micro-Sequencing laboratory, University of Victoria; Biemann1986). Additional analyses using electrospray time of flight(TOF) tandem mass spectrometry (MS–MS; University ofAlberta Mass Spectroscopy laboratory; Biemann 1986) werealso conducted. Finally, matrix assisted laser desorption ion-ization (MALDI) treatment of antifungal material in eitheran α-cyano-4-hydroxycinnaminic acid or a 2,5-dihydroxy-benzoic acid matrix generated ions that were analyzed bypost source decay (PSD) with a TOF mass analyzer (H. vonDohren, Technical University of Berlin; Erhard et al. 1997).In addition to MS analyses of the purified antifungal mate-
© 2002 NRC Canada
Beatty and Jensen 163
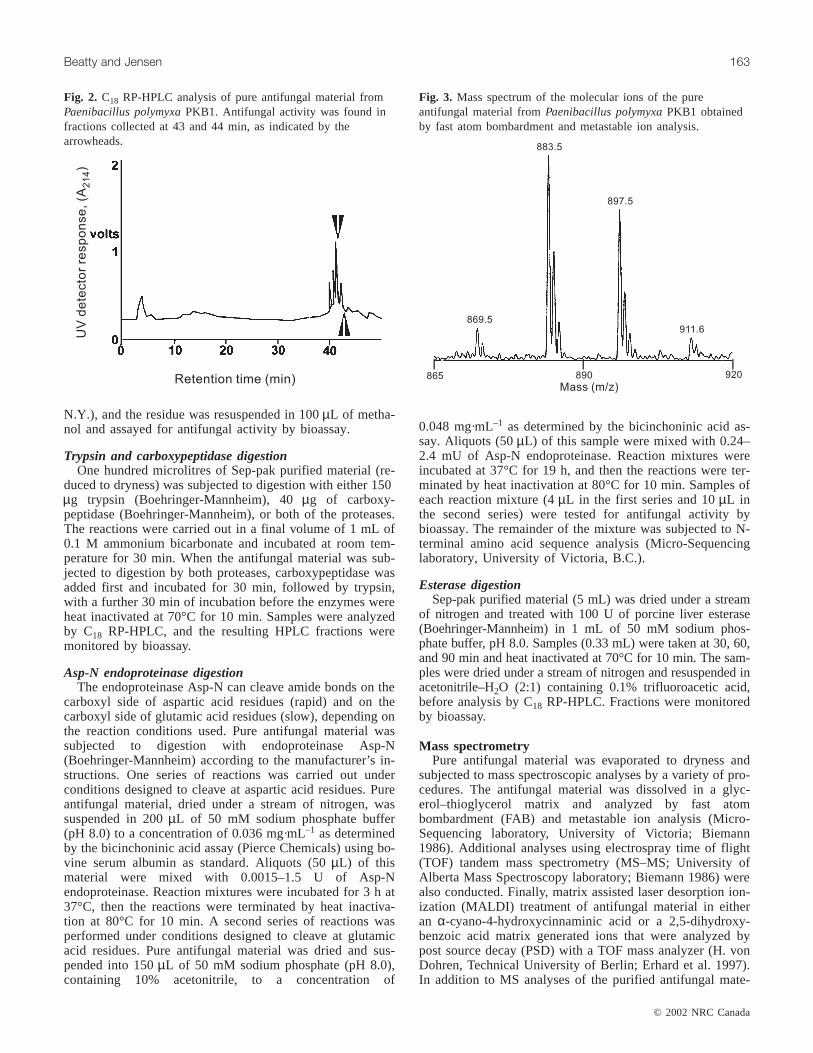
Fig. 2. C18 RP-HPLC analysis of pure antifungal material fromPaenibacillus polymyxaPKB1. Antifungal activity was found infractions collected at 43 and 44 min, as indicated by thearrowheads.
Fig. 3. Mass spectrum of the molecular ions of the pureantifungal material fromPaenibacillus polymyxaPKB1 obtainedby fast atom bombardment and metastable ion analysis.
I:\cjm\cjm48\4802\W02-002.vp27-Feb-2002 10:08:21 AM
Color profile: DisabledComposite Default screen

rial, whole cells of PKB1 were analyzed by MALDI-TOFMS. Cells were mixed with the ionization matrix in organicsolvents to extract secondary metabolites, as described byErhard et al. (1997).
Results and discussion
Identification of PKB1Canola stubble collected from a field near Edmonton, Alta.,
Canada was used to isolate microorganisms with antifungalproperties, and PKB1 was selected as one such isolate withparticularly good antifungal activity againstL. maculans(Kharbanda et al. 1998). PKB1 cells were motile, gram-positive rods approximately 2–4 × 1–1.5µm. The cells de-veloped subterminal endospores that were ellipsoid andrefractile. When grown on PDA medium, PKB1 colonieswere opaque, tan colored, and glistening. The cells also pro-duced a shiny, white, mucoid capsular material.
Biochemical tests were performed, and growth character-istics were observed on a variety of media as described inTable 2. Except for its negative reaction for glucose fermen-tation, the growth characteristics of PKB1 are consistentwith its identification as a strain ofP. polymyxa(Parry et al.1983; Holt et al. 1994). The cellular and colony morphologyof PKB1 were also very similar to that described for the typeculture ofP. polymyxaATCC 842 (Parry et al. 1983; Holt etal. 1994).
In addition to these biochemical and morphological char-acteristics, a 1390-bp region of the 16S rDNA sequence ofPKB1 was determined (deposited under GenBank accessionNo. AF355463) and compared with that of known species ofmicroorganisms. A BLASTn search (Altschul et al. 1997;Peruski and Peruski 1997) for DNA sequences resemblingthe PKB1 16S rDNA gene sequence determined that it wasmost similar to P. polymyxa 16S rDNA (accession No.X60632). A gapped sequence alignment protocol was usedto carry out Ribosomal Database Project (RDP) analysis ofthe 16S rDNA sequence from PKB1. The sequence alignedclosely with P. polymyxaIAM 13419, with 98.8% identity.Based on these observations, the soil isolate PKB1 was iden-tified as P. polymyxaand has since been deposited at the
American Type Culture Collection under the identificationNo. ATCC 202127.
Optimization of P. polymyxaPKB1 growth andantibiotic production
PKB1 was originally isolated by cultivating it on PDAwhere it was seen to inhibit the growth ofL. maculans(Kharbanda et al. 1998). PDA has a pH of about 5 and isused frequently for the cultivation of fungi, but is too acidicfor optimal growth of most bacteria. To optimize growth andantifungal activity, PKB1 was cultivated on a variety ofgrowth media, including complex media, such as brain heartinfusion broth (BHI), PDB, and TCSS, and on chemicallydefined media, such as Katznelson and Lochhead (KL;Kanamori et al. 1987) and Stansly’s medium (Paulus andGray 1964). PDB supported only moderate growth of PKB1compared with BHI and TCSS, but production of antifungalactivity was consistently higher than in any other mediumtested. Furthermore, attempts to increase antifungal activityby supplementation of PDB with various carbon, nitrogen,or mineral sources or by manipulation of the pH producedno better results than did unmodified PDB. Therefore, PDBwas used for all subsequent studies.
Effect of temperature on growth was also examined, andwhereas growth was optimal at 30°C, production ofantifungal activity was maximal at 25°C. Growth at 25°Cwas therefore used for all studies described in this paper, butgrowth and production of antifungal activity was also notedto persist at temperatures as low as 15°C (data not shown).This more closely approximates soil temperatures found incanola growing regions and holds promise for the use ofPKB1 as a biocontrol agent.
Purification of the antifungal antibioticPreliminary studies showed that the antifungal activity
produced by PKB1 was associated with spores harvested inthe early stationary phase of growth (Fig. 1). No antifungalactivity could be recovered from the culture broth. Anti-fungal material was only detected upon the onset ofsporulation, not beforehand. Purification of the antifungalmaterial was carried out using lyophilized spores of PKB1
© 2002 NRC Canada
164 Can. J. Microbiol. Vol. 48, 2002
Fig. 4. The structures of fusaricidins A, B, C, and D. Adapted from Umezawa et al. 1978 and Kajimura and Kaneda 1996 and 1997.
I:\cjm\cjm48\4802\W02-002.vp27-Feb-2002 10:08:22 AM
Color profile: DisabledComposite Default screen
as a starting material as described in Materials and methods.The antifungal material could be extracted from washedspores using methanol. The concentrated crude methanol ex-tract was then passed through a Sephadex LH-20 column,followed by batch-wise purification on a C18 RP cartridge,and finally purification by HPLC on a C18 RP column. Col-umn fractions were monitored for absorbance at 214 nm (noabsorbance at 280 nm was observed) and for bioactivity.When the antifungal material recovered after batch-wise pu-rification on the C18 RP cartridge was subjected to a finalHPLC purification step, the material was resolved into acluster of four closely spaced peaks, and antifungal activitywas associated with the last two peaks of the cluster (Fig. 2).A variety of chromatographic conditions and mobile phases
were tested in an attempt to separate these components, butfurther resolution could not be obtained.
Inhibitory spectrum of antifungal antibiotic producedby P. polymyxaPKB1
Crude methanol extract of the antifungal material fromPKB1 was tested for bioactivity against fungal and bacterialspecies and was found to be active against a variety of fungi,including some responsible for costly agricultural crop dis-eases (Table 3). The antifungal material was ineffectiveagainst most bacteria, although it was found to inhibit somegram-positive actinomycetes, includingMicrococcus luteusandStreptomyces clavuligerus(Beatty 2000). Of the variousantibiotics reported to be produced by strains of
© 2002 NRC Canada
Beatty and Jensen 165
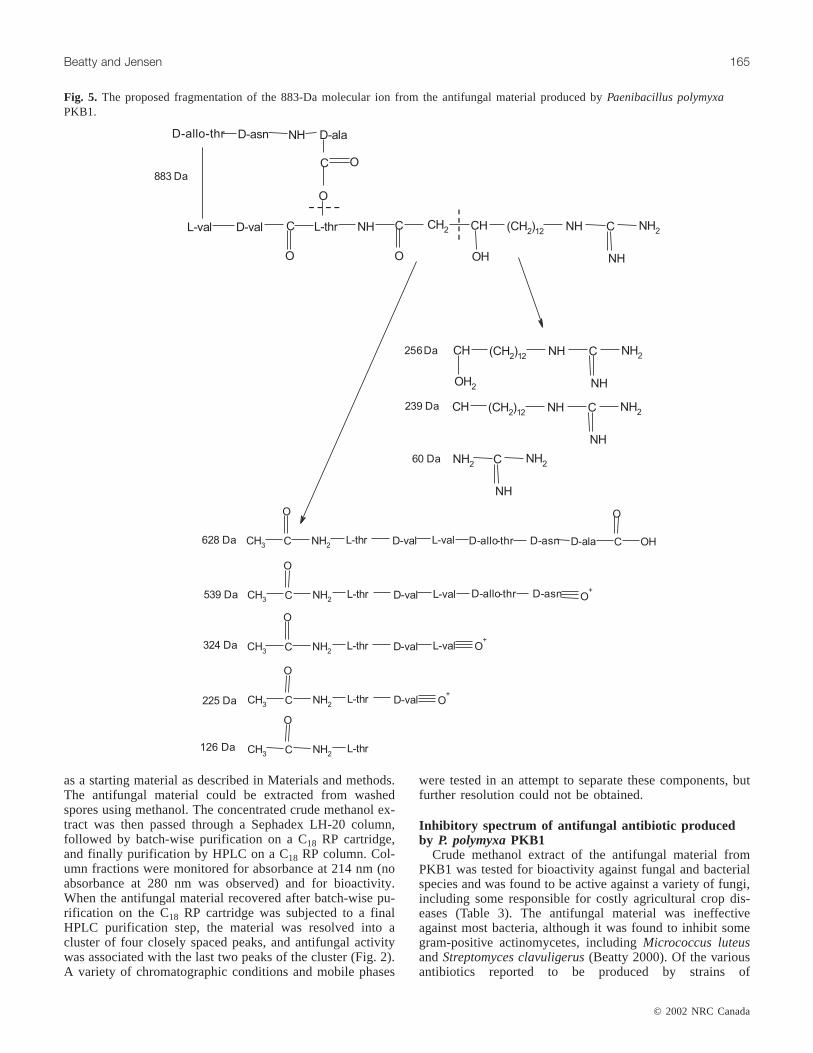
Fig. 5. The proposed fragmentation of the 883-Da molecular ion from the antifungal material produced byPaenibacillus polymyxaPKB1.
I:\cjm\cjm48\4802\W02-002.vp27-Feb-2002 10:08:22 AM
Color profile: DisabledComposite Default screen

P. polymyxa, the polymyxins, polypeptins A and B,jolipeptin, gavaserin, saltavalin, and polyxin are all activeagainst both gram-negative and gram-positive bacteria(Umezawa et al. 1978; Shoji et al. 1977; Ito and Koyama1972a; Pichard et al. 1995; Piuri et al. 1998), and so it ap-peared that PKB1 was not producing any of these metabo-lites under the growth conditions used. However, otherisolates of P. polymyxahave been reported that producegatavalin; antifungal compounds named as LI-F03, LI-F04,LI-F05, LI-F07, and LI-F08; and fusaricidins A, B, C, andD. These metabolites are reported to have spectra of activityagainst fungi and certain gram-positive bacteria that are sim-ilar to what was observed for the antifungal material fromPKB1 (Nakajima et al. 1972; Kurusu et al. 1987; Kajimuraand Kaneda 1996, 1997).
Characteristics of the antifungal materialPure antifungal material was soluble in methanol,n-
butanol, ethanol, acetonitrile, isopropanol, and 0.1% aceticacid; slightly soluble in water; and insoluble inn-hexane,ethyl acetate, and methylene chloride. Sep-pak purifiedantifungal material in 50% methanol could be heated to90°C for 1 h, with no loss of activity, and could also bestored indefinitely at 4°C, with no loss of activity. Acid-hydrolyzed antifungal material (6 N HCl, 100°C, 16 h),lyophilized to dryness and separated by thin-layer chroma-tography on a cellulose thin-layer sheet (mobile phasen-butanol – acetic acid – H2O; 3:1:1), gave several ninhydrin-positive spots. In contrast, unhydrolyzed material did not re-act with ninhydrin, suggesting that the antifungal material ispeptidic in nature, but that the peptide is likely to be cyclicor N-terminally blocked.
The effect of digestion of antifungal antibiotic withtrypsin, carboxypeptidase, and pronase was examined bybioassay and (or) RP-HPLC. No reduction of antifungal ac-tivity was seen after treatment with these proteases nor wasthe HPLC elution profile altered (data not shown). Similarly,treatment of purified antifungal material with endoproteinaseAsp-N under conditions that supported cleavage at asp resi-dues only or at both asp and glu residues did not result in aloss of antifungal activity, as measured by bioassay. The ob-servation that the antifungal material was unaffected bytreatment with any of these proteases again suggested that ifit is a peptide, it must be cyclic, N- and C-terminallyblocked, or composed of unnatural amino acids. Cyclic pep-tides can be resistant to cleavage by proteases because theircyclic structure renders them relatively inflexible, whichmay make cleavage sites inaccessible because of steric hin-drance (Eckart 1994). These peptides often owe their cyclicnature to the presence of ester linkages and are then referredto as depsipeptides.
To examine whether the antifungal material produced byPKB1 might be depsipeptide in nature, pure antifungal mate-rial was tested for esterase sensitivity. The characteristicclosely spaced cluster of peaks seen in the HPLC profile ofpurified antifungal material disappeared, and antifungal ac-tivity was progressively lost as the time of esterase treatmentincreased. The ZOI obtained from HPLC fractions pooledfrom the region where antifungal material elutes were 30,27, 24, and 0 mm for samples treated with esterase for 0, 30,
60, and 90 min, respectively. This suggests that ester bondsessential for antifungal activity are present in the material.Taken together, these results suggest that the antifungal ma-terial produced by PKB1 is a cyclic depsipeptide.
N-Terminal sequence analysis, amino acidcompositional analysis, and mass spectroscopic analysisof the antifungal material from P. polymyxaPKB1
Pure antifungal material was subjected to Edman degrada-tion, but did not yield N-terminal sequence information,again suggesting that the peptide is blocked at the N-terminus or cyclized. Antifungal material subjected to diges-tion with endoproteinase Asp-N was also resistant to Edmandegradation, confirming that the proteinase was unable to at-tack the material.
Antifungal material subjected to total acid hydrolysis (6 NHCl, 100°C, 16 h) was analyzed to reveal the followingamino acid composition: asx (0.5), glx (0.6), thr (0.8), ala(1.0), val (1.6), and an unknown amino acid, designated X(1.0), which did not correspond to any of the 20 proteinamino acids. These ratios are calculated based on an as-sumption of one alanine residue, and glx indicates either gluor gln. When pure antifungal material was subjected to par-tial acid hydrolysis (3 N HCl, 100°C, 3 h) and then toEdman degradation, N-terminal sequence information wasobtained. This analysis indicated the following sequence:NH2-thr-val-val-X-glu-ala-COOH, where X again representsan unknown amino acid.
Further structural information on the antifungal peptidefrom PKB1 was obtained by carrying out mass spectroscopicstudies using two different soft ionization procedures. Thehigh sensitivity of these methods made it possible to obtainstructural confirmation using very small quantities of pureantifungal material, much less than would be required forother types of conformational experimentation techniques,such as nuclear magnetic resonance (NMR). The FAB MSspectrum showed the presence of two parent ions withmasses of 883.5 and 897.5 Da, suggesting that the purifiedantifungal material analyzed was actually a mixture of twocomponents differing in molecular mass by 14 Da (Fig. 3).Antifungal peptides with these characteristic masses havebeen reported previously as products of some strains ofB. polymyxa(now P. polymyxa). Nakajima et al. (1972) re-ported the isolation of aB. polymyxa strain (namedB. polymyxassp.colistinus koyamaand deposited as ATCC21830) with antifungal properties. They isolated and par-tially characterized the antifungal material, which theycalled gatavalin, with a total amino acid composition of asx(1.1), glx (1.0), ala (2.2), thr (3.8), and val (3.8), where asxindicates either asp or asn, and glx indicates either glu orgln. Mass spectroscopic analysis indicated that the materialwas an inseparable mixture of two peptides with masses of883 and 897 Da. Despite the disagreement in the total aminoacid composition, the close agreement in masses suggestedthat PKB1 may be producing a form of a gatavalin-like pep-tide. To investigate this similarity more closely, we obtaineda culture of B. polymyxassp. colistinus koyama(ATCC21830) and cultivated it under the conditions used byNakajima et al. (1972). When cells of that isolate were sub-jected to methanol extraction and the extract purified by the
© 2002 NRC Canada
166 Can. J. Microbiol. Vol. 48, 2002
I:\cjm\cjm48\4802\W02-002.vp27-Feb-2002 10:08:23 AM
Color profile: DisabledComposite Default screen

same procedures used for the antifungal material fromPKB1, a peptide product with a very similar HPLC profileand the same molecular masses as that seen from PKB1 wasobtained. This provides strong indication thatB. polymyxassp.colistinus koyama(ATCC 21830) and PKB1 both pro-duce the same or a similar family of peptide antibiotics.However, no further structural information on the gatavalinfamily of peptides has since appeared.
Kurusu et al. (1987) reported the isolation of anotherstrain ofB. polymyxathat produced a series of peptides withantifungal properties. They designated these peptides LI-F03, LI-F04, LI-F05, LI-F07, and LI-F08. Once again, nofull structural analyses were conducted, but mass spectro-scopic analyses indicated that LI-F04, one of the majorproducts, was actually a mixture of two peptides withmasses of 883 and 897 Da. Most recently, Kajimura andKaneda (1996) have reported the isolation of another strainof B. polymyxa that produces peptide antibiotics withantifungal properties. These peptides were designatedfusaricidins A–D. Unlike the previous studies, completestructural determination of the fusaricidin peptides was con-ducted using a combination of MS and NMR analyses.Fusaricidins A and B have masses of 883 and 897 Da, re-spectively, and structures as shown in Fig. 4. The amino acidcompositions of fusaricidins A and B were consistent withthat observed for the antifungal material produced by PKB1,assuming that the unknown amino acid X encountered in thePKB1 product wasD-allo-threonine. To confirm that as-sumption, we repeated total amino acid analysis of purifiedantifungal material from PKB1, including allo-threonine as astandard. The unknown amino acid X was found to havechromatographic properties identical to those of authenticallo-threonine.
Fusaricidin B contains glutamine in position five of thepeptide, whereas sequence analysis of the material fromPKB1 indicated the presence of a glutamic acid residue.This may represent a difference between fusaricidin B andthe antifungal antibiotic from PKB1. However, the hydroly-sis conditions used could have deamidated a glutamine resi-due, forming the glutamic acid residue that was measured. Insupport of this possibility, the mass of the peptide antibioticfrom PKB1 of 897 Da is consistent with the presence of aglutamine residue instead of glutamic acid. The structure ofthe fusaricidin peptides is also consistent with the observa-tion that partial acid hydrolysis released the previouslyblocked N-terminal group of the PKB1 antifungal material.The N-terminal threonine residue of the fusaricidin peptidesis blocked by a 15-guanidino-3-hydroxypentadecanoic acid(GHPD) moiety that is released when these depsipeptidesare hydrolyzed, leaving a free amino group on the threonineresidue (Kajimura and Kaneda 1997).
The amino acid composition, sequence, and FAB – massspectra of the antifungal material from PKB1 are consistentwith the known sequences and structures of fusaricidins Aand B. Fusaricidins A and B differ only in a single aminoacid residue (A has an asparagine residue in position five,whereas B has a glutamine residue) to give masses of 883and 897 Da, respectively. The antifungal material in thisstudy contains the same two molecular ions, and both asxand glx were present in the total amino acid composition,
along with the other amino acids that comprise fusaricidinsA and B.
The fusaricidin peptides are cyclic, with the ring closedby an ester bond between theβ-hydroxy group of threonineand the C-terminal carboxyl group of alanine and with aGHPD side chain bound to theα-amino group of threonine.This side chain is identical for all four of the cyclic peptidesof the fusaricidin group. Although complete MS analysis ofthe fusaricidins was not reported by Kajimura and Kaneda(1996, 1997), MS was used to determine the nature of theside chain. Total acid hydrolysis of fusaricidins A and Bgave a component that partitioned into organic solvent ratherthan into the aqueous phase. This component was analyzedby FAB–MS and was determined to comprise a 298-Da mo-lecular ion. Further analysis of this ion by a variety of NMRprocedures determined it to have the GHPD structure.
In the present work, a 256-Da ion was abundant in everymass spectrum generated from the antifungal material pro-duced by PKB1 (data not shown). The MALDI–PSD massspectrum of the 883-Da molecular ion showed a peak at256 Da as well as a peak at 628 Da (the difference betweenthe 883-Da molecular ion and the 256-Da fragment). The256-Da ion peak was interpreted to represent the GHPD sidechain that was easily fragmented from the cyclic peptide,and the 628-Da ion peak comprised the remaining cyclicring structure of the peptide. Tandem MS analysis was per-formed on the 256-Da ion, giving a fragmentation patternconsistent with it being part of a GHPD side chain. A frag-mentation pattern for the entire 883-Da antibiotic metabolitefrom PKB1 that is consistent with the mass spectra obtainedafter soft ionization is illustrated in Fig. 5.
In contrast to MS analyses of the purified antifungal mate-rial, when whole cells of PKB1 were analyzed by MALDI–TOF MS, the spectrum showed the presence of two clustersof peaks, one containing molecular ions of 870, 883, 897,and 912 Da and a second containing molecular ions of 947and 961 Da (data not shown). Fusaricidins C and D havemasses of 947 and 961 Da, respectively, and since peakswith these masses were seen previously in MS spectra of thepurified antifungal material from PKB1, we conclude thatfusaricidins C and D and, possibly, other fusaricidin-typepeptides are also made by PKB1. However, bioassays sug-gest that they are not inhibitory towardsL. maculans.In ad-dition to the two clusters of peaks, the MALDI–TOFspectrum also showed the presence of three higher molecularmass ions at 1203, 1225, and 1241 Da. These molecular ionspresumably represent other secondary metabolites that areproduced by PKB1.
Based on information from the literature and on the re-sults of this study, strains ofP. polymyxawould appear toproduce two major groups of peptide antibiotics. One groupcomprises the antibiotics active against both gram-negativeand gram-positive bacteria, including the polymyxins,polypeptins, jolipeptin, gavaserin, and saltavalin. These pep-tide antibiotics all contain 2,4-diaminobutyric acid. Theother group is made up of the peptide antibiotics activeagainst fungi and gram-positive bacteria and includes theantifungal material from PKB1; gatavalin; antifungal com-pounds named as LI-F03, LI-F04, LI-F05, LI-F07, and LI-F08; and fusaricidins A, B, C, and D. This last group of pep-
© 2002 NRC Canada
Beatty and Jensen 167
I:\cjm\cjm48\4802\W02-002.vp27-Feb-2002 10:08:23 AM
Color profile: DisabledComposite Default screen

tide antibiotics has not been characterized structurally aswell as the first group, but they are apparently all similar inmolecular mass and amino acid composition. In fact, basedon these characteristics, the antifungal material from PKB1,gatavalin, and LI-F04 all appear to be mixtures offusaricidins A and B; LI-F03 appears to be a mixture offusaricidins C and D; LI-F05 is a mixture of fusaricidin Aand an uncharacterized fusaricidin-type peptide; and LI-F07and LI-F08 may also be mixtures of other as yetuncharacterized fusaricidin-type peptide antibiotics.
Purification and characterization of the antifungal materialfrom P. polymyxaPKB1 has helped to clarify the relation-ship between these related groups of antifungal peptide anti-biotics produced by various isolates ofP. polymyxa.Furthermore, the mass spectrometric methods described pro-vide a means by which the fusaricidin-producing ability ofenvironmental isolates ofP. polymyxacan be assessed usingonly small amounts of purified material. The antifungal ac-tivity of these peptides againstL. maculansand other dis-ease-causing fungi makes these strains ofP. polymyxapromising candidates for development as biocontrol agentsagainst blackleg of canola and other fungal diseases. Green-house studies and field trials are currently in progress to as-sess the efficacy ofP. polymyxa PKB1 for control ofblackleg disease of canola.
Acknowledgements
This study was supported by grants from the Alberta Ag-riculture Research Institute. P.H.B. was also supported by anindustrial studentship from the Natural Sciences and Engi-neering Research Council of Canada with industrial supportfrom the Alberta Canola Commission.
References
Altschul, S.F., Madden, T.L., Schaffer, A.A., Zhang, J., Zhang, Z.,Miller, W., and Lipman, D.J. 1997. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs.Nucleic Acids Res.25: 3389–3402.
Ash, C., Priest, F.G., and Collins, M.D. 1993. Molecular identifica-tion of rRNA group 3 bacilli (Ash, Farrow, Wallbanks and Col-lins) using a PCR probe test. Proposal for the creation of a newgenusPaenibacillus. Antonie van Leewenhoek,64: 253–260.
Beatty, P.H. 2000. Investigations of an antifungal antibiotic pro-duced by an environmental isolate identified asPaenibacilluspolymyxa. Ph.D thesis, University of Alberta, Edmonton.
Biemann, K. 1986. Mass spectrometric methods for protein se-quencing. Anal. Chem.58: 1288–1300.
Chakraborty, U., Chakraborty B.N., and Basu, K. 1994. Antago-nism of Erwinia herbicola towards Leptosphaeria maculanscausing blackleg disease ofBrassica napus. Lett. Appl.Microbiol. 18: 74–76.
Difco Manual. 1984. Dehydrated culture media and reagents. 10thed. Difco Laboratories, Detroit, Mich.
Dijksterhuis, J., Sanders, M., Gorris, L.G.M., and Smid, E.J. 1999.Antibiosis plays a role in the context of direct interaction duringantagonism of Paenibacillus polymyxa towards Fusariumoxysporum. J. Appl. Microbiol. 86: 13–21.
Doi, R.H., and McGloughlin, M. 1992. Biology ofBacilli. Appli-cations to industry. Butterworth-Heinemann, Toronto, Ont., Can-ada.
Eckart, K. 1994. Mass spectrometry of cyclic peptides. Mass.Spectrom. Rev.13: 23–55.
Erhard, M., von Dohren, H., and Jungblut, P. 1997. Rapid typingand elucidation of new secondary metabolites of intactcyanobacteria using MALDI-TOF mass spectrometry. Nat.Biotechnol.15: 906–909.
Hauben, L., Vauterin, L., Swings, J., and Moore, E.R.B. 1997.Comparison of 16S ribosomal DNA sequences of allXanthomonasspecies. Int. J. Syst. Bacteriol.47: 328–335.
Hediger, H., Stevens, R.L., Bradenberger, H., and Schmid, K.1973. Determination of asparagine, glutamine andpyrrolidonecarboxylic acid in total enzymic hydrolysates of pep-tides and glycopeptides by gas-liquid chromatography. Biochem.J. 133: 551–561.
Heulin, T., Berge, O., Mavingui, P., Gouzou, L., Hebbar, K.P., andBalandreau, J. 1994.Bacillus polymyxaand Rahnella aquatilis,the dominant N2-fixing bacteria associated with wheatrhizosphere in French soils. Eur. J. Soil Biol.30: 35–42.
Holl, F.B., and Chanway, C.P. 1992. Rhizosphere colonization andseedling growth promotion of lodgepole pine byBacilluspolymyxa. Can. J. Microbiol.38: 303–308.
Holl, F.B., Chanway, C.P., Turkington, R., and Radley, R.A. 1988.Response of crested wheatgrass (Agropyron cristatumL.), pe-rennial ryegrass (Lolium perenne) and white clover (Trifoliumrepens L.) to inoculation with Bacillus polymyxa. Soil Biol.Biochem.20: 19–24.
Holt, J.G., N.R. Krieg, P.H.A. Sneath, J.T. Staley, and S.T.Williams, (Editors). 1994. Bergey’s manual of determinativebacteriology. 9th ed. Williams and Wilkins, Philadelphia, Pa.,U.S.A. pp. 98, 99, and 131.
Ito, M., and Koyama, Y. 1972a. Jolipeptin, a new peptide antibi-otic. Isolation, physico-chemical and biological characteristics.J. Antibiot. 25: 304–308.
Ito, M., and Koyama, Y. 1972b. Jolipeptin, a new peptide antibi-otic. The mode of action of jolipeptin. J. Antibiot.25: 309–314.
Kajimura, Y., and Kaneda, M. 1996. Fusaricidin A, a newdepsipeptide antibiotic produced byBacillus polymyxaKT-8,taxonomy, fermentation, isolation, structure elucidation and bio-logical activity. J. Antibiot.49: 129–135.
Kajimura, Y., and Kaneda, M. 1997. Fusaricidins B, C and D, newdepsipeptide antibiotics produced byBacillus polymyxaKT-8,isolation, structure elucidation and biological activity. J.Antibiot. 50: 220–228.
Kanamori, K., Weiss, R.L., and Roberts, J.D. 1987. Ammonia as-similation in Bacillus polymyxa.J. Biol. Chem.262: 11 038 –11 045.
Kharbanda, P.D. 1993. Blackleg of canola in Alberta: investiga-tions on biology, epidemiology and management. Alberta Envi-ronment Centre, Vegreville, Alberta. Report No. AECV93-R5.
Kharbanda, P.D. 1994. Control of blackleg of canola. Alberta Envi-ronment Centre, Vegreville, Alberta.
Kharbanda, P.D. 1999. (Alberta Environmental Centre.) Personalcommunication.
Kharbanda, P.D., and Dahiya, J.S. 1990. A metabolite ofPenicillium verrucosuminhibitory to growth of LeptosphaeriamaculansandRhizoctonia solani. Can. J. Plant Pathol.12: 335.
Kharbanda, P.D., Yang, J., Beatty, P.H., Tewari, J.P., and Jensen,S.E. Effectiveness of a bacterium (Bacillus sp.) to control black-leg of canola. International Congress on Plant Pathology, August8–13, 1998, Edinburgh, Scotland.
Kropp, K.G., Gondalves, J.A., Andersson, J.T., and Fedorak, P.M.1994. Microbially mediated formation of benzonaphthothio-phenes from benzo[b]thiophenes. Appl. Environ. Microbiol.60:3624–3631.
© 2002 NRC Canada
168 Can. J. Microbiol. Vol. 48, 2002
I:\cjm\cjm48\4802\W02-002.vp27-Feb-2002 10:08:24 AM
Color profile: DisabledComposite Default screen

© 2002 NRC Canada
Beatty and Jensen 169
Kurusu, K., Ohba, K., Arai, T., and Fukushima, K. 1987. New pep-tide antibiotics LI-FO3, FO4, FO5, FO7, and FO8, produced byBacillus polymyxaI. Isolation and characterization. J. Antibiot.40: 1506–1514.
Liang, X.Y., Huang, H.C., Yanke, L.J., and Kozub, G.C. 1996.Control of damping-off of safflower by bacterial seed treatment.Can. J. Plant Pathol.18: 43–49.
Maidak, B.L., Olsen, G.J., Larson, N., Overbeek, R., McCaughey,M.J., and Woese, C.R. 1997. The RDP (ribosomal database pro-ject). Nucleic Acids Res.25: 109–110.
Mavingui, P., and Heulin, T. 1994. In vitro chitinase and antifungalactivity of a soil, rhizosphere and rhizoplane population ofBa-cillus polymyxa. Soil Biol. Biochem.26: 801–803.
Nakajima, N., Chihara, S., and Koyama, Y. 1972. A new antibiotic,gatavalin I. Isolation and characterization. J. Antibiot.25: 243–247.
Nakajima, N., Nakatsugawa, S., Koyama, Y., and Chihara, S. 1975.Gatavalin and process for producing same. U.S. Pat. 3923978.
O’Brien, M.C., and Colwell, R.R. 1987. A rapid test for chitinaseactivity that uses 4-methylumbelliferyl-N-acetyl-β-D-glucosa-minide. Appl. Environ. Microbiol.53: 1718–1720.
Oedjijono, M., Line, A., and Dragar, C. 1993. Isolation of bacteriaantagonistic to a range of plant pathogenic fungi. Soil Biol.Biochem.25: 247–250.
Parry, J.M., Turnbull, P.C.B., and Gibson, J.R. 1983. A colour atlasof Bacillus species. Wolfe Medical Publications, Ipswich, Eng-land.
Paulus, H., and Gray, E. 1964. The biosynthesis of polymyxin B bygrowing cultures ofBacillus polymyxa. J. Biol. Chem.239:865–871.
Peruski, L.F.J., and Peruski, A.H. 1997. The internet and the newbiology: tools for genomic and molecular research. AmericanSociety for Microbiology, Washington, D.C. pp. 52–59.
Petersen, D.J., Srinivasan, M., and Chanway, C.P. 1996.Bacillus
polymyxastimulates increasedRhizobium etlipopulations andnodulation when co-resident in the rhizosphere ofPhaseolusvulgaris. FEMS Microbiol. Lett.142: 271–276.
Pichard, B., Larue, J.P., and Thouvenot, D. 1995. Gavaserin andsaltavalin, new peptide antibiotics produced byBacilluspolymyxa. FEMS Microbiol. Lett.133: 215–218.
Piuri, M., Sanchez-Rivas, C., and Ruzal, S.M. 1998. A novelantimicrobial activity of aPaenibacillus polymyxastrain isolatedfrom regional fermented sausages. Lett. Appl. Microbiol.27: 9–13.
Rosado, A., and Seldin, L. 1993. Production of a potentially novelanti-microbial substance byBacillus polymyxa. World J.Microbiol. Biotechnol.9: 521–528.
Seldin, L., Silva de Azevedo, F., Alviano, D.S., and de FreireBastos, M.C. 1999. Inhibitory activity of Paenibacilluspolymyxa SCE2 against human pathogenic micro-organisms.Lett. Appl. Microbiol. 28: 423–427.
Shishido, M., Massicotte, H.B., and Chanway, C.P. 1996. Effect ofplant growth promotingBacillus strains on pine and spruceseedling growth and mycorrhizal infection. Ann. Bot. (London),77: 433–441.
Shoji, J., Kato, T., and Hinoo, H. 1977. The structure of polymyxinT. J. Antibiot. 30: 1042–1048.
Smid, E.J., van Beek, J.A.M, and Gorris, L.G.M. 1993. Biologicalcontrol of Penicillium hirsutumby antagonistic soil bacteria.InBiological control of foliar post-harvest diseases. Vol. 16.Editedby N.J. Kokkema, J. Kohl, and Y. Elad. IOBC WPRS Bulletin,Wageningen, the Netherlands. pp. 190–193.
Umezawa, H., Takita, T., and Shiba, T. 1978. Bioactive peptidesproduced by microorganisms. John Wiley and Sons, Toronto,Ont., Canada.
I:\cjm\cjm48\4802\W02-002.vp27-Feb-2002 10:08:24 AM
Color profile: DisabledComposite Default screen










