Embed Size (px)
Citation preview

Biochem. J. (2009) 418, 463–473 (Printed in Great Britain) doi:10.1042/BJ20080293 463
p/CAF modulates the activity of the transcription factor p48/Ptf1ainvolved in pancreatic acinar differentiationAnnie RODOLOSSE*†1, Maria-Luisa CAMPOS†‡, Ilse ROOMAN†2, Mathieu LICHTENSTEIN*†3 and Francisco X. REAL*†‡4
*Departament de Ciencies Experimentals i de la Salut, Universitat Pompeu Fabra, Doctor Aiguader 88, 08003 Barcelona, Spain, †Unitat de Biologia Cel·lular i Molecular, InstitutMunicipal d’Investigacio Medica, 08003 Barcelona, Spain, and ‡Programa de Patologıa Molecular, Centro Nacional de Investigaciones Oncologicas, Melchor Fernandez Almagro 3,28029 Madrid, Spain
p48, also called Ptf1a (pancreas-specific transcription factor 1a),is a tissue-restricted bHLH (basic helix–loop–helix) transcriptionfactor which is critical for pancreatic commitment during develop-ment and for the activation and maintenance of the acinardifferentiation programme in the exocrine pancreas. High-levelexpression of exocrine digestive enzymes, a hallmark of matureacinar cells, depends largely on the trimeric complex PTF1,formed by p48, RBP-L (recombination signal-binding protein1-like) and a class A bHLH protein. In addition, p48 inducescell-cycle exit by controlling G1/S-phase progression. However,the mechanisms that mediate PTF1-dependent gene activationare poorly understood. In the present study, we report thatp48 increases transcription through two activation domainslocated in its N-terminal region by recruiting transcriptionalco-activators. The histone acetyltransferase cofactor p/CAF{p300/CBP [CREB (cAMP-response-element-binding protein)-binding protein]-associated factor} interacts with p48 in acinar
cells in vivo and is associated with the promoter regionof acinar genes targeted by the PTF1 complex. p/CAFpotentiates PTF1 transcriptional activity by enhancing selectivelythe p48 transactivation activity. p/CAF promotes the nuclearaccumulation of p48 and its in vivo acetylation in Lys200. TheK200R mutation abolishes the transcriptional activity of p48, aswell as its capacity to functionally co-operate with RBP-L toensure effective PTF1-driven transcription, indicating that p/CAF-mediated acetylation of p48 is required for the full transcriptionalactivity of PTF1. In contrast, p/CAF did not co-operate with p48in its growth regulatory effects. These results support a criticaland selective role of p/CAF in PTF1-dependent gene activationduring acinar differentiation.
Key words: acinar cell, differentiation, p48/PTF1a, pancreas,histone acetyltransferase (HAT), recombination signal-bindingprotein 1-like (RBP-L).
INTRODUCTION
The bHLH (basic helix–loop–helix) transcription factor p48 [alsodesignated Ptf1a (pancreas-specific transcription factor 1a) orPtf1α] exerts critical functions during both early and late pan-creatogenesis: it is required for the acquisition of pancreatic fateby undifferentiated foregut endoderm [1] and is also necessaryfor the activation and maintenance of the acinar differentiationprogramme in the exocrine pancreas. In murine pancreatic buds,p48 expression is detected starting at E9.5 (embryonic day 9.5)[2,3] in a progenitor population that gives rise to all acinar andductal cells, as well as the majority of insulin- and glucagon-producing cells, and becomes restricted to the acinar compart-ment from E14 onwards [1,4]. In the absence of p48, progenitorsfrom the dorsal pancreatic bud give rise to the four differentiatedintestinal cell types [1]. Inactivation of p48 by homologousrecombination in the mouse also results in pancreatic andcerebellar agenesis, as well as in defective specification ofGABA (γ -aminobutyric acid)-ergic neurons in the spinal cordand horizontal/amacrine cells in the developing retina [1,2,5–7]. Asimilar pancreatic and cerebellar phenotype is observed in patients
with permanent neonatal diabetes mellitus harbouring mutationsleading to the expression of a truncated form of p48 lacking theC-terminal 32 amino acids [8]. These observations highlight acrucial role of the C-terminal domain of p48 in pancreatic andneuronal specification. Additionally, the C-terminal region ofp48 has been shown to be responsible for the antiproliferativeactivity of the protein ([9], but see [9a]) and to interact withthe mammalian Suppressor of Hairless [RBP (recombinationsignal-binding protein 1)-Jκ] and its paralogue RBP-L (RBP-like)[10,13].
The establishment of the acinar differentiation programmerelies on the formation of transcriptional complexes thought tobe responsible for the activation of expression of all acinar di-gestive enzymes. Thus tissue-specific transcription of the acinargenes at high levels is largely controlled by the pancreas-specifictranscription complex PTF1 [11,12]. PTF1 is a hetero-oligomericcomplex composed of the ubiquitously expressed class A bHLHproteins E2A or HEB, RBP-Jκ or its paralogue RBP-L [10,13],and p48 [14]. In the adult, RBP-L and Ptf1a are expressed mainlyin acinar cells in the pancreas [4,10]. The PTF1-binding site isa bipartite sequence with an E-box and a TC box (TTTCCCA)
Abbreviations used: bHLH, basic helix–loop–helix; CBP, CREB-binding protein; ChIP, chromatin immunoprecipitation; CMV, cytomegalovirus; CPA1,carboxypeptidase A1; CREB, cAMP-response-element-binding protein; Ct, threshold cycle; CTRB, chymotrypsinogen B; E9.5 etc., embryonic day 9.5etc.; Gal4DBD, Gal4 DNA-binding domain; Gcn5, general control non-derepressible 5; GFP, green fluorescent protein; GST, glutathione transferase;HA, haemagglutinin; HAT, histone acetyltransferase; HPRT, hypoxanthine–guanine phosphoribosyltransferase; KLF6, Kruppel-like factor 6; luc, luciferase;mAb, monoclonal antibody; NP40, Nonidet P40; p/CAF, p300/CREB-binding protein-associated factor; PTF1 etc., pancreas-specific transcription factor1 etc.; RBP, recombination signal-binding protein 1; RBP-L, RBP-like; RT-PCR, reverse transcription-PCR; SV40, simian virus 40; HEK-293T cell, humanembryonic kidney cell expressing the large T-antigen of simian virus 40; TK, thymidine kinase.
1 Present address: Institute for Research in Biomedicine, Parc Cientıfic de Barcelona, Josep Samitier 1-5, 08028 Barcelona, Spain.2 Present address: Diabetes Research Center, Vrije Universiteit Brussel, Laarbeeklaan 103, B-1090 Brussels, Belgium.3 Present address: Institut de Neurociencies, Universitat Autonoma de Barcelona, 08193 Bellaterra, Cerdanyola del Valles, Barcelona, Spain.4 To whom correspondence should be addressed (email [email protected]).
c© The Authors Journal compilation c© 2009 Biochemical Society
www.biochemj.org
Bio
chem
ical
Jo
urn
al

464 A. Rodolosse and others
present in the promoter of all of the acinar digestive enzymes genes[11]. An antisense RNA-mediated reduction of p48 synthesis incultured exocrine pancreatic cells led to inhibition of the exocrinetranscription programme [4], and in vitro acinar-to-ductaltransdifferentiation of normal exocrine pancreas is associated witha loss of PTF1 activity and the selective down-regulation of p48and RBP-L expression ([15] and A. Pinho, I. Rooman and F. X.Real, unpublished work). These findings indicate that p48 andRBP-L are essential for maintaining the terminally differentiatedstate in acinar cells. However, the mechanisms supporting PTF1-mediated gene activation are still unknown.
p/CAF {p300/CBP [CREB (cAMP-response-element-bindingprotein)-binding protein]-associated factor} [16] and the closelyrelated Gcn5 (general control non-derepressible 5) (also calledp/CAF-B) [17] are transcriptional co-activators with intrinsicHAT (histone acetyltransferase) activity which participatein transcriptional regulation by modifying chromatin andtranscription factors. Histone acetylation on lysine residuesis associated predominantly with gene activation, but theprecise mechanism by which this post-translational modi-fication augments transcription is not fully understood. It hasbeen proposed that acetylation would weaken the nucleosomalstructure and the disrupt contacts between nucleosomes, therebyfacilitating the access of transcription factors and the progressionof RNA polymerase II on to chromatin [18]. In human cells,p/CAF is part of a large multi-subunit complex of more than20 polypeptides, including TATA box-binding-protein associatedfactors [19] and may associate with the HATs CBP/p300 [16].It is thought that the combination of HATs ensuring the optimaltranscription of a given promoter may be determined at least inpart by their interaction with specific transcription factors boundto this promoter. In addition to histones, a growing number oftranscription factors have been identified as substrates for p/CAF.They include p53 [20], pRb (retinoblastoma protein) [21] and thebHLH transcription factors MyoD [22,23], Tal1 [24], E2A [25]and Beta2 [26]. p/CAF-mediated acetylation of these transcriptionfactors is required for their function, illustrating the participationof p/CAF in proliferation, apoptosis or differentiation processes.
In the present study, we report on the mechanisms throughwhich p48 contributes to transcriptional activation. We demon-strate that the N-terminal of p48 contains activation domains andwe show that p/CAF, through its interaction with p48, contributesto the transcriptional activity of the PTF1 complex. We demon-strate that p48 is acetylated by p/CAF and that p/CAF-mediatedacetylation is required for both p48 and PTF1 transcriptionalactivity. These results reveal a novel mechanism by which p48function is regulated and support the important contribution ofp48 acetylation in the acinar differentiation programme.
EXPERIMENTAL
Cells and cell culture
We used parental AR42J cells, derived from an azaserine-inducedpancreatic acinar tumour [27] and acinar pancreatic 266-6 cells,derived from a mouse tumour induced with an elastase I/SV40(simian virus 40) T-antigen fusion gene [28]. All cells werecultured in Dulbecco’s modified Eagle’s medium (Life Techno-logies) supplemented with 10% (v/v) fetal bovine serum,L-glutamine, non-essential amino acids, penicillin and strep-tomycin (complete medium).
Plasmids
pIRESp48, pIRESN-bHLH, pIRESN + bHLH, pIRESC-bHLH, pIRESC + bHLH expression vectors and 6XA26-luc
(where luc is luciferase) reporter plasmid constructions havebeen described previously ([9], but see [9a]). Plasmids expressingfull-length rat p48/Ptf1a (amino acids 1–326) or the fragments(residues 1–43, residues 38–108, residues 104–138 andresidues 133–168), N−bHLH (N-terminal construct withoutbHLH) (residues 1–168), N + bHLH (N-terminal constructwith bHLH) (residues 1–217), C−bHLH (C-terminal con-struct without bHLH) (residues 211–326) and C + bHLH(C-terminal construct with bHLH) (residues 162–326) wereconstructed by cloning PCR amplification products frompIRES-p48 into the pBXG1 expression vector in frame with theGal4DBD (Gal4 DNA-binding domain). The pBXG1 expressionvector and the Gal4-TKluc (where TK is thymidine kinase)reporter vector containing four copies of the Gal4DBD upstreamof the TK promoter [29] were obtained from Dr P. Munoz(Differentiation and Cancer Programme, Centre de RegulacioGenomica, Barcelona, Spain). The p48GFP (where GFP is greenfluorescent protein) and pFLAGCMV2 (where CMV is cyto-megalovirus) plasmids were constructed by subcloning the ratPtf1a cDNA into pEGFP-C1 (where EGFP is enhanced GFP;Clontech) and pFLAGCMV2 (Sigma) respectively. The p48K-R(p48K196R, p48K200R, p48K252R, p48K286R, p48K289R, p48K298R, p48K307R
and p48K311R) mutants were generated by PCR-based mutagenesis.The amplification products were cloned into pFLAGCMV2,pBXG1 or pIRESneo (Clontech). pCx-FLAG-pCAF and pCx-FLAG-pCAF (HAT−) expression vectors were gifts fromDr H. Santos-Rosa (The Wellcome Trust/Cancer Research UKGurdon Institute, University of Cambridge, Cambridge, U.K.).pIRES-GFP-p48 and pIRES-GFP-p48K200R were obtained bycloning p48 and p48K200R PCR products into pIRES-hrGFP-2a(Stratagene). pFLAGCMV2-Gcn5 was obtained from ProfessorT. Honjo (Graduate School of Medicine, Kyoto University,Kyoto, Japan). The pcDNA3RBP-L and the pcDNA3E47plasmids were provided by Dr R. MacDonald (University ofTexas Southwestern Medical Center, Dallas, TX, U.S.A.) andDr A. Cano (Departamento de Biologıa del Cancer, Institutode Investigaciones Biomedicas ‘Alberto Sols’, Madrid, Spain)respectively. All constructs were DNA sequenced and the expres-sion of the corresponding proteins was assessed by Westernblotting.
Antibodies
The following antibodies were used: affinity-purified rabbitanti-p48 [15], Gal4DBD mAb (monoclonal antibody) (BDBiosciences), anti-FLAG M2 mAb (Sigma), rabbit polyclonalanti-(acetyl-lysine) antibody (Chemicon) and E2A/E12 rabbitpolyclonal antibody (Santa Cruz Biotechnology). The rabbit poly-clonal anti-p/CAF and the affinity-purified rabbit anti-RBP-Lantibodies were provided by Dr Y. Nakatani (Dana-FarberCancer Institute, Boston, MA, U.S.A.) and Professor R. Wagener(Institute for Biochemistry II, University of Cologne, Germany)respectively.
Transient transfection and reporter gene assays
AR42J cells were transfected with the expression vector andreporter plasmids using LipofectamineTM (Invitrogen). Cells werelysed 24 h after transfection and luciferase activity was deter-mined using the Dual Luciferase Reporter Assay System(Promega). The pRL-TK vector (15 ng) was used as an internalcontrol for normalization.
For transactivation assays, cells were co-transfected with 0.2 μgof plasmid expressing p48 or its deletion mutant forms fused to theGal4DBD and 0.1 μg of Gal4-TKluc reporter plasmid. To deter-mine the co-activation induced by p/CAF, 0.1 μg of p48 and
c© The Authors Journal compilation c© 2009 Biochemical Society

p/CAF modulates p48/Ptf1a activity 465
RBP-L expression vectors or the corresponding empty vectorswere co-transfected with increasing amounts of pCX-FLAG-p/CAF or pCX-FLAG-p/CAF (HAT−) and 0.1 μg of 6XA26-luc. Transcriptional activity assays of wild-type and the lysine-mutated forms (p48K-R) of p48 were performed by transfecting0.2 μg of p48 or p48K-R expression plasmids, 0.2 μg of RBP-Lexpression vector and 0.1 μg of 6XA26-luc.
Immunoprecipitation and Western blotting
HEK-293T cells (human embryonic kidney cells expressing thelarge T-antigen of SV40) were transfected with 3 μg of pIRESp48expression vector and 3 μg of FLAG-p/CAF or FLAG-Gcn5plasmids. After 48 h, cells were lysed in 1 ml of immuno-precipitation buffer [50 mM Tris/HCl (pH 8.0), 120 mMNaCl and 0.5 % NP40 (Nonidet P40)] supplemented withCompleteTM protease inhibitor cocktail and lysates were incubatedovernight. Endogenous p/CAF was immunoprecipitated fromAR42J lysates using an anti-p/CAF antibody and Protein G–agarose beads (Roche). FLAG-tagged p/CAF and Gcn5 wereimmunoprecipitated with anti-FLAG M2 affinity gel (Sigma).The interaction of p48 and p48K200R with RBP-L or E47was analysed in HEK-293T cells transfected with 3 μg ofpFLAG-p48 or pFLAG-p48K200R and 3 μg of pcDNA3RBP-L orpCDNA3E47 expression vectors. FLAG-tagged p48 or p48K200R
were immunoprecipitated with anti-FLAG M2 affinity gel asdescribed above. The immune complexes were pelleted by gentlecentrifugation (3000 g for 10 min), washed three times withimmunoprecipitation buffer, resuspended in loading buffer andfractionated by SDS/PAGE. The following antibodies were usedfor Western blotting: affinity-purified rabbit polyclonal anti-p48 antibody (1:800 dilution), rabbit polyclonal anti-E2A/E12antibody (1:500 dilution), affinity-purified rabbit polyclonal anti-RBP-L antibody (1:300 dilution) and anti-FLAG M2 mAb(1:10000 dilution).
In vitro acetylation assays
FLAG-tagged p48 and N + bHLH proteins were immunoprecip-itated from HEK-293T transfected cell extracts using the anti-FLAG M2 affinity gel. After immunoprecipitation, the beads werewashed with immunoprecipitation buffer, divided into aliquotsand stored at −80 ◦C. For acetylation assays, immunoprecipitatedproteins were incubated with 300 ng of recombinant GST (gluta-thione transferase)–p/CAF HAT domain (Upstate) or GST aloneand 10 nmol of acetyl-CoA (Sigma) in HAT buffer [25 mM Tris/HCl (pH 8.0), 2.5% glycerol, 0.5 mM EDTA and 25 mM KCl]at 30◦C for 1 h. Reactions were stopped by adding sample buffer.Proteins in the input samples were visualized by Coomassie Bluestaining and acetylated proteins were detected by immunoblottingusing an anti-(acetyl-lysine) antibody.
In vivo acetylation assays
HEK-293T cells were transfected with 3 μg of FLAG-p48expression vector and 3 μg of pCX-FLAG-p/CAF or pCX-FLAG-p/CAF (HAT−) plasmids. Acetylation of wild-type and K200Rmutated forms of the p48 N + bHLH fragment was assessed bytransfecting 3 μg of the corresponding expression plasmids and3 μg of pCX-FLAG-p/CAF. Lysates were prepared as describedabove and immunoprecipitation was performed with 2.5 μg ofanti-(acetyl-lysine) antibody and 20 μl of Protein G–agarosebeads. For Western blotting, an anti-FLAG M2 mAb (1:10000dilution) was used. In a reciprocal assay, immunoprecipitation wasperformed using the anti-FLAG affinity gel. Acetylated proteins
were detected with an anti-(acetyl-lysine) antibody (1:1000). Todetect the immunoprecipitated proteins, membranes were strippedand re-probed with an anti-FLAG mAb.
Indirect immunofluorescence
COS-7 cells were seeded on to sterile coverslips and transfectedwith 0.2 μg of p48GFP expression vector and 0.2 μg of pCX-FLAG-p/CAF or pCX-FLAG-p/CAF (HAT−) plasmids. Cellswere fixed for 10 min with 4% (w/v) paraformaldehyde andwashed. After blocking with blocking buffer (1% BSA, 0.1%saponin and 0.1% Triton X-100 in PBS) for 30 min, coverslipswere incubated with an anti-FLAG mAb (1:1000 dilution),washed and incubated with a rhodamine-conjugated anti-mouseantibody (1:200 dilution; Dako). Samples were visualized usinga Leica TCS SP2 confocal microscope.
ChIP (chromatin immunoprecipitation)
Acinar pancreatic 266-6 cells were transfected either with 3 μgof FLAG-p/CAF or with 3 μg of FLAG-tagged p48 usingLipofectamineTM. After 48 h, cells were cross-linked with 1%formaldehyde. Nuclear extracts were prepared by lysis in 50 mMTris/HCl (pH 8.0), 2 mM EDTA, 0.1% NP40 and 10% (v/v)glycerol with protease inhibitors. After centrifugation (3000 g for10 min), the pellet was resuspended in SDS lysis solution [50 mMTris/HCl (pH 8.0), 10 mM EDTA and 1 % SDS] with proteaseinhibitors. Lysates were sonicated and pre-cleared with ProteinG–agarose beads for 3 h at 4 ◦C. One half of the chromatin wasimmunoprecipitated overnight at 4◦C with pre-blocked agarosebeads coupled to anti-FLAG M2 mAb, and the other half withpre-blocked agarose coupled to anti-HA (haemagglutinin) mAb(Sigma). After washing, bound chromatin was eluted in 100 mMsodium carbonate with 1% SDS and proteinase K (1 mg/ml)(Sigma). Cross-linking was reversed at 65 ◦C. After treatmentwith RNase A (0.04 mg/ml) (Roche) and proteinase K, theimmunoprecipitated DNA was purified using the GFX PCR DNApurification kit (Amersham Biosciences). Quantitative SYBRGreen PCR (Applied Biosystems) was performed by comparingthe Ct (threshold cycle) values for the anti-HA antibody with theanti-FLAG antibody immunoprecipitates. Results are expressedas the fold increase in FLAG-immunoprecipitated DNA overHA-immunoprecipitated DNA. Two independent experimentswere performed and all samples were analysed in triplicate. Theprimer sequences used are as follows: CTRB (chymotrypsinogenB)1 promoter, 5′-GCTGGCCACTACCAATGTTC-3′ and 5′-CT-GAGGCTCTTTTATGTCCC-3′; CPA1 (carboxypeptidase A1)promoter, 5′-CCATGGTCAAGGGTGAAAGC-3′ and 5′-TCT-GGGGCCTTTTTAAACAC-3′. The transfection efficiency of266-6 cells was <10%, as determined using a p48GFP plasmidand counting GFP-positive cells.
Quantitative RT-PCR (reverse transcription-PCR) analysis ofelastase, carboxypeptidase and chymotrypsinogen
AR42J cells were transfected with 6 μg of pIRES-GFP-p48, pIRES-GFP-p48K200R or pIRES-GFP empty vector usingLipofectamineTM and after 20 h, GFP-positive cells were isolatedby sorting using a FACSvantage SE (Becton Dickinson). Toexamine the effects of glucocorticoids, AR42J cells were treatedwith 100 nM dexamethasone for 72 h. RNA was extracted andtreated with DNAse I using the DNA-Free kit (Ambion). cDNAgenerated by Taqman reverse transcriptase was analysed byquantitative PCR (Applied Biosystems). The relative Ct values
c© The Authors Journal compilation c© 2009 Biochemical Society
466 A. Rodolosse and others
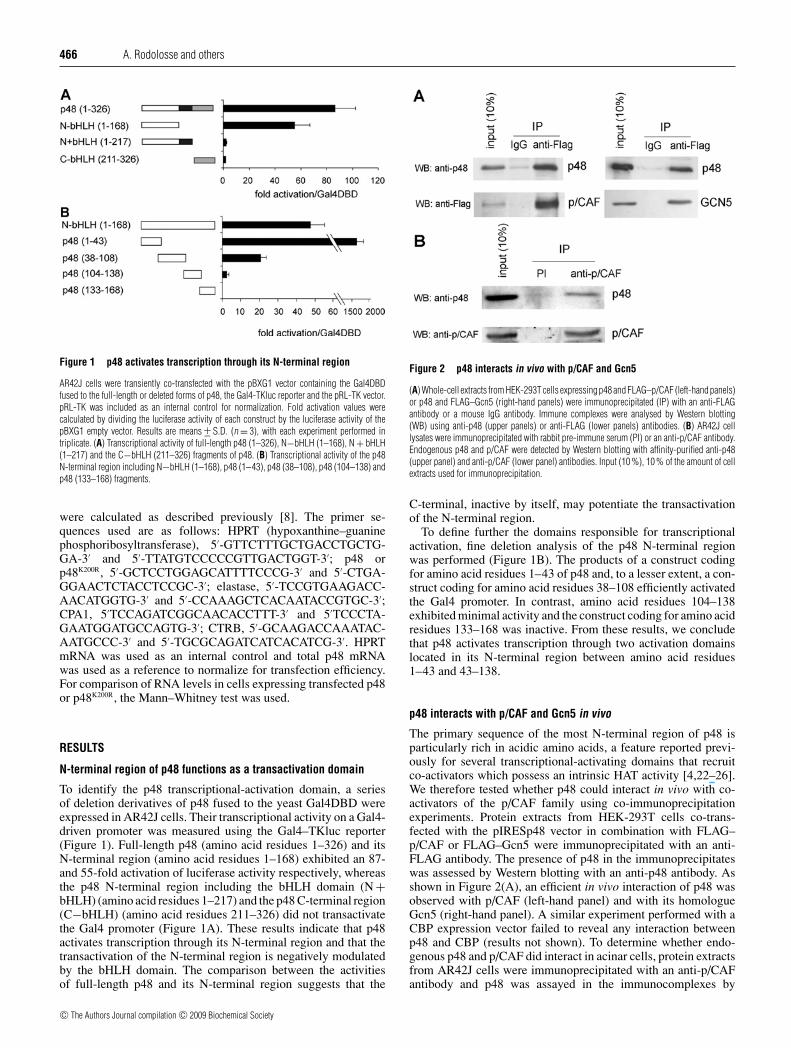
Figure 1 p48 activates transcription through its N-terminal region
AR42J cells were transiently co-transfected with the pBXG1 vector containing the Gal4DBDfused to the full-length or deleted forms of p48, the Gal4-TKluc reporter and the pRL-TK vector.pRL-TK was included as an internal control for normalization. Fold activation values werecalculated by dividing the luciferase activity of each construct by the luciferase activity of thepBXG1 empty vector. Results are means +− S.D. (n = 3), with each experiment performed intriplicate. (A) Transcriptional activity of full-length p48 (1–326), N−bHLH (1–168), N + bHLH(1–217) and the C−bHLH (211–326) fragments of p48. (B) Transcriptional activity of the p48N-terminal region including N−bHLH (1–168), p48 (1–43), p48 (38–108), p48 (104–138) andp48 (133–168) fragments.
were calculated as described previously [8]. The primer se-quences used are as follows: HPRT (hypoxanthine–guaninephosphoribosyltransferase), 5′-GTTCTTTGCTGACCTGCTG-GA-3′ and 5′-TTATGTCCCCCGTTGACTGGT-3′; p48 orp48K200R, 5′-GCTCCTGGAGCATTTTCCCG-3′ and 5′-CTGA-GGAACTCTACCTCCGC-3′; elastase, 5′-TCCGTGAAGACC-AACATGGTG-3′ and 5′-CCAAAGCTCACAATACCGTGC-3′;CPA1, 5′TCCAGATCGGCAACACCTTT-3′ and 5′TCCCTA-GAATGGATGCCAGTG-3′; CTRB, 5′-GCAAGACCAAATAC-AATGCCC-3′ and 5′-TGCGCAGATCATCACATCG-3′. HPRTmRNA was used as an internal control and total p48 mRNAwas used as a reference to normalize for transfection efficiency.For comparison of RNA levels in cells expressing transfected p48or p48K200R, the Mann–Whitney test was used.
RESULTS
N-terminal region of p48 functions as a transactivation domain
To identify the p48 transcriptional-activation domain, a seriesof deletion derivatives of p48 fused to the yeast Gal4DBD wereexpressed in AR42J cells. Their transcriptional activity on a Gal4-driven promoter was measured using the Gal4–TKluc reporter(Figure 1). Full-length p48 (amino acid residues 1–326) and itsN-terminal region (amino acid residues 1–168) exhibited an 87-and 55-fold activation of luciferase activity respectively, whereasthe p48 N-terminal region including the bHLH domain (N +bHLH) (amino acid residues 1–217) and the p48 C-terminal region(C−bHLH) (amino acid residues 211–326) did not transactivatethe Gal4 promoter (Figure 1A). These results indicate that p48activates transcription through its N-terminal region and that thetransactivation of the N-terminal region is negatively modulatedby the bHLH domain. The comparison between the activitiesof full-length p48 and its N-terminal region suggests that the
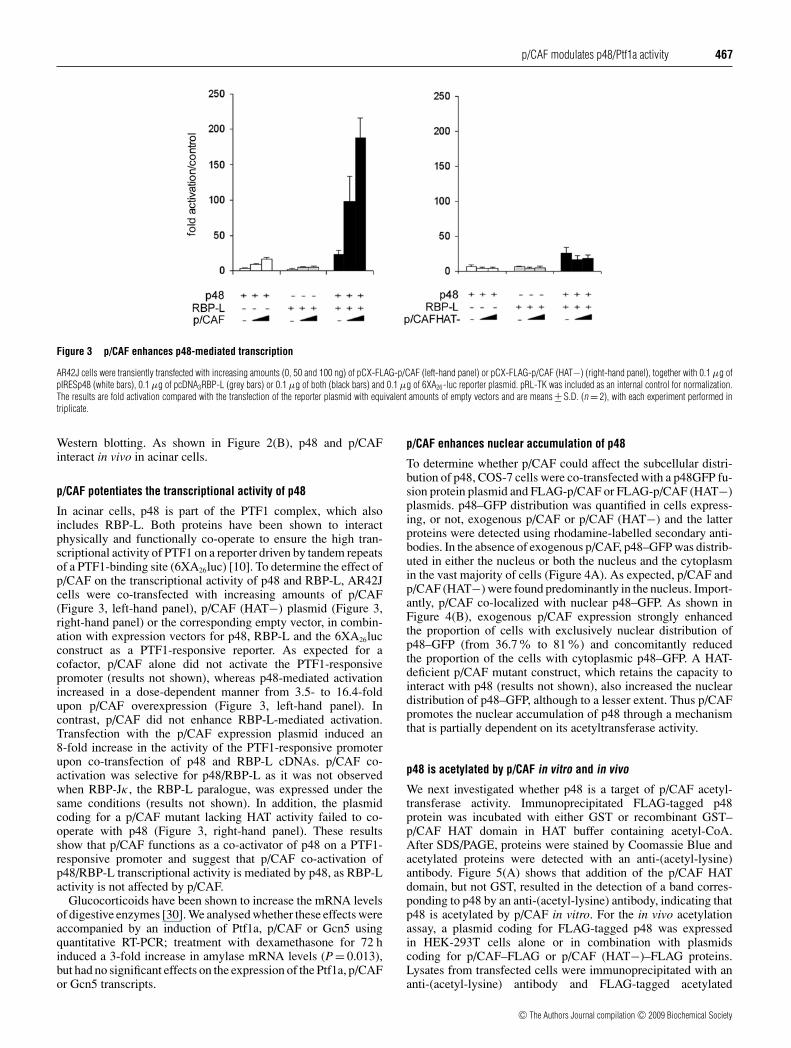
Figure 2 p48 interacts in vivo with p/CAF and Gcn5
(A) Whole-cell extracts from HEK-293T cells expressing p48 and FLAG–p/CAF (left-hand panels)or p48 and FLAG–Gcn5 (right-hand panels) were immunoprecipitated (IP) with an anti-FLAGantibody or a mouse IgG antibody. Immune complexes were analysed by Western blotting(WB) using anti-p48 (upper panels) or anti-FLAG (lower panels) antibodies. (B) AR42J celllysates were immunoprecipitated with rabbit pre-immune serum (PI) or an anti-p/CAF antibody.Endogenous p48 and p/CAF were detected by Western blotting with affinity-purified anti-p48(upper panel) and anti-p/CAF (lower panel) antibodies. Input (10 %), 10 % of the amount of cellextracts used for immunoprecipitation.
C-terminal, inactive by itself, may potentiate the transactivationof the N-terminal region.
To define further the domains responsible for transcriptionalactivation, fine deletion analysis of the p48 N-terminal regionwas performed (Figure 1B). The products of a construct codingfor amino acid residues 1–43 of p48 and, to a lesser extent, a con-struct coding for amino acid residues 38–108 efficiently activatedthe Gal4 promoter. In contrast, amino acid residues 104–138exhibited minimal activity and the construct coding for amino acidresidues 133–168 was inactive. From these results, we concludethat p48 activates transcription through two activation domainslocated in its N-terminal region between amino acid residues1–43 and 43–138.
p48 interacts with p/CAF and Gcn5 in vivo
The primary sequence of the most N-terminal region of p48 isparticularly rich in acidic amino acids, a feature reported previ-ously for several transcriptional-activating domains that recruitco-activators which possess an intrinsic HAT activity [4,22–26].We therefore tested whether p48 could interact in vivo with co-activators of the p/CAF family using co-immunoprecipitationexperiments. Protein extracts from HEK-293T cells co-trans-fected with the pIRESp48 vector in combination with FLAG–p/CAF or FLAG–Gcn5 were immunoprecipitated with an anti-FLAG antibody. The presence of p48 in the immunoprecipitateswas assessed by Western blotting with an anti-p48 antibody. Asshown in Figure 2(A), an efficient in vivo interaction of p48 wasobserved with p/CAF (left-hand panel) and with its homologueGcn5 (right-hand panel). A similar experiment performed with aCBP expression vector failed to reveal any interaction betweenp48 and CBP (results not shown). To determine whether endo-genous p48 and p/CAF did interact in acinar cells, protein extractsfrom AR42J cells were immunoprecipitated with an anti-p/CAFantibody and p48 was assayed in the immunocomplexes by
c© The Authors Journal compilation c© 2009 Biochemical Society
p/CAF modulates p48/Ptf1a activity 467
Figure 3 p/CAF enhances p48-mediated transcription
AR42J cells were transiently transfected with increasing amounts (0, 50 and 100 ng) of pCX-FLAG-p/CAF (left-hand panel) or pCX-FLAG-p/CAF (HAT−) (right-hand panel), together with 0.1 μg ofpIRESp48 (white bars), 0.1 μg of pcDNA3RBP-L (grey bars) or 0.1 μg of both (black bars) and 0.1 μg of 6XA26-luc reporter plasmid. pRL-TK was included as an internal control for normalization.The results are fold activation compared with the transfection of the reporter plasmid with equivalent amounts of empty vectors and are means +− S.D. (n = 2), with each experiment performed intriplicate.
Western blotting. As shown in Figure 2(B), p48 and p/CAFinteract in vivo in acinar cells.
p/CAF potentiates the transcriptional activity of p48
In acinar cells, p48 is part of the PTF1 complex, which alsoincludes RBP-L. Both proteins have been shown to interactphysically and functionally co-operate to ensure the high tran-scriptional activity of PTF1 on a reporter driven by tandem repeatsof a PTF1-binding site (6XA26luc) [10]. To determine the effect ofp/CAF on the transcriptional activity of p48 and RBP-L, AR42Jcells were co-transfected with increasing amounts of p/CAF(Figure 3, left-hand panel), p/CAF (HAT−) plasmid (Figure 3,right-hand panel) or the corresponding empty vector, in combin-ation with expression vectors for p48, RBP-L and the 6XA26lucconstruct as a PTF1-responsive reporter. As expected for acofactor, p/CAF alone did not activate the PTF1-responsivepromoter (results not shown), whereas p48-mediated activationincreased in a dose-dependent manner from 3.5- to 16.4-foldupon p/CAF overexpression (Figure 3, left-hand panel). Incontrast, p/CAF did not enhance RBP-L-mediated activation.Transfection with the p/CAF expression plasmid induced an8-fold increase in the activity of the PTF1-responsive promoterupon co-transfection of p48 and RBP-L cDNAs. p/CAF co-activation was selective for p48/RBP-L as it was not observedwhen RBP-Jκ , the RBP-L paralogue, was expressed under thesame conditions (results not shown). In addition, the plasmidcoding for a p/CAF mutant lacking HAT activity failed to co-operate with p48 (Figure 3, right-hand panel). These resultsshow that p/CAF functions as a co-activator of p48 on a PTF1-responsive promoter and suggest that p/CAF co-activation ofp48/RBP-L transcriptional activity is mediated by p48, as RBP-Lactivity is not affected by p/CAF.
Glucocorticoids have been shown to increase the mRNA levelsof digestive enzymes [30]. We analysed whether these effects wereaccompanied by an induction of Ptf1a, p/CAF or Gcn5 usingquantitative RT-PCR; treatment with dexamethasone for 72 hinduced a 3-fold increase in amylase mRNA levels (P = 0.013),but had no significant effects on the expression of the Ptf1a, p/CAFor Gcn5 transcripts.
p/CAF enhances nuclear accumulation of p48
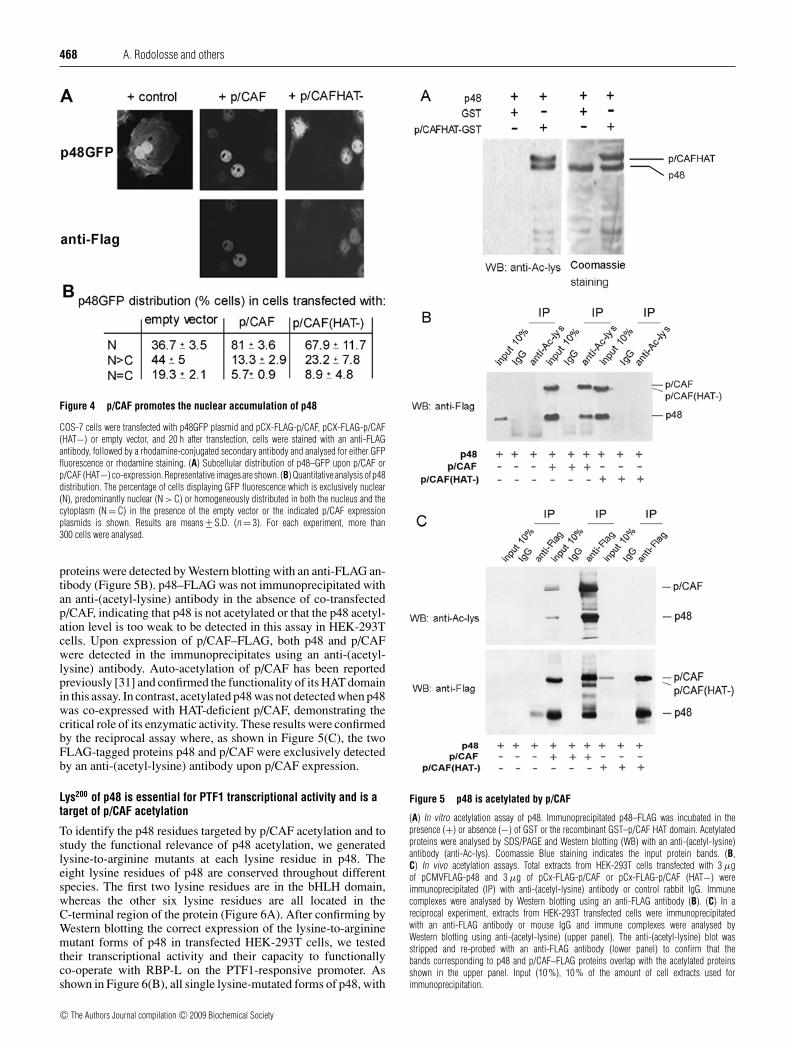
To determine whether p/CAF could affect the subcellular distri-bution of p48, COS-7 cells were co-transfected with a p48GFP fu-sion protein plasmid and FLAG-p/CAF or FLAG-p/CAF (HAT−)plasmids. p48–GFP distribution was quantified in cells express-ing, or not, exogenous p/CAF or p/CAF (HAT−) and the latterproteins were detected using rhodamine-labelled secondary anti-bodies. In the absence of exogenous p/CAF, p48–GFP was distrib-uted in either the nucleus or both the nucleus and the cytoplasmin the vast majority of cells (Figure 4A). As expected, p/CAF andp/CAF (HAT−) were found predominantly in the nucleus. Import-antly, p/CAF co-localized with nuclear p48–GFP. As shown inFigure 4(B), exogenous p/CAF expression strongly enhancedthe proportion of cells with exclusively nuclear distribution ofp48–GFP (from 36.7% to 81%) and concomitantly reducedthe proportion of the cells with cytoplasmic p48–GFP. A HAT-deficient p/CAF mutant construct, which retains the capacity tointeract with p48 (results not shown), also increased the nucleardistribution of p48–GFP, although to a lesser extent. Thus p/CAFpromotes the nuclear accumulation of p48 through a mechanismthat is partially dependent on its acetyltransferase activity.
p48 is acetylated by p/CAF in vitro and in vivo
We next investigated whether p48 is a target of p/CAF acetyl-transferase activity. Immunoprecipitated FLAG-tagged p48protein was incubated with either GST or recombinant GST–p/CAF HAT domain in HAT buffer containing acetyl-CoA.After SDS/PAGE, proteins were stained by Coomassie Blue andacetylated proteins were detected with an anti-(acetyl-lysine)antibody. Figure 5(A) shows that addition of the p/CAF HATdomain, but not GST, resulted in the detection of a band corres-ponding to p48 by an anti-(acetyl-lysine) antibody, indicating thatp48 is acetylated by p/CAF in vitro. For the in vivo acetylationassay, a plasmid coding for FLAG-tagged p48 was expressedin HEK-293T cells alone or in combination with plasmidscoding for p/CAF–FLAG or p/CAF (HAT−)–FLAG proteins.Lysates from transfected cells were immunoprecipitated with ananti-(acetyl-lysine) antibody and FLAG-tagged acetylated
c© The Authors Journal compilation c© 2009 Biochemical Society
468 A. Rodolosse and others
Figure 4 p/CAF promotes the nuclear accumulation of p48
COS-7 cells were transfected with p48GFP plasmid and pCX-FLAG-p/CAF, pCX-FLAG-p/CAF(HAT−) or empty vector, and 20 h after transfection, cells were stained with an anti-FLAGantibody, followed by a rhodamine-conjugated secondary antibody and analysed for either GFPfluorescence or rhodamine staining. (A) Subcellular distribution of p48–GFP upon p/CAF orp/CAF (HAT−) co-expression. Representative images are shown. (B) Quantitative analysis of p48distribution. The percentage of cells displaying GFP fluorescence which is exclusively nuclear(N), predominantly nuclear (N > C) or homogeneously distributed in both the nucleus and thecytoplasm (N = C) in the presence of the empty vector or the indicated p/CAF expressionplasmids is shown. Results are means +− S.D. (n = 3). For each experiment, more than300 cells were analysed.
proteins were detected by Western blotting with an anti-FLAG an-tibody (Figure 5B). p48–FLAG was not immunoprecipitated withan anti-(acetyl-lysine) antibody in the absence of co-transfectedp/CAF, indicating that p48 is not acetylated or that the p48 acetyl-ation level is too weak to be detected in this assay in HEK-293Tcells. Upon expression of p/CAF–FLAG, both p48 and p/CAFwere detected in the immunoprecipitates using an anti-(acetyl-lysine) antibody. Auto-acetylation of p/CAF has been reportedpreviously [31] and confirmed the functionality of its HAT domainin this assay. In contrast, acetylated p48 was not detected when p48was co-expressed with HAT-deficient p/CAF, demonstrating thecritical role of its enzymatic activity. These results were confirmedby the reciprocal assay where, as shown in Figure 5(C), the twoFLAG-tagged proteins p48 and p/CAF were exclusively detectedby an anti-(acetyl-lysine) antibody upon p/CAF expression.
Lys200 of p48 is essential for PTF1 transcriptional activity and is atarget of p/CAF acetylation
To identify the p48 residues targeted by p/CAF acetylation and tostudy the functional relevance of p48 acetylation, we generatedlysine-to-arginine mutants at each lysine residue in p48. Theeight lysine residues of p48 are conserved throughout differentspecies. The first two lysine residues are in the bHLH domain,whereas the other six lysine residues are all located in theC-terminal region of the protein (Figure 6A). After confirming byWestern blotting the correct expression of the lysine-to-argininemutant forms of p48 in transfected HEK-293T cells, we testedtheir transcriptional activity and their capacity to functionallyco-operate with RBP-L on the PTF1-responsive promoter. Asshown in Figure 6(B), all single lysine-mutated forms of p48, with
Figure 5 p48 is acetylated by p/CAF
(A) In vitro acetylation assay of p48. Immunoprecipitated p48–FLAG was incubated in thepresence (+) or absence (−) of GST or the recombinant GST–p/CAF HAT domain. Acetylatedproteins were analysed by SDS/PAGE and Western blotting (WB) with an anti-(acetyl-lysine)antibody (anti-Ac-lys). Coomassie Blue staining indicates the input protein bands. (B,C) In vivo acetylation assays. Total extracts from HEK-293T cells transfected with 3 μgof pCMVFLAG-p48 and 3 μg of pCx-FLAG-p/CAF or pCx-FLAG-p/CAF (HAT−) wereimmunoprecipitated (IP) with anti-(acetyl-lysine) antibody or control rabbit IgG. Immunecomplexes were analysed by Western blotting using an anti-FLAG antibody (B). (C) In areciprocal experiment, extracts from HEK-293T transfected cells were immunoprecipitatedwith an anti-FLAG antibody or mouse IgG and immune complexes were analysed byWestern blotting using anti-(acetyl-lysine) (upper panel). The anti-(acetyl-lysine) blot wasstripped and re-probed with an anti-FLAG antibody (lower panel) to confirm that thebands corresponding to p48 and p/CAF–FLAG proteins overlap with the acetylated proteinsshown in the upper panel. Input (10 %), 10 % of the amount of cell extracts used forimmunoprecipitation.
c© The Authors Journal compilation c© 2009 Biochemical Society
p/CAF modulates p48/Ptf1a activity 469
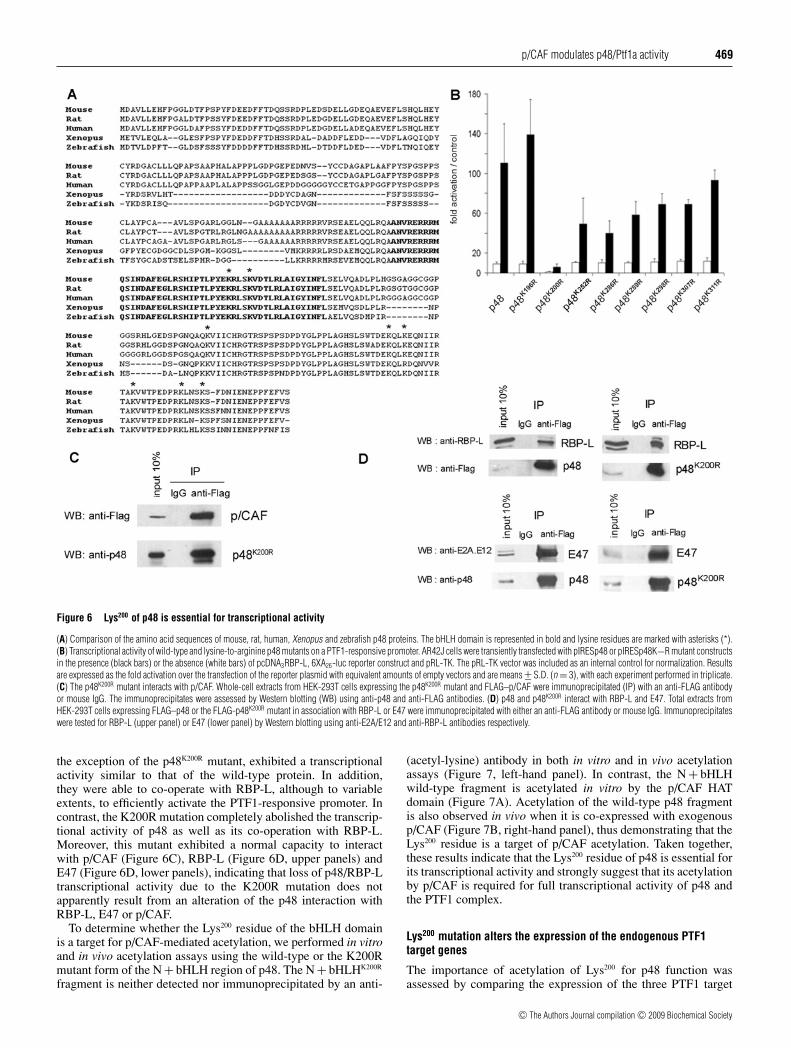
Figure 6 Lys200 of p48 is essential for transcriptional activity
(A) Comparison of the amino acid sequences of mouse, rat, human, Xenopus and zebrafish p48 proteins. The bHLH domain is represented in bold and lysine residues are marked with asterisks (*).(B) Transcriptional activity of wild-type and lysine-to-arginine p48 mutants on a PTF1-responsive promoter. AR42J cells were transiently transfected with pIRESp48 or pIRESp48K−R mutant constructsin the presence (black bars) or the absence (white bars) of pcDNA3RBP-L, 6XA26-luc reporter construct and pRL-TK. The pRL-TK vector was included as an internal control for normalization. Resultsare expressed as the fold activation over the transfection of the reporter plasmid with equivalent amounts of empty vectors and are means +− S.D. (n = 3), with each experiment performed in triplicate.(C) The p48K200R mutant interacts with p/CAF. Whole-cell extracts from HEK-293T cells expressing the p48K200R mutant and FLAG–p/CAF were immunoprecipitated (IP) with an anti-FLAG antibodyor mouse IgG. The immunoprecipitates were assessed by Western blotting (WB) using anti-p48 and anti-FLAG antibodies. (D) p48 and p48K200R interact with RBP-L and E47. Total extracts fromHEK-293T cells expressing FLAG–p48 or the FLAG-p48K200R mutant in association with RBP-L or E47 were immunoprecipitated with either an anti-FLAG antibody or mouse IgG. Immunoprecipitateswere tested for RBP-L (upper panel) or E47 (lower panel) by Western blotting using anti-E2A/E12 and anti-RBP-L antibodies respectively.
the exception of the p48K200R mutant, exhibited a transcriptionalactivity similar to that of the wild-type protein. In addition,they were able to co-operate with RBP-L, although to variableextents, to efficiently activate the PTF1-responsive promoter. Incontrast, the K200R mutation completely abolished the transcrip-tional activity of p48 as well as its co-operation with RBP-L.Moreover, this mutant exhibited a normal capacity to interactwith p/CAF (Figure 6C), RBP-L (Figure 6D, upper panels) andE47 (Figure 6D, lower panels), indicating that loss of p48/RBP-Ltranscriptional activity due to the K200R mutation does notapparently result from an alteration of the p48 interaction withRBP-L, E47 or p/CAF.
To determine whether the Lys200 residue of the bHLH domainis a target for p/CAF-mediated acetylation, we performed in vitroand in vivo acetylation assays using the wild-type or the K200Rmutant form of the N + bHLH region of p48. The N + bHLHK200R
fragment is neither detected nor immunoprecipitated by an anti-
(acetyl-lysine) antibody in both in vitro and in vivo acetylationassays (Figure 7, left-hand panel). In contrast, the N + bHLHwild-type fragment is acetylated in vitro by the p/CAF HATdomain (Figure 7A). Acetylation of the wild-type p48 fragmentis also observed in vivo when it is co-expressed with exogenousp/CAF (Figure 7B, right-hand panel), thus demonstrating that theLys200 residue is a target of p/CAF acetylation. Taken together,these results indicate that the Lys200 residue of p48 is essential forits transcriptional activity and strongly suggest that its acetylationby p/CAF is required for full transcriptional activity of p48 andthe PTF1 complex.
Lys200 mutation alters the expression of the endogenous PTF1target genes
The importance of acetylation of Lys200 for p48 function wasassessed by comparing the expression of the three PTF1 target
c© The Authors Journal compilation c© 2009 Biochemical Society
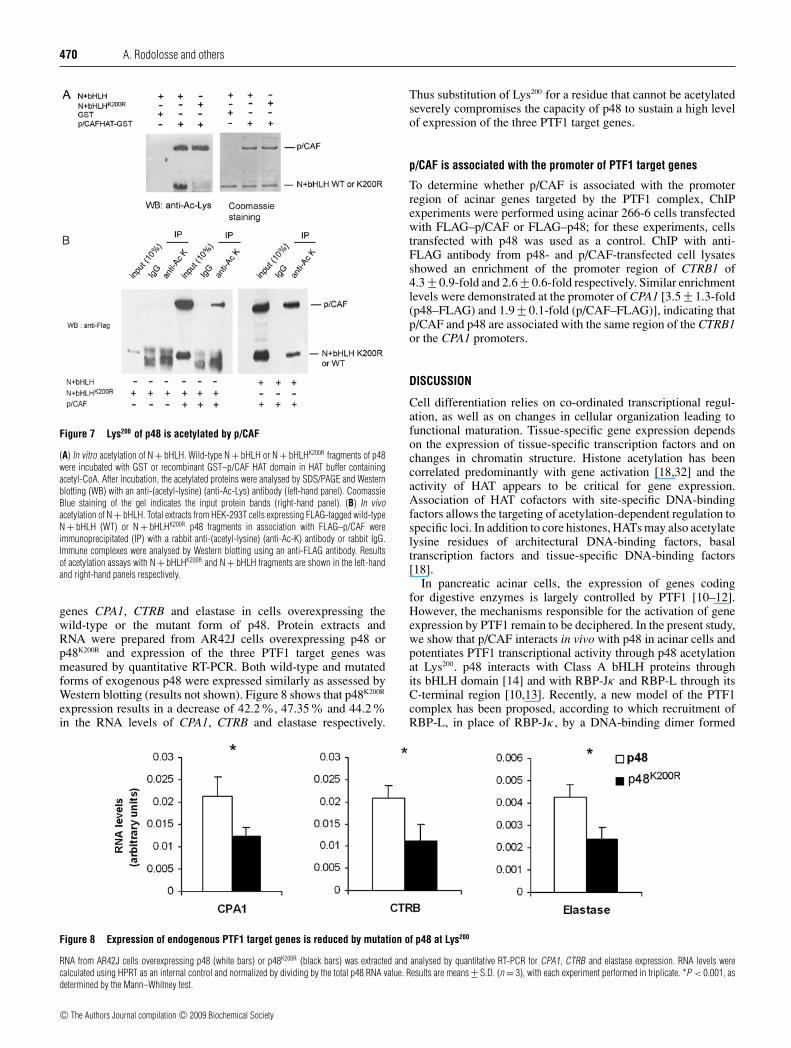
470 A. Rodolosse and others
Figure 7 Lys200 of p48 is acetylated by p/CAF
(A) In vitro acetylation of N + bHLH. Wild-type N + bHLH or N + bHLHK200R fragments of p48were incubated with GST or recombinant GST–p/CAF HAT domain in HAT buffer containingacetyl-CoA. After incubation, the acetylated proteins were analysed by SDS/PAGE and Westernblotting (WB) with an anti-(acetyl-lysine) (anti-Ac-Lys) antibody (left-hand panel). CoomassieBlue staining of the gel indicates the input protein bands (right-hand panel). (B) In vivoacetylation of N + bHLH. Total extracts from HEK-293T cells expressing FLAG-tagged wild-typeN + bHLH (WT) or N + bHLHK200R p48 fragments in association with FLAG–p/CAF wereimmunoprecipitated (IP) with a rabbit anti-(acetyl-lysine) (anti-Ac-K) antibody or rabbit IgG.Immune complexes were analysed by Western blotting using an anti-FLAG antibody. Resultsof acetylation assays with N + bHLHK200R and N + bHLH fragments are shown in the left-handand right-hand panels respectively.
genes CPA1, CTRB and elastase in cells overexpressing thewild-type or the mutant form of p48. Protein extracts andRNA were prepared from AR42J cells overexpressing p48 orp48K200R and expression of the three PTF1 target genes wasmeasured by quantitative RT-PCR. Both wild-type and mutatedforms of exogenous p48 were expressed similarly as assessed byWestern blotting (results not shown). Figure 8 shows that p48K200R
expression results in a decrease of 42.2%, 47.35% and 44.2%in the RNA levels of CPA1, CTRB and elastase respectively.
Thus substitution of Lys200 for a residue that cannot be acetylatedseverely compromises the capacity of p48 to sustain a high levelof expression of the three PTF1 target genes.
p/CAF is associated with the promoter of PTF1 target genes
To determine whether p/CAF is associated with the promoterregion of acinar genes targeted by the PTF1 complex, ChIPexperiments were performed using acinar 266-6 cells transfectedwith FLAG–p/CAF or FLAG–p48; for these experiments, cellstransfected with p48 was used as a control. ChIP with anti-FLAG antibody from p48- and p/CAF-transfected cell lysatesshowed an enrichment of the promoter region of CTRB1 of4.3 +− 0.9-fold and 2.6 +− 0.6-fold respectively. Similar enrichmentlevels were demonstrated at the promoter of CPA1 [3.5 +− 1.3-fold(p48–FLAG) and 1.9 +− 0.1-fold (p/CAF–FLAG)], indicating thatp/CAF and p48 are associated with the same region of the CTRB1or the CPA1 promoters.
DISCUSSION
Cell differentiation relies on co-ordinated transcriptional regul-ation, as well as on changes in cellular organization leading tofunctional maturation. Tissue-specific gene expression dependson the expression of tissue-specific transcription factors and onchanges in chromatin structure. Histone acetylation has beencorrelated predominantly with gene activation [18,32] and theactivity of HAT appears to be critical for gene expression.Association of HAT cofactors with site-specific DNA-bindingfactors allows the targeting of acetylation-dependent regulation tospecific loci. In addition to core histones, HATs may also acetylatelysine residues of architectural DNA-binding factors, basaltranscription factors and tissue-specific DNA-binding factors[18].
In pancreatic acinar cells, the expression of genes codingfor digestive enzymes is largely controlled by PTF1 [10–12].However, the mechanisms responsible for the activation of geneexpression by PTF1 remain to be deciphered. In the present study,we show that p/CAF interacts in vivo with p48 in acinar cells andpotentiates PTF1 transcriptional activity through p48 acetylationat Lys200. p48 interacts with Class A bHLH proteins throughits bHLH domain [14] and with RBP-Jκ and RBP-L through itsC-terminal region [10,13]. Recently, a new model of the PTF1complex has been proposed, according to which recruitment ofRBP-L, in place of RBP-Jκ , by a DNA-binding dimer formed
Figure 8 Expression of endogenous PTF1 target genes is reduced by mutation of p48 at Lys200
RNA from AR42J cells overexpressing p48 (white bars) or p48K200R (black bars) was extracted and analysed by quantitative RT-PCR for CPA1, CTRB and elastase expression. RNA levels werecalculated using HPRT as an internal control and normalized by dividing by the total p48 RNA value. Results are means +− S.D. (n = 3), with each experiment performed in triplicate. *P < 0.001, asdetermined by the Mann–Whitney test.
c© The Authors Journal compilation c© 2009 Biochemical Society

p/CAF modulates p48/Ptf1a activity 471
by the p48/Class A bHLH protein, provides high transcriptionalactivity in the adult pancreas [10,13]. The recruitment of RBP-Lwould result in ensuring the formation of a PTF1 complex with thehighest level of transcriptional activity by excluding RBP-Jκ [10],leading to full acinar cell differentiation. In the present study, weshow that mutation of the p/CAF-acetylated residue Lys200 resultsin the loss of p48 transcriptional activity and of its functional co-operation with RBP-L without affecting its capacity to interactwith RBP-L and the class A bHLH protein E47 (Figure 6C) orp/CAF (Figure 6D). We propose that p/CAF-mediated acetylationof p48 on Lys200 could, by favouring the selective co-operationbetween RBP-L and p48, help to confer to the PTF1 complex thehigh and specific activity required for full differentiation of acinarcells.
p/CAF has been reported to interact with, and to regulatethe function of, several bHLH transcription factors required forcell fate specification or terminal differentiation. For example,MyoD interacts with, and is acetylated by, p/CAF and botheffects are necessary for its transcriptional activity on chromatin-associated templates and for triggering terminal myogenic differ-entiation [22,33]. Beta2/NeuroD, which is essential for the dif-ferentiation of pancreatic islet and neural cells, is an in vivotarget of p/CAF acetylation in pancreatic β-cells. Acetylationof Beta2 affected its transcriptional activity on the insulin genepromoter, supporting the idea that gene activation by Beta2 isregulated by acetylation [26]. The close relation between p/CAF-mediated bHLH transcriptional regulation and cell differentiationis further illustrated by Tal1, a critical bHLH regulator ofhaematopoietic and vascular development. Inhibition of p/CAF-mediated Tal1 acetylation in erythroleukaemia cells resulted inan inhibition of Tal1 transcriptional activity and compromiseddifferentiation [24]. As reported for Beta2 and Tal1, p48 isacetylated in vivo by p/CAF on a lysine residue located in thebHLH domain. Several lysine residues are found at conservedpositions in the helix 2 region of bHLH factors and, in a fewcases, have been demonstrated to be acetylated by p/CAF andessential for transcriptional activation: KRLSKVDTLRLA (p48),KKLSKNEILRLA (Tal1), QKLSKIETLRLA (Beta2/NeuroD),KKLSKYETLQMA (Math1), KKLSKVETLRSA (Mash2),KKLSKIETLTLA (Mist1) and AKLTKVETLRFA (neuro-genin 3) (with the relevant lysine residues underlined).
A comparison of these sequences indicates that p/CAF acetyl-ation takes place in a highly conserved region of bHLH proteins. Itis conceivable that the activity of other bHLH factors required forneuronal and/or pancreatic differentiation, such as Mash2 [34],Mist1 [35,36] or neurogenin 3 [37], could be similarly regulatedby p/CAF acetylation. Whether HAT-mediated acetylation of p48is required for its function in pancreatic and cerebellar organo-genesis remains to be determined.
Despite the fact that the K200R mutation impairs the acetylationof the N-terminal region of p48, the p48K200R mutant is immuno-precipitated by an anti-(acetyl-lysine) antibody upon p/CAF co-transfection (results not shown), supporting the notion that addi-tional lysine residues are targeted by p/CAF in the full-lengthprotein. Interestingly, five out of the eight potentially acetylatedlysine residues are in the far C-terminal part of p48, close tothe interaction domains with RBP-Jκ and RBP-L [10], within aregion shown to be crucial for pancreatic and cerebellar organo-genesis [8] and for p48 anti-proliferative activity ([9], but see[9a]). We have previously described that p48-mediated inhibitionof proliferation is associated with the up-regulation of p21 mRNA,the down-regulation of cyclin D2 mRNA levels and increasedp27 protein levels. Up-regulation of p21 mRNA levels by MyoDhas been shown to depend on the MyoD–p/CAF interactionand p/CAF-mediated acetylation in differentiated muscle cells
[38,39]. Up-regulation of p21 mRNA levels by KLF6 (Kruppel-like factor 6) also required KLF6 acetylation by p/CAF and CBPcofactors [40]. To determine the contribution of p/CAF to theeffects of p48 on cell proliferation, we used cell-cycle analysis,p21 promoter reporter assays and p21 quantitative RT-PCR.p/CAF did not co-operate with p48 in any of these assays (seeSupplementary Figures S1 and S2 at http://www.BiochemJ.org/bj/418/bj4180463add.htm). We cannot rule out a contribution ofendogenous p/CAF; however, these findings indicate a distinctrole of p/CAF in enhancing the transcriptional activation ofacinar genes and other p48-mediated cellular effects. Whetherp/CAF plays a role in a p48-dependent recovery from injury uponinduction of experimental acute pancreatitis also merits furtherstudy [41].
Regarding the mechanisms through which p/CAF contributesto enhanced transcriptional activity, we show that in COS-7 cells,p/CAF promotes the nuclear accumulation of p48, an effectthat has been reported for other transactivators [42]. ShouldHAT activity be required for nuclear targeting or retentionof p48, expression of the p/CAF (HAT−) mutant would notmodify the subcellular distribution of p48 or might competewith endogenous p/CAF, resulting in an increase in cytoplasmicp48. However, we observed that p/CAF (HAT−) also leads,although to a lesser extent, to nuclear p48 accumulation,indicating that HAT activity is not fully required for thiseffect. We therefore propose that the p/CAF-mediated effecton the subcellular distribution of p48 relies mainly on itsinteraction with p48, rather than on its capacity to acetylate it.However, we cannot exclude the possibility that modulation ofp48 distribution results from an indirect effect of p/CAF. It isthought that E2A could, by interacting with p48, be responsiblefor its nuclear import [43]. Obata et al. [3] reported that, inCOS-7 cells, overexpression of E47 (produced from the E2Agene) shifted the predominant localization of Myc-tagged p48from the cytoplasm to the nucleus [3]. However, using a differenttag, we have found that the distribution of GFP–p48 in COS-7cells is predominantly nuclear in 80% of the cells. Additionally,overexpression of E47 or HEB in acinar and non-acinar culturedcells had minor effects on the subcellular localization of p48(M. Lichtenstein, unpublished work). The importance of analys-ing the mechanisms responsible for the nuclear targeting of p48 isunderlined by the study of acinar tumours. We have reported thatin some acinar pancreatic tumours, endogenous p48 was foundexclusively in the cytoplasm and was associated with a reducedexpression of amylase [15]. p48 is also redistributed partiallyto the cytoplasm in Elas CCK2 transgenic mice in associationwith defects in acinar cell differentiation [44]. These findingssuggest that altered subcellular distribution of p48 may contributeto reduced expression of acinar genes and compromised exocrinefunction. Additional experiments need to be performed, usingp/CAF-null mice [45], to determine the p/CAF contribution to thenuclear localization and transcriptional activity of p48.
To assess the role of p/CAF in cell differentiation further,knockdown RNA interference experiments were conducted incultured acinar cells. However, we could not detect any effect,possibly because of a redundancy with Gcn5, a HAT protein, ashas been suggested to be the case in p/CAF-null mice [45]. Gcn5displays a very high amino acid identity with p/CAF and is alsoexpressed in the pancreas (A. Rodolosse, I. Rooman and F.X.Real, unpublished work). Therefore it is possible that the resultsreported for p/CAF may be extended to Gcn5.
Altogether, the present study provides new insights into themechanisms through which the PTF1 complex mediates acinargene expression during exocrine differentiation. Deciphering themechanisms by which p/CAF and/or other HAT cofactors may
c© The Authors Journal compilation c© 2009 Biochemical Society

472 A. Rodolosse and others
regulate p48 functions is an important step toward the understand-ing of acinar differentiation and fate determination in the pancreasand in the nervous system. In addition, it may contribute to a betterunderstanding of the role of acinar transcriptional regulators indisease processes, such as chronic pancreatitis and pancreaticcancer [46].
ACKNOWLEDGEMENTS
We thank Dr R. MacDonald (University of Texas Southwestern Medical Center, Dallas,TX, U.S.A.) for the pcDNA3RBP-L plasmid; Dr A. Cano (Departamento de Biologıa delCancer, Instituto de Investigaciones Biomedicas ‘Alberto Sols’, Madrid, Spain) for thepcDNA3E47 plasmid; Dr P. Munoz (Differentiation and Cancer Programme, Centre deRegulacio Genomica, Barcelona, Spain) for the pBXG1 and Gal4-TKluc vectors; Dr H.Santos-Rosa (The Wellcome Trust/Cancer Research UK Gurdon Institute, University ofCambridge, Cambridge, U.K.) for the pCx-FLAG-pCAF and pCx-FLAG-pCAF (HAT−)expression vectors; Professor T. Honjo (Graduate School of Medicine, Kyoto University,Kyoto, Japan) for the pFLAGCMV2-Gcn5 vector; Dr Y. Nakatani (Dana-Farber CancerInstitute, Boston, MA, U.S.A.) for the rabbit polyclonal anti-p/CAF antibody; and ProfessorR. Wagener (Institute for Biochemistry II, University of Cologne, Germany) for the affinity-purified rabbit anti-RBP-L antibody. We also thank Dr R. MacDonald for discussion ofunpublished work, and S. Peiro and F. Mateo for valuable advice for ChIP experimentsand acetylation assays.
FUNDING
This work was supported in part by the Ministerio de Ciencia y Tecnologıa (PlanNacional de I + D) [grant numbers GEN2001-4748-C01, SAF2004-01137]; the Institutode Salud Carlos III, Ministerio de Sanidad [grant number C03/010]; and the ComissioInterdepartamental de Recerca i Tecnologia, Govern De Catalunya [grant number SGR-00410]. M. L. C. was supported by the Fundacao para a Ciencia e Tecnologia, Portugal[grant number SFRH/BD/17661/2004; POCI2010] and I. R. was the recipient of the MarieCurie Intra-European fellowship.
REFERENCES
1 Kawaguchi, Y., Cooper, B., Gannon, M., Ray, M., MacDonald, R. J. and Wright, C. V. E.(2002) The role of the transcriptional regulator Ptf1a in converting intestinal to pancreaticprogenitors. Nat. Genet. 32, 128–134
2 Krapp, A., Knofler, M., Ledermann, B., Burki, K., Berney, C., Zoerkler, N., Hagenbuchle, O.and Wellauer, P. K. (1998) The bHLH protein PTF1-p48 is essential for the formation ofthe exocrine and the correct spatial organization of the endocrine pancreas. Genes Dev.12, 3752–3763
3 Obata, J., Yano, M., Mimura, H., Goto, T., Nakayama, R., Mibu, Y., Oka, C. andKawaichi, M. (2001) p48 subunit of mouse PTF1 binds to RBP-Jκ/CBF-1, theintracellular mediator of Notch signalling, and is expressed in the neural tube of earlystage embryos. Genes Cells 6, 345–360
4 Krapp, A., Knofler, M., Frutiger, S., Hughes, G. J., Hagenbuchle, O. and Wellauer, P. K.(1996) The p48 DNA-binding subunit of transcription factor PTF1 is a new exocrinepancreas-specific basic helix-loop-helix protein. EMBO J. 15, 4317–4329
5 Hoshino, M., Nakamura, S., Mori, K., Kawauchi, T., Terao, M., Nishimura, Y. V.,Fukuda, A., Fuse, T., Matsuo, N., Sone, M. et al. (2005) Ptf1a, a bHLH transcriptionalgene, defines GABAergic neuronal fates in cerebellum. Neuron 47, 201–213
6 Fujitani, Y., Fujitani, S., Luo, H., Qiu, F., Burlison, J., Long, Q., Kawaguchi, Y., Edlund, H.,MacDonald, R. J., Furukawa, T. et al. (2006) Ptf1a determines horizontal and amacrinecell fates during mouse retinal development. Development 133, 4439–4450
7 Pascual, M., Abasolo, I., Mingorance-Le Meur, A., Martınez, A., Del Rio, J. A., Wright,C. V., Real, F. X. and Soriano, E. (2007) Cerebellar GABAergic progenitors adopt anexternal granule cell-like phenotype in the absence of Ptf1a transcription factorexpression. Proc. Natl. Acad. Sci. U.S.A. 104, 5193–5198
8 Sellick, G. S., Barker, K. T., Stolte-Dijkstra, I., Fleischmann, C., Coleman, R. J., Garrett, C.,Gloyn, A. L., Edghill, E. L., Hattersley, A. T., Wellauer, P. K. et al. (2004) Mutations inPTF1A cause pancreatic and cerebellar agenesis. Nat. Genet. 36, 1301–1305
9 Rodolosse, A., Chalaux, E., Adell, T., Hagege, H., Skoudy, A. and Real, F. X. (2004)PTF1α/p48 transcription factor couples proliferation and differentiation in the exocrinepancreas. Gastroenterology 127, 937–949
9a Erratum (2004) Gastroenterology 127, 165110 Beres, T. M., Masui, T., Swift, G. H., Shi, L., Henke, R. M. and MacDonald, R. J. (2006)
PTF1 is an organ-specific and notch-independent basic helix-loop-helix complexcontaining the mammalian suppressor of hairless (RBP-J) or its paralogue, RBP-L.Mol. Cell. Biol. 26, 117–130
11 Cockell, M., Stevenson, B. J., Strubin, M., Hagenbuchle, O. and Wellauer, P. K. (1989)Identification of a cell-specific DNA-binding activity that interacts with a transcriptionalactivator of genes expressed in the acinar pancreas. Mol. Cell. Biol. 9, 2464–2476
12 Rose, S. D., Swift, G. H., Peyton, M. J., Hammer, R. E. and MacDonald, R. J. (2001) Therole of PTF1-P48 in pancreatic acinar gene expression. J. Biol. Chem. 276, 44018–44026
13 Masui, T., Long, Q., Beres, T. M., Magnusson, M. A. and MacDonald, R. J. (2007) Earlypancreatic development requires the vertebrate suppressor of hairless (RBPJ) in the PTF1bHLH complex. Genes Dev. 21, 2629–2643
14 Roux, E., Strubin, M., Hagenbuchle, O. and Wellauer, P. K. (1989) The cell-specifictranscription factor Ptf1 contains 2 different subunits that interact with the DNA. GenesDev. 3, 1613–1624
15 Adell, T., Gomez-Cuadrado, A., Skoudy, A., Pettengill, O. S., Longnecker, D. S. and Real,F. X. (2000) Role of the basic helix-loop-helix transcription factor p48 in the differentiationphenotype of exocrine pancreas cancer cells. Cell Growth Differ. 11, 137–147
16 Yang, X. J., Ogryzko, V. V., Nishikawa, J., Howard, B. H. and Nakatani, Y. (1996) Ap300/CBP-associated factor that competes with the adenoviral oncoprotein E1A. Nature382, 319–324
17 Smith, E. R., Belote, J. M., Schiltz, R. L., Yang, X. J., Moore, P. A., Berger, S. L.,Nakatani, Y. and Allis, C. D. (1998) Cloning of Drosophila GCN5: conserved featuresamong metazoan GCN5 family members. Nucleic Acids Res. 26, 2948–2954
18 Bannister, A. J. and Miska, E. A. (2000) Regulation of gene expression by transcriptionfactor acetylation. Cell. Mol. Life Sci. 57, 1184–1192
19 Ogryzko, V. V., Kotani, T., Zhang, X. L., Schiltz, R. L., Howard, T., Yang, X. J., Howard,B. H., Qin, J. and Nakatani, Y. (1998) Histone-like TAFs within the PCAF histone acetylasecomplex. Cell 94, 35–44
20 Gu, W. and Roeder, R. G. (1997) Activation of p53 sequence-specific DNA binding byacetylation of the p53 C-terminal domain. Cell 90, 595–606
21 Nguyen, D. X., Baglia, L. A., Huang, S. M., Baker, C. M. and McCance, D. J. (2004)Acetylation regulates the differentiation-specific functions of the retinoblastoma protein.EMBO J. 23, 1609–1618
22 Dilworth, F. J., Seaver, K. J., Fishburn, A. L., Htet, S. L. and Tapscott, S. J. (2004) In vitrotranscription system delineates the distinct roles of the coactivators pCAF and p300during MyoD/E47-dependent transactivation, Proc. Natl. Acad. Sci. U.S.A. 101,11593–11598
23 Sartorelli, V., Puri, P. L., Hamamori, Y., Ogryzko, V., Chung, G., Nakatani, Y., Wang, J. Y. J.and Kedes, L. (1999) Acetylation of MyoD directed by PCAF is necessary for theexecution of the muscle program. Mol. Cell 4, 725–734
24 Huang, S. M., Qiu, Y., Shi, Y. B., Xu, Z. X. and Brandt, S. J. (2000) P/CAF-mediatedacetylation regulates the function of the basic helix-loop-helix transcription factorTAL1/SCL. EMBO J. 19, 6792–6803
25 Bradney, C., Hjelmeland, M., Komatsu, Y., Yoshida, M., Yao, T. P. and Zhuang, Y. (2003)Regulation of E2A activities by histone acetyltransferases in B lymphocyte development.J. Biol. Chem. 278, 2370–2376
26 Qiu, Y., Guo, M., Huang, S. M. and Stein, R. (2004) Acetylation of the BETA2 transcriptionfactor by p300-associated factor is important in insulin gene expression. J. Biol. Chem.279, 9796–9802
27 Jessop, N. W. and Hay, R. J. (1980) Characteristics of two rat pancreatic exocrine celllines from transplantable tumors. In Vitro 16, 212
28 Ornitz, D. M., Palmiter, R. D., Messing, A., Hammer, R. E., Pinkert, C. A. and Brinster, R. L.(1985) Elastase I promoter directs expression of human growth hormone and SV40 Tantigen genes to pancreatic acinar cells in transgenic mice. Cold Spring Harb. Symp.Quant. Biol. 50, 399–409
29 Chakravarti, D., LaMorte, V. J., Nelson, M. C., Nakajima, T., Schulman, I. G., Juguilon, H.,Montminy, M. and Evans, R. M. (1996) Role of CBP/P300 in nuclear receptor signalling.Nature 383, 99–103
30 Logsdon, C. D., Moessner, J., Williams, J. A. and Goldfine, I. D. (1985) Glucocorticoidsincrease amylase mRNA levels, secretory organelles, and secretion in pancreatic acinarAR42J cells. J. Cell Biol. 100, 1200–1208
31 Santos-Rosa, H., Valls, E., Kouzarides, T. and Martinez-Balbas, M. (2003) Mechanisms ofP/CAF auto-acetylation. Nucleic Acids Res. 31, 4285–4292
32 Kuo, M. H. and Allis, C. D. (1998) Roles of histone acetyltransferases and deacetylases ingene regulation. Bioessays 20, 615–626
33 Puri, P. L., Sartorelli, V., Yang, X. J., Hamamori, Y., Ogryzko, V. V., Howard, B. H.,Kedes, L., Wang, J. Y., Graessmann, A., Nakatani, Y. and Levrero, M. (1997) Differentialroles of p300 and PCAF acetyltransferases in muscle differentiation. Mol. Cell 1, 35–45
34 Johnson, J. E., Birren, S. J. and Anderson, D. J. (1990) Two rat homologs of Drosophilaachaete-scute specifically expressed in neuronal precursors. Nature 346, 858–861
35 Lemercier, C., To, R. Q., Swanson, B. J., Lyons, G. E. and Konieczny, S. F. (1997) Mist1:A novel basic helix-loop-helix transcription factor exhibits a developmentally regulatedexpression pattern. Dev. Biol. 182, 101–113
c© The Authors Journal compilation c© 2009 Biochemical Society

p/CAF modulates p48/Ptf1a activity 473
36 Pin, C. L., Rukstalis, J. M., Johnson, C. and Konieczny, S. F. (2001) The bHLHtranscription factor Mist1 is required to maintain exocrine pancreas cell organization andacinar cell identity. J. Cell Biol. 155, 519–530
37 Apelqvist, A., Li, H., Sommer, L., Beatus, P., Anderson, D. J., Honjo, T., de Angelis, M. H.,Lendahl, U. and Edlund, H. (1999) Notch signalling controls pancreatic celldifferentiation. Nature 400, 877–881
38 Mal, A., Sturniolo, M., Schiltz, R. L., Ghosh, M. K. and Harter, M. L. (2001) A role forhistone deacetylase HDAC1 in modulating the transcriptional activity of MyoD: inhibitionof the myogenic program. EMBO J. 20, 1739–1753
39 Puri, P. L., Avantaggiati, M. L., Balsano, C., Sang, N. L., Graessmann, A., Giordano, A.and Levrero, M. (1997) p300 is required for MyoD-dependent cell cycle arrest andmuscle-specific gene transcription. EMBO J. 16, 369–383
40 Li, D., Yea, S., Dolios, G., Martignetti, J. A., Narla, G., Wang, R., Walsh, M. J. andFriedman, S. L. (2005) Regulation of Kruppel-like factor 6 tumor suppressor activity byacetylation. Cancer Res. 65, 9216–9225
41 Molero, X., Adell, T., Skoudy, A., Padilla, M. A., Gomez, J. A., Chalaux, E., Malagelada,J. R. and Real, F. X. (2007) Pancreas transcription factor 1α expression is regulated inpancreatitis. Eur. J. Clin. Invest. 37, 791–801
42 Spilianakis, C., Papamatheakis, J. and Kretsovali, A. (2000) Acetylation by PCAFenhances CIITA nuclear accumulation and transactivation of major histocompatibilitycomplex class II genes. Mol. Cell. Biol. 20, 8489–8498
43 Sommer, L., Hagenbuchle, O., Wellauer, P. K. and Strubin, M. (1991) Nuclear targeting ofthe transcription factor Ptf1 is mediated by a protein subunit that does not bind to the Ptf1cognate sequence. Cell 67, 987–994
44 Bierkamp, C., Bonhoure, S., Mathieu, A., Clerc, P., Fourmy, D., Pradayrol, L., Seva, C. andDufresne, M. (2004) Expression of cholecystokinin-2/gastrin receptor in the murinepancreas modulates cell adhesion and cell differentiation in vivo. Am. J. Pathol. 165,2135–2145
45 Yamauchi, T., Yamauchi, J., Kuwata, T., Tamura, T., Yamashita, T., Bae, N., Westphal, H.,Ozato, K. and Nakatani, Y. (2000) Distinct but overlapping roles of histone acetylasePCAF and of the closely related PCAF-B/GCN5 in mouse embryogenesis. Proc. Natl.Acad. Sci. U.S.A. 97, 11303–11306
46 Hernandez-Munoz, I., Skoudy, A., Real, F. X. and Navarro, P. (2008) Pancreatic ductaladenocarcinoma: cellular origin, signalling pathways and stroma contribution.Pancreatology 8, 462–469
Received 5 February 2008/23 September 2008; accepted 6 October 2008Published as BJ Immediate Publication 6 October 2008, doi:10.1042/BJ20080293
c© The Authors Journal compilation c© 2009 Biochemical Society
Biochem. J. (2009) 418, 463–473 (Printed in Great Britain) doi:10.1042/BJ20080293
SUPPLEMENTARY ONLINE DATAp/CAF modulates the activity of the transcription factor p48/Ptf1ainvolved in pancreatic acinar differentiationAnnie RODOLOSSE*†1, Maria-Luisa CAMPOS†‡, Ilse ROOMAN†2, Mathieu LICHTENSTEIN*†3 and Francisco X. REAL*†‡4
*Departament de Ciencies Experimentals i de la Salut, Universitat Pompeu Fabra Doctor Aiguader, 88, 08003 Barcelona, Spain, †Unitat de Biologia Cel.lular i Molecular, InstitutMunicipal d’Investigacio Medica, 08003 Barcelona, Spain, and ‡Programa de Patologıa Molecular, Centro Nacional de Investigaciones Oncologicas, Melchor Fernandez Almagro 3,28029 Madrid, Spain
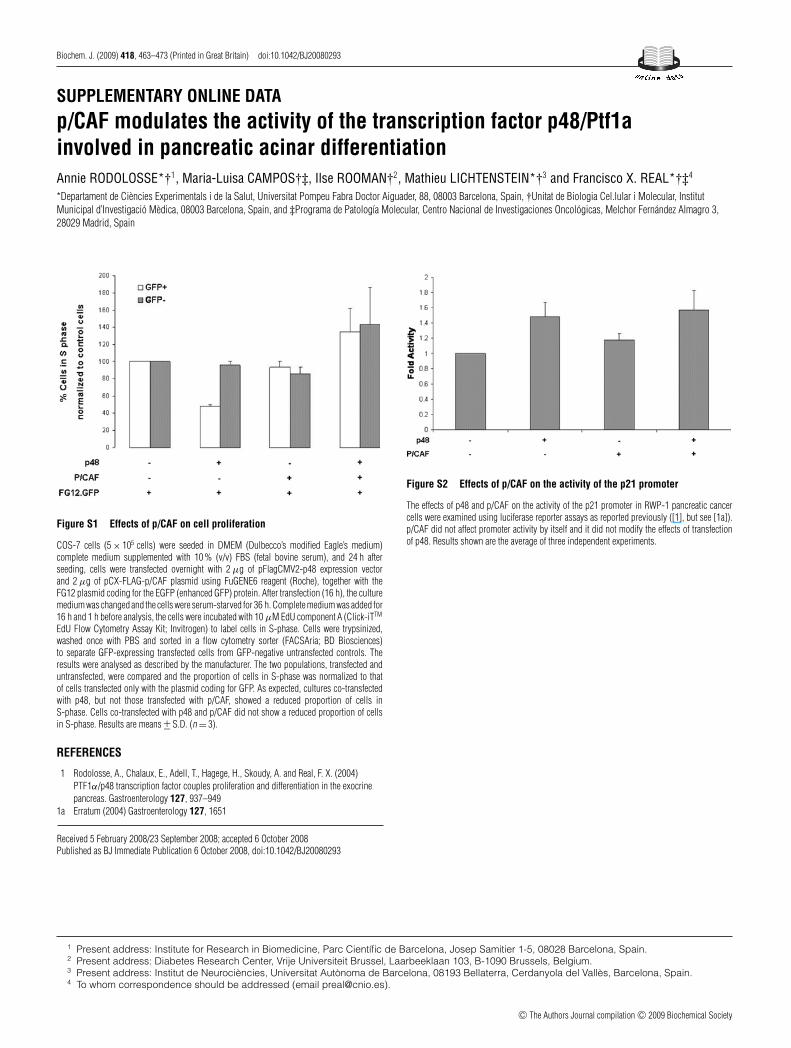
Figure S1 Effects of p/CAF on cell proliferation
COS-7 cells (5 × 105 cells) were seeded in DMEM (Dulbecco’s modified Eagle’s medium)complete medium supplemented with 10 % (v/v) FBS (fetal bovine serum), and 24 h afterseeding, cells were transfected overnight with 2 μg of pFlagCMV2-p48 expression vectorand 2 μg of pCX-FLAG-p/CAF plasmid using FuGENE6 reagent (Roche), together with theFG12 plasmid coding for the EGFP (enhanced GFP) protein. After transfection (16 h), the culturemedium was changed and the cells were serum-starved for 36 h. Complete medium was added for16 h and 1 h before analysis, the cells were incubated with 10 μM EdU component A (Click-iTTM
EdU Flow Cytometry Assay Kit; Invitrogen) to label cells in S-phase. Cells were trypsinized,washed once with PBS and sorted in a flow cytometry sorter (FACSAria; BD Biosciences)to separate GFP-expressing transfected cells from GFP-negative untransfected controls. Theresults were analysed as described by the manufacturer. The two populations, transfected anduntransfected, were compared and the proportion of cells in S-phase was normalized to thatof cells transfected only with the plasmid coding for GFP. As expected, cultures co-transfectedwith p48, but not those transfected with p/CAF, showed a reduced proportion of cells inS-phase. Cells co-transfected with p48 and p/CAF did not show a reduced proportion of cellsin S-phase. Results are means +− S.D. (n = 3).
REFERENCES
1 Rodolosse, A., Chalaux, E., Adell, T., Hagege, H., Skoudy, A. and Real, F. X. (2004)PTF1α/p48 transcription factor couples proliferation and differentiation in the exocrinepancreas. Gastroenterology 127, 937–949
1a Erratum (2004) Gastroenterology 127, 1651
Received 5 February 2008/23 September 2008; accepted 6 October 2008Published as BJ Immediate Publication 6 October 2008, doi:10.1042/BJ20080293
Figure S2 Effects of p/CAF on the activity of the p21 promoter
The effects of p48 and p/CAF on the activity of the p21 promoter in RWP-1 pancreatic cancercells were examined using luciferase reporter assays as reported previously ([1], but see [1a]).p/CAF did not affect promoter activity by itself and it did not modify the effects of transfectionof p48. Results shown are the average of three independent experiments.
1 Present address: Institute for Research in Biomedicine, Parc Cientıfic de Barcelona, Josep Samitier 1-5, 08028 Barcelona, Spain.2 Present address: Diabetes Research Center, Vrije Universiteit Brussel, Laarbeeklaan 103, B-1090 Brussels, Belgium.3 Present address: Institut de Neurociencies, Universitat Autonoma de Barcelona, 08193 Bellaterra, Cerdanyola del Valles, Barcelona, Spain.4 To whom correspondence should be addressed (email [email protected]).
c© The Authors Journal compilation c© 2009 Biochemical Society





























