Embed Size (px)
Citation preview

Human Movement Science 25 (2006) 275–291
www.elsevier.com/locate/humov
Postural fluctuations during pointing froma unilateral or bilateral stance
Ing-Shiou Hwang a,b,*, Chien-Ting Huang a,Rong-Ju Cherng a,b, Chien-Chun Huang c
a Department of Physical Therapy, College of Medicine, National Cheng Kung University, Tainan 701, Taiwanb Institute of Allied Health Sciences, College of Medicine, National Cheng Kung University, Tainan 701, Taiwan
c Department of Electrical Engineering, College of Electrical Engineering and Computer Science,
National Cheng Kung University, Tainan 701, Taiwan
Available online 3 February 2006
Abstract
An experiment was conducted to compare the effects of bilateral and unilateral stance on posturalfluctuations and intralimb coordination during active balance control. Fifteen participants stoodbilaterally and unilaterally while conducting a pointing task with an outstretched arm. Excursionof center of foot pressure (CoP) and limb movements were recorded with a force plate and eightdual-axis accelerometers, respectively. Compared to bilateral stance, unilateral stance resulted inwider CoP trajectories and greater postural fluctuations, especially in the lower limbs. The limb-dependent postural fluctuations during unilateral stance were associated with an increased couplingbetween the upper limb segments and a decreased coupling between the segments of the stance leg.Unilateral stance further resulted in greater regularity and spectral changes in postural fluctuationsof the trunk and lower limb due to increased central oscillations (8–15 Hz). The observed structuraldifferences in postural fluctuations between unilateral and bilateral stance strongly suggested that thepostural control system modulates joint stiffness in a stance-dependent manner. Probably, in unilat-eral stance, attentive control was shifted to the stance leg at the expense of increasing arm stiffness toreduce movement redundancy.� 2005 Elsevier B.V. All rights reserved.
PsycINFO classification: 2330
0167-9457/$ - see front matter � 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.humov.2005.09.009
* Corresponding author. Address: Department of Physical Therapy, College of Medicine, National ChengKung University, Tainan 701, Taiwan. Tel.: +886 6 2353535x5932; fax: +886 6 2370411.
E-mail address: [email protected] (I.-S. Hwang).

276 I.-S. Hwang et al. / Human Movement Science 25 (2006) 275–291
Keywords: Postural coordination patterns; Postural tremor; Motor control; Pointing
1. Introduction
Maintaining upright stance is an important activity in that it is a prerequisite for successin many daily tasks. In fact, standing completely still is impossible: there are always alter-nating movements of center of pressure and adjustments of the limb position againstchanges in gravitational torque (Hodges, Gurfinkel, Brumagne, Smith, & Cordo, 2002;Kantor, Poupard, Le Bozec, & Bouisset, 2001). Stance control is especially critical fortasks in which the participant has to aim at a fixed target from a standing position (suchas in shooting or in pointing at a particular location). In this case, the suprapostural point-ing or aiming task should be effectively integrated into the control of posture, requiringcoordination of a large number of biomechanical degrees of freedom (Bernstein, 1967;Ko, Challis, & Newell, 2003).
In studying the coordinated control of a suprapostural task, that is, a task in which pos-ture has to be maintained to facilitate other actions (Stoffregen, Smart, Bardy, & Pagu-layan, 1999), most researchers have focused on the effects of the focal movement onpostural equilibrium, rather than on the effects of balance constraint on movement control(Pozzo, Stapley, & Papaxanthis, 2002). In particular, little research has been conducted touncover how limb kinematics are controlled in the face of variations in balance con-straints, such as a change in position from bilateral to unilateral stance. Pointing froma standing position requires a movement synergy for minimizing fluctuations in the upperlimb and a postural synergy for maintaining balance in the trunk and the lower limbs(Balasubramaniam, Riley, & Turvey, 2000; Era, Konttinen, Mehto, Saarela, & Lyytinen,1996). An interesting question that arises in this context is which control strategies areused by the central nervous system to allow precise pointing under disadvantageous pos-tures. Unilateral stance is an effective manipulation in this regard, as is evidenced by thefact that it not only introduces postural instability in daily life situations, but also chal-lenges gymnasts when required to perform skilled maneuvers in this particular position.
A classical approach to studying the coordination between posture and movement hasbeen to analyze postural stability during rapid arm movements (Ferry, Martin, Termoz,Cote, & Prince, 2004; Patla, Ishac, & Winter, 2002; Vernazza-Martin, Martin, Cincera,Pedotti, & Massion, 1999). In contrast to rapid arm movements, postural pointing froma standing position induces only minute fluctuations in limb segments (smaller than0.01 rad), which are difficult to measure reliably with traditional motion-based systems(Aramaki et al., 2001; Winter, Patla, Ishac, & Gage, 2003). For this purpose, it is betterto use accelerometry than state-of-the-art 3D movement registration (Moe-Nilssen,1998). Accelerometers can record not only very low frequency body sway (Panzer, Bandi-nelli, & Hallett, 1995) but also high frequency central oscillation (8–15 Hz), as well asmechanical resonant components (1–4 Hz) of the limbs (Morrison & Newell, 2000a,2000b; Newell & Vaillancourt, 2001), all of which are subject to neuromuscular-skeletalconstraints for maintaining posture. Morrison and Newell (2000b) reported that the spec-tral distribution of tremor changes when the number of effective degrees of freedom injoint space is reduced. In a later study, increased amplitude and regularity of tremor were

I.-S. Hwang et al. / Human Movement Science 25 (2006) 275–291 277
observed in a motor task requiring delicate manipulative control and dexterity (Morrison& Keogh, 2001).
The aim of the present study was to investigate the coordinated control of posture andthe performance of a suprapostural task by focusing on the difference in dynamical inter-action between limb segments when pointing with an outstretched arm from a bilateraland a unilateral stance. By analyzing the spectral and temporal features of limb fluctua-tions, we sought to answer the following two questions: (1) does the act of postural point-ing involve different motor synergies for the upper and lower limbs, and (2) does thecoordination between limb segments differ between stance patterns?
2. Methods and materials
2.1. Participants
Fifteen healthy adults (ten male, five female, mean age 25.6 years, SD = 4.8 years) with-out any history of neuromuscular diseases participated in the experiment. All participantswere self-proclaimed right-handers. The participants were not allowed to consume caffeinederivatives on the day of testing because caffeine is known to increase limb fluctuations.Informed consent was obtained from all participants in accordance with institutionalguidelines.
2.2. Instrumentation
Eight dual-axis accelerometers (ADXL202, Analog Devices Inc., Norwood, USA, sen-sitivity = 312 mV/g, measurement range = ±2 g, bandwidth = 0.01 Hz–5 kHz) were posi-tioned on the L4 spinous process and the dominant upper and lower limbs. Fouraccelerometers were placed on the upper limb and three accelerometers were attached tothe lower limb to measure kinematic fluctuations of selected limb segments. The accelero-meters on the upper limb were placed on the following anatomical landmarks (Morrison &Keogh, 2001; Morrison & Newell, 2000a): (1) the index finger: dorsal aspect of the distalindex finger; (2) the hand: middle shaft of the third metacarpal bone; (3) the forearm: bellyof the brachioradialis; (4) the arm: 3 cm lateral to the belly of the biceps brachii. The accel-erometers on the lower limb were placed on: (1) the thigh: 5 cm inferior to the greater tro-chanter; (2) the calf: 5 cm inferior to the fibular head; (3) the foot: peroneal tubercle. Likethe platform data, the fluctuation signals were recorded in anterior–posterior (AP) andmedial–lateral (ML) direction, and amplified with a gain of 450 so that the signal-to-noiseratio was at least 10:1 in all recorded signals. As unnecessary variability could be registeredif the accelerometers were not placed horizontally on the limb segment, the accelerometerswere carefully placed, by adjusting their ‘‘output tilt in degrees’’, in order to minimize thisthreat to measurement validity. The recorded fluctuation data were preprocessed usinganalog low-pass filters (cut-off frequencies set at 40 Hz) within a distribution box and thendigitized at 400 Hz using a specific computer program constructed on a Labview 6.1 plat-form (National Instruments, Austin, USA). A force platform (Kistler 9284; Kistler Instru-ments Ltd, UK) was used to measure the displacement of the center of pressure (CoP)during the pointing tasks. The force-platform signals and accelerometer data weresynchronized with an external trigger, and all data were digitized at 400 Hz. The systemsetup is displayed in Fig. 1.

Fig. 1. Schematic diagram of the apparatus used for data collection.
278 I.-S. Hwang et al. / Human Movement Science 25 (2006) 275–291
2.3. Procedures
Participants performed a suprapostural pointing task with the dominant arm from botha bilateral and a unilateral stance. The participants outstretched their arm and thenpointed the index finger directly ahead, parallel to the ground. The positions of the upperlimb were as follows: the shoulder flexed 90� in the sagittal plane, the elbow fully extendedwith forearm pronated, and the wrist kept at 0� flexion. The fist was closed, but the indexfinger was fully extended. During the bilateral stance, the participants stood on the plat-form barefooted, with their feet about a shoulder-width apart. In the unilateral stance con-dition, they lifted the non-stance (non-dominant) leg in 90� knee and hip flexion. In eachof the two conditions, the participants performed the pointing task three times with eyesopen, for a duration of 20 s. In addition to remaining motionless during stance, the par-ticipants were instructed to look at their index fingers and to minimize the motions of theupper limb. Stance order was randomized across participants, and 2 min rests were sched-uled between trials. Additional breaks were allowed if the participants felt fatigued duringthe experiment. Accelerometer data and force platform data (CoP) were recorded in theML and AP directions.
2.4. Data analysis
Linear trends of the accelerometer and force platform data were removed before thecalculation of the root mean square (RMS) of the limb segment fluctuations and CoPtraces. Based on the RMS values, the degree of fluctuation in unilateral stance relativeto that in bilateral stance (RMS(unilateral)/RMS(bilateral)) was determined for each limb seg-ment. Analysis of the regularity of the fluctuation signal was conducted using approximateentropy (ApEn), which measures the logarithmic likelihood that runs of patterns that are

I.-S. Hwang et al. / Human Movement Science 25 (2006) 275–291 279
close to each other for m observations (within a tolerance r) remain close on the next incre-mental comparisons. The general mathematical expression for ApEn is as follows:
�ApEn ¼ umþ1ðrÞ � umðrÞ;where um is the average value of a natural logarithm for regularity function of given win-dow length m (for details see Pincus, 1995), and r is the tolerance level (r = 0.2 · the stan-dard deviation (SD) of the data points). ApEn values typically range from 0 to 2. Valuesclose to 0 represent greater periodicity, while values near 2 represent higher complexity.
As the accelerometers were placed on different limb segments, the correlation betweenrecorded fluctuations of neighboring limb segments in the time domain indicate that activecontrol is being exerted on the joint between two limb segments. A higher positive corre-lation suggests a stronger mechanical coupling (i.e., a stiffer joint) between the two limbsegments. Prior to cross-correlation analysis, the accelerometer data were first prepro-cessed with a digital 6th Butterworth band pass filter (cut-off frequency = 1 and 4 Hz)to eliminate (1) body sway of very low frequency under 1 Hz (Taguchi, 1977) that mayartificially enhance mechanical coupling between limb segments, and (2) tremor detectedby the accelerometers in the range of 8–15 Hz (Gallasch, Rafolt, Kenner, Konev, & Koz-lovskaya, 1994; Vaillancourt & Newell, 2000). Spectral analysis involved estimation of thepower spectral density of each profile of fluctuation signals using a combined windowingand averaging, the Welch approach. A Hanning window was used, with a window lengthof 2.45 s and an overlap of 0.64 s. Typically, the frequency profile of the accelerometerdata contains two distinct spectral components whose maxima are located in the rangesof 1–4 Hz and 8–15 Hz and are defined as the low frequency peak (LFP) and the highfrequency peak (HFP), respectively. The LFP represents the intensity of the mechanicalresonant response and the HFP the central oscillation of postural tremor recorded in alimb segment.
In relation to intralimb coordination dynamics, the primary interests of this study wereto examine the main effect of stance pattern on temporal and spectral features of the CoPand accelerometer data. For both ML and AP directions, a Hotelling’s T2 statistic wasused to compare both platform data and accelerometer data of all limb segments betweenbilateral and unilateral stance, using the following dependent measures: (1) RMS of CoPexcursions; (2) RMS, RMS(unilateral)/RMS(bilateral) ratio, and ApEn of accelerometer data;(3) spectral features of the accelerometer data, LFP and HFP; and (4) correlation coeffi-cients of accelerometer data between two adjacent limb segments. A significance level of.05 was adopted, and univariate t-tests with Bonferroni correction were used in all posthoc analyses. For practical purposes, complex interaction effects between two directions(AP and ML) and eight limb segments on fluctuation during bilateral and unilateralstances were not accounted for. Signal processing and statistical analyses were completedusing Matlab v. 6.0 (Mathworks Inc. Natick, MA, USA) and the statistical package forSocial Sciences (SPSS) for Windows v. 10.0 (SPSS Inc. Chicago, IL, USA).
3. Results
The results of Hotelling’s T2 statistic indicated that stance pattern significantly affectedthe CoP excursion during the pointing task (Wilks’ K = .061, p < .001), with a larger CoPRMS during unilateral stance than during bilateral stance (ML direction: t = �14.02,p < .001; AP direction: t = �7.87, p < .001) (Fig. 2). Typical accelerometer data exhibiting

COP_ML COP_AP
CO
P R
MS
(m
m)
0
2
4
6
8
10
12 BilateralUnilateral
**
**
**: p < .001
Fig. 2. Contrast of variability of center of pressure (CoP) data during pointing task in different standing postures.CoP_ML and CoP_AP represent RMS of CoP in the mediolateral (ML) and anterioposterior (AP) directions.
ML Direction
Time (Second)
0 5 10 15 20
0.2
0.3
0.4
0.5
Index
Hand
Arm
Forearm
Lumbar
Thigh
Calf
Foot
ML Direction
Time (Second)
0 5 10 15 20
0.0
0.1
0.2
0.3
0.4Index
Hand
Arm
Forearm
Lumbar
Thigh
Calf
Foot
(a) (b)
Fig. 3. Example of a single trial of the raw accelerometer signal from limb segments during pointing tasks for twodifferent standing postures: (a) bilateral stance, (b) unilateral stance.
280 I.-S. Hwang et al. / Human Movement Science 25 (2006) 275–291
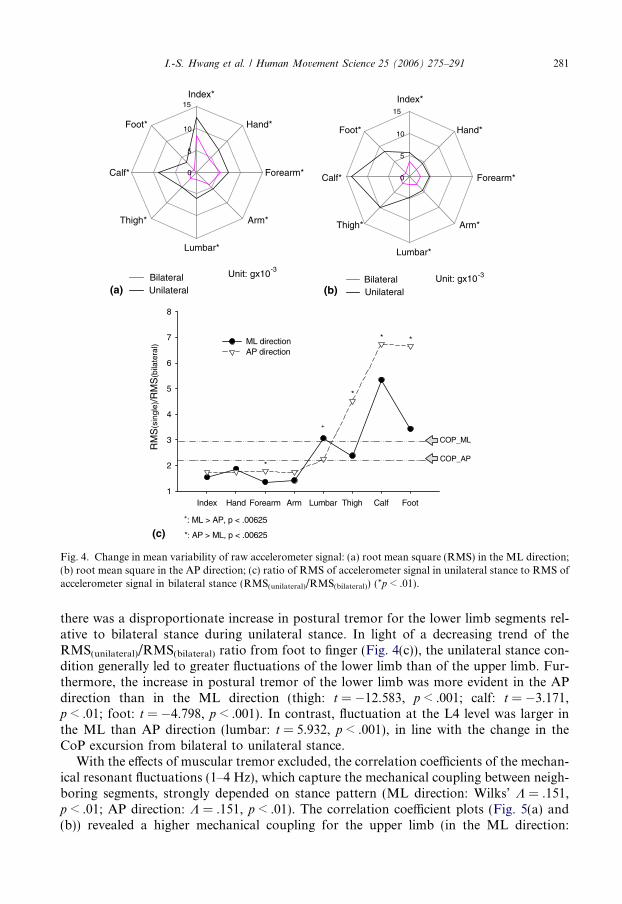
limb fluctuations are shown in Fig. 3. Figs. 4(a) and (b) display population means of limbfluctuations in eight distinct azimuths of a polar plot. In the plot, significant differences inthe amplitude of the postural tremor between the two stance conditions are indicated withan asterisk for each limb segment (*p < .01) (ML direction: Wilks’ K = .125, p < .05; APdirection: Wilks’ K = .053, p < .001). Besides changes in the CoP trajectory, unilateralstance also resulted in greater limb fluctuations than bilateral stance. It is noteworthy that
Forearm*0
5
10
15Index*
Hand*
Arm*
Lumbar*
Thigh*
Calf*
Foot*
BilateralUnilateral
Unit: gx10-3
(a) (b)
Forearm*Calf* 0
5
10
15
Index*
Hand*
Arm*
Lumbar*
Thigh*
Foot*
BilateralUnilateral
Unit: gx10-3
Index Hand Forearm Arm Lumbar Thigh Calf Foot
RM
S(s
ingl
e)/R
MS
(bila
tera
l)
1
2
3
4
5
6
7
8
ML directionAP direction
* *
*
*
+
+: ML > AP, p < .00625
*: AP > ML, p < .00625
COP_ML
COP_AP
(c)
Fig. 4. Change in mean variability of raw accelerometer signal: (a) root mean square (RMS) in the ML direction;(b) root mean square in the AP direction; (c) ratio of RMS of accelerometer signal in unilateral stance to RMS ofaccelerometer signal in bilateral stance (RMS(unilateral)/RMS(bilateral)) (*p < .01).
I.-S. Hwang et al. / Human Movement Science 25 (2006) 275–291 281
there was a disproportionate increase in postural tremor for the lower limb segments rel-ative to bilateral stance during unilateral stance. In light of a decreasing trend of theRMS(unilateral)/RMS(bilateral) ratio from foot to finger (Fig. 4(c)), the unilateral stance con-dition generally led to greater fluctuations of the lower limb than of the upper limb. Fur-thermore, the increase in postural tremor of the lower limb was more evident in the APdirection than in the ML direction (thigh: t = �12.583, p < .001; calf: t = �3.171,p < .01; foot: t = �4.798, p < .001). In contrast, fluctuation at the L4 level was larger inthe ML than AP direction (lumbar: t = 5.932, p < .001), in line with the change in theCoP excursion from bilateral to unilateral stance.
With the effects of muscular tremor excluded, the correlation coefficients of the mechan-ical resonant fluctuations (1–4 Hz), which capture the mechanical coupling between neigh-boring segments, strongly depended on stance pattern (ML direction: Wilks’ K = .151,p < .01; AP direction: K = .151, p < .01). The correlation coefficient plots (Fig. 5(a) and(b)) revealed a higher mechanical coupling for the upper limb (in the ML direction:

Cor
rela
tion
Coe
ffici
ent (
r)
0.0
0.2
0.4
0.6
0.8
1.0BilateralUnilateral.867*
(.058)
.818(.091)
.853(.062)
.022(.082)
.244(.124) .444
(.237)
.114(.207)
.806(.067)
.879*(.046)
.945*(.013)
.201*(.113)
.148(.221)
.600*(.141)
.309*(.175)
(a)
(b) Index-Hand Hand-Forearm Forarm-Arm Arm-Lumbar Lumbar-Thigh Thigh-Calf Calf-Foot
Index-Hand Hand-Forearm Forarm-Arm Arm-Lumbar Lumbar-Thigh Thigh-Calf Calf-Foot
Cor
rela
tion
Coe
ffici
ent (
r)
-0.2
0.0
0.2
0.4
0.6
0.8
1.0BilateralUnilateral
.948*(.030)
.753(.093)
.741*(.095)
.061(.119)
-.285(.300)
.285(.200)
.005(.185)
.888(.058)
.669(.122) .634
(.116)
.191+
(.107)
.061*(.277)
.613*(.112)
.350*(.196)
Fig. 5. Mechanical coupling of neighboring limb segments in terms of mean correlation coefficient ofaccelerometer data (1–4 Hz): (a) in the ML direction; (b) in the AP direction (*p < .01, +p < .02).
0
1
2Index*
Hand*
Arm*
Lumbar*
Thigh*
Calf*
Foot*
Bilateral Unilateral
Forearm
(a)
0
1
2
Index*
Hand*
Arm*
Lumbar*
Thigh*
Calf*
Foot*
Bilateral Unilateral
Forearm*
(b)
Fig. 6. Mean approximate entropy (ApEn) scores for accelerometer data recorded in different limb segments:(a) in the ML direction; (b) in the AP direction (*p < .01).
282 I.-S. Hwang et al. / Human Movement Science 25 (2006) 275–291

I.-S. Hwang et al. / Human Movement Science 25 (2006) 275–291 283
r = .806–.945; in the AP direction: r = .634–.948), but a much weaker coupling for thelower limb (in the ML direction: r = .114–.600; in the AP direction: r = �.285 to .613).In particular, during unilateral stance, correlation coefficients between mechanical fluctu-ations in the upper limbs (index–hand, hand–forearm, and forearm–arm) were enhancedcompared to bilateral stance (*p < .01). In contrast, the mechanical coupling between thelower limb segments was reduced during unilateral stance, except that the coupling of thecalf–foot in the ML direction was intensified (p < .05). ApEn of the limb fluctuationsdepended significantly on stance pattern (ML direction: Wilks’ K = .079, p < .01; APdirection: Wilks’ K = .062, p < .001) (Fig. 6(a) and (b)), and the ApEn in the unilateral
Bilateral_ML
Frequency (Hz)0 5 10 15 20 25 30
Pow
er S
pect
ral D
ensi
ty (
g2 /Hz)
x10
-3
0.0
0.5
1.0
1.5 IndexHandForarmArm
LFPHFP
(a)
Unilateral_ML
Frequency (Hz)0 5 10 15 20 25 30
Pow
er S
pect
ral D
ensi
ty (
g2 /Hz)
x10
-3
0
1
2
3
4
5
6
IndexHandForearmArm
LFP
HFP
(b)
Fig. 7. Spectral plot for accelerometer data in different limb segments illustrating low frequency peak (LFP) andhigh frequency peak (HFP) in the power density function: (a) upper limb segments during bilateral stance in theML direction; (b) upper limb segments during unilateral stance in the ML direction; (c) lower limb segmentsduring bilateral stance in the AP direction; (d) lower limb segments during unilateral stance in the AP direction.

284 I.-S. Hwang et al. / Human Movement Science 25 (2006) 275–291
condition was generally smaller than in the bilateral condition (*p < .01). Comparativedecreases in ApEn of postural tremor in the unilateral condition (16.9–50.8%) indicatednot only more regularity of the tremor signal but also signified enhanced active controlof postural steadiness when participants stood unilaterally on the force plate.
Power spectral analysis of the accelerometer data from a typical case revealed somechanges in the frequency peak of postural tremor, either the LFP (1–4 Hz) or HFP (8–15 Hz), with respect to stance pattern (Fig. 7(a)–(d)). Fig. 8(a)–(d) summarize the differ-ences in LFP and HFP magnitudes between bilateral and unilateral stance. A largerLFP magnitude for all limb segments was evident during unilateral stance than duringbilateral stance (ML direction: Wilks’ K = .162, p < .05; AP direction: Wilks’ K = .148,p < .05) (Fig. 8(a) and (b)). Stance pattern modified the HFP magnitude of the limb
Bilateral_AP
Frequency (Hz)0 5 10 15 20 25 30
Pow
er S
pect
ral D
ensi
ty (
g2 /H
z) x
10-3
0.00
0.05
0.10
0.15
0.20
0.25
0.30
LumbarThighCalfFoot
Unilateral_AP
Frequency (Hz)0 5 10 15 20 25 30
Pow
er S
pect
ral D
ensi
ty (
g2 /H
z) x
10-3
0
2
4
6
8
LumbarThighCalfFoot
LFP
LFP
HFP
HFP
(c)
(d)
Fig. 7 (continued)

ML Direction
Index Hand Forearm Arm Lumbar Thigh Calf Foot
Low
Fre
quen
cy M
agni
tude
(g2
/Hz
x 10
-3)
Low
Fre
quen
cy M
agni
tude
(g2
/Hz
x 10
-3)
0
2
4
6
8
10
12
BilateralUnilateral
*
*
* * **
*
*
(a)
(b)
AP Direction
Index Hand Forearm Arm Lumbar Thigh Calf Foot
0
2
4
6
8
10
12
BilateralUnilateral
*
* ** *
*
*
*
Fig. 8. Comparison of peak frequency magnitudes for accelerometer data in different limb segments duringpointing task in different standing postures: (a) magnitude of low frequency peak in the ML direction; (b)magnitude of low frequency peak in the AP direction; (c) magnitude of high frequency peak in the ML direction;(d) magnitude of high frequency peak in the AP direction (*p < .01).
I.-S. Hwang et al. / Human Movement Science 25 (2006) 275–291 285
segments as well (ML direction: Wilks’ K = .158, p < .05; AP direction: Wilks’ K = .142,p < .05). However, the HFP magnitude in the upper limb during pointing did not differbetween the two stance conditions (p > .05). Only the HFP magnitude in the lower limband the L4 in the unilateral stance condition were significantly enhanced in comparisonwith those in the bilateral stance condition (p < .01) (Fig. 8(c) and (d)).
4. Discussion
The purpose of the present study was to examine, through detailed analyses of limb seg-ment fluctuations and CoP trajectories, if there exists limb-specific coordinated synergies

ML Direction
Index Hand Forearm Arm Lumbar Thigh Calf Foot0
1
2
3
4
5
BilateralUnilateral
*
*
*
*
(c)
(d)
AP Direction
Index Hand Forearm Arm Lumbar Thigh Calf Foot0
1
2
3
4
5
BilateralUnilateral
*
*
*
*
Low
Fre
quen
cy M
agni
tude
(g2
/Hz
x 10
-3)
Low
Fre
quen
cy M
agni
tude
(g2
/Hz
x 10
-3)
Fig. 8 (continued)
286 I.-S. Hwang et al. / Human Movement Science 25 (2006) 275–291
during postural pointing from a quiet stance, and if those coordinated synergies differbetween bilateral and unilateral stance. The present study suggested that two postural sub-systems coexist for the act of postural pointing, a movement strategy to minimize limbfluctuations in the upper limb and a balance strategy in the trunk and leg for stabilitymaintenance. Both control strategies varied significantly with the adopted stance pattern,as evidenced by significant variations in the strength of the mechanical coupling betweenfluctuations of neighboring limb segments.
4.1. Enhanced mechanical coupling of the upper limb in the unilateral stance
A marked increase in the CoP trajectories in both AP and ML direction evidenced thatthe postural constraints on one-legged standing are more severe than on two-legged stand-

I.-S. Hwang et al. / Human Movement Science 25 (2006) 275–291 287
ing (Fig. 3). The postural constraints on unilateral stance were increased due to a reduc-tion in the base of support as well as a different mechanical balance situation for lifting thenon-stance leg forward. In response to the critical postural constraint, the participantstended to enhance the mechanical coupling of the upper limb (Fig. 5(a) and (b)) when pos-tural stability was jeopardized during unilateral stance. The observed pattern of stiffeningthe upper limb segments is reminiscent of the process of freezing up independent func-tional units during the initial stage of motor learning (Bernstein, 1967; Ko, Challis, &Newell, 2001; Newell & Vaillancourt, 2001). It has been proposed that postural coordina-tion is predominantly governed by the principle of maximum efficacy in that the posturalsystem adopts a coordination manner so as to achieve a stable equilibrium state with leastexertion (Ko et al., 2003; Riccio & Stoffregen, 1988). For suprapostural pointing imposedon unilateral stance, the central nervous system is inclined to suppress the redundancy ofthe degrees-of-freedom of the upper limb for ease of postural control, if balance mainte-nance is superior to postural pointing. Through stiffening of the upper limb segments, theparticipants expended less effort on dealing with joint interaction force and movementamplitude.
A common cause of enhanced mechanical coupling is co-activation of antagonist pairsaround a joint (Ghafouri, Archambault, Adamovich, & Feldman, 2002; Humphrey &Reed, 1983). However, coactivation of antagonist muscles may not be an appropriateaccount for intensified joint stiffness of the upper limb in the present study. Morrisonand Newell (2000b) investigated the relationship between posture tremors and joint stiff-ness as a function of antagonist co-activation. They reported that a higher level of jointstiffness was associated with an increase in the frequency peaks at 1–4 Hz and 8–15 Hz.However, we found that only the LFP of postural tremor (1–4 Hz) in the upper limbwas larger for unilateral than for bilateral stance (Fig. 8(a)–(d)). In fact, increasing jointstiffness by means of co-contraction has a high energetic cost (Popescu, Hidler, & Rymer,2003), and often results in greater variability in the position of distal effectors over time(Milner, 2002). Hence, in keeping with the principle of maximum efficacy, co-activationis unlikely to be the cause of the increase in mechanical coupling observed in the upperlimb during unilateral stance. Alternatively, greater joint stability can be achieved byexploiting joint geometry (Flash & Mussa-Ivaldi, 1990; Mussa-Ivaldi, Hogan, & Bizzi,1985), which determines the amount of compression force between articulation surfaces.The participant might slightly flex the elbow in the closely-packed position so that jointstiffness could be maintained economically by passive constraints of surrounding liga-ments without additional muscle contraction. Therefore, muscle contraction vibrationof the upper limb segments, the magnitude of HFP (8–15 Hz) of the postural fluctuation,was independent of change in stance pattern (Fig. 8(c) and (d)).
4.2. Stance-related strategy in the trunk and leg for postural stability
In light of the RMS(unilateral)/RMS(bilateral) ratio, it is worth noting that the fluctuationsof the upper limb were affected much less by the change in stance than those of the lowerlimb (Fig. 4(c)). This new finding suggests that some compensatory actions, or movementstrategies, must be at work in the trunk or lower limb to reduce the postural fluctuations ofthe end-effectors during unilateral stance, as a consequence of disadvantageous enhance-ment of mechanical coupling in the arm (Fig. 5(a) and (b)) that facilitates passive trans-mission of postural fluctuations from the other portions of body segments. Compared

288 I.-S. Hwang et al. / Human Movement Science 25 (2006) 275–291
to postural pointing during bilateral stance, the participants further reduced the mechan-ical coupling in the lower limb and the arm-lumbar site to allow more joint mobility dur-ing unilateral standing. Hence, by reorganizing joint stiffness and the dynamic interactionsbetween the segments, the trunk and the leg are likely to play decisive roles in maintainingpostural stability when the stance pattern changes from bilateral to unilateral.
Compared to bilateral stance, unilateral stance required additional neural control overthe trunk and lower limb, presumably as a consequence of increased balance constraints.During unilateral stance, the participants might have shifted their attention to the controlof the trunk and lower limb, which would explain the increase in regularity (Fig. 6) andamplitude of the HFP of postural fluctuation in these segments (Fig. 8(c) and (d)). Thestructural change in postural fluctuations implied that greater voluntary control was beingexerted to the arm when the participant was requested to aim at a target with a greaterlevel of accuracy (Morrison & Keogh, 2001). In addition, enhanced HFP in the lumbarpostural fluctuation during single stance is compatible with the findings of Henry, Fung,and Horak (2001), who reported increased muscle (EMG) activity of the trunk as stancewidth decreased, acknowledging that the HFP is primarily attributable to synchronizationof motor units and central-driven oscillation (McAuley & Marsden, 2000; Vallbo & Wess-berg, 1993). Modification of trunk control with respect to stance pattern during perfor-mance of the suprapostural pointing task was evidenced by a reduction in themechanical coupling of the arm-lumbar when a subject raised the supporting leg(Fig. 5(a) and (b)). Pearcy and Tibrewal (1984) proposed that narrow stance perturbationsare accompanied by rotation and flexion of the trunk causing a passive stiffness of thespine. For the compensatory release of the trunk mechanical coupling, the power of thelower limb fluctuations was not transmitted to the upper limb segments, so posturalfluctuations of the upper limb segments did not drastically increase (Fig. 4(c)) as wouldbe predicted from an inverted pendulum model (Loram, Kelly, & Lakie, 2001).
Hoogvliet, Van Duyl, De Bakker, Mulder, and Stam (1997) reported that with one leglifted, the threat to postural stability was larger in the ML direction than in the AP direc-tion; this explains why participants exhibited a greater corrective action in the frontalplane. We found that the movement strategy to prevent falling during unilateral stancewas to intensify calf–foot mechanical coupling in the ML direction (the ankle strategy)(Fig. 5(a)), resulting in a decrease in mechanical coupling at the lumbar–thigh, thigh–calf,and calf–foot sites in the AP direction (Fig. 5(b)). Riemann, Myers, and Lephart (2003)claimed that the ankle joint is the greatest source of corrective action during one-leggedstanding. Enhancement of mechanical coupling of the calf–foot during one-legged stand-ing is likely to constrain the ankle movements against any abrupt increase in gravitationaltorque in the frontal plane (Riemann et al., 2003; Tropp & Odenrick, 1988). The unex-pected finding was the release of mechanical coupling in the lower limb in the AP planeduring unilateral stance, since the knee joint might be considered as rigidly in supportof body weight with little transitional movement. However, based on the intralimb fluctu-ations, we conclude that there was a multi-joint coordination strategy for the stance leg toadjust the center of gravity in the vertical direction for better postural stability during uni-lateral stance. Perhaps the release of mechanical coupling in the joints of the lower limb inthe AP direction occurred because participants relied on persistently bending and extend-ing the knee joint within a minute range that coupled with interchanging kinematic com-binations of the thigh and calf. Day, Steiger, Thompson, and Marsden (1993) suggested

I.-S. Hwang et al. / Human Movement Science 25 (2006) 275–291 289
that it would be more effective to control motion in the sagittal plane to provide morepostural stability in the frontal plane.
4.3. Methodological issues for the use of accelerometry
In recent years, many researchers have used accelerometry to measure balance duringstanding (Kamen, Patten, Du, & Sison, 1998; Mayagoitia, Lotters, Veltink, & Hermens,2002). Accelerometers provide good reliability in studying posture, provided that theirinclination is carefully controlled (Moe-Nilssen, 1998). However, the results of this studymay have been influenced by methodological limitations. It is important to note that theestimation of some temporal or spectral parameters, such as ApEn and correlation coef-ficients, could be biased because of insufficient signal-to-noise ratios (S/N ratio) of theaccelerometer data. Although no scientific work has recommended a standard S/N ratioof the accelerometer data for postural studies, our results are unlikely to have been affectedby noise level when S/N ratio generally exceeded 10 (Morrison & Newell, 1996). Forinstance, if noise level did affect the correlation coefficient of fluctuation between neighbor-ing limb segments, one could expect a higher correlation coefficient for the lower limbduring unilateral stance due physically to a higher S/N ratio.
5. Conclusions
Through detailed analysis of postural fluctuations, the current study revealed severalinteresting properties of suprapostural pointing as a function of variations in stance pat-terns. First, the increase in balance constraint from bilateral stance to unilateral stanceresulted in enhanced mechanical coupling of the upper limb to master joint redundancy.Second, when stance pattern varied, the central nervous system reorganized joint mechan-ical couplings, especially that of the trunk and leg, to maintain the equilibrium of thewhole posture system and minimize fluctuation of the upper limb.
Acknowledgements
This research was supported financially by grants from the National Science Council,ROC, under Grant no. NSC 92-2314-B-006-035.
References
Aramaki, Y., Nozaki, D., Masani, K., Sato, T., Nakazawa, K., & Yano, H. (2001). Reciprocal angularacceleration of the ankle and hip joints during quiet standing in humans. Experimental Brain Research, 136,463–473.
Balasubramaniam, R., Riley, M. A., & Turvey, M. T. (2000). Specificity of postural sway to the demands of aprecision task. Gait and Posture, 11, 12–24.
Bernstein, N. A. (1967). The co-ordination and regulation of movement. Oxford: Pergamon Press.Day, B. L., Steiger, M. J., Thompson, P. D., & Marsden, C. D. (1993). Effect of vision and stance width on
human body motion when standing: Implications for afferent control of lateral sway. Journal of Physiology
(London), 469, 479–499.Era, P., Konttinen, N., Mehto, P., Saarela, P., & Lyytinen, H. (1996). Postural stability and skilled performance–
a study on top-level and naive rifle shooters. Journal of Biomechanics, 29, 301–306.

290 I.-S. Hwang et al. / Human Movement Science 25 (2006) 275–291
Ferry, M., Martin, L., Termoz, N., Cote, J., & Prince, F. (2004). Balance control during an arm raising movementin bipedal stance: Which biomechanical factor is controlled. Biological Cybernetics, 91, 104–114.
Flash, T., & Mussa-Ivaldi, F. (1990). Human arm stiffness characteristics during the maintenance of posture.Experimental Brain Research, 82, 315–326.
Gallasch, E., Rafolt, D., Kenner, T., Konev, A., & Kozlovskaya, I. B. (1994). Physiological tremor and control oflimb position in 1 and 0 G. Journal of Gravitational Physiology, 1, 52–54.
Ghafouri, M., Archambault, P. S., Adamovich, S. V., & Feldman, A. G. (2002). Pointing movements may beproduced in different frames of reference depending on the task demand. Brain Research, 929, 117–128.
Henry, S. M., Fung, J., & Horak, F. B. (2001). Effect of stance width on multidirectional postural responses.Journal of Neurophysiology, 85, 559–570.
Hodges, P. W., Gurfinkel, V. S., Brumagne, S., Smith, T. C., & Cordo, P. C. (2002). Coexistence of stability andmobility in postural control: Evidence from postural compensation for respiration. Experimental Brain
Research, 144, 293–302.Hoogvliet, P., Van Duyl, W. A., De Bakker, J. V., Mulder, P. G., & Stam, H. J. (1997). Variations in foot
breadth: Effect on aspects of postural control during one-leg stance. Archives of Physical Medicine and
Rehabilitation, 78, 284–289.Humphrey, D. R., & Reed, D. J. (1983). Separate cortical systems for control of joint movement and joint
stiffness: Reciprocal activation and coactivation of antagonist muscles. Advances in Neurology, 39, 347–372.Kamen, G., Patten, C., Du, C. D., & Sison, S. (1998). An accelerometry-based system for the assessment of
balance and postural sway. Gerontology, 44, 40–45.Kantor, E., Poupard, L., Le Bozec, S., & Bouisset, S. (2001). Does body stability depend on postural chain
mobility or stability area. Neuroscience Letters, 308, 128–132.Ko, Y. G., Challis, J. H., & Newell, K. M. (2001). Postural coordination patterns as a function of dynamics of the
support surface. Human Movement Science, 20, 737–764.Ko, Y. G., Challis, J. H., & Newell, K. M. (2003). Learning to coordinate redundant degrees of freedom in a
dynamic balance task. Human Movement Science, 22, 47–66.Loram, I. D., Kelly, S. M., & Lakie, M. (2001). Human balancing of an inverted pendulum: Is sway size
controlled by ankle impedance? Journal of Physiology, 532(Pt 3), 879–891.Mayagoitia, R. E., Lotters, J. C., Veltink, P. H., & Hermens, H. (2002). Standing balance evaluation using a
triaxial accelerometer. Gait and Posture, 16, 55–59.McAuley, J. H., & Marsden, C. D. (2000). Physiological and pathological tremors and rhythmic central motor
control. Brain, 123, 1545–1567.Milner, T. E. (2002). Contribution of geometry and joint stiffness to mechanical stability of the human arm.
Experimental Brain Research, 143, 515–519.Moe-Nilssen, R. (1998). Test-retest reliability of trunk accelerometry during standing and walking. Archives of
Physical Medicine and Rehabilitation, 79, 1377–1385.Morrison, S., & Keogh, J. (2001). Changes in the dynamics of tremor during goal-directed pointing. Human
Movement Science, 20, 675–693.Morrison, S., & Newell, K. M. (1996). Inter- and intra-limb coordination in arm tremor. Experimental Brain
Research, 110, 455–464.Morrison, S., & Newell, K. M. (2000a). Postural and resting tremor in the upper limb. Clinical Neurophysiology,
111, 651–663.Morrison, S., & Newell, K. M. (2000b). Limb stiffness and postural tremor in the arm. Motor Control, 4, 293–315.Mussa-Ivaldi, F. A., Hogan, N., & Bizzi, E. (1985). Neural, mechanical, and geometric factors subserving arm
posture in humans. Journal of Neuroscience, 5, 2732–2743.Newell, K. M., & Vaillancourt, D. E. (2001). Dimensional change in motor learning. Human Movement Science,
20, 695–715.Panzer, V. P., Bandinelli, S., & Hallett, M. (1995). Biomechanical assessment of quiet standing and changes
associated with aging. Archives of Physical Medicine and Rehabilitation, 76, 151–157.Patla, A. E., Ishac, M. G., & Winter, D. A. (2002). Anticipatory control of center of mass and joint stability
during voluntary arm movement from a standing posture: Interplay between active and passive control.Experimental Brain Research, 143, 318–327.
Pearcy, M. J., & Tibrewal, S. B. (1984). Axial rotation and lateral bending in the normal lumbar spine measuredby three-dimensional radiography. Spine, 9, 582–587.
Pincus, S. (1995). Approximate entropy (ApEn) as a complexity measure. Chaos, 5, 110–117.

I.-S. Hwang et al. / Human Movement Science 25 (2006) 275–291 291
Popescu, F., Hidler, J. M., & Rymer, W. Z. (2003). Elbow impedance during goal-directed movements.Experimental Brain Research, 152, 17–28.
Pozzo, T., Stapley, P. J., & Papaxanthis, C. (2002). Coordination between equilibrium and hand trajectoriesduring whole body pointing movements. Experimental Brain Research, 144, 343–350.
Riccio, G. E., & Stoffregen, T. A. (1988). Affordances as constraint on the control of stance. Human Movement
Science, 4–5, 695–716.Riemann, B. L., Myers, J. B., & Lephart, S. M. (2003). Comparison of the ankle, knee, hip, and trunk corrective
action shown during unilateral-leg stance on firm, foam, and multiaxial surfaces. Archives of Physical
Medicine and Rehabilitation, 84, 90–95.Stoffregen, T. A., Smart, L. J., Bardy, B. G., & Pagulayan, R. J. (1999). Postural stabilization of looking. Journal
of Experimental Psychology: Human Perception and Performance, 25(6), 1641–1658.Taguchi, K. (1977). Spectral analysis of body sway. Journal of Oto-Rhino-Laryngology & its Related Specialties,
39, 330–337.Tropp, H., & Odenrick, P. (1988). Postural control in unilateral-limb stance. Journal of Orthopaedic Research, 6,
833–839.Vaillancourt, D. E., & Newell, K. M. (2000). Amplitude changes in the 8–12, 20–25, and 40 Hz oscillations in
finger tremor. Clinical Neurophysiology, 111, 1792–1801.Vallbo, A. B., & Wessberg, J. (1993). Organization of motor output in slow finger movements in man. Journal of
Physiology (London), 469, 673–691.Vernazza-Martin, S., Martin, N., Cincera, M., Pedotti, A., & Massion, J. (1999). Arm raising in humans under
loaded vs. unloaded and bipedal vs. unipedal conditions. Brain Research, 846, 12–22.Winter, D. A., Patla, A. E., Ishac, M., & Gage, W. H. (2003). Motor mechanisms of balance during quiet
standing. Journal of Electromyography and Kinesiology, 13, 49–56.





























