Embed Size (px)
Citation preview

1
-Scientific Report-
RAGE binds phosphatidylserine and assists in the clearance of apoptotic cells.
Mei He1, Hiroshi Kubo1, Konosuke Morimoto2, Naoya Fujino1, Takaya Suzuki3, Toru
Takahasi4, Mitsuhiro Yamada5, Mutsuo Yamaya1, Tomoyuki Maekawa2, Yasuhiko
Yamamoto6, Hiroshi Yamamoto6.
1Department of Advanced Preventive Medicine for Infectious Disease, Tohoku
University Graduate School of Medicine, Sendai, 980-8575 Japan; 2Department of
Clinical Medicine, Institute of Tropical Medicine, Nagasaki University, Nagasaki,
852-8523 Japan; 3Department of Thoracic Surgery, 4Department of Anesthesiology,
Tohoku University School of Medicine, Sendai, 980-8575 Japan; 5Department of
Infection Control and Laboratory Diagnostics, Tohoku University Graduate School of
Medicine, Sendai, 980-8575 Japan; 6Department of Biochemistry and Molecular
Vascular Biology, Kanazawa University Graduate School of Medical Science,
Kanazawa, 920-8640 Japan.
Corresponding author: Hiroshi Kubo, M.D., Ph.D.
Department of Advanced Preventive Medicine for Infectious Disease
Tohoku University Graduate School of Medicine
2-1 Seiryoumachi, Aobaku, Sendai 980-8575, Japan.
Phone: +81-22-717-7184, FAX: +81-22-717-7576.
E-mail: [email protected]
Published in EMBO reports 12:358-364, 2011 (doi:10.1038/embor.2011.28)

2
Abstract
Clearance of apoptotic cells is necessary for tissue development, homeostasis,
and resolution of inflammation. The uptake of apoptotic cells is initiated by an "eat-
me" signal such as phosphatidylserine (PS) on the cell surface, and phagocytes
recognize the signal using specific receptors. Here we show that the soluble form of
the receptor for advanced glycation end-products (RAGE) bound to PS as well as to
the apoptotic thymocytes. RAGE-deficient (Rage-/-) alveolar macrophages exhibited
impaired phagocytosis of apoptotic thymocytes and defective clearance of apoptotic
neutrophils in Rage-/- mice. Our results indicate that RAGE acts as a PS receptor and
assists in the clearance of apoptotic cells.

3
Introduction
The receptor for advanced glycation end-products (RAGE) is a member of the
immunoglobulin superfamily of cell-surface molecules (Neeper et al, 1992); it is
expressed in diverse cell types, including endothelium, T-lymphocytes and
monocytes/macrophages (Brett et al, 1993). RAGE recognizes a variety of ligands,
such as high-mobility group protein-1 (HMGB1), and the binding of RAGE to such
ligands has been linked to many diseases, including diabetes, atherosclerosis,
neurodegeneration, inflammation, and tumors (Ramasamy et al, 2009). RAGE is
expressed as both a full-length membrane-bound form (mRAGE) and various soluble
forms lacking the transmembrane domain (sRAGE). sRAGE is produced by the
proteolytic cleavage of membrane RAGE and alternative mRNA splicing (Hudson et
al, 2008a; Raucci et al, 2008) and represents a competitive inhibitor of RAGE by
acting as a decoy for RAGE ligands (Santilli et al, 2009).
Rapid removal of apoptotic cells by phagocytes is critical for the maintenance
of tolerance and prevents inflammation and autoimmune responses against
intracellular antigens released from the dying cells (Savill & Fadok, 2000). The
uptake of apoptotic cells is initiated by an “eat-me” signal such as phosphatidylserine
(PS) on the cell surface, and phagocytes recognize the signal using specific receptors
(Fadok et al, 1992; Verhoven et al, 1995). Several classes of receptors have been
implicated in the recognition of apoptotic cells, possibly via direct binding to the
exposed PS, including BAI1, Tim4, and stabilin-2 (Kobayashi et al, 2007; Miyanishi
et al, 2007; Park et al, 2007; Park et al, 2008) or indirect binding through bridging
molecules (MFG-E8, Gas6) (Hanayama et al, 2004; Scott et al, 2001). The
engagement of the PS receptors initiates signaling events within the phagocytes that
lead to activation of the small GTPase Rac, and the subsequent cytoskeletal

4
reorganization of the phagocyte membrane to allow corpse internalization (Albert et
al, 2000). Deficiencies of the receptors that participate in the recognition of PS on the
surface of apoptotic cells lead to a failure to maintain self-tolerance and the
development of autoimmunity (Asano et al, 2004; Cohen et al, 2002).
Although RAGE is a member of the pattern-recognition receptor family (Xie et
al, 2008), whether it participates in the phagocytosis and clearance of apoptotic cells
remains unknown. In this study, we present evidence that RAGE binds to PS and
modulates apoptotic cell phagocytosis. Furthermore, we confirm the role of RAGE in
apoptotic cell clearance in lipopolysaccharide (LPS)-induced acute lung injury in
mice.
Results and Discussion
sRAGE binds to apoptotic thymocytes as well as to PS
To determine whether RAGE identifies its functional ligands on apoptotic cells,
we prepared thymocytes with a proportion of apoptotic cells by dexamethasone
treatment, and we evaluated the binding capacity of RAGE to the apoptotic
thymocytes using Alexa Fluor 660 dye–labeled sRAGE and JC-1 dye (Petit et al,
1995). We found that sRAGE specifically bound to an apoptotic population of
thymocytes (Fig. 1A, black portion), indicating that RAGE binds to a structure highly
expressed on the surface of apoptotic thymocytes. AGE, one of the major RAGE
ligands, was not present on apoptotic thymocytes (Suppl. Fig. 2). Exposure of PS on
the outer-membrane leaflet is a major indicator of apoptotic versus live cells (Fadok et
al, 1992), so we tested whether RAGE can bind to PS. Using a protein–lipid overlay
assay that employed hydrophobic membranes pre-spotted with a concentration
gradient of eight different lipids found in cell membranes (PIP Strip™), we found that

5
sRAGE specifically bound to PS in a concentration-dependent manner, except for a
slight binding to phosphatidylglycerol (Fig. 1B). To determine the binding affinity of
sRAGE to PS, we employed a surface-plasmon resonance analysis (Biacore) using PS
or PC (as a reference) liposomes immobilized on the sensor chip and serial dilutions
of sRAGE as soluble analytes. The sensorgrams showed an increase in response units
(RU) reflective of PS binding, and the binding response was concentration-dependent,
with a KD of 0.563 µM (Fig.1C). These findings indicate that sRAGE specifically
binds to PS.
To investigate whether sRAGE can interact with PS on apoptotic cells, apoptotic
thymocytes were incubated with NBD-PS and Chromeo 546–labeled sRAGE. The
distribution of PS and sRAGE on the cell surface was determined by confocal
imaging, and the interaction of sRAGE and NBD-PS was measured by fluorescence
resonance energy transfer (FRET). As shown in Figure 2A, both PS and sRAGE were
localized to the surface of apoptotic thymocytes. FRET spots, indicating interactions
between sRAGE and PS, appeared on the cell surface with the acceptor (Chromeo
546–sRAGE, red fluorescence) being photo-bleached. In contrast, fewer FRET spots
were observed on the cell surface of the same thymocytes that were not photo-
bleached (Fig. 2A). This observation that RAGE and PS colocalize on the cell
membrane.
Confocal images of alveolar macrophages cocultured with NBD-PS liposomes
revealed that mRAGE expression was concentrated in a pseudopod that formed
toward a PS liposome (Figure 2B). Both FRET analysis and confocal images
demonstrated that PS expressed on apoptotic cells binds to full-length mRAGE
located in macrophages and that this PS–RAGE binding may play a role in apoptosis
recognition and phagocytosis.

6
RAGE deficiency or sRAGE administration impairs the phagocytosis of apoptotic
thymocytes by alveolar macrophages
Because RAGE can bind to PS, we then sought to determine the function of
RAGE in the phagocytosis of apoptotic cells, examining the phagocytic capacity of
both wild-type and Rage-/- alveolar macrophages (Fig. 3A). Alveolar macrophages
were co-cultured with apoptotic thymocytes from wild-type mice. The phagocytic
index of wild-type alveolar macrophages at 2 h was significantly higher than that of
Rage-/- macrophages (p = 0.013) (Fig. 3B), suggesting that alveolar macrophages can
recognize and phagocytose apoptotic cells through the activity of RAGE on their cell
surface. Because sRAGE acts as an endogenous competitive inhibitor of ligand
engagement by cell-surface RAGE (Santilli et al, 2009), we then examined the effect
of sRAGE on macrophage phagocytosis. sRAGE administration significantly
impaired the phagocytosis of apoptotic thymocytes by wild-type alveolar
macrophages, which confirms that RAGE contributes to the phagocytosis of apoptotic
cells. Interestingly, sRAGE administration also induced a significant phagocytosis
defect in Rage-/- alveolar macrophages; it is possible that sRAGE counteracts not only
RAGE-mediated phagocytosis but also other PS receptor–mediated phagocytosis,
such as that mediated by BAI1 and Tim4 These results suggest that RAGE is one of
the PS receptors that recognize apoptotic cells.
PS–RAGE binding is critical for Rac1 activation
Intracellular RAGE signaling pathways involve Rho GTPases (Hudson et al,
2008b), and Rac1, one of Rho GTPases, activation is essential in the PS-recognizing
system that is involved in apoptotic cell removal (Albert et al, 2000). We therefore

7
prepared PS liposomes to mimic apoptotic cells and tested whether PS–RAGE
binding could alter the level of activated Rac1 during uptake by alveolar
macrophages. Wild-type alveolar macrophages showed greater PS liposome–induced
Rac1 activation; in contrast, PS liposomes did not activate Rac1 in Rage-/- alveolar
macrophages. Phosphatidylcholine (PC) liposomes, as a negative control, did not
activate Rac1 in either the wild-type or Rage-/- alveolar macrophages (Fig. 3C). These
data demonstrate that inducible Rac1 activation is due to PS recognition through
RAGE in alveolar macrophages. The level of Rac1 stayed at near-basal levels in both
wild-type and Rage-/- macrophages when PS liposomes were supplied with sRAGE
(Fig. 3C). sRAGE competed with RAGE for PS binding and functioned as a decoy
that abrogated Rac1 activation, supporting the assertion that PS–RAGE binding is a
prerequisite for the induction of Rac1 activation in alveolar macrophages.
Defective clearance of apoptotic neutrophils in LPS-induced acute lung injury in
Rage-/- mice
We next tested whether RAGE participates in apoptotic cell clearance in vivo.
Because RAGE is constitutively expressed at high levels in the lung (Brett et al,
1993), we induced lung injury in Rage-/- and wild-type mice by an intranasal
insufflation of LPS (20 µg/mouse in 100 µl PBS). Intranasal instillation of LPS
induces extensive neutrophil migration within the airspace. Once neutrophils
extravasate, they undergo programmed cell death (Watson et al, 1996), and are then
removed from the airspace by the phagocytosis of alveolar macrophages (Cox et al,
1995). This active program of resolution begins in the first few hours after an
inflammatory response begins (Serhan & Savill, 2005). We performed a
bronchoalveolar lavage (BAL) at hours 0 (baseline), 2, 4, 6, and 24 after LPS

8
administration and examined the apoptotic cells recovered from the BAL fluid. More
apoptotic neutrophils were found in Rage-/- mice at 6 h and 24 h after LPS
administration than in wild-type mice (Fig. 4A). Then we examined the phagocytosis
of both wild-type and Rage-/- alveolar macrophages (Fig. 4B, right). Rage-/- alveolar
macrophages exhibited significantly decreased phagocytosis of apoptotic neutrophils
at 6 h and 24 h compared with wild-type (Fig. 4B, left). Next, we checked whether
this impaired clearance of apoptotic neutrophils would affect the accumulation of
inflammatory cells within the airspace. We found that LPS administration resulted in a
rapid increase of inflammatory cells in BAL fluid at 2, 4, 6, and 24 h in the mice. The
total cell number in BAL fluid recovered from the Rage-/- mice was substantially
greater than that from the wild-type mice at 2, 4, 6, and 24 h, and there was a
statistically significant difference at 6 h between the two groups (p = 0.02) (Fig. 4C,
left). The increased number of inflammatory cells was attributable largely to the
increase in neutrophils (Fig. 4C, right). We also checked KC and MIP-2, which are
the key cytokines associated with neutrophil recruitment (Martin, 2002). The KC and
MIP-2 levels in BAL fluid did not significantly differ between the two types of mice
after LPS administration (Fig. 4D). Taken together, these data show that deletion of
RAGE in macrophages impairs the clearance of apoptotic cells, thereby leading to
increased inflammatory cell accumulation within the airspace in the early phase of
LPS-induced acute lung injury.
In LPS-induced lung injury, HMGB1 plays a key role in the late phase of the
injury (Wang et al, 1999). Liu et al. reported that HMGB1 interacts with PS and
diminishes the phagocytosis of apoptotic cells (Liu et al, 2008). We confirmed the
binding of HMGB1 and PS by protein–lipid overlay assay (Suppl. Fig. 3A) and
surface-plasmon resonance analysis (Biacore) (Suppl. Fig. 3B). However, HMGB1

9
also bound to other lipids, namely phosphatidylglycerol and phosphatidic acid, and its
binding affinity to PS was lower than that of sRAGE (KD of 3.08 µM in HMGB1, KD
of 0.563 µM in sRAGE) according to the Biacore analysis. Nevertheless, there
remains the possibility of a multiprotein complex composed of sRAGE and HMGB1
that binds to PS in vivo, so further work is needed to clarify this possibility.
Apoptotic cell clearance is important for tissue development (Savill & Fadok,
2000), and inefficient phagocytosis of apoptotic cells could lead to the development of
autoimmune disease (Zitvogel et al, 2010). However, Rage-/- mice, which were kept in
a sterile environment, were viable and did not show any overt phenotypes. The lack of
both RAGE and sRAGE may keep the RAGE-null animals in a comparative balance
in which no phenotype is expressed.
The data presented here provide new insights into the process of apoptotic cell
clearance by RAGE. First, RAGE may act as one of the PS receptors that recognize
and initiate apoptotic cell clearance (Fig. 5A). Second, our data raise an important
point regarding an sRAGE/mRAGE balance in overall apoptosis recognition, because
sRAGE may counteract the PS engagement of cell-surface RAGE and other receptors,
such as BAI1, Tim4, or stabilin-2, during phagocytosis (Fig. 5B). sRAGE is produced
by alternative splicing of RAGE mRNA or by proteolytic cleavage of RAGE from the
cell surface in humans, and sRAGE has the same ligand-binding specificity, resulting
in competition with RAGE for ligand binding. Because sRAGE acts as a competitor
for mRAGE binding to PS, the balance of sRAGE and mRAGE could modify
macrophage phagocytotic activity, which might be an important component of
pathophysiological situations in which apoptosis recognition plays a key role.

10
The present finding of RAGE as a PS receptor unveils a new role for RAGE in
apoptosis-induced pathophysiological conditions and identifies a potential new target
for the treatment of relevant human diseases.
Methods
Recombinant sRAGE protein and Rage-/- mice were utilized for this study.
Identification of sRAGE by Flow Cytometry
Alexa Fluor 660 dye–labeled sRAGE or BSA was added to dexamethasone-
treated thymocytes. Apoptotic cells were identified using a MitoProbe JC-1 assay kit
(Molecular Probes). Flow cytometry was performed on a BD FACSCanto II (BD
Biosciences).
PIP Strip™ assay
PIP Strips on which the indicated phospholipids had been spotted were
purchased from Echelon Bioscience, and dot-blot experiments were carried out
according to the manufacturer’s protocol.
Surface-Plasmon Resonance Analysis (Biacore)
The binding affinity of sRAGE to PS was analyzed using a Biacore X100
(Biacore AB) in a single-cycle affinity model. KD was calculated using ka and kd
values by a trivalent analyte model.
Fluorescence resonance energy transfer (FRET) analysis
FRET analysis was performed as previously described (Liu et al, 2008).
Confocal laser scanning microscopy
Alveolar macrophages were grown on cover slips and then preincubated with
5 µM NBD-PS liposome for 2 h, and then mRAGE was detected by indirect

11
immunofluorescence using a rabbit polyclonal anti-RAGE antibody (Abcam) with
Alexa Fluor 647–conjugated goat anti-rabbit IgG (Molecular Probes).
Phagocytosis of apoptotic thymocytes
Phagocytosis was assayed by adding apoptotic thymocytes (2.5 × 106 cells/ml)
with or without sRAGE (3 µg/ml). Phagocytosis was determined as a phagocytic
index under microscopy as previously described (Morimoto et al, 2006). Each
condition was tested in duplicate, and the samples were blindly and independently
analyzed by two researchers (M.H. and K.M.).
Rac1 Activity Assays
Rac1 activity was measured using an ELISA-based Rac1 Activation Assay
Biochem Kit (G-LISA, Cytoskeleton).
LPS-induced lung injury
LPS from Escherichia coli serotype 055:B5 was obtained from Sigma-
Aldrich, and the induction of lung injury was performed as previously described
(Yamada et al, 2004).
Detailed methods are described in the Supplementary Information.

12
Acknowledgments
We thank J. Aoki and A. Inoue for technical advice on preparing PS and PC
liposomes, and M. Kanzaki and H. Hatakeyama for expert technical assistance on
FRET analysis. This work was supported by a grant-in-aid from the Japan Society for
the Promotion of Science (20.08466) and grants from National Institute of Biomedical
Innovation to H.K.

13
References
Albert ML, Kim JI, Birge RB (2000) alphavbeta5 integrin recruits the CrkII-
Dock180-rac1 complex for phagocytosis of apoptotic cells. Nat Cell Biol 2: 899-905
Asano K, Miwa M, Miwa K, Hanayama R, Nagase H, Nagata S, Tanaka M (2004)
Masking of phosphatidylserine inhibits apoptotic cell engulfment and induces
autoantibody production in mice. J Exp Med 200: 459-467
Brett J, Schmidt AM, Yan SD, Zou YS, Weidman E, Pinsky D, Nowygrod R, Neeper
M, Przysiecki C, Shaw A, et al. (1993) Survey of the distribution of a newly
characterized receptor for advanced glycation end products in tissues. Am J Pathol
143: 1699-1712
Cohen PL, Caricchio R, Abraham V, Camenisch TD, Jennette JC, Roubey RA, Earp
HS, Matsushima G, Reap EA (2002) Delayed apoptotic cell clearance and lupus-like
autoimmunity in mice lacking the c-mer membrane tyrosine kinase. J Exp Med 196:
135-140
Cox G, Crossley J, Xing Z (1995) Macrophage engulfment of apoptotic neutrophils
contributes to the resolution of acute pulmonary inflammation in vivo. Am J Respir
Cell Mol Biol 12: 232-237

14
Fadok VA, Voelker DR, Campbell PA, Cohen JJ, Bratton DL, Henson PM (1992)
Exposure of phosphatidylserine on the surface of apoptotic lymphocytes triggers
specific recognition and removal by macrophages. J Immunol 148: 2207-2216
Hanayama R, Tanaka M, Miyasaka K, Aozasa K, Koike M, Uchiyama Y, Nagata S
(2004) Autoimmune disease and impaired uptake of apoptotic cells in MFG-E8-
deficient mice. Science 304: 1147-1150
Hudson BI, Carter AM, Harja E, Kalea AZ, Arriero M, Yang H, Grant PJ, Schmidt
AM (2008a) Identification, classification, and expression of RAGE gene splice
variants. FASEB J 22: 1572-1580
Hudson BI, Kalea AZ, Del Mar Arriero M, Harja E, Boulanger E, D'Agati V, Schmidt
AM (2008b) Interaction of the RAGE cytoplasmic domain with diaphanous-1 is
required for ligand-stimulated cellular migration through activation of Rac1 and
Cdc42. J Biol Chem 283: 34457-34468
Kobayashi N, Karisola P, Pena-Cruz V, Dorfman DM, Jinushi M, Umetsu SE, Butte
MJ, Nagumo H, Chernova I, Zhu B, Sharpe AH, Ito S, Dranoff G, Kaplan GG,
Casasnovas JM, Umetsu DT, Dekruyff RH, Freeman GJ (2007) TIM-1 and TIM-4
glycoproteins bind phosphatidylserine and mediate uptake of apoptotic cells.
Immunity 27: 927-940

15
Liu G, Wang J, Park YJ, Tsuruta Y, Lorne EF, Zhao X, Abraham E (2008) High
mobility group protein-1 inhibits phagocytosis of apoptotic neutrophils through
binding to phosphatidylserine. J Immunol 181: 4240-4246
Martin TR (2002) Neutrophils and lung injury: getting it right. J Clin Invest 110:
1603-1605
Miyanishi M, Tada K, Koike M, Uchiyama Y, Kitamura T, Nagata S (2007)
Identification of Tim4 as a phosphatidylserine receptor. Nature 450: 435-439
Morimoto K, Janssen WJ, Fessler MB, McPhillips KA, Borges VM, Bowler RP, Xiao
YQ, Kench JA, Henson PM, Vandivier RW (2006) Lovastatin enhances clearance of
apoptotic cells (efferocytosis) with implications for chronic obstructive pulmonary
disease. J Immunol 176: 7657-7665
Neeper M, Schmidt AM, Brett J, Yan SD, Wang F, Pan YC, Elliston K, Stern D, Shaw
A (1992) Cloning and expression of a cell surface receptor for advanced glycosylation
end products of proteins. J Biol Chem 267: 14998-15004
Park D, Tosello-Trampont AC, Elliott MR, Lu M, Haney LB, Ma Z, Klibanov AL,
Mandell JW, Ravichandran KS (2007) BAI1 is an engulfment receptor for apoptotic
cells upstream of the ELMO/Dock180/Rac module. Nature 450: 430-434

16
Park SY, Jung MY, Kim HJ, Lee SJ, Kim SY, Lee BH, Kwon TH, Park RW, Kim IS
(2008) Rapid cell corpse clearance by stabilin-2, a membrane phosphatidylserine
receptor. Cell death and differentiation 15: 192-201
Petit PX, Lecoeur H, Zorn E, Dauguet C, Mignotte B, Gougeon ML (1995)
Alterations in mitochondrial structure and function are early events of
dexamethasone-induced thymocyte apoptosis. J Cell Biol 130: 157-167
Ramasamy R, Yan SF, Schmidt AM (2009) RAGE: therapeutic target and biomarker
of the inflammatory response--the evidence mounts. J Leukoc Biol 86: 505-512
Raucci A, Cugusi S, Antonelli A, Barabino SM, Monti L, Bierhaus A, Reiss K, Saftig
P, Bianchi ME (2008) A soluble form of the receptor for advanced glycation
endproducts (RAGE) is produced by proteolytic cleavage of the membrane-bound
form by the sheddase a disintegrin and metalloprotease 10 (ADAM10). FASEB J 22:
3716-3727
Santilli F, Vazzana N, Bucciarelli LG, Davi G (2009) Soluble forms of RAGE in
human diseases: clinical and therapeutical implications. Current medicinal chemistry
16: 940-952
Savill J, Fadok V (2000) Corpse clearance defines the meaning of cell death. Nature
407: 784-788

17
Scott RS, McMahon EJ, Pop SM, Reap EA, Caricchio R, Cohen PL, Earp HS,
Matsushima GK (2001) Phagocytosis and clearance of apoptotic cells is mediated by
MER. Nature 411: 207-211
Serhan CN, Savill J (2005) Resolution of inflammation: the beginning programs the
end. Nature immunology 6: 1191-1197
Verhoven B, Schlegel RA, Williamson P (1995) Mechanisms of phosphatidylserine
exposure, a phagocyte recognition signal, on apoptotic T lymphocytes. J Exp Med
182: 1597-1601
Wang H, Bloom O, Zhang M, Vishnubhakat JM, Ombrellino M, Che J, Frazier A,
Yang H, Ivanova S, Borovikova L, Manogue KR, Faist E, Abraham E, Andersson J,
Andersson U, Molina PE, Abumrad NN, Sama A, Tracey KJ (1999) HMG-1 as a late
mediator of endotoxin lethality in mice. Science 285: 248-251
Watson RW, Redmond HP, Wang JH, Condron C, Bouchier-Hayes D (1996)
Neutrophils undergo apoptosis following ingestion of Escherichia coli. J Immunol
156: 3986-3992
Xie J, Reverdatto S, Frolov A, Hoffmann R, Burz DS, Shekhtman A (2008) Structural
basis for pattern recognition by the receptor for advanced glycation end products
(RAGE). J Biol Chem 283: 27255-27269

18
Yamada M, Kubo H, Kobayashi S, Ishizawa K, Numasaki M, Ueda S, Suzuki T,
Sasaki H (2004) Bone marrow-derived progenitor cells are important for lung repair
after lipopolysaccharide-induced lung injury. J Immunol 172: 1266-1272
Zitvogel L, Kepp O, Kroemer G (2010) Decoding cell death signals in inflammation
and immunity. Cell 140: 798-804

19
Figure 1: sRAGE binds to apoptotic cells and PS. (A) JC-1 dye detects mitochondrial
membrane potential and the red fluorescence decreases in apoptotic cells (black dots).
sRAGE specifically bound to an apoptotic cells (black) compared with a non-
apoptotic population (gray). Data shown are representative of three experiments. (B)
sRAGE bound to phosphatidylserine blotted on PIP Strip™. Data shown are
representative of two experiments. a: 100 pmol/spot; b: 50 pmol/spot; c: 25 pmol/
spot; d: 12.5 pmol/spot; e: 6.25 pmol/spot; f: 3.13 pmol/spot; g: 1.56 pmol/spot. (C)
SPR assays. A serial concentration of sRAGE (0, 4.32, 13.9, 117, and 350 µg/ml) was
flowed over immobilized PS or PC liposomes on the Biacore sensor chip. Response to
PC was utilized as a reference (left). The response curve of PS/sRAGE binding was

20
obtained by subtracting the curve of PS from that of PC as described under the
Methods section (right). Data shown are representative of two experiments.

21
Figure 2: RAGE and PS bind each other on cell surface. (A) Images of the donor
(NBD-PS, green) on apoptotic thymocytes, the acceptor (sRAGE, red), the bleached
area, and the FRET spots are shown. FRET efficiency was significantly increased by
bleaching the fluorescence of the acceptor (sRAGE), demonstrating the direct binding
between PS and RAGE (right graph). Data represent average values with SEM.
Student’s t-test was utilized to analyze the comparisons between two groups, and p
values lower than 0.05 were considered significant. (B) A representative confocal
microscopic image of a macrophage phagocytosing a PS liposome. Membrane-
associated full-length RAGE expression on macrophages was concentrated in a
pseudopod that was formed (arrow) toward a PS liposome (arrowhead). Phagocytosed
NBD-PS was seen in the cytoplasm of the macrophage.

22
Figure 3: Rage-/- alveolar macrophages impair the phagocytosis of apoptotic
thymocytes in vitro and exhibit reduced Rac1 activation in response to PS liposomes.
(A) Representative microscopic images depicting the phagocytosis of wild-type and
Rage-/- alveolar macrophages with or without sRAGE (3 µg/ml) treatment.
Arrowheads indicate phagocytized thymocyte fragments. (B) Wild-type alveolar
macrophages (WT) or Rage-/- alveolar macrophages incubated with apoptotic
thymocytes with or without sRAGE. The number of apoptotic cells per macrophage
uptake was determined as the phagocytic index. Phagocytic indexes were significantly
decreased in sRAGE-treated and RAGE-deficient macrophages. (C)
Phosphatidylserine (PS), not phosphatidylcholine (PC), activated Rac1 in wild-type
macrophages. PS-induced Rac1 activation was not seen in RAGE-deficient
macrophages. sRAGE significantly attenuated PS-induced Rac1 activation. Open bars
represent wild-type; closed bars represent Rage-/-. Data represent average values with
SEM. The data were compared using analysis of variance with the Bonferroni
adjustment, and p values lower than 0.05 were considered significant.

23
Figure 4: Rage-/- alveolar macrophages impair apoptotic cell clearance in vivo. (A)
Cells recovered from BAL fluid after LPS exposure were stained with FITC-
conjugated annexin V and PE-conjugated anti–mouse Ly-6G (GR-1) antibody. Data
show average counts of apoptotic neutrophils (GR-1+/annexin V+) in wild-type mice
and Rage-/- mice (n = 3–5). (B) Alveolar macrophage phagocytosis of apoptotic
neutrophils in LPS-induced BAL was determined as the alveolar macrophage (AM)
phagocytic index (n = 6–9). The image indicates an alveolar macrophage with an
ingested apoptotic neutrophil. (C) Changes in total cell counts in BAL fluid (left) and
neutrophils (right) after LPS exposure in wild-type mice and Rage-/- mice, each
obtained from 5–9 mice at each time point. (D) Changes in KC (left) and MIP-2
(right) levels in BAL fluid during the 24 h after LPS exposure (n = 6–9). Open bars
represent wild-type; closed bars represent Rage-/-. Data represent average values with

24
SEM throughout. The data were compared using analysis of variance with the
Bonferroni adjustment except neutrophil counts, KC and MIP-2. Neutrophil counts,
KC and MIP-2 were analyzed using non-parametric analysis of variance (Kruskal-
Wallis test) with Dunn's Multiple Comparison Test. P values lower than 0.05 were
considered significant.

25
Figure 5: Summary of a possible RAGE and sRAGE signaling pathway in
phagocytosis that involves PS recognition. We propose that RAGE is a PS receptor
that acts similarly to stabilin-2, BAI1 and Tim-4 (A). By binding to PS, sRAGE may
counteract RAGE and other PS receptors, such as BAI1 and stabilin-2 (B).

26
Supplementary Methods Preparation of human soluble RAGE (sRAGE)
Human histidine-tagged sRAGE was prepared in a baculovirus expression
system using Sf9 cells and purified by Ni-NTA chromatography according to
procedures published previously (Park et al, 1998). The purified proteins were then
desalted, buffer-exchanged, concentrated, and analyzed by SDS-PAGE and blue
native-PAGE. The blue native-PAGE experiment was performed with the
NativePAGETM Novex Bis-Tris Gel System with a 4–16% bis-Tris gradient gel
(Invitrogen, CA, USA) according to the manufacture’s instructions. The gels were
stained with Coomassie Brilliant Blue R-250.
Fluorescence labeling
sRAGE or bovine serum albumin (BSA) (Pierce, IL, USA) were labeled with
Alexa Fluor 660–conjugated dye using an Alexa Fluor 660 Protein Labeling Kit
(Molecular Probes, Inc., OR, USA) according to the manufacturer’s instructions. For
FRET analysis, sRAGE and BSA were labeled with Chromeo 546 using a Chromeo
546 NHS-Ester labeling kit (Active Motif, Tokyo, Japan)
Mice
Animal protocols were reviewed and approved according to the standards for
the care and use of laboratory animals of the Tohoku University Animal Experiment
Ethics Committee. C57BL/6 mice were purchased from CLEA Japan, Inc.
(Yokohama, Japan). Rage-/- mice backcrossed into the C57BL/6 strain (F7) were
described previously (Abeyama et al, 2005).

27
Isolation and apoptosis induction in thymocytes
Thymus was harvested from 3- to 4-week-old female wild-type mice (C57BL/
6 strain) and minced to yield a single-cell suspension. To induce apoptosis,
thymocytes (1x106/ml) were suspended with RPMI 1640 containing 10% heat-
inactivated FCS and incubated with a final concentration of 10-6 M dexamethasone
(Sigma, MO, USA) for 6 h.
Identification of sRAGE by Flow Cytometry
To evaluate the ability of RAGE to bind to apoptotic cells, 30 µg/ml Alexa
Fluor 660 dye–labeled sRAGE or BSA was added to dexamethasone-treated
thymocytes and incubated for 30 min at room temperature. To identify apoptotic cells,
the cymocytes were stained using a MitoProbe JC-1 assay kit (Molecular Probes)
according to the manufacturer’s instructions. Mitochondrial depolarization is
indicated by a decrease in the red/green fluorescence intensity ratio. Flow cytometry
was performed on a BD FACSCanto II (BD Biosciences, MD, USA).
Identification AGE by Flow cytometry
To detect advanced glycation end-products on apoptotic thymocytes, the cells
were treated with FcBlock (BD Biosciences) and stained with mouse monoclonal
anti–Ne-carboxymethyl lysine antibody (CML) (clone 6D12, Transgenic Inc., Hyogo,
Japan) or anti-pentosidine antibody (clone PEN12, Transgenic Inc. Japan) in
combination with Alexa Fluor 488–labeled donkey anti-mouse IgG (H+L) antibody
(Invitrogen, Tokyo, Japan) for 20 min at 4ºC in the dark. Cells were washed,
resuspended in 200 ml of 1x Binding buffer from the Annexin V-eFluor 450
Apoptosis Detection Kit (eBioscience, CA, USA), stained with eFluor 450–

28
conjugated Annexin V and 0.2 mg/ml PI (Sigma), and analyzed by FACSAriaII (BD
Bioscience). The data were transferred and reanalyzed with FlowJo software (Tree
Star, Inc., OR, USA).
PIP Strip™ assay
PIP Strips on which the indicated phospholipids had been spotted were
purchased from Echelon Bioscience (UT, USA), and dot-blot experiments were
carried out according to the manufacturer’s protocol. Briefly, strips were blocked in
PBS with 0.1% BSA (Millipore, MA, USA) for 1 hour at room temperature and then
incubated with 100 µg/ml of sRAGE in PBS with 0.1% BSA at 4°C overnight. The
strips were then incubated with a rabbit polyclonal anti-RAGE Ab (ab37647, Abcam,
Tokyo, Japan) for 1 hour at room temperature. The PIP strips were washed three times
in PBST (PBS with 0.1% Tween 20) before incubation with an HRP-conjugated
swine anti-rabbit IgG Ab (P0217, Dako, Tokyo, Japan). Antibody binding was
detected using ECL plus western blotting detection reagents (Amersham GE,
Buckinghamshire, England) and visualized using a Fujifilm LAS-1000 processor. A
recombinant LL5-a PH domain GST-tagged protein (Echelon Bioscience) was used as
a positive control and BSA was used as a negative control. To evaluate HMGB1 and
phospholipids binding, 1 µg/ml HMGB1 was incubated with the PIP strips, and
binding was detected with a rabbit anti-HMGB1 polyclonal antibody diluted 1:1000
(SHINO-TEST Corporation, Tokyo, Japan) in blocking buffer.
Solid-phase ELISA
The solid-phase ELISA for phospholipid binding was carried out as previously
described (Miyanishi et al, 2007). In brief, phospholipid (Avanti Polar Lipids, Inc.,

29
AL, USA) (100 µl, 0.75 µg/ml) in methanol was added to 96-well NUNC-IMMUNO
Plates (Nalge-Nunc, NY, USA) and air-dried. The non-specific binding sites were
blocked by treating the wells with 0.1% BSA (Millipore) in PBS. Serial
concentrations of sRAGE proteins (0, 3, 6, 12, and 24 µg/ml) were diluted to the
appropriate concentrations with PBS containing 0.1% BSA, added to the wells, and
incubated overnight at 4°C. After the wells were washed with PBS containing 0.05%
Tween 20, the wells were then incubated with 100 µL rabbit polyclonal anti-RAGE
Ab diluted 1:2000 in blocking buffer. Finally, the plates were incubated with 100 µL
HRP-conjugated swine anti-rabbit IgG Ab diluted 1:1000 in blocking buffer. The
peroxidase activity was detected using a peroxidase detection kit (Sumitomo Co.,
Kyushu, Japan) and quantified by measuring the absorbance at 490 nm with a Micro-
ELISA reader (SpectraMax 190, Molecular Devices, Tokyo, Japan).
Preparation of liposomes
Small, unilamellar liposomes containing the indicated phospholipids (Avanti
Polar Lipids, Inc.) were made as described previously (Fadok et al, 2001). For PS
liposomes, a mixture of 70% PC and 30% PS (mol/mol) was used unless otherwise
indicated, whereas PC liposomes consisted of pure PC. The lipids were mixed in
chloroform, dried, resuspended in PBS, mixed and sonicated until the solution was
clear. For surface-plasmon resonance analysis, PS liposomes were prepared with 90%
PC and 10% PS (mol/mol) and resuspended in HBS-N buffer (0.01M HEPES, 0.15M
NaCl, pH 7.4). In some experiments, 7-nitro-2-1,3-benzoxadiazol-4 (NBD) \-PS was
used. The liposomes were stored at 4°C for up to 2 weeks.
Surface-Plasmon Resonance Analysis (Biacore)

30
The binding affinity of sRAGE or HMGB1 to PS was analyzed using a
Biacore X100 (Biacore AB, Uppsala, Sweden) in a single-cycle affinity model at
25ºC. HBS-N buffer (0.01M HEPES, 0.15M NaCl, pH 7.4) was used as running
buffer, and 1 mM PS liposome (PS: PC = 1:9) was diluted in running buffer and
immobilized onto flow cell 2 (FL2) of the L1-sensor chip at a flow rate of 5 µl/min for
18 min, whereas PC liposomes were immobilized onto a flow cell 1 (FL1) of the L1-
sensor chip. Approximately 6000 RU of the liposome was immobilized. A threefold
concentration series of sRAGE (0, 4.32, 13, 38.9, 117, and 350 µg/ml) or a twofold
concentration series of HMGB1 (0, 2.5, 5, 10, and 20 µg/ml) diluted with running
buffer was injected over the flow cells at a flow rate of 10 µl/min for 120 s each in the
order of increasing concentration. A concentration series of sRAGE or HMGB1 was
injected in a single-analysis cycle without regeneration of the surface in between
injections. At the end of the cycle, the sensor chip surface was regenerated with 20
mM CHAPS (Dojindo, Kumamoto, Japan). After subtracting out the response to
buffer alone in the flow cells, the response to PS liposome binding curves were
obtained by subtracting the FL2 curve from the FL1 curve and analyzed using BIA-
evaluation software, version 2.0 (Biacore AB). Here, we assumed that the molecular
mass of sRAGE was 120 kDa. We analyzed the sensorgrams by fitting them with
possible binding models. In the case of sRAGE, KD was calculated using ka and kd
values by a trivalent analyte model, whereas that of HMGB1 was evaluated by a 1:1
binding model.
Confocal microscopy and fluorescence resonance energy transfer (FRET) analysis
Following apoptosis induction, 1,2-dioleoyl-sn-glycero-3-phospho-L-serine-
N-(7-nitro-2-1,3-benzoxadiazol-4-yl) (NBD-PS, Avanti Polar Lipid, Inc) (1 µg/ml)

31
and Chromeo546 labeled sRAGE (1 µg/ml) were added to the apoptotic thymocytes
and incubated at 37°C for 3h. The cells were washed and then fixed with 4%
paraformaldehyde in PBS for 10 min and cytospin slides were prepared. FRET
analysis was performed as previously described (Liu et al, 2008). Briefly, the
proximity of cell surface proteins was investigated for FRET measurements by the
acceptor photobleaching technique. An excitation wavelength of 450 nm and an
emission wavelength of 525–550 nm were used for donor probes (NBD), and an
excitation wavelength of 530 nm and an emission wavelength of 570–625 nm were
used for the acceptor (Chromeo 546). FRET-positive pixels and efficiencies were
calculated by directly subtracting images obtained prior to acceptor photobleaching
from images acquired immediately after acceptor photobleaching to reveal the
increased fluorescence of the donor. After an initial confocal image was acquired, a
portion of a single cell was masked, and the acceptor fluorescence (Chromeo 546) in
the masked area was bleached with a pumped dye laser to < 30%. The remaining area
of the same cell was left un-bleached and used as a control. After pixel alignment
between the pre- and post-bleach images was ensured, individual pixel FRET
efficiencies were calculated by expressing the increased donor fluorescence as a
fraction of the post-bleach intensity. The noise level in unbleached areas was
eliminated to identify FRET-positive areas and calculate image statistics.
Primary alveolar macrophage isolation and culture
Mouse alveolar macrophages were collected by BAL as previously described
(He et al, 2007), and more than 95% of the BAL cells were alveolar macrophages.
Cells (5.0 x 105/ml) were plated on baked glass cover slips in 24-well tissue culture
plates (BD Biosciences) or plated in 6-well tissue-culture plates (BD Biosciences).

32
After 1 h of incubation at 37°C, non-adherent cells were removed by a gentle
washing. Alveolar macrophage monolayers were cultured overnight before use in the
Rac1 activity assays or phagocytosis assay.
Confocal laser scanning microscopy
Alveolar macrophages were grown on cover slips and then preincubated with
5 µM NBD-PS liposome for 2 h. Cells were washed with PBS and fixed with 4%
paraformaldehyde in PBS for 10 minutes. After washing with PBS and blocking with
5% BSA in PBS, mRAGE was detected by indirect immunofluorescence using a
rabbit polyclonal anti-RAGE antibody (Abcam) diluted 1:100 in PBS with 5% BSA.
Bound antibodies were visualized with Alexa Fluor 647–conjugated goat anti-rabbit
IgG (Molecular Probes, Invitrogen) diluted 1:1000 in PBS with 5% BSA. Cells were
mounted on glass slides with ProLong Antifade Reagent with DAPI (Molecular
Probes, Inc.) and photographed using a laser confocal fluorescence microscope (Carl
Zeiss, Tokyo, Japan). To assess spatial localizations within the cell and at the surface,
serial optical sections were taken at 0.7-µm intervals throughout the thickness of all
cells examined.
Phagocytosis of apoptotic thymocytes assay
Phagocytosis was assayed by adding apoptotic thymocytes (2.5 × 106 cells/ml)
with or without sRAGE (3 µg/ml) to each well of a 24-well plate containing adherent
alveolar macrophage monolayers at 37°C for 2 h. Wells were washed gently with cold
PBS to remove uningested thymocytes and then fixed and stained with a Diff-Quick
kit (Baxter Healthcare, FL, USA). Phagocytosis was determined as a phagocytic
index under microscopy as previously described (Morimoto et al, 2006) . Each

33
condition was tested in duplicate, and the samples were blindly and independently
analyzed by two researchers (M.H. and K.M.).
Rac1 Activity Assays
Alveolar macrophages were treated with 10 µM PC or PS liposomes or PS liposomes
with 10 µg/ml sRAGE at 37 °C for 10 min, after which Rac1 activity was measured in
the lysates collected from treated alveolar macrophages using an ELISA-based Rac1
Activation Assay Biochem Kit (G-LISA, Cytoskeleton, CO, USA) according to the
manufacturer’s instructions.
LPS-induced lung injury
LPS from Escherichia coli serotype 055:B5 was obtained from Sigma-
Aldrich, and the induction of lung injury was performed as previously described
(Yamada et al, 2004). Briefly, mice (8–12 weeks old) were anesthetized using
ketamine hydrochloride with atropine. While the mice were anesthetized, an
intranasal insufflation was conducted via inhalation of LPS (20 µg/mouse in PBS), or
PBS as a control, placed in one nostril.
Evaluation of apoptotic neutrophils recovered in bronchoalveolar lavage (BAL)
fluid
The procedure for counting total cells recovered in the BAL fluid after LPS-
induced acute lung injury is described above. The cells were immediately stained with
FITC-conjugated annexin V using an apoptosis detection kit according to the
manufacturer’s protocols (Roche Applied Science, Mannheim, Germany). To identify
neutrophils in the BAL, PE-conjugated anti–mouse Ly-6G (GR-1) antibody

34
(eBioscience, Inc) was also included in the staining solution. For each sample, a
negative staining control was prepared using a solution that did not contain annexin
V or anti–GR-1 antibody, to determine background fluorescence. After staining, the
cells were washed and immediately analyzed by flow cytometry. Apoptotic
neutrophils were considered to be the GR-1–positive cells that were annexin V–
positive.
Alveolar macrophage phagocytosis of apoptotic neutrophils in BAL fluid
For the study of alveolar macrophage phagocytosis of apoptotic neutrophils,
the total cells from BAL fluid were prepared for Cytospin and stained with Diff-
Quick. Phagocytosis was determined by visual inspection of the samples and
expressed as the phagocytic index determined microscopy as previously described
(Morimoto et al, 2006).
Assessment of total cell number and differential cell counts in BAL fluid
Mice were given an overdose of halothane, and then BAL was performed.
Total cell number and differential cell counts in BAL fluid were determined as
described previously (He et al, 2007).
ELISA to assess KC and MIP-2 concentrations in BAL fluid
The concentrations of KC and MIP-2 proteins in the BAL fluid supernatant
were measured using commercial ELISA kits (R&D Systems, MN, USA) according
to the manufacturer’s instructions.
Statistical Analysis

35
All values were shown as means ± SEM. The data were compared using analysis of
variance except when specified otherwise. When overall differences were identified,
multiple comparisons with the Bonferroni adjustment were used to identify which
groups were significantly different. Between-group differences in neutrophil counts,
the levels of KC and MIP-2 were analyzed using non-parametric analysis of variance
(Kruskal-Wallis test). When overall differences were identified, Dunn's Multiple
Comparison Test. FRET efficiency between non-bleached and bleached was analyzed
by paired Student’s t-test. Phagocytic index of peritoneal macrophages was analyzed
by unpaired Student’s t-test. P values lower than 0.05 were considered significant.
Statistical analysis was performed with StatView 5.0 software (Abacus Concepts,
Berkeley, CA, USA) and GraphPad Prism 5.0 (GraphPad Software, Inc., CA, USA)

36
References
Abeyama K, Stern DM, Ito Y, Kawahara K, Yoshimoto Y, Tanaka M, Uchimura T, Ida
N, Yamazaki Y, Yamada S, Yamamoto Y, Yamamoto H, Iino S, Taniguchi N,
Maruyama I (2005) The N-terminal domain of thrombomodulin sequesters high-
mobility group-B1 protein, a novel antiinflammatory mechanism. J Clin Invest 115:
1267-1274
Fadok VA, de Cathelineau A, Daleke DL, Henson PM, Bratton DL (2001) Loss of
phospholipid asymmetry and surface exposure of phosphatidylserine is required for
phagocytosis of apoptotic cells by macrophages and fibroblasts. J Biol Chem 276:
1071-1077
He M, Kubo H, Ishizawa K, Hegab AE, Yamamoto Y, Yamamoto H, Yamaya M
(2007) The role of the receptor for advanced glycation end-products in lung fibrosis.
Am J Physiol Lung Cell Mol Physiol 293: L1427-1436
Liu G, Wang J, Park YJ, Tsuruta Y, Lorne EF, Zhao X, Abraham E (2008) High
mobility group protein-1 inhibits phagocytosis of apoptotic neutrophils through
binding to phosphatidylserine. J Immunol 181: 4240-4246
Miyanishi M, Tada K, Koike M, Uchiyama Y, Kitamura T, Nagata S (2007)
Identification of Tim4 as a phosphatidylserine receptor. Nature 450: 435-439
Morimoto K, Janssen WJ, Fessler MB, McPhillips KA, Borges VM, Bowler RP, Xiao
YQ, Kench JA, Henson PM, Vandivier RW (2006) Lovastatin enhances clearance of

37
apoptotic cells (efferocytosis) with implications for chronic obstructive pulmonary
disease. J Immunol 176: 7657-7665
Park L, Raman KG, Lee KJ, Lu Y, Ferran LJ, Jr., Chow WS, Stern D, Schmidt AM
(1998) Suppression of accelerated diabetic atherosclerosis by the soluble receptor for
advanced glycation endproducts. Nat Med 4: 1025-1031
Yamada M, Kubo H, Kobayashi S, Ishizawa K, Numasaki M, Ueda S, Suzuki T,
Sasaki H (2004) Bone marrow-derived progenitor cells are important for lung repair
after lipopolysaccharide-induced lung injury. J Immunol 172: 1266-1272

38
Supplementary Figure 1: Recombinant human sRAGE protein separated by 10%
SDS-PAGE and 4–16% blue native-PAGE. M, Precision plus protein standards
marker (Bio-Rad) in SDS-PAGE and NativeMarkTM unstained protein standard
marker (Invitrogen) in blue native-PAGE.

39
Supplementary Figure 2: RAGE ligands, such as AGE structures of CML and
pentosidine were not detected on apoptotic thymocytes. Dex, dexamethasone; Isotype,
isotype IgG; MFI, mean fluorescent intensity; AU, arbitrary unit.

40
Supplementary Figure 3: (A) 1 µg/ml HMGB1 was incubated with strips spotted
with a concentration gradient of the indicated phospholipids, then washed and
incubated with a rabbit anti-HMGB1 Ab before developing with an anti-rabbit IgG-
HRP. Data shown are representative of two experiments (left). Right panel, a PIP strip
assay only with an anti-rabbit IgG-HRP for a negative control. a: 100 pmol/spot; b: 50
pmol/spot; c: 25 pmol/spot; d: 12.5 pmol/spot; e: 6.25 pmol/spot; f: 3.13 pmol/spot; g:
1.56 pmol/spot. (B) A concentration series of HMGB-1 (0, 2.5, 5, 10, and 20 µg/ml)
was flowed over immobilized PS or PC liposomes on the sensor chip. The response
units are presented as the response to PS liposome binding.
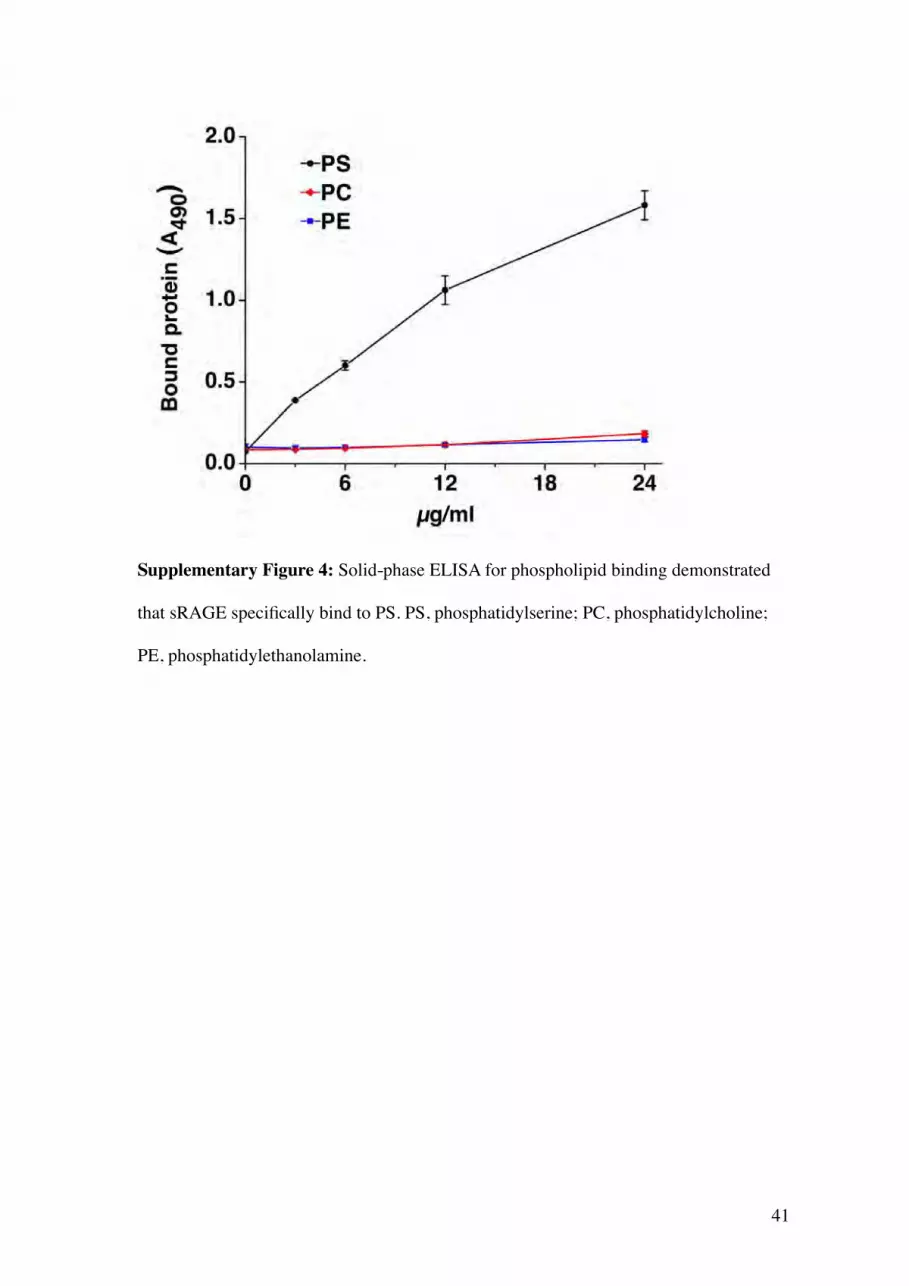
41
Supplementary Figure 4: Solid-phase ELISA for phospholipid binding demonstrated
that sRAGE specifically bind to PS. PS, phosphatidylserine; PC, phosphatidylcholine;
PE, phosphatidylethanolamine.

42
Supplementary Figure 5: Rage-/- peritoneal macrophages impair phagocytosis of
apoptotic thymocytes in vitro. Wild-type or Rage-/- peritoneal macrophages (PM)
incubated with apoptotic thymocytes for 30 min, and the number of apoptotic cells
per macrophage uptake was determined as the phagocytic index. Each condition was
tested three times in duplicate and samples were blindly and independently analyzed
by two researchers (M.H. and K.M.). The average value is shown with SEM. A one-
tailed t test was used to analyze the difference.





























