Embed Size (px)
Citation preview

Recruited Brain Tumor-Derived Mesenchymal StemCells Contribute to Brain Tumor Progression
JINAN BEHNAN,a,b PAULINE ISAKSON,c MRINAL JOEL,a,d CORRADO CILIO,e IVER A. LANGMOEN,a
EINAR O. VIK-MO,a* WIAAM BADNb,e*
Key Words. Glioma • Recruited cells • Mesenchymal stem cells • Cell surface markers • Cellmigration • Multipotential differentiation • Cell culture • Cell adhesion molecules •
Cell biology
ABSTRACT
The identity of the cells that contribute to brain tumor structure and progression remainsunclear. Mesenchymal stem cells (MSCs) have recently been isolated from normal mouse brain.Here, we report the infiltration of MSC-like cells into the GL261 murine glioma model. Thesebrain tumor-derived mesenchymal stem cells (BT-MSCs) are defined with the phenotype (Lin-Sca-11CD91CD441CD1661/2) and have multipotent differentiation capacity. We show thatthe infiltration of BT-MSCs correlates to tumor progression; furthermore, BT-MSCs increased theproliferation rate of GL261 cells in vitro. For the first time, we report that the majority ofGL261 cells expressed mesenchymal phenotype under both adherent and sphere culture condi-tions in vitro and that the non-MSC population is nontumorigenic in vivo. Although the GL261cell line expressed mesenchymal phenotype markers in vitro, most BT-MSCs are recruited cellsfrom host origin in both wild-type GL261 inoculated into green fluorescent protein (GFP)-trans-genic mice and GL261-GFP cells inoculated into wild-type mice. We show the expression of che-mokine receptors CXCR4 and CXCR6 on different recruited cell populations. In vivo, the GL261cells change marker profile and acquire a phenotype that is more similar to cells growing insphere culture conditions. Finally, we identify a BT-MSC population in human glioblastoma thatis CD441CD91CD1661 both in freshly isolated and culture-expanded cells. Our data indicatethat cells with MSC-like phenotype infiltrate into the tumor stroma and play an important rolein tumor cell growth in vitro and in vivo. Thus, we suggest that targeting BT-MSCs could be apossible strategy for treating glioblastoma patients. STEM CELLS 2014;32:1110–1123
INTRODUCTION
Glioblastoma multiforme (GBM) is the mostcommon and aggressive primary tumor of thecentral nervous system tumors. The median sur-vival of patients is less than 15 months despitetumor resection followed by radio- and chemo-therapy [1]. The poor prognosis of GBM is aresult of highly infiltrative cells that spreadaway into the normal brain parenchyma escap-ing the surgical removal of the tumor mass anddevelop a high level of drug resistance throughtheir cellular plasticity and heterogeneity [2].Thus, it is important to understand the biologyof the cells that form the tumor structure andplay a role in tumor progression and invasion.
Mesenchymal stem cells (MSCs) or multi-potent mesenchymal stromal cells are nonhe-matopoietic stem cells characterized with self-renewing and multipotency properties [3].Identification and isolation of MSCs from dif-ferent organs is dependent on a combinationof criteria including morphology, surfacemarker phenotype, and differentiation potential.
These criteria include (1) adherent growth onplastic surface [4], (2) differentiation into osteo-genic, chondrogenic, and adipogenic lineage,and (3) marker expression profile. Although adefined panel of markers has been suggested inhumans [5], so far no standard criteria havebeen proposed in mice. Many investigatorsreported stable identification and isolation ofmurine MSCs using a lineage2, Sca-11, andCD91 phenotype [6, 7]. In addition, CD44 andCD166 are considered as common markersmainly in human MSCs [8]. In glioma, the expres-sion of CD44 was found to be correlated withtumor grade and to affect cell invasion [9, 10].Kijima et al. reported recently that CD166 wasfound to enrich for brain tumor stem cells andpromote invasion [11]. Thus, a combined panelof the following surface markers (Lin-Sca-11CD91CD441CD1661) could be a useful toolto identify the brain tumor-derived mesenchy-mal stem cells (BT-MSCs).
MSCs have traditionally been isolated frombone marrow, but probable pericyte-derivedtissue MSCs have been identified in several
aVilhelm Magnus Laboratory,Institute for SurgicalResearch, CAST-Cancer StemCell Innovation Center andNorwegian Center for StemCell Research, Oslo UniversityHospital, Oslo, Norway;bGlioma ImmunotherapyGroup, Institute for ClinicalSciences, Department ofNeurosurgery, LundUniversity, Lund, Sweden;cDepartment of Biochemistry,Institute for Basic MedicalScience, University of Oslo,Oslo, Norway; dDepartmentof Physiology, Institute forBasic Medical Science,University of Oslo, Oslo,Norway; eDepartment ofClinical Sciences, CellularAutoimmunity Unit, LundUniversity, Malm€o UniversityHospital, Malm€o, Sweden
*contributed equally.
Correspondence: Jinan Behnan,PhD candidate, Vilhelm MagnusLaboratory, Institute for SurgicalResearch, Department ofNeurosurgery, University ofOslo, 0027 Oslo, Norway.Telephone: 14748340740; Fax:14723071397; e-mail:[email protected]
Received May 20, 2013;accepted for publicationOctober 23, 2013; firstpublished online in STEM CELLS
EXPRESS December 3, 2013.
VC AlphaMed Press1066-5099/2014/$30.00/0
http://dx.doi.org/10.1002/stem.1614
Stem Cells 2014;32:1110–1123 www.StemCells.com VC AlphaMed Press 2013
CANCER STEM CELLS

organs. Recently, Kang et al. described the presence of MSCsin the adult murine brain [6, 7, 12]. The homing property ofMSCs to brain tumors has been investigated in many studies,where they showed that intracranial or intra-arterialimplanted bone marrow MSCs (BM-MSCs) possess migratoryability to the site of the tumor with efficient distributionwithin the tumor stroma and integration into the tumor neo-vascularity to act as progenitors of pericytes [13, 14]. However,their role in brain tumor growth is not well understood as theycan both accelerate and inhibit tumor growth [15–17]. Severalreports have classified glioblastoma biopsies according to theirgene expression patterns [18, 19]. Glioblastomas dominated bymesenchymal-gene expression exhibit shorter survival, andupon tumor recurrence, there is a clear shift toward a mesen-chymal signature and loss of neuronal similarity [18]. Here, wecharacterize the cellular composition in the GL261 murine gli-oma model aiming to evaluate the presence of MSCs andexplore their role during tumor progression. We show the infil-tration of endogenous BT-MSCs (Lin-Sca-11CD91CD441
CD1661/2). Interestingly, the cell line GL261 expresses thesemarkers in vitro. The mesenchymal marker negative populationof the GL261 line is nontumorigenic, while infiltrative BT-MSCsare tumorigenic. Together our results suggest that BT-MSCscould play an important role during tumor progression andcould therefore be a separate important therapeutic target.
MATERIALS AND METHODS
Animal and Cell Culture
Eight- to 10-week-old female C57Bl/6 (Scanbur BK AB Sollen-tum, Sweden) and C57BL/6-Tg(UBC-green fluorescent protein[GFP])30Scha/J (Jackson, TN) mice were housed in the animalfacility of BMC, Lund University Hospital and Department ofComparative Medicine, Oslo University Hospital, under stableconditions of humidity and temperature controlled vivariumon a 12:12 hours light–dark cycle with free access to foodand water. All experimental procedures were carried outaccording to the guide of Swedish Board of Animal Researchand the Norwegian National Committee for Medical ResearchEthics. The GL261 mouse glioma cell line was obtained fromthree different suppliers: Dr. Safarany (Budapest, Hungary), Dr.Pellegatta (Milan, Italy), and NCI-Frederick (New York, USA).The GL261 cell line was grown as adherent cells (GL-Ad) inR10 medium: RPMI 1640 (GIBCO-BRL, Paisley, UK) supple-mented with 10% fetal bovine serum (FBS) (Biochrom AB, Ber-lin, Germany), 1 mM sodium pyruvate, 10 mM HEPES, and 50mg/ml gentamicin (Invitrogen AB, Sweden). GL261 cells werealso grown as spheres (GL-Sp) in serum free medium Dulbec-co’s modified Eagle’s medium (DMEM)/F12-GlutaMAX (LifeTechnologies, Sweden) supplemented with penicillin/strepto-mycin sulfate, 2% B-27 (Life Technologies, Sweden), 10 ng/mlbasic fibroblast growth factor (R&D Inc, Minneapolis), and 20ng/ml epidermal growth factor (EGF, R&D Inc), and 2.5 mg/mlheparin (LeoPharma AS). For protocols of lentiviral labeling ofGL261 cells with GFP, isolation and expanding of BM/or com-pact bone (CB)-MSCs please refer to Supporting InformationExperimental Procedures. Fibroblasts (FB) were a generousgift from Dr. Siva (Lund, Sweden). Adipose tissue-derivedMSCs from subvascular fraction (AT-MSCs) were kind gift fromDr. Moskaug (Oslo, Norway). The cells were kept in culture at
37�C in a humidified 5.5% CO2 atm. Population doubling time(PDT) was calculated using the following formula: PDT5 t log2/log Nt2 log No; where t is time period, Nt is number ofcells at time t. Cell senescent state was quantified by X-galstaining to detect senescence-associated b-galactosidase activ-ity (SA-bgal) [20].
Human Samples and Culture
The brain tumor biopsies were acquired from five consentingGBM patients, human GBM (hGBM). The cells were obtainedby ultrasonic aspiration with a Sonoca 300 ultrasonic dissec-tor/aspirator (S€oring, Quickborn, Germany) peroperatively.Cells were washed and spun down to remove erythrocytes.Tumor cells were grown in DMEM/F12-GlutaMAX mediumsupplemented with 1% FBS, 2% B-27, 10 ng/ml basic fibro-blast growth factor, 20 ng/ml TGFa, 10 mM HEPES (Lonza, Bio-Whittaker), and 2.5 mg/ml heparin (LeoPharma AS) [21].
Brain Tumor Model
GL261 or GL261-GFP cells (10,000 cells in a volume 5 ml RPMI1640) were intracranially injected in the right striatum of C57BL/6 mice (0.5 mm forward from bregma, 1.5 mm lateral, and 2.5mm ventral from the dura) using a 10-ml Hamilton syringe with a33-gauge needle. To inhibit the back-flow throw the injectioncanal, the needle was left in the brain for 2 minutes and with-drawn gradually (1 mmwith waiting time for 2 minutes). Animalswere allowed to recover; their health status was closely moni-tored, and immediately killed when showing neurological symp-toms. Animals were scored on a standardized pain score andwere euthanized when one or more of the following symptomswere present: weight loss [mt]15%, lethargy, slow motion,hunched posture, and anorexia. The median survival of tumorbearing mice was around 21 days after cell inoculation.
Flow Cytometry Analysis and Sorting
Freshly isolated cells were first incubated with 2% Fc block toinhibit nonspecific binding for 10 minutes. Subsequently, about3.105 cells were stained with biotinylated or directly conjugatedantibodies at saturating concentrations for 30 minutes at 4�C.Subsequently, cells were washed once with Phosphate bufferedsaline (PBS) and resuspended in 500 ml PBS supplemented with2% Fetal bovine serum (FBS). The following antibodies wereused; anti-mouse antibodies: CD45-Alexa fluor 700, CD11b-FITC,CD31-PE-Cy7, CD166-PE, CD9- FITC, or APC, PDGFR-a-APC, conju-gated PDGFR-b-APC, CXCR4-APC, CD133-PE, m/hCD44-APC(eBioscience, San Deigo, CA), m/h-SSEA-4-APC (R&D systems,Minneapolis, MN ), Sca-1-Pacific blue, CD105-APC, CD106-APC,CD90-APC, CD73-APC (Biolegend, San Diego, CA), rabbit-Nestin(1:500, Millipore, Billerica, MA), rabbit-Glial fibrillary acidicprotein (GFAP) (1:1000, Dako, Glostrup, Denmark), rabbit-CXCR6(Novus Biologicals, Littleton, CO), rabbit-NGFR (abcam, Cam-bridge, UK), and anti-m/hAPC-A2B5 (Miltenyi, Billerica, MA).Anti-human antibodies: APC-Cy7-CD45 (BD Pharmengin), Pacificblue-CD31, PE-CD166, PE-CD9, APC-CXCR4 (eBioscience, SanDeigo, CA), mouse-anti-human Nestin (Millipore, Billerica, MA),mouse-anti-human GFAP (R&D, Minneapolis, MN), and thematched secondary antibodies (anti-rabbit Alexa 594, anti-rabbit, and anti-mouse Alexa 488 1:500, Molecular Probes, Invi-trogen, Junction City, Oregon). To stain the cells with GFAP andNestin, the cells were incubated with primary antibody for 2hours at 4�C, washed, and then incubated with secondary
Behnan, Isakson, Joel et al. 1111
www.StemCells.com VC AlphaMed Press 2013

antibody for 1 hour. Cells were then washed twice before beinganalyzed by flow cytometer LSRII (BD Bioscience). At least10,000 events were counted.
For cell sorting, cells were resuspended in PBS, 4% FBS to afinal concentration (2–4 3 106 cells per milliliter). The followingmarkers were used for Fluorescence-activated cell sorting(FACS) sorting: 7AAD to exclude dead cells, FITC or Pacific blueconjugated anti-Sca.1, PE-Cy7 conjugated anti-CD31, CD45,CD11b, and Terr129, APC anti-m/hCD44, and PE conjugatedanti-CD166. Sorting was performed on FACS Aria I (BD Bio-science, San Jose, California), sometimes, anti-Sca-1 microbeadskit (FITC) for positive selection of murine Sca-1 was used forpopulation enrichment before sorting (Miltenyi Biotec, Cologne,Germany). Single staining and beads were used for compensa-tion, and sorting gates were set according to FluorescenceMinus One controls. Data acquisition was performed using theCell Quest or DiVa software (BD, Franklin Lakes, New Jersey),and FlowJo software was used for data analysis.
In Vitro Characterization of BT-MSCs
Colony Forming Unit Fibroblast Assay. Cells were plated induplicate in six-well plates at a density of 1 3 103 and 5 3 103 incomplete MSC medium; after 10 days, adherent cells werewashed with PBS, fixed with methanol for 5 minutes, and stainedwith 0.5% crystal violet (Sigma) for 1 minute at room tempera-ture (RT). Cluster of�40 cells were considered as colonies.
Differentiation Assays
Osteoblastic Differentiation. The MSC cells were grown inosteogenic differentiation medium (Invitrogen AB, Sweden)for 2 weeks, and Alzarin red staining was used to visualize cal-cium deposition.
Adipogenic Differentiation. Cultured MSCs were grown in adi-pogenic differentiation medium (Invitrogen AB, Sweden). Theinduction of adipocytes was assessed after 2 weeks using oil redO stain (Sigma) as an indicator of intracellular lipid accumulation.
Chondrogenic Differentiation. Chondrogenic induction wasperformed by pelleting cultured cells (5 3 105 cells per pellet)in polypropylene 15 ml tube and place the pellets in 0.5 mlof chondrogenic differentiation medium (Invitrogen AB, Swe-den). Chondrocyte differentiation was evaluated by aggrecanstaining after 4 weeks. In all three assays, the medium wasreplaced with fresh twice a week and controls were grown inMSC-medium. See details in Supporting Information.
Immunohistochemistry and Immunocytochemistry
Ten micrometers of cryostat sections were prepared from fro-zen brain tumor after it has been fixed in 4% paraformaldehydefor 24 hours, and cryoprotected in 20% sucrose for 24 hoursmore. To perform immunohistochemistry (IHC) staining, frozensections were allowed to be defrosted for 15 minutes, rehy-drated 3 3 5 minutes in PBS, and then 2 3 5 minutes in Tris-Buffered Saline Tween-20 (TBST) and incubated with blockingbuffer (5% milk powder in TBST and 10% normal serum of sec-ondary antibody host) for 30 minutes at RT. The following anti-bodies were used: goat anti-mouse Sca-1 (1:40, R&D system,Minneapolis, MN), rat anti-mouse CD45 (1:150), and rabbitanti-GFP (1:200, Abcam, Cambridge, UK). Slides were incubatedovernight at 4�C, washed 3 3 5 minutes in PBS followed by 2 3
5 minutes in TBST, and then incubated with anti-different hostsecondary antibodies conjugated to Alexa Fluor 488, Alexa Fluor594, Cy5 (1:400), and incubated overnight at 4�C, washed 3 3 5minutes in PBS and 1 3 5 minutes in Hoechst (1:1000, SigmaAldrich) before being mounted with Prolong Gold anti-fade rea-gent (Invitrogen). High-resolution images of histological sec-tions were acquired using an automated slide scanner systemand image analysis was performed by MIRAX Viewer (MiraxScan, Carl Zeiss MicroImaging, Jena, Germany).
For confocal microscopy, cells were grown on cover slips(Thermo Scientific), incubated with the indicated primary anti-bodies for 2 hours, and appropriate secondary antibodies for1 hour at RT, gentle washing after each step for 2 3 2minutes in PBS. Cover slips were examined using an OlympusFluoview 1,000 confocal microscope. Image processing andanalysis were carried out with Olympus Fluoview softwareand Adobe Photoshop (Adobe Systems).
Proliferation Assay
GL261 cells were stained with Carboxyfluorescein succinimidylester (CFSE) (0.001 mM) for 5 minutes at 37�C and coculturedwith BT-MSCs with four different percentages (1:1, 2:1, 5:1, and10:1) GL261 to MSCs. Unstained GL261 were used as negativecontrol for FACS analysis. The assay was read after 24, 48, and 72hours (assay in details could be seen in Supporting Information).
Statistical Analysis
Survival analysis and the Mantel-Cox log-rank were performedusing GraphPad Prism 6 (GraphPad Software, LA Jolla, CA).
RESULTS
GL261 Brain Tumors Contain MSC-like Cells that AreColony Forming and Have Multilineage DifferentiationCapacity
To explore the possible presence of MSCs in brain tumors,GL261 cells were inoculated into the brain of syngeneicC57BL/6 mice. Tumors were harvested when mice developedsymptoms after 20–28 days. Analysis of single cell suspensionsshowed that 55%6 2.1% (mean6 SEM; n5 9) of the viablecells were negative for the hematopoietic markers CD45 andCD11b as well as the endothelial cell marker CD31. In thisnegative compartment, 536 3.4% of the cells were positivefor the stem cell marker Sca-1 (Fig. 1A, 1B the third row). Inthe Sca-1 positive compartment, 816 2.8% were CD91,886 3.5% were CD441, and 586 3.2% were CD1661 (Fig.1C–1E the third row). However, CD140a/PDGFRa expressionwas on BT-MSCs, while it was highly expressed on the naiveBM-/CB-MSCs used as positive control (Fig. 1D). The culturedFB used as negative control cells did not show Sca-1 expres-sion at all as expected (Fig. 1B, the fourth row).
Brain tumor Lin-Sca-11 cells were FACS sorted and puta-tive BT-MSCs were expanded on a plastic surface in adherentconditions. Further characterization with wide range of BM-MSCs and AT-MSCs markers was carried out on the sorted BT-MSCs (Supporting Information Table S1). BT-MSCs appearedrelatively distinct from BM- and AT-MSCs, with higher levelsof Sca-1, CXCR4, CD271, and lower level of CD105 and CD106.Overall, the most specific marker for BT-MSCs were theexpression of high levels of Sox2, and the neural stem cell
1112 Brain Tumor-Derived Mesenchymal Stem Cells in GL261
VC AlphaMed Press 2013 STEM CELLS

marker Nestin. This finding suggests that the origin of therecruited BT-MSCs might be another source than bone mar-row or adipose tissue.
The sorted BT-MSCs grew as colonies and had fibroblas-toid morphology similar to CB- and BM-MSCs (Fig. 2A). BT-MSCs were able to differentiate into three lineages, osteo-,chondro-, and adipocyte similar to BM and CB-MSCs, whereasFBs did not differentiate (Fig. 2B–2D). Thus, confirming thepresence of brain tumor derived cells fulfilling the criteria ofMSCs.
BT-MSCs Accumulate upon Late Brain TumorProgression
GL261 tumor growth develops over distinct stages [22]. Toinvestigate whether the accumulation of BT-MSCs is correlatedwith tumor progression, the tumor bearing mice were killed
either at 2 or 3 weeks after inoculation. The infiltration oflin-Sca-11 cells 2-weeks after tumor inoculation was very low,0.56 0.1% (n5 6) (Fig. 3B second row) and significantly dif-ferent compared with 3 weeks grown tumors with 316 3.2%(n5 6) p5 .002 (Fig. 3B third row), whereas in the sham con-trol it was 0.026 0.002% (n5 5) (Fig. 3B first row). Thus,increased infiltration of BT-MSCs upon tumor growth mightreflect an important role of these cells in tumor progression.
The GL261 Cell Line Expresses MSC-like Phenotype inAdherent Cell Culture
The GL261 cell line is commonly cultivated as monolayer cul-tures in medium containing 10% serum (GL-Ad). Unexpectedly,flow cytometric analysis showed that the cell line expressesthe marker profile of BT-MSCs (Lin-Sca-11) 946 1.8%.Although all Sca-11 cells were CD91CD441 only 146 2.9%
Figure 1. Brain tumor-derived mesenchymal stem cells (BT-MSCs) express the mesenchymal surface markers Sca-1, CD9, CD166, CD44,and low CD140a. When animals showed symptoms following tumor inoculation, tumors were isolated and digested. Single cell suspen-sions were stained with AF-700-CD45, PE.Cy-7-CD31, FITC-CD11b, Pacific blue-Sca-1, PE-CD166, APC-CD9, or replacing CD9 with CD140aor APC-CD44 in other panels. Bone marrow (BM)-derived MSC and compact bone (CB)-derived-MSC from adherent culture were used aspositive control and primary cultured fibroblasts were used as negative control. The expression of Sca-1, CD9, CD166, CD44, andCD140a was analyzed in the negative compartment for CD45, CD11b, and CD31 after the exclusion of dead cells. In BT-samples, twosubpopulations were defined according the marker expression within CD45-CD11b-CD312 portion, Sca-11CD91CD441CD1661/orCD1662 (A–D) in third row. All BT samples were analyzed similarly. The whole strategy is represented with sample-1; the same stepswere followed in positive and negative controls. Abbreviations: AF, alexa fluor; BM, bone marrow; BT-MSC, brain tumor-derived mesen-chymal stem cells; CB, compact bone; FB, fibroblasts.
Behnan, Isakson, Joel et al. 1113
www.StemCells.com VC AlphaMed Press 2013

were CD1661. This finding was confirmed in GL261 cell linesobtained from three independent sources. To evaluate theexpression of other stem cell markers related to human gli-oma, the expression of following markers was investigated;CXCR4 2.66 0.3%, CD133 2.46 0.3% (Fig. 4A), SSEA-41.26 0.1%, similar to our in vivo data the mesenchymalmarkers, PDGF-a and PDGF-b were not expressed (SupportingInformation Fig. S1). The expression of neural stem cellmarker Nestin was 476 7%, the glial marker GFAP, and theglial progenitor marker A2B5 was 986 0.4% and 9.06 1.9%,respectively (Supporting Information Fig. S2). This data clearlydisplay a neuroepithelial–mesenchymal dualism in markerexpression of the GL261 cell line.
GL261 Lose Sca-1 and Gain CXCR4 Expression UnderSphere Forming Conditions
Higher biological validity is suggested for primary cells culti-vated under serum free- and growth factor enhanced condi-tions, that mimic primary tumors [23, 24], this might also betrue for established cell lines like GL261 [25]. Interestingly,GL261 cells grown in sphere forming condition (GL-Sp)showed much lower expression of BT-MSC markers. The Lin-Sca-11 population dramatically decreased 35 times to2.76 0.1% compared with GL-Ad. Although the frequency ofCD9 and CD44 in GL-Sp was not changed to 986 0.1% and986 0.5%, respectively, the expression of CD166 was halved
to 6.36 0.5% (Fig. 4A). On the other hand, the expression ofCXCR4 was increased 13 times to 346 1.5%. The expressionof Nestin and A2B5 was increased to 596 11%, 326 5.1%respectively, while the level of GFAP was not changed976 0.5% (Supporting Information Fig. S2).
GL-MSC-Population Have Limited DifferentiationCapacity and Are Nontumorigenic In Vivo
To investigate the mesenchymal phenotype and its role intumor initiation, GL261 cells were sorted into two populationsGL-MSC1 (Lin-Sca-11) and GL-MSC- (Lin-Sca-12) (Fig. 4B).Sorted cells were expanded in culture; GL-MSC1 cells couldrepopulate the general marker profile of GL-Ad with a highfrequency of Sca-11 cells and a small population of Sca-12
cells. In contrast, GL-MSC- cells could not reestablish a Sca-11 population. The PDT of GL-MSC1 and GL-MSC- were notdifferent (296 3.9 hours vs. 316 4.1 hours, respectively). Theviability of cells upon passaging or before tumor inoculationwas not different, 946 1.2% and 946 2.1%, respectively. Invitro multilineage differentiation assays showed that GL-MSC1 cells had the capacity to differentiate into osteo-,adipo-, and chondrocytes, whereas GL-MSC- cells could onlydifferentiate along the osteoid-lineage (Fig. 4C). GL-MSC cellswere not able to form chondrocyte aggregates, thereby pre-cluding immunostaining. Crucially, upon orthotopic transplan-tation only GL-MSC1 cells formed tumors, while GL-MSC cells
Figure 2. Brain tumor-derived mesenchymal stem cells (BT-MSCs) form adherent colonies to plastic surface with fibroblast like shapeand have multilineage differentiation capacity. BT-MSCs were sorted by FACS. Bone marrow (BM) and compact bone (CB)-derived MSCswere used as positive controls, whereas FB cells were used as a negative control. (A): Cells growing in colonies showed a typical spindlefibroblastic-shaped morphology (magnification 340). Cells were cultured in six-well plate at 10 [3] cells per well. After 10 days, culturedcells were stained with crystal violet. (B–D): Multilineage differentiation capacity was determined by culturing MSCs in osteo-, chondro,and adpocyte differentiation medium. (B): Alizarin red staining for osteogenic-induced differentiation of BT-MSCs was compared withBM, CB, and FB. (C): Aggrecan staining on chondrocyte pellet section. The pellets were allowed to differentiate in chondrocyte inductionmedium (chondro-med) for 4 weeks, then fixed and frozen. Later, the pellets were sectioned into 6-mm-thick sections (cryotom). Chon-dropellets were stained with a primary antibody against aggrecan (R&D system) and appropriate AF488 secondary antibodies, in additionto nuclear staining with 4,6-diamidino-2-phenylindole (DAPI). Images were acquired by Olympus BX61 (scale bar5 100 mm, magnification310). (D): Oil red O staining for lipid droplets of adipogenic induced differentiation of BT-MSCs (scale bar5 50 mm, magnification 320).Abbreviations: BM, bone marrow; BT-MSC, brain tumor-derived mesenchymal stem cells; CB, compact bone; FB, fibroblasts.
1114 Brain Tumor-Derived Mesenchymal Stem Cells in GL261
VC AlphaMed Press 2013 STEM CELLS

could not form tumors even after 120 days of observation(Fig. 4D). Thus, the mesenchymal population is necessary fortumor initiation.
The BT-MSCs Are of Host Origin Recruited to theTumor
Following our discovery of the GL-MSC1 population, we askedwhether BT-MSCs could be derived from the host (endogenous)or from the inoculated tumor cells (exogenous). To investigatethe origin of BT-MSCs, we used two strategies: (1) wild-typeGL261 (WT-GL261) cells inoculated into mice that expressenhanced-GFP under the control of UBC-promoter, (2) lentiviraltransduced GL261 cells with GFP (GL-GFP1) inoculated intowild-type mice. Analysis of brain tumor samples in the firststrategy showed that 216 1.3% of the total cells were Lin-Sca-11, where 856 1.6% of these cells were GFP1 derived fromthe host. Of the GL261 (GFP-) derived cells, only 2.66 0.5%were Sca-11. Similar results were obtained using the secondstrategy, the GL-GFP in wild-type mice. This time 376 5.3% ofthe total cells were Lin-Sca-11 and 856 2.8% of this population
were of host origin (GFP2). A small portion of the GFP-Sca-11
population 156 4.8% were Nestin1 and 13.56 2.3% wereCD311. However, only 1.76 0.3% of the GL-GFP1Sca-11 wereNestin1. As much as 126 3% were CD311 (Fig. 5A). IHC stain-ing with anti-GFP antibody confirmed the data obtained by flowcytometry and showed that the majority of the cells that formthe tumor stroma were GFP-Sca-11 (Fig. 5I). The GL-GFP1 cellswere mostly Sca-12 (Fig. 5B; Supporting Information Fig. 3),confirming again that Gl261 cells lose Sca-1 expression in vivo.
CXCR4 is Expressed Mainly by Recruited Cells andPartly by Tumor Inoculated Cells, While CXCR6Expression Is Restricted to Recruited CD451 Cells
Under sphere forming condition in vitro GL261 cells hadincreased CXCR4 expression. CXCR6 has also been suggestedto be important for the recruitment of MSCs into prostatecancer [26]. To explore whether CXCR4 and CXCR6 areexpressed in our glioma model, we did flow cytometry analy-sis on fresh samples derived from transplanted GL-GFP in WT-mice. Our results showed that 176 3.2% of total tumor cells
Figure 3. Increased accumulation of brain tumor-derived mesenchymal stem cells (BT-MSCs) with increased tumor progression. Thepresented FACS plot was gated on the CD45-CD11b2 compartment as presented in (Fig. 1), then gated on CD31-Sca-11(B) to see theexpression of CD9, CD44, and CD166 (C–D). In the sham-injured brains (used as controls), the expression of MSCs markers was low, lessthan 0.026 0.002% (as shown in (B) in the upper row), after 2-weeks (early-stage) of tumor injection, the expression of Sca-1 wasincreased up to 0.56 0.1% (shown in (B) the middle row), and after 3-weeks (late stage, when animals show symptoms) of tumor pro-gression, the Sca-11 cells constitute 436 3.9% of total linage negative compartment (B the third row). Abbreviation: BT-MSC, braintumor-derived mesenchymal stem cells.
Behnan, Isakson, Joel et al. 1115
www.StemCells.com VC AlphaMed Press 2013

Figure 4. GL261s express mesenchymal characteristics and tend to lose Sca-1 and gain CXCR4 expression in sphere forming conditions.Surface marker expression of in vitro growing GL261s in both adherent (GL-Ad) and sphere (GL-Sp) conditions. GL-Ad express the mes-enchymal phenotype of brain tumor-derived mesenchymal stem cells (Sca-11CD441CD91CD1661/2), while GL-Sp lose Sca-1 expres-sion and gain CXCR4. (B): Sorting strategy of two populations: Lin-Sca-11 (GL-MSC1) and Lin-Sca-12 (GL-MSC2). GL-MSC2 wereexpanded in culture and they kept the same phenotype. (C): Both GL-MSC1 and GL-MSC2 showed osteogenic differentiation capacity,while only GL-MSC1 but not GL-MSC2 showed adipogenic differentiation capacity (scale bar5 50 mm, magnification 320). (D): Thelack of mesenchymal phenotype in GL261 turned off the tumorigenicity of the cell line in vivo. The tumorigenicity of sorted GL-MSC2cells was compared with that of GL-MSC1 cells by transplanting similar cell numbers (1 3 104 cells). Animals injected with GL-MSC1cells developed tumors and showed neurological symptoms within the regular time (about 3 weeks), whereas animal injected with GL-MSC2 cells survived 120 days and were killed at the end point of the experiment without showing tumors (nine animals were injectedwith GL-MSC1 and 14 with GL-MSC2 cells). Abbreviations: Ad, adherent; MSC, mesenchymal stem cell; Sp, sphere.
1116 Brain Tumor-Derived Mesenchymal Stem Cells in GL261
VC AlphaMed Press 2013 STEM CELLS
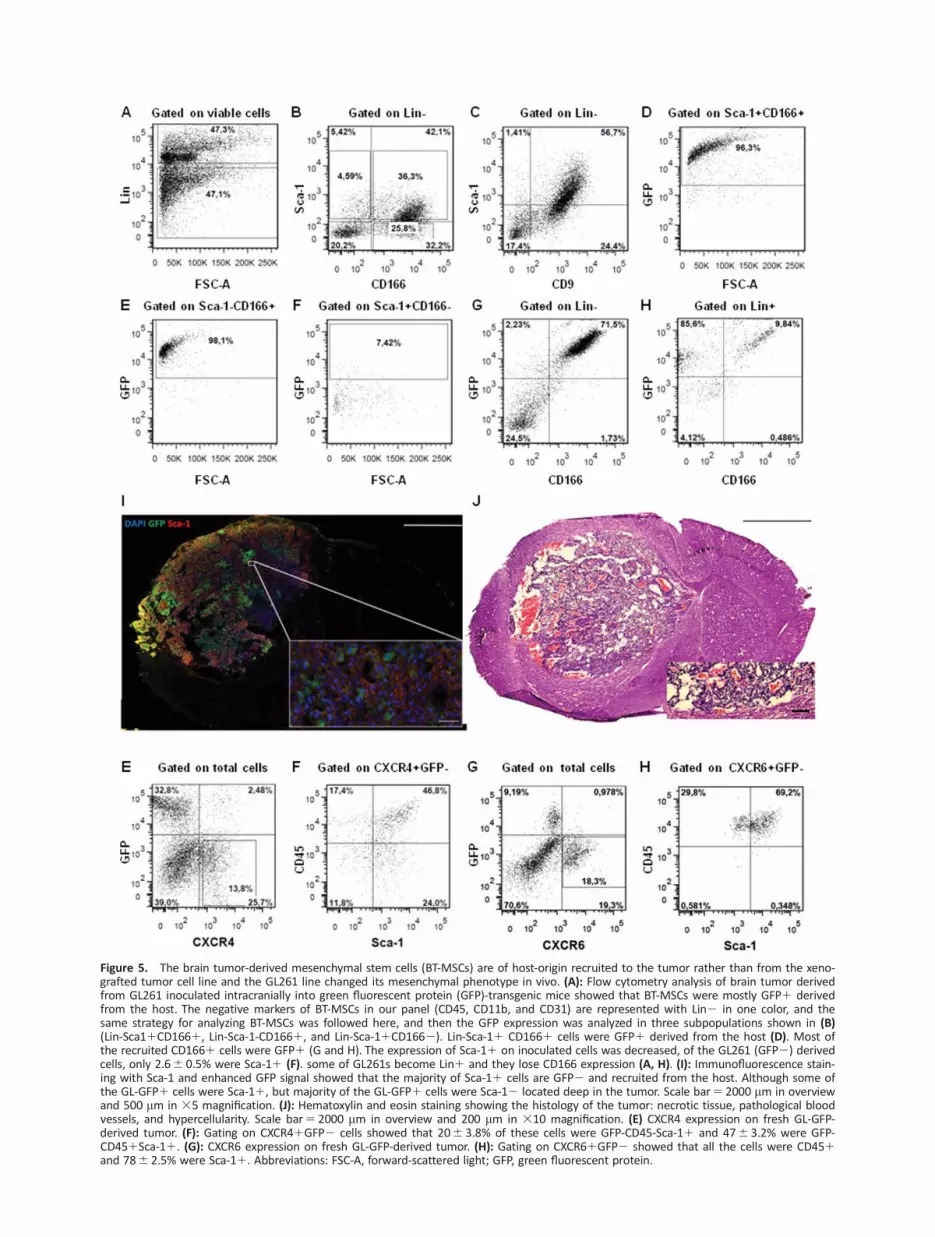
Figure 5. The brain tumor-derived mesenchymal stem cells (BT-MSCs) are of host-origin recruited to the tumor rather than from the xeno-grafted tumor cell line and the GL261 line changed its mesenchymal phenotype in vivo. (A): Flow cytometry analysis of brain tumor derivedfrom GL261 inoculated intracranially into green fluorescent protein (GFP)-transgenic mice showed that BT-MSCs were mostly GFP1 derivedfrom the host. The negative markers of BT-MSCs in our panel (CD45, CD11b, and CD31) are represented with Lin2 in one color, and thesame strategy for analyzing BT-MSCs was followed here, and then the GFP expression was analyzed in three subpopulations shown in (B)(Lin-Sca11CD1661, Lin-Sca-1-CD1661, and Lin-Sca-11CD1662). Lin-Sca-11 CD1661 cells were GFP1 derived from the host (D). Most ofthe recruited CD1661 cells were GFP1 (G and H). The expression of Sca-11 on inoculated cells was decreased, of the GL261 (GFP2) derivedcells, only 2.66 0.5% were Sca-11 (F). some of GL261s become Lin1 and they lose CD166 expression (A, H). (I): Immunofluorescence stain-ing with Sca-1 and enhanced GFP signal showed that the majority of Sca-11 cells are GFP2 and recruited from the host. Although some ofthe GL-GFP1 cells were Sca-11, but majority of the GL-GFP1 cells were Sca-12 located deep in the tumor. Scale bar5 2000 mm in overviewand 500 mm in 35 magnification. (J): Hematoxylin and eosin staining showing the histology of the tumor: necrotic tissue, pathological bloodvessels, and hypercellularity. Scale bar5 2000 mm in overview and 200 mm in 310 magnification. (E) CXCR4 expression on fresh GL-GFP-derived tumor. (F): Gating on CXCR41GFP2 cells showed that 206 3.8% of these cells were GFP-CD45-Sca-11 and 476 3.2% were GFP-CD451Sca-11. (G): CXCR6 expression on fresh GL-GFP-derived tumor. (H): Gating on CXCR61GFP2 showed that all the cells were CD451and 786 2.5% were Sca-11. Abbreviations: FSC-A, forward-scattered light; GFP, green fluorescent protein.

were CXCR41, 206 3.8% of these cells were GFP-CD45-Sca-11 (recruited BT-MSCs), and 476 3.2% were GFP-CD451Sca-11 (recruited hematopoietic stem cells). Resembling in vitrosphere growing condition, GL261 increased CXCR4 expressionin vivo, 196 6.0% of inoculated GL-GFP cells were CXCR41.Interestingly, the expression of CXCR6 was also high in freshsample, 146 1.5%, but its expression was restricted toCD451 cells only, 786 2.5% of these cells were Sca-11,while GL-GFP1 cells and recruited CD45-Sca-11 cells werecompletely negative for CXCR6. These data suggest thatCXCR4 might play important role in recruiting BT-MSCs, whileCXCR6 in concert with CXCR4 might affect the recruitment ofhematopoietic stem cells.
In Vitro Expanded BT-MSCs Have Similar MarkerExpression to Cultured GL261 Cells, with DifferentMorphology and Are Tumorigenic In Vivo
Tumors induced by GL261-GFP cells were sorted for isolationof BT-MSCs, which were expanded in culture and analyzed by
flow cytometry. Cells were kept in culture for up to 4 monthsand analyzed on different passages without changes in themarker expression (Fig. 6A). The proliferation rate was not sig-nificantly different between early and late passages and PDTwas not significantly changed upon a moderate number ofpassages in vitro (P2-5 vs. P10-13), 356 5.2 hours versus456 17 hours, as well, no change in cell viability, 946 1.2%and 946 2.1% respectively. The frequency of senescent cellswas very low (<1%) in sorted BT-MSCs at early passage (P2)and did not increase upon cell expansion in vitro (P10 andP20) (Fig. 6B). BT-MSCs in culture appeared similar to GL-Ad.Compared with GL261, the BT-MSCs were larger, had morecytoplasmic extensions, and some of the cells were multi-nucleated (Fig. 6C).
To explore the possible growth promoting effect of BT-MSCs, these cells were cocultured with CFSE-stained GL261cells. The proliferation rate of the GL261 cells was increasedin a dose-dependent manner (Supporting Information Fig. 4).There was a trend toward shorter survival in mice coinjected
Figure 6. Expanded brain tumor-derived mesenchymal stem cells (BT-MSCs) in vitro have similar phenotype to GL261s, different mor-phology and are tumorigenic. Tumors induced by GL261-green fluorescent protein (GFP) cells were sorted for isolation of BT-MSCs,which were expanded in culture and analyzed by flow cytometry. (A): Expression of Sca-1, CD9, CD166, CD44, CD133, and CXCR4 on BT-MSCs. (B): X-Gal staining for senescence-associated b-galactosidase activity. The frequency of senescent cells was very low (<1%) insorted BT-MSCs at early passage (P2) and did not increase upon cell expansion in vitro (P10 and P20), differentiated Br-MSC were usedas positive control. Scale bar5 250 mm. (C): BT-MSCs and GL261s were fixed and stained with Hoechst, CD45, Sca-1, and analyzed byconfocal microscopy. Scale bar5 10 mm. (D): Survival rate of intracranial transplanted animals with BT-MSCs, GL261s, and BT-MSC1GL261s. Abbreviations: BT-MSC, brain tumor-derived mesenchymal stem cells, Br, brain.
1118 Brain Tumor-Derived Mesenchymal Stem Cells in GL261
VC AlphaMed Press 2013 STEM CELLS

with BT-MSCs and GL261 cells. The median survival of miceinoculated with GL261 cells alone was 246 3.4 days, whilecoadministration of BT-MSCs with GL261 cells (50%) inducedtumor within 20.76 3.1 days, no effect was noticed usinglower dose (20%), 23.56 2.8 days, this last one was not dif-ferent from GL261 alone. Interestingly, upon orthotopic graft-ing of the BT-MSCs alone (P7), tumors developed within thesame time frame as for the GL261 cells. The tumors gener-ated from BT-MSCs displayed a phenotype that was similar toGL261-derived tumors with some differences. Both tumorscontained pleomorphic cells with nuclear atypia, hemorrhages,and neovascularization, but no microvascular proliferation.Tumors did not contain areas of fibrillar cells or reticulin, nordo they show invasive pattern into the brain. No necrotic
areas were found in BT-MSCs induced tumor and thesetumors appeared to have several growth centers (SupportingInformation Fig. S5 and Table S2). Thus, although recruitedfrom the host, these tumor-derived cells acquired tumorigeniccapacity and were able to induce tumor per se.
BT-MSCs Are Present in Human Glioblastoma Biopsiesand Show Changes in the Expression of CD166 andCXCR4 as GL261
Flow cytometry analysis of five GBM samples showed thatboth fresh tumors and culture expanded tumor cells expressthe mesenchymal profile; CD441CD91CD1661 surfacemarkers (Fig. 7A). In fresh primary GBM biopsies, the frequen-cies were: 646 5.9% CD441, 156 2.7% CD91, and
Figure 7. Brain tumor-derived mesenchymal stem cells (BT-MSCs) marker panel is relevant for human glioma and show, as GL261s, shift inCD166 and CXCR4 expression according to culture conditions. Expression of CD44, CD9, CD166, and CXCR4 was analyzed on primary and invitro expanded tumor cells in both adherent and sphere culture conditions. (A): CD441CD91CD1661/CXCR41 population was identifiedin primary tumor cells obtained from human ultrasonic biopsies. (B): Tumor cells CD441 CD91CD1661/CXCR41 (low) were enriched inadherent culture conditions. (C): CD441 CD91CD1661 (low) /CXCR41 were enriched in sphere culture. Five glioblastoma multiforme sam-ples were analyzed, and one representative tumor sample is represented. Abbreviation: FSC-A, forward-scattered light.
Behnan, Isakson, Joel et al. 1119
www.StemCells.com VC AlphaMed Press 2013

6.66 0.9% CD1661, while in culture expanded cells were786 4.2% CD441, 546 5.1% CD91, and 576 4.0% CD1661.Similar to GL261, the expression of CXCR4 was increased insphere culture conditions compared with adherent culturingconditions, from 146 4.8% in adherent culture to 336 2.1%in sphere cultures. The expression of CD166 was on the otherhand decreased from 576 4.0% to 316 19%. Overall, thesedata on human tumor cells are very similar to our data in themouse model.
DISCUSSION
The GL261 cell line is a widely used and well-characterizedglioma model. The orthotopic grafting of the cell line indu-ces brain tumors with complex genetic changes and signal-ing pathways, marker expression and tumor phenotypesimilar to human glioblastomas [27]. It allows a high repro-ducibility of tumor induction, and because of its syngeneicnature, it is possible to study the interaction between thetumor and the immune system. Thus, GL261 has been con-sidered the gold-standard in preclinical glioma immunother-apy [28]. However, surprisingly little is published of thesurface marker phenotype of the GL261-derived tumors orthe cell line itself.
The main goal of this article was to investigate the existenceof MSC like cells in GL261glioma mouse model and study thedetailed characteristics of the cellular content of the inducedtumor compared with human glioma. For this purpose, we haveused a marker panel suggested to identify mouse MSCs fromnormal brain and other sources [6–11]. We have identified thepresence of BT-MSCs (Lin-Sca-11CD91CD441CD1661) withclassical MSC colony formation and differentiation potential.These cells have similar surface marker expression to BM/CB-MSCs, but they lack the well known BM-MSC marker CD140a.Interestingly, as this gene is associated with the proneural glio-blastoma subtype, the low expression in the GL261-derivedtumors supports the notion of this model more resembling themesenchymal or classical glioblastoma subtype [19]. Previouslydescribed genetic changes in GL261s (loss of PTEN, p53, andEGFR amplification) have also been associated with these sub-types [19, 27]. The low expression of CD133 has previously beenidentified by others [29] and lends support to this interpretation[30]. The recently described tumor-derived endothelial cells [31]could also be demonstrated in our model. Finally, the identifica-tion for BT-MSCs (Lin-CD441CD91CD1661) in human gliomasamples suggests that these cells are not model specific, butlikely are clinically relevant.
To our knowledge, our data highlight for the first time themesenchymal characteristics of the GL261 cell line understandard culturing conditions. As cross contamination is amajor concern working with a long established cell line [32],we have used GL261 from three independent sources. Theyall demonstrated the same results. The presence of neuroepi-thelial markers, however, suggests a neuroepithelial–mesen-chymal dualism in the cell line. The changing levels of nestinand A2B5 depending on culturing conditions that we find haspreviously been identified by others using IHC [33]. This lendssupport not only to the authenticity of the cell lines used, butalso fits a pattern that would be expected by neuroeptihelialcells.
Although the GL261 cell line contained a population withthe marker profile of BT-MSCs in vitro, both the WT-GL261transplanted into GFP-transgenic mice and GL261-GFP cellstransplanted into WT-mice showed that Sca-11CD91CD441CD1661/2 cells are mainly recruited fromthe host. The infiltration of MSCs into the tumor area mightbe “a host defense mechanism” in original to suppress tumorprogression; this hypothesis came in line with new datareporting the suppressive effect of hBM-MSCs on hGBMthrough inhibiting tumor angiogenesis [34]. Another reasoncould be that the tumor induces recruitment of proinflamma-tory MSCs where they become immunosuppressive MSCsunder the effect of tumor microenvironment. Recently it wasreported that MSCs can be induced to become either proin-flammatory MSC1 that suppress tumor growth, r immunosup-pressive MSC2 that favor tumor growth upon activation ofdifferent toll-like receptors [35]. The recruitment of MSCs intothe tumor might play important role in tumor progressionand modification of tumor microenvironment. It was foundthat MSCs exert an inhibitory effect on the maturation ofdendritic cells [36] and suppress the proliferation and toxicityof natural killer cells [37, 38], in addition to their inhibitoryeffect on the proliferation and alloreactivity of T-cells [39–41].Thus, the recruitment of MSCs with their immunosuppressivecharacteristics to the tumor with such large number, as wehave shown, might play role in the tumor strategy to escapethe immune system.
However, the inoculated GL261 cells reduced the expres-sion of Sca-1 and CD166. This change in phenotype could beimportant for tumor initiation, as recent data have shownthat the repression of Sca-1 increases tumor propagation andproliferation at early stages of tumor formation [42–44]. Inter-estingly, repression of Sca-1 results in upregulation of inflam-matory chemokines [44].
Culturing in serum-free conditions has been suggested toincrease the validity of traditional cell cultures [25, 29]. Wefound that the reduced expression of Sca-1 in sphere cultureconditions was associated with increased expression of che-mockine receptor CXCR4 and reduced expression of CD166.Both CXCR4 and CD166 increase with grade of malignancy ingliomas and are suggested as glioma stem cells markers [25,45]. However, removal of serum from culturing mediumreduces CD166 protein levels through miR-9 activation [46].Interestingly, miR-9 is highly expressed in glioma supportinginvasiveness and suppressing proliferation [47]. As well, miR-9is necessary for sphere formation and maintenance ofCD1331 cells in an undifferentiated state [48]. Thus, it is pos-sible that the divergency in CXCR4 and CD166 frequency aredue to cells in different states of proliferation and invasion.Supporting our in vitro data, CXCR4 expression was increasedin vivo too, mimicking in that the sphere growing conditionfor GL261. However, much higher expression level for CXCR4was shown on recruited cells of hematopoietic and nonhema-topoietic origin. While the other candidate, CXCR6, recentlyshown to play important role in recruiting MSCs into prostatecancer [26], it had restricted expression to cells of hematopoi-etic origin only in the GL261 model. It was very recentlyshown that CXCL16 and it receptor CXCR6 were highlyexpressed in fresh human gliomas. Furthermore, expanded gli-oma/glioma stem cells in vitro were highly CXCL16 positive,but CXCR6 disappeared in culture conditions usually used to
1120 Brain Tumor-Derived Mesenchymal Stem Cells in GL261
VC AlphaMed Press 2013 STEM CELLS

expand glioma cells and glioma stem cells [49]. Our data sug-gest that the glioma tumor recruits cells of hematopoietic andnon hematopoietic origin and CXCR4/CXCR6 might be behindthe mechanism of recruitment and host contribution to buildthe tumor niche.
We found that the recruited BT-MSCs stimulated GL261proliferation in vitro, and although nonsignificant—that thesecells could have tumor growth enhancing effects in vivo.Another study has shown BM-derived endothelial progenitorscontribute little to glioma vascularization [50]. However, thetumor promoting effects of MSCs are highly controversial asMSCs may have different effects on tumor growth depend-ing on the source of origin. While adipose tissue derived-MSCs appear to have growth promoting properties [51],bone marrow and umbilical cord blood derived-MSCs haveshown inhibitory effects on tumor growth through reducedangiogenesis [17, 52, 53]. Although the source of infiltratedBT-MSCs is not explored here, migratory brain tissuespecific-MSCs recently identified in normal brain are likelycandidates [7]. Interestingly, while only the GL-MSC1 inocu-lated cells were capable of inducing tumor, infiltrated hostderived BT-MSCs could also induce tumors. Although a lowpassage BT-MSCs population was used, the possibility ofmalignant transformation of in vitro is difficult to exclude aseven a low number of passages can induce genomic changesin MSCs [54]. However, the tumors established from BT-MSCs are more similar in phenotype to the establishedGL261 tumors and less similar to the undifferentiated sarco-matic tumors described after malignant transformation inMSCs [54]. Alternatively, the tumorigenicity could be a resultof fusion of MSCs with tumor cells. Such interactionbetween glioma cells and MSCs was reported recently bySchichor et al. where they describe a functional cross-talkvia gap junctions and structural fusion of cells in vitro [55].In line with our results from cocultur of BT-MSCs withGL261, Mercapide et al. have reported an increased prolifer-ation of both GL261s and U87MGs in coculture with MSCs,which was depending on cell–cell interaction and the fusionof GL261 cells with MSCs in vitro [56]. Such fusion of tumorand stromal cells was suggested several decades ago [57].Clear evidence demonstrates the horizontal transfer of malig-nancy from human GBM to hamster cells in vivo wasrecently reported [58]. CXCR4, highly expressed in ourmodel, was one of the three genes that were implicated insuch spontaneous transmission of malignancy [58].
Further studies are clearly needed to understand the MSC– tumor cell interaction. Targeting BT-MSCs could inhibit
tumor growth; reduce tumorigenicity and amelorate thetumor immunosuppressive microenvironment. This approachcould have important clinical outcome in combination witheffective immunotherapy against glioma.
CONCLUSION
Our data indicate that brain tumors recruit MSCs upon tumorprogression. Furthermore, we have shown that the GL261 cellline consists of a dominant population of cells with mesenchy-mal characteristics, and injecting tumor cell lines without thispopulation abolishes tumorigenicity. Additionally, we haveshown that both mouse and human tumor cells undergo ashift in marker expression under cell culture conditions aswell as in vivo. This data in addition to our discovery for theexpression of CXCR4 and CXCR6 on different recruited cellpopulations and identification of BT-MSCs in human gliomasuggest a strategy to treat brain tumor by targeting cellsrecruited to the tumor.
ACKNOWLEDGMENTS
We thank Wayne Murrell for checking the language; PeterSiesjo and his group, Trygve Leergaard, Sveinung Lillehaug,Grazyna Babinska, Andreas Svensson, Johan Bengzon, YanZhang, and Kavitha Siva for the social and technical support.Histological images were acquired at the Norbrain Slidescan-ning Facility at the Institute of Basic Medical Sciences, Univer-sity of Oslo, a resource funded by the Research Council ofNorway.
AUTHOR CONTRIBUTIONS
J.B.: conception and design, data collection and assembly,data analysis and interpretation and manuscript writing; P.I.:data analysis and interpretation; M.J.: manuscript writing;C.C.: financial support; I.A.L.: financial support and manuscriptwriting; E.O.V.-M.: provision of study materials, data analysisand interpretation, manuscript writing; W.B.: conception anddesign, data analysis and interpretation, financial support,manuscript writing; W.B. and E.O.V.-M. contributed equally tothis article.
DISCLOSURE OF POTENTIAL CONFLICTS OF INTEREST
The authors declare that they have no conflict of interest.
REFERENCES
1 Louis DN, Ohgaki H, Wiestler OD et al.The 2007 WHO classification of tumours ofthe central nervous system. Acta Neuropa-thol 2007;114:97–109.
2 Hadjipanayis CG, Van Meir EG. Tumorinitiating cells in malignant gliomas: biologyand implications for therapy. J Mol Med2009;87:363–374.
3 Lagasse E, Shizuru JA, Uchida N et al.Toward regenerative medicine. Immunity2001;14:425–436.
4 A. J. Friedenstein RKC, K. S. Lalykina.The development of fibroblast colonies inmonolayer cultures of guinea-pic bone mar-row and spleen cells. Cell Tissue Kinet 1970;3:393–403.5 Dominici M, Le Blanc K, Mueller I
et al. Minimal criteria for defining multipo-tent mesenchymal stromal cells. The Inter-national Society for Cellular Therapyposition statement. Cytotherapy 2006;8:315–317.6 Morikawa S, Mabuchi Y, Kubota Y et al.
Prospective identification, isolation, and sys-temic transplantation of multipotent mesen-
chymal stem cells in murine bone marrow. JExp Med 2009;206:2483–2496.7 Taichman RS, Wang Z, Shiozawa Y et al.Prospective identification and skeletal local-ization of cells capable of multilineage differ-entiation in vivo. Stem Cells Dev 2010;19:1557–1570.8 Phinney DG, Prockop DJ. Concise review:mesenchymal stem/multipotent stromal cells:the state of transdifferentiation and modesof tissue repair–current views. Stem Cells2007;25:2896–2902.9 Yoshida T, Matsuda Y, Naito Z et al.CD44 in human glioma correlates with histo-
Behnan, Isakson, Joel et al. 1121
www.StemCells.com VC AlphaMed Press 2013

pathological grade and cell migration. PatholInt 2012;62:463–470.10 Jijiwa M, Demir H, Gupta S et al. (2011)CD44v6 Regulates Growth of Brain TumorStem Cells Partially through the AKT-Mediated Pathway. PLoS ONE 2011;6(9).11 Kijima N, Hosen N, Kagawa N et al.CD166/activated leukocyte cell adhesion mol-ecule is expressed on glioblastoma progeni-tor cells and involved in the regulation oftumor cell invasion. Neuro Oncol 2012;14(10):1254–64.12 Kang SG, Shinojima N, Hossain A et al.Isolation and perivascular localization of mes-enchymal stem cells from mouse brain. Neu-rosurgery 2010;67:711–720.13 Nakamizo A, Marini F, Amano T et al.Human bone marrow-derived mesenchymalstem cells in the treatment of gliomas. Can-cer Res 2005;65:3307–3318.14 Bexell DGS, Tormin A, Darabi A et al.Bone marrow multipotent mesenchymalstroma cells act as pericyte-like migratoryvehicles in experimental gliomas. Mol Ther2009;17:183–190.15 Karnoub AE, Dash AB, Vo AP et al. Mes-enchymal stem cells within tumour stromapromote breast cancer metastasis. Nature2007;449:557–63.16 Houghton J, Stoicov C, Nomura S et al.Gastric cancer originating from bonemarrow-derived cells. Science 2004;306:1568–1571.17 Klopp AH, Gupta A, Spaeth E et al. Con-cise review: Dissecting a discrepancy in theliterature: do mesenchymal stem cells sup-port or suppress tumor growth? Stem Cells2011;29(1):11–9.18 Phillips HS, Kharbanda S, Chen R et al.Molecular subclasses of high-grade gliomapredict prognosis, delineate a pattern of dis-ease progression, and resemble stages inneurogenesis. Cancer Cell 2006;9:157–173.19 Verhaak RG, Hoadley KA, Purdom Eet al. Integrated genomic analysis identifiesclinically relevant subtypes of glioblastomacharacterized by abnormalities in PDGFRA,IDH1, EGFR, and NF1. Cancer Cell 2010;17:98–110.20 Debacq-Chainiaux F, Erusalimsky JD,Campisi J et al. Protocols to detectsenescence-associated beta-galactosidase(SA-betagal) activity, a biomarker of senes-cent cells in culture and in vivo. Nat Protoc2006;4:1798–1806.21 Murrell W, Palmero E, Bianco J et al.Expansion of multipotent stem cells from theadult human brain, Plos One 2013;8(8):e71334.22 Zagzag D, Armirnovin R, Greco MA et al.Vascular apoptosis and evolution in gliomasprecede neovascularization: a novel conceptfor glioma growth and angiogenesis. LabInvest 2000;80:837–49.23 Lee J, S Kotliarova, Y Kotliarov, et al.Tumor stem cells derived from glioblastomascultured in bFGF and EGF more closely mir-ror the phenotype and genotype of primarytumors than do serum-cultured cell lines.Cancer Cell 2006;9:391–403.24 Vik-Mo EO, Sandberg C, Olstorn H et al.Brain tumor stem cells maintain overall phe-notype and tumorigenicity after in vitro cul-
turing in serum-free conditions. Neuro Oncol2010;12:1220–30.25 S Pellegatta, P L Poliani, et al. Neuro-spheres enriched in cancer stem-like cells arehighly effective in eliciting a dendritic cell-mediated immune response against malig-nant gliomas. Cancer Res 2006;66:21. 10247–52.26 Jung Y, Kim J, Shiozawa Y et al. Recruit-ment of mesenchymal stem cells into pros-tate tumours promotes metastasis. NatureCommunications 2013;4:1795.27 Newcomb EW, Zagzag D. The MurineGL261 glioma experimental model to assessnovel brain tumor treatments. In: Van MeirEG (ed). Cancer Drug Discovery and Develop-ment. Humana Press: Atlanta, GA 2009; 227–241.28 Maes W, Van Gool SW. Experimentalimmunotherapy for malignant glioma: lessonsfrom two decades of research in the GL261model. Cancer Immunol Immunother 2011;60(2):153–60.29 Wu A, Oh S, Wiesner SM et al. Persist-ence of CD1331 cells in human and mouseglioma cell lines: detailed characterization ofGL261 glioma cells with cancer stem cell-likeproperties. Stem Cells Dev 2008;17:173–184.30 Lottaz C, D Beier, K Meyer, et al. Tran-scriptional profiles of CD1331 and CD133-glioblastoma-derived cancer stem cell linessuggest different cells of origin. Cancer Res2010;70:2030–2040.31 Soda Y, Marumoto T, Friedmann-Morvinski D et al. Transdifferentiation of glio-blastoma cells into vascular endothelial cells.Proc Natl Acad Sci U S A 2010;108:4274–4280.32 Capes-Davis A, Theodosopoulos G, AtkinI, Drexler HG, Kohara A et al. Check your cul-tures! A list of cross-contaminated or misi-dentified cell lines. International Journal OfCancer 2010;127:1–8.33 Rappa G, Mercapide J, Anzanello F et al.Growth of cancer cell lines under stem cell-like conditions has the potential to unveiltherapeutic targets. Exp Cell Res 2008;314:2110–2122.34 Ho IA, Toh HC, Ng WH et al. Humanbone marrow-derived mesenchymal stemcells suppress human glioma growth throughinhibition of angiogenesis. Stem Cells 2013;31(1):146–55.35 Waterman RS, Henkle SL, BetancourtAM. Mesenchymal Stem Cell 1 (MSC1)-BasedTherapy Attenuates Tumor Growth WhereasMSC2-Treatment Promotes Tumor Growthand Metastasis. Plos One 2012;7(9):e45590.36 Nauta AJ, Fibbe WE. Immunomodulatoryproperties of mesenchymal stromal cells.Blood 2007;110:3499–506.37 Spaggiari GM, Capobianco A, Becchetti Set al. Mesenchymal stem cell-natural killercell interactions: evidence that activated NKcells are capable of killing MSCs, whereasMSCs can inhibit IL-2-induced NK-cell prolif-eration. Blood 2006;107:1484–90.38 Sotiropoulou PA, Perez SA, Gritzapis ADet al. Interactions between human mesen-chymal stem cells and natural killer cells.Stem Cells 2006;24:74–85.39 Bartholomew A, Sturgeon C, Siatskas Met al. Mesenchymal stem cells suppress lym-phocyte proliferation in vitro and prolong
skin graft survival in vivo. Exp Hematol 2002;30:42–8.40 Di Nicola M, Carlo-Stella C, Magni Met al. Human bone marrow stromal cells sup-press Tlymphocyte proliferation induced bycellular or nonspecific mitogenic stimuli.Blood 2002;99:3838–43.41 Potian JA, Aviv H, Ponzio NM et al.Veto-like activity of mesenchymal stem cells:functional discrimination between cellularresponses to alloantigens and recall antigens.J Immunol 2003;171:3426–34.42 Welm BE, Tepera SB, Venezia T et al.Sca-1(pos) cells in the mouse mammarygland represent an enriched progenitor cellpopulation. Dev Biol 2002;245:42–56.43 Upadhyay G, Yin Y, Yuan H et al. Stemcell antigen-1 enhances tumorigenicity bydisruption of growth differentiation factor-10(GDF10)-dependent TGF-beta signaling. ProcNatl Acad Sci U S A 2011;108:7820–7825.44 Batts TD, Machado HL, Zhang Y et al.Stem Cell Antigen-1 (Sca-1) Regulates Mam-mary Tumor Development and Cell Migra-tion. Plos One 2011;6(11):e27841.45 Ehtesham M, Min E, Issar NM et al. Therole of the CXCR4 cell surface chemokinereceptor in glioma biology. J Neurooncol2013;14(10):1007/s11060–013–1108-4.46 Wang J, Gu Z, Ni P et al. NF-kappaBP50/P65 hetero-dimer mediates differentialregulation of CD166/ALCAM expression viainteraction with micoRNA-9 after serumdeprivation, providing evidence for a novelnegative auto-regulatory loop. Nucleic AcidsRes 2011;39:6440–6455.47 Tan X, Wang S, Yang B et al. (2012) TheCREB-miR-9 Negative Feedback MinicircuitryCoordinates the Migration and Proliferationof Glioma Cells. Plos One 2012;7(11):e49570.48 Meijer J, Ogink J, Kreike B et al. TheCXCL16-CXCR6 chemokine axis in glialtumors. J Neuroimmunol 2013;260(1-2):47–54.49 Schraivogel D, Weinmann L, Beier Det al. CAMTA1 is a novel tumour suppressorregulated by miR-9/9* in glioblastoma stemcells. EMBO J 2011;30:4309–4322.50 Kim SM, Kang SG, Park NR et al. Pres-ence of glioma stroma mesenchymal stemcells in a murine orthotopic glioma model.Childs Nerv Sys 2011;27(6):911–22.51 Yu JM, ES Jun, YC Bae and JS Jung. Mes-enchymal stem cells derived from humanadipose tissues favor tumor cell growth invivo. Stem Cells Dev 2008;17:463–473.52 Akimoto K, Kimura K, Nagano M et al.Umbilical Cord Blood-Derived MesenchymalStem Cells Inhibit, But Adipose Tissue-Derived Mesenchymal Stem Cells Promote,Glioblastoma Multiforme Proliferation. StemCells Dev 2013;22(9).53 Machein MR, Renninger S, de Lima-Hahn E et al. Minor contribution of bonemarrow-derived endothelial progenitors tothe vascularization of murine gliomas. BrainPathol 2003;13:582–97.54 Zhou YF, Bosch-Marce M, Okuyama Het al. Spontaneous transformation of culturedmouse bone marrow-derived stromal cells.Cancer Res 2006;66:10849–10854.55 Schichor C, Albrecht V, Korte B, BuchnerA et al. Mesenchymal stem cells and glioma
1122 Brain Tumor-Derived Mesenchymal Stem Cells in GL261
VC AlphaMed Press 2013 STEM CELLS

cells form a structural as well as a functionalsyncytium in vitro. Exp Neurol 2012;234(1):208–19.
56 Mercapide J, Rappa G, Anzanello F et al.Primary gene-engineered neural stem/pro-genitor cells demonstrate tumor-selective
migration and antitumor effects in glioma.Int J Cancer 2010;1:126(5):1206–15.
57 Goldenberg DM. €U ber die Progressionder Malignita €at: Eine Hypothese [on theprogression of malignancy: a hypothesis].Klin Wochenschr 1968;46:898–9.
58 Goldenberg DM, Zagzag D, Heselmeyer-Haddad KM et al. Horizontal transmission andretention of malignancy, as well as functionalhuman genes, after spontaneous fusion ofhuman glioblastoma and hamster host cells invivo. Int J Cancer 2012;131(1):49–58.
See www.StemCells.com for supporting information available online.
Behnan, Isakson, Joel et al. 1123
www.StemCells.com VC AlphaMed Press 2013





























