Embed Size (px)
Citation preview

1 23
Agroforestry SystemsAn International Journal incorporatingAgroforestry Forum ISSN 0167-4366 Agroforest SystDOI 10.1007/s10457-012-9505-9
Regeneration pattern of four threatenedtree species in Sudanian savannas ofBurkina Faso
Amadé Ouédraogo & AdjimaThiombiano

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer
Science+Business Media B.V.. This e-offprint
is for personal use only and shall not be self-
archived in electronic repositories. If you
wish to self-archive your work, please use the
accepted author’s version for posting to your
own website or your institution’s repository.
You may further deposit the accepted author’s
version on a funder’s repository at a funder’s
request, provided it is not made publicly
available until 12 months after publication.

Regeneration pattern of four threatened tree speciesin Sudanian savannas of Burkina Faso
Amade Ouedraogo • Adjima Thiombiano
Received: 25 April 2011 / Accepted: 27 February 2012
� Springer Science+Business Media B.V. 2012
Abstract Research was carried out on four threa-
tened tree species across north and south Sudanian
phytogeographical zones in Burkina Faso. A quanti-
tative inventory of regeneration populations was
completed and sapling dynamics were monitored
through surveys of survival and growth in permanent
plots. Laboratory and nursery experiments were
carried out to test germination and the performance
of transplanted seedlings. Results revealed difficulties
that saplings faced during their development. In
natural stands of Sterculia setigera and Afzelia afri-
cana, saplings were rare or irregularly distributed. A
greater number of individuals of Bombax costatum
were observed, which was the only species with
significant density variation between phytogeograph-
ical zones (p = 0.021). Size class showed irregular
population structures. A high mortality between the
first and second year, followed by a stabilization trend,
was observed. The initial height of saplings was highly
correlated to their survival rate. The rate of new
sapling spontaneous recruitment was better in Bombax
costatum and Boswellia dalzielii stands, but heir
growth was unpredictable in all species. Most saplings
grew from lignotubers and root suckers; however, the
potential for sexual reproduction was good. Sterculia
setigera and Bombax costatum seedlings showed
potential for use in silviculture, as evidenced by the
fact that both showed high post-transplantation rates
of survival and growth.
Keywords Germination � Growth �Population dynamics � Regeneration � Tree species
Introduction
The semi-arid Sahelo-Sudanian zone of Africa, which
is one of the world’s dryland ecosystems, is subject to
major impacts from human activities (Rasmussen et al.
2001; Sop et al. 2010). When humans alter the
landscape, as they do through cropping or ranching,
the system become more vulnerable to climatic
variability, particularly droughts (Nicholson et al.
1998). Weak and irregular precipitation causes stress
to woody plants, whose regeneration then becomes
unpredictable. Indeed, these ecosystems experience
dramatic changes in rainfall, which affect vegetation
growth, habitat and human livelihoods (White et al.
2002). For example, annual rainfall has declined since
the 1970s in the Sahel (Agnew and Chappell 1999) and
subsequent droughts have caused high mortality in
many woody species in northern Burkina Faso
A. Ouedraogo (&) � A. Thiombiano
Laboratoire de Biologie et Ecologie Vegetales,
UFR Sciences de la Vie et de la Terre, Universite de
Ouagadougou, 09 BP 848, Ouagadougou 09,
Burkina Faso
e-mail: [email protected];
A. Thiombiano
e-mail: [email protected]
123
Agroforest Syst
DOI 10.1007/s10457-012-9505-9
Author's personal copy

(Ganaba et al. 1998). Because many other species are
associated with these trees, the shift in the structure of
their woodlands has large-scale community conse-
quences (Mueller et al. 2005). The drought has
sometimes affected other areas of the country, which
usually have sufficient rainfall. Some tree species
exhibit intrinsic adaptation strategies of growth and
survival to face dryness (Weber et al. 2008). The
impact of human activities, through extensive agri-
culture and livestock farming, also negatively affect
woody plant dynamics (Ouedraogo et al. 2006b).
Saplings are the most vulnerable stage in the woody
plant population, as these individuals are less sturdy,
particularly in the recruitment stage (Ouedraogo et al.
2006c). Seedlings and young shoots are the first to
wither, and they are also more exposed to livestock
damage, water stress and fire. For many woody species
in savannas, recruitment during the rainy season is
abundant, but very few individuals live through the dry
season (Gijsbers et al. 1994; Setterfield 2002). In the
long term, disturbance factors lead to the selection of
well-adapted species, which in consequence leads to
the reduction of diversity due to the regression of
sensitive species. For example, studies of woody plant
recruitment in the Cerrado savannas of Brazil have
revealed that frequent fires have caused a shift in
species composition, favouring those species that can
propagate vegetatively (Hoffmann 1998). A similar
situation exists in Africa savannas, where vegetative
propagation of woody species is favoured (Zida et al.
2007; Bognounou et al. 2010). Assessing woody
species diversity along a climatic gradient in eastern
Burkina Faso, Ouedraogo (2006) found that the
sapling layer is less rich than the adult one, introducing
a dynamic trend of diversity loss. In addition, those
woody saplings that tolerate semi-arid conditions are
trapped in a continual cycle of growth, then cessation,
and then re-growth (Ouedraogo et al. 2006b). The
advantage of this adaptation is ‘‘self-conservation’’,
rather than the rejuvenation of ageing populations.
This study aims at analysing the structure of the
regeneration population in natural stands and docu-
menting the juvenile individuals’ behaviour as well as
the potential for reproduction by seeds of four tree
species. Such research will contribute to understand-
ing the sapling response to environmental conditions,
which is helpful in supporting future adequate
approaches for conservation of these threatened
woody species. This investigation is especially
critical, as according to Zida et al. (2008), monitoring
of sapling field performance has received very little
attention in Africa, especially in west Africa.
Materials and methods
Study species
Four tree species were chosen, which are Afzelia
africana Sm. (Fabaceae), Bombax costatum Pellegr. &
Vuillet (Malvaceae), Boswellia dalzielii Hutch. (Burs-
eraceae) and Sterculia setigera Delile (Malvaceae).
These species have a high economic value in the west
Africa Sahelo-Sudanian zone, and especially in Burk-
ina Faso (Guinko and Pasgo 1992; Mertz et al. 2001;
Lykke et al. 2004), but they are also threatened (Belem
et al. 2008; Mbayngone and Thiombiano 2011). They
have a wide distribution range across the Sudanian
zone, with ageing population (Sinsin et al. 2004;
Ouedraogo 2006).
Afzelia africana is among the largest trees in the
savannas whose northernmost limit in Burkina Faso is
the south Sahel, at about 13�N (Terrible 1984), where
it is sparsely distributed (Ouedraogo 2006). The
species is mainly used for timber, firewood and forage
(leaves). In the Sudanian zone, its natural stands
exhibit ageing populations (dhp = 35–50 cm) with
many individuals damaged by livestock and pruning.
A similar situation was observed in Benin—to the
southeast of Burkina Faso—where uncontrolled util-
isation leads to the degradation and reduction of its
habitat and populations (Sinsin et al. 2004). Direct
sowing techniques were unsuccessfully tested in the
field as an alternative to relying upon weak natural
regeneration (Bationo et al. 2001a). In natural pro-
tected areas, it is not unusual to observe cohorts of
seedlings underneath parent plants during the rainy
season, but their survival rate is almost nil.
Bombax costatum is a common tree in the Sudanian
zone, where its natural stands are sometimes large
with 6–10 trees per 1,000 m2. Populations are char-
acterised by the predominance of individuals with
5–15 cm-diameter stems in the south Sudanian zone,
while 25–40 cm-diameter trees are most frequent in
the north Sudanian one (Ouedraogo 2006). This
species is mainly used for food and timber. It is
threatened by its apparently poor regeneration (Ou-
edraogo et al. 2006b), and also by the intensive
Agroforest Syst
123
Author's personal copy

harvesting of its flowers for domestic and commercial
use as a vegetable (Belem et al. 2008).
Boswellia dalzielii is a gregarious tree that is widely
distributed and prefers to grow on rocky hills. The
natural groves are quite open, with 6–9 individuals per
1,000 m2, dominated by trees with 20–40 cm-diame-
ter stems (Ouedraogo et al. 2006a). The bark of
Boswellia dalzielii is used for medicinal purposes. It is
declining despite ample fruit production. The inten-
sive de-barking causes high mortality in natural
populations.
Sterculia setigera is a sparsely distributed tree,
rarely forming significant populations. The species
occurs widely, but without an apparent habitat pref-
erence. The few natural populations are rather open
with 4–5 trees per 1,000 m2. Most trees have stems in
the 25–40 cm-diameter class, reflecting old popula-
tions (Ouedraogo 2006). The species is pruned for
forage and also cut for timber. Its natural stands are
characterised by a lack of regeneration, despite ample
fruit production.
Study area
Field work was carried out in eastern Burkina Faso
across two phytogeographical zones, the north Suda-
nian and south Sudanian (Fontes and Guinko 1995).
This area is situated in the Sudanian regional centre of
endemism (White 1983). The rainy season lasts from
May to October, and temperature variation is limited
(Fig. 1). The mean annual rainfall (1974–2004) varies
from 805.30 ± 187 mm to 862.46 ± 158 mm.
According to the FAO classification (WRB 2006),
common soil types in the study area include luvisols,
lixisols, leptosols, cambisols, vertisols and gleysols,
which are common across most of eastern Burkina
Faso.
The vegetation varies from steppes in the north, to
tree and bush savannas in the south. The herbaceous
layer is dominated by grasses, while the woody
vegetation is dominated by shrubs. The region is a
livestock-farming and agricultural area, with exten-
sive land use. Livestock farming is on the increase, and
for over 85 % of farmers natural pasture is the main
source of fodder (Ministere des ressources animales
2004). The extent of cultivated land has also increased,
as cotton cultivation and internal immigration has
expanded.
Field surveys
Field work consisted of inventorying and monitoring
regeneration individuals of each species in natural
stands across the phytogeographical zones. Regener-
ation individuals refer to saplings, with main stems
less than 5-cm diameter at 1.30-m height. Inventories
were carried out in 18–33, 5 9 5 m2 plots depending
on the species (Table 1), randomly chosen within
stands of trees where regeneration individuals were
counted and their height was measured. The monitor-
ing was carried out within five plots, each of 25 m2, at
five different sites per species and for both phytogeo-
graphical zones. The initial stock of saplings in the
plot at the beginning of the survey were marked with
paint and observed for 24 months.
Monitoring was carried out at four periods: begin-
ning of the rainy season (June); early end of rainy
season (end of September); beginning (November)
and end (May) of dry season. The following param-
eters were recorded four times a year: number of
Fig. 1 Ombrothermic
diagrams of the main
localities in each
phytogeographical zone.
Data are from the period
1974 to 2004 (sourceNational Office of
Meteorology, Burkina Faso
2006)
Agroforest Syst
123
Author's personal copy

surviving individuals; number of new recruitments
(these were subsequently marked with a different
colour paint); height and sturdiness of saplings. The
origin of saplings (seedlings, root suckers, lignotubers
or coppice) was distinguished by gently digging
around the base of the trunk. Sapling density was
calculated and the height class distribution was
computed for each species.
Laboratory and nursery trials
Experiments consisted of germination tests which
were carried out for each species using seeds from
different provenances (10 populations 9 3 trees per
population = 30 trees) in each phytogeographical
zone. The experimental design was randomised blocks
with ten repetitions of ten seeds (from the composite
samples of 30 trees) per species. Seeds were sown
without any pre-treatment, in pots filled with a
substrate consisting of two-thirds sand and one-third
humus. Each pot received 250 ml of water daily.
Conditions in the nursery were similar to those of the
surrounding environment (temperature = 25–38 �C;
humidity = 79.5 % ± 11.24). The number of seeds
that germinated or died was recorded daily. Ten
seedlings were marked at germination, and both their
height and number of leaves were recorded every
2 days for a period of 30 days to assess their initial
growth. Afterwards, they were transplanted into larger
pots (70-cm tall 9 25-cm diameter) containing topsoil
to assess their response. Survival rate, diameter and
height growth were measured, and vitality was
visually assessed.
Data analysis
The means of regeneration population parameters
(density, recruitment, survival and growth rates) were
calculated for species in each plot and compared
according to the phytogeographical zones and species.
The growth performance of saplings in natural stands
was compared according to the regeneration mode.
For the germination, both latent period of germi-
nation (LPG, number of days before the first germi-
nation) and speed of germination (SG, number of days
for 50 % germination) were calculated. The increase in
mean height, diameter and number of leaves of
seedlings was calculated to assess the post-germina-
tion and post-transplantation growth performance. For
each parameter, test of equal variance was performed
and multiple comparison test of Tukey was applied to
the series of data through one-way analysis of variance
(ANOVA). The Pearson correlation coefficient was
calculated to measure the degree of linear relationship
between variables for some growth parameters. Sta-
tistical analyses were performed with Minitab 14.
Results
Regeneration populations’ structure
Saplings were rare or absent in Sterculia setigera and
Afzelia africana natural stands. On the other hand,
populations of Bombax costatum and Boswellia dalzi-
elii often contained saplings. Across all inventories,
sapling density showed considerable heterogeneity
according to stands (apart from B. costatum in the north
Table 1 Sapling density per plot and mean number of stems per individual (mean ± SD) in natural stands
Species Number of plots Sapling density (number of individuals/25 m2) Number of stems per individual
NS SS NS SS p-value NS SS p-value
Afzelia africana 16 11 0.93az ± 0.29 1.55ay ± 0.49 0.12 1.15ax ± 0.10 1.09ax ± 0.09 0.66
Bombax costatum 19 14 2.93bx ± 0.31 6.64ax ± 2.10 0.021 1.34ax ± 0.08 1.14bx ± 0.04 0.010
Boswellia dalzielii 12 8 1.92ay ± 0.43 1.88ay ± 0.69 0.96 1.18ax ± 0.08 1.25ax ± 0.16 0.69
Sterculia setigera 12 6 0.75az ± 0.25 0.83az ± 0.31 0.83 1.21ax ± 0.26 1.06ax ± 0.17 0.62
p-value 0.023 0.011 0.112 0.139
Mean values of parameters were compared (ANOVA) for each species according to the phytogeographical zone (lines) and between
species per phytogeographical zones (columns). Values with the same letter are not significantly different at p = 0.05 (Tukey’s test).
First letter is for comparison between the columns and second letter for comparison between the lines. SD standard deviation, NSnorth Sudanian phytogeographical zone, SS south Sudanian
Agroforest Syst
123
Author's personal copy

Sudanian zone). Significant difference in sapling
density between phytogeographical zones was found
only for B. costatum (p = 0.021). This species showed
the highest densities in both north and south Sudanian
zones, whereas S. setigera showed the lowest densities,
although it was not different from the one of A. africana
in the north Sudanian zone (Table 1). The tendency of
saplings to branch at the base was limited. Individuals
rarely had more than one stem, and the mean number of
stems did not vary significantly in species according to
phytogeographical zones, except for B. costatum
(p = 0.010). However, the number was constantly a
little higher in the north Sudanian zone (Table 1).
The height size distribution demonstrated irregular
population structures, characterised by large gaps
between size classes and the notably small number of
individuals in each class (Fig. 2). In general, the
recruitment class (0–0.5 m) had the highest number of
individuals. These population structures reveal prob-
lems of recruitment, survival and growth of saplings.
Only B. costatum showed a relatively good
regeneration population structure with the five height
classes well represented in both north and south
Sudanian zones (Fig. 2). B. dalzielii had a better
structure in the south Sudanian zone.
Regeneration population dynamics
The mean densities of the initial stocks of saplings in
the permanent plots are shown in Table 2. They were
significantly different between phytogeographical
zones for all species, with higher density for B.
costatum in the south Sudanian zone and lower one for
S. setigera in both zones. The saplings of S. setigera in
both zones died or their number decreased consider-
ably from the first to the second year. The maximum
survival rate did not exceed 45.3 %, which was the one
of B. dalzielii’s natural stands in the south Sudanian
zone (Table 2). Across the second and the third years,
survival rates did not vary considerably. There was a
high correlation (r = 0.85, N = 98) between sapling
initial height and their survival rate.
Fig. 2 Height size
distribution of saplings. The
proportion of individuals
from global inventory is
represented per size class for
each species in the north
Sudanian (NS) and the south
Sudanian (SS) zones
Agroforest Syst
123
Author's personal copy

Recruitment in the permanent plots is illustrated by
the cumulative mean number of new saplings in the
stands over 3 years (Fig. 3). A higher recruitment
number was observed for B. costatum in both north
and south Sudanian zones (11.8 ± 3.4 & 16.4 ± 2.7).
A. africana and S. setigera showed the lowest
recruitment rates (no recruitment for S. setigera in
north Sudanian zone). Sapling growth in the perma-
nent plots was unpredictable, as illustrated by the
curves in Fig. 4 (not estimated for S. setigera because
of the lack or weakness of saplings). Increases in
height were irregular across the 3 years of monitoring.
During the rainy season (June–September), height
increased, but it decreased during the dry season
(November–May) due to drought, grazing and
bushfire.
Natural regeneration modes and saplings’
responses to environmental disturbances
The modes of natural regeneration were seedlings,
lignotubers and root suckers. The proportion of each of
these modes of recruitment varied significantly
according to species, but rarely according to the
phytogeographical zone (Table 3). Saplings of S.
Table 2 Survival rates of initial stock of saplings in five permanent plots per species in natural stands
Species Year 1—initial stock Surviving saplings (%)
Number of individuals per plot Height (cm) Year 2 Year 3
NS SS p-
value
NS SS p-
value
NS SS NS SS
Afzeliaafricana
10.85ax ± 4.10 6.21by ± 2.51 0.04 52.60ax ± 8.50 65.60ax ± 9.00 0.27 36.0 38.2 34.0 38.2
Bombaxcostatum
10.87bx ± 2.40 16.74ax ± 1.19 0.02 32.36ay ± 3.7 48.24ay ± 7.10 0.19 20.4 28.1 18.5 28.1
Boswelliadalzielii
9.66ax ± 1.90 6.41by ± 1.50 0.04 43.18bx ± 5.86 60.54ax ± 12.00 0.04 39.6 45.3 36.7 45.3
Sterculiasetigera
2.12by ± 1.20 4.51az ± 0.71 0.02 9.61bz ± 1.40 16.09az ± 1.80 0.01 0.00 20.6 0.00 20.6
p-value 0.022 0.034 0.021 0.047
Mean number and height of individuals were compared (ANOVA) according to the phytogeographical zones (lines) and species
(columns). Values with the same letter are not significantly different at p = 0.05 (Tukey’s test). First letter is for comparison between
the columns and second letter for comparison between the lines. NS north Sudanian phytogeographical zone, SS south Sudanian
Fig. 3 Total mean number of new recruited saplings over
3 years in the two phytogeographical zones in permanent plots
Fig. 4 Fluctuation of sapling growth over 3 years. The month
of June corresponds to the beginning of rainy season, September
to its early end, November to the beginning of dry season and
April to its end
Agroforest Syst
123
Author's personal copy

setigera and A. africana grew mainly from lignotubers
(50.8–61.6 and 60.8–64.7 % respectively), while those
of B. costatum and B. dalzielii originated almost
entirely from sprouting of root suckers (94.3–95.1 and
94.7–97.9 % respectively). The highest proportion of
seedlings was recorded in S. setigera and A. africana
stands, with 38.4–49.1 % and 25.2–26.4 %, respec-
tively. Water stress due to drought, annual bushfires
and repeated grazing affected the survival and growth
of saplings. Saplings of all species except B. dalzielii
were systematically grazed by cattle and goats. The
combined effects of these factors caused very high
mortality among seedlings. Individuals growing from
lignotubers and root suckers had a higher survival rate,
but their growth was periodically reduced. Indeed,
saplings often lost all or part of their stems and
survived in the form of stumps. When environmental
conditions became favourable again (in the rainy
season), new stems grew from lignotubers or roots.
Most individuals in the regeneration stage followed
this cycle of growth re-start, having survived
disturbance.
Trees’ potential for sexual reproduction
The germination rates of freshly harvested seeds were
30 % or more for all species. Those with the highest
germination rates were A. africana (68 and 74 %) and
S. setigera (60 and 62 %) for south and north Sudanian
provenances, respectively. On the other side, average
germination in B. costatum (55 and 36 %) and B.
dalzielii (35 and 30 %) was lower for the north and
south Sudanian provenances, respectively. Differences
in germination rates between provenances were only
significant for A. africana (p = 0.038) and B. costatum
(p = 0.006). All provenances taken together, germi-
nation was rapid for B. dalzielii (6.8 ± 1.2 and
8.5 ± 1.3 days, respectively) for the latent period of
germination (LPG) and the speed of germination (SG),
but relatively slow for A. africana (LPG = 14.8 ± 2.7
and SG = 17.1 ± 3.1), B. costatum (LPG = 17.8 ±
3.9 and SG = 23.2 ± 3.9) and S. setigera (LPG =
14.2 ± 3.9 and SG = 17.0 ± 2.7).
The seedlings after 1 month revealed highly vari-
able growth rates between species. The size of the
seedlings was significantly different according to
species after 30 days (Table 4). Seedlings of A.
africana and S. setigera included the tallest individ-
uals, while those of B. dalzielii being the smallest. TheTa
ble
3P
erce
nta
ge
of
sap
lin
gs
per
spec
ies
inth
etw
op
hy
tog
eog
rap
hic
alzo
nes
(NS
no
rth
Su
dan
ian
,S
Sso
uth
Su
dan
ian
)ac
cord
ing
toth
ere
cru
itm
ent
mo
de
A.
afr
ica
na
B.
cost
atu
mB
.d
alz
ieli
iS
.se
tig
era
NS
SS
pN
SS
Sp
NS
SS
pN
SS
Sp
See
dli
ng
26
.4ay
±7
.22
5.3
ay
±9
.10
.14
0.0
bz
±0
.01
.2az
±0
.70
.04
0.0
3.5
y±
1.3
0.0
13
8.4
ay
±1
0.2
49
.1ax
±1
1.7
0.0
6
Lig
no
tub
er6
0.8
ax
±1
2.6
64
.7ax
±1
4.1
0.0
94
.9ay
±1
.64
.5ay
±1
.70
.49
2.1
ay
±1
.11
.9ay
±0
.60
.07
61
.6ax
±1
5.8
50
.9ax
±1
3.4
0.0
6
Ro
ot
suck
er1
2.7
az
±4
.71
0.0
az
±5
.20
.33
95
.1ax
±1
9.6
94
.3ax
±2
1.3
0.4
29
7.9
ax
±1
8.5
94
.7ax
±1
6.6
0.6
30
.00
.0–
p0
.01
0.0
10
.00
0.0
00
.01
0.0
10
.00
.8
Mea
nv
alu
esw
ere
com
par
ed(A
NO
VA
)fo
rea
chsp
ecie
sac
cord
ing
toth
ere
cru
itm
ent
mo
de
(co
lum
ns)
and
the
ph
yto
geo
gra
ph
ical
zon
es(l
ines
).V
alu
esw
ith
the
sam
ele
tter
are
no
t
sig
nifi
can
tly
dif
fere
nt
atp
=0
.05
(Tu
key
’ste
st).
Fir
stle
tter
isfo
rco
mp
aris
on
bet
wee
nth
eco
lum
ns
and
seco
nd
lett
erfo
rco
mp
aris
on
bet
wee
nth
eli
nes
Agroforest Syst
123
Author's personal copy
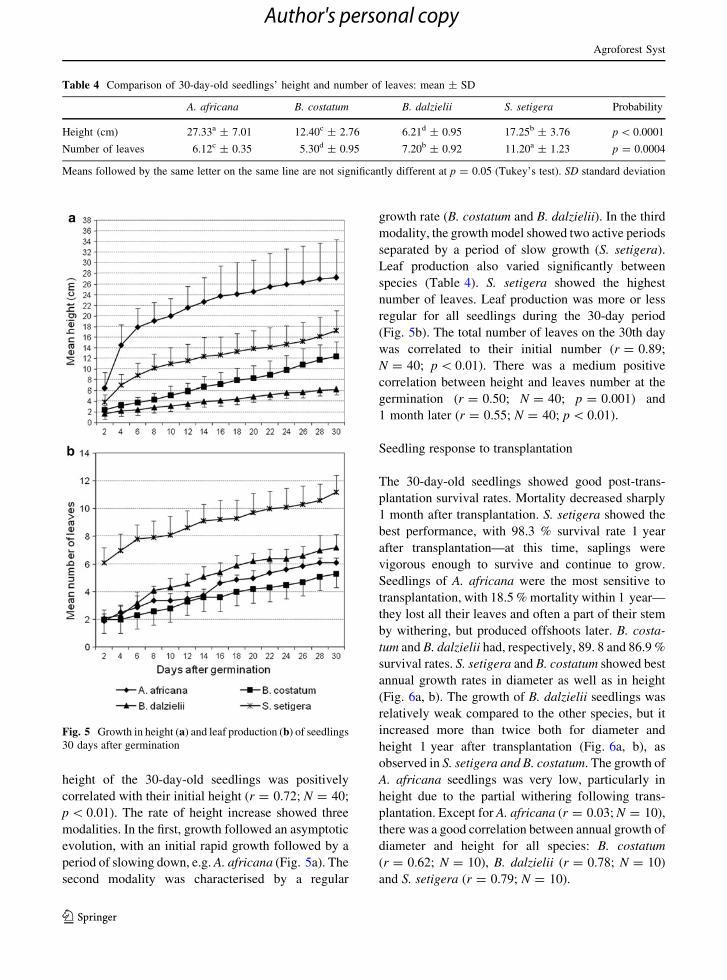
height of the 30-day-old seedlings was positively
correlated with their initial height (r = 0.72; N = 40;
p \ 0.01). The rate of height increase showed three
modalities. In the first, growth followed an asymptotic
evolution, with an initial rapid growth followed by a
period of slowing down, e.g. A. africana (Fig. 5a). The
second modality was characterised by a regular
growth rate (B. costatum and B. dalzielii). In the third
modality, the growth model showed two active periods
separated by a period of slow growth (S. setigera).
Leaf production also varied significantly between
species (Table 4). S. setigera showed the highest
number of leaves. Leaf production was more or less
regular for all seedlings during the 30-day period
(Fig. 5b). The total number of leaves on the 30th day
was correlated to their initial number (r = 0.89;
N = 40; p \ 0.01). There was a medium positive
correlation between height and leaves number at the
germination (r = 0.50; N = 40; p = 0.001) and
1 month later (r = 0.55; N = 40; p \ 0.01).
Seedling response to transplantation
The 30-day-old seedlings showed good post-trans-
plantation survival rates. Mortality decreased sharply
1 month after transplantation. S. setigera showed the
best performance, with 98.3 % survival rate 1 year
after transplantation—at this time, saplings were
vigorous enough to survive and continue to grow.
Seedlings of A. africana were the most sensitive to
transplantation, with 18.5 % mortality within 1 year—
they lost all their leaves and often a part of their stem
by withering, but produced offshoots later. B. costa-
tum and B. dalzielii had, respectively, 89. 8 and 86.9 %
survival rates. S. setigera and B. costatum showed best
annual growth rates in diameter as well as in height
(Fig. 6a, b). The growth of B. dalzielii seedlings was
relatively weak compared to the other species, but it
increased more than twice both for diameter and
height 1 year after transplantation (Fig. 6a, b), as
observed in S. setigera and B. costatum. The growth of
A. africana seedlings was very low, particularly in
height due to the partial withering following trans-
plantation. Except for A. africana (r = 0.03; N = 10),
there was a good correlation between annual growth of
diameter and height for all species: B. costatum
(r = 0.62; N = 10), B. dalzielii (r = 0.78; N = 10)
and S. setigera (r = 0.79; N = 10).
Table 4 Comparison of 30-day-old seedlings’ height and number of leaves: mean ± SD
A. africana B. costatum B. dalzielii S. setigera Probability
Height (cm) 27.33a ± 7.01 12.40c ± 2.76 6.21d ± 0.95 17.25b ± 3.76 p \ 0.0001
Number of leaves 6.12c ± 0.35 5.30d ± 0.95 7.20b ± 0.92 11.20a ± 1.23 p = 0.0004
Means followed by the same letter on the same line are not significantly different at p = 0.05 (Tukey’s test). SD standard deviation
Fig. 5 Growth in height (a) and leaf production (b) of seedlings
30 days after germination
Agroforest Syst
123
Author's personal copy

Morphological behaviour of seedlings
Our observations in the nursery for over 1 year
suggested that seedlings reduced their aerial organs
(all leaves and parts of branches) and specialised their
roots systems to survive water stress during the dry
season. Seedlings of B. costatum, B. dalzielii and S.
setigera hypertrophied their root stock (Fig. 7b–d).
This specialisation allowed their seedlings to store
water and other nutrients in these swollen roots to
Fig. 6 Post-transplantation
growth in height (a) and
diameter (b) of seedlings
A. africana B. costatum B. dalzielii S. setigera
dcba
Fig. 7 Root systems of 6-month-old seedlings: expanded root system (a Afzelia africana) and hypertrophied root systems (b Bombaxcostatum, c Boswellia dalzielii and d Sterculia setigera)
Agroforest Syst
123
Author's personal copy

survive the drought as geophytes. The root system in
A. africana seedlings was not hypertrophied, but much
expanded with long branching roots (Fig. 7a).
Discussion
Regeneration population structure and dynamics
The density of saplings and their size distribution
illustrate the regeneration capacity of woody species
populations. The perceptible reduction of natural adult
populations is certainly the result of a previous long-
term disturbance of regeneration populations. The gap
between the regeneration population and the adult one
will enlarge as the disturbance of reproductive
processes continues. The absent or very small sapling
population in S. setigera and A. africana natural stands
demonstrates this situation. Many authors report the
degraded status of the populations of both species in
the Sahelo-Sudanian zone of West Africa (Lykke
1998; Sinsin et al. 2004; Ouedraogo et al. 2005), as
well as their unpredictable regeneration (Traore 1998;
Bationo et al. 2001a). B. costatum and B. dalzielii had
substantial sapling populations; the variation in den-
sity between both species stands seems to be due to the
intensity of local environmental disturbances. In
similar climatic conditions, Zida et al. (2007) proved
that grazing and fire affect the density of saplings,
largely by reducing the density of single-stemmed
individuals. According to their analysis, the multi-
stemmed state is advantageous in disturbed environ-
ments in terms of chance to conserve aerial organs
alive across the time. They also noticed an increasing
tendency towards multi-stemmed individuals on
grazed sites. Even though the variation is not signif-
icant, the increased number of multi-stemmed saplings
in the north Sudanian sites for all four species could
reflect the environmental pressures in this zone. In
terms of adaptation, B. costatum and B. dalzielii
showed a greater tendency to become multi-stemmed.
Regeneration is affected by the recruitment mech-
anism and the growth conditions of newly recruited
individuals. The small number of saplings and their
irregular height class distribution suggest both recruit-
ment and growth difficulties. The density in the
recruitment class (0–0.5 cm) is too low to ensure
successful regeneration. Indeed, this class is most
vulnerable (Ouedraogo et al. 2006c, 2009) and
requires a very high quantity of individuals to offset
the losses due to disturbances. The low survival rate
between the first and the second year in the initial
stock is mainly attributable to mortality in the
recruitment class. Natural selection due to distur-
bances would favour the tallest saplings which can
hopefully get away from livestock trampling and
bushfire intensity. A positive correlation between
height size and survival rate of saplings is also
reported by Ouedraogo et al. (2006b)and Bellefon-
taine et al. (2000). The better survival rate of A.
africana, compared to B. costatum, could be due to the
origin of its saplings, which were for the most part
‘‘old’’ saplings growing from lignotubers.
Among the four species, B. dalzielii and S. setigera
have a high fruit production in the field, which could
favour an abundant recruitment. However, environ-
mental conditions are limiting factors. For the former
species, there is a relatively high rate of seed
infertility, which accounts for its poor germination
(Ouedraogo et al. 2006a). In addition, the rocky
habitats where the species grows do not favour
recruitment by seeds. The scarcity of S. setigera
seedlings is common across its range, and was also
reported to be the case in Senegal (Traore 1998). In
addition to the difficult terrain, probable predation of
the large seeds, as was reported for A. africana
(Bationo et al. 2001a), may be a problem. For S.
setigera and B. costatum, sexual recruitment potential
is highly reduced by pruning, which reduces fruit
production. Based on the unpredictable recruitment
patterns, the chance for satisfactory regeneration is
weak. Indeed, the upper height classes, resulting from
the survival and growth of the first recruitment, will be
of limited size, as indicated by the gaps in the
population structure. The capacity for rapid growth at
the beginning of the rainy season is an advantage in
semi-arid environments, but the recurrent and some-
times fluctuating drought in the dry season, together
with human and livestock pressures, quash the efforts
of saplings to grow, restricting them to a cycle of
perpetually restarting growth. Considering the pres-
ence of all height classes, and the reverse ‘‘J’’ shape of
their size distribution, the species with the greatest
potential for good regeneration are B. dalzielii and B.
costatum. For these two species, the main threat is
direct and indirect human pressure, as Glele Kakaı
et al. (2009) have observed for Pterocarpus erinaceus,
a common savanna tree that is under threat. A. africana
Agroforest Syst
123
Author's personal copy

and S. setigera appear to be more sensitive to climatic
variability.
Natural regeneration modes and saplings’
responses to harsh environmental conditions
A variety of recruitment modes offer alternative ways
for plant regeneration in difficult environmental
conditions. Facing the unpredictable growth of their
saplings, tree species develop recruitment strategies
that allow them to secure the survival of individuals.
This could explain the highly significant predomi-
nance of recruitment by root suckers for B. costatum
and B. dalzielii, where vegetative propagation is more
common than sexual reproduction. These observations
are in line with those of Bognounou et al. (2010) who
reported a dominance of asexual propagation in five
Combretaceae species in Sahelo-Sudanian zone of
Burkina Faso. The advantages of vegetative propaga-
tion are low-cost renewal of regeneration populations
and rapid growth of saplings, as previously docu-
mented by Bellefontaine et al. (2000). For A. africana
and S. setigera, most saplings originate from lignotu-
bers, which represent in most cases the living ‘‘ves-
tige’’ of several previous years of growth. For an
individual, losing all its aerial parts and conserving
itself as a geophyte during the dry season then growing
early in the rainy season is an effective survival
strategy. Bationo et al. (2001b) observed a similar
behaviour for Detarium microcarpum in Sudanian
savannas of Burkina Faso. Although the annual
renewal of saplings from lignotubers is similar to root
suckers sprouting because of the seasonal growth of
stems, it has a seedling origin. Bognounou et al. (2010)
have distinguished ‘‘true seedlings’’ from ‘‘seedlings
sprouts’’.
New seedlings are more vulnerable than root
suckers and lignotuber shoots. For all species, direct
recruitment by seeds is low, revealing the unsuitability
of this mode of regeneration, given the actual climatic
and environmental conditions. The dominance of
vegetative propagation in natural populations is often
the result of an inefficient recruitment by seeds.
Indeed, vegetative propagation is documented as an
adaptative strategy or response to disturbance and
climatic variation in species with normal sexual
reproduction (Hoffmann 1998; Higgins et al. 1999;
Dickinson et al. 2000). This mode of reproduction
cannot ensure the long-term conservation of natural
populations due to limited dissemination capacity of
individuals (Puig et al. 1998; Setterfield 2002; Tremb-
lay et al. 2002). A wide genetic base in agroforestry
trees is essential to prevent inbreeding depression and
allow adaptation to changing environmental condi-
tions and to altering markets for tree products (Daw-
son et al. 2009).
Species potential for sexual reproduction
and adaptation strategies
Excluding eventual dormancy or parasite attack, the
germination rates of freshly collected seeds reveal the
initial fertility rate. The high germination rates of A.
africana and S. setigera seeds are means to mitigate
the environmental instability in natural habitats.
Indeed, the recruitment of seedlings will be reduced
for species with a low germination rate, under threat of
predation and bushfire (Bationo et al. 2001a; Setter-
field 2002; Danthu et al. 2003; Thiombiano et al.
2003). If the moderate germination rate of B. dalzielii
reflects its seed infertility (Ouedraogo et al. 2006a), as
also observed in Anogeissus leiocarpa (Kambou
1997), then the rate for B. costatum is probably due
to dormancy, since Belem (2009) observed a germi-
nation rate of 80 % following pre-treatment that
consisted of soaking seeds in water for 48 h. Dor-
mancy could also explain the difference of the
germination rates between the two provenances of
both A. africana and B. costatum. Indeed, Ouedraogo
(2006) observed variable seed dormancy rates for
these species according to their north and south
Sudanian origins. The rapid germination in B. dalzielii
is an undeniable advantage for seedlings facing the
constraint of a short rainy season. Compared with the
other three species, the germination in B. dalzielii is
faster giving more time to seedlings to gain vigour and
resist poor weather conditions in the dry season.
Endogenous control of natural seed stock is vital for
plants facing irregular environmental factors (Al-
Helal 1996; Schutz and Milberg 1997), latent periods
that allow avoiding harmful effects in the event of
sporadic rainfall. The orthodox seeds of certain
species can conserve their capacity to germinate for
several years (Danthu et al. 2003). This is a useful
adaptation to survive an extended drought following
fruit production.
After germination, the early growth capacity of
seedlings is decisive for their future in disturbed areas.
Agroforest Syst
123
Author's personal copy

Indeed, observations have shown that shorter saplings
are most vulnerable. Rapid growth permits seedlings
to gain in size during the short rainy season and have a
greater chance of survival. A. africana showed better
height and S. setigera higher leaf number than the
other species 30 days after germination. Thus, the
relatively low germination rate of A. africana and S.
setigera is compensated for by the rapid early growth
of their seedlings. However, for A. africana, seedling
growth slows down early and saplings show limited
development 1 year after transplantation. This obser-
vation agrees with those of Bationo et al. (2001a), who
noticed in the same species a long period of rest after
4–5 weeks of continuous growth. The seedlings of S.
setigera and B. costatum maintained a good growth
rhythm and showed better post-transplantation per-
formances, suggesting good potential for silviculture.
The vigorous growth of B. costatum indicates that the
post-germination height is not bound to have an
influence on subsequent post-transplantation growth.
The development of the root system in seedlings is an
essential factor that determines the difference in their
survival capacity (Dauro et al. 1997). Hypertrophy of
the root system remains an effective survival strategy
because it allows the storage of moisture and nutrients,
which keep the individual alive. Seedlings can thus
lose all their leaves and survive as geophytes until the
next favourable season, as observed in B. costatum and
B. dalzielii. While nutrient storage in hypertrophied
root confers certain survival advantages, root expan-
sion in A. africana is only effective when water is
available in the soil, which could explain its poor
adaptation to drought.
Conclusion
This study provides evidence that the efficiency of
natural regeneration in the four woody species is
uncertain in the actual environmental conditions of
Sudanian savannas. The regeneration is under threat
and its extent varies more according to the species than
to the phytogeographical zones. For Sterculia setigera
and Afzelia africana, the quasi-absence or very low
density of saplings suggests that the conservation of
their natural population is in jeopardy. Although the
sapling densities are relatively high in Bombax cost-
atum and Boswellia dalzielii, their regeneration is also
threatened considering the high mortality during the
dry season. Lack of saplings and high mortality rates
are evidence that the natural populations face a
regression trend in both north Sudanian and south
Sudanian zones. Natural reproductive capacities of
species seem to be surpassed by the worsened harsh
environmental conditions which are characterised by
drought, grazing and bushfire. Thus, the conservation
of these species in parklands would be only possible
through plantations and protection of saplings. The
results have shown some potential for such activities
considering the observed germination capacity, post-
transplantation survival and growth rates. However,
for the purpose of conservation management, the
results from natural regeneration monitoring suggest
that fencing new plantations until saplings reach
sufficient height to be less vulnerable to current
disturbances would be necessary. In addition, further
research should focus on understanding the biological
mechanism of seedlings adaptation to drought in order
to mitigate water stress that is very damageable in
young plantations.
Acknowledgments Our thanks go to the German Federal
Ministry for Education and Research through BIOTA
(Biodiversity Monitoring Transect Analysis in Africa)
program for funding this research. We are very grateful to our
field assistant Nicolas Thiombiano who helped us in data
collection and also to Dr. Ross Bayton for his assistance in
improvement of the written English. We are grateful to the
anonymous reviewers whose pertinent comments considerably
improved the quality of our paper.
References
Agnew CT, Chappell A (1999) Drought in the Sahel. Geo-
Journal 48:299–311
Al-Helal AA (1996) Studies on germination of Rumex dentatusL. seeds. J Arid Environ 33:39–47
Bationo BA, Ouedraogo SJ, Guinko S (2001a) Longevite des
graines et contraintes a la survie des plantules d’Afzeliaafricana Sm. dans une savane boisee du Burkina Faso. Ann
For Sci 58:69–75
Bationo BA, Ouedraogo SJ, Guinko S (2001b) Strategies de
regeneration naturelle de Detarium microcarpum Guill. Et
Perr. dans la foret classee de Nazinon (Burkina Faso). Fruit
56:256–271
Belem B (2009) Ethnobotanique et conservation de Bombaxcostatum Pel. & Vuil. (faux kapokier) dans les systemes de
production agricoles du Plateau Central, Burkina Faso.
Dissertation, University of Ouagadougou
Belem B, Boussim IJ, Bellefontaine R, Guinko S (2008) Stim-
ulation du drageonnage de Bombax costatum par blessure
des racines au Burkina Faso. Bois et Forets des Tropiques
295:71–79
Agroforest Syst
123
Author's personal copy

Bellefontaine R, Edelin C, Ichaou A, du Laurens D, Monssarat
A, Loquai C (2000) Le drageonnage, alternative aux semis
et aux plantations de ligneux dans les zones semi-arides:
protocole de recherches. Secheresse-Science et Change-
ments planetaires. Numero special Desertification
11:221–226
Bognounou F, Tigabu M, Savadogo P, Thiombiano A, Boussim
IJ, Oden PC, Guinko S (2010) Regeneration of five Com-
bretaceae species along a latitudinal gradient in Sahelo-
Sudanien zone of Burkina Faso. Ann For Sci 67:1–10
Danthu P, Ndongo M, Diaou M, Thiam O, Sarr A, Dedhiou B,
Ould Mohamed Vall A (2003) Impact of bushfire on ger-
mination of some West African acacias. For Ecol Manage
173:1–10
Dauro D, Mohamed-Saleem MA, Gintzburger G (1997)
Recruitment and survival of native annual Trifolium spe-
cies in the highlands of Ethiopia. Afr J Ecol 35:1–19
Dawson IK, Lengkeek A, Weber JC, Jamnadass R (2009)
Managing genetic variation in tropical trees: linking
knowledge with action in agroforestry ecosystems for
improved conservation and enhanced livelihoods. Biodi-
vers Conserv 18:969–986
Dickinson MB, Whigham DF, Hermann SM (2000) Tree
regeneration in felling and natural tree fall disturbances in a
semideciduous tropical forest in Mexico. For Ecol Manage
134:137–151
Fontes J, Guinko S (1995) Carte de la vegetation et de l’occu-
pation du sol du Burkina Faso. Notice explicative. Min-
istere de la Cooperation Francaise. Projet Campus
(88 313 101), Toulouse
Ganaba S, Ouadba JM, Bognounou O (1998) Les ligneux a
usage de bois d’energie en region sahelienne du Burkina
Faso: preference des groupes ethniques. Secheresse
9:261–268
Gijsbers HJM, Kessler JJ, Knevel MK (1994) Dynamic and
natural regeneration of woody species in farmed parklands
in the Sahelian region (Province of Passore, Burkina Faso).
For Ecol Manage 64:1–12
Glele Kakaı R, Assogbadjo AE, Sinsin B, Pelz D (2009)
Structure spatiale et regeneration naturelle de Pterocarpuserinaceus Poir. en zone soudanienne au Benin. Revue
Ivoirienne des Sciences et Technologie 13:199–212
Guinko S, Pasgo LJ (1992) Recolte et commercialisation des
produits non ligneux des essences forestieres locales dans
le departement de Zitenga, province d’Oubritenga, Burkina
Faso. Etudes flore et vegetation du Burkina Faso et pays
avoisinants 1:47–56
Higgins IS, Shackleton MC, Robinson RE (1999) Changes in
woody community structure and composition under con-trasting land use systems in semi-arid savanna, South
Africa. J Biogeogr 26:619–627
Hoffmann AW (1998) Postburn reproduction of woody plants in
a neotropical savanna: the relative importance of sexual
reproduction and vegetative reproduction. J Appl Ecol
35:422–433
Kambou S (1997) Etude de la biologie de reproduction de
Anogeissus leiocarpus (DC.) Guill. et Per. au Burkina Faso.
Dissertation, University of Ouagadougou
Lykke AM (1998) Assessment of species composition change in
savanna vegetation by mean of woody plant’s size class
distribution and local information. Biodivers Conserv
7:1261–1275
Lykke AM, Kristensen MK, Ganaba S (2004) Valuation of local
use and dynamics of 56 woody species in the Sahel. Bio-
divers Conserv 13:1961–1990
Mbayngone E, Thiombiano A (2011) Degradation des aires
protegees par l’exploitation des ressources vegetales: cas
de la reserve partielle de faune de Pama, Burkina Faso
(Afrique de l’Ouest). Fruit 66:187–202
Mertz O, Lykke AM, Reenberg A (2001) Importance and sea-
sonality of vegetable consumption and marketing in
Burkina Faso. Econ Bot 55:276–289
Ministere des ressources animales (2004) Deuxieme enquete
nationale sur les effectifs du cheptel. Resultats et analyses.
Direction des Etudes et de la Planification, Ouagadougou
Mueller RC, Scudder CM, Porter ME, Trotter T III, Gehring CA,
Whitham TG (2005) Differential tree mortality in response
to severe drought: evidence for long-term vegetation shifts.
J Ecol 93:1085–1093
Nicholson SE, Tucker CJ, Ba MB (1998) Desertification,
drought, and surface vegetation: an example from the West
African Sahel. Bull Am Meteor Soc 79:815–829
Ouedraogo A (2006) Diversite et dynamique de la vegetation
ligneuse de la partie orientale du Burkina Faso. Disserta-
tion, University of Ouagadougou
Ouedraogo A, Thiombiano A, Hahn-Hadjali K, Guinko S (2005)
Utilisations, etat des peuplements et regeneration de 5 es-
peces ligneuses utilitaires dans l’Est du Burkina Faso.
Homme. In: Boussim IJ, Lykke AM, Nombre I, Nielsen I,
Guinko S (eds) Plantes et Environnement au Sahel Occi-
dental, Serein Occasional Paper 19. Aarhus, pp 173–183
Ouedraogo A, Thiombiano A, Hahn-Hadjali K, Guinko S
(2006a) Regeneration sexuee de Boswellia dalzielii Hutch.,
un arbre medicinal de grande valeur au Burkina Faso. Bois
et Forets des Tropiques 289:41–48
Ouedraogo A, Thiombiano A, Hahn-Hadjali K, Guinko S
(2006b) Diagnostic de l’etat de degradation des peuple-
ments de quatre especes ligneuses en zone soudanienne du
Burkina Faso. Secheresse 17:485–491
Ouedraogo A, Thiombiano A, Hahn-Hadjali K, Guinko S
(2006c) Structure du peuplement juvenile et potentialites
de regeneration des ligneux dans l’Est du Burkina Faso.
Etudes flore et vegetation du Burkina Faso et pays avoisi-
nants 10:17–24
Ouedraogo O, Thiombiano A, Hahn-Hadjali K, Guinko S (2009)
Diversite et dynamique de la vegetation ligneuse du Parc
National d’Arly (Burkina Faso). Candollea 64:257–278
Puig H, Fabre A, Gauquelin T (1998) Distribution spatiale des
plantules et des jeunes plants d’Iryanthera hostmannii(Benth.) Warb. en foret tropicale humide de Guyane
francaise. Sciences de la Vie/Life Sciences 321:429–435
Rasmussen K, Fog B, Madsen JE (2001) Desertification in
reverse? Observations from northern Burkina Faso. Global
Environ Change 11:271–282
Schutz W, Milberg P (1997) Seed germination in Launaea ar-borescens: a continuously flowering semi-desert shrub.
J Arid Environ 36:113–122
Setterfield SA (2002) Seedling establishment in an Australian
tropical savanna: effects of seed supply, soil disturbance
and fire. J Appl Ecol 39:949–956
Agroforest Syst
123
Author's personal copy

Sinsin B, Eyog Matig O, Assogbadjo AE, Gaoue AG, Sina-
douwirou T (2004) Dendrometric characteristics as indi-
cators of pressure of Afzelia africana Sm. dynamic changes
in trees found in different climatic zones of Benin. Biodi-
vers Conserv 13:1555–1570
Sop TK, Oldeland J, Schmieldel U, Ouedraogo I, Thiombiano A
(2010) Population structure of tree woody species in four
ethnic domains of the sub-Sahel of Burkina Faso. Land
Degrad Develop. doi:10.1002/ldr.1026
Terrible M (1984) Essai sur l’ecologie et la sociologie d’arbres
et arbustes de Haute-Volta
Thiombiano A, Wittig R, Guinko S (2003) Conditions de mul-
tiplication sexuee chez les Combretaceae du Burkina Faso.
Rev Ecol (Terre Vie) 58:361–379
Traore SA (1998) Quelques caracteristiques de la regeneration
naturelle des especes ligneuses du Parc National du
Niokolo Koba (Senegal Oriental), In: Ba AT, Madsen JE,
Sambou B (eds) Vegetation et Biodiversite au Sahel. AAU
Report, Aarhus, pp 275–288
Tremblay FM, Bergeron Y, Lalonde D, Maufette Y (2002) The
potential effects of sexual reproduction and seedling
recruitment on the maintenance of red maple (Acer rubrumL.) populations at the northern limit of the species range.
J Biogeogr 29:365–373
Weber JC, Larwanou M, Abasse TA, Kalinganire A (2008)
Growth and survival of Prosopis africana provenances
tested in Niger and related to rainfall gradients in the West
African Sahel. For Ecol Manage 256:585–592
White F (1983) Vegetation map of Africa. A descriptive memoir
to accompany the UNESCO/AETFAT/UNSO vegetation
map of Africa. UNESCO, Paris
White RP, Tunstall D, Henninger N (2002) An ecosystem
approach to drylands: building support for new develop-
ment policies. World Resources Institute. Information
Policy Brief No 1. Washington, DC
WRB (2006) World reference base for soil resources 2006. A
framework for international classification, correlation and
communication. FAO, Rome
Zida D, Sawadogo L, Tigabu M, Tiveau D, Oden PC (2007)
Dynamics of sapling population in savanna woodlands of
Burkina Faso subjected to grazing, early fire and selective
tree cutting for a decade. For Ecol Manage 243:102–115
Zida D, Tigabu M, Sawadogo L, Oden PC (2008) Initial seedling
morphological characteristics and field performance of two
Sudanian savanna species in relation to nursery production
period and watering regimes. For Ecol Manage 225:2151–
2162
Agroforest Syst
123
Author's personal copy




























