Embed Size (px)
Citation preview

Role of proteinase-activated receptor-2 in anti-bacterial andimmunomodulatory effects of interferon-c on human
neutrophils and monocytes
Victoria M. Shpacovitch,1,2,* Micha
Feld,1,* Dirk Holzinger,3,* Makiko
Kido,4,1 Morley D. Hollenberg,5
Francesca Levi-Schaffer,6 Nathalie
Vergnolle,7 Stephan Ludwig,8
Johannes Roth,3 Thomas Luger1
and Martin Steinhoff1,9
1Department of Dermatology, IZKF Munster,
Boltzmann Institute for Immunobiology of the
Skin, University of Munster, 2Department of
Nephrology and Hypertension & Endocrinology
and Metabolic Disease; Otto-von-Guericke
University of Magdeburg, Magdeburg, 3Insti-
tute of Immunology, University of Munster,
Munster, Germany, 4Department of Dermatol-
ogy, Graduate School of Medical Sciences, Kyu-
shu University, Fukuoka, Japan, 5Department
of Pharmacology and Therapeutics, Canadian
Institutes of Health Research Proteinases and
Inflammation Network, University of Calgary,
Calgary, AB, Canada, 6Department of Phar-
macology and Experimental Therapeutics,
School of Pharmacy, the Hebrew University of
Jerusalem, Israel, 7INSERM, U536 Centre de
Physiopathologie de Toulouse Purpan, Tou-
louse, France, 8Department of Molecular Virol-
ogy, ZMBE, University of Munster, Germany,
and 9Department of Dermatology and Surgery,
University of California San Francisco, San
Francisco, CA, USA
doi:10.1111/j.1365-2567.2011.03443.x
Received 15 September 2010; revised 24
February 2011; accepted 15 March 2011.
*These authors contributed equally to this
work.
Correspondence: V. M. Shpacovitch,
Department of Nephrology and
Hypertension & Endocrinology and
Metabolic Disease; Otto-von-Guericke
University of Magdeburg, Leipziger str. 40,
39120 Magdeburg, Germany.
Email: [email protected]
Senior author: Martin Steinhoff,
email: [email protected]
Summary
Recent studies show that proteinase-activated receptor-2 (PAR2) contrib-
utes to the development of inflammatory responses. However, investiga-
tions into the precise role of PAR2 activation in the anti-microbial
defence of human leucocytes are just beginning. We therefore evaluated
the contribution of PAR2 to the anti-microbial response of isolated
human innate immune cells. We found that PAR2 agonist, acting alone,
enhances phagocytosis of Staphylococcus aureus and killing of Escherichia
coli by human leucocytes, and that the magnitude of the effect is similar
to that of interferon-c (IFN-c). However, co-application of PAR2-cAP and
IFN-c did not enhance the phagocytic and bacteria-killing activity of leu-
cocytes beyond that triggered by either agonist alone. On the other hand,
IFN-c enhances PAR2 agonist-induced monocyte chemoattractant protein
1 (MCP-1) secretion by human neutrophils and monocytes. Furthermore,
phosphoinositide-3 kinase and janus kinase molecules are involved in the
synergistic effect of PAR2 agonist and IFN-c on MCP-1 secretion. Our
findings suggest a potentially protective role of PAR2 agonists in the anti-
microbial defence established by human monocytes and neutrophils.
Keywords: innate immunity; monocyte chemoattractant protein-1; phago-
cytosis; proteinase-activated receptor; Staphylococcus aureus
Abbreviations: FCS, fetal calf serum; IFN-c, interferon-c; MFI, mean fluorescence intensity; MCP-1, monocyte chemoattractantprotein-1 (other name is CCL2); PAR2, proteinase-activated receptor-2; PAR2-cAP, PAR2-tc-activating peptide; PAR2-cRP,PAR2-tc-reverse peptide.
� 2011 The Authors. Immunology � 2011 Blackwell Publishing Ltd, Immunology, 133, 329–339 329
I M M U N O L O G Y O R I G I N A L A R T I C L E

Introduction
Proteinase-activated receptor-2 (PAR2) plays a role in the
development of allergic diseases of the skin1 and in cer-
tain inflammatory disorders.2 The impact of PAR2 activa-
tion on inflammation can be pro- or anti-inflammatory,
depending on the stage of disease and the primary cell
type involved in disease progression.2 During receptor
activation, serine protease cleavage of PAR2 unmasks the
N-terminal sequence of the ‘tethered ligand’. This
unmasked sequence further serves as a receptor activator.3
The PAR2 is activated by trypsin and tryptase, and also
by proteases derived from immune cells and pathogens.4
However, serine proteases cause PAR-dependent as well
as PAR-independent effects.5,6 As a result, specific syn-
thetic activating peptides are important probes for investi-
gating the role of PAR activation in different processes.
Interferon-c (IFN-c) is well-known as a mediator that
has a wide range of anti-viral, anti-bacterial and anti-
tumour or immunomodulatory activities.7 We recently
demonstrated in vitro that PAR2 activation on human
monocytes enhances the suppressive effects of IFN-c on
influenza A virus replication.8 Moreover, in vivo studies
have shown that a protective role of PAR2 against influ-
enza infection is also mediated by an IFN-c-dependent
mechanism.9 These studies revealed interplay between
PAR2 activation and IFN-c during the anti-viral response
and raise the intriguing question of whether PAR2 activa-
tion also contributes to anti-bacterial and immunomodu-
latory effects triggered by IFN-c in monocytes and
neutrophils.
Human neutrophils and monocytes are not only ‘pro-
fessional’ phagocytes, they are cells that, when activated,
secrete different chemokines and cytokines. Stimulation of
PAR2 agonist affects chemokine [IFN-inducible protein-
10, interleukin-8 (IL-8)] and cytokine (IL-1b, IL-6) secre-
tion by human neutrophils and monocytes.8,10 Among
the chemokines secreted by neutrophils there is a mole-
cule that appears to link neutrophils and monocytes dur-
ing the time-delayed immune response to local infection.
Monocyte chemoattractant protein-1 (MCP-1) is an
essential mediator for monocyte and macrophage recruit-
ment towards the site of infection.11,12 Neutrophils are a
source of MCP-1 in time-delayed responses,13 and so may
attract monocytes and macrophages. However, MCP-1 is
not only a chemotactic molecule for monocytes and mac-
rophages, it also enhances the engulfment of apoptotic
neutrophils (efferocytosis), thereby helping to resolve
acute inflammation.14 In addition, MCP-1 is involved in
fibroblast activation and influences collagen production,
which makes MCP-1 an important participant in initial
events during systemic scleroderma and skin fibrosis.15
Interferon-c is known to increase the secretion of MCP-1
by human neutrophils 48 hr after stimulation.13 However,
whether PAR2 agonists enhance MCP-1 release or influ-
ence the IFN-c-induced secretion of MCP-1 by human
neutrophils has received little study.
We therefore evaluated the contribution of PAR2 to
the anti-microbial response of isolated human innate
immune cells. We investigated whether PAR2 agonist
acting alone affects the phagocytic and bactericidal activ-
ity of human neutrophils and monocytes. We also inves-
tigated whether IFN-c enhances the effect of PAR2
agonist on the MCP-1 release by human neutrophils and
monocytes, and examined the intracellular signalling
molecules involved in the effects of PAR2 agonist on
MCP-1 secretion.
Materials and methods
Materials
Human recombinant IFN-c was purchased from TebuBio
(Offenbach, Germany). Lipopolysaccharide (LPS) from
Escherichia coli 055:B5 was purchased from Sigma
(Munich, Germany; cat.#L2880). Human PAR2 activating
peptide with the sequence trans-cinnamoyl-LIGRLO-NH2
(cAP) and reverse peptide with sequence trans-cinnamoyl-
OLRGIL-NH2 (cRP) (Peptide Synthesis Facility, Univer-
sity of Calgary, Canada, Director: Dr Denis McMaster; the
web page: http://www.ucalgary.ca/peptides).8,10 Fluorescein-
conjugated killed Staphylococcus aureus was purchased
from Molecular Probes (Karlsruhe, Germany). The E. coli
strain JM109 was obtained from Promega (Mannheim,
Germany). Cell culture reagents were purchased from Bio-
Whittaker (Aachen, Germany), PAA Laboratories (Coelbe,
Germany) and Gibco-Life Technologies (Karlsruhe,
Germany). Cell-permeable inhibitors of intracellular sig-
nalling molecules [SB203580, rottlerin, LY 294002 and
janus kinase (JAK) inhibitor I pyridone 6] were purchased
from Calbiochem (Nottingham, UK).
Cell culture and isolation of human neutrophils andmonocytes
Buffy-coats with blood cells for in vitro experiments with
human neutrophils and monocytes were obtained from
healthy adult volunteers via the German Red Cross (De-
utsches Rotes Kreuz, Munster, Germany). Neutrophils
were isolated by Biocoll (Biochrom, Berlin, Germany)
density gradient centrifugation followed by a hypotonic
shock procedure.10,16 Peripheral blood monocytes were
isolated by leukapheresis as previously described.17 Iso-
lated human monocytes were cultivated in Teflon bags in
McCoy’s medium (Biochrom) supplemented with 15%
fetal calf serum (FCS), 2 mM L-glutamine and 1% non-
essential amino acids. Monocytes were allowed to rest for
24 hr before stimulation. Isolated neutrophils were cul-
tured in RPMI-1640 medium supplemented with 0�9%
FCS, 2 mM L-glutamine and 1% non-essential amino acids
330 � 2011 The Authors. Immunology � 2011 Blackwell Publishing Ltd, Immunology, 133, 329–339
V. M. Shpacovitch et al.

and allowed to recover for 2 hr before stimulation. The
following concentrations of reagents were used for stimu-
lation during experiments: LPS 100 ng/ml; IFN-c 10 or
100 ng/ml; PAR2-cAP 1 · 10)4M. The corresponding
reverse peptide with the reverse-sequence (PAR2-cRP) was
used at a concentration of 1 · 10)4M and served as a
negative control.
Bacterial killing assay
Bacterial killing assay using E. coli (strain JM109) was
performed as described previously18,19 with modifications.
In brief, E. coli bacteria were cultured into Luria broth
medium overnight at 37�. Isolated uninfected human
neutrophils were pre-stimulated with 10)4M PAR2-cAP
and/or IFN-c (100 ng/ml) for 2 hr (37�, 5% CO2).
Unstimulated neutrophils were used as control samples.
After 2 hr incubation of neutrophils with stimuli, the cell
culture medium (RPMI-1640 with 0�9% FCS, 2 mM L-glu-
tamine and 1% non-essential amino acids) with stimuli
was removed and cells were washed. Human neutrophils
(2 · 106 cells) were resuspended in 200 ll RPMI-1640
containing 2 mM L-glutamine, 1% non-essential amino
acids, 0�2% BSA, 0�01% CaCl2 and 0�01% MgCl2 (this
medium was designated the ‘assay medium’). Collected
and washed bacteria (40 · 106 cells) were opsonized for
15 min at 37�. For opsonization, bacteria were incubated
in the assay medium containing 5% human serum from
the same donor from whom the neutrophils were
obtained. After opsonization, bacteria were washed. Neu-
trophils and opsonized bacteria were co-incubated in
assay medium in the absence (for unstimulated control
samples) or presence of stimuli (10)4M PAR2-cAP and/or
100 ng/ml IFN-c) for 1 hr at 37� on a shaker. Neutrophils
were removed from samples by hypotonic lysis (for this
purpose sterile water was added to samples). A series of
dilutions were prepared from the remaining bacteria. Bac-
teria were cultured on Luria broth agar plates without
antibiotic at 37� overnight. Colonies were counted the
next day.
Phagocytosis assay with killed S. aureus
The phagocytosis assay was performed as described previ-
ously.19,20 In brief, FITC-conjugated killed S. aureus (Invi-
trogen, Darmstadt, Germany) was used for assay. The
bacteria were opsonized before the assay. For this pur-
pose, bacteria were incubated with 5% serum (from the
same donor from whom neutrophils were isolated) for
25 min at 37�. Non-infected neutrophils were pre-stimu-
lated with PAR2-cAP 10)4M, PAR2-cRP 10)4
M and/or
IFN-c 100 ng/ml for 2 hr at 37� and 5% CO2. Neutroph-
ils and opsonized bacteria were co-incubated at 1 : 20
ratio (neutrophils : S. aureus). During co-incubation of
bacteria and neutrophils, PAR2-cAP 10)4M, PAR2-cRP
10)4M and/or IFN-c 100 ng/ml were applied in the con-
centrations indicated above. Co-incubation took place in
assay medium on a shaker for 30 min at 37�. The phago-
cytosis assay was stopped by the addition of ice-cold PBS
containing 0�5 mM EDTA (500 ll PBS to 1 ml of sample
medium). Samples were then centrifuged at 169 g and
neutrophil pellets were resuspended in ice-cold PBS con-
taining 0�9% FCS and 2 mM EDTA. Trypan blue quench,
which helps to discriminate adherent and ingested bacte-
ria, was performed as described previously.21 The efficacy
of phagocytosis was estimated using flow cytometry
(FACS analysis). Measurements were performed for the
next 15 min and all samples were kept on ice during
measurements. At least 30 000 cells were analysed with
the FACSCALIBUR and CELL QUEST PRO Software (Becton
Dickinson, Heidelberg, Germany).
Phagocytosis assay with live S. aureus
Bacteria. The S. aureus (SH1000) was kindly provided
by Dr C. Eiff22 and S. aureus was grown for 18 hr in
Mueller–Hinton bouillon at 37�. Bacterial density was
measured spectrophotometrically at 540 nm, after two
PBS washings. The number of bacterial cells was calcu-
lated using a previously determined standard curve
(based on the counts of colony-forming units). Finally,
the concentration of bacteria in PBS was adjusted to
5 · 108 cells/ml. For the purpose of the quantitative
analysis of phagocytosis by flow cytometry, S. aureus was
incubated in PBS containing 0�1% FITC (Sigma Aldrich,
Munich, Germany) for 1 hr at 37�. After being labelled,
bacteria were washed three times before incubation with
pre-treated leucocytes.
Assay. During pre-treatment, human monocytes or neu-
trophils (1 · 106 cells) were cultured in medium either
without stimuli (‘control’) or containing the following
stimuli: 100 ng/ml LPS; 1 · 10)4M PAR2-cAP, 10 ng/ml
or 100 ng/ml of IFN-c. Monocytes or neutrophils were
pre-treated for 2 hr at 37� and subsequently co-incubated
with live FITC-labelled S. aureus at a ratio of 1 : 10
(cells : S. aureus) for 30 min at 37�. During co-incubation
of leucocytes and bacteria, PAR2-cAP and IFN-c were
applied in the same order and the same concentrations as
during the pre-treatment of leucocytes by these stimuli.
The efficacy of phagocytosis was determined by FACS
analysis as described previously.23
Measurement of MCP-1 release. The inhibition of cellsignalling molecules involved in the changes ofMCP-1 release
Non-infected human neutrophils (3�75 · 106 cells) and
monocytes (3 · 106 cells) were treated with PAR2-cAP
� 2011 The Authors. Immunology � 2011 Blackwell Publishing Ltd, Immunology, 133, 329–339 331
PAR2 and anti-microbial defence of human leucocytes

and/or IFN-c for 20 or 28 hr. Cell culture supernatants
were collected and used for MCP-1 ELISA. Concentration
of MCP-1 in the cell culture supernatants was measured
with a human CCL2/MCP-1 (R&D Systems, Wiesbaden-
Nordenstadt, Germany) ELISA kit according to the man-
ufacturer’s instructions.
Specific inhibitors of intracellular signalling molecules
were used to reveal which ones are involved in the effects
of PAR2-cAP and/or IFN-c at MCP-1 secretion by human
neutrophils and monocytes. The inhibitors were used in
the following concentrations: rottlerin [inhibits protein
kinase Cd (PKCd)] 5 lM; LY294002 [inhibits phosphoino-
sitide 3 (PI3) kinase] 50 lM; SB203580 (inhibits p38
kinase) 1 lM; and JAK inhibitor I pyridone 6 (pan-JAK
inhibitor) 500 nM. All inhibitors were dissolved in DMSO,
so the vehicle DMSO (1 : 1000) was used as an additional
control. Human neutrophils and monocytes were pre-
treated with the inhibitors for 30 min and then PAR2-
cAP (1 · 10)4M) alone or in combination with IFN-c
(100 ng/ml) was applied for 28 hr (the maximum effect
of the stimuli at MCP-1 release was noticed at this time-
point). After treatment, cell culture supernatants were
collected and used to measure MCP-1 concentration by
human CCL2/MCP-1 (R&D Systems) ELISA kit.
Statistical analysis
Results are expressed as mean ± SEM. At least three inde-
pendent experiments were performed. Statistical evalua-
tion was performed by paired two-tailed Student’s t-tests.
Significance was set at P < 0�05.
Results
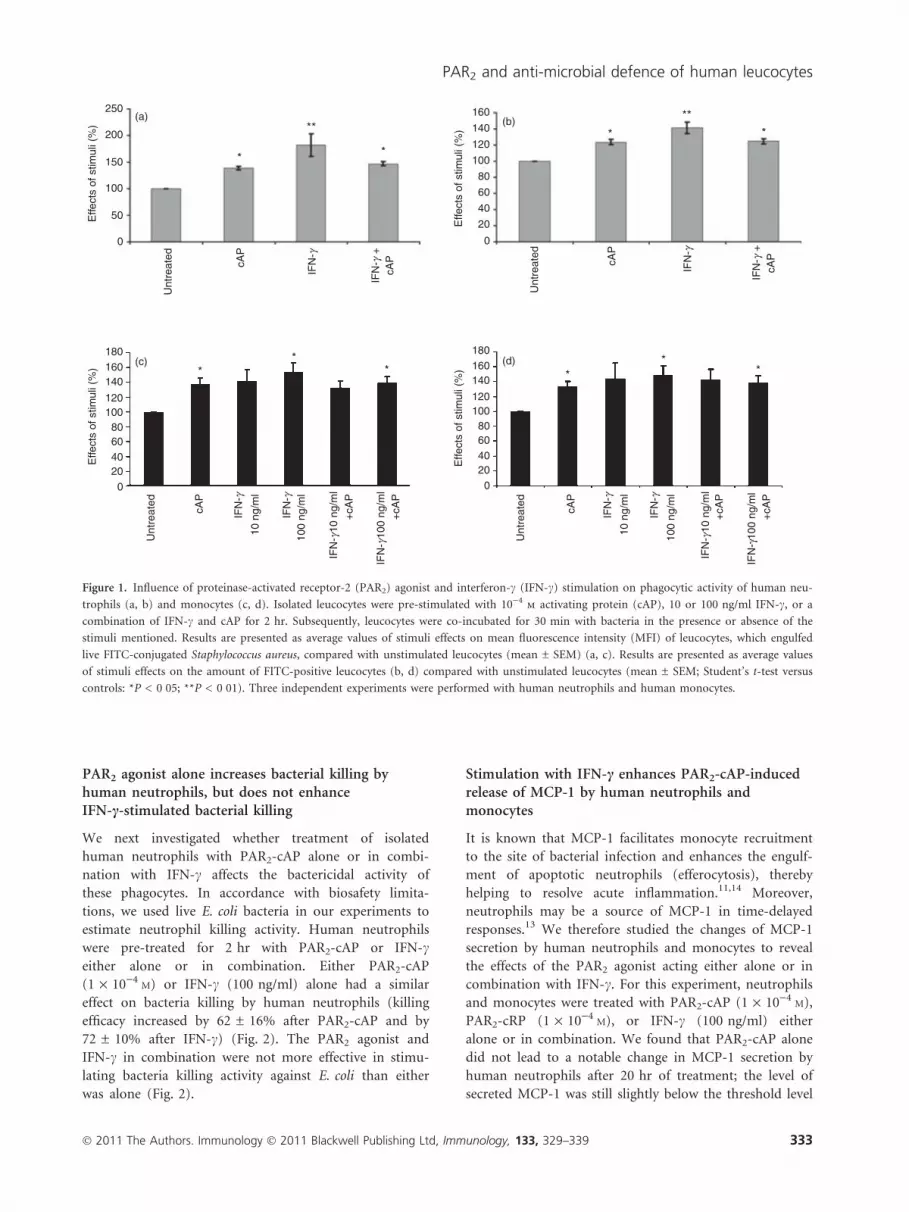
PAR2 agonist alone enhances the phagocytic activityof human neutrophils and monocytes, but does notaffect IFN-c-stimulated phagocytosis
Neutrophils and macrophages from PAR2-deficient mice
have been shown to display a significantly reduced phago-
cytic efficiency of Pseudomonas aeruginosa compared with
cells from wild-type animals.24 However, the ability of
PAR2 agonist to enhance the phagocytic activity of
human neutrophils and monocytes and to affect IFN-c-
stimulated phagocytosis has yet to be evaluated. To inves-
tigate whether PAR2 agonist might potentially enhance
the IFN-c-induced phagocytosis we first carried out the
phagocytosis assay with FITC-conjugated killed S. aureus.
The treatment of human neutrophils with either PAR2-
cAP (1 · 10)4M) or IFN-c (100 ng/ml) alone led to a
similar enhancement of the mean fluorescence intensity
(MFI) of human neutrophils (increased by around
40 ± 7% compared with untreated cells), indicating that
the phagocytic activity of treated neutrophils increased
(see supplementary material, Fig. S1). The combined
action of PAR2-cAP and IFN-c did not enhance the phag-
ocytic activity of neutrophils above that triggered by
either agonist acting alone (combined treatment increased
phagocytic activity by around 51 ± 12% as compared
with untreated cells) (Fig. S1). Treatment with the con-
trol peptide that cannot activate PAR2 (PAR2-cRP:
1 · 10)4M) either alone or together with IFN-c did not
affect the phagocytic activity of human neutrophils (Fig.
S1).
In our next experiments, we used live FITC-conjugated
S. aureus (strain SH1000) to investigate the effect of
PAR2-cAP alone or together with IFN-c on the phagocytic
activity of human monocytes and neutrophils against via-
ble bacteria. We found that PAR2-cAP (1 · 10)4M) or
IFN-c (100 ng/ml) alone enhanced phagocytic activity
(Fig. 1a–d; a,b for neutrophils and c,d for monocytes).
Although IFN-c already appeared to stimulate phagocytic
activity of monocytes at a concentration of 10 ng/ml,
these effects were not statistically significant (Fig. 1c,d).
Interferon-c at a higher concentration (100 ng/ml) also
enhanced phagocytic activity of human monocytes and
neutrophils. The effects of IFN-c at a concentration of
100 ng/ml reached statistical significance (Fig. 1a–d).
Stimulation with IFN-c increased the number of FITC-
positive human monocytes (49 ± 13% of change com-
pared with untreated cells) and FITC-positive human
neutrophils (41 ± 7% of change compared with untreated
cells). The MFI also increased in IFN-c-treated human
monocytes (increased by 53 ± 14%) and neutrophils
(increased by 80 ± 18%) compared with untreated con-
trols. PAR2-cAP led to an increase in the amount of
FITC-positive monocytes (increased by 35 ± 7%) and
FITC-positive neutrophils (increased by 24 ± 4%) com-
pared with untreated samples. The MFI also increased in
monocytes treated with PAR2-cAP (increased by
38 ± 8%) and in neutrophils (increased by 38 ± 4%)
compared with untreated control samples. The combined
action of PAR2-cAP and IFN-c using the same concentra-
tions did not enhance the phagocytic activity of neu-
trophils or monocytes beyond that triggered by either
agonist acting alone (Fig. 1a–d).
Interferon-c is a well-known endogenous modulator of
phagocytic bacteria killing and secretory activity of
human neutrophils and human monocytes.25,26 As an
exogenous activator, LPS also affects phagocytic activity
of both cell types. We wondered whether PAR2-cAP stim-
ulation might interfere with LPS-modulated phagocytic
activity of human neutrophils and monocytes. However,
PAR2-cAP stimulation of human neutrophils as well as
monocytes did not enhance the LPS-induced phagocytic
activity against S. aureus (see supplementary material, Fig.
S2). Hence, despite the fact that PAR2-cAP alone up-reg-
ulates the phagocytic activity of human neutrophils and
monocytes against S. aureus, this agonist failed to enhance
IFN-c-induced and LPS-induced phagocytic activity.
332 � 2011 The Authors. Immunology � 2011 Blackwell Publishing Ltd, Immunology, 133, 329–339
V. M. Shpacovitch et al.
PAR2 agonist alone increases bacterial killing byhuman neutrophils, but does not enhanceIFN-c-stimulated bacterial killing
We next investigated whether treatment of isolated
human neutrophils with PAR2-cAP alone or in combi-
nation with IFN-c affects the bactericidal activity of
these phagocytes. In accordance with biosafety limita-
tions, we used live E. coli bacteria in our experiments to
estimate neutrophil killing activity. Human neutrophils
were pre-treated for 2 hr with PAR2-cAP or IFN-ceither alone or in combination. Either PAR2-cAP
(1 · 10)4M) or IFN-c (100 ng/ml) alone had a similar
effect on bacteria killing by human neutrophils (killing
efficacy increased by 62 ± 16% after PAR2-cAP and by
72 ± 10% after IFN-c) (Fig. 2). The PAR2 agonist and
IFN-c in combination were not more effective in stimu-
lating bacteria killing activity against E. coli than either
was alone (Fig. 2).
Stimulation with IFN-c enhances PAR2-cAP-inducedrelease of MCP-1 by human neutrophils andmonocytes
It is known that MCP-1 facilitates monocyte recruitment
to the site of bacterial infection and enhances the engulf-
ment of apoptotic neutrophils (efferocytosis), thereby
helping to resolve acute inflammation.11,14 Moreover,
neutrophils may be a source of MCP-1 in time-delayed
responses.13 We therefore studied the changes of MCP-1
secretion by human neutrophils and monocytes to reveal
the effects of the PAR2 agonist acting either alone or in
combination with IFN-c. For this experiment, neutrophils
and monocytes were treated with PAR2-cAP (1 · 10)4M),
PAR2-cRP (1 · 10)4M), or IFN-c (100 ng/ml) either
alone or in combination. We found that PAR2-cAP alone
did not lead to a notable change in MCP-1 secretion by
human neutrophils after 20 hr of treatment; the level of
secreted MCP-1 was still slightly below the threshold level
250
200
150
100
50
0 0
0
20
20
40
40
60
60
80
80
100
100
120
120
140
140
160
160180
0
20406080
100120
140160180
Unt
reat
ed
Unt
reat
ed
Unt
reat
edU
ntre
ated
cAP
cAP
cAP
cAP
cAP
cAP
+cA
P
+cA
P
+cA
P
+cA
P
IFN
-g
IFN
-g +
IFN
-g
IFN
-g +
IFN
-γ10
0 ng
/ml
IFN
-g10
ng/
ml
IFN
-g
IFN
-g
IFN
-g10
0 ng
/ml
IFN
- g10
ng/
ml
IFN
- g
IFN
-g10
ng/
ml
10 n
g/m
l
100
ng/m
l
100
ng/m
l
* **
* ***
*
* **
*
**
Effe
cts
of s
timul
i (%
)E
ffect
s of
stim
uli (
%)
Effe
cts
of s
timul
i (%
)E
ffect
s of
stim
uli (
%)
(a) (b)
(c) (d)
Figure 1. Influence of proteinase-activated receptor-2 (PAR2) agonist and interferon-c (IFN-c) stimulation on phagocytic activity of human neu-
trophils (a, b) and monocytes (c, d). Isolated leucocytes were pre-stimulated with 10)4m activating protein (cAP), 10 or 100 ng/ml IFN-c, or a
combination of IFN-c and cAP for 2 hr. Subsequently, leucocytes were co-incubated for 30 min with bacteria in the presence or absence of the
stimuli mentioned. Results are presented as average values of stimuli effects on mean fluorescence intensity (MFI) of leucocytes, which engulfed
live FITC-conjugated Staphylococcus aureus, compared with unstimulated leucocytes (mean ± SEM) (a, c). Results are presented as average values
of stimuli effects on the amount of FITC-positive leucocytes (b, d) compared with unstimulated leucocytes (mean ± SEM; Student’s t-test versus
controls: *P < 0�05; **P < 0�01). Three independent experiments were performed with human neutrophils and human monocytes.
� 2011 The Authors. Immunology � 2011 Blackwell Publishing Ltd, Immunology, 133, 329–339 333
PAR2 and anti-microbial defence of human leucocytes

of the ELISA (Fig. 3a). However, treatment of human
neutrophils with PAR2-cAP for 28 hr resulted in a signifi-
cant increase of MCP-1 secretion by these cells (MCP-1
level in PAR2-cAP stimulated samples was 36 ± 4 pg/ml,
but was undetectable in unstimulated control samples)
(Fig. 3b). Treatment of neutrophils with IFN-c alone did
not affect MCP-1 secretion at the 20 and 28 hr time-
points. The level of secreted MCP-1 was below the thresh-
old level of the ELISA at 20 hr and at 28 hr (Fig. 3a,b).
Surprisingly, the co-application of IFN-c with PAR2-cAP
enhanced the effect of the PAR2 agonist on MCP-1 secre-
tion 20 hr after stimulation (Fig. 3a). This effect was sta-
tistically significant even at 20 hr after stimulation
(Fig. 3a). However, this effect was even more prominent
at 28 hr (MCP-1 level was 284 ± 37 pg/ml versus 36 ±
4 pg/ml in samples treated by PAR2-cAP alone) (Fig. 3b).
Treatment with the PAR2-inactive control peptide
PAR2-cRP (1 · 10)4M) alone or together with IFN-c did
not affect MCP-1 secretion by human neutrophils
(Fig. 3a,b).
We also investigated whether treatment of human
monocytes with PAR2-cAP alone or in combination with
IFN-c affects MCP-1 secretion. Here, we measured the
level of secreted MCP-1 at 28 hr after stimulation of
human monocytes with PAR2-cAP or IFN-c alone or in
combination. We found that stimulation of human neu-
trophils for 28 hr with PAR2-cAP alone, but especially in
combination with IFN-c, led to a statistically significant
increase of MCP-1 secretion. We wondered whether
monocytes would also be responsive to such stimulation
at this time-point. Indeed, PAR2-cAP enhanced MCP-1
secretion by human monocytes (Fig. 3c). The level of
MCP-1 in untreated monocytes was 71 ± 10 pg/ml, but it
was 271 ± 60 pg/ml in monocytes treated with PAR2-
cAP. When monocytes were stimulated with IFN-c alone
0
20
40
60
80
100
120
Con. cAP IFN-g IFN-g+cAP
**
**
**
Live
bac
teria
com
pare
d w
ithco
ntro
l (%
)
Figure 2. Influence of proteinase-activated receptor-2 (PAR2) agonist
and interferon-c (IFN-c) stimulation on bacteria-killing activity of
human neutrophils. Neutrophils were pre-stimulated with 10)4m
activating peptide (cAP), 100 ng/ml IFN-c, or both, for 2 hr before
incubation with living Escherichia coli. The same stimuli were further
used during incubation of neutrophils and bacteria. The number of
bacterial colonies formed after co-incubation with untreated neu-
trophils was considered to be 100%. The results are presented as an
average number of bacterial colonies formed after co-incubation with
stimulated neutrophils ± SEM (results are expressed as percentages).
Student’s t-test versus controls: **P < 0�01. Eight independent exper-
iments were performed.
50
40
30
20
10
0
cont
.
cAP
cRP
IFN
-g
IFN
-g+
cAP
IFN
-g+
cRP
***
MC
P-1
rel
ease
(pg
/ml)
350
300
250
200150
100
50
0
cont
.
cAP
cRP
IFN
-g
IFN
-g+
cAP
IFN
-g+
cRP
***
***
MC
P-1
rel
ease
(pg
/ml)
2500
2000
1500
1000
500
0
cont
.
cAP
IFN
-g
IFN
-g+
cAP
***
**
MC
P-1
rel
ease
(pg
/ml)
(a)
(b)
(c)
Figure 3. Analysis of changes in monocyte chemoattractant protein
1 (MCP-1) secretion by human neutrophils and monocytes after
proteinase-activated receptor-2 activating peptide (PAR2-cAP),
PAR2-reverse peptide (cRP) and interferon-c (IFN-c) stimulation.
Neutrophils were stimulated with 10)4m cAP, 10)4
m cRP, 100 ng/
ml IFN-c alone or in combination for 20 hr (a) or 28 hr (b). Mono-
cytes were stimulated with 10)4m cAP, 100 ng/ml IFN-c alone or in
combination for 28 hr (c). The sensitivity threshold of the assay was
15�6 pg/ml. This level is marked on (a, b) by dotted lines. Results
are presented as the average values of MCP-1 secretion by unstimu-
lated (control) as well as by stimulated leucocytes in pg/ml ± SEM.
Student’s t-test versus controls: **P < 0�01; ***P < 0�005. Five inde-
pendent experiments were performed for human neutrophils and
four independent experiments for human monocytes.
334 � 2011 The Authors. Immunology � 2011 Blackwell Publishing Ltd, Immunology, 133, 329–339
V. M. Shpacovitch et al.

MCP-1 secretion was not notably affected (Fig. 3c). How-
ever, when IFN-c and PAR2-cAP were used together,
MCP-1 secretion was enhanced significantly (1686 ±
335 pg/ml versus 271 ± 60 pg/ml in samples treated by
PAR2-cAP alone) (Fig. 3c).
The inhibition of PI3 kinase and PKCd abolishes theenhancement of MCP-1 secretion after co-stimulationof human neutrophils with IFN-c and PAR2-cAP
We next investigated which intracellular signalling mole-
cules were involved in the effects of PAR2 agonist on
MCP-1 secretion by human neutrophils, when this agonist
was applied alone or in combination with IFN-c. In these
experiments, we investigated the effects of the inhibitors of
different intracellular signalling molecules: rottlerin (inhib-
its PKCd), LY294002 (inhibits PI3 kinase), SB203580
(inhibits p38 kinase), and JAK inhibitor I pyridone 6
(inhibits JAKs). Experiments were performed with neu-
trophils treated for 28 hr with PAR2 agonist alone (PAR2-
cAP 1 · 10)4M) or in combination with IFN-c (100 ng/
ml), because the maximum effect on the MCP-1 secretion
was revealed at that time-point. We found that rottlerin
and LY294002 each completely abolished the effect of
co-application of PAR2-cAP and IFN-c on MCP-1 release
by human neutrophils (Fig. 4a). These results indicate the
crucial role of PI3 kinase and PKCd in enhancing MCP-1
secretion after co-stimulation of human neutrophils with
PAR2-cAP and IFN-c. In addition, treating neutrophils
with either pyridine 6 or SB203580 only weakened the
effect of PAR2-cAP and IFN-c on MCP-1 secretion, which
shows that p38 kinase and JAKs are involved in the com-
bined action of both agonists (Fig. 4a).
We also examined which intracellular signalling mole-
cules are involved in the enhanced secretion of MCP-1 by
human neutrophils after treatment with PAR2-cAP alone
(Fig. 4b). For these experiments, rottlerin, LY294002,
SB203580 and pyridine 6 were used to check whether PI3
kinase, PKCd, p38 kinase and JAKs were involved in the
solo effect of PAR2 agonist on MCP-1 secretion by neu-
trophils. Rottlerin, LY294002 and SB203580 abolished
PAR2-cAP-induced MCP-1 secretion (Fig. 4b), indicating
a crucial role of PI3 kinase, p38 kinase and PKCd on the
effect of PAR2 stimulation. However, pyridine 6 did not
significantly affect the changes in MCP-1 release, indicat-
ing that JAKs do not participate in the effect induced by
PAR2 agonist alone (Fig. 4b).
The inhibition of PI3 kinase and JAKs in humanmonocytes reduces the enhancement of MCP-1secretion caused by co-stimulation withPAR2-cAP and IFN-c
We also investigated whether rottlerin (inhibits PKCd),
LY294002 (inhibits PI3 kinase), SB203580 (inhibits p38
kinase), and JAK inhibitor I pyridone 6 (inhibits JAKs)
affected the induction of MCP-1 secretion after stimula-
tion of human monocytes with PAR2-cAP and IFN-c(Fig. 5a). Experiments were performed with monocytes
treated with PAR2-cAP together with IFN-c for 28 hr. We
found that LY294002 and JAK inhibitor I pyridone 6 each
reduced the effect of co-stimulation with PAR2-cAP and
IFN-c on the MCP-1 secretion by human monocytes
(MCP-1 level was 1686 ± 335 pg/ml in samples treated
with both PAR2-cAP and IFN-c, 333 ± 140 pg/ml in sam-
ples treated with LY294002, and 352 ± 121 pg/ml in sam-
ples treated with JAK inhibitor) (Fig. 5a). SB203580 had
0
50
100
150
200
250
300
350
MC
P-1
pro
tein
(pg
/ml)
Unt
reat
ed
IFN
-g+
cAP
IFN
-g+
cAP
+
IFN
-g+
cAP
+
IFN
-g+
cAP
+
IFN
-g+
cAP
+
IFN
-g+
cAP
+LY
2940
02
rottl
erin
SB
2035
80
pan
JAK
inh.
DM
SO
(a)
***
***
***
0
5
10
15
20
25
30
35
40
45
MC
P-1
pro
tein
(pg
/ml)
Unt
reat
ed
cAP
cAP
+LY
2940
02
cAP
+ro
ttler
in
cAP
+S
B20
3580
cAP
+pa
n JA
K in
h.
cAP
+D
MS
O
(b)
***
******
Figure 4. Inhibition of the signalling molecules during co-applica-
tion of proteinase-activated receptor-2 activating peptide (PAR2-
cAP) and interferon-c (IFN-c) (a) and during application of PAR2-
cAP alone (b) affects monocyte chemoattractant 1 (MCP-1) secretion
by human neutrophils. The following inhibitors were used: rottlerin
(protein kinase-d; PKCd) 5 lm; LY294002 [phosphoinositide-3 (PI3)
kinase] 50 lm; SB203580 (p38 kinase) 1 lm; and janus kinase (JAK)
inhibitor I pyridone 6 (pan JAK) 500 nm. Cells were pre-treated with
inhibitors and vehicle (DMSO; 1 : 1000) as described in the Materi-
als and methods, and further treated with PAR2-cAP 1 · 10)4m
together with IFN-c 100 ng/ml (a) or with PAR2-cAP 1 · 10)4m
alone (b) for 28 hr. Human MCP-1 level in collected samples was
determined by ELISA. Results are presented as the average values of
MCP-1 secretion by unstimulated, as well as by stimulated neutroph-
ils in pg/ml ± SEM. Student’s pairwise t-test: *P < 0�05, **P < 0�01,
***P < 0�005. Four independent experiments were performed.
� 2011 The Authors. Immunology � 2011 Blackwell Publishing Ltd, Immunology, 133, 329–339 335
PAR2 and anti-microbial defence of human leucocytes

no effect on MCP-1 secretion by human monocytes
(Fig. 5a). Surprisingly, rottlerin enhanced the effect of
co-stimulation with PAR2-cAP and IFN-c on MCP-1
secretion by monocytes (Fig. 5a) and also enhanced
PAR2-cAP-induced MCP-1 release when PAR2 agonist
was used alone (Fig. 5b). However, rottlerin did not affect
MCP-1 levels in IFN-c stimulated cells (data not shown).
We were also interested in whether rottlerin alone might
affect MCP-1 secretion by human monocytes and found
that it did increase secretion (Fig. 5c). SB203580 and
JAK inhibitor each did not affect MCP-1 secretion trig-
gered by PAR2-cAP (Fig. 5b). LY294002 slightly reduced
the effect of PAR2-cAP stimulation on MCP-1 secretion
by human monocytes (the level of MCP-1 secretion
after PAR2-cAP application was 271 ± 60 pg/ml and if
LY294002 was also added, the level of MCP-1 was
154 ± 72 pg/ml) (Fig. 5b). In all cases, treatment of
monocytes with DMSO did not affect MCP-1 secretion
(Fig. 5a–c).
Discussion
The most important finding of our study is that PAR2 acti-
vation enhances phagocytic activity against Gram-positive
(S. aureus) bacteria and the killing of Gram-
negative (E. coli) bacteria by human leucocytes. The
magnitude of the bactericidal effect induced by PAR2 ago-
nist was similar to that induced by IFN-c (Figs 1 and 2; see
supplementary material, Fig. S1). Since PAR2 agonist can
synergize with IFN-c in enhancing anti-viral responses,8,9
we investigated whether co-application of PAR2-cAP and
IFN-c led to stronger anti-bacterial responses of innate
immune cells, but found that the response was no greater
than when each compound was used alone (Figs 1 and 2;
Fig. S1). In addition, PAR2 agonist stimulation also failed to
enhance LPS-stimulated phagocytic activity of neutrophils
and monocytes (see supplementary material, Fig. S2).
Hence, PAR2 stimulation might trigger additional mecha-
nisms that enhance the phagocytic activity of innate
3500
3000
2500
2000
1500
1000
800
600
400
200
0
1200
1000
500
0
MC
P-1
pro
tein
(pg
/ml)
3500
3000
2500
2000
1500
1000
500
0
MC
P-1
pro
tein
(pg
/ml)
MC
P-1
pro
tein
(pg
/ml)
Unt
reat
ed
Unt
reat
ed
IFN
-g+
cAP
IFN
-g+
cAP
+
IFN
-g+
cAP
+
IFN
-g+
cAP
+
IFN
-g+
cAP
+
IFN
-g+
cAP
+
LY29
4002
LY29
4002
LY29
4002
rottl
erin
rottl
erin
rottl
erin
SB
2035
80
SB
2035
80
SB
2035
80
pan
JAK
inh.
pan
JAK
inh.
pan
JAK
inh.
DM
SO
DM
SO
DM
SO
cAP
+
cAP
+
cAP
+
cAP
+
cAP
+
cAP
Unt
reat
ed
# #
(a) (b)
(c)
Figure 5. Inhibition of the signalling molecules during co-application of proteinase-activated receptor-2 activating peptide (PAR2-cAP) and inter-
feron-c (IFN-c) (a) and during application of PAR2-cAP alone (b) affects monocyte chemoattractant protein 1 (MCP-1) secretion by human
monocytes. In addition, the effects of inhibitors of the signalling molecules alone on MCP-1 secretion were checked (c). The following inhibitors
were used: rottlerin (protein kinase d; PKCd) 5 lm; LY294002 [phosphoinositide 3 (PI3) kinase] 50 lm; SB203580 (p38 kinase) 1 lm; and janus
kinase (JAK) inhibitor I pyridone 6 (pan JAK) 500 nm. Cells were pre-treated with inhibitors and vehicle (DMSO; 1 : 1000) as described in the
Materials and methods, and further treated with PAR2-cAP 1 · 10)4m together with IFN-c 100 ng/ml (a) or with PAR2-cAP 1 · 10)4
m alone
(b) for 28 hr. Human MCP-1 level in collected samples was determined by ELISA. Results are presented as the average values of MCP-1 secretion
by unstimulated, as well as by stimulated monocytes in pg/ml ± SEM. Student’s pairwise t-test: #P < 0�05. Four independent experiments were
performed.
336 � 2011 The Authors. Immunology � 2011 Blackwell Publishing Ltd, Immunology, 133, 329–339
V. M. Shpacovitch et al.

immune cells, and these mechanisms do not synergize with
IFN-c or LPS-triggered ones. Unfortunately, it remains
problematic to investigate whether the classic PAR2 activa-
tors trypsin and tryptase can affect phagocytic and bacteria-
killing activity of human innate immune cells. Trypsin and
tryptase are known to induce PAR-independent effects.5,6
These effects could confound the data obtained using these
enzymes as PAR2 agonists.
Cytokines and chemokines influence the recruitment of
phagocytes to the site of pathogen infection. The PAR2
agonists reportedly affect the secretion of IFN-inducible
protein-10, IL-8, IL-6 and IL-1b by human neutrophils,
monocytes and endothelial cells.8,10,27 Among chemokin-
es, MCP-1 appears to play a distinct role linking
neutrophils and monocytes during time-delayed inflam-
matory response, and helping to resolve inflammation via
activation of efferocytosis.14 In addition, IFN-c reportedly
enhances time-delayed MCP-1 secretion by human neu-
trophils.13 In our study, we investigated whether PAR2
activation interferes or synergizes with IFN-c-enhanced
MCP-1 secretion. We found that MCP-1 secretion by
human neutrophils and monocytes was enhanced 28 hr
after stimulation with PAR2-cAP (Fig. 3). Moreover, the
treatment of human neutrophils and monocytes with
IFN-c together with PAR2-cAP resulted in a synergistic
action of these agents, and so enhanced secretion of
MCP-1 by innate immune cells (Fig. 3). These findings
indicate that the combination of PAR2-cAP and IFN-c is
apparently effective at enhancing secretion of MCP-1 by
human neutrophils and monocytes.
In our study, we were interested in which intracellular
signalling molecules were involved in the synergetic action
of PAR2-cAP and IFN-c on MCP-1 secretion by human
neutrophils and monocytes. Several signalling molecules
are known to be involved in the regulation of MCP-1
secretion. For example, a serine protease plasmin induces
MCP-1 expression in human monocytes via activation of
p38 kinase and JAK/signal transducer and activator of
transcription (STAT) pathways.28 Inhibitors of PI3 kinase
attenuate IFN-c-induced expression of MCP-1 in macro-
phages.29 Moreover, IFN-c-induced activation of PI3
kinase results in down-stream activation of PKCd.30 Con-
versely, PAR2 induces some effects via signalling cascades
involving PI3 kinase and PKCd.31 Altogether, these facts
led us to hypothesize that p38 kinase, PI3 kinase, PKCdand JAKs were involved in the synergistic effect of PAR2
agonist and IFN-c on MCP-1 secretion by human mono-
cytes and neutrophils. Indeed, our experiments with
inhibitors of these signalling molecules indicate that they
all participate in synergistic effects of PAR2-cAP and IFN-
c on MCP-1 secretion by human neutrophils (Fig. 4a).
Our results show that the enhanced effect of combined
PAR2-cAP and IFN-c treatment on MCP-1 secretion by
human neutrophils appears to be associated with the sig-
nalling pathway JAK–PI3K–PKCd (Fig. 6a). Possibly,
STAT1 could be the next participant in this pathway in
neutrophils. Interferon-c is known to activate the PI3K–
PKCd axis, and activated PKCd, in turn, affects STAT1
phosphorylation.30 The PKCd is involved in a dual mech-
anism by which it participates in regulating IFN-depen-
dent responses: (i) via STAT1 phosphorylation and (ii)
via p38 mitogen-activated protein (MAP) kinase activa-
tion.32 The results of our study strongly suggest that
PKCd is the upstream activator of p38 MAP kinase dur-
ing combined action of PAR2-cAP and IFN-c on MCP-1
secretion by human neutrophils. We found that PKCdinhibition abolished the effect of co-application of PAR2-
cAP and IFN-c on MCP-1 secretion, but that p38 MAP
kinase inhibitor just weakened MCP-1 secretion by
human neutrophils (Fig. 4a). In addition, we found that
the PI3K–PKCd axis plays a crucial role for MCP-1 secre-
tion by human neutrophils stimulated with PAR2-cAP
alone (Fig. 4b).
However, our study shows that human monocytes
demonstrate not only similarities, but also differences, in
the set of signalling molecules involved in the synergistic
effect of PAR2-cAP and IFN-c on MCP-1 secretion
(Fig. 6b). The inhibition of PI3K and JAKs reduced, but
did not abolish, the enhanced MCP-1 secretion, which
was induced after monocytes were treated with PAR2-cAP
together with IFN-c (Fig. 5a). This reduced level of
secreted MCP-1 was similar to the level reached after
monocytes were stimulated with PAR2-cAP alone
(Fig. 5a,b). These data indicate that PAR2-cAP effects on
IFN-γ IFN-γ
JAK JAK
PAR2-cAP PAR2-cAP
PI3K
PKC
p38 kinase
MCP-1 release MCP-1 release
?
PI3K ?
(a) (b)
Figure 6. Suggested signalling events involved in the effects induced
by combined treatment with proteinase-activated receptor-2 activat-
ing peptide (PAR2-cAP) and interferon-c (IFN-c) on monocyte
chemoattractant protein 1 (MCP-1) secretion by human neutrophils
(a) and monocytes (b). Experiments with selective inhibitors of sig-
nalling molecules led us to suggest that the phosphoinositide 3
kinase–protein kinase d (PI3K–PKCd) axis plays a key role in
changes of MCP-1 secretion induced by co-application of PAR2-cAP
and IFN-c in human neutrophils (a). However, the inhibition of the
PI3K molecule in human monocytes just weakened the effect of
combined PAR2-cAP and IFN-c application on MCP-1 secretion.
This indicates the involvement of the PI3K-independent pathway in
the effect of PAR2-cAP and IFN-c co-application on MCP-1 secre-
tion by monocytes (b).
� 2011 The Authors. Immunology � 2011 Blackwell Publishing Ltd, Immunology, 133, 329–339 337
PAR2 and anti-microbial defence of human leucocytes

MCP-1 secretion by human monocytes are mediated not
only via a signalling pathway involving PI3K activation,
but also via another pathway (Fig. 6b). Surprisingly, the
PKCd inhibitor rottlerin enhanced the effect of PAR2-cAP
and IFN-c on MCP-1 release by monocytes (Fig. 5a). Rot-
tlerin also synergized with PAR2-cAP in its action on
MCP-1 secretion (Fig. 5b). Moreover, rottlerin, when
applied alone, enhanced MCP-1 secretion by human
monocytes (Fig. 5c). Treatment with the p38 inhibitor
SB203580 did not influence the increased MCP-1 secre-
tion caused by either PAR2-cAP stimulation or combined
application of PAR2-cAP and IFN-c (Fig. 5a,b). The levels
of secreted MCP-1 after IFN-c stimulation were below
the threshold in the neutrophil samples and could there-
fore not be determined (Fig. 3a,b). The treatment of
human monocytes with IFN-c yielded no significant
changes in MCP-1 levels (Fig. 3c). Hence, the effects of
the inhibitors of signalling molecules at MCP-1 release
were not studied after IFN-c stimulation of human
monocytes and neutrophils. Altogether, the results of our
experiments allowed us to suggest a possible scheme of
signalling events involved in the enhancement of MCP-1
secretion triggered after combined stimulation of human
neutrophils and monocytes with PAR2-cAP and IFN-c(Fig. 6a,b).
In summary, our study demonstrates that PAR2 agonist
acting alone can enhance a bactericidal response of human
neutrophils and monocytes in vitro. However, PAR2
agonist is unable to synergize with IFN-c in the enhance-
ment of the bactericidal response. On the other hand,
PAR2 agonist and IFN-c do synergize to increase MCP-1
secretion by human neutrophils and monocytes during
the late phase (after 24 hr) of the inflammatory response.
This synergistic action of PAR2 agonist and IFN-c on
MCP-1 release apparently involves the activation of PI3
kinase and JAKs in neutrophils and monocytes.
Acknowledgements
The work was supported by grants from the IZKF Mun-
ster (Stei3/034/09), German Research Foundation (SFB
293-A14, STE 1014/2-2), CERIES (Paris), Weston Haven
foundation San Francisco USA (to M.S.), SFB 293 (S.L.),
IMF grant SH 120709 (University of Munster, Germany)
(to V.M.S.), IMF grant FE 110905 (University of Munster,
Germany) (to M.F.) as well as Canadian Institutes of
Health Research (Operating and Proteinases and Inflam-
mation Network grants to M.D.H.), Transregional Collab-
orative Research Centre 34 (C12) (to D.H. and J.R.) and
IMF grant HO 220912 (University of Munster, Germany)
(to D.H.). The position of V.M.S at the Department of
Nephrology and Hypertension, University of Magdeburg,
Germany was supported by German Research Foundation
SFB 854 (SFB854 TP1). The authors are deeply grateful to
Pamela Derish for excellent editorial work.
Disclosures
The authors have no financial conflict of interest.
References
1 Seeliger S, Derian CK, Vergnolle N et al. Proinflammatory role of proteinase-activated
receptor-2 in humans and mice during cutaneous inflammation in vivo. FASEB J 2003;
17:1871–85.
2 Shpacovitch V, Feld M, Hollenberg MD, Luger TA, Steinhoff M. Role of protease-acti-
vated receptors in inflammatory responses, innate and adaptive immunity. J Leukoc Biol
2008; 83:1309–22.
3 Ossovskaya VS, Bunnett NW. Protease-activated receptors: contribution to physiology
and disease. Physiol Rev 2004; 84:579–621.
4 Shpacovitch V, Feld M, Bunnett NW, Steinhoff M. Protease-activated receptors: novel
PARtners in innate immunity. Trends Immunol 2007; 28:541–50.
5 Corteling R, Bonneau O, Ferretti S, Ferretti M, Trifilieff A. Differential DNA synthesis
in response to activation of protease-activated receptors on cultured guinea-pig tracheal
smooth muscle cells. Naunyn Schmiedebergs Arch Pharmacol 2003; 368:10–6.
6 Hollenberg MD. Physiology and pathophysiology of proteinase-activated receptors
(PARs): proteinases as hormone-like signal messengers: PARs and more. J Pharmacol
Sci 2005; 97:8–13.
7 Tsanev R, Ivanov I. Immune interferon: properties and clinical applications. In:
Hollinger MA, ed. Pharmacology and Toxicology Series. Boca Raton, FL: CRC Press LLC,
2002.
8 Feld M, Shpacovitch VM, Ehrhardt C, Kerkhoff C, Hollenberg MD, Vergnolle N, Ludwig
S, Steinhoff M. Agonists of proteinase-activated receptor-2 enhance IFN-c-inducible
effects on human monocytes: role in influenza a infection. J Immunol 2008; 180:6903–10.
9 Khoufache K, LeBouder F, Morello E et al. Protective role for protease-activated recep-
tor-2 against influenza virus pathogenesis via an IFN-c-dependent pathway. J Immunol
2009; 182:7795–802.
10 Shpacovitch VM, Varga G, Strey A et al. Agonists of proteinase-activated receptor-2
modulate human neutrophil cytokine secretion, expression of cell adhesion molecules,
and migration within 3-D collagen lattices. J Leukoc Biol 2004; 76:388–98.
11 Serbina NV, Jia T, Hohl TM, Pamer EG. Monocyte-mediated defense against microbial
pathogens. Annu Rev Immunol 2008; 26:421–52.
12 Melgarejo E, Medina MA, Sanchez-Jimenez F, Urdiales JL. Monocyte chemoattractant
protein-1: a key mediator in inflammatory processes. Int J Biochem Cell Biol 2009;
41:998–1001.
13 Yoshimura T, Takahashi M. IFN-c-mediated survival enables human neutrophils to
produce MCP-1/CCL2 in response to activation by TLR ligands. J Immunol 2007;
179:1942–9.
14 Amano H, Morimoto K, Senba M et al. Essential contribution of monocyte chemoattr-
actant protein-1/C-C chemokine ligand-2 to resolution and repair processes in acute
bacterial pneumonia. J Immunol 2004; 172:398–409.
15 Yamamoto T. Pathogenic role of CCL2/MCP-1 in scleroderma. Front Biosci 2008;
13:2686–95.
16 Shpacovitch VM, Seeliger S, Huber-Lang M et al. Agonists of proteinase-activated
receptor-2 affect transendothelial migration and apoptosis of human neutrophils. Exp
Dermatol 2007; 16:799–806.
17 Zwadlo G, Brocker EB, Von Bassewitz DB, Feige U, Sorg C. A monoclonal antibody to
a differentiation antigen present on mature human macrophages and absent from
monocytes. J Immunol 1985; 134:1487–92.
18 Essin K, Salanova B, Kettritz R et al. Large-conductance calcium-activated potassium
channel activity is absent in human and mouse neutrophils and is not required for
innate immunity. Am J Physiol Cell Physiol 2007; 293:C45–54.
19 Hampton MB, Winterbourn CC. Methods for quantifying phagocytosis and bacterial
killing by human neutrophils. J Immunol Methods 1999; 232:15–22.
20 Perticarari S, Presani G, Banfi E. A new flow cytometric assay for the evaluation of
phagocytosis and the oxidative burst in whole blood. J Immunol Methods 1994;
170:117–24.
21 Van Amersfoort ES, Van Strijp JA. Evaluation of a flow cytometric fluorescence
quenching assay of phagocytosis of sensitized sheep erythrocytes by polymorphonuclear
leukocytes. Cytometry 1994; 17:294–301.
22 Horsburgh MJ, Aish JL, White IJ, Shaw L, Lithgow JK, Foster SJ. SigmaB modulates
virulence determinant expression and stress resistance: characterization of a func-
tional rsbU strain derived from Staphylococcus aureus 8325-4. J Bacteriol 2002;
184:5457–67.
23 Ehrchen J, Sindrilaru A, Grabbe S, Schonlau F, Schlesiger C, Sorg C, Scharffetter-Koch-
anek K, Sunderkotter C. Senescent BALB/c mice are able to develop resistance to Leish-
mania major infection. Infect Immun 2004; 72:5106–14.
338 � 2011 The Authors. Immunology � 2011 Blackwell Publishing Ltd, Immunology, 133, 329–339
V. M. Shpacovitch et al.

24 Moraes TJ, Martin R, Plumb JD et al. Role of PAR2 in murine pulmonary pseudomon-
al infection. Am J Physiol Lung Cell Mol Physiol 2008; 294:L368–77.
25 Ellis TN, Beaman BL. Interferon-c activation of polymorphonuclear neutrophil func-
tion. Immunology 2004; 112:2–12.
26 Fernandez-Boyanapalli R, McPhillips KA, Frasch SC et al. Impaired phagocytosis of
apoptotic cells by macrophages in chronic granulomatous disease is reversed by IFN-c
in a nitric oxide-dependent manner. J Immunol 2010; 185:4030–41.
27 Shpacovitch VM, Brzoska T, Buddenkotte J et al. Agonists of proteinase-activated
receptor 2 induce cytokine release and activation of nuclear transcription factor kappaB
in human dermal microvascular endothelial cells. J Invest Dermatol 2002; 118:380–5.
28 Burysek L, Syrovets T, Simmet T. The serine protease plasmin triggers expression of
MCP-1 and CD40 in human primary monocytes via activation of p38 MAPK and janus
kinase (JAK)/STAT signaling pathways. J Biol Chem 2002; 277:33509–17.
29 Harvey EJ, Li N, Ramji DP. Critical role for casein kinase 2 and phosphoinositide-3-
kinase in the interferon-c-induced expression of monocyte chemoattractant protein-1
and other key genes implicated in atherosclerosis. Arterioscler Thromb Vasc Biol 2007;
27:806–12.
30 Platanias LC. Mechanisms of type-I- and type-II-interferon-mediated signalling. Nat
Rev Immunol 2005; 5:375–86.
31 Van der Merwe JQ, Moreau F, MacNaughton WK. Protease-activated receptor-2 stimu-
lates intestinal epithelial chloride transport through activation of PLC and selective
PKC isoforms. Am J Physiol Gastrointest Liver Physiol 2009; 296:G1258–66.
32 Uddin S, Sassano A, Deb DK et al. Protein kinase C-delta (PKC-d) is activated by type
I interferons and mediates phosphorylation of Stat1 on serine 727. J Biol Chem 2002;
277:14408–16.
Supporting information
Additional Supporting Information may be found in the
online version of this article:
Figure S1. Influence of proteinase-activated receptor-2
(PAR2) agonist and interferon-c (IFN-c) stimulation on
phagocytic activity of human neutrophils against killed
FITC-conjugated Staphylococcus aureus. Isolated neutroph-
ils were pre-stimulated with 10)4M activating peptide
(cAP), 10)4M reverse peptide (cRP), or 100 ng/ml IFN-c,
or a combination of IFN-c and cAP or cRP for 2 hr.
Figure S2. Influence of proteinase-activated receptor-
2(PAR2) agonist and lipopolysaccharide (LPS) stimulation
on phagocytic activity of human neutrophils (a, b) and
monocytes (c, d). Isolated leucocytes were pre-stimulated
with 10)4M activating peptide (cAP), or 100 ng/ml LPS,
or a combination of LPS and cAP for 2 hr. Subsequently,
leucocytes were co-incubated for 30 min with bacteria in
the presence or absence of the stimuli mentioned.
Please note: Wiley-Blackwell are not responsible for the
content or functionality of any supporting materials sup-
plied by the authors. Any queries (other than missing
material) should be directed to the corresponding author
for the article.
� 2011 The Authors. Immunology � 2011 Blackwell Publishing Ltd, Immunology, 133, 329–339 339
PAR2 and anti-microbial defence of human leucocytes





























