Embed Size (px)
Citation preview

lable at ScienceDirect
Food Control 44 (2014) 49e57
Contents lists avai
Food Control
journal homepage: www.elsevier .com/locate/ foodcont
Screening of adhesive lactobacilli with antagonistic activity againstCampylobacter jejuni
Gang Wang a, Yu Zhao a, Fengwei Tian a, Xing Jin a, Haiqin Chen a, Xiaoming Liu a,Qiuxiang Zhang a, Jianxin Zhao a, Yongquan Chen a,b, Hao Zhang a, Wei Chen a,b,*
a State Key Laboratory of Food Science and Technology, School of Food Science and Technology, Jiangnan University, Wuxi 214122, PR Chinab Synergistic Innovation Center for Food Safety and Nutrition, Wuxi 214122, PR China
a r t i c l e i n f o
Article history:Received 1 October 2013Received in revised form16 March 2014Accepted 25 March 2014Available online 2 April 2014
Keywords:LactobacilliCampylobacter jejuniProbioticsAntagonistic activityAdhesion
* Corresponding author. State Key Laboratory of FSchool of Food Science and Technology, Jiangnan UChina. Tel./fax: þ86 510 85912155.
E-mail address: [email protected] (W. C
http://dx.doi.org/10.1016/j.foodcont.2014.03.0420956-7135/� 2014 Elsevier Ltd. All rights reserved.
a b s t r a c t
In the present study, 78 lactic acid bacteria (LAB) strains were screened for the antagonistic activityagainst Campylobacter jejuni and 15 LAB strains with high bactericidal capacity were selected for furtherstudy through the analysis of the inhibition zone of LAB spent culture supernatant on the growth ofC. jejuni. Among these strains, four isolates, Lactobacillus plantarum N8, N9, ZL5 and Lactobacillus caseiZL4 exhibited high adhesion ability to HT-29 cells. All the cell free supernatant (CFS) of these four strainscontained high concentration of organic acid and their inhibition effects against C. jejuni were pH sen-sitive. Furthermore, these four strains could strongly antagonize the adhesion and invasion of C. jejuni toHT-29 cells and showed good tolerance to artificial gastric and small intestinal juices. This study suggeststhat Lactobacillus strains N8, N9, ZL4 and ZL5 could be used as potential probiotics in food applicationsagainst C. jejuni infection.
� 2014 Elsevier Ltd. All rights reserved.
1. Introduction
Campylobacter infections usually lead to watery or bloodydiarrhea and abdominal pain and have been identified as theleading bacterial pathogen causing human enterocolitis in manycountries (Dasti, Tareen, Lugert, Zautner, & Gross, 2010). In somedeveloped countries, Campylobacter infections induce diarrhealdisease 7e12 times as frequently as infections with Salmonellaspecies, Shigella species, or Escherichia coli O157:H7 (Blaser, Wells,Feldman, Pollard, & Allen, 1983; Slutsker et al., 1997). Furthermore,Campylobacter infections might cause serious sequel e the Guil-laineBarré syndrome (GBS), a demyelinating disorder resulting inacute neuromuscular paralysis (Altekruse, Stern, Fields, &Swerdlow, 1999). Campylobacter jejuni is now recognized as oneof the main causes of bacterial food-borne disease in manydeveloped countries (Moore et al., 2005). At present, antibioticsare commonly used as the treatment of diseases caused byCampylobacter. However, the increasing incidence of antibiotic-resistant Campylobacter strains suggests that novel alternative
ood Science and Technology,niversity, Wuxi 214122, PR
hen).
approaches are necessary to be developed (Alfredson & Korolik,2007).
Lactobacilli are inhabitants of the gastrointestinal tract ofhealthy humans and play a pivotal role in normal gut function andmaintaining host health. Beneficial effects conferred by lactobacilliinclude inhibition of microbial pathogens, such as Salmonella(Bernet-Camard et al., 1997), E. coli (Mangell et al., 2002) and Lis-teria monocytogenes (de Waard, Garssen, Bokken, & Vos, 2002;Wang et al., 2014). Some studies have also evidenced the protec-tive effect of lactobacilli in preventing Campylobacter infections.Chaveerach, Lipman, and van Knapen (2004) reported that pro-biotic bacteria isolated from conventional chicken had potentialinhibitory activity against this pathogen, and the culturability ofCampylobacter was under the detection limit after 48 h of incuba-tion with Lactobacillus (P93). The efficacy of mixed probioticpreparations in inhibiting pathogen shedding and colonization ofC. jejuni in chicks has also been reported (Morishita, Aye, Harr,Cobb, & Clifford, 1997; Willis & Reid, 2008). Another studydescribed a 55% reduction in the invasion of C. jejuni into humanintestinal epithelial cells after treatment with Lactobacillus helve-ticus R0052, which also suggested that competitive exclusion couldcontribute to protection by adherent probiotics (Wine, Gareau,Johnson-Henry, & Sherman, 2009). Moreover, Wagner, Johnson,and Kurniasih Rubin (2009) reported that lactobacilli and bifido-bacteria could supplement the colonization resistance of a model

G. Wang et al. / Food Control 44 (2014) 49e5750
human microbiota against C. jejuni enteric persistence in immu-nodeficient and immunocompetent mice. However, the studiesdirectly focused on the ability of potential Lactobacillus strains toinhibit C. jejuni, especially the antagonistic activity of Lactobacillusstrains to reduce adhesion and invasion of C. jejuni in human in-testinal epithelial cells are still not enough.
In this study, to explore new potential probiotics of antimicro-bial activity against C. jejuni, 78 LAB were screened and fourLactobacillus strains with high adhesion capacity to human intes-tinal epithelial cells were selected for further study on their anti-microbial properties. With characterizing by in vitro assay, theseselected Lactobacillus strains may be considered as new anti-campylobacter agent and applied to food to provide an efficientmethod for inhibiting C. jejuni.
2. Materials and methods
2.1. Bacterial strains and culture conditions
78 LAB were isolated from fermented pickles, health infant fecesand fermented dairy products. Lactobacillus rhamnosus GG (LGG,ATCC 533103), C. jejuni NCTC 11168 (ATCC 700819), C. jejuni subsp.jejuni (ATCC 33291), C. jejuni subsp. jejuni (ATCC BAA-1153),L. monocytogenes (ATCC 19114), Enteroinvasive E. coli (ATCC43893), Enterococcus faecalis (ATCC 19433) and other 11 strainswere obtained from the Culture and Information Center of Indus-trial Microorganisms of China Universities, Jiangnan University(CICIM-CU). All Lactobacillus strains were cultured in deMan,Rogosa and Sharpe (MRS) broth at 37 �C for 20 h. C. jejuni strainswere incubated in Mueller-Hinton broth (Oxoid, UK) and culturedon Columbia blood agar base plates (Oxoid) supplemented with 6%sheep blood and C. jejuni selective supplement for 48 h at 37 �Cunder microaerophilic conditions (5% O2, 10% CO2, 85% N2) inanaerobic jars.
For adhesion and inhibition assays, Lactobacillus strains culturedfor 20 h were harvested and washed twice (5000 � g, 10 min, 4 �C)with 0.1 mol/L sterile phosphate-buffered saline (PBS, pH7.2), andfinally resuspended in RPMI-1640 medium (HyClone, USA) at aconcentration of 108 CFU/mL. C. jejuni strains cultured on Columbiablood agar base plates were harvested and washed twice with PBS(4000� g, 15min, 4 �C), the concentration of C. jejuni cultures (ATCC700819 only or equalmixture of ATCC700819, ATCC33291 andATCCBAA-1153) was adjusted to 108 CFU/mL with RPMI-1640 medium.
2.2. Growth inhibition assay
Todetect the antimicrobial activity of the selected LABonC. jejuniNCTC 11168 and other pathogenic bacteria, the oxford cup methodwas used. Campylobacter cultures (250 ml, 108 CFU/mL) were platedon charcoalcefoperazone e deoxycholate (CCDA) agar (Oxoid, UK).Other pathogenic bacteriawere plated on tryptone soy yeast extract(TSYE, L. monocytogenes), lysogeny broth (LB, Enteroinvasive E. coli)or MRS (E. faecalis). Then 100 mL cell free culture supernatant (CFS)from the 20 h LAB liquid culture (8000 � g, 20 min, 4 �C) whichfilteredwith 0.22 mmpore size filter was added into the Oxford cup.Sterilized MRS broth (pH 3.6) was used as a negative control andgentamicin (320 mg/mL; Sangon, Shanghai, China) was used as thepositive control. Plates were incubated for 48 h at 37 �C undermicroaerophilic conditions and the diameter of the inhibition zonearound each well was measured. Assays were performed in tripli-cate. The inhibition values were classified into four categories cor-responding to þþþ (>13 mm diameter), þþ (10e13 mmdiameter), þ (8e10 mm diameter) and � (<8 mm diameter).
Experiments were performed to examine the sensitivity ofLactobacillus strains CFS antimicrobial activity to heat (100 �C,
20 min), pH (pH was adjusted to 6.5) and enzymes. To assess theeffect of enzymes on the inhibitory effect of Lactobacillus strains CFSon C. jejuni growth, the antimicrobial activity of CFS were testedafter incubated at 37 �C for 1 h with trypsin (200 mg/mL; Sangon),proteinase K (200 mg/mL; Sangon) or catalase (1 mg/mL; Sangon).
The growth of C. jejuni co-cultured with Lactobacillus strains wasdetermined by the followingmethod according to Sgouras (Sgouraset al., 2004). The fresh C. jejuni NCTC 11168 cells (107 CFU/mL)suspended in antibiotic-free brain heart infusion broth (BHIB)containing 5% serum were incubated under microaerophilic con-ditions for 48 h at 37 �C in the presence of a 10% volume of livelactobacilli cells (107 CFU/mL). The viability of C. jejuni NCTC 11168was evaluated from the number of viable CFUs of C. jejuni NCTC11168 cultured as described above on C. jejuni-selective plates.
2.3. Organic acid analysis
The organic acids in Lactobacillus strains CFSwere determined byHPLC as previously described by Lin et al. (2009) with some modi-fications. Following protein precipitation with 50% H2SO4, super-natantwas filtered through a 0.22 mmpore size filter and diluted 10-fold with ddH2O2. 10 mL diluted supernatant were injected into a250 � 4.6 mm Synergi Hydro-RP column. Elutionwas performed at30 �C with 5% CH3OH and 0.05% H3PO4 (pH 2.3) at a flow rate of0.8 mL/min. Organic acids were determined by optical density (OD)measurements at 210 nm. Lactic acid, acetic acid, malic acid, citricacid, succinic acid, and fumaric acid were used as the standard.
2.4. Cell lines and adhesion assay
The HT-29 cell line was purchased from the Institute ofBiochemistry and Cell Biology, Shanghai Institutes for BiologicalSciences, Chinese Academy of Sciences, Shanghai, China. Cells werecultured in RPMI-1640 medium supplemented with 10% (v/v) fetalbovine serum (HyClone, USA) and 100 U/mL penicillinestrepto-mycin at 37 �C in an incubator with 5% CO2. For adhesion assays, HT-29 cells were seeded (4 � 105 cells/well) into six-well tissue cultureplates (Gibco, USA). The plates were cultured until monolayers ofcells were formed in each well. Prior to adherence assays, HT-29monolayers were washed twice with PBS. 1 mL lactobacilli sus-pension (108 CFU/mL in RPMI-1640) and 1mL antibiotic-free RPMI-1640 were added to each well and incubated at 37 �C in 5% CO2.After a 2-h incubation, cells werewashed twicewith PBS, fixedwithmethanol, Gram-stained and then examined microscopically underoil immersion. The adherence index was evaluated in 20 randommicroscopic fields of adhering bacteria per 100 cells. Adherenceassays were performed in triplicate.
2.5. Spectrophotometric hydrophobicity assays
Hydrophobicity interaction was carried out by the method ofKos et al. (2003) with modifications. Lactobacillus strains culturedfor 24 h were harvested and re-suspended in 0.1 mol/L KNO3 (pH6.2) to 108 CFU/mL. The absorbance of the cell suspension wasmeasured at 600 nm (A0). 1 mL of dimethylbenzene was added to3 mL of cell suspension. After a 10-min pre-incubation at roomtemperature, the two-phase system was mixed by vortexing for1 min. The aqueous phase was removed after 20 min of incubationat room temperature, and its absorbance at 600 nm (A1) wasmeasured. The percentage of bacterial adhesion to dime-thylbenzene (H%) was represented by the following formulae:
H% ¼ ð1� A1=A0Þ � 100%
G. Wang et al. / Food Control 44 (2014) 49e57 51
2.6. Inhibition of adhesion and invasion of C. jejuni
The inhibition of adhesion and invasion of C. jejuni by Lactoba-cillus strains were examined as previously described by Wine et al.(2009) and Zhang et al. (2010) with somemodifications. HT-29 cellswere seeded into six-well tissue culture plates (Gibco, USA) andcultured until monolayers of cells were formed in each well.Monolayers were washed twice with PBS before inhibition assays.For exclusion assays,1mL Lactobacillus strains suspension (108 CFU/mL) and 1 mL antibiotic-free cell culture medium were added toeach well and incubated for 1 h. Monolayers were washed threetimes with PBS to release unbound bacteria. Then 1 mL C. jejunisuspension (ATCC 700819 only or equal mixture of ATCC 700819,ATCC 33291 and ATCC BAA-1153, 108 CFU/mL) and 1 mL antibiotic-free cell culture mediumwere added to wells. Cells were incubatedfor a further 4 h. Wells containing C. jejuni alone served as controls.For competition assays, 1 mL Lactobacillus strains suspension(108 CFU/mL) and 1 mL C. jejuni suspension (ATCC 700819 only orequal mixture of ATCC 700819, ATCC 33291 and ATCC BAA-1153,108 CFU/mL) were added to each well simultaneously and incu-bated for 4 h. Wells containing C. jejuni alone served as controls. Fordisplacement assays, 1 mL C. jejuni suspension (ATCC 700819 onlyor equal mixture of ATCC 700819, ATCC 33291 and ATCC BAA-1153,108 CFU/mL) and 1 mL antibiotic-free cell culture medium wereadded to wells. After incubation for 4 h, monolayers were washedthree times with PBS to release unbound bacteria. Then 1 mL
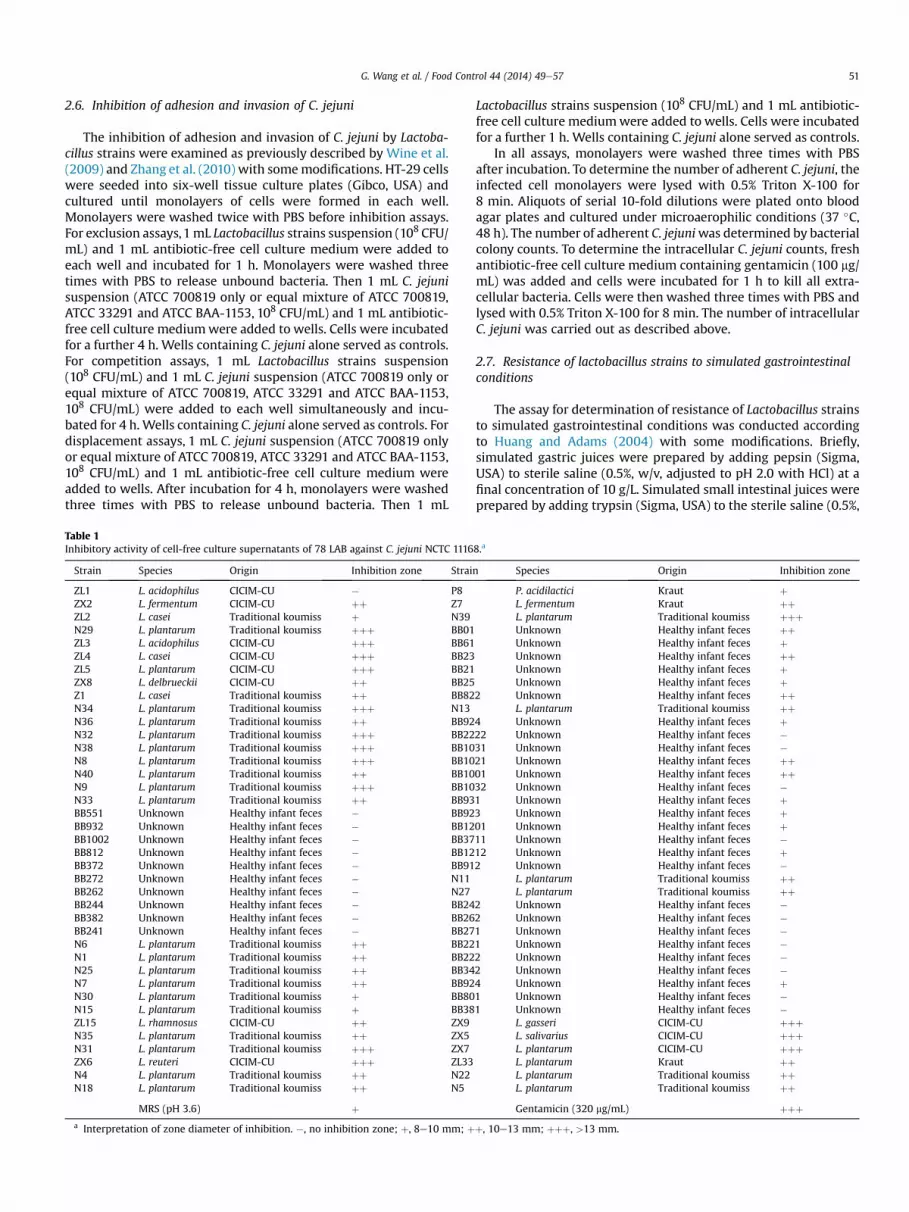
Table 1Inhibitory activity of cell-free culture supernatants of 78 LAB against C. jejuni NCTC 1116
Strain Species Origin Inhibition zone Strai
ZL1 L. acidophilus CICIM-CU � P8ZX2 L. fermentum CICIM-CU þþ Z7ZL2 L. casei Traditional koumiss þ N39N29 L. plantarum Traditional koumiss þþþ BB01ZL3 L. acidophilus CICIM-CU þþþ BB61ZL4 L. casei CICIM-CU þþþ BB23ZL5 L. plantarum CICIM-CU þþþ BB21ZX8 L. delbrueckii CICIM-CU þþ BB25Z1 L. casei Traditional koumiss þþ BB82N34 L. plantarum Traditional koumiss þþþ N13N36 L. plantarum Traditional koumiss þþ BB92N32 L. plantarum Traditional koumiss þþþ BB22N38 L. plantarum Traditional koumiss þþþ BB10N8 L. plantarum Traditional koumiss þþþ BB10N40 L. plantarum Traditional koumiss þþ BB10N9 L. plantarum Traditional koumiss þþþ BB10N33 L. plantarum Traditional koumiss þþ BB93BB551 Unknown Healthy infant feces � BB92BB932 Unknown Healthy infant feces � BB12BB1002 Unknown Healthy infant feces � BB37BB812 Unknown Healthy infant feces � BB12BB372 Unknown Healthy infant feces � BB91BB272 Unknown Healthy infant feces � N11BB262 Unknown Healthy infant feces � N27BB244 Unknown Healthy infant feces � BB24BB382 Unknown Healthy infant feces � BB26BB241 Unknown Healthy infant feces � BB27N6 L. plantarum Traditional koumiss þþ BB22N1 L. plantarum Traditional koumiss þþ BB22N25 L. plantarum Traditional koumiss þþ BB34N7 L. plantarum Traditional koumiss þþ BB92N30 L. plantarum Traditional koumiss þ BB80N15 L. plantarum Traditional koumiss þ BB38ZL15 L. rhamnosus CICIM-CU þþ ZX9N35 L. plantarum Traditional koumiss þþ ZX5N31 L. plantarum Traditional koumiss þþþ ZX7ZX6 L. reuteri CICIM-CU þþþ ZL33N4 L. plantarum Traditional koumiss þþ N22N18 L. plantarum Traditional koumiss þþ N5
MRS (pH 3.6) þa Interpretation of zone diameter of inhibition. �, no inhibition zone; þ, 8e10 mm; þ
Lactobacillus strains suspension (108 CFU/mL) and 1 mL antibiotic-free cell culture mediumwere added to wells. Cells were incubatedfor a further 1 h. Wells containing C. jejuni alone served as controls.
In all assays, monolayers were washed three times with PBSafter incubation. To determine the number of adherent C. jejuni, theinfected cell monolayers were lysed with 0.5% Triton X-100 for8 min. Aliquots of serial 10-fold dilutions were plated onto bloodagar plates and cultured under microaerophilic conditions (37 �C,48 h). The number of adherent C. jejuniwas determined by bacterialcolony counts. To determine the intracellular C. jejuni counts, freshantibiotic-free cell culture medium containing gentamicin (100 mg/mL) was added and cells were incubated for 1 h to kill all extra-cellular bacteria. Cells were then washed three times with PBS andlysed with 0.5% Triton X-100 for 8 min. The number of intracellularC. jejuni was carried out as described above.
2.7. Resistance of lactobacillus strains to simulated gastrointestinalconditions
The assay for determination of resistance of Lactobacillus strainsto simulated gastrointestinal conditions was conducted accordingto Huang and Adams (2004) with some modifications. Briefly,simulated gastric juices were prepared by adding pepsin (Sigma,USA) to sterile saline (0.5%, w/v, adjusted to pH 2.0 with HCl) at afinal concentration of 10 g/L. Simulated small intestinal juices wereprepared by adding trypsin (Sigma, USA) to the sterile saline (0.5%,
8.a
n Species Origin Inhibition zone
P. acidilactici Kraut þL. fermentum Kraut þþL. plantarum Traditional koumiss þþþUnknown Healthy infant feces þþUnknown Healthy infant feces þUnknown Healthy infant feces þþUnknown Healthy infant feces þUnknown Healthy infant feces þ
2 Unknown Healthy infant feces þþL. plantarum Traditional koumiss þþ
4 Unknown Healthy infant feces þ22 Unknown Healthy infant feces �31 Unknown Healthy infant feces �21 Unknown Healthy infant feces þþ01 Unknown Healthy infant feces þþ32 Unknown Healthy infant feces �1 Unknown Healthy infant feces þ3 Unknown Healthy infant feces þ01 Unknown Healthy infant feces þ11 Unknown Healthy infant feces �12 Unknown Healthy infant feces þ2 Unknown Healthy infant feces �
L. plantarum Traditional koumiss þþL. plantarum Traditional koumiss þþ
2 Unknown Healthy infant feces �2 Unknown Healthy infant feces �1 Unknown Healthy infant feces �1 Unknown Healthy infant feces �2 Unknown Healthy infant feces �2 Unknown Healthy infant feces �4 Unknown Healthy infant feces þ1 Unknown Healthy infant feces �1 Unknown Healthy infant feces �
L. gasseri CICIM-CU þþþL. salivarius CICIM-CU þþþL. plantarum CICIM-CU þþþL. plantarum Kraut þþL. plantarum Traditional koumiss þþL. plantarum Traditional koumiss þþGentamicin (320 mg/mL) þþþ
þ, 10e13 mm; þþþ, >13 mm.

Fig. 1. Adhesion of 15 Lactobacillus strains to HT-29 cells in vitro.
G. Wang et al. / Food Control 44 (2014) 49e5752
w/v, pH 6.8) at a final concentration of 10 g/L with 0.3% (w/v) bilesalts. Overnight cultures of the four selected Lactobacillus strainswere harvested and re-suspended in simulated gastric juices orsimulated small intestinal juices for a 2 h incubation at 37 �C. Viablecounts (log CFU/mL) of each strain were determined.
2.8. Statistical analysis
SPSS Inc. software (version 19.0; Chicago, IL, USA) was used toperform statistical analysis. Data were subjected to one-wayANOVA and, where appropriate, the StudenteNewmaneKeulstest was used for comparison of means. P < 0.05 was consideredstatistically significant (*).
3. Results
3.1. Effect of LAB-CFS on the growth of C. jejuni
Antimicrobial activities of 78 LAB were tested with oxford cupmethod. The results showed awide range of antimicrobial activitieswith many of the strains exhibiting weak or no inhibition effect
Fig. 2. Adhesion of different Lactobacillus strains to HT-29 cells as observed by Gram staininHT-29 cells. (B) Adhesion of L. plantarum N9 to HT-29 cells. (C) Adhesion of L. plantarum ZL5GG to HT-29 cells.
against C. jejuni NCTC 11168. Fifteen Lactobacillus strains (ZL3, ZL4,ZL5, N8, N9, N29, N31, N32, N34, N38, N39, ZX5, ZX6, ZX7, ZX9)exhibited higher inhibition capacity on C. jejuni NCTC 11168 thanthe other strains (Table 1). The diameters of the inhibition zones forthe 15 Lactobacillus strains ranged from 13 mm to 16 mm, showingthat these 15 strains strongly inhibited the growth of C. jejuni NCTC11168. Except for Lactobacillus acidophilus, Lactobacillus casei,Lactobacillus gasseri, Lactobacillus reuteri and Lactobacillus salivar-ius, the Lactobacillus strains associated with higher antimicrobialactivity against C. jejuni were predominantly Lactobacillusplantarum.
3.2. Adherence of Lactobacillus strains to HT-29 cells
Fifteen Lactobacillus strains selected with oxford cup methodwere further examined for the ability of adhering to HT-29 cells.LGG with high adherence ability (Tuomola, Ouwehand, &Salminen, 1999; Tuomola & Salminen, 1998) was used as a posi-tive control. The adherence indexes of these 15 strains are shownin Fig. 1. Compared to the other strains, L. casei ZL4 exhibited thehighest adherence capacity to HT-29 cells. Other three strains(L. plantarum N8, N9 and ZL5) were also found to adhere stronglyto HT-29 cell. Strains N34, ZX6 and ZX7 showed no obviousadhesion capacity. Because of their high ability to adhere toepithelial cell, L. plantarum N8, N9, ZL5 and L. casei ZL4 wereselected for further studies. Fig. 2 showed the adhesion of thesefour strains to HT-29 cell observed by Gram staining under a lightmicroscope. Similar to the pattern of adhesion by LGG, all the fourselected Lactobacillus strains showed substantially uniform dis-tribution on the cellular surface with a certain degree of clustersor bacterial aggregates.
3.3. Effect of LAB-CFS on the growth of pathogenic bacteria
Anti-pathogen activities of four selected Lactobacillus strainswith high adherence capacity were tested with oxford cup methodfurther. The results showed that these four Lactobacillus strainsperformed strong antimicrobial activities against C. jejuni (ATCC700819, ATCC 33291 or ATCC BAA-1153) but weak inhibition effectagainst the growth of L. monocytogenes, Enteroinvasive E. coli andE. faecalis (Table 2).
g under a light microscope (magnification 1000�). (A) Adhesion of L. plantarum N8 toto HT-29 cells. (D) Adhesion of L. casei ZL4 to HT-29 cells. (E) Adhesion of L. rhamnosus

Table 2Inhibitory activity of cell-free culture supernatants of four selected Lactobacillus strains against several pathogenic microorganisms.a
Species Medium(37 �C)
Inhibition zone of N8 Inhibition zone of N9 Inhibition zone of ZL4 Inhibition zone of ZL5
Campylobacter jejuni NCTC 11168 (ATCC 700819) CCDA þþþ þþþ þþþ þþþCampylobacter jejuni subsp. jejuni (ATCC 33291) CCDA þþþ þþ þþ þþþCampylobacter jejuni subsp. jejuni (ATCC BAA-1153) CCDA þþ þþ þþ þþListeria monocytogenes (ATCC 19114) TSYE � � � �Enteroinvasive Escherichia coli (ATCC 43893) LB � � � �Enterococcus faecalis (ATCC 19433) MRS � þ � þMRS (pH 3.6) þ/� Gentamicin (320 mg/mL) þþþ (E. faecalis�)
a Interpretation of zone diameter of inhibition. �, no inhibition zone; þ, 8e10 mm; þþ, 10e13 mm; þþþ, >13 mm.
G. Wang et al. / Food Control 44 (2014) 49e57 53
3.4. Hydrophobicity of Lactobacillus strains
Hydrophobicity interaction of four Lactobacillus strains wastested with spectrophotometric hydrophobicity assay. LGG hasbeen previously described as probiotics with high hydrophobicity.As showed in Table 3, the hydrophobicity index of four Lactobacillusstrains was range from 38.55% to 56.20%. L. casei ZL4 showed thehighest index (56.20 � 1.95%), followed by L. plantarum N9(51.97 � 2.37%), L .plantarum N8 (39.07 � 6.27%) and L. plantarumZL5 (38.55 � 5.88%) which possessed similar index.
3.5. Inhibition of C. jejuni growth by co-culturing Lactobacillusstrains
The viability of C. jejuni co-cultured with or without live Lacto-bacillus strains was measured. As showed in Fig. 3, although thenumber of pathogen elevated in all test after a 24-h incubation,compared to the negative control without co-cultured lactobacilli,the amount of C. jejuni NCTC 11168 co-cultured with all fourLactobacillus strains decreased obviously. Furthermore, comparedto the negative control, the pathogen was killed by one order ofmagnitude after the live Lactobacillus strains were added to thegrowing C. jejuni NCTC 11168 for 48 h.
3.6. Sensitivity to pH, enzymes and temperature
The sensitivity of lactobacilli CFS to different treatment wasmeasured (Fig. 4). When the pH of CFS was adjusted to 6.5, a sig-nificant reduction in diameter of the inhibition zone can be foundin all the four Lactobacillus strains. However, the antimicrobialsubstances in the CFS of all four Lactobacillus strains showed strongresistance to heat treatment. Although the antimicrobial activity ofall four Lactobacillus strains CFS treated with proteinase K, trypsinor catalase displayed decrease in some extent, most of the reduc-tion showed no significant different to the untreated CFS (P> 0.05).
3.7. Organic acid in Lactobacillus strains CFS
All four Lactobacillus strains CFS were engaged to analyze theorganic acid byHPLC (Table 4). Compared to the LGG,whose organicacid has been previously described, all four Lactobacillus strains CFS
Table 3Surface hydrophobicity of four Lactobacillus strains.
Strain Hydrophobicity index (%)
L. plantarum N8 39.07 � 6.27L. plantarum N9 51.97 � 2.37L. casei ZL4 56.20 � 1.95L. plantarum ZL5 38.55 � 5.88L. rhamnosus GG 65.27 � 5.38
contained higher concentration of lactic acid than LGG CFS (around220 mM). The concentration of acetic acid in four Lactobacillusstrains CFS was almost same with that in LGG CFS (around 70 mM).Less citric acid (6e10 mM) and succinic acid (15e21 mM) weredetected in lactobacillus strains CFS. No detectable malic acid andfumaric acid could be found in all four Lactobacillus strains CFS.
3.8. Inhibition of adhesion and invasion of C. jejuni by Lactobacillusstrains
The inhibition capacity of four adhesive Lactobacillus strains onadhesion and invasion of C. jejuni to HT-29 cells were examined.The adhesion or invasion of C. jejuni to HT-29 cells without inter-ruption by lactobacilli was assigned as 100%. As shown in Fig. 5,C. jejuni NCTC 11168 adhesion was reduced significantly by fourLactobacillus strains in all assays. The level of adhesion inhibitionobserved in exclusion assays was higher than that observed in thecompetition and displacement assays. In exclusion assays, L. caseiZL4 exhibited higher inhibition activity in adherence of C. jejuniNCTC 11168 compared with the other 3 strains, mediating a sig-nificant reduction (37%) in the adherence of C. jejuni NCTC 11168.Furthermore, L. plantarum ZL5 also showed marked antagonisticeffects, with the number of C. jejuni NCTC 11168 adhered toepithelial cells reduced to 44%. It was also observed that L. casei ZL4and L. plantarum ZL5 reduced the adherence of C. jejuni NCTC 11168more significantly than strains L. plantarum N8 and N9 in compe-tition and displacement assays.
As shown in Fig. 6, the invasion of HT-29 cells by C. jejuni NCTC11168 was also reduced by Lactobacillus strains in the exclusion,competition and displacement assays. However, the levels of inhi-bition varied between strains among these assays. Lactobacillusstrains showed more obvious antagonistic effects against C. jejuni
Fig. 3. Live cell counts of C. jejuni in the co-culture with Lactobacillus strains.

Fig. 4. Effect of different treatments on the antimicrobial activity of the four Lacto-bacillus strains CFS against C. jejuni. Asterisk (*) indicates means which were signifi-cantly different from the untreated CFS (P < 0.05).
Fig. 5. Effect of four adherent Lactobacillus strains on the adhesion of C. jejuni to HT-29cells. Each value shown was the mean � standard error of the three experiments.Means with no common letters differ significantly (P < 0.05).
G. Wang et al. / Food Control 44 (2014) 49e5754
NCTC 11168 in exclusion and displacement assays compared tocompetition assays. Exclusion assays demonstrated that the inva-sion rate of C. jejuni NCTC 11168 interrupted by L. plantarum N8 andN9 was significantly reduced by 44% and 64%, respectively,compared to C. jejuni NCTC 11168 invasion in the absence ofLactobacillus strains. L. casei ZL4 and L. plantarum ZL5 showed highinhibition activity in displacement assays, mediating a reduction inpathogen invasion to 51% and 50% respectively. L. casei ZL4 alsodisplayed excellent inhibition ability of C. jejuni NCTC 11168 inva-sion (49%) in exclusion assays.
As shown in Figs. 7 and 8, these four adhesive Lactobacillusstrains also displayed antagonistic effects against adhesion andinvasion of mixed C. jejuni (equal mixture of ATCC 700819, ATCC33291 and ATCC BAA-1153) to HT-29 cells. Compared with the ef-fect on C. jejuni NCTC 11168 only, these four Lactobacillus strainsperformed an obvious inhibitory activity against the adhesion ofmixed C. jejuni to HT-29 cells but a relatively weak inhibition effectagainst the invasion of pathogens to cells. Moreover, the differencesof antagonistic activity among these four Lactobacillus strainsagainst mixed C. jejuni were also not as obvious as that on C. jejuniNCTC 11168 only.
3.9. Resistance to simulated gastric and small intestinal juices
The effects of simulated gastric juices and small intestinal juiceson the viability of all four Lactobacillus strains were evaluated. Asshown in Table 5, viable counts of all four Lactobacillus strainsdropped about two orders of magnitude when strains wereexposed to simulated gastric juices for 2 h compared with that ofthe unexposed control. All the four Lactobacillus strains weresimulated small intestinal juices resistant because there was noremarkable different in viable counts of all the strains except for aslight drop (0.5 log CFU/mL) of L. plantarum N8.
Table 4Organic acids in four Lactobacillus strains CFS after cultured for 24 h in MRS.
Strain Lactic acid (mM) Acetic acid (mM) Citric acid (m
L. plantarum N8 227.8 � 1.3 71.4 � 1.0 9.3 � 1.5L. plantarum N9 214.2 � 0.8 68.4 � 0.9 10.8 � 1.7L. casei ZL4 219.1 � 1.0 67.6 � 2.1 8.6 � 0.8L. plantarum ZL5 220.3 � 1.4 73.9 � 0.7 6.1 � 0.8L. rhamnosus GG 149.2 � 1.3 69.6 � 1.3 13.8 � 1.0
NA indicates no detection.
4. Discussion
The burden of disease caused by C. jejuni remains considerable(Crushell, Harty, Sharif, & Bourke, 2004). In recent years, resistanceto antibiotics such as macrolides and quinolones has beenincreasing among Campylobacter (Moore et al., 2006; Young, Davis,& Dirita, 2007). Therefore, it is of great importance to develop novelalternative approaches for prevention and treatment of infectionsby this pathogen. As beneficial bacteria, the protective effect oflactobacillus strains on infection by pathogens has attracted a greatattention. The aim of this study was to screen antibacterial lacto-bacillus strains with potential capability of inhibiting the adhesionand invasion to human intestinal epithelial cells by C. jejuni.
In the present study, 15 lactobacillus strains whose CFS exhib-iting high antimicrobial activity against C. jejuni NCTC 11168 wasscreened from 78 LAB. All 78 LAB were isolated from fermenteddairy products, health infant feces and fermented pickles. It isnoteworthy that most of the strains which showed high inhibitoryeffect against C. jejuni NCTC 11168 were isolated from traditionalkoumiss. These koumiss products were collected from the pastoralarea in InnerMongolia, which indicates a rich resource of probioticsantagonizing C. jejuni in these places. On the contrary, strains iso-lated from healthy infant feces showed no obvious antagonisticactivity against C. jejuniNCTC 11168. These indicate the relationshipbetween the organic acid producing ability and antimicrobial ac-tivity of these strains isolated from fermented products. Manyspecies of microbe, such as L. plantarum, Bifidobacterium bifidum,L. acidophilus, L. gasseri, Lactobacillus sakei, Lactococcus lactis,Streptococcus faecium, L. helveticus, Klebsiella pheumoniae, Cit-robacter diversus and L. salivarius, have been found to be effective ininhibiting growth and intestinal colonization of C. jejuni
M) Succinic acid (mM) Malic acid (mM) Fumaric acid (mM)
15.7 � 0.9 NA NA17.2 � 2.3 NA NA21.1 � 1.3 NA NA19.7 � 1.5 NA NA22.8 � 3.7 NA NA

Fig. 6. Effect of four adherent Lactobacillus strains on the invasion of C. jejuni to HT-29cells. Each value shown was the mean � standard error of the three experiments.Means with no common letters differ significantly (P < 0.05).
Fig. 8. Effect of four adherent Lactobacillus strains on the invasion of mixed C. jejuni(ATCC 700819 þ ATCC 33291 þ ATCC BAA-1153) to HT-29 cells. Each value shown wasthe mean � standard error of the three experiments. Means with no common lettersdiffer significantly (P < 0.05).
G. Wang et al. / Food Control 44 (2014) 49e57 55
(Chaveerach et al., 2004; Fernandez, Boris, & Barbes, 2003; Fooks &Gibson, 2002; Jones, Hussein, Zagorec, Brightwell, & Tagg, 2008;Jones, Zagorec, Brightwell, & Tagg, 2009; Morishita et al., 1997;Schoeni & Doyle, 1992; Stern et al., 2006; Wagner et al., 2009;Wine et al., 2009). In this study, several strains (L. plantarum,L. acidophilus, L. casei, L. gasseri, L. reuteri and L. salivarius) whichproved to be effective in antagonizing C. jejuni were also isolated.However, L. plantarum were predominant species which can ex-press high inhibitory effects against C. jejuni in this study. This wasprobably due to its large proportion in fermentation strains ofkoumiss and high ability in producing organic acid. Considering theadhesion of lactobacillus strains to intestinal epithelial cells to be acrucially important factor for probiotics colonization and inhibitionof pathogen adhesion (Dunne et al., 2001), the adhesion index ofthese 15 selected lactobacillus strains is assessed and four strains(L. plantarum N8, N9, ZL5 and L. casei ZL4) showed high adherenceto HT-29 cells. L. plantarum N8 and N9 were obtained from tradi-tional koumiss while the original source of L. plantarum ZL5 andL. casei ZL4 was fermented pickles. The inhibitory effects againstC. jejuni of these adhesive strains were specific because all thesefour strains showed antagonistic activity against C. jejuni (ATCC
Fig. 7. Effect of four adherent Lactobacillus strains on the adhesion of mixed C. jejuni(ATCC 700819 þ ATCC 33291 þ ATCC BAA-1153) to HT-29 cells. Each value shown wasthe mean � standard error of the three experiments. Means with no common lettersdiffer significantly (P < 0.05).
700819, ATCC 33291 and ATCC BAA-1153) but no toL. monocytogenes, Enteroinvasive E. coli and E. faecalis. L. plantarumhas been proved to show high adhesion ability to Caco-2 cells andHT-29 cells in previous studies (Adlerberth et al., 1996; Lin et al.,2011). In this study, the adhesion indexes of these strains rangingfrom 9.4 to 12.5 bacteria/cell are similar to those of adhesiveL. plantarum (from 8.1 to 13.3 bacteria/cell) reported by Dunne et al.(2001). Surface properties of bifidobacterial strains are proved to berelevant to their adherence ability (Perez, Minnaard, Disalvo, & DeAntoni, 1998). It is found bacterial surface hydrophobicity appearedto be necessary for its adhesion to enterocyte-like cells andautoagglutination. Adhesive strains are highly hydrophobic, andthe degree of adherence is tightly dependent on the surface po-tential. In this study, all the four selected adhesive Lactobacillusstrains show relatively high hydrophobicity index ranging from38.55% to 56.20%, which indicates that hydrophobic interactionmight be involved in the adhesion of lactobacilli to HT-29 cells.
The Lactobacillus strains CFS is engaged to different treatment todeterminate the characterization of inhibitory substance. Anobvious reduction of inhibition is found when the pH of CFS isadjusted to neutral. However, the antibacterial activity of CFS isheat stable and insensitive to catalase, proteinase K and trypsin.This result indicates that the antibacterial substance should not tobe bacteriocin. With further HPLC assay, high proportion of lacticacid is found in CFS, which suggests a possible explanation to thecharacter mentioned above. It is reported that undissociatedorganic acid in low pH may be beneficial process for diffusion intothe bacteria cell and cause acidification of the cytoplasm
Table 5Effects of simulated gastric juices and simulated small intestinal juices on theviability of four selected Lactobacillus strains.a
Strain Viable count (log CFU/ml)
Untreated Simulatedgastric, 2 h
Simulatedintestinal, 2 h
L. plantarum N8 9.25 � 0.04 7.12 � 0.08* 8.75 � 0.10*L. plantarum N9 9.21 � 0.08 7.45 � 0.07* 9.15 � 0.04L. casei ZL4 9.20 � 0.02 7.23 � 0.05* 9.18 � 0.04L. plantarum ZL5 9.23 � 0.01 7.05 � 0.04* 9.20 � 0.02
Asterisk (*) indicates means which were significantly different from the controlvalue (P < 0.05).
a Experimental conditions were described in the text. Each value shown was themean � standard error of the three experiments.

G. Wang et al. / Food Control 44 (2014) 49e5756
(Chaveerach, Keuzenkamp, Urlings, Lipman, & van Knapen, 2002).In addition, Makras et al. (2006) also prove that the ability ofLactobacillus strains to inhabit Salmonella typhimurium SL1344 issolely due to the production of lactic acid. Therefore, the antibac-terial activity of these four Lactobacillus strains may probablydepend on the low pH and the production of metabolites, such aslactic acid and antibiotic-like substances. Furthermore, besides CFS,co-culturing bacteria also show significant inhibition of C. jejuniNCTC 11168 growth with one order of magnitude reduction ofviable cells after a 48 h incubation, which indicates excellentbactericidal capacity of these four adhesive Lactobacillus strains.
One important role of probiotics is protecting of the hostgastrointestinal micro-environment. Several reports haveconfirmed that lactobacilli effectively inhibit the cell associationand invasion by pathogens (Bernet, Brassart, Neeser, & Servin,1994;Gopal, Prasad, Smart, & Gill, 2001). The competitive exclusion ofpathogen adhesion to host cells by adherent LAB cells has beensuggested to be important factor enabling LAB to inhibit pathogensinfection. However, information on the ability of lactobacilliinhibiting C. jejuni adherence and invasion in human intestinal cellsis scarce. In this study, examination of colonization resistanceprovided by four selected adhesive Lactobacillus strains againstC. jejuni shows that C. jejuni NCTC 11168 adhesion to HT-29 cells isreduced in all cases in competition, exclusion and displacementassays. It is noteworthy that, in exclusion assay, these four Lacto-bacillus strains, especially L. casei ZL4 and L. plantarum ZL5, causemore significant antagonistic activity against C. jejuni NCTC 11168than that in competition and displacement assays. This indicatesthat exclusion of pathogen adhesion to host intestinal cells byadherent Lactobacillus strains may be a predominant mechanismfor adhesive lactobacilli to inhibit adhesion of C. jejuni. We alsodemonstrate that all four Lactobacillus strains inhibit the invasion toHT-29 cells by C. jejuni NCTC 11168, despite the differences in in-vasion inhibitory levels among Lactobacillus strains in competition,exclusion and displacement assays. L. plantarum N9 shows thegreatest inhibitory activity to C. jejuni NCTC 11168 in exclusionassay, while L. casei ZL4 and L. plantarum ZL5 exhibit ideal antag-onistic activity against C. jejuniNCTC 11168 in both competition anddisplacement assays. As can be seen from the results, incubation ofHT-29 cells with Lactobacillus strains prior to infectionwith C. jejuniNCTC 11168 significantly inhibits the invasion of C. jejuni NCTC11168, whereas the inhibition level decreases when HT-29 cells areincubated with C. jejuni NCTC 11168 preceded or simultaneous withthe incubation of Lactobacillus strains. Several mechanisms ofpathogen adhesion and invasion inhibition by lactobacilli havebeen suggested. Steric hindrance of human enterocytic pathogenreceptors by whole-cell probiotics rather than specific blockade ofreceptors is postulated as the reason for pathogen adhesion andinvasion inhibition (Coconnier et al., 1993). S-layer proteins ofLactobacillus crispatus ZJ001 is also proved to be involved in theadhesion and competitive exclusion of pathogens to HeLa cells(Chen et al., 2007). In addition, organic acids of lactic acid bacteriaare demonstrated to be able to inhibit Helicobacter pylori adhesionand invasion of AGS cells (Lin et al., 2009). In present study, thedifferences between results of anti-invasion assays and anti-adhesion assays appear to be related to the more complex inva-sion mechanism of C. jejuni NCTC 11168 or the different compo-nents in the surface-layer of these Lactobacillus strains. L. casei ZL4exhibits the highest C. jejuni NCTC 11168 adhesion exclusion ca-pacity whereas L. plantarum N9 shows most efficient exclusion ef-fect on pathogen invasion. Taking the fact that adherent C. jejuniNCTC 11168 in anti-adhesion assay include the pathogen on the cellsurface and inside the cell into consideration, the mechanism ofadhesion exclusion by L. casei ZL4 to C. jejuni NCTC 11168 is mostlybecause of nonspecific steric hindrance caused by bacterial cells.
C. jejuniNCTC 11168which touch the surface of HT-29 cells from thegap between the probiotics can invade intestinal cells easily. On thecontrary, L. plantarum N9, which exhibits no prominent effect onpathogen adhesion exclusion, displays the most efficient invasionexclusion effect on C. jejuni NCTC 11168. It suggests that specificblockade of receptors is probably involved in the inhibition onC. jejuniNCTC 11168 invasion by L. plantarumN9. In addition, L. caseiZL4 and L. plantarum ZL5 show high inhibition level to C. jejuniNCTC 11168 in invasion displacement assay but relatively weakeffects on adhesion displacement. This indicates that these twoLactobacillus strains may participate in regulating the signalingpathway of pathogen internalization or exocytosis. Similar to theeffect on C. jejuni NCTC 11168 only, these four adhesive Lactobacillusstrains also displayed antagonistic effects against adhesion andinvasion of mixed C. jejuni (equal mixture of ATCC 700819, ATCC33291 and ATCC BAA-1153) to HT-29 cells. However, although thesefour Lactobacillus strains still performed an obvious inhibitory ac-tivity against the adhesion of mixed C. jejuni to HT-29 cells, theeffects on the invasion of pathogens to cells were relatively weak.Moreover, the differences of antagonistic activity among these fourLactobacillus strains against mixed C. jejuniwere also not as obviousas that on C. jejuni NCTC 11168 only. This is probably because oftheir relatively weak antagonistic activity against C. jejuni subsp.jejuni (ATCC BAA-1153). In addition, different strain of C. jejunimayengage different mechanisms to adhere or invade to human cells.All above make the inhibitory effect of Lactobacillus strains againstmixed pathogens to be not as obvious as that on single C. jejuniNCTC 11168. The mechanisms by which lactobacilli inhibit path-ogen adhesion and invasion of human cells are still not fully un-derstood. Further studies are required to characterize thisantagonistic effect caused by different strains.
The pH value in human stomach is very low ranging from 1.0 to4.0. Besides low pH a variety of intestinal enzymes and bile have awide range of effects on the viability of probiotics. In order tosurvive in and colonize the gastrointestinal tract to perform theirbeneficial effect on health in the intestine, excepting for highadhesion capability to intestinal cell surface probiotic bacteriashould express high tolerance to acid and bile. It is reported thatlactobacilli show consistent tolerance to bile and pH 3, whereasonly a few can survive pH 2.0 for long time incubation (Charteris,Kelly, Morelli, & Collins, 1998; Mishra & Prasad, 2005; Pan, Chen,Wu, Tang, & Zhao, 2009). In our study, although the viable cellcount of four strains drop about two orders of magnitude in arti-ficial gastric juice (pH 2.0) after 2 h of culture, the number of viablecells still maintained at a high level (107 CFU/mL). After a 2 h in-cubation in artificial intestinal juice, number of viable lactic acidbacteria is not changed except for a slight drop (0.5 log CFU/mL) ofL. plantarum N8. Therefore, the selected strains could overcome thebarriers in the gastrointestinal tract and have potential for appli-cations in probiotic foods.
5. Conclusion
In this study, four adhesive Lactobacillus strains L. plantarum N8,N9, ZL5 and L. casei ZL4 selected from 78 LAB were capable ofexerting significant antagonistic activity against C. jejuni in vitro andpromoted effective inhibition of adhesion and invasion of HT-29 cells by C. jejuni. The bactericidal capacity of these four Lacto-bacillus strains may probably be relative to the low pH and theproduction of metabolites, such as lactic acid and antibiotic-likesubstances. Furthermore, all four strains exhibited significantresistance to simulated gastrointestinal conditions. Therefore,further research is required to evaluate the antagonistic activity ofthese Lactobacillus strains against C. jejuni in vivo and investigate thepotential for use as probiotics for prevention of C. jejuni infections.

G. Wang et al. / Food Control 44 (2014) 49e57 57
Acknowledgments
This work was supported by the National Natural ScienceFoundation of China (No. 31301407), the Key Projects in theNationalScience & Technology Pillar Program during the Twelfth Five-YearPlan Period (No. 2013BAD18B01, 2013BAD18B02, 2012BAD28B07,2012BAD28B08), the 111 project B07029, and the Priority AcademicProgram Development of Jiangsu Higher Education Institutions.
References
Adlerberth, I., Ahrne, S., Johansson, M. L., Molin, G., Hanson, L. A., & Wold, A. E.(1996). A mannose-specific adherence mechanism in Lactobacillus plantarumconferring binding to the human colonic cell line HT-29. Applied and Environ-mental Microbiology, 62(7), 2244e2251.
Alfredson, D. A., & Korolik, V. (2007). Antibiotic resistance and resistance mecha-nisms in Campylobacter jejuni and Campylobacter coli. FEMS Microbiology Letters,277(2), 123e132.
Altekruse, S. F., Stern, N. J., Fields, P. I., & Swerdlow, D. L. (1999). Campylobacter jejunie an emerging foodborne pathogen. Emerging Infectious Diseases, 5(1), 28e35.
Bernet, M. F., Brassart, D., Neeser, J. R., & Servin, A. L. (1994). Lactobacillus acidophilusLA 1 binds to cultured human intestinal cell lines and inhibits cell attachmentand cell invasion by enterovirulent bacteria. Gut, 35(4), 483e489.
Bernet-Camard, M. F., Lievin, V., Brassart, D., Neeser, J. R., Servin, A. L., & Hudault, S.(1997). The human Lactobacillus acidophilus strain LA1 secretes a non-bacteriocin antibacterial substance(s) active in vitro and in vivo. Applied andEnvironmental Microbiology, 63(7), 2747e2753.
Blaser, M. J., Wells, J. G., Feldman, R. A., Pollard, R. A., & Allen, J. R. (1983).Campylobacter enteritis in the United States. A multicenter study. Annals ofInternal Medicine, 98(3), 360e365.
Charteris, W. P., Kelly, P. M., Morelli, L., & Collins, J. K. (1998). Development andapplication of an in vitro methodology to determine the transit tolerance ofpotentially probiotic Lactobacillus and Bifidobacterium species in the upperhuman gastrointestinal tract. Journal of Applied Microbiology, 84(5), 759e768.
Chaveerach, P., Keuzenkamp, D. A., Urlings, H. A., Lipman, L. J., & van Knapen, F.(2002). In vitro study on the effect of organic acids on Campylobacter jejuni/colipopulations in mixtures of water and feed. Poultry Science, 81(5), 621e628.
Chaveerach, P., Lipman, L. J., & van Knapen, F. (2004). Antagonistic activities ofseveral bacteria on in vitro growth of 10 strains of Campylobacter jejuni/coli.International Journal of Food Microbiology, 90(1), 43e50.
Chen, X., Xu, J., Shuai, J., Chen, J., Zhang, Z., & Fang, W. (2007). The S-layer proteins ofLactobacillus crispatus strain ZJ001 is responsible for competitive exclusionagainst Escherichia coli O157:H7 and Salmonella typhimurium. InternationalJournal of Food Microbiology, 115(3), 307e312.
Coconnier, M. H., Bernet, M. F., Kerneis, S., Chauviere, G., Fourniat, J., & Servin, A. L.(1993). Inhibition of adhesion of enteroinvasive pathogens to human intestinalCaco-2 cells by Lactobacillus acidophilus strain LB decreases bacterial invasion.FEMS Microbiology Letters, 110(3), 299e305.
Crushell, E., Harty, S., Sharif, F., & Bourke, B. (2004). Enteric campylobacter: purgingits secrets? Pediatric Research, 55(1), 3e12.
Dasti, J. I., Tareen, A. M., Lugert, R., Zautner, A. E., & Gross, U. (2010). Campylobacterjejuni: a brief overview on pathogenicity-associated factors and disease-mediating mechanisms. International Journal of Medical Microbiology, 300(4),205e211.
Dunne, C., O’Mahony, L., Murphy, L., Thornton, G., Morrissey, D., O’Halloran, S., et al.(2001). In vitro selection criteria for probiotic bacteria of human origin: cor-relationwith in vivo findings. American Journal of Clinical Nutrition, 73(2 Suppl.),386Se392S.
Fernandez, M. F., Boris, S., & Barbes, C. (2003). Probiotic properties of human lac-tobacilli strains to be used in the gastrointestinal tract. Journal of AppliedMicrobiology, 94(3), 449e455.
Fooks, L. J., & Gibson, G. R. (2002). In vitro investigations of the effect of probioticsand prebiotics on selected human intestinal pathogens. FEMS MicrobiologyEcology, 39(1), 67e75.
Gopal, P. K., Prasad, J., Smart, J., & Gill, H. S. (2001). In vitro adherence properties ofLactobacillus rhamnosus DR20 and Bifidobacterium lactis DR10 strains and theirantagonistic activity against an enterotoxigenic Escherichia coli. InternationalJournal of Food Microbiology, 67(3), 207e216.
Huang, Y., & Adams, M. C. (2004). In vitro assessment of the upper gastrointestinaltolerance of potential probiotic dairy propionibacteria. International Journal ofFood Microbiology, 91(3), 253e260.
Jones, R. J., Hussein, H. M., Zagorec, M., Brightwell, G., & Tagg, J. R. (2008). Isolationof lactic acid bacteria with inhibitory activity against pathogens and spoilageorganisms associated with fresh meat. Food Microbiology, 25(2), 228e234.
Jones, R. J., Zagorec, M., Brightwell, G., & Tagg, J. R. (2009). Inhibition by Lactobacillussakei of other species in the flora of vacuum packaged raw meats during pro-longed storage. Food Microbiology, 26(8), 876e881.
Kos, B., Suskovic, J., Vukovic, S., Simpraga, M., Frece, J., & Matosic, S. (2003).Adhesion and aggregation ability of probiotic strain Lactobacillus acidophilusM92. Journal of Applied Microbiology, 94(6), 981e987.
Lin, W. H., Lin, C. K., Sheu, S. J., Hwang, C. F., Ye, W. T., Hwang, W. Z., et al. (2009).Antagonistic activity of spent culture supernatants of lactic acid bacteria againstHelicobacter pylori growth and infection in human gastric epithelial AGS cells.Journal of Food Science, 74(6), M225eM230.
Lin, W. H., Wu, C. R., Fang, T. J., Lee, M. S., Lin, K. L., Chen, H. C., et al. (2011). Adherentproperties and macrophage activation ability of 3 strains of lactic acid bacteria.Journal of Food Science, 76(1), M1eM7.
Makras, L., Triantafyllou, V., Fayol-Messaoudi, D., Adriany, T., Zoumpopoulou, G.,Tsakalidou, E., et al. (2006). Kinetic analysis of the antibacterial activity ofprobiotic lactobacilli towards Salmonella enterica serovar Typhimurium revealsa role for lactic acid and other inhibitory compounds. Research in Microbiology,157(3), 241e247.
Mangell, P., Nejdfors, P., Wang, M., Ahrne, S., Westrom, B., Thorlacius, H., et al.(2002). Lactobacillus plantarum 299v inhibits Escherichia coli-induced intestinalpermeability. Digestive Diseases and Sciences, 47(3), 511e516.
Mishra, V., & Prasad, D. N. (2005). Application of in vitro methods for selection ofLactobacillus casei strains as potential probiotics. International Journal of FoodMicrobiology, 103(1), 109e115.
Moore, J. E., Barton, M. D., Blair, I. S., Corcoran, D., Dooley, J. S., Fanning, S., et al.(2006). The epidemiology of antibiotic resistance in Campylobacter. Microbesand Infection, 8(7), 1955e1966.
Moore, J. E., Corcoran, D., Dooley, J. S., Fanning, S., Lucey, B., Matsuda, M., et al.(2005). Campylobacter. Veterinary Research, 36(3), 351e382.
Morishita, T. Y., Aye, P. P., Harr, B. S., Cobb, C. W., & Clifford, J. R. (1997). Evaluation ofan avian-specific probiotic to reduce the colonization and shedding ofCampylobacter jejuni in broilers. Avian Diseases, 41(4), 850e855.
Pan, X., Chen, F., Wu, T., Tang, H., & Zhao, Z. (2009). The acid, bile tolerance and anti-microbial property of Lactobacillus acidophilus NIT. Food Control, 20(6), 598e602.
Perez, P. F., Minnaard, Y., Disalvo, E. A., & De Antoni, G. L. (1998). Surface propertiesof bifidobacterial strains of human origin. Applied and Environmental Microbi-ology, 64(1), 21e26.
Schoeni, J. L., & Doyle, M. P. (1992). Reduction of Campylobacter jejuni colonization ofchicks by cecum-colonizing bacteria producing anti-C. jejuni metabolites.Applied and Environmental Microbiology, 58(2), 664e670.
Sgouras, D., Maragkoudakis, P., Petraki, K., Martinez-Gonzalez, B., Eriotou, E.,Michopoulos, S., et al. (2004). In vitro and in vivo inhibition of Helicobacterpylori by Lactobacillus casei strain Shirota. Applied and Environmental Microbi-ology, 70(1), 518e526.
Slutsker, L., Ries, A. A., Greene, K. D., Wells, J. G., Hutwagner, L., & Griffin, P. M.(1997). Escherichia coli O157:H7 diarrhea in the United States: clinical andepidemiologic features. Annals of Internal Medicine, 126(7), 505e513.
Stern, N. J., Svetoch, E. A., Eruslanov, B. V., Perelygin, V. V., Mitsevich, E. V.,Mitsevich, I. P., et al. (2006). Isolation of a Lactobacillus salivarius strain andpurification of its bacteriocin, which is inhibitory to Campylobacter jejuni in thechicken gastrointestinal system. Antimicrobial Agents and Chemotherapy, 50(9),3111e3116.
Tuomola, E. M., Ouwehand, A. C., & Salminen, S. J. (1999). The effect of probioticbacteria on the adhesion of pathogens to human intestinal mucus. FEMSImmunology and Medical Microbiology, 26(2), 137e142.
Tuomola, E. M., & Salminen, S. J. (1998). Adhesion of some probiotic and dairyLactobacillus strains to Caco-2 cell cultures. International Journal of FoodMicrobiology, 41(1), 45e51.
de Waard, R., Garssen, J., Bokken, G. C., & Vos, J. G. (2002). Antagonistic activity ofLactobacillus casei strain Shirota against gastrointestinal Listeria monocytogenesinfection in rats. International Journal of Food Microbiology, 73(1), 93e100.
Wagner, R. D., Johnson, S. J., & Kurniasih Rubin, D. (2009). Probiotic bacteria areantagonistic to Salmonella enterica and Campylobacter jejuni and influencehost lymphocyte responses in human microbiota-associated immunodeficientand immunocompetent mice. Molecular Nutrition & Food Research, 53(3),377e388.
Wang, G., Ning, J., Zhao, J., Hang, F., Tian, F., Zhao, J., et al. (2014). Partial charac-terisation of an anti-listeria substance produced by Pediococcus acidilactici P9.International Dairy Journal, 34(2), 275e279.
Willis, W. L., & Reid, L. (2008). Investigating the effects of dietary probiotic feedingregimens on broiler chicken production and Campylobacter jejuni presence.Poultry Science, 87(4), 606e611.
Wine, E., Gareau, M. G., Johnson-Henry, K., & Sherman, P. M. (2009). Strain-specific probiotic (Lactobacillus helveticus) inhibition of Campylobacter jejuniinvasion of human intestinal epithelial cells. FEMS Microbiology Letters,300(1), 146e152.
Young, K. T., Davis, L. M., & Dirita, V. J. (2007). Campylobacter jejuni: molecularbiology and pathogenesis. Nature Reviews Microbiology, 5(9), 665e679.
Zhang, Y. C., Zhang, L. W., Tuo, Y. F., Guo, C. F., Yi, H. X., Li, J. Y., et al. (2010). Inhibitionof Shigella sonnei adherence to HT-29 cells by lactobacilli from Chinese fer-mented food and preliminary characterization of S-layer protein involvement.Research in Microbiology, 161(8), 667e672.





























