Embed Size (px)
Citation preview

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, researchlibraries, and research funders in the common goal of maximizing access to critical research.
Seasonal, meteorological, and microhabitat effects on breeding success andoffspring phenotype in the barn swallow, Hirundo rusticaAuthor(s): Roberto Ambrosini, Raffaella Paola Ferrari, Roberta Martinelli, Maria Romano, and NicolaSainoSource: Ecoscience, 13(3):298-307. 2006.Published By: Centre d'études nordiques, Université LavalDOI: http://dx.doi.org/10.2980/i1195-6860-13-3-298.1URL: http://www.bioone.org/doi/full/10.2980/i1195-6860-13-3-298.1
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological, andenvironmental sciences. BioOne provides a sustainable online platform for over 170 journals and books publishedby nonprofit societies, associations, museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance ofBioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiriesor rights and permissions requests should be directed to the individual publisher as copyright holder.

13 (3): 298-xxx (2006)13 (3): 298-307 (2006)
Temporal and spatial variation in ecological conditions can influence reproductive success of organisms (Lack, 1954; McCarty & Winkler, 1999). In birds, the reproduc-tive performance and quality of offspring vary during the breeding season, and part of this variation can be attributed to seasonal changes in extrinsic factors such as availabil-ity of limiting resources (Blancher & Robertson, 1987; McCarty & Winkler, 1999) and weather conditions (Buse et al., 1999; McCarty & Winkler, 1999; Takagi, 2001). General predictions of optimal time for breeding propose
that single-brooded species will breed when conditions are most favourable, while multi-brooded species will make an early first breeding attempt to enhance the quality of the offspring from the second or later broods (Smith, Källander & Nilsson, 1989; Crick, Gibbons & Magrath, 1993). These predictions do not take into account that seasonal variation in ecological conditions may differentially affect compo-nents of parental fitness. For example, the number of fledg-lings may be influenced by food availability, while immune function, which may affect survival prospects, may be affected by specific components of diet (Lochmiller, Vestey & Boren, 1993). In addition, the availability of nutrients critical to immunity may vary seasonally according to a pat-tern different from that of the total amount of food. Finally,
Seasonal, meteorological, and microhabitat effects on breeding success and offspring phenotype in the barn swallow, Hirundo rustica1
Roberto AMBROSINI2, Raffaella Paola FERRARI, Roberta MARTINELLI, Maria ROMANO & Nicola SAINO, Dipartimento di Biologia, Università degli Studi di Milano, via Celoria 26, I-20133 Milano, Italy, e-mail: [email protected]
Abstract: Breeding performance and offspring quality can be affected by temporal and spatial variation in ecological conditions. We analyzed the concomitant effects of timing of breeding, ambient temperature at different stages of the breeding cycle, and microhabitat conditions (presence or absence of livestock farming) on breeding success, morphology, and immunity of the offspring in an aerially insectivorous passerine bird, the barn swallow (Hirundo rustica). A seasonal decline in breeding success was observed, while maximum offspring phenotypic values of different characters were attained at different times in the breeding season, depending on presence or absence of livestock farming. In addition, environmental temperature at different stages of the breeding cycle (pre-incubation, incubation, and nestling stage) of individual pairs affected all the components of nestling phenotypic quality. The results of this correlational study suggest that parents are faced with complex decisions about optimal timing of breeding and microhabitat choice. In addition, different ecological factors seem to interact in influencing breeding success and offspring quality in the barn swallow. Natural and anthropogenic changes in ecological conditions, including ambient temperature and distribution of livestock, may therefore affect distribution and productivity of a population of a declining passerine species.Keywords: Hirundo rustica, immunocompetence, livestock farming, nestling quality, seasonal trends, temperature.
Résumé : Des variations temporelles et spatiales dans les conditions écologiques peuvent avoir un effet sur le succès reproducteur et la qualité des jeunes. Nous avons analysé les effets concomitants du choix du moment de la reproduction, de la température ambiante à différentes étapes du cycle reproducteur et des conditions du micro-habitat (présence ou absence d’élevage d’animaux de ferme) sur le succès reproducteur, la morphologie et l’immunité des jeunes chez un passereau insectivore, l’hirondelle rustique (Hirundo rustica). Un déclin saisonnier du succès reproducteur a été observé alors que les valeurs phénotypiques maximales de différents caractères des jeunes ont été atteintes à différents moments de la saison de reproduction, selon la présence ou l’absence d’élevage d’animaux de ferme. De plus, la température ambiante à différentes étapes du cycle reproducteur (pré-incubation, incubation et élevage des jeunes) de paires d’individus avait un effet sur toutes les composantes de la qualité phénotypique des jeunes. Les résultats de cette étude de corrélation suggèrent que les parents ont des décisions complexes à prendre quant au choix du moment optimal de la reproduction et du micro-habitat. De plus, différents facteurs écologiques semblent interagir dans leur influence sur le succès reproducteur et la qualité des jeunes chez l’hirondelle rustique. Les changements naturels et anthropogéniques des conditions écologiques, incluant la température ambiante et la distribution des animaux d’élevage, peuvent ainsi avoir un effet sur la distribution et la productivité des populations d’un passereau en déclin.Mots-clés : élevage d’animaux de ferme, Hirundo rustica, immunocompétence, qualité des jeunes, température, tendances saisonnières.
Nomenclature: Linnaeus, 1758.
Introduction
1Rec. 2005-06-06; acc. 2005-11-05. Associate Editor: Gary Burness.2Author for correspondence.

ÉCOSCIENCE, vOl. 13 (3), 2006
299
parasite virulence, which can greatly affect the fitness of the hosts, may also vary during the breeding season (Møller, 1991). The optimal time for breeding will therefore be determined by the combined effects of factors that vary sea-sonally on the number of offspring and diverse components of their phenotypic quality.
On a short time scale, ecological conditions may vary unpredictably, because weather conditions can markedly fluctuate over short periods. Environmental temperature, in particular, can greatly affect availability of food (Turner, 1982; Hill & Chambers, 1992) and the costs of thermo-regulation by the nestlings and parents (Turner, 1982), particularly for insectivores. Furthermore, temperature can affect offspring phenotype via other pathways (McCarty & Winkler, 1999). For example, low temperature during egg formation may depress egg quality and therefore negatively affect offspring performance. Such early maternal effects have been documented in a number of animal taxa, includ-ing birds (review in Mousseau & Fox, 1998). Finally, low temperatures may enhance the physiological cost of egg production and incubation by females, thus depressing their condition and capability of egg and nestling attendance (Turner, 1982). At small spatial scales, breeding microhabi-tats often vary considerably in quality, and their conditions can influence reproductive performance (Högstedt, 1980; Dhondt, Kempenaers & Adriaensen, 1992; Seki & Takano, 1998). Birds are therefore predicted to adjust the time of reproduction according to the conditions of the microhabitat where they breed (Seki & Takano, 1998).
The aim of this study was to analyze seasonal varia-tion in offspring number and quality in the barn swallow (Hirundo rustica) and to investigate the effect of mean daily temperatures at different stages of the breeding cycle (pre-incubation, incubation, and nestling periods) on nestling phenotypic traits. We predicted that maximal offspring qual-ity and breeding success are achieved later than first broods are produced and that nestling quality is positively associated with ambient temperature. We measured four components of offspring phenotypic quality, including three morphological characters (body mass, body size, and plumage develop-ment) and T cell–mediated immune response, which is a fundamental component of the acquired immune system and influences ability to resist parasite attack (Saino, Calza & Møller, 1997; Hõrak et al., 1999; Christe et al., 2001). Barn swallows almost exclusively breed synanthropically in buildings, and larger colonies are usually associated with livestock farming (Møller, 1983; 2001; Ambrosini et al., 2002a). Livestock can buffer abrupt ambient temperature variations, thus providing better conditions for nesting in the rooms where it is reared, but it may also affect several com-ponents of barn swallow habitat quality (e.g., availability of insect prey and nesting sites), thus creating discontinuous variation in microhabitat quality (Møller, 2001). We there-fore also analyze the effect of livestock farming on nestling quality and its variation during the breeding season, while predicting that nestlings are, on average, of better quality in rooms where livestock is reared.
Measuring the temperature in the rooms where the indi-vidual nests we studied were located might have provided important information on the effect of thermal microhabitat
conditions on nestling phenotype. However, the nests we studied were located in a total of 248 rooms at 57 different farms. This made it unfeasible to obtain reliable estimates of mean daily temperatures for all nests in our sample throughout the entire nestling period. In any case, these data were not required in the present study as our aim was to test the effect of outdoor temperature on nestling phenotype as probably mediated by an effect on food availability to for-aging parents.
MethodsFIEld prOCEdurES
The study was conducted during spring 2001 in 57 farms in an intensively cultivated area (240 km2) in north-ern Italy (see Ambrosini et al., 2002a for a description of the study area). Nests were visited at least once a week from 7 April until 9 July 2001 to record breeding events and every day around hatching to record hatching date. In our study area the earliest date of laying of the first egg record-ed during the 1993–2003 breeding seasons in 200–400 nests per season was 2 April. As all existing nests were inspected at least once before 15 April, we did not miss any first clutches. Owing to the large number of colonies and pairs considered in this study, we could not capture and indi-vidually mark all breeding adults. This prevented us from identifying second broods, as barn swallows may change nest in consecutive breeding attempts. Hence, we focused our analysis on first broods only. In order to avoid includ-ing second broods in our data, we disregarded all nests in which the first egg occurred after fledging of the brood that hatched first in that colony (Ambrosini et al., 2002a). We sought to measure nestlings when they were 10 d old. However, due to the large number of nests and colonies stud-ied, measuring all the nestlings at exactly the same age was not feasible. Nestlings were therefore measured when they were 9 to 11 d old. This age was chosen because we could confidently assume that nestlings had not already started their decrease in weight which occurs before fledging. Out of the 1155 nestlings considered, 271 (23.5%) were 9 d old, 582 (50.4%) were 10 d old, and 302 (26.1%) were 11 d old. Since nestling phenotypic values may vary in relation to age, we included age at measurement (hereafter “age”) as a covariate in statistical models (see below). We individually marked all nestlings and measured their right tarsometatarsus (hereafter “tarsus” for brevity) with a calliper (precision 0.05 mm, innermost left rectrix with a ruler (precision of 0.5 mm) and body mass with a Pesola spring balance (precision 0.1 g) (PESOLA AG, Baar, Switzerland). We also did a cutaneous test to measure T cell–mediated immune response by inject-ing the right wing web with a lectin (phytohemagglutinin, PHA) following the procedure described in Saino, Calza, and Møller (1997). The PHA skin test has become a standard method to assess T cell–mediated components of immune response in birds in the wild (Saino, Calza & Møller, 1997; Saino et al., 1999; Smits, Bortolotti & Tella, 1999; Tella, Sheuerlein & Ricklefs, 2002). Brood size at the time when measures were taken was considered an index of breeding success as mortality after age 10 d accounts for less than 5% of total nestling mortality (N. Saino, unpubl. data).
At each room where nests were located we recorded the presence of cows, pigs, and horses. This information was

AmbrOSINI et al.: vArIAtION IN NEStlINg quAlIty
300
summarized in a dichotomous variable accounting for pres-ence or absence of livestock in that room (Ambrosini et al., 2002b). Different livestock species may affect barn swal-low reproduction in different ways. However, in a previous study carried on in the same study area, we demonstrated that presence of livestock, no matter what species, is the best predictor of both barn swallow presence and abundance (Ambrosini et al., 2002a). Hence, we included this simple variable in the analyses.
tEmpErAturES
We obtained the mean 24-h temperatures recorded at the weather station of Montanaso Lombardo, a small vil-lage within the study area, from the Ufficio Centrale di Ecologia Agraria of the Italian Ministero delle Politiche Agricole e Forestali (http://www.ucea.it). Our study area is wide (approximately 60 × 6 km), but it is entirely within the Po river plain, ranging in altitude from 40 to 110 m asl, and weather conditions are therefore uniform. Thus, we are confident that data from a single weather station are repre-sentative of conditions in the whole study area. Temperature data were not available for a few days during the period April–July 2001, however. Missing values were replaced with those recorded at the weather station of Linate airport, which is close to our study area (about 20 km away).
StAtIStICAl mEthOdS
Throughout the study, date is expressed as progressive days with 1 April = 1. Preliminary analyses were performed to check for a difference in the number of breeding pairs and in the time of breeding in rooms with or without live-stock. These analyses were performed using Linear Mixed Effects Models (LMEs) where “farm” was included as a clustering variable and “livestock farming” as a fixed fac-tor. The mean within-nest value of nestling phenotypic traits was calculated and used for subsequent analysis. Statistical analyses of brood size and nestling phenotype were per-formed using LMEs with the same clustering variable and factor, while the other variables were included as covariates. The interactions between “livestock farming” and all the covariates were also initially entered in the LMEs. To derive parsimonious models we adopted a backward selection pro-cedure of variables. At each step of the selection procedure we excluded the interaction or main effect with the smallest associated effect size until the model contained only signifi-cant effects. The linear term of breeding date was removed only after the squared term. When the squared term of date was significant, we also tested the significance of variation before and after the vertex, if any, of the fitted function. If a significant interaction between the factor livestock farming and date was found, these analyses were run separately for colonies with or without livestock. For brevity, only final models after stepwise selection procedure are presented in the results. All analyses were performed using the lme pro-cedure of the S-Plus 4.5 statistical package (MathSoft Inc., Seattle, Washington, USA).
IdENtIFICAtION OF rElEvANt brEEdINg StAgES FOr thE ANAlySIS OF tEmpErAturE EFFECt
We sought to test for the effect on nestling morphol-ogy and immunity of mean daily temperature at different
stages in the breeding cycle. We therefore created a data set in which mean daily temperatures up to 40 d before nestling measurement were associated with individual nests. The 40-d period included at least 8 d before the laying of the first egg in a clutch, and thus probably included the period of rapid yolk development (RYD) (Robertson, 1995; Williams & Cooch, 1996). We subdivided the 40-d interval before the day of measurement into three periods: the pre-incubation period (including the pre-laying and laying days), the incu-bation period, and the nestling period.
We called the difference in days between the focal day and that of nestling measurement the “breeding stage”. For each breeding stage we ran analyses in which we included farm as a clustering variable and livestock farming in the room where the nest occurred as a fixed factor. The linear and squared terms of date, age, brood size, and tem-perature were entered as covariates. For each period we identified the breeding stage when mean daily temperature had the largest effect on nestling phenotype using the z-ratio value, which is computed as the parameter estimate divided by its asymptotic standard error. It is also known as the Wald Z; it is a likelihood-based statistic that is valid for large samples. Mean daily temperatures in the 3 d, one for each breeding period, when maximum predictive capa-bility was achieved were then entered in the subsequent analysis of temperature effect.
mOdElS OF brOOd SIzE ANd NEStlINg quAlIty
The effect of the presence of livestock farming on brood size at measurement was investigated in an LME with farm as the clustering variable, livestock farming as a fixed factor, and nestling age as a covariate. Age was included as it could account for the effect of mortality from day 9 to day 11. Variation in nestling phenotypic variables was investigated in models that also included brood size as a covariate.
Seasonal variation in brood size and nestling quality was investigated in LMEs that also included the linear and squared terms of date at nestling measurement (an index of breeding date, hereafter “date” for simplicity).
The effect of temperature on the components of nestling quality was analyzed in LMEs where we simultaneously entered mean daily temperatures in the 3 d when maximum predictive capability was achieved (see above).
In all cases we finally derived parsimonious models by excluding the non-significant terms according to their z-ratio (see above).
COmpArISON bEtwEEN dAtE At mAxImum FItNESS COmpO-NENtS ANd ACtuAl dAtE At rEprOduCtION
We were interested in assessing the date when brood size or components of nestling quality reached a maximum. Therefore, in all cases we calculated when the fitted func-tions reached a maximum in the period considered by the present study (13 May–19 July). To compare date at maxi-mum nestling phenotypic quality and actual date of repro-duction, as indicated by date at measurement, we relied on a bootstrap resampling approach (Sokal & Rohlf, 1995; Siriwardena et al., 1998) following the procedure described
ÉCOSCIENCE, vOl. 13 (3), 2006
301
in Ambrosini et al. (2002b). Briefly, 1000 replicates, each containing the same number of nests as the original data set, was produced by randomly resampling, with replacement, the observed data. The same model fitted to the original data set was then fitted to each replicate, and the date (within the range of measurement dates relevant to this study, i.e., 3 May–19 July) when the maximum of the focal phenotypic trait was achieved was computed. The significance of the difference between the date at maximum phenotypic value and the mean breeding date was calculated as the proportion of replicates in which the date at maximum occurred earlier or later than mean breeding date. The difference was consid-ered significant when this proportion was < 0.05.
ResultsOur analyses were based on 248 first broods with a total
of 1155 nestlings. Due to accidental reasons, the tarsus length of the nestlings in one brood and the immunocompetence index of the nestlings in three broods were not available.
The number of breeding pairs was larger in rooms with livestock compared to those without livestock (z = 4.229, P < 0.001; mean number of breeding pairs in rooms with live-stock = 8.030 ± 1.237 [SE], without livestock = 3.715 ± 0.414 [SE]). The date at measurement, which reflects date at repro-duction, did not differ significantly between rooms with and without livestock (z = –0.7518, P = 0.452; mean date in rooms with livestock = 65.426 ± 1.639 [SE], without live-stock = 67.191 ± 1.497 [SE]).
mICrOhAbItAt CONdItIONS ANd SEASONAl vArIAtION IN brOOd SIzE ANd NEStlINg phENOtypIC quAlIty
Brood size and components of nestling phenotypic quality did not differ in rooms with or without livestock, as indicated by the fact that the “livestock farming” factor never achieved statistical significance (z = 1.040, P ≥ 0.298 in all cases, details not shown).
Body mass and central rectrix length significantly increased with nestling age (body mass: z = 4.379, P < 0.001, b = 7.452 ± 1.702 [SE]; central rectrix length: z = 7.065, P < 0.001, b = 27.489 ± 3.891 [SE]), but was unaffected by brood size. Tarsus length positively covaried with both brood size (z = 3.010, P = 0.003, b = 0.791 ± 0.263 [SE]) and age (z = 2.597, P = 0.010, b = 0.953 ± 0.367 [SE]). Immunocompetence index negatively covaried with brood size (z = -3.011, P = 0.003, b = -7.927 ± 2.633 [SE]) but was unaffected by nestling age.
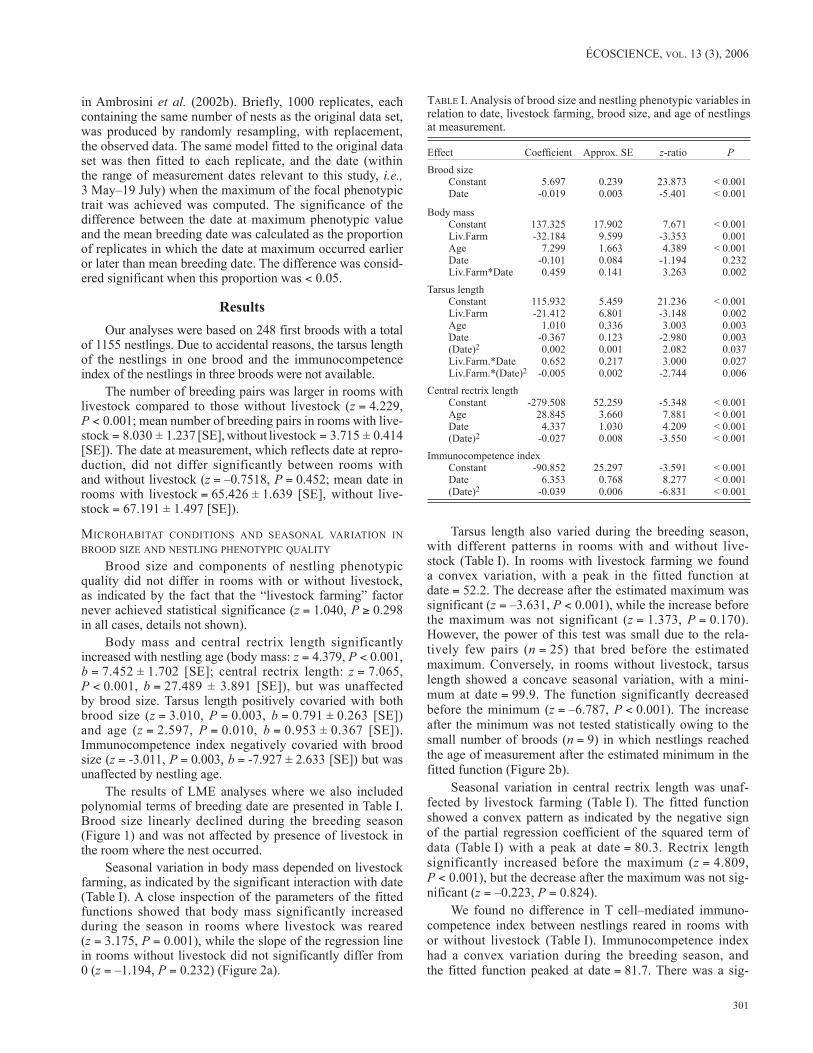
The results of LME analyses where we also included polynomial terms of breeding date are presented in Table I. Brood size linearly declined during the breeding season (Figure 1) and was not affected by presence of livestock in the room where the nest occurred.
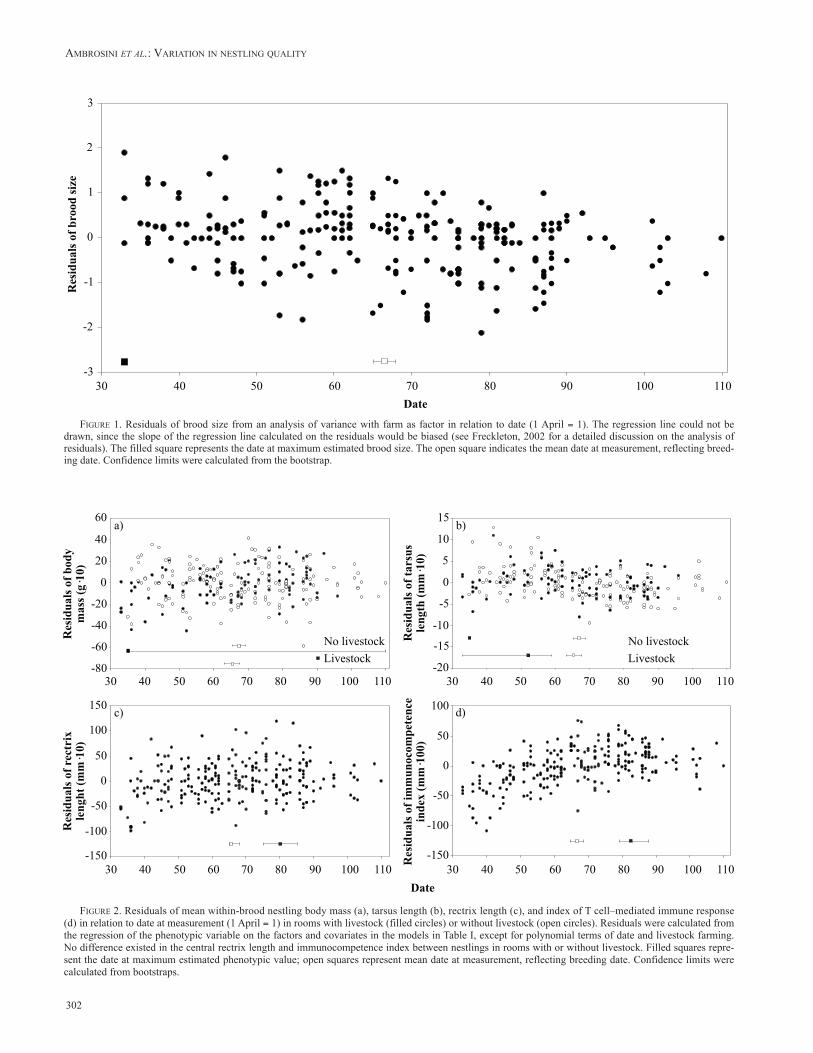
Seasonal variation in body mass depended on livestock farming, as indicated by the significant interaction with date (Table I). A close inspection of the parameters of the fitted functions showed that body mass significantly increased during the season in rooms where livestock was reared (z = 3.175, P = 0.001), while the slope of the regression line in rooms without livestock did not significantly differ from 0 (z = –1.194, P = 0.232) (Figure 2a).
Tarsus length also varied during the breeding season, with different patterns in rooms with and without live-stock (Table I). In rooms with livestock farming we found a convex variation, with a peak in the fitted function at date = 52.2. The decrease after the estimated maximum was significant (z = –3.631, P < 0.001), while the increase before the maximum was not significant (z = 1.373, P = 0.170). However, the power of this test was small due to the rela-tively few pairs (n = 25) that bred before the estimated maximum. Conversely, in rooms without livestock, tarsus length showed a concave seasonal variation, with a mini-mum at date = 99.9. The function significantly decreased before the minimum (z = –6.787, P < 0.001). The increase after the minimum was not tested statistically owing to the small number of broods (n = 9) in which nestlings reached the age of measurement after the estimated minimum in the fitted function (Figure 2b).
Seasonal variation in central rectrix length was unaf-fected by livestock farming (Table I). The fitted function showed a convex pattern as indicated by the negative sign of the partial regression coefficient of the squared term of data (Table I) with a peak at date = 80.3. Rectrix length significantly increased before the maximum (z = 4.809, P < 0.001), but the decrease after the maximum was not sig-nificant (z = –0.223, P = 0.824).
We found no difference in T cell–mediated immuno-competence index between nestlings reared in rooms with or without livestock (Table I). Immunocompetence index had a convex variation during the breeding season, and the fitted function peaked at date = 81.7. There was a sig-
tAblE I. Analysis of brood size and nestling phenotypic variables in relation to date, livestock farming, brood size, and age of nestlings at measurement.
Effect Coefficient Approx. SE z-ratio PBrood size Constant 5.697 0.239 23.873 < 0.001 Date -0.019 0.003 -5.401 < 0.001
Body mass Constant 137.325 17.902 7.671 < 0.001 Liv.Farm -32.184 9.599 -3.353 0.001 Age 7.299 1.663 4.389 < 0.001 Date -0.101 0.084 -1.194 0.232 Liv.Farm*Date 0.459 0.141 3.263 0.002Tarsus length Constant 115.932 5.459 21.236 < 0.001 Liv.Farm -21.412 6.801 -3.148 0.002 Age 1.010 0.336 3.003 0.003 Date -0.367 0.123 -2.980 0.003 (Date)2 0.002 0.001 2.082 0.037 Liv.Farm.*Date 0.652 0.217 3.000 0.027 Liv.Farm.*(Date)2 -0.005 0.002 -2.744 0.006Central rectrix length Constant -279.508 52.259 -5.348 < 0.001 Age 28.845 3.660 7.881 < 0.001 Date 4.337 1.030 4.209 < 0.001 (Date)2 -0.027 0.008 -3.550 < 0.001Immunocompetence index Constant -90.852 25.297 -3.591 < 0.001 Date 6.353 0.768 8.277 < 0.001 (Date)2 -0.039 0.006 -6.831 < 0.001
AmbrOSINI et al.: vArIAtION IN NEStlINg quAlIty
302
FIgurE 2. Residuals of mean within-brood nestling body mass (a), tarsus length (b), rectrix length (c), and index of T cell–mediated immune response (d) in relation to date at measurement (1 April = 1) in rooms with livestock (filled circles) or without livestock (open circles). Residuals were calculated from the regression of the phenotypic variable on the factors and covariates in the models in Table I, except for polynomial terms of date and livestock farming. No difference existed in the central rectrix length and immunocompetence index between nestlings in rooms with or without livestock. Filled squares repre-sent the date at maximum estimated phenotypic value; open squares represent mean date at measurement, reflecting breeding date. Confidence limits were calculated from bootstraps.
FIgurE 1. Residuals of brood size from an analysis of variance with farm as factor in relation to date (1 April = 1). The regression line could not be drawn, since the slope of the regression line calculated on the residuals would be biased (see Freckleton, 2002 for a detailed discussion on the analysis of residuals). The filled square represents the date at maximum estimated brood size. The open square indicates the mean date at measurement, reflecting breed-ing date. Confidence limits were calculated from the bootstrap.

ÉCOSCIENCE, vOl. 13 (3), 2006
303
nificant increase in immunocompetence index before the maximum (z = 8.658, P < 0.001), but also in this case the decrease after the maximum was not significant (z = –0.081, P = 0.935) (Figure 2d).
bOOtStrAp ANAlySIS OF dAtE At mAxImum NEStlINg quAlIty ANd mEAN dAtE At mEASurEmENt
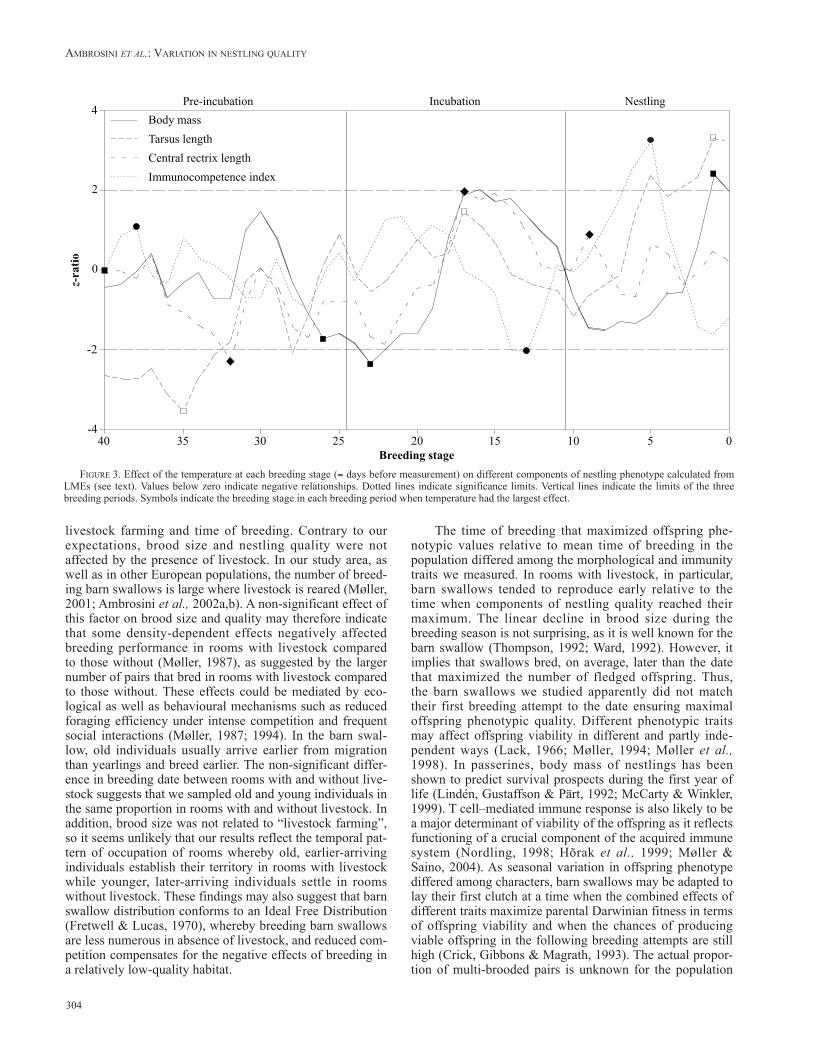
Mean date of breeding occurred later than the date when maximum brood size occurred (Table II; Figure 1). In rooms with livestock farming, maximum body mass was achieved significantly later than the mean time of reproduc-tion of barn swallows, while no difference existed between the date at maximum body mass and mean time of repro-duction in rooms without livestock (Table II; Figure 2a). Maximum tarsus length occurred significantly earlier than mean time of reproduction both in rooms with and without livestock (Table II; Figure 2b). Date at maximum rectrix length occurred significantly later than the mean date of breeding. Finally, barn swallows were found to breed signif-icantly earlier than the date that determined the maximum immunocompetence of nestlings (Table II; Figure 2d).
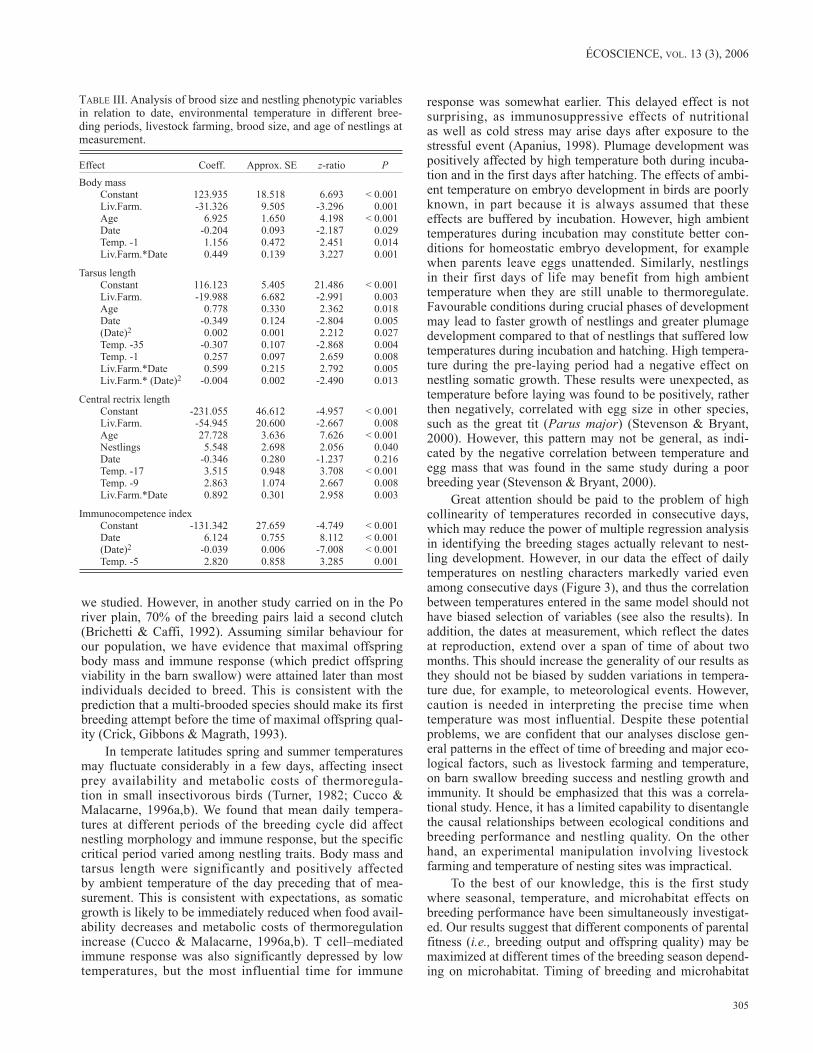
EFFECt OF tEmpErAturE
The effect of temperature at each breeding stage on brood size and components of nestling phenotypic quality is represented in Figure 3. Final models are summarized in Table III. Correlation coefficients between temperatures simultaneously entered in our models ranged between 0.23 and 0.53, with the only exception being the correlation coef-ficient between temperature at breeding stages -26 and -23 simultaneously entered in the model of body mass, where it was 0.71. This value was larger than 0.7, which has been suggested as the threshold above which collinearity may bias multiple regression models (Green, 1979; Fielding & Haworth, 1995). Hence, in the model of body mass we checked for possible biases in the backward selection proce-dure of variables by changing the order of variable removal. However, alternative models always converged to the final model of Table III (details not shown) and therefore collin-earity of independent variables should have not biased our analysis.
Temperatures included in the analysis were recorded over a long period of time, as they are related to time of reproduc-tion. Mean date at measure, which reflects date at reproduction, was 66.5 ± 1.11 (mean ± SE), which corresponds to 5 June.
The minimum and maximum dates were respectively 33 (3 May) and 110 (19 July), while quartiles were respectively 53 (23 May), 67 (6 June), and 80 (19 June). Hence, tem-perature in the same breeding stage for different nests can be recorded in days as far apart in time as two months.
Temperature of the day preceding that of measurement (stage -1) had a significant and positive effect on body mass. Date, age of nestlings, and livestock farming significantly affected body mass, implying that seasonal effects existed independently of temperature variation. A significant effect of the interaction between date and livestock farming also persisted (see also Table I). Temperature of the day preced-ing that of measurement (stage -1) and during the pre-laying period (stage -35) had significant effects on tarsus length. Specifically, temperature at stage -1 had a positive effect on tarsus length, while temperature at stage -35 had a negative effect. The general pattern of variation in tarsus length dur-ing the breeding season did not change after controlling for the effect of environmental temperature (see also Table I). Temperature during the laying period (stage -17) and in the nestling period (stage -9), had significant and reciprocally independent positive effects on central rectrix length. The inclusion of the effect of temperatures led to a change in the patterns of variation in central rectrix length in relation to date (see also Table I) and to the inclusion of livestock farming and brood size in the final model. In particular, there was a linear increase in rectrix length with date in rooms with livestock and a linear decrease in rooms without livestock. A close inspection of the parameters of the model showed that there was a significant difference in the slopes of these regression lines, but neither the linear increase in rooms with livestock nor the linear decrease in rooms with-out livestock was significantly different from 0 (livestock present: z = 1.706, P = 0.088; livestock absent: z = –1.237, P = 0.216). Finally, temperature recorded 5 d before the immune response was measured had a significant positive effect on nestling immunocompetence. The seasonal pattern of variation did not change after the inclusion of this term in the model (see also Table I).
DiscussionWe found complex patterns of variation in brood size,
nestling morphology, and immune response in relation to
tAblE II. Average date at measurement (an index of breeding date) and date at maximum phenotypic values of nestlings. 5% and 95% confi-dence limits were estimated using bootstraps.
Date at measurement Maximum of the fitted function Observed Observed confidence limits Min Max confidence limits Min Max PbrOOd SIzE 66.5 65.1 67.9 33 33 33 < 0.001
bOdy mASS No Livestock 67.2 65.3 69.2 35 35 110 .ns Livestock 65.4 63.1 67.5 96 96 96 < 0.001
tArSuS lENgth No Livestock 67.0 65.2 69.0 35 35 35 < 0.001 Livestock 65.4 63.2 67.5 52.2 33 58.8 < 0.001
rECtrIx lENgth 66.5 65.1 67.9 80.2 75.1 85.2 < 0.001
ImmuNOCOmpEtENCE 66.7 65.3 68.1 81.7 79.3 85.5 < 0.001
AmbrOSINI et al.: vArIAtION IN NEStlINg quAlIty
304
livestock farming and time of breeding. Contrary to our expectations, brood size and nestling quality were not affected by the presence of livestock. In our study area, as well as in other European populations, the number of breed-ing barn swallows is large where livestock is reared (Møller, 2001; Ambrosini et al., 2002a,b). A non-significant effect of this factor on brood size and quality may therefore indicate that some density-dependent effects negatively affected breeding performance in rooms with livestock compared to those without (Møller, 1987), as suggested by the larger number of pairs that bred in rooms with livestock compared to those without. These effects could be mediated by eco-logical as well as behavioural mechanisms such as reduced foraging efficiency under intense competition and frequent social interactions (Møller, 1987; 1994). In the barn swal-low, old individuals usually arrive earlier from migration than yearlings and breed earlier. The non-significant differ-ence in breeding date between rooms with and without live-stock suggests that we sampled old and young individuals in the same proportion in rooms with and without livestock. In addition, brood size was not related to “livestock farming”, so it seems unlikely that our results reflect the temporal pat-tern of occupation of rooms whereby old, earlier-arriving individuals establish their territory in rooms with livestock while younger, later-arriving individuals settle in rooms without livestock. These findings may also suggest that barn swallow distribution conforms to an Ideal Free Distribution (Fretwell & Lucas, 1970), whereby breeding barn swallows are less numerous in absence of livestock, and reduced com-petition compensates for the negative effects of breeding in a relatively low-quality habitat.
The time of breeding that maximized offspring phe-notypic values relative to mean time of breeding in the population differed among the morphological and immunity traits we measured. In rooms with livestock, in particular, barn swallows tended to reproduce early relative to the time when components of nestling quality reached their maximum. The linear decline in brood size during the breeding season is not surprising, as it is well known for the barn swallow (Thompson, 1992; Ward, 1992). However, it implies that swallows bred, on average, later than the date that maximized the number of fledged offspring. Thus, the barn swallows we studied apparently did not match their first breeding attempt to the date ensuring maximal offspring phenotypic quality. Different phenotypic traits may affect offspring viability in different and partly inde-pendent ways (Lack, 1966; Møller, 1994; Møller et al., 1998). In passerines, body mass of nestlings has been shown to predict survival prospects during the first year of life (Lindén, Gustaffson & Pärt, 1992; McCarty & Winkler, 1999). T cell–mediated immune response is also likely to be a major determinant of viability of the offspring as it reflects functioning of a crucial component of the acquired immune system (Nordling, 1998; Hõrak et al., 1999; Møller & Saino, 2004). As seasonal variation in offspring phenotype differed among characters, barn swallows may be adapted to lay their first clutch at a time when the combined effects of different traits maximize parental Darwinian fitness in terms of offspring viability and when the chances of producing viable offspring in the following breeding attempts are still high (Crick, Gibbons & Magrath, 1993). The actual propor-tion of multi-brooded pairs is unknown for the population
FIgurE 3. Effect of the temperature at each breeding stage (= days before measurement) on different components of nestling phenotype calculated from LMEs (see text). Values below zero indicate negative relationships. Dotted lines indicate significance limits. Vertical lines indicate the limits of the three breeding periods. Symbols indicate the breeding stage in each breeding period when temperature had the largest effect.
ÉCOSCIENCE, vOl. 13 (3), 2006
305
we studied. However, in another study carried on in the Po river plain, 70% of the breeding pairs laid a second clutch (Brichetti & Caffi, 1992). Assuming similar behaviour for our population, we have evidence that maximal offspring body mass and immune response (which predict offspring viability in the barn swallow) were attained later than most individuals decided to breed. This is consistent with the prediction that a multi-brooded species should make its first breeding attempt before the time of maximal offspring qual-ity (Crick, Gibbons & Magrath, 1993).
In temperate latitudes spring and summer temperatures may fluctuate considerably in a few days, affecting insect prey availability and metabolic costs of thermoregula-tion in small insectivorous birds (Turner, 1982; Cucco & Malacarne, 1996a,b). We found that mean daily tempera-tures at different periods of the breeding cycle did affect nestling morphology and immune response, but the specific critical period varied among nestling traits. Body mass and tarsus length were significantly and positively affected by ambient temperature of the day preceding that of mea-surement. This is consistent with expectations, as somatic growth is likely to be immediately reduced when food avail-ability decreases and metabolic costs of thermoregulation increase (Cucco & Malacarne, 1996a,b). T cell–mediated immune response was also significantly depressed by low temperatures, but the most influential time for immune
response was somewhat earlier. This delayed effect is not surprising, as immunosuppressive effects of nutritional as well as cold stress may arise days after exposure to the stressful event (Apanius, 1998). Plumage development was positively affected by high temperature both during incuba-tion and in the first days after hatching. The effects of ambi-ent temperature on embryo development in birds are poorly known, in part because it is always assumed that these effects are buffered by incubation. However, high ambient temperatures during incubation may constitute better con-ditions for homeostatic embryo development, for example when parents leave eggs unattended. Similarly, nestlings in their first days of life may benefit from high ambient temperature when they are still unable to thermoregulate. Favourable conditions during crucial phases of development may lead to faster growth of nestlings and greater plumage development compared to that of nestlings that suffered low temperatures during incubation and hatching. High tempera-ture during the pre-laying period had a negative effect on nestling somatic growth. These results were unexpected, as temperature before laying was found to be positively, rather then negatively, correlated with egg size in other species, such as the great tit (Parus major) (Stevenson & Bryant, 2000). However, this pattern may not be general, as indi-cated by the negative correlation between temperature and egg mass that was found in the same study during a poor breeding year (Stevenson & Bryant, 2000).
Great attention should be paid to the problem of high collinearity of temperatures recorded in consecutive days, which may reduce the power of multiple regression analysis in identifying the breeding stages actually relevant to nest-ling development. However, in our data the effect of daily temperatures on nestling characters markedly varied even among consecutive days (Figure 3), and thus the correlation between temperatures entered in the same model should not have biased selection of variables (see also the results). In addition, the dates at measurement, which reflect the dates at reproduction, extend over a span of time of about two months. This should increase the generality of our results as they should not be biased by sudden variations in tempera-ture due, for example, to meteorological events. However, caution is needed in interpreting the precise time when temperature was most influential. Despite these potential problems, we are confident that our analyses disclose gen-eral patterns in the effect of time of breeding and major eco-logical factors, such as livestock farming and temperature, on barn swallow breeding success and nestling growth and immunity. It should be emphasized that this was a correla-tional study. Hence, it has a limited capability to disentangle the causal relationships between ecological conditions and breeding performance and nestling quality. On the other hand, an experimental manipulation involving livestock farming and temperature of nesting sites was impractical.
To the best of our knowledge, this is the first study where seasonal, temperature, and microhabitat effects on breeding performance have been simultaneously investigat-ed. Our results suggest that different components of parental fitness (i.e., breeding output and offspring quality) may be maximized at different times of the breeding season depend-ing on microhabitat. Timing of breeding and microhabitat
tAblE III. Analysis of brood size and nestling phenotypic variables in relation to date, environmental temperature in different bree-ding periods, livestock farming, brood size, and age of nestlings at measurement.
Effect Coeff. Approx. SE z-ratio PBody mass Constant 123.935 18.518 6.693 < 0.001 Liv.Farm. -31.326 9.505 -3.296 0.001 Age 6.925 1.650 4.198 < 0.001 Date -0.204 0.093 -2.187 0.029 Temp. -1 1.156 0.472 2.451 0.014 Liv.Farm.*Date 0.449 0.139 3.227 0.001
Tarsus length Constant 116.123 5.405 21.486 < 0.001 Liv.Farm. -19.988 6.682 -2.991 0.003 Age 0.778 0.330 2.362 0.018 Date -0.349 0.124 -2.804 0.005 (Date)2 0.002 0.001 2.212 0.027 Temp. -35 -0.307 0.107 -2.868 0.004 Temp. -1 0.257 0.097 2.659 0.008 Liv.Farm.*Date 0.599 0.215 2.792 0.005 Liv.Farm.* (Date)2 -0.004 0.002 -2.490 0.013
Central rectrix length Constant -231.055 46.612 -4.957 < 0.001 Liv.Farm. -54.945 20.600 -2.667 0.008 Age 27.728 3.636 7.626 < 0.001 Nestlings 5.548 2.698 2.056 0.040 Date -0.346 0.280 -1.237 0.216 Temp. -17 3.515 0.948 3.708 < 0.001 Temp. -9 2.863 1.074 2.667 0.008 Liv.Farm.*Date 0.892 0.301 2.958 0.003
Immunocompetence index Constant -131.342 27.659 -4.749 < 0.001 Date 6.124 0.755 8.112 < 0.001 (Date)2 -0.039 0.006 -7.008 < 0.001 Temp. -5 2.820 0.858 3.285 0.001

AmbrOSINI et al.: vArIAtION IN NEStlINg quAlIty
306
choice may thus reflect adaptive strategies whereby indi-viduals maximize the combination of different components of their Darwinian fitness rather than a single component (e.g., number of fledged offspring).
Great emphasis has recently been given to the effect of anthropogenic environmental changes, including global warming and changes in agricultural practices, on bird popu-lations (Forchhammer, Post & Stenseth, 1998; Siriwardena et al., 1998; Ormerod & Watkinson, 2000). Natural or anthro-pogenic variation in ecological conditions such as livestock farming practices and temperature may therefore generate a mismatch between the time of breeding and the time when conditions for reproduction are optimal and thus negatively affect the breeding performance of passerines.
AcknowledgementsWe are grateful to the farm owners who allowed us into their
barns and houses to collect data. F. Addario, S. Arieni, B. Fabbo, M. Paba, and D. Tosi greatly contributed to field work. This study was funded by the Parco Regionale Adda Sud.
Literature citedAmbrosini, R., A. M. Bolzern, L. Canova, S. Arieni, A. P. Møller
& N. Saino, 2002a. The distribution and colony size of barn swallows in relation to agricultural land use. Journal of Applied Ecology, 39: 524–534.
Ambrosini, R., A. M. Bolzern, L. Canova & N. Saino, 2002b. Latency in response of barn swallow Hirundo rustica popu-lations to changes in breeding habitat conditions. Ecology Letters, 5: 640–647.
Apanius, V., 1998. Stress and immune response. Pages 133–150 in A. P. Møller, M. Milinski & P. J. B. Slater (eds.). Advances in the Study of Behavior. Volume 27. Academic Press, San Diego, California.
Blancher, P. J. & R. J. Robertson, 1987. Effect of food supply on the breeding biology of Western Kingbirds. Ecology, 68: 723–732.
Brichetti, P & M. Caffi, 1992. Biologia riproduttiva di una popo-lazione di rondine Hirundo rustica, nidificante in un cascinale della padania. Rivista Piemontese di Scienze Naturali, 13: 73–87 (in Italian with English summary).
Buse, A., S. J. Dury, R. J. W. Woodburn, C. M. Perrins & J. E. G. Good, 1999. Effects of elevated temperature on multi-species interactions: The case of Peduncolate Oak, Winter Moth and Tits. Functional Ecology, 13: 74–82.
Christe, P., F. de Lope, G. Gonzàlez, N. Saino & A. P. Møller, 2001. The influence of weather conditions and ectoparasites on immune response, morphology and recapture probability of nestling house martins (Delichon urbica). Oecologia, 126: 333–338.
Crick, H. Q. P., D. W. Gibbons & R. D. Magrath, 1993. Seasonal changes in clutch size in British birds. Journal of Animal Ecology, 62: 263–273.
Cucco, M. & G. Malacarne, 1996a. Effect of food availability on nestling growth and fledging success in manipulated pallid swift broods. Journal of Zoology, 240: 141–151.
Cucco, M. & G. Malacarne, 1996b. Reproduction of the pallid swift (Apus pallidus) in relation to weather and aerial insect abundance. Italian Journal of Zoology, 63: 247–253.
Dhondt, A. A., B. Kempenaers & F. Adriaensen, 1992. Density dependent clutch size determined by habitat heterogeneity. Journal of Animal Ecology, 61: 643–648.
Fielding, A. H. & P. F. Haworth, 1995. Testing the generality of bird-habitat models. Conservation Biology, 9: 1466–1481.
Forchhammer, M. C., E. Post & N. C. Stenseth, 1998. Breeding phenology and climate. Nature, 391: 29–30.
Freckleton, R. P., 2002. On the misuse of residuals in ecology: Regression of residuals versus multiple regression. Journal of Animal Ecology, 71: 542–546.
Fretwell, S. D. & H. L. Lucas, 1970. On territorial behaviour and other factors influencing habitat distribution in birds. I. Theoretical development. Acta Biotheoretica, 19: 16–36.
Green, R., 1979. Sampling Design and Statistical Methods for Environmental Biologists. Wiley, New York, New York.
Hill, J. R. & L. Chambers, 1992. Purple Martin weather deaths during the summer of ’92. Purple Martin Update, 4: 6–10.
Högstedt, G., 1980. Evolution of clutch size in birds: Adaptive vari-ation in relation to territory quality. Science, 210: 1148–1150.
Hõrak, P., L. Tegelmann, I. Ots & A. P. Møller, 1999. Immune function and survival of great tit nestlings in relation to growth conditions. Oecologia, 121: 316–322.
Lack, D., 1954. The Natural Regulation of Animal Numbers. Oxford University Press, Oxford.
Lack, D., 1966. Population Studies of Birds. Clarendon Press, Oxford.
Lindén, M., L. Gustaffson & T. Pärt, 1992. Selection on fledging mass in the collared flycatcher and the great tit. Ecology, 73: 336–343.
Linnaeus, C., 1758. Systema Naturae per Regna Tria Naturae secundum Classes, Ordinus, Genera, Species cum Characteribus, Differentiis Synonymis, Locis. 10e édition. Volume. 1. Holmiae.
Lochmiller, R. L., M. R. Vestey & J. C. Boren, 1993. Relationship between protein nutritional status and immunocompetence in northern bobwhite chicks. Auk, 110: 503–510.
McCarty, J. P. & D. W. Winkler, 1999. Relative importance of environmental variables in determining the growth of nestling Tree Swallows Tachycineta bicolor. Ibis, 141: 286–296.
Møller, A. P., 1983. Breeding habitat selection in the Swallow Hirundo rustica. Bird Study, 30: 134–142.
Møller, A. P., 1987. Advantages and disadvantages of coloniality in the swallow, Hirundo rustica. Animal Behaviour, 35: 819–832.
Møller, A. P., 1991. Parasites, sexual ornaments and mate choice in the barn swallows. Pages 328–348 in J. E. Loye & M. Zuk (eds.). Bird-Parasite Interactions. Ecology, Evolution and Behaviour. Oxford University Press, Oxford.
Møller, A. P., 1994. Sexual Selection and the Barn Swallow. Oxford University Press, Oxford.
Møller, A. P., 2001. The effect of dairy farming on barn swallow Hirundo rustica abundance, distribution and reproduction. Journal of Applied Ecology, 38: 378–389.
Møller, A. P. & N. Saino, 2004. Immune response and survival. Oikos, 104: 299–304.
Møller, A. P., P. Christe, J. Erritzøe & J. Mavarez, 1998. Condition, disease and immune defence. Oikos, 83: 301–306.
Mousseau, T. A. & C. W. Fox, 1998. Maternal Effects. Oxford University Press, New York, New York.
Nordling, D., 1998. Trade-offs Between Life-History Traits and Immune Defence in the Collared Flycatcher Ficedula albicol-lis. Ph.D. thesis, Uppsala University, Uppsala.
Ormerod, S. J. & A. R. Watkinson, 2000. Editors’ Introduction: Birds and Agriculture. Journal of Applied Ecology, 37: 699–705.
Robertson, G. J., 1995. Annual variation in common eider egg size: Effects of temperature, clutch size, laying date, and laying sequence. Canadian Journal of Zoology, 73: 1579–1587.

ÉCOSCIENCE, vOl. 13 (3), 2006
307
Saino, N., S. Calza. & A. P. Møller, 1997. Immunocompetence of nestling barn swallow in relation to brood size and parental effort. Journal of Animal Ecology, 66: 827–836.
Saino, N., S. Calza, P. Ninni & A. P. Møller, 1999. Barn swallows trade survival against offspring condition and immunocompe-tence. Journal of Animal Ecology, 68: 999–1009.
Seki, S.-I. & H. Takano, 1998. Caterpillar abundance in the terri-tory affects the breeding performance of great tit Parus major minor. Oecologia, 114: 514–521.
Siriwardena, G. M., S. R. Baillie, S. T. Buckland, R. M. Fewster, J. H. Marchant & J. D. Wilson, 1998. Trends in abundance of far-mland birds: A quantitative comparison of smoothed Common Bird Census indices. Journal of Applied Ecology, 35: 24–43.
Smith, H. G., H. Källander & J.-Å. Nilsson, 1989. The trade-off between offspring number and quality in the great tit Parus major. Journal of Animal Ecology, 58: 383–401.
Smits, J. E., G. R. Bortolotti & J. L. Tella, 1999. Simplifying the phytohemagglutinin skin-testing technique in studies of avian immunocompetence. Functional Ecology, 13: 567–572.
Sokal, R. R. & F. J. Rohlf, 1995. Biometry: The Principle and Practice of Statistics in Biological Research. 3rd Edition. Freeman, New York, New York.
Stevenson, I. R. & D. M. Bryant, 2000. Climate change and constraints on breeding. Nature, 406: 366–367.
Takagi, M., 2001. Some effects of inclement weather conditions on the survival and condition of bull-headed shrike nestlings. Ecological Research, 16: 55–63.
Tella, J. L., A. Sheuerlein & R. E. Ricklefs, 2002. Is cell-mediated immunity related to the evolution of life-history strategies in birds? Proceedings of the Royal Society of London, Series B, 269: 1059–1066.
Thompson, M. L. P., 1992. Reproductive success and survival of swallows (Hirundo rustica). Effect of age and breeding condi-tions. Ph.D. thesis, University of Stirling, Stirling.
Turner, A. K., 1982. Timing of laying by swallows (Hirundo rus-tica) and sand martins (Riparia riparia). Journal of Animal Ecology, 51: 29–46.
Ward, S., 1992. Energetic of Laying and Incubation in Birds. Studies of Swallows Hirundo rustica, Dippers Cinclus cinclus and Japanese Quail Coturnix coturnix japonica. Ph.D. thesis, University of Stirling, Stirling.
Williams, T. D. & E. G. Cooch, 1996. Egg size, temperature and laying sequence: Why do snow geese lay big eggs when it’s cold? Functional Ecology, 10: 112–118.





























