Embed Size (px)
Citation preview

http://hol.sagepub.com/The Holocene
http://hol.sagepub.com/content/24/8/1009The online version of this article can be found at:
DOI: 10.1177/0959683614534743
2014 24: 1009 originally published online 21 May 2014The HoloceneXimena S Villagran and Paulo CF Giannini
Shell mounds as environmental proxies on the southern coast of Brazil
Published by:
http://www.sagepublications.com
can be found at:The HoloceneAdditional services and information for
http://hol.sagepub.com/cgi/alertsEmail Alerts:
http://hol.sagepub.com/subscriptionsSubscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.com/journalsPermissions.navPermissions:
http://hol.sagepub.com/content/24/8/1009.refs.htmlCitations:
What is This?
- May 21, 2014OnlineFirst Version of Record
- Jul 15, 2014Version of Record >>
by guest on July 17, 2014hol.sagepub.comDownloaded from by guest on July 17, 2014hol.sagepub.comDownloaded from

The Holocene2014, Vol. 24(8) 1009 –1016© The Author(s) 2014Reprints and permissions:sagepub.co.uk/journalsPermissions.navDOI: 10.1177/0959683614534743hol.sagepub.com
IntroductionShell mounds, or sambaquis, are accumulations of shells and fish bones mixed with natural sediments, organic debris, and other refuse, including artifacts, made by prehistoric hunter–gatherer–fisher populations. On the coast of Brazil, sites date back to c. 8700 cal. yr BP (Lima et al., 2002). The large volume and the frequent concentration of human burials inside the mounds lead researchers to interpret their formation as building endeavors, although function is still a matter of debate (Afonso and DeBlasis, 1994; DeBlasis et al., 1998; Figuti and Klokler, 1996; Gaspar et al., 2008; Klokler, 2008).
Dating carbonate shells collected from the base of the shell mounds has been traditionally used for reconstructing relative-sea-level (RSL) curves on the coast of Brazil (Bigarella, 1965; Fairbridge, 1976; Martin and Suguio, 1976; Suguio et al., 1992). Although many authors have argued the geological and cultural constraints of using shell mounds for building RSL curves and for paleo-shoreline reconstruction (see Angulo and Lessa, 1997; Angulo et al., 2006; Giannini et al., 2005; Scheel-Ybert et al., 2009), at least one of the premises behind this use is still present in modern archaeology. The premise refers to the chosen location for shell mound construction: always near large bodies of water containing a diversity of fish species, the main subsistence item among coastal groups (Figuti, 1992; Klokler, 2008), as well as intertidal and subtidal resources (Barbosa, 2007; DeBlasis et al., 2007; Gaspar et al., 2008; Klokler, 2008; Kneip, 2004).
Furthermore, recent archaeological studies show that the evo-lution of prehistoric settlement patterns in the states of Rio Grande do Sul, Santa Catarina, São Paulo, and Rio de Janeiro has been
directly influenced by changes in coastal landscapes (Barbosa, 2007; Calippo, 2010; DeBlasis et al., 2007; Giannini et al., 2010; Kneip, 2004; Perreti, 2009), which ultimately result from RSL fluctuations after the maximum Holocene transgression, among other factors. Thus, even if shell mounds are not precise indica-tors of paleo-shorelines and RSL changes, their constant proxim-ity to water sources means that they can still hold evidences of the paleo-geographical changes in coastal areas during the Holocene, for which RSL is only one of the controlling variables.
Mollusks in shell-bearing sites (e.g. shell middens, shell mounds, etc.) do not necessarily reflect the adjacent aquatic habi-tat (Claassen, 1991; Stein, 1992a; Waselkov, 1987), but the ecol-ogy of the predominant mollusk species can be used as a paleo-environmental proxy whenever there are multiple records available for comparison (geological, botanical, etc.; see exam-ples in Cortez-Sanchez et al., 2008; Fa, 2008; Morey and Croth-ers, 1998; Morey et al., 2002; Sandweiss, 2003; Stein, 1992b; Van der Schriek et al., 2007, 2008). In this respect, the sediments con-tained in shell-bearing sites have been less used as an environ-mental proxy. However, considering the basket-load action of collecting and transporting mollusks to the sites, whether for
Shell mounds as environmental proxies on the southern coast of Brazil
Ximena S Villagran1 and Paulo CF Giannini2
AbstractDespite the multiple constraints of using shell mounds for building relative-sea-level (RSL) curves, one of the premises behind this use is still valid in modern archaeological research. This refers to the sites being continuously built near bodies of water rich in fish species and mollusk beds. Studies that combine the evolution of settlement patterns with the geological evolution of coastal areas in Brazil reach analogous results: the distribution of shell mounds in time and space follows the landscape transformations induced by RSL variations through the Holocene. Although shell mounds are not precise indicators of RSL, they provide evidences of the paleo-geographical changes during the Holocene, of which RSL is one of the many control variables. By collecting and transporting mollusks to the shell mounds, humans inevitably carry sediments from the substrate where mollusks live, for example, the beaches and lagoons near the sites. In this work, the geoarchaeological study of three shell mounds located in the southern coast of Santa Catarina State, combined with mollusk taxa identification, show the direct association of shell mound content with the changing landscape. The predominance of Ostrea sp. versus Anomalocardia brasiliana, the presence of colluvial versus lagoonal sediments, and the occurrence of echinoid spines versus muddy aggregates with diatoms, gastropods, and glauconitic clay characterize the distinction between sites built before and after the maximum Holocene transgression, respectively. This attests the potential of geoarchaeological analyses in shell mounds as a complementary proxy for paleo-environmental reconstructions.
Keywordsgeoarchaeology, maximum Holocene transgression, micromorphology, sambaqui
Received 28 November 2013; revised manuscript accepted 9 April 2014
1Eberhard Karls Universität Tübingen, Germany2Universidade de São Paulo, Brazil
Corresponding author:Ximena S Villagran, Institute for Archaeological Sciences, Eberhard Karls Universität Tübingen, Rümelinstrasse 23, Tübingen 72072, Germany. Email: [email protected]
534743 HOL0010.1177/0959683614534743The HoloceneVillagran and Gianniniresearch-article2014
Research report
by guest on July 17, 2014hol.sagepub.comDownloaded from
1010 The Holocene 24(8)
consumption or as prime material for mound building, remnants of the substrate where the mollusks live (i.e. the beaches or lagoons that existed nearby) are inevitably carried to the sites. Likewise, since some Brazilian shell mounds show evidences of being built after reworking of debris deposited close by (Villagran et al., 2009, 2010), natural sediments from the surroundings could also be carried together. Thus, the sediments in the shell mounds can potentially provide information on the paleo-geography near the sites and can, therefore, be used as another environmental proxy. Based on the dynamics of shell mound formation, we pres-ent the geoarchaeological study of three Brazilian shell mounds and their potential for reconstructing paleo-geography, by com-bining the traditional identification of mollusk species with geo-archaeological analyses of shell mound sediments.
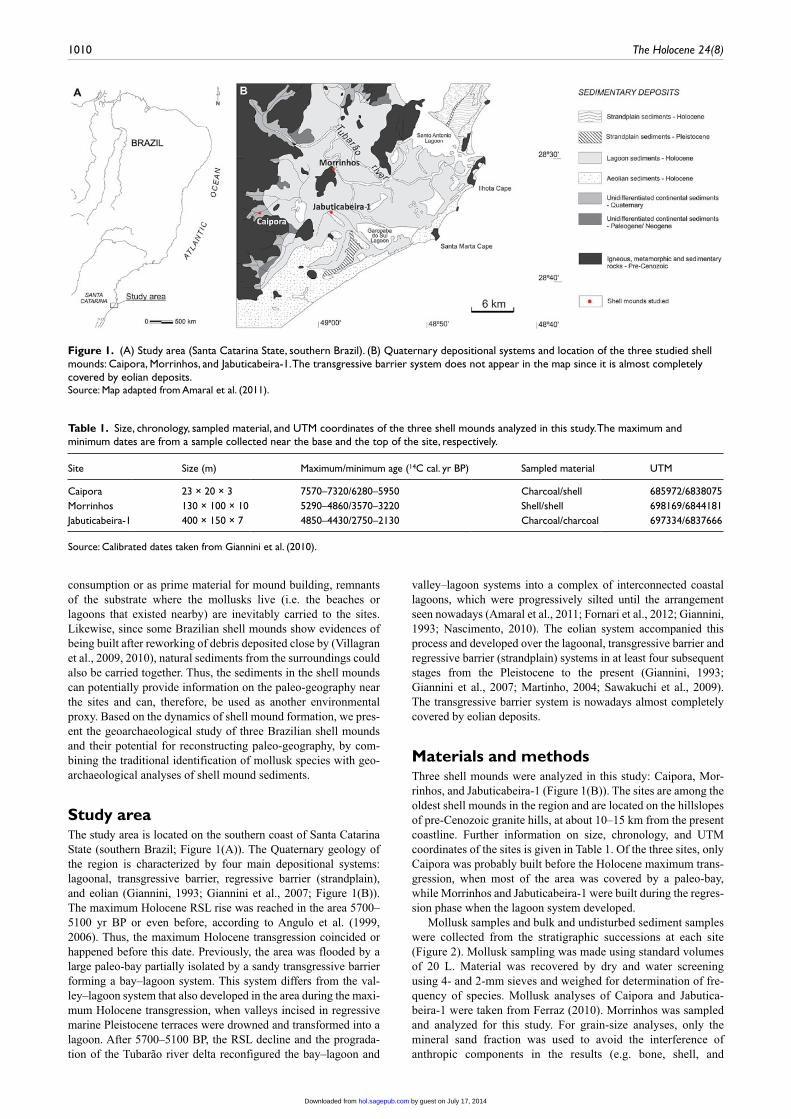
Study areaThe study area is located on the southern coast of Santa Catarina State (southern Brazil; Figure 1(A)). The Quaternary geology of the region is characterized by four main depositional systems: lagoonal, transgressive barrier, regressive barrier (strandplain), and eolian (Giannini, 1993; Giannini et al., 2007; Figure 1(B)). The maximum Holocene RSL rise was reached in the area 5700–5100 yr BP or even before, according to Angulo et al. (1999, 2006). Thus, the maximum Holocene transgression coincided or happened before this date. Previously, the area was flooded by a large paleo-bay partially isolated by a sandy transgressive barrier forming a bay–lagoon system. This system differs from the val-ley–lagoon system that also developed in the area during the maxi-mum Holocene transgression, when valleys incised in regressive marine Pleistocene terraces were drowned and transformed into a lagoon. After 5700–5100 BP, the RSL decline and the prograda-tion of the Tubarão river delta reconfigured the bay–lagoon and
valley–lagoon systems into a complex of interconnected coastal lagoons, which were progressively silted until the arrangement seen nowadays (Amaral et al., 2011; Fornari et al., 2012; Giannini, 1993; Nascimento, 2010). The eolian system accompanied this process and developed over the lagoonal, transgressive barrier and regressive barrier (strandplain) systems in at least four subsequent stages from the Pleistocene to the present (Giannini, 1993; Giannini et al., 2007; Martinho, 2004; Sawakuchi et al., 2009). The transgressive barrier system is nowadays almost completely covered by eolian deposits.
Materials and methodsThree shell mounds were analyzed in this study: Caipora, Mor-rinhos, and Jabuticabeira-1 (Figure 1(B)). The sites are among the oldest shell mounds in the region and are located on the hillslopes of pre-Cenozoic granite hills, at about 10–15 km from the present coastline. Further information on size, chronology, and UTM coordinates of the sites is given in Table 1. Of the three sites, only Caipora was probably built before the Holocene maximum trans-gression, when most of the area was covered by a paleo-bay, while Morrinhos and Jabuticabeira-1 were built during the regres-sion phase when the lagoon system developed.
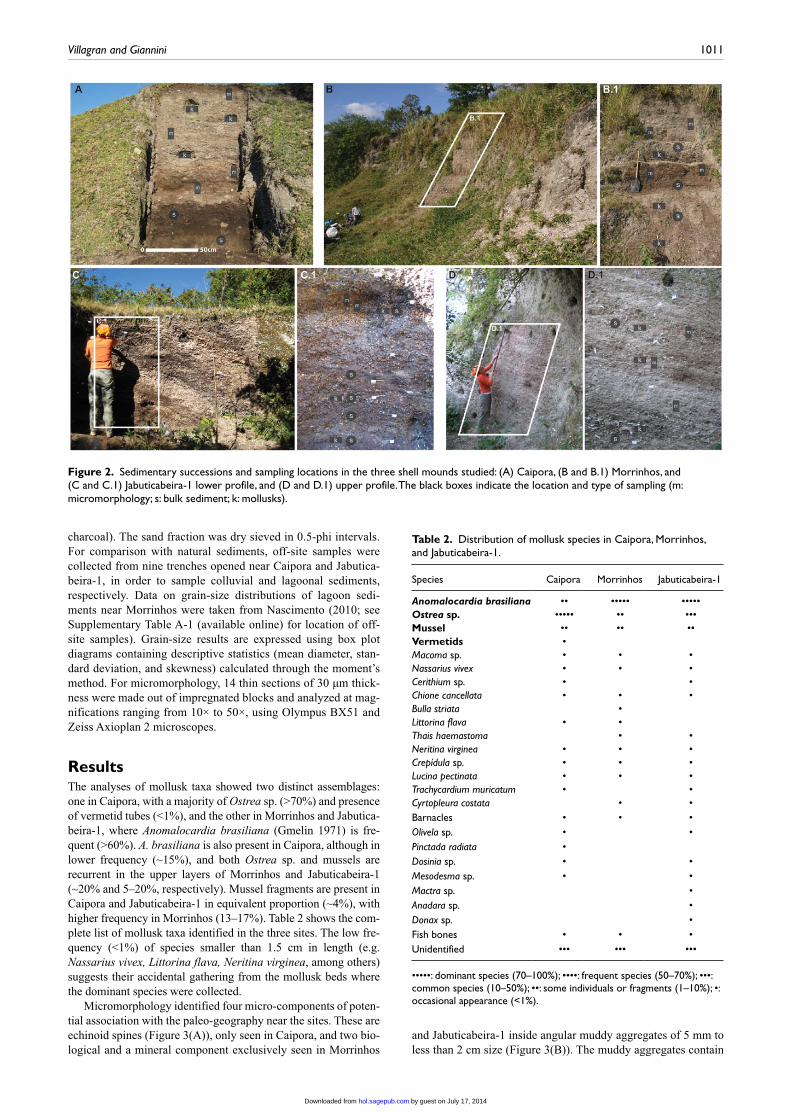
Mollusk samples and bulk and undisturbed sediment samples were collected from the stratigraphic successions at each site (Figure 2). Mollusk sampling was made using standard volumes of 20 L. Material was recovered by dry and water screening using 4- and 2-mm sieves and weighed for determination of fre-quency of species. Mollusk analyses of Caipora and Jabutica-beira-1 were taken from Ferraz (2010). Morrinhos was sampled and analyzed for this study. For grain-size analyses, only the mineral sand fraction was used to avoid the interference of anthropic components in the results (e.g. bone, shell, and
Figure 1. (A) Study area (Santa Catarina State, southern Brazil). (B) Quaternary depositional systems and location of the three studied shell mounds: Caipora, Morrinhos, and Jabuticabeira-1. The transgressive barrier system does not appear in the map since it is almost completely covered by eolian deposits.Source: Map adapted from Amaral et al. (2011).
Table 1. Size, chronology, sampled material, and UTM coordinates of the three shell mounds analyzed in this study. The maximum and minimum dates are from a sample collected near the base and the top of the site, respectively.
Site Size (m) Maximum/minimum age (14C cal. yr BP) Sampled material UTM
Caipora 23 × 20 × 3 7570–7320/6280–5950 Charcoal/shell 685972/6838075Morrinhos 130 × 100 × 10 5290–4860/3570–3220 Shell/shell 698169/6844181Jabuticabeira-1 400 × 150 × 7 4850–4430/2750–2130 Charcoal/charcoal 697334/6837666
Source: Calibrated dates taken from Giannini et al. (2010).
by guest on July 17, 2014hol.sagepub.comDownloaded from
Villagran and Giannini 1011
charcoal). The sand fraction was dry sieved in 0.5-phi intervals. For comparison with natural sediments, off-site samples were collected from nine trenches opened near Caipora and Jabutica-beira-1, in order to sample colluvial and lagoonal sediments, respectively. Data on grain-size distributions of lagoon sedi-ments near Morrinhos were taken from Nascimento (2010; see Supplementary Table A-1 (available online) for location of off-site samples). Grain-size results are expressed using box plot diagrams containing descriptive statistics (mean diameter, stan-dard deviation, and skewness) calculated through the moment’s method. For micromorphology, 14 thin sections of 30 µm thick-ness were made out of impregnated blocks and analyzed at mag-nifications ranging from 10× to 50×, using Olympus BX51 and Zeiss Axioplan 2 microscopes.
ResultsThe analyses of mollusk taxa showed two distinct assemblages: one in Caipora, with a majority of Ostrea sp. (>70%) and presence of vermetid tubes (<1%), and the other in Morrinhos and Jabutica-beira-1, where Anomalocardia brasiliana (Gmelin 1971) is fre-quent (>60%). A. brasiliana is also present in Caipora, although in lower frequency (~15%), and both Ostrea sp. and mussels are recurrent in the upper layers of Morrinhos and Jabuticabeira-1 (~20% and 5–20%, respectively). Mussel fragments are present in Caipora and Jabuticabeira-1 in equivalent proportion (~4%), with higher frequency in Morrinhos (13–17%). Table 2 shows the com-plete list of mollusk taxa identified in the three sites. The low fre-quency (<1%) of species smaller than 1.5 cm in length (e.g. Nassarius vivex, Littorina flava, Neritina virginea, among others) suggests their accidental gathering from the mollusk beds where the dominant species were collected.
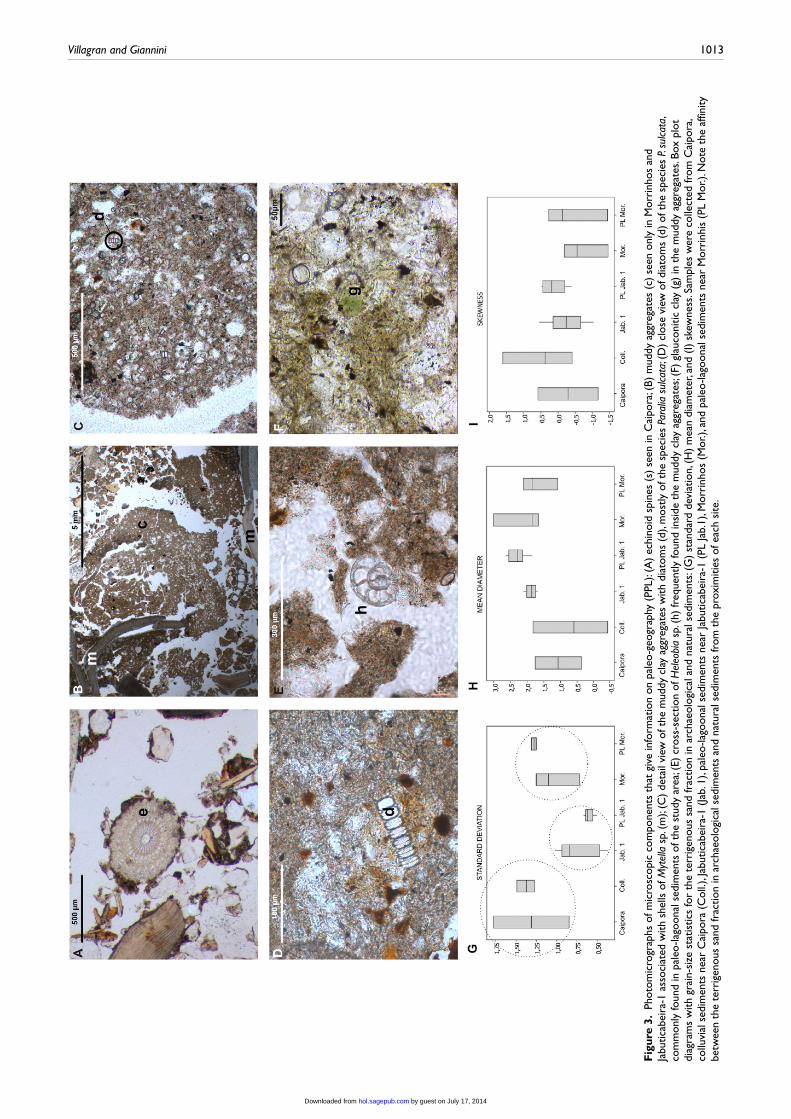
Micromorphology identified four micro-components of poten-tial association with the paleo-geography near the sites. These are echinoid spines (Figure 3(A)), only seen in Caipora, and two bio-logical and a mineral component exclusively seen in Morrinhos
and Jabuticabeira-1 inside angular muddy aggregates of 5 mm to less than 2 cm size (Figure 3(B)). The muddy aggregates contain
Figure 2. Sedimentary successions and sampling locations in the three shell mounds studied: (A) Caipora, (B and B.1) Morrinhos, and (C and C.1) Jabuticabeira-1 lower profile, and (D and D.1) upper profile. The black boxes indicate the location and type of sampling (m: micromorphology; s: bulk sediment; k: mollusks).
Table 2. Distribution of mollusk species in Caipora, Morrinhos, and Jabuticabeira-1.
Species Caipora Morrinhos Jabuticabeira-1
Anomalocardia brasiliana •• ••••• •••••Ostrea sp. ••••• •• •••Mussel •• •• ••Vermetids • Macoma sp. • • •Nassarius vivex • • •Cerithium sp. • •Chione cancellata • • •Bulla striata • Littorina flava • • Thais haemastoma • •Neritina virginea • • •Crepídula sp. • • •Lucina pectinata • • •Trachycardium muricatum • •Cyrtopleura costata • •Barnacles • • •Olivela sp. • •Pinctada radiata • Dosinia sp. • •Mesodesma sp. • •Mactra sp. •Anadara sp. •Donax sp. •Fish bones • • •Unidentified ••• ••• •••
•••••: dominant species (70–100%); ••••: frequent species (50–70%); •••: common species (10–50%); ••: some individuals or fragments (1–10%); •: occasional appearance (<1%).
by guest on July 17, 2014hol.sagepub.comDownloaded from

1012 The Holocene 24(8)
diatoms (Figure 3(C) and (D)), a gastropod genus identified as Heleobia sp. (Figure 3(E)), and grains of glauconitic clay miner-als (Figure 3(F)). In Jabuticabeira-1, the aggregates show evi-dences of anthropic heating like carbonization and rubefaction (Figure 4(A)). Besides the changes induced by heating, the dia-toms and gastropods are still visible in the aggregates (Figure 4(B) and (C)). The identification of glauconitic clay was done by its optical properties: grains of cryptocrystalline green clay of 20–30 µm, sub-equidimensional and subrounded shape, and first-order interference color (maximum estimated birefringence around 0.020). XRD analysis to confirm this mineralogy could not be made because of the difficulty to concentrate material that happens in such low quantity. Besides shells, bone, and charcoal fragments, the coarse fraction in the three sites is made of variable proportions of quartz grains, feldspars, and rock fragments, with rock fragments being predominant only in Caipora.
Box plots of grain-size statistics (mean diameter, standard deviation, and skewness; see Supplementary Table A-2 (available online) for grain-size data) showed the affinity of shell mound sediments with the natural sediments in close proximity: colluvial deposits in Caipora and lagoon sediments in Morrinhos and Jabu-ticabeira 1 (Figure 3(G)–(I)). For the three statistical parameters, the interquartile range for natural sediments (colluvium in the case of Caipora and paleo-lagoonal for Jabuticabeira-1 and Mor-rinhos) is contained in the variation range for shell mound sediments.
DiscussionThe data set allowed differentiating between the sedimentary sig-natures of shell mounds built before and after the maximum Holocene transgression. This difference expresses the interplay between the changing environment and the preferences of coastal groups.
Pre-maximum Holocene transgression shell moundThe faunal assemblage in Caipora is characterized by the predom-inance of Ostrea sp. with frequent vermetid tubes attached to the surfaces of the shells (vermetid fragments also appear loose in the sediments). Two species of Oyster sp. are found on the southern coasts of Brazil: Ostrea equestris (Say 1834) and Ostrea puel-chana (Orbigny 1841). Both are euryhaline intertidal and subtidal marine species that live on rocky bottoms (De Souza et al., 2011; Rios, 1985), while vermetids are marine gastropods that live on rocky shores exposed to wave action (Laborel and Laborel-Deguen, 1996). The predominance of Ostrea sp. in Caipora and the exclusive presence of vermetid tubes in this site suggest that before the maximum Holocene transgression, coastal populations were gathering mollusks from rocky shores exposed to marine waters. The echinoid spines (625–645 µm, basal section) seen in thin sections corroborate this. Echinoids are marine subtidal invertebrates that make up the biological assemblage in rocky coasts (Laborel and Laborel-Deguen, 1996). Its presence in the shell mound sediments indicates marine influence in the body of water near the site where resources were being captured. Echinoid spines have also been described in thin sections from other shell-bearing sites located on marine shores in South Africa (Goldberg, 2000), the San Juan Islands (United States; Stein et al., 2011), and Rio de Janeiro (Brazil; Corrêa et al., 2013). The body of water near Caipora would correspond to the large paleo-bay that formed in the mid-Holocene transgression flooding. The chronology of Caipora, the predominance of Ostrea sp., and the presence of echinoids suggest that rocky shores were readily exploited for their proximity to the site (Figure 5(A)).
The comparison of grain-size statistics of Caipora and nearby natural sediments (colluvium) shows close affinity (Figure
3(G)–(I)). This, and the visual evaluation of the mineral coarse fraction in thin section (with coarse angular quartz grains, sub-angular feldspars and granule sized angular rock fragments), con-firms the association of shell mound sediments with colluvial deposits. Besides collecting mollusks from the rocky shores nearby, colluvial sediments from the surroundings were also being brought to the mounds, mixed with faunal and other organic components. This supports the hypothesis of shell mounds being built after reworking of debris previously left near the sites. The natural deposition of colluvial sediments on the shell mound is precluded by the higher topography of the mound in relation to the surroundings.
Post-maximum Holocene transgression shell moundsBoth Morrinhos and Jabuticabeira-1 are mainly composed of A. brasiliana, an intertidal euryhaline species that lives in muddy and muddy sand beaches, commonly bays and estuaries (Rios, 1985). It burrows 5 cm deep in quiet waters with little distur-bance of the bottom deposits and is easily collected during low tides (Narchi, 1972). This species proliferated in the region with the development of the lagoon system. However, the presence of Ostrea sp. in the upper layers of both sites attests continuity in the exploitation of this resource with a diminishment in inten-sity. By the time Morrinhos and Jabuticabeira-1 were active, Ostrea sp. could be collected from rocky shores within the lagoon system or brought from the maritime rocky shores located more than 5 km away (Figure 5(B)). The intermixing of layers rich in A. brasiliana and Ostrea sp. is seen in other sites of the region (Klokler, 2008; Kneip, 2004). The switch in spe-cies could be indicative of a foraging behavior intended to pre-vent the over-exploitation of the shell beds (see Mannino and Thomas, 2002).
Besides the shells of Ostrea sp. and A. brasiliana, sediments in Morrinhos and Jabuticabeira-1 contain quartz sands associated with lagoon sediments, as demonstrated by comparison of statisti-cal parameters between natural and cultural sediments (Figure 3(G)–(I)). Muddy aggregates are another common component in the shell mound sediments. Micromorphological analyses showed that the muddy aggregates contain diatoms, gastropods and glau-conitic clay, and, in Jabuticabeira-1, the aggregates were the sub-strate of hearths lit near or over the shell mound.
The most abundant diatom species found in the muddy aggre-gates is Paralia sulcata. Other species like Triceratium favus, Actinoptychus vulgaris, and fragments of Biddulphia pulchella and Coscinodiscus sp. appear in less abundance. Both P. sulcata and B. pulchella are marine eurytropic species, which means they tolerate variations in water salinity. They have optimal develop-ment in eusaline conditions (salinity between 30‰ and 40‰), but can also grow in meso-polysaline (salinity between 5‰ and 30‰) and even metasaline waters (salinity above 40‰). T. favus and A. vulgaris are marine stenotopic species, with low tolerance to vari-ations in salinity that live only in eusaline conditions. Thus, the diatom assembly in the muddy aggregates suggests meso to eusa-line conditions (salinity from 5–40‰) associated with estuarine–lagoonal sediments.
This hypothesis is reinforced by the fine grain size of the aggregates and the high frequency of P. sulcata, whose predomi-nance has already been detected in lagoonal sediments of the area (Amaral et al., 2011). Likewise, the muddy aggregates in Mor-rinhos and Jabuticabeira-1 contain many specimens of Heleobia sp. (Stimpson 1865), a gastropod commonly found in muddy sandy substrate associated with estuarine–lagoonal environments (Rios, 1985). This gastropod was only identified at genus level because shape alone is not sufficient to determine the species. However, the species Heleobia australis is well known in the area and lives nowadays in the Garopaba do Sul lagoon (Netto et al.,
by guest on July 17, 2014hol.sagepub.comDownloaded from
Villagran and Giannini 1013
Fig
ure
3. P
hoto
mic
rogr
aphs
of m
icro
scop
ic c
ompo
nent
s th
at g
ive
info
rmat
ion
on p
aleo
-geo
grap
hy (
PPL)
: (A
) ec
hino
id s
pine
s (s
) se
en in
Cai
pora
; (B)
mud
dy a
ggre
gate
s (c
) se
en o
nly
in M
orri
nhos
and
Ja
butic
abei
ra-1
ass
ocia
ted
with
she
lls o
f Myt
ella
sp.
(m
); (C
) de
tail
view
of t
he m
uddy
cla
y ag
greg
ates
with
dia
tom
s (d
), m
ostly
of t
he s
peci
es P
aral
ia s
ulca
ta; (
D)
clos
e vi
ew o
f dia
tom
s (d
) of
the
spe
cies
P. s
ulca
ta,
com
mon
ly fo
und
in p
aleo
-lago
onal
sed
imen
ts o
f the
stu
dy a
rea;
(E)
cros
s-se
ctio
n of
Hel
eobi
a sp
. (h)
freq
uent
ly fo
und
insi
de t
he m
uddy
cla
y ag
greg
ates
; (F)
gla
ucon
itic
clay
(g)
in t
he m
uddy
agg
rega
tes.
Box
plot
di
agra
ms
with
gra
in-s
ize
stat
istic
s fo
r th
e te
rrig
enou
s sa
nd fr
actio
n in
arc
haeo
logi
cal a
nd n
atur
al s
edim
ents
: (G
) st
anda
rd d
evia
tion,
(H
) m
ean
diam
eter
, and
(I)
skew
ness
. Sam
ples
wer
e co
llect
ed fr
om C
aipo
ra,
collu
vial
sed
imen
ts n
ear
Cai
pora
(C
oll.)
, Jab
utic
abei
ra-1
(Ja
b. 1
), pa
leo-
lago
onal
sed
imen
ts n
ear
Jabu
ticab
eira
-1 (
PL Ja
b.1)
, Mor
rinh
os (
Mor
.), a
nd p
aleo
-lago
onal
sed
imen
ts n
ear
Mor
rinh
is (
PL M
or.).
Not
e th
e af
finity
be
twee
n th
e te
rrig
enou
s sa
nd fr
actio
n in
arc
haeo
logi
cal s
edim
ents
and
nat
ural
sed
imen
ts fr
om t
he p
roxi
miti
es o
f eac
h si
te.
by guest on July 17, 2014hol.sagepub.comDownloaded from

1014 The Holocene 24(8)
2012), as well as in other coastal lagoons of Brazil, Uruguay, and Argentina (Martínez et al., 2006).
The glauconitic clay minerals in the muddy aggregates testify the history of the formation of the lagoon deposits. Glauconite is a 2:1 clay mineral formed in eodiagenesis, under the influence of marine waters in low sedimentation rates (see Odin, 1988; Odin and Fullagar, 1988). Glauconite is commonly found today in the continental platform (Dillenburg et al., 2000). Its presence in the muddy aggregates, associated with estuarine–lagoonal sediments (fresh to brackish waters) according to the diatom and mollusk assembly, suggests the mechanical reworking of marine sedi-ments from the shelf during the formation of the lagoon system.
A clear association of mussel shells and the muddy aggregates was seen in thin section, which could serve as an indirect indica-tor of the mussel species. Two mussels are common in Brazilian shell mounds: Mytella guayanensis (Lamarck 1819) and Brachi-odontes sp. (Swaison 1840; Figuti, 1993). The mussel M. guaya-nensis is an euryhaline intertidal bivalve that lives in muddy lagoon substrates and intertidal flats, even attached to mangrove roots (Bacon, 1975; De Souza et al., 2011), while Brachiodontes sp. lives only on rocky substrate (De Souza et al., 2011). The lagoon origin of the muddy aggregates and their frequent associa-tion with mussel valves suggests that M. guayanensis, collected from the lagoons nearby, is present in the sites.
The muddy aggregates and the predominant mollusk assem-blage in Morrinhos and Jabuticabeira-1 indicate the exploitation of lagoon resources. The affinity seen between the paleo-lagoonal sands and shell mound sands confirm this.
ConclusionStudies on the relationship of coastal groups and the changing landscape of the Holocene have been done in different states of Brazil (Rio Grande do Sul, São Paulo, and Rio de Janeiro). By contrasting site location and chronology with the RSL curves, studies have shown that shell mounds’ distribution in time and space followed the landscape transformations induced by Holo-cene RSL variations. This is because of the constant construc-tion of shell mounds near bodies of water, sources of fish, and intertidal resources. In several coastal areas of the world, the ecology of the predominant mollusk taxa, in combination with other records, has been used for paleo-environmental recon-structions. In this work, geoarchaeological analyses in three shell mounds of Santa Catarina state show that archaeological sediments can also record the paleo-geographical changes in coastal settings. The accidental carrying of natural sediments from the shell beds to the shell mounds is the human action behind such use of archeological sediments as an environmen-tal proxy.
In the study area, the sediments and mollusk assemblages in sites built prior to and after the maximum Holocene transgression reflect the paleo-geography near the sites, determined partially by RSL changes. The prehistoric occupations near the large paleo-bay before 5100 yr BP (minimum age of the Holocene maximum transgression) and the exploitation of its resources are evidenced in shell mounds containing Ostrea sp., with vermetids, echinoid spines, and colluvial sediments from the hilly surroundings. The
Figure 4. Photomicrographs of muddy aggregates in Jabuticabeira-1 with signs of heating (PPL): carbonized (ca) and rubefied (ra) aggregates in-between shell fragments. Note (A) Heleobia sp. (h) within the rubefied clay, (B) rubefied muddy aggregate with diatom species (d) Paralia sulcata, and (C) carbonized muddy aggregate with diatoms.
Figure 5. Landscape reconstruction before and after the maximum Holocene transgression with diagnostic macro-mollusks and micro-components: (A) the Caipora was built when the area was covered by a large paleo-bay and (B) the Morrinhos and Jabuticabeira-1 sites were first settled between 5500 and 4000 BP, already in the regression phase near the newly formed lagoonal system.Source: Maps adapted from Giannini et al. (2010) and Fornari et al. (2012).
by guest on July 17, 2014hol.sagepub.comDownloaded from

Villagran and Giannini 1015
development of the lagoon system after the Holocene transgres-sion and the exploitation of its resources is evidenced in the shell mounds made of A. brasiliana and M. guayanensis and containing paleo-lagoonal sediments (from sands to clay aggregates with dia-toms and gastropods). The analyses of shell mound sediments show the constant choice of locating settlements near the bodies of water and the continuity in shell mound building practices, besides landscape transformations.
AcknowledgementsThe authors thank Professor Paulo DeBlasis (MAE/USP) coor-dinator of the ‘Sambaquis and Landscape’ research project. The authors also thank Professor Daniela Klokler, Professor Milene Fornari, Dr Paula G. C. Amaral, Kiristen Bright, Julie Boreham, and Tobias Sprafke. Analyses were done at the laboratories of sedimentology and sedimentary petrology of the Institute of Geo-sciences, University of São Paulo. Two anonymous reviewers are also thanked for their helpful comments.
FundingThe doctoral research of Ximena S. Villagran was supported by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP, proc. 08/51264-0). The study was supported by a research grant of the FAPESP (proc. 04/11038-0).
ReferencesAfonso MC and DeBlasis P (1994) Aspectos da formação de
um grande sambaqui: alguns indicadores em Espinheiros II, Joinville. Revista do Museu de Arqueologia e Etnologia 4: 21–30.
Amaral PGCG, Giannini PCF, Sylvestre F et al. (2011) Paleoenvi-ronmental reconstruction of a Late Quaternary lagoon system in southern Brazil (Jaguaruna region, Santa Catarina state) based on multi-proxy analysis. Journal of Quaternary Sci-ence 27(2): 181–191.
Angulo R and Lessa G (1997) The Brazilian sea-level curves: A critical review with emphasis on the curves from the Para-naguá and Cananéia regions. Marine Geology 140(1–2): 141–166.
Angulo R, Giannini PCF, Suguio K et al. (1999) Relative sea-level change during the last 5500 years in the Laguna-Imbituba region (Santa Catarina, Brazil), based on vermetid radiocar-bon ages. Marine Geology 159: 323–339.
Angulo R, Lessa G and Souza M (2006) A critical review of mid- to late-Holocene sea-level fluctuations on the eastern Brazilian coastline. Quaternary Science Reviews 25(5–6): 486–506.
Bacon PR (1975) Shell form, byssal development and habitat of Mytella Guayanensis (Lamarck) and M. Falcata (Orbigny) (Pellecipoda: Mytilidae) in Trinidad, West Indies. Journal of Molluscan Studies 41(6): 511–520.
Barbosa M (2007) A ocupação pré-colonial da região dos lagos, RJ: sistema de assentamento e relações intersocietais entre grupos sambaquianos e grupos ceramistas tupinambá da tradiçao Una. PhD Thesis, Universidade de São Paulo.
Bigarella JJ (1965) Subsídios para o estudo das variações de nível oceânico no Quaternário brasileiro. Anais Da Academia Brasileira De Ciencias 37: 263–278.
Calippo FR (2010) Sociedade sambaquieira, comunidades marítimas. PhD Thesis, Universidade de São Paulo.
Claassen C (1991) Normative thinking in shell-bearing sites. Archaeological Method and Theory 3: 249–298.
Corrêa GR, Schaefer CE and Gilkes RJ (2013) Phosphate loca-tion and reaction in an archaeoanthrosol on shell-mound in the Lakes Region, Rio de Janeiro State, Brazil. Quaternary International 315(27): 16–23.
Cortez-Sanchez M, Morales-Muniz A, Simon-Vallejo M et al. (2008) Palaeoenvironmental and cultural dynamics of the coast of Málaga (Andalusia, Spain) during the Upper Pleis-tocene and early Holocene. Quaternary Science Reviews 27(23–24): 2176–2193.
DeBlasis P, Fish SK, Gaspar MD et al. (1998) Some references for the discussion of complexity among the sambaqui mound-builders from the southern shores of Brazil. Revista de Arque-ologia Americana 15: 75–105.
DeBlasis P, Kneip A, Scheel-Ybert R et al. (2007) Sambaquis e paisagem: dinâmica natural e arqueologia regional no litoral sul do Brasil. Revista de Arqueología Sudamericana 3(1): 29–61.
De Souza RCCL, Lima TA and da Silva E (2011) Conchas marinhas de sambaquis do Brasil. Rio de Janeiro: Technical Books.
Dillenburg SR, Laybauer L, Mexias AS et al. (2000) Significado estratigráfico de minerais glauconíticos da planície coste-ira do Rio Grande do Sul, região da Laguna de Tramandaí. Revista Brasileira de Geociências 30(4): 649–654.
Fa D (2008) Effects of tidal amplitude on intertidal resource avail-ability and dispersal pressure in prehistoric human coastal populations: The Mediterranean–Atlantic transition. Quater-nary Science Reviews 27: 2194–2209.
Fairbridge RW (1976) Shellfish-eating preceramic indians in coastal Brazil. Science 191(4225): 353–359.
Ferraz T (2010) Estudo comparativo dos sambaquis Caipora, Lageado e Jaboticabeira I: interpretações acerca da mudança de material construtivo ao longo do tempo. Master’s Disserta-tion, Universidade de São Paulo.
Figuti L (1992) Les sambaquis COSIPA (4200 à 1200 ans BP): étude de la subsistance chez les peuples préhistoriques de pêcheurs-ramasseurs de bivalves de la côte centrale de l’état de São Paulo, Brésil. PhD Dissertation, Museum National d’Histoire Naturelle.
Figuti L (1993) O homem pré-histórico, o molusco e o sambaqui: considerações sobre a subsistência dos povos sambaquieiros. Revista do Museu de Arqueologia e Etnologia 3: 67–80.
Figuti L and Klokler D (1996) Resultados preliminares dos vestígios zooarqueológicos do sambaqui Espinheiros II (Joinville, SC). Revista do Museu de Arqueologia e Etno-logia 6: 169–188.
Fornari M, Giannini PCF and Nascimento DR (2012) Facies asso-ciations and controls on the evolution from a coastal bay to a lagoon system, Santa Catarina Coast, Brazil. Marine Geology 323–325: 56–68.
Gaspar MD, DeBlasis P, Fish SK et al. (2008) Sambaqui (Shell Mound) societies of coastal Brazil. In: Silverman H and Isbell WH (eds) Handbook of South American Archaeology. New York: Springer, pp. 319–337.
Giannini PCF (1993) Sistemas Deposicionais no Quaternário Costeiro entre Jaguaruna e Imbituba, SC. PhD Thesis, Uni-versidade de São Paulo.
Giannini PCF, Deblasis P, Sawakuchi AO et al. (2005) Processos e materiais geológicos e a construção de sambaquis no litoral sul de. In: Anais do Congresso da Associaçao Brasileira de Estudos do Quaternário (ABEQUA), Guarapari 9–16 Octo-ber 2005.
Giannini PCF, Sawakuchi AO, Martinho CT et al. (2007) Eolian depositional episodes controlled by Late Quaternary relative sea level changes on the Imbituba–Laguna coast (southern Brazil). Marine Geology 237(3–4): 143–168.
Giannini PCF, Villagran XS, Fornari M et al. (2010) Interações entre evolução sedimentar e ocupação humana pré-histórica na costa centro-sul de Santa Catarina, Brasil. Bolteim do Museu Paraense Emílio Goeldi. Série Ciências Humanas 5(1): 105–128.
by guest on July 17, 2014hol.sagepub.comDownloaded from

1016 The Holocene 24(8)
Goldberg P (2000) Micromorphology and site formation at Die Kelders Cave 1, South Africa. Journal of Human Evolution 38: 43–90.
Klokler DM (2008) Food for Body and Soul: Mortuary Ritual in Shell Mounds (Laguna – Brazil). Tucson, AZ: University of Arizona.
Kneip A (2004) O povo da lagoa: uso do SIG para modelamento e simulaçao na área arqueológica do Camacho. PhD Thesis, Universidade de São Paulo.
Laborel J and Laborel-Deguen F (1996) Biological indicators of Holocene sea-level and climatic variations on rocky coasts of tropical and subtropical regions. Quaternary International 31: 53–60.
Lima TA, Macario KD, Anjos RM et al. (2002) The antiquity of the prehistoric settlement of the central-south Brazilian coast. Radiocarbon 44(3): 733–738.
Mannino MA and Thomas KD (2002) Depletion of a resource? The impact of prehistoric human foraging on intertidal mollusc communities and its significance for human settle-ment, mobility and dispersal. World Archaeology 33(3): 452–474.
Martin L and Suguio K (1976) O Quaternário marinho do lito-ral do Estado de São Paulo. In: Anais do XXIX Congresso Brasileiro de Geologia, Ouro Preto: Sociedade Brasileira de Geologia, pp. 281–294.
Martínez S, Rojas A, Ubilla M et al. (2006) Molluscan assem-blages from the marine Holocene of Uruguay: Composition, geochronology, and paleoenvironmental signals. Ameghini-ana 43(2): 385–397.
Martinho CT (2004) Morfodinâmica e sedimentología de campos de dunas transgressivas da região de Jaguaruna-Imbituba, Santa Catarina. São Paulo: Universidade de São Paulo.
Morey DF and Crothers GM (1998) Clearing up clouded waters: Palaeoenvironmental analysis of freshwater mussel assem-blages from the Green River shell middens, western Ken-tucky. Journal of Archaeological Science 25(9): 907–926.
Morey DF, Crothers GM, Stein JK et al. (2002) The fluvial and geomorphic context of Indian Knoll, an Archaic shell midden in West-Central Kentucky. Geoarchaeology 17(6): 521–553.
Narchi W (1972) Comparative study of the functional morphol-ogy of Anomalocardia brasiliana (Gmelin 1791) and Tivela mactroides (Born, 1778) (Bivalvia, Veneridae). Bulletin of Marine Science 22: 643–670.
Nascimento DR (2010) Evoluçao sedimentar holocênica do delta do Rio Tubarão, estado de Santa Catarina. PhD Thesis, Uni-versidade de São Paulo.
Netto SA, Domingos AM and Kurtz MN (2012) Effects of arti-ficial breaching of a temporarily open/closed estuary on ben-thic macroinvertebrates (Camacho Lagoon, Southern Brazil). Estuaries and Coasts 35(4): 1069–1081.
Odin GS (1988) Conclusion to the study of green marine clays. In: Odin GS (ed.) Green Marine Clays. Amsterdam: Elsevier, pp. 399–418.
Odin GS and Fullagar PD (1988) Geological significance of the glaucony facies. In: Odin GS (ed.) Green Marine Clays. Amsterdam: Elsevier, pp. 337–397.
Perreti G (2009) Sambaquis da barreira da Itapeva. Uma per-spectiva geoarqueológica. PhD Thesis, Pontifícia Universi-dade Católica do Rio Grande do Sul.
Rios EC (1985) Seashells of Brazil. Rio Grande: Fundação Cidade do Rio Grande, Fundação Universidade do Rio Grande Museu Oceanográfico.
Sandweiss DH (2003) Terminal Pleistocene through mid-Holocene archaeological sites as paleoclimatic archives for the Peruvian coast. Palaeogeography, Palaeoclimatology, Palaeoecology 194(1–3): 23–40.
Sawakuchi AO, Giannini PCF, Martinho CT et al. (2009) Grain size and heavy minerals of the Late Quaternary eolian sedi-ments from the Imbituba–Jaguaruna coast, Southern Brazil: Depositional controls linked to relative sea-level changes. Sedimentary Geology 222(3–4): 226–240.
Scheel-Ybert R, Afonso MC, Barbosa M et al. (2009) Considera-ções sobre o papel dos sambaquis como indicadores do nível do mar. Quaternary and Environmental Geosciences 01: 3–9.
Stein JK (1992a) The analysis of shell middens. In: Stein JK (ed.) Deciphering a Shell Midden. San Diego, CA: Academic Press, pp. 1–24.
Stein JK (1992b) Deciphering a Shell Midden. New York: Aca-demic Press.
Stein JK, Green D and Sherwood S (2011) Sediment analysis. In: Taylor A and K SJ (eds) Is It a House? Archaeological excavations at English Camp, San Juan Island, Washington. Seattle, WA: University of Washington Press, pp. 45–63.
Suguio K, Martin L and Flexor JM (1992) Paleoshorelines and the sambaquis of Brazil. In: Johnson LL and Stright M (eds) Paleoshorelines and Prehistory. Boca Raton, FL: CRC Press, pp. 83–99.
Van der Schriek T, Passmore DG, Franco F et al. (2008) Holocene palaeoecology and floodplain evolution of the Muge tribu-tary, Lower Tagus Basin, Portugal. Quaternary International 189: 135–151.
Van der Schriek T, Passmore DG, Stevenson AC et al. (2007) The palaeogeography of Mesolithic settlement-subsistence and shell midden formation in the Muge valley, Lower Tagus Basin, Portugal. The Holocene 17(3): 369–385.
Villagran XS, Giannini PCF and DeBlasis P (2009) Archaeofacies analysis: Using depositional attributes to identify anthropic processes of deposition in a monumental shell mound of Santa Catarina State, southern Brazil. Geoarchaeology 24(3): 311–335.
Villagran XS, Klokler D, Nishida P et al. (2010) Lecturas estrati-graficas. arquitetura funeraria y depositación de residuos en el sambaquí Jabuticabeira II. Latin American Antiquity 21(2): 195–216.
Waselkov GA (1987) Shellfish gathering and shell midden archaeology. Advances in Archaeological Method and Theory 10: 93–210.
by guest on July 17, 2014hol.sagepub.comDownloaded from






![some mounds of the Rainy River aspect [by] Lloyd A. Wilford](https://img.pdfslide.net/doc/110x75/635038be2653ab3a9d08dfc6/some-mounds-of-the-rainy-river-aspect-by-lloyd-a-wilford.jpg)






















