Embed Size (px)
Citation preview

SPECIALIZED JUNCTIONS INVOLVED IN ELECTRICAL TRANSMISSION BETWEEN NEURONS. *
George D. Pappasl- and Michael V. L. Bennett? Departments of Anatomy and Neurology,
Columbia University. College of Physicians and Surgeons,
New York, N . Y . and the Marine Biological Laboratory,
Woods Hole, Mass.
Electron microscopic studies of the synaptic junctions have revealed, among other features, that the pre- and postsynaptic processes are separated by an extra- cellular space of 200-250 A (Palay, 1958; De Robertis, 1958). This gap may be slightly wider than that found separating neuronal and glial processes through- out the central nervous system and may contain a material of intermediate density to that of the pre- and postsynaptic membranes (cf. De Robertis, 1964; and Pap- pas, 1966). The synaptic cleft substance sometimes appears as a condensed line or band parallel to the pre- and postsynaptic membranes, or as a series of filaments or striations perpendicular to these membranes. It has been suggested that the synaptic cleft substance is made up of mucopolysaccharide material (Pappas & Purpura, 1966). Mucopolysaccharide material in the synaptic gap might be one reason for the strong cohesion between pre- and postsynaptic membranes, as well as play a role in diffusion and dissipation of transmitter substances.
Electrical transmission has been demonstrated in the central nervous system of several species of fish (Bennett et al., 1966, a,b,c,d; Bennett & Pappas, 1965; Bennett et al., 1963, 1964). Where this mode of transmission occurs, neurons form distinctive morphological junctions (Pappas et al., 1965). For example, the spinal electromotor neurons of the Mormyrid electric fish are electrotonically coupled (Bennett et a]., 1963). In the light microscope, short thick dendrites connect the cells, and these appear to form cytoplasmic continuity of adjacent neurons (at arrows in FIGURE 1). However, using the higher resolving power of the electron microscope, a definite septum between the two dendritic processes can be demonstrated (FIGURE 2). The junction of the two apposing dendrites is formed by the fusion of the apposing plasma membranes, obliterating the extra- cellular space at these sites. The overall measurement of the fused membranes is about 140 A (inset, FIGURE 2) . At high magnification, the fused membranes can be identified as being similar to the “external compound membrane” described by Robertson originally in the myelin sheath and later at club endings on Mauth- ner cells (see Robertson, 1966). These neuronal junctions formed by fusion of adjacent plasma membranes are not found in adjacent areas of the spinal cord where there is no special reason to believe that neurons are electrotonically coupled.
In the electric catfish, two bilaterally-located giant neurons in the medulla control electric organ discharge, and they are closely coupled electrically (Bennett
Supported in part by Public Health Service grants from the National Institute of Neuro- logical Diseases and Blindness (NB-02314-07, NB-03448-05, NB-037-28, and NB-032-70), and the United Cerebral Palsy and Educational Foundation; the National Science Foundation (GB-2940); the Muscular Dystrophy Associations of America; and the Air Force Office of Scientific Research (AF-AFOSR-550-64).
‘r This investigation was supported in part by Public Health Service research career program awards (1-K3-NB-21, 947-01) and (2-K3-GM-5823) from the National Institute of Neuro- logical Diseases and Blindness.
495

496 Annals New York Academy of Sciences
et al., 1966 c). These neurons are separated and apparently do not form direct soma-soma or dendro-dendritic contacts (Bennett et al., 1966 c ) . However, axo- somatic junctions on the giant neurons formed by the fusion of the apposing membranes are of common occurrence (FIGURE 3) . In tangential sections of the junctions, ladderlike striae about 90 A apart as well as an intermediate dense layer or line may be observed in the fused membrane complex (inset, FIGURE 3). The physiological evidence confirms that transmission from excitatory afferents is electrotonic. In addition to these junctions, typical axo-somatic synapses having an intercellular space between the pre- and postsynaptic membranes are present (cf. A2 in FIGURE 3) . These synapses probably mediate certain postsynaptic po- tentials in the giant cells that are apparently inhibitory (Bennett et al., 1966 c) . Since no direct contact between the giant cells can be demonstrated, it is reason-
M FIG [om
URE 1. Light micrograph showing electromotor neurons (N) in the spinal cord iyrid electric fish. Short, thick dendrites connect two of the neurons (at arrows). x
of a 800.

Pappas & Bennett: Specialized Junctions 497
FIGURE 2. Electron micrograph of a section from the spinal cord of a Mormyrid electric fish showing the junction formed by two thick dendrites (D) of electromotor neurons. The two apposing membranes are fused, obliterating the extracellular space at this site. A portion out- lined by the rectangle is shown at higher magnification in the inset. The junctional complex is about 140 A thick (at arrows). An intermediate dense line or layer can be seen as a com- ponent of the fused membrane complex. M = mitochondria. x 42,000; inset = x 300,000.
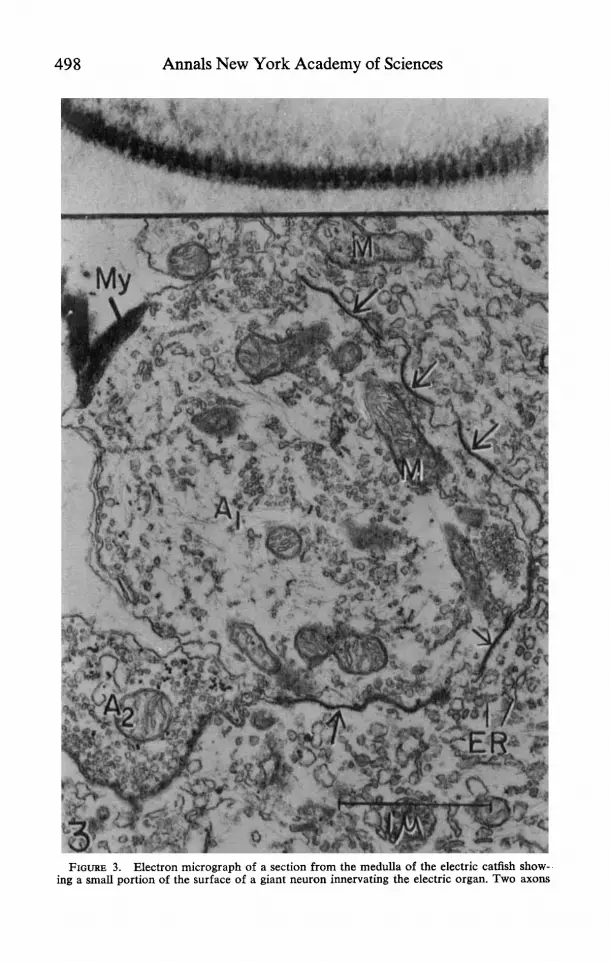
498 Annals New York Academy of Sciences
FIGURE 3. Electron micrograph of a section from the medulla of the electric catfish show- ing a small portion of the surface of a giant neuron innervating the electric organ. Two axons

Pappas & Bennett: Specialized Junctions 499
able to suggest that afferent fibers which form electrotonic junctions with both neurons couple them.
The spinal neurons that innervate the electric organ in the electric eel are electrotonically coupled (Bennett et ul., 1964). The motoneurons are not re- stricted in distribution to a few segments of the spinal cord as are those of the Mormyrids, but rather are found over most of its length. AS in the Mormyrid, the neurons lie in the midline dorsal to the central canal. However, in the electric eel no direct soma-soma or dendro-dendritic contacts are found between them. As in the case of the giant motoneurons of the electric catfish, axo-somatic junc- tions formed by membrane fusion are present. The coupling thus appears to arise by way of the presynaptic fibers, and physiological data indicate that transmission at these junctions is electrotonic. In FIGURE 5, an axon forms a junction with two processes, a dendrite (D) in the upper portion of the micrograph and a neuronal soma (N) in the lower portion. This relationship provides a morphological basis for electrotonic coupling of the two postjunctional elements.
Motoneurons of the toadfish (Opsunus) which innervate the swim bladder muscle fire synchronously at 100-200/ sec for contractions generating sound (Bennett & Pappas, 1965 cf. FIGURE 9 ) . These neurons form a spindle-shaped nucleus in the midline of the spinal cord which is just ventral to the central canal at the level of the first spinal segment. The morphological relations in the nucleus are somewhat more complicated than in the preceding examples. Typical axo- somatic synapses can be found, where a distinct gap between the pre- and post- synaptic membranes is present ( FIGURE 5 ) . Also, axo-somatic junctions are present (FIGURE 6) where the apposing axonal membrane is fused with the plasma membrane of the cell body, obliterating the extracellular space. The fused mem- brane complex is about 140 A thick, and at higher magnification the character- istic appearance of an intermediate line or layer can be seen (inset, FIGURE 6 ) . There are also junctions between cell bodies and processes that appear to be dendrites (FIGURE 5 ) .
The physiological evidence for electrotonic coupling of the swim bladder moto- neurons is somewhat indirect. In graded antidromic stimulation of either swim bladder nerve, short latency graded potentials are evoked in motoneurons, the axons of which do not have a very low threshold (FIGURE 7’). This result is to be expected in a coupled pool of neurons provided the degree of coupling is not very great, for the spike in an antidromically stimulated cell will produce a small depolarization in its inactive neighbors. The possibility that afferents produce these potentials seems excluded, because they persist following dorsal root section and the amplitude is closely correlated with the size of the antidromic volley. That they are electrotonically mediated is indicated by their short latency, which may be determined by subtraction of the extracellular potential obtained at a given stimulus strength from the corresponding intracellular potential (cf, FIGURE 7A). This latency is about 0.3 to 0.4 msec. from the onset of the antidromic spikes, significantly shorter than at known chemically transmitting junctions in cold-blooded forms (see Bennett, 1966).
are abutting onto the neuron. Portions of the plasma membrane of the axon A,, are fused with the apposing neuronal membrane, obliterating the extracellular space at these sites (at arrows). In an oblique section of such a junction ( inse t ) , ladderlike striae about 90 A apart, as well as an intermediate dense layer, can be observed in the fused membrane complex. Axon A2 probably forms a typical synaptic junction with the neuronal soma, where extracellular space is maintained. The synaptic gap is not clearly delineated in this oblique section. Note that the vesicles in axon A, are accumulated at the appositional region. ER = endoplasmic reticulum; My = myelin. M = mitochondria x 32,000; inset X 330,000.

500 Annals New York Academy of Sciences
FIGURE 4. Electron micrograph of a portion of the spinal cord containing neurons that innervate the electric organ of the electric eel. Three elements, an axon (A), a dendrite (D),

Pappas & Bennett: Specialized Junctions 501
FIGURE 5 . Electron micrograph of a small portion of the surface of a spinal motoneuron (N) from the swim bladder nucleus of a toadfish. A dense junction, at the arrow, is formed by the fusion of the apposing plasma membranes of a dendrite (D) and of the neuronal soma (N). The extracellular space is obliterated at this site. In addition, an axon (A) forms a synaptic junction with the neuronal soma (N). This junction is characterized by the presence of extracellular space between the pre- and postsynaptic membranes and by the definite clump- ing of the synaptic vesicles close to the desmosome-like thickening of the presynaptic mem- brane. x 30,000.
It is also possible to record from the prejunctional fibers within the nucleus. These units are identified as afferent fibers, because they fire before the moto- neurons in the orthodromically initiated burst responses that can be evoked by tetanic stimulation of the caudal spinal cord (FIGURE 8B,B’). Very little delay occurs between the fiber spikes and the onset of the postjunctional potentials in the motoneurons, but accurate measurement would be difficult because of the large number of fibers and uncertainty about their course within the nucleus. While the fiber spike is immediately followed by onset of the postjunctional potential in the motoneurons, the threshold of the motoneuron is not reached until after about 1 msec. The prejunctional fibers can also be activated by anti- dromic stimulation of the swim bladder nerves. These responses, however, never have as short a latency as the antidromic responses of the motoneurons. In gen- eral, they appear to be initiated by a graded depolarization that is smaller than in the motoneurons. Frequently, a maximal stimulus to both nerves or paired maximal stimuli to one must be used to excite the fibers. It can also be shown that the level at which the impulse appears to arise is below the firing level of the fiber at the recording site, indicating that the impulse is initiated some distance and a neuronal soma (N) are linked together. The axon forms junctions with both the dendrite and the neuronal soma by the fusion of their apposing membranes (at arrows). GI = glycogen, ER = endoplasmic reticulum. x 40,000. (Taken by Y. Nakajima.)

502 Annals New York Academy of Sciences
from the penetrating electrode. That prefibers may be excited by antidromic activity of the motoneurons is most simply explained by electrotonic coupling of the fibers to the motoneurons. That potentials produced in the fibers by different motoneurons can summate is indicated by the fact that paired stimuli may be required for excitation. These two observations therefore suggest that at least some coupling of the motoneurons should occur by way of the prejunctional pathway. The existence of a direct pathway as well would, however, be difficult to establish
FIGURE 6 . Tissue taken from the same region as FIGURE 5 . An axon containing many mitochondria and vesicles (V) is abutting onto the surface of a neuronal soma (N) . At arrow, the apposing membranes are fused, obliterating the extracellular space. This junction is shown enlarged (at borrom) and is identical in structure to those described in the pre- ceeding electron micrographs. ER = endoplasmic reticulum. X 36,000; bottom inset X 280,000.

Pappas & Bennett: Specialized Junctions 503
- 1 ,4 msec
FIGURE 7. Evidence for electrotonic coupling of swim bladder motoneurons of toadfish. Upper traces: antidromic volleys monopolarly recorded at the cord margin (dorsal roots sectioned). Lower truces: intracellular recordings. Several superimposed sweeps in each record. A . Antidromic stimuli threshold for the penetrated cell. When the antidromic spike did not occur, there was a brief positive-negative potential followed by a prolonged positivity. Photographically superimposed on these records is the potential obtained just extracellular to this cell at the same strength of stimulation. The extracellular record showed only a brief positive-negative potential. Subtraction of this from the intracellular potential gives the trans- membrane potential. Thus when the penetrated cell did not fire, a depolarization of its mem- brane began at a latency about 0.35 msec. longer than that of the antidromic spike. B. As in A, but a t a slower sweep speed to show the time course of the antidromic depolarization. There is also one sweep without stimulation to give the base line. C. As in B, but with graded stimuli to show the graded nature of the antidromic depolarization.
electrophysiologically. While certain physiological observations suggest a dual pathway between cells (cf. Bennett, 1966), the direct pathway is primarily a morphological observation.
There are also hyperpolarizing inhibitory postsynaptic potentials (i.p.s.p.’s) in the motoneurons. The burst responses are preceded by i.p.s.p.’s, and each re- sponse starts from a level of maintained hyperpolarization (FIGURE 8A,B). Hy- perpolarization can also be recorded from some afferent fibers during these bursts (A’), another indication that the fibers are electrotonically coupled to the moto- neurons. The occurrence of i.p.s.p.’s correlates with the presence of typical vesi- cle-packed boutons terminaux on the motoneurons where extracellular space be- tween cells is maintained.
1.p.s.p.’~ can also be evoked in motoneurons by antidromic stimulation of either ipsi- or contralateral swim bladder nerve. In FIGURE 9A a pair of stimuli to the ipsilateral nerve evoked antidromic spikes that were not followed by hyperpolari- zation. The spikes in responses to a pair of maximal stimuli (B) were followed by a hyperpolarization that from A could not have been an after-potential and must therefore have been i.p.s.p. There was also a large efferent volley in the contra- lateral nerve (middle trace). Maximal stimuli to the contralateral nerve produced only small depolarizations when the interval between stimuli was fairly long (C). With a shorter interval between stimuli, the depolarization in response to the

504 Annals New York Academy of Sciences
second stimulus was increased and was followed by a hyperpolarization (D). There was also a small volley in the ipsilateral nerve (middle trace), although the penetrated cell was not excited. The possibility of excitation of the afferent fibers by antidromic activity complicates identification of the pathway mediating the apparently recurrent inhibition, since either collaterals of the afferent fibers or of the motoneurons could be responsible. The threshold for the antidromically evoked inhibition is much higher than observed with Renshaw cell inhibition (cf. Eccles, 1964) and is in the same range as that observed for excitation of the afferents (cf. FIGURE SC’) . These data suggest, therefore, that the apparently recurrent inhibition does involve collaterals of the afferents. A similar situation exists in the electric catfish (Bennett et al., 1966 c ) .
From the foregoing examples, it can be concluded that electrical transmission between neurons takes place by way of morphologically distinct junctions. More- over, differences in the properties of neurons in respect to their electrotonic coupling is reflected in differences in the frequency and distribution of these specialized junctions (cf. Bennett et al., 1966 a,b,c,d).
In the instances of electrical transmission cited here, the junctions serve to synchronize discharge whether coupling is direct or by a prejunctional pathway. It is interesting to note that where electrotonic junctions are formed by axonal processes on cell bodies or dendrites, there are often desmosome-like appositions as well as prejunctional vesicles of the same size-range as those found in junctions where transmission is presumably chemical. However, the vesicles are usually neither as numerous nor as closely clustered to the desmosome-like regions of the
FIGURE 8. Responses of motoneurons and prejunctional fibers to orthodromic and anti- dromic stimulation (toadfish). Upper truces: monopolar recording from the cord margin ipsilateral to the motoneuron, dorsal roots sectioned. Middle truces in A to C: recording from the contralateral swim bladder nerve. Lower truces: intracellular recordings from a motoneuron ( A to C ) or afferent fiber (A’ to C‘), not simultaneous but from the same experiment. A , A’. Tetanic stimuli were given to the caudal spinal cord causing the large artifacts at the beginning of the sweeps. The subsequent burst responses had a frequency of about 185/sec. and could be recorded in both cells, at the cord margin, and in the efferent nerve. There were somewhat more spikes in the response in A . The motoneuron was slightly depolarized during the stimulus and then hyperpolarized ( A ) . The first spike arose from a hyperpolarized level and was preceded by an abortive response. A smaller amount of hyper- polarization was recorded in the afferent fiber ( A ’ ) . B , B’. Expanded sweep of burst responses as in A and A’. In A, a sweep without stimulation was included to show the base line. The motoneuronal spikes arose from a hyperpolarized level. The spikes in the afferent fiber arose at a constant interval before the efferent volley recorded at the cord margin as indicated by the vertical lines. The depolarization in the motoneurons began at about the same time as determined by measurement from the efferent volley (vertical lines). Firing of the moto- neuron was delayed by about 1 msec. from the onset of the depolarization. C . Antidromic stimulation of the motoneuron. C‘. Maximal antidromic activation of the contralateral nerve produced a small depolarization in the fiber. When this stimulus was preceded by a maximal stimulus to the ipsilateral nerve, the fiber was excited.

Pappas & Bennett: Specialized Junctions
A C I I
505
k - 4 I: 20 msec
FIGURE 9. 1.p.s.p.'~ evoked in swim bladder motoneurons by antidromic stimulation (toad- fish). Upper traces: antidromic volley recorded monopolarly at the ipsilateral cord margin. Middle traces, recording from the efferent nerve on the contralateral (A, B) or ipsilateral side ( C , D). Lower traces: intracellular recording from a motonueron. A. A pair of weak antidromic stimuli to the ipsilateral nerve evoked two spikes that were not followed by hyper- polarization. B. A pair of maximal antidromic stimuli were followed by a hyperpolarization, and there was an efferent volley in the contralateral nerve, indicating that previously inactive motoneurons had been excited. C. A pair of maximal antidromic stimuli were given to the contralateral nerve. Each produced a depolarization, the second of which was somewhat larger than the first. D. As in C, but with a shorter interval between stimuli. The response to the second stimulus consisted of a larger depolarization followed by a large prolonged hyper- polarization. There was also an efferent volley in the ipsilateral nerve.
axonal membrane (FIGURES 3, 4, 5, and 6). At such junctions in the catfish (Bennett et al., 1966c), Gymnotids (Bennett et al., 1966d) and probably the Mauthner cell (Furshpan, 1964; see also Bennett et al., 1966d), the physiological evidence indicates that transmission is solely electrical. At these junctions, no functional significance can yet be assigned to the structures associated with chemi- cal transmission elsewhere.
In the ciliary ganglion of the newly-hatched chick, transmission appears to have both chemically and electrically mediated components (Martin & Pilar, 1963). It is interesting that this unique mixed mode of transmission is functionally transitory, for within a few weeks, the electrical component is adequate to initiate impulses in all cells. The persistence of the chemically mediated components has not been determined. Although junctions formed by fused apposing membranes have been described in the newly-hatched chick (Hama, personal communica- tion), such junctions are not obvious in the adult (Hess, 1965). Because of the fact that myelin encloses both the calyx and postjunctional cell in the adult, a low resistance pathway between cells may not be necessary for effective electrical coupling (cf. discussion in Bennett et al., 1966d).
Electrotonic junctions occur not only between neurons of the central nervous system but also in a wide variety of cells (cf. Bennett et al., 1966d). In epithelia, such a junction has been called zonulu occludens (Farquhar & Palade, 1963), and in smooth and cardiac muscle, nexus (Dewey & Barr, 1962, 1964) or fucia occludens (Richardson, 1964).
Wherever measurements have been made, i.e., on neural tissues, cardiac mus- cle, and epithelia, (cf. Bennett, 1966), it has been found that electrotonic junctions are of lower resistivity than the neighboring membrane. The nature of the membrane modification that leads to decreased resistivity remains to be

506 Annals New York Academy of Sciences
FIGURE 10. Passage of fluorescein across the septum of the crayfish septate axon. Micro- graphs of a dorsal view of the abdominal nerve cord and ganglia taken with a fluorescence microscope. The rostral sides of the preparations are on the left of the micrographs. A . A normal preparation. Fluorescein was injected in the caudal axon, A,,, about 3 mm. from the septum, and allowed to diffuse for about one hour. No effect on the action potential was observed. Fluorescein crossed the septum and diffused along the rostral axon, A., although the concentration did not reach as high a level as in the caudal axon. The septum, S , is visible as a demarcation between the two cells. The process labeled C passes caudomedially

Pappas & Bennett: Specialized Junctions 5 07
determined. One approach is to measure the permeability to various ions at these junctions, and a preliminary series of experiments can be mentioned here. Enough fluorescein may be injected into the crayfish septate axon to be visible under ordinary illumination with little effect on impulse conduction. A small amount of this dye, but enough to be readily detected by fluorescent techniques, will pass across the septum in normal preparations (FIGURE 10a, cf. Loewenstein & Kanno for related study on a morphologically very different junction, also discussed in Bennett, 1966). This result is not an artifact of injury. If a segment of an axon is cut open midway between septae, and the preparation allowed to stand, conduction will ultimately fail at both neighboring septae. Under these con- ditions, greatly reduced or even no permeability to fluorescein is noted (FIGURE lob) , and this effect does not depend on whether the dye is injected into the intact or the injured cell. The resistivity of the junctional membranes under these various conditions remains to be determined. The observed permeability to fluorescein appears to be due to an increased permeability to large molecules and not to a general increase in permeability of the same selectivity as the non- junctional membrane. In the latter case, the concentration of fluorescein would decrease along the axon with a space-constant equal to that for electrotonic potentials. In fact, much greater diffusion is observed. The molecular weight of the fluorescein ion is 332, and if the molecule remains planar in solution, its greatest dimension should be just under 10 A. Phenol red, another ionic dye of an only slightly greater molecular weight (346), does not cross the septum in quantities detectable under ordinary illumination and i? only weakly fluores- cent. Whether the permeability is lower than for fluorescein or if the dye is only less detectable has not yet been determined.
The size of the fluorescein molecule is such that pores of sufficient diameter to permit passage should be visible with the electron microscope. However, the density of these pores may be very low. At any rate, the preliminary results indi- cate that the problem could well be pursued with the use of small electron opaque particles. As noted above, cross striations are observed at electrotonic junctions (see FIGURE 3 ) . The permeability of the junctions to such large molecules as fluorescein perhaps lends support to the as yet tenuous hypothesis that these striations represent water-filled channels through the membranes. However, that the striations even represent transverse structures in the membrane has recently been questioned (Robertson, 1966). Moreover, molecules such as fluorescein may not cross the junctions in the same manner as ionic currents, and another mode of communication between cells may occur in these regions.
References BENNETT, M. V. L., E. ALJURE, Y . NAKAJIMA & G. D. PAPPAS. 1963. Electrotonic junctions
between teleost spinal neurons: electrophysiology and ultrastructure. Science 141 : 262. BENNETT, M. V. L., M. GIMENEZ, Y. NAKAJIMA & G. D. PAPPAS. 1964. Spinal and medullary
nuclei controlling electric organ in the eel, Electrophorus. Biol. Bull. 127: 362. BENNETT, M. V. L. & G. D. PAPPAS. 1965. Neurophysiology and ultrastructure of a syn-
chronously firing nucleus in toadfish Opsanus. Fed. Proc. 24: 462.
to form the commissural junction between the right and left axons. B. Preparation in which the membranes of the rostral axons, A,, had been destroyed 2 mm. rostral to the septum about an hour before injection of fluorescein. On the right of the preparation (top of the figure) the uninjured axon caudal to the injured axon was injected. On the left of the preparation (bottom of the figure) the injured axon was injected. Both sites of injection were about 2 mm. from the ganglion. In neither case did a detectable amount of fluorescein cross the septum. The uninjured axons can be seen as faintly outlined dark areas. The commissural process, C, is visible only in the anterior axon that was injected. x 800.

508 Annals New York Academy of Sciences BENNETT, M. V. L., Y . NAKAJIMA & G. D. PAPPAS.
BENNETT, M. V. L., G. D. PAPPAS, E. ALJURE & Y. NAKAJIMA.
1 9 6 6 ~ . Physiology and ultrastructure of electrotonic junctions. I. The supramedullary neurons. (In preparation.)
19666. Physiology and ultra- structure of electrotonic junctions. 11. Spinal and medullary electromotor nuclei in Mormyrid fish. (In preparation.)
1 9 6 6 ~ . Physiology and ultrastructure of electronic junctions. 111. The giant electromotor neurons of Malupterurus electricus. (In preparation.)
1966d. Physiology and ultrastructure of electrotonic junctions. IV. Medullary electromotor nuclei in Gymnotid
BENNETT, M. V. L., Y. NAKAJIMA & G. D. PAPPAS.
BENNETT, M. V. L., G. D. PAPPAS, M. GIMENEZ & Y . NAKAJIMA.
fish. (In preparation.) 1966. BENNETT, M. V. L. Physiology of electrotonic junctions. Ann. New York Acad. Sci.
(This volume.) DE ROBERTIS. E. D. P. 1958. Submicroscouic moroholoav and function of the svnause. - -_ ~~
Exp. CeliRes., Suppl. 5: 347. DE ROBERTIS, E. D. P. 1964. Histophysiology of Synapses and Neurosecretion . Pergamon
Press, New York, N. Y. DEWEY, M. M. & L. BARR.
the nexus. Science 137: 670. DEWEY, M. M. & L. BARR.
J. Cell Biol. 23: 553 . ECCLES, J. C. 1964. The Physiology of Synapses. Academic Press, New York, N. Y. FARQUHAR, M. G. & G. E. PALADE. Junctional complexes in various epithelia, J. Cell
Biol. 17: 375. FURSHPAN, E. J. 1964. Electrical transmission at an excitatory synapse in a vertebrate brain.
Science 144: 878. HESS, A. 1965. Developmental changes in the structure of the synapse on the myelinated cell
bodies of the chick ciliary ganglion. J. Cell Biol. 25: 1. LOEWENSTEIN, W. R. & Y. KANNO. 1964. Studies on an epithelial (gland) cell junction. I.
Modifications of surface membrane permeability. J. Cell Biol. 22: 565-598. MARTIN, A. R. & G. PILAR. 1963. Dual mode of synaptic transmission in the avian ciliary
ganglion. J. Physiol. 168: 443. PALAY, S. L. The morphology of synapses in the central nervous system. Exp. Cell
Res., Suppl. 5: 275. PAPPAS, G. D., Y . NAKAJIMA & M. V. L. BENNETT. The fine structure of electrically
transmitting junctions in the CNS. Anat. Rec. 151 (3) : 397. PAPPAS, G. D. 1966. Electron microscopy of neuronal junctions involved in transmission in
the central nervous system. I n : Nerve As A Tissue. K. Rodahl, Ed. Harper and Row, New York, N. Y. (In press.)
Distribution of colloidal particles in extracellular space and synaptic cleft substance of mammalian cerebral cortex. Nature (In press).
The fine structure of the albino rabbit iris with special reference to the identification of adrenergic and cholinergic nerves and nerve endings in its intrinsic muscles. Am. J. Anat. 114: 173.
Current problems of unit membranes and contact relationships In: Nerve As A Tissue. K. Rodahl, Ed. Harper and Row, New York, N. Y. (In press.)
The transverse density artifact in unit membranes. Ann. New York Acad. Sci. (This volume.)
1962. Intracellular connection between smooth muscle cells:
1964. A study of the structure and distribution of the nexus.
1963.
1958.
1965.
PAPPAS, G. D. & D. P. PURPURA.
RICHARDSON, K. C.
1966.
1964.
ROBERTSON, J. D.
ROBERTSON, J. D.
1966.
1966.





























