Embed Size (px)
Citation preview

Specific Changes in Somatosensory EvokedMagnetic Fields during Recovery from
Sensorimotor StrokeHeidi Wikstrom, MD,*† Risto O. Roine, MD, PhD,† Hannu J. Aronen, MD, PhD,*‡§,
Oili Salonen, MD, PhD,‡ Janne Sinkkonen, MSc,*i Risto J. Ilmoniemi, PhD,*and Juha Huttunen, MD, PhD*¶
We studied recovery-induced changes in the responsiveness of the primary somatosensory cortex in stroke patients withsensory and/or motor symptoms. Somatosensory evoked magnetic fields, in response to median nerve stimulation, wererecorded in 14 patients with their first symptomatic unilateral stroke 1 to 15 days from the first symptoms and again 2to 3 months later. Neuronal activity at the contralateral primary somatosensory cortex was modeled with equivalentcurrent dipoles at the peak latencies of the first two cortical deflections at about 20 msec (N1m) and at 28 to 91 msec(P1m). Twenty-three age-matched healthy volunteers, 9 of whom were tested also in serial recordings, served as controlsubjects. At follow-up, 6 patients showed a significant increase of P1m amplitude, whereas N1m increased only in 1.Clinical improvement of two-point discrimination ability, but not of other basic somatosensory skills, was significantlycorrelated with the increase of P1m. We conclude that the recovery of discriminative touch after stroke is paralleled bythe growth of the P1m somatosensory evoked magnetic field deflection, and we propose that this may reflect re-establishment of lateral inhibitory functions at the primary somatosensory cortex.
Wikstrom H, Roine RO, Aronen HJ, Salonen O, Sinkkonen J, Ilmoniemi RJ, Huttunen J. Specific changes insomatosensory evoked magnetic fields during recovery from sensorimotor stroke. Ann Neurol 2000;47:353–360
The neuronal mechanisms leading to functional recov-ery after cerebral lesion have recently been clarified byseveral experiments in animal models.1–4 In additionto edema resorption and tissue debris removal, “active”neuronal events, such as axonal sprouting5,6 and un-masking of latent connections,7 may underlie allevia-tion of the initial defects. In human stroke patients,functional imaging studies have indicated that fingermovements may be partially overtaken by areas (eg, inthe ipsilateral hemisphere) that are not normally crucialfor lateralized movements.8,9
In the present study, we investigated whether clinicalimprovement of somatosensory functions, initiallycompromised by stroke, is paralleled by changes in theactivation pattern of the somatosensory cortex. Toachieve this, we recorded somatosensory evoked mag-netic fields (SEFs) from the primary somatosensorycortex (SI) in the acute phase of sensorimotor strokeand 2 to 3 months later. In a previous study, weshowed that in the acute phase, abnormalities of theearly SEFs from SI were associated with clinical de-
rangements in somatosensory functions, most notablyin two-point discrimination.10
Patients and MethodsPatients and Healthy ControlsFourteen inpatients at the Stroke Unit of the Helsinki Uni-versity Central Hospital, with their first symptomatic unilat-eral stroke affecting sensorimotor functions, were enrolled inthe study (age, 25–77 years; median age, 56 years; 2 womenand 12 men; 2 left-handed and 12 right-handed; Tables1–3). Patients with known neurological illness before thestroke, severe dysphasia, poor cooperation, or medically un-stable condition were not included; 2 patients with type IIdiabetes mellitus were included. All patients were managedin accordance to the guidelines of the American Heart Asso-ciation for the treatment of acute ischemic stroke; no specialrehabilitation programs were used. Informed consent was ob-tained from all patients; the study protocol was approved bythe Ethics Committees of the Departments of Clinical Neu-rosciences and Radiology of Helsinki University CentralHospital. Control data were obtained from 23 age-matched
From the *BioMag Laboratory, Medical Engineering Centre, and†Departments of †Clinical Neurosciences, ‡Radiology, and ¶Clini-cal Neurophysiology, Helsinki University Central Hospital, Hel-sinki; §Department of Clinical Radiology, Kuopio University Hos-pital, Kuopio; and iCognitive Brain Research Unit, Department ofPsychology, University of Helsinki, Helsinki, Finland.
Received Apr 17, 1999, and in revised form Aug 30 and Nov 10.Accepted for publication Nov 11, 1999.
Address correspondence to Dr Wikstrom, BioMag Laboratory, POBox 508, FIN-00029 HYKS, Finland.
Copyright © 2000 by the American Neurological Association 353

healthy subjects (age, 28–68 years; median age, 59 years; 13women and 10 men; 1 left-handed and 22 right-handed).11
T1- and T2-weighted magnetic resonance imaging (MRI)scans were obtained from each patient, 1 to 16 days afterdisease onset, with a 1.5-T scanner (Siemens Vision 1.5 T;Siemens, Erlangen, Germany). Acute infarcts were character-ized by decreased signal intensity in axial T1-weighted im-ages. In all except 1 patient with a pontine lesion, the in-farcts were in the territory of the middle cerebral artery,comprising both subcortical and cortical–subcortical lesions(see Tables 1–3). Two patients had old asymptomatic lacu-nar lesions. The lesion volumes were calculated from T1-weighted images by using software developed at the Bio-Mag Laboratory. MRI scans were also obtained in allcontrols; 5 initially recruited controls were excluded be-cause of silent lacunar infarcts. All MRI scans were judgedby a neuroradiologist.
Before each SEF recording, the patient was clinically eval-uated. Two-point discrimination ability was tested at thethenar bilaterally. Values exceeding the mean value on theunaffected side by at least 2.5 SDs (35 mm; 2 patients withbilaterally missing two-point discrimination ability were ex-cluded before calculating the mean) were considered abnor-mal. Pain, temperature, vibration, and joint-position senseswere classified as normal or affected. Motor paresis of thehand was classified as present or none. Barthel, ScandanavianStroke Scale, and Rankin indices were determined.
SEF RecordingsEach patient was investigated 1 to 15 days from the firstsymptoms and again 2 to 3 months later. In 9 control sub-jects, stability of the SEFs was studied by recordings in twosessions separated by at least 2 weeks. The recordings wereperformed with a helmet-shaped neuromagnetometer arrayconsisting of 122 planar gradiometers (Neuromag; Helsinki,Finland)12; the subject sat inside a magnetically shieldedroom (Euroshield; Eura, Finland). Both median nerves werealternately stimulated at the wrist with square-wave constantcurrent pulses. Stimulus intensity was just above the motorthreshold of the abductor pollicis brevis muscle. The inter-stimulus interval (ISI) was 3 seconds per nerve. The subjectwatched a video film throughout the recording session. The
signals were bandpass filtered at 0.03 to 310 Hz and sampledat 940 Hz. The analysis time of 400 msec included a 50-msec prestimulus baseline. Two separate averaged responsescontaining 100 to 200 sweeps each were obtained. Theelectro-oculogram was monitored to detect and automaticallyreject sweeps with signals exceeding 150 mV.
Source ModelingIn each subject, the brain was modeled with a sphere bestfitting the inner surface of the skull in the vicinity of thecentral sulcus. The locations, orientations, and activationstrengths of the neuronal generators of the SEFs were esti-mated with equivalent current dipoles (ECDs), which weredetermined with the least squares method, using data from18 to 30 channels covering both field extremes. The result-ing ECD was accepted as a model of local cortical activity ifthe goodness of fit13 of the model with the measured dataexceeded 80%.
Data AnalysisThe first two SEF deflections over SI were analyzed. Theearliest of these, with anteriorly directed current, peaked atabout 20 msec and is designated N1m; the second deflec-tion, with posteriorly directed current, peaked at 28 to 91msec and is designated P1m. The ECD strengths (dipolemoment in nAm) and peak latencies as well as changes inthese parameters between the acute and follow-up phaseswere considered abnormal if they differed from the corre-sponding mean of the control group by more than 2.5 SDs.Differences in dipole location ~Dl 5 ÎDx2 1 Dy2 1 Dz2!between the acute and follow-up MEG recordings were con-sidered abnormal if they exceeded the variability (SD) in di-pole locations between two MEG recordings in the controlgroup by a factor of 2.5 or more. (Table 4; also see Wik-strom and colleagues11). A change in the ECD strength wasnormalized with respect to the larger amplitude value in thetwo sessions. In statistical evaluation, we used the sign testand the V 2 test, which is a modification of the x2 test forsmall sample sizes.
Table 1. Clinical and Experimental Findings in Patients in Whom Two-Point Discrimination Recovered
PatientNo.
Age(yr) Sex Side Days Barthel SSS Rankin
Vol(cm3) Thalamus
InternalCapsule
ParietalLobe
FrontalLobe Other
QN1m QP1m
2-p(mm)
SensorySymptoms
MotorParesisH D H D P/T Vibr Pos
1 Acute 51 M R 3 100 58 2 ,1 x 30 0 29 0 24/` 1 2 2 2Follow-up 100 58 2 20 0 22 25 20/35 1 2 2 2
2 Acute 64 M L 11 55 16 4 76 x xa x Cerebellum 18 0 36 0 9/` 1 1 1 1Follow-up 75 34 4 26 11 33 17 10/35 1 1 1 1
3 Acute 64 M R 3 65 27 3 100 xa x 17 23 44 26 15/50 1 2 2 1Follow-up 100 56 1 18 15 47 59 13/13 2 2 2 1
4 Acute 77 M L 4 70 40 3 9 xb x 23 12 33 15 10/50 1 2 2 1Follow-up NA 50 3 18 18 23 40 20/12 1 2 2 1
5 Acute 63 M R 5 90 54 3 71 xb x x Basal gangliab 12 9 18 15 13/40 1 2 2 1Follow-up 100 58 2 19 18 34 45 15/10 2 2 2 1
6 Acute 48 M L 2 30 NA 4 59 x x x 17 25 19 23 20/` 1 1 1 2Follow-up 100 NA 2 32 15 59 19 15/15 1 1 2 2
Data in the acute phase are presented on the first line and data at the follow-up on the second line. Days 5 time interval from the first symptoms to the initial examination; Vol 5 the infarct volume;x 5 affected areas; a 5 lesion in the primary somatosensory cortex hand area; b 5 old lesion in the left hemisphere; 2-p 5 two-point discrimination ability, nonaffected/affected side; ` 5 completeinability to distinguish between two points; P/T 5 pain/temperature; Vibr 5 vibration; Pos 5 position; 2 5 not affected; 1 5 affected; NA 5 nonavailable data; Q N1m and Q P1m 5 the equivalentcurrent dipole (ECD) strengths of N1m and P1m, respectively, in the healthy (H) and diseased (D) hemispheres. Data in boldface indicate abnormal change in ECD strength between acute phase andfollow-up.
354 Annals of Neurology Vol 47 No 3 March 2000

ResultsThe patients were divided into three groups accordingto clinical impairments: those showing a recovery of aninitially disturbed two-point discrimination (6 patients;see Table 1), those with no recovery of two-point dis-crimination (3 patients; see Table 2), and those inwhom 2-point discrimination was not affected at all (5patients; see Table 3).
In the first group, 4 of the 6 patients had large fron-toparietal infarcts, 1 had a posterior thalamic, and 1 afrontal white matter lesion extending to below the cen-tral sulcus. In 2 patients, the lesion extended into theSI hand area. All these patients had additional somato-sensory defects, and 4 also had motor weakness. At thetime of follow-up, 5 patients showed a significant in-
crease of the initially absent or reduced P1m amplitude(Figs 1–4); in 1 patient, an absent N1m also recov-ered, whereas in the remainder N1m amplitude didnot change. In 1 patient, neither deflection changedsignificantly in the diseased hemisphere, but P1m grewonly in the unaffected hemisphere (in response to stim-ulation of the unaffected hand).
In the second group, 2 of the 3 patients had largefrontoparietal lesions including the internal capsule andextending into the SI hand area in 1, and 1 had a smallinfarct in the frontoparietal junction. In addition totwo-point discrimination, all had additional somato-sensory defects, and 2 also had motor weakness. Onthe affected side, two-point discrimination remainedseverely impaired; in 2 patients, however, the initiallyimpaired two-point discrimination on the “unaffected”side did recover. In 1 of these, P1m grew significantlyin the unaffected hemisphere, and in the other in theaffected hemisphere. The latter patient was the onlyone in whom P1m grew in the affected hemisphere,although two-point discrimination on the affected sidedid not recover. N1m did not change in any of thesepatients.
In the third group with intact two-point discrimina-tion, the lesions were relatively small and subcorticallylocated, affecting the internal capsule in 3, the thala-mus in 2, and the basal ganglia in 4; in 1, only a smallpontine lesion was found. Two patients had exclusively
Table 2. Clinical and Experimental Findings in Patients in Whom Two-Point Discrimination Did Not Recover
Patient No.Age(yr) Sex Side Days Barthel SSS Rankin
Vol(cm3) Thalamus
InternalCapsule
ParietalLobe
FrontalLobe Other
Q N1m Q P1m
2-p(mm)
Sensory Symptoms
MotorParesisH D H D P/T Vibr Pos
7 Acute 65 M L 4 100 54 2 7 x x 57 36 38 25 20/` 1 1 NA 2Follow-up 100 54 1 45 41 22 34 8/` 1 2 2 2
8 Acute 48 M L 2 NA NA 4 50 x xa x 15 0 27 0 `/` 1 1 1 1Follow-up 90 48 3 32 0 76 0 11/ ` 1 1 1 1
9 Acute 56 M R 6 25 18 4 31 x x x 23 0 37 13 `/` 1 1 1 1Follow-up 85 49 3 26 0 37 29 20/` 1 1 1 1
Data in the acute phase are presented on the first line and data at the follow-up on the second line. Days 5 time interval from the first symptoms to the initial examination. Vol 5 the infarct volume;x 5 affected areas; a 5 lesion in the primary somatosensory cortex hand area; 2-p 5 two-point discrimination ability, nonaffected/affected side; ` 5 complete inability to distinguish between two points;P/T 5 pain/temperature; Vibr 5 vibration; Pos 5 position; 2 5 not affected; 1 5 affected; NA 5 nonavailable data; Q N1m and Q P1m 5 the equivalent current dipole (ECD) strengths of N1m andP1m, respectively, in the healthy (H) and diseased (D) hemispheres. Data in boldface indicate an abnormal change in the ECD strength.
Table 3. Clinical and Experimental Findings in Patients in Whom Two-Point Discrimination Was Not Affected
Patient No.Age(yr) Sex Side Days Barthel SSS Rankin
Vol(cm3) Thalamus
InternalCapsule
ParietalLobe
FrontalLobe Other
Q N1m Q P1m
2-p(mm)
SensorySymptoms
MotorParesisH D H D P/T Vibr Pos
10 Acute 56 M L 15 70 47 2 9 x Basal ganglia 16 21 22 16 12/5 2 2 2 1Follow-up 100 52 2 17 17 19 18 16/10 2 2 2 1
11 Acute 48 F L 2 100 58 1 3 x Basal ganglia 3 NA 30 26 16/12 2 2 2 1Follow-up 100 58 1 NA 11 40 34 NA/NA 2 2 2 1
12 Acute 61 M R 4 65 50 3 13 x x x Basal ganglia 9 11 12 0 20/20 1 1 2 1Follow-up 100 56 2 11 10 15 0 20/15 1 1 2 1
13 Acute 52 M L 2 100 58 1 ,1 Pons 8 10 11 16 20/25 1 1 2 1Follow-up 100 58 0 8 4 21 19 12/15 2 2 2 1
14 Acute 26 F L 13 70 42 4 14 x x Basal ganglia 22 16 26 27 10/15 1 2 1 1Follow-up 100 49 2 21 15 37 39 14/15 2 2 2 1
Data in the acute phase are presented on the first line and data at the follow-up on the second line. Days 5 time interval from the first symptoms to the initial examination; Vol 5 the infarct volume;x 5 affected areas; 2-p 5 two-point discrimination ability, nonaffected/affected side; ` 5 complete inability to distinguish between two points; P/T 5 pain/temperature; Vibr 5 vibration; Pos 5position; 2 5 not affected; 1 5 affected; NA 5 nonavailable data; Q N1m and Q P1m 5 the equivalent current dipole (ECD) strengths of N1m and P1m, respectively, in the healthy (H) and diseased(D) hemispheres. There were no significant changes in the ECD strengths in any of these patients.
Table 4. Normal Variability of the N1m and P1m ECDStrength (DQ R), Location (Dl), and Latency (Dt) Changes inSerial Recordings
DQR
(%) Dl (mm) Dt (msec)
N1m 59 16 3P1m 50 17 5
Data are expressed as mean 1 2.5 SD values. Q R was calculated bydividing the absolute value of the difference in dipole moments by thelarger dipole moment.
ECD 5 equivalent current dipole.
Wikstrom et al: SEFs and Recovery from Stroke 355
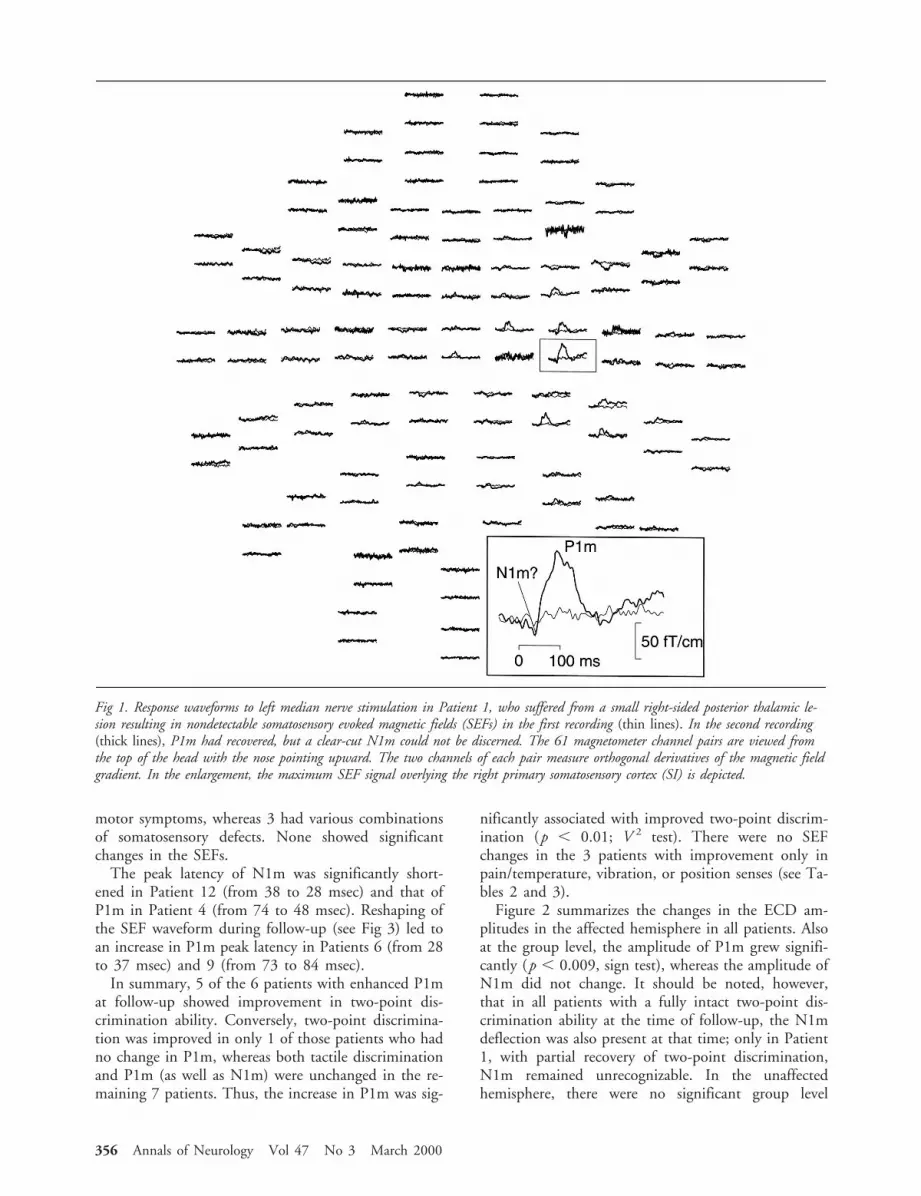
motor symptoms, whereas 3 had various combinationsof somatosensory defects. None showed significantchanges in the SEFs.
The peak latency of N1m was significantly short-ened in Patient 12 (from 38 to 28 msec) and that ofP1m in Patient 4 (from 74 to 48 msec). Reshaping ofthe SEF waveform during follow-up (see Fig 3) led toan increase in P1m peak latency in Patients 6 (from 28to 37 msec) and 9 (from 73 to 84 msec).
In summary, 5 of the 6 patients with enhanced P1mat follow-up showed improvement in two-point dis-crimination ability. Conversely, two-point discrimina-tion was improved in only 1 of those patients who hadno change in P1m, whereas both tactile discriminationand P1m (as well as N1m) were unchanged in the re-maining 7 patients. Thus, the increase in P1m was sig-
nificantly associated with improved two-point discrim-ination (p , 0.01; V 2 test). There were no SEFchanges in the 3 patients with improvement only inpain/temperature, vibration, or position senses (see Ta-bles 2 and 3).
Figure 2 summarizes the changes in the ECD am-plitudes in the affected hemisphere in all patients. Alsoat the group level, the amplitude of P1m grew signifi-cantly (p , 0.009, sign test), whereas the amplitude ofN1m did not change. It should be noted, however,that in all patients with a fully intact two-point dis-crimination ability at the time of follow-up, the N1mdeflection was also present at that time; only in Patient1, with partial recovery of two-point discrimination,N1m remained unrecognizable. In the unaffectedhemisphere, there were no significant group level
Fig 1. Response waveforms to left median nerve stimulation in Patient 1, who suffered from a small right-sided posterior thalamic le-sion resulting in nondetectable somatosensory evoked magnetic fields (SEFs) in the first recording (thin lines). In the second recording(thick lines), P1m had recovered, but a clear-cut N1m could not be discerned. The 61 magnetometer channel pairs are viewed fromthe top of the head with the nose pointing upward. The two channels of each pair measure orthogonal derivatives of the magnetic fieldgradient. In the enlargement, the maximum SEF signal overlying the right primary somatosensory cortex (SI) is depicted.
356 Annals of Neurology Vol 47 No 3 March 2000

changes in either N1m or P1m amplitude (p , 0.4and 0.6, respectively; sign test).
In all except 1 patient (Patient 11; see Table 3) atleast one of the three overall clinical indices improved.At the time of follow-up, 11 patients were independentof outside help in daily tasks (Rankin 0–2). Therewere no significant correlations between improvementin any of the indices and the recovery of either SEFs orsomatosensory abilities.
We did not observe changes in the lateralization ofthe main SEF components during follow-up: no ab-normal deflections appeared at the ipsilateral (unaffect-ed) SI when the affected side was stimulated. In thecontrols, the average ECD location difference~ÎDx2 1 Dy2 1 Dz2! at the SI between the two sessionswas 7.6 mm (SD, 3.3 mm) for N1m and 8.5 mm (SD,3.4 mm) for P1m. In 3 patients, the ECD locationchanges within the affected hemisphere exceeded thenormal limits (P1m, Patients 4, 6, and 9; N1m, Pa-tient 4; see Table 4 for normal values). The directionsof these changes varied without any recognizable asso-ciation with the clinical symptoms and/or type of thelesion, and the ECDs were located close to theV-shaped convolution that characterizes the hand areain the MRI scans.14
DiscussionIn the present study, we monitored poststroke recoveryfrom a relatively acute stage of the disease by means ofclinical evaluation and magnetoencephalographic re-cordings. Although the studied cohort was heteroge-neous with respect to lesion location and size, the sen-sorimotor system was affected in all patients asevidenced by the clinical symptoms. We showed thatthe recovery of discriminative touch was associatedwith an increase in the P1m SEF deflection from SI.Previously, few studies have looked at longitudinalchanges in somatosensory evoked potentials (SEPs) af-ter stroke. In one study, a P45 SEP deflection wasfound to increase along with the recovery of somato-sensory impairments.15 The neuronal generators of theSEPs were not determined in that study; however, thelatency of the P45 SEP deflection that is similar to thatof the P1m SEF, and its opposite polarity comparedwith the N20 SEP deflection, suggest that the P45 SEPand P1m SEF deflections could reflect similar neuronalevents.
The N1m and P1m SEFs are generated in pyramidalcells of Brodmann area 3b.11,16–18 N1m, the first cor-tical deflection, probably reflects excitatory postsynap-tic potentials generated through thalamocortical affer-ents mainly in layer IV neurons.19 The generation
Fig 2. The dipole moments (Q) of the equivalent current di-poles (ECDs) corresponding to N1m and P1m in the acuteand follow-up recordings. P1m grew significantly duringfollow-up (p , 0.009; sign test), whereas N1m did not(p , 0.8). Two-point discrimination recovered in Patients 1to 6 (thick lines) during the follow-up period.
Fig 3. Response waveforms on the affected side in all Patients(1–14). The channel with the strongest signal over the pri-mary somatosensory cortex (SI) in the first recording (thinline) and in the second recording (thick line) is shown. Casesare numbered according to Tables 1 to 3. * 5 two-point dis-crimination ability improved during follow-up. A difference insignal waveform does not necessarily mean a significant differ-ence in the equivalent current dipole (ECD) strengths; differ-ences paralleled by significant ECD strength changes areshaded.
Wikstrom et al: SEFs and Recovery from Stroke 357

mechanisms of P1m are less clear. The source locationof P1m differs slightly from that of N1m.17,19 Further-more, the two deflections have functional differences,eg, in the ISI dependence. P1m starts to diminish withshortening of ISI in the range of seconds, whereasN1m remains stable until the ISI is reduced to a fewhundred milliseconds.19 This ISI dependence of P1mis similar to the ISI behavior of inhibitory postsynapticpotentials (IPSPs) in the somatosensory cortex.20–22
The direction of current flow during P1m is consistentwith IPSPs generated near the somata of area 3b pyra-midal neurons in deep cortical layers.19 We have there-fore suggested that P1m reflects postexcitatory IPSPs inarea 3b.19
Experimentally induced small cortical lesions in themonkey area 3b have been reported to cause rearrange-ments in the scale of hundreds of micrometers in thelocations of neuron populations that can be activatedby stimulating a given skin area.23 Also, the relearningof sensorimotor skills after removal of the task-relevantpart of area 3b was reported to be associated with ma-jor rearrangements in S1, including the doubling ofarea 1 representation of behaviorally important skinsurfaces.4 Corresponding recovery-related plasticity inhuman somatosensory cortical areas has not been dem-onstrated. However, the sizes of cortical representationsof different body parts in human SI have been sug-gested to change both in physiological conditions24
and after arm amputation.25 In the present study, weobserved significant ECD location shifts in 3 patients.In 2 of these, however, major tissue swelling waspresent in the first recording, which probably had sig-nificant influence on ECD coordinates. Thus, we can-not consider the ECD location shifts to result from thereshaping of sensory maps. In addition to shifts in re-ceptive field locations, cortical reorganization could
also consist of concentric enlargements of receptivefields. When modeling neuronal activity with an ECDin such a case, the enlargement of activated corticalarea would be reflected as an increase in ECDstrength.26 It is possible that in the patients with en-hanced P1m, regardless of whether P1m reflects inhi-bition or excitation, the cortical area generating P1mhad, in fact, been increased.
In our patients, the SI cortex was either directly in-volved in the ischemic lesion (3 patients) or was par-tially deafferented (11 patients) by lesions extending inthe subcortical white matter, the thalamus, or the pons.Several studies have shown that both deafferenta-tion6,27,28 and ischemia3,29,30 acutely lead to impair-ment of inhibition in the affected cortical region. Hy-perexcitability is also typical of cerebral cortex adjacentto a lesion during the first week after lesion induc-tion.1,2 Clinically, pathological reflexes and mass move-ments, as well as the increased incidence of epilepsyafter stroke, can be seen as manifestations of defectiveinhibitory functions. Discriminative touch, measuredas two-point discrimination ability, requires small exci-tatory receptive fields in the cortical neurons responsi-ble for this function, such as those found in area 3bpyramidal neurons.31 The maintenance of small recep-tive fields, in turn, relies on properly functioning lat-eral inhibitory connections.32 Hence, a defect in (lat-eral) inhibition is expected to cause impairments indiscriminative sensory functions, the recovery of whichcould depend on successful re-establishment of inhibi-tion. In the present study, the normalization of P1m inpatients with improved discriminative touch could bea neurophysiological sign of the re-establishment ofIPSPs in the deafferented or partially lesioned SI.
The processes underlying the normalization of P1m,however, may not alone suffice to produce recovery of
Fig 4. Isofield contour maps and magnetic resonance imaging (MRI) scans showing the equivalent current dipole (ECD) locationscorresponding to P1m in the first (circle) and in the follow-up recording (square) in Patient 5, in whom the right parietal cortexand white matter immediately caudal to the central sulcus were damaged. Dashed lines represent magnetic flux entering and solidlines exiting the head. The contour line separation is 20 fT/cm. In the field map, the arrow illustrates the location and orientationof the ECD. In the MRI slices, the arrow points to the infarcted area.
358 Annals of Neurology Vol 47 No 3 March 2000

two-point discrimination ability. In Patient 6, the ini-tially normal P1m did not grow despite clinical im-provement; and in Patient 9, P1m increased withoutrecovery of two-point discrimination ability. In bothpatients, the lesions extended to the opercular cortex,which contains several areas responsive to somatic stim-ulation.33 Opercular lesions have, indeed, been shownto produce disturbances in somatosensory task perfor-mance.34 It is possible that both SI and opercular so-matosensory areas are needed for an intact two-pointdiscrimination. In Patient 6, the clinical recovery couldhave been because of plastic changes in the survivedportion of the operculum; whereas in Patient 9, oper-cular areas may have been permanently severed so thatthe recovery at SI (P1m) was not sufficient to improveperformance. Taken together, the cases of Patients 6and 9 indicate that considerable interindividual varia-tion may occur after cerebral damage.
Recent functional imaging studies have emphasizedthe possibility of some lateralized motor functions tobe overtaken by the undamaged hemisphere.8,9,35 Thepresent study does not provide evidence to extendthese findings to the somatosensory system, because noabnormal SEFs were observed from the ipsilateral SIwhen the affected hand was stimulated. However, ourfinding that P1m grew significantly in the undamagedhemisphere of 2 patients (when the unaffected handwas stimulated) implies that changes of excitability mayoccur also on the “intact” side. These findings are inaccord with reports of enhanced SEPs in the unaffectedhemisphere in occasional stroke patients.36 Both of ourpatients had relatively large cortical–subcortical in-farcts. Although we cannot determine what caused theexcitability change in the unaffected SI, one possibilityis that in these patients the callosal connections be-tween SI hand areas were severed more than in theother patients, possibly leading to changes in tonic in-tercallosal signaling.
ConclusionThe present results showed a correlation between achange in an electrophysiological parameter reflectingSI function and in somatosensory task performance.Magnetoencephalography allows for a relatively accu-rate determination of source currents at SI. Althoughthe clinical correlates of SEFs are not yet fully under-stood, SEF monitoring may prove valuable in the eval-uation of specific rehabilitation programs designed torestore somatosensory functions.
This study was supported by the Instrumentarium Science Founda-tion, Klingendahl Foundation, the Academy of Finland, the Radio-logical Society of Finland, Duodecim Foundation, Paavo NurmiFoundation, and research grants (TYH-8102 and TYH-9102) fromthe Helsinki University Central Hospital.
We thank Suvi Heikkila for excellent technical assistance and JussiVirkkala for software development.
References1. Buchkremer-Ratzmann I, Witte OW. Extended brain disinhi-
bition following small photothrombotic lesions in rat frontalcortex. Neuroreport 1997;8:519–522
2. Hagemann G, Redecker C, Neumann-Haefelin T, et al. In-creased long-term potentiation in the surround of experimen-tally induced focal cortical infarction. Ann Neurol 1998;44:255–258
3. Sober SJ, Stark JM, Yamasaki DS, Lytton WW. Receptive fieldchanges after strokelike cortical ablation: a role for activationdynamics. J Neurophysiol 1997;78:3438–3443
4. Xerri C, Merzenich MM, Peterson BE, Jenkins W. Plasticity ofprimary somatosensory cortex paralleling sensorimotor skill re-covery from stroke in adult monkeys. J Neurophysiol 1998;79:2119–2148
5. Raisman G, Field PM. A quantitative investigation of the de-velopment of contralateral reinnervation after partial deafferen-tation of the septal nuclei. Brain Res 1973;50:241–264
6. Wall PD, Egger MD. Formation of new connexions in adultrat brains after partial deafferentation. Nature 1971;232:542–545
7. Wall PD. Mechanisms of plasticity of connection followingdamage in adult mammalian nervous system. In: Bach-y-Rita P,ed. Recovery of function: theoretical considerations for braininjury rehabilitation. Bern: Hans Huber, 1980:91–106
8. Weiller C, Chollet F, Friston KJ, et al. Functional reorganiza-tion of the brain in recovery from striatocapsular infarction inman. Ann Neurol 1992;31:463–472
9. Cramer SC, Nelles G, Benson RR, et al. A functional MRIstudy of subjects recovered from hemiparetic stroke. Stroke1997;28:2518–2527
10. Wikstrom H, Roine RO, Salonen O, et al. Somatosensoryevoked magnetic fields from the primary somatosensory cortex(SI) in acute stroke. Clin Neurophysiol 1999;110:916–923
11. Wikstrom H, Roine RO, Salonen O, et al. Somatosensoryevoked magnetic fields to median nerve stimulation: interhemi-spheric differences in a normal population. ElectroencephalogrClin Neurophysiol 1997;104:480–487
12. Ahonen AI, Hamalainen MS, Knuutila JET, et al. 122-channelSQUID instrument for investigating the magnetic signals fromthe human brain. Physica Scripta 1993;T49:198–205
13. Hamalainen M, Hari R, Ilmoniemi RJ, et al. Magnetoencepha-lography: theory, instrumentation, and applications to noninva-sive studies of the working human brain. Rev Mod Phys 1993;65:413–497
14. Puce A, Constable RT, Luby ML, et al. Functional magneticresonance imaging of sensory and motor cortex: comparisonwith electrophysiological localization. J Neurosurg 1995;83:262–270
15. Macdonell RA, Donnan GA, Bladin PF. Serial changes in so-matosensory evoked potentials following cerebral infarction.Electroencephalogr Clin Neurophysiol 1991;80:276–283
16. Allison T, McCarthy G, Wood CC, et al. Human cortical po-tentials evoked by stimulation of the median nerve. I. Cytoar-chitectonic areas generating short-latency activity. J Neuro-physiol 1989;62:694–710
17. Tiihonen J, Hari R, Hamalainen M. Early deflections of cere-bral magnetic responses to median nerve stimulation. Electro-encephalogr Clin Neurophysiol 1989;74:290–296
18. Wood CC, Cohen D, Cuffin BN, et al. Electrical sources inhuman somatosensory cortex: identification by combined mag-netic and potential recordings. Science 1985;227:1051–1053
19. Wikstrom H, Huttunen J, Korvenoja A, et al. Effects of inter-
Wikstrom et al: SEFs and Recovery from Stroke 359

stimulus interval on somatosensory evoked magnetic fields(SEFs): a hypothesis concerning SEF generation at the primarysensorimotor cortex. Electroencephalogr Clin Neurophysiol1996;100:479–487
20. Deisz RA, Prince DA. Frequency-dependent depression of inhi-bition in guinea-pig neocortex in vitro by GABAB receptor feed-back on GABA release. J Physiol (Lond) 1989;412:513–541
21. Hellweg F-C, Schultz W, Creutzfeldt OD. Extracellular and in-tracellular recordings from cat’s cortical whisker projection area:thalamocortical response transformation. J Neurophysiol 1977;40:463–479
22. Nacimiento AC, Lux HD, Creutzfeldt OD. PostsynaptischePotentiale von Nervenzellen des motorischen Cortex nach elec-trischer Reizung spezifischer und unspezifischer Thalamuskerne.Pflugers Arch 1964;281:152–169
23. Jenkins WM, Merzenich MM. Reorganization of neocorticalrepresentations after brain injury: a neurophysiological model ofthe bases of recovery from stroke. Prog Brain Res 1987;71:249–266
24. Elbert T, Pantev C, Wienbruch C, et al. Increased cortical rep-resentation of the fingers of the left hand in string players. Sci-ence 1995;270:305–307
25. Ramachandran VS. Behavioral and magnetoencephalographiccorrelates of plasticity in the adult human brain. Proc NatlAcad Sci USA 1993;90:10413–10420
26. Okada Y. Discrimination of localized and distributed currentdipole sources and localized single and multiple sources. In:Weinberg H, Stroink G, Katila T, ed. Biomagnetism: applica-tions and theory. New York: Pergamon Press, 1985:266–272
27. Dykes RW, Lamour Y. An electrophysiological laminar analysisof single somatosensory neurons in partially deafferented rat
hindlimb granular cortex subsequent to transection of the sci-atic nerve. Brain Res 1988;449:1–17
28. Ziemann U, Hallett M, Cohen LG. Mechanisms of deaffer-entation-induced plasticity in human motor cortex. J Neurosci1998;18:7000–7007
29. Krnjevic K, Xu YZ, Zhang L. Anoxic block of GABAergicIPSPs. Neurochem Res 1991;3:279–284
30. Luhmann HJ, Kral T, Heinemann U. Influence of hypoxia onexcitation and GABAergic inhibition in mature and developingrat neocortex. Exp Brain Res 1993;97:209–224
31. Kaas JH. The functional organization of somatosensory cortexin primates. Ann Anat 1993;175:509–518
32. Dykes RW, Landry P, Metherate R, Hicks TP. Functional roleof GABA in cat primary somatosensory cortex: shaping recep-tive fields of cortical neurons. J Neurophysiol 1984;52:1066–1093
33. Burton H, Robinson CJ. Organization of the SII parietalcortex: multiple representations within and near second somaticsensory area of cynomolgus monkeys. In: Woolsey CN, ed.Cortical sensory organization. Clifton, NJ: Humana Press,1981:67–114
34. Caselli RJ. Ventrolateral and dorsomedial somatosensory asso-ciation cortex damage produces distinct somesthetic syndromesin humans. Neurology 1993;43:762–771
35. Cao Y, D’Olhaberriague L, Vikingstad EM, et al. Pilot study offunctional MRI to assess cerebral activation of motor functionafter poststroke hemiparesis. Stroke 1998;29:112–122
36. Nakashima K, Kanba M, Fujimoto K, et al. Somatosensoryevoked potentials over the non-affected hemisphere in patientswith unilateral cerebrovascular lesions. J Neurol Sci 1985;70:117–127
360 Annals of Neurology Vol 47 No 3 March 2000





























