Embed Size (px)
Citation preview

Stimulated Tyrosine Phosphorylation of Phosphatidylinositol3-Kinase Causes Acidic pH-Induced Contraction inSpontaneously Hypertensive Rat Aorta
DILEEP KUMAR ROHRA, TOHRU YAMAKUNI, KEN-ICHI FURUKAWA, NOE ISHII, TAKASHI SHINKAWA,TOSHIAKI ISOBE, and YASUSHI OHIZUMI
Department of Pharmaceutical Molecular Biology, Graduate School of Pharmaceutical Sciences, Tohoku University, Sendai, Japan (D.K.R.,T.Y., N.I., Y.O.); Department of Pharmacology, Hirosaki University School of Medicine, Hirosaki, Japan (K.I.F.); Integrated Proteomics SystemProject, Poineer Research on Genome the Frontier, MEXT, c/o Department of Chemistry, Graduate School of Science, Tokyo MetropolitanUniversity, Tokyo, Japan (T.S., T.I.); and Department of Chemistry, Graduate School of Science, Tokyo Metropolitan University, Tokyo, Japan(T.I.)
Received July 16, 2002; accepted September 4, 2002
ABSTRACTAcidic pH induced a contraction (APIC) in isolated aortas fromspontaneously hypertensive (SHR) and Wistar Kyoto rats, butfailed to produce any response in age-matched Wistar rataorta. This study was conducted to test the hypothesis thattyrosine phosphorylation of proteins is a molecular mechanismunderlying the APIC. Tyrosine kinase inhibitors, genistein andtyrphostin 23 inhibited the APIC in a concentration-dependentmanner. APIC was inhibited by phosphatidylinositol 3-kinase(PI3-kinase) inhibitors, LY-294002 [2-(4-morpholinyl)-8-phenyl-4H-1-benzopyran-4-one hydrochloride] and wortmannin. Con-sistent with the results from tension measurement experiments,Western blot analysis showed that acidic pH induced an ap-preciable increment of tyrosine phosphorylation of 85-kDa pro-tein (p85) in SHR aorta, which was completely inhibited bytyrphostin 23, whereas in Wistar rat aorta, the protein tyrosinephosphorylation was not observed. Further investigations usingimmunoprecipitation followed by Western blotting confirmed
an increase in the tyrosine phosphorylation of p85. Analysis bySDS-polyacrylamide gel electrophoresis followed by silverstaining of the gel revealed that amounts of multiple proteinswith molecular sizes of 120, 130, 210, and 225 kDa wereincreased at acidic pH, which were immunoprecipitated withanti-phosphotyrosine antibody. Western blotting using a spe-cific anti-PI3-kinase antibody identified the p85 as the regula-tory subunit of PI3-kinase, whereas 120-, 130-, and 225-kDaproteins were identified by mass spectrometry as pro-�2 (I)collagen, collagen �1 (I) chain, and fibernectin I, respectively.As assayed by Western blotting using anti-myosin light chain(MLC) antibody, acidic pH induced a stimulation of MLC phos-phorylation, and the stimulated MLC phosphorylation was abol-ished by tyrphostin 23 and LY-294002. These results suggestthat acidic pH induces an increase in tyrosine phosphorylationof PI3-kinase, resulting in the MLC phosphorylation-dependentcontraction of SHR aorta.
Multiple signaling pathways have been shown to be in-volved in the mechanism of vasoconstriction (Somlyo andSomlyo, 1998). Ca2� is a major determinant of contractileforce in all types of vascular smooth muscle (VSM). Apartfrom that, there is accumulating evidence for additional reg-ulatory mechanism(s) in smooth muscle contraction (for re-view, see Horowitz et al., 1996). Tyrosine kinase-mediated
pathways have also been shown to cause agonist-stimulatedVSM contractile processes (Khalil et al., 1995; Banes et al.,1999). Consistently, tyrosine kinase inhibitors have beenshown to attenuate vasoconstriction caused by a number ofagents, including noradrenaline and angiotensin II in intactarteries (Di Salvo et al., 1993, 1994; Laniyonu et al., 1994;Carmines et al., 2001). Recent studies have shown that thevoltage-dependent Ca2� channels (VDCCs), activated bymany vasoactive agents, are inhibited by tyrosine kinaseinhibitors via tyrosine kinases (Wijetunge et al., 1992; Wije-tunge et al., 1998; Lagaud et al., 1999). In addition to theeffects on Ca2� channels, tyrosine kinases may regulate
This work was supported in part by a grant-in-aid for Scientific Researchfrom the Ministry of Education, Science and Culture of Japan.
Article, publication date, and citation information can be found athttp://jpet.aspetjournals.org.
DOI: 10.1124/jpet.102.041475.
ABBREVIATIONS: VSM, vascular smooth muscle; VDCC, voltage-dependent calcium channel; APIC, acidic pH-induced contraction; SHR,spontaneously hypertensive rat; WKY, Wistar Kyoto; PI3-kinase, phosphatidylinositol 3-kinase; PSS, physiological salt solution; BSA, bovineserum albumin; PAGE, polyacrylamide gel electrophoresis; HRP, horseradish peroxidase; MLC, myosin light chain; DTT, dithiothreitol; MS/MS,tandem mass spectrometry; PE, phenylephrine; p85, 85-kDa protein; LY-294002, 2-(4-morpholinyl)-8-phenyl-4H-1-benzopyran-4-one hydrochlo-ride.
0022-3565/02/3033-1255–1264$7.00THE JOURNAL OF PHARMACOLOGY AND EXPERIMENTAL THERAPEUTICS Vol. 303, No. 3Copyright © 2002 by The American Society for Pharmacology and Experimental Therapeutics 41475/1026544JPET 303:1255–1264, 2002 Printed in U.S.A.
1255
at ASPE
T Journals on A
ugust 12, 2016jpet.aspetjournals.org
Dow
nloaded from

other mechanisms controlling the contractile state of smoothmuscle cells. For example, mitogen-activated protein kinase,which itself is partly activated by tyrosine phosphorylation,has been suggested to modulate the activity of the actin- andmyosin-binding protein caldesmon (Adam et al., 1995).
The contractile state of the VSM can be modified by severalfactors, including pH (Chen and Rembold, 1995). Ischemiaand metabolic disorders, such as diabetes mellitus, causevarious disturbances, including hypoxia and acidosis in thecirculatory system (Levine, 1993; Leach et al., 1998; Komu-kai et al., 1998). During ischemia or hypoxia, alterations inpH may play a significant role in the changes in vessel wallfunction (Nagesetty and Paul, 1994; Nishiyama et al., 1999).Although physiological pH of the body fluids is maintained ataround 7.4, ischemia easily decreases the pH value signifi-cantly (Butwell et al., 1993). We have described previouslythat acidic pH induces a contraction (APIC) in spontaneouslyhypertensive (SHR) and Wistar Kyoto (WKY) rat aortas (Fu-rukawa et al., 1996); however, the molecular mechanismunderlying the APIC is not clear. We have also observed thatacidic pH fails to produce any contractile response in Wistarrat aorta. This study was conducted to test the hypothesisthat tyrosine phosphorylation of proteins may be a differencebetween SHR and Wistar aortas in the sensitivity towardacidic pH. In this article, we present the first direct evidencethat acidic pH stimulates the tyrosine phosphorylation ofphosphatidylinositol 3-kinase (PI3-kinase) in isolated aortafrom SHR but not in that from Wistar rat, and this enhancedtyrosine phosphorylation is the major mechanism underlyingthe contractile response to acidic pH.
Materials and MethodsMeasurement of Isometric Tension. Male SHR and WKY, both
NCrj strain, and Wistar rats (all strains 13–14 weeks old) werestunned and killed by cervical dislocation. Thoracic aorta was re-moved and placed in ice-cold physiological salt solution (PSS) of thefollowing composition: 120 mM NaCl, 4.8 mM KCl, 1.3 mM MgSO4,1.2 mM CaCl2, 25.5 mM NaHCO3, 5.8 mM glucose, 1.2 mM KH2PO4,and 20 mM HEPES. Aorta was dissected into helical strips (0.2 �1.7–1.8 cm). The endothelial cell layer was removed by rubbing theluminal side of the vessel with a moistened cotton swab. Tissueswere placed in organ baths filled with warmed (37°C) and aerated(95% O2, 5% CO2) PSS. pH of the bathing solution was strictlymaintained at 7.4. One end of the preparation was attached to astainless steel rod and the other was attached to a force transducer(Nihon Kohden, Tokyo, Japan). The tissues were then placed underresting tension of 10 mN. Changes in isometric force were recordedon a polygraph. After equilibration for 1 h, the lack of the endothe-lium was checked by observing the failure of 1 �M carbachol toinduce relaxation in tissues contracted with 1 �M phenylephrine.After washing multiple times, the tissues were equilibrated by con-tracting with 64.8 mM KCl three times and the last contractileresponse was taken as a standard and later all the contractions werenormalized with it. Acidic solutions were prepared by the addition ofHCl to PSS and pH of the bathing solution was changed from 7.4 todesired pH by simply replacing the bathing PSS. For experiments inwhich inhibitors of tyrosine kinase and PI3-kinase were used, tissueswere incubated with these compounds for 20 min before the induc-tion of contraction with acidic pH or other agonists. In control, 0.1%dimethyl sulfoxide was used instead. When using different concen-trations of the drugs, separate tissue was used for each concentra-tion.
Tyrosine Phosphorylation Assay. Analyses of the expression oftyrosine-phosphorylated proteins in SHR and Wistar rat aorta were
conducted by Western blotting as follows. Helical strips (0.3 � 2.5cm) of aorta with wet weight of approximately 12 mg were treatedaccording to the same procedure as for tension recording experi-ments. Thereafter, the aortic strips were immediately frozen in liq-uid nitrogen and kept at �80°C until subsequently used.
Segments of aorta were homogenized in 10 mM HEPES buffer, pH7.5, containing 1 mM EDTA, protease inhibitors [leupeptin, anti-paine, pepstatin A, chymostatin, and phosphoramidon (all 10 �gml�1), and 250 �M phenylmethylsulfonyl fluoride] and phosphataseinhibitors (0.32 �M okadaic acid, 10 nM calyculin A, 10 mM sodiumfluoride, 50 �M bisperoxo(1,10-phenanthroline)oxovanadate, and 1mM sodium orthovanadate) at 4°C using a glass-glass tissue homog-enizer. Then 10% SDS was added in the homogenate (final concen-tration of SDS was 1%), mixed, and boiled for 5 min. After centrifu-gation (18,000g) for 30 min, the supernatant was used as a tissueextract. Protein content was determined by BCA protein analysis kit(Pierce Chemical, Rockford, IL) using bovine serum albumin (BSA)as a standard. Proteins (50 �g) in each sample were separated bySDS-PAGE (7.5% gel) and transferred to polyvinylidene difluoridemembrane (Bio-Rad, Hercules, CA). Blotted membrane was blockedin 5% BSA in Tris-buffered saline containing 0.1% Tween 20 for 2 hat room temperature. The membrane was incubated with the anti-phosphotyrosine antibody (Zymed Laboratories, South San Fran-cisco, CA), in a dilution of 1:1000 overnight at 4°C. After washing,the membrane was reacted with anti-rabbit IgG antibody linked tohorseradish peroxidase (HRP) (Cell Signaling Technology Inc., Bev-erly, MA) in a dilution of 1:2000 for 1 h at room temperature. Afterincubation with enhanced chemiluminescence reagent (Pierce Chem-ical), immunoreactive proteins were visualized by exposing the blotsto Hyperfilm ECL (Amersham Biosciences, Buckinghamshire, UK).
Myosin Light Chain (MLC) Phosphorylation Assay. MLCphosphorylation assay using glycerol-PAGE followed by Westernblotting was performed as follows. The frozen tissues were treatedwith dry ice-cold acetone containing 10% trichloroacetic acid and 10mM dithiothreitol (DTT). Trichloroacetic acid was washed out withacetone-10 mM DTT. The tissues were mixed with urea-samplebuffer (20 mM Tris, 22 mM glycine, 10 mM DTT, 8.3 M urea, and0.1% bromphenol blue) and allowed to stand at room temperature for1 h, while vigorously shaking the mixture intermittently. The sam-ples were then filtered through a 0.45 �M membrane filter (MilliporeCorporation, Bedford, MA), and the filtrates were subjected to gly-cerol-PAGE. Proteins were transferred to polyvinylidene difluoridemembrane. The blotted membrane was blocked in 1% BSA in phos-phate-buffered saline containing 0.1% Tween 20 for 2 h at roomtemperature. Later, the blot was incubated with monoclonal anti-MLC antibody (Sigma-Aldrich, St. Louis, MO) in a dilution of 1:200overnight at 4°C. After washing with phosphate-buffered saline con-taining 0.1% Tween 20, the blot was incubated with anti-mouse IgGlinked to HRP (Sigma-Aldrich) in a dilution of 1:2000 for 90 min atroom temperature. Immunoreactive proteins were visualized as de-scribed above.
Immunoprecipitation Followed by Western Blotting. Anti-phosphotyrosine antibody (5 �g) was added to tissue extract contain-ing 150 �g of protein and mixed by rotation at room temperature for2 h. Then, 40 �l of protein A-Sepharose (50% slurry) was added toeach sample and mixed by rotation at room temperature for 1 h. Thesamples were centrifuged (5000g) for 1 min and the supernatant wasdiscarded. The pellet was washed with 10 mM HEPES containing 1%Nonidet P-40 five times, mixed with sample buffer, and heated at100°C for 5 min. The mixture was centrifuged (12,000g) for 5 minand the supernatant was subjected to SDS-PAGE, as describedabove. The blot was blocked in blocking solution (5% skimmed milkin Tris-buffered saline containing 0.05% Tween 20 for PI3-kinaseand 3% skimmed milk in phosphate-buffered saline for cortactin) for20 min at room temperature and incubated with either 0.8 �g/mlanti-PI3-kinase p85� antibody (Santa Cruz Biotechnology, Inc.,Santa Cruz, CA), or 0.5 �g/ml anti-cortactin (p80/85), clone 4F11antibody (Upstate Biotechnology, Lake Placid, NY), for 2 h at room
1256 Rohra et al.
at ASPE
T Journals on A
ugust 12, 2016jpet.aspetjournals.org
Dow
nloaded from
temperature. After washing, the blot was incubated with eitheranti-rabbit IgG for PI3-kinase or anti-mouse IgG for cortactin, bothlinked to HRP, in a dilution of 1:2000 for 45 min at room tempera-ture. The immunoreactive proteins were visualized as describedabove.
Identification of Proteins by Mass Spectrometry. To identifythe proteins undergoing tyrosine phosphorylation at acidic pH bymass spectrometry, tissue extract immunoprecipitated with anti-phosphotyrosine antibody was subjected to SDS-PAGE and the gelwas silver-stained. Silver-stained protein bands were excised fromthe gel, in-gel digested with trypsin, and subjected to nano-flowliquid chromatography-tandem mass spectrometry (MS/MS) analy-sis for protein identification. The chromatography was performed ona nano ESI column (inside diameter, 120 �m � 30 mm) packed witha C18 reversed phase medium (Mightysil-C18; Kanto Chemicals,Tokyo, Japan) using a linear gradient from 0 to 70% acetonitrile in0.1% formic acid at 100 nl/min, and the separated peptides weredirectly sprayed into a hybrid mass spectrometer equipped with anelectrospray source (Q-T of 2; Micromass, Manchester, UK). Electro-spray ionization was carried out at a voltage of 1.5 kV, and MS/MSspectra were automatically acquired in data-dependent mode duringthe entire run. All MS/MS spectra were correlated by search engine,Mascot program (Matrixscience, London, UK), against the nonre-
dundant protein sequence database at the National Center for Bio-technology Information (National Institutes of Health). Each high-scoring peptide sequence was confirmed by manual inspection of thecorresponding MS/MS spectrum to ensure that the match was cor-rect.
Materials. SHR and WKY were purchased from Charles River(Kanagawa, Japan) and Wistar rats were from Kumagai (Sendai,Japan). Tyrphostin 23 and phenylephrine were from Sigma-Aldrich.Genistein, LY-294002, and wortmannin were obtained from WakoPure Chemicals (Osaka, Japan), BIOMOL Research Laboratories(Plymouth Meeting, PA), and Calbiochem (San Diego, CA), respec-tively. Genistein, tyrphostin 23, LY-294002, and wortmannin weredissolved in dimethyl sulfoxide, whereas phenylephrine was dis-solved in water. Other reagents used for this study were all ofbiochemical grade.
Statistics. All values are expressed as mean � S.E.M. n repre-sents the number of experiments as indicated in figure legends.Statistical significance was evaluated using Students’ t test, whentwo groups were compared and in case of more than two groupsDunnett’s multiple comparison test was used. The results were con-sidered significant at p � 0.05. The density of the signals correspond-ing to tyrosine-phosphorylated proteins and MLC was quantitativelyanalyzed by densitometry (Advanced American Biotechnology, Ful-
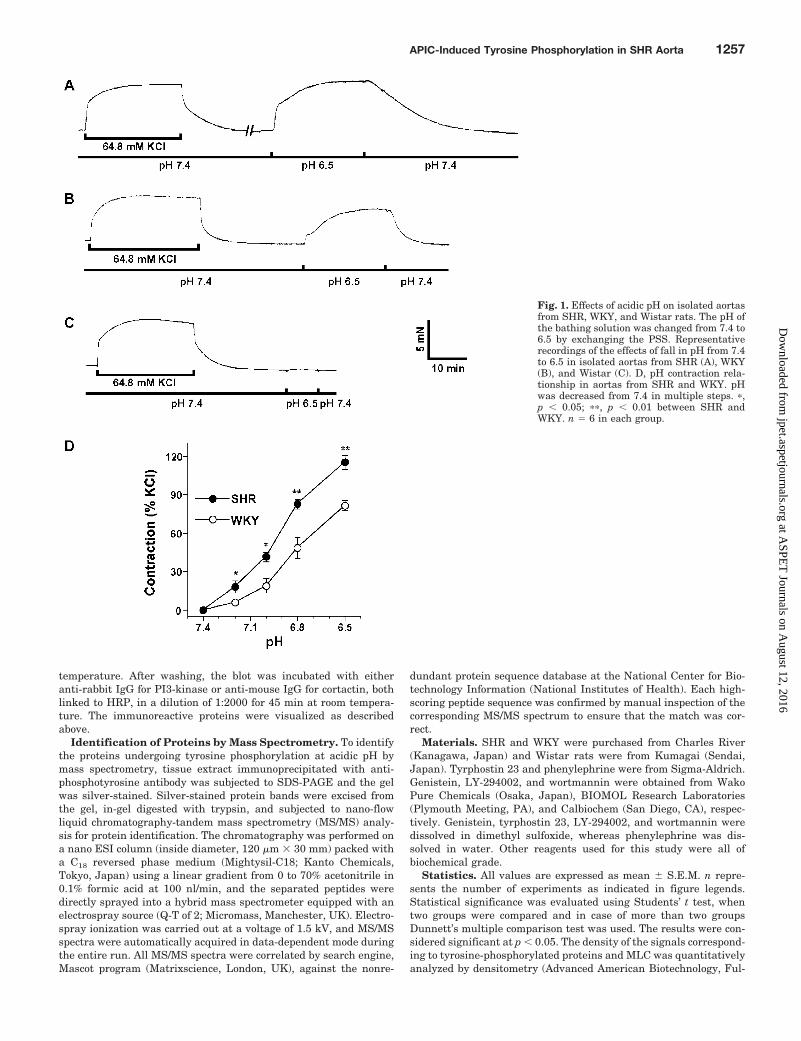
Fig. 1. Effects of acidic pH on isolated aortasfrom SHR, WKY, and Wistar rats. The pH ofthe bathing solution was changed from 7.4 to6.5 by exchanging the PSS. Representativerecordings of the effects of fall in pH from 7.4to 6.5 in isolated aortas from SHR (A), WKY(B), and Wistar (C). D, pH contraction rela-tionship in aortas from SHR and WKY. pHwas decreased from 7.4 in multiple steps. �,p � 0.05; ��, p � 0.01 between SHR andWKY. n � 6 in each group.
APIC-Induced Tyrosine Phosphorylation in SHR Aorta 1257
at ASPE
T Journals on A
ugust 12, 2016jpet.aspetjournals.org
Dow
nloaded from
lerton, CA). Tyrosine phosphorylation of proteins was expressed asthe -fold of control, whereas MLC phosphorylation was quantifiedwith the formula [monophosphorylated MLC/(nonphosphorylatedMLC � monophosphorylated MLC)].
ResultsEffect of Acidic pH on Isolated Aortas from SHR,
WKY, and Wistar Rats. Decreasing pH of the bathing me-dium from 7.4 to 6.5 by exchanging the PSS produced per-sistent contractions in isolated aortas from SHR (Fig. 1A)and WKY (Fig. 1B) that were 122.0 � 4.9% (n � 12) and82.4 � 3.8% of the 64.8 mM KCl-induced contraction, respec-tively. Interestingly, unlike SHR and WKY, acidic pH failedto produce any contractile response in aorta from Wistar rat(Fig. 1C). In separate experiments, pH-contractile responseshowed a direct relationship with the level of decrease in pH
in both SHR and WKY; however, at each pH level, the con-tractile response in SHR was significantly higher than thatin WKY (Fig. 1D).
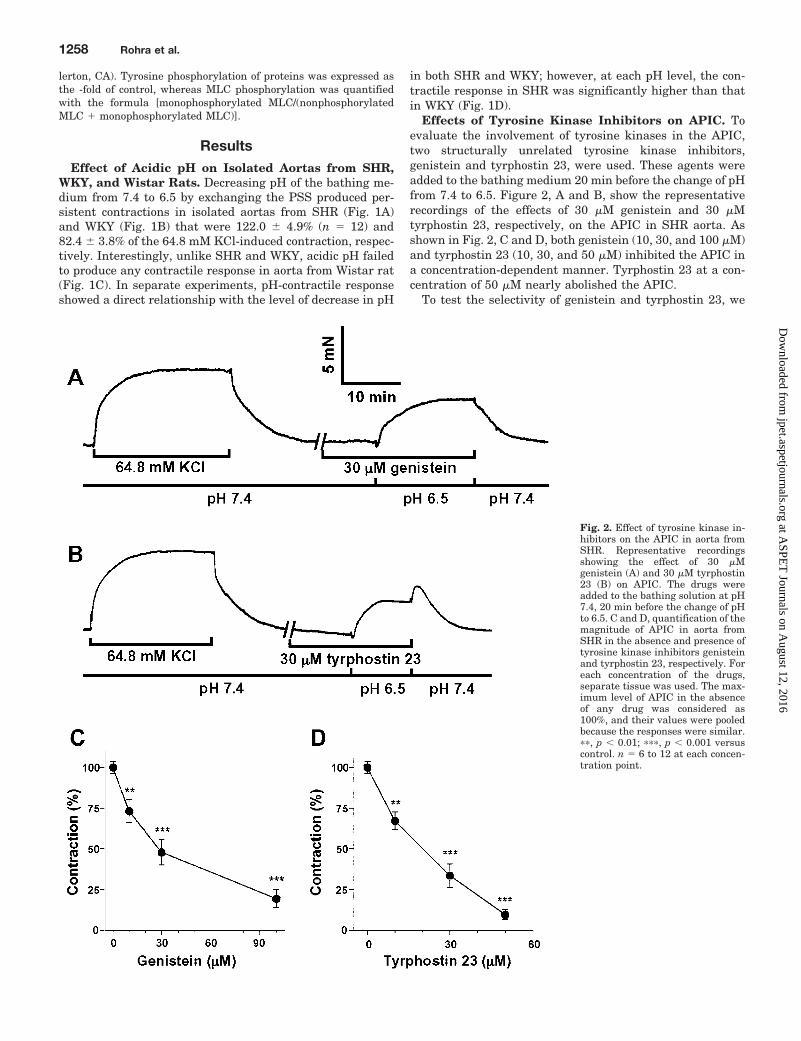
Effects of Tyrosine Kinase Inhibitors on APIC. Toevaluate the involvement of tyrosine kinases in the APIC,two structurally unrelated tyrosine kinase inhibitors,genistein and tyrphostin 23, were used. These agents wereadded to the bathing medium 20 min before the change of pHfrom 7.4 to 6.5. Figure 2, A and B, show the representativerecordings of the effects of 30 �M genistein and 30 �Mtyrphostin 23, respectively, on the APIC in SHR aorta. Asshown in Fig. 2, C and D, both genistein (10, 30, and 100 �M)and tyrphostin 23 (10, 30, and 50 �M) inhibited the APIC ina concentration-dependent manner. Tyrphostin 23 at a con-centration of 50 �M nearly abolished the APIC.
To test the selectivity of genistein and tyrphostin 23, we
Fig. 2. Effect of tyrosine kinase in-hibitors on the APIC in aorta fromSHR. Representative recordingsshowing the effect of 30 �Mgenistein (A) and 30 �M tyrphostin23 (B) on APIC. The drugs wereadded to the bathing solution at pH7.4, 20 min before the change of pHto 6.5. C and D, quantification of themagnitude of APIC in aorta fromSHR in the absence and presence oftyrosine kinase inhibitors genisteinand tyrphostin 23, respectively. Foreach concentration of the drugs,separate tissue was used. The max-imum level of APIC in the absenceof any drug was considered as100%, and their values were pooledbecause the responses were similar.��, p � 0.01; ���, p � 0.001 versuscontrol. n � 6 to 12 at each concen-tration point.
1258 Rohra et al.
at ASPE
T Journals on A
ugust 12, 2016jpet.aspetjournals.org
Dow
nloaded from
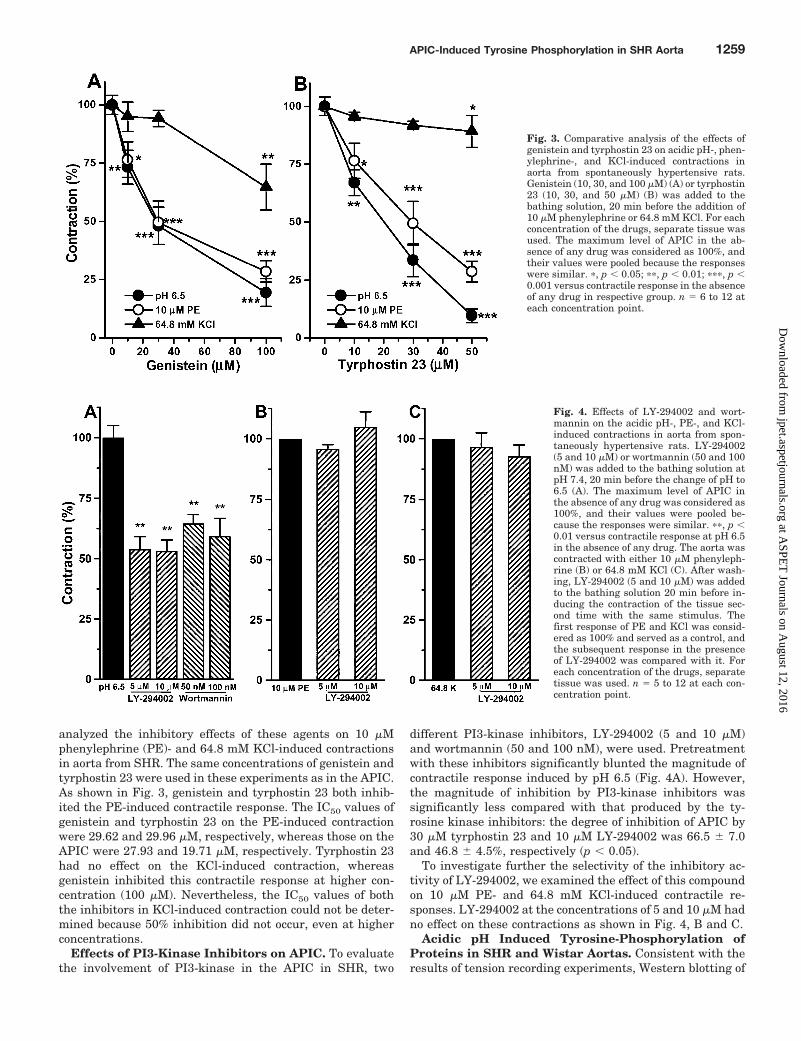
analyzed the inhibitory effects of these agents on 10 �Mphenylephrine (PE)- and 64.8 mM KCl-induced contractionsin aorta from SHR. The same concentrations of genistein andtyrphostin 23 were used in these experiments as in the APIC.As shown in Fig. 3, genistein and tyrphostin 23 both inhib-ited the PE-induced contractile response. The IC50 values ofgenistein and tyrphostin 23 on the PE-induced contractionwere 29.62 and 29.96 �M, respectively, whereas those on theAPIC were 27.93 and 19.71 �M, respectively. Tyrphostin 23had no effect on the KCl-induced contraction, whereasgenistein inhibited this contractile response at higher con-centration (100 �M). Nevertheless, the IC50 values of boththe inhibitors in KCl-induced contraction could not be deter-mined because 50% inhibition did not occur, even at higherconcentrations.
Effects of PI3-Kinase Inhibitors on APIC. To evaluatethe involvement of PI3-kinase in the APIC in SHR, two
different PI3-kinase inhibitors, LY-294002 (5 and 10 �M)and wortmannin (50 and 100 nM), were used. Pretreatmentwith these inhibitors significantly blunted the magnitude ofcontractile response induced by pH 6.5 (Fig. 4A). However,the magnitude of inhibition by PI3-kinase inhibitors wassignificantly less compared with that produced by the ty-rosine kinase inhibitors: the degree of inhibition of APIC by30 �M tyrphostin 23 and 10 �M LY-294002 was 66.5 � 7.0and 46.8 � 4.5%, respectively (p � 0.05).
To investigate further the selectivity of the inhibitory ac-tivity of LY-294002, we examined the effect of this compoundon 10 �M PE- and 64.8 mM KCl-induced contractile re-sponses. LY-294002 at the concentrations of 5 and 10 �M hadno effect on these contractions as shown in Fig. 4, B and C.
Acidic pH Induced Tyrosine-Phosphorylation ofProteins in SHR and Wistar Aortas. Consistent with theresults of tension recording experiments, Western blotting of
Fig. 3. Comparative analysis of the effects ofgenistein and tyrphostin 23 on acidic pH-, phen-ylephrine-, and KCl-induced contractions inaorta from spontaneously hypertensive rats.Genistein (10, 30, and 100 �M) (A) or tyrphostin23 (10, 30, and 50 �M) (B) was added to thebathing solution, 20 min before the addition of10 �M phenylephrine or 64.8 mM KCl. For eachconcentration of the drugs, separate tissue wasused. The maximum level of APIC in the ab-sence of any drug was considered as 100%, andtheir values were pooled because the responseswere similar. �, p � 0.05; ��, p � 0.01; ���, p �0.001 versus contractile response in the absenceof any drug in respective group. n � 6 to 12 ateach concentration point.
Fig. 4. Effects of LY-294002 and wort-mannin on the acidic pH-, PE-, and KCl-induced contractions in aorta from spon-taneously hypertensive rats. LY-294002(5 and 10 �M) or wortmannin (50 and 100nM) was added to the bathing solution atpH 7.4, 20 min before the change of pH to6.5 (A). The maximum level of APIC inthe absence of any drug was considered as100%, and their values were pooled be-cause the responses were similar. ��, p �0.01 versus contractile response at pH 6.5in the absence of any drug. The aorta wascontracted with either 10 �M phenyleph-rine (B) or 64.8 mM KCl (C). After wash-ing, LY-294002 (5 and 10 �M) was addedto the bathing solution 20 min before in-ducing the contraction of the tissue sec-ond time with the same stimulus. Thefirst response of PE and KCl was consid-ered as 100% and served as a control, andthe subsequent response in the presenceof LY-294002 was compared with it. Foreach concentration of the drugs, separatetissue was used. n � 5 to 12 at each con-centration point.
APIC-Induced Tyrosine Phosphorylation in SHR Aorta 1259
at ASPE
T Journals on A
ugust 12, 2016jpet.aspetjournals.org
Dow
nloaded from
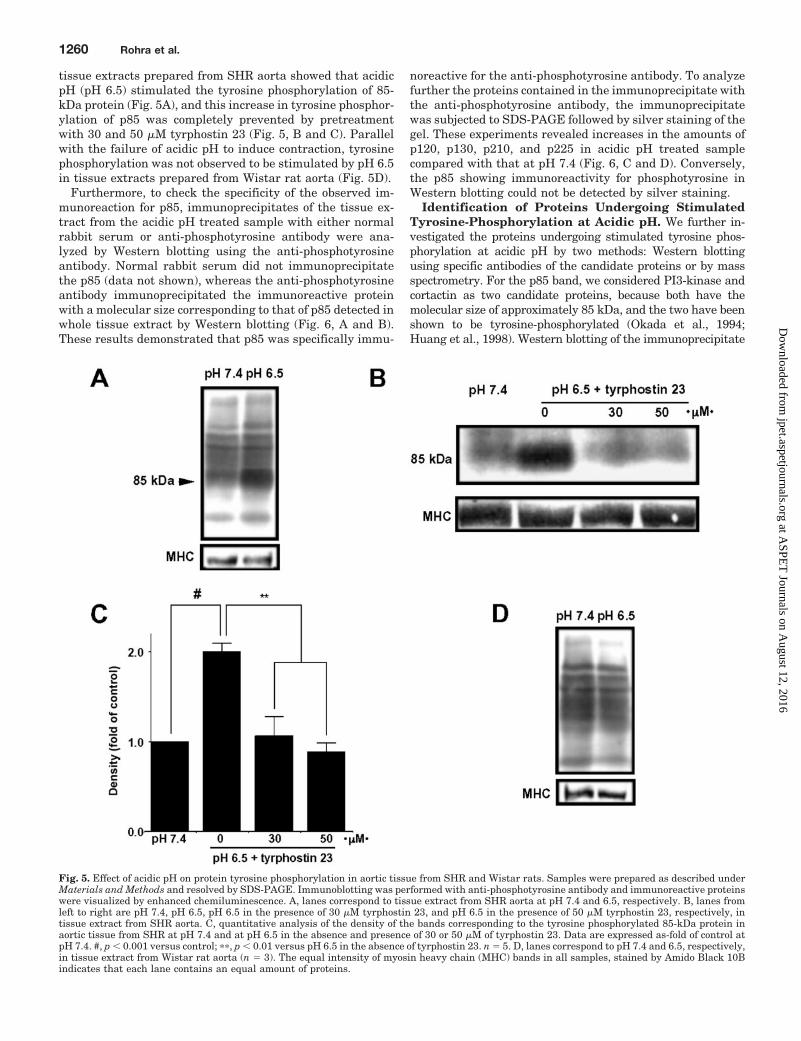
tissue extracts prepared from SHR aorta showed that acidicpH (pH 6.5) stimulated the tyrosine phosphorylation of 85-kDa protein (Fig. 5A), and this increase in tyrosine phosphor-ylation of p85 was completely prevented by pretreatmentwith 30 and 50 �M tyrphostin 23 (Fig. 5, B and C). Parallelwith the failure of acidic pH to induce contraction, tyrosinephosphorylation was not observed to be stimulated by pH 6.5in tissue extracts prepared from Wistar rat aorta (Fig. 5D).
Furthermore, to check the specificity of the observed im-munoreaction for p85, immunoprecipitates of the tissue ex-tract from the acidic pH treated sample with either normalrabbit serum or anti-phosphotyrosine antibody were ana-lyzed by Western blotting using the anti-phosphotyrosineantibody. Normal rabbit serum did not immunoprecipitatethe p85 (data not shown), whereas the anti-phosphotyrosineantibody immunoprecipitated the immunoreactive proteinwith a molecular size corresponding to that of p85 detected inwhole tissue extract by Western blotting (Fig. 6, A and B).These results demonstrated that p85 was specifically immu-
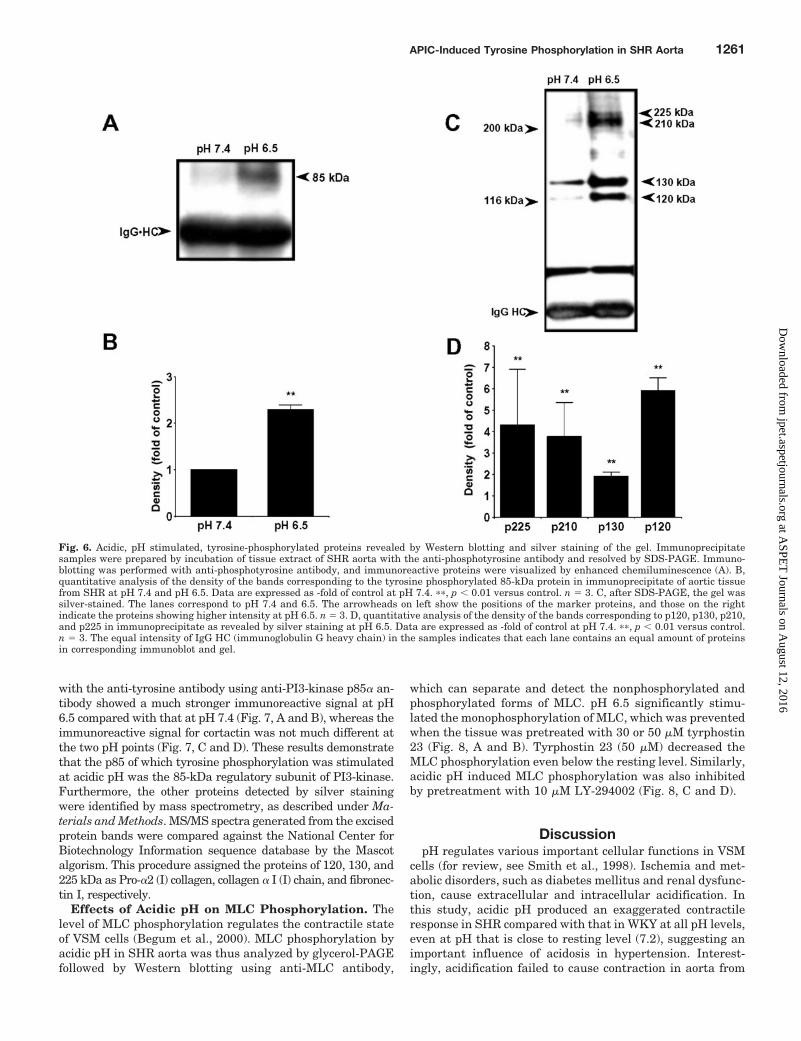
noreactive for the anti-phosphotyrosine antibody. To analyzefurther the proteins contained in the immunoprecipitate withthe anti-phosphotyrosine antibody, the immunoprecipitatewas subjected to SDS-PAGE followed by silver staining of thegel. These experiments revealed increases in the amounts ofp120, p130, p210, and p225 in acidic pH treated samplecompared with that at pH 7.4 (Fig. 6, C and D). Conversely,the p85 showing immunoreactivity for phosphotyrosine inWestern blotting could not be detected by silver staining.
Identification of Proteins Undergoing StimulatedTyrosine-Phosphorylation at Acidic pH. We further in-vestigated the proteins undergoing stimulated tyrosine phos-phorylation at acidic pH by two methods: Western blottingusing specific antibodies of the candidate proteins or by massspectrometry. For the p85 band, we considered PI3-kinase andcortactin as two candidate proteins, because both have themolecular size of approximately 85 kDa, and the two have beenshown to be tyrosine-phosphorylated (Okada et al., 1994;Huang et al., 1998). Western blotting of the immunoprecipitate
Fig. 5. Effect of acidic pH on protein tyrosine phosphorylation in aortic tissue from SHR and Wistar rats. Samples were prepared as described underMaterials and Methods and resolved by SDS-PAGE. Immunoblotting was performed with anti-phosphotyrosine antibody and immunoreactive proteinswere visualized by enhanced chemiluminescence. A, lanes correspond to tissue extract from SHR aorta at pH 7.4 and 6.5, respectively. B, lanes fromleft to right are pH 7.4, pH 6.5, pH 6.5 in the presence of 30 �M tyrphostin 23, and pH 6.5 in the presence of 50 �M tyrphostin 23, respectively, intissue extract from SHR aorta. C, quantitative analysis of the density of the bands corresponding to the tyrosine phosphorylated 85-kDa protein inaortic tissue from SHR at pH 7.4 and at pH 6.5 in the absence and presence of 30 or 50 �M of tyrphostin 23. Data are expressed as-fold of control atpH 7.4. #, p � 0.001 versus control; ��, p � 0.01 versus pH 6.5 in the absence of tyrphostin 23. n � 5. D, lanes correspond to pH 7.4 and 6.5, respectively,in tissue extract from Wistar rat aorta (n � 3). The equal intensity of myosin heavy chain (MHC) bands in all samples, stained by Amido Black 10Bindicates that each lane contains an equal amount of proteins.
1260 Rohra et al.
at ASPE
T Journals on A
ugust 12, 2016jpet.aspetjournals.org
Dow
nloaded from
with the anti-tyrosine antibody using anti-PI3-kinase p85� an-tibody showed a much stronger immunoreactive signal at pH6.5 compared with that at pH 7.4 (Fig. 7, A and B), whereas theimmunoreactive signal for cortactin was not much different atthe two pH points (Fig. 7, C and D). These results demonstratethat the p85 of which tyrosine phosphorylation was stimulatedat acidic pH was the 85-kDa regulatory subunit of PI3-kinase.Furthermore, the other proteins detected by silver stainingwere identified by mass spectrometry, as described under Ma-terials and Methods. MS/MS spectra generated from the excisedprotein bands were compared against the National Center forBiotechnology Information sequence database by the Mascotalgorism. This procedure assigned the proteins of 120, 130, and225 kDa as Pro-�2 (I) collagen, collagen � I (I) chain, and fibronec-tin I, respectively.
Effects of Acidic pH on MLC Phosphorylation. Thelevel of MLC phosphorylation regulates the contractile stateof VSM cells (Begum et al., 2000). MLC phosphorylation byacidic pH in SHR aorta was thus analyzed by glycerol-PAGEfollowed by Western blotting using anti-MLC antibody,
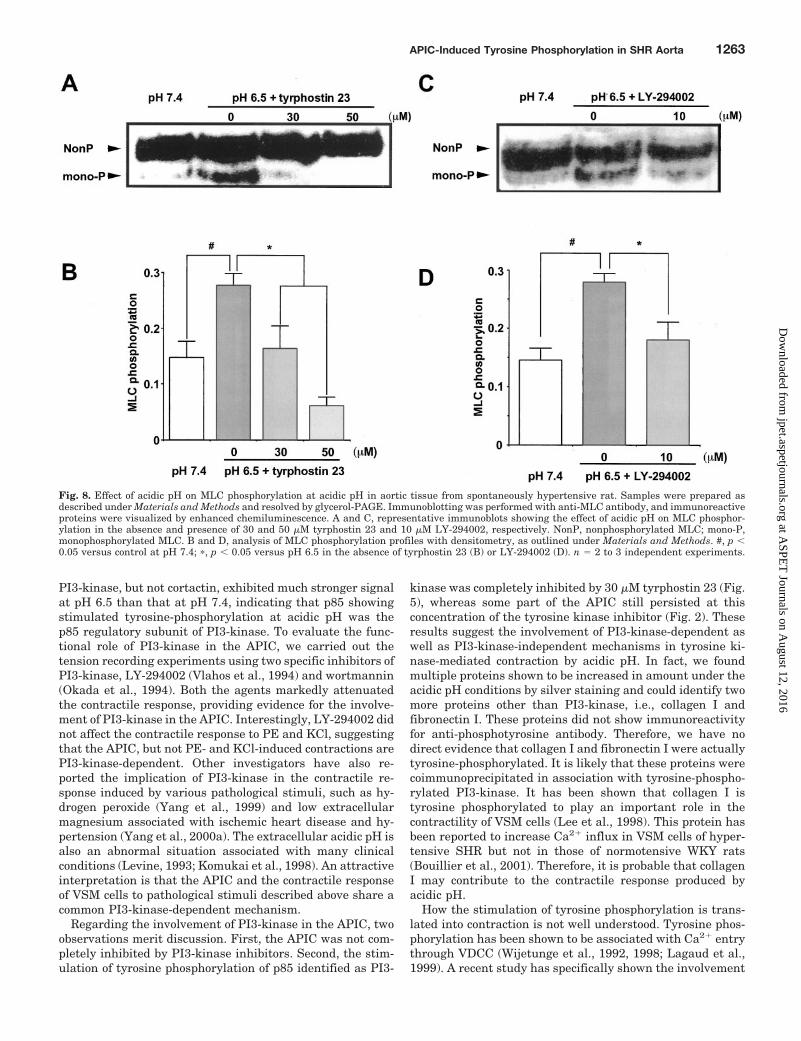
which can separate and detect the nonphosphorylated andphosphorylated forms of MLC. pH 6.5 significantly stimu-lated the monophosphorylation of MLC, which was preventedwhen the tissue was pretreated with 30 or 50 �M tyrphostin23 (Fig. 8, A and B). Tyrphostin 23 (50 �M) decreased theMLC phosphorylation even below the resting level. Similarly,acidic pH induced MLC phosphorylation was also inhibitedby pretreatment with 10 �M LY-294002 (Fig. 8, C and D).
DiscussionpH regulates various important cellular functions in VSM
cells (for review, see Smith et al., 1998). Ischemia and met-abolic disorders, such as diabetes mellitus and renal dysfunc-tion, cause extracellular and intracellular acidification. Inthis study, acidic pH produced an exaggerated contractileresponse in SHR compared with that in WKY at all pH levels,even at pH that is close to resting level (7.2), suggesting animportant influence of acidosis in hypertension. Interest-ingly, acidification failed to cause contraction in aorta from
Fig. 6. Acidic, pH stimulated, tyrosine-phosphorylated proteins revealed by Western blotting and silver staining of the gel. Immunoprecipitatesamples were prepared by incubation of tissue extract of SHR aorta with the anti-phosphotyrosine antibody and resolved by SDS-PAGE. Immuno-blotting was performed with anti-phosphotyrosine antibody, and immunoreactive proteins were visualized by enhanced chemiluminescence (A). B,quantitative analysis of the density of the bands corresponding to the tyrosine phosphorylated 85-kDa protein in immunoprecipitate of aortic tissuefrom SHR at pH 7.4 and pH 6.5. Data are expressed as -fold of control at pH 7.4. ��, p � 0.01 versus control. n � 3. C, after SDS-PAGE, the gel wassilver-stained. The lanes correspond to pH 7.4 and 6.5. The arrowheads on left show the positions of the marker proteins, and those on the rightindicate the proteins showing higher intensity at pH 6.5. n � 3. D, quantitative analysis of the density of the bands corresponding to p120, p130, p210,and p225 in immunoprecipitate as revealed by silver staining at pH 6.5. Data are expressed as -fold of control at pH 7.4. ��, p � 0.01 versus control.n � 3. The equal intensity of IgG HC (immunoglobulin G heavy chain) in the samples indicates that each lane contains an equal amount of proteinsin corresponding immunoblot and gel.
APIC-Induced Tyrosine Phosphorylation in SHR Aorta 1261
at ASPE
T Journals on A
ugust 12, 2016jpet.aspetjournals.org
Dow
nloaded from
Wistar rat. These results point to the importance of acidicpH-contraction coupling mechanism in SHR and WKY,which is lacking in Wistar rat. From all these findings, twoconclusions can be drawn. First, the magnitude of the APICis dependent on blood pressure in the case of SHR and WKYand is consistent with our previous report (Furukawa et al.,1996). Second, APIC is strain-specific because acidic pH didnot induce any contraction in Wistar rat aorta.
Other important findings from the tension recording ex-periments are that tyrosine kinase inhibitors genistein andtyrphostin 23 both caused a marked inhibition of the APIC,suggesting the involvement of tyrosine kinases in the APIC.Genistein and tyrphostin 23 are two structurally unrelatedtyrosine kinase inhibitors that act through different mecha-nisms: genistein exerts its inhibitory action by binding to theATP-binding site (Akiyama et al., 1987), whereas tyrphostin23 interacts with the substrate-binding site of the enzymes(Levitzki and Gazit, 1995). In the present study, genisteinand tyrphostin 23 showed a differential inhibitory effect onthe acidic pH , PE-, and KCl-induced contractions, that is, thedrugs inhibited the acidic pH and PE-induced contractionsmarkedly compared with that induced by KCl. Similar relax-ant effects of tyrosine kinase inhibitors, including tyrphostin23 and genistein, on a variety of agonists-induced contrac-tions in vascular tissues have been reported through inhibi-tion of tyrosine kinases (Di Salvo et al., 1993; Jin et al., 1996;Watts et al., 1996; Yang et al., 2000b; Carmines et al., 2001).
Consistent with the results of the tension recording experi-ments, Western blotting of the tissue extract from acidic pHtreated aorta from SHR demonstrated the stimulation oftyrosine phosphorylation at acidic pH, which was preventedby pretreatment with tyrphostin 23. Because tyrosine phos-phorylation has been implicated in several signaling path-ways, one may argue that the contraction and stimulation oftyrosine phosphorylation observed at acidic pH in SHR mightbe unrelated to each other. Therefore, to confirm the paral-lelism between the two events, tissue extract from Wistar rataorta was used in which APIC failed to occur. Acidic pHfailed to induce tyrosine phosphorylation in aorta fromWistar rat. These observations demonstrate that the tyrosinekinase-mediated protein phosphorylation is mainly responsi-ble for the contractile response induced by acidic pH in SHRaorta and that failure of acidic pH to produce contraction inWistar rat is in parallel with the lack of protein tyrosinephosphorylation.
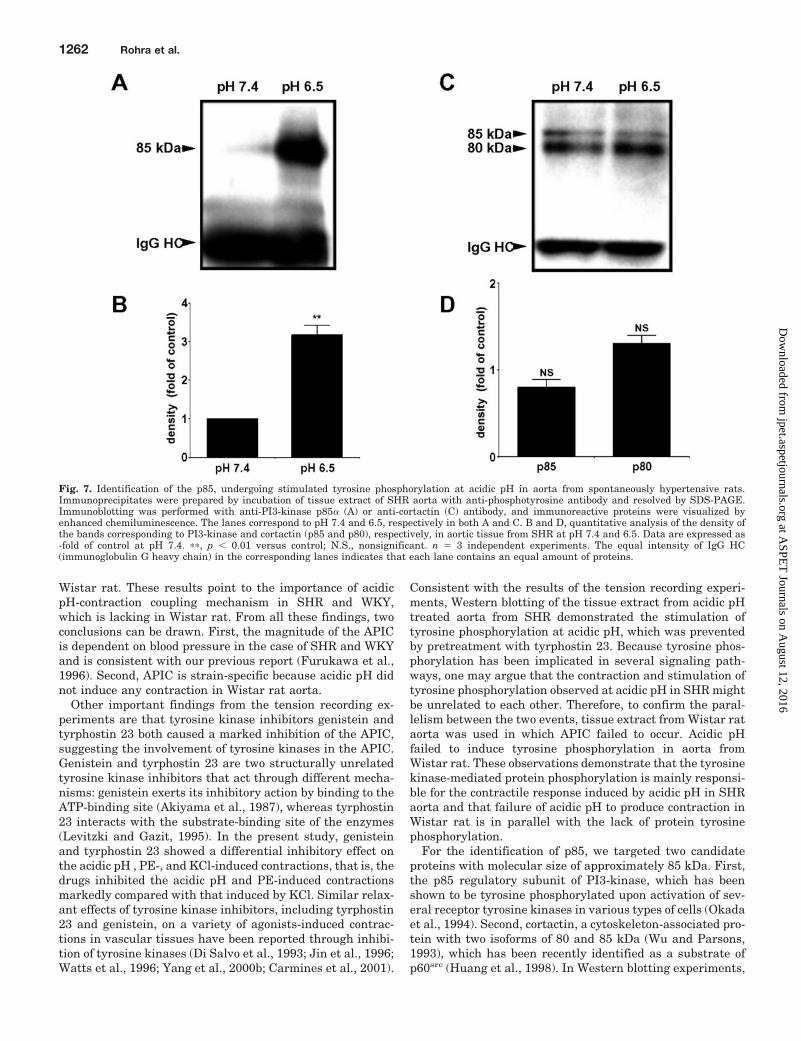
For the identification of p85, we targeted two candidateproteins with molecular size of approximately 85 kDa. First,the p85 regulatory subunit of PI3-kinase, which has beenshown to be tyrosine phosphorylated upon activation of sev-eral receptor tyrosine kinases in various types of cells (Okadaet al., 1994). Second, cortactin, a cytoskeleton-associated pro-tein with two isoforms of 80 and 85 kDa (Wu and Parsons,1993), which has been recently identified as a substrate ofp60src (Huang et al., 1998). In Western blotting experiments,
Fig. 7. Identification of the p85, undergoing stimulated tyrosine phosphorylation at acidic pH in aorta from spontaneously hypertensive rats.Immunoprecipitates were prepared by incubation of tissue extract of SHR aorta with anti-phosphotyrosine antibody and resolved by SDS-PAGE.Immunoblotting was performed with anti-PI3-kinase p85� (A) or anti-cortactin (C) antibody, and immunoreactive proteins were visualized byenhanced chemiluminescence. The lanes correspond to pH 7.4 and 6.5, respectively in both A and C. B and D, quantitative analysis of the density ofthe bands corresponding to PI3-kinase and cortactin (p85 and p80), respectively, in aortic tissue from SHR at pH 7.4 and 6.5. Data are expressed as-fold of control at pH 7.4. ��, p � 0.01 versus control; N.S., nonsignificant. n � 3 independent experiments. The equal intensity of IgG HC(immunoglobulin G heavy chain) in the corresponding lanes indicates that each lane contains an equal amount of proteins.
1262 Rohra et al.
at ASPE
T Journals on A
ugust 12, 2016jpet.aspetjournals.org
Dow
nloaded from
PI3-kinase, but not cortactin, exhibited much stronger signalat pH 6.5 than that at pH 7.4, indicating that p85 showingstimulated tyrosine-phosphorylation at acidic pH was thep85 regulatory subunit of PI3-kinase. To evaluate the func-tional role of PI3-kinase in the APIC, we carried out thetension recording experiments using two specific inhibitors ofPI3-kinase, LY-294002 (Vlahos et al., 1994) and wortmannin(Okada et al., 1994). Both the agents markedly attenuatedthe contractile response, providing evidence for the involve-ment of PI3-kinase in the APIC. Interestingly, LY-294002 didnot affect the contractile response to PE and KCl, suggestingthat the APIC, but not PE- and KCl-induced contractions arePI3-kinase-dependent. Other investigators have also re-ported the implication of PI3-kinase in the contractile re-sponse induced by various pathological stimuli, such as hy-drogen peroxide (Yang et al., 1999) and low extracellularmagnesium associated with ischemic heart disease and hy-pertension (Yang et al., 2000a). The extracellular acidic pH isalso an abnormal situation associated with many clinicalconditions (Levine, 1993; Komukai et al., 1998). An attractiveinterpretation is that the APIC and the contractile responseof VSM cells to pathological stimuli described above share acommon PI3-kinase-dependent mechanism.
Regarding the involvement of PI3-kinase in the APIC, twoobservations merit discussion. First, the APIC was not com-pletely inhibited by PI3-kinase inhibitors. Second, the stim-ulation of tyrosine phosphorylation of p85 identified as PI3-
kinase was completely inhibited by 30 �M tyrphostin 23 (Fig.5), whereas some part of the APIC still persisted at thisconcentration of the tyrosine kinase inhibitor (Fig. 2). Theseresults suggest the involvement of PI3-kinase-dependent aswell as PI3-kinase-independent mechanisms in tyrosine ki-nase-mediated contraction by acidic pH. In fact, we foundmultiple proteins shown to be increased in amount under theacidic pH conditions by silver staining and could identify twomore proteins other than PI3-kinase, i.e., collagen I andfibronectin I. These proteins did not show immunoreactivityfor anti-phosphotyrosine antibody. Therefore, we have nodirect evidence that collagen I and fibronectin I were actuallytyrosine-phosphorylated. It is likely that these proteins werecoimmunoprecipitated in association with tyrosine-phospho-rylated PI3-kinase. It has been shown that collagen I istyrosine phosphorylated to play an important role in thecontractility of VSM cells (Lee et al., 1998). This protein hasbeen reported to increase Ca2� influx in VSM cells of hyper-tensive SHR but not in those of normotensive WKY rats(Bouillier et al., 2001). Therefore, it is probable that collagenI may contribute to the contractile response produced byacidic pH.
How the stimulation of tyrosine phosphorylation is trans-lated into contraction is not well understood. Tyrosine phos-phorylation has been shown to be associated with Ca2� entrythrough VDCC (Wijetunge et al., 1992, 1998; Lagaud et al.,1999). A recent study has specifically shown the involvement
Fig. 8. Effect of acidic pH on MLC phosphorylation at acidic pH in aortic tissue from spontaneously hypertensive rat. Samples were prepared asdescribed under Materials and Methods and resolved by glycerol-PAGE. Immunoblotting was performed with anti-MLC antibody, and immunoreactiveproteins were visualized by enhanced chemiluminescence. A and C, representative immunoblots showing the effect of acidic pH on MLC phosphor-ylation in the absence and presence of 30 and 50 �M tyrphostin 23 and 10 �M LY-294002, respectively. NonP, nonphosphorylated MLC; mono-P,monophosphorylated MLC. B and D, analysis of MLC phosphorylation profiles with densitometry, as outlined under Materials and Methods. #, p �0.05 versus control at pH 7.4; �, p � 0.05 versus pH 6.5 in the absence of tyrphostin 23 (B) or LY-294002 (D). n � 2 to 3 independent experiments.
APIC-Induced Tyrosine Phosphorylation in SHR Aorta 1263
at ASPE
T Journals on A
ugust 12, 2016jpet.aspetjournals.org
Dow
nloaded from

of PI3-kinase in the mediation of Ca2� influx through L-typeCa2� channels in vascular myocytes (Quignard et al., 2001).Indeed, we have also reported that a major component of theAPIC is associated with depolarization and Ca2� entrythrough VDCC (Furukawa et al., 1996). Therefore, it is rea-sonable to assume that the acidic pH induced tyrosine phos-phorylation results in the Ca2� influx via VDCC and hencethe contraction.
It is well known that the contractile state of the VSM cellsis usually dependent upon the phosphorylated level of MLC(Karaki, 1987). In the present study, MLC phosphorylationwas shown to be stimulated at the acidic pH, and both tyr-phostin 23 and LY-294002 inhibited the phosphorylationlevel of MLC. Tyrphostin 23 at a concentration of 50 �Mcompletely prevented the stimulation of MLC phosphoryla-tion, complementing the results of tension recording experi-ments where tyrphostin 23 nearly abolished the APIC. Fromthese observations, it is suggested that, the acidic pH acti-vates tyrosine kinases and causes enhancement of MLCphosphorylation, and thus the contraction of SHR aorta.
In summary, the present study is the first to demonstratethat acidic pH stimulates tyrosine kinases to enhance ty-rosine phosphorylation of PI3-kinase, resulting in the MLCphosphorylation-dependent contraction of isolated aortafrom SHR but not from Wistar.
ReferencesAdam LP, Franklin MT, Raff GJ, and Hathaway DR (1995) Activation of mitogen-
activated protein kinase in porcine carotid arteries. Circ Res 76:183–190.Akiyama T, Ishida J, Nakagawa S, Ogawara H, Watanabe S, Itoh N, Shibuya M, and
Fukami Y (1987) Genistein, a specific inhibitor of tyrosine-specific protein kinases.J Biol Chem 262:5592–5595.
Banes A, Florian JA, and Watts SW (1999) Mechanism of 5-hydroxytryptamine2Areceptor activation of the mitogen-activated protein kinase pathway in vascularsmooth muscle. J Pharmacol Exp Ther 291:1179–1187.
Begum N, Duddy N, Sandu O, Reinzie J, and Ragolia L (2000) Regulation ofmyosin-bound protein phosphatase by insulin in vascular smooth muscle cells:evaluation of the role of Rho kinase and phosphatidylinositol-3-kinase-dependentsignaling pathways. Mol Endocrinol 14:1365–1376.
Bouillier H, Samain E, Rucker-martin C, Renaud JF, Safar M, and Dagher G (2001)Effect of extracellular matrix elements on angiotensin II-induced calcium releasein vascular smooth muscle cells from normotensive and hypertensive rats. Hyper-tension 37:1465–1472.
Butwell NA, Ramasamy R, Lazar I, Sherry AD, and Malloy CR (1993) Effect oflidocaine on contracture, intracellular sodium and pH in ischaemic rat hearts.Am J Physiol 264:H1884–H1889.
Carmines PK, Fallet RW, Che Q, and Fujiwara K (2001) Tyrosine kinase involve-ment in renal arteriolar constrictor responses to angiotensin II. Hypertension37:569–573.
Chen X-L and Rembold CM (1995) pHi, [Ca2�]i and myosin phosphorylation inhistamine- and NH4
�-induced swine carotid artery contraction. Hypertension 25:482–489.
Di Salvo J, Pfitzer G, and Semenchuk LA (1994) Protein tyrosine phosphorylation,cellular Ca2� and Ca2� sensitivity for contraction of smooth muscle. Can J PhysiolPharmacol 72:1434–1439.
Di Salvo J, Steusloff A, Semenchuk L, Satoh S, Kolquist K, and Pfitzer G (1993)Tyrosine kinase inhibitors suppress agonist-induced contraction in smooth muscle.Biochem Biophys Res Commun 190:968–974.
Furukawa KI, Komaba J, Sakai H, and Ohizumi Y (1996) The mechanism of acidicpH induced contraction in aortae from SHR and WKY rats enhanced by increasingblood pressure. Br J Pharmacol 118:485–492.
Horowitz A, Menice CB, Laporte R, and Morgan KG (1996) Mechanism of smoothmuscle contraction. Physiol Rev 76:967–1003.
Huang C, Liu J, Haudenschild CC, and Zhan X (1998) The role of tyrosine phosphor-
ylation of cortactin in the locomotion of endothelial cells. J Biol Chem 273:25770–25776.
Jin N, Siddiqui RA, English D, and Rhoades RA (1996) Communication betweentyrosine kinase pathway and myosin light chain kinase pathway in smooth mus-cle. Am J Physiol 271:H1348–H1355.
Karaki H (1987) Use of tension measurements to delineate the mode of action ofvasodilators. J Pharmacol Methods 18:1–21.
Khalil RA, Menice CB, Wang CLA, and Morgan KG (1995) Phosphotyrosine-dependent targeting of mitogen-activated protein kinase in differentiated contrac-tile vascular cells. Circ Res 76:1101–1108.
Komukai K, Ishikawa T, and Kurihara S (1998) Effects of acidosis on Ca2� sensi-tivity of contractile elements in intact ferret myocardium. Am J Physiol 274:H147–H154.
Lagaud GJL, Randriamboavonjy V, Roul G, Stoclet JC, and Andriantsitohaina R(1999) Mechanism of Ca2� release and entry during contraction elicited by nor-epinephrine in rat resistance arteries. Am J Physiol 276:H300–H308.
Laniyonu AA, Saifeddine M, Yang SG, and Hollenberg MD (1994) Tyrosine kinaseinhibitors and the contractile action of G-protein linked vascular agonists. CanJ Physiol Pharmacol 72:1075–1085.
Leach RM, Sheehan DW, Chacko VP, and Sylvester JT (1998) Effects of hypoxia onenergy state and pH in resting pulmonary and femoral arterial smooth muscles.Am J Physiol 275:L1051–L1060.
Lee KM, Tsai KY, Wang N, and Ingber DE (1998) Extracellular matrix and pulmo-nary hypertension: control of vascular smooth muscle cell contractility. Am JPhysiol 274:H76–H82.
Levine RL (1993) Ischaemia: from acidosis to oxidation. FASEB J 7:1242–1246.Levitzki A and Gazit A (1995) Tyrosine kinase inhibition: an approach to drug
development. Science (Wash DC) 267:1782–1788.Nagesetty R and Paul RJ (1994) Effects of pHi on isometric force and [Ca2�]i in
porcine coronary artery smooth muscle. Circ Res 75:990–998.Nishiyama A, Miyatake A, Aki Y, Fukui T, Rahman M, Kimura S, and Abe Y (1999)
Adenosine A1 receptor antagonist KW-3902 prevents hypoxia-induced renal vaso-constriction. J Pharmacol Exp Ther 291:988–993.
Okada T, Kawano Y, Sakakibara T, Hazeki O, and Ui M (1994) Essential role ofphosphatidylinositol 3-kinase in insulin-induced glucose transport and antilypoly-sis in rat adipocytes. Studies with a selective inhibitor wortmannin. J Biol Chem269:3568–3573.
Quignard JF, Mironneau J, Carricaburu V, Fournier B, Babich A, Nurnbergs B,Mironneau C, and Marcrez N (2001) Phosphoinositide 3-kinase � mediates angio-tensin II-induced stimulation of L-type calcium channels in vascular myocytes.J Biol Chem 276:32545–32551.
Somlyo AP and Somlyo AV (1998) From pharmacomechanical coupling to G-proteinsand myosin phosphatase. Acta Physiol Scan 164:437–448.
Smith GL, Austin C, Crichton C, and Wray S (1998) A review of the actions andcontrol of intracellular pH in vascular smooth muscle. Cardiovasc Res 38:316–331.
Vlahos C, Matter WF, Hui KY, and Brown RF (1994) A specific inhibitor of phos-phatidylinositol 3-kinase, 2-(4-morpholinyl)-8-phenyl-4H-1-benzopyran-4-one(LY294002). J Biol Chem 269:5241–5248.
Watts SW, Yeum CH, Campbell G, and Webb RC (1996) Serotonin stimulates proteintyrosyl phosphorylation and vascular contraction via tyrosine kinase. J Vasc Res33:288–298.
Wijetunge S, Aalkjaer C, Schachter M, and Hughes AD (1992) Tyrosine kinaseinhibitors block calcium channel currents in vascular smooth muscle cells. Bio-chem Biophys Res Commun 189:1620–1623.
Wijetunge S, Lymn JS, and Hughes AD (1998) Effect of inhibition of tyrosinephosphatases on voltage-operated calcium channel currents in rabbit isolated earartery cells. Br J Pharmacol 124:307–316.
Wu H and Parsons JT (1993) Cortactin, an 80/85-kilodalton pp60src substrate, is afilamentous actin-binding protein enriched in the cell cortex. J Cell Biol 120:1417–1426.
Yang ZW, Wang J, Altura BT, and Altura BM (2000a) Extracellular magnesiumdeficiency induces contraction of arterial muscle: role of PI3-kinases and MAPKsignaling pathways. Pfluegers Arch 439:240–247.
Yang ZW, Wang J, Zheng T, Altura BT, and Altura BM (2000b) Low [Mg2�]o inducescontraction of cerebral arteries: roles of tyrosine and mitogen-activated proteinkinases. Am J Physiol 279:H185–H194.
Yang ZW, Zheng T, Wang J, Zhang A, Altura BT, and Altura BM (1999) Hydrogenperoxide induces contraction and raises [Ca2�]i in canine cerebral arterial smoothmuscle: participation of cellular signaling pathways. Naunyn-Schmiedeberg’s ArchPharmacol 360:646–653.
Address correspondence to: Yasushi Ohizumi, Department of Pharmaceu-tical Molecular Biology, Graduate School of Pharmaceutical Sciences, TohokuUniversity, Aoba, Aramaki, Aoba-ku, Sendai, 980-8575, Japan. E-mail:[email protected]
1264 Rohra et al.
at ASPE
T Journals on A
ugust 12, 2016jpet.aspetjournals.org
Dow
nloaded from





























