Embed Size (px)
Citation preview

ELSEVIER PII S0031-9384(96) 00063-7
Physiology & Behavior, Vol. 60, No. 2, pp. 595-610, 1996 Copyright © 1996 Elsevier Science Inc. Printed in the USA. All rights reserved
0031-9384/96 $15.00 + .00
Stimulus Specificity in Flavor Acceptance Learning
I S R A E L R A M I R E Z
Monell Chemical Senses Center, Philadelphia, PA 19104-3308 USA
R e c e i v e d 14 M a r c h 1995
RAMIREZ, I. Stimulus specificity inflavor acceptance learning. PHYSIOL BEHAV 60(2) 595-610, 1996.--Infusing rats with 6% carbohydrate whenever they drank a test fluid stimulated their intake of 0.03-0.3% saccharin, 0.9% sodium chloride, and 0.5% oligosaccharide solutions within 1 to 3 days, but did not greatly increase intake of cherry-flavored water, 2% saccharin, 0.45% sodium chloride, monosodium glutamate, 0.025% sucrose octaacetate, or 0.4% maltodextrin, when food was available ad lib. In general, it was more difficult to detect an effect with aversive stimuli, but it was possible to detect a significant effect with one such aversive flavor by either increasing the number of animals to permit detection of a small effect, or by using a preference test. For most stimuli, however, rats that had been trained with food available and then tested when food deprived showed significant increases in fluid intake, compared to rats that had been given the same flavor without carbohydrate infusions, even when plain water was infused in all rats during the food deprivation test. The following forms of evidence indicate that the increased intake is due to a form of Pavlovian conditioning. It tends to increase over repeated trials and to persist for a day after the carbohydrate infusions have been discontinued. Indeed, in one experiment, increased intake even persisted for a day when the infusion conditions were reversed, such that control rats were infused with carbohydrate and experimental rats were infused with water. Increased ingestion is attenuated by preexposure to the infusion in the absence of an appropriate taste solution. Increased ingestion is attenuated by preexposure to the taste solution in the absence of infusions (i.e., latent inhibition). Although increased intake is a form of associative conditioning, it is unlike malaise-conditioned taste aversions because taste aversions were conditioned to monosodium glutamate, cherry flavor, and saccharin with equal ease. The ease with which infusion increased intake is influenced by at least three different factors, degree of aversiveness, stimulus intensity, and rats' innate predisposition to acquire certain stimulus-response relationships more readily than others.
Learning Intragastric feeding Appetite Carbohydrates Sugars Taste
TASTE/FLAVOR preferences can be increased by repeatedly pair- ing nutrients with ingestion of the taste/flavor, apparently via a form of Pavlovian conditioning in which the nutrient reinforces the in- gestive response to the flavor (5,21,22,33,34,36,41,56,60,61). Nu- trient-conditioned taste/flavor preferences are sometimes thought to be analogous to, or the converse of, malaise-condit ioned taste/ flavor aversions (7 ,34) . If these two forms of condit ioning are indeed related, it should be possible to demonstrate analogous behavioral changes with each.
It therefore seemed odd that these two forms of conditioning have usually been assessed in different ways. Most studies of f lavor- nutrient conditioning have used a discrimination procedure in which a CS+ flavor is paired with a reinforcer and a C S - flavor is paired with a control treatment (5,21,22,33,34,36,41,56,60,61). A few studies employed single-stimulus acceptance tests, but these seemed to suggest that special conditions were required to obtain conditioning, such as food deprivation (34) or the use of very concentrated nutrients (17) . Malaise-condit ioned aversions, on the other hand, are commonly assessed by a single stimulus tech- nique (3,16,32,37). When aversion studies employed preference tests, the second stimulus was usually plain water rather than a flavored C S - (3,16,32,37).
A recent report showed that dilute carbohydrate infusions stimulated intake of saccharin but not of cherry-flavored water (50) . This result seemed surprising because the same cherry fla-
vor serves as an adequate stimulus for discriminative condition- ing (19,21,59) .
Part I of this article describes the results of several attempts to condition increased ingestion of cherry-flavored water. Sub- sequent sections examine other tastes (Part II) , the possible role of associative mechanisms (Part III) , and aversive conditioning (Part IV) .
The methods are similar those employed in previous studies in this laboratory (50,51 ). Nutrients were delivered via intragas- tric infusions because this provides better experimental control than does giving nutrients by mouth.
GENERAL METHOD
Animals
Female CD rats (Crl :CD BR, Charles River Breeding Labs., Wilmington, M A ) arrived in the laboratory when they were about 8 weeks of age. They were housed individually, maintained on a 12/12 h l ight /dark cycle at 21-25°C, and given Purina Labo- ratory Chow (#5001) ad lib, except where indicated. When not being tested, the rats were given tap water in sipper bottles to drink ad lib; the rats always had some fluid to drink. The rats were maintained in the laboratory for at least 1 week after arrival to permit them to acclimate to their surroundings before surgery began.
595
596 RAMIREZ
Data from some rats were discarded because of mechanical or electrical problems (leaks, electrical shorts, clogged tubing, etc.) or, rarely, complications from surgery. The number of rats reported is the number that provided usable data.
Surgery
The catheters were like those described by Young and Deutsch (64) except that the parts entering the stomach were made of 2-mm-diameter silicone and the parts exiting from the head were made of polyvinyl chloride. They were implanted in a small hole placed in the forestomach, sutured to both the stom- ach and the abdominal wall, exited the body through an incision in the back of the neck, and were anchored to the back of the neck with a stainless steel button. During surgery, rats were an- esthetized with ketamine (90 mg/kg) and acepromazine (1 mg/ kg), supplemented with ether as needed. The rats were allowed to recover for at least 8 days after surgery before they were put in the test chambers.
Apparatus
Rats were housed in hanging stainless steel cages until they had recovered from surgery. Training was carried out in Plexiglas test chambers containing Plexiglas rods with a 0.6-cm-diameter well filled with test fluid. Whenever a rat drank fluid from the well, a level detector activated a peristaltic pump that refilled the well and injected approximately the same amount of fluid into the rat's gastric catheter. The pumping rate was approximately 3.5 ml/min but the pumps were typically on for only a few sec- onds at a time.
Most chambers had only one fluid source, or well. Preference conditioning was conducted with a modified apparatus having two fluid sources approximately 10 cm apart. Preference tests involving rats having no gastric catheters were done in the steel home cages.
Test Procedure
When the rats were placed in the test apparatus, their catheters were connected to an overhead tether with a swivel joint (Instech, Plymouth Meeting, PA). For the first 3 days, the rats were ha- bituated to the apparatus. During this period, they were infused with plain water whenever they drank plain water. On subsequent days, the rats were offered flavored solutions to drink and infused with nutrient solutions or plain water whenever they drank the test fluids. In most experiments, the rats were trained with the test fluid for at least 3 consecutive days; the dependent variable of interest being oral fluid intake.
Substances Offered
Unsweetened cherry Kool-Aid® (Kraft General Foods, White Plains, NY) flavor was used at the same concentration (0.05%) as that used by previous investigators (19,21,59). The carbohy- drate infused in most studies was Maltrin M200 (Grain Process- ing Corp., Muscatine, IA); this maltodextrin is a glucose polymer similar to Polycose® (49). All mixtures were formulated on a weight basis because intake was measured by weighing. Deion- ized water was used to make up solutions.
Statistics
All group data are given as means _ SE. Intake of rats in the test apparatus is reported as grams intake by mouth; the rats re- ceived an equal amount of fluid via their gastric catheters. Be- tween- and within-group comparisons were done for each exper-
imental day by analysis of variance (ANOVA) and t-tests. To avoid any questions about the validity of the statistical tests, probability values were computed by a nonparametric method, variously known as permutation, randomization, or resampling (18,40). The significance of tests involving multiple compari- sons was evaluated by the method recommended by Saville (54). All probability values are two-tailed.
PART I: SACCHARIN VS. CHERRY FLAVOR
EXPERIMENT 1
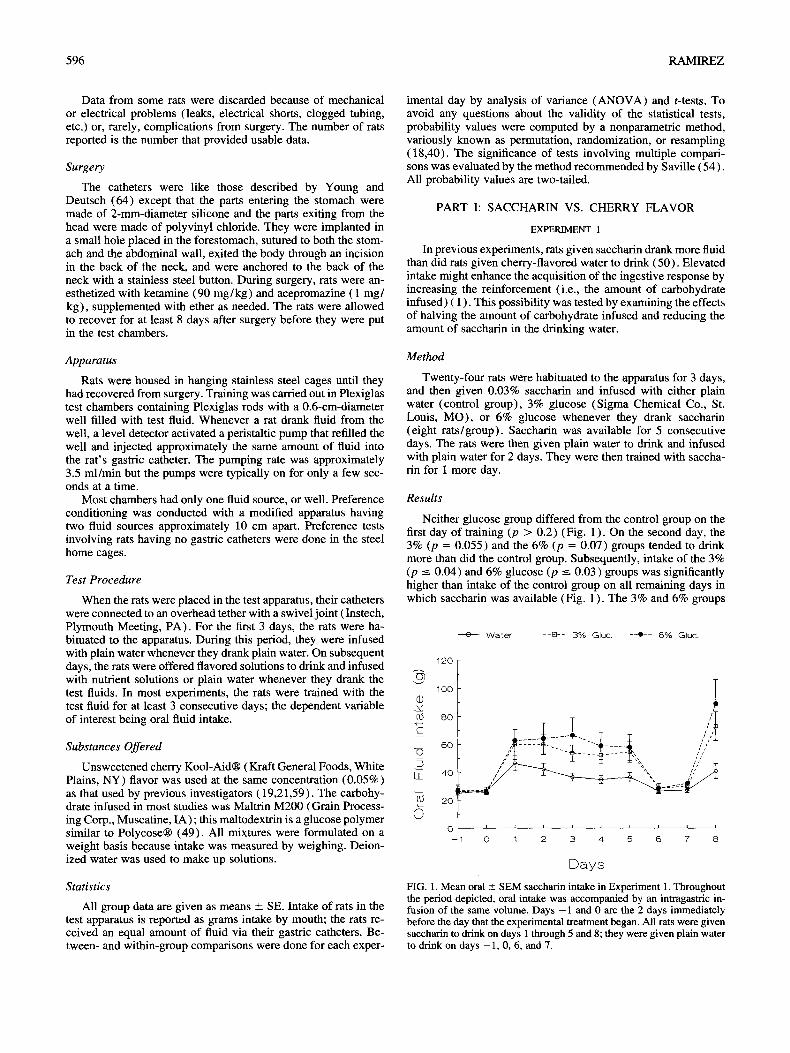
In previous experiments, rats given saccharin drank more fluid than did rats given cherry-flavored water to drink (50). Elevated intake might enhance the acquisition of the ingestive response by increasing the reinforcement (i.e., the amount of carbohydrate infused) ( 1 ). This possibility was tested by examining the effects of halving the amount of carbohydrate infused and reducing the amount of saccharin in the drinking water.
Method
Twenty-four rats were habituated to the apparatus for 3 days, and then given 0.03% saccharin and infused with either plain water (control group), 3% glucose (Sigma Chemical Co., St. Louis, MO), or 6% glucose whenever they drank saccharin (eight rats/group). Saccharin was available for 5 consecutive days. The rats were then given plain water to drink and infused with plain water for 2 days. They were then trained with saccha- rin for 1 more day.
Results
Neither glucose group differed from the control group on the first day of training (p > 0.2) (Fig. 1 ). On the second day, the 3% (p = 0.055) and the 6% (p = 0.07) groups tended to drink more than did the control group. Subsequently, intake of the 3% (p --- 0.04) and 6% glucose (p --- 0.03) groups was significantly higher than intake of the control group on all remaining days in which saccharin was available (Fig. 1 ). The 3% and 6% groups
0 W a t e r --19-- 3 % Gluc. -- l iP-- 6 % Gluc.
1 2 0
03 v
1 0 0
0 2£
80
6 0
kl_ 4O
-~ 20 I
o
T "/I i I , '
J
1 2
I I I I I i
3 4 5 6 7 8
D a y s
FIG. 1. Mean oral _ SEM saccharin intake in Experiment 1. Throughout the period depicted, oral intake was accompanied by an intragastric in- fusion of the same volume. Days - 1 and 0 are the 2 days immediately before the day that the experimental treatment began. All rats were given saccharin to drink on days 1 through 5 and 8; they were given plain water to drink on days -1, 0, 6, and 7.
STIMULUS SPECIFICITY: SACCHARIN VS, CHERRY 597
did not differ on any day (all p > 0.2). Overall ANOVAs were significant on the third, fourth, and last training days (p -< 0.05 ).
Discussion
Halving the amount of carbohydrate infused had remarkably little effect on carbohydrate-induced stimulation of saccharin in- take. Both carbohydrate concentrations increased fluid intake by 16 g or more beginning on the second day of training (Fig. 1 ). This result suggests, at least when 6% carbohydrate is infused, that saccharin's ability to promote intake, and thereby increase the amount of carbohydrate infused, does not greatly contribute to difference between saccharin and cherry flavor.
EXPERIMENTS 2A AND 2B
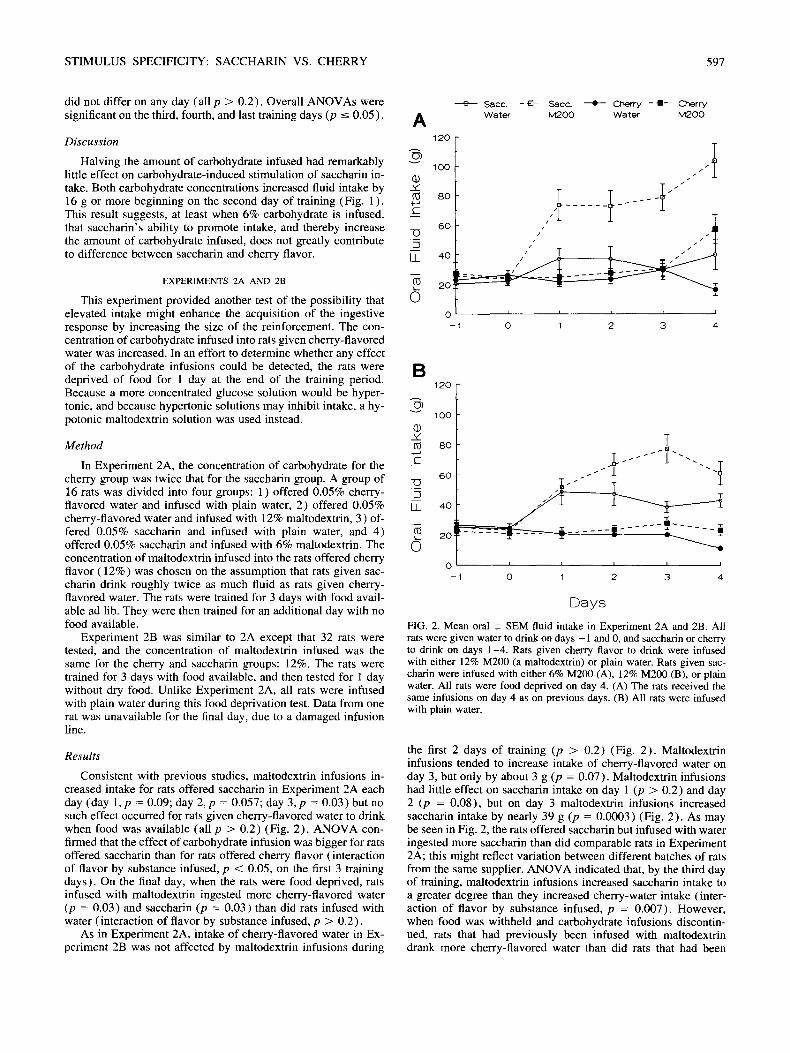
This experiment provided another test of the possibility that elevated intake might enhance the acquisition of the ingestive response by increasing the size of the reinforcement. The con- centration of carbohydrate infused into rats given cherry-flavored water was increased. In an effort to determine whether any effect of the carbohydrate infusions could be detected, the rats were deprived of food for l day at the end of the training period. Because a more concentrated glucose solution would be hyper- tonic, and because hypertonic solutions may inhibit intake, a hy- potonic maltodextrin solution was used instead.
Method
In Experiment 2A, the concentration of carbohydrate for the cherry group was twice that for the saccharin group. A group of 16 rats was divided into four groups: 1 ) offered 0.05% cherry- flavored water and infused with plain water, 2) offered 0.05% cherry-flavored water and infused with 12% maltodextrin, 3) of- fered 0.05% saccharin and infused with plain water, and 4) offered 0.05% saccharin and infused with 6% maltodextrin. The concentration of maltodextrin infused into the rats offered cherry flavor (12%) was chosen on the assumption that rats given sac- charin drink roughly twice as much fluid as rats given cherry- flavored water. The rats were trained for 3 days with food avail- able ad lib. They were then trained for an additional day with no food available.
Experiment 2B was similar to 2A except that 32 rats were tested, and the concentration of maltodextrin infused was the same for the cherry and saccharin groups: 12%. The rats were trained for 3 days with food available, and then tested for 1 day without dry food. Unlike Experiment 2A, all rats were infused with plain water during this food deprivation test. Data from one rat was unavailable for the final day, due to a damaged infusion line.
Results
Consistent with previous studies, maltodextrin infusions in- creased intake for rats offered saccharin in Experiment 2A each day (day 1, p = 0.09; day 2, p = 0.057; day 3, p = 0.03) but no such effect occurred for rats given cherry-flavored water to drink when food was available (all p > 0.2) (Fig. 2). ANOVA con- finned that the effect of carbohydrate infusion was bigger for rats offered saccharin than for rats offered cherry flavor (interaction of flavor by substance infused, p < 0.05, on the first 3 training days). On the final day, when the rats were food deprived, rats infused with maltodextrin ingested more cherry-flavored water (p = 0.03) and saccharin (p = 0.03) than did rats infused with water (interaction of flavor by substance infused, p > 0.2).
As in Experiment 2A, intake of cherry-flavored water in Ex- periment 2B was not affected by maltodextrin infusions during
A
03 v
¢
(5 (-
i
-o
I i
(5
6
1 2 0
1 0 0
8O
6O
40
20
0
0 Sacc . - ~ - Sacc . • C h e r r y - I - C h e r r y
W a t e r M 2 0 0 W a t e r M 2 0 0
T
T ~ _ _
, T / • / / / z
a .
i i i i I
0 1 2 3 4
B 1 2 0
03 v 1 0 0
@ .2£ (5 80
4-a c - I
60 73 i
I
IJ 40
20 ! 6
o
/ / T
i i i i i
0 1 2 3 4
D a y s
FIG. 2. Mean oral _+ SEM fluid intake in Experiment 2A and 2B. All rats were given water to drink on days - 1 and 0, and saccharin or cherry to drink on days 1-4. Rats given cherry flavor to drink were infused with either 12% M200 (a maltodextrin) or plain water. Rats given sac- charin were infused with either 6% M200 (A), 12% M200 (B), or plain water. All rats were food deprived on day 4. (A) The rats received the same infusions on day 4 as on previous days. (B) All rats were infused with plain water.
the first 2 days of training (p > 0.2) (Fig. 2). Maltodextrin infusions tended to increase intake of cherry-flavored water on day 3, but only by about 3 g (p = 0.07). Maltodextrin infusions had little effect on saccharin intake on day 1 (p > 0.2) and day 2 (p = 0.08), but on day 3 maltodextrin infusions increased saccharin intake by nearly 39 g (p = 0.0003) (Fig. 2). As may be seen in Fig. 2, the rats offered saccharin but infused with water ingested more saccharin than did comparable rats in Experiment 2A; this might reflect variation between different batches of rats from the same supplier. ANOVA indicated that, by the third day of training, maltodextrin infusions increased saccharin intake to a greater degree than they increased cherry-water intake (inter- action of flavor by substance infused, p = 0.007). However, when food was withheld and carbohydrate infusions discontin- ued, rats that had previously been infused with maltodextrin drank more cherry-flavored water than did rats that had been

598 RAMIREZ
previously infused with plain water (p = 0.0002) ; this effect did not reach a conventional significance level for rats offered sac- charin (p = 0.07, interaction of flavor by substance infused, p > 0.2).
Discussion
Consistent with Experiment 1, increasing the amount of mat- todextrin infused did not make it easier to observe an effect for rats given cherry-flavored water. Indeed, infusing more malto- dextrin may have hindered the appearance of the effect in the rats offered saccharin. This is probably due to the satiating effects of the concentrated carbohydrate. The amount of maltodextrin infused in the rats offered saccharin in Experiment 2B was sub- stantial, averaging 25 kcal or more per day. Given the well- known satiating effects of carbohydrates, it is remarkable that the rats infused with maltodextrin drank so much saccharin on the third day of training in Experiment 2B.
When food was available ad lib, maltodextrin infusions did not greatly affect intake of cherry-flavored water. When food was withdrawn, rats that had been infused with maltodextrin drank more than did rats that had been infused with water. This oc- curred even when carbohydrate infusions were discontinued dur- ing the food deprivation test. It appears that the rats learned some- thing about the consequences of ingesting the cherry-flavored water even though they did not show large changes in ingestion during the training period.
EXPERIMENT 3
In Experiment 2 there was a nonsignificant trend for malto- dextrin infusions to stimulate intake of cherry-flavored water (Fig. 2). It is therefore not clear whether the maltodextrin infu- sions had no effect at all, or merely a very weak effect. This possibility could be evaluated by examining a more sensitive behavioral technique, preference.
Method
Discussion
These results demonstrate that maltodextrin infusions can in- fluence response to cherry flavor; the failure to demonstrate a significant effect in previous experiments is at least partly due to the use of a less sensitive technique. With a longer training period and a higher concentration of maltodextrin, it may be possible to conditioned an actual preference for Kool-Aid® flavors over plain water (19,59).
EXPERIMENT 4
This experiment evaluated high and low saccharin concentra- tions to evaluate the possible roles of stimulus intensity and un- conditioned preference. The higher concentration used (2%) was slightly aversive ( 12,15 ). The lower concentration used (0.01%) is close to the threshold for stimulation of fluid intake (20,28). Certain aspects of the procedure differ from those employed in other experiments in this series because this experiment was con- ducted before any of the others. These procedural differences do not alter the main conclusions.
Method
A total of 28 rats were trained according to the following sequence; 1 day of habituation in which only water was available, 1 day of training, 1 day in which only water was available, and another day of training. Thirteen rats were offered 0.01% sac- charin to drink. Fourteen were offered 2% saccharin. Half of the rats were infused with 6% glucose whenever they drank saccha- rin and half of the rats were infused with plain water whenever they drank saccharin.
Results
Twenty-one rats were tested in a modified apparatus having two drinking wells. The rats were connected to two infusion pumps via two-channel swivels. The first 3 habituation days were the same as described above; plain water was infused into the rats' stomachs whenever the rats drank plain water. For the next 3 days, 10 rats were infused with 12% maltodextrin whenever they drank 0.05% cherry flavor and 11 rats were infused with 12o F plain water whenever they drank 0.05% cherry flavor. Plain water was infused into the rats' stomachs whenever the rats drank plain "~ v 1 to water.
On the first training day, both wells contained cherry flavor; this was done to insure that all rats drank at least some cherry- ~ so I- flavored water. On the next 2 days, cherry-flavored water was e-
L available from one well and plain water was available from the ~3 60 other well. The lef t - r ight position of the well containing cherry 5 flavor was reversed on successive days. Food was always avail- U_ 40 able ad lib.
Results
Consistent with Experiment 2, total fluid intake was not in- fluenced by maltodextrin infusions (all p > 0.2, data not shown). However, rats infused with maltodextrin showed a greater pref- erence for cherry-flavored water than did the rats infused with water on the second (55 ± 13% vs. 23 ± 10%, p = 0.06) and third training days (52 ± 13 vs. 18 ± 7%, p = 0.02).
For rats given 0.01% saccharin, glucose infusions signifi- cantly increased saccharin intake (p = 0.03 on the first day of training and p = 0.008 on the second day of training) (Fig. 3). However, for rats given 2% saccharin no such effect appeared (both p -> 0.19) (Fig. 3). ANOVA revealed a significant inter- action of saccharin concentration by substance infused for the
0.01% I 0.01% ~ 2% ~ 2% Water Glucose Water Glucose
0 20
o Train 1 Tra in 2
FIG. 3. Mean oral ± SEM saccharin intake for rats given either 0.01% or 2% saccharin in Experiment 4. Training days 1 and 2 are the second and fourth days of the experiment; the rats were given plain water on the first and third days.

STIMULUS SPECIFICITY: SACCHARIN VS. CHERRY 599
second training day (p = 0.009) but not for the first training day (p = 0.07).
Discussion
This experiment is suggests that the unconditioned hedonic effects of the conditioned stimulus may influence the degree to which carbohydrate infusions stimulate intake. These results do not support a role for stimulus intensity because glucose infu- sions did not increase the intake of 2% saccharin, even though it must have a very intense taste.
E X P E R I M E N T 5
The previous experiment suggested that a very dilute saccha- rin solution may not stimulate intake by itself, yet strongly stim- ulate intake when paired with carbohydrate infusions. Experi- ment 5 was originally intended to confirm this. Although this expectation was not confirmed, some unexpected findings pro- vided useful information.
Method
Forty-nine rats were habituated to the apparatus for 3 days according to the usual procedure and then divided into four groups. Twenty-six of the rats were offered 0.01% saccharin; the remaining 23 were offered 0.05% cherry flavor. Half of the rats in each flavor group were infused with plain water and half of the rats were infused with 6% maltodextrin whenever they drank the test flavor. The rats were trained for 3 days with food avail- able. On the next day, the flavors continued to be available but food was then withheld and all rats were infused with plain water.
Results
The rats offered 0.01% saccharin and infused with plain water ingested approximately the same amount of fluid on the first day of training as they did the day before training (p > 0.2). How- ever, carbohydrate infusions did not greatly stimulate saccharin intake in rats given food ad lib during the first 2 days of training (p > 0.2) and had only a moderate effect on the third day (p = 0.053 ) (Fig. 4). When food was withheld, the group infused with maltodextrin drank significantly more saccharin than did the group infused with water (p = 0.006).
The rats offered cherry-flavored water, and infused with plain water, drank significantly less fluid on the first day of training than they did the day before training (p = 0.02). Maltodextrin infusions had no significant effect on intake of cherry-flavored water on the first and second days of training (p --> 0.09) but did increase intake on the third day (p = 0.02) and when food was withheld (p = 0.02) (Fig. 4).
The magnitude of the elevation in fluid intake was similar in rats given 0.01% saccharin- or cherry-flavored water (all p -> 0.18).
Discussion
The results of this experiment would appear to contradict Ex- periment 2 in which 12% maltodextrin infusions had no signifi- cant effect on intake of cherry flavor. However, there was a non- significant trend for 12% maltodextrin infusions to increase intake in Experiment 2. Data from rats given cherry-flavored wa- ter Experiments 2A, 2B, and 5 were pooled and tested with a factorial A N O V A involving 50 rats. Rats infused with 12% mal- todextrin did not differ from the rats in Experiment 5 on any training day (all p > 0.2). Thus, Experiment 5 does not contra- dict Experiments 2A and 2B. Interestingly, the pooled data
0 Sacc. --El-- Sacc. • Cher ry - -m- - Cherry Water M200 Water M20Q
120 ol
IO0 o
2£ 80
6 O -(3 5 ~_ 40
-~ 20 I
(5 0
T T T
i r r I i
0 1 2 3 4
D a y s
FIG. 4. Mean oral _ SEM fluid intake in Experiment 5. On days 1-4, the rats were given either 0.01% saccharin or 0.05% cherry flavor to drink. The maltodextrin (M200) concentration was 6%. All rats were food deprived and infused with water on day 4. For other details, see the caption for Fig. 2.
showed a significant effect for maltodextrin infusions on the sec- ond (p = 0.03) and third day of training (p = 0.01 ). Although pooling data improved statistical significance, it did not change the magnitude of the effect; the increase on the second day was only about 4 g/rat.
The observation that maltodextrin infusions increased intake of cherry-flavored water supports the conclusion from Experi- ment 2 that if a sufficiently sensitive technique is used, an effect can be detected. In this case, high sensitivity was attained by using a relatively large sample size.
It is not known why maltodextrin infusions had a much stronger effect on intake of 0.01% saccharin in Experiment 4 than in this experiment. Potential reasons include the differences in the procedure, and variation between different shipments of rats in sensitivity to saccharin. If this concentration is close to the threshold for reliable conditioning, then high variability between experiments would be expected.
E X P E R I M E N T 6
Experiment 6 tests two explanations for why carbohydrate infusions had a much bigger effect for saccharin solutions than for cherry-flavored water aversiveness and stimulus intensity. If the aversiveness of cherry flavor is the critical attribute, then adding cherry flavor to a saccharin solution should inhibit car- bohydrate-induced stimulation of saccharin intake. If, on the other hand, the crucial attribute is stimulus intensity, then adding cherry flavor to a saccharin solution might enhance conditioning or else have no effect. The amount of cherry flavor used was sufficient to block saccharin preference. In a pilot experiment, nine rats were offered a choice of water vs. a mixture of 0.05% saccharin and 0.1% cherry flavor. Mean 24-h preference for the saccharin/cherry mixture was 47 _ 8% and 56 ___ 8% on 2 suc- cessive days.
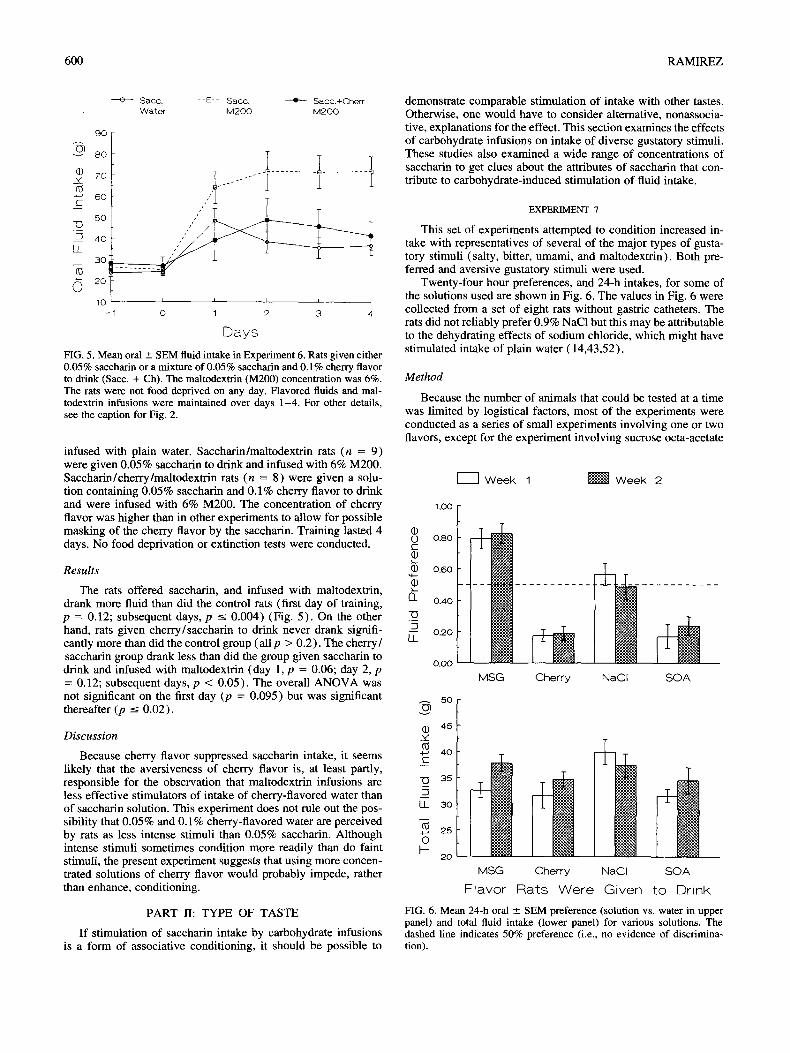
Method
Twenty-seven rats were habituated to the apparatus for 3 days according to the usual procedure and then divided into groups. Control rats (n = 10) were given 0.05% saccharin to drink and
600 RAMIREZ
0 Sacc. - - B - - Sacc. • Secc,+Cherr Wa te r M 2 0 0 M 2 0 0
90 A
(D 70 T . . . . . . . . . . . . . . . . . . 1--- i
~3 -~ 60
~ / , / /
~ 4 0
__ 3 0 N
(~ 2o 10 i ~ I i
- 0 1 2 3 4
[Days
FIG. 5. Mean oral __ SEM fluid intake in Experiment 6. Rats given either 0.05% saccharin or a mixture of 0.05% saccharin and 0.1% cherry flavor to drink (Sacc. + Ch). The maltodextrin (M200) concentration was 6%. The rats were not food deprived on any day. Flavored fluids and mal- todextrin infusions were maintained over days 1-4. For other details, see the caption for Fig. 2.
infused with plain water. Saccharin/maltodextrin rats (n = 9) were given 0.05% saccharin to drink and infused with 6% M200. Saccharin/cherry/maltodextrin rats (n = 8) were given a solu- tion containing 0.05% saccharin and 0.1% cherry flavor to drink and were infused with 6% M200. The concentration of cherry flavor was higher than in other experiments to allow for possible masking of the cherry flavor by the saccharin. Training lasted 4 days. No food deprivation or extinction tests were conducted.
Results
The rats offered saccharin, and infused with maltodextrin, drank more fluid than did the control rats (first day of training, p = 0.12; subsequent days, p < 0.004) (Fig. 5). On the other hand, rats given cherry/saccharin to drink never drank signifi- cantly more than did the control group (a l lp > 0.2). The cherry/ saccharin group drank less than did the group given saccharin to drink and infused with maltodextrin (day 1, p = 0.06; day 2, p = 0.12; subsequent days, p < 0.05). The overall ANOVA was not significant on the first day (p = 0.095) but was significant thereafter (p --< 0.02).
Discussion
Because cherry flavor suppressed saccharin intake, it seems likely that the aversiveness of cherry flavor is, at least partly, responsible for the observation that maltodextrin infusions are less effective stimulators of intake of cherry-flavored water than of saccharin solution. This experiment does not rule out the pos- sibility that 0.05% and 0.1% cherry-flavored water are perceived by rats as less intense stimuli than 0.05% saccharin. Although intense stimuli sometimes condition more readily than do faint stimuli, the present experiment suggests that using more concen- trated solutions of cherry flavor would probably impede, rather than enhance, conditioning.
PART II: TYPE OF TASTE
If stimulation of saccharin intake by carbohydrate infusions is a form of associative conditioning, it should be possible to
demonstrate comparable stimulation of intake with other tastes. Otherwise, one would have to consider alternative, nonassocia- rive, explanations for the effect. This section examines the effects of carbohydrate infusions on intake of diverse gustatory stimuli. These studies also examined a wide range of concentrations of saccharin to get clues about the attributes of saccharin that con- tribute to carbohydrate-induced stimulation of fluid intake.
E X P E R I M E N T 7
This set of experiments attempted to condition increased in- take with representatives of several of the major types of gusta- tory stimuli (salty, bitter, umami, and maltodextrin). Both pre- ferred and aversive gustatory stimuli were used.
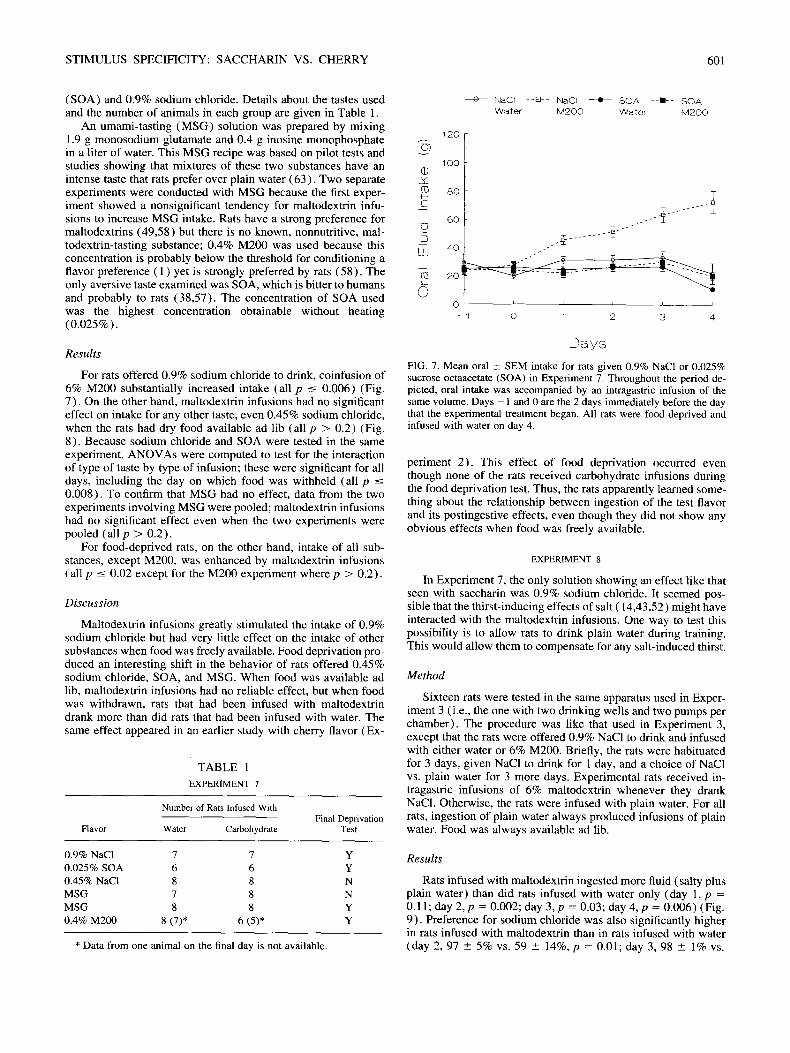
Twenty-four hour preferences, and 24-h intakes, for some of the solutions used are shown in Fig. 6. The values in Fig. 6 were collected from a set of eight rats without gastric catheters. The rats did not reliably prefer 0.9% NaC1 but this may be attributable to the dehydrating effects of sodium chloride, which might have stimulated intake of plain water (14,43,52).
Method
Because the number of animals that could be tested at a time was limited by logistical factors, most of the experiments were conducted as a series of small experiments involving one or two flavors, except for the experiment involving sucrose octa-acetate
r ~ W e e k 1 ~ Week 2
EL
-O 5
LL
1.00
(1) o 0.80 C
0 .60
0 .40
0 . 2 0
0 . 0 0
5O
i1) 4 5 ~,.
4--' 4 0 C
'0 35 5 LL 3o
~ 25
2 0
MSG Cherry NaCI SOA
MSG Cherry NaCI S O A
Flavor Rats Were Given to Drink
FIG. 6. Mean 24-h oral +__ SEM preference (solution vs. water in upper panel) and total fluid intake (lower panel) for various solutions. The dashed line indicates 50% preference (i.e., no evidence of discrimina- tion).
STIMULUS SPECIFICITY: SACCHARIN VS. CHERRY 601
( S O A ) and 0.9% sodium chloride. Details about the tastes used and the number of animals in each group are given in Table 1.
An umami-tast ing ( M S G ) solution was prepared by mixing 1.9 g monosodium glutamate and 0.4 g inosine monophosphate in a liter of water. This MSG recipe was based on pilot tests and studies showing that mixtures of these two substances have an intense taste that rats prefer over plain water (63) . Two separate experiments were conducted with MSG because the first exper- iment showed a nonsignificant tendency for maltodextrin infu- sions to increase M S G intake. Rats have a strong preference for maltodextrins (49,58) but there is no known, nonnutritive, mal- todextrin-tasting substance; 0.4% M200 was used because this concentration is probably below the threshold for conditioning a flavor preference ( 1 ) yet is strongly preferred by rats (58) . The only aversive taste examined was SOA, which is bitter to humans and probably to rats (38,57) . The concentration of SOA used was the highest concentration obtainable without heating (0 .025%).
Results
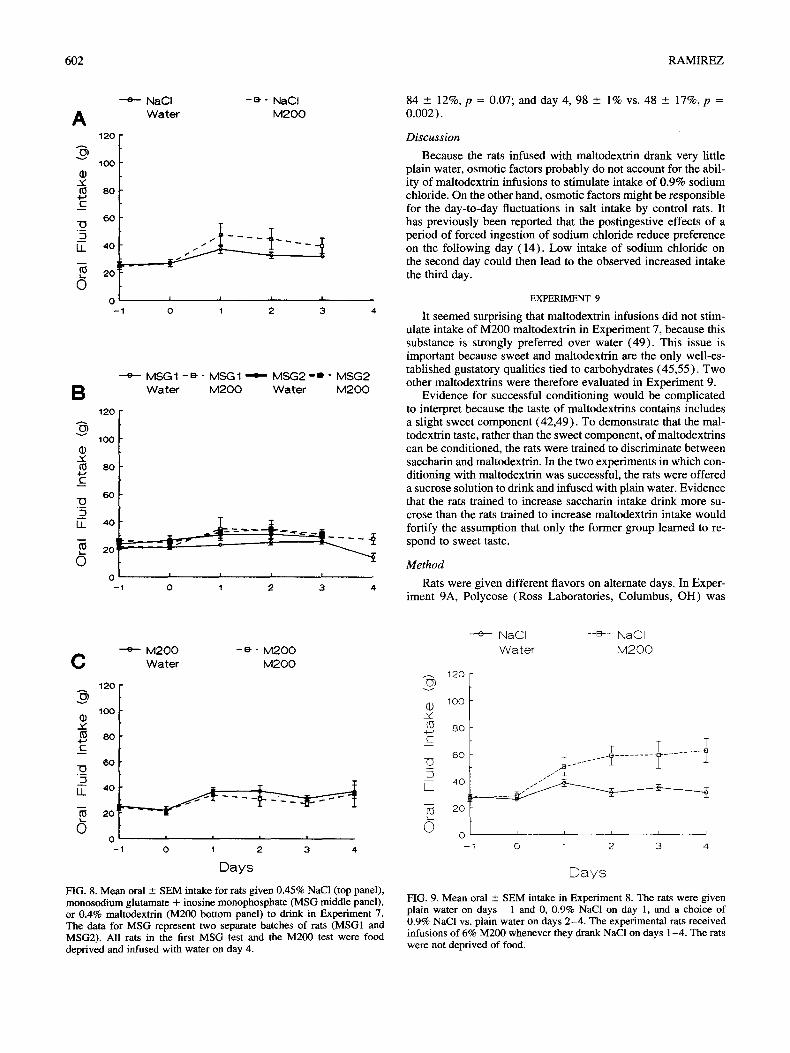
For rats offered 0.9% sodium chloride to drink, coinfusion of 6% M200 substantially increased intake (all p ----- 0.006) (Fig. 7) . On the other hand, maltodextrin infusions had no significant effect on intake for any other taste, even 0.45% sodium chloride, when the rats had dry food available ad lib (all p > 0.2) (Fig. 8) . Because sodium chloride and SOA were tested in the same experiment, A N O V A s were computed to test for the interaction of type of taste by type of infusion; these were significant for all days, including the day on which food was withheld (all p -< 0.008). To confirm that M S G had no effect, data from the two experiments involving MSG were pooled; maltodextrin infusions had no significant effect even when the two experiments were pooled (all p > 0.2).
For food-deprived rats, on the other hand, intake of all sub- stances, except M200, was enhanced by maltodextrin infusions (all p -< 0.02 except for the M200 experiment where p > 0.2).
Discussion
Maltodextr in infusions greatly stimulated the intake of 0.9% sodium chloride but had very little effect on the intake of other substances when food was freely available. Food deprivation pro- duced an interesting shift in the behavior of rats offered 0.45% sodium chloride, SOA, and MSG. When food was available ad lib, maltodextrin infusions had no reliable effect, but when food was withdrawn, rats that had been infused with maltodextrin drank more than did rats that had been infused with water. The same effect appeared in an earlier study with cherry flavor (Ex-
TABLE 1
EXPERIMENT 7
Number of Rats Infused With Final Deprivation
Flavor Water Carbohydrate Test
0.9% NaCI 7 7 Y 0.025% SOA 6 6 Y 0.45% NaCI 8 8 N MSG 7 8 N MSG 8 8 Y 0.4% M200 8 (7)* 6 (5)* Y
* Data from one animal on the final day is not available.
o NaCI --El-- NaCI • SOA - - i - - SOA Water K4200 Water M20Q
120
(33
100 0 2d
8o £
6 0 ~D
~ 4o
20 !
5 0
T .5 . . . . . . . . ±
"0 n I I p ~
0 1 2 3 4
D a y s
FIG. 7. Mean oral + SEM intake for rats given 0.9% NaC1 or 0.025% sucrose octaacetate (SOA) in Experiment 7. Throughout the period de- picted, oral intake was accompanied by an intragastric infusion of the same volume. Days - 1 and 0 are the 2 days immediately before the day that the experimental treatment began. All rats were food deprived and infused with water on day 4.
periment 2) . This effect of food deprivation occurred even though none of the rats received carbohydrate infusions during the food deprivation test. Thus, the rats apparently learned some- thing about the relationship between ingestion of the test flavor and its postingestive effects, even though they did not show any obvious effects when food was freely available.
EXPERIMENT 8
In Experiment 7, the only solution showing an effect like that seen with saccharin was 0.9% sodium chloride. It seemed pos- sible that the thirst-inducing effects of salt (14,43,52) might have interacted with the maltodextrin infusions. One way to test this possibility is to allow rats to drink plain water during training. This would allow them to compensate for any salt-induced thirst.
Method
Sixteen rats were tested in the same apparatus used in Exper- iment 3 (i.e., the one with two drinking wells and two pumps per chamber) . The procedure was like that used in Experiment 3, except that the rats were offered 0.9% NaC1 to drink and infused with either water or 6% M200. Briefly, the rats were habituated for 3 days, given NaC1 to drink for 1 day, and a choice of NaC1 vs. plain water for 3 more days. Experimental rats received in- tragastric infusions of 6% maltodextrin whenever they drank NaC1. Otherwise, the rats were infused with plain water. For all rats, ingestion of plain water always produced infusions of plain water. Food was always available ad lib.
Results
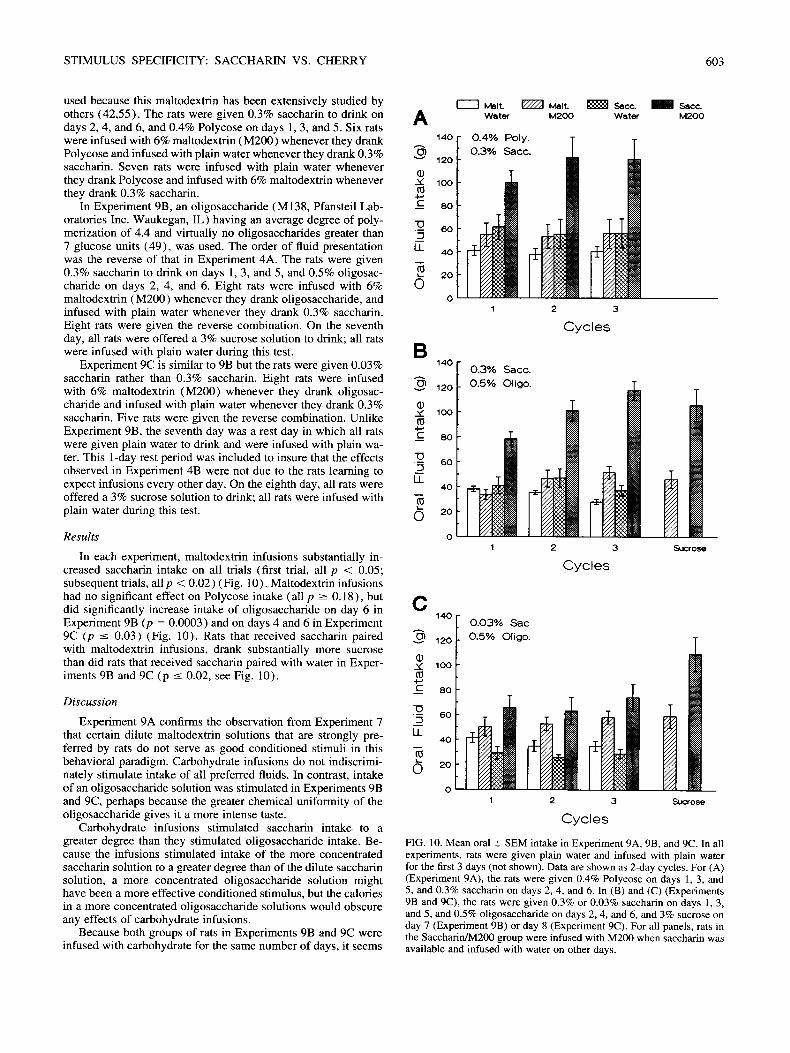
Rats infused with maltodextrin ingested more fluid (salty plus plain water) than did rats infused with water only (day 1, p = 0.11; day 2, p = 0.002; day 3, p = 0.03; day 4, p = 0.006) (Fig. 9) , Preference for sodium chloride was also significantly higher in rats infused with maltodextrin than in rats infused with water (day 2, 97 _+ 5% vs. 59 + 1 4 % , p = 0.01; day 3, 98 _+ 1% vs.
602 RAMIREZ
A A
loo ¢ x~
80
E
120
B
5 E 4o
"~ 2o I
o
120
loo 0
~ 8 0
c -
60 13 5 LL 40
-~ 20 ~
(3 o
o NaCl -B - - NaCI Wa te r M 2 0 0
I I I I
o I 2 3
o M S G I - e " M S G I - " ~ M S G 2 - • " MSG2 W a t e r M 2 0 0 Wate r M 2 0 0
_
~ t - - - { -
I I I I
o 1 2 3 4
84 _ 1 2 % , p = 0.07; and day 4, 98 ___ 1% vs. 48 4- 17%, p = 0.002).
Discussion
Because the rats infused with maltodextrin drank very little plain water, osmotic factors probably do not account for the abil- ity of maltodextfin infusions to stimulate intake of 0.9% sodium chloride. On the other hand, osmotic factors might be responsible for the day-to-day fluctuations in salt intake by control rats. It has previously been reported that the postingestive effects of a period of forced ingestion of sodium chloride reduce preference on the following day (14) . Low intake of sodium chloride on the second day could then lead to the observed increased intake the third day.
EXPERIMENT 9
It seemed surprising that maltodextrin infusions did not stim- ulate intake of M200 maltodextrin in Experiment 7, because this substance is strongly preferred over water (49) . This issue is important because sweet and maltodextrin are the only well-es- tablished gustatory qualities tied to carbohydrates (45,55) . Two other maltodextrins were therefore evaluated in Experiment 9.
Evidence for successful condit ioning would be complicated to interpret because the taste of maltodextfins contains includes a slight sweet component (42,49) . To demonstrate that the mal- todextrin taste, rather than the sweet component, of maltodextrins can be conditioned, the rats were trained to discriminate between saccharin and maltodextrin. In the two experiments in which con- ditioning with maltodextrin was successful, the rats were offered a sucrose solution to drink and infused with plain water. Evidence that the rats trained to increase saccharin intake drink more su- crose than the rats trained to increase maltodextrin intake would fortify the assumption that only the former group learned to re- spond to sweet taste.
Method
Rats were given different flavors on alternate days. In Exper- iment 9A, Polycose (Ross Laboratories, Columbus, OH) was
o M 2 0 0 - e - M 2 0 0 C Wate r M 2 0 0
1 2 0
1 0 0 ¢ l
80
"0 '5
"~ 20 ]
(3 0 i i i !
- 0 1 2 3 4
Days FIG. 8. Mean oral _+ SEM intake for rats given 0.45% NaCl (top panel), monosodium glutamate + inosine monophosphate (MSG middle panel), or 0.4% maltodextrin (M200 bottom panel) to drink in Experiment 7. The data for MSG represent two separate batches of rats (MSG1 and MSG2). All rats in the first MSG test and the M200 test were food deprived and infused with water on day 4.
o NaCI --B-- NaCI
W a t e r M 2 0 0
120 O3
v
(D 1 O0
2£ 8O
60
- - 40 II
~ 2O
© o
T T . . . . . ~, T . . . . . . . I . . . . . . . . iN - - l
t 1 t i i
0 1 2 3 4
D a y s
FIG. 9. Mean oral __ SEM intake in Experiment 8. The rats were given plain water on days - 1 and 0, 0.9% NaC1 on day 1, and a choice of 0.9% NaC1 vs. plain water on days 2-4. The experimental rats received infusions of 6% M200 whenever they drank NaC1 on days 1-4. The rats were not deprived of food.
STIMULUS SPECIFICITY: SACCHARIN VS. CHERRY 603
used because this maltodextrin has been extensively studied by others (42,55) . The rats were given 0.3% saccharin to drink on days 2, 4, and 6, and 0.4% Polycose on days 1, 3, and 5. Six rats were infused with 6% maltodextrin ( M 2 0 0 ) whenever they drank Polycose and infused with plain water whenever they drank 0.3% saccharin. Seven rats were infused with plain water whenever they drank Polycose and infused with 6% maltodextrin whenever they drank 0.3% saccharin.
In Experiment 9B, an oligosaccharide (M138, Pfansteil Lab- oratories Inc. Waukegan, IL) having an average degree of poly- merization of 4.4 and virtually no oligosaccharides greater than 7 glucose units (49) , was used. The order of fluid presentation was the reverse of that in Experiment 4A. The rats were given 0.3% saccharin to drink on days 1, 3, and 5, and 0.5% oligosac- charide on days 2, 4, and 6. Eight rats were infused with 6% maltodextrin ( M 2 0 0 ) whenever they drank oligosaccharide, and infused with plain water whenever they drank 0.3% saccharin. Eight rats were given the reverse combination. On the seventh day, all rats were offered a 3% sucrose solution to drink; all rats were infused with plain water during this test.
Experiment 9C is similar to 9B but the rats were given 0.03% saccharin rather than 0.3% saccharin. Eight rats were infused with 6% maltodextrin (M200) whenever they drank oligosac- charide and infused with plain water whenever they drank 0.3% saccharin. Five rats were given the reverse combination. Unlike Experiment 9B, the seventh day was a rest day in which all rats were given plain water to drink and were infused with plain wa- ter. This 1-day rest period was included to insure that the effects observed in Experiment 4B were not due to the rats learning to expect infusions every other day. On the eighth day, all rats were offered a 3% sucrose solution to drink; all rats were infused with plain water during this test.
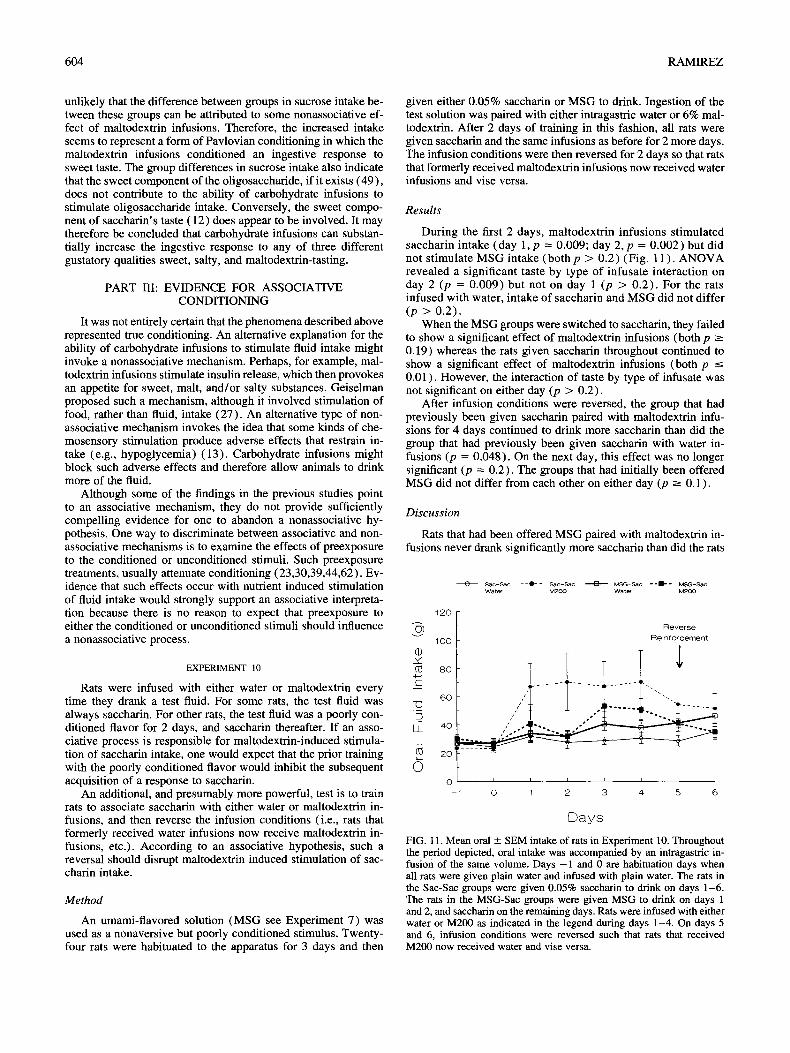
Results
In each experiment, maltodextrin infusions substantially in- creased saccharin intake on all trials (first trial, all p < 0.05; subsequent trials, a l lp < 0.02) (Fig. 10). Mattodextr in infusions had no significant effect on Polycose intake (all p >- 0.18) , but did significantly increase intake of oligosaccharide on day 6 in Experiment 9B (p = 0.0003) and on days 4 and 6 in Experiment 9C (p -< 0.03) (Fig. 10). Rats that received saccharin paired with maltodextrin infusions, drank substantially more sucrose than did rats that received saccharin paired with water in Exper- iments 9B and 9C (p -< 0.02, see Fig. 10).
Discussion
Experiment 9A confirms the observation from Experiment 7 that certain dilute maltodextrin solutions that are strongly pre- ferred by rats do not serve as good conditioned stimuli in this behavioral paradigm. Carbohydrate infusions do not indiscrimi- nately stimulate intake of all preferred fluids. In contrast, intake of an oligosaccharide solution was stimulated in Experiments 9B and 9C, perhaps because the greater chemical uniformity of the oligosaccharide gives it a more intense taste.
Carbohydrate infusions stimulated saccharin intake to a greater degree than they stimulated oligosaccharide intake. Be- cause the infusions stimulated intake of the more concentrated saccharin solution to a greater degree than of the dilute saccharin solution, a more concentrated oligosaccharide solution might have been a more effective condit ioned stimulus, but the calories in a more concentrated oligosaccharide solutions would obscure any effects of carbohydrate infusions.
Because both groups of rats in Experiments 9B and 9C were infused with carbohydrate for the same number of days, it seems
A A
v
,¢ 63
. 4 . a
E
-(3
LL
63
6
B
._Y. 63 c"
"(3 5
LL
6
I
140
120
lOO
80
60
40
20
o
140
120
100
80
60
40
20
0
r - - - - i Melt. ~ Malt. Water M200
0.4% Poly, 0.3% S a t e
1
Sac¢. Water
2 3
C y c l e s
0.3% Sacc. 0.5% Oligo.
1 2 3
C y c l e s
m
M200
C 140
~ 120
(9 ._~ IO0
C 8 0
-O '-- 60
kL 40
m (~ 2 o
o
0.03% Sac 0.5% Oligo.
1 2 3
C y c l e s
1 FIG. 10. Mean oral + SEM intake in Experiment 9A, 9B, and 9C. In all experiments, rats were given plain water and infused with plain water for the first 3 days (not shown). Data are shown as 2-day cycles. For (A) (Experiment 9A), the rats were given 0.4% Polycose on days 1, 3, and 5, and 0.3% saccharin on days 2, 4, and 6. In (B) and (C) (Experiments 9B and 9C), the rats were given 0.3% or 0.03% saccharin on days 1, 3, and 5, and 0.5% oligosaccharide on days 2, 4, and 6, and 3% sucrose on day 7 (Experiment 9B) or day 8 (Experiment 9C). For all panels, rats in the Saccharin/M200 group were infused with M200 when saccharin was available and infused with water on other days.
604 RAMIREZ
unlikely that the difference between groups in sucrose intake be- tween these groups can be attributed to some nonassociative ef- fect of maltodextrin infusions. Therefore, the increased intake seems to represent a form of Pavlovian conditioning in which the maltodextrin infusions conditioned an ingestive response to sweet taste. The group differences in sucrose intake also indicate that the sweet component of the oligosaccharide, if it exists (49) , does not contribute to the ability of carbohydrate infusions to stimulate oligosaccharide intake. Conversely, the sweet compo- nent of saccharin 's taste (12) does appear to be involved. It may therefore be concluded that carbohydrate infusions can substan- tially increase the ingestive response to any of three different gustatory qualities sweet, salty, and maitodextrin-tasting.
PART III: EVIDENCE FOR ASSOCIATIVE CONDITIONING
It was not entirely certain that the phenomena described above represented true conditioning. An alternative explanation for the ability of carbohydrate infusions to stimulate fluid intake might invoke a nonassociative mechanism. Perhaps, for example, mal- todextrin infusions stimulate insulin release, which then provokes an appetite for sweet, malt, and /o r salty substances. Geiselman proposed such a mechanism, al though it involved stimulation of food, rather than fluid, intake (27) . An alternative type of non- associative mechanism invokes the idea that some kinds of che- mosensory stimulation produce adverse effects that restrain in- take (e.g., hypoglycemia) (13) . Carbohydrate infusions might block such adverse effects and therefore allow animals to drink more of the fluid.
Al though some of the findings in the previous studies point to an associative mechanism, they do not provide sufficiently compelling evidence for one to abandon a nonassociative hy- pothesis. One way to discriminate between associative and non- associative mechanisms is to examine the effects of preexposure to the conditioned or uncondit ioned stimuli. Such preexposure treatments, usually attenuate conditioning (23,30,39,44,62). Ev- idence that such effects occur with nutrient induced stimulation of fluid intake would strongly support an associative interpreta- tion because there is no reason to expect that preexposure to either the conditioned or uncondit ioned stimuli should influence a nonassociative process.
EXPERIMENT 10
Rats were infused with either water or maltodextrin every time they drank a test fluid. For some rats, the test fluid was always saccharin. For other rats, the test fluid was a poorly con- ditioned flavor for 2 days, and saccharin thereafter. If an asso- ciative process is responsible for maltodextrin-induced stimula- tion of saccharin intake, one would expect that the prior training with the poorly condit ioned flavor would inhibit the subsequent acquisition of a response to saccharin.
An additional, and presumably more powerful, test is to train rats to associate saccharin with either water or maltodextrin in- fusions, and then reverse the infusion conditions (i.e., rats that formerly received water infusions now receive maltodextrin in- fusions, etc.). According to an associative hypothesis, such a reversal should disrupt maltodextrin induced stimulation of sac- charin intake.
Method
An umami-flavored solution ( M S G see Experiment 7) was used as a nonaversive but poorly conditioned stimulus. Twenty- four rats were habituated to the apparatus for 3 days and then
given either 0.05% saccharin or M S G to drink. Ingestion of the test solution was paired with either intragastric water or 6% mal- todextrin. After 2 days of training in this fashion, all rats were given saccharin and the same infusions as before for 2 more days. The infusion conditions were then reversed for 2 days so that rats that formerly received maitodextrin infusions now received water infusions and vise versa.
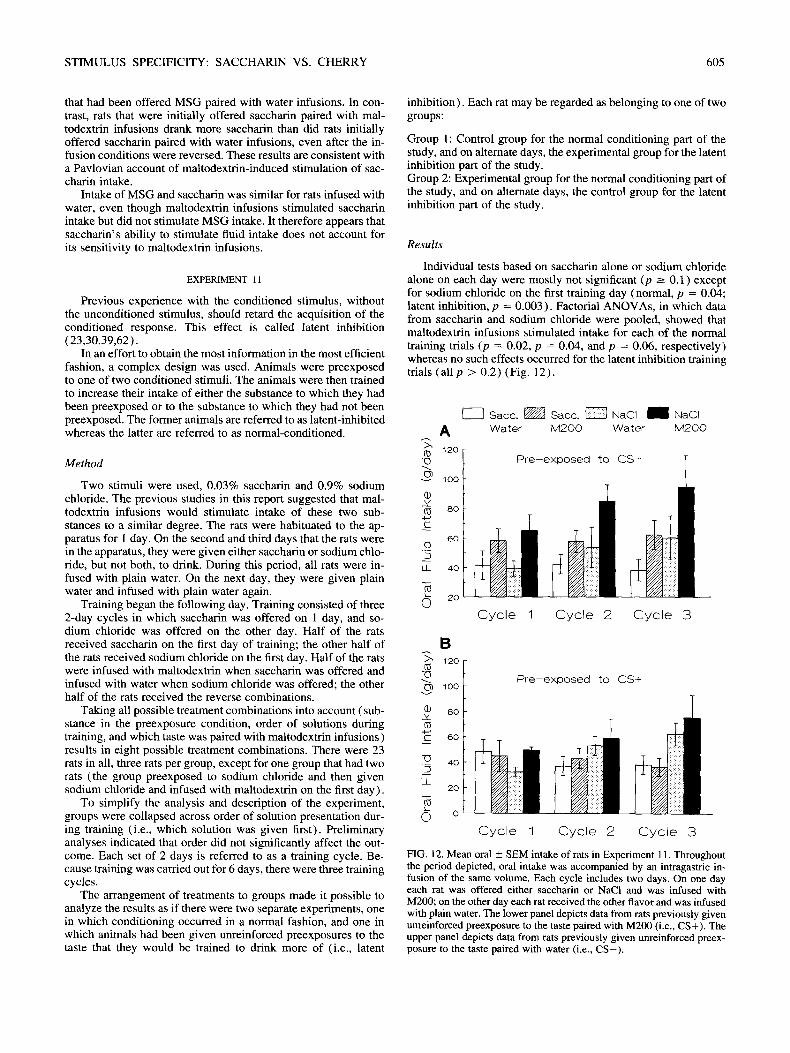
Results
During the first 2 days, mal todext r in infus ions s t imulated sacchar in in take (day 1, p = 0.009; day 2, p = 0 .002) but did not s t imulate M S G intake (bo th p > 0.2) (Fig. 11 ). A N O V A revealed a s ignif icant taste by type of infusate in terac t ion on day 2 (p = 0 .009) but not on day 1 (p > 0 .2) . For the rats infused with water, intake of sacchar in and M S G did not differ (p > 0 .2 ) .
When the M S G groups were switched to saccharin, they failed to show a significant effect of maltodextrin infusions (both p 0.19) whereas the rats given saccharin throughout continued to show a significant effect of maltodextrin infusions (both p -< 0.01 ). However, the interaction of taste by type of infusate was not significant on either day (p > 0.2).
After infusion conditions were reversed, the group that had previously been given saccharin paired with maltodextrin infu- sions for 4 days continued to drink more saccharin than did the group that had previously been given saccharin with water in- fusions (p = 0.048). On the next day, this effect was no longer significant (p = 0.2) . The groups that had initially been offered MSG did not differ from each other on either day (p >-- 0.1 ).
Discussion
Rats that had been offered M S G paired with maltodextrin in- fusions never drank significantly more saccharin than did the rats
0 Sac-Sac - - ~ - Sac-Sac FI MSG-SaC - - I I - - MSG-Sac Water M200 wa t t " t¢200
1 2 0
03 1 0 0
@ 2d 6~ 8o c
6 0 7:3
L 40
-~ 20
0
R e v e r s e
R e i n f o r c e m e n t
i L I i i i i
0 1 2 3 4 5 6
D a y s
FIG. 11. Mean oral _+ SEM intake of rats in Experiment 10. Throughout the period depicted, oral intake was accompanied by an intragastric in- fusion of the same volume. Days - 1 and 0 are habituation days when all rats were given plain water and infused with plain water. The rats in the Sac-Sac groups were given 0.05% saccharin to drink on days 1-6. The rats in the MSG-Sac groups were given MSG to drink on days 1 and 2, and saccharin on the remaining days. Rats were infused with either water or M200 as indicated in the legend during days 1-4. On days 5 and 6, infusion conditions were reversed such that rats that received M200 now received water and vise versa.
STIMULUS SPECIFICITY: SACCHARIN VS. CHERRY 605
that had been offered MSG paired with water infusions. In con- trast, rats that were initially offered saccharin paired with mal- todextrin infusions drank more saccharin than did rats initially offered saccharin paired with water infusions, even after the in- fusion conditions were reversed. These results are consistent with a Pavlovian account of maltodextrin-induced stimulation of sac- charin intake.
Intake of M S G and saccharin was similar for rats infused with water, even though maltodextrin infusions stimulated saccharin intake but did not stimulate MSG intake. It therefore appears that saccharin 's ability to stimulate fluid intake does not account for its sensitivity to maltodextrin infusions.
EXPERIMENT 11
Previous experience with the condit ioned stimulus, without the uncondit ioned stimulus, should retard the acquisition of the condit ioned response. This effect is called latent inhibit ion (23,30,39,62).
In an effort to obtain the most information in the most efficient fashion, a complex design was used. Animals were preexposed to one of two condit ioned stimuli. The animals were then trained to increase their intake of either the substance to which they had been preexposed or to the substance to which they had not been preexposed. The former animals are referred to as latent-inhibited whereas the latter are referred to as normal-conditioned.
Method
Two stimuli were used, 0.03% saccharin and 0.9% sodium chloride. The previous studies in this report suggested that mal- todextrin infusions would stimulate intake of these two sub- stances to a similar degree. The rats were habituated to the ap- paratus for 1 day. On the second and third days that the rats were in the apparatus, they were given either saccharin or sodium chlo- ride, but not both, to drink. During this period, all rats were in- fused with plain water. On the next day, they were given plain water and infused with plain water again.
Training began the following day. Training consisted of three 2-day cycles in which saccharin was offered on 1 day, and so- dium chloride was offered on the other day. Half of the rats received saccharin on the first day of training; the other half of the rats received sodium chloride on the first day. Half of the rats were infused with maltodextrin when saccharin was offered and infused with water when sodium chloride was offered; the other half of the rats received the reverse combinations.
Taking all possible treatment combinat ions into account (sub- stance in the preexposure condition, order of solutions during training, and which taste was paired with maltodextrin infusions) results in eight possible treatment combinations. There were 23 rats in all, three rats per group, except for one group that had two rats ( the group preexposed to sodium chloride and then given sodium chloride and infused with maltodextrin on the first day) .
To simplify the analysis and description of the experiment, groups were collapsed across order of solution presentation dur- ing training (i.e., which solution was given first). Preliminary analyses indicated that order did not significantly affect the out- come. Each set of 2 days is referred to as a training cycle. Be- cause training was carried out for 6 days, there were three training cycles.
The arrangement of treatments to groups made it possible to analyze the results as if there were two separate experiments, one in which condit ioning occurred in a normal fashion, and one in which animals had been given unreinforced preexposures to the taste that they would be trained to drink more of (i.e., latent
inhibi t ion) . Each rat may be regarded as belonging to one of two groups:
Group 1: Control group for the normal conditioning part of the study, and on alternate days, the experimental group for the latent inhibit ion part of the study. Group 2: Experimental group for the normal condit ioning part of the study, and on alternate days, the control group for the latent inhibition part of the study.
Results
Individual tests based on saccharin alone or sodium chloride alone on each day were mostly not significant (p --> 0.1 ) except for sodium chloride on the first training day (normal, p = 0.04; latent inhibition, p = 0.003). Factorial ANOVAs, in which data from saccharin and sodium chloride were pooled, showed that maltodextrin infusions stimulated intake for each of the normal training trials (p = 0.02, p = 0.04, and p = 0.06, respectively) whereas no such effects occurred for the latent inhibition training trials ( a l l p > 0.2) (Fig. 12).
F~ A 120
-0
1 0 0
(b N 80
c 6 0
K? 5 LL 4 0
@ 2o
] Sacc. ~ Sacc. ~ NaCI I NaCI Water M 2 O O Water M200
Pre-exposed to C!-
C y c l e 1 C y c l e 2 C y c l e 3
B 120
. ~ 100
(1) 8o i
~ 60
~ 4o 5 i
kl_ 2O
6 o
Pre-exposed to CS+
C y c l e 1 C y c l e 2 C y c l e 3
FIG. 12. Mean oral _+ SEM intake of rats in Experiment 11. Throughout the period depicted, oral intake was accompanied by an intragastric in- fusion of the same volume. Each cycle includes two days. On one day each rat was offered either saccharin or NaCI and was infused with M200; on the other day each rat received the other flavor and was infused with plain water. The lower panel depicts data from rats previously given unreinforced preexposure to the taste paired with M200 (i.e., CS +). The upper panel depicts data from rats previously given unreinforced preex- posure to the taste paired with water (i.e., CS-) .

606 RAMIREZ
To maximize statistical power, data from the three training cycles were averaged. Maltodextrin infusions stimulated fluid in- take for the normal training trials (p = 0.02) but not for the latent inhibition trials (p > 0.2). An analysis based on the average of the days in which maltodextrin was infused showed that intake was greater for rats trained in the normal fashion than for rats having previous unreinforced experience with the test fluid (i.e., latent-inhibition, p = 0.054).
Discussion
The results fit expectations based on an associative mecha- nism. Maltodextrin infusions did not reliably stimulate intake in animals preexposed to the training fluid. There is no compelling reason to assume that stimulus preexposure should have damp- ened nonassociative effects.
Although the normally conditioned rats showed a moderately large effect, it was not quite as large as in other experiments in this study. The rats given saccharin with maltodextrin infusions ingested only 46% more fluid than did the rats given saccharin and water infusions. Inspection of the means and SEs of this and of previous experiments suggests that this is merely a statistical accident resulting from the use of a small numbers of animals per group.
PART IV: AVERSIVE CONDITIONING
Because malaise conditioned taste/flavor aversion is presum- ably the converse of carbohydrate conditioned taste/flavor ac- ceptance, the literature on conditioned aversions might provide useful insights about why conditioning occurs more readily with some stimuli than with others. Studies of conditioned taste/flavor aversions have occasionally identified tastes/flavors that condi- tion poorly or unusually well (4,9,11,31 ). The concept of sali- ence has been invoked to account for this phenomenon (35). It might therefore be hypothesized that the infusion experiments inadvertently employed the some of most salient and least salient stimuli.
The present experiments attempted to determine whether some of the stimuli used in these studies differ in their salience for malaise-conditioned aversions. The procedures used are based on those typically used for conditioned flavor aversions. The first experiment compared saccharin and cherry-flavored wa- ter. The second experiment compared saccharin and an umami- flavored (MSG) fluid.
EXPERIMENT 12
Method
Twenty-five rats were maintained and tested as described in previous reports (46,47). All tests were conducted on Mondays, Wednesdays, and Fridays during the first hour of the dark part of the light/dark cycle. The rats were given plain water ad lib from Saturday through Sunday each week. To encourage the rats to drink promptly, the only illumination during these tests was provided by dim red lights.
In the first two trials, the rats were habituated to the procedure by depriving them of water for 24 h and then giving them deion- ized water to drink overnight. On the third trial, the rats were water deprived overnight and given either plain water (nine rats), 0.05% saccharin (eight rats), or 0.05% cherry-flavored Kool-Aid (eight rats) to drink for 60 min. The rats were then injected IP with 1 ml 0.15 M lithium chloride per 100 g body weight. The group given plain water is a control for nonspecific effects of lithium chloride treatment. The rats were given tap water to drink 30-60 min after the lithium chloride injection.
After the training trial, the rats were given two additional ha- bituation trials in which they were water deprived, and then of- fered deionized water. On the next test, the rats were water de- prived for 24 h once again, and then given either cherry-flavored water or saccharin solution for 30 rain during the first hour of the dark part of the light/dark cycle. Beginning 60 min after this test, the rats were given plain water to drink overnight, and were then water deprived for 24 h once again. The rats were then retested according to the same procedure as for the previous test but the rats that were previously given cherry flavor were given saccharin and vise versa.
Results
Rats trained to avoid saccharin, drank less saccharin on the test days than did the control rats (p = 0.003) and the rats trained to avoid cherry-flavored water (p = 0.01; overall ANOVA, p = 0.008) (Fig. 13). Conversely, rats trained to avoid cherry flavor drank less cherry-flavored water on the test days than did the control rats (p = 0.004) and the rats trained to avoid saccharin (p = 0.001; overall ANOVAp = 0.002).
The absolute magnitude of the aversion was not greatly influ- enced by the taste used. Rats trained to avoid saccharin ingested 3 g less saccharin than did the control rats, whereas rats trained to avoid cherry-flavored water ingested 4 g less cherry flavor than did the control rats (Fig. 13).
Discussion
It appears that 0.05% cherry flavor is reasonably salient in a flavor aversion experiment. Significant conditioning was ob- served in a single training trial even though presentation of the unconditioned stimulus occurred 60 min after presentation of the conditioned stimulus, a fairly long delay. Training produced a moderate degree of aversion, which was not so great that a base- ment effect, or so strong that a ceiling effect, might have ob- scured differences between the two stimuli (Fig. 13).
EXPERIMENT 13
In Experiment 12, control rats were trained with plain water and received the unconditioned stimulus. However, in most of
Substance Rats Were Trained to Avoid
Control ~ Saccharin ~ Cherry
15 v
(1)
10
- - 5 LL
-8 O o
iiil Sacchar in
Fluid
Cher ry
O f f e r e d
FIG. 13. Mean ___ SEM fluid intake in Experiment 12. Rats that had been trained to avoid saccharin, cherry flavor, or nothing (control) were of- fered saccharin solution or cherry-flavored water for 30 rain.
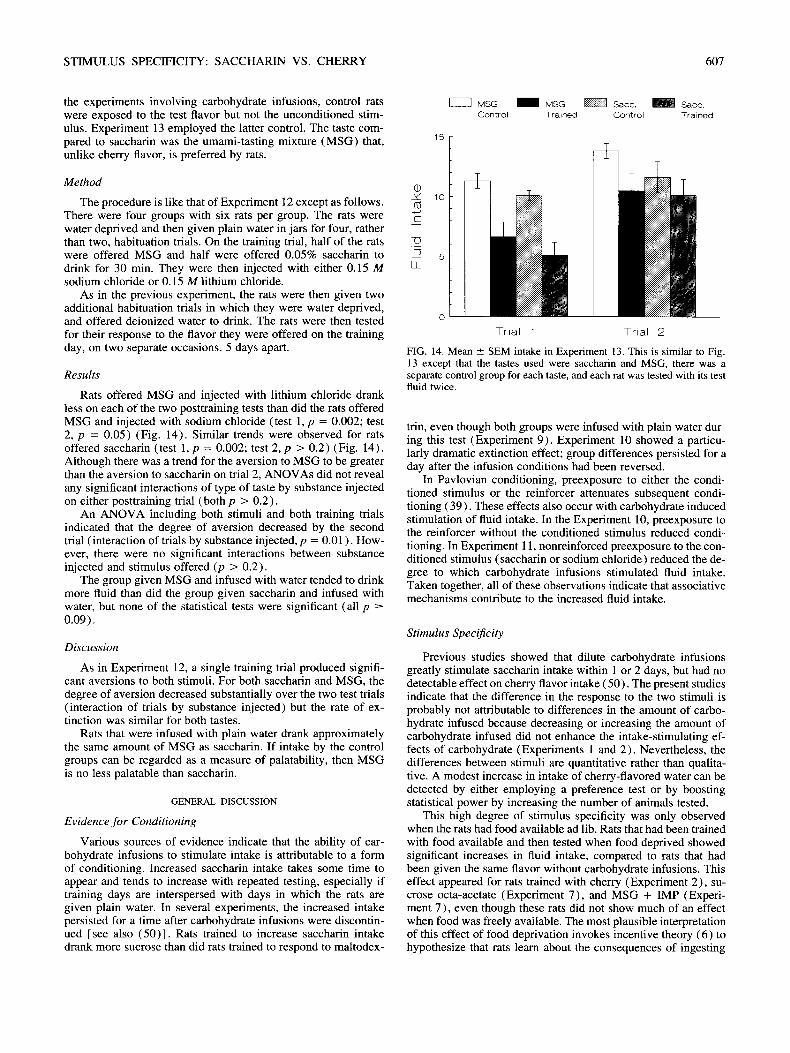
the experiments involving carbohydrate infusions, control rats were exposed to the test flavor but not the uncondit ioned stim- ulus. Experiment 13 employed the latter control. The taste com- pared to saccharin was the umami-tast ing mixture ( M S G ) that, unlike cherry flavor, is preferred by rats.
I I MSG I MSG ~ Sate. ~ Sate. Control Trained Control Trained
Method
The procedure is like that of Experiment 12 except as follows. There were four groups with six rats per group. The rats were water deprived and then given plain water in jars for four, rather than two, habituation trials. On the training trial, half of the rats were offered M S G and half were offered 0.05% saccharin to drink for 30 min. They were then injected with either 0.15 M sodium chloride or 0.15 M lithium chloride.
As in the previous experiment, the rats were then given two additional habituation trials in which they were water deprived, and offered deionized water to drink. The rats were then tested for their response to the flavor they were offered on the training day, on two separate occasions, 5 days apart.
Results
Rats offered M S G and injected with l i thium chloride drank less on each of the two posttraining tests than did the rats offered M S G and injected with sodium chloride (test 1, p = 0.002; test 2, p = 0.05) (Fig. 14). Similar trends were observed for rats offered saccharin (test 1, p = 0.002; test 2, p > 0.2) (Fig. 14). Al though there was a trend for the aversion to MSG to be greater than the aversion to saccharin on trial 2, ANOVAs did not reveal any significant interactions of type of taste by substance injected on either posttraining trial (both p > 0.2).
An A N O V A including both stimuli and both training trials indicated that the degree of aversion decreased by the second trial ( interaction of trials by substance injected, p = 0.01 ). How- ever, there were no significant interactions between substance injected and stimulus offered (p > 0.2).
The group given MSG and infused with water tended to drink more fluid than did the group given saccharin and infused with water, but none of the statistical tests were significant (all p -> 0.09).
Discussion
As in Experiment 12, a single training trial produced signifi- cant aversions to both stimuli. For both saccharin and MSG, the degree of aversion decreased substantially over the two test trials ( interaction of trials by substance injected) but the rate of ex- tinction was similar for both tastes.
Rats that were infused with plain water drank approximately the same amount of MSG as saccharin. If intake by the control groups can be regarded as a measure of palatability, then MSG is no less palatable than saccharin.
GENERAL DISCUSSION
Evidence for Conditioning
Various sources of evidence indicate that the ability of car- bohydrate infusions to stimulate intake is attributable to a form of conditioning. Increased saccharin intake takes some time to appear and tends to increase with repeated testing, especially if training days are interspersed with days in which the rats are given plain water. In several experiments, the increased intake persisted for a t ime after carbohydrate infusions were discontin- ued [see also ( 5 0 ) ] . Rats trained to increase saccharin intake drank more sucrose than did rats trained to respond to maltodex-
15
4-
Tr ia l 1
@ k / 10
-(3
- - 5
U_
STIMULUS SPECIFICITY: SACCHARIN VS. CHERRY 607
Tr ia l 2
FIG. 14. Mean _+ SEM intake in Experiment 13. This is similar to Fig. 13 except that the tastes used were saccharin and MSG, there was a separate control group for each taste, and each rat was tested with its test fluid twice.
trin, even though both groups were infused with plain water dur- ing this test (Exper iment 9) . Experiment 10 showed a particu- larly dramatic extinction effect; group differences persisted for a day after the infusion conditions had been reversed.
In Pavlovian conditioning, preexposure to either the condi- tioned stimulus or the reinforcer attenuates subsequent condi- t ioning (39) . These effects also occur with carbohydrate induced stimulation of fluid intake. In the Experiment 10, preexposure to the reinforcer without the conditioned stimulus reduced condi- tioning. In Experiment 11, nonreinforced preexposure to the con- ditioned stimulus (saccharin or sodium chloride) reduced the de- gree to which carbohydrate infusions stimulated fluid intake. Taken together, all of these observations indicate that associative mechanisms contribute to the increased fluid intake.
Stimulus Specificity
Previous studies showed that dilute carbohydrate infusions greatly stimulate saccharin intake within 1 or 2 days, but had no detectable effect on cherry flavor intake (50) . The present studies indicate that the difference in the response to the two stimuli is probably not attributable to differences in the amount of carbo- hydrate infused because decreasing or increasing the amount of carbohydrate infused did not enhance the intake-stimulating ef- fects of carbohydrate (Experiments 1 and 2) . Nevertheless, the differences between stimuli are quantitative rather than qualita- tive. A modest increase in intake of cherry-flavored water can be detected by either employing a preference test or by boosting statistical power by increasing the number of animals tested.
This high degree of stimulus specificity was only observed when the rats had food available ad lib. Rats that had been trained with food available and then tested when food deprived showed significant increases in fluid intake, compared to rats that had been given the same flavor without carbohydrate infusions. This effect appeared for rats trained with cherry (Exper iment 2) , su- crose octa-acetate (Exper iment 7) , and MSG + IMP (Experi- ment 7) , even though these rats did not show much of an effect when food was freely available. The most plausible interpretation of this effect of food deprivation invokes incentive theory (6) to hypothesize that rats learn about the consequences of ingesting

608 RAMIREZ
various fluids rather than merely acquiring an ingestive response to a taste (21).
Theoretical Analysis of Stimulus Specificity
The differential effects of carbohydrate infusions on intake of different fluids was unforeseen. One might have expected that a ceiling effect would make it easier to increase ingestion of a moderately aversive flavor than to increase ingestion of a palat- able flavor, just as a basement effect would make it easier to observe a conditioned aversion with an innately preferred than with an innately aversive stimulus. This logic may be the reason that preferred concentrations of saccharin, sucrose, and sodium chloride have been the most popular conditioned stimuli in stud- ies of conditioned aversions (53). On occasion, such basement effects have made it difficult for researchers to analyze aversions to aversive stimuli (24,48).
On the other hand, it is interesting, in retrospect, that the observed degree of stimulus specificity was not predicted from arguments based on natural selection. It has long been thought that natural selection has favored animals capable of rapidly learning relationships that regularly occur in nature, such as flavor and illness (25,26) . A seemingly obvious, but never discussed, corollary of this concept is that some tastes should condit ion aversions more readily than others. Under natural conditions, sweet taste should not be a good predictor of the ability of a substance to cause illness because sweetness in- dicates the presence of sugars (45) . Bitter taste, on the other hand, is a good indicator of potential toxicity (29) . One might therefore expect that natural selection would favor animals that readily learn avoidance of bitter substances but poorly learn a,~oidance of sweet substances. The present results are generally in accord with this line of reasoning in that tastes present in nutritious foods (sweet, maltodextrin and salty) condit ioned more readily than did other tastes.
Differences Between Conditioned Acceptance and Aversion
Although the pattern of stimulus specificity for conditioned acceptance fits expectations based on evolutionary theory, no such pattern has ever been reported for malaise-conditioned aver- sions (4,9,11,31 ). The present studies suggest that salience for one type of conditioning does not predict salience for the other kind of conditioning. Cherry flavor and MSG are roughly as sa- lient as 0.05% saccharin with regard to their ability to condition an aversion (Experiments 12 and 13) but not with regard to their ability to condition fluid acceptance (Experiments 1, 2, 5, 7, and 10). There might be subtle differences in the salience of cherry flavor, MSG, and saccharin that the present experiments were not sufficiently sensitive to detect, but such subtle differences would not easily account for the large and reproducible differences be- tween these stimuli in carbohydrate conditioned stimulation of intake.
The apparent difference between the two forms of condition- ing might reflect some fundamental difference in the underlying mechanisms. Alternatively, the observed difference might be due to differences in the training procedures. In most studies, aver- sions were conditioned with one or only a few pairings of malaise and taste in discrete trials employing water-deprived rats. Ele- vated fluid acceptance, on the other hand, was conditioned by infusing nondeprived rats every time they drank (i.e., continuous training). Because every drinking bout was a training trial, the number of conditioned stimulus/unconditioned stimulus pairings was much greater than in a typical flavor aversion study. Fur-
thermore, increased intake was conditioned by infusing amounts of carbohydrate that were proportional to the amount ingested, whereas in most studies of malaise-conditioned aversions, the amount of drug given was not related to the amount of fluid ingested.
The protocols differ because potent drugs are available, ca- pable of producing intense malaise that rapidly conditions a strong behavioral response, whereas the rewarding effects of in- tragastric carbohydrates are more feeble. Because mild toxins are probably more abundant in nature than severe nonlethal toxins, it would be interesting to determine whether the degree of stim- ulus specificity for malaise-conditioned aversions varies with the degree of malaise and the type of training protocol.
CONCLUSIONS
Conditioning is much more readily observed with some tastes/flavors (saccharin, sodium chloride, oligosaccharide)than with others (cherry flavor, sucrose octa-acetate, and MSG) . Al- though such large differences between test stimuli would lead one to suspect that nonassociative mechanisms may be involved, all the evidence points to an associative mechanism (i.e., con- ditioning). At least three factors account for the differences be- tween various tastes/flavors.
Some tastes and flavors impede the development of increased fluid intake by virtue of their aversiveness (see Part I). However, it is not sufficient for a taste to be preferred; MSG and 0.4% maltodextrin are preferred over water, yet condition poorly.
Some of the differences between stimuli can be attributed to stimulus intensity. Sodium chloride provides the clearest evi- dence for this because no conditioning appeared in rats trained with 0.45% sodium chloride (when they were fed ad lib) whereas conditioning reliably appeared in rats trained with 0.9% sodium chloride (Experiments 7 and 8). This factor may also account for the poor conditioning observed with 0.01% saccharin and 0.4% maltodextrin in Experiments 5 and 7.
Other differences between stimuli are not readily attribut- able to either aversiveness or lack of salience. The absence of carbohydrate-condit ioned stimulation of intake with MSG is particularly interesting because MSG is preferred over water (Fig. 6) , is ingested as much as saccharin (Experiments 10 and 13 ), and appears to be as salient as saccharin as a stimulus for malaise-condit ioned aversion (Exper iment 13). It is pro- posed that rats are predisposed to acquire certain s t imulus - response relationships more readily than others. Thus, rats learn to associate the postingest ive effects of carbohydrates with sweet very easily, but do not learn as easily to associate the postingestive effects of carbohydrates with umami, bitter, and sour (cherry f lavor) .
Finally, some theorists have sharply divided conditioned and unconditioned responses to flavors, and put great emphasis on how conditioning can overpower unconditioned responses (8,10,57). This series of experiments indicates that such a na- ture/nurture dichotomy provides an incomplete or misleading ac- count of the factors controlling ingestion, just as inadequate as for other behaviors (2) . Carbohydrate intake can only be under- stood by analyzing how these factors interact. A better under- standing of such interactions could help us determine when con- ditioning influences behavior in real life and when it does not.
ACKNOWLEDGEMENTS
This work was supported by NIH ( 1 R01 DC01107, BRSG S07-RR- 05825-11 ) and the Monell Chemical Senses Center.

S T I M U L U S SPECIFICITY: S A C C H A R I N VS. C H E R R Y 609
REFERENCES
1. Ackroff, K.; Sclafani, A. Flavor preferences conditioned by intra- 26. gastric infusions of dilute Polycose solutions. Physiol. Behav. 55:957-962; 1994.
2. Bateson, P. P. G. Specificity and the origins of behavior. Adv. Study 27. Behav. 6:1-20; 1976.
3. Batsell, W. R. J.; Best, M. R. One bottle too many? Method of testing determines the detection of overshadowing and retention of taste aversions. Anita. Learn. Behav. 21:154-158; 1993. 28.
4. Bernstein, I. L.; Goehler, L. E.; Fenner, D. P. Learned aversions to proteins in rats on a dietary self-selection regimen. Behav. Neurosci. 98:1065-1072; 1984. 29.
5. Birch, L. L. Conditioned flavor preferences in young children. Phys- iol. Behav. 47:501-505; 1990. 30.
6. Bolles, R. C. Theory of motivation. New York: Harper & Row; 1967.
7. Booth, D. A. Food-conditioned eating preferences and aversions 31. with interoceptive elements: Conditioned appetites and satieties. Ann. NY Acad. Sci. 443:22-41; 1985. 32.
8. Booth, D. A. Learned role of tastes in eating motivation. In: Capaldi, E. D.; Powley, T. L., eds. Taste, experience, and feeding. Washing- ton: American Psychological Association; 1990:179-194. 33.
9. Brackbill, R. M.; Rosenbush, S. N.; Brookshire, K. H. Acquisition and retention of conditioned taste aversions as a function of the taste 34. quality of the CS. Learn. Motiv. 2:341-350; 1971.
10. Breslin, P. A. S.; Davidson, T. L.; Grill, H. J. Conditioned reversal 35. of reactions to normally avoided tastes. Physiol. Behav. 47:535- 538; 1990.
11. Brot, M. D.; Braget, D. J.; Bernstein, I. L. Flavor, not postingestive, 36. cues contribute to the salience of proteins as targets in aversion con- ditioning. Behav. Neurosci. 101:683-689; 1987.
12. Dess, N. K. Saccharin's aversive taste in rats: Evidence and impli- cations. Neurosci. Biobehav. Rev. 17:359-372; 1993. 37.
13. Deutsch, R. Conditioned hypoglycemia: Amechanism for saccharin- induced sensitivity to insulin in the rat. J. Comp. Physiol. Psychol. 86:350-358; 1974. 38.
14. Devenport, L. D. Aversion to a palatable saline solution in rats: Interactions of physiology and experience. J. Comp. Physiol. Psy- chol. 83:98-105; 1973. 39.
15. Domjan, M.; Gillan, D. Role of novelty in the aversion for increas- ingly concentrated saccharin solutions. Physiol. Behav. 16:537- 40. 542; 1976.
16. Dragoin, W.; McCleary, G. E.; McCleary, P. A comparison of two 41. methods of measuring conditioned taste aversions. Behav. Res. Methods Instrum. 3:309-310; 1971.
17. Drucker, D. B.; Ackroff, K.; Sclafani, A. Nutrient-conditioned flavor 42. preference and acceptance in rats: Effects of deprivation state and nonreinforcement. Physiol. Behav. 56:701-707; 1994.
18. Edgington, E. S. Randomization tests. 2nd ed. New York: Dekker; 43. 1987.
19. Elizalde, G.; Sclafani, A. Flavor preferences conditioned by intra- gastric polycose infusions: A detailed analysis using an electronic 44. esophagus preparation. Physiol. Behav. 47:63-77; 1990.
20. Emits, T.; Corbit, J. D. Taste as a diposegenic stimulus. J. Comp. Physiol. Psychol. 83:27-31; 1973.
21. Fedorchak, P. M.; Bolles, R. C. Hunger enhances the expression of 45. calorie- but not taste-mediated conditioned flavor preferences. J. Exp. Psychol. Anim. Behav. 13:73-79; 1987. 46.
22. Fedorchak, P. M.; Bolles, R. C. Nutritive expectancies mediate cho- lecystokinin's suppression-of-intake effect. Behav. Neurosci. 47. 102:451-455; 1988.
23. Franchina, J. J.; Slank, K. L. Salience and the effects of CS preex- 48. posure on aversion conditioning. Behav. Neural Biol. 50:367-373; 1988. 49.
24. Frank, M. E.; Nowlis, G. H. Learned aversions and taste qualities in hamsters. Chem. Senses 14:379-394; 1989. 50.
25. Garcia, J.; Brett, L. P.; Rusiniak, K. W. Limits of darwinian condi- tioning. In: Klein, S. B.; Mowrer, R. R., eds. Contemporary learning 51. theories: Instrumental conditioning theory and the impact of bio- logical constraints on learning. Hillsdale, NJ: Lawrence Erlbaum 52. Associates; 1989:181-203.
Garcia, J.; McGowan, B. K.; Ervin, F. R.; Koelling, R. A. Cues: Their relative effectiveness as a function of the reinforcer. Science 160:794-795; 1968. Geiselman, P. J. Carbohydrates do not always produce satiety: An explanation of the appetite- and hunger-stimulating effects of carbohydrates. Prog.Psychobiol. Physiol. Psychol. 12:1-46; 1987. Gilbert, R. M.; Sherman, I. P, Palatability-induced polydipsia: Sac- charin, sucrose, and water intake in rats, with and without food dep- rivation. Psychol. Rep. 27:319-325; 1970. Glendinning, J. I. Is the bitter rejection response always adaptive. Physiol. Behav. 56:1217-1227; 1994. Grahame, N. J.; Bamet, R. C.; Gunther, L. M.; Miller, R. R. Latent inhibition as a performance deficit resulting from CS-context asso- ciations. Anim. Learn. Behav. 22:395-408; 1994. Green, K. F.; Churchill, P. A. An effect of flavors on strength of conditioned aversions. Psychon. Sci. 21 : 19-20; 1970. Grote, F. W.; Brown, R. T. Conditioned taste aversions: Two-stim- ulus tests are more sensitive than one-stimulus tests. Behav. Res. Methods Instrum. 3:311-312; 1971. Holman, E. W. Immediate and delayed reinforcers for flavor pref- erences in rats. Learn. Motiv. 6:91-100; 1975. Holman, G. L. Intragastric reinforcement effect. J. Comp. Physiol. Psychol. 69:432-441; 1969. Kalat, J. W.; Rozin, P. "Salience": A facor which can override temporal contiguity in taste-aversion learning. J. Comp. Physiol. Psychol. 71:192-197; 1970. Kern, D. L.; McPhee, L.; Fisher, J.; Johnson, S.; Birch, L. L. The postingestive consequences of fat condition preferences for fla- vors associated with high dietary fat. Physiol. Behav. 54:71-76; 1993. Klein, S. B.; Domato, G. C.; Hallstead, C.; Stephens, I.; Mikulka, P. J. Acquisition of a conditioned aversion as a function of age and measurement technique. Physiol. Psychol. 3:379-384; 1975. Kratz, C. M.; Levitsky, D. A.; Lustick, S. Differential effects of quinine and sucrose octa acetate on food intake in the rat. Physiol. Behav. 20:665-667; 1978. Mackintosh, N. J. Conditioning and Associative Learning. New York: Oxford University Press; 1983. Manly, B. F. J. Randomization and Monte Carlo methods in biology. London: Chapman & Hall; 1991. Mehiel, R. Hedonic-shift conditioning with calories. In: Capaldi, E. D.; Powley, T. L., eds. Taste, experience, and feeding. Washing- ton: American Psychological Association; 1991:107-126. Nissenbaum, J. W.; Sclafani, A. Qualitative differences in polysac- charide and sugar tastes in the rat: A two-carbohydrate taste model. Neurosci. Biobehav. Rev. 11:187-196; 1987. Rabe, E. F.; Corbet, J. D. Postingestional control of sodium chloride solution drinking in the rat. J. Comp. Physiol. Psychol. 84:268-274; 1973. Rabin, B. M.; Hunt, W. A.; Lee, J. Attenuation and cross-attenuation in taste aversion learning in the rat: studies with ionizing radiation, lithium chloride and ethanol. Pharmacol. Biochem. Behav. 31:909- 918; 1989. Ramirez, I. Why do sugars taste good? Neurosci. Biobehav. Rev. 14:125-134; 1990. Ramirez I. Is starch flavor unitary? Evidence from studies of cooked starch. Physiol. Behav. 52:535-540; 1992. Ramirez I. Rats discriminate between starch and other substances having a similar texture. Physiol. Behav. 53:373-377; 1993. Ramirez I. Chemosensory similarities among oils: Does viscosity play a role? Chem. Senses 19:155-168; 1994. Ramirez I. Glucose polymer taste is not unitary for rats. Physiol. Behav. 55:355-360; 1994. Ramirez I. Stimulation of fluid intake by carbohydrates: Interaction of taste and calories. Am. J. Physiol. 266:R682-R687; 1994. Ramirez I. Stimulation of fluid intake by maltodextrins and starch. Physiol. Behav. 57:687-692; 1995. Richter, C. P. Maximum sodium chloride intake and thirst in domesti- cated and wild Norway rats. Am. J. Physiol. 176:213-222; 1954.

610 RAMIREZ
53. Riley, A. L.; Baril, L. L. Conditioned taste aversions: A bibliogra- phy. Anim. Learn. Behav. 4:1S-13S; 1976.
54. Saville, D. J. Multiple comparsion procedures: The practical solu- tion. Am. Stat. 44:174-180; 1990.
55. Sclafani, A. Carbohydrate taste, appetite, and obesity: An overview. Neurosci. Biobehav. Rev. 11:131-153; 1987.
56. Sclafani, A. Nutritionally based learned flavor preferences in rats. In: Capaldi, E, D.; Powley, T. L., eds. Taste, experience, and feeding. Washington, DC: American Psychological Association; 1990:139- 156.
57. Sclafani, A. Conditioned food preferences. Bull. Psychonom. Soc. 29:256-260; 1991.
58. Sclafani, A.; Nissenbaum, J. W. Taste preference thresholds for Po- lycose, maltose, and sucrose in rats. Neurosci. Biobehav. Rev. 11:181-185; 1987.
59. Sclafani, A.; Nissenbaum, J. W. Robust conditioned flavor prefer- ence produced by intragastric starch infusions in rats. Am. J. Physiol. 255:R672-R675; 1988.
60. Tordoff, M. G. Metabolic basis of learned food preferences. In: Friedman, M. I.; Tordoff, M. G.; Kare, M. R., eds. Chemical senses volume 4: Appetite and nutrition. New York: Marcel Dekker; 1991:239-260.
61. Warwick, Z. S.; Weingarten, H. P. Dissociation of palatability and calorie effects in learned flavor preferences. Physiol. Behav. 55:501-504; 1994.
62. Weingarten, H. P.; Kulikovsky, O. T. Taste-to-postingestive con- sequence conditioning: Is the rise in sham feeding with repeated experience a learning phenomenon? Physiol. Behav. 45:471-476; 1989.
63. Yamamoto, T.; Matsuo, R.; Fujimoto, Y.; Fukunaga, I.; Miyasaka, A.; Imoto, T. Electrophysiological and behavioral studies on the taste of umami substances in the rat. Physiol. Behav. 49:919-925; 1991.
64. Young, W. G.; Deutsch, J. A. The construction, surgical implanta- tion, and use of gastric catheters and a pyloric cuff. J. Neurosci. Methods 3:377-384; 1981.





























