Embed Size (px)
Citation preview

Stress Response Inhibits the Nephrotoxicity of Cisplatin
Marie H. Hanigan1, Mei Deng2, Lei Zhang1, Peyton T. Taylor Jr. 2 and Maia G. Lapus1
1Department of Cell Biology, University of Oklahoma Health Sciences Center, Oklahoma City, OK
73104 and 2Department of Obstetrics and Gynecology, Division of Gynecologic Oncology,
University of Virginia Health Sciences Center, Charlottesville, VA 22908
Running Head: Stress Response and Cisplatin
Corresponding Author: Marie H. Hanigan, Ph.D., Biomedical Research Center Room 264, 975
N.E. 10th Street, Oklahoma City, Oklahoma 73104, USA (E-mail: [email protected]).
Telephone: 405-271-3832; FAX 405-271-3813
1
Articles in PresS. Am J Physiol Renal Physiol (September 7, 2004). doi:10.1152/ajprenal.00041.2003
Copyright © 2004 by the American Physiological Society.

Abstract. Salt-loading and saline hydration are used to protect patients from cisplatin-induced
nephrotoxicity. The mechanism by which salt exerts its protective effect is unknown. As part of an
ongoing study of cisplatin nephrotoxicity, an in vitro assay system was developed that models the in
vivo exposure and response of proximal tubule cells to cisplatin. In this study, it was discovered that
the toxicity of cisplatin toward LLC-PK1 cells varied dramatically according to the tissue culture
media used for the 3 hr cisplatin exposure. Further experiments revealed that minor variations in the
sodium concentration among standard tissue culture media modulated cisplatin nephrotoxicity.
Sodium chloride has been shown to protect against cisplatin-induced nephrotoxicity in vivo, but has
never before been demonstrated in vitro. NaCl did not alter the cellular accumulation of cisplatin.
NaCl altered the osmolarity of the external media and its effect was replicated by substituting
equiosmolar concentrations of impermeant anions or cations. The change in osmolarity triggered
a stress response within the cell that modulated sensitivity to cisplatin. These data resolve several
long-standing controversies regarding the mechanism by which salt-loading protects the kidney
from cisplatin-induced nephrotoxicity.
Key Words: kidney, cis-diamminedichloroplatinum, toxicity, proximal tubule, LLC-PK1 cells
2

INTRODUCTION
The discovery of the potent antitumor activity of cisplatin provided an effective drug for the
treatment of germ cell tumors and ovarian cancer (11). It is also used as a radiosensitizer for
cervical cancer and as a component of consolidation therapy for many types of solid tumors. The
efficacy of cisplatin is limited, however, by the cumulative nature of its dose-limiting
nephrotoxicity. Cisplatin is toxic to the renal proximal tubules (8). The drug is excreted in the
urine in a diphasic manner; most of the drug is excreted within the first three hours (6). Renal
toxicity, as determined by elevated levels of blood urea nitrogen, is not evident until 72 to 96 hours
after the initial administration (8). Studies in rodents revealed that administering cisplatin in
hypertonic salt reduced its nephrotoxicity (21). Hydration with saline was shown to be protective in
dogs (24). These data have been applied in the clinic. The drug is re-constituted in 0.9% sodium
chloride. Vigorous hydration with saline and simultaneous administration of mannitol before,
during, and after cisplatin administration significantly reduced the cisplatin-induced nephrotoxicity
and is now the accepted standard of care (4).
Despite the wide-spread use of salt, the mechanism by which salt protects the kidney has never
been identified. Several theories have been proposed to explain the protective effect of hydration
against cisplatin nephrotoxicity. Some investigators have suggested that hypertonic saline and
saline hydration protect the kidney by increasing the rate of cisplatin excretion (9). Others have
proposed that the salt provides a high concentration of chloride ions which prevents the dissociation
of the chloride ions from the platinum molecule, thereby reducing the formation of the reactive,
aquated species of cisplatin (5; 10). There has been debate as to whether the sodium or the chloride
ion is responsible for the protective effect. Litterst's mouse data indicated that the sodium ion is the
3

important molecule, while Earhart and coworkers reported data from rat studies suggesting that the
chloride is the protective ion (10; 21).
As part of ongoing studies on the mechanism of cisplatin nephrotoxicity, we developed an in vitro
model system of cisplatin-induced toxicity toward proximal tubule cells(33). Confluent monolayers
of cells are treated for 3 hours with cisplatin. The drug is removed and cell viability is assessed at 3
days. This treatment mimics the in vivo exposure. In this system, cisplatin toxicity is dose- and
time-dependent. During the course of our studies we found that treating the cells with cisplatin in
RPMI-1640 medium was more toxic than treating the cells with cisplatin in Dulbecco's Modified
Eagle Medium (DMEM). Both RPMI-1640 medium and DMEM are commonly used to maintain
mammalian cells in culture. Their formulations are very similar. In this study we analyzed the
effect of four commonly used tissue culture media on the toxicity of cisplatin. We tested individual
components and found that the NaCl concentration in the media modulated cisplatin toxicity. The
robust effect of NaCl on cisplatin toxicity in this in vitro system is similar to that observed in vivo.
This model system enabled us to test directly several of the hypotheses regarding the mechanism by
which NaCl protects the kidney from cisplatin toxicity.
MATERIALS AND METHODS
Cell Culture
LLC-PK1 (ATCC CRL 1392), a pig kidney cell line, was purchased from the American Type
Culture Collection (Manassas,VA). LLC-PK1 cells were grown in DMEM supplemented with 4
mM L-glutamine (DMEM, GIBCO/BRL, Grand Island, NY), 5% fetal bovine serum (FBS, Hyclone
4

Laboratories, Logan, UT), penicillin (5 U/ml), and streptomycin (5 µg/ml, GIBCO/BRL). Cells
were maintained in 5% CO2 and 95% air at 37oC. Subconfluent cultures were passaged every 3 to 4
days. For toxicity experiments, LLC-PK1 cells were seeded in 96 well plates at 5x103 cells/well.
The cells formed confluent monolayers on the 3rd day after plating. The medium was replaced with
fresh medium on the 4th day and the cells were used for experiments on the 7th day after plating.
Cisplatin Toxicity
Cisplatin (Sigma Chemical Co, St. Louis, MO) was prepared as a 3.3 mM stock solution in 0.9%
saline daily. RPMI-1640 Medium (RPMI, Cat. No. 31800); DMEM (Cat. No. 12800); Leibovitz's
L-15 Medium (L-15, Cat. No. 41300); Nutrient Mixture Ham's F-12 (F-12, Cat. No. 21700); and
Hank's Balanced Salt Solution (HBSS, Cat. No.11201) were prepared from powder (GIBCO/BRL).
L-15 was supplemented with 18 mM HEPES and 0.2% BSA. The pH of all solutions and media
was adjusted to 7.4. For toxicity experiments, the media on the cells was changed to the test
solution and cisplatin was added to the wells. Cells treated with only the test solution served as
controls. The cells were incubated at 37oC in 5% CO2 , 95% air, except cells in L-15 medium which
were incubated at 37oC without CO2. After 3 hrs, the cisplatin-containing solution was removed
from the cells and replaced with DMEM containing 5% FBS and antibiotics. The cells were
incubated for an additional 69 hrs at 37oC in 5% CO2. The number of viable cells was then
determined with the MTT assay (23). A standard curve showed a direct correlation between the
number of viable LLC-PK1 cells and the OD570 in the MTT assay.
Effect of NaCl Concentration and Osmolality on Cisplatin Toxicity
Solutions containing the inorganic salt components of L-15, F-12, RPMI, HBSS, and DMEM were
prepared according to their formulations in the GIBCO/BRL catalogue, but without sodium
5

bicarbonate and the pH adjusted to pH 7.4. Saline was prepared with 2mM sodium phosphate
buffer , pH 7.4. Cisplatin was diluted in each test solution. The cells were exposed to cisplatin for 3
hrs at 37oC in an air atmosphere. Cells treated with the test solution that did not contain cisplatin
served as controls. After 3 hrs, the cisplatin-containing solutions were removed from the cells and
replaced with DMEM containing 5% FBS and antibiotics. Cell viability was assayed 69 hrs after
the cisplatin-solution was removed.
Solutions of NaCl were prepared with 5 mM HEPES, pH 7.4. Cells were exposed to 100 µM
cisplatin in the NaCl solutions for 3 hrs at 37oC without CO2. Cells incubated in the NaCl solutions
without cisplatin served as controls for each concentration of NaCl. Cisplatin was prepared in 0.9%
NaCl. The solution for each set of triplicate wells was made separately and constituted such that the
NaCl concentration in the cisplatin-containing solution was the same as its corresponding control.
Solutions containing NaCl and mannitol or NaCl and N-methyl-D-glucamine were prepared with 5
mM HEPES, pH 7.4. Equiosmolar solutions were prepared by decreasing the concentrations
mannitol or N-methyl-D-glucamine as the concentration of NaCl increased to maintain the
osmolality of the solution at 288 mosmol/kg H2O. The osmolality of the tissue culture media and
salt solutions was determined with a Wescor 5520 VAPRO Vapor Pressure Osmometer (Logan,
UT).
Cisplatin Uptake
LLC-PK1 cells were seeded in 24-well plates at 5 x104 cells/well. On the 5th day, 100 µM cisplatin
in 125 to 150 mM NaCl/5 mM HEPES, pH 7.4, was added to the confluent monolayers and
incubated at 37°C without CO2 for 3 hrs. The cells were then washed twice with phosphate-buffered
saline (PBS). The cells in each well were digested in 150 µL concentrated HNO3 at 70°C for 4 hr.
6

The concentration of HNO3 was adjusted to 3.25 M with distilled water, and Triton X-100 was
added to a final concentration of 1%. Platinum levels were quantified by flameless atomic
absorption spectrometry (FAAS). A Varian SPECTRAA-220Z Graphite Furnace Double Beam
Atomic Absorption Spectrophotometer (Houston, TX) with Zeeman background correction was
used. The platinum standard included equivalent amounts of HNO3 and Triton X-100. The number
of cells in triplicate wells was determined as previously described (16).
Pretreatment with Salt
DMEM was prepared according to the formulation in the GIBCO/BRL catalogue with the exception
of NaCl, which was omitted from the stock solution so that the concentration could be varied within
the experiment. Three hrs prior to cisplatin treatment, the media was changed to fresh DMEM
containing 5% FBS, antibiotics and various concentrations of NaCl. After 3 hrs the media was
removed and the cells treated with 100 µM cisplatin in DMEM. At the end of the 3 hr cisplatin
exposure the cisplatin was removed and cells returned to DMEM containing 5% FBS and
antibiotics for 69 hrs. The number of viable cells was determined with the MTT assay.
DNA Platination
LLC-PK1 cells were seeded in P-60 plates at 5.7x105 cells/plate. On the 5th day, the confluent cells
were exposed to 100 µM cisplatin in 125 to150 mM NaCl with 5 mM HEPES, pH 7.4, at 37°C
without CO2 for 3 hrs. The cells were washed with PBS and trypsinized. DNA was purified by the
method of McKeage and coworkers (22). Briefly, cells were lysed (10 mM Tris, 10 mM EDTA,
0.15 M NaCl, and 0.5% SDS, pH 8.0) in the presence of 1 mg/ml proteinase K. The lysis mixture
was incubated at 60°C for 10 min and then 37°C overnight. DNA was purified by phenol extraction
and ethanol precipitation. The DNA pellet was resuspended in 10 mM Tris, 1 mM EDTA, pH 8.0,
7

with addition of 100 µg/ml RNase and incubation at 37°C for 1 hr. DNA was then reprecipitated
and hydrolyzed in 0.2% HNO3. DNA concentration was estimated by measuring the absorbance at
260 nm, and the platinum content was measured by FAAS as described above.
Data analysis
Each experiment was done a minimum of three times. Each data point within the experiment
represents the mean value of three to six replicate wells. To detect statistically significant effects of
medium or inorganic salt solutions on cisplatin nephrotoxicity, the toxicity of each concentration of
cisplatin in each of the solutions was compared to the toxicity of cisplatin in L-15 by a t-test.
Significant correlations between the salt concentration in the solution and cisplatin toxicity,
platinum accumulation or platinum-DNA binding were detected by One-Way ANOVA and post test
for a linear trend. Statistically significant difference between curves were detected by an F-test. All
statistical analyses were done using GraphPad Prism version 3 (GraphPad Software Inc. San Diego,
CA).
RESULTS
Effect of Medium on Cisplatin Toxicity
Tissue culture media modulated the toxicity of cisplatin toward proximal tubule cells. LLC-PK1
cells were grown to confluence and maintained in DMEM with 5% FBS and antibiotics. The cells
were treated with cisplatin for 3 hrs, then returned to complete DMEM. Toxicity was assessed at 72
hrs. The media on the cells in the four experimental groups varied only during the 3 hr cisplatin
exposure. Prior to and following cisplatin treatment, all cells were in complete DMEM. The data in
8

Fig. 1A show that cisplatin in RPMI or F-12 media was significantly more toxic to LLC-PK1 cells
than equimolar cisplatin in DMEM or L-15 media. A dose-dependent toxicity of cisplatin was
observed with RPMI and F-12 media containing cisplatin. At 100 µM and 150 µM, cisplatin was
significantly less toxic in DMEM or L-15 media (p< 0.001).
To determine which components of the media modulated cisplatin toxicity, components of the
media were tested with the same protocol used for the media. The cells were exposed to cisplatin in
the test solutions for 3 hr and toxicity was measured at 72 hrs. Data from experiments in which the
inorganic salt components of the media were tested showed that the salt solutions modulated
cisplatin toxicity (Fig 1B). The composition of the solutions tested is shown in Table I. There was a
direct correlation between the concentration of NaCl in the solution and the reduction in cisplatin
toxicity: saline>L-15>HBSS>F-12>DMEM>RPMI . The higher the concentration of NaCl the less
toxic cisplatin was to the cells. The concentrations of the other components did not correlate with
cisplatin toxicity. It should be noted that DMEM reduced the toxicity of cisplatin (Fig. 1A).
However, when the inorganic salt solution of DMEM was tested it potentiated cisplatin toxicity
(Fig. 1B). DMEM contains 44 mM NaHCO3, which is higher than the amount in F-12 or RPMI (14
mM and 24 mM respectively). NaHCO3 increased the Na+ ion concentration in DMEM media by
39 mM in comparison to the Na+ ion concentration DMEM inorganic salt solution tested. The
inorganic salt fraction of L-15 is also somewhat less protective than L-15 medium. There may be
additional components in L-15 medium that modulate cisplatin toxicity.
To directly analyze the effect of NaCl on the toxicity of cisplatin, cells were incubated for 3 hrs in a
100 µM solution of cisplatin containing 130 mM to 155 mM NaCl and 5 mM HEPES, pH 7.4. As
shown in Fig.2, increasing the concentration of NaCl from 130 mM to 155 mM increased cell
9

survival in 100 µM cisplatin from 22+3% to 98+3 % (p<0.001). In the absence of cisplatin, the 3 hr
incubation in the NaCl solutions had no effect on cell viability (p=0.6).
Effect of Sodium Chloride Concentration on Cisplatin Accumulation
Sodium chloride did not alter the accumulation of cisplatin in LLC-PK1 cells. Platinum levels were
measured in confluent monolayers treated with cisplatin solutions containing NaCl. Increasing the
NaCl concentration from 125 mM to 155 mM had no significant effect on platinum accumulation
(Fig.3), despite the dramatic effect on toxicity (Fig. 2).
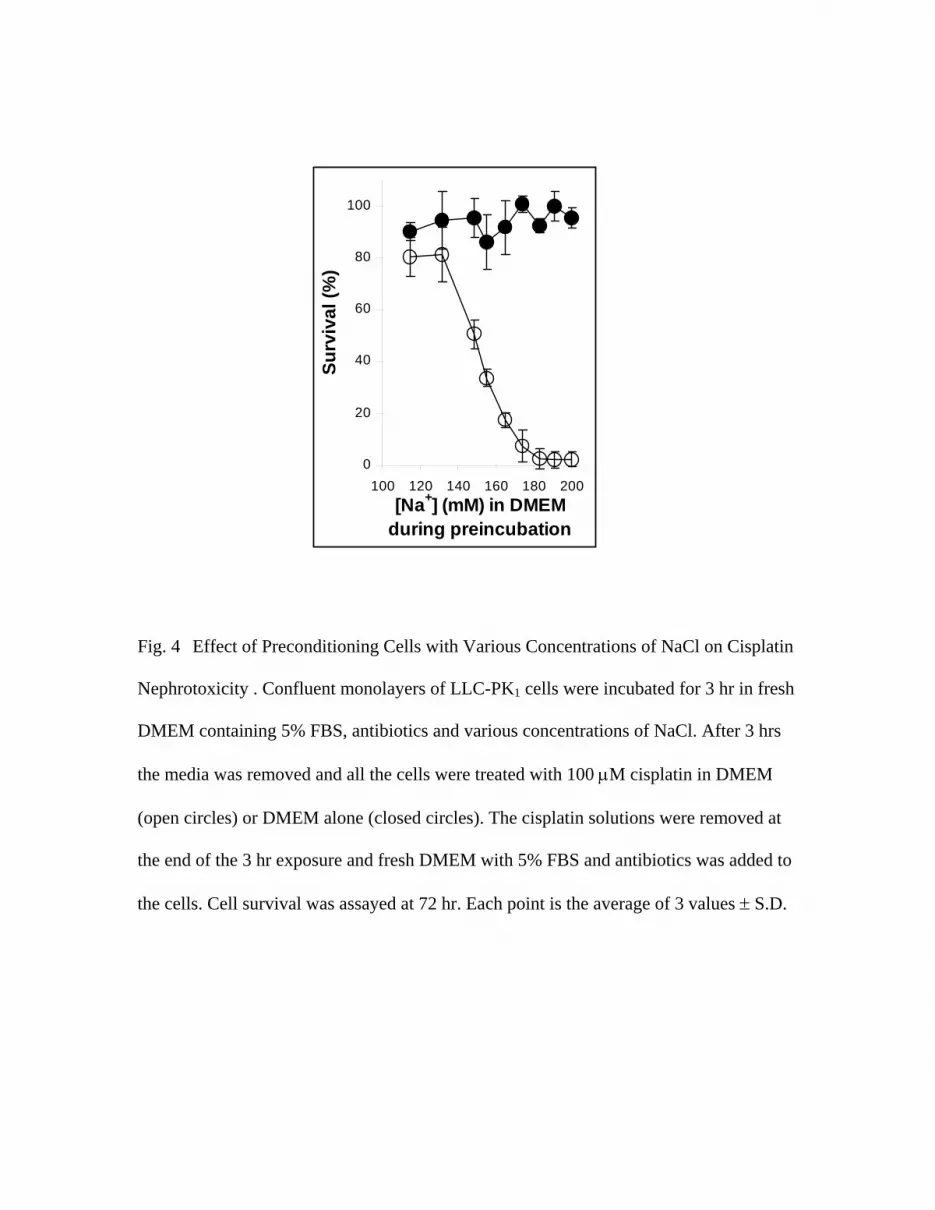
Preconditioning Cells with Various Concentrations of NaCl
A series of experiments were undertaken to determine whether the effect of NaCl on cisplatin
toxicity was due to interaction of the NaCl with the cisplatin or due to an effect of the salt on the
cells. The cells were incubated in media with various concentrations of NaCl for 3 hrs before the
cisplatin treatment. The preincubation media consisted of DMEM formulated with various
concentrations of NaCl plus 5% FBS. At the end of the preincubation period, the media was
removed and all the cells were treated with 100 µM cisplatin in DMEM containing the standard
amount of NaCl. After the cisplatin treatment, the cisplatin-containing solution was removed and all
cells were incubated for an additional 69 hrs in DMEM with 5% FBS and antibiotics. The data in
Fig. 4 show that the concentration of NaCl in the preincubation medium had a dramatic effect on
the toxicity of cisplatin. Pretreatment of the cells with higher concentrations of Na+ ions in DMEM
was correlated with greater cisplatin toxicity. Cell survival decreased from 80.4 + 7.6% to 2.4 +
2.9%. The linear trend was highly significant (p<0.0001). All of the cells were treated with the
same cisplatin solution. These data show that the NaCl concentration is modulating cisplatin
toxicity through its effect on the cell rather than through a direct interaction with the cisplatin.
10

Controls showed that the pretreatment with the media alone (no cisplatin) had no significant effect
on cell viability (p=0.18).
Higher concentrations of NaCl during the pretreatment period resulted in increased sensitivity to
cisplatin toxicity (Fig. 4), while higher NaCl concentrations during the cisplatin exposure were
protective (Fig. 2). These data suggest a correlation between osmotic induction of changes in cell
volume and sensitivity to cisplatin. In the experiment shown in Fig. 2, all of the cells were
incubated in DMEM, which contains 156 mM sodium, prior to the cisplatin exposure. The medium
was then changed to solutions with 130 mM to 155 mM NaCl during the cisplatin exposure. The
decrease in extracellular salt concentration may have caused cell swelling. The data show that there
was a dose-response relationship between the increase in cisplatin susceptibility and the decrease in
extracellular salt. Conversely, in the experiment shown in Fig. 4, the extracellular NaCl was varied
for 3 hrs prior to the cisplatin exposure. All of the cells were then treated with 100 µM cisplatin in a
solution containing 156 mM sodium. Cells that were pretreated with media containing less than 156
mM sodium experienced an increase in extracellular salt when switched to the cisplatin-solution.
These cells may have responded by cell shrinkage which protected them from cisplatin toxicity.
The cells that had been pretreated with media containing more than 156 mM sodium experienced a
decrease in extracellular salt when switched to the cisplatin-solutions. These cells may have
responded by swelling which made them more susceptible to cisplatin toxicity. The data in Fig. 3
demonstrate that the changes in extracellular salt concentration alters the sensitivity of the cell to
cisplatin without altering the accumulation of platinum in the cell. The changes in cell volume may
have modulated stress responses within the cell that altered sensitivity to cisplatin.
Effect of Osmolality on Cisplatin Toxicity
11

An analysis of the osmolality of the four tissue culture media tested in Fig. 1A revealed a
correlation between the osmolality of the media and the toxicity of cisplatin (Table II). These data
suggested that the NaCl concentration in the extracellular solution may be modulating cisplatin
toxicity through an osmotic effect. To compare the effect of NaCl vs osmolality on cisplatin
toxicity, one set of cells was treated for 3 hrs in a 100 µM solution of cisplatin containing 115 mM
to 155 mM NaCl and 5 mM HEPES, pH 7.4. Two additional sets of cells were incubated in the
same solutions balanced with mannitol or N-methyl-D-glucamine such that the osmolality of each
solution was 288 mosmol/kg H2O. As shown in Fig. 5, maintaining the osmolality of the solution
with either mannitol or N-methyl-D-glucamine protected the cells from cisplatin toxicity indicating
that the osmotic effect of NaCl modulated cisplatin toxicity. In the absence of cisplatin, the 3 hr
incubation in the salt solutions had no effect on cell viability (data not shown).
Four compounds were tested for their ability to modulate cisplatin toxicity by modulating the
osmolality. Cells were incubated for 3hr in solutions containing 100 µM solution of cisplatin, 115
mM NaCl, 5 mM HEPES, pH 7.4 and increasing concentrations of mannitol (a sugar), sucrose (a
sugar), N-methyl-D-glucamine (an impermeant cation) or gluconate (an impermeant anion). At
equivalent osmolality, all four compounds reduced cisplatin toxicity to a similar level (Fig 6). In
the absence of cisplatin, the 3 hr incubation in the solutions had no effect on cell viability (data not
shown). The osmolality range tested in these experiments (214 to 288 mosm/kg H2O) is equivalent
to the osmolality range of the NaCl solutions tested in Fig 5 (open diamonds, 115 to 155 mM).
These data show that an osmotic stress response modulates the sensitivity of renal cells to cisplatin
toxicity.
Effect of Osmolality on Cisplatin Accumulation
12

Platinum levels were measured in confluent monolayers of cisplatin-treated cells incubated in
115mM NaCl with mannitol. Increasing the mannitol concentration from 19 mM to 75 mM
increased the osmolality of the solution from 232 to 288 mosm/kg H2O. Over this concentration
range there appears to be a decrease in platinum accumulation per cell (Fig 7A). However, when
the data are graphed with a volume correction to account for the osmolarity-induced change in cell
volume, the data show that the platinum accumulation does not change over this osmolality range
(Fig 7B).
Effect of NaCl on Platinum bound to DNA
Increasing the NaCl concentration resulted in a decrease in the amount of platinum bound to DNA.
Increasing the NaCl concentration from 125 mM to 150 mM decreased the level of DNA platination
from 153 ± 11 to 98 ± 15 ng/mg DNA, a 36% decrease (Fig. 8, p<0.0001). The decrease in DNA
platination is less than the 65% decrease in cisplatin toxicity over this same range of extracellular
NaCl. However, altered DNA platination may be part of the mechanism by which changes in
extracellular NaCl exerts its protective effect.
DISCUSSION
Small variations in the osmolality among tissue culture media account for their modulation of
cisplatin nephrotoxicity. Sodium chloride and mannitol have been shown to protect against
cisplatin-induced nephrotoxicity in vivo, but not in vitro (5). Our data have shown that the NaCl
does not alter the accumulation of cisplatin. The salt did not exert its effect by interacting directly
13

with cisplatin, but rather by altering the osmolality of the extracellular solution, thereby modulating
a stress response within the cell.
It has been proposed that volume repletion of patients with isotonic saline protects against cisplatin-
induced nephrotoxicity by flushing the cisplatin out of the body, limiting uptake into the proximal
tubule cells (9). Patients treated with cisplatin are generally given up to 6 L of saline intravenously
per day (28; 34). The diuretic mannitol is also commonly used to decrease the nephrotoxicity of
cisplatin. In our assay system NaCl and mannitol protected the cells from cisplatin toxicity. The
amount of cisplatin in the medium was constant during the 3h treatment; therefore, the protective
effect of these compounds was not due to decreased exposure of the cells to the drug.
The in vitro studies presented here demonstrated a large effect of NaCl on cisplatin toxicity over a
narrow concentration range (130 to 155 mM). The normal reference range for sodium in serum is
135-145mM. Sodium concentrations below 120 mM or above 160 mM are considered pathologic
(13). Physiologic saline (0.85% NaCl) is 145 mM NaCl. The sodium concentration in the tissue
culture media we tested ranged from 138 mM to 156 mM. In calculating the sodium and the
chloride concentration in each medium, the total amount of each ion included the ions contributed
by the sodium salts of amino acids and the sodium hydroxide or hydrochloric acid solutions used to
adjust the pH. Studies analyzing the protective effect of a variety of compounds on cisplatin
toxicity often do not take into account the amount of sodium added when the sodium salt of the
compound is used. Sodium selenite protected rats against cisplatin-induced nephrotoxicity without
changing the pharmacokinetics of the cisplatin (35). The investigators demonstrated that selenite
does not react with the cisplatin and speculated that the selenite requires bioactivation to exert its
protective effect. It is possible that the sodium rather than the selenite is responsible for the
14

protection. This scenario has been demonstrated with aminoglycosides which are also toxic to the
proximal tubules. Ticarcillin has been shown to protect against gentamicin nephrotoxicity. Sabra
and Branch demonstrated that the salt load associated with ticarcillin administration was the
primary mechanism by which ticarcillin protected against aminoglycoside nephrotoxicity (31).
Clinical protocols have administered cisplatin in solutions containing as much as 3% NaCl (2; 28).
Some investigators have proposed that the salt provides a high concentration of chloride ions which
prevents the dissociation of the chloride ions from the platinum molecule, thereby reducing the
formation of the reactive, aquated species of cisplatin (5). In vivo NaCl must be injected shortly
before or in the same solution as the cisplatin to exert its nephroprotective effect (5). Our studies in
which the NaClconcentration was varied prior to cisplatin treatment showed that the extracellular
NaCl is affecting the cell and not interacting directly with the cisplatin.
Both NaCl and mannitol protected against cisplatin-induced toxicity. At equiosmolar
concentrations they were equally protective. Studies of platinum accumulation showed that
increasing the extracellular concentration of NaCl did not change the total amount of platinum in
cells at 3h. In contrast, increasing the extracellular concentration of mannitol decreased cellular
platinum accumulation. These data indicate that total platinum accumulation is not the mechanism
by which osmolality alters cisplatin toxicity, as these two compounds differed in their effect on
accumulation while having the same effect on toxicity. There have been conflicting data regarding
the mechanism by which cisplatin is taken up into cells (1). Recently, two groups have shown that
cisplatin can be transported into the cell by the copper transporter Ctr1, an energy and sodium-
independent transporter (17; 19; 20). Permeant ions such as sodium and chloride and impermeant
ions such as mannitol have different effects on membrane permeability and transport systems (12).
15

We propose that osmotic stress responses are induced in proximal tubule cells by increased or
decreased osmolality of the extracellular solution. These responses may be triggered by cell
swelling or cell shrinkage as the osmolality of the extracellular solution changes. The volume
changes are transient. Epithelial cells can re-adjust their volume by regulatory volume decrease or
regulatory volume increase, active processes involving ion channels and other cotransporter
mediated mechanisms (3; 25; 37). These transient changes in cell volume may affect cisplatin
nephrotoxicity by any of several mechanisms. Changes in cell volume have been shown to modulate
cell survival and apoptotic pathways (32). Cisplatin kills LLC-PK1 cells by a caspase-3 dependent
apoptosis (18; 26; 38). Resistance to cisplatin has been associated with induction of proteins
associated with other stress responses. Wachsberger and coworkers reported that mammalian cells
grown at pH 6.7 had elevated levels of HSP27 and were resistant to cisplatin (36). Changes in the
osmolality of the extracellular medium have also been shown to alter chromatin structure and free-
radical induced DNA-protein crosslinks (27; 30). Alteration of chromatin structure may alter the
accessibility of platinum to DNA. The data in this study showed increased platinum-DNA binding
at low NaCl concentrations, the conditions under which the cells were the most sensitive to cisplatin
toxicity. Alternatively, an osmotic response may be modulating components of the pathway
through which cisplatin is metabolized to a nephrotoxin. We have shown that the nephrotoxicity of
cisplatin is due to the conjugation of cisplatin to glutathione and the subsequent metabolism of that
conjugate to a cysteinyl-glycine-conjugate, then to a cysteine-conjugate, and finally to a reactive
thiol which is toxic to the cell (14; 15; 33). This pathway is analogous to the metabolic activation of
nephrotoxic alkenes (7). In this pathway the glutathione-conjugates are formed intracellularly then
excreted. The metabolism of the glutathione-conjugate and the cysteinyl-glycine conjugate occur
extracellularly, catalyzed by the cell surface enzymes GGT and aminodipeptidase. The cysteine-
16

conjugates are metabolized to reactive thiols intracellularly by cysteine-S-conjugate beta-lyase. The
osmotic stress response could be affecting any of several steps in this pathway.
In the clinic, hydration by intravenous infusion of saline is the method most commonly used to
prevent cisplatin-induced nephrotoxicity (29). Our ongoing studies into the molecular mechanism
of this stress response will elucidate this pathway and provide novel strategies to block the
nephrotoxic effects of cisplatin.
ACKNOWLEDGEMENT
We gratefully acknowledge Dr. Siribhinya Benyajati in the Department of Physiology at The
University of Oklahoma Health Sciences Center for her helpful discussions during the preparation
of this manuscript. This work was supported by RO1 CA57530 (M.H.H.) from The National
Cancer Institute, NIH and a grant from the Presbyterian Health Foundation, Oklahoma City, OK.
17

REFERENCES
Reference List
1. Andrews PA. Cisplatin accumulation. In: Platinum-Based Drugs in Cancer Therapy, edited
by Kelland LR and Farrell NP. Totowa, New Jersey: Humana Press, 2000, p. 89-113.
2. Bajorin D, Bosl GJ and Fein R. Phase-I Trial of Escalating Doses of Cisplatin in
Hypertonic Saline. J Clin Oncol 5: 1589-1593, 1987.
3. Choi JY, Shah M, Lee MG, Schultheis PJ, Shull GE, Muallem S and Baum M. Novel
amiloride-sensitive sodium-dependent proton secretion in the mouse proximal convoluted
tubule. J Clin Invest 105: 1141-1146, 2000.
4. Cornelison TL and Reed E. Nephrotoxicity and hydration management for cisplatin,
carboplatin, and ormaplatin. Gynecol Oncol 50: 147-158, 1993.
5. Daley-Yates PT and McBrien DCH. A study of the protective effect of chloride salts on
cisplatin nephrotoxicity. Biochem Pharmacol 34: 2363-2369, 1985.
6. DeConti RC, Toftness BR, Lange RC and Creasey WA. Clinical and pharmacological
studies with cis-diamminedichloroplatinum(II). Cancer Res 33: 1310-1315, 1973.
7. Dekant W, Vamvakas S and Anders MW. Formation and fate of nephrotoxic and
cytotoxic glutathione S-conjugates: Cysteine conjugate beta-lyase pathway. In: Advances in
18

Pharmacology, Vol.27, edited by Anders MW and Dekant W. New York: Academic Press
Inc., 1995, p. 115-162.
8. Dobyan DC, Levi J, Jacobs C, Kosek J and Weiner MW. Mechanism of cis-platinum
nephrotoxicity: II. morphological observations. J Pharmacol Exp Ther 213: 551-556, 1980.
9. Dumas M, de Gislain C, d'Athis P, Chadoint-Noudeau V, Escousse A, Guerrin J and
Autissier N. Influence of hydration on ultrafilterable platinum kinetic and kidney function
on patients treated with cis-diamminedichloroplatinum (II). Cancer Chemother Pharmacol
26: 278-282, 1990.
10. Earhart RH, Martin PA, Tutsch KD, Erturk E, Wheeler RH and Bull FE. Improvement
in the therapeutic index of cisplatin (NSC 119875) by pharmacologically induced
chloruresis in the rat. Cancer Res 43: 1187-1194, 1983.
11. Einhorn LH. Curing metastatic testicular cancer. Proc Natl Acad Sci U S A 99: 4592-4595,
2002.
12. Filipovic D and Sackin H. Stretch- and volume-activated channels in isolated proximal
tubule cells. Am J Physiol 262: F857-F870, 1992.
13. Finley PR, Grady HJ, Olsowka ES and Tilzer LL. Chemistry. In: Laboratory Test
Handbook, edited by Jacobs DS, DeMott WR, Finley PR, Horvat RT, Kasten BL and Tilzer
LL. Hudson,OH: Lexi-Comp, 1993, p. 94-350.
19

14. Hanigan MH, Gallagher BC, Taylor PTJr and Large MK. Inhibition of gamma-glutamyl
transpeptidase activity by acivicin in vivo protects the kidney from cisplatin-induced
toxicity. Cancer Res 54: 5925-5929, 1994.
15. Hanigan MH, Lykissa ED, Townsend DM, Ou CN, Barrios R and Lieberman MW.
Gamma-glutamyl transpeptidase-deficient mice are resistant to the nephrotoxic effects of
cisplatin. Am J Pathol 159: 1889-1894, 2001.
16. Hanigan MH and Pitot HC. Isolation of gamma-glutamyl transpeptidase positive
hepatocytes during the early stages of hepatocarcinogenesis in the rat. Carcinogenesis 3:
1349-1354, 1982.
17. Ishida S, Lee J, Thiele DJ and Herskowitz I. Uptake of the anticancer drug cisplatin
mediated by the copper transporter Ctr1 in yeast and mammals. Proc Natl Acad Sci U S A
99: 14298-14302, 2002.
18. Kruidering M, van de WB, Zhan Y, Baelde JJ, Heer E, Mulder GJ, Stevens JL and
Nagelkerke JF. Cisplatin effects on F-actin and matrix proteins precede renal tubular cell
detachment and apoptosis in vitro. Cell Death Differ 5: 601-614, 1998.
19. Lee J, Pena MM, Nose Y and Thiele DJ. Biochemical characterization of the human
copper transporter Ctr1. J Biol Chem 277: 4380-4387, 2002.
20. Lin X, Okuda T, Holzer A and Howell SB. The copper transporter CTR1 regulates
cisplatin uptake in Saccharomyces cerevisiae. Mol Pharmacol 62: 1154-1159, 2002.
20

21. Litterst CL. Alterations in the toxicity of cis-dichlorodiammineplatinum-II and in tissue
localization of platinum as a function of NaCl concentration in the vehicle of administration.
Toxicol Appl Pharmacol 61: 99-108, 1981.
22. McKeage MJ, Abel G, Kelland LR and Harrap KR. Mechanism of action of an orally
administered platinum complex [ammine bis butyrato cyclohexylamine dichloroplatinum
(IV) (JM221)] in intrinsically cisplatin-resistant human ovarian carcinoma in vitro. Br J
Cancer 69: 1-7, 1994.
23. Mosmann T. Rapid colorimetric assay for cellular growth and survival:Application to
proliferation and cytotoxicity assays. J Immunol Methods 65: 55-63, 1983.
24. Ogilvie GK, Krawiec DR, Gelberg HB, Twardock AR, Reschke RW and Richardson
BC. Evaluation of a short-term saline diuresis protocol for the administration of cisplatin.
Am J Vet Res 49: 1076-1078, 1988.
25. Okada Y, Maeno E, Shimizu T, Dezaki K, Wang J and Morishima S. Receptor-mediated
control of regulatory volume decrease (RVD) and apoptotic volume decrease (AVD). J
Physiol 532: 3-16, 2001.
26. Okuda M, Masaki K, Fukatsu S, Hashimoto Y and Inui K. Role of apoptosis in cisplatin-
induced toxicity in the renal epithelial cell line LLC-PK1. Implication of the functions of
apical membranes. Biochem Pharmacol 59: 195-201, 2000.
21

27. Oleinick NL, Chiu SM, Ramakrishnan N and Xue LY. The formation, identification, and
significance of DNA-protein cross-links in mammalian cells. Br J Cancer Suppl 8: 135-140,
1987.
28. Ozols RF, Corden BJ, Jacob J, Wesley MN, Ostchega Y and Young RC. High-dose
cisplatin in hypertonic saline. Ann Intern Med 100: 19-24, 1984.
29. Pinzani V, Bressolle F, Haug IJ, Galtier M and Blayac JP. Cisplatin induced renal
toxicity and toxicity-modulating strategies: a review. Cancer Chemother Pharmacol 35: 1-9,
1994.
30. Proft M and Struhl K. MAP kinase-mediated stress relief that precedes and regulates the
timing of transcriptional induction. Cell 118: 351-361, 2004.
31. Sabra R and Branch RA. Role of sodium in protection by extended-spectrum penicillins
against tobramycin-induced nephrotoxicity. Antimicrob Agents Chemother 34: 1020-1025,
1990.
32. Terada Y, Inoshita S, Hanada S, Shimamura H, Kuwahara M, Ogawa W, Kasuga M,
Sasaki S and Marumo F. Hyperosmolality activates Akt and regulates apoptosis in renal
tubular cells. Kidney Int 60: 553-567, 2001.
33. Townsend DM, Deng M, Zhang L, Lapus MG and Hanigan MH. Metabolism of
Cisplatin to a nephrotoxin in proximal tubule cells. J Am Soc Nephrol 14: 1-10, 2003.
22

34. Townsend DM and Hanigan MH. Inhibition of gamma-glutamyl transpeptidase or
cysteine S-conjugate beta-lyase activity blocks the nephrotoxicity of cisplatin in mice. J
Pharmacol Exp Ther 300: 142-148, 2002.
35. Vermeulen NP, Baldew GS, Los G, McVie JG and De Goeij JJ. Reduction of cisplatin
nephrotoxicity by sodium selenite. Lack of interaction at the pharmacokinetic level of both
compounds. Drug Metab Dispos 21: 30-36, 1993.
36. Wachsberger PR, Landry J, Storck C, Davis K, O'Hara MD, Owen CS, Leeper DB
and Coss RA. Mammalian cells adapted to growth at pH 6.7 have elevated HSP27 levels
and are resistant to cisplatin. Int J Hyperthermia 13: 251-255, 1997.
37. Wu KL, Khan S, Lakhe-Reddy S, Wang L, Jarad G, Miller RT, Konieczkowski M,
Brown AM, Sedor JR and Schelling JR. Renal tubular epithelial cell apoptosis is
associated with caspase cleavage of the NHE1 Na+/H+ exchanger. Am J Physiol Renal
Physiol 284: F829-F839, 2003.
38. Zhan Y, van de WB, Wang Y and Stevens JL. The roles of caspase-3 and bcl-2 in
chemically-induced apoptosis but not necrosis of renal epithelial cells. Oncogene 18: 6505-
6512, 1999.
23

__________________________________________________________________________________________ Table I. Composition of Salt Solutions Tested in Fig 1B __________________________________________________________________________________________ Medium Component Saline L-15 HBSS F12 DMEM RPMI __________________________________________________________________________________________ CaCl2 - 1.3mM* 1.3 0.3 1.8 - KCl - 5.4 5.4 3.0 5.4 5.4 KH2PO4 - - 0.4 - - - MgCl2 - 1.0 - 0.6 - - MgSO4 - 0.8 0.8 - 0.8 0.4 NaCl 145.4 136.9 136.9 130.0 109.5 102.7 Na2HPO4 1.5 1.3 0.3 1.0 3.9 5.6 NaH2PO4 0.5 - - - 1.2 - HCl to adjust pH - 0.2 - 0.1 - 1.0 NaOH to adjust pH 0.3 - 0.3 - 0.6 - Na+ 149.2 139.5 137.8 132.0 119.1 113.9 Cl- 145.4 147.1 144.9 134.9 118.5 109.1 ___________________________________________________________________________________________ * all concentrations are mM

______________________________________ Table II. Osmolality of Tissue Culture Media ______________________________________ Osmolality Medium (mosmol/kg H2O) ______________________________________ RPMI 286 F-12 298 L-15 330 DMEM 350 ______________________________________

Fig. 1 Nephrotoxicity of Cisplatin in Tissue Culture Media (A) and Inorganic salt
Solutions (B). Panel A:Confluent monolayers of LLC-PK1 cells were incubated for 3 hrs
in cisplatin in RPMI (open squares), F-12 (closed triangles), DMEM (open diamonds ) or
L-15 medium with 18 mM HEPES and 0.2% BSA (closed circles). Panel B: Confluent
monolayers of LLC-PK1 cells were incubated for 3 hrs in cisplatin in the solutions
containing the inorganic salt component of RPMI (open squares), DMEM with 5mM
NaPO4 (open diamonds), F-12 (closed triangles), HBSS ((horizontal bars), L-15 (closed
circles) and saline buffered with 2mM NaPO4 (Xs). All solutions were adjusted to pH
7.4. For experiments in both panel A and B, the cisplatin solutions were removed at the
end of the 3 hr exposure and fresh DMEM medium with 5% FBS and antibiotics was
added to the cells. Cell survival was assayed at 72 hrs. Each point is the average of 3 to
six values + S.D. Values that differed significantly from the survival in L-15 medium
(panel A) or L-15 inorganic salt solution (Panel B) are indicated by * (p<0.01) or ** (p<
0.001).
0
20
40
60
80
100
0 50 100 150
Cisplatin (mM)
Su
rviv
al(
%)
0
20
40
60
80
100
120
140
0 50 100 150
Cisplatin (mM)S
urv
iva
l(%
)
A B
**
** **
**
***

0
20
40
60
80
100
125 135 145 155
NaCl (mM)
Su
rviv
al(%
)
Fig. 2 Influence of Sodium Chloride Concentration on Cisplatin Nephrotoxicity.
Confluent monolayers of LLC-PK1 cells were incubated for 3 hrs in solutions containing
100 mM cisplatin, with 5 mM HEPES, pH 7.4 and varying amounts of NaCl (open
circles) or in the HEPES/NaCl solutions without cisplatin (closed circles). The test
solutions were removed at the end of the 3 hr exposure and fresh DMEM medium with
5% FBS and antibiotics was added to the cells. Cell survival was assayed at 72 hrs. Each
point is the average of 3 values + S.D.

0
5
10
15
20
25
120 130 140 150 160
NaCl (mM)
Pt (n
g/10
5 cel
ls)
Fig. 3 The Effect of NaCl on the Accumulation of Platinum. Confluent monolayers of
LLC-PK1 cells were incubated for 3 hrs in solutions containing 100 µM cisplatin, with 5
mM HEPES, pH 7.4 and varying amounts of NaCl. At the end of 3 hrs the cisplatin was
rinsed off the cells and the amount of platinum that had accumulated in the cells was
determined by FAAS. Each point is the average of the mean value from 3 experiments ±
S.E.
0
20
40
60
80
100
100 120 140 160 180 200[Na+] (mM) in DMEM
during preincubation
Surv
ival
(%)
Fig. 4 Effect of Preconditioning Cells with Various Concentrations of NaCl on Cisplatin
Nephrotoxicity . Confluent monolayers of LLC-PK1 cells were incubated for 3 hr in fresh
DMEM containing 5% FBS, antibiotics and various concentrations of NaCl. After 3 hrs
the media was removed and all the cells were treated with 100 µM cisplatin in DMEM
(open circles) or DMEM alone (closed circles). The cisplatin solutions were removed at
the end of the 3 hr exposure and fresh DMEM with 5% FBS and antibiotics was added to
the cells. Cell survival was assayed at 72 hr. Each point is the average of 3 values ± S.D.

0
20
40
60
80
100
110 120 130 140 150 160
NaCl (mM)
Surv
ival
(%)
Fig. 5 Effect of NaCl vs. Osmolality on Cisplatin Toxicity. Confluent monolayers of
LLC-PK1 cells were incubated for 3 hr in100 µM cisplatin in 5 mM HEPES containing
NaCl alone (open diamonds), NaCl and mannitol (closed circles), or NaCl and N-methyl-
D-glucamine (open squares). The varying amounts of NaCl were balanced with mannitol
or N-methyl-D-glucamine such that the final osmolality of each solution was 288
mosmol/kg H2O. The cisplatin solutions were removed at the end of the 3 hr exposure
and fresh DMEM with 5% FBS and antibiotics was added to the cells. Cell survival was
assayed at 72 hr. Each point is the average of 3 values ± S.D.

0
20
40
60
80
100
200 220 240 260 280 300
Osmolality (mosmol/kg H2O)
surv
ival
(%)
Fig. 6. Effect of Osmolality on Cisplatin Toxicity. Various concentrations of mannitol
(closed circles), sucrose (closed triangles), N-methyl-D-glucamine (open squares) and
sodium gluconate (closed bar) were added to 115 mM NaCl with 5 mM HEPES.
Confluent monolayers of LLC-PK1 cells were incubated for 3 hr in each of the solutions
containing 100 µM cisplatin. The cisplatin solutions were removed at the end of the 3 hr
exposure and fresh DMEM medium with 5% FBS and antibiotics was added to the cells.
Cell survival was assayed at 72 hr. Each point is the average of 3 values ± S.D.

0
5
10
15
20
220 240 260 280 300
Osmolality (mOsm)
Pt (n
g/10
5 cel
ls)
0
500
1000
1500
2000
2500
3000
3500
4000
220 240 260 280 300
Osmolality (mOsm)
accu
mul
atio
n/(1
/osm
olal
ity)
A B
Fig. 7. The Effect of Osmolality on the Accumulation of Platinum. Confluent
monolayers of LLC-PK1 cells were incubated for 3 hrs in solutions containing 100 µM
cisplatin, with 115mM NaCl, 5 mM HEPES, pH 7.4. Mannitol was used to vary the
osmolarity of the solutions. At the end of 3 hrs the cisplatin was rinsed off the cells and
the amount of platinum that had accumulated in the cells was determined by FAAS. Each
point is the average of the mean value from three to six values + S.D. The data are
expressed as the platinum accumulation per cell (A) and platinum accumulation per cell
with a volume correction (B).

0
50
100
150
120 140 160
NaCl (mM)
Pt (
ng/m
g D
NA
)
`
Fig. 8. The Effect of NaCl on Platinum Bound to DNA. Confluent monolayers of LLC-
PK1 cells were incubated for 3 hrs in solutions containing 100 µM cisplatin, with 5 mM
HEPES, pH 7.4 and varying amounts of NaCl. At the end of 3 hrs the cisplatin was rinsed
off the cells, DNA was isolated and the amount of platinum bound to the DNA was
determined by FAAS. Each point is the average of 3 values + S.D.





























