Embed Size (px)
Citation preview

S
CDMa
b
c
a
ARRAA
KSCNNRO
1
thb1thaaanCia(t
0d
Toxicology Letters 192 (2010) 278–285
Contents lists available at ScienceDirect
Toxicology Letters
journa l homepage: www.e lsev ier .com/ locate / tox le t
ulforaphane protects against cisplatin-induced nephrotoxicity
arlos Enrique Guerrero-Beltrána , Mariel Calderón-Olivera , Edilia Tapiab , Omar N. Medina-Camposa ,olores Javier Sánchez-Gonzálezc, Claudia María Martínez-Martínezc, Karla Mariana Ortiz-Vegab,artha Francob, José Pedraza-Chaverri a,∗
Departamento de Biología, Facultad de Química, Universidad Nacional Autónoma de México, Edificio F, 04510 Mexico City, MexicoDepartamento de Nefrología, Instituto Nacional de Cardiología Ignacio Chávez, 04510 Mexico City, MexicoDepartamento de Biología Celular, Escuela Médico Militar, Universidad del Ejército y Fuerza Aérea, Mexico City, Mexico
r t i c l e i n f o
rticle history:eceived 5 October 2009eceived in revised form 30 October 2009ccepted 2 November 2009vailable online 12 November 2009
eywords:ulforaphaneisplatin
a b s t r a c t
Cisplatin (cis-diamminedichloroplatinum II, CDDP) is a chemotherapeutic agent that induces nephro-toxicity associated with oxidative/nitrosative stress. Sulforaphane (SFN) is an isothiocyanate producedby the enzymatic action of myrosinase on glucorophanin, a glucosinolate contained in cruciferousvegetables. SFN is able to induce cytoprotective enzymes through the transcription factor Nrf2. The pur-pose of this study was to evaluate whether SFN induces a cytoprotective effect on the CDDP-inducednephrotoxicity. Preincubation of LLC-PK1 cells with 0.5–5 �M SFN by 24 h was able to prevent, in aconcentration-dependent way, CDDP-induced cell death. Immunofluorescent staining confirmed thenuclear translocation of Nrf2 after treatment with SFN. In the in vivo studies, CDDP was given to Wis-
ephrotoxicityrf2enoprotectionxidative/nitrosative stress
tar rats as a sole i.p. injection at a dose of 7.5 mg/kg. SFN (500 �g/kg i.v.) was given two times (24 hbefore and 24 after CDDP-injection). Animals were killed three days after CDDP-injection. SFN attenuatedCDDP-induced renal dysfunction, structural damage, oxidative/nitrosative stress, glutathione depletion,enhanced urinary hydrogen peroxide excretion and the decrease in antioxidant enzymes (catalase, glu-tathione peroxidase and glutathione-S-transferase). The renoprotective effect of SFN on CDDP-inducednephrotoxicity was associated with the attenuation in oxidative/nitrosative stress and the preservation
of antioxidant enzymes.. Introduction
Cisplatin (cis-diamminedichloroplatinum II, CDDP) is an impor-ant chemotherapeutic agent in the clinical treatment of severaluman cancers including those of the head, neck, testis, ovary,reast, bladder, esophageal and cervical (Lebwohl and Canetta,998). Unfortunately, it has several side effects such as nephro-oxicity (Pabla and Dong, 2008). In vivo and in vitro evidencesave suggested that reactive oxygen species (ROS) like superoxidenion (O2
•−), hydrogen peroxide (H2O2), hydroxyl radical (OH•)nd peroxynitrite (ONOO−), depletion of antioxidant enzymesnd enhanced oxidative/nitrosative are involved in CDDP-inducedephrotoxicity (Chirino and Pedraza-Chaverri, 2009). In addition,DDP inhibits mitochondrial function and antioxidant activity,
nduces DNA damage and glutathione (GSH) depletion whichre strongly associated with the renal toxicity of this compoundChirino and Pedraza-Chaverri, 2009; Rios et al., 2009). In this con-ext, it has been shown that free radical scavengers and natural
∗ Corresponding author. Tel.: +52 55 5622 3878; fax: +52 55 5622 3878.E-mail address: [email protected] (J. Pedraza-Chaverri).
378-4274/$ – see front matter © 2009 Elsevier Ireland Ltd. All rights reserved.oi:10.1016/j.toxlet.2009.11.007
© 2009 Elsevier Ireland Ltd. All rights reserved.
and synthetic antioxidants ameliorate or prevent CDDP-inducednephrotoxicity (Chirino and Pedraza-Chaverri, 2009; Weijl et al.,2004; Ali and Al Moundhri, 2006; Chirino et al., 2004, 2008a,b;Pérez-Rojas et al., 2009).
Sulforaphane (SFN) is a naturally occurring isothiocyanateproduced by the enzymatic action of myrosinase on glucoro-phanin, a glucosinolate contained in cruciferous vegetables suchas broccoli, cabbage, brussel sprouts, etc (Fahey and Talalay, 1999;Dinkova-Kostova, 2008). It has been shown that SFN is an indi-rect antioxidant that acts by inducing Nrf2-dependent phase 2enzymes through the transcription NF-E2-related factor 2 (Nrf2).Under basal conditions cytoplasmic Nrf2 is bound to the Kelch-likeECH-associated protein 1 (Keap1). However, SFN can dissociate theNrf2/Keap1 complex, thereby permitting Nrf2 to translocate intothe nucleus (Juge et al., 2007; Zhang, 2006). After translocationinto nucleus, Nrf2 forms a heterodimer with other transcriptionfactors such as small Maf, which in turn binds to the 5′-upstream
cis-acting regulatory sequence, termed antioxidant response ele-ment (ARE) or electrophile response elements (EpRE) located inthe promoter region of genes encoding various antioxidant andphase 2 detoxifying enzymes, including heme oxygenase-1 (HO-1),NAD(P)H: quinone oxidoreductase 1 (NQO1), glutathione reductase
xicolog
(Sda
tcii
2
2
PUo3(2oaMp(1GmpamAJb(aLAPH(a
2
2
oCucaa(N5w5tm
2
tcrBtinfsflTT
C.E. Guerrero-Beltrán et al. / To
GR), glutathione peroxidase (GPx), catalase (CAT) and glutathione--transferase (GST). Phase 2 enzymes play a major role in theetoxification of ROS produced by xenobiotics (Dinkova-Kostovand Talalay, 2008).
Based on the above information, we hypothesized that SFNreatment may be protective against cisplatin-induced nephrotoxi-ity. The purpose of the present study was to evaluate whether SFNnduces a cytoprotective effect on the CDDP-induced nephrotoxic-ty in LLC-PK1 cells and in rats.
. Materials and methods
.1. Reagents
Porcine renal epithelial cells Lily Laboratory Culture Porcine Kidney (LLC-K1) were obtained from American Type Culture Collection (Rockville, MD,SA). SFN (Cat. no. S8044, batch 26815401) was purchased from LKT lab-ratories (St. Paul, MN, USA). CDDP (Cat. no. P4394-1Gm batch 087K1349),-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium (MTT), dimethyl sulfoxideDMSO), p-nitrophenyl-N-acetyl-�-d-glucosaminide, nitroblue tetrazolium (NBT),,4-dinitrophenylhydrazine (DNPH), chloro-2,4,dinitrobenzene (CDNB), GST, Flu-romount, triton X-100, poly-l-lysine (P-1524), paraformaldehyde, bovine serumlbumin (BSA) and glycine were purchased from Sigma Chemical Co. (St. Louis,O, USA). Monochlorobimane and Hoechst 33258 stain (Cat. no. 14530) were
urchased by Fluka (Schnelldorf, Germany). Dulbecco’s modified eagle mediumDMEM), fetal bovine serum (FBS), trypsin, antibiotic (10,000 U/ml penicillin and0,000 �g/ml streptomycin) and other tissue culture reagents were obtained fromibco. Cell culture plates were obtained by Nunc (Roskilde, Denmark). Mouseonoclonal anti-4-hydroxy-2-nonenal (4-HNE) antibody (Cat. no. 24325) was
urchased from Oxis International Inc. (Portland, OR, USA). Mouse monoclonalnti-3-nitrotyrosine (3-NT) antibody (Cat. no. 189542) was purchased from Cay-an Chemical Co. (Ann Arbor, MI, USA). Secondary antibody biotin SP-conjugatedffiniPure donkey anti-mouse IgG (Cat. no. 715-065-151) was purchased from
ackson ImmunoResearch Laboratories Inc. (West Grove, PA, USA), anti-Nrf2 anti-odies (Cat. no. sc-722) were purchased from Santa Cruz Biotechnology Inc.Santa Cruz, CA, USA), fluorescein isothiocyanate (FITC) conjugated secondaryntibodies (Cat. no. 111-095-144) were purchased from Jackson Immunoresearchaboratories (West Grove, PA, USA). Declere was from Cell Marque (Hot Springs,R, USA). ABC-kit Vectastain was from Vector Laboratories (Orton Southgate,eterborough, UK). Diaminobenzidine substrate (Cat. no. K3466) and Mayer’sematoxylin (Lillie’s Modification) (Cat. no. S3309) were from DAKO Corporation
Carpinteria, CA, USA). All other chemicals were reagent grade and commerciallyvailable.
.2. In vitro studies
.2.1. Cell culture and viabilityLLC-PK1 cells were maintained in DMEM supplemented with 10% FBS and 1%
f antibiotic. Cells were cultured under permissive conditions: 37 ◦C and 5% CO2.ells were seeded at a density of 4 × 104 cells/cm2 onto 12-well plates and weresed for experiments on the following day. Cells were exposed to different con-entrations of CDDP (0–50 �M) or SFN (0–100 �M) for 24 h. The cell viability wasssessed by reduction of MTT to formazan as previously described (Orozco-Ibarra etl., 2009). After treatment, cells were washed twice with phosphate-buffered salinePBS, pH 7.4), which was followed by the addition of MTT (0.125 mg/ml of DMEM).ext, the cells were incubated at 37 ◦C for 1 h in humidified air supplemented with% CO2. Medium was then discarded and the formazan crystals, formed in cells,ere dissolved in isopropanol acid (500 �l/well). The absorbance was measured at
70 nm. In additional experiments bright field images were obtained by phase con-rast microscopy Nikon Eclipse TS100F (Nikon Co., Tokyo, Japan) to compare cell
orphology in all experimental conditions.
.2.2. Immunocytochemical localization of Nrf2The cells were seeded on 12-well plates containing 18 mm glass coverslips-
reated with 0.025% poly-l-lysine and growth for 24 h. SFN was added for 3 h, andells were washed with PBS and fixed with 4% paraformaldehyde for 10 min atoom temperature, permeabilized with 1% triton X-100 for 20 min, blocked with 1%SA–1% triton X-100–0.3 M glycine and incubated with anti-Nrf2 (in 1% BSA–1% tri-on X-100) for 2.5 h at room temperature. The coverslips were incubated overnightn the dark at 4 ◦C with FITC conjugated secondary antibody and washed with PBS. A
uclear counterstaining was made with a solution of 0.2 �g/ml Hoechst 33258 stainor 1 min and mounting on a slide using Fluoromount. The coverslips edges wereealed with nail polish to prevent drying and movement under microscopy. Theuorescence was visualized in an inverted fluorescence microscope (Nikon EclipseS-100F) using B-2A/C filter for FITC fluorescence and UV-2A filter for Hoechst signal.he images were acquired with a Nikon Digital Sight DS-Fi 1 camera.
y Letters 192 (2010) 278–285 279
2.3. In vivo studies
2.3.1. Experimental designMale Wistar rats with an initial body weight of 230–260 g were used. Exper-
imental work followed the guidelines of Norma Official Mexicana Guide forthe use and care of laboratory animals (NOM-062-ZOO-1999) and for the dis-posal of biological residues (NOM-087-ECOL-1995). Four groups of rats werestudied (n = 8–12/group): (1) CT, control injected via jugular vein with isotonicsaline solution and DMSO/isotonic saline solution by intraperitoneal injection;(2) SFN, SFN was suspended in isotonic saline solution and injected at a dose of500 �g/kg (Yoon et al., 2008) via jugular vein two times (24 h before and 24 hafter DMSO/isotonic saline solution injection). (3) CDDP, CDDP in isotonic salinesolution was administered to rats by a single intraperitoneal injection (7.5 mg/kg)(Chirino et al., 2008a). (4) CDDP + SFN, SFN was suspended in isotonic saline solu-tion and injected at a dose of 500 �g/kg (Yoon et al., 2008) via jugular veintwo times (24 h before and 24 h after 7.5 mg/kg CDDP-injection). Rats had freeaccess to water and food and were maintained in metabolic cages to collect24 h urine for the measurement of proteinuria and excretion of N-acetyl-�-d-glucosaminidase (NAG), as markers of nephrotoxicity, and H2O2 as a marker of ROSproduction.
Animals were anesthetized with ether 72 h after CDDP or vehicle injectionand blood was obtained via aorta using a syringe and a needle #18 at roomtemperature. Blood plasma was separated and stored at −20 ◦C until the follow-ing markers of renal damage, serum creatinine, blood urea nitrogen (BUN) andplasma GPx were measured (Pedraza-Chaverri et al., 2000). Creatinine clearancewas calculated with the standard formula. One kidney was quickly removed toobtain samples for histological, immunohistochemical and morphometric studiesand to measure the activity of GPx, CAT and glutathione-S-transferase (GST). Theother kidney was removed and stored at −80 ◦C until protein carbonyl contentand 4-HNE, markers of protein oxidation and lipid peroxidation, respectively, weremeasured.
2.3.2. Analytical methodsBUN and creatinine were measured using commercial kits according to the
instructions of the manufacturers. Proteinuria was measured by a turbidimetricmethod using 12.5% trichloroacetic acid at 420 nm and the data were expressedas mg/24 h (Pedraza-Chaverri et al., 2008). Urinary NAG activity was determinedat 405 nm using p-nitrophenyl-N-acetyl-�-d-glucosaminide as substrate and datawere expressed as U/24 h (Pedraza-Chaverri et al., 2008). The urinary excretionof H2O2 was performed by a previously described method using xylenol orange;the colorimetric complex was detected at 560 nm (Long and Halliwell, 2001).GSH levels were measured using monochlorobimane as previously described byFernández-Checa and Kaplowitz (1990). The fluorescence was measured usingexcitation and emission wavelengths 385 and 478 nm, respectively, using aSynergy HT multi-mode microplate reader (Biotek Instruments Inc., Winooski,VT, USA).
2.3.3. Histopathological and immunohistochemical studiesKidney sections were fixed in 10% neutral buffered formalin solution and
embedded in paraffin. Histopathological and immunohistochemical studies wereperformed as previously described (Pérez-Rojas et al., 2009; Chirino et al., 2008b;Pedraza-Chaverri et al., 2008).
2.3.4. Protein carbonyl and 4-hydroxy-2-nonenal contentProtein carbonyl content in the kidney tissue was determined as previously
described (Pedraza-Chaverri et al., 2008). 4-HNE was measured according to apreviously described method (Gérard-Monnier et al., 1998). The reaction mixturewas incubated at 45 ◦C for 30 min and was measured upon 586 nm absorbance.The final 586 nm absorbance was a linear function of the concentrations of 4-hydroxyalkenals. An apparent molar extinction coefficient of 110,000 M−1 cm−1 wasobtained with the diethylacetals of 4-HNE.
2.3.5. Antioxidant enzymesCAT activity was assayed by a method based on the disappearance of H2O2 at
240 nm and GPx activity was measured by the disappearance of NADPH at 340 nmas previously described method (Pedraza-Chaverri et al., 2000). GST activity wasassayed in a mixture containing GSH, and CDNB as previously described (Pedraza-Chaverri et al., 2008).
2.4. Statistical analyses
Results were expressed as mean ± SEM. Data were analyzed by one-way ANOVAfollowed by Bonferroni’s multiple comparisons test using software Prism 2.01(GraphPad, San Diego, CA, USA). A p-value <0.05 was considered statistically sig-nificant.

280 C.E. Guerrero-Beltrán et al. / Toxicology Letters 192 (2010) 278–285
F ed with CDDP (0–50 �M) or SFN (0–100 �M) for 24 h and cell viability was determinedu
3
3
3
c5mc4w
3
mC((t5pi
Fig. 2. SFN prevents CDDP-induced cells death. LLC-PK1 cells were treated withSFN (0–5 �M) for 24 h and then CDDP (40 �M) was added for additional 24 h. Cell
F2
ig. 1. Effect of (A) CDDP and (B) SFN on cell viability. LLC-PK1 cells were incubatsing an MTT assay. Data are mean ± SEM. *p < 0.001 vs. CT.
. Results
.1. In vitro studies
.1.1. Effect of CDDP or SFN on renal epithelial cells viabilityAs shown in Fig. 1, CDDP decreased cell viability in a
oncentration-dependent manner. Exposure to CDDP (from 20 to0 �M) produced a significant reduction in cell viability. Further-ore, SFN (from 20 to 100 �M) produced a significant decrease in
ell viability (Fig. 1). In the next experiments a concentration of0 �M of CDDP, which reduced cell viability in approximately 50%,as used.
.1.2. SFN protects against CDDP-induced cell deathThe next experiment was designed to find out if SFN pretreat-
ent was able to prevent CDDP-induced LLC-PK1 cell damage.DDP treatment significantly reduced cell viability to 52.1 ± 1.99%p < 0.001) of the control value (Fig. 2). Pretreatment with SFN
0.5–5 �M) resulted in a substantial concentration-dependent pro-ection against CDDP-induced damage. Viability increased from2.1 ± 1.99% in CDDP-treated cells to 90.1 ± 4.7% with 5 �M SFNretreatment in CDDP + SFN treated cells. In a separate exper-ment, cell morphology under all experimental conditions, was
ig. 3. Bright field images obtained by phase contrast microscopy. LLC-PK1 cells were tre4 h. The arrows indicate dead cells. 40×.
viability was then determined using the MTT assay. Data are mean ± SEM. *p < 0.001vs. CT; **p < 0.001 vs. CDDP.
analyzed under phase contrast microscopy (Fig. 3). The infor-
mation obtained under these conditions (Fig. 3) confirmed theconcentration–response protection induced by SFN with the MTTmethod (Fig. 2).ated with SFN (0–5 �M) for 24 h and then CDDP (40 �M) was added for additional

C.E. Guerrero-Beltrán et al. / Toxicology Letters 192 (2010) 278–285 281
Fd
Fv
ig. 4. SFN induces translocation of Nrf2 in LLC-PK1 cells. LLC-PK1 cells were incubated wetermined by immunofluorescent staining for Nrf2 and counterstained with Hoechst 33
ig. 5. Markers of nephrotoxicity in the groups of rats studied. (A) Serum creatinine, (B) cres. CT; #p < 0.001 vs. CDDP. Values are mean ± SEM.
ith SFN (10 and 25 �M) for 3 h. Nuclear translocation of Nrf2 by SFN treatment was258.
atinine clearance, (C) BUN and (D) GPx activity. *p < 0.001 vs. CT and SFN. **p < 0.001

282 C.E. Guerrero-Beltrán et al. / Toxicology Letters 192 (2010) 278–285
ts stu
3
r
3
3
fwI
Fai
Fig. 6. Urinary excretion of (A) total protein and (B) NAG in the groups of ra
.1.3. SFN induces nuclear Nrf2 translocation in LLC-PK1 cellsFig. 4 shows that incubation with two concentrations of SFN
esulted in a significant translocation of Nrf2 to the nucleus.
.2. In vivo studies
.2.1. Effect of SFN on CDDP-induced nephrotoxicityIn the CDDP group, creatinine and BUN increased 3.6- and 3.2-
old, respectively, and creatinine clearance decreased 76% whichere prevented by SFN treatment in the CDDP + SFN group (Fig. 5).
n addition, CDDP-induced a decrease in plasma GPx activity to 16%
ig. 7. Histopathological and immunohistochemical analysis in the four groups of rat stund (+) indicates necrosis observed like eosinophilic areas with vacuoles and destructionnside the tubular lumen. Rats were studied after 72 h of CDDP treatment (7.5 mg/kg).
died. *p < 0.001 vs. CT and SFN; #p < 0.001 vs. CDDP. Values are mean ± SEM.
of control value and SFN treatment prevented this decrease in theCDDP + SFN group (Fig. 5). Moreover, SFN treatment was able toprevent the CDDP-induced increase of urinary excretion of totalprotein and NAG of 3.3- and 5.1-fold, respectively (Fig. 6). All theabove mentioned parameters were unaffected in the SFN group.
3.2.2. Histopathological studiesHistological evaluation was performed using H&E staining
and PAS, in order to demonstrate polysaccharides inside thetubuli (Fig. 7). Light microscopy studies revealed that kidneysisolated from animals after 72 h of CDDP administration pre-
dies. Asterisks indicate apoptosis observed clearly in cellular nuclei of tubular cellsof the cytoplasmic morphology of tubular cells and arrows show numerous casts

C.E. Guerrero-Beltrán et al. / Toxicology Letters 192 (2010) 278–285 283
F f rat st(
sacsglipa
3
mlan
FS
ig. 8. Quantitative data of the immunohistochemical studies (%) in four groups oB) 3-NT. *p < 0.001 vs. CT and SFN; **p < 0.01 vs. CT and SFN; #p < 0.001 vs. CDDP.
ented epithelial tubular injury, most of cortical tubuli showedpoptosis (asterisks), necrosis and vacuolization in epithelialells; and tubular casts (arrows) observed clearly with PAStaining (Fig. 7). In contrast, kidneys isolated from CDDP + SFNroup, revealed a nearly regular morphology of epithelial tubu-ar cells. Renal damage was significantly attenuated with SFNn the CDDP + SFN group (affected area: 21.2 ± 2% vs. 11.2 ± 0.8%,< 0.001). Slices from the control and SNF groups showed normalrchitecture.
.2.3. Oxidative/nitrosative stress markers
In addition, renal immunohistochemistry of 3-NT and 4-HNE asarkers of nitrosative and oxidative stress, respectively, were ana-yzed in kidney slides from studied groups. As shown in Figs. 7 and 8,weak immunostaining for 3-NT and 4-HNE was observed in kid-ey sections of control group. In contrast, a strong staining for
ig. 9. Oxidative stress markers in the groups of rats studied. (A) 4-HNE, (B) protein carbFN; #p < 0.001 vs. CDDP. Values are mean ± SEM.
udies. Rats were studied after 72 h of CDDP treatment (7.5 mg/kg). (A) 4-HNE and
3-NT and 4-HNE was observed in CDDP-treated rats (Figs. 7 and 8).SFN significantly attenuated the kidney of nitrosative and oxidativestress; a weak immunoreactivity for 3-NT and 4-HNE was observedin CDDP + SFN group (Figs. 7 and 8).
Furthermore, 4-HNE levels, protein carbonyl content andurinary excretion of H2O2 increased 1.6-, 1.5- and 5.2-fold,respectively, and GSH levels decreased to 53% of the controlvalues in CDDP-treated rats (Fig. 9). These changes were sig-nificantly prevented by SFN treatment in the CDDP + SFN group(Fig. 9).
3.2.4. Antioxidant enzymesThe renal activity of CAT, GPx and GST decreased 35%, 46%, and
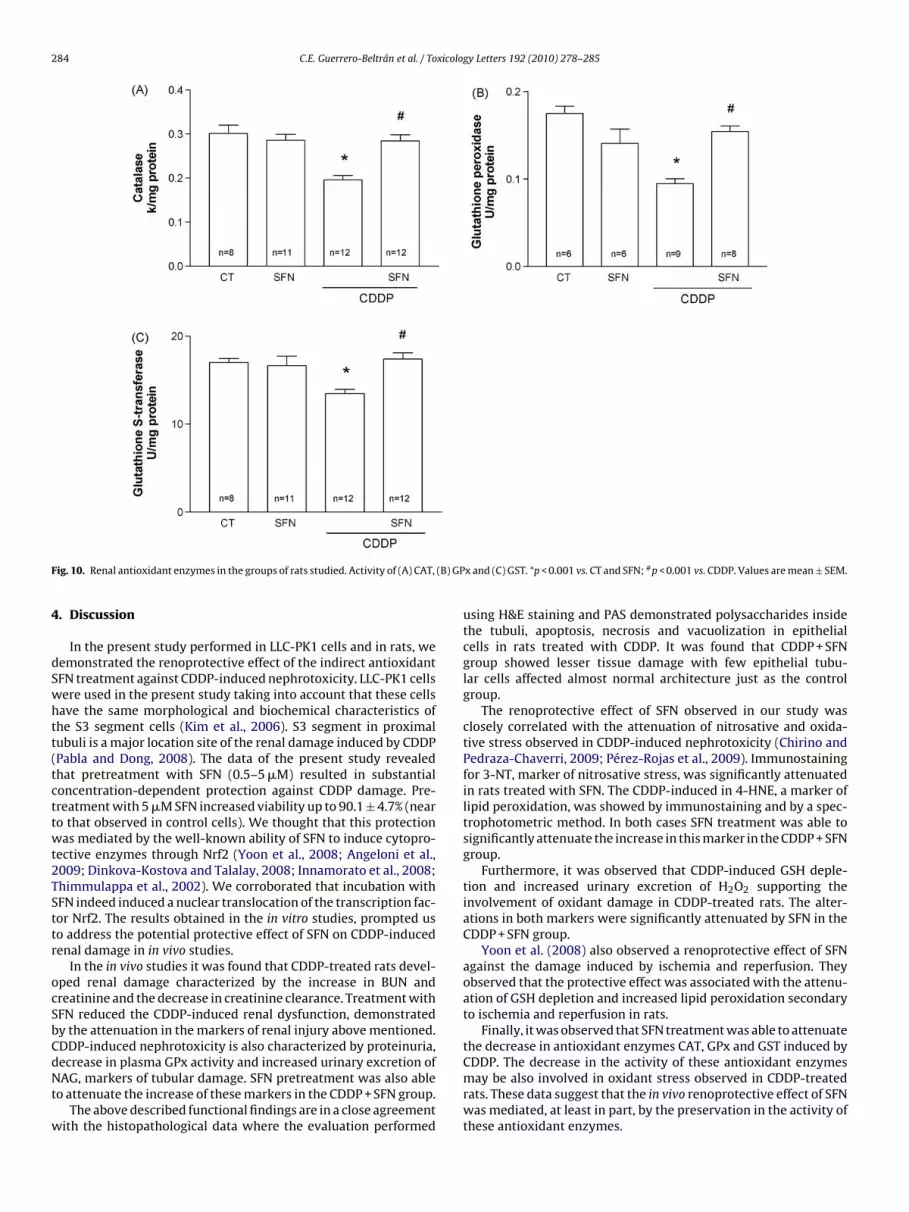
21%, respectively, in CDDP-treated rats (Fig. 10). These changeswere completely prevented by SFN treatment in the CDDP + SFNgroup (Fig. 10).
onyl content, (C) GSH content and (D) urinary H2O2 excretion. *p < 0.05 vs. CT and
284 C.E. Guerrero-Beltrán et al. / Toxicology Letters 192 (2010) 278–285
F (B) GP
4
dSwhtt(tcttwt2TSttr
ocSbCdNt
w
ig. 10. Renal antioxidant enzymes in the groups of rats studied. Activity of (A) CAT,
. Discussion
In the present study performed in LLC-PK1 cells and in rats, weemonstrated the renoprotective effect of the indirect antioxidantFN treatment against CDDP-induced nephrotoxicity. LLC-PK1 cellsere used in the present study taking into account that these cellsave the same morphological and biochemical characteristics ofhe S3 segment cells (Kim et al., 2006). S3 segment in proximalubuli is a major location site of the renal damage induced by CDDPPabla and Dong, 2008). The data of the present study revealedhat pretreatment with SFN (0.5–5 �M) resulted in substantialoncentration-dependent protection against CDDP damage. Pre-reatment with 5 �M SFN increased viability up to 90.1 ± 4.7% (nearo that observed in control cells). We thought that this protectionas mediated by the well-known ability of SFN to induce cytopro-
ective enzymes through Nrf2 (Yoon et al., 2008; Angeloni et al.,009; Dinkova-Kostova and Talalay, 2008; Innamorato et al., 2008;himmulappa et al., 2002). We corroborated that incubation withFN indeed induced a nuclear translocation of the transcription fac-or Nrf2. The results obtained in the in vitro studies, prompted uso address the potential protective effect of SFN on CDDP-inducedenal damage in in vivo studies.
In the in vivo studies it was found that CDDP-treated rats devel-ped renal damage characterized by the increase in BUN andreatinine and the decrease in creatinine clearance. Treatment withFN reduced the CDDP-induced renal dysfunction, demonstratedy the attenuation in the markers of renal injury above mentioned.DDP-induced nephrotoxicity is also characterized by proteinuria,
ecrease in plasma GPx activity and increased urinary excretion ofAG, markers of tubular damage. SFN pretreatment was also ableo attenuate the increase of these markers in the CDDP + SFN group.The above described functional findings are in a close agreement
ith the histopathological data where the evaluation performed
x and (C) GST. *p < 0.001 vs. CT and SFN; #p < 0.001 vs. CDDP. Values are mean ± SEM.
using H&E staining and PAS demonstrated polysaccharides insidethe tubuli, apoptosis, necrosis and vacuolization in epithelialcells in rats treated with CDDP. It was found that CDDP + SFNgroup showed lesser tissue damage with few epithelial tubu-lar cells affected almost normal architecture just as the controlgroup.
The renoprotective effect of SFN observed in our study wasclosely correlated with the attenuation of nitrosative and oxida-tive stress observed in CDDP-induced nephrotoxicity (Chirino andPedraza-Chaverri, 2009; Pérez-Rojas et al., 2009). Immunostainingfor 3-NT, marker of nitrosative stress, was significantly attenuatedin rats treated with SFN. The CDDP-induced in 4-HNE, a marker oflipid peroxidation, was showed by immunostaining and by a spec-trophotometric method. In both cases SFN treatment was able tosignificantly attenuate the increase in this marker in the CDDP + SFNgroup.
Furthermore, it was observed that CDDP-induced GSH deple-tion and increased urinary excretion of H2O2 supporting theinvolvement of oxidant damage in CDDP-treated rats. The alter-ations in both markers were significantly attenuated by SFN in theCDDP + SFN group.
Yoon et al. (2008) also observed a renoprotective effect of SFNagainst the damage induced by ischemia and reperfusion. Theyobserved that the protective effect was associated with the attenu-ation of GSH depletion and increased lipid peroxidation secondaryto ischemia and reperfusion in rats.
Finally, it was observed that SFN treatment was able to attenuatethe decrease in antioxidant enzymes CAT, GPx and GST induced by
CDDP. The decrease in the activity of these antioxidant enzymesmay be also involved in oxidant stress observed in CDDP-treatedrats. These data suggest that the in vivo renoprotective effect of SFNwas mediated, at least in part, by the preservation in the activity ofthese antioxidant enzymes.
xicolog
awa
C
A
apTia
R
A
A
C
C
C
C
D
D
F
F
C.E. Guerrero-Beltrán et al. / To
In summary, our data show the renoprotective effect of SFNgainst the CDDP-induced nephrotoxicity. This protective effectas associated with the attenuation in nitrosative/oxidant stress
nd with the preservation in antioxidant enzymes.
onflict of interest
The authors declare that there are no conflicts of interest.
cknowledgements
This work was supported by DGAPA IN207007 and IN201910nd by CONACYT 89641. We thank Arturo Ruiz-García for his sup-ort with some analytical determinations. We also thank Dr. Ismaelorres-Saldana and Dr. Enrique Pinzón-Estrada (Facultad de Medic-na, UNAM) for their continuous support with the experimentalnimals.
eferences
li, B.H., Al Moundhri, M.S., 2006. Agents ameliorating or augmenting the nephro-toxicity of cisplatin and other platinum compounds: a review of some recentresearch. Food Chem. Toxicol. 44, 1173–1183.
ngeloni, C., Leoncini, E., Malaguti, M., Angelini, S., Hrelia, P., Hrelia, S., 2009. Modu-lation of phase II enzymes by sulforaphane: implications for its cardioprotectivepotential. J. Agric. Food Chem. 57, 5615–5622.
hirino, Y.I., Hernández-Pando, R., Pedraza-Chaverri, J., 2004. Peroxynitritedecomposition catalyst ameliorates renal damage and protein nitration incisplatin-induced nephrotoxicity in rats. BMC Pharmacol. 4, 20.
hirino, Y.I., Sánchez-González, D.J., Martínez-Martínez, C.M., Cruz, C., Pedraza-Chaverri, J., 2008a. Protective effects of apocynin against cisplatin-inducedoxidative stress and nephrotoxicity. Toxicology 245, 18–23.
hirino, Y.I., Trujillo, J., Sanchez-Gonzalez, D.J., Martinez-Martinez, C.M., Cruz, C.,Bobadilla, N.A., Pedraza-Chaverri, J., 2008b. Selective iNOS inhibition reducesrenal damage induced by cisplatin. Toxicol. Lett. 176, 48–57.
hirino, Y.I., Pedraza-Chaverri, J., 2009. Role of oxidative and nitrosative stress incisplatin-induced nephrotoxicity. Exp. Toxicol. Pathol. 61, 223–242.
inkova-Kostova, A.T., 2008. Phytochemicals as protectors against ultraviolet radi-ation: versatility of effects and mechanisms. Planta Med. 74, 1548–1559.
inkova-Kostova, A.T., Talalay, P., 2008. Direct and indirect antioxidant proper-
ties of inducers of cytoprotective proteins. Mol. Nutr. Food Res. 52 (Suppl. 1),S128–S138.ahey, J.W., Talalay, P., 1999. Antioxidant functions of sulforaphane: a potent inducerof phase II detoxication enzymes. Food Chem. Toxicol. 37, 973–979.
ernández-Checa, J.C., Kaplowitz, N., 1990. The use of monochlorobimane to deter-mine hepatic GSH levels and synthesis. Anal. Biochem. 190, 212–219.
y Letters 192 (2010) 278–285 285
Gérard-Monnier, D., Erdelmeier, I., Régnard, K., Moze-Henry, N., Yadan, J.C.,Chaudière, J., 1998. Reactions of 1-methyl-2-phenylindole with malondialde-hyde and 4-hydroxyalkenals. Analytical applications to a colorimetric assay oflipid peroxidation. Chem. Res. Toxicol. 11, 1176–1183.
Innamorato, N.G., Rojo, A.I., García-Yagüe, A.J., Yamamoto, M, de Ceballos, M.L.,Cuadrado, A., 2008. The transcription factor Nrf2 is a therapeutic target againstbrain inflammation. J. Immunol. 181, 680–689.
Juge, N., Mithen, R.F., Traka, M., 2007. Molecular basis for chemoprevention by SFN:a comprehensive review. Cell Mol. Life Sci. 64, 1105–1127.
Kim, Y.H., Kim, Y.W., Oh, Y.J., Back, N.I., Chung, S.A., Chung, H.G., Jeong, T.S., Choi, M.S.,Lee, K.T., 2006. Protective effect of the ethanol extract of the roots of Brassica rapaon cisplatin-induced nephrotoxicity in LLC-PK1 cells and rats. Biol. Pharm. Bull.29, 2436–2441.
Lebwohl, D., Canetta, R., 1998. Clinical development of platinum complexes in cancertherapy: an historical perspective and an update. Eur. J. Cancer 34, 1522–1534.
Long, L.H., Halliwell, B., 2001. Antioxidant and prooxidant abilities of foods andbeverages. Methods Enzymol. 335, 181–190.
Orozco-Ibarra, M., Estrada-Sánchez, A.M., Massieu, L., Pedraza-Chaverrí, J., 2009.Heme oxygenase-1 induction prevents neuronal damage triggered during mito-chondrial inhibition: role of CO and bilirubin. Int. J. Biochem. Cell Biol. 41,1304–1314.
Pabla, N., Dong, Z., 2008. Cisplatin nephrotoxicity: mechanisms and renoprotectivestrategies. Kidney Int. 73, 994–1007.
Pedraza-Chaverri, J., Maldonado, P., Medina-Campos, O.N., Olivares-Corichi, I.M.,Granados-Silvestre, M.A., Hernández-Pando, R., Ibarra-Rubio, M.E., 2000. Garlicameliorates gentamicin nephrotoxicity: relation to antioxidant enzymes. FreeRadic. Biol. Med. 29, 602–611.
Pedraza-Chaverri, J., Yam-Canul, P., Chirino, Y.I., Sánchez-González, D.J., Martínez-Martínez, C.M., Cruz, C., Medina-Campos, O.N., 2008. Protective effects of garlicpowder against potassium dichromate-induced oxidative stress and nephrotox-icity. Food Chem. Toxicol. 46, 619–627.
Pérez-Rojas, J.M., Cruz, C., García-López, P., Sánchez-González, D.J., Martínez-Martínez, C.M., Ceballos, G., Espinosa, E., Meléndez-Zajgla, J., Pedraza-Chaverri,J., 2009. Renoprotection by �-mangostin is related to the attenuation in renaloxidative/nitrosative stress induced by cisplatin nephrotoxicity. Free Radic. Res.43, 1122–1132.
Rios, A., de, O., Antunes, L.M.G., Bianchi, M., de, L.P., 2009. Bixin and lycopene mod-ulation of free radical generation induced by cisplatin–DNA interaction. FoodChem. 113, 1113–1118.
Thimmulappa, R.K., Mai, K.H., Srisuma, S., Kensler, T.W., Yamamoto, M., Biswal, S.,2002. Identification of Nrf2-regulated genes induced by the chemopreventiveagent sulforaphane by oligonucleotide microarray. Cancer Res. 62, 5196–5203.
Weijl, N.I., Elsendoorn, T.J., Lentjes, E.G., Hopman, G.D., Wipkink-Bakker, A., Zwin-derman, A.H., Cleton, F.J., Osanto, S., 2004. Supplementation with antioxidantmicronutrients and chemotherapy-induced toxicity in cancer patients treatedwith cisplatin-based chemotherapy: a randomised, double-blind, placebo-
controlled study. Eur. J. Cancer 40, 1713–1723.Yoon, H.Y., Kang, N.I., Lee, H.K., Jang, K.Y., Park, J.W., Park, B.H., 2008. Sulforaphaneprotects kidneys against ischemia-reperfusion injury through induction of theNrf2-dependent phase 2 enzyme. Biochem. Pharmacol. 75, 2214–2223.
Zhang, D.D., 2006. Mechanistic studies of the Nrf2-Keap1 signaling pathway. DrugMetab. Rev. 38, 769–789.





























