Embed Size (px)
Citation preview

doi:10.1016/j.jmb.2006.10.017 J. Mol. Biol. (2007) 367, 303–309
COMMUNICATION
Structural Insights into Fibronectin Type IIIDomain-mediated Signaling
Sompop Bencharit1,2, Cai Bin Cui3, Adnan Siddiqui3
Escher L. Howard-Williams4, John Sondek2,5, Kheir Zuobi-Hasona6
and Ikramuddin Aukhil3,6⁎
1Department of Prosthodontics,School of Dentistry, Universityof North Carolina, Chapel Hill,NC 27599, USA2Department of Pharmacology,School of Medicine, Universityof North Carolina, Chapel Hill,NC 27599, USA3Department of Periodontology,School of Dentistry, Universityof North Carolina, Chapel Hill,NC 27599, USA4Department of Medicine,School of Medicine, Chapel Hill,NC 27599, USA5Department of Biochemistryand Biophysics, School ofMedicine, University of NorthCarolina, Chapel Hill,NC 27599, USA6Department of Periodontology,College of Dentistry, Universityof Florida, Gainesville,FL 32610, USAAbbreviations used: FN, fibronectiE-mail address of the correspondi
0022-2836/$ - see front matter © 2006 E
The alternatively spliced type III extradomain B (EIIIB) of fibronectin (FN)is expressed only during embryogenesis, wound healing and tumorigen-esis. The biological function of this domain is unclear. We describe herethe first crystal structure of the interface between alternatively splicedEIIIB and its adjacent FN type III domain 8 (FN B-8). The opened CC′loop of EIIIB, and the rotation and tilt of EIIIB allow good access to theFG loop of FN-8, which is normally hindered by the CC′ loop of FN-7. Inaddition, the AGEGIP sequence of the CC′′ loop of EIIIB replaces theNGQQGN sequence of the CC′ loop of FN-7. Finally, the CC′′ loop ofEIIIB forms an acidic groove with FN-8. These structural findings warrantfuture studies directed at identifying potential binding partners for FN B-8 interface, linking EIIIB to skeletal and cartilaginous development,wound healing, and tumorigenesis, respectively.
© 2006 Elsevier Ltd. All rights reserved.
*Corresponding author
Keywords: fibronectin; EIIIB; neovascularization; structureFibronectin (FN) is a multifunctional proteinserving major roles in the adhesion, migration,differentiation and proliferation of cells with impli-cations in embryonic development, wound healing,and tumorigenesis.1 The significance of FN inembryonic development has been documented bythe embryonic lethality seen in mice when the FNgene is disrupted.2 FN is a high molecular mass
n; FN-III, FN type-III.ng author:
lsevier Ltd. All rights reserve
dimeric glycoprotein with disulfide-linked subunits,each with a molecular mass of approximately 220–250 kDa. FN interacts with several other proteins inthe extracellular matrix, including collagen, heparin,fibrin, and cell membrane receptors. The amino acidcomposition of FN reveals it to be a large modularglycoprotein composed of homologous repeats ofthree prototypical types of domains known as typesI, II and III.3 Of these domains, FN type III (FN-III)repeats are composed of ≈90 amino acid residues,making them the largest and most common of theFN subdomains.4 One of the best-characterizedinteractions between FN and cell membrane recep-
d.
304 Fibronectin Type III Domain-mediated Signaling
tors is the interaction between the RGD sequence ofthe FN-III domain 10 (FN-10) and the extracellulardomain of β1-integrin.5–7 This interaction is believed
Figure 1 (legend
to regulate integrin signaling and, therefore, mayplay an important role in integrin-mediated celldifferentiation.8–10
on next page)

305Fibronectin Type III Domain-mediated Signaling
Among the type III domains of FN that span thecentral part of the FN molecule, three regionsreferred to as EIIIA, EIIIB and IIICS, respectively,are subject to alternative splicing of a primarytranscript.3–11 Of these spliced domains, EIIIA andEIIIB are either included or excluded by exonskipping and their alternative splicing is regulatedin a tissue-specific and developmental stage-depen-dent manner.11,12 The amino acid sequence ofalternatively spliced exons EIIIA and EIIIB is wellconserved, suggesting their functional impor-tance.13 Initial studies showed EIIIA and EIIIB splicevariants of FN expressed prominently arounddeveloping blood vessels during embryonic deve-lopment.14,15 These splice variants are absent fromnormal adult tissues, except during wound healingand neoplastic vascularization. However, EIIIB-nulland EIIIA-null mice develop normally with onlysome small differences in wound healing, athero-sclerosis and life-span in the EIIIA-null mice.13
Fibroblasts cultured from EIIIB-null mice growslowly in vitro and deposit less FN in the pericellularmatrix compared to fibroblasts cultured from wild-type mice.12 EIIIB expression is found predomi-nantly in breast and colorectal carcinomas. It wasbelieved that tumor cells express EIIIB FN to recruitendothelial cells in order to supply the tumor masswith oxygen and nutrients. Thus, EIIIB may be oneof a few clinical markers of neovascularization.16
Antibodies and variations of recombinant antibo-dies against EIIIB were proposed to be used indetection and treatment of several tumors, includingoral and mammary carcinomas.17,18
The primary structure and electromicrographssuggested that FN-III is an extended moleculecomposed of repeated sequence motifs describedas “beads on a string”.19,20 The crystal structure ofFN type III domains 7 to 10 (FN7-10) that includesthe RGD loop in domain 10 and the PHSRN synergysite in domain 9 has been published.4 The structureof FN-EIIIB alone has been solved by NMR.21 Theindividual FN type-III domains are very similar toeach other, and each domain consists of two βstrands, one of four strands (G, F, C and C′) and oneof three strands (A, B, and E).4 The domains in FN-III have a low level of sequence identity (approxi-mately 30% or lower), but they all have highlyconserved secondary structural frameworks.4,21 Themajor variable factor between each FN-III domain isthe interdomain interface, which appears to bespecific to each domain pair.4 It is of interest thatthe interdomain buried surface area between type IIIdomains 7 to 10 varies, and the maximum buriedsurface area is between domains 7 and 8.4 Similarly,
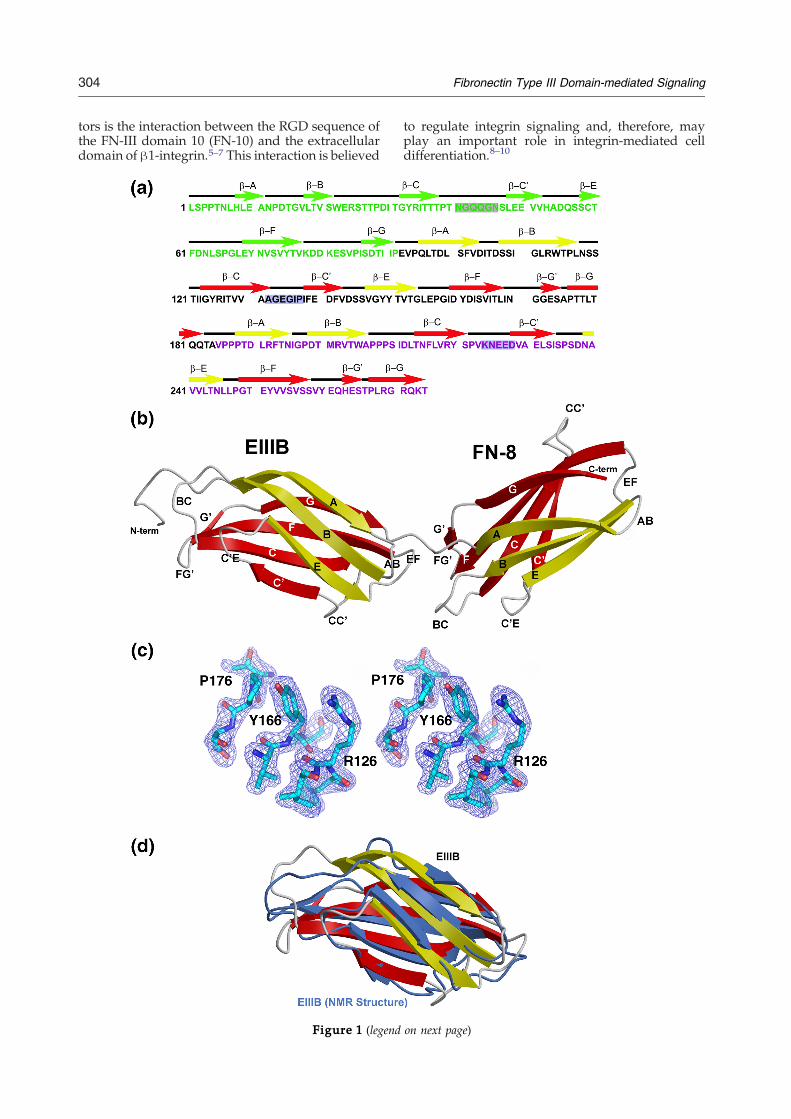
Figure 1. Crystal structure of FN-B8. (a) The primary an(green), B (black), and 8 (magenta). The sequences of CC′MOLSCRIPT diagram of the FN B-8 structure. The β sandwicannealing omit Fobs–Fcalc electron density map contouring at 3.these amino acids and a sphere of 1.0 Å around them omitted b125–127), β-F (residues 165–167) and β-G (residues 175–177) strstructure with previously solved NMR structure (RSCB Data Bfolds.
the tilt between type III domains 7 to 10 of FN varieswith the maximum tilt (52°) between domains 7 and8. There is unfortunately no general rule in theorientations between adjacent domains. EIIIB inser-tion between type III domains 7 and 8 undoubtedlyalters the interface, and possibly the overall mole-cular polarity as a consequence of domain flips. As aconsequence of the insertion of EIIIB between type IIIdomains 7 and 8, it was hypothesized that thesestructural conformational changes of fibronectinwould alter its interactions with other proteins,thereby modulating extracellular signaling cascadesin embryogenesis, wound healing, and neovascular-ization. To understand the role of EIIIB insertion andthe interface between FN type III domain extradomain B (EIIIB) and domain 8 (FN-8), we solved a2.0 Å resolution crystal structure of the FN-IIIfragment of EIIIB and FN-8. The structure reveals aunique interdomain relationship and provides struc-tural insights into possible biological roles of EIIIB.
The overall structure of FN-B8
We initially attempted to crystallize the recombi-nant fragment of FN-III domain 7 (FN-7), EIIIB, andFN-8 (the primary structure is shown in Figure 1(a)).However, the crystals were too small and failed toyield sufficient X-ray diffraction. Since the FN 7-10structure is known,4 we changed our strategy tomake two shorter constructs; FN 7-B (FN-7 andEIIIB), and FN B-8 (EIIIB and FN-8). FN 7-B grewvery small, needle-like crystals that failed to yieldsufficient X-ray diffraction. On the other hand, theFN B-8 fragment crystallized readily in 10–15%(w/v) PEG-3350, 0.15–0.2 mM diammonium citrate((NH4)2HC6H5O7)) and the crystals diffracted well.The crystal structure of FN B-8 was determined bymolecular replacement of previously solved FN-8(RSCB code 1fnf). The structure was refined to2.01 Å (Table 1) with R=24.9% and Rfree=30.1%(Figure 1(a)–(c)). We were unable to trace comple-tely the N-terminal histidine tag and several aminoacid residues thereafter. This highly flexible N-ter-minal portion may contribute to slightly increased Rvalues. However, these refinement statistic valuesare mimicking those of the 2.0 Å crystal structure oftype III fibronectin solved previously.4
FN B-8 appears to be a monomeric structure in onecrystallographic asymmetric unit. Like other crystalstructures in the fibronectin family, the overallstructure of FN B-8 is composed of two copies of βsandwich globular domains (Figure 1(b)). Each βsandwich domain is formed by an intimate anti-parallel embrace of three anti-parallel β-strands
d secondary structures of type III fibronectin domain 7loops of FN-7, FN-B and FN-8 are highlighted. (b) A
hes are shown in yellow and red. (c) The 2.01 Å simulated0 σ. The mapwas calculated via |Fobs|–|Fcalc|, φcalc, withefore annealing and map calculation. Part of β-C (residuesands of EIII-B are shown. (d) Superimposition of our EIII-Bank accession code 2fnb) demonstrates the similar overall

Table 1. Crystallographic statistics for human FN-B8structures
Resolution (Å) 50–2.0 (2.13–2.01)Space group P212121Cell constants
a (Å) 43.41b (Å) 55.29c (Å) 74.90
Total reflections 23,182Unique reflections 12,558Mean redundancy 6.2Rsym
a (%) 8.2 (48.3)Completeness (%) 99.6 (98.5)Mean I/σ 36.6 (6.0)Rcryst
b (%) 24.9Rfree
c (%) 30.1Number of protein atoms 1443Number of solvent sites 384
Values in parentheses are for the highest resolution shell. Arecombinant fragment of human type III fibronectin FN-B andFN-8 (FN-B8) was amplified by PCR and subcloned into thepET15b expression vector. The protein was expressed andpurified as described.4,25 FN-B8 was concentrated to 6 mg ml−1
in 20 mM Tris–HCl (pH 8.0), and crystallized using the sitting-drop, vapor-diffusion method at 22 °C. Crystals of 200–300 μm insize grew in 10% (w/v) PEG-3350, 0.15–0.2 mM diammoniumcitrate ((NH4)2HC6H5O7). The crystals were cryo-protected in 30–40% (v/v) glycerol plus 60–70% (v/v) mother liquor before flash-freezing in liquid nitrogen. Diffraction data were collected at theX-ray Facility at the University of North Carolina-Chapel Hill.The experiments were performed at 100 K using cryo-cooledcrystals, and were processed and reduced using DENZO,SCALEPACK, and HKL2000.26 The FN-B8 structure was deter-mined by molecular replacement (AMoRe)27 using the structureof FN-8 (RBSC code 1fn1)4 as a searchmodel. Residues 87–274 (188residues) of the 223 residues were traced. Structures were refinedusing torsion angle dynamics in CNS,28 with the maximumlikelihood function target, and included an overall anisotropic B-factor anda bulk solvent correction.About 10%of the observeddatawere set aside for cross-validation usingRfree before any refinement.Manual adjustments were performed using the program O29 andσA-weighted30 electron density maps. Simulated annealing omitand σA-weighted difference density maps were used. Finalstructures exhibit good geometry with no Ramachandran outlier.Molecular graphic Figures were created with MOLSCRIPT,31
BobScript,32 Raster3D,33 and Pymol [http://www.pymol.org].a Rsym=∑|I–<I>|/∑I, where I is the observed intensity and
<I> is the average intensity of multiple symmetry-relatedobservations of that reflection.
b Rcryst=∑||Fobs|–|Fcalc||/∑|Fobs|, where Fobs and Fcalcare the observed and calculated structure factors, respectively.
c Rfree=∑||Fobs|–|Fcalc||/∑|Fobs| for 10% of the data notused at any stage of structural refinement.
306 Fibronectin Type III Domain-mediated Signaling
(A, B, and E) on one side, and four anti-parallelβ-strands (C′, C, F, and G) on the opposite side.The general framework of EIIIB is almost identical
with the NMR structure of FN-B (RSCB code 2fnb;Figure 1(d)).21 Compared to the NMR structure,however, the crystal structure showed clearly theformation of longer anti-parallelβ-strands. The rmsdbetween the Cα positions of our crystal structure andthe NMR structure is as close as 2.5 Å. This allows usto validate our phasing information, sincewe did notuse the NMR structure for our molecular replace-ment. We believe that the molecular arrangementand packing of the crystal may contribute to the tightβ strand to β strand relationship in our structurecompared to that in the NMR structure.
The overall fold of FN-8 is similar to that of theprevious crystal structure (rmsd=4.2 Å; Figure 1(d)).The only obvious change in our FN-8 structure is theformation of an extra β-G′ strand. While thisalteration is possibly caused by crystal packing, itis clear that the additional extra domain B has littleeffect on the overall structural arrangement of FN-8.
FN-B structurally alters the macromolecularstructure of fibronectin
We further compared our EIIIB structure with thatof the previously solved FN-7. As expected, theoverall frameworks of EIIIB and FN-7 are verysimilar (rmsd=3.3 Å). The only noticeable alterationin the structural framework is that the β-E strand ofEIIIB seems to be longer and more structured thanthat of FN-7 (Figure 2(a)). While the general archi-tectures of EIIIB and FN-7 are alike, the interactionsbetween FN-7 and FN-8 are distinctively differentfrom those of EIIIB and FN-8.When we superimposed FN-8 of our structure and
FN-8 of the 1fnf structure, we discovered three dis-tinctions between the FN-7/FN-8 and FN-B/FN-8structures (Figure 2(a)). First, the bending angle be-tween EIIIB and FN-8 in our structure appears to beapproximately 63°, or about 11° more than the anglebetween FN-7 andFN-8 (previously reported as 52°).4
Secondly, the rotating angle between EIIIB and FN-8is approximately 117°, or about 5° different from thatbetween FN-7 and FN-8 (previously reported as112°).4 Finally, and perhaps most importantly, thelocations and conformations of the AB, CC′, and EFloops of FN-B are different from those of FN-7.
FN-B and FN-8 interface
The location and conformation of EF loops are themost conserved among other loops in the interface.The more acute bending of loop AB in domain Ballows the dimer to bend further, which coincideswith the rotation of domain B and the opening ofloop CC′. The interdomain linker between EIIIB andFN-8 buries 416 Å2 of surface area, while theinterdomain linker between FN-7 and FN-8 buries587 Å2 of surface area. The insertion of EIIIB clearlyopens up the buried surface over 170 Å2. The buriedinterface of EIIIB and FN-8 (416 Å2) is also less thanthat between FN-8 and FN-9 (527 Å2), but more thanthat between FN-9 and FN-10 (333 Å2). Therefore,the interface of EIIIB and FN-8 is the second smallestburied interface among the known FN-III interdo-main interfaces. The smallest FN-III buried interfacebelonging to FN-9 and FN-10 is believed to allow theRGD sequence of FN-10 and the synergy sequence(PHSRN) of FN-9 to function together in the inter-action with integrin.4,7 This alteration in the buriedinterface and the size of the FN B-8 interface suggestthat it is possible that loop CC′ between the interfaceof EIIIB and FN-8may behave like the RGD of FN-10and the synergy region of FN-9.We further examined the electrostatic potential
surface of FN B-8 (Figure 2(b)). The CC′ loop of EIIIB

Figure 2. EIIIB alters the fibronectin structure. (a) Superimposition of EIIIB-8 of FN B-8 structure (shown in yellow/red) with FN 7–8 previously solved, RSBC code 1fn1 (shown in green), demonstrates that the differences in torsion andbending angles of FN B-8 compared with the FN 7–8; and the conformational differences of the AB, EF, and CC′ loops inEIIIB and FN-7. The two orientations are related by a rotation of 180°. (b) Molecular electrostatic potential surfacerepresentation of FN B-8 is colored by charge distribution (red, negative; white, neutral; blue, positive) using programPymol [http://www.pymol.org].
307Fibronectin Type III Domain-mediated Signaling

308 Fibronectin Type III Domain-mediated Signaling
forms an acidic groove with FN-8. In addition, TheAGEGIP sequence of the CC′ loop of EIIIB is distinctfrom theNGQQGNsequence of the CC′ loop of FN-7(Figure 1(a)). An equivalent acidic groove, formed byloop CC′ and its adjacent domain was identified inanother FN-III-like structure, the FN-III domains ofthe cytoplasmic tail of integrin β4.22 The sizes ofthese two homologous acidic grooves are almost thesame, approximately 20 Å×10 Å. The acidic grooveand loop CC′were proposed to be potential protein–protein interacting sites.22 We found that the neigh-boring molecule in the crystal structure packs nicelyinto this acidic groove. The opened CC′ loop of EIIIBand the rotation and tilt of EIIIB domain allow goodaccess to the FG loop of FN-8, which is normallyhindered by the CC′ loop of FN-7 (Figure 2(a)).
Defining the potential protein-binding site
We have determined the high-resolution crystalstructure of the FN type III domains EIIIB and 8, andcompared it with the known structure of FN type IIIdomains 7 and 8. It is of interest to consider theburied interdomain interface between EIIIB and FN-8, and its comparison to that of FN-7 and FN-8. Ourcrystal structure of FN B-8 shows that the interfacebetween EIIIB and FN-8 is the second smallest of theknown FN-III structures and the CC′ loop of EIIIBforms a unique acidic groove with FN-8. The grooveis large enough to allow the neighboring molecule inthe crystal to pack. In addition, the bending anglebetween EIIIB and FN-8 in our structure appears tobe tilted about 11°more than the angle between FN-7and FN-8. The rotating angle between EIIIB and FN-8 is also approximately 5° different from that betweenFN-7 and FN-8. Given the rotation angles between thevarious type-III domains of FN (7–10) and thebiological significance attached to it (e.g. RGD andPHSRN loops in domains 10 and 9, respectively), itwas speculated that insertion of EIIIB betweendomains 7 and 8 could alter the downstream structureby causing the domains to be flipped. Our datasuggest that insertion of EIIIB may not have anydownstream effect with respect to alteration ofdomain polarity. Finally, and perhaps most signifi-cantly, the locations and conformations of loops AB,CC′, and EF of FN-B are different from those of FN-7.
Proposed potential protein-binding site
Although the expression of spliced domain EIIIB ishighly regulated during embryonic development,wound healing and tumorigenesis, there is no well-defined function for this spliced domain of FN.Interestingly, even the EIIIB-null mice fail to reveal aclear function for domain EIIIB, although embryonicfibroblasts prepared from the EIIIB–/– mice showedslower proliferation and the FN fibrils assembled bythese embryonic fibroblasts were shorter and thinnercompared to those deposited by the wild-typefibroblasts.12 These findings suggest that FN EIIIBmay play a specific role in the regulation of particularmatrix assembly and FN matrix-dependent cell
growth. It has been shown that EIIIB-containing FNproduced by cultured chondrocytes is incorporatedpreferentially in the cartilage matrix. EIIIB+FN hasbeen described in the walls of smaller blood vessels,in the smooth muscle of the gastro-intestinal andrespiratory tracts, and in cartilaginous structures.15
FN is known to utilize short-peptide surface loopsin engaging its interactions with other proteins in thesignaling pathway; for example, the RDG sequenceof FN-10,6 the PHSRN sequence of FN-9,23 and theKNEED sequence of FN-8.24 On the basis of ourcrystal structure, it is tempting to speculate that theAGEGIP sequence of loop CC′ of EIIIB may be apotential site for protein binding. However, there isno evidence at present that AGEGIP is the protein-binding site and studies are in progress to examinethis potential binding site. Preliminary analysis of celllysates showed several proteins (data not shown)binding to FN 7-EIIIB-8 and not FN 7–8 in a re-producible manner (affinity chromatography and 2Dgel analysis). Further characterization of proteinsbinding to the FN 7-EIIIB-8 is in progress.
Protein Data Bank accession number
The structural coordinate has been deposited inthe RCSB Protein Data Bank with accession number2GEE.
Acknowledgements
We thank Laurie Betts, Kent Rossman, and BrantHamel for experimental assistance. The researchwas supported by the N.I.H. DE-014394 (to I.A.), theUniversity of North Carolina-Research CouncilGrant (to S.B.), and the UNC-School of Dentistry-Faculty Creativity Fund (to S.B.).
References
1. Hynes, R. O. (1990). Fibronectins. Springer Verlag, NewYork.
2. George, E. L., Georges-Labouesse, E. N., Patel-King,R. S., Rayburn, H. & Hynes, R. O. (1993). Defects inmesoderm, neural tube and vascular development inmouse embryos lacking fibronectin. Development(Camb.), 119, 1079–1091.
3. Kornblihtt, A. R., Vibe-Pedersen, K. & Baralle, F. E.(1984). Human fibronectin: cell specific alternativemRNA splicing generates polypeptide chains differ-ing in the number of internal repeats. Nucl. Acids Res.12, 5853–5868.
4. Leahy, D. J., Aukhil, I. & Erickson, H. P. (1996). 2.0 Åcrystal structure of a four-domain segment of humanfibronectin encompassing the RGD loop and synergyregion. Cell, 84, 155–164.
5. Pierschbacher, M. D. & Ruoslahti, E. (1984). Cellattachment activity of fibronectin can be duplicated bysmall synthetic fragments of the molecule.Nature, 309,30–33.
6. Obara, M., Kang, M. S. & Yamada, K. M. (1988). Site-

309Fibronectin Type III Domain-mediated Signaling
directed mutagenesis of the cell-binding domain ofhuman fibronectin: separable, synergistic sites med-iate adhesive function. Cell, 53, 649–657.
7. Takagi, J., Strokovich, K., Springer, T. A. & Walz, T.(2003). Structure of integrin alpha5beta1 in complexwith fibronectin. EMBO J. 22, 4607–4615.
8. Hynes, R. O. (2002). Integrins: bidirectional, allostericsignaling machines. Cell, 110, 673–687.
9. Takagi, J. (2004). Structural basis for ligand recogni-tion by RGD (Arg-Gly-Asp)-dependent integrins.Biochem. Soc. Trans. 32, 403–406.
10. Hynes, R. O. (2005). The emergence of integrins: apersonal and historical perspective. Matrix Biol. 23,333–340.
11. Schwarzbauer, J. E., Patel, R. S., Fonda, D. & Hynes,R. O. (1997). Multiple sites of alternative splicing of therat fibronectin gene transcript. EMBO J. 6, 2573–2580.
12. Fukuda, T., Yoshida, N., Kataoka, Y., Manabe, R.,Mizuno-Horikawa, Y., Sato, M. et al. (2002). Micelacking the ED-B segment of fibronectin developnormally but exhibit reduced cell growth and fibro-nectin matrix assembly in vitro. Cancer Res. 62,5603–5610.
13. Astrof, S., Crowley, D., George, E. L., Fukuda, T.,Sekiguchi, K., Hanahan, D. & Hynes, R. O. (2004).Direct test of potential roles of EIIIA and EIIIBalternatively spliced segments of fibronectin in phy-siological and tumor angiogenesis. Mol. Cell Biol. 24,8662–8670.
14. Ffrench-Constant, C. & Hynes, R. O. (1989). Alter-native splicing of fibronectin is temporally andspatially regulated in the chicken embryo. Develop-ment (Camb.), 106, 375–388.
15. Peters, J. H., Chen, G. E. & Hynes, R. O. (1996).Fibronectin isoform distribution in the mouse. II.Differential distribution of the alternatively splicedEIIIB, EIIIA, and V segments in the adult mouse. CellAdhes. Commun. 4, 127–148.
16. Castellani, P., Viale, G., Dorcaratto, A., Nicolo, G.,Kaczmarek, J., Querze, G. & Zardi, L. (1994). Thefibronectin isoform containing the ED-B oncofetaldomain: a marker of angiogenesis. Int. J. Cancer, 59,612–618.
17. Kaczmarek, J., Castellani, P., Nicolo, G., Spina, B.,Allemanni, G. & Zardi, L. (1994). Distribution ofoncofetal fibronectin isoforms in normal, hyperplasticand neoplastic human breast tissues. Int. J. Cancer, 59,11–16.
18. Mandel, U., Gaggero, B., Reibel, J., Therkildsen, M. H.,Dabelsteen, E. & Clausen, H. (1994). Oncofetalfibronectins in oral carcinomas: correlation of twodifferent types. APMIS, 102, 695–702.
19. Engel, J., Odermatt, E., Engel, A., Madri, J. A.,Furthmayr, H., Rohde, H. & Timpl, R. (1981). Shapes,
domain organizations and flexibility of laminin andfibronectin, two multifunctional proteins of the extra-cellular matrix. J. Mol. Biol. 150, 97–120.
20. Erickson, H. P., Carrell, N. & McDonagh, J. (1981).Fibronectin molecule visualized in electron micro-scopy: a long, thin, flexible strand. J. Cell Biol. 91,673–678.
21. Fattorusso, R., Pellecchia, M., Viti, F., Neri, P., Neri, D.& Wuthrich, K. (1999). NMR structure of the humanoncofoetal fibronectin ED-B domain, a specific markerfor angiogenesis. Structure, 7, 381–390.
22. De Pereda, J. M., Wiche, G. & Liddington, R. C. (1999).Crystal structure of a tandem pair of fibronectin typeIII domains from the cytoplasmic tail of integrinalpha6beta4. EMBO J. 18, 4087–4095.
23. Aota, S., Nomizu, M. & Yamada, K. M. (1994). Theshort amino acid sequence Pro-His-Ser-Arg-Asn inhuman fibronectin enhances cell-adhesive function.J. Biol. Chem. 269, 24756–24761.
24. Wong, J. Y., Weng, Z., Moll, S., Kim, S. & Brown, C. T.(2002). Identification and validation of a novel cell-recognition site (KNEED) on the 8th type III domain offibronectin. Biomaterials, 23, 3865–3870.
25. Aukhil, I., Joshi, P., Yan, Y. & Erickson, H. P. (1993).Cell- and heparin-binding domains of the hexabra-chion arm identified by tenascin expression proteins.J. Biol. Chem. 268, 2542–2553.
26. Otwinowski, Z. &Minor, W. (1993).Data Collection andProcessing. Daresbury Laboratories, Warrington, UK.
27. Navaza, J. (2001). Implementation of molecularreplacement in AMoRe. Acta Crystallog. sect. D, 57,1367–1372.
28. Brunger, A. T., Adams, P. D., Clore, G. M., DeLano,W. L., Gros, P., Grosse-Kunstleve, R. W. et al. (1998).Crystallography and NMR system: a new softwaresuite for macromolecular structure determination.Acta Crystallog. sect. D, 54, 905–921.
29. Jones, T. A., Zou, J. Y., Cowan, S. W. & Kjeldgaard, M.(1991). Improved methods for building protein modelsin electron density maps and the location of errors inthese models. Acta Crystallog. sect. A, 47, 110–119.
30. Read, R. J. (1986). Improved Fourier coefficients formaps using phases from partial structures with errors.Acta Crystallog. sect. A, 42, 140–149.
31. Kraulis, P. J. (1991). MOLSCRIPT: a program toproduce both detailed and schematic plots of proteinstructures. J. Appl. Crystallog. 24, 946–950.
32. Esnouf, R. M. (1999). Further additions to MolScriptversion 1. 4, including reading and contouring ofelectron-density maps. Acta Crystallog. sect. D, 55,938–940.
33. Merritt, E. A. & Murphy, M. E. P. (1991). Raster3Dversion 2. 0 – a program for photorealistic moleculargraphics. Acta Crystallog. sect. D, 50, 869–873.
Edited by R. Huber
(Received 2 June 2006; received in revised form 27 September 2006; accepted 5 October 2006)Available online 11 October 2006





























