Embed Size (px)
Citation preview


!!!
Subolesin/Akirin:/Expression/of/ recombinant/ antigens/
and/characterization/of/different/formulations/for/the/
control/of/hematophagous/ arthropod/ vectors/!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!
Juan/Antonio/Moreno?Cid/Mora//
Doctoral/Thesis/


Subolesin/Akirin:!Expression!of!recombinant!antigens!and!
characterization!of!different!formulations!for!the!control!of!
hematophagous!arthropod!vectors!!!!!!
Akirina/Subolesina:!Expresión!del!antígeno!recombinante!y!
ensayo!de!diferentes!formulaciones!para!el!control!de!las!
infestaciones!por!ectoparásitos!hematófagos!!!!!!
Trabajo!de!investigación! desarrollado!por!el!ingeniero!Juan!Antonio!MorenoECid!Mora!
para!optar!al!grado!de!Doctor!por!la!Universidad!de!CastillaELa!Mancha!!!!!!!!DEPARTAMENTO:!
!!Ciencia!y!Tecnología!Agroforestal!y!Genética!
CENTRO:!
Instituto!de!Investigación!en!Recursos!Cinegéticos,!IREC.!!!Grupo!de!Sanidad!y!Biotecnología,!SaBio!(Health!&!Biotechnology)!
PROGRAMA!DE!DOCTORADO:!
Investigación!Básica!y!Aplicada!a!Recursos!Cinegéticos!!!!!!DIRECTOR:!
!!Dr.!José!de!la!Fuente!García!

!!!!!!!!!!!
Vº!Bº!del!Director!!!!!!!!!!!!
Fdo.!Dr.!José!de!la!Fuente!García!!!!!!!!!!!!!
Universidad!de!CastillaELa!Mancha!!!!!
Instituto!de!Investigación!en!Recursos!Cinegéticos,!IREC!
(CSICEUCLMEJCCM)!

!!!!!!!
La!realización!de!este!trabajo!de!investigación!ha!sido!posible!gracias!a!
la!financiación!del!proyecto:!!!!!“Tecnología! para! la! producción! de! proteínas! recombinantes! mediante!
fermentación! extractiva! en! un! sistema! de! dos! fases! acuosas”,! Nº! Expediente!
PEII09E0118E8907,! de! la! Consejerıa! de! Educacio! n! y! Ciencia,! Junta! de!Comunidades! de! Castilla! la! Mancha! (JCCM),! cofinanciado! con! Fondos!
Europeos!en!el!marco!del!Programa!de!Potenciación!de!Recursos!Humanos!del!
Plan!Regional! de! Investigación! Científica,! Desarrollo! Tecnológico! e! Innovación!
2005E!2010!(PRINCET).!!!
!
Consejería de Educación, Cultura y DeportesDirección General de Universidades, Investigación e Innovación
Página 4 de 13
ANEXO II INFORME DEL DIRECTOR DE INVESTIGACIÓNIP: DE LA FUENTE GARCÍA, JOSÉ DE JESÚSNº de expediente del proyecto: PEII09-0118-8907Nombre candidato: MORENO-CID MORA, JUAN ANTONIO
Informe del Director de Investigación sobre el trabajo realizado por el solicitante en los últimos 12 meses.(añadir las hojas necesarias)
Durante este periodo el doctorando desarrolló diferentes actividades de investigación dentro del
proyecto de tesis y completó los cursos de formación requeridos, incluyendo la culminación del
Máster Universitario. Los resultados alcanzados hasta el momento han avanzado nuestro
conocimiento sobre los procesos y la fisiología de Pichia pastoris para la expresión de proteínas
recombinantes. Estos avances son básicos para el desarrollo de procesos biotecnológicos más
eficientes y rentables utilizando este microorganismo, lo cual constituye uno de los objetivos de
trabajo de nuestro grupo. El doctorando mostró una gran capacidad y dedicación al trabajo y
realizó aportaciones esenciales para el desarrollo de los experimentos, contribuyendo a varios
de los proyectos que se desarrollan en el grupo.
Estos resultados se reflejarón en 7 publicaciones (5 publicadas y dos en proceso de revisión) en
revistas de alto impacto en las que el doctorando es primer autor o coautor de las mismas.
Además el doctorando completó una estancia de tres meses en la Universidad de Iowa, EEUU,
donde incorporó conocimientos y técnicas esenciales para el desarrollo del trabajo del grupo.
Por todo esto valoro muy positivamente el trabajo realizado por el doctorando durante este
periodo.


!!!!!!!!!!!!!!!!!
Para Eva, Celia y venideros

La ciencia tiene las raíces amargas, pero muy dulces frutos.
Aristóteles


&217(1767+(6,6�6758&785(����������������������������������������������������������������������������������������������������������������
&+$37(5�,��,1752'8&7,21��������������������������������������������������������������������������������������������������
7DUJHWLQJ� DUWKURSRG� VXEROHVLQ�DNLULQ� IRU� WKH� GHYHORSPHQW� RI� D� XQLYHUVDO�YDFFLQH�IRU
FRQWURO�RI�YHFWRU�LQIHVWDWLRQV�DQG�SDWKRJHQ�WUDQVPLVVLRQ������������������������������������������������������������������������
+<327+(6,6�$1'�$,06��������������������������������������������������������������������������������������������������������
&+$37(5�,,��(;35(66,21��352'8&7,21���������������������������������������������������������������������
3URGXFWLRQ�RI�UHFRPELQDQW�$HGHV�DOERSLFWXV�DNLULQ�LQ�3LFKLD�SDVWRULV�XVLQJ�DQ�DTXHRXV
WZR�SKDVH�VHPLFRQWLQXRXV�IHUPHQWDWLRQ�SURFHVV���������������������������������������������������������������������������������������
&+$37(5�,,,���$17,*(1�&+$5$&7(5,=$7,21������������������������������������������������������������
&KDUDFWHUL]DWLRQ� RI� $HGHV� DOERSLFWXV� DNLULQ� IRU� WKH� FRQWURO� RI� PRVTXLWR� DQG�VDQG�IO\
LQIHVWDWLRQV�����������������������������������������������������������������������������������������������������������������������������������������������������������
%DFWHULDO�PHPEUDQHV�HQKDQFH� WKH� LPPXQRJHQLFLW\�RI� WKH� VXUIDFH� H[SRVHG�WLFN�6XEROHVLQ�
$QDSODVPD�PDUJLQDOH�063�D�FKLPHULF�DQWLJHQ������������������������������������������������������������������������
&+$37(5�,9��9$&&,1$7,21�����������������������������������������������������������������������������������������������
&RQWURO� RI� PXOWLSOH� DUWKURSRG� YHFWRU� LQIHVWDWLRQV� ZLWK� VXEROHVLQ�DNLULQ�YDFFLQHV�����
&RQWURO� RI� WLFN� LQIHVWDWLRQV� LQ� FDWWOH� YDFFLQDWHG� ZLWK� EDFWHULDO� PHPEUDQHV�FRQWDLQLQJ�����������������
VXUIDFH�H[SRVHG�WLFN�SURWHFWLYH�DQWLJHQV���������������������������������������������������������������������������������������������������
&+$37(5�9��',6&866,21�����������������������������������������������������������������������������������������������������
6XEROHVLQ�$NLULQ�YDFFLQHV�IRU�WKH�FRQWURO�RI�DUWKURSRG�YHFWRUV�DQG�YHFWRU�ERUQH�SDWKRJHQV��������
&21&/86,216������������������������������������������������������������������������������������������������������������������������6�
6800$5<��������������������������������������������������������������������������������������������������������������������������������9

2!

3
THESIS&STRUCTURE&
The main objective of this work was to establish and develop technologies for the
production and characterization of SUB/AKR recombinant antigens using fermentations
with different microorganisms, and to evaluate their effect as protective antigens against
hematophagous ectoparasites by conducting vaccination trials. Akirin (AKR) and
Subolesin (SUB) are evolutionary conserved ortholog proteins that affect the
expression of signal transduction and innate immune response genes in vertebrates and
invertebrates, thus providing the opportunity to develop a universal vaccine for the
control of multiple arthropod infestations and their associated pathogens. In Chapter I, a
briefly review is given on the use of the Aedes albopictus AKR and the tick protective
antigen SUB for the development of vaccines for the control of arthropod vectors.
Expression, production, development of technologies and characterization were
established in the first part of this thesis. The development of novel technologies were
established (a) in Chapter II by the expression of recombinant AKR using the yeast
Pichia pastoris conducted by a semicontinuous extractive bioconversion in an aqueous
two-phase system (ATPS), and (b) in the Chapter III by a novel, simple and cost-
effective approach for the production of tick protective antigens by surface display
of the antigenic protein chimera SUB-MSP1a expressed on the Escherichia coli
membrane. Evaluation of the protective capacity of the antigens against vectors,
vaccination and infestation trials are described in the second part of this work.
Chapter IV details several experiments conducted to evaluate the effect of recombinant
SUB/AKR: (a) to test its ability to control multiple arthropod vectors infestation and the
effect of chimeric constructions combining protective epitopes from the mosquito AKR
and the tick SUB as a potential universal vaccine, mice were vaccinated with SUB, AKR
and the chimeric constructions (Q38 and Q41) then challenged with A. albopictus,
Phlebotomus perniciosus and Ixodes ricinus; and (b) to evaluate the efficacy of the
chimeric system SUB-MSP1a a vaccination trial was conducted in cattle infested with the
ticks Rhipicephalus microplus and Rhipicephalus annulatus.
In Chapter V vaccination trials for the control of arthropod vectors and vector borne
pathogens with vaccines based in SUB/AKR are described and discussed.
Finally, overall conclusions and a summary of this research are detailed.

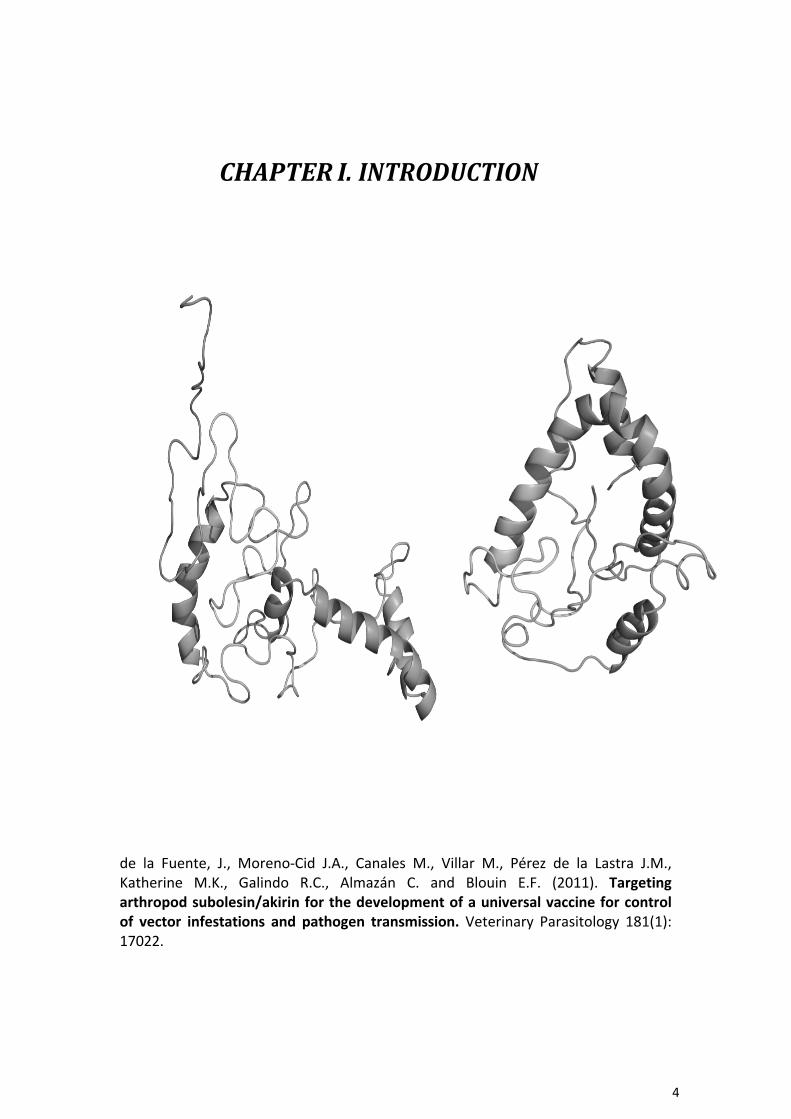
4!
CHAPTER&I.&INTRODUCTION&
de! la! Fuente,! J.,! Moreno3Cid! J.A.,! Canales! M.,! Villar! M.,! Pérez! de! la! Lastra! J.M.,!Katherine! M.K.,! Galindo! R.C.,! Almazán! C.! and! Blouin! E.F.! (2011).! Targeting)arthropod)subolesin/akirin)for)the)development)of)a)universal)vaccine)for)control)of) vector) infestations) and) pathogen) transmission.! Veterinary! Parasitology! 181(1):!17022.!


TARGETING'ARTHROPOD'SUBOLESIN/AKIRIN'FOR'
THE' DEVELOPMENT' OF' A' UNIVERSAL' VACCINE'
FOR' CONTROL' OF' VECTOR' INFESTATIONS' AND'
PATHOGEN'TRANSMISSION'

6!
&KDSWHU�,
Contents lists available at ScienceDirect
Veterinary Parasitology
jo u rn a l hom epa ge : www.elsevier .com/locate /vetpar
Targeting arthropod subolesin/akirin for the development of a universal vaccine for control of vector infestations and pathogen transmission
José de la Fuente a,b,∗", Juan A. Moreno-Cid a , Mario Canales a , Margarita Villar a , José M. Pérez de la Lastra a , Katherine M. Kocan b , Ruth C. Galindo a , Consuelo Almazán c , Edmour F. Blouin ba Instituto de Investigación en Recursos Cinegéticos IREC (CSIC-UCLM-JCCM), Ronda de Toledo s/n, 13005 Ciudad Real, Spain b Department of Veterinary Pathobiology, Center for Veterinary Health Sciences, Oklahoma State University, Stillwater, OK 74078, USA c Facultad de Medicina Veterinaria y Zootecnia, Universidad Autónoma de Tamaulipas, Km. 5 carretera Victoria-Mante, CP 87000 Ciudad Victoria, Tamaulipas, Mexico
a r t i c l e i n f o a b s t r a c t
Keywords: Tick Mosquito Sandfly Mite Vaccine RNA interference Akirin Subolesin
Diseases caused by arthropod-borne pathogens greatly impact on human and animal health. Recent research has provided evidence that tick protective antigens can be used for devel- opment of vaccines with the dual target of controlling arthropod infestations and reducing their vector capacity for pathogens. As reviewed herein, protective antigens such as sub- olesin/akirin, which are highly conserved across vector species, show promise for use in development of a universal vaccine for the control of arthropod infestations and the reduc- tion of pathogen transmission. However, further research is needed in critical areas towards achieving this goal.
© 2011 Elsevier B.V. All rights reserved.
1. Introduction
Diseases caused by arthropod-borne pathogens greatly impact on human and animal health, accounting for over 20% of all emerging infectious diseases recorded between 1940 and 2004 (Jones et al., 2008). In particular, insects, such as mosquitoes and sand flies, are considered to be the most important vectors of human diseases worldwide (Halstead, 2007; Chappuis et al., 2007; Jones et al., 2008), while ticks are considered to be second worldwide to mosquitoes as vectors of pathogens that cause human dis- eases. In addition, ticks are the most important vectors of
∗" Corresponding author at: Instituto de Investigación en Recursos Cinegéticos IREC (CSIC-UCLM-JCCM), Ronda de Toledo s/n, 13005 Ciudad Real, Spain.
E-mail addresses: jose [email protected], [email protected] (J. de la Fuente).
pathogens that cause disease in cattle (Peter et al., 2005; de la Fuente et al., 2008a).
With the exception of a few diseases such as yel- low fever, vaccines against vector-transmitted pathogens have not been successfully developed nor implemented. Furthermore, the intense use of insecticides and/or chemotherapy has resulted in an increasing number of insecticide-resistant vectors and drug-resistant pathogens (de la Fuente and Kocan, 2003; Kishore et al., 2006; Speranc a and Capurro, 2007).
The efficacy of tick vaccines for the reduction of tick infestations and prevention of the transmission of some tick-borne pathogens (de la Fuente et al., 1998, 2007a,b; de la Fuente and Kocan, 2003; Willadsen, 2004; Rodríguez Valle et al., 2004) and preliminary results obtained in insect vector species (Valenzuela et al., 2001; Lal et al., 2001; Almeida and Billingsley, 2002; Suneja et al., 2003; Milleron et al., 2004; Titus et al., 2006; Kedzierski et al., 2006; Saul, 2007; Canales et al., 2009a) have provided evidence that
0304-4017/$ – see front matter © 2011 Elsevier B.V. All rights reserved. doi:10.1016/j.vetpar.2011.04.018

&KDSWHU�,
7!
protective antigens could be used for the development of vaccines with the dual target of controlling arthropod infestations and reducing the vector capacity for pathogens which impact on human and animal health.
Particularly relevant towards this goal is the identi- fication of protective antigens such as cement proteins (Trimnell et al., 2005) and subolesin/akirin (de la Fuente et al., 2006b) that are conserved across vector species, thus providing the opportunity to develop a universal vac- cine for the control of multiple arthropod infestations and their associated pathogens. Thus, in principle, a universal vaccine would bypass the need to develop a vaccine for individual pathogens and vectors, and these vaccines could also be combined with pathogen-specific antigens and other control measures such as sterile-arthropod methods (Alphey et al., 2010) for a more effective overall control of vector-borne diseases.
Herein, we review recent research on subolesin/akirin and the possibilities of using these antigens for the devel- opment of a universal vaccine for the control of multiple arthropod infestations and the reduction of their vector capacity to transmit pathogens that impact on human and animal health.
2. Evolution and function of arthropodsubolesin/akirin
Akirins constitute a recently renamed group of evo- lutionarily conserved proteins in insects and vertebrates (de la Fuente et al., 2006b; Goto et al., 2008; Macqueen and Johnston, 2009; Mangold et al., 2009). Subolesin, the ortholog of akirin in ticks, was discovered as a tick protec- tive antigen in Ixodes scapularis (Almazán et al., 2003). Most vertebrates have two closely related akirin homologues, akirin1 and akirin2 (Goto et al., 2008). However, up to 8 akirin family members have been described in some verte- brates (Macqueen et al., 2010). In insects and ticks, only one subolesin/akirin gene has been identified which is evolu- tionarily and functionally closer to mammalian akirin2 (de la Fuente et al., 2006b; Goto et al., 2008; Macqueen and Johnston, 2009; Galindo et al, 2009; Mangold et al., 2009).
The proposed function of akinrins is as transcription factors required for NF-kB-dependent gene expression in fruit flies and mice (Goto et al., 2008; Galindo et al., 2009), and in the regulation of the innate immune response in fruit flies (Goto et al., 2008). Additionally, metazoan akirins have been shown to regulate myogenesis and carcinogen- esis (Gonzalez and Baylies, 2005; Marshall et al., 2008; Komiya et al., 2008; Salerno et al., 2009). In ticks, sub- olesin was proposed to be involved in the regulation of NF-kB-dependent and independent gene expression (de la Fuente et al., 2008b; Galindo et al., 2009). Subolesin/akirin regulate gene expression through interactions with inter- mediate regulatory proteins such as GI, GII, 14-3-3beta and other as yet unidentified proteins (Komiya et al., 2008; de la Fuente et al., 2008b; Macqueen et al., 2010). These inter- mediate proteins interact with NF-kB, bind DNA or remodel chromatin to regulate gene expression (Goto et al., 2008; de la Fuente et al., 2008b, 2010a; Macqueen et al., 2010). Interestingly, the results of RNA interference experiments in the tick, I. scapularis, suggested that NF-kB participates
in the transcription of subolesin while subolesin may be involved in the regulation of NF-kB expression, thus sug- gesting a possible function for subolesin in self regulation (Galindo et al., 2009). Additionally, subolesin is differ- entially expressed and involved in tick innate immunity in response to pathogen infection (Zivkovic et al., 2010; de la Fuente et al., 2010a). This broad function of sub- olesin/akirin as transcription factors explains the profound effect of its silencing on organism physiology and devel- opment (Maeda et al., 2001; de la Fuente et al., 2006a,b, 2008b; Nijhof et al., 2007; Kocan et al., 2007, 2009; Goto et al., 2008; Galindo et al., 2009; Canales et al., 2009a).
3. Effect of vaccination with subolesin/akirin onarthropod vector infestations and pathogentransmission
Immunization trials using recombinant subolesin pro- tected hosts against tick infestations by reducing tick survival, weight and oviposition (Table 1) and decreased the vector capacity of I. scapularis ticks for Anaplasma phagocytophilum (Almazán et al., 2005a,b, 2010; de la Fuente et al., 2006c, 2010b; Canales et al., 2009a).
Recent results revealed a reduction in the survival and/or fertility of mosquitoes (Anopheles atroparvus, Aedes caspius and Culex pipiens), sand flies (Phlebotomus perni- ciosus) and poultry red mites (Dermanyssus gallinae) after being fed in vitro with antibodies against the recombinant Aedes albopictus akirin (Canales et al., 2009a; Harrington et al., 2009). Vaccination with recombinant A. albopictus akirin also reduced tick infestations (Canales et al., 2009a,b) and the oviposition by A. albopictus mosquitoes and sand flies, P. perniciosus (Moreno-Cid et al., 2011).
As demonstrated previously for Bm86-based vaccines (de la Fuente et al., 1998, 2007a,b; de la Fuente and Kocan, 2003; Willadsen, 2004; Rodríguez Valle et al., 2004), these results suggested that vaccination with subolesin/akirin may reduce the transmission of vector-borne pathogens by decreasing arthropod vector populations, as well as by affecting vector capacity for pathogens.
4. Putative mechanism of subolesin/akirinvaccine protection
Immunization with intracellular proteins such as subolesin/akirin, cytoplasmic nucleotidase, interfase cyto- plasmic foci protein 45, heat shock and ribosomal proteins has been effective against ticks and other invertebrate organisms (Elad and Segal, 1995; Almazán et al., 2003, 2005a,b, 2010; de la Fuente et al., 2010b; Canales et al., 2009a,b; Harrington et al., 2009). The protection elicited by anti-tick vaccines (de la Fuente et al., 1998; Willadsen, 2004; Trimnell et al., 2005), and likely by other anti- arthropod vaccines (Tellam et al., 2001; Lal et al., 2001; Milleron et al., 2004), is based on the production of anti- bodies in vaccinated hosts that interact with and affect the function of the target antigen in feeding arthropods. Host antibodies may interact with arthropod intracellu- lar proteins through a two-step process: first, antibodies are specifically transported across the gut barrier into the hemolymph and then they should enter into arthro-
&KDSWHU�,
8!
Table 1 Inhibition of tick infestations and vaccine efficacy in subolesin-vaccinated hosts.
Tick species Stage Inhibition of tick infestations (%)a
Vaccine efficacy against adult ticks (%)b
References
Ixodes scapularis Larvae 61 – Almazán et al. (2005a,b), Nymphs Adults
35 10–58
– 71
Canales et al. (2009a)
Amblyomma americanum Nymphs 12–17 – de la Fuente et al. Adults 7 66 (2010b) Adults 43 51 Almazán et al. (2010) Adults 18 60 Almazán et al. (2010)
Rhipicephalus microplus R. annulatus Dermacentor variabilis Nymphs 22 – Almazán et al. (2005a) a The percent inhibition of tick infestation was calculated with respect to the saline/adjuvant control group. b Vaccine efficacy was calculated considering the reduction in the number of adult female ticks, oviposition and egg fertility as compared to the control
group as reported previously (de la Fuente et al., 2010b).
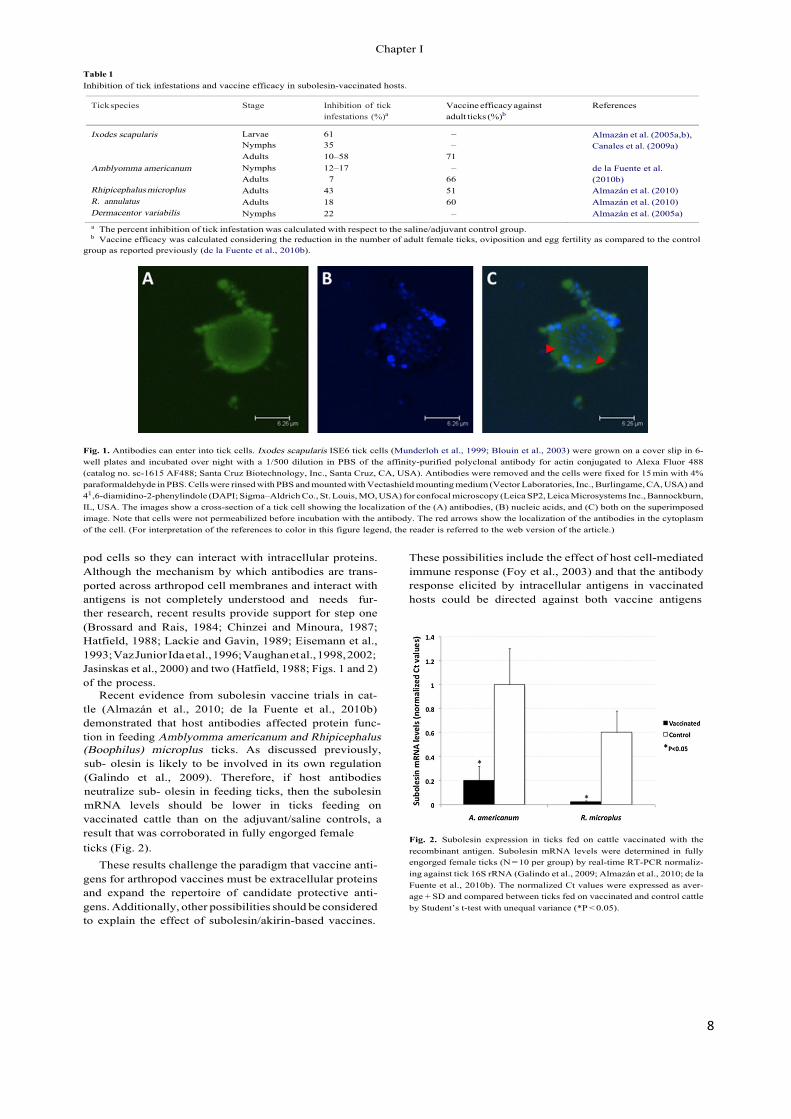
Fig. 1. Antibodies can enter into tick cells. Ixodes scapularis ISE6 tick cells (Munderloh et al., 1999; Blouin et al., 2003) were grown on a cover slip in 6-well plates and incubated over night with a 1/500 dilution in PBS of the affinity-purified polyclonal antibody for actin conjugated to Alexa Fluor 488 (catalog no. sc-1615 AF488; Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA). Antibodies were removed and the cells were fixed for 15 min with 4% paraformaldehyde in PBS. Cells were rinsed with PBS and mounted with Vectashield mounting medium (Vector Laboratories, Inc., Burlingame, CA, USA) and 41",6-diamidino-2-phenylindole (DAPI; Sigma–Aldrich Co., St. Louis, MO, USA) for confocal microscopy (Leica SP2, Leica Microsystems Inc., Bannockburn, IL, USA. The images show a cross-section of a tick cell showing the localization of the (A) antibodies, (B) nucleic acids, and (C) both on the superimposed image. Note that cells were not permeabilized before incubation with the antibody. The red arrows show the localization of the antibodies in the cytoplasm of the cell. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of the article.)
pod cells so they can interact with intracellular proteins. Although the mechanism by which antibodies are trans- ported across arthropod cell membranes and interact with antigens is not completely understood and needs fur- ther research, recent results provide support for step one (Brossard and Rais, 1984; Chinzei and Minoura, 1987; Hatfield, 1988; Lackie and Gavin, 1989; Eisemann et al., 1993; Vaz Junior Ida et al., 1996; Vaughan et al., 1998, 2002; Jasinskas et al., 2000) and two (Hatfield, 1988; Figs. 1 and 2) of the process.
Recent evidence from subolesin vaccine trials in cat- tle (Almazán et al., 2010; de la Fuente et al., 2010b) demonstrated that host antibodies affected protein func- tion in feeding Amblyomma americanum and Rhipicephalus (Boophilus) microplus ticks. As discussed previously, sub- olesin is likely to be involved in its own regulation (Galindo et al., 2009). Therefore, if host antibodies neutralize sub- olesin in feeding ticks, then the subolesin mRNA levels should be lower in ticks feeding on vaccinated cattle than on the adjuvant/saline controls, a result that was corroborated in fully engorged female ticks (Fig. 2).
These results challenge the paradigm that vaccine anti- gens for arthropod vaccines must be extracellular proteins and expand the repertoire of candidate protective anti- gens. Additionally, other possibilities should be considered to explain the effect of subolesin/akirin-based vaccines.
These possibilities include the effect of host cell-mediated immune response (Foy et al., 2003) and that the antibody response elicited by intracellular antigens in vaccinated hosts could be directed against both vaccine antigens
Fig. 2. Subolesin expression in ticks fed on cattle vaccinated with the recombinant antigen. Subolesin mRNA levels were determined in fully engorged female ticks (N = 10 per group) by real-time RT-PCR normaliz- ing against tick 16S rRNA (Galindo et al., 2009; Almazán et al., 2010; de la Fuente et al., 2010b). The normalized Ct values were expressed as aver- age + SD and compared between ticks fed on vaccinated and control cattle by Student’s t-test with unequal variance (*P < 0.05).

&KDSWHU�,
9!
and cross-reactive epitopes in other arthropod proteins (Trimnell et al., 2005).
5. Towards development of a universal vaccinefor the control of vector infestations andpathogen transmission
Analysis of subolesin/akirin ortholog sequences in mosquitoes and ticks revealed a high degree of sequence conservation among these organisms and suggested the presence of conserved antigenic epitopes (Canales et al., 2009a). Immune cross-reactivity between I. scapularis and A. albopictus subolesin/akirin ortholog proteins sug- gested that common antigenic epitopes might be used to elicit a protective response in immunized hosts (Canales et al., 2009a). Linear B-cell epitopes, con- formational epitopes, and conformational discontinuous epitopes were identified in I. scapularis and A. albopic- tus subolesin/akirin ortholog proteins by combining the results of peptide and phage display libraries scan anal- ysis with sera from vaccinated and protected animals and computational modeling (Prudencio et al., 2010). The determination of conserved protective epitopes in subolesin/akirin orthologs may lead to the development of a chimeric-epitope universal vaccine directed at the control of both arthropod infestations and reduction of their vector capacity. Although positive results have been obtained with epitope-based vaccines (Szalai et al., 2008), possible drawbacks of vaccinating with chimeric- epitope vaccines should be considered and include the induction of antibodies with specificities different from those induced by the native protein and the resultant antibody, because it is oligoclonal, can more read- ily select for escape mutants than polyvalent serum, although this may not be a problem if mutation is con- strained by functional requirements of the protein (Caoili, 2010).
As discussed previously (Canales et al., 2009a), the con- servation of subolesin/akirin sequences among arthropod vectors and vertebrate hosts raises the question of safety when using subolesin/akirin for immunization because of the potential of inducing autoimmune responses that may be damaging to the host. However, it is expected that the antibody response would be primarily directed against non-self epitopes thus reducing the possibility of detrimental effects to the host. Additionally, immu- nization of vertebrates with intracellular proteins such as akirin/subolesin has been effective against ticks and other invertebrate organisms without inducing autoim- mune responses in vertebrate hosts (Elad and Segal, 1995; Almazán et al., 2003, 2005a,b, 2010; de la Fuente et al., 2010b; Canales et al., 2009a,b; Harrington et al., 2009).
Finally, although less advanced than other research areas, development of cost-effective processes to produce recombinant antigens is essential for the successful pro- duction of arthropod vaccines (Canales et al., 2010). This research should be directed towards reducing production costs while increasing yield and efficacy of vaccine antigens (Canales et al., 2009b,c, 2010).
6. Conclusions and future directions
The results discussed herein provide evidence for the use of subolesin/akirin orthologs for the control of acarine and insect infestations and the reduction of their vector capacity. However, the development of a subolesin/akirin-based universal vaccine to control mul- tiple arthropod vector species and the transmission of pathogens is still in its infancy. The efficacy obtained so far with subolesin/akirin-based vaccines in the con- trol of vector infestations and pathogen infection needs to be improved through the use of antigen combinations and enhanced immunogenicity of vaccine formulations for a more effective overall control of vector-borne dis- eases. Future experiments may include, but should not be limited to: (a) the study of subolesin/akirin sequence divergence among individuals within and between geo- graphical regions and between species, (b) characterization of subolesin/akirin regulation and function in different organisms, (c) cloning of subolesin/akirin orthologs in other vector species of public and veterinary health impor- tance such as fleas, lice, black flies, tsetse flies and triatomine bugs, (d) development of improved vaccine for- mulations through antigen combinations and enhanced immunogenicity, (e) characterization of subolesin/akirin recombinant proteins, chimeric-epitope antigens and anti- gen combinations for the control of multiple arthropod species and their associated pathogens, (f) development of cost-effective processes and formulations for vaccine pro- duction, and (g) design of effective strategies for vaccine administration to vector-borne pathogen reservoir hosts and animal populations at risk in order to reduce arthro- pod infestations and vector capacities. Collectively, these advances will likely contribute to the control of vector infestations and pathogen transmission to vertebrate hosts.
Conflict of interest statement
None declared.
Acknowledgements
We thank Ruchira Mitra (OSU) for performing the exper- iments to visualize antibodies inside tick cells. We thank other members of our laboratories for fruitful discussions and technical assistance. This work was supported by the Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (INIA), Spain (project FAU2008-00014-00-00) and the Consejería de Educación y Ciencia, JCCM, Spain (project PEII09-0118-8907). J.A. Moreno-Cid is a recipient of a JCCM fellowship. M. Canales was funded by the Well- come Trust under the “Animal Health in the Developing World” initiative (project 0757990). M. Villar and Ruth C. Galindo were funded by the JAE-DOC program (CSIC-FSE) and MEC, Spain, respectively.
References
Almazán, C., Kocan, K.M., Bergman, D.K., Garcia-Garcia, J.C., Blouin, E.F., de la Fuente, J., 2003. Identification of protective antigens for the con- trol of Ixodes scapularis infestations using cDNA expression library immunization. Vaccine 21, 1492–1501.

&KDSWHU�,
10!
Almazán, C., Blas-Machado, U., Kocan, K.M., Yoshioka, J.H., Blouin, E.F., Mangold, A.J., de la Fuente, J., 2005a. Characterization of three Ixodes scapularis cDNAs protective against tick infestations. Vaccine 23, 4403–4416.
Almazán, C., Kocan, K.M., Blouin, E.F., de la Fuente, J., 2005b. Vaccination with recombinant tick antigens for the control of Ixodes scapularis adult infestations. Vaccine 23, 5294–5298.
Almazán, C., Lagunes, R., Villar, M., Canales, M., Rosario-Cruz, R., Jongejan, F., de la Fuente, J., 2010. Identification and characterization of Rhipi- cephalus (Boophilus) microplus candidate protective antigens for the control of cattle tick infestations. Parasitol. Res. 106, 471–479.
Almeida, A.P., Billingsley, P.F., 2002. Induced immunity against the mosquito Anopheles stephensi (Diptera: Culicidae): effects of cell fraction antigens on survival, fecundity, and Plasmodium berghei (Eucoccidiida: Plasmodiidae) transmission. J. Med. Entomol. 39, 207–214.
Alphey, L., Benedict, M., Bellini, R., Clark, G.G., Dame, D.A., Service, M.W., Dobson, S.L., 2010. Sterile-insect methods for control of mosquito- borne diseases: an analusis. Vector-Borne Zoonotic Dis. 10, 295–311.
Blouin, E.F., Saliki, J.T., de la Fuente, J., Garcia-Garcia, J.C., Kocan, K.M., 2003. Antibodies to Anaplasma marginale major surface proteins 1a and 1b inhibit infectivity for cultured tick cells. Vet. Parasitol. 111, 247–260.
Brossard, M., Rais, O., 1984. Passage of hemolysins through the midgut epithelium of female Ixodes ricinus L. fed on rabbits infested or rein- fested with ticks. Experientia 40, 561–563.
Canales, M., Naranjo, V., Almazán, C., Molina, R., Tsuruta, S.A., Szabó, M.P.J., Manzano-Roman, R., Pérez de la Lastra, J.M., Kocan, K.M., Jiménez, M.I., Lucientes, J., Villar, M., de la Fuente, J., 2009a. Conservation and immunogenicity of the mosquito ortholog of the tick protective anti- gen, subolesin. Parasitol. Res. 105, 97–111.
Canales, M., Labruna, M.B., Soares, J.F., Prudencio, C.R., de la Fuente, J., 2009b. Protective efficacy of bacterial membranes containing surface-exposed BM95 antigenic peptides for the control of cattle tick infestations. Vaccine 27, 7244–7248.
Canales, M., Ballesteros, C., Moreno-Cid, J.A., Espinosa, A.M., Villar, M., de la Fuente, J., 2009c. Extractive bioconversion to produce the Aedes albopictus akirin in an aqueous two-phase system support- ing Pichia pastoris growth and protein secretion. Biochem. Eng. J. 46, 105–114.
Canales, M., Moreno-Cid, J.A., Almazán, C., Villar, M., de la Fuente, J., 2010. Bioprocess design and economics ofrecombinant BM86/BM95 antigen production for anti-tick vaccines. Biochemical Engineering Journal 52, 79–90.
Caoili, S.E., 2010. Benchmarking B-cell epitope prediction for the design of peptide-based vaccines: problems and prospects. J. Biomed. Biotech- nol. 2010, 910524.
Chappuis, F., Sundar, S., Hailu, A., Ghalib, H., Rijal, S., Peeling, R.W., Alvar, J., Boelaert, M., 2007. Visceral leishmaniasis: what are the needs for diagnosis, treatment and control? Nat. Rev. Microbiol. 5, 873–882.
Chinzei, Y., Minoura, H., 1987. Host immunoglobulin G titre and antibody activity in haemolymph of the tick Ornithodoros moubata. Med. Vet. Entomol. 1, 409–416.
de la Fuente, J., Kocan, K.M., 2003. Advances in the identification and characterization of protective antigens for development of recom- binant vaccines against tick infestations. Expert Rev. Vaccines 2, 583–593.
de la Fuente, J., Rodríguez, M., Redondo, M., Montero, C., García-García, J.C., Méndez, L., Serrano, E., Valdés, M., Enríquez, A., Canales, M., Ramos, E., de Armas, C.A., Rey, S., Rodríguez, J.L., Artiles, M., García, L., 1998. Field studies and cost-effectiveness analysis of vaccination with GavacTM
against the cattle tick Boophilus microplus. Vaccine 16, 366–373. de la Fuente, J., Almazán, C., Naranjo, V., Blouin, E.F., Meyer, J.M., Kocan,
K.M., 2006a. Autocidal control of ticks by silencing of a single gene by RNA interference. Biochem. Biophys. Res. Commun. 344, 332–338.
de la Fuente, J., Almazán, C., Blas-Machado, U., Naranjo, V., Mangold, A.J., Blouin, E.F., Gortazar, C., Kocan, K.M., 2006b. The tick protective anti- gen, 4D8, is a conserved protein involved in modulation of tick blood ingestion and reproduction. Vaccine 24, 4082–4095.
de la Fuente, J., Almazán, C., Blouin, E.F., Naranjo, V., Kocan, K.M., 2006c. Reduction of tick infections with Anaplasma marginale and A. phagocy- tophilum by targeting the tick protective antigen subolesin. Parasitol. Res. 100, 85–91.
de la Fuente, J., Almazán, C., Canales, M., Pérez de la Lastra, J.M., Kocan, K.M., Willadsen, P., 2007a. A ten-year review of commercial vaccine performance for control of tick infestations on cattle. Anim. Health Res. Rev. 8, 23–28.
de la Fuente, J., Kocan, K.M., Blouin, E.F., 2007b. Tick vaccines and the transmission of tick-borne pathogens. Vet. Res. Commun. 31 (Suppl. 1), 85–90.
de la Fuente, J., Estrada-Pen a, A., Venzal, J.M., Kocan, K.M., Sonenshine, D.E., 2008a. Overview: ticks as vectors of pathogens that cause disease in humans and animals. Front. Biosci. 13, 6938–6946.
de la Fuente, J., Maritz-Olivier, C., Naranjo, V., Ayoubi, P., Nijhof, A.M., Almazán, C., Canales, M., Pérez de la Lastra, J.M., Galindo, R.C., Blouin, E.F., Gortazar, C., Jongejan, F., Kocan, K.M., 2008b. Evidence of the role of tick subolesin in gene expression. BMC Genomics 9, 372.
de la Fuente, J., Kocan, K.M., Blouin, E.F., Zivkovic, Z., Naranjo, V., Almazán, C., Esteves, E., Jongejan, J., Daffre, S., Mangold, A.J., 2010a. Functional genomics and evolution of tick-Anaplasma interactions and vaccine development. Vet. Parasitol. 167, 175–186.
de la Fuente, J., Manzano-Roman, R., Naranjo, V., Kocan, K.M., Zivkovic, Z., Blouin, E.F., Canales, M., Almazán, C., Galindo, R.C., Step, D.L., Villar, M., 2010b. Identification of protective antigens by RNA interference for control of the lone star tick. Amblyomma americanum. Vaccine 28, 1786–1795.
Eisemann, C.H., Pearson, R.D., Donaldson, R.A., Cadogan, L.C., Vuocolo, T., 1993. Uptake and fate of specific antibody in feeding lar- vae of the sheep blowfly, Lucilia cuprina. Med. Vet. Entomol. 7, 177–185.
Elad, D., Segal, E., 1995. Immunogenicity in calves of a crude riboso- mal fraction of Trichophyton verrucosum: a field trial. Vaccine 13, 83–87.
Foy, B.D., Magalhaes, T., Injera, W.E., Sutherland, I., Devenport, M., Thanawastien, A., Ripley, D., Cárdenas-Freytag, L., Beier, J.C., 2003. Induction of mosquitocidal activity in mice immunized with Anopheles gambiae midgut cDNA. Infect. Immun. 71, 2032–2040.
Galindo, R.C., Doncel-Pérez, E., Zivkovic, Z., Naranjo, V., Gortazar, C., Man- gold, A.J., Martín-Hernando, M.P., Kocan, K.M., de la Fuente, J., 2009. Tick subolesin is an ortholog of the akirins described in insects and vertebrates. Dev. Comp. Immunol. 33, 612–617.
Gonzalez, K., Baylies, M., 2005. Bhringi: a novel twist co-regulator. A Dros. Res. Conf. 46, 320B.
Goto, A., Matsushita, K., Gesellchen, V., El Chamy, L., Kuttenkeuler, D., Takeuchi, O., Hoffmann, J.A., Akira, S., Boutros, M., Reichhart, J.M., 2008. Akirins are highly conserved nuclear proteins required for NF-kappaB-dependent gene expression in drosophila and mice. Nat. Immunol. 9, 97–104.
Halstead, S.B., 2007. Dengue. Lancet 370, 1644–1652. Harrington, D., Canales, M., de la Fuente, J., de Luna, C., Robinson, K., Guy,
J., Sparagano, O., 2009. Immunisation with recombinant proteins sub- olesin and Bm86 for the control of Dermanyssus gallinae in poultry. Vaccine 27, 4056–4063.
Hatfield, P.R., 1988. Detection and localization of antibody ingested with a mosquito bloodmeal. Med. Vet. Entomol. 2, 339–345.
Jasinskas, A., Jaworski, D.C., Barbour, A.G., 2000. Amblyomma americanum: specific uptake of immunoglobulins into tick hemolymph during feed- ing. Exp. Parasitol. 96, 213–221.
Jones, K.E., Patel, N.G., Levy, M.A., Storeygard, A., Balk, D., Gittleman, J.L., Daszac, P., 2008. Global trends in emerging infectious diseases. Nature 451, 990–994.
Kedzierski, L., Zhu, Y., Handman, E., 2006. Leishmania vaccines: progress and problems. Parasitol. 133, S87–S112.
Kishore, K., Kumar, V., Kesari, S., Dinesh, D.S., Kumar, A.J., Das, P., Bhat- tacharya, S.K., 2006. Vector control in leishmaniasis. Indian J. Med. Res. 123, 467–472.
Kocan, K.M., Manzano-Roman, R., de la Fuente, J., 2007. Transovarial silencing of the subolesin gene in three-host ixodid tick species after injection of replete females with subolesin dsRNA. Parasitol. Res. 100, 1411–1415.
Kocan, K.M., Zivkovic, Z., Blouin, E.F., Naranjo, V., Almazán, C., Mitra, R., de la Fuente, J., 2009. Silencing of genes involved in Anaplasma marginale-tick interactions affects the pathogen developmental cycle in Dermacentor variabilis. BMC Dev. Biol. 9, 42.
Komiya, Y., Kurabe, N., Katagiri, K., Ogawa, M., Sugiyama, A., Kawasaki, Y., Tashiro, F., 2008. A novel binding factor of 14-3-3beta functions as a transcriptional repressor and promotes anchorage- independent growth, tumorigenicity, and metastasis. J. Biol. Chem. 283, 18753–18764.
Lackie, A.M., Gavin, S., 1989. Uptake and persistence of ingested anti- body in the mosquito Anopheles stephensi. Med. Vet. Entomol. 3, 225–230.
Lal, A.A., Patterson, P.S., Sacci, J.B., Vaughan, J.A., Paul, C., Collins, W.E., Wirtz, R.A., Azad, A.F., 2001. Anti-mosquito midgut antibodies block development of Plasmodium falciparum and Plasmodium vivax in mul- tiple species of Anopheles mosquitoes and reduce vector fecundity and survivorship. Proc. Natl. Acad. Sci. U.S.A. 98, 5228–5233.
Macqueen, D.J., Johnston, I.A., 2009. Evolution of the multifaceted eukary- otic akirin gene family. BMC Evol. Biol. 9, 34.

&KDSWHU�,
11!
Macqueen, D.J., Kristjánsson, B.K., Johnston, I.A., 2010. Salmonid genomes have a remarkably expanded akirin family, co-expressedwith genes from conserved pathways governing skeletal muscle growth and- catabolism. Physiol. Genomics 42, 134–148.
Maeda, I., Kohara, Y., Yamamoto, M., Sugimoto, A., 2001. Large-scale anal- ysis of gene function in Caenorhabditis elegans by high-throughput RNAi. Curr. Biol. 11, 171–176.
Mangold, A.J., Galindo, R.C., de la Fuente, J., 2009. Response to the com- mentary of D. Macqueen on: Galindo, R.C., Doncel-Pérez, E., Zivkovic, Z., Naranjo, V., Gortazar, C., Mangold, A.J, et al. Tick subolesin is an ortholog of the akirins described in insects and vertebrates [Dev. Comp. Immunol. 33 (2009) 612–617]. Dev. Comp. Immunol. 33, 878–879.
Moreno-Cid, J.A., Jiménez, M., Cornelie, S., Molina, R., Alarcón, P., Lacroix, M.-N., Pinal, R., Delacour, S., Lucientes, J., Canales, M., Pérez de la Las- tra, J.M., Villar, M., de la Fuente, J., 2011. Characterization of Aedes albopictus akirin for the control of mosquitoand sand fly infestations. Vaccine 29, 77–82.
Marshall, A., Salerno, M.S., Thomas, M., Davies, T., Berry, C., Dyer, K., Brace- girdle, J., Watson, T., Dziadek, M., Kambadur, R., Bower, R., Sharma, M., 2008. Mighty is a novel promyogenic factor in skeletal myogenesis. Exp. Cell Res. 314, 1013–1029.
Milleron, R.S., Ribeiro, J.M., Elnaime, D., Soong, L., Lanzaro, G., 2004. Neg- ative effect of antibodies against maxadilan on the fitness of the sand fly vector of American visceral leishmaniasis. Am. J. Trop. Med. Hyg. 70, 278–285.
Munderloh, U.G., Jauron, S.D., Fingerle, V., Leitritz, L., Hayes, S.F., Hautman, J.M., Nelson, C.M., Huberty, B.W., Kurtti, T.J., Ahlstrand, G.G., Greig, B., Mellencamp, M.A., Goodman, J.L., 1999. Invasion and intracellular development of the human granulocytic ehrlichiosis agent in tick cell culture. J. Clin. Microbiol. 37, 2518–2524.
Nijhof, A.M., Taoufik, A., de la Fuente, J., Kocan, K.M., de Vries, E., Jonge- jan, F., 2007. Gene silencing of the tick protective antigens, Bm86, Bm91 and subolesin, in the one-host tick Boophilus microplus by RNA interference. Int. J. Parasitol. 37, 653–662.
Peter, R.J., Van den Bossche, P., Penzhorn, B.L., Sharp, B., 2005. Tick, fly, and mosquito control—lessons from the past, solutions for the future. Vet. Parasitol. 132, 205–215.
Prudencio, C.R., Pérez de la Lastra, J.M., Canales, M., Villar, M., de la Fuente, J., 2010. Mappingprotective epitopes in the tick and mosquitosub- olesin ortholog proteins. Vaccine 28, 5398–5406.
Rodríguez Valle, M., Mèndez, L., Valdez, M., Redondo, M., Espinosa, C.M., Vargas, M., Cruz, R.L., Barrios, H.P., Seoane, G., Ramirez, E.S., Boue, O., Vigil, J.L., Machado, H., Nordelo, C.B., Pin eiro, M.J., 2004. Integrated control of Boophilus microplus ticks in Cuba based on vaccination with the anti-tick vaccine Gavac. Exp. Appl. Acarol. 34, 375–382.
Salerno, M.S., Dyer, K., Bracegirdle, J., Platt, L., Thomas, M., Siriett, V., Kam- badur, R., Sharma, M., 2009. Akirin1 (Mighty), a novel promyogenic
factor regulates muscle regeneration and cell chemotaxis. Exp. Cell Res. 315, 2012–2021.
Saul, A., 2007. Mosquito stage, transmission blocking vaccines for malaria. Curr. Opin. Infect. Dis. 5, 476–481.
Speranc a, M.A., Capurro, M.L., 2007. Perspectives in the control of infec- tious diseases by transgenic mosquitoes in the post-genomic era – a review. Mem. Inst. Oswaldo Cruz 102, 425–433.
Suneja, A., Gulia, M., Gakhar, S.K., 2003. Blocking of malaria parasite devel- opment in mosquito and fecundity reduction by midgut antibodies in Anopheles stephensi (Diptera: Culicidae). Arch. Insect Biochem. Physiol. 52, 63–70.
Szalai, K., Jensen-Jarolim, E., Pali-Schöll, I., 2008. Vaccination strategies based on the mimotope concept. G. Ital. Dermatol. Venereol. 143, 95–104.
Tellam, R.L., Eisemann, C.H., Vuocolo, T., Casu, R., Jarmey, J., Bowles, V., Pearson, R., 2001. Role of oligosaccharides in the immune response of sheep vaccinated with Lucilia cuprina larval glycoprotein, peritrophin- 95. Int. J. Parasitol. 31, 798–809.
Titus, R.G., Bishop, J.V., Mejia, J.S., 2006. The immunomodulatory factors of arthropod saliva and the potential for these factors to serve as vac- cine targets to prevent pathogen transmission. Parasite Immunol. 4, 131–141.
Trimnell, A.R., Davies, G.M., Lissina, O., Hails, R.S., Nuttall, P.A., 2005. A cross-reactive tick cement antigen is a candidate broad-spectrum tick vaccine. Vaccine 23, 4329–4341.
Valenzuela, J.G., Belkaid, Y., Garfield, M.K., Mendez, S., Kamhawi, S., Rowton, E.D., Sacks, D.L., Ribeiro, J.M., 2001. Toward a defined anti- Leishmania vaccine targeting vector antigens: characterization of a protective salivary protein. J. Exp. Med. 194, 331–342.
Vaughan, J.A., Thomas, R.E., Silver, G.M., Wisnewski, N., Azad, A.F., 1998. Quantitation of cat immunoglobulins in the hemolymph of cat fleas (Siphonaptera: Pulicidae) after feeding on blood. J. Med. Entomol. 35, 404–409.
Vaughan, J.A., Sonenshine, D.E., Azad, A.F., 2002. Kinetics of ingested host immunoglobulin G in hemolymph and whole body homogenates during nymphal development of Dermacentor variabilis and Ixodes scapularis ticks (Acari: Ixodidae). Exp. Appl. Acarol. 27, 329–340.
Vaz Junior Ida, S., Martinez, R.H., Oliveira, A., Heck, A., Logullo, C., Gonzales, J.C., Dewes, H., Masuda, A., 1996. Functional bovine immunoglob- ulins in Boophilus microplus hemolymph. Vet. Parasitol. 62, 155–160.
Willadsen, P., 2004. Anti-tick vaccines. Parasitology 129, S367–S874. Zivkovic, Z., Torina, A., Mitra, R., Alongi, A., Scimeca, S., Kocan, K.M.,
Galindo, R.C., Almazán, C., Blouin, E.F., Villar, M., Nijhof, A.M., Mani, R., La Barbera, G., Caracappa, S., Jongejan, F., de la Fuente, J., 2010. Subolesin expression in response to pathogen infection in ticks. BMC Immunol. 11, 7.


HIPÓTESIS&Y&OBJETIVOS&
HYPOTHESIS&AND&AIMS&

13
HIPÓTESIS
/a expresión y la producción de antígenos protectores contra ectoparásitos hematófagos para
control inmunológico de las infestaciones y las enfermedades que transmiten� HV� SRVLEOH
mediante el desarrollo de vacunas formuladas con subunidades proteicas altamente
conservadas en vectores.
OBJETIVOS
1. ExpresDU del antígeno Akirina (AKR) del mosquito Aedes albopictus y su ortólogo en�
garrapata, el antígeno Subolesina (SUB), mediante diferentes sistemas de expresión de�
proteínas recombinantes.
2. CaracterizaU los antígenos AKR y SUB expresados en diferentes sistemas.
3. 5HDOL]DU�HnsayRV de vacunación con el antígeno AKR/SUB recombinante para el
control�ectoparásitos hematófagos.

14
HYPOTHESIS
7he expression and production of protective antigens for immunological control of
hematophagous ectoparasites infestations and the diseases they transmit� LV�SRVVLEOH through the
development of vaccines based in highly conserved protein subunits vectors.
OBJECTIVES
1. (xpress mosquito Aedes albopictus A kirin antigen (AKR) and its ortholog in� ticks,
Subolesin antigen (SUB) by using different expression systems of recombinant�proteins.
2. &haracterL]H of expressed AKR/SUB antigens in different systems.
3. 3HUIRUP�Yaccination trials with recombinant AKR/SUB antigen for control of
hematophagous�ectoparasites.

CHAPTER&II.&EXPRESSION&&&PRODUCTION&
Moreno3Cid,! J.! A.,! Canales! M.! and! de! la! Fuente! J.! (2012).! Production* of*recombinant* Aedes% albopictus* akirin* in* Pichia% pastoris* using* an* aqueous*two:*phase* semicontinuous* fermentation*process.*Biochemical! Engineering!Journal!68(0):!1143119.!


PRODUCTION' OF' RECOMBINANT' AEDES& ALBOPICTUS'
AKIRIN'IN'PICHIA&PASTORIS'USING'AN'AQUEOUS'TWO9
PHASE''SEMICONTINUOUS''FERMENTATION''PROCESS'

17!
&KDSWHU�,,
Contents lists available at SciVerse ScienceDirect
Biochemical Engineering Journal
journa l h o me pa ge: www.elsevier .com/locate /bej
Production of recombinant Aedes albopictus akirin in Pichia pastoris using an aqueous two-phase semicontinuous fermentation process
Juan A. Moreno-Cid a , Mario Canales a , José de la Fuente a,b,∗
a Instituto de Investigación en Recursos Cinegéticos IREC-CSIC-UCLM-JCCM, Ronda de Toledo s/n, 13005 Ciudad Real, Spain b Department of Veterinary Pathobiology, Center for Veterinary Health Sciences, Oklahoma State University, Stillwater, OK 74078, USA
a r t i c l e i n f o a b s t r a c t
Article history: Received 28 May 2012 Received in revised form 3 July 2012 Accepted 23 July 2012 Available online 31 July 2012
Keywords: Pichia pastoris Akirin Subolesin Fermentation Biotechnology Vaccine
The yeast, Pichia pastoris, has been successfully used as an efficient system to produce heterologous proteins. The secretion of the recombinant Aedes albopictus mosquito akirin (AKR) from P. pastoris using an extractive bioconversion in an aqueous two-phase system (ATPS) fed-batch fermentation process allowed the conduction of vaccination trials to assay its protective efficacy against different arthropod vector species. Herein, we report the development of a semicontinuous process for the extractive bioconversion in an ATPS coupled with a simple and easy to scale up process for polymer recycling as an effective alternative to improve production of recombinant A. albopictus AKR in P. pastoris. Six repetitive batches of the extractive bioconversion were conducted in a semicontinuous process by replacing 2/3 of the culture and recycling the top polymeric phase every 72 h. Overall process in a 5-L fermentor yielded a protein concentration in the bottom phase of 0.46 ± 0.13 mg mL−1 and 6 g of total purified protein in 490 h with a reduction of 25% of the operation time and 27% of the operation cost when compared to the fed-batch ATPS process. These results improve the production process for mosquito AKR and suggest the possibilities to develop similar processes for the expression of other recombinant proteins in P. pastoris.
© 2012 Elsevier B.V. All rights reserved.
1. Introduction
Akirin (AKR) and Subolesin (SUB) are evolutionaryconserved ortholog proteins that affect the expression of signal transduction and innate immune response genes in vertebrates and inverte- brates [1–4]. The broad function of SUB/AKR as transcription factors explains the profound effect of gene knockdown by RNA inter- ference on tick and insect physiology and development and on gene expression in ticks [2,3,5,4,6–8]. Recent experiments with recombinant SUB/AKR have shown the effect of vaccination and/or antigen-specific antibodies on several arthropod vectors including hard and soft ticks, mosquitoes, sand flies, poultry red mites and sea lice [9–14]. These results have encouraged designing a cost- effective process to produce recombinant Aedes albopictus AKR for the development of a universal vaccine against multiple arthropod vector species [9–14].
Pichia pastoris has emerged as an efficient expression system for the production of recombinant proteins [15–17]. The productivity and economics of the expression system not only depends on several genetic and physiological factors of the host cell but also on the optimization of the fermentation process. The fermentation process
∗ Corresponding author at: Instituto de Investigación en Recursos CinegéticosIREC-CSIC-UCLM-JCCM, Ronda de Toledo s/n, 13005 Ciudad Real, Spain.
E-mail address: jose [email protected] (J. de la Fuente).
including a glycerol or glucose batch phase followed by a methanol induction throughout a fed-batch stage has been accepted as a stan- dard protocol to produce recombinant proteins in P. pastoris [18]. In general, this strategy allows obtaining high protein expression levels and high cell densities [18]. However, these fermentation processes are time consuming due to the low P. pastoris growth rate in methanol [19].
The gene encoding for the mosquito A. albopictus AKR was recently cloned to produce the recombinant protein in P. pastoris using an extractive bioconversion process in an aqueous two- phase system (ATPS) [20]. ATPS are formed when two or more hydrophilic polymers or a salt and a polymer in water solution are dissolved together above certain concentrations and have been widely used in biotechnology for the extraction and purification of biomolecules including recombinant proteins, with high potential in downstream processing as a large-scale continuous operation [21–31]. In addition, extractive bioconversions using ATPS have proved to be a promising alternative to conventional fermenta- tion processes for simultaneous production and purification of proteins and biomolecules [22–24,27,28,32,33]. The extractive bio- conversion using ATPS allowed combining AKR fermentation and purification processes in a single step [20].
In extractive bioconversion, polymer recycling is of paramount importance in process optimization and economics [34–36]. It has been previously demonstrated that the productivity of fer- mentation processes with P. pastoris can be improved by using
1369-703X/$ – see front matter © 2012 Elsevier B.V. All rights reserved. http://dx.doi.org/10.1016/j.bej.2012.07.020

18!
Fig. 1. Bioprocess flowchart showing the semicontinuous operation for the extractive bioconversion in an ATPS coupled with polymer recycling for the production of recombinant A. albopictus AKR in P. pastoris. The schematic diagram shows two integrated processes, with and without PEG4000 recycling.
a semicontinuous system as an approximation to a continuous system [37–40]. Herein, we report the development of a semicon- tinuous process for the extractive bioconversion in an ATPS coupled with a simple and easy to scale up process for polymer recycling as an effective alternative to improve production of recombinant A. albopictus AKR in P. pastoris.
2. Materials and methods2.1. Chemicals, media and solutions
Unless otherwise indicated, all reagents used in this work were purchased either from Sigma–Aldrich (St Louis, MO, USA) or VWR International Eurolab S.L. (Mollet del Vallés, Barcelona, Spain). Yeast extract and yeast extract peptone (YP) medium having 10 g L−1 yeast extract and 20 g L−1 peptone were purchased from Laboratorios CONDA S.A. (Madrid, Spain).
The ATPS was composed of 23% PEG4000 and 12% salts and was obtained after preparing a solution containing 46.1% of the saline medium (SM) at 16.7% (w/v) plus 75.5% of PBS buffer at 35% (w/v); 11.8% of a glucose solution at 40% (w/v) and 46% of a PEG4000 solu- tion at 50% (w/v) at pH 5.0. The PEG4000-salts system was chosen at a stability ratio [41,42] that avoids reaching the continuous phase of the system below the binodial curve according to our previous work [20].
The composition of the SM used in ATPS media for the expres- sion of the recombinant A. albopictus AKR was 12.9 g L−1 KH2PO4, 11.25 g L−1 (NH4)2SO4, 4.5 g L−1 MgSO4, 0.56 g L−1 CaCl2·2H2O, 5 g L−1 urea and 30 g L−1 glucose; supplemented with 5 mL L−1
of both a Trace Element Solution (TES) containing: 2.0 g L−1
ZnSO4·7H2O, 0.02 g L−1 CuSO4·5H2O, 0.08 g L−1 KI, 0.3 g L−1
MnSO4·H2O, 0.19 g L−1 Na2MoO4·H2O, 0.02 g L−1 H3BO3, 2.9 g L−1
FeCl3. A vitamin solution (VT) containing 0.4 g L−1 calcium pan- tothenate, 0.4 g L−1 tyamine, 0.4 g L−1 myo-inositol, 0.1 g L−1
nicotinic acid, 0.4 g L−1 pyridoxine and 0.4 g L−1 biotin was also periodically supplemented to the culture at a rate of 5 mL L−1 every 24 h.
2.2. Cell propagation
The A. albopictus AKR (Genbank accession number EU637024) was cloned into P. pastoris strain X33M84D8-2 for secretion of the recombinant protein as previously reported [20]. Pre-inoculums and inoculums for Erlenmeyer flasks or bioreactor cultures were grown in a shaker at 30 ◦C and 250 rpm. Two 100-mL long-term stock vials from the Master Cell Bank were seeded in 1 mL YP medium, grown for 12 h and transferred into 4× 50-mL tubes containing 5 mL of YP medium with 20 g L−1 glycerol. After 24 h, cultures were mixed again and 5 mL were used to inoculate the 2-L Erlenmeyer flasks containing 250 mL of YP medium with 20 g L−1
glycerol. Cell growth was resumed 24 h later and cultures were used to seed the 5-L bioreactor.
2.3. Semicontinuous fermentation processes in a 5-L bioreactor
Cells from propagation steps in 2× 250-mL YPD medium were seeded into a 5-L working volume Biostat B bioreactor (B. Braun Biotech, Melsungen, Germany) containing 2.5 L of the ATPS media supplemented with 30 g L−1 glucose. During the whole process, pH was kept at 5.0 by adding NH4OH or H3PO4, temperature was kept at 30 ◦C and dissolved oxygen was maintained at 30% saturation by regulating agitation and aeration rates.
A standard two-phase cultivation protocol was used during the first stage. This protocol included a 24 h batch stage in glucose followed by a 466 h methanol fed-batch stage. Upon exhaustion of glucose, indicated by a sharp increase in dissolved oxygen, methanol induction was made following the P. pastoris Fermentation Process Guideline [19]. Methanol concentration in the media was monitored using a sensor probe (Raven Biotech, Inc.) and flow rates were adjusted to keep a final concentration between 1 and 3 g L−1. When cell growth in methanol was finished, two thirds of the culture broth was withdrawn from the bioreactor and replaced with sterile ATPS to carry out the semicontinuous fermentation process. Fig. 1 describes the main operation units of the process. Throughout the entire fermentation process, supplements of TES and VT solutions were added to the culture medium every 24 h.
&KDSWHU�,,

19!
Tab
le 1
T
hrou
ghpu
ts f
rom
the
sem
icon
tinu
ous
extr
acti
ve b
ioco
nver
sion
in A
TPS
usin
g a
5-L
fer
men
tor
to p
rodu
ce A
. alb
opic
tus
AK
R in
P. pa
stor
is.
Ste
p C
arbo
n so
urce
C
arbo
n so
urce
co
msu
ptio
n (g
) S
peci
fic
grow
th
rate
, µsp
(h−
1 )
Bio
mas
s/su
bstr
ate
yiel
d (g
g−
1 )
Aki
rin
conc
entr
atio
n (m
g m
L−
1 )
Pro
tein
/bio
mas
s yi
eld
(mg
g−1 )
PEG
4000
co
nsum
ptio
n (g
) R
ecyc
led
PEG
4000
co
nsum
ptio
n (g
) P
rodu
ctiv
ity
Bio
mas
s (g
L−
1 h−
1 )
0.64
0.
39
0.52
0.
34
0.48
0.
33
0.51
0.
43 ±
0.0
8
Pro
tein
(m
g L−
1 h−
1 )
– 4.34
6.
26
4.42
8.
48
5.24
7.
71
6.08
± 1
.73
1 2 3 4 5 6 7 Ove
rall
pro
cess
Glu
cose
M
eOH
M
eOH
M
eOH
M
eOH
M
eOH
M
eOH
75
356.
4 36
0.36
38
0.16
59
4 47
5.2
546.
48
452
± 1
02
0.15
0.
01
0.02
0.
01
0.02
0.
01
0.02
0.
02 ±
0.0
0
0.20
0.
08
0.09
0.
07
0.06
0.
07
0.07
0.
07 ±
0.0
1
– 0.31
0.
39
0.36
0.
64
0.50
0.
56
0.46
± 0
.13
– 10.9
8 11
.97
12.8
6 17
.65
15.6
7 15
.21
14.0
6 ±
2.5
3
575
575
400
500
– – – 492
± 8
8
– – – – 458
458
458
458
± 0
The
ave
rage
± S
D o
f th
e m
ain
par
amet
ers
of th
e pr
oces
s fo
r A
KR
pro
duct
ion
wer
e ca
lcul
ated
ove
r m
etha
nol s
tep
s 2–
7. S
teps
2–5
wer
e do
ne w
ith
fre
sh o
rgan
ic p
hase
and
ste
ps
6 an
d 7
usin
g re
cycl
ed P
EG.
2.4. Biomass analysis
Time-course samples were withdrawn from the bioreactor at regular intervals to check growth rate and protein concentration. Cell density was measured as grams of wet weight per liter of broth, which was obtained by centrifuging the samples for 10 min at 12,500 × g (Allegr X-22R centrifuge, rotor F2402H; Beckman- Coulter, Palo Alto, CA, USA). Samples were diluted ten folds to disrupt the ATPS and to decrease the viscosity with the purpose of favoring cell precipitation during centrifugation. In previous works [45 and unpublished results], data of dry weight was measured in a moisture analyzer MA30 (Sartorius AG, Gottingen, Germany) where wet weight and the O.D.600 nm of samples from more than 35 fermentation processes in PM medium (N = 650) were statistically analyzed by a simple regression analysis and the linear correlations between parameters were obtained with regression coefficients higher than 0.99.
In this work, biomass was also measured by O.D.600 nm and expressed as:
O.D.600 nm = 1.39 × wet weight (g L−1) − 27.26
or as dry weight by using the experimental equation:
dry weight (g L−1) = 0.18 wet weight (g L−1) − 3.79
2.5. Phase separation and protein recovery
Fermentation broth from the 5-L bioreactor was separated for 30 min at 8000 rpm and 4 ◦C in a Beckman JLA-10.500 centrifuge. Solids were rejected and supernatant was carefully recovered and placed in a separating funnel. Phases were then separated by grav- ity settling at 4 ◦C for 1 h. Bottom phase was collected and filtered through a 0.22 µm filtration system StericupTM (Millipore, Bedford, MA, USA) to recover the recombinant A. albopictus AKR.
2.6. Purification and characterization of PEG4000 from the upper phase
The upper phase was sterilized at 121 ◦C for 20 min and placed in a separating funnel to recover the PEG4000. After gravity settling at 25 ◦C for 1 h, the PEG4000 rich phase was collected when reached 60 ◦C, filtered through a 0.22 µm filtration system StericupTM (Mil- lipore) and analyzed. Recovered PEG4000 purity was determined by using a previously reported protocol [44]. Briefly, 5 mL of 0.5 M perchloric acid were added to 1 mL of PEG4000-containing protein solution. After 15 min the mixture was centrifuged for 10 min at 4000 × g. Then, 1 mL 5% bariumchloride solution and 0.4 mL 0.1 M iodine solution were added to 4 mL of the supernatant. After stand- ing for 15 min, extinction coefficient was measured at O.D.535 nm.
2.7. Recombinant protein characterization
Total protein concentration was quantified using the Bradford method with BSA as standard [45]. The expression of recombinant AKR was determined by densitometric analysis of Coomassie based Instant Blue-stained SDS-polyacrylamide gels using ImageJ 1.44p (National Institute of Health, USA).
Samples from bottom phase were dialyzed against water to remove the high salts concentration from ATPS. Samples were analyzed using denaturing SDS-12% polyacrylamide gel elec- trophoresis (PAGE-gel Inc., San Diego, CA, USA) under reducing conditions. The bands were visualized by Coomassie Brilliant Blue R250 staining. Samples were treated with DTT reducer (PAGE-gel Inc.), heated in boiling water for 5 min before load- ing onto gels and electrophoresed for 80 min at 90 mA constant
&KDSWHU�,,
20!
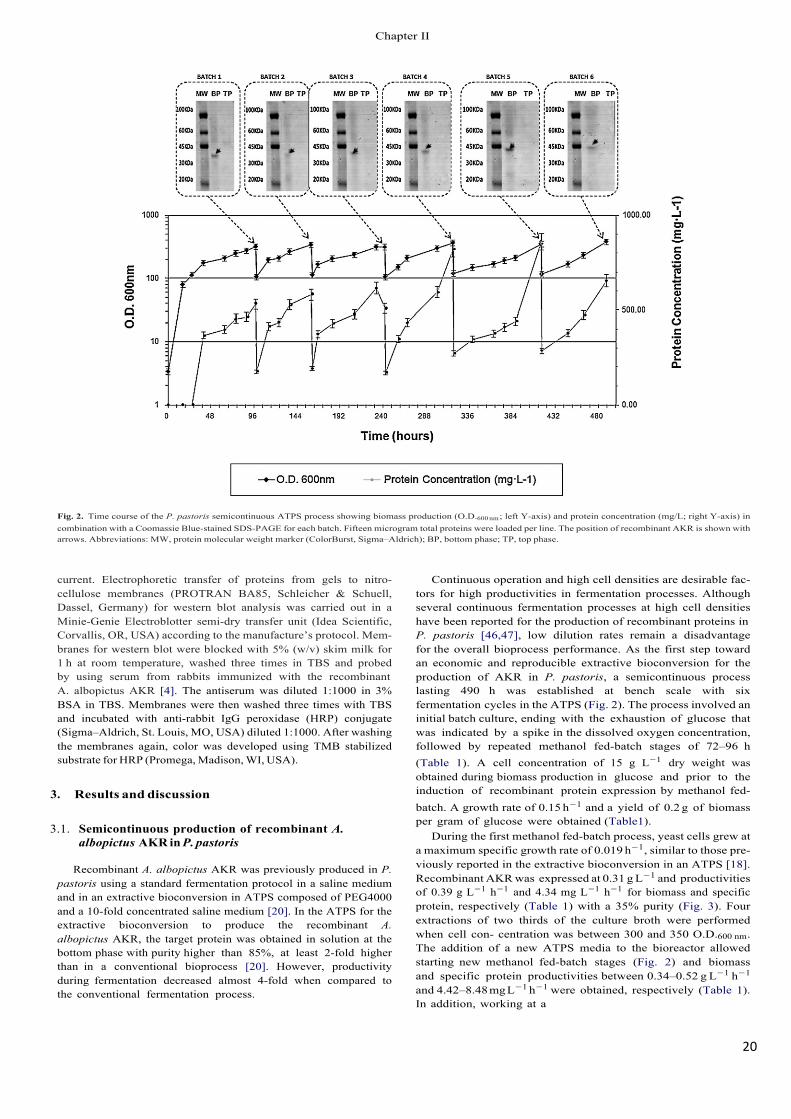
Fig. 2. Time course of the P. pastoris semicontinuous ATPS process showing biomass production (O.D.600 nm ; left Y-axis) and protein concentration (mg/L; right Y-axis) in combination with a Coomassie Blue-stained SDS-PAGE for each batch. Fifteen microgram total proteins were loaded per line. The position of recombinant AKR is shown with arrows. Abbreviations: MW, protein molecular weight marker (ColorBurst, Sigma–Aldrich); BP, bottom phase; TP, top phase.
current. Electrophoretic transfer of proteins from gels to nitro- cellulose membranes (PROTRAN BA85, Schleicher & Schuell, Dassel, Germany) for western blot analysis was carried out in a Minie-Genie Electroblotter semi-dry transfer unit (Idea Scientific, Corvallis, OR, USA) according to the manufacture’s protocol. Mem- branes for western blot were blocked with 5% (w/v) skim milk for 1 h at room temperature, washed three times in TBS and probed by using serum from rabbits immunized with the recombinant A. albopictus AKR [4]. The antiserum was diluted 1:1000 in 3% BSA in TBS. Membranes were then washed three times with TBS and incubated with anti-rabbit IgG peroxidase (HRP) conjugate (Sigma–Aldrich, St. Louis, MO, USA) diluted 1:1000. After washing the membranes again, color was developed using TMB stabilized substrate for HRP (Promega, Madison, WI, USA).
3. Results and discussion
3.1. Semicontinuous production of recombinant A. albopictus AKR in P. pastoris
Recombinant A. albopictus AKR was previously produced in P. pastoris using a standard fermentation protocol in a saline medium and in an extractive bioconversion in ATPS composed of PEG4000 and a 10-fold concentrated saline medium [20]. In the ATPS for the extractive bioconversion to produce the recombinant A. albopictus AKR, the target protein was obtained in solution at the bottom phase with purity higher than 85%, at least 2-fold higher than in a conventional bioprocess [20]. However, productivity during fermentation decreased almost 4-fold when compared to the conventional fermentation process.
Continuous operation and high cell densities are desirable fac- tors for high productivities in fermentation processes. Although several continuous fermentation processes at high cell densities have been reported for the production of recombinant proteins in P. pastoris [46,47], low dilution rates remain a disadvantage for the overall bioprocess performance. As the first step toward an economic and reproducible extractive bioconversion for the production of AKR in P. pastoris, a semicontinuous process lasting 490 h was established at bench scale with six fermentation cycles in the ATPS (Fig. 2). The process involved an initial batch culture, ending with the exhaustion of glucose that was indicated by a spike in the dissolved oxygen concentration, followed by repeated methanol fed-batch stages of 72–96 h (Table 1). A cell concentration of 15 g L−1 dry weight was obtained during biomass production in glucose and prior to the induction of recombinant protein expression by methanol fed-batch. A growth rate of 0.15 h−1 and a yield of 0.2 g of biomass per gram of glucose were obtained (Table1).
During the first methanol fed-batch process, yeast cells grew at a maximum specific growth rate of 0.019 h−1, similar to those pre- viously reported in the extractive bioconversion in an ATPS [18]. Recombinant AKR was expressed at 0.31 g L−1 and productivities of 0.39 g L−1 h−1 and 4.34 mg L−1 h−1 for biomass and specific protein, respectively (Table 1) with a 35% purity (Fig. 3). Four extractions of two thirds of the culture broth were performed when cell con- centration was between 300 and 350 O.D.600 nm. The addition of a new ATPS media to the bioreactor allowed starting new methanol fed-batch stages (Fig. 2) and biomass and specific protein productivities between 0.34–0.52 g L−1 h−1
and 4.42–8.48 mg L−1 h−1 were obtained, respectively (Table 1). In addition, working at a
&KDSWHU�,,

21!
Fig. 3. Characterization of recombinant AKR produced in the semicontinuous ATPS process. Coomassie Blue-stained SDS-PAGE (A) and Western Blot (B) analysis of 15 µg total proteins from bottom phase in fermentation cycles 1–6. Samples were dialyzed for 24 h at 4 ◦ C against 10 water volumes to remove salts before electrophoresis. The position of recombinant AKR is shown with arrows. Abbreviation: MW, protein molecular weight marker (ColorBurst, Sigma–Aldrich).
security ratio of 0.25 [20] allowed obtaining well-separated phases after centrifugation of the removed culture broth during the whole semicontinuous process in spite of methanol consumption that ranged between 5.9 and 9.6 mL h−1 per culture liter. After phase separation, recombinant A. albopictus AKR partitioned to the bot- tom phase only as previously reported [20] and demonstrated here by SDS-PAGE and Western blot (Figs. 2 and 3).
When compared to the ATPS in batch [20], the semicontinu- ous process increased specific protein productivity and purity by 1.5–2.2 and 1.07–1.43 fold, respectively.
3.2. Semicontinuous production of recombinant A. albopictus AKR in P. pastoris using recycled PEG4000
Two repetitive batches were conducted by recycling the organic phase (top phase), to achieve an economic and environmentally friendly process. Determination of PEG4000 concentration from the top phase suggested that PEG4000 did not degrade during the whole process, recovering always a 23% of PEG4000 in the top phase. The loss of PEG4000 during recovery operation was less than 1% (w/w), indicating the possibility to reuse the whole organic phase during the semicontinuous operation.
The expression of recombinant AKR and specific protein productivity with recycled PEG4000 were 4% and 10% �higher, respectively than in the process with fresh organic phase (Table 1). Protein purity was similar with recycled PEG400 and fresh organic phase. These results suggested an improvement in the production process, probably by providing a culture medium better adapted to the microorganism. Additionally, the fact of performing serial medium withdrawals increased the accumulation of recombinant protein in the fermentation medium.
In the ATPS fed-batch fermentation process for the production of recombinant AKR, the cost of PEG4000 was estimated as 27% of all raw materials used during extractive bioconversion, being the total operation costs for a secretable protein in P. pastoris of 2% and 68% for raw materials and fermentation process, respec- tively [48]. Therefore, the semicontinuous process using recycled PEG4000 improved the economics of the process by reducing 25% of the operation time and 27% of the operation cost when compared to the fed-batch ATPS process.
4. Conclusions
In summary, the main advantages of the semicontinuous pro- cess for the production of recombinant AKR� using recycled PEG4000 were (a) the easy and effective recycling and recovery of PEG4000 using common techniques applicable to any bioprocess without the need for additional or special investments, (b) the higher recombi- nant protein production when compared to the process with fresh organic phase, and (c) the reduction of the operation time and
production costs by 25% and 27%, respectively as a result of PEG4000 recycling and semicontinouos operation. The process described here for the production of recombinant AKR is easy to scale up with high process integration capability and biocom- patibility, thus potentially applicable to the production of other recombinant proteins in P. pastoris.
Acknowledgements
This research was supported by projects PAI06-0046-5285 and FAU2008-00014-00-00.
References
[1] D.J. Macqueen, I.A. Johnston, Evolution of the multifaceted eukaryotic akirin gene family, BMC Evol. Biol. 9 (2009) 34.
[2] R.C. Galindo, E. Doncel-Pérez, Z. Zivkovic, V. Naranjo, C. Gortazar, A.J. Mangold, M.P. Martín-Hernando, K.M. Kocan, J. de la Fuente, Tick subolesin is an ortholog of the akirins described in insects and vertebrates, Dev. Comp. Immunol. 33 (2009) 612–617.
[3] A.J. Mangold, R.C. Galindo, J. de la Fuente, Response to the commentary of D. Macqueen on: R.C. Galindo, E. Doncel-Pérez, Z. Zivkovic, V. Naranjo, C. Gortazar, A.J. Mangold, et al. Tick subolesin is an ortholog of the akirins described in insects and vertebrates [Dev. Comp. Immunol. 33 (2009) 612–617], Dev. Comp. Immunol. 33 (2009) 878–879.
[4] A. Goto, K. Matsushita, V. Gesellchen, L. El Chamy, D. Kuttenkeuler, O. Takeuchi, J.A. Hoffmann, S. Akira, M. Boutros, J.M. Reichhart, Akirins are highly con- served nuclear proteins required for NF-kappaB-dependent gene expression in drosophila and mice, Nat. Immunol. 9 (2008) 97–104.
[5] J. de la Fuente, C. Almazán, U. Blas-Machado, V. Naranjo, A.J. Mangold, E.F. Blouin, C. Gortazar, K.M. Kocan, The tick protective antigen, 4D8, is a con- served protein involved in modulation of tick blood ingestion and reproduction, Vaccine 24 (2006) 4082–4095.
[6] J. de la Fuente, C. Maritz-Olivier, V. Naranjo, P. Ayoubi, A.M. Nijhof, C. Almazán, M. Canales, J.M. Pérez de la Lastra, R.C. Galindo, E.F. Blouin, C. Gortazar, F. Jonge- jan, K.M. Kocan, Evidence of the role of tick subolesin in gene expression, BMC Genomics 9 (2008) 372.
[7] S.J. Nowak, H. Aihara, K. Gonzalez, Y. Nibu, M.K. Baylies, Akirin links twist- regulated transcription with the Brahma chromatin remodeling complex during embryogenesis, PLoS Genet. 8 (2012) 1002547.
[8] K.M. Kocan, Z. Zivkovic, E.F. Blouin, V. Naranjo, C. Almazán, R. Mitra, J. de la Fuente, Silencing of genes involved in Anaplasma marginale-tick interactions affects the pathogen developmental cycle in Dermacentor variabilis, BMC Dev. Biol. 9 (2009) 42.
[9] M. Canales, V. Naranjo, C. Almazán, R. Molina, S.A. Tsuruta, M.P.J. Szabó, R. Manzano-Roman, J.M. Pérez de la Lastra, K.M. Kocan, M.I. Jiménez, J. Lucientes, M. Villar, J. de la Fuente, Conservation and immunogenicity of the mosquito ortholog of the tick protective antigen, subolesin, Parasitol. Res. 105 (2009) 97–111.
[10] J. de la Fuente, J.A. Moreno-Cid, M. Canales, M. Villar, J.M. Pérez de la Lastra, K.M. Kocan, R.C. Galindo, C. Almazán, E.F. Blouin, Targeting arthropod sub- olesin/akirin for the development of a universal vaccine for control of vector infestations and pathogen transmission, Vet. Parasitol. 181 (2011) 17–22.
[11] J.A. Moreno-Cid, M. Jiménez, S. Cornelie, R. Molina, P. Alarcón, M.-N. Lacroix, R. Pinal, S. Delacour, J. Lucientes, M. Canales, J.M. Pérez de la Lastra, M. Villar, J. de la Fuente, Characterization of Aedes albopictus akirin for the control of mosquito and sand fly infestations, Vaccine 29 (2011) 77–82.
[12] D. Harrington, M. Canales, J. de la Fuente, C. de Luna, K. Robinson, J. Guy, O. Sparagano, Immunisation with recombinant proteins subolesin and Bm86 for the control of Dermanyssus gallinae in poultry, Vaccine 27 (2009) 4056–4063.
&KDSWHU�,,

22!
[13] R. Manzano-Román, V. Díaz-Martín, A. Oleaga, M. Siles-Lucas, R. Pérez-Sánchez, Subolesin/akirin orthologs from Ornithodoros spp. soft ticks: cloning, RNAi gene silencing and protective effect of the recombinant proteins, Vet. Parasitol. 185 (2012) 248–259.
[14] Y. Carpio, L. Basabe, J. Acosta, A. Rodríguez, A. Mendoza, A. Lisperger, E. Zamorano, M. González, M. Rivas, S. Contreras, D. Haussmann, J. Figueroa, V.N. Osorio, G. Asencio, J. Mancilla, G. Ritchie, C. Borroto, M.P. Estrada, Novel gene isolated from Caligus rogercresseyi: a promising target for vaccine development against sea lice, Vaccine 29 (2011) 2810–2820.
[15] J.M. Cregg, T.S. Vedvick, W.C. Raschke, Recent advances in the expression of foreign genes in Pichia pastoris, Biotechnology 11 (1993) 905–910.
[16] G.P. Lin Cereghino, J.M. Cregg, Applications of yeast in biotechnology: pro- tein production and genetic analysis, Curr. Opin. Biotechnol. 10 (1999) 422–427.
[17] S. Macauley-Patrick, M.L. Fazenda, B. McNeil, L.M. Harvey, Heterologous pro- tein production using the Pichia pastoris expression system, Yeast 22 (2005) 249–270.
[18] Invitrogen user’s manuals K1710-01 and K1750-01, http://www.invitrogen. com.
[19] P. Geoff, J. Lin Cereghino, C. Ilgen, J.M. Cregg, Production of recombinant pro- teins in fermenter cultures of the yeast Pichia pastoris, Curr. Opin. Biotechnol. 13 (2002) 329–332.
[20] M. Canales, C. Ballesteros, J.A. Moreno-Cid, A.M. Espinosaa, M. Villar, J. de la Fuente, Extractive bioconversion to produce the Aedes albopictus akirin in an aqueous two-phase system supporting Pichia pastoris growth and protein secretion, Biochem. Eng. J. 46 (2009) 105–114.
[21] G.M. Zijlstra, C.D. de Gooijer, J. Tramper, Extractive bioconversions in aqueous two-phase systems, Curr. Opin. Biotechnol. 9 (1998) 171–176.
[22] A.M. Azevedo, A.G. Gomes, P.A.J. Rosa, I.F. Ferreira, A.M.M.O. Pisco, M.R. Aires- Barros, Partitioning of human antibodies in polyethylene glycol–sodium citrate aqueous two-phase systems, Sep. Purif. Technol. 65 (2009) 14–21.
[23] C.A. Sodré da Silva, J.S.R. Coimbra, E.E.G. Rojas, L.A. Minim, L.H. Mendes da Silva, Partitioning of caseinomacropeptide in aqueous two-phase systems, J. Chromatogr. B 858 (2007) 205–210.
[24] Z.D.V.L. Mayerhoff, I.C. Roberto, T.T. Franco, Purification of xylose reductase from Candida moggi in aqueous two-phase systems, Biochem. Eng. J. 18 (2004) 217–223.
[25] M.G. Antov, Partitioning of pectinase produced by Polyporus squamosus in aque- ous two-phase system polyethylene glycol 4000/crude dextran at different initial pH values, Carbohydr. Polym. 56 (2004) 295–300.
[26] N.D. Srinivas, R.S. Barhate, K.S.M.S. Raghavarao, Aqueous two-phase extrac- tion in combination with ultrafiltration for downstream processing of Ipomoea peroxidise, J. Food Eng. 54 (2002) 1–6.
[27] H.S. Mohamadi, E. Omidinia, Purification of phenylalanine dehydrogenase by partitioning in aqueous two-phase systems, J. Chromatogr. B 854 (2007) 273–278.
[28] M.T. Cunha, M.J.L. Costa, C.R.C. Calado, L.P. Fonseca, M.R. Aires-Barros, J.M.S. Cabral, Integration of production and aqueous two-phase systems extraction of extracellular Fusarium solani pisi cutinase fusion proteins, J. Biotechnol. 100 (2003) 55–64.
[29] N. Kulkarni, A. Vaidya, M. Rao, Extractive cultivation of recombinant Escherichia coli using aqueous two-phase systems for production and sepa- ration of extracellular xylanase, Biochem. Biophys. Res. Commun. 255 (1999) 274–278.
[30] A.S. Schmidt, B.A. Andrews, J.A. Asenjo, Correlations for the partition behaviour of proteins in aqueous two-phase systems: effect of overall protein concentra- tion, Biotechnol. Bioeng. 50 (1996) 617–626.
[31] M. Rito-Palomares, Practical application of aqueous two-phase partition to pro- cess development for the recovery of biological products, J. Chromatogr. B 807 (2004) 3–11.
[32] J. Sinha, P.K. Dey, T. Panda, Extractive fermentation for improved production of endoglucanase by an intergeneric fusant of Trichoderma reesei/Sacharomyces cerevisiae using aqueous two-phase system, Biochem. Eng. J. 6 (2000) 163–175.
[33] Y.J. Kwon, R. Kaul, B. Mattiason, Extractive lactic acid fermentation in poly(ethyleneimine)-based aqueous two-phase system, Biotechnol. Bioeng. 50 (1996) 280–290.
[34] M. Rito-Palomares, A. Lyddiatt, Practical implementation of aqueous two-phase process for protein recovery from yeast, J. Chem. Technol. Biotechnol. 75 (2000) 632–638.
[35] E. Andersson, B. Hahn-Hägerdal, Bioconversions in aqueous two-phase sys- tems, Enzyme Microb. Technol. 12 (1990) 242–254.
[36] H. Hustedt, K.H. Kroner, U. Menge, M.R. Kula, Protein recovery using two-phase systems, Trends Biotechnol. 3 (1985) 139–144.
[37] P.J. Wesgate, A.H. Emery, Approximation of continuous fermentation by semi continuous culture, Biotechnol. Bioeng. 35 (1990) 437–453.
[38] G. Feijoo, C. Dosoretz, J.M. Lema, Production of lignin peroxidase by Phane- rochaete chrysosporium in a packed bed bioreactor operated in semi-continuous mode, J. Biotechnol. 42 (1995) 247–253.
[39] M.E. Bushell, M. Rowe, C.A. Avignone-Rossa, J.N. Wardell, Cyclic fed-batch cul- ture for production of human serum albumin in Pichia pastoris, Biotechnol. Bioeng. 82 (2003) 678–683.
[40] T. Ohya, M. Ohyama, K. Kobayashi, Optimization of human serum albumin production in methylotrophic yeast Pichia pastoris by repeated fed-batch fer- mentation, Biotechnol. Bioeng. 90 (2005) 876–887.
[41] A. Kaul, R.A.M. Pereira, J.A. Asenjo, J.C. Merchuk, Kinetics of phase separation for polyethylene glycol-phosphate two-phase systems, Biotechnol. Bioeng. 48 (1995) 246–256.
[42] J.A. Asenjo, R.E. Turner, S.L. Mistry, A. Kaul, Separation and purification of recombinant proteins from Escherichia coli with aqueous two-phase systems, J. Chromatogr. A 668 (1994) 129–137.
[43] M. Canales, A. Enriquez, E. Ramos, D. Cabrera, H. Dandie, A. Soto, V. Falcon, M. Rodriguez, J. de la Fuente, Large-scale production in Pichia pastoris of the recombinant vaccine GavacTM against cattle ticks, Vaccine 15 (1997) 414–422.
[44] F. Hasko, R. Vazileva, L. Halasz, Solubility of plasma proteins in the presence of polyethylene glycol, Biotechnol. Bioeng. 24 (1982) 1931–1939.
[45] M. Bradford, A rapid and sensitive method for the quantitation of micro- gram quantities of protein utilizing the principle of protein-dye binding, Anal. Biochem. 72 (1976) 248–254.
[46] Y. Chen, J. Krol, J. Cino, D. Freedman, Continuous production of Thrombomod- ulin from a Pichia pastoris fermentation, J. Chem. Technol. Biotechnol. 67 (1996) 143–148.
[47] W. Zhang, C.-P. Liu, M. Inan, M.M. Meagher, Optimization of cell density and dilution rate in Pichia pastoris continuous fermentations for production of recombinant proteins, J. Ind. Microbiol. Biotechnol. 31 (2004) 330–334.
[48] M. Canales, J.A. Moreno-Cid, C. Almazán, M. Villar, J. de la Fuente, Bioprocess design and economics of recombinant BM86/BM95 antigen production for anti- tick vaccines, Biochem. Eng. J. 52 (2010) 79–90.
&KDSWHU�,,


CHAPTER(III.((ANTIGEN(CHARACTERIZATION(
Moreno3Cid!J.A.,!Jiménez!M.,!Sylvie!C.,!Molina!R.,!Alarcón!P.,!Lacroix!M.N.,!Pinal!R.,!Delacour! S.,! Lucientes! J.,! Canales! M.,! Pérez! de! la! Lastra! J.M.,! Villar! M.,! de! la!Fuente! J.! (2010).! Characterization* of* Aedes% albopictus* akirin* for* the* control* of*mosquito*and*sand*fly*infestations.!Vaccine!29(1):!77382.!
Moreno3Cid! J.A.,! Domingos! A.,! Pérez! de! la! Lastra! J.M.,! Canales! M.,! Sánchez! E.,!Díez3Delgado! I.,! Boadella! M.,! Gortázar! C.,! de! la! Fuente! J.! Bacterial* membranes*enhance* the* immunogenicity* of* the* surface* exposed* tick* Subolesin@Anaplasma%marginale%MSP1a*chimeric*antigen.**Journal!of!Biotechnology.*“Submitted”*


CHARACTERIZATION' OF' AEDES( ALBOPICTUS' AKIRIN'
FOR' THE' CONTROL' OF' MOSQUITO' AND' SAND' FLY'
INFESTATIONS'

25!
Contents lists available at ScienceDirect
Vaccine
journal homepage: www.elsevier .com/locate /vaccine
Characterization of Aedes albopictus akirin for the control of mosquito and sand fly infestations
Juan A. Moreno-Cid a , Maribel Jiménez b , Sylvie Cornelie c , Ricardo Molina b , Pedro Alarcón d , Marie-Noelle Lacroix c , Rocío Pinal d , Sarah Delacour d , Javier Lucientes d , Mario Canales a , José M. Pérez de la Lastra a , Margarita Villar a , José de la Fuente a,e,∗
a Instituto de Investigación en Recursos Cinegéticos IREC (CSIC-UCLM-JCCM), Ronda de Toledo s/n, 13005 Ciudad Real, Spain b Unidad de Entomología Médica, Servicio de Parasitología, Centro Nacional de Microbiología, Instituto de Salud Carlos III, 28220 Majadahonda, Madrid, Spain c Caractérisation et Contrôle des Populations de Vecteurs, IRD/UR 016, Montpellier, France d Departamento de Patología Animal, Facultad de Veterinaria, C/Miguel Servet 177, 50013 Zaragoza, Spain e Department of Veterinary Pathobiology, Center for Veterinary Health Sciences, Oklahoma State University, Stillwater, OK 74078, USA
a r t i c l e i n f o a b s t r a c t
Article history: Received 12 May 2010 Received in revised form 24 September 2010 Accepted 6 October 2010 Available online 20 October 2010
Keywords: Akirin Arthropod Mosquito Sand fly Subolesin Tick Vaccine
The control of arthropod vectors of pathogens that affect human and animal health is important for the eradication of vector-borne diseases. Recent evidences showed a reduction in the survival and/or fertility of mosquitoes, sand flies and poultry red mites fed in vitro with antibodies against the recombinant Aedes albopictus akirin. These experiments were the first step toward the development of a multi-target arthro- pod vaccine. In this study, we showed that the oviposition of A. albopictus and Phlebotomus perniciosus fed on mice vaccinated with recombinant A. albopictus akirin was reduced by 17% and 31%, respectively when compared to controls. However, Aedes aegypti mosquitoes were not affected after feeding on vaccinated mice. These results showed that recombinant A. albopictus akirin could be used to vaccinate hosts for the control of mosquito and sand fly infestations and suggested new experiments to develop improved vaccine formulations.
© 2010 Elsevier Ltd. All rights reserved.
1. Introduction
Diseases caused by vector-borne pathogens greatly impacthuman and animal health, accounting for over 20% of all emerg- ing infectious diseases recorded between 1940 and 2004 [1]. In particular, insects such as mosquitoes and sand flies are consid- ered the most important vectors of human diseases worldwide [1]. Aedes spp. (Diptera: Culicidae) are vectors of many viruses that cause human disease, including the exposure of over 2.5 billion people in tropical and subtropical countries to dengue fever virus [2]. Phlebotomine sand flies (Diptera: Psychodidae) are also vec- tors of several pathogens including Leishmania spp., which causes various forms of human leishmaniasis [3].
With the exception of a few diseases such as yellow fever, vaccines against vector-transmitted pathogens have not been
∗ Corresponding author at: Instituto de Investigación en Recursos CinegéticosIREC (CSIC-UCLM-JCCM), Ronda de Toledo s/n, 13005 Ciudad Real, Spain.
E-mail addresses: jose [email protected], [email protected], [email protected] (J. de la Fuente).
successfully developed nor implemented, and intense use of insec- ticides and/or chemotherapy have resulted in an increasing number of insecticide-resistant vectors and drug-resistant pathogens [4–6].
The effect of anti-tick vaccines on the reduction of tick infes- tations and the transmission of some tick-borne pathogens [4,7,8] and preliminary results obtained in insect vector species [9–17] have provided evidence that protective antigens may be used for development of vaccines with the dual target control of both arthropod infestations and reduction of vector capacity to transmit pathogens that impact human and animal health.
Subolesin, the ortholog of akirin in ticks, was discovered as a tick protective antigen in Ixodes scapularis [18]. Subolesin was shown by RNAi gene knockdown and immunization trials using the recombinant protein to protect hosts against tick infesta- tions, reduce tick survival and reproduction, cause degeneration of guts, salivary glands, reproductive tissues and embryos and to decrease the vector capacity of ticks for Anaplasma marginale and A. phagocytophilum [18–24]. In addition, subolesin was shown to be evolutionary conserved and similar in structure and function to insect and vertebrate akirins, which control NF-kB-dependent
0264-410X/$ – see front matter © 2010 Elsevier Ltd. All rights reserved. doi:10.1016/j.vaccine.2010.10.011
&KDSWHU�,,,

26!
and independent gene expression that impact innate immunity [21,25–28].
Recent evidences showed a reduction in the survival and/or fertility of mosquitoes, sand flies and poultry red mites fed in vitro with antibodies against the recombinant Aedes albopictus akirin [17,29]. Vaccination with recombinant A. albopictus akirin also reduced tick infestations [17,30]. These results suggested the experiments reported here to characterize the effect of A. albopictus akirin for the control of mosquito and sand fly infestations in mice vaccinated with the recombinant protein. These results support the role of mosquito akirin as a protective antigen for the control of mosquito and sand fly infestations.
2. Materials and methods
2.1. Mosquitoes and sand flies Mosquitoes, A. albopictus, from the Baix Llobregat (Barcelona,
Spain) colony were used to establish a colony at the University of Zaragoza and used for this study. The A. albopictus strain has been maintained in the laboratory for approximately one year (12 gen- erations). Mosquitoes were reared in an environmental cabinet at 25 ± 1 ◦C, 75 ± 5% relative humidity (RH), and a 12 h light:12 h dark photoperiod.
Mosquitoes, Aedes aegypti, Bora-Bora reference strain originat- ing from Bora-Bora (French Polynesia) were used for this study. The A. aegypti strain has been maintained in the laboratory for 12 years (approximately 150 generations). Mosquitoes were main- tained in the IRD-Monpellier insectary under controlled conditions of 27 ± 2 ◦C, 80% RH and a 17 h light:7 h dark photoperiod.
Sand flies, Phlebotomus perniciosus, used in this study were from an autochthonous (Madrid Province, Spain) colony established at the Medical Entomology Unit of the Instituto de Salud Carlos III (Spain). The P. perniciosus strain has been maintained in the labo- ratory for approximately 161 generations. Flies were reared in an environmental cabinet at 28 ◦C, 95–100% RH, and a 17 h light:7 h dark photoperiod.
For all insect species, females were mixed with males after emergence until females were selected for the experiment. Under our rearing conditions, most of the insects mate as demonstrated by ovaries dissection and examination in previous experiments. After emergence, insects were supplied absorbent cotton saturated with 20–30% sucrose solution during four days. At day five, they were starved 5 h before mice infestation [31]. Then, females were main- tained in new cages without males until the end of the experiment.
2.2. Vaccine formulation The expression and purification of recombinant A. albopictus
akirin was conducted as previously reported using an extractive bioconversion process in an aqueous two-phase system support- ing Pichia pastoris growth and protein secretion [32]. Recombinant antigen or saline were adjuvated in Montanide ISA 50 V2 (Seppic, Paris, France).
2.3. Mouse vaccination and infestation Three experiments were conducted to evaluate the effect of
recombinant A. albopictus akirin vaccination on mice infested with A. albopictus, A. aegypti and P. perniciosus. These experiments were independently conducted at the University of Zaragoza (Spain), the Centre IRD de Montpellier (France) and the Instituto de Salud Carlos III (Spain), respectively. All mice were immunized three times at weeks 0, 3 (4 for the experiment with A. aegypti), and 6 with 1 ml (0.2 ml for the experiment with P. perniciosus) doses injected subcutaneously in the dorsum near the base of the tail
using a 1-ml tuberculin syringe and a 27.5-G needle. Vaccinated and control mice were injected with recombinant A. albopictus akirin (50 µg/dose) and adjuvant/saline, respectively. Immuniza- tions, insect collections and evaluations were done blinded and the key to the experimental groups was not opened until the end of the experiment. Animals were cared for in accordance with standards specified in the Guide for Care and Use of Laboratory Animals and approved by ethical committees for animal care and experimenta- tion.
2.4. Vaccination against A. albopictus mosquito infestations
Five female 5 weeks old Balb/c mice were used for each of the vaccinated and control groups. Each mouse was restrained in a nylon cage, and exposed to A. albopictus bites at week 8 of the exper- iment. Twenty female mosquitoes aged 3–5 days old were fed on each mouse during 30 min. After exposure, fed females were trans- ferred to individual vials for oviposition. Eggcups were removed seven days post-infestation and eggs were counted. The mortality rate seven days post-infestation was also evaluated. Four weeks after the last immunization, blood samples were collected from each mouse.
2.5. Vaccination against A. aegypti mosquito infestations Four female 8–10 weeks old Balb/c mice were used for each of
the vaccinated and control groups. Infestations were done at week 8. Each mouse was restrained in a perforated tube, placed on awarming plate and exposed to A. aegypti bites during 15 min. Blood samples were collected approximately 48 h before each immuniza- tion and infestation, and seven days after infestation by saphenous vein or retro-orbital bleeding. Thirty female mosquitoes were fed on each mouse during 15 min. An oviposition cup was placed into each cage and females were allowed to oviposit. Eggcups were removed seven days post-infestation and eggs were counted. The mortality rate seven days post-infestation was also evaluated.
2.6. Vaccination against sand fly infestations Five female 6–8 weeks old CD-1 mice were used for each of
the vaccinated and control groups. The effect on mortality and fertil- ity of P. perniciosus sand flies fed on vaccinated and control mice was evaluated. Two weeks after the last immunization, 25 female sand flies were fed for 20–30 min on each mouse anaesthetized with a blend of ketamine (100 mg/kg) and xylazine (10 mg/kg) by intraperitoneal injection [33]. After sand fly exposure, 10 fed females per group were transferred to individual flasks for analy- sis of oviposition. Eggs from females fed on the same mouse were transferred to a pot for larval rearing to study the development to adult flies as described previously [17]. Four weeks after the last immunization, blood samples were collected from each mouse.
2.7. Characterization of the immune response in immunized mice by ELISA
Serum was separated in blood samples by centrifugation and individually stored at −20 ◦C. An indirect ELISA test was per- formed to detect antibodies against akirin in mouse serum samples. High absorption capacity polystyrene microtiter plates were coated with 50 µl (0.02 µg/ml solution of purified akirin; [32]) per well in carbonate–bicarbonate buffer (Sigma, Barcelona, Spain). After an overnight incubation at 4 ◦C, coated plates were blocked with 200 µl/well of blocking solution (5% skim milk in PBS). Serum sam- ples or PBS as negative control were diluted (1:20, v/v) in blocking solution and 100 µl/well were added into duplicate wells of the antigen-coated plates. After a 1 h incubation period at RT, the plates were washed three times with a washing solution (PBS contain- ing 0.05% Tween 20). A goat anti-mouse IgG-peroxidase conjugate
&KDSWHU�,,,
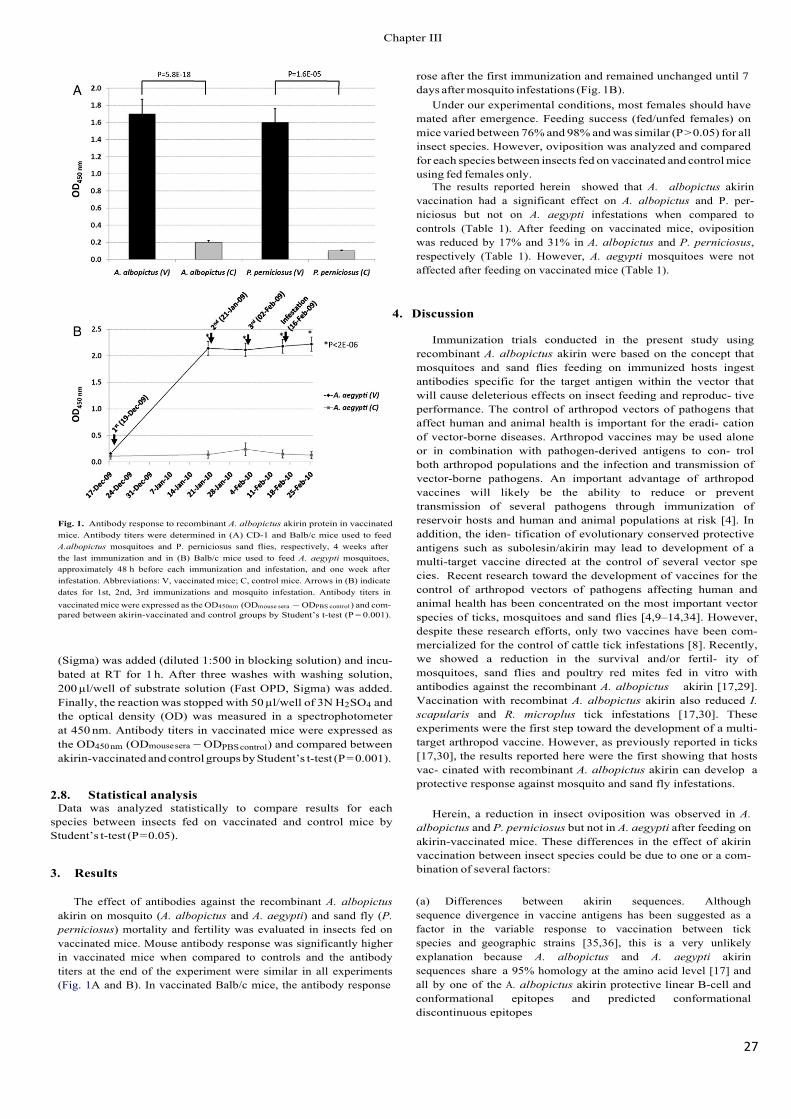
27!
rose after the first immunization and remained unchanged until 7 days after mosquito infestations (Fig. 1B).
Under our experimental conditions, most females should have mated after emergence. Feeding success (fed/unfed females) on mice varied between 76% and 98% and was similar (P > 0.05) for all insect species. However, oviposition was analyzed and compared for each species between insects fed on vaccinated and control mice using fed females only.
The results reported herein showed that A. albopictus akirin vaccination had a significant effect on A. albopictus and P. per- niciosus but not on A. aegypti infestations when compared to controls (Table 1). After feeding on vaccinated mice, oviposition was reduced by 17% and 31% in A. albopictus and P. perniciosus, respectively (Table 1). However, A. aegypti mosquitoes were not affected after feeding on vaccinated mice (Table 1).
4. Discussion
Fig. 1. Antibody response to recombinant A. albopictus akirin protein in vaccinated mice. Antibody titers were determined in (A) CD-1 and Balb/c mice used to feed A.albopictus mosquitoes and P. perniciosus sand flies, respectively, 4 weeks after the last immunization and in (B) Balb/c mice used to feed A. aegypti mosquitoes, approximately 48 h before each immunization and infestation, and one week after infestation. Abbreviations: V, vaccinated mice; C, control mice. Arrows in (B) indicate dates for 1st, 2nd, 3rd immunizations and mosquito infestation. Antibody titers in vaccinated mice were expressed as the OD450nm (ODmouse sera − ODPBS control ) and com- pared between akirin-vaccinated and control groups by Student’s t-test (P = 0.001).
(Sigma) was added (diluted 1:500 in blocking solution) and incu- bated at RT for 1 h. After three washes with washing solution, 200 µl/well of substrate solution (Fast OPD, Sigma) was added. Finally, the reaction was stopped with 50 µl/well of 3N H2SO4 and the optical density (OD) was measured in a spectrophotometer at 450 nm. Antibody titers in vaccinated mice were expressed as the OD450 nm (ODmouse sera − ODPBS control) and compared between akirin-vaccinated and control groups by Student’s t-test (P = 0.001).
2.8. Statistical analysis Data was analyzed statistically to compare results for each
species between insects fed on vaccinated and control mice by Student’s t-test (P = 0.05).
3. Results
The effect of antibodies against the recombinant A. albopictusakirin on mosquito (A. albopictus and A. aegypti) and sand fly (P. perniciosus) mortality and fertility was evaluated in insects fed on vaccinated mice. Mouse antibody response was significantly higher in vaccinated mice when compared to controls and the antibody titers at the end of the experiment were similar in all experiments (Fig. 1A and B). In vaccinated Balb/c mice, the antibody response
Immunization trials conducted in the present study using recombinant A. albopictus akirin were based on the concept that mosquitoes and sand flies feeding on immunized hosts ingest antibodies specific for the target antigen within the vector that will cause deleterious effects on insect feeding and reproduc- tive performance. The control of arthropod vectors of pathogens that affect human and animal health is important for the eradi- cation of vector-borne diseases. Arthropod vaccines may be used alone or in combination with pathogen-derived antigens to con- trol both arthropod populations and the infection and transmission of vector-borne pathogens. An important advantage of arthropod vaccines will likely be the ability to reduce or prevent transmission of several pathogens through immunization of reservoir hosts and human and animal populations at risk [4]. In addition, the iden- tification of evolutionary conserved protective antigens such as subolesin/akirin may lead to development of a multi-target vaccine directed at the control of several vector spe�cies. 5HFHQW�research toward the development of vaccines for the control of arthropod vectors of pathogens affecting human and animal health has been concentrated on the most important vector species of ticks, mosquitoes and sand flies [4,9–14,34]. However, despite these research efforts, only two vaccines have been com- mercialized for the control of cattle tick infestations [8]. Recently, we showed a reduction in the survival and/or fertil- ity of mosquitoes, sand flies and poultry red mites fed in vitro with antibodies against the recombinant A. albopictus akirin [17,29]. Vaccination with recombinat A. albopictus akirin also reduced I. scapularis and R. microplus tick infestations [17,30]. These experiments were the first step toward the development of a multi-target arthropod vaccine. However, as previously reported in ticks [17,30], the results reported here were the first showing that hosts vac- cinated with recombinant A. albopictus akirin can develop a protective response against mosquito and sand fly infestations.
Herein, a reduction in insect oviposition was observed in A. albopictus and P. perniciosus but not in A. aegypti after feeding on akirin-vaccinated mice. These differences in the effect of akirin vaccination between insect species could be due to one or a com- bination of several factors:
(a) Differences between akirin sequences. Although sequence divergence in vaccine antigens has been suggested as a factor in the variable response to vaccination between tick species and geographic strains [35,36], this is a very unlikely explanation because A. albopictus and A. aegypti akirin sequences share a 95% homology at the amino acid level [17] and all by one of the A. albopictus akirin protective linear B-cell and conformational epitopes and predicted conformational discontinuous epitopes
&KDSWHU�,,,
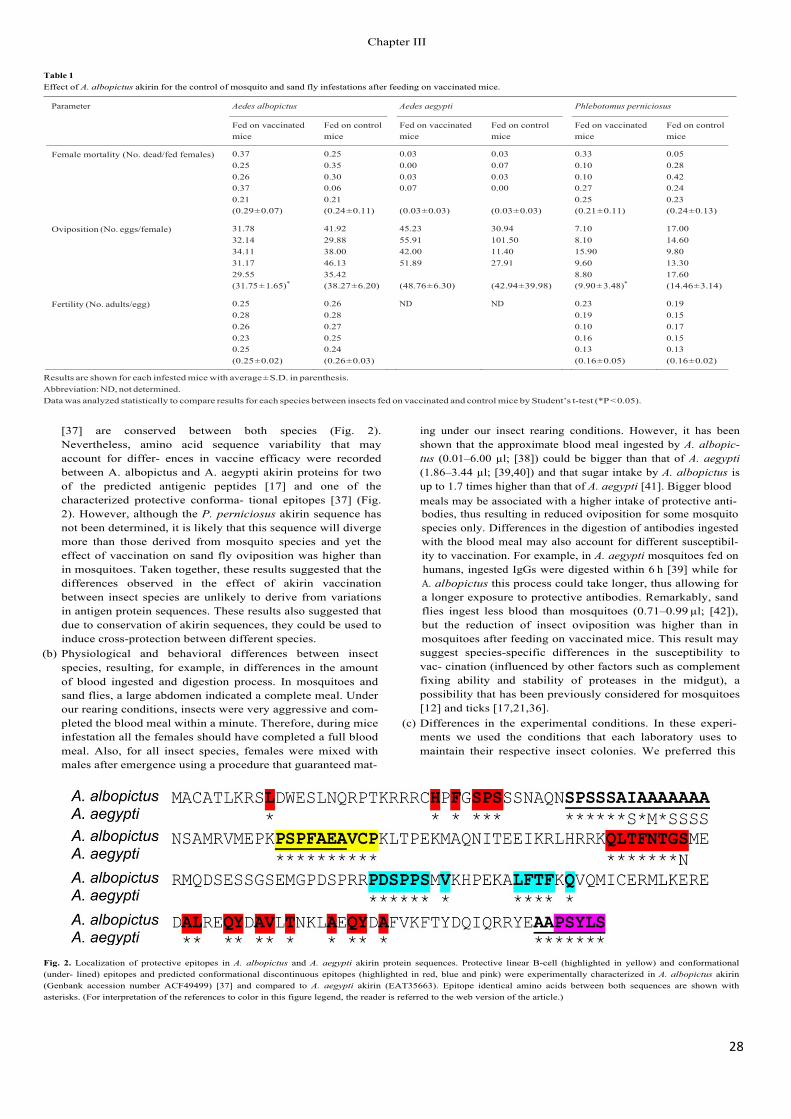
28!
Table 1 Effect of A. albopictus akirin for the control of mosquito and sand fly infestations after feeding on vaccinated mice.
Parameter Aedes albopictus Aedes aegypti Phlebotomus perniciosus
Fed on vaccinated mice
Fed on control mice
Fed on vaccinated mice
Fed on control mice
Fed on vaccinated mice
Fed on control mice
Female mortality (No. dead/fed females) 0.37 0.25 0.03 0.03 0.33 0.05 0.25 0.35 0.00 0.07 0.10 0.28 0.26 0.30 0.03 0.03 0.10 0.42 0.37 0.06 0.07 0.00 0.27 0.24 0.21 0.21 0.25 0.23 (0.29 ± 0.07) (0.24 ± 0.11) (0.03 ± 0.03) (0.03 ± 0.03) (0.21 ± 0.11) (0.24 ± 0.13)
Oviposition (No. eggs/female) 31.78 41.92 45.23 30.94 7.10 17.00 32.14 29.88 55.91 101.50 8.10 14.60 34.11 38.00 42.00 11.40 15.90 9.80 31.17 46.13 51.89 27.91 9.60 13.30 29.55 (31.75 ± 1.65)*
35.42 (38.27 ± 6.20) (48.76 ± 6.30) (42.94 ± 39.98)
8.80 (9.90 ± 3.48)*
17.60 (14.46 ± 3.14)
Fertility (No. adults/egg) 0.25 0.26 ND ND 0.23 0.19 0.28 0.28 0.19 0.15 0.26 0.27 0.10 0.17 0.23 0.25 0.16 0.15 0.25 0.24 0.13 0.13 (0.25 ± 0.02) (0.26 ± 0.03) (0.16 ± 0.05) (0.16 ± 0.02)
Results are shown for each infested mice with average ± S.D. in parenthesis. Abbreviation: ND, not determined. Data was analyzed statistically to compare results for each species between insects fed on vaccinated and control mice by Student’s t-test (*P < 0.05).
[37] are conserved between both species (Fig. 2). Nevertheless, amino acid sequence variability that may account for differ- ences in vaccine efficacy were recorded between A. albopictus and A. aegypti akirin proteins for two of the predicted antigenic peptides [17] and one of the characterized protective conforma- tional epitopes [37] (Fig. 2). However, although the P. perniciosus akirin sequence has not been determined, it is likely that this sequence will diverge more than those derived from mosquito species and yet the effect of vaccination on sand fly oviposition was higher than in mosquitoes. Taken together, these results suggested that the differences observed in the effect of akirin vaccination between insect species are unlikely to derive from variations in antigen protein sequences. These results also suggested that due to conservation of akirin sequences, they could be used to induce cross-protection between different species.
(b) Physiological and behavioral differences between insect species, resulting, for example, in differences in the amount of blood ingested and digestion process. In mosquitoes and sand flies, a large abdomen indicated a complete meal. Under our rearing conditions, insects were very aggressive and com- pleted the blood meal within a minute. Therefore, during mice infestation all the females should have completed a full blood meal. Also, for all insect species, females were mixed with males after emergence using a procedure that guaranteed mat-
ing under our insect rearing conditions. However, it has been shown that the approximate blood meal ingested by A. albopic- tus (0.01–6.00 µl; [38]) could be bigger than that of A. aegypti (1.86–3.44 µl; [39,40]) and that sugar intake by A. albopictus is up to 1.7 times higher than that of A. aegypti [41]. Bigger blood meals may be associated with a higher intake of protective anti- bodies, thus resulting in reduced oviposition for some mosquito species only. Differences in the digestion of antibodies ingested with the blood meal may also account for different susceptibil- ity to vaccination. For example, in A. aegypti mosquitoes fed on humans, ingested IgGs were digested within 6 h [39] while for A. albopictus this process could take longer, thus allowing for a longer exposure to protective antibodies. Remarkably, sand flies ingest less blood than mosquitoes (0.71–0.99 µl; [42]), but the reduction of insect oviposition was higher than in mosquitoes after feeding on vaccinated mice. This result may suggest species-specific differences in the susceptibility to vac- cination (influenced by other factors such as complement fixing ability and stability of proteases in the midgut), a possibility that has been previously considered for mosquitoes [12] and ticks [17,21,36].
(c) Differences in the experimental conditions. In these experi- ments we used the conditions that each laboratory uses to maintain their respective insect colonies. We preferred this
Fig. 2. Localization of protective epitopes in A. albopictus and A. aegypti akirin protein sequences. Protective linear B-cell (highlighted in yellow) and conformational (under- lined) epitopes and predicted conformational discontinuous epitopes (highlighted in red, blue and pink) were experimentally characterized in A. albopictus akirin (Genbank accession number ACF49499) [37] and compared to A. aegypti akirin (EAT35663). Epitope identical amino acids between both sequences are shown with asterisks. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of the article.)
&KDSWHU�,,,

29!
option, instead of trying to homogenize conditions for all tri- als, to prevent an effect of changing conditions used at each laboratory to maintain insect colonies that may affect insect mortality and fitness and will be unrelated to akirin vaccina- tion. Additionally, for each experiment, a mice control group injected with adjuvant/saline alone was used to compare the results between vaccinated and control groups, those allowing comparison between insects raised under similar conditions. However, some variations in the experimental conditions used here could have contributed to the differences in the effect of akirin vaccination between insect species. For example, the out- bred mouse strain, CD-1, was used in the experiments with A. albopictus and P. perniciosus, while inbred Balb/c mice were used to feed A. aegypti. Although the antibody levels obtained after vaccination were similar between CD-1 and Balb/c mice, the protective response to vaccination (for example in the IgG1/IgG2 ratio; [43]) may be genetically restricted [44]. In the previous experiment in which mosquitoes and sand flies were fed in vitro with anti-A. albopictus rabbit antibodies, increased insect mortality was recorded [17]. However, in the experi- ments reported here, insect mortality was not affected after feeding on vaccinated mice, a result similar to previous reports in mosquitoes [12]. This difference could be attributed to differences in insect susceptibility to anti-akirin antibodies when fed in vitro and in vivo (due, for example, to a higher antibody con- centration when fed in vitro) and/or differences in the antibody response elicited by rabbits and mice. In previous experiments we have shown that rabbit and mouse antibodies may recog- nize different epitopes in the protein antigen after vaccination�>��@Antibodies against intracellular proteins such as sub- olesin/akirin have been effective against ticks, mosquitoes and other invertebrate organisms [7,17–21,29,30,46]. The protection elicited by anti-tick vaccines [7,8,34,47], and likely by other anti-arthropod vaccines [10,13,48], is based on the production of antibodies in vaccinated hosts that interact with and affect the function of the target antigen in feeding arthropods. For subolesin/akirin, the mode of action of the antibodies is not completely understood but may be the result of one or a combination of several factors such as inhibiting feeding, digestion, metabolism, immune response, tissue development and function, reproduction and embryo GHYHdevelopment [20–24,28]. The anti-insect effects achieved so far by us and others [10–13,17,49,50] are small when compared to anti-tick vaccines [4,7,8,18,19,30,34,36,47]. However, this fact can be explained by differences in the digestive physiology of ticks and mosquitoes as well as time spent on the host. Although the size of the blood meal and digestion time varies between different blood-sucking arthropods being much higher in ticks than in mosquitoes and sand flies, several results demonstrate that even with small amounts of antibodies ingested, their effect is of sufficient duration to reduce vector survival and/or fitness [10,17]. In fact, the reduction in insect oviposition obtained here for A. albopictus (17%) and P. perniciosus (31%) fed on mice vaccinated with recombinant A. albopictus akirin was within the range reported previously in Anopheles stephensi mosquitoes fed with midgut antibodies [12]. The mechanism by which antibodies are transported across arthropod cell membranes and interact with antigens is not completely understood. However, antibodies are specifically trans- ported across gut barrier into the hemolymph and then inside arthropod cells so they can interact with intracellular proteins [50–56]. As discussed previously [17], the conservation of akirin sequences among arthropod vectors and vertebrate hosts may raise the question of safety when using akirin for immunization with
the potential of inducing autoimmune responses damaging to the host. However, it is expected that the antibody response would be primarily directed against non-self epitopes thus reducing the possibility of detrimental effects to the host. Additionally, immu- nization with intracellular proteins such as akirin/subolsin has been effective in ticks and other invertebrate organisms and suggests a low risk to induce autoimmune responses in vertebrate hosts [18,19,21,46].
In summary, the results reported here showed that recombinant A. albopictus akirin could be used to vaccinate hosts for the con- trol of mosquito and sand fly infestations. Reduction in oviposition would result in the reduction of insect populations in subsequent generations, as previously shown for anti-tick vaccines [8]. If effective, the vaccine could be used to immunize reservoir hosts to reduce vector populations and thus the risk of pathogen transmission to humans and animals. However, although obtained results are promising for development of an anti-arthropod vector akirin vaccine, several questions remain to be addressed. These questions include characterization of vaccine efficacy using a larger number of vaccinated and infested individuals, differences in vaccine susceptibility between different arthropod species and in host response to vaccination that may require the inclusion of multiple akirin anti- gens and development of improved vaccine formulations. Finally, as shown for tick subolesin [22], the effect of akirin vaccination on pathogen infection and transmission needs to be tested.
Acknowledgments
Diego Ayala is acknowledged for valuable suggestions. This research was supported by the Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (INIA), Spain (project FAU2008- 00014-00-00) and the Consejería de Educación y Ciencia, JCCM, Spain (project PEII09-0118-8907). J.A. Moreno-Cid is a recipient of a JCCM fellowship. M. Canales was funded by the Wellcome Trust under the “Animal Health in the Developing World” initiative (project 0757990). M. Villar was funded by the JAE-DOC program (CSIC-FSE), Spain.
References
[1] Jones KE, Patel NG, Levy MA, Storeygard A, Balk D, Gittleman JL, et al. Global trends in emerging infectious diseases. Nature 2008;451:990–4.
[2] Halstead SB. Dengue. Lancet 2007;370:1644–52. [3] Chappuis F, Sundar S, Hailu A, Ghalib H, Rijal S, Peeling RW, et al. Visceral
leishmaniasis: what are the needs for diagnosis, treatment and control? Nat Rev Microbiol 2007;5:873–82.
[4] de la Fuente J, Kocan KM. Advances in the identification and characterization of protective antigens for development of recombinant vaccines against tick infestations. Expert Rev Vaccines 2003;2:583–93.
[5] Kishore K, Kumar V, Kesari S, Dinesh DS, Kumar AJ, Das P, et al. Vector control in leishmaniasis. Indian J Med Res 2006;123:467–72.
[6] Speranc a MA, Capurro ML. Perspectives in the control of infectious diseases by transgenic mosquitoes in the post-genomic era – a review. Mem Inst Oswaldo Cruz 2007;102:425–33.
[7] de la Fuente J, Rodríguez M, Redondo M, Montero C, García-García JC, Méndez L, et al. Field studies and cost-effectiveness analysis of vaccination with GavacTM
against the cattle tick Boophilus microplus. Vaccine 1998;16:366–73. [8] de la Fuente J, Almazán C, Canales M, Pérez de la Lastra JM, Kocan KM, Willadsen
P. A ten-year review of commercial vaccine performance for control of tick infestations on cattle. Anim Health Res Rev 2007;8:23–8.
[9] Valenzuela JG, Belkaid Y, Garfield MK, Mendez S, Kamhawi S, Rowton ED, et al. Toward a defined anti-Leishmania vaccine targeting vector antigens: char- acterization of a protective salivary protein. J Exp Med 2001;194:331–42.
[10] Lal AA, Patterson PS, Sacci JB, Vaughan JA, Paul C, Collins WE, et al. Anti- mosquito midgut antibodies block development of Plasmodium falciparum and Plasmodium vivax in multiple species of Anopheles mosquitoes and reduce vec- tor fecundity and survivorship. Proc Natl Acad Sci USA 2001;98:5228–33.
[11] Almeida AP, Billingsley PF. Induced immunity against the mosquito Anophe- les stephensi (Diptera: Culicidae): effects of cell fraction antigens on survival, fecundity, and Plasmodium berghei (Eucoccidiida: Plasmodiidae) transmission. J Med Entomol 2002;39:207–14.
[12] Suneja A, Gulia M, Gakhar SK. Blocking of malaria parasite development in mosquito and fecundity reduction by midgut antibodies in Anopheles stephensi (Diptera: Culicidae). Arch Insect Biochem Physiol 2003;52:63–70.
&KDSWHU�,,,

30!
[13] Milleron RS, Ribeiro JM, Elnaime D, Soong L, Lanzaro G. Negative effect of anti- bodies against maxadilan on the fitness of the sand fly vector of American visceral leishmaniasis. Am J Trop Med Hyg 2004;70:278–85.
[14] Titus RG, Bishop JV, Mejia JS. The immunomodulatory factors of arthropod saliva and the potential for these factors to serve as vaccine targets to prevent pathogen transmission. Parasite Immunol 2006;4:131–41.
[15] Kedzierski L, Zhu Y, Handman E. Leishmania vaccines: progress and problems. Parasitol 2006;133:S87–112.
[16] Saul A. Mosquito stage, transmission blocking vaccines for malaria. Curr Opin Infect Dis 2007;5:476–81.
[17] Canales M, Naranjo V, Almazán C, Molina R, Tsuruta SA, Szabó MPJ, et al. Con- servation and immunogenicity of the mosquito ortholog of the tick protective antigen, subolesin. Parasitol Res 2009;105:97–111.
[18] Almazán C, Blas-Machado U, Kocan KM, Yoshioka JH, Blouin EF, Mangold AJ, et al. Characterization of three Ixodes scapularis cDNAs protective against tick infestations. Vaccine 2005;23:4403–16.
[19] Almazán C, Kocan KM, Blouin EF, de la Fuente J. Vaccination with recombi- nant tick antigens for the control of Ixodes scapularis adult infestations. Vaccine 2005;23:5294–8.
[20] de la Fuente J, Almazán C, Naranjo V, Blouin EF, Meyer JM, Kocan KM. Autocidal control of ticks by silencing of a single gene by RNA interference. Biochem Biophys Res Commun 2006;344:332–8.
[21] de la Fuente J, Almazán C, Blas-Machado U, Naranjo V, Mangold AJ, Blouin EF, et al. The tick protective antigen, 4D8, is a conserved protein involved in mod- ulation of tick blood ingestion and reproduction. Vaccine 2006;24:4082–95.
[22] de la Fuente J, Almazán C, Blouin EF, Naranjo V, Kocan KM. Reduction of tick infections with Anaplasma marginale and A. phagocytophilum by targeting the tick protective antigen subolesin. Parasitol Res 2006;100:85–91.
[23] Kocan KM, Manzano-Roman R, de la Fuente J. Transovarial silencing of the sub- olesin gene in three-host ixodid tick species after injection of replete females with subolesin dsRNA. Parasitol Res 2007;100:1411–5.
[24] Nijhof AM, Taoufik A, de la Fuente J, Kocan KM, de Vries E, Jongejan F. Gene silencing of the tick protective antigens, Bm86, Bm91 and subolesin, in the one-host tick Boophilus microplus by RNA interference. Int J Parasitol 2007;37:653–62.
[25] de la Fuente J, Maritz-Olivier C, Naranjo V, Ayoubi P, Nijhof AM, Almazán C, et al. Evidence of the role of tick subolesin in gene expression. BMC Genomics 2008;9:372.
[26] Galindo RC, Doncel-Pérez E, Zivkovic Z, Naranjo V, Gortazar C, Mangold AJ, et al. Tick subolesin is an ortholog of the akirins described in insects and vertebrates. Dev Comp Immunol 2009;33:612–7.
[27] Mangold AJ, Galindo RC, de la Fuente J. Response to the commentary of D. Macqueen on: Galindo RC, Doncel-Pérez E, Zivkovic Z, Naranjo V, Gortazar C, Mangold AJ, et al. Tick subolesin is an ortholog of the akirins described in insects and vertebrates [Dev. Comp. Immunol. 33 (2009) 612–617]. Dev Comp Immunol 2009;33:878–9.
[28] Zivkovic Z, Torina A, Mitra R, Alongi A, Scimeca S, Kocan KM, et al. Subolesin expression in response to pathogen infection in ticks. BMC Immunol 2010;11:7.
[29] Harrington D, Canales M, de la Fuente J, de Luna C, Robinson K, Guy J, et al. Immunisation with recombinant proteins subolesin and Bm86 for the control of Dermanyssus gallinae in poultry. Vaccine 2009;27:4056–63.
[30] Canales M, Labruna MB, Soares JF, Prudencio CR, de la Fuente J. Protective efficacy of bacterial membranes containing surface-exposed BM95 antigenic peptides for the control of cattle tick infestations. Vaccine 2009;27:7244–8.
[31] Molina R. Laboratory adaptation of an autochthonous colony of Phleboto- mus perniciosus Newstead, 1911 (Diptera: Psychodidae). Res Rev Parasitol 1991;51:87–9.
[32] Canales M, Ballesteros C, Moreno-Cid JA, Espinosa AM, Villar M, de la Fuente J. Extractive bioconversion to produce the Aedes albopictus akirin in an aque- ous two-phase system supporting Pichia pastoris growth and protein secretion. Biochem Eng J 2009;46:105–14.
[33] Molina R, Amela C, Nieto J, San-Andrés M, González F, Castillo JA, et al. Infectivity of dogs naturally infected with Leishmania infantum to colonized Phlebotomus perniciosus. Trans R Soc Trop Med Hyg 1994;88:491–3.
[34] Willadsen P. Anti-tick vaccines. Parasitol 2004;129:S367–874.
[35] García-García JC, González IL, González DM, Valdés M, Méndez L, Lamberti J, et al. Sequence variations in the Boophilus microplus Bm86 locus and implications for immunoprotection in cattle vaccinated with this antigen. Exp Appl Acarol 1999;23:883–95.
[36] Canales M, Almazán C, Naranjo V, Jongejan F, de la Fuente J. Vaccination with recombinant Boophilus annulatus Bm86 ortholog protein, Ba86, protects cattle against B. annulatus and B. microplus infestations. BMC Biotechnol 2009;9:29.
[37] Prudencio CR, Pérez de la Lastra JM, Canales M, Villar M, de la Fuente J. Map- ping protective epitopes in the tick and mosquito subolesin ortholog proteins. Vaccine 2010;28:5398–406.
[38] Konishi E. Size of blood meals of Aedes albopictus and Culex tritaeniorhynchus (Diptera: Culicidae) feeding on an unrestrained dog infected with Dirofilaria immitis (Spirurida: Filariidae). J Med Entomol 1989;26:535–8.
[39] Chen WJ, Chen ER. Fate of ingested immunoglobulin G in the mosquito Aedes aegypti (Diptera: Culicidae). Gaoxiong Yi Xue Ke Xue Za Zhi 1990;6:606–13.
[40] Knaus RM, Foil LD, Issel CJ, Leprince DJ. Insect blood meal studies using radiosodium 24Na and 22Na. J Am Mosq Control Assoc 1993;9:264–8.
[41] Dutra Nunes R, Lourenc o de Oliveira R, Cardoso Braz GR. A novel method for measuring fructose ingestion by mosquitoes. J Vector Ecol 2008;33:225–31.
[42] Daba S, Daba A, Shehata MG, El Sawaf BM. A simple micro-assay method for estimating blood meal size of the sand fly, Phlebotomus langeroni (Diptera: Psychodidae). J Egypt Soc Parasitol 2004;34:173–82.
[43] Rodríguez M, Montero C, Machado H, Joglar M, de la Fuente J, Garcia-Garcia JC. The evaluation of yeast derivatives as adjuvants for the immune response to the Bm86 antigen in cattle. BMC Biotechnol 2001;1:2 [Erratum in BMC Biotechnol 2001;1:3].
[44] Jaffe RI, Lowell GH, Gordon DM. Differences in susceptibility among mouse strains to infection with Plasmodium berghei (ANKA clone) sporozoites and its relationship to protection by gamma-irradiated sporozoites. Am J Trop Med Hyg 1990;42:309–13.
[45] Garcia-Garcia JC, de la Fuente J, Kocan KM, Blouin EF, Halbur T, Onet VC, et al. Mapping of B-cell epitopes in the N-terminal repeated peptides of Anaplasma marginale major surface protein 1a and characterization of the humoral immune response of cattle immunized with recombinant and whole organism antigens. Vet Immunol Immunopathol 2004;98:137–51.
[46] Elad D, Segal E. Immunogenicity in calves of a crude ribosomal fraction of Trichophyton verrucosum: a field trial. Vaccine 1995;13:83–7.
[47] Trimnell AR, Davies GM, Lissina O, Hails RS, Nuttall PA. A cross-reactive tick cement antigen is a candidate broad-spectrum tick vaccine. Vaccine 2005;23:4329–41.
[48] Tellam RL, Eisemann CH, Vuocolo T, Casu R, Jarmey J, Bowles V, et al. Role of oligosaccharides in the immune response of sheep vaccinated with Lucilia cuprina larval glycoprotein, peritrophin-95. Int J Parasitol 2001;31:798–809.
[49] Alger NE, Cabrera EJ. An increase in death rate of Anopheles stephensi fed on rabbit immunized with mosquito antigen. J Econ Entomol 1972;65:165–8.
[50] Hatfield PR. Detection and localization of antibody ingested with a mosquito bloodmeal. Med Vet Entomol 1988;2:339–45.
[51] Chinzei Y, Minoura H. Host immunoglobulin G titre and antibody activ- ity in haemolymph of the tick, Ornithodoros moubata. Med Vet Entomol 1987;1:409–16.
[52] Lackie AM, Gavin S. Uptake and persistence of ingested antibody in the mosquito Anopheles stephensi. Med Vet Entomol 1989;3:225–30.
[53] Eisemann CH, Pearson RD, Donaldson RA, Cadogan LC, Vuocolo T. Uptake and fate of specific antibody in feeding larvae of the sheep blowfly, Lucilia cuprina. Med Vet Entomol 1993;7:177–85.
[54] Vaz Junior Ida S, Martinez RH, Oliveira A, Heck A, Logullo C, Gonzales JC, et al. Functional bovine immunoglobulins in Boophilus microplus hemolymph. Vet Parasitol 1996;62:155–60.
[55] Vaughan JA, Sonenshine DE, Azad AF. Kinetics of ingested host immunoglobulin G in hemolymph and whole body homogenates during nymphal development of Dermacentor variabilis and Ixodes scapularis ticks (Acari: Ixodidae). Exp Appl Acarol 2002;27:329–40.
[56] Jasinskas A, Jaworski DC, Barbour AG. Amblyomma americanum: specific uptake of immunoglobulins into tick hemolymph during feeding. Exp Parasitol 2000;96:213–21.
&KDSWHU�,,,


BACTERIAL'MEMBRANES'ENHANCE'THE' IMMUNOGENICITY'OF'
THE' SURFACE' EXPOSED' TICK' SUBOLESIN9ANAPLASMA'
MARGINALE!MSP1A'CHIMERIC'ANTIGEN!

32
Bacterial membranes enhance the immunogenicity of the
surface exposed tick Subolesin-Anaplasma marginale
MSP1a chimeric antigen
Juan A. Moreno-Cid1, Ana Domingos2, José M. Pérez de la Lastra1, Mario Canales1,*, Emilio Sánchez1, Iratxe Díez-Delgado1, Mariana Boadella1, Christian Gortázar1, José de la Fuente1,3,§
1SaBio. Instituto de Investigación en Recursos Cinegéticos IREC-CSIC-UCLM-JCCM, Ronda de Toledo s/n, 13005 Ciudad Real, Spain. 2Centro de Malária e Outras Doenças Tropicais, Instituto de Higiene e Medicina Tropical, Rua da Junqueira 100, 1349-008 Lisboa, Portugal. 3Department of Veterinary Pathobiology, Center for Veterinary Health Sciences, Oklahoma State University, Stillwater, OK 74078, USA.
*Present address: ProBioVet S.L., Instituto Regional de Investigación Científica Aplicada (IRICA),Laboratorio 3.06, Universidad de Castilla-La Mancha, Avenida Camilo José Cela, S/N, 13071 Ciudad Real, Spain.
§Corresponding author: Instituto de Investigación en Recursos Cinegéticos IREC-CSIC-UCLM-JCCM,Ronda de Toledo s/n, 13005 Ciudad Real, Spain. Phone: +34 926295450. E-mail: [email protected]
ABSTRACT
Ticks are vectors of diseases that affect humans and animals worldwide. Ticks are difficult to control and vaccines using recombinant tick antigens have been proposed as a cost-effective and environmentally sound alternative for tick control. The tick Subolesin (SUB)-MSP1a chimeric antigen was produced in Escherichia coli as membrane-bound and exposed protein and vaccination with bacterial membranes containing SUB-MSP1a resulted in the control of cattle tick infestations. In this research, we characterized the role of E. coli membranes in enhancing antigen immunogenicity and the potential problems associated with E. coli membrane proteins and bacterial endotoxins. The results showed that bacterial membranes enhance the immunogenicity of the SUB-MSP1a antigen in mouse and pig vaccination models. Furthermore, using the simple and cost-effective process developed for the purification of membrane-bound SUB-MSP1a, endotoxin levels were bellow the maximum level accepted for recombinant vaccines. These results provide further support for the development of tick vaccines using E. coli membranes exposing chimeric antigens composed of tick polypeptides fused to MSP1a such as SUB-MSP1a. The production of these membrane-bound antigens is simple and cost- effective and results in antigen preparations with increased immunogenicity and endotoxin levels within the limits acceptable for recombinant vaccine formulations.
Keywords: vector, immunology, antigenicity, subolesin, chimera, vaccine, MSP1a
&KDSWHU�,,,

33
1. Introduction
Diseases caused by arthropod-borne pathogens greatly impact human and animal health (Jones et al., 2008). Insects such as mosquitoes and sand flies are considered to be the most important vectors of human diseases worldwide (Halstead, 2007; Chappuis et al., 2007; Jones et al., 2008) while ticks are considered to be second to mosquitoes as vectors of pathogens to humans and the most important vectors of pathogens that cause disease in cattle (Willadsen et al., 2005; de la Fuente et al., 2008). Control of tick infestations has been based primarily on the use of chemical acaricides that has resulted in selection of acaricide-resistant ticks and environmental pollution (de la Fuente et al., 1999; de la Fuente et al., 2007). Vaccination with the Rhipicephalus (Boophilus) microplus BM85/BM95 gut antigen emerged as an alternative for tick control that has shown the advantage of being cost-effective while reducing acaricide applications and the drawbacks associated with their use (de la Fuente et al., 1998; de la Fuente et al., 2007). Subolesin (SUB), the ortholog of insect and vertebrate Akirins (AKR) is an evolutionary conserved protein that was recently discovered in Ixodes scapularis as a tick protective antigen. (Almazán et al., 2003; de la Fuente et al., 2006a; Mangold et al., 2009; de la Fuente et al., 2011). SUB is involved in tick innate immune response and in other molecular pathways including those required for feeding, reproduction and pathogen infection and multiplication (de la Fuente et al., 2006a; de la Fuente et al., 2006b; Kocan et al., 2009; Zivkovic et al., 2010; de la Fuente et al., 2011). Vaccination with recombinant SUB/AKR showed a reduction in tick, mosquito, sand fly and poultry red mite infestations by reducing ectoparasite numbers, weight and/or oviposition and tick infection with different pathogens such as Anaplasma phagocytophilum, A. marginale, Babesia bigemina and Borrelia burgdorferi (Moreno- Cid et al., 2013). These results suggested that SUB/AKR is a good candidate antigen for the development of vaccines for the control of
multiple arthropod vectors (Moreno-Cid et al., 2013). Proteins exposed on the cell membrane such as major surface proteins are good targets for vaccine development (Blouin et al., 2003). Recently, recombinant chimeras comprising tick proteins such as BM95 immunogenic peptides and SUB fused to the N-terminal region of the Anaplasma marginale major surface protein 1a (MSP1a) (BM95-MSP1a and SUB-MSP1a chimeras, respectively) were produced in Escherichia coli as membrane- bound and exposed proteins (Canales et al., 2008; Almazán et al., 2012). Furthermore, the use of bacterial membranes containing the membrane-bound BM95-MSP1a or SUB- MSP1a proteins as vaccines resulted in the control of R. microplus and R. annulatus infestations in cattle (Canales et al., 2009; Almazán et al., 2012). This system provides a novel, simple and cost-effective approach for the production of tick protective antigens by surface displaying antigenic protein chimera on the E. coli membrane and demonstrated the possibility of using recombinant bacterial membrane fractions in vaccine preparations to protect cattle against tick infestations (Almazán et al. 2012; Canales et al., 2009; Canales et al., 2010). However, important issues still need to be addressed before E. coli membranes with surface-exposed antigens such as the SUB- MSP1a chimera could be used in commercial vaccine formulations. Some of these issues were addressed in this research and include the characterization of the role of E. coli membranes in enhancing antigen immunogenicity and the potential problems associated with E. coli membrane proteins and bacterial endotoxins.
2. Materials and Methods2.1. Production of the recombinant SUB-
MSP1a chimeric antigen in E. coli
Unless otherwise indicated, all reagents used in this work were purchased either from Sigma-
&KDSWHU�,,,

34
Aldrich (St Louis, MO, USA) or VWR International Eurolab S.L. (Mollet del Vallés, Barcelona, Spain). For the production of the membrane-bound R. microplus SUB-MSP1a chimera, recombinant E. coli JM109 cells transformed with the pMBXAF3 expression vector were used (Almazán et al., 2012). Recombinant E. coli were propagated in 1 L flasks containing 250 mL Luria–Bertani (LB) broth supplemented with 10 g·L-1 tryptone, 5 g·L-1 yeast extract, 10 g·L-1 NaCl, 50 µg/ml ampicillin and 0.4% glucose (Laboratorios CONDA S.A., Madrid, Spain) for 2 h at 37 ºC and 200 rpm and then for 5.5 h after addition of 0.5 mM final concentration of isopropyl-β- d-thiogalactopyranoside (IPTG) for induction of recombinant protein production as previously described (Canales et al., 2008). Cell growth was monitored by measuring O.D.600 nm. Protein concentration was determined using bicinchoninic acid (BCA). The cells were harvested by centrifugation at 10,000 x g for 15 min at 4 ºC and then 1 g of cell pellet was resuspended in 5 mL of disruption buffer (100mM Tris–HCl, pH 7.5, 150mM NaCl, 1mM PMSF, 5mM MgCl2·6H2O and 0.1% (v/v) Triton X-100) and disrupted using a cell sonicator (Model MS73; Bandelin Sonopuls, Berlin, Germany). After disruption, the insoluble protein fraction containing the membrane-bound SUB-MSP1a was collected by centrifugation at 21,500 x g
for 15 min at 4 ◦C and stored at −20 ◦C untilused for characterization and vaccine formulations.
2.2. Characterization of recombinant SUB- MSP1a
The recombinant SUB-MSP1a was analyzed by SDS-PAGE and Western blot. Ten micrograms of total proteins were loaded onto a 12% SDS-polyacrylamide gel (Criterion XT precast gels, Bio-Rad, Hercules, CA, USA) and either stained with Coomassie Brilliant Blue or transferred to a nitrocellulose membrane for Western blot analysis. The percent of total cell proteins corresponding to recombinant SUB-MSP1a was determined in the stained SDS-polyacrilamide gel by densitometric analysis.
For Western blot analysis, proteins in the gel were transferred to a nitrocellulose membrane during 1 h at 12V in a Minie-Genie Electroblotter semi-dry transfer unit (Idea Scientific, Corvallis, OR, USA). The membrane was blocked with 5% skim milk for 1 h at room temperature, washed three times in TBS and probed with rabbit antibodies. Serum from rabbits immunized with recombinant R. microplus SUB (Almazán et al., 2010) was diluted 1:500 in 3% BSA in TBS and the membrane was incubated with the diluted sera for 1 h at room temperature, and washed three times with TBS. The membrane was then incubated with an anti-rabbit horseradish peroxidase (HRP) conjugate (Sigma, St. Louis, MO, USA) diluted 1:1000 in TBS. The membrane was washed three times with TBS and finally developed with TMB stabilized substrate for HRP (Promega) for 20 min.
2.3. Vaccine formulations
The membrane-bound insoluble protein fraction containing over 50% of total proteins corresponding to the SUB-MSP1a chimera was resuspended in PBS, pH 7.4. To prepare cell membrane-free SUB-MSP1a, 250 µg of the membrane-bound SUB-MSP1a were loaded onto a preparative 12% SDS-polyacrylamide gel, separated by SDS-PAGE and stained with Coomassie Brilliant Blue. The band corresponding to the SUB-MSP1a chimera was cut off the gel, crushed using a sterile razor blade and incubated in 1 mL 150 mM Tris buffer, pH 7.4, containing 0.1% SDS, with
gentle agitation overnight at 4 ◦C. After
centrifugation for 15 min at 4 ◦C, themembrane-free SUB-MSP1a protein was collected in the supernatant. Both membrane- bound and membrane-free SUB-MSP1a proteins were adjuvated in Montanide ISA 50 V2 (Seppic, Paris, France) at a concentration of 125 µg SUB-MSP1a per mL (Almazán et al., 2012).
2.4. Endotoxin quantitation
The endotoxin levels in the membrane-bound SUB-MSP1a protein fraction were determined by standard kinetic chromogenic Limulus Amebocyte Lysate (LAL) assay (Thermo
&KDSWHU�,,,

35
Scientific, Hudson, NH, USA) following the manufacturer’s instructions. The reaction was stopped by the addition of 25% acetic acid and the absorbance was measured at 405 nm.
2.5. Immunizations in mice
Three groups of five female 5 weeks old
pigs were serially diluted from 1:10 to 1:1,000 in PBST (PBS/0.5% Tween 20, pH 7.2) and 10% fetal bovine serum (Sigma). The plates were incubated with the diluted sera for 1h at
37◦C and then incubated with 1:10,000 rabbitanti-mouse or anti-pig IgG–HRP conjugates
for 1h at 37◦C. The plates were washed withPBST, the color reaction was developed with SigmaFast OPD following the manufacturer’s
Balb/c mice per group were each immunized three times at weeks 0, 3 and 6 with 0.2 mL doses (25 µg) injected subcutaneously in the dorsum near the base of the tail using a 1-ml tuberculin syringe and a 27.5-G needle. Mice
recommendations and the OD determined.
3. Results and Discussion
!450nm was
were immunized with membrane-bound SUB- MSP1a, membrane-free SUB-MSP1a or adjuvant/saline alone as control. Blood was collected at weeks 0, 6 and at the end of the experiment (week 8) and used for serum preparation. Mice were housed at the Instituto de Higiene e Medicina Tropical, Universidade Nova de Lisboa, Portugal (Permit No. 023357).
2.6. Oral immunization in pigs
Ten 2-3 month-old domestic Iberian piglets
3.1. Production of recombinant SUB- MSP1a
The expression of the SUB-MSP1a chimera was characterized in E.coli grown in 250 mL working volume at bench-top scale. The yield in terms of biomass production and SUB- MSP1a protein levels were similar to that previously reported for BM95-MSP1a (Canales et al., 2008) (Fig.1A). Cells grew on 1% glucose as sole carbon source exponentially at the maximum growth rate of 2.3 h-1, reaching an optical density of 0.8±0.4
from a farm in Castilla-La Mancha, Spain were randomly separated into two groups of five O.D.600nm before induction with IPTG for
animals each and orally immunized at weeks 0 and 4 with 5 mL PBS, pH 7.4 containing 5 mg recombinant R. microplus SUB (Almazán et al., 2010) or 5 mg membrane-bound SUB- MSP1a. Blood was collected for serum preparation before immunization at weeks 0 and 4 and at the end of the experiment at week 8. Animals handling procedures and samplingfrequency were designed to reduce stress and health risks for subjects, according to European (86/609) and Spanish legislation (R.D. 223/1988, R.D. 1021/2005). The protocol was approved by the Committee on the Ethics of Animal Experiments of the Regional Agriculture Authority.
2.7. Determination of antibody titers in mice and pigs by ELISA
Purified recombinant SUB (Almazán et al., 2010) (0.1 mg/well) were used to coat ELISA
plates overnight at 4◦C. Sera from mice and
expression of the recombinant protein.
Kinetics was conducted to determine the optimal induction time for SUB-MSP1a production (Fig. 1B). The SUB–MSP1a chimera started to accumulate after 1 h of IPTG induction and its concentration reached the maximum of 9.5% of total cell proteins 5.5 h after induction (Fig. 1B). A simple process previously reported for the bacterial membrane containing the chimeric BM95-MSP1a antigen (Canales et al., 2008) was used for SUB- MSP1a recovery. This process involved cell disruption and phase separation by centrifugation, yielding an enriched membrane fraction with a total recovery of 90% of the recombinant protein and purity higher than 50% in the final fraction (Figs. 1C and 1D). The overall operating time in the downstream process was 79% less when compared to intracellular SUB production in E. coli (Almazán et al., 2010) (Table 1). The productivity, calculated as the total amount of recombinant protein obtained per culture
&KDSWHU�,,,

36
volume and unit time, was compared between SUB-MSP1a (186.7 mg/Lh) and SUB (1.7 mg/Lh) production processes showing 91% higher productivity for SUB-MSP1a. The protein recovery, calculated as protein concentration in the purified final fraction divided by total protein concentration, was also
higher for SUB-MSP1a (90% recovery) than for SUB (30% recovery). These results reinforced the advantages and cost-effectiveness of the production of surface- exposed membrane-bound chimeric antigens (Canales et al., 2010; Almazán et al., 2012).
Table 1. Estimated time per unit operation for the production and recovery of recombinant antigens in E.coli.
Estimated time per operation (h)
Process ��operation Intracellular SUB Membrane-bound SUB-MSP1a
12 12
7 7
0.25 0.5
1 1
0.5 0.5
0.25 0.5
1 1
1 NR
1 NR
12 NR
Seed I
Multiplication Cell
harvesting I
Homogenization I
Cell disruption Cell
+arvesting II
Homogenization II
Microfiltration
Chromatography
Dialysis
Total time 35 21
Abbreviation: NR, not required for antigen production.
&KDSWHU�,,,
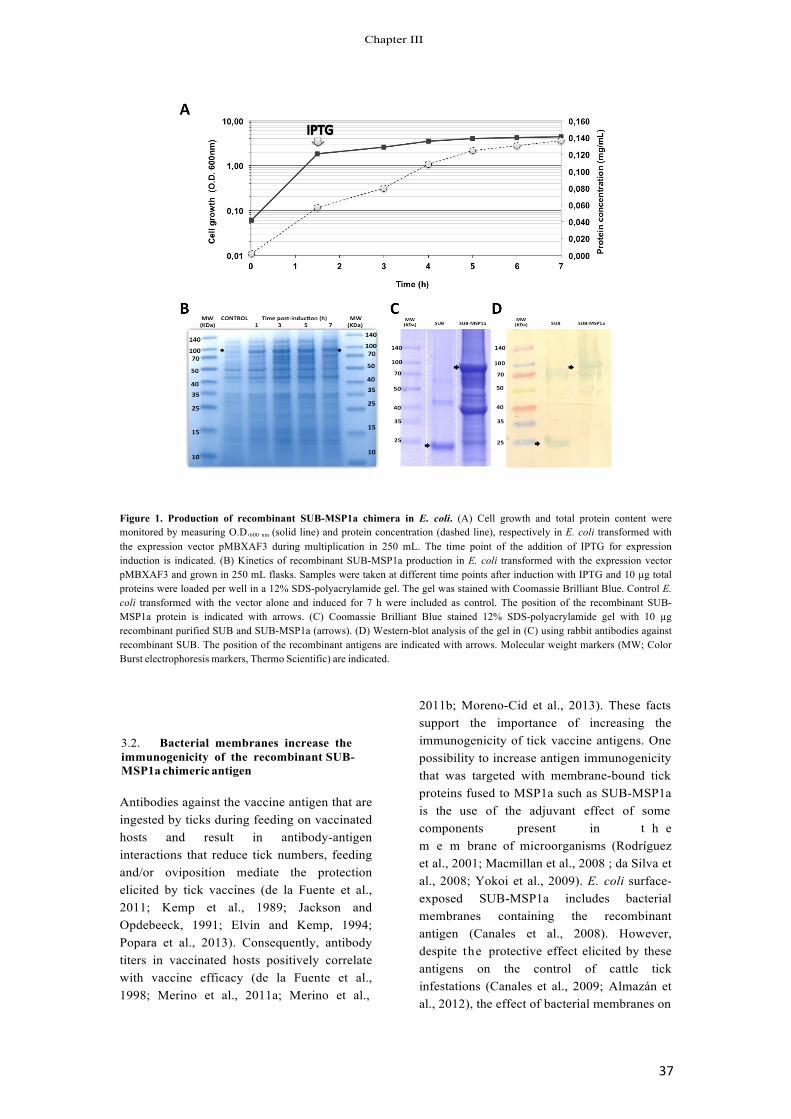
37
Figure 1. Production of recombinant SUB-MSP1a chimera in E. coli. (A) Cell growth and total protein content were monitored by measuring O.D.600 nm (solid line) and protein concentration (dashed line), respectively in E. coli transformed with the expression vector pMBXAF3 during multiplication in 250 mL. The time point of the addition of IPTG for expression induction is indicated. (B) Kinetics of recombinant SUB-MSP1a production in E. coli transformed with the expression vector pMBXAF3 and grown in 250 mL flasks. Samples were taken at different time points after induction with IPTG and 10 µg total proteins were loaded per well in a 12% SDS-polyacrylamide gel. The gel was stained with Coomassie Brilliant Blue. Control E. coli transformed with the vector alone and induced for 7 h were included as control. The position of the recombinant SUB- MSP1a protein is indicated with arrows. (C) Coomassie Brilliant Blue stained 12% SDS-polyacrylamide gel with 10 µg recombinant purified SUB and SUB-MSP1a (arrows). (D) Western-blot analysis of the gel in (C) using rabbit antibodies against recombinant SUB. The position of the recombinant antigens are indicated with arrows. Molecular weight markers (MW; Color Burst electrophoresis markers, Thermo Scientific) are indicated.
3.2. Bacterial membranes increase the immunogenicity of the recombinant SUB-MSP1a chimeric antigen
Antibodies against the vaccine antigen that are ingested by ticks during feeding on vaccinated hosts and result in antibody-antigen interactions that reduce tick numbers, feeding and/or oviposition mediate the protection elicited by tick vaccines (de la Fuente et al., 2011; Kemp et al., 1989; Jackson and Opdebeeck, 1991; Elvin and Kemp, 1994; Popara et al., 2013). Consequently, antibody titers in vaccinated hosts positively correlate with vaccine efficacy (de la Fuente et al., 1998; Merino et al., 2011a; Merino et al.,
2011b; Moreno-Cid et al., 2013). These facts support the importance of increasing the immunogenicity of tick vaccine antigens. One possibility to increase antigen immunogenicity that was targeted with membrane-bound tick proteins fused to MSP1a such as SUB-MSP1a is the use of the adjuvant effect of some components present in t h e m e m brane of microorganisms (Rodríguez et al., 2001; 0DFPLOODQ�HW�DO����������da Silva et al., 2008; Yokoi et al., 2009). E. coli surface-exposed SUB-MSP1a includes bacterial membranes containing the recombinant antigen (Canales et al., 2008). However, despite the protective effect elicited by these antigens on the control of cattle tick infestations (Canales et al., 2009; Almazán et al., 2012), the effect of bacterial membranes on
&KDSWHU�,,,
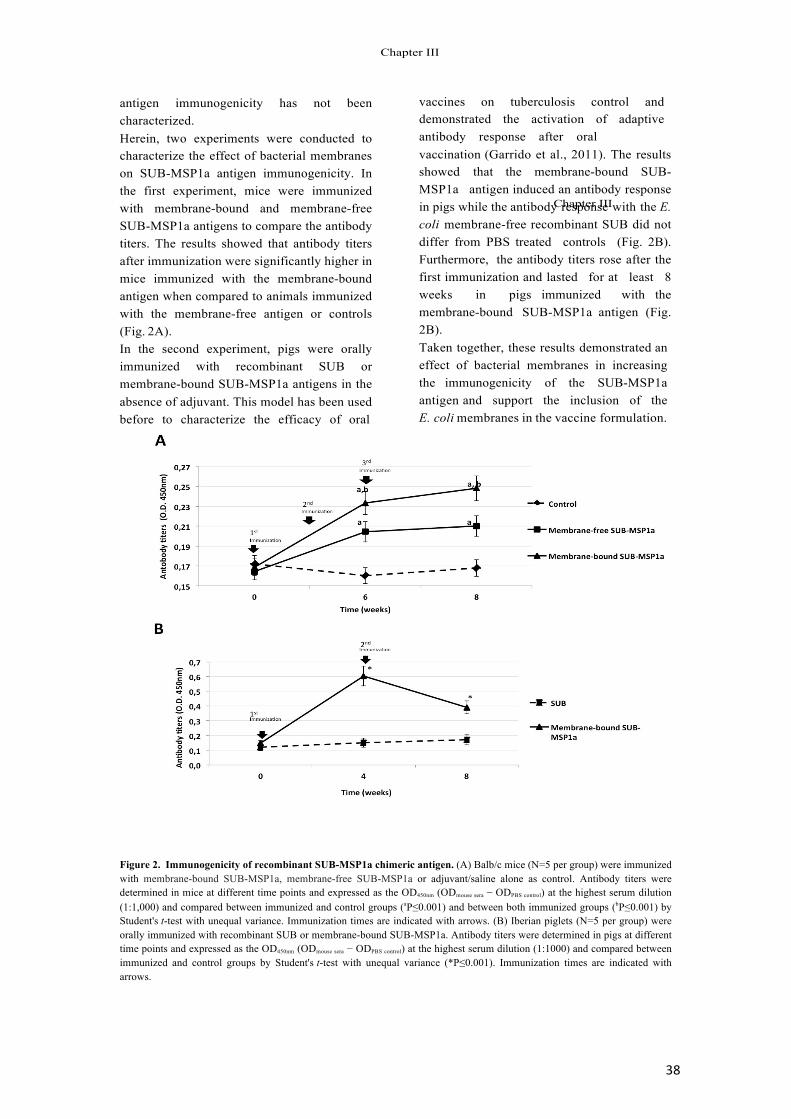
38
antigen immunogenicity has not been characterized. Herein, two experiments were conducted to characterize the effect of bacterial membranes on SUB-MSP1a antigen immunogenicity. In the first experiment, mice were immunized with membrane-bound and membrane-free SUB-MSP1a antigens to compare the antibody titers. The results showed that antibody titers after immunization were significantly higher in mice immunized with the membrane-bound antigen when compared to animals immunized with the membrane-free antigen or controls (Fig. 2A). In the second experiment, pigs were orally immunized with recombinant SUB or membrane-bound SUB-MSP1a antigens in the absence of adjuvant. This model has been used before to characterize the efficacy of oral
vaccines on tuberculosis control and demonstrated the activation of adaptive antibody response after oral vaccination (Garrido et al., 2011). The results showed that the membrane-bound SUB- MSP1a antigen induced an antibody response in pigs while the antibody response with the E. coli membrane-free recombinant SUB did not differ from PBS treated controls (Fig. 2B). Furthermore, the antibody titers rose after the first immunization and lasted for at least 8 weeks in pigs immunized with the membrane-bound SUB-MSP1a antigen (Fig. 2B). Taken together, these results demonstrated an effect of bacterial membranes in increasing the immunogenicity of the SUB-MSP1a antigen and support the inclusion of the E. coli membranes in the vaccine formulation.
Figure 2. Immunogenicity of recombinant SUB-MSP1a chimeric antigen. (A) Balb/c mice (N=5 per group) were immunized with membrane-bound SUB-MSP1a, membrane-free SUB-MSP1a or adjuvant/saline alone as control. Antibody titers were determined in mice at different time points and expressed as the OD450nm (ODmouse sera − ODPBS control) at the highest serum dilution (1:1,000) and compared between immunized and control groups (aP≤0.001) and between both immunized groups (bP≤0.001) by Student's t-test with unequal variance. Immunization times are indicated with arrows. (B) Iberian piglets (N=5 per group) were orally immunized with recombinant SUB or membrane-bound SUB-MSP1a. Antibody titers were determined in pigs at different time points and expressed as the OD450nm (ODmouse sera − ODPBS control) at the highest serum dilution (1:1000) and compared between immunized and control groups by Student's t-test with unequal variance (*P≤0.001). Immunization times are indicated with arrows.
&KDSWHU�,,,
&KDSWHU�,,,

39
3.3. Endotoxin content is the membrane- bound SUB-MSP1a antigen is within the limits acceptable for recombinant vaccines
Considering the effect of bacterial membranes in the SUB-MSP1a vaccine formulation, a balance should be reached between this positive effect and the detrimental effect of endotoxins associated with the membrane fraction that may raise safety issues. To address this issue, herein we used the LAL assay to determine the endotoxin content in the purified membrane-bound SUB-MSP1a preparation. The LAL assay has been used since the early 1970s for the detection of endotoxins and provides a reliable and validated tool for these studies (Brito and Singh, 2010). Several approaches has been used to obtain endotoxin-free antigens produced in E. coli (Canales et al., 2010), including endotoxin removal using non-ionic detergents (Aida et al., 1990). The process developed to recover the E. coli membrane fraction containing the exposed SUB-MSP1a antigen contains 0.1% Triton X-100 (Almazán et al., 2012). The residual endotoxin levels obtained in the crude fraction after cell centrifugation was equal to 173±20 endotoxin units (EU)/mg. However, after cell disruption and treatment with Triton X-100, endotoxin levels decreased to 85±5 EU/mg. Therefore, in the final vaccine formulation for cattle containing 0.1 mg antigen per dose (Almazán et al., 2012), endotoxin levels were equivalent to 8.5±0.5 EU/dose. This endotoxin level is lower than the maximum recommended level of 20 EU/dose for recombinant vaccines (Brito and Singh, 2010). These results showed that the use of 0.1% Triton X-100 was successful in reducing endotoxins to acceptable levels for vaccine formulations based on the membrane-bound SUB-MSP1a using a simple a cost-effective production system (Canales et al., 2010).
4. ConclusionsIn summary, the results of this study provide further support for the development of tick vaccines using E. coli membranes exposing chimeric antigens composed of tick polypeptides fused to MSP1a such as SUB- MSP1a. The production of these membrane- bound antigens is simple and cost-effective (Canales et al., 2010) and results in antigen preparations with increased immunogenicity and endotoxin levels within the limits acceptable for recombinant vaccine formulations. However, additional experiments are required to validate the efficacy of the vaccine containing the SUB-MSP1a antigen preparation in field trials.
Acknowledgements This research was supported by EU FP7 Grant WildTBVac and Plan Nacional I+D+i AGL2011-30041 from Ministerio de Economía y Competitividad (MINECO), Spain and FEDER. J.A. Moreno-Cid was funded by the Consejería de Educación y Ciencia, JCCM, Spain (project PEII09-0118-8907), as a recipient of a JCCM fellowship.
&KDSWHU�,,,

40
References
Aida, Y., Pabst, M.J. 1990. Removal of endotoxin from protein solutions by phase separation using Triton X-114. J. Immunol. Methods. 132, 191–195.
Almazán, C., Kocan, K.M., Bergman, D.K., Garcia- Garcia, J.C., Blouin, E.F., de la Fuente, J. 2003. Identification of protective antigens for the control of Ixodes scapularis infestations using cDNA expression library immunization. Vaccine. 21, 1492–501.
Almazán, C., Lagunes, R., Villar, M., Canales, M., Rosario-Cruz, R., Jongejan, F., de la Fuente, J. 2010. Identification and characterization of Rhipicephalus (Boophilus) micropluscandidate protective antigens for the control of cattle tick infestations. Parasitology Research. 106, 471-479.
Almazán, C., Moreno-Cantú, O., Moreno-Cid, J.A., Galindo, R.C., Canales, M., Villar, M., de la Fuente, J. 2012. Control of tick infestations in cattle vaccinated with bacterial membranes containing surface-exposed tick protective antigens. Vaccine. 30, 265-272.
Blouin, E. F., Saliki, J. T., de la Fuente, J., Garcia-Garcia, J. C., Kocan, K. M. 2003. Antibodies to Anaplasma marginale major surface proteins 1a and 1b inhibit infectivity for cultured tick cells. Veterinary Parasitology. 111, 247-260.
Brito, L.A., Singh, M. 2010. Acceptable Levels of Endotoxin in Vaccine Formulations During Preclinical Research Novartis Vaccines and Diagnostics. Journal of Pharmaceutical Science. 100, 34-37.
Canales, M., Almazán, C., Pérez de la Lastra, J.M., de la Fuente, J. 2008. Anaplasma marginale major surface protein 1a directs cell surface display of tick BM95 immunogenic peptides on Escherichia coli. J. Biotechnol. 135, 326-332.
Canales, M., Labruna, M.B., Soares, J.F., Prudencio, C.R., de la Fuente, J. 2009. Protective efficacy of bacterial membranes containing surface-exposed BM95 antigenic peptides for the control of cattle tick infestations. Vaccine. 27, 7244-7248.
Canales, M., Moreno-Cid, J.A., Almazán, C., Villar, M. and de la Fuente, J. 2010. Bioprocess design and economics of recombinant BM86/BM95 antigen production for anti-tick vaccines. Biochemical Engineering Journal. 52, 79-90.
Chappuis, F., Sundar, S., Hailu, A., Ghalib, H., Rijal, S., Peeling, R.W., Alvar, J. and Boelaert, M. 2007. Visceral leishmaniasis: what are the needs for diagnosis, treatment and control? NatRev Microbiol. 5, 873–82.
Elvin, C.M. and Kemp, D.H. 1994. Generic approaches to obtaining efficacious antigens from vector arthropods. Int J Parasitol. 24, 67-79.
de la Fuente, J., Rodríguez, M., Redondo, M., Montero, C., García-García, J.C., Méndez, L., Serrano, E., Valdés, M., Enríquez, A., Canales, M., Ramos, E., de Armas, C.A., Rey, S., Rodríguez, J.L., Artiles, M. and García, L. 1998. Field studies and cost-effectiveness analysis of vaccination with GavacTM against the cattle tick Boophilus microplus. Vaccine. 16, 366-373.
de la Fuente, J., Rodriguez, M., Montero, C., Redondo, M., Garcia-Garcia, J.C., Mendez, L., Serrano, E., Valdes, M., Enriquez, A., Canales, M., Ramos, E., Boué, O., Machado, H. and Lleonart, R. 1999. Vaccination against ticks (Boophilus spp.): the experience with the Bm86-based vaccine GavacTM, Genet. Anal. Biomol. Eng. 15, 143– 148.
de la Fuente, J., Garcia-Garcia, J.C., Blouin, E.F. and Kocan, K.M. 2001. Major surface protein 1a effects tick infection and transmission of the ehrlichial pathogen Anaplasma marginale, Int. J. Parasitol. 31, 1705–1714.
de la Fuente, J., Almazán, C., Blas-Machado, U., Naranjo, V., Mangold, A.J., Blouin, E.F., Gortazar, C. and Kocan, K. 2006a. The tick protective antigen, 4D8, is a conserved protein involved in modulation of tick blood ingestion and reproduction. Vaccine. 24, 4082–95.
de la Fuente, J., Almazán, C., Naranjo, V., Blouin, E.F., Meyer, J.M., Kocan, K.M. 2006b. Autocidal control of ticks by silencing of a single gene by RNA interference. Biochem. Biophys. Res. Commun. 344, 332–8.
de la Fuente, J., Almazán, C., Canales, M., Perez de la Lastra, J.M., Kocan, K.M., Willadsen, P. 2007. A ten-year review of commercial vaccine performance for control of tick infestations on cattle. Animal Health Res. Rev. 8, 23– 28.
de la Fuente, J., Maritz-Olivier, C., Naranjo, V., Ayoubi, P., Nijhof, A.M., Almazán, C., Canales, M., Perez de la Lastra, J.M., Galindo, R.C., Blouin, E.F., Gortazar, C., Kocan, K.M. 2008. Evidence of the role of tick subolesin in gene expression. BMC Genomics. 9, 372.
de la Fuente, J., Moreno-Cid, J.A., Canales, M., Villar, M., Pérez de la Lastra, J.M., Kocan, K.M., Galindo, R.C., Almazán, C., Blouin, E.F. 2011. Targeting arthropod subolesin/akirin for the development of a universal vaccine for control of vector infestations and pathogen transmission. Vet Parasitol. 181, 17–22.
&KDSWHU�,,,
Garrido, J.M., Sevilla, I., Beltrán-Beck, B., Minguijón, E., Ballesteros, C., Galindo, R.C., Boadella, M., Lyashchenko, K.P., Romero, B., Geijo, M.V., Ruiz-Fons, J.F., Aranaz, A., Juste, R., Vicente, J., de la Fuente, J., Gortázar, C. 2011. Protection against tuberculosis in Eurasian wild boar vaccinated with heat-inactivated Mycobacterium bovis. PLoS ONE 6: e24905.

41
Halstead, S.B. 2007. Dengue. Lancet. 370, 1644–52.
Jackson, L.A., Opdebeeck, J.P. 1991. Humoral immune responses of Hereford cattle vaccinated with midgut antigens of the cattle tick B. microplus. Parasite Immunol. 13, 141-147.
Jones, K.E., Patel, N.G., Levy, M.A., Storeygard, A., Balk, D., Gittleman, J.L., Daszak, P. 2008. Global trends in emerging infectious diseases. Nature. 451, 990–4.
Kemp, D.H., Pearson, R.D., Gough, J.M., Willadsen, P. 1989. Vaccination against B. microplus: localisation of antigens on ticks gut cells and their interaction with the host immune system. Exp Appl Acarol. 7, 43-58
Kocan, K.M., Zivkovic, Z., Blouin, E.F., Naranjo,V., Almazán, C., Mitra R., de la Fuente, J. 2009. Silencing of genes involved in Anaplasma marginale-tick interactions affects the pathogen developmental cycle in Dermacentor variabilis. BMC Dev Biol. 9, 42.
Macmillan, H., Norimine, J., Brayton, K.A., Palmer, G.H., Brown, W.C. 2008. Physical linkage of naturally complexed bacterial outer membrane proteins enhances immunogenicity. Infect. Immun. 76, 1223-1229.
Mangold, A.J., Galindo, R.C., de la Fuente, J. 2009. Response to the commentary of D. Macqueen on: Galindo RC, Doncel-Pérez E, Zivkovic Z, Naranjo V, Gortazar C, Mangold AJ, et al. Tick subolesin is an ortholog of the akirins described in insects and vertebrates [2009. Dev. Comp. Immunol. 33, 612–617]. Dev Comp Immunol. 33, 878–9.
Merino, O., Almazán, C., Canales, M., Villar, M., Moreno- Cid, J.A., Estrada-Peña, A., Kocan, K.M., de la Fuente, J. 2011a Control of Rhipicephalus (Boophilus) microplus infestations by the combination of subolesin vaccination and tick autocidal control after subolesin gene knockdown in ticks fed on cattle. Vaccine. 29, 2248-2254.
Merino, O., Almazán, C., Canales, M., Villar, M., Moreno- Cid, J.A., Galindo, R.C., de la Fuente, J. 2011b. Targeting the tick protective antigen subolesin reduces vector infestations and pathogen infection by Anaplasma marginale and Babesia bigemina. Vaccine. 29, 8575–9.
Moreno-Cid, J.A., Pérez de la Lastra, J.M., Villar, M., Jiménez, M., Pinal, R., Estrada-Peña, A., Alarcón, P., Delacour, S., Oropeza, V., Ruiz, I., Molina, R., Lucientes, J., Prudencio, C.R., Galindo, R.C., Almazán, C., Nijhof,
A.M., Mangold, A.J., Gortázar, C., de la Fuente, J. 2013. Control of multiple arthropod vector infestations with subolesin/akirin vaccines. Vaccine. 31, 1187-1196.
Popara, M., Villar, M., Mateos-Hernández, L., Fernández de Mera, I.G., Marina, A., del Valle, M., Almazán, C., Domingos, A., de la Fuente, J. 2013. Lesser protein degradation machinery correlates with higher BM86 tick vaccine efficacy in Rhipicephalus annulatus when compared to R. microplus. Vaccine. In press.
Rodríguez, M., Montero, C., Machado, H., Joglar, M., de la Fuente, J., Garcia-Garcia J.C. 2001. The evaluation of yeast derivatives as adjuvants for the immune response to the Bm86 antigen in cattle. BMC Biotechnol. 1: 2. Erratum in BMC Biotechnol. 1: 3
Yokoi, S., Niizeki, H., Iida, H., Asada, H., Miyagawa, S. 2009. A djuvant effect of lipopolysaccharide on the induction of contact hypersensitivity to haptens in mice. J Dermatol Sci. 53, 120-128.
da Silva R.A., Conceição, F.R., Grassmann, A.A., Lagranha, V.L., Dellagostin, O.A. 2008. B subunit of Escherichia coli heat-labile enterotoxin as adjuvant of humoral immune response in recombinant BCG vaccination. Can J Microbiol. 54, 677-686.
Willadsen, P. 2005. Vaccination Against Ticks and the Control of Ticks and Tick-borne Disease. Applications of Gene-Based Technologies for Improving Animal Production and Health in Developing Countries. Springer Netherlands, pp. 313-321.
Zivkovic, Z., Torina, A., Mitra, R., Alongi, A., Scimeca, S., Kocan, K.M., Galindo R.C., Almazán C., Blouin E.F., Villar, M., Nijhof, A.M., Mani, R., La Barbera G., Caracappa, S., Jongejan F., de la Fuente, J.. 2010. Subolesin expression in response to pathogen infection in ticks. BMC Immunol. 11, 7.
&KDSWHU�,,,


CHAPTER(IV.(VACCINATION(
Almazan!C.,!Moreno3Cantu!O.,!Moreno3Cid!J.A.,!Galindo!R.C.,!Canales!M.,!Villar!M.,!de! la! Fuente! J.! (2012).! Control' of' tick' infestations' in' cattle' vaccinated' with'bacterial' membranes' containing' surface7exposed' tick' protective' antigens.!Vaccine!30!(2):!2653272!
Moreno3Cid! J.A.,! Pérez! de! la! Lastra! J.M.,! Villar! M.,! Pinal! R.,! Estrada3Peña! A.,!Molina! R.,! Lucientes! J.,! Gortázar! C.,! de! la! Fuente! J.! (2013).! Control' of' multiple'arthropod' vector' infestations' with' subolesin/akirin' vaccines.! Vaccine! ! 31(8):!118731196.!


CONTROL'OF'MULTIPLE'ARTHROPOD'
VECTOR'INFESTATIONS'WITH'
SUBOLESIN/AKIRIN'VACCINES'

44!
Contents lists available at SciVerse ScienceDirect
Vaccine
jou rn a l h om epa ge: www.elsevier .com/locate /vaccine
Control of multiple arthropod vector infestations with subolesin/akirin vaccines
Juan A. Moreno-Cid a , José M. Pérez de la Lastra a , Margarita Villar a , Maribel Jiménez b , Rocío Pinal c , Agustín Estrada-Pen a d, Ricardo Molina b, Javier Lucientes c, Christian Gortázar a, José de la Fuente a,e,∗, the SUB/AKR Vaccine Study Group a Instituto de Investigación en Recursos Cinegéticos IREC-CSIC-UCLM-JCCM, Ronda de Toledo s/n, 13005 Ciudad Real, Spain b Unidad de Entomología Médica, Servicio de Parasitología, Centro Nacional de Microbiología, Instituto de Salud Carlos III, 28220 Majadahonda, Madrid, Spain c Departamento de Patología Animal, Facultad de Veterinaria, C/Miguel Servet 177, 50013 Zaragoza, Spain d Department of Parasitology, Veterinary Faculty, Miguel Servet 177, 50013 Zaragoza, Spain e Department of Veterinary Pathobiology, Center for Veterinary Health Sciences, Oklahoma State University, Stillwater, OK 74078, USA
a r t i c l e i n f o a b s t r a c t
Article history: Received 5 October 2012 Received in revised form 20 December 2012 Accepted 23 December 2012 Available online 3 January 2013
Keywords: Akirin Arthropod Mosquito Sand fly Subolesin Tick Vaccine Immunology
Diseases transmitted by arthropod vectors such as mosquitoes, ticks and sand flies greatly impact human and animal health and thus their control is important for the eradication of vector-borne diseases (VBD). Vaccination is an environmentally friendly alternative for vector control that allows control of several VBD by targeting their common vector. Recent results have suggested that subolesin/akirin (SUB/AKR) is good candidate antigens for the control of arthropod vector infestations. Here we describe the comparative effect of vaccination with SUB, AKR and Q38 and Q41 chimeras containing SUB/AKR conserved protective epitopes on tick, mosquitoes and sand flies vector mortality, molting, oviposition and/or fertility. We demonstrated that SUB vaccination had the highest efficacy (E) across all vector species (54–92%), Q41 vaccination had the highest vaccine E in mosquitoes (99%) by reducing female survival and fertility, and Q38 vaccination had the highest effect on reducing mosquito (28%) and sand fly (26%) oviposition. The effect of vaccination on different developmental processes in several important arthropod vectors encourages the development of SUB/AKR universal vaccines for the control of multiple vector infestations and reduction of VBD.
© 2012 Elsevier Ltd. All rights reserved.
1. Introduction
Diseases transmitted by arthropod vectors such as mosquitoes, ticks and sand flies greatly impact human and animal health [1]. Ixodes spp. ticks (Acari: Ixodidae) are important vectors of pathogens that cause disease in humans and domestic animals worldwide, including Anaplasma phagocytophilum (human, equine and canine granulocytic anaplasmosis and tick-borne fever of ruminants), Borrelia burgdorferi (Lyme disease) and tick-borne encephalitis virus [2]. Aedes spp. mosquitoes (Diptera: Culicidae) are vectors of viruses that cause disease in humans such as dengue fever [3]. Phlebotomine sand flies (Diptera: Psychodidae) are also
∗ Corresponding author at: Instituto de Investigación en Recursos CinegéticosIREC-CSIC-UCLM-JCCM, Ronda de Toledo s/n, 13005 Ciudad Real, Spain. Tel.: +34 926295450.
E-mail addresses: [email protected], jose [email protected] (J. de la Fuente).
1 See Appendix A.
vectors of several pathogens including Leishmania spp., which causes various forms of human and canine leishmaniasis [4].
Despite the impact of vector-borne diseases (VBD) on human and animal health, few VBD such as yellow fever have effective vaccines [5]. Thus, the control of arthropod vectors is important for the eradication of VBD [6–10]. Vaccination is an environmentally friendly alternative for vector control that allows control of several VBD by targeting their common vector [9,11,12]. Vaccination with arthropod vector antigens has been proposed as an alternative for the control of vector infestations and reduction of vector capac- ity to transmit pathogens that impact human and animal health [7,11–21].
Subolesin (SUB) and akirin (AKR) are ortholog proteins in ticks and insects [22–25] that affect the expression of signal transduc- tion and innate immune response genes as well as positive and negative transcriptional regulators [24–26]. These proteins inter- act with NF-KB and other regulatory proteins, bind DNA or remodel chromatin to regulate gene expression [24–28]. This broad function of SUB/AKR as transcription factors explains the profound effect of gene knockdown on tick and insect physiology, development and gene expression [22,27,29,30]. SUB expression in infected ticks
0264-410X/$ – see front matter © 2012 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.vaccine.2012.12.073
&KDSWHU�,9

45!
suggested that SUB might be functionally important for tick innate immunity to pathogens [31]. In addition, SUB has a role in other molecular pathways including those required for tissue develop- ment and function and for pathogen infection and multiplication in ticks [22,29–31].
SUB was discovered as a tick protective antigen in Ixodes scapu- laris [32]. Vaccination with recombinant SUB showed an effective control of tick infestations by reducing tick numbers, weight and oviposition and tick infection with different tick-borne pathogens [12,33]. Subsequent experiments with recombinant SUB/AKR have shown an effect of antigen-specific antibodies on several arthro- pod vectors including hard and soft ticks, mosquitoes, sand flies, poultry red mites and sea lice [21,33–37]. Here we provide the first report showing the comparative effect of vaccination with SUB, AKR and SUB/AKR chimeras containing conserved protective epitopes on ticks, mosquitoes and sand flies.
2. Materials and methods
2.1. Ticks, mosquitoes and sand flies
Ticks, Ixodes ricinus, originally collected in Neûchatel, Switzerland and maintained as a colony at the University of Neûchatel were kindly provided by Lise Gern and used in this study. Ten engorged females were transferred to the University of Zaragoza (UZ, Spain) and allowed to oviposit at 24 ◦C. Resulting larvae were kept in the dark at 22 ◦C and 90% relative humidity (RH).
Mosquitoes, Aedes albopictus, from the Baix Llobregat (Barcelona, Spain) colony maintained for 31 generations at the UZ were used for this study. Mosquitoes were reared in an environmental cabinet at 25 ± 1 ◦C, 75 ± 5% RH, and a 12 h light:12 h dark photoperiod.
Sand flies, Phlebotomus perniciosus, used in this study were from an autochthonous (Madrid Province, Spain) colony established at the Medical Entomology Unit of the Instituto de Salud Carlos III (ISCIII, Spain). The P. perniciosus strain has been maintained in the laboratory for over 200 generations. Flies were reared in an envi- ronmental cabinet at 28 ◦C, 95–100% RH, and a 17 h light:7 h dark photoperiod.
PepSurf at the Pepitope Server [39]. The three-dimensional models were created by FirstGlance in Jmol (http://firstglance.jmol.org).
Analysis of SUB/AKR sequences
SUB and AKR protein sequences from different tick and insect species and geographic strains (Table S1) were used for phylo- genetic analysis and to characterize conservation of protective epitopes included in Q38 and Q41 chimeras (Supplementary methods).
A fragment of the P. perniciosus (ISCIII colony) AKR was cloned from unfed adult females (Supplementary methods) and submitted to GenBank (JX193855).
2.2. Vaccine formulations
The expression and purification of secreted recombinant SUB (Rhipicephalus microplus Susceptible Media Joya strain; DQ159966) and A. albopictus AKR (EU637024) was conducted in Pichia pastoris as previously reported [40,41]. Recombinant SUB/AKR chimeras Q38 and Q41 were expressed in Escherichia coli from synthetic genes (JX193856 and JX193857) optimized for codon usage in E. coli and purified by Ni affinity chromatography (Genscript Corporation, Piscataway, NJ, USA) (Supplementary methods). Recombinant anti- gens or saline were adjuvanted in Montanide ISA 50 V2 (Seppic, Paris, France) [34].
2.3. Mouse vaccination and infestation
Three experiments were conducted to evaluate and compare the effect of vaccination with recombinant tick SUB, mosquito AKR and SUB/AKR Q38 and Q41 chimeras on mice infested with I. ricinus, A. albopictus and P. perniciosus. These experiments were independently conducted at the UZ for ticks and mosquitoes and the ISCIII for sand flies. Six female 5 weeks old Balb/c mice per group were each immunized three times at weeks 0, 3 and 6 with 0.2 ml doses injected subcutaneously in the dorsum near the base of the tail using a 1-ml tuberculin syringe and a 27.5-G needle. Vaccinated and control mice were injected with recombinant vac- cines (25 µ,g/dose) and adjuvant/saline, respectively, and infested with arthropod vectors at week 8. Immunizations, arthropods col- lections and evaluations were done blinded and the key to the experimental groups was not opened until the end of the experi- ment. For ticks, mortality (no. of dead larvae/total no. of larvae used for infestation) and molting (no. of nymphs/no. of replete larvae) were evaluated. For insects, mortality (no. of dead/fed females), oviposition (no. of eggs/female) and fertility (fertility 1 = no. of lar-
For both insect species, females were mixed with males after emergence until females were selected for the experiment. Under our rearing conditions, most of the insects mate as demonstrated by ovaries dissection and examination in previous experiments [34]. After emergence, insects were supplied absorbent cotton saturated with 10–30% sucrose solution during four days. At day five, they were starved for 5 h before mice infestation [34]. Then, females were maintained in new cages without males until the end of the H[SHULPHQW�
9DFFLQH�efficacy (E) was calculated as E = 100��O��Ȇ����������ZKHUH�DN���������vae/egg and fertility 2 = no. of adults/eJJ��ZKHUH�HYDOXDWHG�
ak=1 �����k
Q
2.4. Design and characterization of SUB/AKR chimeras
The SUB/AKR chimeras, Q38 and Q41, were designed by combin- ing linear B-cell, conformational and conformational discontinuous protective epitopes identified with sera from sheep and rabbits immunized with I. scapularis SUB and A. albopictus AKR [21,38]. Chimera Q38 was designed by combining selected SUB and AKR protective epitopes and introducing the GGGS amino acids spacer to better expose these epitopes. Chimera Q41 was designed by adding cysteins between selected SUB and AKR protective epitopes to form disulphide bridges and aid in epitope conformation. Amino acids R and K were also added to facilitate trypsin digestion and pep- tide analysis in future experiments. Protein sequences for Q38 and Q41 chimeras were analyzed using the epitope-mapping algorithm
UHSresent the reduction in the studied developmental processes (k) in vectors fed on vaccinated mice as compared to the controls fed on adjuvant/saline injected mice [32]. Data was analyzed statisti- cally to compare results for each species between individuals fed on vaccinated and control mice by x 2-test (p = 0.005) and Student’s t- test (p = 0.05). Animals were cared for in accordance with standards specified in the Guide for Care and Use of Laboratory Animals and approved by ethical committees for animal care and experimenta- tion (ISCIII CBBA/4.2-PA 225/08 and UZ PI12/10). 2.5. Tick infestations
Larvae derived from batches of 100 tick eggs were used for each mouse infestation. Mice were kept at 22 ◦C and 80–90% RH. Engorged larvae were collected three times per day during 7 days, two days after last replete larvae dropped off the host. Collected larvae were counted and incubated for molting at 24 ◦C and 90% RH.
&KDSWHU�,9
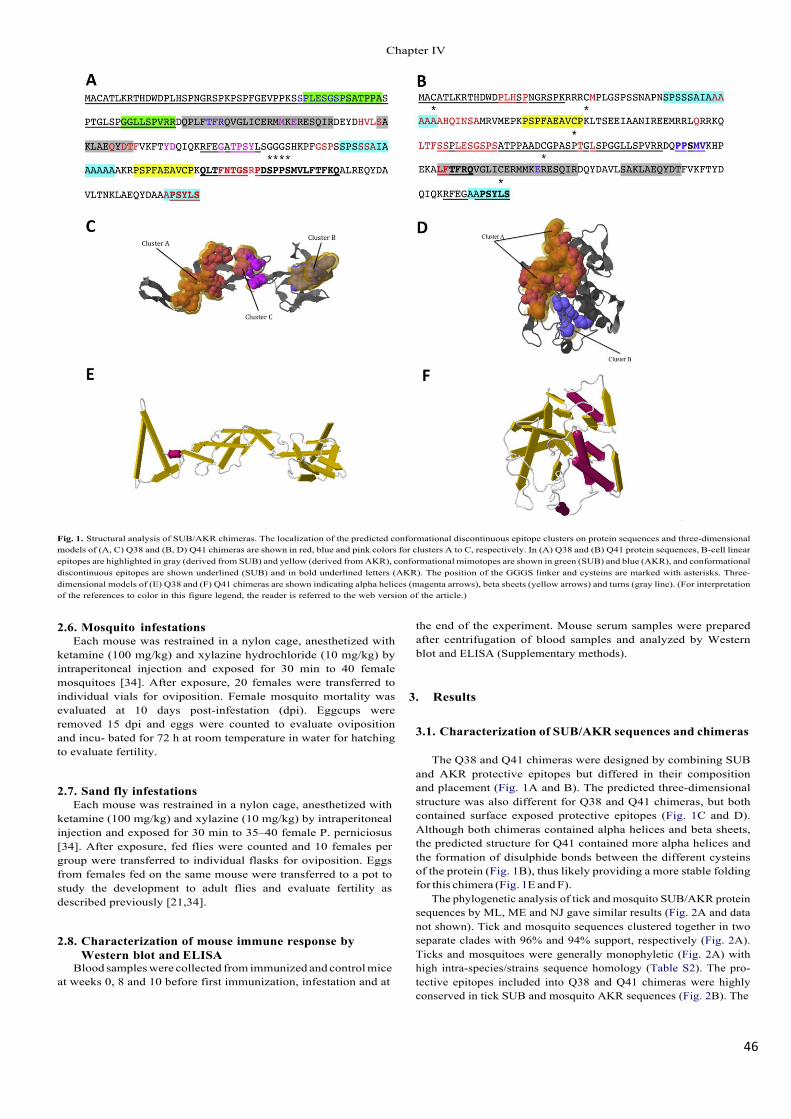
46!
Fig. 1. Structural analysis of SUB/AKR chimeras. The localization of the predicted conformational discontinuous epitope clusters on protein sequences and three-dimensional models of (A, C) Q38 and (B, D) Q41 chimeras are shown in red, blue and pink colors for clusters A to C, respectively. In (A) Q38 and (B) Q41 protein sequences, B-cell linear epitopes are highlighted in gray (derived from SUB) and yellow (derived from AKR), conformational mimotopes are shown in green (SUB) and blue (AKR), and conformational discontinuous epitopes are shown underlined (SUB) and in bold underlined letters (AKR). The position of the GGGS linker and cysteins are marked with asterisks. Three- dimensional models of (E) Q38 and (F) Q41 chimeras are shown indicating alpha helices (magenta arrows), beta sheets (yellow arrows) and turns (gray line). (For interpretation of the references to color in this figure legend, the reader is referred to the web version of the article.)
2.6. Mosquito infestations Each mouse was restrained in a nylon cage, anesthetized with
ketamine (100 mg/kg) and xylazine hydrochloride (10 mg/kg) by intraperitoneal injection and exposed for 30 min to 40 female mosquitoes [34]. After exposure, 20 females were transferred to individual vials for oviposition. Female mosquito mortality was evaluated at 10 days post-infestation (dpi). Eggcups were removed 15 dpi and eggs were counted to evaluate oviposition and incu- bated for 72 h at room temperature in water for hatching to evaluate fertility.
2.7. Sand fly infestations Each mouse was restrained in a nylon cage, anesthetized with
ketamine (100 mg/kg) and xylazine (10 mg/kg) by intraperitoneal injection and exposed for 30 min to 35–40 female P. perniciosus [34]. After exposure, fed flies were counted and 10 females per group were transferred to individual flasks for oviposition. Eggs from females fed on the same mouse were transferred to a pot to study the development to adult flies and evaluate fertility as described previously [21,34].
2.8. Characterization of mouse immune response by Western blot and ELISA
Blood samples were collected from immunized and control mice at weeks 0, 8 and 10 before first immunization, infestation and at
the end of the experiment. Mouse serum samples were prepared after centrifugation of blood samples and analyzed by Western blot and ELISA (Supplementary methods).
3. Results
3.1. Characterization of SUB/AKR sequences and chimeras
The Q38 and Q41 chimeras were designed by combining SUB and AKR protective epitopes but differed in their composition and placement (Fig. 1A and B). The predicted three-dimensional structure was also different for Q38 and Q41 chimeras, but both contained surface exposed protective epitopes (Fig. 1C and D). Although both chimeras contained alpha helices and beta sheets, the predicted structure for Q41 contained more alpha helices and the formation of disulphide bonds between the different cysteins of the protein (Fig. 1B), thus likely providing a more stable folding for this chimera (Fig. 1E and F).
The phylogenetic analysis of tick and mosquito SUB/AKR protein sequences by ML, ME and NJ gave similar results (Fig. 2A and data not shown). Tick and mosquito sequences clustered together in two separate clades with 96% and 94% support, respectively (Fig. 2A). Ticks and mosquitoes were generally monophyletic (Fig. 2A) with high intra-species/strains sequence homology (Table S2). The pro- tective epitopes included into Q38 and Q41 chimeras were highly conserved in tick SUB and mosquito AKR sequences (Fig. 2B). The
&KDSWHU�,9

47!
Fig. 2. Analysis of SUB/AKR protein sequences. (A) Phylogenetic analysis of tick and mosquito protein sequences inferred using the Maximum Likelihood method. The tree with the highest log likelihood (−2034.4263) is shown. The percentage of trees in which the associated taxa clustered together is shown next to the branches. (B) Conservation of SUB/AKR protective epitopes included into Q38 and Q41 chimeras. Identical amino acids to tick (highlighted in gray) and mosquito (in bold underlined letters) consensus sequences are shown. (C) Sequence of sand fly AKR fragment. Sequence alignment was performed with I. scapularis (AAV67031), I. ricinus (ABA62325), A. albopictus (ACF49499) and P. perniciosus (JX193855) SUB/AKR protein sequences. Identical amino acids to I. scapularis reference sequence are shown with asterisks. Identical amino acids between sand fly and mosquito sequences are marked with crosses. The sequence of the SUB/AKR protective epitope included in both Q38 and Q4 chimeras is underlined.
&KDSWHU�,9

48!
sequence obtained for the sand fly AKR protein fragment had 30–31% and 66% homology to Ixodes spp. tick and A. albopictus mosquito sequences, respectively, and showed 100% homology to one of the protective epitopes included in both Q38 and Q41 chimeras (Fig. 2C).
3.2. Mouse antibody response to vaccination
Mouse antibody response was significantly higher in vaccinated mice when compared to controls but the antibody titers were higher for Q38 and Q41 chimeras (Fig. 3B and C). Similar results were obtained in all vaccination experiments. Mouse antibodies against SUB, AKR, Q38 and Q41 were cross-reactive between them (Fig. 3A and C), suggesting conservation of the immunogenicity of SUB and AKR protective epitopes expressed in the chimeras.
3.3. Effect of vaccination on arthropod vector infestations The effect of antibodies against recombinant SUB, AKR and
SUB/AKR chimeras on tick, mosquito and sand fly vector mortality, molting, oviposition and/or fertility was evaluated in arthropods fed on vaccinated mice.
In tick larvae, mortality was not significantly affected (Table 1), despite the reduction observed in SUB and Q38 vaccinated mice when compared to controls (Fig. 4A). Larval molting to nymphs was significantly reduced by 14–48% in all vaccinated groups when compared to controls, with the highest effect in larvae derived from SUB vaccinated mice (Table 1; Fig. 4A).
In mosquitoes, only vaccination with Q41 and Q38 significantly reduced female survival (12%) and oviposition (28%), respectively, when compared to controls (Table 1; Fig. 4B). However, fertility was highly reduced (79–91% for Fertility 1 and 62–85% for Fertility 2) in mosquitoes fed on SUB, AKR and Q41 vaccinated mice, reaching the maximum values for the Q41 chimera (Table 1; Fig. 4B).
Only vaccination with SUB showed a significant effect on all developmental stages evaluated in sand flies with 20%, 16% and 58% reduction in female survival, oviposition and fertility, respectively, when compared to controls (Table 1; Fig. 4C). Vaccination with all antigens resulted in a significant reduction in sand fly oviposition (16–26%) when compared to controls (Table 1; Fig. 4C).
Vaccine E is a parameter that accounts for all effects on vector infestation observed for a given antigen. This parameter cannot be used to compare the effect of vaccination between different vector species because the developmental processes evaluated may differ for each vector. However, vaccine E is a good parameter to compare the effect of different antigens on vector infestations. The analysis of vaccine E showed that SUB gave the best results for ticks (54%) and sand flies (72%) (Fig. 4D). In mosquitoes, vaccine E was higher for SUB (92%), AKR (97%) and Q41 (99%) (Fig. 4D).
A positive correlation was shown between statistically signifi- cant effects of vaccination on vector infestations and antibody titers in vaccinated and control mice (Fig. 5A–H). Furthermore, average vaccine E also positively correlated with average antibody titers in all groups (Fig. 5I).
The results reported here showed that vaccination with SUB, AKR, Q38 and Q41 had a significant effect on I. ricinus, A. albopictus and P. perniciosus infestations when compared to controls (Table 1; Fig. 4A–D). Interestingly, vaccination with Q38 was the only result- ing in reduced oviposition in both mosquitoes (28%) and sand flies (26%) fed on vaccinated mice (Table 1; Fig. 4B and C).
4. Discussion
The control of arthropod vectors of pathogens that affect human and animal health is important for the eradication of VBD. An
important advantage of arthropod vaccines will likely be the abil- ity to reduce or prevent transmission of several pathogens through immunization of reservoir hosts and human and animal popula- tions at risk [6,9,12]. In addition, the identification of evolutionary conserved protective antigens such as SUB/AKR and the construc- tion of chimeric antigens containing conserved protective epitopes like thus shown herein may lead to development of a multi- target vaccine directed at the control of several arthropod vector species. As in previous studies [11,33,34], the hypothesis behind these experiments is that blood-sucking arthropods feeding on immunized hosts ingest antibodies specific for the target antigen that will cause deleterious effects on their feeding, developmental and reproductive performance. The exact mechanisms by which anti-SUB/AKR antibodies affect vector infestations and fertility are unknown, but may include reduction of SUB/AKR biological activ- ity and/or interaction with conserved epitopes in other proteins [33,34].
In previous experiments we demonstrated the effect of anti-AKR antibodies on the reduction in survival, oviposition and/or fertility of ticks, mosquitoes, sand flies and poultry red mites [21,33–35]. The comparison between SUB, AKR and SUB/AKR Q38 and Q41 chimeras on tick, mosquito and sad fly infestations showed that (a) SUB vaccination had the highest E across all vector species, (b) Q41 vaccination had the highest vaccine E in mosquitoes by reducing female survival and fertility, and (c) Q38 vaccination had the highest effect on reducing mosquito and sand fly oviposition. The E of SUB and AKR vaccines for the control of vector infesta- tions was expected due to previous results and the high sequence conservation for these proteins in ticks, mosquitoes and sand flies shown herein and in previous studies [21–25,33,38]. However, the distinctive effect of vaccination with Q38 and Q41 chimeras on insect fertility and/or oviposition supports the rationale for design- ing SUB/AKR chimeras to improve vaccine E against different vector species. Tick vaccines have shown that reduction in vector oviposi- tion and fertility are critical toward reducing vector populations with possible impact on VBD [6,9,42]. Additionally, vaccination with SUB has shown a reduction in A. phagocytophilum, A. marginale and Babesia bigemina infection levels in ticks fed on vaccinated hosts, thus suggesting an effect on tick vector capacity [12,43]. As previously discussed [34], differences in vaccine E between dif- ferent vector species may be due to physiological and behavioral characteristics that condition species-specific differences in the susceptibility to vaccination [16,21,44].
The vaccine E achieved in this experiment was similar to that obtained with tick vaccines [6,9,11,12,33,42,44–47] and higher than the anti-insect effects achieved in previous experiments [14–17,21,34,48,49]. Although field experiments are necessary to validate the effect of SUB/AKR vaccines on vector population con- trol, commercial use of BM86-based tick vaccines have shown that efficacies higher than 50% are sufficient to achieve tick control and reduce acaricide applications in the field [6,9,50–54].
When antibody titers in mice at time of vector infestations (week 8) were compared with vaccination effects, statistically significant correlations were obtained, strongly suggesting as in previous experiments with BM86 and SUB in cattle and deer [9,12,49,54], that the reduction in vector infestations was the result of antibodies produced against protective antigens in vaccinated mice.
The issue of SUB/AKR vaccine safety due to sequence conser- vation between arthropod vectors and vertebrate hosts has been discussed previously [21,34]. Herein, as in previous experiments with different hosts [12,21,34,43,47,54], physiological or patho- logical alterations were not observed in vaccinated mice, thus suggesting that as expected, the antibody response was directed against non-self epitopes thus reducing the possibility of detrimen- tal effects to the host. The only mouse that died in the experiment
&KDSWHU�,9
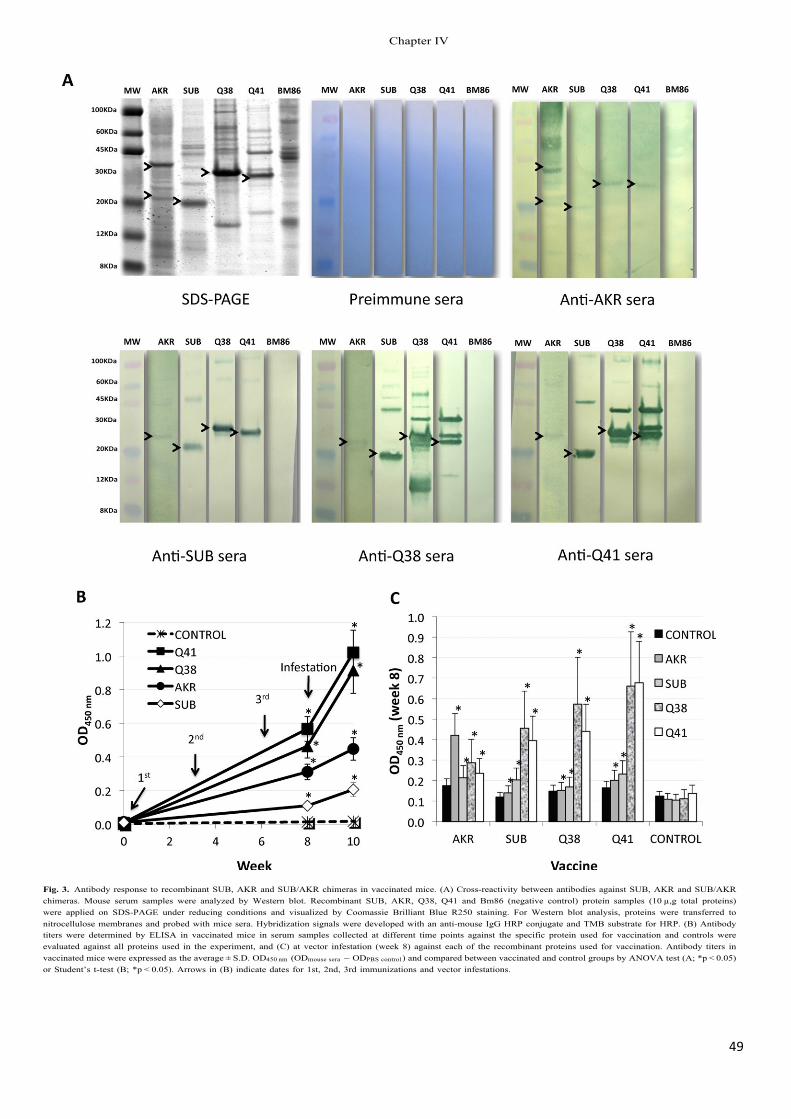
49!
Fig. 3. Antibody response to recombinant SUB, AKR and SUB/AKR chimeras in vaccinated mice. (A) Cross-reactivity between antibodies against SUB, AKR and SUB/AKR chimeras. Mouse serum samples were analyzed by Western blot. Recombinant SUB, AKR, Q38, Q41 and Bm86 (negative control) protein samples (10 µ,g total proteins) were applied on SDS-PAGE under reducing conditions and visualized by Coomassie Brilliant Blue R250 staining. For Western blot analysis, proteins were transferred to nitrocellulose membranes and probed with mice sera. Hybridization signals were developed with an anti-mouse IgG HRP conjugate and TMB substrate for HRP. (B) Antibody titers were determined by ELISA in vaccinated mice in serum samples collected at different time points against the specific protein used for vaccination and controls were evaluated against all proteins used in the experiment, and (C) at vector infestation (week 8) against each of the recombinant proteins used for vaccination. Antibody titers in vaccinated mice were expressed as the average ± S.D. OD450 nm (ODmouse sera − ODPBS control ) and compared between vaccinated and control groups by ANOVA test (A; *p < 0.05) or Student’s t-test (B; *p < 0.05). Arrows in (B) indicate dates for 1st, 2nd, 3rd immunizations and vector infestations.
&KDSWHU�,9
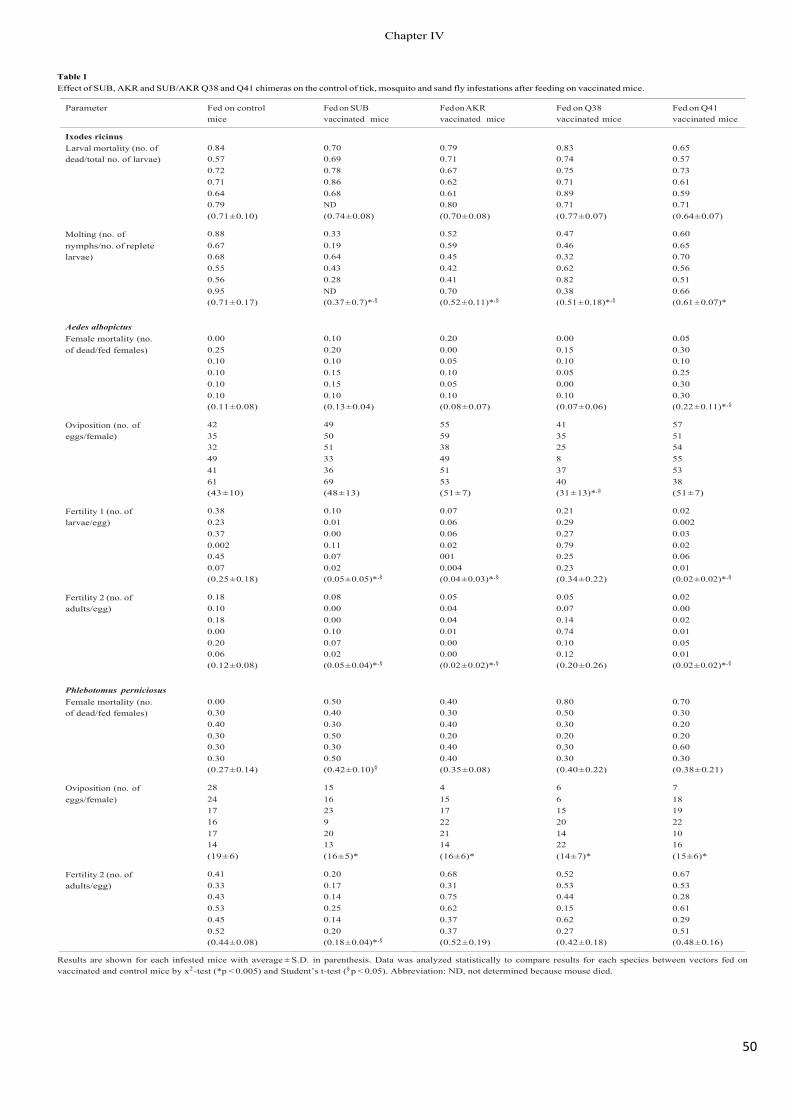
50!
Table 1 Effect of SUB, AKR and SUB/AKR Q38 and Q41 chimeras on the control of tick, mosquito and sand fly infestations after feeding on vaccinated mice.
Parameter Fed on control mice
Fed on SUB vaccinated mice
Fed on AKR vaccinated mice
Fed on Q38 vaccinated mice
Fed on Q41 vaccinated mice
Ixodes ricinus Larval mortality (no. of 0.84 0.70 0.79 0.83 0.65 dead/total no. of larvae) 0.57 0.69 0.71 0.74 0.57
0.72 0.78 0.67 0.75 0.73 0.71 0.86 0.62 0.71 0.61 0.64 0.68 0.61 0.89 0.59 0.79 ND 0.80 0.71 0.71 (0.71 ± 0.10) (0.74 ± 0.08) (0.70 ± 0.08) (0.77 ± 0.07) (0.64 ± 0.07)
Molting (no. of 0.88 0.33 0.52 0.47 0.60 nymphs/no. of replete 0.67 0.19 0.59 0.46 0.65 larvae) 0.68 0.64 0.45 0.32 0.70
0.55 0.43 0.42 0.62 0.56 0.56 0.28 0.41 0.82 0.51 0.95 (0.71 ± 0.17)
ND (0.37 ± 0.7)*,§
0.70 (0.52 ± 0.11)*,§
0.38 (0.51 ± 0.18)*,§
0.66 (0.61 ± 0.07)*
Aedes albopictus Female mortality (no. 0.00 0.10 0.20 0.00 0.05 of dead/fed females) 0.25 0.20 0.00 0.15 0.30
0.10 0.10 0.05 0.10 0.10 0.10 0.15 0.10 0.05 0.25 0.10 0.15 0.05 0.00 0.30 0.10 (0.11 ± 0.08)
0.10 (0.13 ± 0.04)
0.10 (0.08 ± 0.07)
0.10 (0.07 ± 0.06)
0.30 (0.22 ± 0.11)*,§
Oviposition (no. of 42 49 55 41 57 eggs/female) 35 50 59 35 51
32 51 38 25 54 49 33 49 8 55 41 36 51 37 53 61 (43 ± 10)
69 (48 ± 13)
53 (51 ± 7)
40 (31 ± 13)*,§
38 (51 ± 7)
Fertility 1 (no. of 0.38 0.10 0.07 0.21 0.02 larvae/egg) 0.23 0.01 0.06 0.29 0.002
0.37 0.00 0.06 0.27 0.03 0.002 0.11 0.02 0.79 0.02 0.45 0.07 001 0.25 0.06 0.07 (0.25 ± 0.18)
0.02 (0.05 ± 0.05)*,§
0.004 (0.04 ± 0.03)*,§
0.23 (0.34 ± 0.22)
0.01 (0.02 ± 0.02)*,§
Fertility 2 (no. of 0.18 0.08 0.05 0.05 0.02 adults/egg) 0.10 0.00 0.04 0.07 0.00
0.18 0.00 0.04 0.14 0.02 0.00 0.10 0.01 0.74 0.01 0.20 0.07 0.00 0.10 0.05 0.06 (0.12 ± 0.08)
0.02 (0.05 ± 0.04)*,§
0.00 (0.02 ± 0.02)*,§
0.12 (0.20 ± 0.26)
0.01 (0.02 ± 0.02)*,§
Phlebotomus perniciosus Female mortality (no. 0.00 0.50 0.40 0.80 0.70 of dead/fed females) 0.30 0.40 0.30 0.50 0.30
0.40 0.30 0.40 0.30 0.20 0.30 0.50 0.20 0.20 0.20 0.30 0.30 0.40 0.30 0.60 0.30 (0.27 ± 0.14)
0.50 (0.42 ± 0.10)§
0.40 (0.35 ± 0.08)
0.30 (0.40 ± 0.22)
0.30 (0.38 ± 0.21)
Oviposition (no. of 28 15 4 6 7 eggs/female) 24 16 15 6 18
17 23 17 15 19 16 9 22 20 22 17 20 21 14 10 14 13 14 22 16 (19 ± 6) (16 ± 5)* (16 ± 6)* (14 ± 7)* (15 ± 6)*
Fertility 2 (no. of 0.41 0.20 0.68 0.52 0.67 adults/egg) 0.33 0.17 0.31 0.53 0.53
0.43 0.14 0.75 0.44 0.28 0.53 0.25 0.62 0.15 0.61 0.45 0.14 0.37 0.62 0.29 0.52 (0.44 ± 0.08)
0.20 (0.18 ± 0.04)*,§
0.37 (0.52 ± 0.19)
0.27 (0.42 ± 0.18)
0.51 (0.48 ± 0.16)
Results are shown for each infested mice with average ± S.D. in parenthesis. Data was analyzed statistically to compare results for each species between vectors fed on vaccinated and control mice by x2 -test (*p < 0.005) and Student’s t-test (§ p < 0.05). Abbreviation: ND, not determined because mouse died.
&KDSWHU�,9
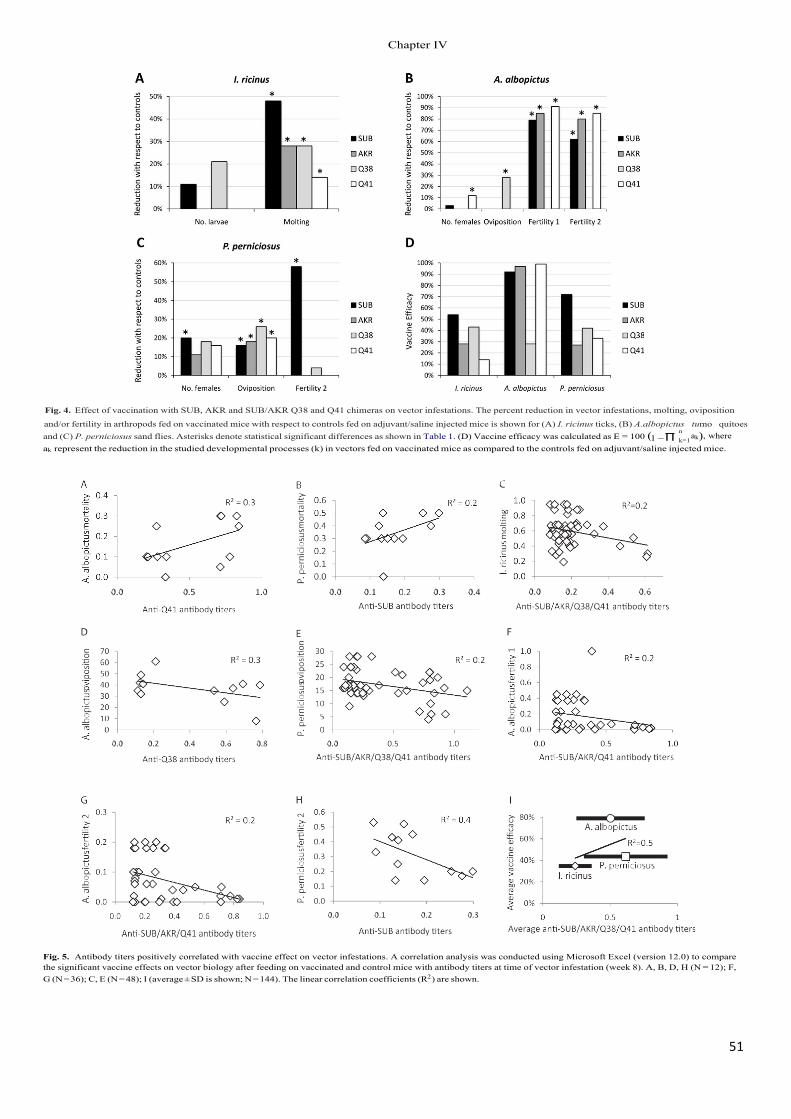
51!
Fig. 4. Effect of vaccination with SUB, AKR and SUB/AKR Q38 and Q41 chimeras on vector infestations. The percent reduction in vector infestations, molting, oviposition
and/or fertility in arthropods fed on vaccinated mice with respect to controls fed on adjuvant/saline injected mice is shown for (A) I. ricinus ticks, (B) A.albopLFWXV� tumo quitoesand (C) P. perniciosus sand flies. Asterisks denote statistical significant differences as shown in Table 1. (D) Vaccine efficacy was calculated as E = 100�� l −ˉ
nak�, where
k=1 ak represent the reduction in the studied developmental processes (k) in vectors fed on vaccinated mice as compared to the controls fed on adjuvant/saline injected mice.
Fig. 5. Antibody titers positively correlated with vaccine effect on vector infestations. A correlation analysis was conducted using Microsoft Excel (version 12.0) to compare the significant vaccine effects on vector biology after feeding on vaccinated and control mice with antibody titers at time of vector infestation (week 8). A, B, D, H (N = 12); F, G (N = 36); C, E (N = 48); I (average ± SD is shown; N = 144). The linear correlation coefficients (R2 ) are shown.
&KDSWHU�,9

52!
corresponded to a SUB-vaccinated mouse that died after blood sampling (Table 1).
5. Conclusions
In summary, the results reported here showed that recombi- nant SUB, AKR and SUB/AKR chimeras could be used to develop vaccines for the control of tick, mosquito and sand fly infestations with possible impact on the control of other arthropod vectors. As previously shown for tick vaccines [11], reduction in insect ovipo- sition obtained with Q38 chimera would result in the reduction of insect populations in subsequent generations. If effective, these vaccines could be used to immunize reservoir hosts to reduce vector populations and thus the risk of pathogen transmission to humans and animals.
Acknowledgments
We thank all collaborators who provided tick and mosquito samples for analysis. This research was supported by grants FAU2008-00014-00-00 and PEII09-0118-8907 to JF. M. Villar was funded by the JAE-DOC program (CSIC-FSE), Spain.
Appendix A. SUB/AKR Vaccine Study Group
Pedro M. Alarcón-Elbal, Sarah Delacour, Vladimir Oropeza, Ignacio Ruiz (Departamento de Patología Animal, Facultad de Veterinaria, C/Miguel Servet 177, 50013 Zaragoza, Spain), Car- los R. Prudencio (Laboratório de Nanobiotecnologia, Instituto de Genética e Bioquímica, Universidade Federal de Uberlândia-UFU, Campus Umuarama 2E-248, 38400-902 Uberlândia, MG, Brazil), Ruth C. Galindo (Instituto de Investigación en Recursos Cinegéti- cos IREC-CSIC-UCLM-JCCM, Ronda de Toledo s/n, 13005 Ciudad Real, Spain), Consuelo Almazán (Facultad de Medicina Veterinaria y Zootecnia, Universidad Autónoma de Tamaulipas, Km. 5 car- retera Victoria-Mante, CP 87000 Cd. Victoria, Tamaulipas, Mexico), Ard M. Nijhof (Institute for Parasitology and Tropical Veterinary Medicine, Freie Universität Berlin, Königsweg 67, 14163 Berlin, Germany), and Atilio J. Mangold (Instituto Nacional de Tecnología Agropecuaria, Estación Experimental Agropecuaria Rafaela, CC 22, CP 2300 Rafaela, Santa Fe, Argentina).
Appendix B. Supplementary data
Supplementary data associated with this article can be found, in the online version, at http://dx.doi.org/10.1016/ j.vaccine.2012.12.073.
References
[1] Jones KE, Patel NG, Levy MA, Storeygard A, Balk D, Gittleman JL, et al. Global trends in emerging infectious diseases. Nature 2008;451:990–4.
[2] Estrada-Pen a A, Ayllón N, de la Fuente J. Impact of climate trends on tick-borne pathogen transmission. Front Physiol 2012;3:64.
[3] Halstead SB. Dengue. Lancet 2007;370:1644–52. [4] Chappuis F, Sundar S, Hailu A, Ghalib H, Rijal S, Peeling RW, et al. Visceral
leishmaniasis: what are the needs for diagnosis, treatment and control? Nat Rev Microbiol 2007;5:873–82.
[5] Hayes EB. Is it time for a new yellow fever vaccine? Vaccine 2010;28:8073–6. [6] de la Fuente J, Kocan KM. Advances in the identification and characterization
of protective antigens for development of recombinant vaccines against tick infestations. Expert Rev Vaccines 2003;2:583–93.
[7] Kishore K, Kumar V, Kesari S, Dinesh DS, Kumar AJ, Das P, et al. Vector control in leishmaniasis. Indian J Med Res 2006;123:467–72.
[8] Speranc a MA, Capurro ML. Perspectives in the control of infectious diseases by transgenic mosquitoes in the post-genomic era – a review. Mem Inst Oswaldo Cruz 2007;102:425–33.
[9] de la Fuente J, Rodríguez M, Redondo M, Montero C, García-García JC, Méndez L, et al. Field studies and cost-effectiveness analysis of vaccination with GavacTM�
against the cattle tick Boophilus microplus. Vaccine 1998;16:366–73.
[10] Karunamoorthi K. Vector control: a cornerstone in the malaria elimination campaign. Clin Microbiol Infect 2011;17:1608–16.
[11] de la Fuente J, Almazán C, Canales M, Pérez de la Lastra JM, Kocan KM, Willadsen P. A ten-year review of commercial vaccine performance for control of tick infestations on cattle. Anim Health Res Rev 2007;8:23–8.
[12] Merino O, Almazán C, Canales M, Villar M, Moreno-Cid JA, Galindo RC, et al. Targeting the tick protective antigen subolesin reduces vector infestations and pathogen infection by Anaplasma marginale and Babesia bigemina. Vaccine 2011;29:8575–9.
[13] Valenzuela JG, Belkaid Y, Garfield MK, Mendez S, Kamhawi S, Rowton ED, et al. Toward a defined anti-Leishmania vaccine targeting vector antigens: charac- terization of a protective salivary protein. J Exp Med 2001;194:331–42.
[14] Lal AA, Patterson PS, Sacci JB, Vaughan JA, Paul C, Collins WE, et al. Anti- mosquito midgut antibodies block development of Plasmodium falciparum and Plasmodium vivax in multiple species of Anopheles mosquitoes and reduce vector fecundity and survivorship. Proc Natl Acad Sci USA 2001;98: 5228–33.
[15] Almeida AP, Billingsley PF. Induced immunity against the mosquito Anophe- les stephensi (Diptera: Culicidae): effects of cell fraction antigens on survival, fecundity, and Plasmodium berghei (Eucoccidiida: Plasmodiidae) transmission. J Med Entomol 2002;39:207–14.
[16] Suneja A, Gulia M, Gakhar SK. Blocking of malaria parasite development in mosquito and fecundity reduction by midgut antibodies in Anopheles stephensi (Diptera: Culicidae). Arch Insect Biochem Physiol 2003;52:63–70.
[17] Milleron RS, Ribeiro JM, Elnaime D, Soong L, Lanzaro G. Negative effect of anti- bodies against maxadilan on the fitness of the sand fly vector of American visceral leishmaniasis. Am J Trop Med Hyg 2004;70:278–85.
[18] Titus RG, Bishop JV, Mejia JS. The immunomodulatory factors of arthropod saliva and the potential for these factors to serve as vaccine targets to prevent pathogen transmission. Parasite Immunol 2006;4:131–41.
[19] Kedzierski L, Zhu Y, Handman E. Leishmania vaccines: progress and problems. Parasitology 2006;133:S87–112.
[20] Saul A. Mosquito stage, transmission blocking vaccines for malaria. Curr Opin Infect Dis 2007;5:476–81.
[21] Canales M, Naranjo V, Almazán C, Molina R, Tsuruta SA, Szabó MPJ, et al. Con- servation and immunogenicity of the mosquito ortholog of the tick protective antigen, subolesin. Parasitol Res 2009;105:97–111.
[22] de la Fuente J, Almazán C, Blas-Machado U, Naranjo V, Mangold AJ, Blouin EF, et al. The tick protective antigen, 4D8, is a conserved protein involved in mod- ulation of tick blood ingestion and reproduction. Vaccine 2006;24:4082–95.
[23] Macqueen DJ, Johnston IA. Evolution of the multifaceted eukaryotic akirin gene family. BMC Evol Biol 2009;9:34.
[24] Galindo RC, Doncel-Pérez E, Zivkovic Z, Naranjo V, Gortazar C, Mangold AJ, et al. Tick subolesin is an ortholog of the akirins described in insects and vertebrates. Dev Comp Immunol 2009;33:612–7.
[25] Mangold AJ, Galindo RC, de la Fuente J. Response to the commentary of D. Macqueen on: Galindo RC, Doncel-Pérez E, Zivkovic Z, Naranjo V, Gortazar C, Mangold AJ, et al. Tick subolesin is an ortholog of the akirins described in insects and vertebrates [Dev. Comp. Immunol. 33 (2009) 612–617]. Dev Comp Immunol 2009;33:878–9.
[26] Goto A, Matsushita K, Gesellchen V, El Chamy L, Kuttenkeuler D, Takeuchi O, et al. Akirins are highly conserved nuclear proteins required for NF- kappaB-dependent gene expression in drosophila and mice. Nat Immunol 2008;9:97–104.
[27] de la Fuente J, Maritz-Olivier C, Naranjo V, Ayoubi P, Nijhof AM, Almazán C, et al. Evidence of the role of tick subolesin in gene expression. BMC Genomics 2008;9:372.
[28] Nowak SJ, Aihara H, Gonzalez K, Nibu Y, Baylies MK. Akirin links twist-regulated transcription with the Brahma chromatin remodeling complex during embryo- genesis. PLoS Genet 2012;8:e1002547.
[29] de la Fuente J, Almazán C, Naranjo V, Blouin EF, Meyer JM, Kocan KM. Autocidal control of ticks by silencing of a single gene by RNA interference. Biochem Biophys Res Commun 2006;344:332–8.
[30] Kocan KM, Zivkovic Z, Blouin EF, Naranjo V, Almazán C, Mitra R, et al. Silencing of genes involved in Anaplasma marginale-tick interactions affects the pathogen developmental cycle in Dermacentor variabilis. BMC Dev Biol 2009;9:42.
[31] Zivkovic Z, Torina A, Mitra R, Alongi A, Scimeca S, Kocan KM, et al. Subolesin expression in response to pathogen infection in ticks. BMC Immunol 2010;11:7.
[32] Almazán C, Kocan KM, Bergman DK, Garcia-Garcia JC, Blouin EF, de la Fuente J. Identification of protective antigens for the control of Ixodes scapularis infestations using cDNA expression library immunization. Vaccine 2003;21:1492–501.
[33] de la Fuente J, Moreno-Cid JA, Canales M, Villar M, Pérez de la Lastra JM, Kocan KM, et al. Targeting arthropod subolesin/akirin for the development of a uni- versal vaccine for control of vector infestations and pathogen transmission. Vet Parasitol 2011;181:17–22.
[34] Moreno-Cid JA, Jiménez M, Cornelie S, Molina R, Alarcón P, Lacroix M-N, et al. Characterization of Aedes albopictus akirin for the control of mosquito and sand fly infestations. Vaccine 2011;29:77–82.
[35] Harrington D, Canales M, de la Fuente J, de Luna C, Robinson K, Guy J, et al. Immunisation with recombinant proteins subolesin and Bm86 for the control of Dermanyssus gallinae in poultry. Vaccine 2009;27:4056–63.
[36] Manzano-Román R, Díaz-Martín V, Oleaga A, Siles-Lucas M, Pérez-Sánchez R. Subolesin/akirin orthologs from Ornithodoros spp. soft ticks: cloning, RNAi gene silencing and protective effect of the recombinant proteins. Vet Parasitol 2012;185:248–59.
&KDSWHU�,9

53!
[37] Carpio Y, Basabe L, Acosta J, Rodríguez A, Mendoza A, Lisperger A, et al. Novel gene isolated from Caligus rogercresseyi: a promising target for vaccine devel- opment against sea lice. Vaccine 2011;29:2810–20.
[38] Prudencio CR, Pérez de la Lastra JM, Canales M, Villar M, de la Fuente J. Map- ping protective epitopes in the tick and mosquito subolesin ortholog proteins. Vaccine 2010;28:5398–406.
[39] Mayrose I, Penn O, Erez E, Rubinstein ND, Shlomi T, Freund NT, et al. Pepitope: epitope mapping from affinity-selected peptides. Bioinformatics 2007;23:3244–6.
[40] Canales M, Pérez de la Lastra JM, Naranjo V, Nijhof AM, Hope M, Jongejan F, et al. Expression of recombinant Rhipicephalus (Boophilus) microplus R. annu- latus and R. decoloratus Bm86 orthologs as secreted proteins in Pichia pastoris. BMC Biotechnol 2008;8:14.
[41] Canales M, Ballesteros C, Moreno-Cid JA, Espinosa AM, Villar M, de la Fuente J. Extractive bioconversion to produce the Aedes albopictus akirin in an aque- ous two-phase system supporting Pichia pastoris growth and protein secretion. Biochem Eng J 2009;46:105–14.
[42] Willadsen P. Anti-tick vaccines. Parasitology 2004;129:S367–874. [43] de la Fuente J, Almazán C, Blouin EF, Naranjo V, Kocan KM. Reduction of tick
infections with Anaplasma marginale and A. phagocytophilum by targeting the tick protective antigen subolesin. Parasitol Res 2006;100:85–91.
[44] Canales M, Almazán C, Naranjo V, Jongejan F, de la Fuente J. Vaccination with recombinant Boophilus annulatus Bm86 ortholog protein, Ba86, protects cattle against B. annulatus and B. microplus infestations. BMC Biotechnol 2009;9:29.
[45] Almazán C, Kocan KM, Blouin EF, de la Fuente J. Vaccination with recombi- nant tick antigens for the control of Ixodes scapularis adult infestations. Vaccine 2005;23:5294–8.
[46] Trimnell AR, Davies GM, Lissina O, Hails RS, Nuttall PA. A cross-reactive tick cement antigen is a candidate broad-spectrum tick vaccine. Vaccine 2005;23:4329–41.
[47] Carreón D, Pérez de la Lastra JM, Almazán C, Canales M, Ruiz-Fons F, Boadella M, et al. Vaccination with BM86, subolesin and akirin protective antigens for the control of tick infestations in white tailed deer and red deer. Vaccine 2012;30:273–9.
[48] Alger NE, Cabrera EJ. An increase in death rate of Anopheles stephensi fed on rabbit immunized with mosquito antigen. J Econ Entomol 1972;65:165–8.
[49] Hatfield PR. Detection and localization of antibody ingested with a mosquito bloodmeal. Med Vet Entomol 1988;2:339–45.
[50] Canales M, Enriquez A, Ramos E, Cabrera D, Dandie H, Soto A, et al. Large-scale production in Pichia pastoris of the recombinant vaccine GavacTM against cattle ticks. Vaccine 1997;15:414–22.
[51] Labarta V, Rodríguez M, Penichet M, Lleonart R, Lorenzo Luaces L, de la Fuente J. Simulation of control strategies for the cattle tick Boophilus microplus employ- ing vaccination with a recombinant Bm86 antigen preparation. Vet Parasitol 1996;63:131–60.
[52] Lodos J, Ochogavía ME, Rodríguez M, de la Fuente J. A simulation study of the effects of acaricides and vaccination on Boophilus cattle-tick populations. Prev Vet Med 1999;38:47–63.
[53] Lodos J, Boué O, de la Fuente J. A model to simulate the effect of vaccination against Boophilus ticks on cattle. Vet Parasitol 2000;87:315–26.
[54] Merino O, Almazán C, Canales M, Villar M, Moreno-Cid JA, Estrada-Pen a A, et al. Control of Rhipicephalus (Boophilus) microplus infestations by the combi- nation of subolesin vaccination and tick autocidal control after subolesin gene knockdown in ticks fed on cattle. Vaccine 2011;29:2248–54.
&KDSWHU�,9


CONTROL'OF'TICK'INFESTATIONS'IN'CATTLE'VACCINATED'
WITH'BACTERIAL'MEMBRANES'CONTAINING'SURFACE9
EXPOSED'TICK'PROTECTIVE'ANTIGENS

55!
Contents lists available at SciVerse ScienceDirect
Vaccine
jou rn a l h om epa ge: www.elsevier .com/locate /vaccine
Control of tick infestations in cattle vaccinated with bacterial membranes containing surface-exposed tick protective antigens
Consuelo Almazán a , Orlando Moreno-Cantú a , Juan A. Moreno-Cid b , Ruth C. Galindo b , Mario Canales b,c , Margarita Villar b , José de la Fuente b,d,∗
a Facultad de Medicina Veterinaria y Zootecnia, Universidad Autónoma de Tamaulipas, Km. 5 carretera Victoria-Mante, CP 87000 Ciudad Victoria, Tamaulipas, Mexico b Instituto de Investigación en Recursos Cinegéticos IREC-CSIC-UCLM-JCCM, Ronda de Toledo s/n, 13005 Ciudad Real, Spain c Instituto de Higiene e Medicina Tropical, Universidade Nova de Lisboa, Rua da Junqueira 100, Lisboa 1349-008, Portugal d Departmentof Veterinary Pathobiology, Center forVeterinary Health Sciences, Oklahoma State University, Stillwater, OK 74078, USA
a r t i c l e i n f o a b s t r a c t
Article history: Received 9 June 2011 Received in revised form 22 October 2011 Accepted 31 October 2011 Available online 12 November 2011
Keywords: Tick Boophilus Anaplasma MSP1a Vaccine Bm86 Bm95 Subolesin Ubiquitin Elongation factor
Vaccines containing the Rhipicephalus (Boophilus) microplus BM86 and BM95 antigens protect cattle against tick infestations. Tick subolesin (SUB), elongation factor 1a (EF1a) and ubiquitin (UBQ) are new candidate protective antigens for the control of cattle tick infestations. Previous studies showed that R. microplus BM95 immunogenic peptides fused to the Anaplasma marginale major surface protein (MSP) 1a N-terminal region (BM95-MSP1a) for presentation on the Escherichia coli membrane were protec- tive against R. microplus infestations in rabbits. In this study, we extended these results by expressing SUB-MSP1a, EF1a-MSP1a and UBQ-MSP1a fusion proteins on the E. coli membrane using this system and demonstrating that bacterial membranes containing the chimeric proteins BM95-MSP1a and SUB-MSP1a were protective (>60% vaccine efficacy) against experimental R. microplus and Rhipicephalus annulatus infestations in cattle. This system provides a novel, simple and cost-effective approach for the production of tick protective antigens by surface display of antigenic protein chimera on the E. coli membrane and demonstrates the possibility of using recombinant bacterial membrane fractions in vaccine preparations to protect cattle against tick infestations.
© 2011 Elsevier Ltd. All rights reserved.
1. IntroductionThe cattle ticks, Rhipicephalus (Boophilus) microplus and Rhipi-
cephalus annulatus, are distributed in tropical and subtropical regions of the world [1–5]. Infestations with R. microplus and R. annulatus economically impact cattle industry by reducing weight gain and milk production, and by transmitting pathogens that cause babesiosis (Babesia bovis and Babesia bigemina) and anaplasmosis (Anaplasma marginale) [2,4,6].
Although acaricides constitute a major component of integrated cattle tick control strategies, their use has had limited efficacy in reducing tick infestations and is often accompanied by seri- ous drawbacks, including the selection of acaricide-resistant ticks, environmental contamination and contamination of milk and meat products with drug residues [7]. All of these considerations rein- force the need for alternative approaches to control of cattle tick
∗ Corresponding author.E-mail addresses: jose [email protected], [email protected]
(J. de la Fuente).
infestations such as the use of hosts with natural resistance to ticks, pheromone-impregnated decoys for attracting and killing ticks, biological control agents and vaccines [8–10].
In the early 1990s, vaccines containing the recombinant R. microplus BM86 and BM95 gut antigens that induced immuno- logical protection against cattle tick infestations were developed and commercialized [10–19]. These vaccines reduce the number of engorging female ticks, their weight and reproductive capacity. Thus the greatest vaccine effect was the reduction of larval infesta- tions in subsequent generations. Vaccine field trials demonstrated that control of cattle ticks by vaccination has the advantages of being cost-effective, reducing environmental contamination and preventing the selection of drug resistant ticks that result from repeated acaricide application [15,16]. In addition, these vaccines may also prevent or reduce transmission of pathogens by reducing tick populations and/or affecting tick vector capacity [15–17].
Despite the demonstrated effect of commercial BM86/BM95 vaccines for the control of cattle tick infestations, they show tick strain-to-strain variations in vaccine efficacy and the pro- duction of recombinant antigens is expensive and technologically complex [8,10,14,15,20–24]. Adding new tick protective antigens
0264-410X/$ – see front matter © 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.vaccine.2011.10.102
&KDSWHU�,9

56!
to vaccine formulations and developing production systems that will reduce the cost while increasing the immunogenicity of recombinant antigens could overcome these limitations of current cattle tick vaccines.
In a recent work, we demonstrated that a recombinant pro- tein comprising the BM95 immunogenic peptides fused to the A. marginale MSP1a N-terminal region (BM95-MSP1a) is surface- exposed on the Escherichia coli membrane, resulting in a simple and cost-effective process for the production of a vaccine prepara- tion that was protective against R. microplus infestations in rabbits [24–26]. These studies suggested the possibility of using recombinant bacterial membrane fractions containing the surface-exposed BM95-MSP1a chimeric antigen as a cheaper preparation for vaccination against cattle tick infestations [24–26]. However, these vaccination experiments were conducted as a preliminary study in rabbits that are not the natural host for Rhipicephalus (Boophilus) spp.
In the study reported herein, we conducted a vaccination trial in cattle to demonstrate the efficacy of recombinant E. coli mem- branes containing the BM95-MSP1a chimera for the control of R. microplus and R. annulatus infestations and to extend to other can- didate protective antigens the possibility of using recombinant bacterial membrane fractions containing surface-expose chimera as a simple and cost-effective approach for the production of tick vaccines.
2. Material and methods
2.1. Construction and expression of tick protective antigen chimeras
The construction and expression of the BM95 chimera was previously reported [25]. The BM95-MSP1a chimera encoded a protein containing three BM95 (Genbank accession number AAD38381) immunogenic and protective peptides fused to A. marginale major surface protein 1a (MSP1a) N-terminal region to surface expose the BM95 peptides when expressed in E. coli [25]. For the construction of tick subolesin (SUB-MSP1a), elongation fac- tor 1a (EF1a-MSP1a) and ubiquitin (UBQ-MSP1a) chimeras, protein coding regions (amino acids 1–146, 1–462, and 45–132, for SUB (DQ159964.1), EF1a (EU436163.1) and UBQ (AF506022), respec- tively) were amplified by PCR using oligonucleotide primers (SUB: pMB4D8R 51�-GATGGAATTCTGTTCTGCGAGTTTGGTAGATAG-31 and
pMB4D8L 51 -CTCCTCGAGATGGCTTGCGCAACATTAAAGCGAAC-31 , EF1a: GIIR 51 -GGCTCGAGAGGCCCACGGACAAACCCCTC-31 and GII-L 51 -GGGAATTCTCAGCGGCCTTGGTGACCTTGCC-31 , UBQ: UBIQ2-L 51 -GGCTCGAGATGCAAATCTTCGTTAAGACA-31 and UBIQ-R 51 - GGGAATTCTCGAGGGTACGCCCATCTTCTAG-31 ) that introduced XhoI and EcoRI restrictions sites for cloning into the pFLAG-GTC expression plasmid [25]. In these constructs, as for the BM95- MSP1a chimera, the inserted coding region is fused to MSP1a and is under the control of the inducible tac promoter [25].
2.2. Recombinant vaccine production For production of recombinant BM95-MSP1a, SUB-MSP1a, EF1a-
MSP1a and UBQ-MSP1a fusion proteins, E. coli were propagated in 10 ml Luria–Bertani (LB) broth containing 50 µg/ml ampicillin and 0.4% glucose (LBAG) for 2 h at 37 ◦C and 200 rpm and then used to inoculate 250 ml cultures. The 250 ml cultures were grown under the same conditions for 4 h to reach an OD600 nm = 1 before inoculation of a 4 l working volume Biostat B bioreactor (B. Braun Biotech, Melsungen, Germany). Fermentation was done as described previously [25] but continuing fermentation during 5.5 h after addition of 0.5 mM final concentration of isopropyl- í3-d-thiogalactopyranoside (IPTG) for induction of recombinant protein expression. Cell growth was monitored by measuring OD600 nm throughout both propagation and fermentation steps. The cells were harvested and disrupted using a cell sonicator (Model MS73; Bandelin Sonopuls, Berlin, Germany) described previously [25]. After disruption, membrane-bound insoluble protein frac- tions were separated by centrifugation at 21,500 × g for 15 min at 4 ◦C and stored at −20 ◦C. Protein concentrations were determinedusing the Nanodrop 1000 (Thermo Scientific, Wilmington, DE, USA). For vaccine formulation, the amount of membrane-bound insoluble protein fraction was adjusted to contain 120 µg of the recombinant chimera that were adjuvated in Montanide ISA 50 V2 (Seppic, Paris, France) in 1 ml doses.
2.3. Recombinant protein characterization
The expression of recombinant proteins was analyzed by SDS-polyacrylamide gel electrophoresis (SDS-PAGE) and Western blot (Fig. 1). Ten micrograms of total proteins were loaded onto a 12% SDS-polyacrylamide precast gel (Expedeon Protein Solutions, Harston Cambridgeshire, UK) and electrophoresed for 3 h at 90 mA
Fig. 1. Expression of recombinant UBQ-MSP1a, BM95-MSP1a, EF1a-MSP1a and SUB-MSP1a fusion proteins in E. coli. Recombinant E. coli were induced for 5.5 h with IPTG for the expression of chimeric proteins. 10 µg of total proteins (pellet 1 in Fig. 2) were loaded per well in a 12% SDS-polyacrylamide gel. ColorBurst electrophoresis markers (Sigma) were used as molecular weight (MW) markers in the electrophoresis (lanes 1 and 10). Proteins were stained with Coomassie based Instant Blue (lanes 2, 4, 6 and 8) and transferred to a nitrocellulose membrane and probed with rabbit anti-MSP1a antibodies and developed with a goat anti-rabbit HRP conjugate (lanes 3, 5, 7 and 9).Recombinant UBQ-MSP1a, BM95-MSP1a, EF1a-MSP1a and SUB-MSP1a chimeric proteins are indicated with arrows.
&KDSWHU�,9
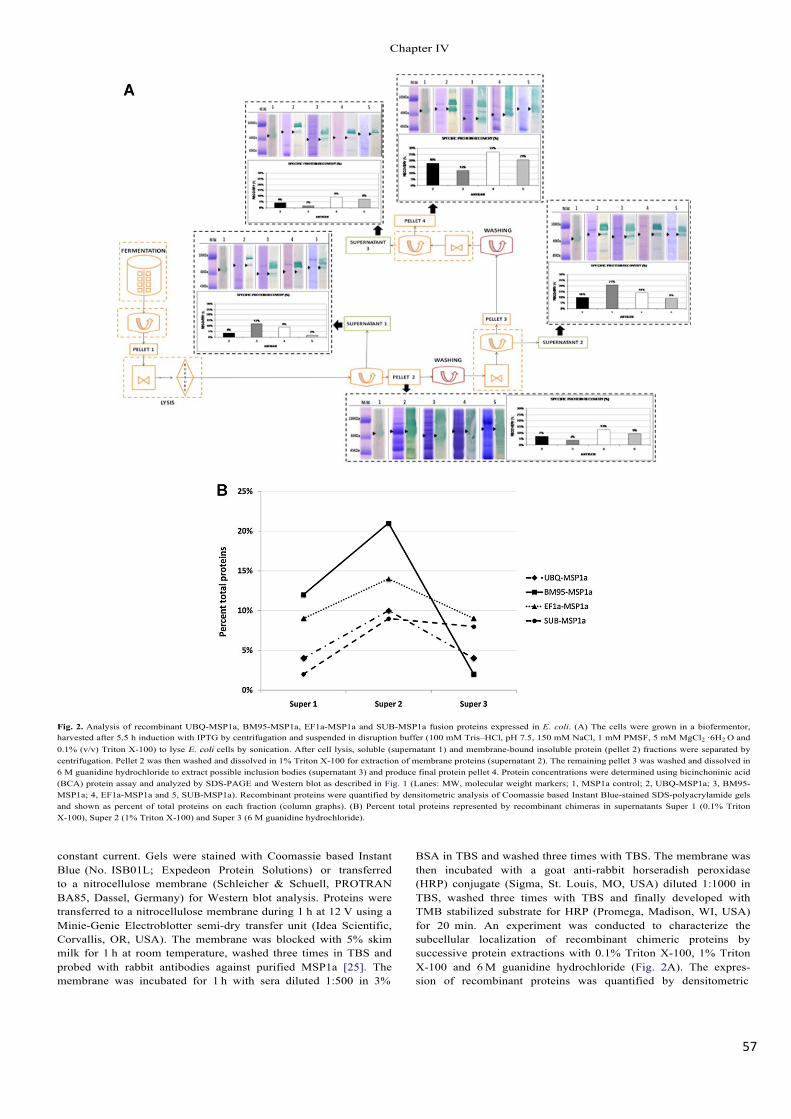
57!
Fig. 2. Analysis of recombinant UBQ-MSP1a, BM95-MSP1a, EF1a-MSP1a and SUB-MSP1a fusion proteins expressed in E. coli. (A) The cells were grown in a biofermentor, harvested after 5.5 h induction with IPTG by centrifugation and suspended in disruption buffer (100 mM Tris–HCl, pH 7.5, 150 mM NaCl, 1 mM PMSF, 5 mM MgCl2 ·6H2 O and 0.1% (v/v) Triton X-100) to lyse E. coli cells by sonication. After cell lysis, soluble (supernatant 1) and membrane-bound insoluble protein (pellet 2) fractions were separated by centrifugation. Pellet 2 was then washed and dissolved in 1% Triton X-100 for extraction of membrane proteins (supernatant 2). The remaining pellet 3 was washed and dissolved in 6 M guanidine hydrochloride to extract possible inclusion bodies (supernatant 3) and produce final protein pellet 4. Protein concentrations were determined using bicinchoninic acid (BCA) protein assay and analyzed by SDS-PAGE and Western blot as described in Fig. 1 (Lanes: MW, molecular weight markers; 1, MSP1a control; 2, UBQ-MSP1a; 3, BM95-MSP1a; 4, EF1a-MSP1a and 5, SUB-MSP1a). Recombinant proteins were quantified by densitometric analysis of Coomassie based Instant Blue-stained SDS-polyacrylamide gels and shown as percent of total proteins on each fraction (column graphs). (B) Percent total proteins represented by recombinant chimeras in supernatants Super 1 (0.1% Triton X-100), Super 2 (1% Triton X-100) and Super 3 (6 M guanidine hydrochloride).
constant current. Gels were stained with Coomassie based Instant Blue (No. ISB01L; Expedeon Protein Solutions) or transferred to a nitrocellulose membrane (Schleicher & Schuell, PROTRAN BA85, Dassel, Germany) for Western blot analysis. Proteins were transferred to a nitrocellulose membrane during 1 h at 12 V using a Minie-Genie Electroblotter semi-dry transfer unit (Idea Scientific, Corvallis, OR, USA). The membrane was blocked with 5% skim milk for 1 h at room temperature, washed three times in TBS and probed with rabbit antibodies against purified MSP1a [25]. The membrane was incubated for 1 h with sera diluted 1:500 in 3%
BSA in TBS and washed three times with TBS. The membrane was then incubated with a goat anti-rabbit horseradish peroxidase (HRP) conjugate (Sigma, St. Louis, MO, USA) diluted 1:1000 in TBS, washed three times with TBS and finally developed with TMB stabilized substrate for HRP (Promega, Madison, WI, USA) for 20 min. An experiment was conducted to characterize the subcellular localization of recombinant chimeric proteins by successive protein extractions with 0.1% Triton X-100, 1% Triton X-100 and 6 M guanidine hydrochloride (Fig. 2A). The expres- sion of recombinant proteins was quantified by densitometric
&KDSWHU�,9
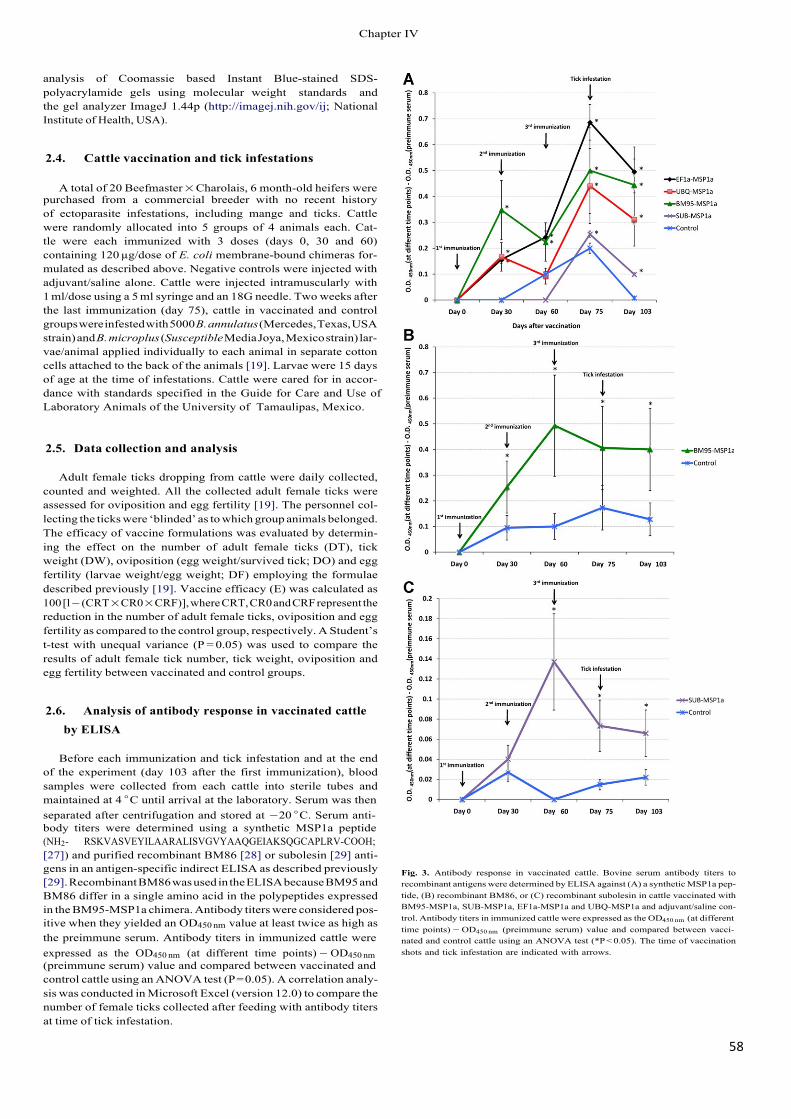
58!
analysis of Coomassie based Instant Blue-stained SDS- polyacrylamide gels using molecular weight standards and the gel analyzer ImageJ 1.44p (http://imagej.nih.gov/ij; National Institute of Health, USA).
2.4. Cattle vaccination and tick infestations
A total of 20 Beefmaster × Charolais, 6 month-old heifers were purchased from a commercial breeder with no recent history of ectoparasite infestations, including mange and ticks. Cattle were randomly allocated into 5 groups of 4 animals each. Cat- tle were each immunized with 3 doses (days 0, 30 and 60) containing 120 µg/dose of E. coli membrane-bound chimeras for- mulated as described above. Negative controls were injected with adjuvant/saline alone. Cattle were injected intramuscularly with 1 ml/dose using a 5 ml syringe and an 18G needle. Two weeks after the last immunization (day 75), cattle in vaccinated and control groups were infested with 5000 B. annulatus (Mercedes, Texas, USA strain) and B. microplus (Susceptible Media Joya, Mexico strain) lar- vae/animal applied individually to each animal in separate cotton cells attached to the back of the animals [19]. Larvae were 15 days of age at the time of infestations. Cattle were cared for in accor- dance with standards specified in the Guide for Care and Use of Laboratory Animals of the University of �Tamaulipas, Mexico.
2.5. Data collection and analysis
Adult female ticks dropping from cattle were daily collected, counted and weighted. All the collected adult female ticks were assessed for oviposition and egg fertility [19]. The personnel col- lecting the ticks were ‘blinded’ as to which group animals belonged. The efficacy of vaccine formulations was evaluated by determin- ing the effect on the number of adult female ticks (DT), tick weight (DW), oviposition (egg weight/survived tick; DO) and egg fertility (larvae weight/egg weight; DF) employing the formulae described previously [19]. Vaccine efficacy (E) was calculated as 100 [l − (CRT × CR0 × CRF)], where CRT, CR0 and CRF represent the reduction in the number of adult female ticks, oviposition and egg fertility as compared to the control group, respectively. A Student’s t-test with unequal variance (P = 0.05) was used to compare the results of adult female tick number, tick weight, oviposition and egg fertility between vaccinated and control groups.
2.6. Analysis of antibody response in vaccinated cattle by ELISA
Before each immunization and tick infestation and at the end of the experiment (day 103 after the first immunization), blood samples were collected from each cattle into sterile tubes and maintained at 4 ◦C until arrival at the laboratory. Serum was then separated after centrifugation and stored at −20 ◦C. Serum anti- body titers were determined using a synthetic MSP1a peptide (NH2- RSKVASVEYILAARALISVGVYAAQGEIAKSQGCAPLRV-COOH; [27]) and purified recombinant BM86 [28] or subolesin [29] anti- gens in an antigen-specific indirect ELISA as described previously [29]. Recombinant BM86 was used in the ELISA because BM95 and BM86 differ in a single amino acid in the polypeptides expressed in the BM95-MSP1a chimera. Antibody titers were considered pos- itive when they yielded an OD450 nm value at least twice as high as the preimmune serum. Antibody titers in immunized cattle were expressed as the OD450 nm (at different time points) − OD450 nm(preimmune serum) value and compared between vaccinated and control cattle using an ANOVA test (P = 0.05). A correlation analy- sis was conducted in Microsoft Excel (version 12.0) to compare the number of female ticks collected after feeding with antibody titers at time of tick infestation.
Fig. 3. Antibody response in vaccinated cattle. Bovine serum antibody titers to recombinant antigens were determined by ELISA against (A) a synthetic MSP1a pep- tide, (B) recombinant BM86, or (C) recombinant subolesin in cattle vaccinated with BM95-MSP1a, SUB-MSP1a, EF1a-MSP1a and UBQ-MSP1a and adjuvant/saline con- trol. Antibody titers in immunized cattle were expressed as the OD450 nm (at different time points) − OD450 nm (preimmune serum) value and compared between vacci- nated and control cattle using an ANOVA test (*P < 0.05). The time of vaccination shots and tick infestation are indicated with arrows.
&KDSWHU�,9
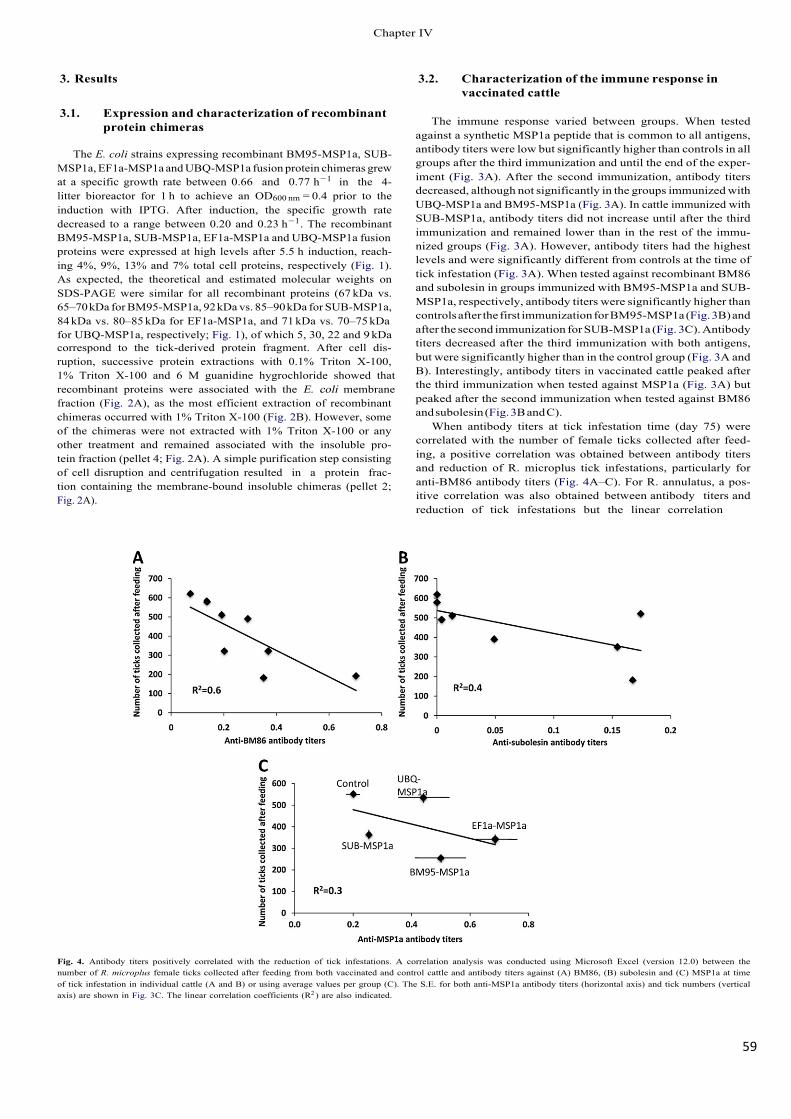
59!
3. Results
3.1. Expression and characterization of recombinant protein chimeras
The E. coli strains expressing recombinant BM95-MSP1a, SUB- MSP1a, EF1a-MSP1a and UBQ-MSP1a fusion protein chimeras grew at a specific growth rate between 0.66 and 0.77 h−1 in the 4- litter bioreactor for 1 h to achieve an OD600 nm = 0.4 prior to the induction with IPTG. After induction, the specific growth rate decreased to a range between 0.20 and 0.23 h−1. The recombinant BM95-MSP1a, SUB-MSP1a, EF1a-MSP1a and UBQ-MSP1a fusion proteins were expressed at high levels after 5.5 h induction, reach- ing 4%, 9%, 13% and 7% total cell proteins, respectively (Fig. 1). As expected, the theoretical and estimated molecular weights on SDS-PAGE were similar for all recombinant proteins (67 kDa vs. 65–70 kDa for BM95-MSP1a, 92 kDa vs. 85–90 kDa for SUB-MSP1a, 84 kDa vs. 80–85 kDa for EF1a-MSP1a, and 71 kDa vs. 70–75 kDa for UBQ-MSP1a, respectively; Fig. 1), of which 5, 30, 22 and 9 kDa correspond to the tick-derived protein fragment. After cell dis- ruption, successive protein extractions with 0.1% Triton X-100, 1% Triton X-100 and 6 M guanidine hygrochloride showed that recombinant proteins were associated with the E. coli membrane fraction (Fig. 2A), as the most efficient extraction of recombinant chimeras occurred with 1% Triton X-100 (Fig. 2B). However, some of the chimeras were not extracted with 1% Triton X-100 or any other treatment and remained associated with the insoluble pro- tein fraction (pellet 4; Fig. 2A). A simple purification step consisting of cell disruption and centrifugation resulted in a protein frac- tion containing the membrane-bound insoluble chimeras (pellet 2; Fig. 2A).
3.2. Characterization of the immune response in vaccinated cattle
The immune response varied between groups. When tested against a synthetic MSP1a peptide that is common to all antigens, antibody titers were low but significantly higher than controls in all groups after the third immunization and until the end of the exper- iment (Fig. 3A). After the second immunization, antibody titers decreased, although not significantly in the groups immunized with UBQ-MSP1a and BM95-MSP1a (Fig. 3A). In cattle immunized with SUB-MSP1a, antibody titers did not increase until after the third immunization and remained lower than in the rest of the immu- nized groups (Fig. 3A). However, antibody titers had the highest levels and were significantly different from controls at the time of tick infestation (Fig. 3A). When tested against recombinant BM86 and subolesin in groups immunized with BM95-MSP1a and SUB- MSP1a, respectively, antibody titers were significantly higher than controls after the first immunization for BM95-MSP1a (Fig. 3B) and after the second immunization for SUB-MSP1a (Fig. 3C). Antibody titers decreased after the third immunization with both antigens, but were significantly higher than in the control group (Fig. 3A and B). Interestingly, antibody titers in vaccinated cattle peaked after the third immunization when tested against MSP1a (Fig. 3A) but peaked after the second immunization when tested against BM86 and subolesin (Fig. 3B and C).
When antibody titers at tick infestation time (day 75) were correlated with the number of female ticks collected after feed- ing, a positive correlation was obtained between antibody titers and reduction of R. microplus tick infestations, particularly for anti-BM86 antibody titers (Fig. 4A–C). For R. annulatus, a pos- itive correlation was also obtained between antibody titers and reduction of tick infestations but the linear correlation
Fig. 4. Antibody titers positively correlated with the reduction of tick infestations. A correlation analysis was conducted using Microsoft Excel (version 12.0) between the number of R. microplus female ticks collected after feeding from both vaccinated and control cattle and antibody titers against (A) BM86, (B) subolesin and (C) MSP1a at time of tick infestation in individual cattle (A and B) or using average values per group (C). The S.E. for both anti-MSP1a antibody titers (horizontal axis) and tick numbers (vertical axis) are shown in Fig. 3C. The linear correlation coefficients (R2 ) are also indicated.
&KDSWHU�,9
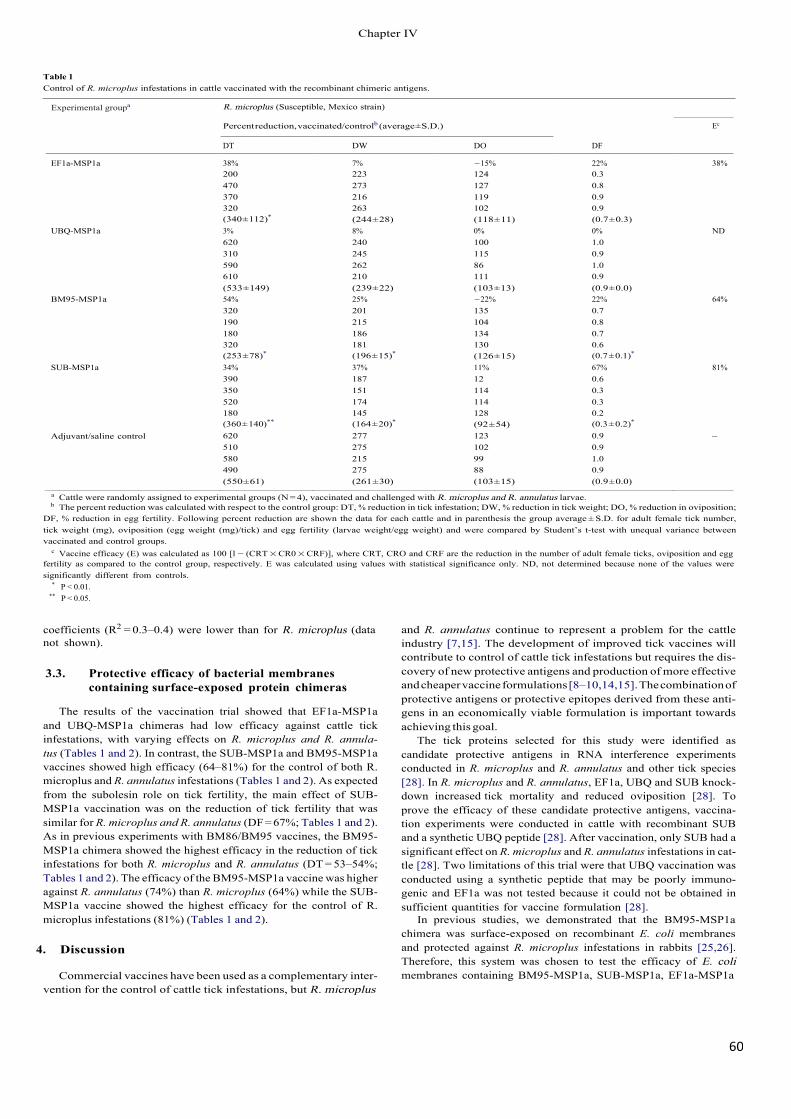
60!
Table 1 Control of R. microplus infestations in cattle vaccinated with the recombinant chimeric antigens.
Experimental groupa R. microplus (Susceptible, Mexico strain)
Percent reduction, vaccinated/controlb (average ± S.D.) Ec
DT DW DO DF
EF1a-MSP1a 38% 7% −15% 22% 38% 200 223 124 0.3 470 273 127 0.8 370 216 119 0.9 320 263 102 0.9
UBQ-MSP1a (340 ± 112)*
3% (244 ± 28) 8%
(118 ± 11) 0%
(0.7 ± 0.3) 0% ND
620 240 100 1.0 310 245 115 0.9 590 262 86 1.0 610 210 111 0.9
BM95-MSP1a (533 ± 149) 54%
(239 ± 22) 25%
(103 ± 13) −22%
(0.9 ± 0.0) 22% 64%
320 201 135 0.7 190 215 104 0.8 180 186 134 0.7 320 181 130 0.6
SUB-MSP1a (253 ± 78)*
34% (196 ± 15)*
37% (126 ± 15) 11%
(0.7 ± 0.1)*
67% 81% 390 187 12 0.6 350 151 114 0.3 520 174 114 0.3 180 145 128 0.2
Adjuvant/saline control (360 ± 140)**
620 (164 ± 20)*
277 (92 ± 54) 123
(0.3 ± 0.2)*
0.9 – 510 275 102 0.9 580 215 99 1.0 490 275 88 0.9 (550 ± 61) (261 ± 30) (103 ± 15) (0.9 ± 0.0)
a Cattle were randomly assigned to experimental groups (N = 4), vaccinated and challenged with R. microplus and R. annulatus larvae. b The percent reduction was calculated with respect to the control group: DT, % reduction in tick infestation; DW, % reduction in tick weight; DO, % reduction in oviposition;
DF, % reduction in egg fertility. Following percent reduction are shown the data for each cattle and in parenthesis the group average ± S.D. for adult female tick number, tick weight (mg), oviposition (egg weight (mg)/tick) and egg fertility (larvae weight/egg weight) and were compared by Student’s t-test with unequal variance between vaccinated and control groups.
c Vaccine efficacy (E) was calculated as 100 [l − (CRT × CR0 × CRF)], where CRT, CRO and CRF are the reduction in the number of adult female ticks, oviposition and egg fertility as compared to the control group, respectively. E was calculated using values with statistical significance only. ND, not determined because none of the values were significantly different from controls.
* P < 0.01. ** P < 0.05.
coefficients (R2 = 0.3–0.4) were lower than for R. microplus (data not shown).
3.3. Protective efficacy of bacterial membranes containing surface-exposed protein chimeras
The results of the vaccination trial showed that EF1a-MSP1a and UBQ-MSP1a chimeras had low efficacy against cattle tick infestations, with varying effects on R. microplus and R. annula- tus (Tables 1 and 2). In contrast, the SUB-MSP1a and BM95-MSP1a vaccines showed high efficacy (64–81%) for the control of both R. microplus and R. annulatus infestations (Tables 1 and 2). As expected from the subolesin role on tick fertility, the main effect of SUB- MSP1a vaccination was on the reduction of tick fertility that was similar for R. microplus and R. annulatus (DF = 67%; Tables 1 and 2). As in previous experiments with BM86/BM95 vaccines, the BM95- MSP1a chimera showed the highest efficacy in the reduction of tick infestations for both R. microplus and R. annulatus (DT = 53–54%; Tables 1 and 2). The efficacy of the BM95-MSP1a vaccine was higher against R. annulatus (74%) than R. microplus (64%) while the SUB- MSP1a vaccine showed the highest efficacy for the control of R. microplus infestations (81%) (Tables 1 and 2).
4. Discussion
Commercial vaccines have been used as a complementary inter- vention for the control of cattle tick infestations, but R. microplus
and R. annulatus continue to represent a problem for the cattle industry [7,15]. The development of improved tick vaccines will contribute to control of cattle tick infestations but requires the dis- covery of new protective antigens and production of more effective and cheaper vaccine formulations [8–10,14,15]. The combination of protective antigens or protective epitopes derived from these anti- gens in an economically viable formulation is important towards achieving this goal.
The tick proteins selected for this study were identified as candidate protective antigens in RNA interference experiments conducted in R. microplus and R. annulatus and other tick species [28]. In R. microplus and R. annulatus, EF1a, UBQ and SUB knock- down increased tick mortality and reduced oviposition [28]. To prove the efficacy of these candidate protective antigens, vaccina- tion experiments were conducted in cattle with recombinant SUB and a synthetic UBQ peptide [28]. After vaccination, only SUB had a significant effect on R. microplus and R. annulatus infestations in cat- tle [28]. Two limitations of this trial were that UBQ vaccination was conducted using a synthetic peptide that may be poorly immuno- genic and EF1a was not tested because it could not be obtained in sufficient quantities for vaccine formulation [28].
In previous studies, we demonstrated that the BM95-MSP1a chimera was surface-exposed on recombinant E. coli membranes and protected against R. microplus infestations in rabbits [25,26]. Therefore, this system was chosen to test the efficacy of E. coli membranes containing BM95-MSP1a, SUB-MSP1a, EF1a-MSP1a
&KDSWHU�,9
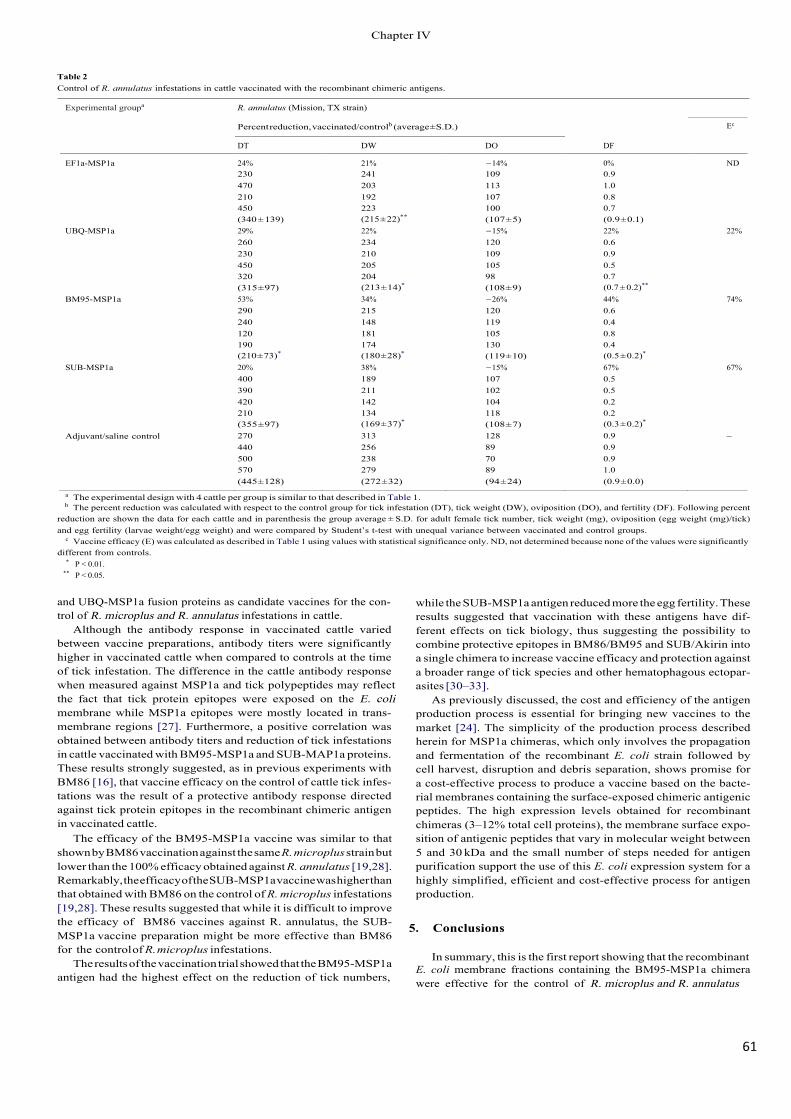
61!
Table 2 Control of R. annulatus infestations in cattle vaccinated with the recombinant chimeric antigens.
Experimental groupa R. annulatus (Mission, TX strain)
Percent reduction, vaccinated/controlb (average ± S.D.) Ec
DT DW DO DF
EF1a-MSP1a 24% 21% −14% 0% ND 230 241 109 0.9 470 203 113 1.0 210 192 107 0.8 450 223 100 0.7
UBQ-MSP1a (340 ± 139) 29%
(215 ± 22)**
22% (107 ± 5) −15%
(0.9 ± 0.1) 22% 22%
260 234 120 0.6 230 210 109 0.9 450 205 105 0.5 320 204 98 0.7
BM95-MSP1a (315 ± 97) 53%
(213 ± 14)*
34% (108 ± 9) −26%
(0.7 ± 0.2)**
44% 74% 290 215 120 0.6 240 148 119 0.4 120 181 105 0.8 190 174 130 0.4
SUB-MSP1a (210 ± 73)*
20% (180 ± 28)*
38% (119 ± 10) −15%
(0.5 ± 0.2)*
67% 67% 400 189 107 0.5 390 211 102 0.5 420 142 104 0.2 210 134 118 0.2
Adjuvant/saline control (355 ± 97) 270
(169 ± 37)*
313 (108 ± 7) 128
(0.3 ± 0.2)*
0.9 – 440 256 89 0.9 500 238 70 0.9 570 279 89 1.0 (445 ± 128) (272 ± 32) (94 ± 24) (0.9 ± 0.0)
a The experimental design with 4 cattle per group is similar to that described in Table 1. b The percent reduction was calculated with respect to the control group for tick infestation (DT), tick weight (DW), oviposition (DO), and fertility (DF). Following percent
reduction are shown the data for each cattle and in parenthesis the group average ± S.D. for adult female tick number, tick weight (mg), oviposition (egg weight (mg)/tick) and egg fertility (larvae weight/egg weight) and were compared by Student’s t-test with unequal variance between vaccinated and control groups.
c Vaccine efficacy (E) was calculated as described in Table 1 using values with statistical significance only. ND, not determined because none of the values were significantly different from controls.
* P < 0.01. ** P < 0.05.
and UBQ-MSP1a fusion proteins as candidate vaccines for the con- trol of R. microplus and R. annulatus infestations in cattle.
Although the antibody response in vaccinated cattle varied between vaccine preparations, antibody titers were significantly higher in vaccinated cattle when compared to controls at the time of tick infestation. The difference in the cattle antibody response when measured against MSP1a and tick polypeptides may reflect the fact that tick protein epitopes were exposed on the E. coli membrane while MSP1a epitopes were mostly located in trans- membrane regions [27]. Furthermore, a positive correlation was obtained between antibody titers and reduction of tick infestations in cattle vaccinated with BM95-MSP1a and SUB-MAP1a proteins. These results strongly suggested, as in previous experiments with BM86 [16], that vaccine efficacy on the control of cattle tick infes- tations was the result of a protective antibody response directed against tick protein epitopes in the recombinant chimeric antigen in vaccinated cattle.
The efficacy of the BM95-MSP1a vaccine was similar to that shown by BM86 vaccination against the same R. microplus strain but lower than the 100% efficacy obtained against R. annulatus [19,28]. Remarkably, the efficacy of the SUB-MSP1a vaccine was higher than that obtained with BM86 on the control of R. microplus infestations [19,28]. These results suggested that while it is difficult to improve the efficacy of BM86 vaccines against R. annulatus, the SUB-MSP1a vaccine preparation might be more effective than BM86 for the control of R. microplus infestations.
The results of the vaccination trial showed that the BM95-MSP1a antigen had the highest effect on the reduction of tick numbers,
while the SUB-MSP1a antigen reduced more the egg fertility. These results suggested that vaccination with these antigens have dif- ferent effects on tick biology, thus suggesting the possibility to combine protective epitopes in BM86/BM95 and SUB/Akirin into a single chimera to increase vaccine efficacy and protection against a broader range of tick species and other hematophagous ectopar- asites [30–33].
As previously discussed, the cost and efficiency of the antigen production process is essential for bringing new vaccines to the market [24]. The simplicity of the production process described herein for MSP1a chimeras, which only involves the propagation and fermentation of the recombinant E. coli strain followed by cell harvest, disruption and debris separation, shows promise for a cost-effective process to produce a vaccine based on the bacte- rial membranes containing the surface-exposed chimeric antigenic peptides. The high expression levels obtained for recombinant chimeras (3–12% total cell proteins), the membrane surface expo- sition of antigenic peptides that vary in molecular weight between 5 and 30 kDa and the small number of steps needed for antigen purification support the use of this E. coli expression system for a highly simplified, efficient and cost-effective process for antigen production.
5. Conclusions
In summary, this is the first report showing that the recombinantE. coli membrane fractions containing the BM95-MSP1a chimera were effective for the control of R. microplus and R. annulatus
&KDSWHU�,9

62!
infestations in cattle. The results of this study demonstrated that the SUB-MSP1a chimera was also protective against cat- tle tick infestations. Additionally, the E. coli expression system allowed the expression of different tick polypeptides fused to MSP1a, suggesting the possibility of developing tick vaccines based on recombinant E. coli membrane fractions containing chimera composed of a combination of surface-exposed protective anti- gens/epitopes. These new vaccine formulations would have the advantage of being cost-effective by using a simple and cheap pro- cess for the production of antigens for the control of cattle tick infestations [24].
Acknowledgments
This research was supported by FOMIX–TAMPS-2008-C17- 107392, the Spanish Ministerio de Ciencia e Innovación (MICINN) project BFU2008-01244/BMC, the Consejería de Educación y Cien- cia, JCCM, Spain (project PEII09-0118-8907), the Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (INIA), Spain (project FAU2008-00014-00-00), and Probiovet S.L., Spain. M. Vil- lar and M. Canales were funded by JAE-DOC program (CSIC-FSE) and Junta de Comunidades de Castilla-La Mancha (Program FSE 2007- 2013), Spain, respectively. J.A. Moreno-Cid is a recipient of a JCCM fellowship.
References
[1] Barker SC, Murrell A. Systematics and evolution of ticks with a list of valid genus and species names. Parasitology 2004;129:S15–36.
[2] Estrada-Pen a A, Bouattour A, Camicas JL, Guglielmone A, Horak I, Jongejan F, et al. The known distribution and ecological preferences of the tick sub- genus Boophilus (Acari: Ixodidae) in Africa and Latin America. Exp Appl Acarol 2006;38:219–35.
[3] Olwoch JM, Van Jaarsveld AS, Scholtz CH, Horak IG. Climate change and the genus Rhipicephalus (Acari: Ixodidae) in Africa. Onderstepoort J Vet Res 2007;74:45–72.
[4] Perez de Leon AA, Strickman DA, Knowles DP, Fish D, Thacker E, de la Fuente J, et al. For the group for emerging babesioses and one health research and devel- opment in the US. One health approach to identify research needs in bovine and human babesioses: workshop report. Parasite Vector 2010;3:36.
[5] Madder M, Thys E, Achi L, Touré A, De Deken R. Rhipicephalus (Boophilus) microplus: a most successful invasive tick species in West-Africa. Exp Appl Acarol 2011;53:139–45.
[6] Peter RJ, Van den Bossche P, Penzhorn BL, Sharp B. Tick, fly, and mosquito control-lessons from the past, solutions for the future. Vet Parasitol 2005;132:205–15.
[7] Graf JF, Gogolewski R, Leach-Bing N, Sabatini GA, Molento MB, Bordin EL, et al. Tick control: an industry point of view. Parasitology 2004;129:S427–42.
[8] de la Fuente J, Kocan KM. Strategies for development of vaccines for control of ixodid tick species. Parasite Immunol 2006;28:275–83.
[9] Sonenshine DE, Kocan KM, de la Fuente J. Tick control: further thoughts on a research agenda. Trends Parasitol 2006;22:550–1.
[10] Willadsen P. Tick control: thoughts on a research agenda. Vet Parasitol 2006;138:161–8.
[11] Willadsen P, Riding GA, McKenna RV, Kemp DH, Tellam RL, Nielsen JN, et al. Immunological control of a parasitic arthropod: identification of a protective antigen from Boophilus microplus. J Immunol 1989;143:1346–51.
[12] Rand KN, Moore T, Sriskantha A, Spring K, Tellam R, Willadsen P, et al. Cloning and expression of a protective antigen from the cattle tick Boophilus microplus. Proc Natl Acad Sci USA 1989;86:9657–61.
[13] Rodríguez M, Rubiera R, Montesinos R, Cremata JA, Falcón V, Sanchez G, et al. High level expression of the B. microplus Bm86 antigen in the yeast P. pastoris forming highly inmunogenic particles for cattle. J Biotechnol 1994;33:135–46.
[14] de la Fuente J, Kocan KM. Advances in the identification and characterization of protective antigens for development of recombinant vaccines against tick infestations. Expert Rev Vaccines 2003;2:583–93.
[15] de la Fuente J, Almazán C, Canales M, Pérez de la Lastra JM, Kocan KM, Willadsen P. A ten-year review of commercial vaccine performance for control of tick infestations on cattle. Anim Health Res Rev 2007;8:23–8.
[16] de la Fuente J, Rodríguez M, Redondo M, Montero C, García-García JC, Méndez L, et al. Field studies and cost-effectiveness analysis of vaccination with GavacTM�
against the cattle tick Boophilus microplus. Vaccine 1998;16:366–73. [17] Rodríguez Valle M, Méndez L, Valdez M, Redondo M, Espinosa CM, Vargas M,
et al. Integrated control of Boophilus microplus ticks in Cuba based on vaccina- tion with the anti-tick vaccine Gavac. Exp Appl Acarol 2004;34:375–82.
[18] García-García JC, Montero C, Redondo M, Vargas M, Canales M, Boué O, et al. Control of ticks resistant to immunization with Bm86 in cattle vaccinated with the recombinant antigen Bm95 isolated from the cattle tick, Boophilus microplus. Vaccine 2000;18:2275–87.
[19] Canales M, Almazán C, Naranjo V, Jongejan F, de la Fuente J. Vaccination with recombinant Boophilus annulatus Bm86 ortholog protein, Ba86, protects cattle against B. annulatus and B. microplus infestations. BMC Biotechnol 2009;9:29.
[20] de la Fuente J, Rodríguez M, Garcia-Garcia JC. Immunological control of ticks through vaccination with Boophilus microplus gut antigens. Ann N Y Acad Sci 2000;916:617–21.
[21] de Vos S, Zeinstra L, Taoufik O, Willadsen P, Jongejan F. Evidence for the utility of the Bm86 antigen from Boophilus microplus in vaccination against other tick species. Exp Appl Acarol 2001;25:245–61.
[22] Canales M, Enriquez A, Ramos E, Cabrera D, Dandie H, Soto A, et al. Large-scale production in Pichia pastoris of the recombinant vaccine GavacTM against cattle ticks. Vaccine 1997;15:414–22.
[23] Canales M, Pérez de la Lastra JM, Naranjo V, Nijhof AM, Hope M, Jongejan F, et al. Expression of recombinant Rhipicephalus (Boophilus) microplus, R. annu- latus and R. decoloratus Bm86 orthologs as secreted proteins in Pichia pastoris. BMC Biotechnol 2008;8:14.
[24] Canales M, Moreno-Cid JA, Almazán C, Villar M, de la Fuente J. Bioprocess design and economics of recombinant BM86/BM95 antigen production for anti-tick vaccines. Biochem Eng J 2010;52:79–90.
[25] Canales M, Almazán C, Pérez de la Lastra JM, de la Fuente J. Anaplasma marginale major surface protein 1a directs cell surface display of tick BM95 immunogenic peptides on Escherichia coli. J Biotechnol 2008;135:326–32.
[26] Canales M, Labruna MB, Soares JF, Prudencio CR, de la Fuente J. Protective efficacy of bacterial membranes containing surface-exposed BM95 antigenic peptides for the control of cattle tick infestations. Vaccine 2009;27:7244–8.
[27] Garcia-Garcia JC, de la Fuente J, Kocan KM, Blouin EF, Halbur T, Onet VC, et al. Mapping of B-cell epitopes in the N-terminal repeated peptides of Anaplasma marginale major surface protein 1a and characterization of the humoral immune response of cattle immunized with recombinant and whole organism antigens. Vet Immunol Immunopathol 2004;98:137–51.
[28] Almazán C, Lagunes R, Villar M, Canales M, Rosario-Cruz R, Jongejan F, et al. Identification and characterization of Rhipicephalus (Boophilus) microplus can- didate protective antigens for the control of cattle tick infestations. Parasitol Res 2010;106:471–9.
[29] Merino O, Almazán C, Canales M, Villar M, Moreno-Cid JA, Estrada-Pen a A, et al. Control of Rhipicephalus (Boophilus) microplus infestations by the combi- nation of subolesin vaccination and tick autocidal control after subolesin gene knockdown in ticks fed on cattle. Vaccine 2011;29:2248–54.
[30] Prudencio CR, Pérez de la Lastra JM, Canales M, Villar M, de la Fuente J. Map- ping protective epitopes in the tick and mosquito subolesin ortholog proteins. Vaccine 2010;28:5398–406.
[31] Canales M, Naranjo V, Almazán C, Molina R, Tsuruta SA, Szabó MPJ, et al. Con- servation and immunogenicity of the mosquito ortholog of the tick protective antigen, subolesin. Parasitol Res 2009;105:97–111.
[32] Moreno-Cid JA, Jiménez M, Cornelie S, Molina R, Alarcón P, Lacroix M-N, et al. Characterization of Aedes albopictus akirin for the control of mosquito and sand fly infestations. Vaccine 2011;29:77–82.
[33] Harrington D, Canales M, de la Fuente J, de Luna C, Robinson K, Guy J, et al. Immunisation with recombinant proteins subolesin and Bm86 for the control of Dermanyssus gallinae in poultry. Vaccine 2009;27:4056–63.
&KDSWHU�,9


CHAPTER(V.(DISCUSSION(
de! la!Fuente! J.,!Moreno3Cid! J.!A.,!Villar!M.,!Galindo!R.!C.,!Almazán!C.,!Kocan!K.!M.,!Merino! O.,! Pérez! de! la! Lastra! J.! M.,! Estrada3Peña! A.! and! Blouin! E.! F.! (2013)!Subolesin/Akirin. Vaccines. for. the. Control. of. Arthropod.Vectors. and.Vectorborne.Pathogens..Transboundary!and!Emerging!Diseases..“In#Press”!


SUBOLESIN/AKIRIN' VACCINES' FOR' THE' CONTROL' OF'
ARTHROPOD'VECTORS'AND'VECTOR9BORNE'PATHOGENS'

65!
Subolesin/Akirin vaccines for the control of arthropod vectors and
vector-borne pathogens
José de la Fuente1,2,§, Juan A. Moreno-Cid1, Margarita Villar1, Ruth C. Galindo1, Consuelo Almazán3, Katherine M. Kocan2, Octavio Merino1, José M. Pérez de la Lastra1, Agustín Estrada-Peña4, Edmour F. Blouin2
1Instituto de Investigación en Recursos Cinegéticos IREC-CSIC-UCLM-JCCM, Ronda de Toledo s/n, 13005 Ciudad Real, Spain. 2Department of Veterinary Pathobiology, Center for Veterinary Health Sciences, Oklahoma State University, Stillwater, OK 74078, USA. 3Facultad de Medicina Veterinaria y Zootecnia, Universidad Autónoma de Tamaulipas, Km. 5 carretera Victoria-Mante, CP 87000 Ciudad Victoria, Tamaulipas, Mexico. 4Department of Parasitology. Veterinary Faculty. Miguel Servet 177. 50013-Zaragoza, Spain.
§Correspondence: José de la Fuente, Instituto de Investigación en Recursos Cinegéticos IREC-CSIC- UCLM-JCCM, Ronda de Toledo s/n, 13005 Ciudad Real, Spain. Email: [email protected]
ABSTRACT
Diseases transmitted by arthropod vectors such as mosquitoes, ticks and sand flies greatly impact human and animal health and therefore their control is important for the eradication of vector-borne diseases (VBD). Vaccination is an environmentally friendly alternative for vector control that allows control of several VBD by targeting their common vector. Recent results have suggested that subolesin (SUB) and its ortholog in insects, akirin (AKR) are good candidate antigens for the control of arthropod vector infestations and pathogen infection. SUB was discovered as a tick protective antigen in Ixodes scapularis. Vaccination trials with recombinant SUB/AKR demonstrated effective control of arthropod vector infestations in various hard and soft tick species, mosquitoes, sand flies, poultry red mites and sea lice by reducing their numbers, weight, oviposition, fertility and/or molting. SUB/AKR vaccination also reduced tick infection with tick-borne pathogens, Anaplasma phagocytophilum, A. marginale, Babesia bigemina and Borrelia burgdorferi. The effect of vaccination on different hosts, vector species, developmental stages and vector-borne pathogen infections demonstrated the feasibility of SUB/AKR universal vaccines for the control of multiple vector infestations and for reduction of VBD.
Keywords: akirin, subolesin, tick, mosquito, fly, mite, lice
&KDSWHU�9

66!
1. Introduction
Diseases transmitted by arthropod vectors greatly impact human and animal health, accounting for over 20% of all emerging infectious diseases recorded between 1940 and 2004 (Jones et al., 2008). Although human vaccines for yellow fever, Japanese encephalitis and tick-borne encephalitis are available, on-going vaccination efforts are insufficient to prevent infection (Hayes, 2010; Andresson et al., 2010). For other vector-borne diseases (VBD), vaccines have not been developed or successfully implemented. Therefore, control of arthropod vectors is important for the eradication of VBD (de la Fuente and Kocan, 2003; Sperança and Capurro, 2007; Karunamoorthi, 2011; Coller et al., 2012). Vaccination is an environmentally friendly alternative for the control of vector infestations and pathogen infections that allows control of several VBD by targeting their common vector (de la Fuente et al., 2007; 2011). Recent results have suggested that subolesin (SUB) and its ortholog in insects, akirin (AKR) may be good candidate antigens for the control of arthropod vector infestations and VBD (de la Fuente et al., 2011). In this paper, we review results obtained with recombinant SUB/AKR vaccines for the control of arthropod vector infestations and vector-borne pathogen infections. These results suggest that SUB/AKR are good candidate antigens for the development of universal vaccines for the control of multiple vector infestations and reduction of VBD.
2. Identification of candidate vector protective antigens
The limiting step in the development of vector vaccines is the identification of protective antigens (de la Fuente and Kocan, 2003). Particularly relevant towards this goal is the identification of protective antigens such as SUB/AKR that are conserved across vector species, thus providing the opportunity to develop a universal vaccine for the control of multiple arthropod infestations and their associated pathogens (de la Fuente et al., 2011).
Traditional control methods for arthropod vector infestations are based on the use of chemical pesticides with associated drawbacks such as selection of arthropod-resistant strains and contamination of both the environment and animal products (de la Fuente and Kocan, 2003). Vaccines have several advantages including (a) no contamination of the environment and animal products, (b) avoiding selection of pesticide resistant arthropod vectors, (c) targeting a broad but selective range of vector species, and (d) reducing vector capacity for pathogen transmission. However, the experience with the only commercial vaccines available for the control of ectoparasite infestations, TickGard and Gavac, demonstrated that these vaccines are difficult to introduce into the market because of the absence of immediate effect on vector infestations and the application in combination with other control measures (de la Fuente and Kocan, 2003; 2006; Willadsen, 2004; de la Fuente et al., 2007). In 1994, Elvin and Kemp proposed the conditions necessary and sufficient for an effective vector protective antigen. These conditions are still valid, with the addition of an effect on pathogen transmission: (a) host antibodies should be able to gain access to the vector antigen, (b) sufficient antibodies must gain access to the target antigen, (c) the formation of antibody-antigen complex should disrupt the normal function of the vector protein, (d) the antigen should share conserved structural/sequence motifs among vector species to protect against multiple vector infestations, and (e) the vaccine should affect vector infestations and vector capacity for transmitted pathogens. SUB, initially named 4D8, was discovered as a tick protective antigen in a mouse model of Ixodes scapularis infestations using cDNA expression library immunization (ELI) combined with expressed sequence tags (EST) analysis (Almazán et al., 2003). This method demonstrated the possibilities of using high throughput screening technologies for the identification of candidate tick protective antigens. Recently, other approaches using RNA interference (RNAi) (de la Fuente et al., 2005;
&KDSWHU�9

67!
2010; Almazán et al., 2010; Kocan et al., 2011) and capillary feeding (Almazán et al., 2005a; Canales et al., 2009a; Rodriguez-Valle et al., 2012; Gonsioroski et al., 2012) have been proposed for the screening of tick protective antigens. Recent developments in both vectors and vector- borne pathogen genomics and the application of systems biology to study of vector-pathogen and vector-host interactions have advanced our understanding of the genetic factors and molecular pathways involved at the host-vector- pathogen interface (de la Fuente and Estrada- Peña, 2012; de la Fuente, 2012). These technologies are generating extensive information, but algorithms are needed to use these data for advancing knowledge on basic biological questions and the development of improved vaccines for the control of vectors and VBD. Holistic approaches using ‘omics’ technology are required to study interactions between components of biological systems and how these interactions give rise to the function and behavior of that system. However, reductionist approaches focusing on specific components of the system are also valuable to address vector and vector-borne pathogen genetics, physiology and other biological questions that would ultimately impact on the development of new vaccines and other control strategies (de la Fuente and Estrada-Peña, 2012; de la Fuente, 2012).
3. Subolesin/Akirin vaccines for the control ofarthropod vectors and vector-bornepathogens
SUB and AKR are ortholog proteins in ticks and insects that affect the expression of signal transduction and innate immune response genes, as well as positive and negative transcriptional regulators (Goto et al., 2008; Macqueen and Johnston, 2009; Galindo et al., 2009; Mangold et al., 2009; de la Fuente et al., 2011; Nowak et al., 2012). These intermediate proteins interact with NF-kB and other regulatory proteins, bind DNA or remodel chromatin to regulate gene expression (Goto et al., 2008; Galindo et al., 2009; de la Fuente et al., 2011; Nowak et al.,
2012). This broad function of SUB/AKR as transcription factors explains the profound effect of gene knockdown by RNAi on tick and insect physiology, as well as on development and on gene expression in ticks. SUB and AKR are functionally important for arthropod innate immunity to pathogens and, at least in ticks, for other molecular pathways, including those required for tissue development and function and for pathogen infection and multiplication (de la Fuente et al., 2011). Additionally, SUB/AKR share common structural and protective epitopes in different arthropod vector species (Canales et al., 2009a; Prudencio et al., 2010). These results strongly suggested the possibility of using SUB/AKR for development of vaccines for control of arthropod vector infestations and pathogen infections. SUB/AKR vaccines were designed using recombinant tick spp.-derived SUB and Ae. albopictus mosquito-derived AKR expressed in Escherichia coli and Pichia pastoris (Canales et al., 2009b; Almazán et al., 2010; 2012; Moreno- Cid et al., 2012) (Tables 1 and 2). SUB/AKR vaccines have been effective for the control of both arthropod infestations (Table 1) and pathogen infection (Table 2). In these experiments, vaccination with SUB/AKR provided control for hard (Ixodes spp., Rhipicephalus spp., Amblyomma americanum, Dermacentor variabilis) and soft (Ornithodoros spp.) ticks, mosquitoes (Aedes albopictus), sand flies (Phlebotomus perniciosus), poultry red mites (Dermanyssus gallinae) and sea lice (Caligus rogercresseyi) infestations and tick infections with Anaplasma phagocytophilum, A. marginale, Babesia bigemina and Borrelia burgdorferi (Tables 1 and 2). The efficacy of vaccination with SUB/AKR was demonstrated on different arthropod developmental stages and in various hosts, including domestic and wild mammals, birds and fishes (Fig. 1; Tables 1 and 2). These results demonstrated the efficacy of SUB/AKR vaccines in different parts of the world for the control of vectors and pathogens that affect human and animal health and suggested the possibility of vaccinating animal species that are hosts and/or affected by these vectors and pathogens. Nevertheless, additional
&KDSWHU�9
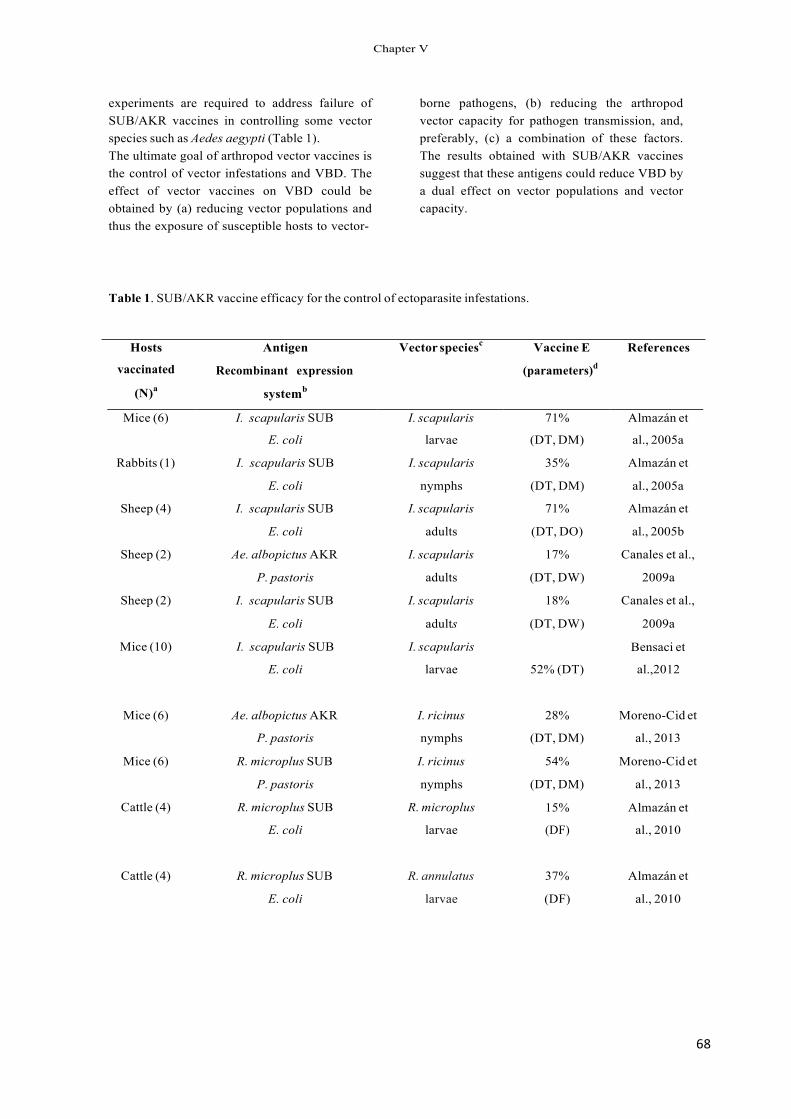
68!
experiments are required to address failure of SUB/AKR vaccines in controlling some vector species such as Aedes aegypti (Table 1). The ultimate goal of arthropod vector vaccines is the control of vector infestations and VBD. The effect of vector vaccines on VBD could be obtained by (a) reducing vector populations and thus the exposure of susceptible hosts to vector-
borne pathogens, (b) reducing the arthropod vector capacity for pathogen transmission, and, preferably, (c) a combination of these factors. The results obtained with SUB/AKR vaccines suggest that these antigens could reduce VBD by a dual effect on vector populations and vector capacity.
Table 1. SUB/AKR vaccine efficacy for the control of ectoparasite infestations.
Hosts
vaccinated
(N)a
Antigen
Recombinant expression
systemb
Vector speciesc Vaccine E
(parameters)d
References
Mice (6) I. scapularis SUB I. scapularis 71% Almazán et
E. coli larvae (DT, DM) al., 2005a
Rabbits (1) I. scapularis SUB
E. coli
I. scapularis
nymphs
35%
(DT, DM)
Almazán et
al., 2005a
Sheep (4) I. scapularis SUB I. scapularis 71% Almazán et
E. coli adults (DT, DO) al., 2005b
Sheep (2) Ae. albopictus AKR I. scapularis 17% Canales et al.,
P. pastoris adults (DT, DW) 2009a
Sheep (2) I. scapularis SUB I. scapularis 18% Canales et al.,
E. coli adults (DT, DW) 2009a
Mice (10) I. scapularis SUB
E. coli
I. scapularis
larvae 52% (DT)
Bensaci et
al.,2012
Mice (6) Ae. albopictus AKR I. ricinus 28% Moreno-Cid et
P. pastoris nymphs (DT, DM) al., 2013
Mice (6) R. microplus SUB I. ricinus 54% Moreno-Cid et
P. pastoris nymphs (DT, DM) al., 2013
Cattle (4) R. microplus SUB
E. coli
R. microplus
larvae
15%
(DF)
Almazán et
al., 2010
Cattle (4) R. microplus SUB R. annulatus 37% Almazán et
E. coli larvae (DF) al., 2010
&KDSWHU�9
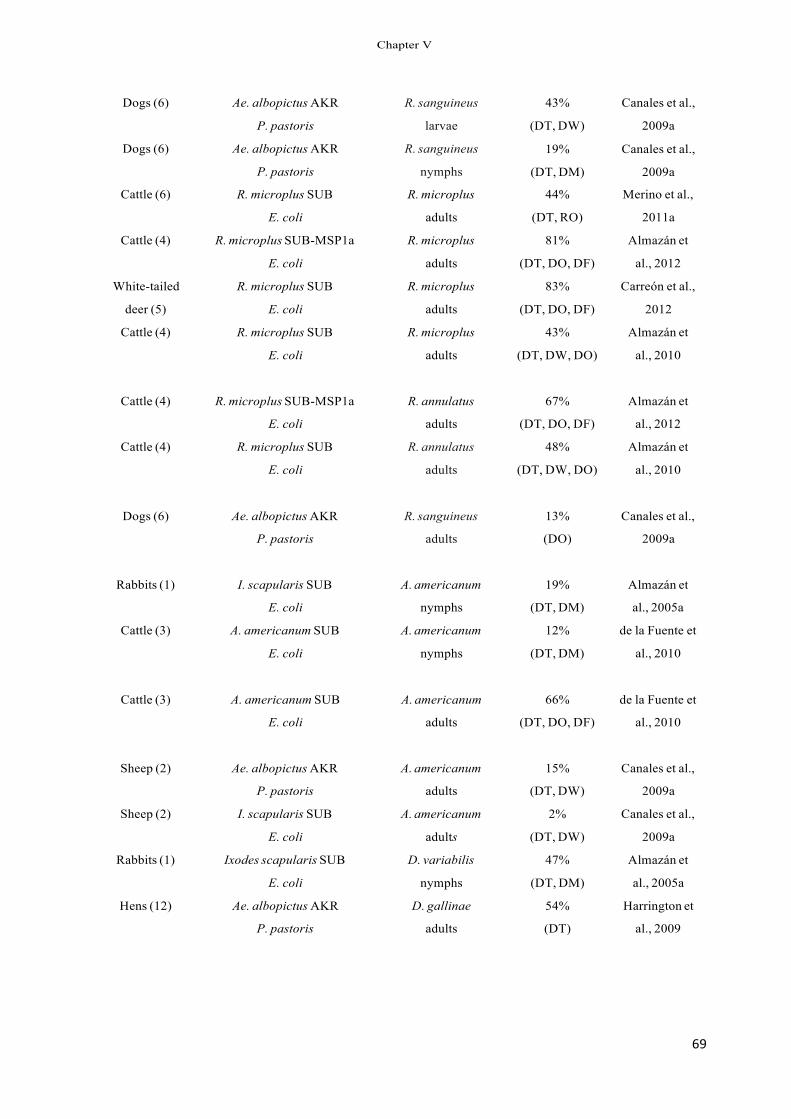
69!
Dogs (6)
Dogs (6)
Ae. albopictus AKR
P. pastoris
Ae. albopictus AKR
P. pastoris
R. sanguineus
larvae
R. sanguineus
nymphs
43%
(DT, DW)
19%
(DT, DM)
Canales et al.,
2009a
Canales et al.,
2009a
Cattle (6) R. microplus SUB R. microplus 44% Merino et al.,
E. coli adults (DT, RO) 2011a
Cattle (4) R. microplus SUB-MSP1a R. microplus 81% Almazán et
E. coli adults (DT, DO, DF) al., 2012
White-tailed R. microplus SUB R. microplus 83% Carreón et al.,
deer (5) E. coli adults (DT, DO, DF) 2012
Cattle (4) R. microplus SUB R. microplus 43% Almazán et
E. coli adults (DT, DW, DO) al., 2010
Cattle (4) R. microplus SUB-MSP1a R. annulatus 67% Almazán et
E. coli adults (DT, DO, DF) al., 2012
Cattle (4) R. microplus SUB R. annulatus 48% Almazán et
E. coli adults (DT, DW, DO) al., 2010
Dogs (6) Ae. albopictus AKR R. sanguineus 13% Canales et al.,
P. pastoris adults (DO) 2009a
Rabbits (1) I. scapularis SUB A. americanum 19% Almazán et
E. coli nymphs (DT, DM) al., 2005a
Cattle (3) A. americanum SUB A. americanum 12% de la Fuente et
E. coli nymphs (DT, DM) al., 2010
Cattle (3) A. americanum SUB
E. coli
A. americanum
adults
66%
(DT, DO, DF)
de la Fuente et
al., 2010
Sheep (2) Ae. albopictus AKR A. americanum 15% Canales et al.,
P. pastoris adults (DT, DW) 2009a
Sheep (2) I. scapularis SUB A. americanum 2% Canales et al.,
E. coli adults (DT, DW) 2009a
Rabbits (1) Ixodes scapularis SUB D. variabilis 47% Almazán et
Hens (12)
E. coli
Ae. albopictus AKR
nymphs
D. gallinae
(DT, DM)
54%
al., 2005a
Harrington et
P. pastoris adults (DT) al., 2009
&KDSWHU�9
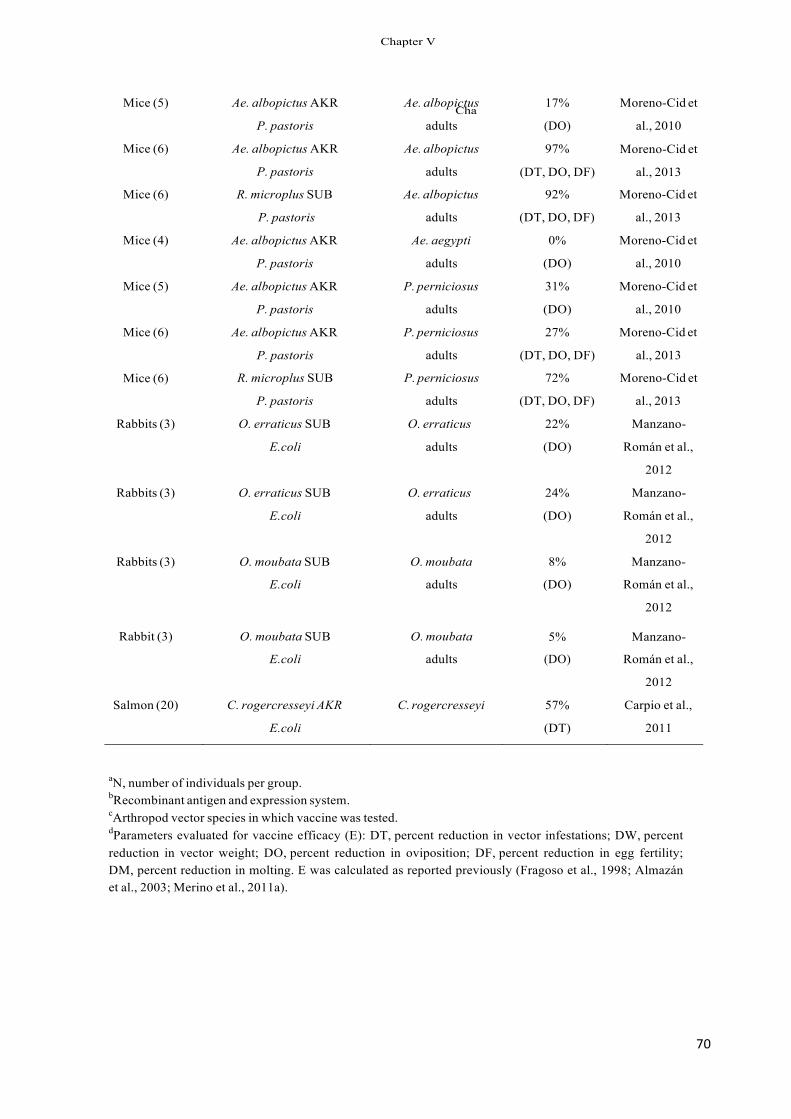
70!
Mice (5)
Mice (6)
Ae. albopictus AKR
P. pastoris
Ae. albopictus AKR
P. pastoris
Ae. albopictus
adults
Ae. albopictus
adults
17%
(DO)
97%
(DT, DO, DF)
Moreno-Cid et
al., 2010
Moreno-Cid et
al., 2013
Mice (6) R. microplus SUB Ae. albopictus 92% Moreno-Cid et
P. pastoris adults (DT, DO, DF) al., 2013
Mice (4) Ae. albopictus AKR Ae. aegypti 0% Moreno-Cid et
P. pastoris adults (DO) al., 2010
Mice (5) Ae. albopictus AKR P. perniciosus 31% Moreno-Cid et
P. pastoris adults (DO) al., 2010
Mice (6) Ae. albopictus AKR P. perniciosus 27% Moreno-Cid et
Mice (6)
P. pastoris
R. microplus SUB
adults
P. perniciosus
(DT, DO, DF)
72%
al., 2013
Moreno-Cid et
P. pastoris adults (DT, DO, DF) al., 2013
Rabbits (3) O. erraticus SUB O. erraticus 22% Manzano-
E.coli adults (DO) Román et al.,
2012
Rabbits (3) O. erraticus SUB O. erraticus 24% Manzano-
E.coli adults (DO) Román et al.,
Rabbits (3) O. moubata SUB O. moubata 8%
2012
Manzano-
E.coli adults (DO) Román et al.,
2012
Rabbit (3) O. moubata SUB
E.coli
O. moubata
adults
5%
(DO)
Manzano-
Román et al.,
2012
Salmon (20) C. rogercresseyi AKR C. rogercresseyi 57% Carpio et al.,
E.coli (DT) 2011
aN, number of individuals per group. bRecombinant antigen and expression system. cArthropod vector species in which vaccine was tested. dParameters evaluated for vaccine efficacy (E): DT, percent reduction in vector infestations; DW, percent reduction in vector weight; DO, percent reduction in oviposition; DF, percent reduction in egg fertility; DM, percent reduction in molting. E was calculated as reported previously (Fragoso et al., 1998; Almazán et al., 2003; Merino et al., 2011a).
&KD
&KDSWHU�9
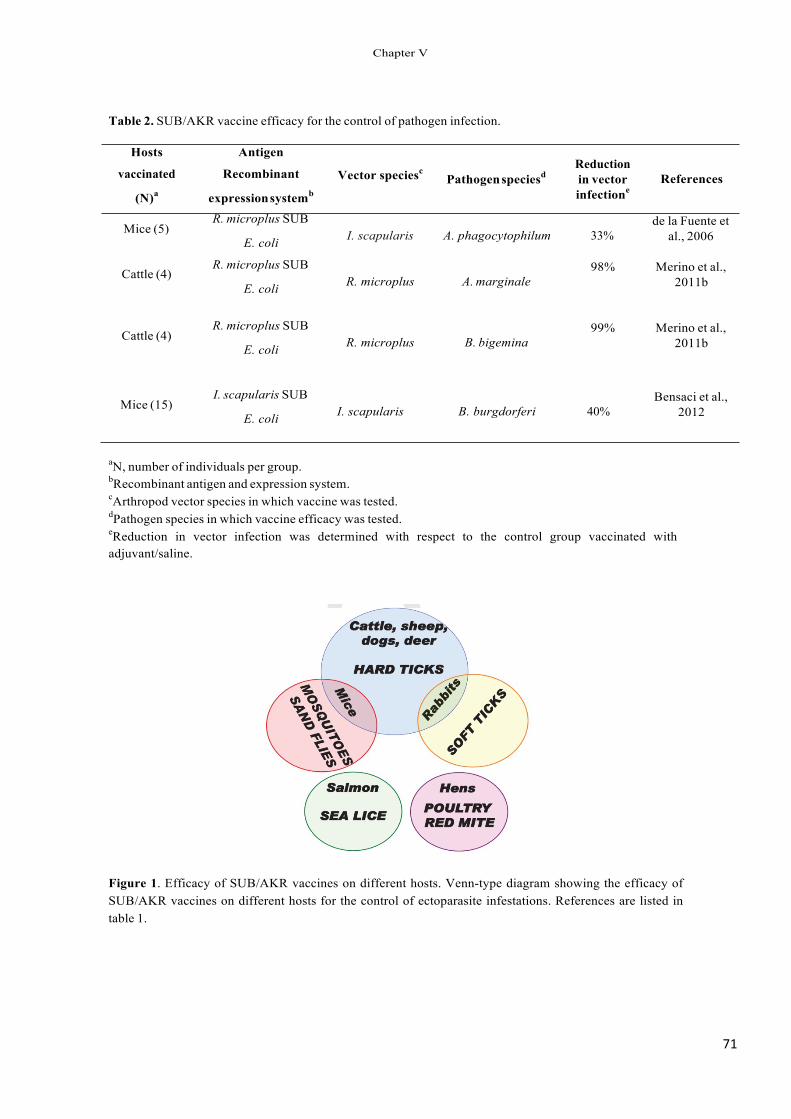
71!
Mice (5)
Cattle (4)
Cattle (4)
Mice (15)
Table 2. SUB/AKR vaccine efficacy for the control of pathogen infection.
Hosts
vaccinated
(N)a
Antigen
Recombinant
expression systemb
Vector speciesc Pathogen speciesd
Reduction in vector infectione
References
R. microplus SUB
E. coli I. scapularis A. phagocytophilum 33% de la Fuente et
al., 2006
R. microplus SUB
E. coli R. microplus A. marginale 98% Merino et al.,
2011b
R. microplus SUB
E. coli R. microplus B. bigemina 99% Merino et al.,
2011b
I. scapularis SUB
E. coli I. scapularis B. burgdorferi 40% Bensaci et al.,
2012
aN, number of individuals per group. bRecombinant antigen and expression system. cArthropod vector species in which vaccine was tested. dPathogen species in which vaccine efficacy was tested. eReduction in vector infection was determined with respect to the control group vaccinated with adjuvant/saline.
Figure 1. Efficacy of SUB/AKR vaccines on different hosts. Venn-type diagram showing the efficacy of SUB/AKR vaccines on different hosts for the control of ectoparasite infestations. References are listed in table 1.
&KDSWHU�9

72!
The exact mechanisms by which SUB/AKR vaccines affect vector infestations, fertility and pathogen infection are unknown. Blood-sucking arthropods feeding on immunized hosts ingest antibodies specific for SUB/AKR that could reduce their levels and biological activity and/or interact with conserved epitopes in other proteins resulting in reduced feeding, developmental and reproductive performance (de la Fuente et al., 2011; Moreno-Cid et al., 2010). The issue of SUB/AKR vaccine safety due to sequence conservation between arthropod vectors and vertebrate hosts has been discussed previously (Canales et al., 2009a; Moreno-Cid et al., 2010). The absence of physiological or pathological alterations in vaccinated hosts suggests that, as expected, the antibody response is directed against non-self epitopes thus reducing the possibility of detrimental effects to the vertebrate host.
4. Conclusions
Taken together, these results present considerable evidence toward use of SUB/AKR for the development of universal vaccines for the control of multiple arthropod vectors and VBD. Using the postulates established for effective vector protective antigens (adapted from Elvin and Kemp, 1994), vaccination trials with SUB/AKR have demonstrated that sufficient host antibodies gain access to the antigen to disrupt its biological function. Furthermore, these antigens share conserved structural/sequence motifs among vector species to protect against multiple vector infestations and pathogen infection. Future controlled pen and field trials should be directed to defining the efficacy of SUB/AKR vaccines alone and in combination with other vector- and pathogen- derived antigens on multiple arthropod vector species and, importantly, on pathogen infection and transmission by mosquitoes, sand flies and ticks for the control of VBD of global importance such as malaria, leishmania and Lyme disease. The effect of SUB/AKR vaccines on vector-borne virus infection and transmission
should also be addressed. Research on basic biological questions concerning SUB/AKR regulation and function and the effect of vaccination on vector biology and host-vector- pathogen interactions is essential to improve vaccine efficacy and design effective strategies for the control of vector populations and VBD using SUB/AKR vaccines.
Acknowledgments
This research was supported by grants FAU2008-00014-00-00 and PEII09-0118-8907 to JF and the Walter R. Sitlington Endowed Chair for Food Animal Research to KMK. M. Villar was funded by the JAE-DOC program (CSIC-FSE), Spain. O. Merino is an Early Stage Researcher supported by the POSTICK ITN (Postgraduate training network for capacity building to control ticks and tick-borne diseases) within the FP7-PEOPLE – ITN programme (EU Grant No. 38511).
References
Almazán, C., K. M. Kocan, D.K. Bergman, J.C. García- García, E.F. Blouin and J. de la Fuente, 2003: Identification of protective antigens for the control of Ixodes scapularis infestations using cDNA expression library immunization. Vaccine 21, 1492-1501.
Almazán, C., U. Blas-Machado, K.M. Kocan, J.H. Yoshioka, E.F. Blouin, A.J. Mangold, and J. de la Fuente, 2005a: Characterization of three Ixodes scapularis cDNAs protective against tick infestations. Vaccine 23, 4403-4416.
Almazán, C., K.M. Kocan, E.F. Blouin and J. de la Fuente, 2005b: Vaccination with recombinant tick antigens for the control of Ixodes scapularis adult infestations. Vaccine 23, 5294-5298.
Almazán, C., R. Lagunes, M. Villar, M. Canales, R. Rosario- Cruz, F. Jongejan and J. de la Fuente, 2010: Identification and characterization of Rhipicephalus (Boophilus) microplus candidate protective antigens for the control of cattle tick infestations. Parasitol. Res. 106, 471-479.
Almazán, C., O. Moreno-Cantú, J.A. Moreno-Cid, R.C. Galindo, M. Canales, M. Villar, and J. de la Fuente, 2012: Control of tick infestations in cattle vaccinated with bacterial membranes containing surface-exposed tick protective antigens. Vaccine 30, 265-272.
&KDSWHU�9

73!
Andersson, C.R., S. Vene, M. Insulander, L. Lindquist, A. Lundkvist, G. Günther, 2010: Vaccine failures after active immunisation against tick-borne encephalitis. Vaccine 28, 2827-2831.
Bensaci, M., D. Bhattacharya, R. Clark and L.T. Hu, 2012: Oral vaccination with vaccinia virus expressing the tick antigen subolesin inhibits tick feeding and transmission of Borrelia burgdorferi. Vaccine 30, 6040-6046.
Canales, M., V. Naranjo, C. Almazán, R. Molina, S.A. Tsuruta, M.P.J. Szabó, R. Manzano-Roman, J.M. Pérez de la Lastra, K.M. Kocan, M.I. Jiménez, J. Lucientes, M. Villar and J. de la Fuente, 2009a: Conservation and immunogenicity of the mosquito ortholog of the tick protective antigen, subolesin. Parasitol. Res. 105, 97-111.
Canales, M., C. Ballesteros, J.A. Moreno-Cid, A.M. Espinosa, M. Villar and J. de la Fuente, 2009b: Extractive bioconversion to produce the Aedes albopictus akirin in an aqueous two-phase system supporting Pichia pastoris growth and protein secretion. Biochem. Eng. J. 46, 105-114.
Carpio, Y, L. Basabe, J. Acosta, A. Rodríguez, A. Mendoza, A. Lisperger, E. Zamorano, M. González, M. Rivas, S. Contreras, D. Haussmann, J. Figueroa, V.N. Osorio, G. Asencio, J. Mancilla, G. Ritchie, C. Borroto and M.P. Estrada, 2011: Novel gene isolated from Caligus rogercresseyi: A promising target for vaccine development against sea lice. Vaccine 29, 2810-2820.
Carreón, D., J. M. Pérez de la Lastra, C. Almazán, M. Canales, F. Ruiz-Fons, M. Boadella, J.A. Moreno- Cid, M. Villar, C. Gortázar, M. Reglero, R. Villarreal, J. de la Fuente, 2012: Vaccination with BM86, subolesin and akirin protective antigens for the control of tick infestations in white tailed deer and red deer. Vaccine 30, 273-279.
Coller, B.A., D.E. Clements, T. Martyak, M. Yelmene, M. Thorne and D.E. Parks, 2012: Advances in flavivirus vaccine development. IDrugs 13, 880- 884.
de la Fuente, J. 2012. Vaccines for vector control: Exciting possibilities for the future. The Vet. J. 194, 139- 140.
de la Fuente, J. and K.M. Kocan, 2003: Advances in the identification and characterization of protective antigens for development of recombinant vaccines against tick infestations. Expert Rev. Vaccines 2, 583-593.
de la Fuente, J. and K.M. Kocan, 2006: Strategies for development of vaccines for control of ixodid tick species. Parasite Immunol. 28, 275-283.
de la Fuente, J. and A. Estrada-Peña, 2012: Ticks and tick- borne pathogens on the rise. Ticks Tick-Borne Dis. 3, 115-116.
de la Fuente, J., C. Almazán, E.F. Blouin, V. Naranjo and K.M. Kocan, 2005: RNA interference screening in ticks for identification of protective antigens. Parasitol. Res. 96, 137-141.
de la Fuente, J., C. Almazán, E.F. Blouin, V. Naranjo and K.M. Kocan, 2006: Reduction of tick infections with Anaplasma marginale and A. phagocytophilum by targeting the tick protective antigen subolesin. Parasitol. Res. 100, 85-91.
de la Fuente, J., C. Almazán, M. Canales, J.M. Pérez de la Lastra, K.M. Kocan, P. Willadsen, 2007: A ten- year review of commercial vaccine performance for control of tick infestations on cattle. Anim. Health Res. Rev. 8, 23-28.
de la Fuente, J., R. Manzano-Roman, V. Naranjo, K.M. Kocan, Z. Zivkovic, E.F. Blouin, M. Canales, C. Almazán, R.C. Galindo, D.L. Step, M. Villar, 2010: Identification of protective antigens by RNA interference for control of the lone star tick, Amblyomma americanum. Vaccine 28, 1786- 1795.
de la Fuente, J., J.A. Moreno-Cid, M. Canales, M. Villar, J.M. Pérez de la Lastra, K.M. Kocan, R.C. Galindo, C. Almazán and E.F. Blouin, 2011: Targeting arthropod subolesin/akirin for the development of a universal vaccine for control of vector infestations and pathogen transmission. Vet. Parasitol. 181, 17-22.
Elvin, C.M. and D.H. Kemp, 1994: Generic approaches to obtaining efficacious antigens from vector arthropods. Int. J. Parasitol. 24, 67-79.
Fragoso, H., P. Hoshmand Rad, M. Ortiz, M. Rodríguez, M. Redondo, L. Herrera, and J. de la Fuente, 1998: Protection against Boophilus annulatus infestations in cattle vaccinated with the B. microplus Bm86-containing vaccine Gavac. Vaccine 16, 1990-1992.
Galindo, R.C., E. Doncel-Pérez, Z. Zivkovic, V. Naranjo, C. Gortazar, A.J. Mangold, M.P. Martín-Hernando, K.M. Kocan, and J. de la Fuente, 2009: Tick subolesin is an ortholog of the akirins described in insects and vertebrates. Dev. Comp. Immunol. 33, 612-617.
Gonsioroski, A.V., I.A. Bezerra, K.U. Utiumi, D. Driemeier, S.E. Farias, I.Jr da Silva Vaz and A. Masuda, 2012: Anti-tick monoclonal antibody applied by artificial capillary feeding in Rhipicephalus (Boophilus) microplus females. Exp. Parasitol. 130, 359-363.
Goto, A., K. Matsushita, V. Gesellchen, L. El Chamy, D. Kuttenkeuler, O. Takeuchi, J.A. Hoffmann, S. Akira, M. Boutros, J.M. Reichhart, 2008: Akirins are highly conserved nuclear proteins required for NF-kappaB-dependent gene expression in drosophila and mice. Nat. Immunol. 9, 97-104.
Harrington, D., M. Canales, J. de la Fuente, C. de Luna, K. Robinson, J. Guy and O. Spargano, 2009: Immunisation with recombinant proteins subolesin and Bm86 for the control of Dermanyssus gallinae in poultry. Vaccine 27, 4056-4063.
Hayes, E.B., 2012: Is it time for a new yellow fever vaccine? Vaccine 28, 8073-8076. Jones K.E., N.G. Patel, M.A. Levy, A. Storeygard, D. Balk, J.L. Gittleman and P.
&KDSWHU�9

74!
Daszac, 2008: Global trends in emerging infectious diseases. Nature 451, 990-994.
Karunamoorthi, K., 2011: Vector control: a cornerstone in the malaria elimination campaign. Clin. Microbiol. Infect. 17, 1608-1616.
Kocan, K. M., E. Blouin, and J. de la Fuente, 2011: RNA interference in ticks. J. Visualized Experiments 47, e2474. http://www.jove.com/details.stp?id=2474
Macqueen, D.J. and I.A. Johnston, 2009: Evolution of the multifaceted eukaryotic akirin gene family. BMC Evol. Biol. 9, 34.
Mangold, A.J., R.C. Galindo and J. de la Fuente, 2009: Response to the commentary of D. Macqueen on: Galindo R.C., Doncel-Pérez E., Zivkovic Z., Naranjo V., Gortazar C., Mangold A.J., Martín- Hernando, M.P., Kocan, K.M. and de la Fuente, J., 2009: Tick subolesin is an ortholog of the akirins described in insects and vertebrates [Dev. Comp. Immunol. 33, 612-617]. Dev. Comp. Immunol. 33, 878-879.
Manzano-Román, R., V. Díaz-Martín, A. Oleaga, M. Siles- Lucas and R. Pérez-Sánchez, 2012: Subolesin/akirin orthologs from Ornithodoros spp. soft ticks: Cloning, RNAi gene silencing and protective effect of the recombinant proteins. Vet. Parasitol. 185, 248-259.
Merino, O., C. Almazán, M. Canales, M. Villar, J.A. Moreno-Cid, A. Estrada-Peña, K.M. Kocan and J. de la Fuente, 2011a: Control of Rhipicephalus (Boophilus) microplus infestations by the combination of subolesin vaccination and tick autocidal control after subolesin gene knockdown in ticks fed on cattle. Vaccine 29, 2248-2254.
Merino, O., C. Almazán, M. Canales, M. Villar, J.A. Moreno-Cid, R.C. Galindo and J. de la Fuente, 2011b: Targeting the tick protective antigen subolesin reduces vector infestations and pathogen
Perspectives in the control of infectious diseases by transgenic mosquitoes in the post-genomic era--a review. Mem. Inst. Oswaldo Cruz 102, 425-433.
Willadsen, P., 2004: Anti-tick vaccines. Parasitol. 129, S367- S874.
infection by Anaplasma marginale and Babesia bigemina. Vaccine 29, 8575-8579.
Moreno-Cid, J. A., M. Jiménez, S. Cornelie, R. Molina, P. Alarcón, M-N Lacroix, R. Pinal, S. Delacour, J. Lucientes, M. Canales, J.M. Pérez de la Lastra, M. Villar and J. de la Fuente, 2010: Characterization of Aedes albopictus akirin for the control of mosquito and sand fly infestations. Vaccine 29, 77-82.
Moreno-Cid, J.A., M. Canales and J. de la Fuente, 2012: Production of recombinant Aedes albopictus akirin in Pichia pastoris using an aqueous two- phase semicontinuous fermentation process. Biochem. Eng. J. 68, 114-119.
Moreno-Cid, J.A., Pérez de la Lastra J.M., Villar M., Jiménez M., Pinal R., Estrada-Peña A., Alarcón P., Delacour S., Oropeza V., Ruiz I., Molina R., Lucientes J., Prudencio C.R., Galindo R.C., Almazán C., Nijhof A.M., Mangold A.J., Gortázar C. and de la Fuente J., 2013: Control of multiple arthropod vector infestations with subolesin/akirin vaccines. Vaccine (In Press).
Nowak, S.J., H. Aihara, K. Gonzalez, Y. Nibu, M.K. Baylies, 2012: Akirin links Twist-regulated transcription with the Brahma chromatin remodeling complex during embryogenesis. PLoS Genet. 8(3), e1002547.
Prudencio, C.R., J.M. Pérez de la Lastra, M. Canales, M. Villar, J. de la Fuente, 2010: Mapping protective epitopes in the tick and mosquito subolesin ortholog proteins. Vaccine 28, 5398-5406.
Rodriguez-Valle, M., M. Vance, P.M. Moolhuijzen, X. Tao and A.E. Lew-Tabor, 2012: Differential recognition by tick-resistant cattle of the recombinantly expressed Rhipicephalus microplus serine protease inhibitor-3 (RMS-3). Ticks Tick Borne Dis. 3, 159-169.
Sperança, M.A. and M.L. Capurro, 2007:
&KDSWHU�9


CONCLUSIONES(((
CONCLUSIONS(


76
CONCLUSIONES
1. El sistema semicontinuo en dos fases fases acuosas (ATPS) utilizando la levadura
P. pastoris para la producción de proteínas recombinantes es factible, facilmente
escalable e integrable en proceso, biocompatible y además permite recircular y
recuperar corriente de proceso no consumida. El sistema ATPS semicontinuo logra
altos niveles de producción de proteínas recombinantes comparado con un proceso
sin recirculación.
2. La producción de antígenos quiméricos utilizando E.coli como sistema de
expression, formado por polipeptidos de garrapata fusionados a MSP1a y
constituido por fracciones de membrana de E.coli con antígenos/epítopos expuestos
sobre superficie de membrana da como resultado un método económico e
inmunogénico para producción de vacunas protectoras contra garrapatas con
niveles aceptables de endotoxinas.
3. El antigeno Akirina (AKR) del mosquito A. Albopictus podría ser utilizado para
vacunar hospedadores y controlar las infestaciones de mosquitos y flebótomos. El
efecto de reducción en la oviposición lograría un progresivo control generacional
en las poblaciones de insectos.
4. Las fracciones de membrana de E.coli recombinante compuesta por la quimera
SUB-MSP1a, confiere protección frente a las infestaciones por garrapatas.
5. Los antígenos recombinantes SUB, AKR y quimeras Q38, Q41 de SUB/AKR,
podrían ser utilizados para el desarrollo de una vacuna “universal” para el control
multiple de artrópodos vectores como garrapatas, mosquitos y flebótomos. Estas
vacunas podrían ser usadas para inmunizar hospedadores reservorios y reducir las
poblaciones de vectores y por tanto el riesgo de transmisión de patógenos a
humanos y animales.

77
CONCLUSSIONS
1. The semicontinuous aqueous two-phase system (ATPS) using the yeast P. pastoris
for the production of recombinant proteins is feasible, easy to scale up with high
process integration, capability, biocompatibility and effective stream recycling and
recovery. The ATPS can reach higher recombinant protein production when
compared to the process without stream recycling.
2. The E. coli expression system for the production of chimeric antigens composed of
tick polypeptides fused to MSP1a is a cost-effective process for the development
of tick vaccines based on recombinant E. coli membrane fractions containing
surface-exposed protective antigens/epitopes with acceptable endotoxin levels and
enhanced immunogenicity.
3. Recombinant A. albopictus Akirin (AKR) could be used to vaccinate hosts for the
control of mosquito and sand fly infestations. Reduction in oviposition would
result in the reduction of insect populations in subsequent generations.
4. The recombinant E. coli membrane fractions containing the SUB-MSP1a chimera
confer protection against cattle tick infestations.
5. Recombinant SUB, AKR and SUB/AKR chimeras Q38 and Q41 could be used to
develop “universal” vaccines for the control of multiple arthropod vectors such as
ticks, mosquitoes and sand flies. These vaccines could be used to immunize
reservoir hosts and populations at risk to reduce vector populations and thus the
risk of pathogen transmission to humans and animals.



RESUMEN((
SUMMARY(


79!
RESUMEN
Las enfermedades causadas por patógenos transmitidos por vectores como garrapatas, afectan a humanos y animales en regiones de clima tropical y subtropical, con un registro del 20% de todas la enfermedades infecciosas emergentes de las últimas decadas e importante impacto económico en la industrLa ganadera. El método de control más utilizado para reducir las infestaciones por garrapatas ha sido el químico, método que desarrolla garrapatas químicamente resistentes y genera residuos. El método de control inmunológico mediante vacunación ha emergido como una alternativa más económica, ambientalmente amigable y que evita la generación de garrapatas quimiorresistentes por el tratamiento continuo de acaricidas. Además, el empleo de vacunas contra garrapatas utilizando multiples antígenos logra protección frente un amplio rango de especies y previene o reduce la transmisión de patógenos (capítuOo I). Resultados experimentales han demostrado que antígHnos protectores contra garrapatas pueden ser utilizados para el desarrollo de vacunas con el objetivo dual de controlar las infestaciones por artrópodos, y además reducir su capacidad como vectores para la transmisión de patógenos. Los antígenos protectores Subolesina (SUB) y Akirina (AKR), muy conservados entre diferentes especies de vectores, han demostrado ser excelentes candidatos para el desarrollo de una vacuna control “universal” para este fin. El objetivo principal de esta tesis de investigación está enfocado en el desarrollo de vacunas para el control inmunológico de las infestaciones por ectoparisitos hematófagos y las enfermedades que transmiten.
La levadura Pichia pastoris es un eficiente sistema para producir proteínas heterólogas (capítulo II). En este capítulo se ha establecido y desarrollado por primera vez en P. pastoris un sistema de producción de antígenos recombinantes mediante tecnología de bioconversion extractiva en sistema de dos fases acuosas (ATPS) en proceso semicontinuo. La producción de AKR secretada en P. pastoris mediante fermentación extractiva en ATPS, permitió ensayos de vacunación a gran escala para evaluar la capacidad protectora del antígeno frente diferentes especies de artrópodos vectores. La tecnología desarrollada en este capítulo aporta un novedoso y eficiente proceso en la levadura para producir AKR de mosquito, además de la posiblidad de aplicar este proceso para producir de manera eficiente otras proteinas recombinantes secretadas en P.�pastoris.
En el capítulo III se desarrollaron ensayos de vacunación para FDUFDWHUL]DU los antígenos. Como primer trabajo, se vacunaron ratones con AKR recombinante producida mediante tecnología ATPS y se evaluó su capacidad protectora por infestación de los ratones con las especies de artrópodos Aedes albopictus� $�� DHJ\SWL� and Phlebotomus perniciosus. Como resultado destacable, la vacunación con AKR redujo la oviposición de A.albopictus y P. perniciosus en un 17% y 31% frente el grupo control, respectivamente. Además, la fertilidad (Nº de adultos/huevo) se redujo en un 60% en flebotomos alimentados sobre ratones vacunados. En cambio, no se observaron resultados significativos sobre los mosquitos A. aegypti DOLPHQWDGRV��Vobre ratones vacunados. Estos resultados preliminares demRstraron la posibilidad de utilizar el antígeno AKR como vacuna para el control de las infestaciones por mosquitos y flebotomos.

80!
&RPR� VHJXQGR� WUDEDMR� GHQWUR� GHO� FDStWXOR� ,,,�� VH� FDUDFWHUL]y� � HO� VLVWHPD� TXLPpULFR� GH�H[SUHVLyQ� HQ� PHPEUDQD� GH� (�FROL� 68%�063�D�� � (O� VLVWHPD� HVWi� IRUPDGR� SRU� S SWLGRV�DQ tJHQRV� SURWHFWRUHV� IXVLRQDGRV� D� OD� SURWHtQD� GH� VXSHUILFLH� PD\RULWDULD� $QDSODVPD�PDUJLQDOH��063��HQ�(�FROL��(VWH�VLVWHPD�GLy�FRPR�UHVXOWDGR�XQD�WHFQRORJtD�HFRQyPLFD�SDUD�SURGXFLU�YDFXQDV��PHGLDQWH�XQ�VLVWHPD�GH�H[SUHVLyQ�HQ�PHPEUDQD�GH�(�FROL�TXH�DXPHQWy�OD�LQPXQRJHQHLFLGDG�GHO�DQWtJHQR�HQ�HVWXGLRV�FRQ�UDWRQHV�\�FHUGRV�FRPR�PRGHORV��GDQGR�FRPR�UHVXOWDGR�XQ�HILFD]�\�D�EDMR�FRVWR�VLVWHPD�\�FRQ�QLYHOHV�DFHSWDEOHV�GH�HQGRWR[LQDV�SDUD�HVWH�WLSR�GH�YDFXQDV��
En el capítulo IV�� VH� HYDOXy� OD� FDSDFLGDG� SURWHFWRUD� GH� ORV� DQWtJHQRV�� 3DUD� HO� FRQWURO�PXOWLSOH�GH�DUWUySRGRV�YHFWRUHV��VH�FRPSDUy�HO�HIHFWR�GH�YDFXQDFLyQ�FRQ�68%��$.5�\�ORV�DQWtJHQRV� TXLPpULFRV� 4��� \� 4��� FRQ� HStWRSRV� SURWHFWRUHV� FRQVHUYDGRV� GH� 68%�$.5�IUHQWH�D�JDUUDSDWDV��PRVTXLWRV�\� IOHERWRPRV�HYDOXDQGR� OD� HILFDFLD�HQ�HO� FRQWURO�JOREDO�(�VHJ~Q� ORV� SDUiPHWURV� PRUWDOLGDG�� PXGD�� RYLSRVLFLyQ� \�R� IHUWLOLGDG�� 6H� REWXYR� TXH� HO�DQWtJHQR� 68%� ORJUy� OD� PD\RU� HILFLDFLD� JOREDO� �(�� HQWUH� WRGDV� ODV� HVSHFLHV� HVWXGLDGDV�����������OD�YDFXQDFLyQ�FRQ�4���GLy�XQD�PD\RU�(�HQ�PRVTXLWRV�������SRU�UHGXFFLyQ�GH�OD�VXSHUYLYHQFLD�\�OD�IHUWLOLGDG��\�OD�YDFXQDFLyQ�FRQ�4���WXYR�OD�PD\RU�(�GH�FRQWURO�VREUH�OD� RYLSRVLFLyQ� VREUH� PRVTXLWR� \� IOHERWRPRV� FRQ� XQD� UHGXFFLyQ� GHO� ���� \� HO� �����UHVSHFWLYDPHQWH�� (Q� XQ� VHJXQGR� WUDEDMR� GHQWUR� GHO� FDStWXOR� ,9�� se HYDOXy la capacidad protectora GHO� � VLVWHPD� TXLPpULFR� HQ� PHPEUDQD� 68%�063�D�� DGHPiV� GHO� IDFWRU� GH�HORQJDFLyQ� �()�D�063�D�� \� XELTXLWLQD� �8%4�063�D�� GH� JDUUDSDWD�� &RPR� UHVXOWDGR�GHVWDFDEOH�GHO�FDStWXOR�� OD�TXLPHUD�68%�063�D�ORJUy�XQD�HILFDFLD�JOREDO�(�SRU�HQFLPD�GHO����� HQ� HQVD\RV� GH� YDFXQDFLyQ� HQ� ERYLQRV� LQIHVWDGRV� FRQ� JDUUDSDWDV� GH� OD� HVSHFLH�5LSKLFHSKDOXV� PLFURSOXV�� GHPRVWUDQGR� OD� HILFDFLD� GHO� HFRQyPLFR� VLVWHPD� SDUD� SURGXFLU�YDFXQDV�SURWHFWRUDV�FRQWUD�JDUUDSDWDV�
Los resultados de este trabajo de investigación han demostrado la eficacia empleando los ortólogos SUB/AKR en el control de las infestaciones e infección por artrópodos hematófagos (capítulo V). Sin embargo, el control m~ltiple total de artrópodos vectores utilizando una vacuna universal basada en los antígenos SUB/AKR está aún en fase inicial, siendo necesario continuar los trabajos de investigación para este fin.

81
SUMMARY
DiseaseV caused by vector-borne pathogens greatly impact human and animal health in tropical and sub-tropical regions of the world, accounting for over 20% of all emerging infectious diseases recorded in recent decades, they are also the main cause of production losses in the cattle industry. Control of cattle tick infestations has been based primarily on the application of acaricides resulting in the selection of resistant ticks and environmental pollution. Vaccination against cattle ticks has emerged as an alternative, with the advantage of being cost-effective and avoiding environmental contamination and selection of drug-resistant ticks resulting from repeated acaricide treatments. In addition, the development of vaccines against ticks using multiple antigens could target a broad range of tick species and may also prevent or reduce transmission of pathogens (chapter I). Experiments have provided evidence that tick protective antigens can be used for the development of vaccines with the dual target of controlling arthropod infestations and reducing their vector capacity for pathogens. The protective antigens Subolesin (SUB) and Akirin (AKR), which are highly conserved across vector species, are promising targets for the development of a universal vaccine for the control of arthropod infestations and the reduction of pathogen transmission. The aim of this research thesis ZDV focused on the development of vaccines for immunological control of hematophagous ectoparasites infestation and the diseases they transmit.
The yeast Pichia pastoris has been successfully used as an efficient system to produce heterologous proteins ( chapter II). In this chapter, ZH��establishHG a n d d e v e l o pHG��IRU�� WKH��ILUVW�� WLPH an extractive semicontinuous bioconversion in an aqueous two-phase system (ATPS) by using P.pastoris for the recombinant antigens production. The efficient secretion of the recombinant AKR from P. pastoris by extractive fermentation in ATPS process has allowed conducting large-scale vaccination trials to assay its protective capacity against different arthropod vector species. The technology developed in this chapter provides a novel and efficient process for the production process for mosquito AKR by using the yeast, and suggest the potential to develop similar processes for the expression of other recombinant proteins�
In chapter III, vaccination trials were conducted to evaluate the protective capacity of the antigens. To test the effect of recombinant AKR produced by ATPS technology���vaccination trials on mice infested with Aedes albopictus, A� aegypti and Phlebotomus perniciosus��ZHUH��SHUIRUPHG� Oviposition of A. albopictus and P. perniciosus fed on mice vaccinated with recombinant AKR was reduced by 17% and 31%, respectively when compared with controls. In addition, fertility (number of adults/egg) was reduced by 60% in sand flies fed on vaccinated mice. Antibody responses were significantly higher in vaccinated animals when compared with controls. Nevertheless, Aedes aegypti mosquitoes were not affected after feeding on vaccinated mice. These results showed� WKDW� UHFRPELQDQW� $.5� FRXOG� EH� XVHG� WR� YDFFLQDWH� hosts for the control of mosquito and sand fly infestations. ,QWR� WKH� FKDSWHU� ,,,�� DV� VHFRQG� H[SHULPHQW� WKH�FKDUDFWHUL]DWLRQ�RI� D� QRYHO� H[SUHVVLRQ� V\VWHP� IRU� WLFN� SURWHFWLYH� DQWLJHQV�ZDV� SHUIRUPHG��7KH�V\VWHP� LV� EDVHG� DV� LPPXQRJHQLF� SHSWLGHV�DQWLJHQV� IXVHG� WR� WKH� $QDSODVPD� PDUJLQDOH� PDMRU�VXUIDFH� SURWHLQ� �D� �063�D�� 1�WHUPLQDO� UHJLRQ� IRU� SUHVHQWDWLRQ� RQ� WKH� (VFKHULFKLD� FROL�PHPEUDQH�

82
7KLV�V\VWHP�SURYLGHV�D�QRYHO��VLPSOH�DQG�FRVW�HIIHFWLYH�DSSURDFK�IRU�WKH�SURGXFWLRQ�RI�DQWLJHQV�E\�VXUIDFH�GLVSOD\�RI�DQWLJHQLF�SURWHLQ�FKLPHUDV�RQ�WKH�(��FROL�PHPEUDQH�DQG�GHPRQVWUDWHV�WKH�SRVVLELOLW\� RI� XVLQJ� UHFRPELQDQW� EDFWHULDO� PHPEUDQH� IUDFWLRQV�� ZKLFK� HQKDQFH� WKH�LPPXQRJHQLFLW\�RI�WKH�68%�063�D�DQWLJHQ��UHVXOWV�SURYLGHG�E\�YDFFLQDWLRQ�WULDOV��LQ�PRXVH�DQG�SLJ�PRGHOV��7KH�SURGXFWLRQ�RI�WKHVH�PHPEUDQH�ERXQG�DQWLJHQV�LV�VLPSOH�DQG�FRVW�HIIHFWLYH�DQG�UHVXOWV� LQ� DQWLJHQ�SUHSDUDWLRQV�ZLWK� LQFUHDVHG� LPPXQRJHQLFLW\� DQG� HQGRWR[LQ� OHYHOV�ZLWKLQ� WKH�OLPLWV�DFFHSWDEOH�IRU�UHFRPELQDQW�YDFFLQH�IRUPXODWLRQV�
In the chapter IV��ZH�HYDOXDWHG�WKH�SURWHFWLYH�FDSDFLW\�RI�WKH�DQWLJHQV�IRU�WKH�PXOWLSOH�FRQWURO�RI�DUWKURSRG�YHFWRUV��:H�GHVFULEHG�WKH�FRPSDUDWLYH�HIIHFW�RI�YDFFLQDWLRQ�ZLWK�68%��$.5�DQG�4���DQG�4��� FKLPHUDV� FRQWDLQLQJ�68%�$.5�FRQVHUYHG�SURWHFWLYH� HSLWRSHV� RQ� WLFN��PRVTXLWRHV� DQG�VDQG� IOLHV� RQ� YHFWRU�PRUWDOLW\��PROWLQJ�� RYLSRVLWLRQ� DQG�RU� IHUWLOLW\��:H� GHPRQVWUDWHG� WKDW� 68%�YDFFLQDWLRQ�KDG�WKH�KLJKHVW�HIILFDF\��(��DFURVV�DOO�YHFWRU�VSHFLHV����±������4���YDFFLQDWLRQ�KDG�WKH� KLJKHVW� YDFFLQH� (� LQ�PRVTXLWRHV� ������ E\� UHGXFLQJ� IHPDOH� VXUYLYDO� DQG� IHUWLOLW\�� DQG� 4���YDFFLQDWLRQ�KDG� WKH� KLJKHVW� HIIHFW� RQ� UHGXFLQJ�PRVTXLWR� ������ DQG� VDQG� IO\� ������RYLSRVLWLRQ�� 7KH�VHFRQG�H[SHULPHQW�LQWR�WKH�FKDSWHU�,9�ZDV�FRQGXFWHG�WR�HYDOXDWH�WKH�SURWHFWLYH�FDSDFLW\�RI� WKH tick Subolesin (SUB-MSP1a) chimeric antigen, as well as the ticks Elongation factor (EF1a-MSP1a) and Ubiquitin (UBQ-MSP1a) fusion proteins on the E. coli membrane system and demonstrating that bacterial membranes containing the chimeric proteins are protective� $V�D�KLJKOLJKW� UHVXOW SUB-MSP1a were protective (>60% vaccine efficacy) against experimental R. microplus in cattle�� � DQG� GHPRQVWUDWLQJ� WKH� HIILFDF\� RI� WKH� FRVW�HIIHFWLYH� V\VWHP� IRU� YDFFLQH�SUHSDUDWLRQV�WR�SURWHFW�FDWWHO�DJDLQVW�WLFN�LQIHVWDWLRQV�
The results REWDLQHG in this research work provide evidence for the use of subolesin/akirin orthologs for the control of acarine and insect infestations and infection capacity (chapter V). However, the development of a subolesin/akirin-based universal vaccine for total multiple control of arthropod vector species and the transmission of pathogens is still in i t s i n f a n c y , therefore, further research is needed LQ�RUGHU�WR�achieYH this goal.


AGRADECIMIENTOS
$�PL�IDPLOLD ��SRU�VX�DSR\R�\�SDFLHQFLD��(YD�\�&HOLD��FRPR�GLMR�2VFDU�:LOGH��ODV�
PXMHUHV� HVWiQ� SDUD� VHU� DPDGDV�� QR� FRPSUHQGLGDV��� $� PLV� SDGUHV�� -XDQ�
$QWRQLR�\�7RxL��SRU�VX�D\XGD�\�DSR\R�\�QR�GHMDUPH�QXQFD�FROJDGR�D�SHVDU�GH�
FXDOTXLHUD�GH�PLV�GHFLVLRQHV��D�PLV�KHUPDQRV��OD�'UD��0DULSL��HO�'U��)UDQFLVFR��
(OLVD� \� )UDQFLVTXLQKR�� TXH� SHVH� D� OD� GLVWDQFLD� VLHPSUH� KDQ� HVWDGR� SUHVHQWHV�
�DGHPiV� GH� ORV� VREULQRV� YHQLGHURV«��� PL� 7DWD� \� VXV� WRUWLOODV� GH� SDWDWDV�� HO��
UHFXHUGR� GH� PL� DEXHOR� $QWRQLR� \� /D� 'HOLFLRVD�� D� PL V � DEXHORV�� )UDQFLVFR� \�
5H\HV��SRU�SUHRFXSDUVH�VLHPSUH�GH�PL��$�PL�IDPLOLD�0XUFLDQLFD��'LHJR��,VDEHO��
)UDQFLVFR��$QD��9LFWRU�\�.LNH���<�FRPR�QR��/HR��
$� PLV� DPLJRV� \� FRPSDxHURV� GHO� ,5(&� SRU� DPHQL]DU� ODV� ODUJDV� KRUDV� GH�
ODERUDWRULR��D�1LHYHV�\�'DYLG�\�VXV�WDUWDV�GH���FKRFRODWHV��PL�HVWLPDGR�2FWDYLR�
\�VXV�PDGUXJRQHV��ODV�5HEHFDV��0DULQHOD�\�VXV�LQWHUPLQDEOHV�3&5V��*UDFL�\�VXV�
DOJDV�� 0DULVD� \� 0DULD� /XLVD��� 0DULQD� \� VX� SHUIHFWR� HVSDxRO�� /RXUGHV� \� VXV�
YHQJDQ]DV�� ,VD� \� VXV� EURPDV�� 3LOL� \� VXV� WUDGXFFLRQHV�� -RVH� 0DQXHO� \� VX�
WUDQTXLOLGDG�\�0DUJD�\�VX�GHVSDFKR�ODERUDWRULR��$�PL�DPLJR�SRUWXJXpV�0DULR�
GD�&RVWD�\�VXV�HQVDODGDV�GH�SROOR��$�'DYLG�\�VXV�LQWHUPLQDEOHV�JHVWLRQHV�H�
LQWHUPLQDDDEOHV� OODPDGDV� WHOHIyQLFDV�$�(QFDUQL�\� VXV�SHWLFLFLRQHV� IDOOLGDV�GH�
WLUDPLV~��$�)UDQ�SRU�VX�ELHQ�HVWDU��
$� ORV� DPLJRV� GHO� 0DVWHU� GHO� ,5(&�� $QGUpV�� -RVH�� -DYLYL�� (VSH�� ,VD�� 3DEOR�
0RUDOHV��3DEOLWR��0DULEHO��\�%HD��OD�PHMRU�FRQWULEXFLyQ�GHO�0DVWHU��
$�3HSH�SRU�VX�DSRUWDFLyQ�FLHQWtILFD�\�UHDOL]DFLyQ�GH�HVWD�WHVLV��$�0DULR�&DQDOHV�
FRPR�PL�LQLFLDGRU�HQ�OD�ELRWHFQRORJtD�\�SUHRFXSDUVH�GH�PL�IRUPDFLyQ�LQLFLDO��\�
VX�FRQH[LyQ�FRQ�$UQROG�'HPDLQ�SDUD�FRQRFHU�HO�&%%�\�OD�8QLYHUVLGDG�GH�,RZD��
3RU� VXSXHVWR�� DO� &%%� HQ� ,RZD� \� VX� FDOXURVD� DFRJLGD� GXUDQWH� PL� HVWDQFLD��
JUDFLDV� DO� SURIHVRU�0DQL� 6XEUDPDQLDQ�� OD� DWHQFLyQ� GH� 6KULGDU� *RSLVKHWW\� \�
D\XGD�GH� 6KXYHQGX��5\DQ�� -LQJ\LQJ�\�0LWFK� �WKDQNV� IRU�\RXU� IULHQGVKLS�� IRU�
OHWWLQJ�PH�ZRUN�RQ�\RXU�ODE�DQG�EH�SDUW�RI�\RXU�UHVHDUFK�WHDP���$�PL�DPLJD�
&KHOR�SRU�VXV�FRQVHMRV�
(Q� JHQHUDO� D� WRGR� HO� ,5(&�� GRQGH� OD� UHODFLyQ� FRQ� WRGRV� VXV� FRPSRQHQHWHV� KD�
VLGR� HVWXSHQGD� �$OPXGHQD�� /XFtD�� -RUJH�� -XDQ� &DUORV�� $UWXUR��\� &DUROLQD�� \�
72'26�/26�'(0É6�,5(48,$126��
-�$�0�&�0






























