Embed Size (px)
Citation preview

ELSEVIER
SUCCESSFUL IN VITRO PRODUCTION OF EMBRYOS IN THE RED DEER (Cervus elaphus) AND THE SIKA DEER (Cervus nippon)
P. Comizzoli,l,2 . 2a 2 1 P. Mermfllod, Y. Cogni6, N. Chai, X. Legendre 1 and R. Mauget 1
1MNHN, Conservation des Esp6ces Animales, Parc Zoologique - 75012 - Paris, France
2INRA, Oocyte and Development Unit - 37380 - Nouzilly, France
Received for publication: March 9, 2000 Accepted: June 21, 2000
ABSTRACT
The aim of our study was to define the conditions for IVM and IVF of oocytes in 2 common deer species as models for endangered related subspecies. Immature oocytes were recovered during the breeding season from postmortem ovaries (red deer) or by repeated laparoscopic follicular aspiration (sika deer). Oocytes were cultured for 24 h in IVM medium supplemented with EGF or FSH and follicular fluid. Stag semen was collected by electroejaculation (both species) or by epididymal flushing (red deer) and cryopreserved. For IVF, oocytes were exposed to different concentrations of thawed spermatozoa in a modified Tyrode albumin lactate pyruvate medium supplemented with 20% (v/v) estrus sheep serum for 18 h. After IVF, presumptive zygotes were allowed to develop in vitro for 7 days in synthetic oviduct fluid (SOF) supplemented with fetal calf serum (10%, v/v) . In both species, the presence of ovine FSH and follicular fluid improved the in vitro maturation rate. In the sika deer, the optimal
sperm concentration for IVF was 106/mL and some fertilized oocytes reached the early morula stage (20 to 25 cells). In the red deer, after IVF with ejaculated or epididymal
spermatozoa (2.0 x 106/mL), 20% of zygotes developed to the blastocyst stage (50 to 80 cells). © 2001 by Elsevier Science Inc.
Key words: IVM, IVF, ovum pickup, Cervus elaphus, Cervus nippon, blastocyst
Acknowledgments The authors thank the staff of the Parc Animalier de la Haute Touche and Y. Gu6rin for his assistance. We are also indebted to Etablissement Gourault, Fine Venaison and Vauventriers, for red deer ovaries and testes. P. Comizzoli was supported by the Conseil R6gional de la R6gion Centre (grant No 97 298 035).
'~Correspondence and reprint requests.
Theriogenology 55:649-659, 2001 O093-691X/O1/$-see front matter © 2001 Elsevier Science Inc. PII: S0093-691 X(01)00433-2

650 Theriogenology
INTRODUCTION
Many species of deer are rare, endangered or threatened in their natural habitat (13). Such assisted reproduction techniques as superovulation, artificial insemination and embryo transfer are possible alternatives for the propagation of small populations (16, 29) and have been used for the production of red deer, fallow deer and Eld's deer (1, 16, 23). Additionally, as shown in semidomestic species (17, 25), successful in vitro production (IVP) has the potential to produce large number of embryos from two selected genitors. Few manipulations of the animals are required; thus, stressful effects of repeated handling and anesthesia are reduced. Morulae or blastocysts can be then transferred to synchronized recipient females or frozen and stored.
To define the conditions for IVP in endangered deer species both red deer (Cervus elaphus) and sika deer (Cervus nippon) are potentially good models. These species are widespread and several related subspecies are threatened (e.g., Cervus elaphus bactrianus, Cervus nippon pseudaxis, Cervus nippon taiouanus). As in other species, deer oocytes may be recovered from slaughtered or hunted animals (4, 7). Ovum pickup (OPU) by laparoscopic follicular aspiration is a useful technique to repeatedly recover immature oocytes from live animals (27). The use of this procedure has not been described in wild deer, but it could be adapted to collect oocytes from individuals of endangered populations raised in semicaptivity. Previous studies have shown that stag semen can be cryopreserved after collection by electroejaculation (3, 4) or postmortem epididymal flushing (3, 7, 12, 32). The IVP of blastocysts from red deer oocytes fertilized by epididymal spermatozoa has not been reported.
Previous studies of IVP of embryos in deer species (3, 4, 6, 7, 11, 18) have used protocols similar to those widely used in domestic animals, but in vitro development has been poor and inconstant. In the sika deer, there have been few studies on assisted breeding (30, 31) and we are not aware of any data on IVP of embryos.
The objectives of the present study were to 1) develop a method for laparoscopic oocyte recovery in sika deer and 2) evaluate some of factors affecting IVM, IVF of oocytes and in vitro development of embryos in both red deer after postmortem oocyte recovery and sika deer after OPU. Specifically, we compared 1) the effect of EGF vs. FSH and either ovine (red deer and sika deer) or caprine (red deer only) follicular fluid supplementation on IVM of red deer and sika deer oocytes (Experiment 1); and 2) the effect of the follicular fluid source during IVM and sperm concentration used for IVF on fertilization rates, incidence of polyspermy (Experiment 2) and in vitro development (Experiment 3).

Theriogenology 651
MATERIALS AND METHODS
Gamete Recovery
Oocyte recovery. Ovaries of adult red deer hinds were harvested at a local slaughterhouse during the breeding season. Immature oocytes were recovered from antral follicles with an aspiration needle (19 1/2 gauge) using gentle vacuum (-30 mm Hg). Immature oocytes of anesthesied adult sika deer hinds (2 to 6 yr old) were recovered by laparoscopic follicular aspiration as described in a preliminary study (21). Sixteen females were randomly allocated in 4 groups, and oocytes were collected every 2 wk from November to March. Only good-quality oocytes (homogenous cytoplasm, surrounded by more than 2 cell layers) were used for IVP.
Semen collection and cryopreservation. During the breeding season (October and November), semen was collected by electroejaculation from stags (3 to 8 yr old) restrained in a drop-floor deer cradle. In the red deer, epididymal spermatozoa were obtained postmortem by retrograde flushing of the ductus deferens of 18- to 24-mo old stags. Individual semen samples were diluted in a citrate-based extender (8) containing 10% egg yolk (v/v), 7% skimmed milk powder (wt/vol), 4% lactose (wt/vol) and 2%
glycerol (v/v). Diluted samples were loaded into 0.25-mL straws (50 x 10 ~ spermatozoa/straw), and placed in liquid nitrogen vapor (about -85°C) for 8 min before being plunged into liquid nitrogen. Spermatozoa from a single ejaculate (red deer and sika deer) or a single epididymal collection (red deer only) were used in all experiments.
IVM/IVF and in vitro development
In Experiment 1, immature oocytes (3 replicates) were cultured in M199 a medium
supplemented with EGF a (10 ng/mL) or ovine FSH b (oFSH, 50 ng/mL) + 10% (v/v) follicular fluid (FF) for 24 h at 38.5°C under 5% CO 2 in humidified air. Follicular fluid was obtained from goats after eCG stimulation (cFF) and from sheep (oFF) by aspiration of ovarian follicles (> 5 mm diameter) and centrifuged; the supernatant was stored at -20°C. The chromatin status of oocytes after 24 h was assessed with
epifluorescence microscopy c after fixation in ethanol and staining with Hoechst 33342 a.
a ,
Sigma Chemical Co., St Louis, MO
bovagen, ICP, New Zealand C ,
Nlkon, Tokyo, Japan

652 Theriogenology
In Experiment 2, the effects of supplementation of oocyte maturation medium
with either sheep or goat FF and sperm concentration (0.5, 1.0 or 2.0 x 106/mL ) were evaluated for the purpose of maximizing normal fertilization rates and minimizing polyspermy. Cumulus-oocyte complexes (4 replicates) were vortexed after IVM to remove cumulus cells before IVF in a modified Tyrode albumin lactate pyruvate (TALP) (26) medium supplemented with 20% (v/v) estrous sheep serum (SS) at 38.5°C under 5% CO 2 in humidified air. After thawing, motile spermatozoa (ejaculated or
epididymal) were selected by a Percoll a gradient and inseminated at a concentration of
0.5, 1.0 or 2.0 x 106/mL. Presumptive zygotes were fixed at 15 to 18 h postinsemination and labeled with Hoechst 33342 to assess the number of pronuclei per oocyte using epifluorescence microscopy.
In Experiment 3, some of the presumptive zygotes from Experiment 2 were allowed to develop in vitro for 7 days in synthetic oviduct fluid (SOF) supplemented
with amino acids and 10% (v/v) fetal calf serum b (FCS) at 38.5°C under 5% CO2, 5% O2 and 90% N 2. Cleavage rates were determined at 20 and 28 h postinsemination. After 7 d of culture, the cell number of blastocysts was assessed by Hoechst 33342 staining after ethanol fixation.
Data from each replicate were pooled to calculate percentages based on the total number of oocytes subjected to WM. Differences between values were analyzed by using the Chi-square test.
RESULTS
Immature Oocyte Recovery
Of a total of 431 ovaries from red deer hinds, an average of 4.5 oocytes was obtained per ovary by aspiration of antral follicles. An average of 3.4 (75%) good- quality oocytes was recovered per ovary. The results of laparoscopic OPU in sika deer hinds are shown in Table 1. No health problems were associated with repeated OPU in the 4 groups of hinds. The oocyte recovery rate was 57% (150 oocytes from 263 follicles), and the percentage of good-quality oocytes was 70% (105 good-quality oocytes of 150 total oocytes).
aPharmacia, Uppsala, Sweden
bsigma

Theriogenology 653
Table 1. Results of ovum pickup in sika deer hinds Hinds, n Procedures, n Procedures/hind, mean (range) Follicles aspirated, n Total oocytes recovered, n Good quality oocytes recovered, n Total oocytes/hind per procedure, mean (range) Good quality oocytes/hind per procedure, mean
16 70
4.4 (2-10) 263 150 105
2.1 (0-6) 1.5
Experiment I
The percentage of oocytes in Metaphase II after 24-h IVM was increased (P<0.05) in the presence of oFSH and FF for both species as compared with the medium supplemented with EGF (Table 2). The IVM rate of red deer oocytes cultured in medium with cFF (85%) was not different (P>0.05) from that of oocytes cultured in oFF (78%).
Table 2.
Species Red deer
Results of IVM in M199 supplemented with EGF, oFSH + ovine FF (oFF) or oFSH + eCG stimulated goat FF (cFF)
Medium Oocytes (n) M199 + EGF 54
M199 + oFSH + oFF 89
M199 + oFSH + cFF 99
Metaphase II (%)
25 a 78 b
85 b
Sika deer M199 + EGF 14 14 a
M199 + oFSH + oFF 25 76 b
Values with a vs. b superscripts differ significantly (P<0.05, Chi-square test).
Experiment 2
As shown in Table 3, fertilization rates were lower (P<0.05) using 0.5 x 106
spermatozoa/mL. Fertilization rates were similar for oocytes of both species w h e n 106
spermatozoa /mL were used for IVF. At 1.0 x 10 6 spermatozoa/mL, an increased incidence (P<0.05) of polyspermy (22% of fertilized oocytes with 3 to 5 pronuclei per oocyte) was observed in sika deer. Since no polyspermy was observed in the red deer
with 1.0 x 10 6 spermatozoa/mL, further experiments were carried out to improve the fertilization rate. Increasing the sperm concentration (either ejaculated or epididymal) for IVF or IVM in medium supplemented with cFF did not improve the fertilization rate
of red deer oocytes. However, at a sperm concentration of 2.0 x 106/mL a higher incidence of polyspermy occurred. In fertilized oocytes, the presence of 2 pronuclei of

654 Theriogenology
different size (probably male and female pronuclei, Figure la) shows that the spontaneous activation of the oocytes was unlikely.
Table 3. Effects of follicular fluid source (FF) during IVM and sperm concentration on in vitro fertilization rates
Species FF source Sperm/mL Oocytes Fertilization Zygotes with > 2 during IVM ~ (106) (n) (%)b pronuclei (%)
Red deer Ovine 0.5 87 35 ~ 0
Ovine 1.0 91 55 a 0
Ovine 2.0 40 65 d 9
Caprine 2.0 70 63 a 11
Caprine 2.0 epid. 44 46 cd 7
Sika deer Ovine 0.5 27 26 ~ 3 ~
Ovine 1.0 43 56 a 22 d
~With ovine FSH.
bThis includes both normally fertilized and polyspermic oocytes. Within a column, values with c vs. d superscripts differ significantly (P<0.05, Chi- square test).
Experiment 3
Development of red deer and sika deer oocytes after IVM in medium
supplemented with oFF or cFF, IVF with 1 or 2.0 x 106 spermatozoa /mL and in vitro culture for 7 days is shown in Table 4.
Table 4. Development of red deer and sika deer embryos produced after IVM/IVF and 7 days of in vitro culture
Species FF source Sperm/mL Oocytes Cleaved Morulae Blastocysts (%) during IVM a (106) (n) (%)b (%)b Total Cleaved c
Red deer Ovine 1.0 53 47 11 Ovine 2.0 88 52 5 9 17
Caprine 2.0 143 57 3 10 17 Caprine 2.0 epid. 118 45 2 11 24
Sika deer Ovine 1.0 37 49 3
aWith ovine FSH.
bpercentages are based on the total number of oocytes subjected to IVM.
CValues within the column did not differ significantly (P>0.05, Chi-square test).
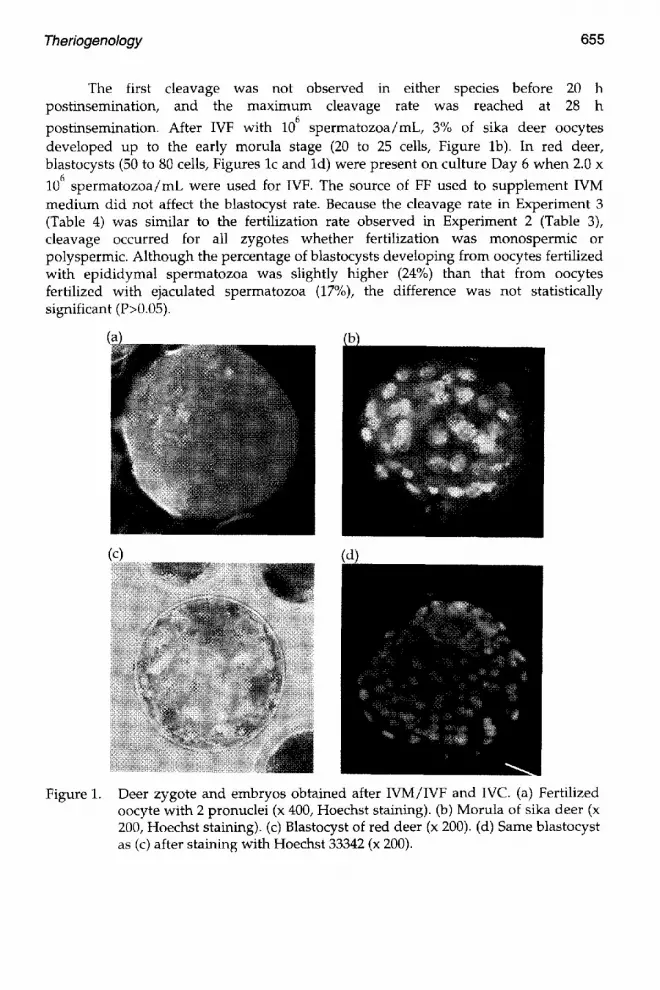
Theriogenology 655
The first cleavage was not observed in either species before 20 h postinsemination, and the maximum cleavage rate was reached at 28 h
postinsemination. After IVF w i t h 10 6 spermatozoa/mL, 3% of sika deer oocytes developed up to the early morula stage (20 to 25 cells, Figure lb). In red deer, blastocysts (50 to 80 cells, Figures lc and ld) were present on culture Day 6 when 2.0 x
106 spermatozoa/mL were used for IVF. The source of FF used to supplement IVM medium did not affect the blastocyst rate. Because the cleavage rate in Experiment 3 (Table 4) was similar to the fertilization rate observed in Experiment 2 (Table 3), cleavage occurred for all zygotes whether fertilization was monospermic or polyspermic. Although the percentage of blastocysts developing from oocytes fertilized with epididymal spermatozoa was slightly higher (24%) than that from oocytes fertilized with ejaculated spermatozoa (17%), the difference was not statistically significant (P>0.05).
(c) (d)
Figure 1. Deer zygote and embryos obtained after WM/IVF and IVC. (a) Fertilized oocyte with 2 pronuclei (x 400, Hoechst staining). (b) Morula of sika deer (x 200, Hoechst staining). (c) Blastocyst of red deer (x 200). (d) Same blastocyst as (c) after staining with Hoechst 33342 (x 200).

656 Theriogenology
DISCUSSION
Viable gametes can be recovered from red deer postmortem and on multiple occasions from individual sika deer. In both species, IVM was improved by the presence of oFSH and FF in the medium. Red deer oocytes fertilized by ejaculated or epididymal spermatozoa can develop in vitro to the blastocyst stage.
Ultrasound-guided transvaginal recovery of oocytes has been used previously in red deer, a large semidomestic species (5), but this approach was not suitable for sika deer because of their small size and susceptibility to stress. The oocyte recovery rate of sika deer by laparoscopy was similar to that reported in sheep, with or without eCG stimulation (27). However, the number of oocytes recovered per hind per procedure was lower in sika deer than in sheep because fewer follicles were aspirated. Although the number of oocytes recovered per ewe was increased after eCG stimulation and anti- eCG treatment (27), ovarian stimulation was not used in sika deer hinds, to minimize the number of variables affecting the in vitro development. Accordingly, too few sika deer oocytes were recovered for us to evaluate specific factors affecting in vitro development as was done with red deer oocytes.
In our IVM conditions, the supplementation with EGF was not as effective as previously shown in the axis deer (7). However, IVM rates were increased by the presence of oFSH and FF (about 80%), as previously reported in sheep (9, 28) and were higher than that obtained in deer oocytes by Fukui et al. (11).
The presence of SS allowed sperm capacitation to occur as previously shown by Berg et al. (4), although they used SOF medium, rather than TALP medium, for IVF.
When a sperm concentration of 1.0 x 106/mL was used for IVF, the incidence of polyspermy was lower in red deer than in sika deer. Possible reasons for this species specificity include 1) different mechanism for preventing polyspermic penetration (2) or 2) different sensitivity to freezing (19), with the result that at a given sperm concentration the actual concentration of viable spermatozoa was dissimilar between the 2 species. It is known that in vitro development is compromised in polyspermic embryos (15, 24); therefore, we suggest that the failure to obtain in vitro blastocyst development in sika deer was at least partially due to a higher incidence of polyspermy. Spermatozoa obtained by electroejaculation have been used previously for IVF of red
deer oocytes at lower concentrations (0.3 x 106 or 1.0 x 106/mL) (3, 6, 11). Our study provides further evidence that postmortem epididymal flushing can be a source of viable spermatozoa, as previously shown in other studies (3, 7, 12, 32).
Fertilization and developmental rates were not different after IVM of the oocytes with oFF or cFF. In the red deer, the increase of the sperm concentration had a positive effect on the subsequent embryo development as previously shown by Long et al. (20) in cattle. Interestingly, the presence of accessory spermatozoa during fertilization could produce a stimulatory effect on development (20) in this species. In the bovine species,

Theriogenology 657
the in vitro blastocyst rate is positively correlated with the interval from insemination to the first cleavage. Blastocyst formation is higher in zygotes that cleave before 28 h postinsemination (14). We found that first cleavage occurred at a similar interval postinsemination in deer zygotes, but the blastocyst rate was lower than reported in bovine species. A shorter interval from insemination to the first cleavage may be necessary in deer species to insure good embryo development. In previous studies in red deer (3, 11), fertilized oocytes developed to the 16-cell stage. The present study is the first to report IVP of sika deer embryos and IVP of blastocysts in red deer after using frozen/thawed epididymal spermatozoa. Under our IVC conditions, fertilized oocytes reached the morula (25 to 30 cells, both species) or the blastocyst stage (50 to 80 cells, red deer only). Since the medium used in this study (SOF) has been shown to be suitable for supporting in vitro development of deer embryos (6), we suggest that the low rate of blastocyst formation was due at least in part to a lack of developmental competence of the oocytes as previously reported in domestic species by Mermillod et al. (22). The genotype of the males used for IVF could also influence the blastocyst rate, as shown in the bovine species (10).
In conclusion, OPU is a useful technique to repeatedly recover immature oocytes from live sika deer. Red deer and sika deer embryos can be produced in vitro using similar IVP conditions. Blastocysts were produced in vitro in red deer after IVF with spermatozoa recovered from live and dead males. Similar methods could possibly be extended to closely related endangered subspecies, and we are doing further experiments to improve the development rate of the embryos.
REFERENCES
1. Asher GW, Fisher MW, Fennessy PF, Mackintosh CG, Jabbour HN, Morrow CJ. Oestrous synchronization, semen collection and artificial insemination of farmed red deer (.Cervus elaphus) and fallow deer (Dama dama). Anita Reprod Science 1993;33: 241-265.
2. Baccetti B, Burrini AG, Collodel G, Falugi C, Moretti E, Piomboni P. Localisation of two classes of acetylcholine receptor-like molecules in sperms of different animal species. Zygote 1995;3: 207-217.
3. Bainbridge DRJ, Catt SL, Evans G, Jabbour HN. Successful in vitro fertilization of in vivo matured oocytes aspirated laparoscopically from red deer hinds (Cervus elaphus). Theriogenology 1999;51: 891-898.
4. Berg DK, Asher GW, Pugh PA, Tervit HR, Thompson JG. Pregnancies following the transfer of in vitro matured and fertilized red deer (Cervus elaphus) oocytes. Theriogenology 1995;43:166 abstr.
5. Berg DK, Berg MC, Hull AD, Bertram MF. Development of transvaginal oocyte pick-up for farmed red deer CCervus elaphus). Theriogenology 2000;53:326 abstr.
6. Berg DK, Thompson JG, Pugh PA, Tervit HR, Asher GW. Successful in vitro culture of early cleavage stage embryos recovered from superovulated red deer (Cervus elaphus). Theriogenology 1995,44: 247-254.

658 Theriogenology
7. Chapman SA, Keller DL, Westhusin ME, Drew ML, Kraemer DC. In vitro production of axis deer (Axis axis) embryos, a preliminary study. Theriogenology 1999;51:280 abstr.
8. Chemineau P, Cogni6 Y, Gu6rin Y, Orgeur P, Vallet JC. Collection and preservation of spermatozoa. In: Training Manual on Artificial Insemination in Sheep and Goats. Rome: FAO Animal Production and Health, 1991;83: 115-130.
9. Cogni6 Y, Poulin N, Pisselet C, Monniaux D. Effect of atresia on the ability of follicular fluid to support developmental competence of sheep oocytes in vitro. Theriogenology 1995;43:188 abstr.
10. Comizzoli P, Marquant-Le Guienne B, Heyman Y, Renard JP. Onset of the S-phase is determined by a paternal effect during the Gl-phase in bovine zygotes. Biol Reprod 2000;62: 1677-1684.
11. Fukui Y, McGowan LT, James RW, Asher GW, Tervit HR. Effects of culture duration and time of gonadotropin addition on in vitro maturation and fertilization of red deer (Cervus elaphus) oocytes. Theriogenology 1991;35: 499- 512.
12. Garde JJ, Ortiz N, Garcia AJ, Gallego L, Landete-Castillejos T, Lopez A. Postmortem assessment of sperm characteristics of the red deer during the breeding season. Arch Androl 1998;41: 195-202.
13. Goombridge B. IUCN Red List of Threatened Animals. Gland, Switzerland: Union Internationale pour la Conservation de la Nature. 1994.
14. Grisart B, Massip A, Dessy F. Cinematographic analysis of bovine embryo development in serum-free oviduct-conditioned medium. J Reprod Fertil 1994;101:257-264.
15. Han YM, Abeydeera LR, Kim JH, Moon HB, Cabot RA, Day BN, Prather RS. Growth retardation of inner cell mass cells in polyspermic porcine embryos produced in vitro. Biol Reprod 1999;60: 1110-1113.
16. Jabbour HN, Hayssen V, Bruford W. Conservation of deer: contributions from molecular biology, evolutionary ecology, and reproductive physiology. J Zool Lond 1997;243: 461-484.
17. Johnston LA, Parrish JJ, Monson R, Leibfried-Rutledge L, Susko-Parrish JL, Simmons LG. Oocyte maturation, fertilization and embryo development in vitro and in vivo in the gaur (Bos gaurus). J Reprod Fertil 1994;100: 131-136.
18. Krogenaes A, Ropstad E, Thomasssen R, Pedersen O, Forsberg M. In vitro maturation and fertilization of oocytes from Norwegian semi-domestic reindeer (Rangifer tarandus). Theriogenology 1994;41: 371-377.
19. Leibo SP. Sperm freezing - species specific? Fertil Steril 1999;72: 747-748. 20. Long CR, Damiani P, Pinto-Correia C, MacLean RA, Duby RT, Robl JM.
Morphology and subsequent development in culture of bovine oocytes matured in vitro under various conditions of fertilization. J Reprod Fertil 1994;102: 361-369.
21. Mauget R, Legendre X, Comizzoli P, Mermillod P. Assisted reproductive technology in sika deer: a programme to preserve endangered deer subspecies. In: Zomborsky Z (ed), Advances in deer biology. Kasposvar: Proc 4 th Int Deer Biol Cong, 1998;185-186.

Theriogenology 659
22. Mermillod P, Oussaid B, Cogni6 Y. Aspects of follicular and oocyte maturation that affect the developmental potential of embryos. J Reprod Fertil Suppl 1999;54: 449-460.
23. Monfort SL, Asher GW, Wildt DE, Wood TC, Schiewe MC, Williamson LR, Bush M, Rall WF. Successful intrauterine insemination of Eld's deer (Cervus eldi thamin) with frozen-thawed spermatozoa. J Reprod Fertil 1993;99: 459-465.
24. Niwa K. Effectiveness of in vitro maturation and in vitro fertilization techniques in pigs. J Reprod Fertil Suppl 1993;48: 49-59.
25. Palta P, Chauhan MS. Laboratory production of buffalo (Bubalus bubalis) embryos. Reprod Fertil Dev 1998;10: 379-391.
26. Parrish JJ, Susko-Parrish JL, Leibfried ML, Critse ES, Eyestone WH, First NL. Bovine in vitro fertilization with frozen-thawed semen. Theriogenology 1986; 25:591-600.
27. Stangl M, Ki~hholzer B, Besenfelder U, Brem G. Repeated endoscopic ovum pick- up in sheep. Theriogenology 1999;52: 709-716.
28. Sun FJ, Holm P, Irvine B, Seamark RF. Effect of sheep and human follicular fluid on the maturation of sheep oocytes in vitro. Theriogenology 1994;41: 981-988.
29. Wildt DE, Rall WF, Critser JK, Monfort SL, Seal US. Genome resource banks: living collections for biodiversity conservation. Bioscience 1997;47:
30. Willard ST, Flores-Foxworth G, Chapman S, Drew ML, Hughes DM, Neuendorff DA, Randel RD. Hybridization between wapiti (Cervus elephus manitobensis) and sika deer (Cervus nippon): a comparison of two artificial insemination techniques. J Zoo Wildl Med 1998;29: 295-299.
31. Willard ST, Hughes Jr DM, Bringans M, Sasser RG, White DR, Jaques JT, Godfrey RW, Welsh Jr, Randel RD. Artificial insemination, hybridization and pregnancy detection in sika deer (Cervus nippon). Theriogenology 1996;46: 779-789.
32. Zomborszky Z, Zubor T, Toth J, Horn P. Sperm collection from shot red deer stags (Cervus elaphus) and the utilization of sperm frozen subsequently thawed. Acta Vet Hung 1999;47: 263-270.





























