Embed Size (px)
Citation preview

TGF-β1 is a negative regulator of lymphatic regeneration
1
TGF-β1 is a negative regulator of lymphatic regeneration during wound
repair
Running head: TGF-B is a negative regulator of lymphatic regeneration
Key words: Lymphangiogenesis, scar, lymphedema, VEGF-C, TGF-β1, excisional wound healing, myofibroblast, fibrosis
Nicholas W. Clavin, Tomer Avraham, John Fernandez, Sanjay V. Daluvoy, Marc A. Soares, Arif Chaudhry, and Babak J. Mehrara
From: The Division of Plastic and Reconstructive Surgery and The Department of Surgery, Memorial Sloan-Kettering Cancer Center, New York, NY 10021
Correspondence: Babak J. Mehrara, MD FACS
1275 York Avenue Room MRI 1005 New York, NY 10065 212-639-8639 212-717-3677 (Fax) [email protected]
Articles in PresS. Am J Physiol Heart Circ Physiol (October 10, 2008). doi:10.1152/ajpheart.00879.2008
Copyright © 2008 by the American Physiological Society.

TGF-β1 is a negative regulator of lymphatic regeneration
2
Disclosure None of the authors have any commercial associations or financial relationships that
would create a conflict of interest with the work presented in this article.

TGF-β1 is a negative regulator of lymphatic regeneration
3
Abstract
Introduction: Although clinical studies have identified scarring/fibrosis as significant risk
factors for lymphedema, the mechanisms by which lymphatic repair is impaired remain
unknown. TGF-β1 is a critical regulator of tissue fibrosis/scarring and may therefore also play a
role in the regulation of lymphatic regeneration. The purpose of this study was therefore to
assess the role of TGF-β1 on scarring/fibrosis and lymphatic regeneration in a mouse tail model.
Methods: Acute lymphedema was induced in mouse tails by full thickness skin excision and
lymphatic ligation. TGF-β1 expression and scarring were modulated by repairing wounds with
or without topical collagen gel. Lymphatic function and histologic analyses were performed at
various time points. Finally, the effects of TGF-β1 on lymphatic endothelial cell (LEC) in vitro
were evaluated. Results: Wound repair with collagen gel significantly reduced the expression of
TGF-β1, decreased scarring/fibrosis, and significantly accelerated lymphatic regeneration. The
addition of recombinant TGF-β1 to the collagen gel negated these effects. Improved lymphatic
regeneration secondary to TGF-β1 inhibition was associated with increased infiltration and
proliferation of LECs and macrophages. TGF-β1 caused a dose-dependent significant decrease
in cellular proliferation and tubule formation of isolated LECs without changes in the expression
of VEGF-C/D. Finally, increased expression of TGF-β1 during wound repair resulted in
lymphatic fibrosis and co-expression of α-SMA and LYVE-1 in regenerated lymphatics.
Conclusions: Inhibition of TGF-β1 expression significantly accelerates lymphatic regeneration
during wound healing. Increased TGF-β1 expression inhibits LEC proliferation and function,
and promotes lymphatic fibrosis. These findings imply that clinical interventions that diminish
TGF-β1 expression may be useful in promoting more rapid lymphatic regeneration.
Key words: Lymphangiogenesis, scar, lymphedema, VEGF-C, excisional wound healing, fibrosis

TGF-β1 is a negative regulator of lymphatic regeneration
4
Introduction
Lymphedema is a debilitating disorder that occurs commonly after axillary lymph node
dissection (ALND) for breast cancer. Approximately 30% of the two million breast cancer
survivors in the US have some degree of lymphedema.(49) Despite the profound impact of
lymphedema on quality of life, very few treatment options exist.(52) There is currently no
medical or surgical cure. Understanding the molecular mechanisms responsible for abnormal
lymphatic regeneration and subsequent lymphedema is an important step in the development of
targeted therapeutic and preventative strategies.
Recent studies have elucidated several key molecular mechanisms that regulate
lymphangiogenesis in wound healing, inflammation, and in tumors. These studies have
demonstrated critical roles for growth factors such as vascular endothelial growth factor C and D
(VEGF-C, VEGF-D) as well as inflammatory cells such as macrophages.(38, 41) Coordinated
expression of these mediators results in the formation of new lymphatic vessels and lymphatic
repair. Although these mechanisms are well described, the defects that prevent normal
lymphatic regeneration and result in subsequent lymphedema remain largely unknown.
Clinical studies have identified extensive surgery, infections, and radiation therapy as
important risk factors for the development of lymphedema. (17, 39) The combination of
radiation and surgery increases the risk of lymphedema 4-8 fold.(1, 53) The common
denominator in these risk factors is soft tissue fibrosis and scarring, suggesting that increased
scarring is associated with abnormal lymphatic regeneration. This contention is supported by the
finding that the induction of lymphedema in animal models requires extensive scarring, often
including full thickness skin resection in combination with radiation therapy.(46)

TGF-β1 is a negative regulator of lymphatic regeneration
5
The molecular mechanisms responsible for soft tissue fibrosis have been the topic of
intense investigation. These studies have shown that TGF-β1 is an important regulator of
scarring and fibrosis both during wound repair as well as in abnormal organ fibrosis.(2, 4, 12, 40,
51) For example, TGF- β1 expression is increased in abnormal wound healing such as keloids
or hypertrophic scars as well as in liver, lung, and renal fibrosis.(16, 20, 21) Similarly, radiation
therapy results in chronically increased TGF-β1 expression that can last several years after
injury.(23) Inhibition of TGF-β1 signaling decreases scarring during wound repair and reduces
organ fibrosis in the lung, liver, and kidney.(9, 16, 19) Down-regulation of TGF-β1 expression
after radiation therapy also decreases soft tissue fibrosis.(13, 54) Thus, although it is known that
fibrosis is associated with lymphedema and up-regulation of TGF-β1 expression, the effects of
this growth factor on lymphatic regeneration during wound repair remain essentially unknown.
A recent study has shown that TGF-β1 has direct inhibitory effects on isolated lymphatic
endothelial cells and inhibits lymphangiogenesis in tumors.(30) We therefore hypothesized that
inhibition of scarring would decrease TGF-β1 expression and improve lymphatic regeneration.
We demonstrate that decreasing TGF-β1 expression during wound repair using a topical collagen
gel treatment accelerates lymphatic regeneration and is associated with an earlier accumulation
of lymphatic endothelial cells (LECs), reduced myofibroblast proliferation, and diminished
capillary lymphatic fibrosis. In contrast, increased TGF-β1 expression during abnormal wound
repair or with addition of supplemental growth factor significantly delayed lymphatic
regeneration. Furthermore, we show that TGF-β1 has direct effects on isolated LECs by
inhibiting proliferation and tubule formation without altering VEGF-C or VEGF-D expression.
Collectively, our results suggest that inhibition of TGF-β1 signaling during wound repair could

TGF-β1 is a negative regulator of lymphatic regeneration
6
be a viable means of improving lymphatic regeneration and decrease the incidence of
postoperative lymphedema.
Materials and Methods
Mouse Tail Excision Model and Surgical Preparation
Female 6-8 week old C57B6 mice (The Jackson Laboratory, Bar Harbor, ME) were used
for all experiments after approval by Resource Animal Research Center at Memorial Sloan
Kettering Cancer Center. For creation of acute post-surgical lymphedema, a 2mm full thickness
skin and subcutaneous excision (thereby removing the superficial lymphatic network) was
performed at a distance of 15mm from the base of the tail as previously described.(6) In
addition, collecting lymphatics running adjacent to the lateral tail veins were identified and
ligated using a surgical microscope (Leica, StereoZoom SZ-4, Wetzlar, Germany). The mice
were divided into two groups. In group 1 (+ collagen gel; n= 16) the skin defect was filled with
0.3% type I rat collagen gel (BD Biosciences, Bedford, MA) and the wound was covered with
Bacitracin Zinc Ointment USP (Fougera® Melville, NY), Xeroform Petrolatum Dressing
(Cardinal Health, Dublin, OH) and Tegaderm™ (3M, St. Paul, MN) to prevent desiccation.
Based on previous experiments we hypothesized that this treatment would decrease scar
formation and therefore inhibit TGF-β1 expression.(8) Animals in group 2 (- collagen gel; n=16)
were treated surgically, and had their wounds covered only with ointment and dressing to
prevent desiccation. In order to further delineate the role of TGF-β1 during lymphatic repair,
additional animals underwent identical procedures with skin and lymphatic ablation and wound
coverage with 0.3% type I rat collagen impregnated with a physiologic dose of recombinant
TGF-β1 (50ng), a physiologic dose (+collagen gel/+TGF-β1; n=16). (45) The recombinant

TGF-β1 is a negative regulator of lymphatic regeneration
7
TGF-β1 used was mature human TGF-β1which has cross reactivity with mouse and shares 99%
amino acid identity with mouse TGF-β1 (R&D Systems, Minneapolis, MN).
Specimen Preparation and Histology
Ten millimeter tail sections centered on the repair site were harvested after 1, 2, 4, and 6
weeks, fixed in 4% paraformaldehyde, decalcified, embedded in paraffin and sectioned at 5
microns. Histochemical staining for scar tissue was performed using picro-sirius red (Direct Red
80, Sigma St. Louis, MO) and hematoxylin and eosin stains as previously described.(12)
Specimens were examined using polarized light microscopy (Leica TCS AOBSSP2) and the scar
index was calculated as previously described.(12) The scar index is based on the fact that
normal skin is characterized by thin, randomly oriented collagen fibers demonstrating yellow-
green birefringence while scarring is associated with deposition of thick parallel collagen
bundles with orange-red birefringence.(10) To calculate the scar index, representative 70,000
µm2 dermal images within and distal to the wound bed of specimens harvested after 4 or 6 weeks
were analyzed using Metamorph™ Offline software (Molecular Devices Corporation,
Sunnyvale, CA) and the ratio of the number of orange-red pixels to the number of yellow-green
pixels was calculated in three high power fields.(12)
Tail volume measurements and lymphoscintigraphy
To calculate the degree of post procedural acute lymphedema, tail volumes distal to the
site of lymphatic excision were calculated 1, 2, 4, and 6 weeks post procedure (n=4/group) using
the truncated cone formula and compared to baseline.(47)
Lymphoscintigraphy was performed using unfiltered 99mTc-sulfur colloid (100nm particle
size; 400-800µCi in ~50 µl) injected intradermally at the tip of the tail. Dynamic planar gamma
camera images were acquired for 2.5 hours post injection using the X-SPECT™ (Gamma

TGF-β1 is a negative regulator of lymphatic regeneration
8
Medica, Northridge, CA) fitted with low-energy parallel-hole collimators. The resulting images
were imported into ASIPro™ (CTI Molecular Imaging, Knoxville, Tennessee) for analysis.
Region-of-interest (ROI) analysis was performed to derive the time-activity data for the injection
site and lymph nodes at the base of the tail. Decay adjusted isotope uptake in the lymph node
basin of the tail was determined.
Immunofluorescence and Immunohistochemistry
Immunohistochemical and immunofluorescent staining were performed as previously
described.(26) Immunofluorescence was used to identify lymphatic vessel endothelial receptor 1
(LYVE-1; Rabbit polyclonal, Abcam, Cambridge, MA); proliferating cell nuclear antigen
(PCNA; Rabbit polyclonal, Abcam), and alpha smooth muscle actin (SMA; Rabbit Polyclonal,
Abcam). Fluorescence detection was performed with fluorescein (R&D Systems, Minneapolis,
MN) or Cy3 Zymed (Invitrogen Molecular Probes, Carlsbad, CA) and visualized using a
fluorescent microscope (Leica TCS).
Immunohistochemistry was used to label TGF-β1 (Rabbit polyclonal, Santa Cruz
Biotechnology, Santa Cruz, CA) and F4/80 (Acris, Hiddenhausen Germany). Secondary antibody
was from VECTASTAIN® ABC Kit (Vector, Burlingame, CA). Images were obtained after
DAB staining using bright-field microscopy (Leica TCS). For all immunolocalization
experiments, negative controls included tissues treated with no primary antibody or no secondary
antibody.
Quantitation of TGF-β1 Expression
Relative expression of TGF-β1 was assessed using a visual analog scale after
immunostatining (0=no staining, 3=intense staining) by two blinded reviewers. A minimum of

TGF-β1 is a negative regulator of lymphatic regeneration
9
three (3) high powered fields (100X magnification) from the distal margins of the wound were
analyzed for each animal (n=4) at each time point. Average staining score was calculated.
In order to confirm the changes in expression of TGF-β1 in group 1 (+collagen) and
group 2 (- collagen) animals, tail excision and wound repairs were performed as described above
in 3 animals per group. The wounds were then harvested 2 weeks postop (the time point
corresponding to the highest level of TGF-β1 expression by immunohistochemistry) and
quantitative PCR was performed using our previously published techniques.(43). Briefly, the
wounded section and an area spanning approximately 1mm proximal and 1mm distal to the
wound were harvested, rapidly frozen in liquid nitrogen, homogenized in Trizol Reagent
(Invitrogen Molecular Probes, Carlsbad, CA), and total cellular RNA was harvested using the
manufacturer’s protocol. RNA quantity and quality of the purified samples was analyzed using a
NanoDrop ND 1000 Spectrophotometer. Reverse transcription was performed using the
TaqMan Reverese Trasncription reagents Kit (Applied Biosystems, Foster City, CA) using 1 μg
RNA. qRT-PCR was then performed using the LightCycler thermocycler and the TaqMan
Universal Mastermix (Roche Diagnostics Corp., Indianapolis, IN), according to manufacturer’s
directions. A standard curve was prepared using serially diluted control samples and was plotted
against cycle numbers obtained at log-linear phase of the reaction. Expression of TGF-β1
(Applied Biosystems) was normalized to 18S RNA expression measured concurrently.
Experiments were performed in triplicate.
Total cellular protein was isolated from the region surrounding the wounds at 2 weeks
following surgery. Briefly, the wounded section and an area spanning approximately 1mm
proximal and 1mm distal to the wound were harvested, rapidly frozen in liquid nitrogen,
homogenized in Trizol Reagent, and total cellular protein was harvested using the

TGF-β1 is a negative regulator of lymphatic regeneration
10
manufacturer’s protocol. Total protein was quantified using the Bradford method. Western blot
analysis for TGF-β1 was performed using a polyclonal antibody mouse TGF-β1 (Santa Cruz
Antibodies) and normalized to actin levels. Immunoreactivity was determined using the ECL
chemiluminescence detection system (Amersham, Arlington Heights, Il).
Lymphatic endothelial cell (LEC) quantification and identification of proliferating cells
LECs in the distal portions of the wound were identified with LYVE-1 antibodies and
counted by 2 blinded reviewers using confocal laser florescence microscopy in 3 random high-
powered fields (200x) for 4 animals per group at each time point. Individual cells which had
both DAPI stained nuclei and LYVE-1 stained cell membranes were identified and counted. To
identify proliferating LECs, sections were double stained with PCNA and LYVE-1. Individual
proliferating cells were identified using confocal laser microscopy to identify the nuclear staining
of PCNA and membrane associated LYVE-1. Cells were quantified in 3 random high powered
fields (200x) at the distal margin of the wound by 2 blinded observers for each animal at the 2
week time point.
Cell Culture and Reagents
Human Lymphatic Endothelial Cells (HLEC) were obtained from PromoCell
(Heidelberg, Germany) and cultured in ECGM-MV containing 0.4% ECGS/H, 5% FCS, EGF
(10ng/ml), hydrocortisone (1μg/ml), and Penicillin-Streptomycin (50U/ml) (Invitrogen) and
passaged every 48 hours. LEC morphology was confirmed with immunofluorescent staining for
LYVE-1 and prox-1 (not shown). Early passage cells (<10) were used for all experiments.
Proliferation and Matrigel Tubule Formation Assay
LECs were plated at a density of 2 x 103 cells in 96-well plates and incubated with 0, 1,
or 10ng/ml of TGF-β1. Relative cell number was measured using the CellTiter96 Aqueous

TGF-β1 is a negative regulator of lymphatic regeneration
11
Assay (Promega, Madison, WI) according to manufacturer’s instructions. The mitochondrial
conversion of MTS to formazan (directly proportional to the number of living cells in the
culture) was recorded at 490nm using a plate reader (EL 312e Biokinetics reader, Biotek
Instruments, Winooski, VT). For lymphatic tubule formation, LECs were cultured for 3 days in
media supplemented with 0, 1, or 10ng/ml of recombinant human TGF-β1. Cells were
subsequently trypsinized and tubule formation was observed 24 hours after plating 1x103
cells/well in 96-well plates coated with growth factor reduced matrigel matrix (BD Biosciences).
Images were acquired using an inverted brightfield microscope (Nikon Eclipse TS-100 Tokyo,
Japan) and digitally photographed (Nikon Coolpix p5100). All experiments were performed in
quadruplicate.
Human VEGFC/D ELISA
Sandwich ELISA kits from R&D systems were used to determine the levels of expressed
VEGFC and D in cultured human LECs 24 hours after exposure to varying concentrations of
TGF-β1 (0, 1, 10ng/ml). Optical density was measured at 450nm and corrected with OD
measurement at 570nm. The values obtained were reported in picograms/mL using a standard
curve.
Statistical analysis
Multi-group comparison was performed using one-way ANOVA with the Tukey-Kramer
post-hoc test. Students T-test was used for analyzing differences between 2 groups. Data are
presented as mean ± standard deviation or standard error as noted with p<0.05 considered
significant.
Results

TGF-β1 is a negative regulator of lymphatic regeneration
12
Treatment of mouse tail skin and lymphatic excision with topical collagen gel inhibits TGF-β1
expression and results in decreased fibrosis.
All mouse tails were grossly stable after surgery without evidence of arterial
insufficiency. All mice demonstrated a variable period of acute lymphedema without any wound
complications.
Collected tissue specimens were assayed for TGF- β1 expression using
immunohistochemistry. Wounds collected from group 1 animals (+collagen gel) demonstrated
little extracellular matrix (ECM) staining for TGF-β1 although spindle shaped cells consistent
with fibroblasts present deep to and adjacent to the wound stained positively. Furthermore, the
low level TGF-β1 expression noted in the early time points of the group 1 wounds all but
disappeared by the later time points. In contrast, wounds from group 2 animals (-collagen gel)
demonstrated a marked increase in TGF-β1 expression in the ECM, in fibroblasts surrounding
the excision sites, and in spindle shaped cells within the granulation tissue. The intensity of
TGF-β1 staining was highest in the early time points (1 and 2 weeks) in this group and persisted
through the later time points (Figure 1A,B). Staining for TGF-β1 in group 2 animals (-collagen
gel) was demonstrably more intense than group 1 animals (+collagen) at all time points
evaluated. In order to further demonstrate decreased TGF-β1 expression in group 1 animals
quantitative PC and western blot analysis were performed 2 weeks after surgery. These analyses
demonstrated significant decreased expression of TGF-β1 as a result of collagen gel treatment
(Figure 1C,D).
When we evaluated the wound sections histologically 1 week after wounding, we noted
inflammation, dermal hypercellularity, spongiosis, and dilated lymphatic vessels distal to the
excision site in all 3 treatment groups (not shown). By 2 weeks, animals in group 2 (-collagen

TGF-β1 is a negative regulator of lymphatic regeneration
13
gel) continued to exhibit hypercellularity and distal lymphatic dilatation, while group 1 animals
(+collagen gel) showed decreased cellularity, inflammation, and resolution of distal lymphedema
(not shown). The addition of 50ng TGF-β1 to the collagen gel (+collagen/+TGF-β1) resulted in
a histological appearance that was very similar to animals treated without collagen gel and
completely reversed the beneficial effects of collagen gel dressing on wound repair. At later
time points evaluated, group 1 (+collagen gel) animals continued to heal rapidly demonstrating
decreased subdermal thickness and collapsed lymphatics, reaching near normal histology by six
weeks (Figure 2A.). In contrast, animals in group 2 (-collagen gel) and those treated with
supplemental TGF-β1 remained hypercellular with a thickened dermal/subdermal tissues and
persistent edema distally even at the 6 week time point. Thus, topical collagen treatment
inhibited TGF-β1 expression and was associated with decreased inflammation and cellularity. In
contrast, exogenous application of TGF-β1 to the collagen gel resulted in increased inflammation
and recapitulated the histological characteristics of the animals treated without collagen gel.
Decreased inflammation, cellularity, and TGF-β1 expression in group 1 animals
(+collagen gel) translated to decreased scarring as demonstrated by picro-sirius red staining. The
collagen gel treated animals had a more random (i.e. normal) organization of green-yellow
birefringent collagen fibers and nearly 3-fold lower scar index than the excision without collagen
gel group and nearly 6-fold lower index than the collagen gel supplemented with TGF-β1 group
4 and 6 weeks after surgery (p<0.01; Figure 2B,C).
Inhibition of TGF-β1 expression and scarring is associated with increased functional
lymphatic transport and decreased acute lymphedema
In order to evaluate the effect of variable scarring on lymphatic regeneration, we
quantified 99mTc-sulfur colloid lymphatic transport using lymphoscintigraphy by calculating

TGF-β1 is a negative regulator of lymphatic regeneration
14
decay corrected base of tail lymph node uptake relative to initial injection at the distal tail tip
(Figure 3 A,B.). Although uptake was limited in all groups 1 week after surgery, it was
significantly higher in group 1 animals (+collagen gel; nearly 6 fold increase; p<0.05). Similar
trends were noted after 2 and 4 weeks with group 1 animals demonstrating more than threefold
higher technetium uptake as compared with group 2 animals (-collagen gel; p< 0.01 at each time
point). Interestingly, at the 6 week time point the radio-nucleotide uptake in group 1 animals
(+collagen gel) returned to baseline levels but remained markedly abnormal in the animals
treated without collagen gel and in those in which collagen gel was supplemented with TGF-β1.
This finding demonstrates that increased TGF-β1 expression during wound repair inhibits
lymphatic regeneration.
Increased lymphatic transport noted on lymphoscintigraphy was associated with
decreased tail volumes and more rapid resolution of acute lymphedema in group 1 (+collagen
gel) animals as compared with group 2 (-collagen gel) and in animals treated with recombinant
TGF-β1. Even 2 weeks after surgery, the increase in tail volume of the group 1 animals was
nearly 4 fold less than that observed in group 2 animals as well as animals treated with
recombinant TGF-β1 (p<0.05, Figure 3C). By 4 weeks postoperatively, tail volumes in group 1
returned to baseline where as group 2 animals still had a 40% increase in tail volume. Similarly,
the addition of TGF-β1 to the wound reversed the beneficial effects of collagen gel, and resulted
in change in tail volumes almost indistinguishable from group 2 animals. This trend was
maintained at the 6 week time point with only minimal improvement in tail volumes in group 2
and the supplemental TGF-β1 group. In addition, histologic sections of wounds and the distal
portions of the tails obtained 6 weeks after treatment with TGF-β1 demonstrated changes
consistent with chronic lymphedema (i.e. fat deposition, thickened dermis/subdermis, and

TGF-β1 is a negative regulator of lymphatic regeneration
15
subdermal fibrosis; Figure 4). Thus, inhibition of TGF-β1 expression during wound repair is
associated with improved lymphatic function and decreased acute lymphedema. In contrast,
increased TGF-β1 expression either endogenously (i.e. group 2) or as a result of TGF-β1
supplementation is associated with delayed lymphatic regeneration and prolonged acute
lymphedema.
Increased expression of TGF- β1 during wound repair is associated with delayed recruitment
and decreased proliferation of lymphatic endothelial cells
In order to evaluate the cellular mechanisms by which increased TGF-β1 expression
during wound repair can inhibit lymphatic regeneration, we quantified the number and
proliferation of lymphatic endothelial cells (LEC) present in the wound. This was based on
previous work by Boardman et al demonstrating that the recruitment of LECs beginning distally
and migrating proximally is crucial for lymphatic regeneration in the mouse tail model. (35)
Thus, LECs were quantified and microscopic lymphatic regeneration in the distal portions of the
wound was analyzed using LYVE-1 immunofluorescence (Figure 5A). At all time points
evaluated, animals in group 1 (+collagen gel) had more LECs present (3-5x increase) in the distal
portion of the wound than group 2 (-collagen gel) or animals treated with supplemental TGF-β1
(p<0.01 at all time points). There was no statistical difference between group 2 and supplemental
TGF-β1 treated animals in LEC number.
The reduced number of LECs in group 2 wounds and in animals treated with
supplemental TGF-β1 was due in part to inhibition of LEC proliferation (Figure 5B,C). When
the number of proliferating LECs was calculated by co-localization of PCNA and LYVE-1, we
noted that animals in group 1 (+collagen gel) had nearly 6 times more proliferating LECs in the
distal portion of the wound as compared to group 2 (-collagen gel) and animals treated with

TGF-β1 is a negative regulator of lymphatic regeneration
16
supplemental TGF- β1 (p<0.01). There was no statistical difference in the number of
proliferating LECs between group 2 and supplemental TGF-β1 treated animals.
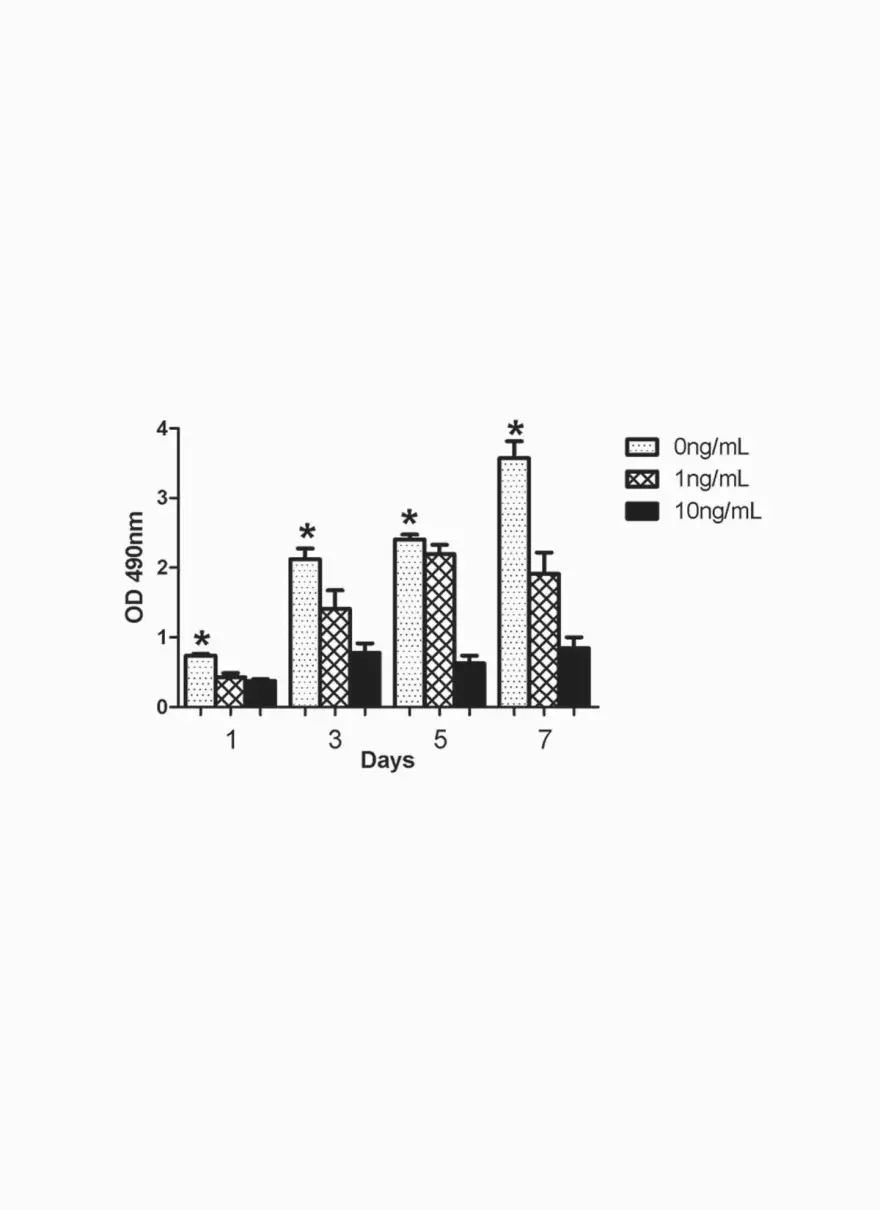
To evaluate the effects of TGF-β1 on isolated LECs, purified LECs were cultured with
varying amounts of TGF-β1 and cellular proliferation was assessed (Figure 5D). This analysis
demonstrated a dose dependent and statistically significant decrease in LEC proliferation in vitro
thus suggesting that increased TGF-β1 expression during abnormal wound repair may directly
inhibit lymphatic regeneration by inhibiting LEC proliferation.
Increased TGF-β1 during wound repair is associated with abnormal lymphatic architecture
and dilated lymphatics.
LYVE-1 staining was used to evaluate the architecture of the newly formed lymphatics in
each group (Figure 6 A). As expected, in group 1 animals (+ collagen gel), lymphatic capillary
formation was noted at the earliest time points with infiltration and coalescence of LECs. By 2
weeks, the newly formed lymphatics demonstrated a capillary and cylindrical architecture with
mild dilation noted in some vessels. In contrast, group 2 animals (- collagen gel) and animals
treated with supplemental TGF-β1 demonstrated very little capillary lymphatic regeneration at
this time point. The few lymphatics that had formed were markedly abnormal, dilated, and
ectatic in appearance. Similar findings were noted in these groups at the 4 and 6 week time
points evaluated with only an occasional normal, collapsed cylindrical capillary lymphatic
present. In contrast, the capillary architecture of group 1 (+collagen gel) animals was similar to
normal skin by the 6 week time point with cylindrical, small collapsed capillary lymphatics.
In order to evaluate the effect of TGF-β1 on LEC tubule formation and function, we
cultured isolated LECs in varying doses of TGF-β1 and evaluated tubule formation using a
matrigel assay (27). LECs cultured in media supplemented with TGF-β1 demonstrated a

TGF-β1 is a negative regulator of lymphatic regeneration
17
significant and dose-dependent decrease in the potential for lymphatic tubule formation (Figure
6B). In the absence of supplemental TGF-β1, lymphatic tubule formation occurred rapidly and
multiple small tubules were observed after 24 and 48 hours. In contrast, large, ectatic and dilated
lymphatics similar to those noted in group 2 (-collagen gel) and supplemental TGF-β1 treated
animals were noted in the low dose TGF-β1 group. Treatment with high dose TGF-β1,
(10ng/ml) all but abolished tubule formation in vitro. It should be noted that the high dose of
TGF-β1 used in our study is still thought to be within the physiologic range of this growth factor
in vitro.(45) Thus, increased TGF- β1 during abnormal wound repair significantly attenuates
LEC function and tubule formation and may contribute to delayed lymphatic regeneration and
increased acute lymphedema.
In order to determine whether increased TGF-β1 causes alterations in LEC proliferation
and tubule formation by changing VEGF-C or D expression, we analyzed the expression of these
growth factors in isolated LECs exposed to various doses of TGF-β1 (Figure 6 C,D).
Interestingly, TGF-β1 did not alter the expression of VEGF-C or D significantly in vitro. This
finding is somewhat surprising given that we and others have shown that TGF-β1 is a potent
regulator of VEGF-A expression (32, 37). Thus, it is likely that the differences we noted in
LEC proliferation or tubule formation are not due to changes in VEGF-C or VEGF-D expression
by LECs. This conclusion is supported by the finding that little difference in VEGF-C expression
was noted between group 1 (+collagen gel) and group 2 (-collagen gel) animal wounds as
assessed by immunohistochemistry in vivo (not shown). It is possible; however, that TGF-β1
can interact indirectly with VEGF-C/D through alterations in the expression of VEGF-receptors
or other mechanisms.

TGF-β1 is a negative regulator of lymphatic regeneration
18
Increased TGFβ1 expression during wound repair is associated with increased
myofibroblastic infiltration and lymphatic fibrosis
TGF-β1 is a known inducer of myofibroblast proliferation and differentiation (28). In
addition, recent studies have demonstrated that endothelial-mesenchymal (ie. fibroblast)
differentiation occurs in a number of scenarios (e.g. peri-tumoral fibrosis, renal fibrosis) and that
this effect is mediated by TGF-β1 (29, 31). In these cases, direct differentiation of microvascular
endothelial cells to fibroblastic phenotype (i.e. expression of fibroblast markers such as alpha-
smooth muscle actin (SMA) leads to tissue fibrosis. Lymphatic fibrosis has been shown in
anatomic and histologic studies of patients with lymphedema.(48) Therefore, we analyzed alpha
SMA expression in wound sections in order to evaluate the possibility of LEC-fibroblast
differentiation as a molecular mechanism of lymphatic fibrosis resulting from increased TGF-β1
expression during abnormal wound. This analysis demonstrated that abnormal wound repair as
noted in group 2 (-collagen gel) was associated with a marked increase in the number of
myofibroblasts in the wound bed (Figure 7A). In addition, we noted that numerous LECs
present in the capillary lymphatics (ie LYVE-1 positive) co-expressed alpha-SMA at the later
time points in group 2. Wounds treated with collagen gel supplemented with TGF-β1
demonstrated an even greater degree of double staining for LYVE-1 and alpha-SMA (Figure
7B,C). Consistent with previous histologic studies of lymphatic fibrosis, the capillary lymphatics
that co-expressed lymphatic and fibroblast markers appeared to be thickened and fibrotic
indicating that endothelial-fibroblast differentiation may be present. In contrast, wounds
harvested from group 1 animals (+collagen gel) demonstrated normal lymphatic architecture and
no evidence of lymphatic fibrosis or co-expression of LYVE-1 and α-SMA. It is important to
note that capillary lymphatics, unlike collecting lymphatics, are not associated with pericytes, a

TGF-β1 is a negative regulator of lymphatic regeneration
19
cell type known to express alpha-SMA. (50) Therefore, expression of this fibroblastic marker by
the LECs lining the normally thin walled capillary lymphatics vessels is an abnormal finding.
These observations suggest that increased TGF-β1 expression during abnormal wound repair
may promote lymphatic fibrosis possibly as a result of direct LEC to fibroblast
transdifferentiation. This finding is important and may represent a molecular mechanism for
lymphatic fibrosis observed clinically in patients with lymphedema. In addition, fibrosis may be
responsible for lymphatic dysfunction by physically preventing dilation and collapse of
lymphatic channels.
Decreased scarring is associated with increased infiltration of F4/80 positive macrophages
Since macrophages are known to be important regulators of lymphangiogenesis,
differences in macrophage migration were evaluated using F4/80 immunohistochemistry.
Wounds in group 1 animals (+collagen gel) demonstrated early infiltration of macrophages, with
most of the cells localized to the distal margin of the wound in the subdermal tissues. Some of
the macrophages could clearly be seen migrating across the margin of the wound into the wound
bed itself. At the 2 and 4 week time points, large numbers of macrophages could be observed in
the wound distally infiltrating the distal margin in the deep dermal tissues (Figure 8).
Macrophage number appeared to decrease by 6 weeks. In contrast, group 2 animals (-collagen
gel) demonstrated very few macrophages in the distal wound margin in the early time points.
The few macrophages present were localized to the deep dermal structures at the junction of the
wound and normal tissues and macrophages invading into the wound were seldom noted (Figure
8). Thus, macrophage migration to the distal wound margin and into the wound bed from this
margin appeared to be delayed significantly by increased scarring and fibrosis.
Discussion

TGF-β1 is a negative regulator of lymphatic regeneration
20
Clinical studies have identified a number of risk factors for lymphedema including: 1.
Treatment related factors (e.g. surgery, radiation); 2. Disease related factors (eg. stage, nodal
status, number of lymph nodes removed); and 3. Patient factors (eg. age, obesity,
infection/inflammation, and excessive arm use). (33) It is clear that some of these factors are
related, and that soft tissue scarring and fibrosis are the common denominator.
Abnormal wound repair and scarring is known to be associated with marked alterations in
the expression of profibrotic growth factors such as TGF-β1. Many of the clinical conditions
that predispose patients to develop lymphedema, most notably radiation therapy, are known to be
associated with increased TGF-β1 expression. (5, 23, 42) Furthermore, inhibition of TGF-β1 or
its intermediaries (e.g. SMAD-3), decreases scarring caused by fibrotic stimuli and radiation, and
improves wound healing. (12-14) Consistent with this, we demonstrated that treatment of
excisional wounds with collagen resulted in significantly decreased scarring and down-regulation
of TGF-β1 expression. Furthermore, we show that diminished TGF-β1 expression is correlated
both temporally and spatially with decreased scarring and accelerated lymphatic regeneration.
The addition of TGF-β1 to the collagen gel recapitulated the fibrosis and deficiencies in
lymphatic regeneration noted in the group treated without collagen gel. Thus, inhibition of TGF-
β1 expression in the collagen gel treated animals was associated with near normal lymphatic
function by 4 weeks postoperatively, whereas lymphatic regeneration in the no collagen group or
the group treated with collagen supplemented with a small dose of TGF-β1 remained markedly
delayed with continued lymphedema even 6 weeks after surgery.
TGF-β1 inhibition of lymphatic regeneration may occur as a consequence of direct
inhibitory effects of TGF-β1 on lymphatic endothelial cells. We found that even low doses of
this growth factor significantly attenuate the proliferation of isolated LECs in vitro. This in vitro

TGF-β1 is a negative regulator of lymphatic regeneration
21
finding is correlated with significant decreases in the number of LECs and the number of
proliferating LECs present in the animals treated without collagen gel (with resultant increase in
TGF-β1) and those treated with supplemental TGF-β1 as compared with animals treated with
collagen gel. In addition, TGF-β1 may have direct negative effects on the potential of LECs to
coalesce into lymphatic channels as evidenced by significant inhibition of tubule formation in
vitro, and large ectatic vessels in vivo. These findings, together with a recent publication by Oka
et al demonstrating a direct inhibitory role for TGF-β1 during inflammatory and tumor
associated lymphangiogenesis (30) suggest that this pro-fibrotic molecule can directly inhibit
lymphatic repair by inhibiting assembly of lymphatic vessels, reducing LEC proliferation, and
inhibiting LEC migration. Our results add to the findings of Oka et al by demonstrating a dose-
dependent inhibition of LEC proliferation and function, as well as demonstrating a negative role
for TGF-β1 during wound repair. These findings are important since they suggest that anti-
scarring strategies in general and inhibition of TGF-β1 signaling in particular, may lead to
improved lymphatic regeneration and possibly represent a potential preventative treatment.
Given that no preventative strategies for lymphedema are currently available, and that this
debilitating condition affects a large number of patients who undergo lymph node dissection for
cancer, our study provides an important first step towards a potential treatment to prevent
abnormal lymphatic regeneration.
Anatomic studies on cadavers treated with axillary lymph node dissection have recently
demonstrated that capillary lymphatics become obliterated due to fibrosis and narrowing.(48)
Fibrosis contributes to lymphatic dysfunction by preventing physiologic dilatation of capillary
lymphatic vessels necessary for lymphatic fluid uptake when fluid accumulates within tissues.
Consistent with this finding we have demonstrated that increased scarring is associated with

TGF-β1 is a negative regulator of lymphatic regeneration
22
lymphatic fibrosis and an increased number of alpha-SMA positive myofibroblasts. In addition,
we found that in contrast to the collagen gel treated animals, animals treated without collagen gel
or those treated with collagen gel supplemented with recombinant TGF-β1 demonstrated
numerous capillary lymphatic endothelial cells that co-expressed LYVE-1 and alpha-SMA. This
finding implies that direct LEC-mesenchymal cell transdifferentiation may occur in the setting of
increased TGF-β1 expression and scarring and may be a putative mechanism for the clinically
observed phenomena of lymphatic vessel fibrosis and obliteration in patients with lymphedema
or lymphatic dysfunction. This hypothesis is supported by the fact that TGF-β1 causes
microvascular endothelial to mesenchymal differentiation both in vivo and in isolated
microvascular and macrovascular bovine endothelial cells resulting in the induction of alpha
SMA-expression.(3, 31, 55) In addition, similar to our study, these in vitro and in vivo studies
have demonstrated so-called “transitional” cells co-expressing endothelial and fibroblastic
markers (alpha-SMA in most cases).(3, 7, 31, 55) Finally, our hypothesis that TGF-β1 causes
lymphatic fibrosis is further validated by the findings of a recent study demonstrating that the
expression of TGF-β1 by regulatory T-cells in the cortical portions of lymph nodes is responsible
for lymph node fibrosis after simian immunodeficiency virus infection.(11) Clearly, however,
additional work is required to determine conclusively if abnormal TGF-β1 expression results in
LEC transdifferentiation. These studies are currently underway in our laboratory.
Interestingly, we did not find differences in VEGF-C expression as a consequence of
changes in TGF-β1 expression during wound repair. VEGF-C is a critical regulator of
lymphangiogenesis by promoting LEC proliferation, migration, and tubule formation.(22) Thus,
it may be that increased TGF-β1 expression and resultant soft tissue fibrosis/scarring do not alter
VEGF-C expression but instead change other factors in the wound that promote lymphatic

TGF-β1 is a negative regulator of lymphatic regeneration
23
regeneration. This is supported by our finding that TGF-β1 had no effect on VEGF-C or VEGF-
D expression in isolated LECs. Similarly, Oka et al found no differences in the expression of
VEGF-C by peritoneal macrophages after inhibition of TGF-β1 activity in their inflammatory
model of lymphangiogenesis.(30) The lack of changes in VEGF-C/D expression further
supports a direct inhibitory role for TGF-β1 on lymphatic regeneration. It is possible however,
that TGF-β1 reacts indirectly with VEGF-C/D by altering the expression of VEGF-receptor 2 or
3 thereby inhibiting lymphatic regeneration during wound repair. (15) These interactions require
further study.
Soft tissue fibrosis and abnormal scarring may also have indirect effects on lymphatic
regeneration. For example, abnormal wound healing may alter interstitial flow which has been
recently identified as an important regulator of tissue morphogenesis and lymphatic
regeneration.(36) In a series of elegant experiments, Boardman et al demonstrated that interstitial
flow is an important mediator of LEC migration leading to re-connection to proximal
lymphatics.(6) Although undoubtedly interstitial flow is important to normal lymphatic
regeneration, it is unlikely that changes in interstitial flow alone could explain our findings
during abnormal wound healing. This conclusion is supported by our finding that abnormal
wound repair did not result in significant changes in VEGF-C expression where as Swartz et al
have shown that interstitial flow dependent lymphatic regeneration in the tail model is dependent
on VEGF-C expression and VEGF-R3 signaling.(34) In addition, it is unlikely that there would
be significant differences in interstitial flow in animals treated with collagen as compared with
those treated with collagen supplemented with TGF-β1.
An additional mechanism by which soft tissue fibrosis may delay or inhibit lymphatic
regeneration is alterations in the recruitment of macrophages. Numerous studies have implicated

TGF-β1 is a negative regulator of lymphatic regeneration
24
macrophages as critical regulators of lymphangiogenesis.(18) Macrophage expression of VEGF-
C is a major regulator of lymphangiogenesis in some tumors and in inflammation.(25, 44)
Decreased macrophage number and function is associated with diminished lymphangiogenesis
and impaired wound healing in diabetic mice.(24) In addition, macrophages are thought to
directly contribute to lymphatic vessel formation by trans-differentiating into LECs. We
demonstrated that macrophages were more prominently present in the +collagen gel beginning as
early as 1 week postoperatively. More importantly, we found that more F4/80 stained
macrophages infiltrated the distal and proximal portions of the wound in the group 1 animals
(+collagen gel) at the early time points as compared with group 2 animals (-collagen gel).
In conclusion, we have shown for the first time that the rate of lymphatic regeneration is
inversely proportional to the expression of TGF-β1 and amount of scarring that is present in a
wound. Further, we have shown that this effect appears to be independent of VEGF-C/D.
Increased TGF-β1 expression appears to have direct inhibitory effects on lymphatic repair by
decreasing LEC recruitment, proliferation, and tubule formation. In addition, TGF-β1 expressed
during abnormal wound repair is associated with lymphatic fibrosis and possibly LEC-fibroblast
transdifferentiation. These results are critical to the understanding of lymphatic repair after
surgical disruption, implying that methods which decrease scarring/fibrosis or the expression of
TGF-β1 may play a role in the prevention of lymphatic dysfunction and subsequent secondary
lymphedema.

TGF-β1 is a negative regulator of lymphatic regeneration
25
Figure Legends
Figure 1. Treatment of mouse tail skin and lymphatic excision with topical collagen gel
inhibits TGF-β1 expression.
A. Representative photomicrographs of TGF-β1 immunostaining in group 1 (+collagen; left,
open arrow) and group 2 (-collagen gel; right, closed arrow) wounds at various times after
surgery (100X). Note marked increase in TGF-β1expression both in the ECM and in spindle-
shaped cells surrounding the excision sites. Note also that TGF-β1 expression in group 2
persisted in the ECM even at the 6 week time point. B. Relative quantitation of TGF-β1
staining. Note the significant increase in TGF-β1 expression at all time points in the group 2 (-
collagen gel) wounds as compared with group 1 (+ collagen gel) animals (*p<0.05). C.
Quantitative PCR for TGF-B1 from total cellular RNA harvested from wounds of mouse in
group 1 and group 2 two weeks postoperatively (*p<0.002). D. Western blot of TGF-B1
protein expression from wounds of mice in group 1 and group 2 two weeks postoperatively.
Note significant increase in TGF-B1 expression in group 2 (-collagen) animals.
Figure 2. Collagen gel treatment and decreased TGF-β1 expression are associated with
decreased scarring and fibrosis.
A. Representative H&E low-power images (50X) of group 1 (+collagen gel; upper left) and
group 2 (-collagen gel; upper middle), and supplemental TGF-β1 treated animals (+collagen
gel/+TGF-β1; upper right) 6 weeks after surgery. Note increased inflammation, hypercellularity,
and tissue edema in group 2 and TGF-β1 treated wounds. B. Representative high-power images
(200x) of picro-sirius red staining in group 1 (lower left), group 2 (lower middle) and TGFβ1-
treated (lower right) wounds 6 weeks after surgery. Note increased orange/red birefringence

TGF-β1 is a negative regulator of lymphatic regeneration
26
(closed arrow) and more parallel deposition of fibers (circle) in group 2 and supplemental TGF-
β1 treated wounds when compared to group 1 indicating more scarring and fibrosis. C. Scar
indices of wounds in group 1, group 2, and supplemental TGF-β1 treated animals. Note
significant decrease in scarring in the group 1 (+collagen) as compared with other groups
(*p<0.05). Data are presented as mean + SEM of quantification from 3 random high powered
fields per animal (n=4) at each time point.
Figure 3. Inhibition of TGF-β1 expression and scarring is associated with increased
functional lymphatic transport and decreased acute lymphedema.
A. Static representative images of nodal uptake of 99mTc-sulfur colloid in group 1 (+collagen
gel; upper panels), group 2 (-collagen gel; middle panel), and supplemental TGFβ1 treated
(+collagen gel/+TGF-β1; lower panel) animals. Note radioactive uptake (red hue) in lymph
nodes (arrow) at the base of the tail in group 1 (+collagen gel) animals beginning at 2 weeks. In
contrast, note minimal uptake in group 2 and supplemental TGF-β1 groups even after 6 weeks.
Arrow marks the tip of the tail. B. Quantitation of radio-nucleotide uptake after
lymphoscintigraphy 6 weeks after surgery. Note significant increase in both the rate (i.e. slope
of the line) and amount of radio-nucleotide uptake in group 1 animals demonstrating more rapid
lymphatic regeneration (data are presented as mean + SEM of uptake calculations from 4 animals
per time point; *p<0.05). In contrast, note severe dysfunction in lymphoscintigraphic uptake in
the supplemental TGF-β1 treated animals. C. Tail volume measurements after surgery. Note
significant decrease in tail volume and return to normal by 4 weeks in group 1 animals (*p <0.05
as compared to all other groups). In contrast, note increase in tail volume measurements and
lymphedema in group 2 and supplemental TGF-β1 treated animals even 6 weeks postoperatively.

TGF-β1 is a negative regulator of lymphatic regeneration
27
Figure 4. Treatment of wounds with supplemental TGF-β1 results in chronic lymphedema
changes distal to the site of surgery.
Representative low-power photomicrographs of wounds treated with collagen gel with (left
panels) or collagen gel supplemented with 50ng of recombinant human TGF-β1 (right panels).
Note subdermal thickening (brackets in lower figures), dilated lymphatics (open arrow), fibrosis,
and hypercellularity in supplemental TGF-β1 treated wounds. Also, note fat deposition (filled
black arrow upper right panel) in the cross sections of the tail harvested distal to the surgical site
and treated with TGF-β1.
Figure 5. TGF-β1 inhibits LEC recruitment and proliferation.
A. Number of LYVE-1 positive cells (LECs) at the distal margin of the wound 2, 4, and 6 weeks
after surgery. Note significant increase in the number of LECs present in the distal portion of
group 1 (+collagen gel) as compared with group 2 (-collagen gel) and supplemental TGF-β1
treated (+collagen gel/+TGF-β1) animals (mean + standard deviation of 3 high powered
fields/animal (n=4) per time point). B. Identification of proliferating LECs (LYVE-1 and PCNA
double positive stain) in wound sections 2 weeks postoperatively (representative high powered
figures shown at 400x magnification). Note red LYVE-1 membrane staining (left), green PCNA
nuclear staining (middle), and double positive cells indicating proliferating LECs (Right; open
arrows). C. Quantification of proliferating LECs (+PCNA +LYVE) in wound sections 2 weeks
postop. Note nearly 6 fold increase in proliferating LECs present in group 1 (+collagen gel) as
compared to group 2 (-collagen) and supplemental TGF-β1 (+collagen gel/+TGF-β1) treated
animals (*p<0.01; data are presented as mean + standard deviation of 3 high powered
fields/animal (n=4 per group). D. TGF-β1 inhibits LECs proliferation in vitro. Note

TGF-β1 is a negative regulator of lymphatic regeneration
28
significant, dose dependent decrease in cellular proliferation with exposure to increasing doses of
TGF-β1 (0, 1, 10ng/ml; Mean + SD of triplicate experiments; *p<0.05).
Figure 6. Increased TGF-β1 during wound repair is associated with abnormal lymphatic
architecture and dilated lymphatics but does not alter VEGF-C or VEGF-D expression.
A. Representative florescent photomicrographs for LYVE-1 in group 1 (+collagen gel; left),
group 2 (-collagen gel; middle), and supplemental TGF-β1 (+collagen/+TGF-β1; right) at
various time points after surgery (50X magnification). Note lymphatic capillary formation in
group 1 animals at the earliest time points with cylindrical architecture and mild dilatation
progressing to collapse lymphatics by 6 weeks (small circle). In contrast, note little capillary
lymphatic regeneration in group 2 and supplemental TGF-β1 animals with the presence of
markedly dilated, ectatic lymphatics (large circles). B. TGF-β1 inhibits LEC tubule formation in
matrigel tubule formation assay in vitro. Representative images (20x) of LECs treated with
various doses of TGF-β1 (0, 1, 10ng/ml) for 3 days then plated on Matrigel coated plates for 24
hours. Note dose dependent inhibition of tubule formation in the TGF-β1 treated LECs. Also,
not large ectatic tubule formation in the TGF-β1 treated groups reminiscent of our in vivo
findings (below panels: 400X representative images). All experiments were performed in
quadruplicate. C, D. Expression of VEGF-C and VEGF-D protein by LECs 24 hours after
treatment with TGF-β1 (0, 1, 10ng/ml). Note no significant difference in VEGF-C/D expression
by ELISA (mean + SD; p=NS).
Figure 7. TGF-β1 induces expression of alpha-SMA and results in lymphatic fibrosis and
co-expression of lymphatic and fibroblast markers by LECs in-vivo.

TGF-β1 is a negative regulator of lymphatic regeneration
29
A. Representative photomicrographs (200x) of alpha-SMA staining in group 1 (+ collagen gel)
and group 2 (- Collagen gel) animals 6 weeks after surgery. Note significant increase in alpha-
SMA expression in group 2 animals. Also note few tubular structures in group 2 animals with
alpha-SMA staining. B. Localization LYVE-1 (red; upper panels) and alpha-SMA (green;
middle panel) in group 1 (+ ollagen; left), group 2 (-collagen gel; middle), and supplemental
TGF-β1 (+collagen gel/+TGF-β1; right) 6 weeks postoperatively. Co-localization (lower panels)
is shown as orange/yellow staining). Note numerous double stained lymphatic vessels (LYVE-1
and alpha-SMA) in group 2 and supplemental TGF-β1 treated animals as compared to group 1.
Also, note that double stained lymphatics demonstrate enlarged lumens and thickened walls
consistent with lymphatic fibrosis (large ovals). C. A high powered image of a lymphatic
composed of cells expressing both LYVE and alpha-SMA (open arrow) as opposed to a
lymphatic with no alpha-SMA expression (open arrowhead).
Figure 8. Decreased scarring is associated with increased infiltration of F4/80 positive
macrophages.
Representative images the distal portions of group 1 and 2 animals at 4 weeks after surgery
(distal aspect of the wound is towards the right of the figures). Upper row demonstrates low-
power (50X) and lower row demonstrates magnified insert (200X). Note large numbers of F4/80
positive macrophages infiltrating into the distal margin of the group 1 section. In contrast, note
paucity of cells in the group 2 section.

TGF-β1 is a negative regulator of lymphatic regeneration
30
References
1. Abu-Rustum NR, Alektiar K, Iasonos A, Lev G, Sonoda Y, Aghajanian C, Chi DS, and Barakat RR. The incidence of symptomatic lower-extremity lymphedema following treatment of uterine corpus malignancies: a 12-year experience at Memorial Sloan-Kettering Cancer Center. Gynecol Oncol 103: 714-718, 2006. 2. Ammann AJ, Beck LS, DeGuzman L, Hirabayashi SE, Lee WP, McFatridge L, Nguyen T, Xu Y, and Mustoe TA. Transforming growth factor-beta. Effect on soft tissue repair. Annals of the New York Academy of Sciences 593: 124-134, 1990. 3. Arciniegas E, Frid MG, Douglas IS, and Stenmark KR. Perspectives on endothelial-to-mesenchymal transition: potential contribution to vascular remodeling in chronic pulmonary hypertension. Am J Physiol Lung Cell Mol Physiol 293: L1-8, 2007. 4. Ask K, Bonniaud P, Maass K, Eickelberg O, Margetts PJ, Warburton D, Groffen J, Gauldie J, and Kolb M. Progressive pulmonary fibrosis is mediated by TGF-beta isoform 1 but not TGF-beta3. Int J Biochem Cell Biol, 2007. 5. Ask K, Martin GE, Kolb M, and Gauldie J. Targeting genes for treatment in idiopathic pulmonary fibrosis: challenges and opportunities, promises and pitfalls. Proc Am Thorac Soc 3: 389-393, 2006. 6. Boardman KC and Swartz MA. Interstitial flow as a guide for lymphangiogenesis. Circulation research 92: 801-808, 2003. 7. Camino AM, Atorrasagasti C, Maccio D, Prada F, Salvatierra E, Rizzo M, Alaniz L, Aquino JB, Podhajcer OL, Silva M, and Mazzolini G. Adenovirus-mediated inhibition of SPARC attenuates liver fibrosis in rats. J Gene Med 10: 993-1004, 2008. 8. De Vries HJ, Zeegelaar JE, Middelkoop E, Gijsbers G, Van Marle J, Wildevuur CH, and Westerhof W. Reduced wound contraction and scar formation in punch biopsy wounds. Native collagen dermal substitutes. A clinical study. The British journal of dermatology 132: 690-697, 1995. 9. Decologne N, Kolb M, Margetts PJ, Menetrier F, Artur Y, Garrido C, Gauldie J, Camus P, and Bonniaud P. TGF-beta1 Induces Progressive Pleural Scarring and Subpleural Fibrosis. J Immunol 179: 6043-6051, 2007. 10. Ehrlich HP, Desmouliere A, Diegelmann RF, Cohen IK, Compton CC, Garner WL, Kapanci Y, and Gabbiani G. Morphological and immunochemical differences between keloid and hypertrophic scar. Am J Pathol 145: 105-113, 1994. 11. Estes JD, Wietgrefe S, Schacker T, Southern P, Beilman G, Reilly C, Milush JM, Lifson JD, Sodora DL, Carlis JV, and Haase AT. Simian immunodeficiency virus-induced lymphatic tissue fibrosis is mediated by transforming growth factor beta 1-positive regulatory T cells and begins in early infection. J Infect Dis 195: 551-561, 2007. 12. Flanders KC, Major CD, Arabshahi A, Aburime EE, Okada MH, Fujii M, Blalock TD, Schultz GS, Sowers A, Anzano MA, Mitchell JB, Russo A, and Roberts AB. Interference with transforming growth factor-beta/ Smad3 signaling results in accelerated healing of wounds in previously irradiated skin. Am J Pathol 163: 2247-2257, 2003. 13. Flanders KC, Sullivan CD, Fujii M, Sowers A, Anzano MA, Arabshahi A, Major C, Deng C, Russo A, Mitchell JB, and Roberts AB. Mice lacking Smad3 are protected against cutaneous injury induced by ionizing radiation. Am J Pathol 160: 1057-1068, 2002.

TGF-β1 is a negative regulator of lymphatic regeneration
31
14. Gauldie J, Kolb M, Ask K, Martin G, Bonniaud P, and Warburton D. Smad3 signaling involved in pulmonary fibrosis and emphysema. Proc Am Thorac Soc 3: 696-702, 2006. 15. Goldman J, Rutkowski JM, Shields JD, Pasquier MC, Cui Y, Schmokel HG, Willey S, Hicklin DJ, Pytowski B, and Swartz MA. Cooperative and redundant roles of VEGFR-2 and VEGFR-3 signaling in adult lymphangiogenesis. FASEB J 21: 1003-1012, 2007. 16. Haviv YS, Takayama K, Nagi PA, Tousson A, Cook W, Wang M, Lam JT, Naito S, Lei X, Carey DE, and Curiel DT. Modulation of renal glomerular disease using remote delivery of adenoviral-encoded solubletype II TGF-beta receptor fusion molecule. J Gene Med 5: 839-851, 2003. 17. Hinrichs CS, Watroba NL, Rezaishiraz H, Giese W, Hurd T, Fassl KA, and Edge SB. Lymphedema secondary to postmastectomy radiation: incidence and risk factors. Annals of surgical oncology 11: 573-580, 2004. 18. Kerjaschki D. The crucial role of macrophages in lymphangiogenesis. The Journal of clinical investigation 115: 2316-2319, 2005. 19. Kim KH, Kim HC, Hwang MY, Oh HK, Lee TS, Chang YC, Song HJ, Won NH, and Park KK. The antifibrotic effect of TGF-beta1 siRNAs in murine model of liver cirrhosis. Biochem Biophys Res Commun 343: 1072-1078, 2006. 20. Leask A and Abraham DJ. TGF-beta signaling and the fibrotic response. FASEB J 18: 816-827, 2004. 21. Liu W, Cai Z, Wang D, Wu X, Cui L, Shang Q, Qian Y, and Cao Y. Blocking transforming growth factor-beta receptor signaling down-regulates transforming growth factor-beta1 autoproduction in keloid fibroblasts. Chin J Traumatol 5: 77-81, 2002. 22. Lohela M, Saaristo A, Veikkola T, and Alitalo K. Lymphangiogenic growth factors, receptors and therapies. Thrombosis and haemostasis 90: 167-184, 2003. 23. Martin M, Lefaix J, and Delanian S. TGF-beta1 and radiation fibrosis: a master switch and a specific therapeutic target? International journal of radiation oncology, biology, physics 47: 277-290, 2000. 24. Maruyama K, Asai J, Ii M, Thorne T, Losordo DW, and D'Amore PA. Decreased macrophage number and activation lead to reduced lymphatic vessel formation and contribute to impaired diabetic wound healing. Am J Pathol 170: 1178-1191, 2007. 25. Maruyama K, Ii M, Cursiefen C, Jackson DG, Keino H, Tomita M, Van Rooijen N, Takenaka H, D'Amore PA, Stein-Streilein J, Losordo DW, and Streilein JW. Inflammation-induced lymphangiogenesis in the cornea arises from CD11b-positive macrophages. The Journal of clinical investigation 115: 2363-2372, 2005. 26. Mehrara BJ, Mackool RJ, McCarthy JG, Gittes GK, and Longaker MT. Immunolocalization of basic fibroblast growth factor and fibroblast growth factor receptor-1 and receptor-2 in rat cranial sutures. Plastic and reconstructive surgery 102: 1805-1817; discussion 1818-1820, 1998. 27. Navarro A, Perez RE, Rezaiekhaligh M, Mabry SM, and Ekekezie, II. T1{alpha}/podoplanin is essential for capillary morphogenesis in lymphatic endothelial cells. Am J Physiol Lung Cell Mol Physiol, 2008. 28. Oberringer M, Meins C, Bubel M, and Pohlemann T. In vitro wounding: effects of hypoxia and transforming growth factor beta1 on proliferation, migration and myofibroblastic differentiation in an endothelial cell-fibroblast co-culture model. J Mol Histol 39: 37-47, 2008.

TGF-β1 is a negative regulator of lymphatic regeneration
32
29. Oberringer M, Meins C, Bubel M, and Pohlemann T. In vitro wounding: effects of hypoxia and transforming growth factor beta(1) on proliferation, migration and myofibroblastic differentiation in an endothelial cell-fibroblast co-culture model. J Mol Histol 39: 37-47, 2008. 30. Oka M, Iwata C, Suzuki HI, Kiyono K, Morishita Y, Watabe T, Komuro A, Kano MR, and Miyazono K. Inhibition of endogenous TGF-{beta} signaling enhances lymphangiogenesis. Blood, 2008. 31. Okada H, Danoff TM, Kalluri R, and Neilson EG. Early role of Fsp1 in epithelial-mesenchymal transformation. The American journal of physiology 273: F563-574, 1997. 32. Pertovaara L, Kaipainen A, Mustonen T, Orpana A, Ferrara N, Saksela O, and Alitalo K. Vascular endothelial growth factor is induced in response to transforming growth factor-beta in fibroblastic and epithelial cells. The Journal of biological chemistry 269: 6271-6274, 1994. 33. Petrek JA, Pressman PI, and Smith RA. Lymphedema: current issues in research and management. CA Cancer J Clin 50: 292-307; quiz 308-211, 2000. 34. Pytowski B, Goldman J, Persaud K, Wu Y, Witte L, Hicklin DJ, Skobe M, Boardman KC, and Swartz MA. Complete and specific inhibition of adult lymphatic regeneration by a novel VEGFR-3 neutralizing antibody. J Natl Cancer Inst 97: 14-21, 2005. 35. Rutkowski JM, Boardman KC, and Swartz MA. Characterization of lymphangiogenesis in a model of adult skin regeneration. American journal of physiology 291: H1402-1410, 2006. 36. Rutkowski JM and Swartz MA. A driving force for change: interstitial flow as a morphoregulator. Trends in cell biology 17: 44-50, 2007. 37. Saadeh PB, Mehrara BJ, Steinbrech DS, Dudziak ME, Greenwald JA, Luchs JS, Spector JA, Ueno H, Gittes GK, and Longaker MT. Transforming growth factor-beta1 modulates the expression of vascular endothelial growth factor by osteoblasts. The American journal of physiology 277: C628-637, 1999. 38. Saaristo A, Tammela T, Timonen J, Yla-Herttuala S, Tukiainen E, Asko-Seljavaara S, and Alitalo K. Vascular endothelial growth factor-C gene therapy restores lymphatic flow across incision wounds. Faseb J 18: 1707-1709, 2004. 39. Sakorafas GH, Peros G, Cataliotti L, and Vlastos G. Lymphedema following axillary lymph node dissection for breast cancer. Surgical oncology 15: 153-165, 2006. 40. Sato M, Muragaki Y, Saika S, Roberts AB, and Ooshima A. Targeted disruption of TGF-beta1/Smad3 signaling protects against renal tubulointerstitial fibrosis induced by unilateral ureteral obstruction. The Journal of clinical investigation 112: 1486-1494, 2003. 41. Schledzewski K, Falkowski M, Moldenhauer G, Metharom P, Kzhyshkowska J, Ganss R, Demory A, Falkowska-Hansen B, Kurzen H, Ugurel S, Geginat G, Arnold B, and Goerdt S. Lymphatic endothelium-specific hyaluronan receptor LYVE-1 is expressed by stabilin-1+, F4/80+, CD11b+ macrophages in malignant tumours and wound healing tissue in vivo and in bone marrow cultures in vitro: implications for the assessment of lymphangiogenesis. The Journal of pathology 209: 67-77, 2006. 42. Schmid P, Itin P, Cherry G, Bi C, and Cox DA. Enhanced expression of transforming growth factor-beta type I and type II receptors in wound granulation tissue and hypertrophic scar. Am J Pathol 152: 485-493, 1998. 43. Schonmeyr BH, Wong AK, Soares M, Fernandez J, Clavin N, and Mehrara BJ. Ionizing radiation of mesenchymal stem cells results in diminution of the precursor pool and

TGF-β1 is a negative regulator of lymphatic regeneration
33
limits potential for multilineage differentiation. Plastic and reconstructive surgery 122: 64-76, 2008. 44. Schoppmann SF, Birner P, Stockl J, Kalt R, Ullrich R, Caucig C, Kriehuber E, Nagy K, Alitalo K, and Kerjaschki D. Tumor-associated macrophages express lymphatic endothelial growth factors and are related to peritumoral lymphangiogenesis. Am J Pathol 161: 947-956, 2002. 45. Shim KS, Kim KH, Han WS, and Park EB. Elevated serum levels of transforming growth factor-beta1 in patients with colorectal carcinoma: its association with tumor progression and its significant decrease after curative surgical resection. Cancer 85: 554-561, 1999. 46. Shin WS, Szuba A, and Rockson SG. Animal models for the study of lymphatic insufficiency. Lymphatic research and biology 1: 159-169, 2003. 47. Sitzia J. Volume measurement in lymphoedema treatment: examination of formulae. European journal of cancer care 4: 11-16, 1995. 48. Suami H, Pan WR, and Taylor GI. Changes in the lymph structure of the upper limb after axillary dissection: radiographic and anatomical study in a human cadaver. Plastic and reconstructive surgery 120: 982-991, 2007. 49. Szuba A, Achalu R, and Rockson SG. Decongestive lymphatic therapy for patients with breast carcinoma-associated lymphedema. A randomized, prospective study of a role for adjunctive intermittent pneumatic compression. Cancer 95: 2260-2267, 2002. 50. Tammela T, Saaristo A, Holopainen T, Lyytikka J, Kotronen A, Pitkonen M, Abo-Ramadan U, Yla-Herttuala S, Petrova TV, and Alitalo K. Therapeutic differentiation and maturation of lymphatic vessels after lymph node dissection and transplantation. Nature medicine 13: 1458-1466, 2007. 51. Theoret CL, Barber SM, Moyana TN, and Gordon JR. Expression of transforming growth factor beta(1), beta(3), and basic fibroblast growth factor in full-thickness skin wounds of equine limbs and thorax. Vet Surg 30: 269-277, 2001. 52. Tobin MB, Lacey HJ, Meyer L, and Mortimer PS. The psychological morbidity of breast cancer-related arm swelling. Psychological morbidity of lymphoedema. Cancer 72: 3248-3252, 1993. 53. Warren AG and Slavin SA. Scar lymphedema: fact or fiction? Annals of plastic surgery 59: 41-45, 2007. 54. Xavier S, Piek E, Fujii M, Javelaud D, Mauviel A, Flanders KC, Samuni AM, Felici A, Reiss M, Yarkoni S, Sowers A, Mitchell JB, Roberts AB, and Russo A. Amelioration of radiation-induced fibrosis: inhibition of transforming growth factor-beta signaling by halofuginone. The Journal of biological chemistry 279: 15167-15176, 2004. 55. Zeisberg EM, Potenta S, Xie L, Zeisberg M, and Kalluri R. Discovery of endothelial to mesenchymal transition as a source for carcinoma-associated fibroblasts. Cancer research 67: 10123-10128, 2007.











0
10
20
30
+Collagen Gel +TGFB1-Collagen Gel+Collagen Gel
2 4 6Weeks
** *
LE
C N
um
ber