Embed Size (px)
Citation preview

Plant Molecular Biology38: 609–622, 1998.© 1998Kluwer Academic Publishers. Printed in the Netherlands.
609
The Arabidopsis Athb-8, -9and -14genes are members of a small genefamily coding for highly related HD-ZIP proteins
Giovanna Sessa1,†, Corinna Steindler1,†, Giorgio Morelli2 and Ida Ruberti1,∗1Centro di studio per gli Acidi Nucleici, c/o Dipartimento di Genetica e Biologia Molecolare, Universit`a diRoma La Sapienza, P.le Aldo Moro 5, 00185 Rome, Italy (∗author for correspondence).2Unita di NutrizioneSperimentale, Istituto Nazionale della Nutrizione, Via Ardeatina 546, 00178 Rome, Italy;†The first two authorscontributed equally to this work
Received 4 February 1998; accepted in revised form 23 April 1998
Key words: binding site selection, gene expression, homeobox-leucine zipper genes, protein-DNA interaction
Abstract
We report the isolation and characterization of twoArabidopsishomeobox genes highly related to theAthb-8gene.The full-length cDNAs encode proteins of 841 and 852 amino acids which we have designated Athb-9 and -14,respectively. Athb-8,−9 and -14 are members of a small family of HD-Zip proteins (HD-ZIP III) characterized bya HD-Zip motif confined to the N-terminus of the polypeptide. The spatial organization of the HD-Zip domain ofAthb-8, -9 and -14 is different from that of the Athb-1 (a member of the HD-ZIP I family) and Athb-2 (a member ofthe HD-ZIP II family) HD-Zip domains. DNA binding analysis performed with random-sequence DNA templatesshowed that the Athb-9 HD-Zip (HD-Zip-9) domain, but not the Athb-9 HD alone, binds to DNA. The HD-Zip-9 domain recognizes a 11 bp pseudopalindromic sequence (GTAAT(G/C)ATTAC), as determined by selectinghigh-affinity binding sites from random-sequence DNA. Moreover, gel retardation assays demonstrated that theHD-Zip-9 domain binds to DNA as a dimer. These data support the notion that the HD-ZIP III domain interactswith DNA recognition elements in a fashion similar to the HD-ZIP I and II domains.
Introduction
Homeodomain-leucine zipper (HD-Zip) proteins areputative transcription factors identified only in plants.A number of HD-Zip proteins identified inArabidop-sis are thought to constitute a large class of proteins[2, 4, 12, 14, 17-19, 21, 23, 27]. We have tentativelygrouped these HD-Zip proteins into four different fam-ilies, named HD-ZIP I, II, III and IV [23]. Expressionanalysis inArabidopsisand in transgenic plants ofAthb-1 and -7 (HD-ZIP I) [1, 28],Athb-2 (HD-ZIPII) [4, 5, 20, 29], Athb-8 (HD-ZIP III) [2], Athb-10/GLABRA2(Athb-10/GL2) and ATML1 (HD-ZIPIV) [6, 12, 13, 17] suggests that the HD-Zip proteinsmight control important aspects of plant development.
The nucleotide sequence data reported will appear in theEMBL, GenBank and DDBJ Nucleotide Sequence Databases underthe accession numbers Y10922 (Athb-9gene) and Y11122 (Athb-14gene).
The presence of a putative leucine zipper closelylinked to a homeodomain suggested that the HD-Zipproteins could, via dimerization of the leucine zip-pers, recognize dyad-symmetric DNA sequences [18].DNA binding studies on several HD-Zip proteins sup-port this model. We have proved that the HD-Zipdomain of Athb-1, a member of the HD-ZIP I fam-ily, and that of Athb-2, a member of the HD-ZIP IIfamily, bind DNA as dimers [22, 24]. We found thatthe HD-Zip-1 and -2 binding sites are pseudopalin-dromes of two 5 bp half-sites that overlap at a centralposition, CAAT(A/T)ATTG and CAAT(G/C)ATTG,respectively [22, 23]. Moreover, we showed that thespacing between the zipper and the helix-3 is criticalfor function; in fact, insertion of two amino acids atthe beginning of the Athb-1 leucine zipper resulted ina HD-Zip complex incapable of DNA binding [22].Recently, we have also demonstrated that the leucinezipper of Athb-10/GL2, a member of the HD-ZIP IV

610
family, although less conserved, is active in dimeriza-tion and can functionally replace that of Athb-2 in anin vitro DNA-binding assay [6]. However, the pres-ence in Athb-8 (HD-ZIP III family) of a HD-Zip motifwith a different architecture [2] prevented us fromgeneralizing our DNA binding model for the HD-Zipproteins.
In this paper, we describe the isolation and char-acterization ofAthb-9 and -14, two genes encodingHD-Zip proteins highly related to Athb-8. We showthat the HD-Zip-9 domain interacts with DNA recog-nition elements in a fashion similar to the HD-Zip-1and -2 domains.
Material and methods
Plant material
Arabidopsis thalianaColumbia ecotype plants weregrown as previously described [4]. Roots were ob-tained from plants grown for three weeks in liquidmedium. Young seedlings were obtained from vernal-ized seeds germinated on solid medium and grown for5 days in the light. Soil-grown plants were taken toanalyse later stages. Whole plants were collected at 2and 3 weeks and the whole aerial portion at 4 weeks.Green siliques were harvested from plants 5 to 6 weeksold.
Isolation of cDNA and genomic clones
Athb-9Several genomic clones were isolated in a screeningperformed with a degenerate oligonucleotide (HB-2) corresponding to a conserved 6 amino acid se-quence from the helix-3 region of Athb-1 and -2.DNA sequence analysis of the homeobox region re-vealed that five of them code for HD-Zip proteins[23]. One of these clones, designatedAthb-9, wasfurther characterized. Genomic sequences flankingthe homeobox region were cloned and sequencedby standard techniques. [32P]-labelled genomic frag-ments spanning theAthb-9-coding region were usedto screen anA. thalianaColumbia cDNA library (5′Stretch, Clontech). Four clones (λHB-9c/1, /2, /3and /4, see Figure 1A) were obtained. TheAthb-9-hybridizing EcoRI fragments were subcloned intopBluescript KSII (Stratagene) and sequenced. No dif-ferences in DNA sequence were observed between theoverlapping regions of different cDNA inserts, with
the exception of an insertion of 88 nucleotides in theλHB-9c/4 clone. Two oligonucleotides, Athb-9 un12and Athb-9 un15 (see Table 1), in combination withthe Athb-9 rev8 (see Table 1) downstream primer wereused to map the 5′ end of the cDNA. An amplifica-tion product was obtained only with the Athb-9 un12primer. The fragment generated by PCR was sub-cloned (pHb-9c/amp2, see Figure 1A) into pBluescriptKSII, sequenced and found to extend 62 nucleotides5′ to the cDNA isolated (λHB-9c/1). No hybridizationsignal was detectable in northern blots containing stempoly(A)+ RNA probed with genomic fragments span-ning a 1.9 kb 5′ end region (5′ Athb-9/2 and /3, seeFigure 1A). DNA sequence comparison of the deducedfull-length cDNA with the genomic clone showed thatthe Athb-9 gene is split by 17 introns and indicatedthat the insertion present in the cDNA ofλHB-9c/4corresponds to the 14th intron. The significance of thepresence of an intron in one of the cDNAs isolatedwas investigated by performing synthesis of cDNAfrom Arabidopsistotal RNA and PCR amplification.A single RT-PCR product was obtained with the Athb-9 PCRun1 and Athb-9 PCRrev1 primers (see Table 1).DNA sequence analysis of this subcloned fragment(pHB-9c/amp1, see Figure 1A) revealed that the 14thintron is indeed not present in this cDNA.
Athb-14During the screening of theAthb-9 cDNA we iso-lated two cDNAs (λHB-14c/1 and -c/2, see Figure 1B)whose sequence analysis revealed a high degree ofsimilarity with the HD-Zip coding region ofAthb-9.Using theλHB-14c/1 insert as a probe, five genomicclones were isolated by screening 30 000 plaques ofan Arabidopsisgenomic library (Clontech). One ofthese, designatedAthb-14, was further characterized.Genomic sequences flanking the homeobox regionwere cloned and sequenced by standard techniques.The 3′ portion of theAthb-14 cDNA was obtainedby PCR cloning. Synthesis of cDNA from 1µg ofstem total RNA with random hexamers was performedwith the GeneAmp RNA PCR kit (Perkin Elmer-Cetus). The 1158 bp RT-PCR product obtained withthe Athb-14 PCRun1 and Athb-14 PCRrev1 primers(see Table 1) was subcloned (pHB-14c/amp1, see Fig-ure 1B) into pBluescript KSII and sequenced. The3′ end of the cDNA was cloned by performing 3′RACE (rapid amplification of cDNA ends) accordingto Frohmanet al. [7], with minor modifications. TheXSSpolyT adapter primer [6] was used to reverse-transcribe stem poly(A)+ RNA (0.3 µg). The cDNA

61
1
Table 1. DNA sequence of the oligonucleotide.
Athb-9 un 15 5′-GAATTCAACCTACTCTCCC -3′ (position 84–102)
of the genomic sequence
Athb-9 un 12 5′-CTGCCTCGCTTTTGCTC -3′ (position 262–278)
Athb-9 5D 5′-CCCGGGATCCGCCATGGACGATAGAGACTC -3′ (position 544–562)
Athb-9 A5D 5′-CCCGGGATCCGCCATGGATTCCGGCAAGTAC -3′ (position 579–593)
Athb-9 A3D 5′-CCCGGGAATTCCTGTCTGAAGACGAGCTGA -3′ (position 1285–1303)
Athb-9 rev8 5′-AGCTTGTTCATAGCACT -3′ (position 1318–1334)
Athb-9 B3D 5′-CCCGGGAATTCAGCAGTGTGGATTCG -3′ (position 1408–1419/1550–1552)
Athb-9 3D 5′-CCCGGGAATTCCTGTGGTCGTCCCAGAAG -3′ (position 1551–1568)
Athb-9 C3D 5′-CCCGGGAATCATAAAGAGGCCTGAGGAC -3′ (position 2727–2746)
Athb-9 PCRun1 5′-TGGAGGTTGTGCTCAGCTTG -3′ (position 3786–3805)
Athb-9 PCRrev1 5′-GAACTCGGAGCAAAGAGCTC -3′ (position 4687–4706)
Athb-14 un26 5′-GTACAAATATGGACACAAAG -3′ (position 2–21)
Athb-14 un10 5′-GTTCTTGCAAGTCTTTCT -3′ (position 165–182)
Athb-14 rev12 5′-GCGCGAGAAAGACTTGC -3′ (position 171–187)
Athb-14 un13 5′-CGTCTGAGATAAAGACCT -3′ (position 267–284)
Athb-14 rev18 5′-GGACCATCATCATAGCTC -3′ (position 602–619)
Athb-14 5D 5′-CCCGGGATCCGCCATGAACAGAGAGTCGCCGG -3′ (position 640–658)
Athb-14 rev4 5′-GTGGCCATTCTCATAGAC -3′ (position 1523–1540)
Athb-14 3D 5′-CCCGGGAATTCTCTGTGGTCGTCCCAGA -3′ (position 1656–1672)
Athb-14 PCRun1 5′-CTATGAATCATCGAAGATTC -3′ (position 2782–2801)
Athb-14 PCRun2 5′-GCTTCAATGGCACGTCAG -3′ (position 4217–4234)
Athb-14 PCRrev1 5′-CAAGATTGCATCAGAGTGAT -3′ (position 4547–4566)
Athb-8 5D 5′-CCCGGGATCCGCCATGGACAACGGGAAGTAC -3′ (position 937–954)
Athb-8 3D 5′-CCCGGGAATTCCTGTGGTAGCCAAATTCC -3′ (position 1445–1462)
BS-9 5′-GTAAAACGGCCAGTGAATTCAGATCTGTAATCATTACAGAGGATCCCTCGAGGTCGTGACTGGGAAAAC-3′

612
Figure 1. Organization of theAthb-9and-14genes. A. A sketch of the exon-intron organization of theAthb-9gene is shown on top. Filled andopen boxes represent coding and non coding exons, respectively. The dotted and cross-hatched boxes indicate the homeodomain (HD) and theleucine-zipper (Zip), respectively. Thin lines represent introns. cDNA clones and DNA probes are indicated by thick lines. B,BglII, C, ClaI,E, EcoRI, P,PstI, S, SacI (other EcoRI andPstI sites are present but not shown). B. A sketch of the exon-intron organization of theAthb-14gene is shown on top. Filled and open boxes represent coding and non coding exons, respectively. The dotted and cross-hatched boxes indicatethe homeodomain (HD) and the leucine-zipper (Zip), respectively. Thin lines represent introns. cDNA clones and DNA probes are indicated bythick lines. H,HindIII, C, ClaI (otherHindIII andClaI sites are present but not shown).

613
pool was then amplified with the XSS adapter primerand the Athb-14 PCRun2 (see Table 1) gene-specificprimer. The amplified product was subcloned (pHB-14c/amp2, see Figure 1B) into pBluescript KSII andsequenced. To determine if genomic sequences lo-cated 5′ to theAthb-14cDNA isolated are transcribed,synthesis of cDNA from stem total RNA and PCRamplification were performed. Two oligonucleotides,Athb-14 un13 and Athb-14 un10 (see Table 1), weredesigned from the genomic sequence and used in com-bination with Athb-14 rev4 (see Table 1). The 712 bpPCR product obtained with the Athb-14 un13 andAthb-14 rev4 primers was purified and re-amplifiedusing Athb-14 un13KpnI, containing a restriction sitefor cloning, and Athb-14 rev4 primers. The PCR prod-uct was restricted withKpnI andXhoI and the resulting460 bp fragment was subcloned (pHB-14/amp3, seeFigure 1B) into pBluescript KSII and sequenced. Nohybridization signal was detectable in northern blotscontaining stem poly(A)+ RNA probed with genomicfragments spanning a 1 kb 5′ end region (5′ Athb-14/2and 14/3, see Figure 1B).
DNA sequencing
Sequencing of the double stranded DNA was car-ried out with Sequenase 2 (USB) according to man-ufacturer’s instructions. Oligonucleotides were syn-thesized on a Applied Biosystem 394 synthesizerand purified according to manufacturer’s instructions.Comparative sequence analysis were performed withthe PILEUP program from the Genetics ComputerGroup.
DNA and RNA gel blot analyses
Genomic DNA isolation, DNA restriction analysis,gel electrophoresis and Southern blot onto Hybond-Nmembrane (Amersham) were done as previously de-scribed [4]. Fragments corresponding to the HD-Zipcoding region of theAthb-8(Hb-8),-9 (Hb-9) and-14(Hb-14) cDNAs were used as probes. [32P] labellingof the probes and hybridization conditions were as pre-viously described [4]. The Hb-8, -9 and -14 fragmentswere obtained by amplification of cDNA subcloneswith Athb-8 5D and Athb-8 3D, Athb-9 5D and Athb-9 3D, and Athb-14 5D and Athb-14 3D (see Table 1),respectively.
RNA was isolated as previously described [4].Poly(A)+ RNA was isolated from total RNA withDynabeads Oligo(dT)25 (Dynal AS) according to
manufacturer’s instructions. For northern blot analy-sis 3 µg of poly(A)+ RNA from A. thaliana wasdenatured, electrophoresed on 1.2% agarose-1.9%formaldehyde gels and transferred onto Hybond-C ex-tra membranes (Amersham). The amount ofAthb-9,-14and-8 transcripts was quantitated by scanning theX-ray films (different exposures) with a laser densit-ometer (Imaging Densitometer GS-670; BioRad) andnormalized to the amount of total RNA by transcriptquantification of the nuclear gene encoding theβ sub-unit of the mitochondrial ATPase [4]. To evaluate therelative abundance ofAthb-9, -8 and -14 transcripts,we performed three parallel hybridizations of rootpoly(A)+ RNA using as probes 5′Athb-9/1, 5′Athb-14/1 and 5′Athb-8 [2], respectively. To normalize forprobe length and specific activity, a Southern blot withthe corresponding genomic fragment was included ineach hybridization. The 5′Athb-14/1 and -/2 fragmentswere obtained by amplification of a genomic subclonewith Athb-14 un13 and Athb-14 rev18, and Athb-14 un26 and Athb-14 rev12 primers (see Table 1),respectively.
Construction of GST fusion proteins
The DNA fragments corresponding to the HD andHD-Zip domains of Athb-9 were obtained by PCRamplification. The PCR reactions were performed aspreviously described [22]. TheAthb-9cDNA was am-plified with primers containing restriction sites forcloning (Athb-9 5D, -9 A3D, -9 3D, see Table 1). TheDNA fragments were ligated in frame into theBamHI-EcoRI sites of the glutathioneS-transferase (GST)expression vector pGEX-2T (Pharmacia) [26]. Thefusion proteins derived from Athb-9 were designatedGST-HD-9 and GST-HD-Zip-9. Expression constructswere transformed intoEscherichia coliK12 strainJM109 (Pharmacia), and preparation ofE. coli proteinextracts was performed as previously described [22].
Binding site selection
The D3 oligonucleotide used for binding site selec-tion and the primers A and B used for amplificationof D3 have been previously described [22]. Double-stranded DNA templates were labeled to a specificactivity of ca. 2×108 cpm/µg by PCR reactions asdescribed by Blackwell and Weintraub [3]. Selectionof DNA, specifically bound by the GST fusion protein,was done by subsequent rounds of DNA binding, iso-lation of bound DNA by an affinity chromatographyassay and PCR amplification as previously described

614
[22]. The E. coli protein extracts utilized for bind-ing contained either GST (ca. 120 ng), or GST-HD-9(ca. 150 ng), or GST-HD-Zip-9 (ca. 200 ng). After7 rounds of selection, double-stranded 70-mer wasdigested withXhoI and EcoRI and ligated into sim-ilarly digested pBluescript II KS. 25 single colonieswere picked and plasmid DNA prepared (Qiagen tip100; Diagen). Sequencing was carried out on double-stranded DNA using Sequenase 2 (USB) according tothe manufacturer’s instructions. DNA sequences wereobtained with the M13(-20) primer.
In vitro transcription and translation
For mixing experiments, aBamHI-EcoRI DNA frag-ment slightly smaller (HD-ZipS-9) than that clonedinto pGEX-2T, and a DNA fragment containing, inaddition to the HD-Zip-coding region, a region cod-ing for a portion C-terminal to the HD-Zip domainof Athb-9 (HD-ZipL-9), were cloned into pGEM-4Z(Promega). The DNA fragments were obtained byPCR amplification of theAthb-9cDNA with primersAthb-9 A5D and Athb-9 B3D, and Athb-9 5D andAthb-9 C3D, respectively (see Table 1). The syn-thetic oligonucleotides Athb-9 5D and Athb-9 A5Dinclude a translational start site.In vitro transcriptionand translation reactions (Promega) were performed aspreviously described [22].
Gel mobility shift assay with GST fusion proteins
The sequence of the BS-9 DNA fragment used ingel retardation assays is shown in Table 1. Double-stranded32P-labelled DNA (3 ng, 6×104 cpm) wasincubated withE. coliprotein extract (4µl), in bindingbuffer [22] containing 1µg poly(dI-dC) (Boehringer-Mannheim) as non-specific competitor DNA. TheE. coli protein extracts utilized for gel retardationassays contained either GST (ca. 100 ng) or GST-HD-Zip-9 (ca. 160 ng). Incubation was performed in avolume of 20µl for 1 h at 20◦C. After the incuba-tion step, the mixture was immediately loaded onto4.5% polyacrylamide gels (acrylamide/bisacrylamide,29:1). Electrophoresis was performed in 0.5× TBE(1× TBE is 89 mM Tris-borate, 2 mM EDTA pH 8.3)for 2 h at 14 mA. Prior to loading, gels were pre-runfor 15 min at 14 mA. Dried gels were subjected toautoradiography at−80◦C with intensifying screens.
Gel mobility shift assay within vitro translatedproteins
The BS-9 duplex was cloned into pBluescript II KS,excised as a 36 bp fragment, and32P-end-labelled to aspecific activity of ca. 5×108 cpm/µg using Klenowfragment (Promega).32P-labelled DNA (ca. 0.1 ng)was incubated with the translated product (1µl), inbinding buffer [22] containing 1µg poly(dI-dC) asnon-specific competitor DNA. Incubation was per-formed in a volume of 20µl for 1 h at 4◦C. Formixing experiments, the RNAs were translated sep-arately, and 1µl of each translation reaction wasadded to the DNA-binding reaction mixture withoutthe DNA probe. After a 1 h incubation at 4◦C, theDNA probe was added to initiate the binding reaction.After the incubation step, the mixture was imme-diately loaded onto 6% polyacrylamide gels (acry-lamide/bisacrylamide, 29:1). Electrophoresis was per-formed in 0.5× TBE for 90 min at 25 mA. Driedgels were subjected to autoradiographyat−80◦C withintensifying screens.
Results
Cloning and characterization of theArabidopsisAthb-9and-14genes
In earlier studies, several genomic clones were iso-lated in a screen performed with a degenerate oligonu-cleotide corresponding to a conserved six amino acidsequence from the helix-3 region of Athb-1 and -2.DNA sequence analysis of the homeobox region re-vealed that two of them,Athb-8 and -9, code for aHD-Zip motif with an architecture different from thatof the Athb-1 and -2 HD-Zip domains [23].Athb-8hasbeen completely sequenced, and found to encode a 95kDa protein [2].
To characterize theAthb-9 gene, genomic se-quences flanking theAthb-9 homeobox region werecloned and sequenced by standard techniques. Usinga combination of cDNA and RT-PCR cloning, severalcDNAs were obtained (see Materials and methods).The analysis of the cDNA fragments revealed thatthe longestAthb-9 transcript is 2978 bp long. DNAsequence comparison of the full-length cDNA withthe genomic sequence showed that theAthb-9gene issplitted by 17 introns (Figure 1A). During the screen-ing of the Athb-9 cDNA we isolated two cDNAswhose sequence analysis revealed a high degree ofsimilarity with the HD-Zip coding region ofAthb-9.

615
Figure 2. Athb-8, -9 and -14 are members of a small gene family.Southern blot ofA. thaliana genomic DNA (3µg) digested withEcoRI and probed, under low- and high-stringency hybridizationconditions (L and H), with the region coding the HD-Zip domain ofAthb-8 (Hb-8), Athb-9 (Hb-9), and Athb-14 (Hb-14). The numbersindicate the size, in kb, of DNA molecular weight markers.
The corresponding genomic clone, designatedAthb-14, was cloned and sequenced by standard techniques.Several cDNA fragments were generated by RT-PCRand RACE techniques, cloned and sequenced (seeMaterials and methods). Alignment of the cDNA se-quence with the genomic sequence showed that thegene is splitted by 16 introns (Figure 1B).
Southern blot analysis ofA. thalianaDNA showedthat the HD-Zip coding regions ofAthb-9(Hb-9) and-14 (Hb-14) hybridize to singleEcoRI fragments of ca.3.6 and 3.5 kb, respectively (Figure 2). Under mod-erate hybridization and washing conditions, each ofthe probes hybridized to four different genomic frag-ments, two of which corresponding to theAthb-9and-14genes. The same analysis performed with the HD-Zip-coding region ofAthb-8(Hb-8) as a probe showedthat one of the four genomic fragments detectable un-der low-stringency hybridization conditions with theHb-9 and Hb-14 probes corresponds to theAthb-8gene (Figure 2). Southern analysis also suggested thata fourth gene related toAthb-8, -9 and-14 is containedin theArabidopsisgenome.
To investigateAthb-9and -14 gene expression inplants, RNA blot hybridization was used to study theabundance ofAthb-9and -14 transcripts in different
Figure 3. Northern blot analysis ofAthb-9and -14 expression. ADNA fragment corresponding to the 5′-untranslated region of theAthb-9cDNA (5′Athb-9/1, see Figure 1A) was used to probe 3µg ofpoly(A)+RNA isolated from green seedlings (GS, lane 1), 2-week(2W, lane 2), 3-week (3W, lane 3) and 4-week old (4W, lane 4)Ara-bidopsisplants, stems (S, lane 5), leaves (L, lane 6), and flowers (F,lane 7) of 5 week old plants, roots of 3-week-old plants (R, lane 8)and developing green siliques (Sil, lane 9). Plant growth conditionsare detailed in Materials and methods. The same blot was probedwith a DNA fragment corresponding to the 5′-untranslated regionof the Athb-14cDNA (5′Athb-14/1, see Figure 1B), with a DNAfragment corresponding to the 5′-untranslated region of theAthb-8cDNA [2], and with a cDNA for theβ-subunit ofN. plumbaginifoliamitochondrial ATPase (β-ATPase). The numbers indicate the size,in bases, of the ribosomal RNAs.
Arabidopsistissues. A blot containing poly(A)+ RNAfrom entire plants of different age and individual or-gans was probed with a 5′ end fragment of theAthb-9cDNA or with a 5′ end fragment of theAthb-14cDNA(Figure 3). For comparison is also showed the north-ern blot analysis performed with a 5′ end fragmentof the Athb-8cDNA as a probe (Figure 3) [2]. Afternormalization, we calculated that bothAthb-9and-14are slightly enriched (ca. 2-fold) in 2-week, 3-weekand 4-week old plants (Figure 3, lanes 2–4) relativeto green seedlings (Figure 3, lane 1). The analysisof RNA isolated from different organs of 5-week oldflowering plants revealed a different expression patternfor Athb-9and-14. Athb-9transcripts are slightly en-riched (2-fold) in stem with respect to leaf and flower(Figure 3, lanes 5–7) whereasAthb-14transcripts are7-fold enriched in stem (Figure 3, lane 5) and 2.5-foldin flower (Figure 3, lane 7) relative to leaf (Figure 3,lane 6). The lowest expression for bothAthb-9and-14is found in siliques (Figure 3, lane 9). In addition, hy-bridization of RNA isolated from roots of plants grownon liquid medium for 4 weeks showed that theAthb-

616
14 gene is expressed at a higher steady-state level inthis organ than in the leaf (6.5-fold enriched, Figure 3,lane 8) whereasAthb-9is expressed in root essentiallyat the same level as in leaf (Figure 3, lanes 6 and8). The expression pattern ofAthb-14during plant de-velopment is similar to that ofAthb-8(Figure 3) [2].However, theAthb-14steady-state mRNA level is ca.2-fold lower than that ofAthb-8(data not shown, seeMaterials and methods).
Athb-9and-14encode HD-Zip proteins highlyrelated toAthb-8
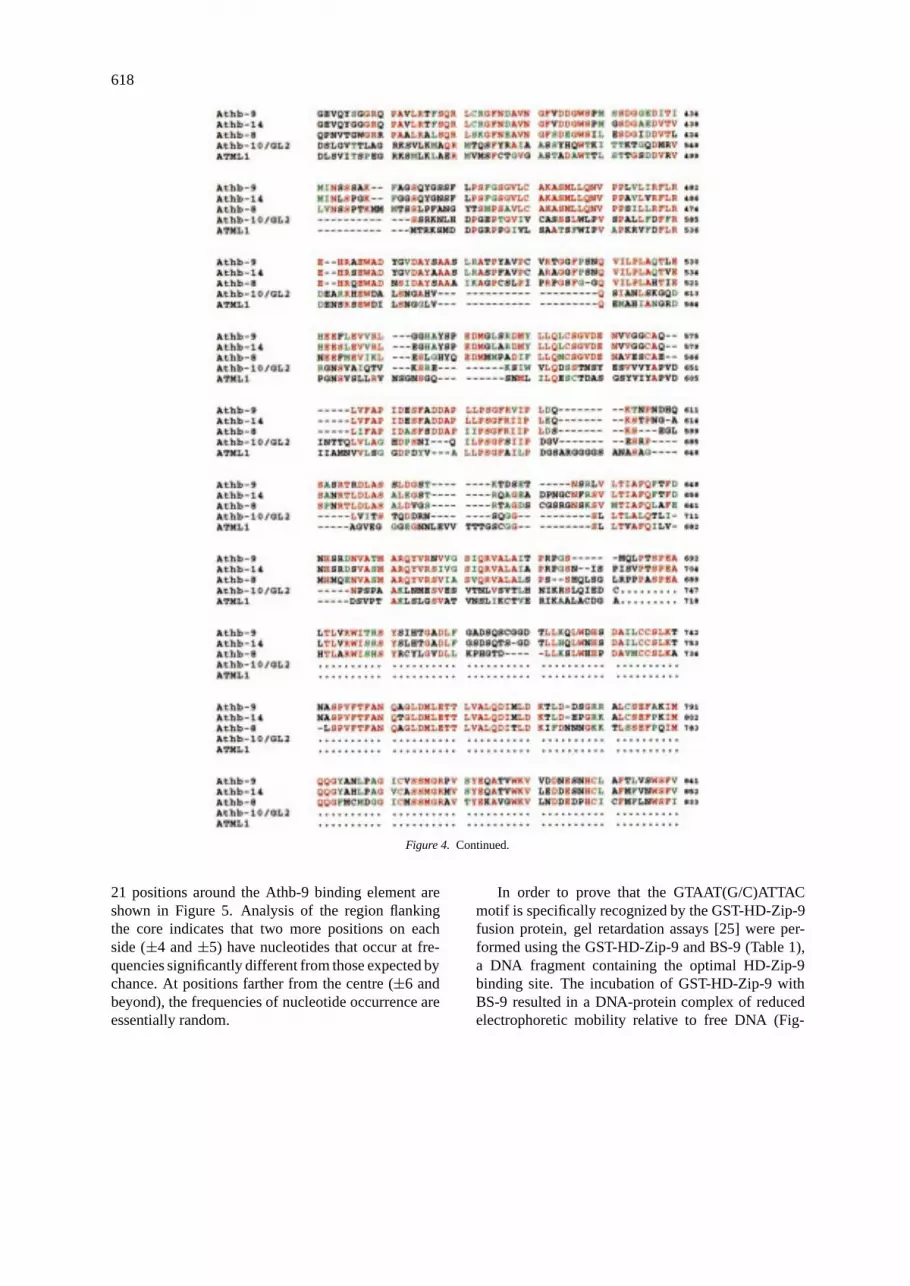
The predicted Athb-9 and -14 proteins contain 841amino acids with a calculated molecular mass of 93.7kDa and 852 amino acids with a calculated molecularmass of 94.9 kDa, respectively (Figure 4). Alignmentof amino acid sequences of Athb-9 and -14 revealed astrong homology throughout their entire length (90%similarity with 85% identity) (Figure 4). In addition,pairwise amino acid comparisons of Athb-8, -9 and -14 showed 60% identity between Athb-8 and -9 and62% between Athb-8 and -14 (Figure 4). Amino acidcomparison between Athb-9 and the proteins encodedby the open reading frames contained in the EST data-bases revealed a high degree of similarity with thepeptide encoded by the T88239 cDNA ofA. thaliana[15]. T88239 is likely to be a partial cDNA, andencodes for a peptide of 131 amino acids. Pairwiseamino acid comparisons showed that T88239 shares69% identity with Athb-8 (amino acids 658–786 inFigure 4), 54% with Athb-9 (amino acids 665–794 inFigure 4), and 53% with Athb-14 (amino acids 673–805 in Figure 4) (data not shown). On the basis ofthese analysis, we propose to include Athb-14 andT88239 together with Athb-8 and -9 in the HD-ZIPIII protein family [23]. Finally, amino acid compar-isons with other HD-Zip proteins outside the HD-Zipregion showed that proteins of the HD-ZIP III fam-ily share some homologies with Athb-10/GL2 (31%amino acid identity) and ATML1 (27% amino acididentity) (Figure 4).Athb-10/GL2, encoding for a HD-ZIP IV protein, is expressed in specific epidermal cellsin the shoot and in the root, and is required for regula-tion of trichome development and root hair formation[6, 13, 17].ATML1, encoding for another HD-ZIP IVprotein, is specifically expressed in the protodermalcells of the 16-cell embryo, and in the L1 layer ofthe meristem from the very early stages of meristempatterning throughout the complete diploid life cycleof the plant [12].
Amino acid sequence analysis also revealed a par-ticular organization of the Athb-9 and -14 HD-Zipdomains, which is identical to that observed for Athb-8 [2]. The Athb-9 and -14 HDs contain the fourinvariant amino acid residues (W48, F49, N51, R53)and seven out of eight highly conserved residues (Q12,L16, Y20, I40, V47, R55, K57) of the homeodomainconsensus sequence [8]. This conservation could beobserved only by looping out four amino acids be-tween the putative helices 2 and 3 of the Athb-9 and-14 HDs (data not shown). Moreover, as observedfor Athb-8 [2], the exact spatial register between thehomeodomain and leucine zipper found in all theproteins of the HD-ZIP I and HD-ZIP II families char-acterized so far is not conserved in the Athb-9 and -14HD-Zip domains (data not shown). The best align-ment of Athb-9 and -14 with the prototype Athb-1HD-Zip domain could be obtained by looping out fouramino acids between the homeodomain and the firstamino acid residue of the zipper (data not shown).These observations led us to investigate the DNA bind-ing properties of a member of the HD-ZIP III proteinfamily in vitro.
DNA-binding properties of the HD-Zip domain ofAthb-9
To determine whether the HD-Zip domain of Athb-9 has the ability to interact with DNA, we performedbinding site selection experiments [3, 22]. To this endthe HD and HD-Zip domains of Athb-9 (HD-9 andHD-Zip-9) were expressed in bacteria using the pGEXexpression system (see Materials and methods).E. coliextracts containing either glutathioneS-transferase(GST)-HD-Zip-9 or GST-HD-9 or GST were testedfor their ability to select binding site sequences fromthe DNA template D3, containing a 15 bp randomcore sequence [22]. GST-HD-Zip-9 binds to D3 toa significant extent with respect to GST and GST-HD-9 (data not shown). Following seven rounds ofselection and amplification, the DNA was cloned intopBluescript II KS. The sequences of 25 independentlyderived, affinity-selected DNA molecules are shownin Figure 5. Strikingly, 21 of these contain a com-mon 7 bp dyad-symmetric sequence, AAT(G/C)ATT,and the remaining 4 contain sequences differing atonly one of the seven positions. This 7 bp core se-quence was used to align all 25 sequences. By usingthe alignment in which a purine residue in the centralposition (indicated as 0 in the figure) has been cho-sen arbitrarily, the frequencies of each of the bases at

617
Figure 4. Comparison of Athb-9, Athb-14, Athb-8, Athb-10/GL2, and ATML1. The alignment was done using the PileUp program. Dasheswere introduced to maximize sequence alignment. Identical amino acids are red, conserved amino acids are green. The GenBank/EMBLaccession numbers ofAthb-9, Athb-14, Athb-8, Athb-10/GL2andATML1are Y10922, Y11122, Z50851, Z54356 and U37589, respectively.
618
Figure 4. Continued.
21 positions around the Athb-9 binding element areshown in Figure 5. Analysis of the region flankingthe core indicates that two more positions on eachside (±4 and±5) have nucleotides that occur at fre-quencies significantly different from those expected bychance. At positions farther from the centre (±6 andbeyond), the frequencies of nucleotide occurrence areessentially random.
In order to prove that the GTAAT(G/C)ATTACmotif is specifically recognized by the GST-HD-Zip-9fusion protein, gel retardation assays [25] were per-formed using the GST-HD-Zip-9 and BS-9 (Table 1),a DNA fragment containing the optimal HD-Zip-9binding site. The incubation of GST-HD-Zip-9 withBS-9 resulted in a DNA-protein complex of reducedelectrophoretic mobility relative to free DNA (Fig-

619
Figure 5. Tabulation and analysis of the HD-Zip-9 binding sitesequences present within affinity-selected DNA molecules. Thenucleotide sequences of DNA molecules cloned after the seventhround of selection are shown as aligned by the method described inthe text. Only the selected sequence of each DNA is shown. Revindicates that the reverse complementary of the selected sequence isshown. The most common nucleotides for each position between−5and+5 are indicated. The number of occurrences for the four nu-cleotides in each position is shown. SUM indicates the total numberof nucleotides in each position.
ure 6A, lanes 1 and 3). The formation of the retardedDNA-protein complex was specific: it was resistantagainst a 100-fold molar excess of unselected start-ing DNA template (D3, Figure 6A, lane 5), whereasa 100-fold molar excess of unlabelled BS-9 abol-ished the formation of the complex (Figure 6A, lane4). No DNA-protein complex was seen in the assayperformed with the labelled DNA and GST alone in-dicating that the HD-Zip-9 component of the fusionprotein was responsible for DNA binding.
To demonstrate that the HD-Zip-9 domain bindsto DNA as a dimeric complex, the ability of Athb-9 derivatives with different lengths to form mixeddimers was tested by gel retardation assay [10]. Whenthe Athb-9 derivatives (HD-ZipL-9 and HD-ZipS-9,see Materials and methods) were mixed after sep-arate translations (Figure 6B, lane 4), the peptidesgave an additional protein-DNA complex migrating
between the complexes corresponding to HD-ZipL-9(Figure 6B, lane 2) and HD-ZipS-9 (Figure 6B, lane3). This result indicates that the HD-Zip-9 domainbinds to DNA as a dimer.
Discussion
In this study, we report the isolation and characteri-zation of twoArabidopsishomeobox genes,Athb-9and -14, encoding HD-Zip proteins of 841 and 852amino acids, respectively.Athb-9and-14share a highdegree of sequence identity withAthb-8, a HD-Zipprotein encoding gene specifically expressed in pro-cambial cells of embryo and developing organs [2].Athb-9 and -14, like Athb-8, also share some simi-larities with Athb-10/GL2 [6, 17] and ATML1 [12],two proteins belonging to the HD-ZIP IV family.Athb-10/GL2is expressed in specific epidermal cells in theshoot and in the root, and is required for regulationof trichome development and root hair formation [6,13, 17]. ATML1 is specifically expressed in the L1layer of the shoot apical meristem [12]. Taken to-gether, these data strongly suggest that members ofthe HD-ZIP IV family might be involved in the reg-ulation of epidermal cell fate and differentiation. Boththe overall similarity of the HD-ZIP III and IV pro-teins and the presence of small conserved domainsoutside of the homeodomain suggest that they couldhave evolved from a common ancestor protein. Weare tempting to speculate that L1-, L2- and L3-layer-specific regulatory factors could have evolved fromthis primordial factor. It will be interesting to inves-tigate the expression pattern ofAthb-9and-14, and todetermine whether members of the HD-ZIP III familyare involved in the regulation of vascular cell fate anddifferentiation.
Like Athb-8, Athb-9 and -14 are characterized bya HD-Zip domain whose spatial organization is differ-ent from that of the Athb-1 and -2 HD-Zip domains[2, 18]. Therefore, we analysed the DNA bindingproperties of the Athb-9 HD-Zip domain. We foundthat the HD-Zip-9 domain binds as a dimer. Further-more, we determined that the HD-Zip-9 binding site isa pseudopalindrome of two 6 bp half-sites that over-lap at a central position. Our results indicate that theHD-Zip-9 domain interacts with DNA recognition el-ements in a fashion similar to the HD-Zip-1 and -2domains, and provide further experimental support tothe model of DNA binding by the HD-Zip proteinspreviously proposed [22].

620
Figure 6. The HD-Zip-9 domain specifically binds to DNA as a dimer. A. The GTAATGATTAC motif is specifically bound by the HD-Zip-9domain. Gel retardation analysis was performed as described in Materials and methods with a DNA fragment bearing the GTAATGATTACmotif (BS-9). The retardation assays were performed either with the GST protein (lane 2), or with the GST-HD-Zip-9 protein (lanes 3–5). To thereaction mixtures shown in lanes 4 and 5 a 100-fold molar excess of unlabeled competitor DNA (BS-9 in lane 4 and D3 in lane 5) was added.B. Mixed-dimer formation by derivatives of Athb-9 with different length. The Athb-9 derivatives were assayed for binding to the BS-9 probeindividually (lanes 2 and 3) and with the two products mixed after separate translation (lane 4). Note that mixed-dimer formation is observedas an additional band with intermediate mobility (lane 4). The control (lacking added RNA) is shown in lane 1.
Figure 7. Comparison of the HD-Zip domains with the HD andb-Zip domains. A. Comparison of the putative Athb-1 and -2 recog-nition helices with those of the Antp HD [16] and Fos b-Zip domain[9]. The amino acids of Antp and Fos that make the base contactsand the putative base-contacting amino acids of the Athb-1 and -2HD-Zip domains [22] are indicated in bold. B. Alignment of theputative Athb-8, -9 and -14 probe helices with those of Antp andFos. Residues of the probe helix [30] are indicated by numbers.The amino acid residues at positions 2, 7, 9, 11 and 12, whichmight be used for binding to phosphates, are boxed. The aminoacid residues which are predicted to interact with DNA bases arehighlighted in bold. The position of the intron in the Athb-8, -9, and-14 recognition helix is indicated by an arrowhead.
DNA binding studies on Athb-1 and -2 HD-Zipdomains suggested that the angle between the recog-nition helix and the dimerization domain is larger withrespect to that observed in the b-Zip-DNA complex[22]. By superimposing the recognition helices suchthat Asn-51 and Ala-11 are located in the same spa-tial orientation, we deduced the putative contacts inthe HD-Zip-DNA complex (Figure 7A) [22]. We pro-posed that conserved amino acids of helix-3 whichmake specific base contacts in HD complexes alsomake base contacts in the HD-Zip-DNA complex. Wealso suggested that Arg-55 of one of the two HD-Zipmonomers, as Arg-15 in the b-Zip-DNA complex, isresponsible for recognition of the central position ofthe pseudopalindromic site in the HD-Zip-DNA com-plex (Figure 7A) [22]. These hypothesis have beenstrongly reinforced by a mutational analysis of theAthb-2 HD-Zip domain [24].
In Athb-8, -9 and -14, the insertion of 4 aminoacids between the recognition helix and the dimer-ization domain results in a spatial organization of theHD-Zip domain identical to that of b-Zip proteins. Ithas been proposed that eukaryotic transcription factorssuch as b-Zip, HD and b-HLH proteins, which use anα-helix for DNA recognition, all share a similar 12

621
residue segment, named the probe helix [30]. Aminoacid residues at particular positions of the probe helixshould bind either to phosphates or to bases (Fig-ure 7B). Amino acid comparison of the Athb-8, -9 and-14 HD-Zip domains with the probe helix of HD andb-Zip proteins suggested that the putative Athb-8, -9and -14 probe helix is a chimeric recognition helix. Infact, most of the amino acid residues homologous toHD proteins are located in the N-terminal part of theAthb-8, -9 and -14 recognition helix, whereas aminoacid residues homologous to b-Zip proteins are locatedin the C-terminal part of the helix (Figure 7B). It isnoteworthy that an intron splits the recognition helixof all HD-ZIP III family members between Arg-53,which is invariant in the HD domains, and C-54 whichis conserved in several b-Zip proteins (Figure 7B) [11].Four out of the five putative base-contacting aminoacids identified in the probe helix of the HD-Zip-8, -9and -14 domains are conserved in the HD-Zip-1 and-2 domains and coincide with those previously pro-posed as the amino acids responsible for base contactsin the HD-Zip-DNA complex (Figure 7A). It seemslikely that, as in the HD-Zip-2 domain, the aminoacids at positions 47, 50, 51 and 55 of the HD-Zip-9 domain are essential for DNA recognition. Furtherstudies using mutant proteins will be needed to deter-mine whether the cysteine present at position 54 of theHD-Zip-9 domain plays a role in DNA binding and, ifso, whether it is involved in the recognition of position±5 of the BS-9 site.
Acknowledgements
We wish to thank Roberto Gargamelli for photo-graphic assistance. This work was supported, in part,by the Fondazione Istituto Pasteur-Fondazione CenciBolognetti, Università di Roma La Sapienza and byPiano Nazionale ‘Biotecnologie Vegetali’, Ministerodelle Risorse Agricole, Alimentari e Forestali (I.R.)and by the European Union Biotechnology Program,Contract No. BIO4-CT960217 (G.M.). G.S. and C.S.are fellows of the National Research Council of Italy.
References
1. Aoyama T, Dong C-H, Wu Y, Carabelli M, Sessa G, Ruberti I,Morelli G, Chua N-H: Ectopic expression of theArabidopsistranscriptional activator Athb-1 alters leaf cell fate in tobacco.Plant Cell 7: 1773–1785 (1995).
2. Baima S, Nobili F, Sessa G, Lucchetti S, Ruberti I, Morelli G:The expression of theAthb-8homeobox gene is restricted toprovascular cells inArabidopsis thaliana. Development 121:4171–4182 (1995).
3. Blackwell TK, Weintraub H: Differences and similarities inDNA-binding preferences of MyoD and E2A protein com-plexes revealed by binding site selection. Science 250: 1104–1110 (1990).
4. Carabelli M, Sessa G, Baima S, Morelli G, Ruberti I: TheArabidopsis Athb-2and -4 genes are strongly induced byfar-red-rich light. Plant J 4: 469–479 (1993).
5. Carabelli M, Morelli G, Whitelam G, Ruberti I: Twilight-zoneand canopy shade induction of theAthb-2homeobox gene ingreen plants. Proc Natl Acad Sci USA 93: 3530–3535 (1996).
6. Di Cristina M, Sessa G, Dolan L, Linstead P, Baima S, RubertiI, Morelli G: TheArabidopsisAthb-10 (GLABRA2) is an HD-Zip protein required for regulation of root hair development.Plant J 10: 393–402 (1996).
7. Frohman MA, Dush MK, Martin GR: Rapid production offull-length cDNAs from rare transcripts: amplification usinga single gene-specific oligonucleotide primer. Proc Natl AcadSci USA 85: 8998–9002 (1988).
8. Gehring WJ, Muller M, Affolter M, Percival-Smith A, Bil-leter M, Qian YQ, Otting G, Wuthrich K: The structure of thehomeodomain and its functional implications. Trends Genet 6:323–329 (1990).
9. Glover JNM, Harrison SC: Crystal structure of the het-erodimeric bZIP transcription factor c-Fos-c-Jun bound toDNA. Nature 373: 257–261 (1995).
10. Hope IA, Struhl K: GCN4, a eukaryotic transcriptional acti-vator protein, binds as a dimer to target DNA. EMBO J 6:2781–2784 (1987).
11. Hurst H: Protein Profile. Leucine Zippers Transcription Fac-tors, 3rd ed. Academic Press, New York (1996).
12. Lu P, Porat R, Nadeau JA, O’Neil SD: Identification of ameristem L1 layer-specific gene inArabidopsisthat is ex-pressed during embryonic pattern formation and define a newclass of homeobox genes. Plant Cell 8: 2155–2168 (1996).
13. Masucci JD, Rerie WG, Foreman DR, Zhang M, Galway ME,Marks MD, Schiefelbein JW: The homeobox geneGLABRA2is required for position-dependent cell differentiation in theroot epidermis ofArabidopsis thaliana. Development 122:1253–1260 (1996).
14. Mattsson J, Soderman E, Svenson M, Borkird C, EngstromP: A new homeobox-leucine zipper gene fromArabidopsisthaliana. Plant Mol Biol 18: 1019–1022 (1992).
15. Newman T, de Bruijn FJ, Green P, Keegstra K, KendeH, McIntosh L, Ohlrogge J, Raikhel N, Somerville S,Thomashow M, Retzel E, Somerville C: Genes galore: a sum-mary of methods for accessing results from large-scale partialsequencing of anonymousArabidopsiscDNA clones. PlantPhysiol 106: 1241–1255 (1994).
16. Otting G, Qian YQ, Billeter M, Muller M, Affolter M, GehringWJ, Wuthrich K: Protein-DNA contacts in the structure of ahomeodomain-DNA complex determined by nuclear magneticresonance spectroscopy in solution. EMBO J 9: 3085–3092(1990).
17. Rerie WG, Feldmann KA, Marks MD: TheGLABRA2geneencodes a homeodomain protein required for normal trichomedevelopment inArabidopsis. Genes Devel 8: 1388–1399(1994).
18. Ruberti I, Sessa G, Lucchetti S, Morelli G: A novel class ofplant proteins containing a homeodomain with a closely linkedleucine zipper motif. EMBO J 10: 1787–1791 (1991).

622
19. Schena M, Davis RW: HD-Zip proteins: members of anAra-bidopsishomeodomain protein superfamily. Proc Natl AcadSci USA 89: 3894–3898 (1992).
20. Schena M, Lloyd AM, Davis RW: TheHAT4 gene ofAra-bidopsisencodes a developmental regulator. Genes Devel 7:367–379 (1993).
21. Schena M, Davis RW: Structure of homeobox-leucine zippergenes suggests a model for the evolution of gene families. ProcNatl Acad Sci USA 91: 8393–8397 (1994).
22. Sessa G, Morelli G, Ruberti I: The Athb-1 and -2 HD-Zipdomains homodimerize forming complexes of different DNAbinding specificities. EMBO J 12: 3507–3517 (1993).
23. Sessa G, Carabelli M, Ruberti I, Lucchetti S, Baima S,Morelli G: Identification of distinct families of HD-Zip pro-teins inArabidopsis thaliana. In: Puigdomenèch P, Coruzzi G(eds) Molecular-Genetic Analysis of Plant Development andMetabolism, pp. 411–426. Springer Verlag, Berlin (1994).
24. Sessa G, Morelli G, Ruberti I: DNA-binding specificity of thehomeodomain-leucine zipper domain. J Mol Biol 274: 303–309 (1997).
25. Singh H, Sen R, Baltimore D, Sharp PA: A nuclear factor thatbinds to a conserved sequence motif in transcriptional con-
trol elements of immunoglobulin genes. Nature 319: 154–158(1986).
26. Smith DB, Johnson KS: Single-step purification of polypep-tides expressed inEscherichia colias fusions with glutathioneS-transferase. Gene 67: 31–40 (1988).
27. Soderman E, Mattsson J, Svenson M, Borkird C, Engstrom P:Expression patterns of novel genes encoding homeodomain-leucine zipper proteins inArabidopsis thaliana. Plant MolBiol 26: 145–154 (1994).
28. Soderman E, Mattsson J, Engstrom P: TheArabidopsishome-obox geneAthb-7 is induced by water-deficit and by abscisicacid. Plant J 10: 275–281 (1996).
29. Steindler C, Carabelli M, Borello U, Morelli G, Ruberti I:Phytochrome A, phytochrome B and other phytochrome(s)regulateATHB-2gene expression in etiolated and greenAra-bidopsisplants. Plant Cell Envir 20: 759–763 (1997).
30. Suzuki M: Common features in DNA recognition helicesof eukaryotic transcription factors. EMBO J 12: 3221–3226(1993).





























