Embed Size (px)
Citation preview

Tansley review
Modulation of plant growth by HD-Zipclass I and II transcription factors inresponse to environmental stimuli
Author for correspondence:John C. Harris
Tel: +61 883037448
Email: [email protected]
Received: 19 January 2011
Accepted: 18 March 2011
John C. Harris, Maria Hrmova, Sergiy Lopato and Peter Langridge
Australian Centre for Plant Functional Genomics, PMB 1, Glen Osmond, SA 5064, Australia
New Phytologist (2011)doi: 10.1111/j.1469-8137.2011.03733.x
Key words: abiotic stress, HD-Zip,phytohormone, plant development,transcription factor.
Summary
Plant development is adapted to changing environmental conditions for optimizing
growth. This developmental adaptation is influenced by signals from the environ-
ment, which act as stimuli and may include submergence and fluctuations in water
status, light conditions, nutrient status, temperature and the concentrations of
toxic compounds. The homeodomain-leucine zipper (HD-Zip) I and HD-Zip II
transcription factor networks regulate these plant growth adaptation responses
through integration of developmental and environmental cues. Evidence is emerg-
ing that these transcription factors are integrated with phytohormone-regulated
developmental networks, enabling environmental stimuli to influence the geneti-
cally preprogrammed developmental progression. Dependent on the prevailing
conditions, adaptation of mature and nascent organs is controlled by HD-Zip I and
HD-Zip II transcription factors through suppression or promotion of cell multiplica-
tion, differentiation and expansion to regulate targeted growth. In vitro assays
have shown that, within family I or family II, homo- and ⁄ or heterodimerization
between leucine zipper domains is a prerequisite for DNA binding. Further, both
families bind similar 9-bp pseudopalindromic cis elements, CAATNATTG, under in
vitro conditions. However, the mechanisms that regulate the transcriptional
activity of HD-Zip I and HD-Zip II transcription factors in vivo are largely unknown.
The in planta implications of these protein–protein associations and the similarities
in cis element binding are not clear.
Contents
Summary 1
I. Introduction 2
II. The role of HD-Zip transcription factors in plant growthadaptation to environmental changes and thephytohormone network
3
III. Dissecting the common cis element, dimerizationand cell specificity of HD-Zip I and HD-Zip IItranscription factors
8
IV. Conclusions 12
Acknowledgements 12
References 12
NewPhytologist Review
� 2011 Australian Centre for Plant Functional Genomics
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 1www.newphytologist.com

I. Introduction
1. Plant development is adapted to the environment
Plants face many environmental challenges during develop-ment, and they must optimize growth to adapt to theprevailing circumstances. They may also be limited by theavailability of resources in their immediate environment.Consequently, plants are sensitive to these abiotic stressesand adapt their growth to variations in water status, lightconditions, nutrient status, temperature, concentrations oftoxic compounds and submergence. These developmentaladaptations are stimulated by signals from the environment(Sultan, 2010) and involve regulation of cell multiplica-tion, differentiation and expansion through the action ofphytohormones (Wolters & Jurgens, 2009; Perrot-Rechenmann, 2010; Skirycz & Inze, 2010). Phytohormones,in turn, affect development through antagonistic and coop-erative pathways that integrate endogenous developmentaland exogenous environmental signals (Achard et al., 2006;Santner et al., 2009; Wolters & Jurgens, 2009; Perrot-Rechenmann, 2010; Stamm & Kumar, 2010). The effectsof phytohormones on growth are transduced through sig-nalling networks that induce changes in the transcriptomeand proteome (Santner et al., 2009; Stamm & Kumar,2010). The changes to the transcriptome are achievedthrough transcription factors (TFs), which repress or acti-vate suites of genes to modulate plant growth (Nakashimaet al., 2009). TFs of homeodomain-leucine zipper (HD-Zip) families I and II contribute to the plasticity of plantgrowth and are responsible for modulating plant develop-ment in response to environmental stimuli (Agalou et al.,2008). These stimuli are involved in changes that occur tothe transcriptome, together with phytohormones, throughthe control of cell differentiation, division and expansion.The transcriptional mechanisms by which the HD-Zip Iand HD-Zip II TFs manipulate growth are currentlyunknown, although it is known that members of these fam-ilies form homo- and heterodimers exclusively with othermembers of their own family as a precursor to DNA bind-ing. Also, dimers have been shown to target similar ciselements under in vitro conditions. However, the way inwhich the dimerization of the HD-Zip I and HD-Zip IITFs and the similarity of the target cis element sequencestranslate into function remains largely unexplored.
In this review, we present our account of the currentknowledge that relates to the roles of the HD-Zip Iand HD-Zip II TFs during plant adaptation underchanging environmental conditions. We also brieflydescribe how these TFs integrate with phytohormone-mediated responses, and indicate the limits of the currentknowledge that relate to the mechanism of transcriptionalactivity and address the issue of how to overcome theselimitations.
2. Discovery of the homeodomain transcription factors
Homeosis is the abnormal conversion of one body part intoanother and is the result of aberrations in the normal devel-opmental program of an organism (Eckardt, 2003). Whilehomeotic abnormalities have been observed in many differ-ent organisms (Eckardt, 2003), the first characterization ofa homeotic gene was made in Drosophila melanogaster. TheDrosophila antennapedia allele is responsible for thedevelopment of antennae in the position of the second legpair, when the gene is expressed (Garber et al., 1983).Further study established that a common region of DNA,named the homeobox (HB), is present in many genesinvolved in the developmental program of Drosophila. It isnow known that the HB encodes a DNA-binding domaintermed the homeodomain (HD). The first characterizedHB-containing gene in plants was KNOTTED1, which wasidentified in maize (Zea mays) as a mutated gene(Vollbrecht et al., 1991). The gain-of-function allele kn1induces areas of irregular cell division along secondary veinsof the lamina and the displacement of the ligule (Hakeet al., 1989). It was also found through mutagenesis studiesthat the HB-containing TENDRIL-LESS gene of Pisumsativum inhibits lamina development and is responsible forthe conversion from leaflet to tendril (Hofer et al., 2009). Astrategy used to isolate HB genes from the nematodeCaenorhabditis elegans was developed by Burglin et al.(1989) and was used to isolate HB sequences from plantcDNA libraries. This strategy was also used to isolate thefirst HB genes from Arabidopsis, which encoded a proteinwith an HD and a closely associated leucine zipper of theHD-Zip family; this association of an HD and leucine zip-per motif is unique to plants (Ruberti et al., 1991). ManyHD-Zip sequences were further isolated from rice (Oryzasativa; Meijer et al., 2000), the resurrection plantCraterostigma plantagineum (Deng et al., 2002) and wheat(Triticum aestivum; Lopato et al., 2006), using the yeastone hybrid system. With the increasing abundance ofsequenced plant genomes it is now possible to identify anentire complement of plant HD-Zip TF sequences(Mukherjee et al., 2009).
Higher plants contain four families of HD-Zip proteins(HD-Zip I–IV). An analysis of available plant genomesequences has identified these genes in flowering plants andmosses. These analyses showed that these genes are absentfrom the genomes of unicellular green and red algae(Mukherjee et al., 2009). Individual families can be distin-guished by conservation within the HD-Zip domain, othermotifs in the amino acid sequences, and also specific intronand exon positions (Ariel et al., 2007). When the phylogenyof each family is scrutinized in rice and Arabidopsis, itbecomes evident that subgroups of genes associate withineach family. These subgroups form paralogous gene pairsthat have arisen through genome duplication, as they are
2 Review Tansley reviewNewPhytologist
� 2011 Australian Centre for Plant Functional Genomics
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011)
www.newphytologist.com

associated with duplicated regions of chromosomes in theArabidopsis and rice genomes (Henriksson et al., 2005;Agalou et al., 2008; Ciarbelli et al., 2008). The phylogenyand structures of the HD-Zip I–IV genes and proteins haverecently been investigated by Henriksson et al. (2005),Agalou et al. (2008) and Ciarbelli et al. (2008).
3. Structure of the homeodomain helix-turn-helix,the role of the leucine zipper in HD-Zip proteins andthree-dimensional structures of nonplant HD andleucine zippers
The HB is a 180-nucleotide region of DNA encoding a60-amino-acid-long HD that consists of three characteristica-helices (Fig. 1; Gehring et al., 1994). a-helix 3, consid-ered the recognition helix, is the most conserved across HDproteins (Gehring et al., 1994) and is responsible for thespecificity of protein–DNA interactions. a-helix 3 typicallylies within the major groove of DNA next to the coresequence ATTA (or TAAT), bound by most HD proteins(Gehring et al., 1994). In contrast, a leucine zipper is ana-helix comprised of seven amino acid (heptad) repeats.The residues of the heptad are designated an, bn....gn (nbeing the number of the heptad), where residue d is a leu-cine. During dimerization through the monomeric a-helices, the structure forms a coiled coil (Fig. 1). Residues aand d form the hydrophobic interface of the coiled coil,while residues at positions e and g form complementarycharge interactions that permit or inhibit dimerizationbetween two monomers (Szilak et al., 1997). The leucinezipper of HD-Zip proteins is immediately downstream ofthe HD and enables dimerization of HD-Zip proteins,which is a prerequisite for DNA binding. Many HD
proteins bind strongly as monomers to DNA, but HD-Zipproteins possess a very weak affinity for DNA as monomersand require dimerization for increased DNA-binding effi-ciency (Palena et al., 1999). If the spacing between the HDand leucine zipper components of HD-Zip I and HD-ZipII proteins is modified, DNA binding is disrupted, indicat-ing that dimerization is essential for the correct spacing ofthe HDs with regard to their position on the DNA (Sessaet al., 1993). While structural information on HD-Zip pro-teins is currently unavailable, three-dimensional structuresof HDs and leucine zippers (bZIP proteins) from varioussources, mainly insect, yeast and mammalian systems, areavailable (Fig. 1). The advantage of the tandem bindingarrangements of HDs is that they provide the opportunityfor a larger and more specific DNA sequence read-out.These dimeric arrangements could create specific structuralpatterns that are recognized by DNAs.
II. The role of HD-Zip transcription factors inplant growth adaptation to environmentalchanges and the phytohormone network
In this section we will review studies of the functions ofHD-Zip I and HD-Zip II TFs, which have suggested thatthese TFs play a role in the plant response to water deficit,ABA, salinity stress and changes in light conditions.Further, the integration of HD-Zip I and HD-Zip II TFswith phytohormone pathways and plant development willbe summarized. It appears that HD-Zip I and HD-Zip IITFs contribute to the regulation of plant cell expansion,division and differentiation. The default body plan can thusbe influenced by external parameters through HD-Zip Iand HD-Zip II TFs that induce adaptation of establishedand nascent organs in response to prevailing conditions.The role of HD-Zip I and HD-Zip II TFs was determinedlargely through work with the model species Arabidopsisand rice. Making comparisons of the characteristics ofhomologous genes from these two model species has proveddifficult. Although Agalou et al. (2008) grouped closelyaligned HD-Zip I proteins from Arabidopsis, C.plantagineum and rice into clades that were established byHenriksson et al. (2005) for Arabidopsis, comparison offunctional characterizations of assumed homologs revealsdivergent roles and expression patterns (Henriksson et al.,2005; Agalou et al., 2008). Phylogenetic trees forArabidopsis and rice have been produced by combiningthe HD-Zip I and HD-Zip II proteins with the addition ofrelevant sequences from other species (Fig. 2). The cladespreviously established by Henriksson et al. (2005) andAgalou et al. (2008) for HD-Zip I and by Ciarbelli et al.(2008) for HD-Zip II have been used to help shed light onsimilar or distinct characters of the HD-Zip I and HD-ZipII paralogs and homologs within and between each species,respectively.
(a) (b)
HD-A
HD-B
bZIP-A
bZIP-B
Fig. 1 Three-dimensional structures of Drosophila and yeast proteinscontaining homeodomains (HDs) and leucine zippers (bZIPs). (a) Atandem arrangement of two HDs (HD-A and HD-B) in the Eve HD-DNA complex from Drosophila, in which each HD represents a helix-loop-helix-turn-helix structure and where both HDs bind an AT-richAATTAAATTC oligonucleotide (Hirsch & Aggarwal, 1995) (ProteinData Bank (PDB) accession 1JGG). a-helix 3 can be seen aligningwith the major groove of the B-DNA. (b) A GCN4 (General ControlNonderepressible 4) bZIP dimer from yeast (bZIP-A and bZIP-B)illustrating the interaction of the two monomeric a-helices. The basicregions of the bZip proteins bind the pseudo-palindromeAATGACTCAT ⁄ TACTGAGTA, centered on a GC base pair (in bold)(Ellenberger et al., 1992) (PDB accession 1YSA).
NewPhytologist Tansley review Review 3
� 2011 Australian Centre for Plant Functional Genomics
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011)
www.newphytologist.com

1. The role of HD-Zip transcription factors in water def-icit, salinity stress and ABA-modulated development
Plants are dependent on their environment for access towater for heat relief, photosynthesis and growth through cellexpansion (Skirycz & Inze, 2010). When a plant senses thatwater is becoming limited, it will reallocate this valuableresource by restricting transpiration and growth and willfrequently flower earlier (Skirycz & Inze, 2010). As summa-rized in this section, the role of HD-Zip I TFs in thedrought response is to mediate ABA signaling and controlgrowth by limiting cell expansion.
HD-Zip I Several specific HD-Zip I family members havebeen characterized with respect to their roles in regulatingdrought responses. Of the Arabidopsis c-clade (Fig. 2),AtHB7 and AtHB12 transcripts are expressed in youngexpanding tissues and transcript levels are dramaticallyincreased, especially in the vasculature, upon application of
exogenous ABA or water deficit (Olsson et al., 2004). Whenover-expressed in Arabidopsis, AtHB7 and AtHB12 confer areduced growth phenotype typical of water-limiting condi-tions (Olsson et al., 2004). Conversely, it has been observedthat athb12 mutant plants have slight but reproducibleincreases in stem length (Son et al., 2010). Hjellstrom et al.(2003) have shown that retardation of stem growth inAtHB7 over-expressing plants is caused by decreased cellelongation. In AtHB12 over-expression lines, stem retarda-tion is associated with down-regulation of GA20 oxidase 1activity, which is responsible for GA-stimulated cell elong-ation (Son et al., 2010). It has been proposed that the roleof AtHB7 and AtHB12 is to reduce growth under water defi-cit, and the inhibition of GA biosynthesis is consistent withthis role (Hjellstrom et al., 2003; Olsson et al., 2004; Sonet al., 2010).
The sunflower (Helianthus annuus) HB4 (HaHB4) geneclusters within the drought ⁄ ABA inducible c-clade (Fig. 2)and is strongly induced by water deficit and ABA (Gago
0.1
OsHOX26
OsHOX18
HAT14
OsHOX11
OsHOX27
1000
OsHOX2
OsHOX28
999
533
OsHOX17
OsHOX1AtHB4HAT3
1000
AtHB2/HAT4
HAT1HAT2
1000
719
840668
840
HaH
B10
CpH
B2
HAT
22
HAT
91000
704
992
OsH
OX
15
OsH
OX19
986
528
OsH
OX3
AtHB17
AtH
B18
903
995
OsHOX7
CpHB1
729
877
699976
AtHB1/HAT5
CpHB5
883
OsHOX25
OsHOX16
OsHOX5
703672
1000
CpHB4
AtHB5
AtHB16
AtHB6 1000 664
1000AtHB13
AtHB23
1000
OsHOX21
OsHOX23
1000
982
AtHB20AtHB3/HAT7
1000
1000
AtHB52
AtHB54
946
CpHB6CpHB7
1000
520
HaHB4AtHB12
AtHB7
1000
MtHB1
NaHD20
969
995
999
OsHOX6
OsHOX22
OsH
OX24
999
586884
AtHB53AtHB21
AtHB40
999
999
OsHOX12
OsHOX14
9991000
AtHB22AtHB51
1000
OsHOX13
OsHOX8
931
OsHOX20OsHOX4
1000
1000
1000
999
1000
δδϕϕ
α
β
β
ζ
ε
γ
αβ
γδ
Fig. 2 An unrooted radial phylogenetic tree of the selected homeodomain-leucine zipper (HD-Zip) I and HD-Zip II transcription factors. Thetree is based on full-length amino acid sequences. The alignment was created using CLUSTALX (Thompson et al., 1997) and branch lengths aredrawn to scale. HD-Zip I clades are circled and identified as a, b1, b2, c, d, f, u1 and u2 and HD-Zip II clades as a, b, c and d, as previouslyestablished (Henriksson et al., 2005; Agalou et al., 2008; Ciarbelli et al., 2008). Two-letter prefixes for sequence identifiers indicate speciesof origin. At, Arabidopsis thaliana; Cp, Craterostigma plantagineum; Mt, Medicago truncatula; Na, Nicotiana attenuata; Os, Oryza sativa;HB, homeobox; HOX, homeobox. The tree was bootstrapped using the N-J algorithm (Thompson et al., 1997) and bootstrap values forreproducibility out of 1000 are shown at the confluence of the clusters, for which these values were > 500. The bar indicates substitutions per site.
4 Review Tansley reviewNewPhytologist
� 2011 Australian Centre for Plant Functional Genomics
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011)
www.newphytologist.com

et al., 2002). Work with two forms of the HaHB4 pro-moter identified an ABA response element (ABRE) thatresponds to ABA application and also an ABRE thatresponds to NaCl treatment via an ABA-independentmechanism (Manavella et al., 2008a). When over-expressedin Arabidopsis, HaHB4 confers a similar reduced growthphenotype as seen when AtHB7 and AtHB12 are over-expressed (Olsson et al., 2004; Dezar et al., 2005). Whentested under drought stress, Arabidopsis plants expressingHaHB4 constitutively or under a drought-inducible pro-moter showed increased survival through a mechanism thatinhibited drought-related senescence (Dezar et al., 2005;Manavella et al., 2006). Arabidopsis plants over-expressingHaHB4 that were grown under well-watered conditions alsoshowed a delay in the developmental senescence that occursat the end of the life cycle (Manavella et al., 2006).Microarray and Q-PCR data suggested that the role ofHaHB4 in Arabidopsis and H. annuus is to suppress theexpression of ethylene-related genes and delay senescence. Itwas then found that the increasing levels of HaHB4 expres-sion in sunflower correlated with the developmental age ofleaves, as older leaves approached senescence (Manavellaet al., 2006). It was concluded that HaHB4 was delayingthe onset of senescence in Arabidopsis under drought stressand also acting as an antagonist to developmental senes-cence in both species. A delay in senescence has not beenreported in transgenic Arabidopsis over-expressing AtHB7and AtHB12 genes under well-watered or water-deficit con-ditions, making it difficult to compare the roles of thesegenes in the two species.
The Medicago truncatula HD-Zip I member HB1(MtHB1), belonging to the c clade, (Fig. 2) is induced byABA and salinity stress in roots and plays a role in lateralroot emergence (Ariel et al., 2010). When MtHB1 is over-expressed in the roots of composite plants, the primary rootis longer and lateral root emergence is reduced, a phenotypetypically seen in wild-type plants exposed to severe saltstress. Two TILLING-derived mthb1 mutant lines showeda reciprocal phenotype and had shorter roots with anincreased number of emerged lateral roots (Ariel et al.,2010). The molecular mechanism responsible for the lateralroot emergence phenotypes involves suppression of LOB-binding domain 1 (LBD1) by MtHB1. The lateral organboundaries (LOB) domain TFs of Arabidopsis and rice playa role in auxin-regulated lateral root initiation and adventi-tious root formation, respectively (Liu et al., 2005;Okushima et al., 2007). It is proposed that the role ofMtHB1 is to inhibit lateral root emergence, when roots areexposed to adverse conditions, which reduces the surfacearea exposed to soil stress (Ariel et al., 2010).
NaHD20, a Nicotiana attenuata c-clade homolog, is alsoinduced in roots and leaves by exogenous ABA and soilwater deficit (Re et al., 2011). It was observed that underwater stress N. attenuata plants with reduced NaHD20
transcripts, through virus-induced gene silencing, hadreduced levels of ABA, N. attenuata 9-cis-epoxycarotenoiddioxygenase 1 (NaNCED1; an ABA biosynthesis gene) andN. attenuata osmotin 1 (NaOSM1) but that N. attenuatalipid transfer protein 1 (NaLTP1) induction was notaffected (Re et al., 2011).
OsHOX6 (O. sativa Homeobox 6), OsHOX22 andOsHOX24, which are considered the rice homologs ofAtHB7 and AtHB12 (Fig. 2), are also up-regulated by waterdeficit but there are no reports clarifying the functional rolesof the corresponding proteins in the rice drought adaptationresponse (Agalou et al., 2008). These rice genes differ fromtheir Arabidopsis counterparts in their basal expression pat-terns and there are also differences between the rice paralogsthemselves. Under well-watered conditions, OsHOX6 has arelatively high basal level of expression in all tissues, whileOsHOX22 transcript is expressed mainly in the blade andpanicles. Further, OsHOX24 is strongly expressed in pani-cles and weakly in other tissues (Agalou et al., 2008).Under extended drought, OsHOX22 and OsHOX24 tran-scripts are increasingly up-regulated in leaf tissues in bothdrought-resistant and drought-sensitive cultivars, whereasOsHOX6 is only slightly up-regulated in a drought-sensitivecultivar (Agalou et al., 2008).
The Arabidopsis HD-Zip I b-clade gene AtHB6 (Fig. 2)is ubiquitously expressed in all tissues of mature plants, butin leaves it is detected predominantly in the vasculature(Soderman et al., 1999). AtHB6 is thought to play a role inthe regulation of cell division or differentiation as its expres-sion in developing leaves declines basipetally, withdrawingwith the wave of epidermal cell differentiation, but AtHB6still remains in the guard cells (Soderman et al., 1999).Expression of AtHB6 in Arabidopsis is increased by waterdeficit and ABA, but remains within the same cell types asunder untreated conditions (Soderman et al., 1999). WhenAtHB6 is over-expressed in transgenic Arabidopsis, plantsshow reduced stomatal closure and diminished inhibitionof germination by ABA (Himmelbach et al., 2002); theseare also characteristics of the ABA-insensitive mutants abi1and abi2 (Leung et al., 1997). These data imply thatAtHB6 plays a role as a negative regulator in the ABAresponse under water deficit (Deng et al., 2006).
AtHB5 of the b clade (Fig. 2) shows a similar expressionpattern to AtHB6 and is found in all major tissues(Soderman et al., 1994; Henriksson et al., 2005). TheAtHB5 promoter is active in the hypocotyl of germinatingseedlings, but is suppressed by ABA application. After ABAtreatment its expression is restricted to a discrete band inthe transition zone between the hypocotyl and root(Johannesson et al., 2003). Transgenic Arabidopsis withectopically increased levels of AtHB5 are more sensitive toroot growth inhibition by ABA at the seedling stage and togermination inhibition. The two observations, that AtHB5is down-regulated by ABA and that increased levels of ABA
NewPhytologist Tansley review Review 5
� 2011 Australian Centre for Plant Functional Genomics
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011)
www.newphytologist.com

enhance ABA-specific responses, have led the authors tosuggest a role in seedling development under short-termwater-limiting conditions that is not sustained throughlong-term water deficit (Johannesson et al., 2003).
The HD-Zip I TFs of C. plantagineum that group withthis b clade under phylogenetic analysis (Fig. 2), CpHB4and CpHB5, are down-regulated in C. plantagineum leavesand roots upon exposure to water deficit, whereas CpHB6and CpHB7 are up-regulated (Deng et al., 2002). Undernormal growth conditions, CpHB7 promoter activity inArabidopsis is considered comparable to that of AtHB6. TheAtHB6 over-expression phenotype is also paralleled by ecto-pic expression of CpHB7 in Arabidopsis (Deng et al., 2006).
OsHOX4 of the rice f clade (Fig. 2) is also droughtresponsive but the level of mRNA transcripts decreasesupon exposure to water deficit (Agalou et al., 2008). Thereare contrasting reports regarding the characteristics of trans-genic rice over-expressing OsHOX4. While transgenicplants are shorter than wild-type plants, this phenotype isreported to be attributable to a decrease in the number of cellsper internode (Agalou et al., 2008) or to a reduction in cellelongation in the stem (Dai et al., 2008). The mechanismbehind stem height reduction is thought to involve OsHOX4up-regulation of YAB1 (YABBY1, encodes a protein with aYABBY domain) expression, a negative regulator of the GAresponse (Dai et al., 2008). These results, when interpretedin isolation, imply that under water deficit OsHOX4 isdown-regulated, leading to a decrease in YAB1 expression,which enables a stronger plant response to GA. However, theimplication of a drought response mechanism involvingOsHOX4 in the adaptation of rice development is unclear.
The transcript levels of the Arabidopsis d-clade genes(Fig. 2) AtHB21, AtHB40 and AtHB53 are up-regulatedupon exposure to ABA and salinity stress. However, no rolein environmental adaptation has been established for theseHD-Zip I TFs, although a role in ovule development hasbeen proposed for all three genes (Skinner & Gasser, 2009).
HD-Zip II There is little functional evidence to suggest arole for HD-Zip II TFs in plant growth adaptationresponses to water deficit. However, expression studiesusing microarrays have shown that HAT2 (Homeobox fromArabidopsis thaliana 2) and HAT22 expression is up-regu-lated under drought in Arabidopsis (Huang et al., 2008).
In rice, OsHOX11 transcripts are dramatically decreasedupon drought exposure in a drought-resistant cultivar andOsHOX27 is up-regulated under mild drought, but tran-script levels decrease as the severity of the water deficitincreases in both drought-resistant and drought-sensitivecultivars (Agalou et al., 2008). OsHOX19 expression isincreased by the imposition of drought stress in bothdrought-sensitive and drought-resistant cultivars.
Transcripts of HD-Zip II TFs from C. plantagineum arealso regulated by water availability and ABA. CpHB1 and
CpHB2 show tissue-specific differences in expression underwater deficit and ABA treatment. CpHB1 expression isinduced in leaves by water deficit but not by ABA, and thelevel of CpHB2 transcript is up-regulated in the roots bywater deficit and ABA (Deng et al., 2002).
2. The role of HD-Zip transcription factors in plantgrowth adaptation to light and their expression andfunction
Light stimulates germinating seedlings to de-etiolate andtriggers the change to a light-harvesting photoautotroph(Franklin & Quail, 2010). Once a plant is harvesting light,the red : far-red ratio (R : FR) is an indicator of shade orcompetition for light from neighboring plants (Franklin,2008). During this post-germinative stage, FR-rich lightacts as a stimulus that is interpreted by the phytochromesignaling system to induce the shade avoidance response(Stamm & Kumar, 2010). Blue light is perceived by crypto-chromes and phototropins, which regulate stem elongation,floral induction, the circadian clock and phototropism (Lin& Shalitin, 2003; Inoue et al., 2010; Moglich et al., 2010).
HD-Zip I Of the four a-clade HD-Zip I TFs (Fig. 2),neither AtHB13 nor AtHB20 transcripts were detected inplants grown in darkness (Henriksson et al., 2005) but theyappear to have distinct roles. AtHB13 is implicated insucrose-responsive development (Hanson et al., 2001),while AtHB20 is involved in vascular patterning and inte-gration of light with internal ABA signals during seedgermination (Mattsson et al., 2003; Barrero et al., 2010).AtHB20 is involved in vascular patterning in leaves and isassociated with areas around emerging procambial strandsin very early leaf primordia. Expression of AtHB20 in devel-oping leaves mirrors that of auxin localization and, as leavesmature, transcripts are localized to the vasculature and even-tually to the fascicular cambium (Mattsson et al., 2003). Ithas also been found that AtHB20 is involved in germinationand is thought to inhibit the action of ABA near the micro-pyle (Barrero et al., 2010). During germination, themicropyle weakens and allows the root cap to emerge, andABA, which inhibits germination, inhibits this weakening.AtHB20 was shown to be expressed in the endosperm at themicropylar end and also in the root cap (Barrero et al.,2010). When seeds were germinated under light, there wasan increase in the transient expression of AtHB20 3 h afterimbibition. Barrero et al. (2010) suggested that AtHB20acts as an integrator of internal ABA signaling and externallight signaling, which is required for germination ofArabidopsis seeds.
Like all members of the Arabidopsis b1 and b2 clades(Fig. 2), AtHB1 and AtHB16 transcripts are expressed ubiq-uitously, but unlike AtHB5 and AtHB6 they are up-regulated when Arabidopsis plants experience periods of
6 Review Tansley reviewNewPhytologist
� 2011 Australian Centre for Plant Functional Genomics
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011)
www.newphytologist.com

darkness (Henriksson et al., 2005). When ectopicallyover-expressed, AtHB1 overrides the etiolation response ofdark-grown seedlings and results in defects in palisade cellformation, suggesting a role in leaf cell fate determination(Aoyama et al., 1995). AtHB16 negatively affects hypocotylelongation in response to blue light. When mRNA over-expression or antisense transgenic lines are exposed to bluelight, the transcript levels are negatively correlated with thehypocotyl elongation response, when compared with wild-type plants (Wang et al., 2003). AtHB16 expression, how-ever, is not affected by blue light or CRYPTOCHROME1(CRY1) and CRY2; neither does it affect CRY1 or CRY2expression. Under white light (WL) conditions, AtHB16 ishighly expressed in leaves and acts as a negative regulator ofleaf cell expansion. This response was confirmed by anincrease in leaf size in both directions upon mRNA anti-sense transcript suppression (Wang et al., 2003).
The Arabidopsis HD-Zip I gene AtHB52, for whichthere is little functional information, also shows a strongup-regulation of transcript level in response to blue lightand especially darkness (Henriksson et al., 2005). Large-scale expression analyses under different light regimes havenot been reported for the rice HD-Zip I family.
HD-Zip II Many of the HD-Zip I genes appear to be regu-lated by water deficit or light conditions. By contrast, five ofthe Arabidopsis HD-Zip II genes are known to respond tochanges in light, but there is little evidence to imply a role inadaptation to water stress. Little is known about the environ-mental conditions regulating the remaining five HD-Zip IImembers other than that they are not responsive to changesin light (Ciarbelli et al., 2008). Of the three c-cladeArabidopsis HD-Zip II genes (Fig. 2), AtHB2 ⁄ HAT4 hasbeen investigated most thoroughly. This gene is highlyexpressed in dark-grown etiolated seedlings (Carabelli et al.,1996) and is strongly inhibited upon exposure to either R orFR light. AtHB2 ⁄ HAT4 mRNA antisense and over-expres-sion lines have shorter and longer hypocotyls, respectively.The biological role of the corresponding protein during ger-mination appears to be in the regulation of hypocotylelongation, which is dependent upon seed reserves, untillight is sensed, at which time elongation can cease and theplant develops autotrophic capabilities. Over-expressionlines show smaller cotyledons through decreased cell expan-sion, which supports a role in determining developmentaldifferences between heterotrophic and autotrophic habits(Steindler et al., 1999). Once development has proceededpast germination, AtHB2 ⁄ HAT4 is expressed at all stages ofleaf development and increases with leaf age (Ciarbelli et al.,2008). When plants are grown under WL, AtHB2 ⁄ HAT4 isstrongly, but reversibly, induced by low R : FR light condi-tions (Steindler et al., 1997), which are typical under canopyshade. This leads in turn to the induction of a shade avoid-ance response, with plant characteristics such as shoot
elongation and a reduction of leaf size. When over-expressedunder WL conditions, AtHB2 ⁄ HAT4 decreases leaf size byrepressing its cell division.
Ectopic over-expression of the two remainingArabidopsis c-clade HD-Zip II genes, HAT1 and HAT2(Fig. 2), results in a phenotype similar to that seen inAtHB2 ⁄ HAT4 over-expression lines (Steindler et al., 1999;Sawa et al., 2002; Ciarbelli et al., 2008). Of the three mem-bers of the c-clade, HAT1 is more closely related to HAT2than AtHB2 ⁄ HAT4, but functionally has more similaritywith AtHB2 ⁄ HAT4. Both the AtHB2 ⁄ HAT4 and HAT1genes are rapidly up-regulated by FR-rich light, presumablythrough the direct action of the phytochrome system(Ciarbelli et al., 2008). The HAT2 gene is also up-regulatedby low R : FR light but is delayed, compared withAtHB2 ⁄ HAT4, as a result of regulation by an auxin-mediated pathway that is induced by FR-rich light(Ciarbelli et al., 2008). HAT1 over-expressing lines sharean identical phenotype with AtHB2 ⁄ HAT4 over-expressors,and HAT2 over-expressors have a similar phenotype butshow additional features that are characteristic of IAA over-producing mutants such as epinastic cotyledons (Sawaet al., 2002). The reduction in leaf size seen in the over-expression phenotype of the three c-clade genes has beenattributed to a reduction in the number of cells via reducedcell division (Ciarbelli et al., 2008).
The sunflower HaHB10 gene is strongly up-regulated indark-grown seedlings and is expressed mainly in maturephotosynthetic organs of sunflower (Rueda et al., 2005).HaHB10 was considered to be closely related toAtHB2 ⁄ HAT4 by Ariel et al. (2007), because of similarresponses to light and their apparent roles in adaptation tochanges in light conditions. However, the extreme C-termi-nal end of the HD-Zip II protein shares a near-identicalamino acid sequence with HAT22 and HAT9 of theArabidopsis HD-Zip II b clade (our own observation),although the expression patterns of these genes show noresponse to changes in light quality (Ciarbelli et al., 2008).
As with members of the c clade, the d-clade genes AtHB4and HAT3 (Fig. 2) are also induced by an increase in FRlight (Ciarbelli et al., 2008). The phenotype of Arabidopsisover-expressing AtHB4 under continuous WL is also similarto that of c-clade over-expression lines (Sorin et al., 2009).However, unlike the response of plants over-expressing thec-clade genes, it was shown that over-expression of AtHB4can inhibit FR-induced hypocotyl elongation, when com-pared with the wild type.
3. Integration of endogenous and environmentalsignaling through phytohormones and the HD-Zip Iand HD-Zip II transcription factors
Coordination of plant development involves the integrationof many factors, including those involved in interactions of
NewPhytologist Tansley review Review 7
� 2011 Australian Centre for Plant Functional Genomics
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011)
www.newphytologist.com

plant phytohormones and HD-Zip I and HD-Zip II TFs.This coordination involves regulation, at different levels, ofphytohormones and HD-Zip TFs, separately or in combi-nation. The complexity of phytohormone regulation of anHD-Zip I TF is seen in the induction process of AtHB53,which is up-regulated by NaCl and ABA in seedlings andin the root meristem and elongation zone by auxin. Up-regulation in the roots by ABA is inhibited by co-applica-tion of cytokinin (Son et al., 2005). Demonstrating theinfluence of HD-Zip I TFs on phytohormone signaling andintegration with environmental signals, OsHOX4 is sup-pressed under water deficit and over-expression results inectopic expression of YAB1, a negative regulator of the GAresponse (Dai et al., 2008). Through ABA, the HD-Zip Idrought-inducible c clade has been shown to influencewater deficit by inhibiting the action of GA and auxin.AtHB12 is up-regulated by ABA and down-regulates GA20oxidase 1 in Arabidopsis (Son et al., 2010). Medicagotruncatula HB1 is induced by ABA and NaCl in roots andsuppresses auxin-mediated LBD1 expression (Ariel et al.,2010). Integrating water deficit, through ABA, with ethyl-ene signaling, HaHB4 suppresses senescence under drought(Manavella et al., 2006). By integrating pathways via planthormones that are considered to be abiotic and biotic stressrelated, HaHB4 expression could also be induced by appli-cation of methyl jasmonate, while NaHD20 is induced byABA, jasmonic acid, salicylic acid and ethephon (whichreleases ethylene; Manavella et al., 2008b; Re et al., 2011).Of the HD-Zip II TFs involved in de-etiolation and theshade avoidance response, many TFs were shown to requireor inhibit the action of phytohormones. To fulfill its role inthe shade avoidance response, the AtHB2 gene requires theauxin transport system for cell elongation in the hypocotyl(Steindler et al., 1999), and HAT2 is induced under FR-rich light conditions but via an auxin-mediated pathway(Sawa et al., 2002). AtHB4 regulates hypocotyl length inthe seedling shade avoidance response and down-regulatesauxin and ⁄ or brassinosteroid induction of genes that arealso up-regulated by FR-rich light (Sorin et al., 2009).HaHB10 is also involved in plant growth adaptation tolight conditions and expression is strongly induced in thehypocotyls of dark-grown seedlings and by GA (Ruedaet al., 2005). There is also evidence that HaHB10 is up-regulated by the biotic stress-related salicylic acid and that itdown-regulates salicylic acid biosynthetic genes, althoughlevels of the phytohormone itself are not affected undercontrol conditions (Dezar et al., 2010).
III. Dissecting the common cis element,dimerization and cell specificity of HD-Zip Iand HD-Zip II transcription factors
HD-Zip I and HD-Zip II TFs cis are involved in adaptingplant growth to changes in environmental conditions. The
mechanism by which they exert their control over plantdevelopment has not yet been identified. HD-Zip I andHD-Zip II TFs are perceived to bind common cis elementsthat were revealed using in vitro methods (Frank et al.,1998; Meijer et al., 2000; Deng et al., 2002; Lopato et al.,2006). It was also shown in vitro that dimerization is a pre-requisite for DNA binding and it is now assumed that HD-Zip I and HD-Zip II TFs can heterodimerize within theirown families (discussed in section III). These observationssuggest that a large network exists, where dimerization part-ners confer different transcriptional characters and twofamilies compete over similar cis elements. This wouldenable environmental and endogenous signals to regulatefluxes in the network to establish developmental programsthrough differential gene expression (Fig. 3). We haverecently found that two members of the HD-Zip I andHD-Zip II families can antagonistically regulate a group ofgenes involved in the same developmental process (S.Lopato et al., unpublished data). Whether this is direct orindirect regulation through the same cis element remains tobe demonstrated. However, to date, very few data have beenpresented that demonstrate the effects of the protein–DNA
Cell expansion, division, differentiation
Adaptation of plant growth to the environment
Post-translational modification1
CAATNATTG Downstream gene
HD-Zip I and HD-Zip II TFs
homo- and heterodimerise strictly within
families
Modulation of TF expression
Developmental stimuli
Abiotic stress
Other TFs
Environmental stimuli
Fig. 3 The proposed plant developmental regulatory networkinvolving homeodomain-leucine zipper (HD-Zip) I and HD-Zip IItranscription factors (TFs). Black arrows indicate an interaction. Theblack dashed line represents a promoter, the cross-hatched boxrepresents cis elements downstream of other trans-acting factors,the CAATNATTG box represents the HD-Zip I and HD-Zip II cis
elements, the right-angled arrow represents the transcriptional startsite, and the ‘Downstream gene’ box represents genes whosetranscription will be either suppressed or activated. 1The arrowedbox represents post-translational modification that affects the abilityof the HD-Zip TFs to dimerize and ⁄ or bind DNA (Himmelbach et al.,2002; Tron et al., 2002).
8 Review Tansley reviewNewPhytologist
� 2011 Australian Centre for Plant Functional Genomics
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011)
www.newphytologist.com

and protein–protein interactions in planta. Problems hin-dering validation of this network will be addressed here.
1. Defining target cis elements within downstreamgenes
When the specificity of HD-Zip I and HD-Zip II ciselement-binding interactions has been investigated in vitro,the results have generally yielded the sequence CAAT(A ⁄ T)ATTG (binding site 1 (BS1)) or CAAT(C ⁄ G)ATTG (BS2). It is considered that BS1 and BS2 arecomposed of two overlapping yet different HD-related ciselements composed of 5¢-TAATTG-3¢ and 3¢-GTTATT-5¢, or 5¢-TGATTG-3¢ and 3¢-GTTACT-5¢, respectively(the difference in the central nucleotide is highlighted inbold). The consequence of this is that each HD of a dimeris interacting with a different BS, depending on the orienta-tion of the dimer relative to the DNA. Also, only one HDof any dimer makes specific contacts with the central nucle-otide and each monomer has specific preferences for theorientation of a cis element (Tron et al., 2005). Key aminoacid residues in the HD which contribute to binding at thecentral nucleotide are conserved in members within eachfamily (Sessa et al., 1997). However, it is now evident thatthese conservations are not solely responsible for the distinc-tion between the BSs. There are many examples ofmembers of each family that bind to both BS1 and BS2sequences in vitro, albeit with varying degrees of efficiency(Frank et al., 1998; Meijer et al., 2000; Deng et al., 2002;Lopato et al., 2006). Also, AtHB2 is known to regulate itsown promoter and DNA footprinting assays confirmed thatit can interact strongly with TAATCATTA andTAATTATTA, but also weakly with TAATCATCT (devia-tions from the typical BS are underlined; Ohgishi et al.,2001). Therefore, while it is considered that there are twodifferent cis elements, clearly there are ambiguities. A moredefinitive study has been performed on the DNA-bindingproperties of MtHB1, which negatively regulates LBD1expression (Ariel et al., 2010). Initial gel shift assays con-firmed that MtHB1 interacted with both BS1 and BS2.Mutation analysis of BS1, found in the promoter of theLBD1 gene, showed that BS1 was necessary to maintain theusual spatial expression pattern regulated by MtHB1 (Arielet al., 2010). ChIP-PCR (Chromatin immuno precipita-tion PCR) assays confirmed that MtHB1-histidine fusionproteins could indeed associate with the LBD1 promoter.Establishing the promoters that are directly regulated byHD-Zip I and HD-Zip II TFs will enable the precise iden-tification of target cis elements. This can be achievedthrough transcriptomic approaches that identify genes thatare co-regulated with HD-Zip I and HD-Zip II transcriptsunder specific induction conditions, or through up-regula-tion of potential target genes by an ectopically expressedHD-Zip I or HD-Zip II TF. Once potential downstream
genes have been revealed, investigations of their promoterregions can be carried out to identify these cis elements andvalidate them using mutations and promoter activationstudies. Performing ChIP-PCR will confirm that the HD-Zip TF of interest interacts with the suspected promoterunder in vivo conditions.
Clearly, plant HD-Zip I and II proteins have similarDNA-binding selectivity but current models that defineHD-Zip–DNA interactions are based on comparisonsbetween HD proteins from insect or mammalian sources,structures of which have been determined by X-ray crystal-lography and NMR spectroscopy (Kissinger et al., 1990;Qian et al., 1993). The atomic structure of a fruit-fly HDprovided the rationale for site-directed mutagenesis experi-ments in plant dimeric HD-Zip TFs. Targeted amino acidresidue substitutions of the N-terminal arm led to signifi-cant changes in DNA-binding affinities of HaHB4 (Palenaet al., 2001) and enabled determination of the molecularbasis of DNA binding that is assigned to helix III (recogni-tion helix) and to the loop between helix I and helix II ofHaHB4 (Tron et al., 2004). Access to the atomic structureof this particular HD-Zip TF would provide more accuratealterations in DNA-binding affinities and would shed lighton the structural basis of oligomerization patterns. Moreatomic structures, in particular those of plant HD-Zip TFs,are needed to establish the precise roles of the structuralelements that guide DNA binding and dimerization.
2. Regulation of cis element binding through post-translational modification
It has been shown using in vitro assays that binding of targetcis elements by HD-Zip I and HD-Zip II TFs may be regu-lated by post-translational modification. The redox status ofa plant cell changes to an oxidative environment understress. Implicating cellular redox status in post-translationalcontrol of HD-Zip II TF DNA binding, the conservedcysteines at a2, a3 and g2 of the leucine zipper and the con-served CPSCE motif (a highly conserved string of fiveamino acids consisting of Cys, Pro, Ser, Cys and Glu foundin the C-terminus) inhibit DNA binding in an oxidativeenvironment (Tron et al., 2002). The phosphorylationstatus of AtHB7 and AtHB12 has been suggested as a formof post-translational control that may explain the inabilityof these HD-Zip I TFs to bind either BS1 or BS2 in vitro(Johannesson et al., 2001). However, AtHB7 is able to ho-modimerize and AtHB7 and AtHB12 can activate areporter construct with six repeats of BS1 (Johannessonet al., 2001; Henriksson et al., 2005). Although Son et al.(2010) found that they could not determine AtHB12 bind-ing to the promoter of GA20 oxidase 1 in vitro, AtHB12over-expression did inhibit activation of the GA20 oxidase1 promoter in transient assays. Himmelbach et al. (2002)found that ABI1 (ABA Insensitive), a serine ⁄ threonine
NewPhytologist Tansley review Review 9
� 2011 Australian Centre for Plant Functional Genomics
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011)
www.newphytologist.com
phosphatase, interacts with AtHB6 in yeast-2-hybrid assays.A role for dephosphorylation in activating AtHB6 transcrip-tional activity was revealed by the observation thatphosphorylation of AtHB6 by protein kinase A inhibitedDNA-binding activity. Similarly, in transient assays, AtHB6activated a promoter with four repeats of the HD-Zip bind-ing sequence more strongly when ABA was applied,suggesting dephosphorylation by an ABA-dependent factor(Himmelbach et al., 2002). These in vitro observationshave demonstrated that post-translational regulation caninfluence DNA binding.
3. Dimerization and the roles of members of the HD-Zip I and HD-Zip II families in the cell- and condition-specific interactome
Currently it is assumed that each HD-Zip I and HD-Zip IITF is able to heterodimerize only with other members of its
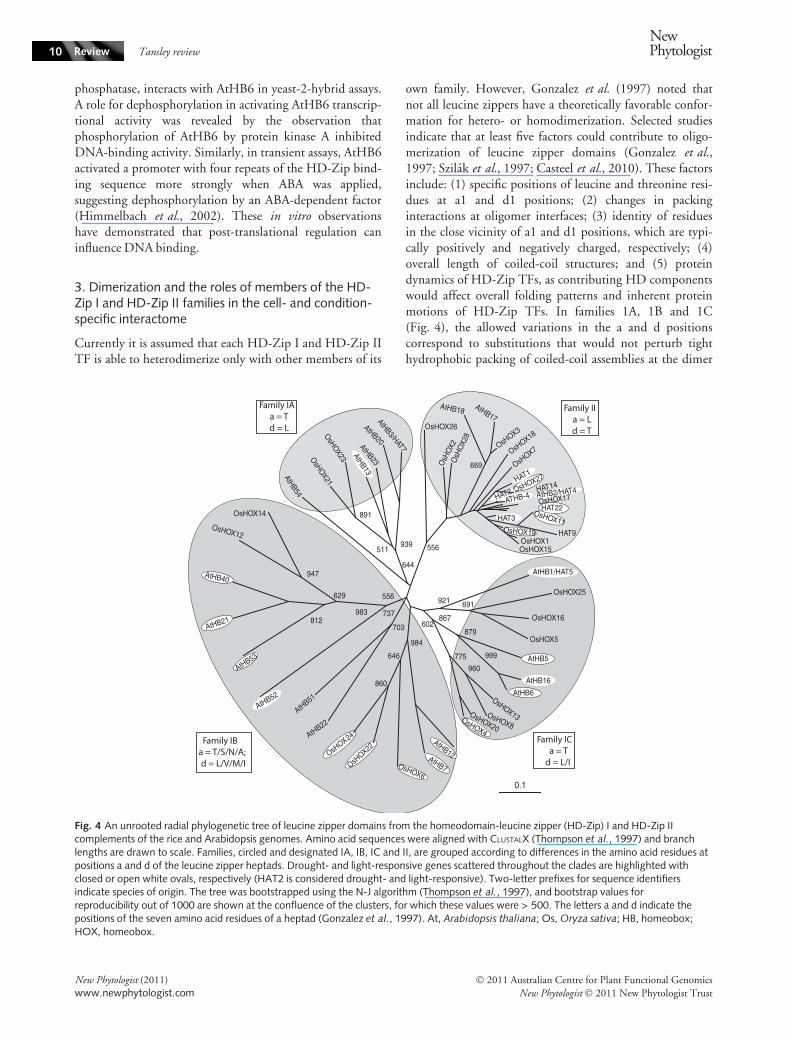
own family. However, Gonzalez et al. (1997) noted thatnot all leucine zippers have a theoretically favorable confor-mation for hetero- or homodimerization. Selected studiesindicate that at least five factors could contribute to oligo-merization of leucine zipper domains (Gonzalez et al.,1997; Szilak et al., 1997; Casteel et al., 2010). These factorsinclude: (1) specific positions of leucine and threonine resi-dues at a1 and d1 positions; (2) changes in packinginteractions at oligomer interfaces; (3) identity of residuesin the close vicinity of a1 and d1 positions, which are typi-cally positively and negatively charged, respectively; (4)overall length of coiled-coil structures; and (5) proteindynamics of HD-Zip TFs, as contributing HD componentswould affect overall folding patterns and inherent proteinmotions of HD-Zip TFs. In families 1A, 1B and 1C(Fig. 4), the allowed variations in the a and d positionscorrespond to substitutions that would not perturb tighthydrophobic packing of coiled-coil assemblies at the dimer
0.1
OsHOX25
OsHOX16
OsHOX5
691921
602
999
867
OsHOX13OsHOX8
775
OsHOX20
960
879
AtHB12AtHB7
984
OsHOX6OsH
OX22OsH
OX24
860
646
703
AtHB22
737812
629
OsHOX12
OsHOX14
947
983
556
AtHB54
OsHOX21
OsHOX23
891
AtHB23
511
AtHB3/HAT7
939
644
OsHOX26
OsH
OX2
OsH
OX2
8
AtHB18AtHB17
OsHOX3
669
OsHOX18
OsHOX7
HAT14
HAT9OsHOX1OsHOX15556
AtHB6
AtHB5
OsHOX4
OsHOX11
OsHOX27
OsHOX19
AtHB51
AtHB21
AtHB40
AtHB53
AtHB13AtHB20
AtHB1/HAT5
AtHB16
AtHB52
AtHB2/HAT4HAT1
ATHB-4
HAT22HAT3
HAT2OsHOX17
Fig. 4 An unrooted radial phylogenetic tree of leucine zipper domains from the homeodomain-leucine zipper (HD-Zip) I and HD-Zip IIcomplements of the rice and Arabidopsis genomes. Amino acid sequences were aligned with CLUSTALX (Thompson et al., 1997) and branchlengths are drawn to scale. Families, circled and designated IA, IB, IC and II, are grouped according to differences in the amino acid residues atpositions a and d of the leucine zipper heptads. Drought- and light-responsive genes scattered throughout the clades are highlighted withclosed or open white ovals, respectively (HAT2 is considered drought- and light-responsive). Two-letter prefixes for sequence identifiersindicate species of origin. The tree was bootstrapped using the N-J algorithm (Thompson et al., 1997), and bootstrap values forreproducibility out of 1000 are shown at the confluence of the clusters, for which these values were > 500. The letters a and d indicate thepositions of the seven amino acid residues of a heptad (Gonzalez et al., 1997). At, Arabidopsis thaliana; Os, Oryza sativa; HB, homeobox;HOX, homeobox.
10 Review Tansley reviewNewPhytologist
� 2011 Australian Centre for Plant Functional Genomics
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011)
www.newphytologist.com

interface. More information is needed on the interactionprofile of HD-Zip I and HD-Zip II TFs and the propertiesthat specific interaction partners contribute to cis elementbinding. Such studies have been successful in defining thecharacteristics of the MADS box TFs (de Folter et al.,2005).
The strength of the DNA–protein interaction is alsodependent on the dimers that make up the unit as demon-strated in vitro with OsHOX4 and OsHOX5, which bindDNA more strongly as heterodimers than as homodimers(Meijer et al., 2000). It was also shown that the presence ofstimulus-specific factors can modulate the DNA-bindingproperties of HD-Zip I and HD-Zip II TFs. A co-activatorof transcription, which was specifically induced in responseto a biotic stress, Solanum tuberosum multiprotein bridgingfactor 1, increased the DNA-binding affinity of two sun-flower HD-Zip TFs from different families under in vitroconditions (Zanetti et al., 2004).
Some members of the HD-Zip I and HD-Zip II familiesare widely expressed in plant tissues throughout plantdevelopment, while others have specific spatial, temporal,developmental and ⁄ or inducible expression characteristics.Over-expression of different HD-Zip genes may result inphenotypes that are not physiologically relevant when ana-lyzed (Supporting Information Table S1). The presence ofan HD-Zip I or HD-Zip II TF, when ectopically expressed,may interfere with the network that is operating within aspecific tissue or cell when dimerization occurs. The sucroseconcentration-dependent small cotyledon phenotypeobserved in Arabidopsis ectopically expressing AtHB13reveals this anomalous effect, as a similar response is notseen in wild-type plants (Hanson et al., 2001). In this case,AtHB13 is thought to interfere with the role of an HD-ZipTF that is involved in cotyledon expansion but is unrelatedto sucrose concentration. Contrary to this idea that an HD-Zip I or HD-Zip II TF can interfere with the interactomewhen ectopically expressed, over-expression of a single HD-Zip I or HD-Zip II TF can result in little overall change ofa plant phenotype. This suggests that other spatial, tempo-ral or conditional factors necessary for activity of the ectopicTF are not present. This has been observed with ectopicover-expression of OsHOX1 in Arabidopsis and rice, wherea distinct effect on leaf shape or no other discerniblechanges in phenotype were observed, respectively (Scarpellaet al., 2000). It was proposed that OsHOX1 is only tran-scriptionally active only in a transient cell type in zones ofvascular differentiation; these observations imply that, out-side of this environment, the TF has little or no discernibleeffect on rice phenotype. A further example demonstratesthat the contribution of other specific conditional or spatialfactors is required for HD-Zip I or HD-Zip II TF activity.Under control conditions, root growth of Arabidopsis over-expressing AtHB7 or AtHB12 remains similar to that ofwild-type plants, whereas leaves and stems are smaller.
However, treating the roots with ABA results in increasedinhibition of root growth compared with that seen in wild-type plants, suggesting that an ABA-inducible factor isnecessary for the response in the roots (Olsson et al., 2004).An example of a developmentally specific role comes fromArabidopsis plants over-expressing AtHB5. Arabidopsisplants over-expressing AtHB5 were found to have anincreased sensitivity to ABA, which was characterized byinhibited germination and seedling root growth with noobservable effect seen in adult plants (Johannesson et al.2003). Johannesson et al. (2003) argued that AtHB5 modu-lates growth in response to water deficit strictly at theseedling stage and that at other developmental stages addi-tional factors required for this response are not present.With different roles in two developmentally different tissuetypes, AtHB2 ⁄ HAT4 over-expression induces hypocotylelongation and represses cotyledon expansion in transgenicplants. This suggests that AtHB2 ⁄ HAT4 plays distinct rolesin specific tissues in response to changes in light quality.
Clearly, HD-Zip I and HD-Zip II TFs are able to dimer-ize within their respective families and to bind a commoncis element under in vitro conditions, which suggests a largenetwork. To investigate the influence that any given HD-Zip I or HD-Zip II TF may have on this proposed network,as a first step, the presence of viable dimerization partnersmust be confirmed. Information on cell- and condition-specific transcriptomes accumulating in public databaseswill aid these efforts. The ability to dimerize with the part-ners that are present would then indicate involvement withthe HD-Zip I and HD-Zip II network, which could betested by analysis of downstream target genes.
4. Redundancies in the roles of HD-Zip I and HD-Zip IItranscription factor paralogs
Arabidopsis and rice HD-Zip families I and II consist ofmany related genes that have presumably arisen throughgenome and gene duplications, although this phenomenonwill need to be assessed in other species as complete genomesequences arise. These duplications can lead to variations inexpression patterns and differences in the roles that each ofthe paralogous genes plays. As is evident in the HD-Zip Iand HD-Zip II families of rice and Arabidopsis, this has ledto functional redundancies, which make it difficult to deter-mine a role for a single member of the families throughdown-regulation (Table S1). The degree of stem retarda-tion seen in AtHB7 and AtHB12 over-expressing plants isadditive, when both are over-expressed in concert. Thissuggests that the effect on the genes targeted by AtHB7 andAtHB12 is additive or, more likely, that they act on thesame target genes (Olsson et al., 2004). Duplication of theHD-Zip TFs would have relieved pressure to adhere to astrict functional role and provided an opportunity for newfunctionality. Transcription of both AtHB7 and AtHB12 is
NewPhytologist Tansley review Review 11
� 2011 Australian Centre for Plant Functional Genomics
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011)
www.newphytologist.com

strongly induced by ABA in all tissues, but the kinetics ofinduction differs and these genes are also regulated differ-ently by ABI1 and ABI2 (Lee & Chun, 1998). It was alsosuggested that AtHB5 may not play an exclusive role, asArabidopsis lines homozygous for an AtHB5 allele with aT-DNA insertion show no differences in phenotype fromwild-type plants (Johannesson et al., 2003). In light of thephylogenetic grouping within the b2 clade, it was suggestedthat AtHB6 and AtHB16 may share redundancies withAtHB5 (Johannesson et al., 2003). This remains to be veri-fied, especially as these genes play distinct roles and possessunique induction characteristics, as discussed in section II.
As noted above, the closely related genes HAT1, HAT2,HAT3, AtHB2 and AtHB4 share many similar characteris-tics though they have different expression patterns(Ciarbelli et al., 2008). All are transcriptionally induced bychanges in light quality, although HAT2 by an auxin-mediated path, over-expression of each gene results in asimilar phenotype, and no phenotype is observed in knockout lines of individual genes (Sorin et al., 2009). However,double knock-out lines have revealed that hat1 ⁄ hat2 plantsshow reduced hypocotyl elongation in response to FR light,and in athb4 ⁄ hat3 plants hypocotyl elongation in responseto FR light is abolished (Sorin et al., 2009).
OsHOX4 of the rice f clade is thought to share redundan-cies in function with its paralog OsHOX20. These genes aresimilar in structure and, although they possess differenttissue-specific expression patterns, both are down-regulatedin leaf blades upon exposure to drought stress (Agalouet al., 2008). When OsHOX4 expression was reduced intransgenic rice, using RNA interference, no phenotypic dif-ferences were observed. It is evident through mRNA knock-down experiments that there are redundancies in the func-tions of HD-Zip I and HD-Zip II TFs that cannot alwaysbe extended to redundancies in expression. These examplesof the facets of redundancy found within the HD-Zip I andHD-Zip II families make it difficult to clarify the role ofgenes in isolation. Where redundancies exist betweenrelated members, paralogs would need to be analyzed usingsingle, double and even triple mutant knockout lines totease apart both redundant and specific roles.
IV. Conclusions
Plants cope with a variety of environmental stresses by mod-ifying their growth pattern to minimize the impacts of stressor to escape damage. The HD-Zip I and HD-Zip II TFsplay an integral role in the signaling network that is trig-gered by endogenous and external stimuli, which leads tothe modified growth characteristic of stressed plants(Fig. 3). This growth adaptation is achieved through theregulation of cell differentiation, division and expansion byHD-Zip I and HD-Zip II TFs. However, there are stillmechanistic and functional aspects of the HD-Zip I and
HD-Zip II network that we need to understand before weare able to characterize the roles of each family in plantgrowth adaptation to environmental stresses. We know verylittle of the downstream genes that are ultimately regulatedby the HD-Zip I and HD-Zip II TFs. Identification of theirtarget genes is needed to build a comprehensive view of theregulatory pathways and will enable validation of the suiteof promoters controlled by HD-Zip I and HD-Zip II pro-teins to define the cis-acting elements. More information isneeded on the interaction profiles of HD-Zip I and HD-Zip II TFs, and the properties that specify interaction part-ners which contribute to cis element binding. The increasingamount of cell-specific microarray data available in publicdatabases will also enable determination of potential interac-tion partners that are dependent upon expression in thesame cell. If transcriptional regulation by the HD-Zip I andHD-Zip II TFs is conferred through a common cis element,that implies that there are factors determining the specificityof transcriptional activity, as transgenic plants over- orunder-expressing HD-Zip I and HD-Zip II TFs have vari-ant phenotypes. Elements contributing to the specificity ofaction remain elusive. Obtaining data on the three-dimen-sional structures of the HD-Zip I and HD-Zip II TFs willalso decisively contribute to our understanding of the natureof the protein–protein and protein–DNA interactions.
Acknowledgements
J.C.H. is a recipient of the Australian Postgraduate Award.
References
Achard P, Cheng H, De Grauwe L, Decat J, Schoutteten H, Moritz T,
Van der Straeten D, Peng JR, Harberd NP. 2006. Integration of plant
responses to environmentally activated phytohormonal signals. Science311: 91–94.
Agalou A, Purwantomo S, Overnaes E, Johannesson H, Zhu X, Estiati A,
de Kam RJ, Engstrom P, Slamet-Loedin IH, Zhu Z et al. 2008. A
genome-wide survey of HD-Zip genes in rice and analysis of drought-
responsive family members. Plant Molecular Biology 66: 87–103.
Aoyama T, Dong CH, Wu Y, Carabelli M, Sessa G, Ruberti I, Morelli G,
Chua NH. 1995. Ectopic expression of the Arabidopsis transcriptional
activator ATHB-1 alters leaf cell fate in tobacco. The Plant Cell 7:
1773–1785.
Ariel F, Diet A, Verdenaud M, Gruber V, Frugier F, Chan R, Crespi M.
2010. Environmental regulation of lateral root emergence in Medicagotruncatula requires the HD-Zip I transcription factor HB1. The PlantCell 22: 2171–2183.
Ariel FD, Manavella PA, Dezar CA, Chan RL. 2007. The true story of the
HD-Zip family. Trends in Plant Science 12: 419–426.
Barrero JM, Millar AA, Griffiths J, Czechowski T, Scheible WR, Udvardi
M, Reid JB, Ross JJ, Jacobsen JV, Gubler F. 2010. Gene expression
profiling identifies two regulatory genes controlling dormancy and ABA
sensitivity in Arabidopsis seeds. Plant Journal 61: 611–622.
Burglin TR, Finney M, Coulson A, Ruvkun G. 1989. Caenorhabditiselegans has scores of homeobox-containing genes. Nature 341:
239–243.
12 Review Tansley reviewNewPhytologist
� 2011 Australian Centre for Plant Functional Genomics
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011)
www.newphytologist.com

Carabelli M, Morelli G, Whitelam G, Ruberti I. 1996. Twilight-zone
and canopy shade induction of the Athb-2 homeobox gene in green
plants. Proceedings of the National Academy of Sciences, USA 93:
3530–3535.
Casteel DE, Smith-Nguyen EV, Sankaran B, Roh SH, Pilz RB, Kim C.
2010. A crystal structure of the cyclic GMP-dependent Protein Kinase I
beta dimerization ⁄ docking domain reveals molecular details of
isoform-specific anchoring. Journal of Biological Chemistry 285:
32684–32688.
Ciarbelli AR, Ciolfi A, Salvucci S, Ruzza V, Possenti M, Carabelli M,
Fruscalzo A, Sessa G, Morelli G, Ruberti I. 2008. The Arabidopsis
Homeodomain-leucine Zipper II gene family: diversity and redundancy.
Plant Molecular Biology 68: 465–478.
Dai M, Hu Y, Ma Q, Zhao Y, Zhou DX. 2008. Functional analysis of rice
HOMEOBOX4 (Oshox4) gene reveals a negative function in gibberellin
responses. Plant Molecular Biology 66: 289–301.
Deng X, Phillips J, Brautigam A, Engstrom P, Johannesson H,
Ouwerkerk PB, Ruberti I, Salinas J, Vera P, Iannacone R et al. 2006.
A homeodomain leucine zipper gene from Craterostigma plantagineumregulates abscisic acid responsive gene expression and physiological
responses. Plant Molecular Biology 61: 469–489.
Deng X, Phillips J, Meijer AH, Salamini F, Bartels D. 2002.
Characterization of five novel dehydration-responsive homeodomain
leucine zipper genes from the resurrection plant Craterostigmaplantagineum. Plant Molecular Biology 49: 601–610.
Dezar CA, Gago GM, Gonzalez DH, Chan RL. 2005. Hahb-4, a
sunflower homeobox-leucine zipper gene, is a developmental regulator
and confers drought tolerance to Arabidopsis thaliana plants. TransgenicResearch 14: 429–440.
Dezar CA, Giacomelli JI, Manavella PA, Re DA, Alves-Ferreira M,
Baldwin IT, Bonaventure G, Chan RL. 2010. HAHB10, a sunflower
HD-Zip II transcription factor, participates in the induction of
flowering and in the control of phytohormone-mediated responses to
biotic stress. Journal of Experimental Botany 62: 1061–1076.
Eckardt NA. 2003. MADS monsters: controlling floral organ identity. ThePlant Cell 15: 803–805.
Ellenberger TE, Brandl CJ, Struhl K, Harrison SC. 1992. The GCN4
basic region leucine zipper binds DNA as a dimer of uninterrupted a-
helices: crystal structure of the protein-DNA complex. Cell 71:
1223–1237.
de Folter S, Immink RGH, Kieffer M, Parenicova L, Henz SR, Weigel D,
Busscher M, Kooiker M, Colombo L, Kater MM et al. 2005.
Comprehensive interaction map of the Arabidopsis MADS box
transcription factors. The Plant Cell 17: 1424–1433.
Frank W, Phillips J, Salamini F, Bartels D. 1998. Two dehydration-
inducible transcripts from the resurrection plant Craterostigmaplantagineum encode interacting homeodomain-leucine zipper proteins.
Plant Journal 15: 413–421.
Franklin KA. 2008. Shade avoidance. New Phytologist 179: 930–944.
Franklin KA, Quail PH. 2010. Phytochrome functions in Arabidopsisdevelopment. Journal of Experimental Botany 61: 11–24.
Gago GM, Almoguera C, Jordano J, Gonzalez DH, Chan RL. 2002.
Hahb-4, a homeobox-leucine zipper gene potentially involved in abscisic
acid-dependent responses to water stress in sunflower. Plant, Cell &Environment 25: 633–640.
Garber RL, Kuroiwa A, Gehring WJ. 1983. Genomic and cDNA clones
of the homeotic locus ANTENNAPEDIA in Drosophila. The EMBOJournal 2: 2027–2036.
Gehring WJ, Qian YQ, Billeter M, Furukubotokunaga K, Schier AF,
Resendez-Perez D, Affolter M, Otting G, Wuthrich K. 1994.
Homeodomain-dna recognition. Cell 78: 211–223.
Gonzalez DH, Valle EM, Gago G, Chan RL. 1997. Interaction between
proteins containing homeodomains associated to leucine zippers from
sunflower. Biochimica et Biophysica Acta, Gene Structure and Expression1351: 137–149.
Hake S, Vollbrecht E, Freeling M. 1989. Cloning knotted, the dominant
morphological mutant in maize using ds2 as a transposon tag. TheEMBO Journal 8: 15–22.
Hanson J, Johannesson H, Engstrom P. 2001. Sugar-dependent
alterations in cotyledon and leaf development in transgenic plants
expressing the HDZhdip gene ATHB13. Plant Molecular Biology 45:
247–262.
Henriksson E, Olsson ASB, Johannesson H, Johansson H, Hanson J,
Engstrom P, Soderman E. 2005. Homeodomain leucine zipper class I
genes in Arabidopsis. Expression patterns and phylogenetic
relationships. Plant Physiology 139: 509–518.
Himmelbach A, Hoffmann T, Leube M, Hohener B, Grill E. 2002.
Homeodomain protein ATHB6 is a target of the protein phosphatase
ABI1 and regulates hormone responses in Arabidopsis. The EMBOJournal 21: 3029–3038.
Hirsch JA, Aggarwal AK. 1995. Structure of even skipped homeodomain
complexed to AT rich DNA: New perspectives on homeodomain
specificity. The EMBO Journal 14: 6280–6291.
Hjellstrom M, Olsson ASB, Engstrom O, Soderman EM. 2003.
Constitutive expression of the water deficit-inducible homeobox gene
ATHB7 in transgenic Arabidopsis causes a suppression of stem
elongation growth. Plant, Cell & Environment 26: 1127–1136.
Hofer J, Turner L, Moreau C, Ambrose M, Isaac P, Butcher S, Weller J,
Dupin A, Dalmais M, Le Signor C et al. 2009. Tendril-less regulates
tendril formation in pea leaves. Plant Cell 21: 420–428.
Huang DQ, Wu WR, Abrams SR, Cutler AJ. 2008. The relationship of
drought-related gene expression in Arabidopsis thaliana to hormonal and
environmental factors. Journal of Experimental Botany 59: 2991–3007.
Inoue S, Takemiya A, Shimazaki K. 2010. Phototropin signaling and
stomatal opening as a model case. Current Opinion in Plant Biology 13:
587–593.
Johannesson H, Wang Y, Engstrom P. 2001. DNA-binding and
dimerization preferences of Arabidopsis homeodomain-leucine zipper
transcription factors in vitro. Plant Molecular Biology 45: 63–73.
Johannesson H, Wang Y, Hanson J, Engstrom P. 2003. The Arabidopsisthaliana homeobox gene ATHB5 is a potential regulator of abscisic acid
responsiveness in developing seedlings. Plant Molecular Biology 51:
719–729.
Kissinger CR, Liu BS, Martin-Blanco E, Kornberg TB, Pabo CO. 1990.
Crystal-structure of an engrailed homeodomain-dna complex at 2.8-a
resolution – a framework for understanding homeodomain-dna
interactions. Cell 63: 579–590.
Lee YH, Chun JY. 1998. A new homeodomain-leucine zipper gene from
Arabidopsis thaliana induced by water stress and abscisic acid treatment.
Plant Molecular Biology 37: 377–384.
Leung J, Merlot S, Giraudat J. 1997. The Arabidopsis ABSCISIC ACID-INSENSITIVE2 (ABI2) and ABI1 genes encode homologous protein
phosphatases 2C involved in abscisic acid signal transduction. The PlantCell 9: 759–771.
Lin CT, Shalitin D. 2003. Cryptochrome structure and signal
transduction. Annual Review of Plant Biology 54: 469–496.
Liu HJ, Wang SF, Yu XB, Yu J, He XW, Zhang SL, Shou HX, Wu P.
2005. ARL1, a LOB-domain protein required for adventitious root
formation in rice. Plant Journal 43: 47–56.
Lopato S, Bazanova N, Morran S, Milligan AS, Shirley N, Langridge P.
2006. Isolation of plant transcription factors using a modified yeast one-
hybrid system. Plant Methods 2: 3–17.
Manavella PA, Arce AL, Dezar CA, Bitton F, Renou JP, Crespi M, Chan
RL. 2006. Cross-talk between ethylene and drought signalling pathways
is mediated by the sunflower Hahb-4 transcription factor. Plant Journal48: 125–137.
NewPhytologist Tansley review Review 13
� 2011 Australian Centre for Plant Functional Genomics
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011)
www.newphytologist.com

Manavella PA, Dezar CA, Ariel FD, Chan RL. 2008a. Two ABREs, two
redundant root-specific and one W-box cis-acting elements are
functional in the sunflower HAHB4 promoter. Plant Physiology andBiochemistry 46: 860–867.
Manavella PA, Dezar CA, Bonaventure G, Baldwin IT, Chan RL. 2008b.
HAHB4, a sunflower HD-Zip protein, integrates signals from the
jasmonic acid and ethylene pathways during wounding and biotic stress
responses. Plant Journal 56: 376–388.
Mattsson J, Ckurshumova W, Berleth T. 2003. Auxin signaling in
Arabidopsis leaf vascular development. Plant Physiology 131:
1327–1339.
Meijer AH, de Kam RJ, d’Erfurth I, Shen W, Hoge JHC. 2000. HD-Zip
proteins of families I and II from rice: interactions and functional
properties. Molecular and General Genetics 263: 12–21.
Moglich A, Yang XJ, Ayers RA, Moffat K. 2010. Structure and function
of plant photoreceptors. Annual Review of Plant Biology 61: 21–47.
Mukherjee K, Brocchieri L, Burglin TR. 2009. A comprehensive
classification and evolutionary analysis of plant homeobox genes.
Molecular Biology and Evolution 26: 2775–2794.
Nakashima K, Ito Y, Yamaguchi-Shinozaki K. 2009. Transcriptional
regulatory networks in response to abiotic stresses in Arabidopsis and
grasses. Plant Physiology 149: 88–95.
Ohgishi M, Oka A, Morelli G, Ruberti I, Aoyama T. 2001. Negative
autoregulation of the Arabidopsis homeobox gene ATHB-2. PlantJournal 25: 389–398.
Okushima Y, Fukaki H, Onoda M, Theologis A, Tasaka M. 2007. ARF7
and ARF19 regulate lateral root formation via direct activation of
LBD ⁄ ASL genes in Arabidopsis. The Plant Cell 19: 118–130.
Olsson AS, Engstrom P, Soderman E. 2004. The homeobox genes
ATHB12 and ATHB7 encode potential regulators of growth in response
to water deficit in Arabidopsis. Plant Molecular Biology 55: 663–677.
Palena CM, Gonzalez DH, Chan RL. 1999. A monomer-dimer
equilibrium modulates the interaction of the sunflower homeodomain
leucine-zipper protein Hahb-4 with DNA. Biochemical Journal 341:
81–87.
Palena CM, Tron AE, Bertoncini CW, Gonzalez DH, Chan RL. 2001.
Positively charged residues at the N-terminal arm of the homeodomain
are required for efficient DNA binding by homeodomain-leucine zipper
proteins. Journal of Molecular Biology 308: 39–47.
Perrot-Rechenmann C. 2010. Cellular responses to auxin: division versus
expansion. Cold Spring Harbor Perspectives in Biology 2: a001446.
Qian YQ, Otting G, Billeter M, Muller M, Gehring W, Wuthrich K.
1993. Nuclear-magnetic-resonance spectroscopy of a DNA complex
with the uniformly c-13-labeled antennapedia homeodomain and
structure determination of the DNA-bound homeodomain. Journal ofMolecular Biology 234: 1070–1083.
Re DA, Dezar CA, Chan RL, Baldwin IT, Bonaventure G. 2011.
Nicotiana attenuata NaHD20 plays a role in leaf ABA accumulation
during water stress, benzylacetone emission from flowers, and the timing
of bolting and flower transitions. Journal of Experimental Botany 62:
155–166.
Ruberti I, Sessa G, Lucchetti S, Morelli G. 1991. A novel class of plant-
proteins containing a homeodomain with a closely linked leucine zipper
motif. The EMBO Journal 10: 1787–1791.
Rueda EC, Dezar CA, Gonzalez DH, Chan RL. 2005. Hahb-10, a
sunflower homeobox-leucine zipper gene, is regulated by light quality
and quantity, and promotes early flowering when expressed in
Arabidopsis. Plant and Cell Physiology 46: 1954–1963.
Santner A, Calderon-Villalobos LIA, Estelle M. 2009. Plant hormones are
versatile chemical regulators of plant growth. Nature Chemical Biology 5:
301–307.
Sawa S, Ohgishi M, Goda H, Higuchi K, Shimada Y, Yoshida S,
Koshiba T. 2002. The HAT2 gene, a member of the HD-Zip gene
family, isolated as an auxin inducible gene by DNA microarray
screening, affects auxin response in Arabidopsis. Plant Journal 32:
1011–1022.
Scarpella E, Rueb S, Boot KJM, Hoge JHC, Meijer AH. 2000. A role for
the rice homeobox gene Oshox1 in provascular cell fate commitment.
Development 127: 3655–3669.
Sessa G, Morelli G, Ruberti I. 1993. The ATHB-1 and -2 HD-Zip
domains homodimerize forming complexes of different DNA-binding
specificities. The EMBO Journal 12: 3507–3517.
Sessa G, Morelli G, Ruberti I. 1997. DNA-binding specificity of the
homeodomain leucine zipper domain. Journal of Molecular Biology 274:
303–309.
Skinner DJ, Gasser CS. 2009. Expression-based discovery of candidate
ovule development regulators through transcriptional profiling of ovule
mutants. BMC Plant Biology 9. doi:10.1186 ⁄ 1471-2229-9-29
Skirycz A, Inze D. 2010. More from less: plant growth under limited
water. Current Opinion in Biotechnology 21: 197–203.
Soderman E, Hjellstrom M, Fahleson J, Engstrom P. 1999. The HD-Zip
gene ATHB6 in Arabidopsis is expressed in developing leaves, roots and
carpels and up-regulated by water deficit conditions. Plant MolecularBiology 40: 1073–1083.
Soderman E, Mattsson J, Svenson M, Borkird C, Engstrom P. 1994.
Expression patterns of novel genes encoding homeodomain leucine-
zipper proteins in Arabidopsis thaliana. Plant Molecular Biology 26:
145–154.
Son O, Cho HY, Kim MR, Lee H, Lee MS, Song E, Park JH, Nam KH,
Chun JY, Kim HJ et al. 2005. Induction of a homeodomain-leucine
zipper gene by auxin is inhibited by cytokinin in Arabidopsis roots.
Biochemical and Biophysical Research Communications 326: 203–209.
Son O, Hur YS, Kim YK, Lee HJ, Kim S, Kim MR, Nam KH, Lee MS,
Kim BY, Park J et al. 2010. ATHB12, an ABA-inducible
homeodomain-leucine zipper (HD-Zip) protein of arabidopsis,
negatively regulates the growth of the inflorescence stem by decreasing
the expression of a gibberellin 20-oxidase gene. Plant and Cell Physiology51: 1537–1547.
Sorin C, Salla-Martret M, Bou-Torrent J, Roig-Villanova I, Martinez-
Garcia JF. 2009. ATHB4, a regulator of shade avoidance, modulates
hormone response in Arabidopsis seedlings. Plant Journal 59: 266–277.
Stamm P, Kumar PP. 2010. The phytohormone signal network regulating
elongation growth during shade avoidance. Journal of ExperimentalBotany 61: 2889–2903.
Steindler C, Carabelli M, Borello U, Morelli G, Ruberti I. 1997.
Phytochrome A, phytochrome B and other phytochrome(s) regulate
ATHB-2 gene expression in etiolated and green Arabidopsis plants.
Plant, Cell & Environment 20: 759–763.
Steindler C, Matteucci A, Sessa G, Weimar T, Ohgishi M, Aoyama T,
Morelli G, Ruberti I. 1999. Shade avoidance responses are mediated by
the ATHB-2 HD-Zip protein, a negative regulator of gene expression.
Development 126: 4235–4245.
Sultan SE. 2010. Plant developmental responses to the environment:
eco-devo insights. Current Opinion in Plant Biology 13: 96–101.
Szilak L, Moitra J, Vinson C. 1997. Design of a leucine zipper coiled coil
stabilized 1.4 kcal mol(-1) by phosphorylation of a serine in the e
position. Protein Science 6: 1273–1283.
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG.
1997. The CLUSTAL_X windows interface: flexible strategies for
multiple sequence alignment aided by quality analysis tools. NucleicAcids Research 25: 4876–4882.
Tron AE, Bertoncini CW, Chan RL, Gonzalez DH. 2002. Redox
regulation of plant homeodomain transcription factors. Journal ofBiological Chemistry 277: 34800–34807.
Tron AE, Comelli RN, Gonzalez DH. 2005. Structure of homeodomain-
leucine zipper ⁄ DNA complexes studied using hydroxyl radical
cleavage of DNA and methylation interference. Biochemistry 44:
16796–16803.
14 Review Tansley reviewNewPhytologist
� 2011 Australian Centre for Plant Functional Genomics
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011)
www.newphytologist.com

Tron AE, Welchen E, Gonzalez DH. 2004. Engineering the loop region
of a homeodomain-leucine zipper protein promotes efficient binding to
a monomeric DNA binding site. Biochemistry 43: 15845–15851.
Vollbrecht E, Veit B, Sinha N, Hake S. 1991. The developmental gene
Knotted-1 is a member of a maize homeobox gene family. Nature 350:
241–243.
Wang Y, Henriksson E, Soderman E, Henriksson KN, Sundberg E,
Engstrom P. 2003. The Arabidopsis homeobox gene, ATHB16,
regulates leaf development and the sensitivity to photoperiod in
Arabidopsis. Developmental Biology 264: 228–239.
Wolters H, Jurgens G. 2009. Survival of the flexible: hormonal growth
control and adaptation in plant development. Nature Reviews Genetics10: 305–317.
Zanetti ME, Chan RL, Godoy AV, Gonzalez DH, Casalongue CA. 2004.
Homeodomain-leucine zipper proteins interact with a plant homologue
of the transcriptional co-activator multiprotein bridging factor 1. Journalof Biochemistry and Molecular Biology 37: 320–324.
Supporting Information
Additional supporting information may be found in theonline version of this article.
Table S1 Phenotypes of transgenic or mutant plants withaltered transcript levels of homeodomain-leucine zipper(HD-Zip) I and HD-Zip II transcription factors as reportedin the literature
Please note: Wiley-Blackwell are not responsible for thecontent or functionality of any supporting informationsupplied by the authors. Any queries (other than missingmaterial) should be directed to the New Phytologist CentralOffice.
NewPhytologist Tansley review Review 15
� 2011 Australian Centre for Plant Functional Genomics
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011)
www.newphytologist.com





























