Embed Size (px)
Citation preview

The biogenic origin of needle fibre calcite
GUILLAUME CAILLEAU*, ERIC P. VERRECCHIA*, OLIVIER BRAISSANT�and LAURENT EMMANUEL�*Institut de Geologie et Paleontologie, Universite de Lausanne, Anthropole, CH-1015 Lausanne,Switzerland (E-mail: [email protected])�Universitat Basel, Biozentrum-Pharmazentrum, Klingelbergstrasse 50-70, CH-4012 Basel, Switzerland�Laboratoire de Biomineralisations et Paleoenvironnements, Universite Pierre & Marie Curie, CasePostale 116, 4 Place Jussieu, 75252 Paris Cedex 05, France
ABSTRACT
Needle fibre calcite is one of the most ubiquitous habits of calcite in vadose
environments (caves deposits, soil pores, etc.). Its origin, either through
inorganic, indirect or direct biological processes, has long been debated. In this
study, investigations at 11 sites in Europe, Africa and Central America support
arguments for its biogenic origin. The wide range of needle morphologies is the
result of a gradual evolution of the simplest type, a rod. This rod is the
elementary brick which, by aggregation and welding, builds more complex
needles. The absence of cross-welded needles implies that they are welded in a
mould, or under a longitudinal and unidirectional constraint, before being
released inside the soil pores. The difference between the lengthening of the
needles and the c axis can be explained by the existence of needles observed
under a scanning electron microscope in organic sleeves, which can act as a
mould during rod growth. Complex morphologies with epitaxial outgrowths
on straight rods cannot have grown entirely inside organic microtubes; they
must result from soil diagenesis after the release of straight rods in a soil-free
medium. Whisker crystals are interpreted as the result of growth and
coalescence of euhedral crystals on a rod. Rhomb chains are considered to
be the consequence of successive epitaxial growth steps on a needle during
variations in growth conditions. Isotopic signatures for needle fibre calcite
vary from )16Æ63& to +1Æ10& and from )8Æ63& to )2Æ25& for d13C and d18O,
respectively. The absence of high d18O values for needle fibre calcite precludes
a purely physicochemical origin (evaporative) for this particular habit of
calcite. As epitaxial growth cannot precipitate in the same conditions as initial
needles, needle fibre calcite stable isotopic signatures should be used with
caution as a proxy for palaeoenvironmental reconstructions. In addition, it is
suggested that the term needle fibre calcite should be kept for the original
biogenic form. The other habit should be referred to as epitaxial forms of
needle fibre calcite.
Keywords Biomineralization, calcite, crystal growth, stable isotopes, vadosezone.
INTRODUCTION
Observed at various latitudes, needle fibre calcite(NFC) is one of the most ubiquitous habits ofcalcite in vadose environments. Nevertheless, itsorigin remains controversial. The origin of NFC
has been attributed to purely physicochemical,indirect biological or direct biological processesaccording to different authors. The aim of thispaper is to demonstrate, through an extensivereview of NFC morphologies and isotopicsignatures, that NFC is biogenic in origin. This
Sedimentology (2009) 56, 1858–1875 doi: 10.1111/j.1365-3091.2009.01060.x
1858 � 2009 The Authors. Journal compilation � 2009 International Association of Sedimentologists

assertion is also based on the relationshipbetween NFC and the environments in which itforms, using examples from Africa, Europe andCentral America. In other words, because NFC iswidespread, and possibly related to multiplebiogenic vectors, it should not be consideredsignificant by itself. Nevertheless, its isotopicsignature can be interpreted in terms of pedogenicmicroenvironments, emphasizing its huge poten-tial for reconstruction of microscale palaeoenvi-ronmental conditions.
State of the art
Calcite in the shape of a needle was first observedassociated with ‘moonmilk’ (secondary cave car-bonate accumulations; Gessner, 1555). Describedfor the first time in the beginning of the 20thCentury (Lacroix, 1901) as ‘needle fibre calcite’, italso has been called ‘lublinite’ (Iwanoff, 1906)although this term is no longer used. Morerecently, other terms were attributed to needles ofcalcitic crystals such as ‘whisker crystals’ (Supko,1971), ‘calcite en echelon’ (Stoops, 1976), ‘needlefibres of calcite’ (James, 1972) and ‘rhomb chains’(Jones & Ng, 1988). Except for moonmilk, all theother terms are related strictly to needle morpho-logies with lengths from 4 to 104 times their width.According to Verrecchia & Verrecchia (1994), onlyNFC should be considered as monocrystalline toprevent confusion in terminology.
Needle fibre calcite was first thought to be ofinorganic origin (Jones & Ng, 1988; Jones & Kahle,1993; Borsato et al., 2000). The inorganic pro-cesses involve highly supersaturated solutionsobtained by rapid evaporation, producing crystalswith abnormal forms such as dendrites, needlesand ‘whisker crystals’ (James, 1972; Riche et al.,1982). This interpretation is justified by the factthat the needles in soil are always above thehighest level of the water table.
The argument for an indirect biological origin issupported by the fact that micro-organisms dis-solve the parent material and provide Ca2+andCO3
2) ions for subsequent precipitation (Borsatoet al., 2000). Roots allow the parent soil solutionto reach a high level of supersaturation, leading toprecipitation of calcite crystals with long habits(Harrison, 1977). The direct biological origin forNFC has been discussed by Callot et al. (1985a,b),Philips & Self (1987), Jones (1988), Verrecchia &Verrecchia (1994), Becze-Deak et al. (1997), Loisyet al. (1999) and Verrecchia (2000). Needles areassumed to precipitate through bacterial activityor inside fungal walls, from where they are
released during organic matter decay or soilmoisture deficiency (leading to the ‘bursting’ ofthe fungal hyphae).
SITE DESCRIPTION
In order to compare characteristics of NFC, 11sites were sampled in Europe, Africa and CentralAmerica. In Europe, NFC has been sampled nearReims (Northern France, data from Loisy, 1997),in Burgundy near Dijon (Eastern France), in theGorges de la Loue (French Jura Mountains), nearVilliers (Swiss Jura Mountains), in a cave at theCornettes de Bises (Swiss Alps), in the Perigord(South-western France), in Montpellier (SouthernFrance) and in Almerıa (South-eastern Spain). InAfrica, two areas were sampled: in Ivory Coast,near the village of Biga and in Cameroon, near thefishing village of Mankare. Finally, NFC has beensampled in Mexico Valley, Central America. Thelocations were chosen for the diversity of theirenvironmental conditions; they are also the loca-tions for various research projects including somerelated to carbon storage in carbonate-free envi-ronments. The sample sites have various soilparent rocks (Table 1). Samples from Reims weretaken in calci-magnesium soils on Cretaceouschalk. In the Swiss Jura, French Jura, Perigordand Burgundy, soils developed on Late Quater-nary scree slopes of Mesozoic marine limestones(Fig. 1A). Soil pore size varies from large at theSwiss Jura site (centimetres to millimetres) tosmall at all the other sites (millimetres). The NFCoccurs as millimetre to centimetre cotton ball-likefeatures in the soil pores (Fig. 1B) or as a thincoating around the gravels and sand grains.Samples from Montpellier were observed in aQuaternary travertine. In South-eastern Spain,NFC has been sampled in a soil developed on acalcareous Plio-Pleistocene conglomerate. TheNFC from Africa has been sampled in varioustypes of orthox soils developed on calc-alkalinegranite parent rocks (Ivory Coast) and in soilsdeveloped on quartziferous fluvial deposits of theMbam River (Cameroon). These different soilswere modified by the presence of iroko trees,which induce large secondary biogenic carbonateaccumulations (Carozzi, 1967; Braissant et al.,2004; Cailleau et al., 2004, 2005). Samples fromMexico were taken from indurated soils devel-oped on volcanic ash deposits (tepetates) inwhich various carbonate accumulations occur.The NFC texture appears as millimetre-sizedcotton ball-like clusters in soil pores as well as
Biogenic origin of needle fibre calcite 1859
� 2009 The Authors. Journal compilation � 2009 International Association of Sedimentologists, Sedimentology, 56, 1858–1875

thin layers coating pore walls. Moonmilk wassampled on the wall of a cave in the Swiss Alps. Itappears as a soft wall coating whose thicknesscan be up to 50 cm (Fig. 1C). This mammilatedand slimy coating is present in a cave cavity to adepth of 20 m.
MATERIALS AND METHODS
At each site, the calcite needles were sampledunder a binocular microscope with tweezers onoutcrops, as well as in the laboratory, and stored atroom temperature after drying at 80 �C for 24 h. Inthe laboratory, optical observations wereperformed using a Leitz Aristoplan microscope(Leica Microsystemes, Rueil-Malmaison, France)with a 100· magnifying oil-immersion objective in
order to characterize the potential relationshipbetween needles and organic matter. Observationsof needles were made using a Phillips XL30 FieldEmission Gun Scanning Electron Microscope(FEG-SEM; FEI Company, Eindhoven, The Nether-lands). The XL30 was also used in its LTSEMcryogenic mode (low temperature scanning elec-tron microscopy). The SEM sample preparationincludes storage in a cold room (4 �C) and drying inthe presence of silica gel. A mesh of needlesdeposited on a SEM stub (with a carbon glue film)was gold coated (10 to 15 nm thick). Some sampleswere embedded in epoxy resin in a vacuumchamber and ultra-thin sections (60 to 150 nm
thick) were made using a Reichert Ultracut S (LeicaMicrosystems (Schweiz) AG, Heerbrugg, Switzer-land) microtome with a diamond knife. Non-prepared samples and ultra-thin sections were
Table 1. Site description and NFC types observed in associated soils.
Sitelocation
Geographicalcoordinates NFC type(s) observed Site description Bundles
Samplingseason
Reims 49�14¢ N, 4�12¢ E Not observed Calcimagnesic soilson Cretaceous chalk
Notobserved
Unknown
Burgundy 47�20¢ N, 4�59¢ E 4; 5; 6; 17 Soils developed on LateQuaternary scree slopesconstituted by Mesozoicmarine limestone cryoclasts
Yes Wet season
French JuraMountains
47�04¢ N, 6�11¢ E 4; 6 Soils developed on LateQuaternary scree slopesconstituted by Mesozoicmarine limestone cryoclasts
No Wet season
Swiss JuraMountains
47�04¢ N, 6�59¢ E 3; 4; 6; 13; 21; 22 Soils developed on LateQuaternary scree slopesconstituted by Mesozoicmarine limestone cryoclasts
No Wet season
Swiss Alps 46�19¢ N, 16�48¢ E 6; 7; 17 Wall of a cave No Dry season
Perigord 44�29¢ N, 0�58¢ E 4; 6; 7; 10; 13; 14; 15;16; 22; 24; 29; 30
Soils developed on LateQuaternary scree slopesconstituted by Mesozoicmarine limestone cryoclasts
Yes Wet season
Montpellier 43�39¢ N, 3�52¢ E 1; 3; 4; 6; 8; 11; 12; 13;20; 21; 23; 24; 25; 26;27; 29; 30; 31; 32; 33
Quaternary travertine Yes Dry season
Almerıa 36�51¢ N, 2�22¢ W 4; 5; 6; 18; 19;28; 29; 30
Calcareous Plio-Pleistoceneconglomerate
No Dry season
Ivory Coast 6�39¢ N, 6�23¢ W 2; 6; 7; 13 Orthox soils developed oncalco-alkaline granite
Yes Dry season
Cameroon 5�27¢ N, 11�03¢ E 2; 6; 13 Quartziferous fluvial deposits No Dry season
Mexico 20�05¢ N, 102�25¢ W 3; 4; 9; 14; 15; 16; 17 Indurated soils developed onvolcanic ash deposits
No Unknown
1860 G. Cailleau et al.
� 2009 The Authors. Journal compilation � 2009 International Association of Sedimentologists, Sedimentology, 56, 1858–1875

observed using a Phillips CM-200 TransmissionElectron Microscope (TEM; FEI Company) with anacceleration voltage of 200 kV.
A systematic procedure has been applied inorder to decipher the isotopic signature of NFC.Firstly, the carbonate parent rocks were sampled.As stipulated above, at African sites, NFC hasbeen sampled in pseudo-orthox (oxic-ferralitic)soils developed on carbonate-free parent rocks.Nevertheless, in some of these soils, bio-inducedcarbonates are present (Carozzi, 1967; Cailleauet al., 2005) and have to be considered as theparent rock. In Mexico, a powder of the bulk soilhas been used as tepetate parent rock. In thesesoils, NFC is not the dominant form of calciumcarbonate. Secondly, for each site, cotton ball-likestructures of NFC were sampled because they arethe most original macromorphological features ofNFC that can be easily analysed. All the mesheswere sampled with tweezers to avoid any con-tamination with other materials. Samples weredried and crushed in an agate mortar after which5 mg of prepared sample was stored under vac-uum overnight. The following day, in the pres-ence of orthophosphoric acid, CO2 was producedand analysed. The isotopic compositions of allsamples (except for needles from Reims) weredetermined with a VG Micromass 602 massspectrometer (Micromass UK Ltd, Altrincham,UK). The stable isotopic composition of NFC fromReims was measured with a VG Prism2 spectro-meter (Micromass UK Ltd, Manchester, UK; data
from Loisy, 1997). All isotopic signatures ofcalcium carbonate are presented using notationaccording to the Vienna Pee Dee Belemnite stan-dard. For most of the samples, isotopic analyseswere performed in triplicate.
RESULTS
Relationships between organic fabricsand calcite rods
An exhaustive analysis has been conducted onfilamentous organisms present in the mesh of NFCin samples from two sites (Burgundy and Mont-pellier) in which samples are fresh and abundant.In Burgundy, the comparison between plane andcross-polarized light observations of a filamentusing a 100· magnification oil-immersion objec-tive show small polarized fibres inside livingorganic matter (Fig. 2A and B). A hydrochloricacid test confirmed the presence of calcium car-bonate while X-ray diffraction analysis determinedthis to be calcite. This result excludes a possiblemisinterpretation of polarizing fibres such as cel-lulose, for example. The SEM observations of thissample show a needle of calcite (MB type ofVerrecchia & Verrecchia, 1994) inside a microtubeof organic matter with a diameter around 1Æ5 lm(Fig. 2C). These microtubes also exist withoutneedles (Fig. 2D). The NFC can be observed pop-ping out of the outer membrane of the same
A C
BFig. 1. (A) Calci-magnesium soildeveloped on periglacial scree inthe quarry at Villier (Swiss JuraMountains). The presence of NFC isshown by white arrows. (B) Cottonball aggregate of NFC found in theVillier quarry (white arrows). (C)Moonmilk coating a wall near thecave entrance at the Swiss Alps site(Valais). Note the tweezers plantedin the soft and slimy structure.
Biogenic origin of needle fibre calcite 1861
� 2009 The Authors. Journal compilation � 2009 International Association of Sedimentologists, Sedimentology, 56, 1858–1875

A CB
D E
F G
Fig. 2. (A) and (B) Optical photographs of a living fungal filament. Cross-polarized light observations (B) show rodsof calcite inside the organic matter and fungal wall microtubes. (C) SEM photograph showing a serrated-edged needleinside a fungal microtube. (D) SEM photograph of an empty fungal organic microtube. The non-calcitic nature ofthese microtubes has been checked with back-scattered analysis and microdiffraction. (E) SEM photograph of anorganic filament similar to those shown in (A) and (B). This wide view shows needles popping out of the organicmatter (white arrows). (F) SEM photograph of a deteriorated organic tube covering needles. Serrated-edged outlinesof the rod are visible (white arrow) even covered by the organic layer due to the lack of sample preparation (i.e.fixation and hardening of organic matter). It can be estimated that there are two or three rods in a sleeve. (G) SEMphotograph of a dichotomous structure composed of an organic sleeve with rods inside. Note that the serrated-edgedrod outline is still visible despite the tube thickness (white arrows). Cracks inside the organic sleeve are due to theabsence of sample fixation during SEM vacuum (white arrow with black outline).
1862 G. Cailleau et al.
� 2009 The Authors. Journal compilation � 2009 International Association of Sedimentologists, Sedimentology, 56, 1858–1875

structures present in Fig. 2A and B (Fig. 2E, whitearrows), i.e. possibly fungal organic matter.
The same observations were made on samplesfrom Montpellier, confirming the relationshipbetween NFC and organic structures: for instance,needles (sometimes only one) were observedcovered by, or inside, an organic sleeve (Fig. 2F,white arrow). In addition, branched structureshave also been detected in an NFC mesh. In eachsegment of this particular structure, the serratededge of a rod is still visible through the organicmatter (Fig. 2G, white arrow).
Needle morphologies
The SEM observations of NFC mesh were con-ducted in order to describe the variety of theirmorphologies. There are a multitude of shapes dueto small variations between needles which super-ficially seem similar. This similarity precludes anyexhaustive description of all morphological typesof NFC. This statement implies that a nomencla-ture should present not an exhaustive catalogue,but a representative picture of the most frequentshapes found in all the samples analysed (Fig. 3).
The most complex morphologies were found atthe Perigord and Mexico sites (Fig. 4); theyexhibit two different directions of excrescenceon a straight principal rod. Sometimes a biggercrystal forms at the end of these outgrowths(Fig. 4B). Some of these excrescences can bebranched. Another type of rod shows outgrowthsdeveloped along a plane in continuity with smallirregularities, crenellating specific planes on thestraight rod (Fig. 4C and D, white arrows). Otheratypical shapes seem to be made of two compo-nents, a single rod or a complex rod bearing bigcrystals which can be scalenohedral, rhombohe-dral or a chain of coalescing subhedral crystals ofcalcite (Fig. 3, needle types 16 and 17, Fig. 4Eand F). The TEM microdiffraction performed onthe subhedral crystals and the rod bearing themindicates a common crystallographic orientation.In addition, rhomb chains (Jones & Ng, 1988)were observed in a thin crust around soil grains inBurgundy and the Swiss Jura Mountains(Fig. 5A). These features do not occur in cot-ton ball-like structures and they are present whenserrated edged needles are the principal rod type.Serrated planes are often extremely well-devel-oped in these crusts on soil grains (Fig. 5B and C),even to the point of the disappearance of theserrated-edge teeth and, in the extreme case,display crystal faces similar to ‘en echelon’ calcite(Fig. 5D, white arrows).
Mesostructures
Although a random mesh is the main structureobserved, some needle bundles have also beenfound (Fig. 6A) with an average apparent dia-meter of 8 lm. Sometimes the morphologies oftwo juxtaposed rods seem to fit together (Fig. 6B).Observations of the moonmilk in SEM low tem-perature mode reveal a structure composed offour major components: (i) fungi; (ii) a compactmesh of very flexible nanofibres; (iii) principallysmooth and sometimes serrated-edged rods; and(iv) several clusters of micritic crystals supportedby the mesh of nanofibres. Roughly, the NFCcontent represents less than one-third of thecarbonate content of the moonmilk. Therefore,sampling specifically for NFC inside the moon-milk is unfeasible.
TEM dark field observations
The homogeneity of light in the ‘planes’ (the term‘plane’ is used here to describe morphology)suggests that they have the same crystallographicorientation throughout and on each side of thecentral darker area (Fig. 7A). Moreover, TEMmicrodiffraction analysis of a single rod revealsonly a single crystal. The TEM observationsprovide a close-up view of serrated-edged planes,single or coupled, growing on rods. The observa-tion of only one side of a plane shows thesuperposition of two sub-planes forming anirregular edge (Fig. 7B, black arrows). Theseedges correspond to the overlapping sub-planesdrawn in Figure 3 (needle no. 13) that create theserrated edge (Fig. 7C to F). These sub-planes arevisible in cross-section on some needles (Fig. 7G)but are missing on others (Fig. 7H and I).
Carbon and oxygen isotopic data
Due to field sampling constraints, or the qualityand nature of samples, only nine of the elevensites were analysed for a comparison betweenNFC and parent rock. Almerıa samples were onlyused for electron microscopy and no isotopicanalysis was performed on them. Moreover, forthe Swiss Alps samples (i.e. the moonmilk), NFCwas impossible to separate from the other calciticphases. Consequently, these samples have beenanalysed only to add supplementary data. Inaddition, moonmilk parent rock has not beenanalysed for the samples from the Swiss Alps.
Isotopic signatures for NFC (Fig. 8) vary from)16Æ63& to +1Æ10& and from )8Æ63& to )2Æ25&
Biogenic origin of needle fibre calcite 1863
� 2009 The Authors. Journal compilation � 2009 International Association of Sedimentologists, Sedimentology, 56, 1858–1875

for d13C and d18O, respectively. At three sites(Cameroon, Reims and Swiss Jura), at least twoanalyses were performed because of the abun-dance of material. The standard deviations inSwiss Jura, Cameroon and Reims NFC are 0Æ42&,0Æ87& and 2Æ66& for d13C, and 0Æ61&, 0Æ23& and0Æ61& for d18O, respectively (Table 2).
A general trend is detected between 13Csignatures of NFC crystals and their associatedparent rock. Following this trend, NFC generally
displays lower values than the parent rock,except for the Montpellier, Cameroon and Mex-ico sites, in which d13C values are fairly similar.The shifts in d13C values between NFC andparent rocks vary from 3Æ63& (Burgundy) to12Æ58& (French Jura). At the Cameroon site,values for NFC are fairly similar to the bio-induced carbonates (Braissant et al., 2004) con-sidered to be the parent rock (Fig. 8). Similarly,samples from Mexico and Montpellier show the
Fig. 3. A non-exhaustive synthesis of needle morphologies. Numbers correspond to different shapes, as representedby cross-section or three-dimensional morphology. Three groups are distinguished. (i) Serrated-edged types: Types 1,11, (12), 13, 19, (20), 21 to 27, (28), 29 to 33. (ii) Smooth types: Types (2), 3 to 8, 10, 18. (iii) Complex types: Types 9,14 to 17. The detail for Type 31 shows a relationship between planes and serrated-edge morphology.
1864 G. Cailleau et al.
� 2009 The Authors. Journal compilation � 2009 International Association of Sedimentologists, Sedimentology, 56, 1858–1875

A B
C D
E F
Fig. 4. (A) to (F) SEM photographs of some complex needle shapes. (A) Typical needle shapes from Africa, com-posed of more than four juxtaposed needles (without serrated-edges). (B) Three-dimensional rake needle from thetepetate site in Mexico. Note the constant angle between the principal direction and the short and long outgrowths.(C) and (D) Details of the small irregularities present along this type of needle. There are one or two lines ofirregularities on each side plane of these needles. Thin long outgrowths and bigger crystals show growth directionsparallel to these irregularities. These photographs correspond to types 15 and 14, respectively. (E) SEM photographshowing a needle bearing scalenohedra. (F) SEM photograph of big rhombohedra growing on a complex needle(white arrow).
Biogenic origin of needle fibre calcite 1865
� 2009 The Authors. Journal compilation � 2009 International Association of Sedimentologists, Sedimentology, 56, 1858–1875

A B
C D
Fig. 5. (A) to (D) SEM photographs of thin crust developed on soil grains. (A) Rhomb chain (white arrow) observed inSwiss Jura Mountains samples. (B) Other examples of needles found in these crusts showing well-developed sep-arated outgrowths (white arrow) and coalescent excrescences (black arrow with white outline) on rods. (C) Anotherexample of coalescent excrescences (black arrow with white outline) on rod. Coalescence of excrescences can lead toa particular type of rod (star) which is shown in detail in (D). (D) Close-up of a needle whose primary shape is stillvisible (black arrow with white outline). Note the shape of the edge that seems to develop a rhomb chain morphology(white arrows).
A B
Fig. 6. (A) SEM photograph showing in detail a bundle of needles from Montpellier (France). Note that most of theneedles have serrated edges. (B) SEM photograph of a bundle in which rods seem to fit together, like pieces of a puzzle.
1866 G. Cailleau et al.
� 2009 The Authors. Journal compilation � 2009 International Association of Sedimentologists, Sedimentology, 56, 1858–1875

A
G H I
B C
D E F
Fig. 7. (A) TEM dark field photograph of a calcite needle without pronounced serrated edge planes. Each plane has ahomogenous white colour (note that this section is of the type 33 or 21 in Fig. 3). This homogeneity implies that thesetwo planes have the same crystallographic orientation. The calcitic material at the junction between the two planesinduces a different transmission pattern, revealed by a dark area (black arrows). This effect should not be interpretedas a different crystallographic orientation of these areas, but rather as the loss of diffracted electrons in a thick area.(B), (C), (D) and (E) TEM bright field observations of a serrated edge needle (the star and the plus sign identify thesame plane). (B) Close-up view of serrated edge planes showing the overlapping of sub-planes (black arrows). Notethe apparent divergence of overlapping directions between opposite sub-planes. (C), (D) and (E) Relationships be-tween cross-section and longitudinal views given by a succession of single tilt observations. The apparent divergenceof overlapping directions between the opposite sub-planes is due to the observation of external (star) and internal(plus) sides of planes as explained in (F). (G), (H) and (I) TEM bright field observations on an ultra-thin section (120 to150 nm) of resin-embedded needles in transverse section. The rough appearance of all material present is due to thecarbon coating. (G) Detail of a rod close to type 13 (or 23). Note the superposition of growth planes (arrow). (H) Cross-section of a type 25 (26 or 27) needle. (I) Malta cross-like needle section, similar to a rod of type 15 (or 9).
Biogenic origin of needle fibre calcite 1867
� 2009 The Authors. Journal compilation � 2009 International Association of Sedimentologists, Sedimentology, 56, 1858–1875

same pattern, with parent rock values close toNFC values.
Comparison between 18O signatures of NFCand parent rocks also shows a trend to negativevalues, except for the Cameroon, Perigord andBurgundy sites. At the first two sites, d18O
values of NFC and parent rocks are similarbut, in Burgundy, a small 18O-enrichment hasbeen measured in the needles. The 18O ofrainwater is also given in order to measure itspossible contribution to the isotopic signature ofNFC.
A
B
Fig. 8. (A) Synthesis of isotopic data. When more than two values are available for parent rocks, a mean point islabelled on the sketch. Arrows represent the shift between parent rock and needle values. (B) Range of the mean andmean d18O values of the rain water compared to the d18O values of NFC from various environments. Shifts areobserved in both directions.
1868 G. Cailleau et al.
� 2009 The Authors. Journal compilation � 2009 International Association of Sedimentologists, Sedimentology, 56, 1858–1875

Table 2. C and O isotopic signatures of NFC sampled at each site, as well as their carbonate parent rock. Some of theparent rocks are bio-induced calcium carbonate which possibly acts as a depleted carbon 13 source. At each site, NFCand parent rock samples were collected at the same depth ±30 cm.
Samplingarea d13C & d18O & d13C parent rock & d18O parent rock &
Cameroon )12Æ08 )2Æ62 )12Æ08 )2Æ28*)11Æ99 )2Æ85 )11Æ55* )1Æ74*)11Æ19 )2Æ67 )11Æ79* )2Æ58*)9Æ86 )2Æ25 )11Æ34* )1Æ91*)10Æ82 )2Æ90 )12Æ38* )2Æ73*)10Æ63 )2Æ90 )11Æ77* )3Æ21*)10Æ03 )2Æ63 )12Æ00* )2Æ90*)10Æ94 (0Æ87)§ )2Æ69 (0Æ23)§ )13Æ55* )1Æ23*
)12Æ40* )3Æ27*)11Æ34* )3Æ12*)10Æ61* )3Æ35*)10Æ88* )2Æ95*)9Æ78* )3Æ77*)9Æ33* )3Æ75*)10Æ54* )3Æ67*)7Æ99* )3Æ29*)6Æ86* )2Æ77*)10Æ64* )2Æ96*)10Æ07* )2Æ81*)11Æ46* )3Æ12*)10Æ14* )2Æ74*)8Æ20* )2Æ58*)9Æ10* )1Æ88*)10Æ69 (1Æ60)§ )2Æ81 (0Æ65)§
Swiss Alps )5Æ70 )5Æ32
Swiss Jura )7Æ12 )8Æ63 1Æ02 )5Æ96)6Æ31 )7Æ85)6Æ91 )7Æ80)6Æ29 )7Æ13)6Æ66 (0Æ42)§ )7Æ85 (0Æ61)§
Ivory Coast )16Æ63 )8Æ02 )11Æ32* )2Æ53*)8Æ93* )1Æ95*)6Æ46* )2Æ17*)6Æ96* )1Æ87*)6Æ53* )2Æ60*)8Æ74* )2Æ19*)8Æ16 (1Æ89)§ )2Æ22 (0Æ30)§
French Jura )11Æ45 )7Æ17 1Æ13 )4Æ391 to 2Æ5� )2 to )4�
Mexico )1Æ53 )8Æ60 )1Æ51 )7Æ71
Montpellier )10Æ14 )4Æ99 )9Æ72 )4Æ52)9Æ91 )4Æ89 )9Æ70 )4Æ85
Burgundy )10Æ79 )5Æ88 )7Æ16 )6Æ36
Perigord )8Æ54 )4Æ27 1Æ09 )4Æ07
Reims )7Æ57� )5Æ17� 2Æ43 )2Æ89)8Æ51� )4Æ81�1Æ10� )2Æ82�)1Æ80� )4Æ20�)4Æ10� )4Æ10�)5Æ00� )4Æ70�)1Æ90� )4Æ20�)3Æ00� )4Æ00�
Biogenic origin of needle fibre calcite 1869
� 2009 The Authors. Journal compilation � 2009 International Association of Sedimentologists, Sedimentology, 56, 1858–1875
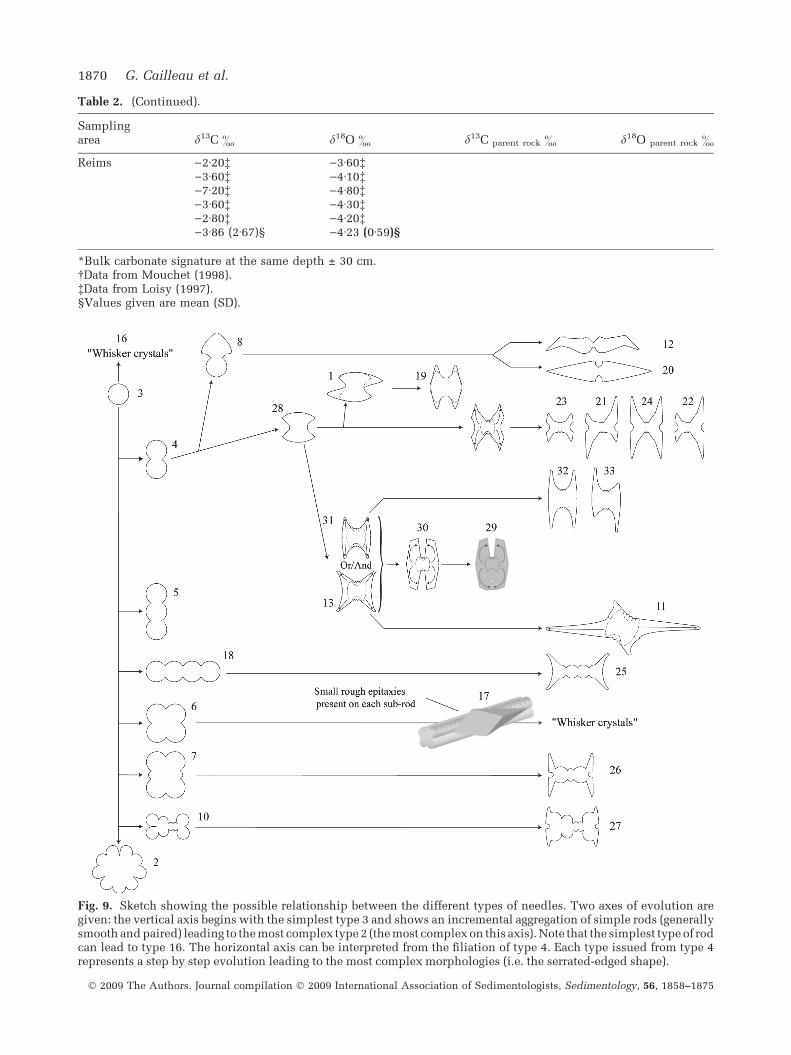
Table 2. (Continued).
Samplingarea d13C & d18O & d13C parent rock & d18O parent rock &
Reims )2Æ20� )3Æ60�)3Æ60� )4Æ10�)7Æ20� )4Æ80�)3Æ60� )4Æ30�)2Æ80� )4Æ20�)3Æ86 (2Æ67)§ )4Æ23 (0Æ59)§
*Bulk carbonate signature at the same depth ± 30 cm.�Data from Mouchet (1998).�Data from Loisy (1997).§Values given are mean (SD).
Fig. 9. Sketch showing the possible relationship between the different types of needles. Two axes of evolution aregiven: the vertical axis begins with the simplest type 3 and shows an incremental aggregation of simple rods (generallysmooth and paired) leading to the most complex type 2 (the most complex on this axis). Note that the simplest type of rodcan lead to type 16. The horizontal axis can be interpreted from the filiation of type 4. Each type issued from type 4represents a step by step evolution leading to the most complex morphologies (i.e. the serrated-edged shape).
1870 G. Cailleau et al.
� 2009 The Authors. Journal compilation � 2009 International Association of Sedimentologists, Sedimentology, 56, 1858–1875

DISCUSSION
Crystal morphology and genesis
Analysis of the various rod types suggests thatthe wide range of needle morphologies could bethe result of a gradual evolution, starting from thesimplest type 3 (see Fig. 3), as suggested by Jones& Kahle (1993) and Verrecchia & Verrecchia(1994). The evolution of needle morphology fromthe initial rod (shape 3, Fig. 9) can be summa-rized using two axes as shown in Fig. 9. Thisinterpretation is based initially on the presence ofshapes, which can be built by simply putting twosingle rods side by side, as shown along thevertical axis in Fig. 9. Small longitudinal mor-phological variations are used to describe rela-tionships between shapes, and this is translatedalong the horizontal axis (Fig. 9). A step by stepmorphological evolution along the horizontal axisis proposed for a simple needle such as type 4(Fig. 9). All the simple types are related toserrated-edged shapes, given as the end memberalong the horizontal axis. The aggregation ofsimple rods along the vertical axis produces thejuxtaposition of sub-rods, side by side, but nevercross-welded needles. The formation of theseneedles implies that they must have been weldedin a mould, or under a longitudinal and uni-directional constraint. This process is a strongargument for aggregation inside an organic mould(as shown in Fig. 2D) before needles are releasedinside the free soil pore medium.
One typical morphological crystallographicproperty of NFC is the presence of serrated-edgedplanes built by superimposed sub-planes (Fig. 3,needle type 13, and Fig. 7B to G). The mono-crystalline nature of the whole needle impliesthat serrated-edged planes must be considered assyntaxial crystal growths. Serrated edges can alsobe interpreted as the result of syntaxial growth on asimpler rod shape inside an organic sleeve. Thisinterpretation is supported by the observation oftype 11 needles that are composed of two differentplanes. One plane is less developed, with aserrated-edge, and the other is more developed,with a straight and smooth edge. The presence ofstraight and smooth edges must be interpreted asthe result of a constraint during the growth in alimited space, such as an organic sleeve. The sameinterpretation can be proposed for needle types 12and 20 (Fig. 3). Moreover, it is not possible toconclude that all the complex needles, for instancethose shown in Fig. 4B to F, have grown inside anorganic sleeve. The epitaxial outgrowths on
straight rods, such as thin and elongated or stubbymorphologies, or large euhedral shapes, cannotform inside organic microtubes because of theirsize and complex arrangement.
The SEM observations made on rhomb chainsand whisker crystals inspire new hypotheses fortheir origin and growth conditions. As shown inFig. 4E and F, whisker crystals can easily beinterpreted as the result of growth and coales-cence of euhedral crystals (scalenohedra and/orrhombohedra) on a rod (smooth or complex).Rhomb chains can be the consequence of succes-sive epitaxial growth steps on a needle asobserved in Fig. 5A to D. Rhombohedra fromrhomb chains, which are subhedral, cannot resultfrom the same growth conditions as the crystalsobserved on whisker crystals (compare Fig. 4Eand F and Fig. 5A and D). Nevertheless, these twotypes of needles begin as rods. The big euhedralcrystals are interpreted commonly as a product ofslow growth from a slightly supersaturated parentsolution. The rhombs forming rhomb chains seemto have been precipitated following successivesyntaxial growth along a single plane (forming athick blade; Fig. 5D), which displays a rhombo-hedral habit with time. Initially, this growth mustbe the result of precipitation from a supersatu-rated solution. Later, the growth slows down dueto a less supersaturated solution, leading to asubhedral crystalline habit. In addition, smallsyntaxial crystals, extremely abundant at theMexico site (Fig. 4B), are interpreted commonlyas the result of fast growth under highly super-saturated conditions.
Conventionally, needle growth is explained bya high growth rate obtained during high evapora-tion (Jones & Ng, 1988; Jones & Kahle, 1993;Borsato et al., 2000). In this case, it seems possi-ble to precipitate needles in evaporative environ-ments where fast growth rates can be reached.Consequently, oxygen isotopic signatures shouldbe enriched in 18O, i.e. d18O must be fairly high, atleast over 0, but it also depends on the initialrainwater signature. The measurements shown inTable 2 do not support this hypothesis. Data inStrong et al. (1992) and Bajnoczi & Kovacs-Kis(2006) also question a potential origin by evapo-ration. Moreover, the hypothesis of a purelyphysicochemical origin for needle growth doesnot explain the difference between the lengthen-ing of the needles and the c axis which is 40� to50� (Iwanoff, 1906), 30� to 40� (Mugge, 1914), 32�to 39� (Stoops, 1976) or 45� (Verges et al., 1982).
In order to understand the preferential andunusual direction of the c axis compared to that
Biogenic origin of needle fibre calcite 1871
� 2009 The Authors. Journal compilation � 2009 International Association of Sedimentologists, Sedimentology, 56, 1858–1875

of crystal lengthening, the role of organic mattermust be considered seriously. The organic mem-brane is composed of carboxyl groups that con-stitute preferential sites for calcite nucleation(Mitterer & Cunningham, 1985). During growth,the crystal propagates in a direction potentiallydifferent from that of the needle lengthening. Thishypothesis can explain the constant angle (from30� to 50�) between the c axis and the needledirection.
Isotopic signature of needle fibre calcite
In order to understand carbon isotopic fraction-ation in NFC, it is necessary to define the variouspotential sources of this element. Two mainsources can be identified: the carbonate parentrock and the soil CO2. Regarding the parent rocksignature, it depends on the nature of thecarbonate origin and its diagenetic evolution(marine, freshwater, etc.). Regarding the soilCO2, it originates from soil respiration (soilorganic matter decay and root respiration). This
source of CO2 induces a specific signature forpedogenic carbonates as demonstrated by Cerling(1984, 1999). In addition, the nature of the plantcover influences the signature of the organicmatter as well as those of the pedogenic carbon-ates. The organic matter signature is about )12&
for d13C under a solely C4 vegetation cover. Thiscarbon source induces a d13C value of ca +2& forpedogenic carbonates. Regarding the solely C3vegetation cover, the mean value for organic d13Cis about )26&, resulting in a signature of ca)12& for pedogenic carbonates. The potentialsource of C for NFC is a combination of twosources, the parent rock and the soil CO2. As nosoil CO2 has been sampled at the studied sites,the discussion related to the isotopic signature ofNFC is only based on the plants observed on thesites as well as their theoretical C4/C3 signatures.Nevertheless, it must be noted that only theMexican site is associated with C4 plants. Theauthors would like to point out that this reason-ing is an approximation that does not take intoaccount the soil type and its possible conse-
A B
Fig. 10. Classification of all theenvironments involved in this studyhas been performed using anascendant cluster analysis. Thevariables used are: mean annualprecipitations, mean cold and warmseasonal temperatures, number ofdays of frost per year, mean locald18O isotopic signature of rains, andaltitude. Clusters determined fromthis analysis were drawn on thescatter plot of NFC isotopicsignature. These stable isotopicsignatures are well-discriminatedby the environmental variables asstated above.
1872 G. Cailleau et al.
� 2009 The Authors. Journal compilation � 2009 International Association of Sedimentologists, Sedimentology, 56, 1858–1875

quences on the soil atmosphere composition andbehaviour. As discussed above in the Crystalmorphology and genesis paragraph, an origin ofNFC through a high evaporation rate is pre-cluded. Therefore, either NFC is precipitateddirectly from parent rock and/or soil CO2 sourcesas a purely physicochemical product, or itundergoes a further fractionation due to biogenicprocesses. At this stage of isotopic investigation,it is difficult to conclude whether the isotopicsignature of NFC is related to biogenic fraction-ation or not. Nevertheless, the absence of anynegative shift between parent rock and NFCsignatures in some samples is related to thepresence of epitaxial features on NFC (see abovein the Crystal morphology and genesis para-graph). Epitaxial growth probably occurs underpurely physicochemical conditions, i.e. not re-lated to an organic template or a direct biogenicinfluence. Based on this interpretation, theabsence of a negative shift can be explained bythe disturbance of the original NFC isotopicsignature by physicochemical epitaxial over-growths. In Mexico, the same line of reasoningleads to the same conclusion: epitaxial growthhas a direct influence on NFC isotopic signatures,inducing the absence of a negative shift betweenparent rock and NFC. The fact that Mexicansamples have the highest values in d13C isexplained by the potential influence of C4 plants.
In addition, a cluster analysis based on envi-ronmental parameters of the studied areas showsa clear segregation between groups. This segrega-tion implies that the environmental factors (meanannual precipitations, mean cold and warm sea-sonal temperatures, number of days of frost peryear, mean local 18O isotopic signature of rains,and altitude) (Fig. 10) should influence in someway the stable isotope signatures of NFC. Never-theless, the sites where the evaporation rate is thehighest (i.e. African tropical sites due to highmean annual temperatures, or cold sites such asthe Jura Mountains because of cryo-fractionation)do not exhibit the highest d18O values. This point,the negative values of d18O data, and the exis-tence of a shift between these values and those ofthe rainwater source (expected to be the majorcontributor of the d18O values in the parentsolution), contradicts the assumption that theneedle morphology results from growth duringhigh evaporation. In conclusion, the use of NFCisotopic signature as a proxy for palaeoenviron-ments remains a difficult challenge. Diagenesis ofNFC, which is a fast and efficient process due tothe large surface area of crystals and the com-
plexity of the soil environment, easily erases theoriginal environmental fingerprint.
CONCLUSIONS
Needle fibre calcite (NFC) is often described inthe literature as the possible result of a highcrystal growth rate obtained during severe evap-oration, because of its occurrence above the watertable in soils. Nevertheless, the light oxygenisotopic signature measured for NFC contradictsthis hypothesis. In addition, severe evaporationcannot explain the divergence angle between thec axis and the needle lengthening, an anglebetween 30� and 50�. Observations of needlesinside organic microtubes provide strong evi-dence to explain rod shapes, because organicsleeves can act as a mould inducing this atypicalcrystal shape. The role of carboxyl groups, abun-dant in membrane molecules, can be suspected aspreferential nucleation sites, inducing a favouredorientation to the crystal nucleus. During thegrowth of the needle, the initial orientation is
Fig. 11. Synthetic sketch of NFC formation and evo-lution during early diagenesis in soils. Its origin andearly stages are related to organic biomineralization,whereas, once released in the soil medium, NFC canundergo outgrowths related to pedo-diagenetic fluids.
Biogenic origin of needle fibre calcite 1873
� 2009 The Authors. Journal compilation � 2009 International Association of Sedimentologists, Sedimentology, 56, 1858–1875

propagated along the crystal and the divergencebetween the c axis and the direction of needlelengthening remains constant if the initial orien-tation is relatively constant.
New observations have shed light on somerelated shapes (such as ‘whisker crystals’ and‘rhomb chains’) often attributed to needle fibrecalcite, but which are slightly different frompreviously described NFC morphologies. Theseneedles have a rod as their original template. Forexample, whisker crystals are constituted by a rodbearing large euhedral crystals that can coalesce;they are interpreted as the result of slow growthfrom a slightly supersaturated parent solution.Rhomb chains are constituted by successive epi-taxial steps of growth. At first, the growth rate mustbe relatively rapid but progressively slows down,forming crystal faces. These two features are theresult of early diagenetic processes in soil pores.
The isotopic comparison between NFC andcarbonate parent rock shows an almost systematicshift to more negative values of d13C. The light Cisotopic signature of needles cannot only beexplained by soil CO2 resulting from organicmatter decay. The C isotopic signature of needlefibre calcite depends on its morphology andenvironmental factors. The shift to more negativevalues of d13C between NFC and carbonate parentrock has to be interpreted as a possible biologicalfractionation. As demonstrated, environmentalfactors must also control part of the stable isotopesignatures but NFC, as a proxy for environmentalconditions, should be used with caution. Inconclusion, the biogenic origin of NFC, in itsoriginal form (Fig. 11), has been clarified and thepurely physicochemical hypothesis should berejected. This study shows that NFC forms insideorganic sleeves, although the micro-organismsresponsible for its formation have still not beenidentified. Nevertheless, epitaxial growth canoccur during diagenesis in the free soil medium(Fig. 11), the diagenesis being purely or partiallyphysicochemical. Consequently, it is suggestedthat the term ‘NFC’ be kept for the originalbiogenic form. The other habit should be referredto as ‘epitaxial forms of NFC’.
ACKNOWLEDGEMENTS
The authors would like to thank Dr C. Durlet forMexican samples, Prof. C. Dupraz for valuablediscussions, Dr M. Dadras, M. Vlimant andM. Leboeuf for SEM and TEM support, and M.Lettingue for field work. Many thanks to two
anonymous reviewers and Professor Peter Swartfor their substantial help in improving this man-uscript. This research has been funded by theSwiss National Foundation for scientific research,grants no. 2153-065174 and 205320-101564.
REFERENCES
Bajnoczi, B. and Kovacs-Kis, V. (2006) Origin of pedogenic
needle-fibre calcite revealed by micromorphology and stable
isotope composition – a case study of a Quaternary paleosol
from Hungary. Chem. Erde, 66, 203–212.
Becze-Deak, J., Langohr, R. and Verrecchia, E.P. (1997) Small
scale secondary CaCO3 accumulations in selected sections
of the European loess belt. Morphological forms and
potential for paleoenvironmental reconstruction. Geoder-
ma, 76, 221–252.
Borsato, A., Frisia, S., Jones, B. and van der Borg, K. (2000)
Calcite moonmilk: crystal morphology and environment of
formation in caves in the Italian Alps. J. Sed. Res., 70, 1179–
1190.
Braissant, O., Cailleau, G., Aragno, M. and Verrecchia, E.P.(2004) Biologically induced mineralization in the treeMilicia excelsa (Moraceae): its causes and consequences to
the environment. Geobiology, 2, 59–66.
Cailleau, G., Braissant, O. and Verrecchia, E.P. (2004) Bio-
mineralization in plants as a long-term carbon sink. Natur-
wissenschaften, 91, 191–194.
Cailleau, G., Braissant, O., Dupraz, C., Aragno, M. and
Verrecchia, E.P. (2005) Biologically induced accumu-
lations of CaCO3 in orthox soils of Biga, Ivory Coast.
Catena, 59, 1–17.
Callot, G., Guyon, A. and Mousain, D. (1985a) Inter-relations
entre aiguilles de calcite et hyphes myce1iens. Agronomie,
5, 209–216.
Callot, G., Mousain, D. and Plassard, C. (1985b) Concentra-
tions de carbonate de calcium sur les parois des hyphes
myceliens. Agronomie, 5, 143–150.
Carozzi, A.V. (1967) Recent calcite-cemented sandstone gen-
erated by the equatorial tree iroko (Chlorophora excelsa),
Daloa, Ivory Coast. J. Sed. Petrol., 37, 597–600.
Cerling, T.E. (1984) Stable isotope composition of modern soil
carbonate and its relationship to climate. Earth Planet. Sci.Lett., 71, 229–240.
Cerling, T.E. (1999) Stable carbon isotopes in paleosol car-
bonates. IAS Spec. Publ., 27, 43–60.
Gessner, K. (1555) Descriptio montis fracti sive Montis Pilat
ut vulgo nominat, juxta Lucernam in Helvetia. Lucernae,
1555.
Harrison, R.S. (1977) Caliches profiles: indicator of nearsur-
face subaerial diagenesis, Barbados West Indies. Bull. Can.
Petrol. Geol., 25, 123–173.
Iwanoff, L.L. (1906) Ein Wasserhaltiges Calcium Carbonat
Aussen Umgebungen von Nowo-Alexandria (guv. Lublin).
Ann. Geol. Mineral. Russland, 8, 23–25.
James, N.P. (1972) Holocene and Pleistocene calcareous crust
(caliche) profiles: criteria for subaerial exposure. J. Sed.
Petrol., 42, 817–836.
Jones, B. (1988) The influence of plants and micro-organisms
on diagenesis in caliche: example from the Pleistocene
ironshore formation on Cayman Brac, British West Indies.
Bull. Can. Petrol. Geol., 36, 191–201.
1874 G. Cailleau et al.
� 2009 The Authors. Journal compilation � 2009 International Association of Sedimentologists, Sedimentology, 56, 1858–1875

Jones, B. and Kahle, C.F. (1993) Morphology, relationship, and
origin of fibre and dendrite calcite crystals. J. Sed. Petrol.,
63, 1018–1031.
Jones, B. and Ng, K.-C. (1988) The structure and diagenesis of
rhizoliths from Cayman Brac, British West Indies. J. Sed.
Petrol., 58, 457–467.
Lacroix, A. (1901) Mineralogie de la France et de ses Anciens
Territoires d’Outre-Mer, Vol. 3. Librairie A. Blanchard,
Paris, 816 pp.
Loisy, C. (1997) Les encroutements carbonates Quaternaires
en substrat crayeux (Est du Bassin de Paris): role de la di-agenese et des actions biologiques. These de l’Universite de
Reims Champagne-Ardennes, 387 pp.
Loisy, C., Verrecchia, E.P. and Dufour, P.. (1999) Microbial
origin for pedogenic micrite associated with a carbon-
ate paleosol (Champagne, France). Sed. Geol., 126, 193–
204.
Mitterer, R.M. and Cunningham, R. (1985) The interaction of
natural organic matter with grain surfaces: implications for
calcium carbonate precipitation. In: Carbonate Cements
(Eds N. Schneidermann and P.M. Harris), SEPM Spec. Publ.,
36, 17–31.
Mouchet, P.O.J. (1998) Stratigraphy and mineralostratigraphy
of the Kimmeridgian in the central Jura Mountains of Swit-
zerland and eastern France. Eclogae Geol. Helv., 91, 53–68.
Mugge, O. (1914) Uber die lublinit genannte, augeblich neve
modification des kohlemsavsen kalkes. Zbl. Min. Geol.
Palaontol., 1914, 673–675.
Philips, S.E. and Self, P.G. (1987) Morphology, crystallogra-
phy and origin of needle-fibre calcite in Quaternary pedo-
genic calcretes of south Australia. Aust. J. Soil Res., 25, 429–
444.
Riche, G., Rambaud, D. and Riera, M. (1982) Etude morpho-
logique d’un encroutement calcaire, Region d’Irece, Bahia,
Bresil. Cah. Orstom, Serie Pedol., 19, 257–270.
Stoops, G.J. (1976) On the nature of ‘‘Lublinite’’ from Hollanta
(Turkey). Am. Mineral., 61, 172.
Strong, G.E., Giles, J.R.A. and Wright, V.P. (1992) A Holocene
calcrete from North Yorkshire, England: implications for
interpreting paleoclimates using calcretes. Sedimentology,
39, 333–347.
Supko, P.R. (1971) ‘‘Whisker’’ crystal cement in a Bahamian
rock. In: Carbonate Cements (Ed. O.P. Bricker), pp. 143–146.
Johns Hopkins University Press, Studies in Geology, 19,
Baltimore, MD.
Verges, V., Madon, M., Bruand, A. and Bocquier, G. (1982)
Morphologie et crystallogenese de monocristaux supergenes
de calcite en aiguilles. Bull. Mineral., 105, 351–356.
Verrecchia, E.P. (2000) Fungi and sediments. In: MicrobialSediments (Eds R. Riding and S.M. Awramik), pp. 68–75.
Springer Verlag, New York.
Verrecchia, E.P. and Verrecchia, K.E. (1994) Needle fiber
calcite: a critical review and a proposed classification.
J. Sed. Res., 64, 650–664.
Manuscript received 21 March 2008; revision accepted29 January 2009
Biogenic origin of needle fibre calcite 1875
� 2009 The Authors. Journal compilation � 2009 International Association of Sedimentologists, Sedimentology, 56, 1858–1875





























