Embed Size (px)
Citation preview

The importance of matrixmetalloproteinase-3 inrespiratory disordersExpert Rev. Respir. Med. 8(4), 411–421 (2014)
Cory M Yamashita1,Derek C Radisky2,Yael Aschner3,4 andGregory PDowney*3–5
1Department of Medicine, University of
Western Ontario, London, ON, Canada2Mayo Clinic Cancer Center,
Jacksonville, FL, USA3Departments of Medicine and
Pediatrics, Division of Pulmonary,
Critical Care, and Sleep Medicine,
National Jewish Health, Denver, CO,
USA4Department of Medicine, Division of
Pulmonary Sciences and Critical Care
Medicine, University of Colorado
Denver, Aurora, CO, USA5Department of Immunology, University
of Colorado Denver, Aurora, CO, USA
*Author for correspondence:
Tel.: +1 303 398 1436
Fax: +1 303 270 2243
The matrix metalloproteinases (MMPs) encompass a family of zinc-dependent endopeptidasesthat are secreted into the extracellular environment or remain bound to the cell surface. WhileMMPs were initially identified based on their ability to degrade collagen and other componentsof the extracellular matrix, recent studies indicate that their non-degradative functions arephysiologically paramount. In particular, MMPs are now known to participate in diversephysiological processes that control key aspects of inflammatory and immune responses andneoplasia, in part by selective triggering of cellular signaling pathways via limited proteolyticprocessing of extracellular and membrane-associated proteins, including cytokines and cellsurface receptors. Herein, we focus on the unique roles of MMP-3 (stromelysin-1) in acute lunginjury and repair, pulmonary fibrosis, and lung cancer.
KEYWORDS: acute lung injury • acute respiratory distress syndrome • epithelial mesenchymal transformation
• extracellular matrix • lung cancer • proteinase • proteolytic processing • pulmonary fibrosis • signal transduction
The matrix metalloproteinases (MMPs) includea family of more than 20 structurally relatedzinc-dependent endopeptidases that were ini-tially recognized for their ability to degrade col-lagen and other components of the extracellularmatrix (ECM), processes that play a criticalphysiological role in tissue remodeling inwound repair. More recent evidence, however,indicates that their nondegradative functionsmay be physiologically dominant [1,2]. MMPshave been subdivided based on their substratespecificity and structural properties into sub-groups that include gelatinases (MMP-2 and 9);stromelysins (MMP–3, 10 and 11); collagenases(MMP-1, 8 and 13); matrilysins (MMP-7 and26) and membrane-type MMPs (MMP-14, 15,16, 17, 24 and 25) [3]. It should be noted, how-ever, that classifying MMPs according to sub-strate specificity is largely based on in vitroobservations and may not reflect in vivo func-tion. As mentioned above, in addition to theirability to remodel the ECM, MMPs are able toposttranslationally modify soluble and mem-brane-associated proteins by a process termedlimited proteolytic processing, which can resultin activation or inactivation of cytokines,growth factors and cell surface receptors [4]. Byvirtue of their multiple biological activities,
MMPs are believed to participate in a variety ofphysiological processes such as repair of tissuesafter injury, inflammation and immune responses[5] and are key contributors to a diverse range ofpathological processes including rheumatoidarthritis and fibrosis of the liver, kidneys, heartand lungs [6–10]. Notably, MMP-3 (stromelysin-1) has been directly implicated in epithelial-mesenchymal transition (EMT), a phenotypicalteration in which epithelial cells detach fromneighboring cells and the underlying basementmembrane and become more motile and inva-sive [11,12]. Importantly, EMT may contribute tothe pathogenesis of neoplasia [13] and pulmonaryfibrosis [14], although the latter remains controver-sial [15–17].
There is mounting evidence that MMPshave pleiotropic effects in the context of tissueremodeling and repair that extend beyondtheir roles in matrix remodeling. In thisregard, an emerging literature suggests thatMMPs, including MMP-3, can regulate tissuerepair by altering the activity of other nonma-trix proteins including cytokines and mem-brane receptors [18,19], thus facilitating cellularsignaling through limited proteolytic process-ing and triggering activation of pathways lead-ing to EMT [13]. This phenomenon, whereby
informahealthcare.com 10.1586/17476348.2014.909288 � 2014 Informa UK Ltd ISSN 1747-6348 411
Review
Exp
ert R
evie
w o
f R
espi
rato
ry M
edic
ine
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Ron
a W
illia
mso
n on
07/
16/1
4Fo
r pe
rson
al u
se o
nly.

a class of tissue degrading enzymes can paradoxically promotethe deposition of excess tissue matrix (e.g., collagen), may beattributable in part to these nondegradative signaling functions.The current manuscript will review the involvement of MMPs,specifically MMP-3, in three pathological processes involvingthe lungs: acute lung injury (ALI) and repair, pulmonary fibro-sis and lung cancer. Although the mechanisms supporting therole of MMP-3 in the pathobiology of lung injury, repair andfibrosis may be more apparent, several lines of evidence pointtoward shared mechanisms underlying the development oftumor growth and cancer biology that will be highlighted inthe last section of this review.
Role of MMP-3 in acute lung injury & repairALI and the acute respiratory distress syndrome (ARDS) repre-sent a spectrum of diseases characterized by the development ofnoncardiogenic pulmonary edema, accompanied by an intenseneutrophilic pulmonary inflammatory response resulting from abroad spectrum of both direct (i.e., gastric aspiration, severepneumonia) and indirect lung insults (e.g., sepsis, multitrauma,transfusion of blood products). ARDS represents a large burdenof disease in the intensive care unit (ICU) population, withover 200,000 cases annually and a high mortality of approxi-mately 40% [20]. Several multicenter randomized clinical trialshave demonstrated improved outcomes in ARDS by the use ofpositive end-expiratory pressure (PEEP) [21], low tidal volumeventilation [22] and conservative fluid management strategies [23].Notably, while the use of high levels of PEEP has not beenshown to improve overall survival in ARDS when compared tolow levels of PEEP, the use of high levels of PEEP was associ-ated with decreased mortality in the ICU in the subgroup ofpatients with most severe lung injury (‘ARDS’) and improvedoxygenation status [24,25].
Recently, the definition of ARDS has been revised to reflectdifferences in blood oxygenation that allows greater predictivevalue in terms of morbidity, while the term ‘ALI’ has beenremoved from clinical diagnostic criteria [26]. Nonetheless, theterm ALI continues to be used in animal studies that areintended to model human ARDS, and for the purpose of thisreview, ALI/ARDS will be utilized interchangeably to reflecttheir common pathophysiology. The pathogenesis of ARDS/ALI is complex and involves neutrophil-dependent lung endo-thelial and epithelial cell injury, resulting in accumulation ofprotein-rich pulmonary edema as well as impaired removal ofedema fluid and resolution of inflammation [20,27].
The pleiotropic effects of MMPs have made them an attrac-tive area of focus in the context of ARDS. In the clinical realm,elevated levels of several MMPs have been identified in bron-choalveolar lavage fluid from patients with ARDS [28–30] and animbalance between MMPs and their endogenous inhibitors, thetissue inhibitors of MMPs (TIMPs), has been hypothesized toplay a role in disease pathogenesis [31]. Elevated levels ofMMP-1 and/or MMP-3 were found to be predictive of mortal-ity in patients with an established diagnosis of ARDS [28]. Stud-ies utilizing animal models have also provided evidence of the
importance of MMP-3 in the pathogenesis of ALI. For example,mice genetically deficient in MMP-3 or MMP-9 were protectedfrom lung injury triggered by intratracheal administration ofIgG immune complexes or macrophage inflammatory protein-2,a potent neutrophil chemotactic factor [32,33]. The MMP-3-defi-cient animals exhibited reduced neutrophil recruitment into thelungs, a finding that was not recapitulated in MMP-9 knockoutmice, suggesting that MMP-3 is specifically required for neutro-phil recruitment into the lungs and highlighting the concept thatthe mechanisms of involvement in ALI differ between individualMMPs. Coupled with the evidence that MMP-3-deficient ani-mals are protected from bleomycin-induced pulmonary fibro-sis [34], these observations support the hypothesis thatMMP-3 plays an important role in both lung injury and repair.Further studies in this area, in particular, those employing geneti-cally altered species of animals such as MMP-3-deficient mice areneeded to delineate how compensatory changes in other MMPsand/or endogenous inhibitors such as TIMPs may be impacted.Although the TIMPs have also been implicated in lung injury [35]
and repair [36], specific interactions between MMP-3 and endoge-nous TIMPs in the setting of ALI represents an area ofongoing investigation.
Although the mechanisms by which MMP-3 contributes tothe pathogenesis of ALI are incompletely understood, a numberof possibilities can be hypothesized from studies investigatingits role in disease models of other organs (FIGURE 1). MMP-3 isable to induce neutrophil chemoattractant activity in interleu-kin-1b-stimulated intestinal epithelial cells (Caco-3 cells) by amechanism involving proteolytic cleavage of chemotacticallyinert platelet basic protein yielding the active neutrophilchemokine (C-X-C motif) ligand-7 (neutrophil activating pep-tide 2) [37]. Additional studies have shown that chondrocyte-derived MMP-3 was capable of generating a macrophage che-motactic factor implicated in disc degeneration in a model ofdisc herniation resorption [38]. The specific mechanisms bywhich MMP-3 itself could promote neutrophil migration, ashas been observed in MMP-3-deficient models of ALI, areunknown and further studies characterizing the role ofMMP-3 in generating neutrophil-specific chemoattractants arerequired [39].
In addition to the intense neutrophilic and proinflammatoryresponses implicated in the pathogenesis of ALI/ARDS, breachof the alveolar capillary barrier facilitates the movement ofprotein-rich plasma into the alveoli and represents a hallmarkfeature of ARDS (FIGURE 1). Prior studies in mouse mammaryepithelial cells revealed that treatment with MMP-3 resulted inloss of intact E-cadherin, a key transmembrane protein in theformation of interepithelial cell (adherens) junctions [13]. Whilethis mechanism has been implicated in malignant transforma-tion of mammary epithelial cells in breast cancer, it remainsuncertain if MMP-3-mediated cleavage of E-cadherin contrib-utes to lung epithelial cell injury and thus to the pathogenesisof ALI.
A recent study further highlights the complexity of the localeffects of MMP-3 within the airspaces of the distal lung. In
Review Yamashita, Radisky, Aschner & Downey
412 Expert Rev. Respir. Med. 8(4), (2014)
Exp
ert R
evie
w o
f R
espi
rato
ry M
edic
ine
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Ron
a W
illia
mso
n on
07/
16/1
4Fo
r pe
rson
al u
se o
nly.

this study, shedding of the receptor foradvanced glycation end products (RAGE)from cultured rat alveolar epithelial cellsvia MMP-3- and MMP-13-mediated pro-teolysis yielded bioactive soluble products(sRAGE) [40]. Furthermore, a significantcorrelation was found between levels ofMMP-3 and MMP-13 and sRAGE inhuman ALI/ARDS bronchoalveolar lavagesamples [40]. Although elevated levels ofboth alveolar and circulating sRAGE cor-relate with adverse outcomes in the settingof ALI/ARDS, the possibility has beenraised that both MMP-3 and MMP-13could limit proinflammatory gene expres-sion in the lungs through decoy receptoractivities of sRAGE, thus providing a neg-ative feedback mechanism [41]. Furtherstudies will be required to delineate theprecise proteolytic processing activities ofMMP-3 with alveolar epithelial basedRAGE and how this interaction couldimpact pro- or anti-inflammatory responsesin the lungs.
Recently, the contribution of injuriousmechanical ventilation to lung injury inARDS has received considerable attention[42–44]. Evidence suggests that excessivemechanical stress can induce both macro-and microscopic injury to the lungs,resulting in proinflammatory responsesthat contribute to adverse patient out-comes. With this in mind, the involve-ment of MMPs in the pathogenesis oflung injury resulting from the effects ofmechanical stress has been an area ofrecent interest, with several recent studiesproviding an interesting perspective onthe role of MMPs in ventilator-inducedlung injury. These studies highlight a potentially complex inter-action between stress- and mechanical stretch-mediated releaseof MMPs that may individually promote acute inflammatoryresponses [45] or, conversely, protect against neutrophilic inflam-mation via modulation of airspace cytokine responses [46].Although these studies suggest a mechanism by whichMMP-3 could modulate the development of ALI in responseto ventilator-induced lung injury, specific studies addressingthis aspect of ALI are lacking and this remains an area requir-ing additional investigation.
In summary, multiple observations suggest the role ofMMP-3 in the pathogenesis of ALI via mechanisms such asneutrophil recruitment, epithelial cell injury and promotion ofacute inflammatory responses. MMP-3 may also play a role inresolution of ALI. However, it remains unclear whether theseenzymes represent nonspecific markers of tissue injury or
biomarkers of disease severity, or whether they play a moredirect role in disease initiation and/or progression and thusmight represent a relevant therapeutic target. Further investiga-tion and validation, including in models of direct and indirectALI, will be critical in determining the exact mechanisms bywhich MMP-3 promotes lung injury.
Role of MMP-3 in pulmonary fibrosis
Fibrotic lung diseases represent a diverse group of progressiveand ultimately fatal disorders collectively termed idiopathicinterstitial pneumonias, of which idiopathic pulmonary fibrosis(IPF) is the most common [47–50]. Clinically, IPF is character-ized by progressive dyspnea and a restrictive physiologicaldefect that is attributable to excessive and disorganized deposi-tion of ECM including collagen and fibronectin within thelung parenchyma [51]. Despite an improved understanding of
Edemafluid
Alveolus
Neutrophil chemotaxis
Tight andadherensjunctions
VE-Cadherin
E-Cadherin
ATI cell
Basementmembrane
MMP3
ATII cell
sRAGE
↑ Endothelialpermeability
↑ Endothelialpermeability Platelets CXCL7
Neutrophil
Pulmonarycapillary
Pulmonaryendothelium
Figure 1. The role of matrix metalloproteinase-3 in acute lung injury/acuterespiratory distress syndrome. Elevated levels of MMP-3 observed in the setting ofacute lung injury/acute respiratory distress syndrome have been demonstrated to provide achemotactic gradient for neutrophil recruitment to the lung interstitium and alveolar space,disrupt junctional proteins along vascular endothelial cells and degrade basementmembrane and cleave adherens junctions (i.e., E-cadherin) between alveolar epithelial celljunctions.ATI cell: Alveolar type I epithelial cell; ATII: Alveolar type II epithelial cell;CXCL-7: Chemokine (C-X-C motif) ligand 7; MMP-3: Matrix metalloproteinase 3;sRAGE: Soluble receptor for advanced glycation end products.
The importance of matrix metalloproteinase-3 in respiratory disorders Review
informahealthcare.com 413
Exp
ert R
evie
w o
f R
espi
rato
ry M
edic
ine
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Ron
a W
illia
mso
n on
07/
16/1
4Fo
r pe
rson
al u
se o
nly.

specific biological mechanisms that propagate this uncontrolledfibrotic process, limited progress has been made in the treat-ment of this devastating condition [47,50,52]. Both genetic andenvironmental factors appear to contribute to the pathogenesisof IPF [51,53–58]. The incidence of IPF increases with age [59]
possibly attributable to cumulative age-related genetic modifica-tions, which impair the ability of the lung to repair itself afterrepeated injury. This leads to release of profibrotic cytokinesand growth factors such as TGF-b and CTGF that inducefibroblast and myofibroblast accumulation, excessive depositionof ECM and progressive widespread lung fibrosis [50,51,60].
While it is evident that MMPs could participate in the deg-radation of the ECM during the injury phase, it is less intuitivethat a class of tissue degrading enzymes could also participate inexcessive deposition of ECM. Consistent with this concept,recent evidence from our group and others indicates thatMMPs can modify the composition of the ECM both directly
and via modulation of the activity of sol-uble or matrix-bound proteins such ascytokines and growth factors of mem-brane receptors, and through the genera-tion of signaling mediators derivedthrough these processes (FIGURE 2). Previousstudies from our laboratory demonstratedthat human fibroblasts selectively secreteMMP-3 (compared to other MMPs)when stimulated with the profibroticmediator IL-1b under conditions wherefocal adhesions are formed [61]. Ourgroup has also provided strong evidencefor the pathophysiological importance ofMMP-3 in the development of pulmo-nary fibrosis [34]. In this regard, wereported (FIGURE 3) that explanted lung tis-sues from patients with IPF expressedgreater levels of MMP-3 compared tocontrols [34]. Further in vivo studies dem-onstrated that the intratracheal administra-tion of recombinant MMP-3 viaadenoviral-mediated gene delivery eliciteda robust profibrotic response in rats, andconversely mice genetically deficient inMMP-3 were resistant to the developmentof bleomycin-induced pulmonary fibrosis(FIGURE 3) [34].
Excessive fibroblast accumulation(FIGURE 2), a hallmark feature of pulmonaryfibrosis, has been proposed to occurthrough a variety of mechanisms includ-ing proliferation of local pulmonaryfibroblasts; recruitment of circulatingfibrocytes and conversion of epithelialcells to a fibroblast phenotype, termedEMT. With respect to the latter process,Konigshoff and colleagues demonstrated
that activation of the canonical Wnt-b-catenin signaling path-way is a key regulator of alveolar type II cell hyperplasia andplasticity leading to EMT [62,63]. Additional studies from ourlaboratory revealed that exogenous MMP-3-induced activationof the Wnt-1 inducible signaling pathway protein, a targetgene of the Wnt-b-catenin pathway, in primary lung epithelialcells [34]. Furthermore, we demonstrated that bleomycin-exposed MMP-3-deficient mice exhibited reduced expression ofcyclin D1 (a target gene of the Wnt-b-catenin pathway)compared to wild-type control animals. Taken together, thesefindings support the role of MMP-3 in the development ofpulmonary fibrosis via mechanisms involving mesenchymaltransition of lung epithelial cells. It is noteworthy, however,that the importance of EMT as a source for the excess fibro-blasts in murine models of pulmonary fibrosis is controver-sial [16] and additional studies are needed to reconcile theseapparently discrepant observations.
Pulmonarycapillary
Myofibroblast
MMP3
↑ Collagen and fibronectin
ECMdegradation
Fibroblast
ECM
Alveolus
ATI cell
ATII cell
E-cadherin degradation
? EMT
Macrophage
Macrophage
Neutrophil
Pro-fibroticgrowth factors
NeutrophilFibrocyte
Monocyte
Figure 2. The role of matrix metalloproteinase-3 in pulmonary fibrosis. Elevatedlevels of MMP-3 observed in the setting of progressive diffuse pulmonary fibrosis such asidiopathic pulmonary fibrosis have been demonstrated to induce mesenchymal transitionof lung epithelial cells, induce myofibroblast differentiation of fibroblasts, degradecomponents of the basement membrane and thus induce profibrotic responsiveness offibroblasts and cleave adherens junctions (i.e., E-cadherin) between alveolar epithelialcell junctions.ATI cell: Alveolar type I epithelial cell; ATII: Alveolar type II epithelial cell; ECM: Extracellularmatrix; EMT: Epithelial-mesenchymal transition; MMP-3: Matrix metalloproteinase 3.
Review Yamashita, Radisky, Aschner & Downey
414 Expert Rev. Respir. Med. 8(4), (2014)
Exp
ert R
evie
w o
f R
espi
rato
ry M
edic
ine
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Ron
a W
illia
mso
n on
07/
16/1
4Fo
r pe
rson
al u
se o
nly.

More recently, Rajshankar and colleagues have provided furtherinsight into the potential mechanisms by which MMP-3 mayenhance a profibrotic milieu in the context of pulmonary fibro-sis [64]. In this study, proteolysis of the fibronectin matrix byMMP-3 yielded degradation products that promoted IL-1b-mediated cell-dependent anchorage and adhesion. Althoughexperiments conducted in this study were performed primarilyusing human gingival fibroblasts, such findings support thenotion that in addition to MMP-3-mediated triggering of
b-catenin-dependent signaling pathways, MMP-3-mediated alter-ations in the ECM can lead to profibrotic changes in the pheno-typic behavior of fibroblasts by modifying cell–matrix interactionspresumably by signaling through integrins and associated signal-ing molecules in the context of focal adhesions.
In summary, strong evidence from in vitro animal and humanstudies supports the importance of MMP-3 in the pathogenesisof pulmonary fibrosis. Nonetheless, many questions remain to beanswered such as whether enhanced expression of MMP-3 is
Ave
rag
e ex
pre
ssio
n IP
F
Average expression controlsR
elat
ive
MM
P3
exp
ress
ion
0
MMP9MMP7
MMP1
MMP11
MMP10
MMP13
MMP1
1
2
3
4
5
Control
0WT MMP-3
Null
NS BLM
WT MMP-3Null
Co
llag
en m
g/m
l
20
40
60
IPF
Con
trol
IPF
Alveoli Airway
4X PS stain 40X PS stain 40X H and E stain
WT
mou
seM
MP
3 nu
ll m
ouse
100
μm
100
μm10
0 μm
100
μm
A B
D E
C
Figure 3. Matrix metalloproteinase-3 is a critical mediator of idiopathic pulmonary fibrosis. (A) Gene expression array analysisof human IPF/UIP lung tissue samples demonstrates a significant increase in MMP-3 mRNA expression (q = 0) compared with non-IPF con-trol samples. Relative expression of other MMPs (i.e., MMP-1, -7, -9, -10, -11 and -13) are shown for comparison. (B) Quantitative RT-PCR analysis of explanted surgical lung specimens demonstrates a greater than fourfold increase in MMP-3 mRNA levels in IPF versusnon-IPF lung specimens (*p < 0.05 IPF vs control). (C) Immunohistochemical analysis of human control (top) and IPF (bottom) lung sec-tions to demonstrate cellular expression of MMP-3 (original magnification, 40). Representative sections from control lung demonstratethat MMP-3 staining (brown) is largely confined to alveolar macrophages (solid arrow). By contrast, in sections from IPF lung, there isstaining of alveolar macrophages (solid arrowheads, bottom left), alveolar epithelial cells (open arrowheads, bottom left and right), airwayepithelial cells (solid arrowheads, bottom right) and intravascular leukocytes (solid arrow, bottom right). Scale bars: 100 mm. (D) Lungsections (top) from wild-type (WT) and MMP-3-null mice treated with bleomycin (original magnification, 4 and 40 as indicated), stainedwith Picrosirius Red (PS) or with H&E demonstrating an increase in collagen (red staining) deposition in WT compared with MMP-3-nullmice. (E) Sircol assay of lung collagen content illustrating that bleomycin-treated MMP-3-null mice have significantly less collagen thanbleomycin-treated WT controls (*p < 0.05 WT BLM vs MMP-3-null BLM).BLM: Bleomycin; IPF: Idiopathic pulmonary fibrosis; MMP-3: Matrix metalloproteinase 3; NS: Normal saline.Data reprinted with permission from [34].
The importance of matrix metalloproteinase-3 in respiratory disorders Review
informahealthcare.com 415
Exp
ert R
evie
w o
f R
espi
rato
ry M
edic
ine
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Ron
a W
illia
mso
n on
07/
16/1
4Fo
r pe
rson
al u
se o
nly.

sufficient to induce clinical disease in humans in addition toidentifying the specific cellular sources of MMP-3 involved inthese fibrogenic processes. Until the discrete origins of diseasepathogenesis become more clearly defined and the precise mecha-nisms that result in the release of MMP-3 become evident, itremains uncertain whether therapeutic targeting of MMP-3 is aviable option to prevent pathological fibrosis.
Role of MMP-3-induced EMT in lung cancer progression
Lung cancer represents a major healthcare burden for bothdeveloped and developing nations and it is one that isincreasing in prevalence [65]. Lung cancers are the secondleading cause of morbidity and mortality globally, and amongthese cancers non-small-cell lung cancer represents the vastmajority (>80%) [65–67]. The principal environmental risk fac-tor for lung cancer is cigarette smoke exposure, althoughother environmental and genetic factors also pose significantrisk [65]. Increased expression of MMP-3 has been linked totumor growth and metastasis in human breast, colon, cervicaland lung cancers [68–73], but the mechanism by whichMMP-3 acts in human cancer development remains uncer-tain (FIGURE 4). In cell culture models, MMPs have been identi-fied as key inducers of EMT: MMP-3, MMP-7 andMMP-28 have all been shown to induce EMT in A549 lungadenocarcinoma cells [74–76]. Furthermore, MMP-3 has beenshown to induce EMT in human and mouse mammary epi-thelial cells [13,77,78] and in normal murine lung epithelialcells [34]. The role of MMP-3-induced EMT in cigarette
smoke-associated lung cancer development has recently beenidentified [79].
Insight into how MMPs could stimulate EMT was providedby transgenic mouse models with inducible expression ofMMP-3 in lung alveolar cells. Induction of MMP-3 in thesemice led to increased expression of Rac1b, an alternativelyspliced isoform of Rac1, that has previously been shown to reg-ulate actin cytoskeleton organization, membrane trafficking,proliferation and gene expression [80–83]. Rac1b results frominclusion of the 57 nucleotide exon 3b, which leads to a19-amino acid insertion in the protein sequence [84,85]. Theresulting Rac1b isoform accelerates GDP/GTP exchange andimpaired GTP hydrolysis [86–88]. Rac1b has been identified inmalignant colorectal [84], papillary thyroid [89] and breast can-cers [85,90]. Additional studies using cultured cells demonstratedthat Rac1b could stimulate oncogenic activities through theinduction of reactive oxygen species production and consequentpromotion of EMT [13,91,92]. Further studies implicated theinduction of Rac1b by MMP-3 as essential for MMP-3-in-duced EMT and acquisition of malignant characteristics in cul-tured lung epithelial cells [79].
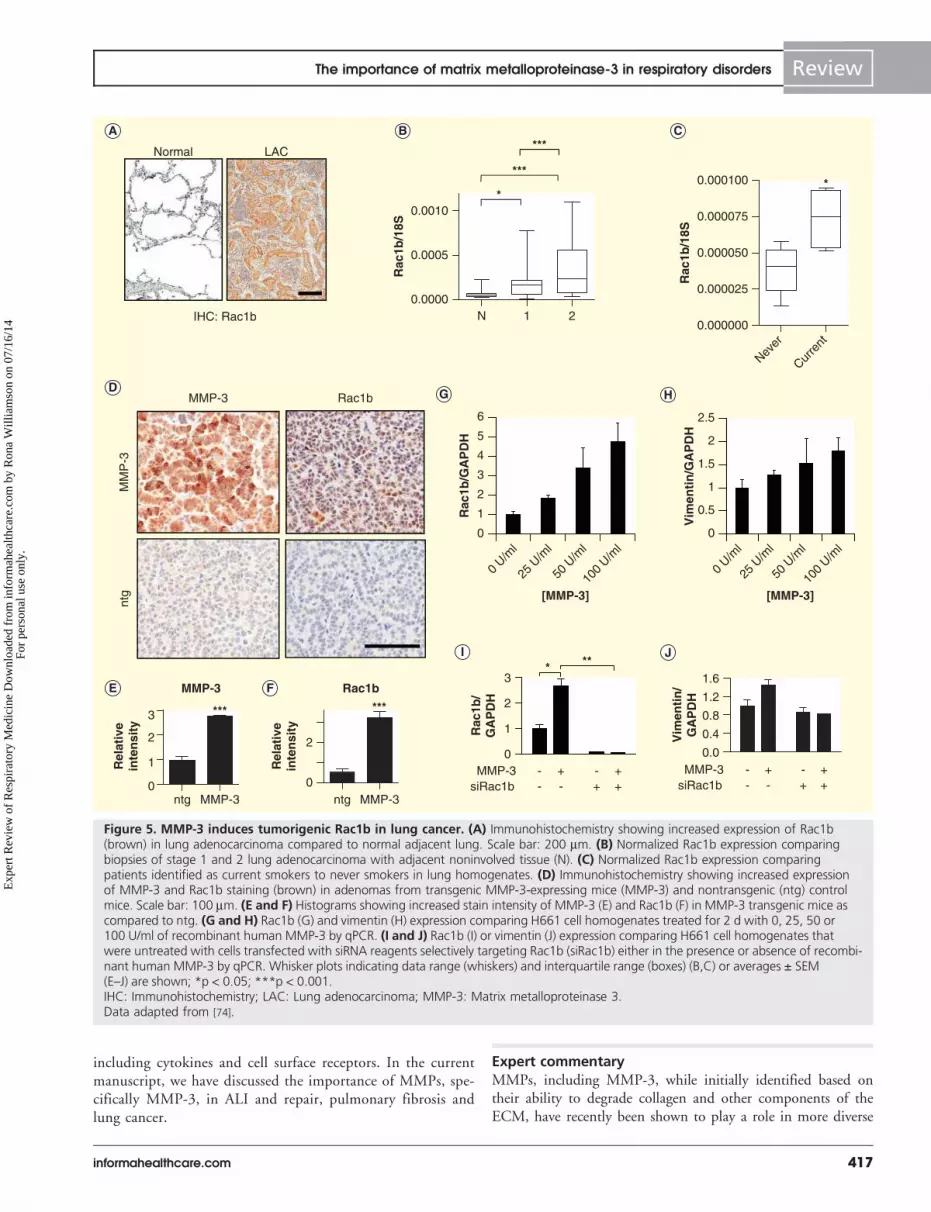
These results identified MMP-3-induced Rac1b as a poten-tial pathway for lung cancer development. In support of thispossibility, investigation of human biopsy tissue samplesrevealed that Rac1b was upregulated both in stages 1 and 2 oflung tumor progression and in response to cigarette smokeexposure (FIGURE 5). Additionally, analyses of transcriptional pro-files induced in lung epithelial cells exposed to MMP-3 showedsignificant similarities to lung tissue from smokers [79], impli-cating a possible MMP-Rac1b signaling axis in cigarettesmoke-associated lung cancer development. These observationsindicate a novel point of therapeutic intervention: while inhibi-tors of MMPs have not been successful anticancer agents [93],due in part to the requirement of these proteins in normalphysiological functions, Rac1b has been identified uniquely intumors and tumor-derived cell lines and has no known physio-logical functions. Thus, targeted inhibition of Rac1b functionis a potential therapeutic approach to attenuate MMP-3-in-duced EMT. Such a strategy would have multiple potentialbenefits as the cellular changes associated with EMT-like transi-tions alter EGFR-dependent signaling pathways and have beenlinked to lung tumor cell, xenograft and patient insensitivity toselective EGFR tyrosine kinase inhibitors [94–96]. BlockingMMP-3/Rac1b-induced EMT could therefore potentially sensi-tize tumor cells and enhance responses to existing therapieswhile independently reducing EMT-associated tumor progres-sion and metastasis of lung cancer.
Concluding remarksMMPs are key participants in diverse physiological and path-ophysiological processes that control key aspects of inflamma-tory and immune responses and neoplasia. In addition todegradation and remodeling of ECM proteins, MMPs caninitiate cellular signaling pathways via limited proteolyticprocessing of extracellular and membrane-associated proteins
MMP-3
GF;ROSTumor
formation
Fibrosis-,angiogenesis-
inducing factors
A
B
CD
E
Figure 4. The role of matrix metalloproteinase-3 in tumorformation. (A) MMP-3 triggers alveolar epithelial cells toincrease expression of Rac1b and consequently (B) undergoepithelial-mesenchymal transition, producing an activated cellthat can (C) generate GF and ROS that further disrupt normallung tissue structure, (D) fibrosis- and angiogenesis-inducing fac-tors that support developing tumors and also (E) directly lead totumor formation.GF: Growth factor; ROS: Reactive oxygen species; MMP-3: Matrixmetalloproteinase 3.
Review Yamashita, Radisky, Aschner & Downey
416 Expert Rev. Respir. Med. 8(4), (2014)
Exp
ert R
evie
w o
f R
espi
rato
ry M
edic
ine
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Ron
a W
illia
mso
n on
07/
16/1
4Fo
r pe
rson
al u
se o
nly.
including cytokines and cell surface receptors. In the currentmanuscript, we have discussed the importance of MMPs, spe-cifically MMP-3, in ALI and repair, pulmonary fibrosis andlung cancer.
Expert commentaryMMPs, including MMP-3, while initially identified based ontheir ability to degrade collagen and other components of theECM, have recently been shown to play a role in more diverse
A
D G
I
E F
B C
MMP-3
LAC
Rac1b
*** ***
0
2
1
ntg MMP-3 ntg MMP-3
3
MMP-3 Rac1b
0
2
Rel
ativ
ein
ten
sity
Rel
ativ
ein
ten
sity
MM
P-3
ntg
*
***
***
N 1 20.0000
0.0005
0.0010
Rac
1b/1
8S
Normal
IHC: Rac1b
Never
Curre
nt0.000000
0.000025
0.000050
0.000075
0.000100
[MMP-3][MMP-3]
Rac
1b/1
8S
*
Vim
enti
n/
GA
PD
H
0.0
0.4
0.8
1.2
1.6
Rac
1b/
GA
PD
H
0
1
2
3
siRac1bMMP-3
* **
siRac1b+ --MMP-3- +
++-
+ --- +
++-
H
J
0
1
2
3
4
5
6
0 U/m
l
25 U
/ml
50 U
/ml
100
U/ml
0 U/m
l
25 U
/ml
50 U
/ml
100
U/ml
Rac
1b/G
AP
DH
0
0.5
1
1.5
2
2.5
Vim
enti
n/G
AP
DH
Figure 5. MMP-3 induces tumorigenic Rac1b in lung cancer. (A) Immunohistochemistry showing increased expression of Rac1b(brown) in lung adenocarcinoma compared to normal adjacent lung. Scale bar: 200 mm. (B) Normalized Rac1b expression comparingbiopsies of stage 1 and 2 lung adenocarcinoma with adjacent noninvolved tissue (N). (C) Normalized Rac1b expression comparingpatients identified as current smokers to never smokers in lung homogenates. (D) Immunohistochemistry showing increased expressionof MMP-3 and Rac1b staining (brown) in adenomas from transgenic MMP-3-expressing mice (MMP-3) and nontransgenic (ntg) controlmice. Scale bar: 100 mm. (E and F) Histograms showing increased stain intensity of MMP-3 (E) and Rac1b (F) in MMP-3 transgenic mice ascompared to ntg. (G and H) Rac1b (G) and vimentin (H) expression comparing H661 cell homogenates treated for 2 d with 0, 25, 50 or100 U/ml of recombinant human MMP-3 by qPCR. (I and J) Rac1b (I) or vimentin (J) expression comparing H661 cell homogenates thatwere untreated with cells transfected with siRNA reagents selectively targeting Rac1b (siRac1b) either in the presence or absence of recombi-nant human MMP-3 by qPCR. Whisker plots indicating data range (whiskers) and interquartile range (boxes) (B,C) or averages ± SEM(E–J) are shown; *p < 0.05; ***p < 0.001.IHC: Immunohistochemistry; LAC: Lung adenocarcinoma; MMP-3: Matrix metalloproteinase 3.Data adapted from [74].
The importance of matrix metalloproteinase-3 in respiratory disorders Review
informahealthcare.com 417
Exp
ert R
evie
w o
f R
espi
rato
ry M
edic
ine
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Ron
a W
illia
mso
n on
07/
16/1
4Fo
r pe
rson
al u
se o
nly.

physiological processes through triggering of multiple cellularsignaling pathways via limited proteolytic processing of extra-cellular and membrane-associated proteins, including cytokines,growth factors and cell surface receptors. These pleiotropiceffects make MMP-3 a fascinating potential mediator in pul-monary pathological processes, including ALI/ARDS, fibrosisand cancer, which together make up a significant burden ofrespiratory disease. The ability of MMP-3 to produce activemediators of signaling pathways that regulate functions asdiverse as neutrophil recruitment, epithelial-to-mesenchymaltransition and inflammation, among others, may underliethe pathogenesis of these various disease states. In addition,recent work has shown that MMPs have the ability to alterthe underlying matrix, which may further enhance theproinflammatory and profibrotic milieu lead to dysfunc-tional repair resulting in fibrosis or neoplasia. Further workwill be required to more clearly define the role of MMPs,MMP-3 in particular, in respiratory disease. However, theseenzymes represent attractive potential therapeutic targetsthat are much needed in the treatment of these commonand often fatal lung diseases.
Five-year viewThe next 5 years will bring further mechanistic investigationinto the role of MMPs in human respiratory disease as well asin the pathogenesis of diseases of other organ systems. We willlikely see additional animal model studies of ALI that further
clarify how MMP-3 participates in neutrophil chemotaxis andbreakdown of the alveolar epithelial barrier. Work in the fieldof pulmonary fibrosis will need to focus on the contributionsof MMPs to the phenomenon of EMT as well as to the altera-tions in the matrix that are induced by proteolytic cleavage bythese enzymes and the way that these changes influence down-stream signaling cascades. Studies of lung cancer biology mayfocus on the potential for therapeutic blocking of EMTinduced by MMP-3-mediated formation of Rac1b. It will beuseful to determine whether MMPs represent direct players indisease initiation and progression, or whether they are morenonspecific markers of tissue injury. While human trials ofinhibitors of MMPs in the treatment of respiratory disease arelikely beyond the scope of the next 5 years, it may be possibleto see animal studies of these compounds initiated withinthis timeframe.
Financial & competing interests disclosure
C Yamashita has received grant funding from the Canadian Institute of
Health. D Radisky, R Zemans and G Downey have received grant fund-
ing from the National Institute of Health. The authors have no other rele-
vant affiliations or financial involvement with any organization or entity
with a financial interest in or financial conflict with the subject matter or
materials discussed in the manuscript. This includes employment, consul-
tancies, honoraria, stock ownership or options, expert testimony, grants or
patents received or pending or royalties.
No writing assistance was utilized in the production of this manuscript.
Key issues
• The matrix metalloproteinases (MMPs) are a family of enzymes with the ability to degrade components of the extracellular matrix
(ECM). They play critical roles in tissue remodeling and wound repair.
• MMPs also participate in limited proteolytic processing, resulting in activation and inactivation of soluble and membrane-associated
proteins, such as cytokines, growth factors and cell surface receptors. These functions may allow MMPs to paradoxically promote depo-
sition of excess ECM.
• MMP-3 has been implicated in a range of pathological processes, including acute lung injury (ALI), pulmonary fibrosis and lung cancer.
• MMP-3 levels have been shown to be elevated in clinical specimens of patients with ALI, and in animal studies and mice
genetically deficient in MMP-3 are protected in ALI models. This may be due to impaired neutrophil recruitment in
MMP-3 knockout mice.
• MMP-3 may also promote breakdown of alveolar epithelial barriers and acute inflammatory responses, particularly in the setting of
ventilator-induced lung injury: both mechanisms could contribute to the development of ALI.
• Genetic deletion of MMP-3 in mice confers protection from bleomycin-induced fibrosis, while transient overexpression of MMP-3 results
in profibrotic responses in rat lungs. This likely occurs by induction of the Wnt-b-catenin pathway by MMP-3, resulting in epithelial-
mesenchymal transition of lung alveolar epithelial cells.
• MMP-3-mediated degradation of the ECM enhances a profibrotic environment, which may affect the phenotype of fibroblasts and
promote further deposition of ECM and fibrosis.
• MMP-3 may promote progression and metastasis of lung cancer by increasing expression of Rac1b, an isoform of Rac1, which
has been implicated in various cancers through induction of reactive oxygen species and promotion of epithelial-mesenchymal
transition.
• Targeting of MMP-3 may be a potential therapeutic strategy for the treatment of multiple respiratory disease processes, including ALI,
fibrosis and lung cancer.
Review Yamashita, Radisky, Aschner & Downey
418 Expert Rev. Respir. Med. 8(4), (2014)
Exp
ert R
evie
w o
f R
espi
rato
ry M
edic
ine
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Ron
a W
illia
mso
n on
07/
16/1
4Fo
r pe
rson
al u
se o
nly.

References
Papers of special note have been highlighted as:
• of interest
•• of considerable interest
1. Page-McCaw A, Ewald AJ, Werb Z. Matrix
metalloproteinases and the regulation of
tissue remodelling. Nat Rev Mol Cell Biol
2007;8(3):221-33
2. Hadler-Olsen E, Fadnes B, Sylte I, et al.
Regulation of matrix metalloproteinase
activity in health and disease. FEBS J 2011;
278(1):28-45
3. Chakraborti S, Mandal M, Das S, et al.
Regulation of matrix metalloproteinases:
an overview. Mol Cell Biochem 2003;
253(1-2):269-85
4. Sternlicht MD, Werb Z. How matrix
metalloproteinases regulate cell behavior.
Annu Rev Cell Dev Biol 2001;17:463-516
5. Khokha R, Murthy A, Weiss A.
Metalloproteinases and their natural
inhibitors in inflammation and immunity.
Nat Rev Immunol 2013;13(9):649-65
6. Green MJ, Gough AK, Devlin J, et al.
Serum MMP-3 and MMP-1 and
progression of joint damage in early
rheumatoid arthritis. Rheumatology
(Oxford) 2003;42(1):83-8
7. Heymans S, Lupu F, Terclavers S, et al.
Loss or inhibition of uPA or
MMP-9 attenuates LV remodeling and
dysfunction after acute pressure overload in
mice. Am J Pathol 2005;166(1):15-25
8. Rosas IO, Richards TJ, Konishi K, et al.
MMP1 and MMP7 as potential peripheral
blood biomarkers in idiopathic pulmonary
fibrosis. PLoS Med 2008;5(4):e93
9. Uchinami H, Seki E, Brenner DA,
D’Armiento J. Loss of MMP 13 attenuates
murine hepatic injury and fibrosis during
cholestasis. Hepatology 2006;44(2):420-9
10. Zuo F, Kaminski N, Eugui E, et al. Gene
expression analysis reveals matrilysin as a
key regulator of pulmonary fibrosis in mice
and humans. Proc Natl Acad Sci USA
2002;99(9):6292-7
11. Radisky DC. Epithelial-mesenchymal
transition. J Cell Sci 2005;118(Pt 19):
4325-6
12. Weinberg RA. Twisted epithelial-
mesenchymal transition blocks senescence.
Nat Cell Biol 2008;10(9):1021-3
13. Radisky DC, Levy DD, et al. Rac1b and
reactive oxygen species mediate
MMP-3-induced EMT and genomic
instability. Nature 2005;436(7047):123-7
14. Selman M, Pardo A, Kaminski N.
Idiopathic pulmonary fibrosis: aberrant
recapitulation of developmental programs?
PLoS Med 2008;5(3):e62
15. Bartis D, Mise N, Mahida RY, et al.
Epithelial-mesenchymal transition in lung
development and disease: does it exist and is
it important? Thorax 2013; doi:10.1136/
thoraxjnl-2013-204608
16. Rock JR, Barkauskas CE, Cronce MJ, et al.
Multiple stromal populations contribute to
pulmonary fibrosis without evidence for
epithelial to mesenchymal transition. Proc
Natl Acad Sci USA 2011;108(52):E1475-83
17. Yamada M, Kuwano K, Maeyama T, et al.
Dual-immunohistochemistry provides little
evidence for epithelial-mesenchymal
transition in pulmonary fibrosis. Histochem
Cell Biol 2008;129(4):453-62
18. Maeda S, Dean DD, Gomez R, et al. The
first stage of transforming growth factor
beta1 activation is release of the large latent
complex from the extracellular matrix of
growth plate chondrocytes by matrix vesicle
stromelysin-1 (MMP-3). Calcif Tissue Int
2002;70(1):54-65
19. Lochter A, Galosy S, Muschler J, et al.
Matrix metalloproteinase
stromelysin-1 triggers a cascade of molecular
alterations that leads to stable
epithelial-to-mesenchymal conversion and a
premalignant phenotype in mammary
epithelial cells. J Cell Biol 1997;139(7):
1861-72
20. Matthay MA, Zemans RL. The acute
respiratory distress syndrome: pathogenesis
and treatment. Annu Rev Pathol 2011;6:
147-63
21. Mercat A, Richard JC, Vielle B, et al.
Positive end-expiratory pressure setting in
adults with acute lung injury and acute
respiratory distress syndrome: a randomized
controlled trial. JAMA 2008;299(6):646-55
22. Ventilation with lower tidal volumes as
compared with traditional tidal volumes for
acute lung injury and the acute respiratory
distress syndrome. The Acute Respiratory
Distress Syndrome Network. N Engl J Med
2000;342(18):1301-8
23. Wiedemann HP, Wheeler AP, Bernard GR,
et al. Comparison of two fluid-management
strategies in acute lung injury. N Engl J
Med 2006;354(24):2564-75
24. Briel M, Meade M, Mercat A, et al. Higher
vs lower positive end-expiratory pressure in
patients with acute lung injury and acute
respiratory distress syndrome: systematic
review and meta-analysis. JAMA 2010;
303(9):865-73
25. Santa Cruz R, Rojas JI, Nervi R, et al.
High versus low positive end-expiratory
pressure (PEEP) levels for mechanically
ventilated adult patients with acute lung
injury and acute respiratory distress
syndrome. Cochrane Database Syst Rev
2013;6:CD009098
26. ARDS Definition Task Force. Ranieri VM,
Rubenfeld GD, Thompson BT, et al. Acute
respiratory distress syndrome: the Berlin
Definition. JAMA 2012;307(23):2526-33
27. Matthay MA, Ware LB, Zimmerman GA.
The acute respiratory distress syndrome.
J Clin Invest 2012;122(8):2731-40
28. Fligiel SE, Standiford T, Fligiel HM, et al.
Matrix metalloproteinases and matrix
metalloproteinase inhibitors in acute lung
injury. Hum Pathol 2006;37(4):422-30
29. O’Kane CM, McKeown SW, Perkins GD,
et al. Salbutamol up-regulates matrix
metalloproteinase-9 in the alveolar space in
the acute respiratory distress syndrome. Crit
Care Med 2009;37(7):2242-9
30. Ricou B, Nicod L, Lacraz S, et al. Matrix
metalloproteinases and TIMP in acute
respiratory distress syndrome. Am J Respir
Crit Care Med 1996;154(2 Pt 1):346-52
31. Lanchou J, Corbel M, Tanguy M, et al.
Imbalance between matrix
metalloproteinases (MMP-9 and MMP-2)
and tissue inhibitors of metalloproteinases
(TIMP-1 and TIMP-2) in acute respiratory
distress syndrome patients. Crit Care Med
2003;31(2):536-42
32. Warner RL, Beltran L, Younkin EM, et al.
Role of stromelysin 1 and gelatinase B in
experimental acute lung injury. Am J Respir
Cell Mol Biol 2001;24(5):537-44
• An animal model study showed that
compared to wild type, mice genetically
deficient in MMP-9 and MMP-3 were
protected from the development of acute
lung injury in an experimental model of
acute lung injury. MMP-3-deficient mice
also showed impaired neutrophil
recruitment in response to injury.
33. Nerusu KC, Warner RL, Bhagavathula N,
et al. Matrix metalloproteinase-3
(stromelysin-1) in acute inflammatory tissue
injury. Exp Mol Pathol 2007;83(2):169-76
34. Yamashita CM, Dolgonos L, Zemans RL,
et al. Matrix metalloproteinase 3 is a
mediator of pulmonary fibrosis. Am J
Pathol 2011;179(4):1733-45
•• Expression of MMP-3 is enhanced in the
lungs of patients with idiopathic
pulmonary fibrosis. Animal model
experiments revealed increased fibrosis in
the setting of transient adenoviral
vector-mediated expression of
recombinant MMP-3, while genetic
The importance of matrix metalloproteinase-3 in respiratory disorders Review
informahealthcare.com 419
Exp
ert R
evie
w o
f R
espi
rato
ry M
edic
ine
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Ron
a W
illia
mso
n on
07/
16/1
4Fo
r pe
rson
al u
se o
nly.

deletion of MMP-3 in mice protected
against bleomycin-induced pulmonary
fibrosis. In vitro experiments suggest that
MMP-3 induces epithelial-mesenchymal
transition via cleavage of E-cadherin and
subsequent activation of b-catenin
signaling pathways.
35. Martin EL, McCaig LA, Moyer BZ, et al.
Differential response of TIMP-3 null mice
to the lung insults of sepsis, mechanical
ventilation, and hyperoxia. Am J Physiol
Lung Cell Mol Physiol 2005;289(2):
L244-51
36. Gill SE, Huizar I, Bench EM, et al. Tissue
inhibitor of metalloproteinases 3 regulates
resolution of inflammation following acute
lung injury. Am J Pathol 2010;176(1):
64-73
37. Kruidenier L, MacDonald TT, Collins JE,
et al. Myofibroblast matrix
metalloproteinases activate the neutrophil
chemoattractant CXCL7 from intestinal
epithelial cells. Gastroenterology 2006;
130(1):127-36
38. Haro H, Crawford HC, Fingleton B, et al.
Matrix metalloproteinase-7-dependent
release of tumor necrosis factor-alpha in a
model of herniated disc resorption. J Clin
Invest 2000;105(2):143-50
39. Van Lint P, Libert C. Chemokine and
cytokine processing by matrix
metalloproteinases and its effect on
leukocyte migration and inflammation.
J Leukoc Biol 2007;82(6):1375-81
40. Yamakawa N, Uchida T, Matthay MA,
Makita K. Proteolytic release of the receptor
for advanced glycation end products from
in vitro and in situ alveolar epithelial cells.
Am J Physiol Lung Cell Mol Physiol 2011;
300(4):L516-25
• In vitro experiments showed that the
release of soluble receptor for advanced
glycation end products (RAGE) from
cultured alveolar epithelial cells is
mediated by proteolysis by MMP-3 and
MMP-13 following LPS treatment.
Soluble RAGE expression in rat
bronchoalveolar lavage was increased by
MMP-3 and MMP-13 proteolysis and in
humans antigen levels of these enzymes
were positively correlated with RAGE
levels in pulmonary edema fluid.
41. Hergrueter AH, Nguyen K, Owen CA.
Matrix metalloproteinases: all the RAGE in
the acute respiratory distress syndrome. Am
J Physiol Lung Cell Mol Physiol 2011;
300(4):L512-15
42. Uhlig S, Ranieri M, Slutsky AS. Biotrauma
hypothesis of ventilator-induced lung injury.
Am J Respir Crit Care Med 2004;169(2):
314-15; author reply 315
43. Tremblay LN, Slutsky AS.
Ventilator-induced lung injury: from the
bench to the bedside. Intensive Care Med
2006;32(1):24-33
44. Fan E, Villar J, Slutsky AS. Novel
approaches to minimize ventilator-induced
lung injury. BMC Med 2013;11:85
45. Albaiceta GM, Gutierrez-Fernandez A,
Garcıa-Prieto E, et al. Absence or inhibition
of matrix metalloproteinase-8 decreases
ventilator-induced lung injury. Am J Respir
Cell Mol Biol 2010;43(5):555-63
46. Albaiceta GM, Gutierrez-Fernandez A,
Parra D, et al. Lack of matrix
metalloproteinase-9 worsens
ventilator-induced lung injury. Am J Physiol
Lung Cell Mol Physiol 2008;294(3):
L535-43
47. Raghu G, Collard HR, Egan JJ, et al. An
official ATS/ERS/JRS/ALAT statement:
idiopathic pulmonary fibrosis:
evidence-based guidelines for diagnosis and
management. Am J Respir Crit Care Med
2011;183(6):788-824
48. Maher TM. Idiopathic pulmonary fibrosis:
pathobiology of novel approaches to
treatment. Clin Chest Med 2012;33(1):
69-83
49. Nathan SD, Shlobin OA, Weir N, et al.
Long-term course and prognosis of
idiopathic pulmonary fibrosis in the new
millennium. Chest 2011;140(1):221-9
50. King TE Jr, Pardo A, Selman M. Idiopathic
pulmonary fibrosis. Lancet 2011;378(9807):
1949-61
51. Wynn TA. Integrating mechanisms of
pulmonary fibrosis. J Exp Med 2011;
208(7):1339-50
52. Downey GP. Resolving the scar of
pulmonary fibrosis. N Engl J Med 2011;
365(12):1140-1
53. Xaubet A, Marin-Arguedas A, Lario S, et al.
Transforming growth factor-beta1 gene
polymorphisms are associated with disease
progression in idiopathic pulmonary
fibrosis. Am J Respir Crit Care Med 2003;
168(4):431-5
54. Seibold MA, Wise AL, Speer MC, et al.
A common MUC5B promoter
polymorphism and pulmonary fibrosis. N
Eng J Med 2011;364(16):1503-12
55. Armanios MY, Chen JJ, Cogan JD, et al.
Telomerase mutations in families with
idiopathic pulmonary fibrosis. N Engl J
Med 2007;356(13):1317-26
56. Alder JK, Cogan JD, Brown AF, et al.
Ancestral mutation in telomerase causes
defects in repeat addition processivity and
manifests as familial pulmonary fibrosis.
PLoS Genet 2011;7(3):e1001352
57. Tsakiri KD, Cronkhite JT, Kuan PJ, et al.
Adult-onset pulmonary fibrosis caused by
mutations in telomerase. Proc Natl Acad Sci
USA 2007;104(18):7552-7
58. Fingerlin TE, Murphy E, Zhang W, et al.
Genome-wide association study identifies
multiple susceptibility loci for pulmonary
fibrosis. Nat Genet 2013;45(6):613-20
59. Raghu G, Weycker D, Edelsberg J, et al.
Incidence and prevalence of idiopathic
pulmonary fibrosis. Am J Respir Crit Care
Med 2006;174(7):810-16
60. Sisson TH, Mendez M, Choi K, et al.
Targeted injury of type II alveolar epithelial
cells induces pulmonary fibrosis. Am J
Respir Crit Care Med 2010;181(3):254-63
61. Wang Q, Rajshankar D, Laschinger C,
et al. Importance of protein-tyrosine
phosphatase-alpha catalytic domains for
interactions with SHP-2 and
interleukin-1-induced matrix
metalloproteinase-3 expression. J Biol Chem
2010;285(29):22308-17
• In vitro experiments showed that the
release of soluble receptor for advanced
glycation end products (RAGE) from
cultured alveolar epithelial cells is
mediated by proteolysis by MMP-3 and
MMP-13 following LPS treatment.
Soluble RAGE expression in rat
bronchoalveolar lavage was increased by
MMP-3 and MMP-13 proteolysis and in
humans antigen levels of these enzymes
were positively correlated with RAGE
levels in pulmonary edema fluid.
62. Konigshoff M, Kramer M, Balsara N, et al.
WNT1-inducible signaling
protein-1 mediates pulmonary fibrosis in
mice and is upregulated in humans with
idiopathic pulmonary fibrosis. J Clin Invest
2009;119(4):772-87
63. Konigshoff M, Balsara N, Pfaff EM, et al.
Functional Wnt signaling is increased in
idiopathic pulmonary fibrosis. PLoS One
2008;3(5):e2142
64. Rajshankar D, Downey GP, McCulloch CA.
IL-1beta enhances cell adhesion to degraded
fibronectin. FASEB J 2012;26(11):4429-44
•• In vitro experiments demonstrated that
IL-1b-mediated cell adhesion is promoted
by proteolytic degradation of the matrix
by MMP-3.
Review Yamashita, Radisky, Aschner & Downey
420 Expert Rev. Respir. Med. 8(4), (2014)
Exp
ert R
evie
w o
f R
espi
rato
ry M
edic
ine
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Ron
a W
illia
mso
n on
07/
16/1
4Fo
r pe
rson
al u
se o
nly.

65. Alberg AJ, Brock MV, Ford JG, et al.
Epidemiology of lung cancer: diagnosis and
management of lung cancer, 3rd ed:
American College of Chest Physicians
evidence-based clinical practice guidelines.
Chest 2013;143(5 Suppl):e1S-29S
66. Molina JR, Yang P, Cassivi SD, et al.
Non-small cell lung cancer: epidemiology,
risk factors, treatment, and survivorship.
Mayo Clin Proc 2008;83(5):584-94
67. Reck M, Heigener DF, Mok T, et al.
Management of non-small-cell lung cancer:
recent developments. Lancet 2013;
382(9893):709-19
68. Lievre A, Milet J, Carayol J. Genetic
polymorphisms of MMP1, MMP3 and
MMP7 gene promoter and risk of colorectal
adenoma. BMC Cancer 2006;6:270
69. Hagemann T, Bozanovic T, Hooper S,
et al. Molecular profiling of cervical cancer
progression. Br J Cancer 2007;96(2):321-8
70. Fang S, Jin X, Wang R, et al.
Polymorphisms in the MMP1 and
MMP3 promoter and non-small cell lung
carcinoma in North China. Carcinogenesis
2005;26(2):481-6
71. Liu D, Guo H, Li Y, et al. Association
between polymorphisms in the promoter
regions of matrix metalloproteinases
(MMPs) and risk of cancer metastasis:
a meta-analysis. PLoS One 2012;7(2):
e31251
72. Mendes O, Kim HT, Stoica G. Expression
of MMP2, MMP9 and MMP3 in breast
cancer brain metastasis in a rat model. Clin
Exp Metastasis 2005;22(3):237-46
73. Sun T, Gao Y, Tan W, et al. Haplotypes in
matrix metalloproteinase gene cluster on
chromosome 11q22 contribute to the risk of
lung cancer development and progression.
Clin Cancer Res 2006;12(23):7009-17
74. Radisky DC, Przybylo JA. Matrix
metalloproteinase-induced fibrosis and
malignancy in breast and lung. Proc Am
Thorac Soc 2008;5(3):316-22
• Transgenic expression of MMP-3 in mice
results in fibrosis and tumor formation,
which is due to the induction of EMT, a
process which is dependent on the
sgeneration of reactive oxygen species.
75. Illman SA, Lehti K, Keski-Oja J, Lohi J.
Epilysin (MMP-28) induces TGF-beta
mediated epithelial to mesenchymal
transition in lung carcinoma cells. J Cell Sci
2006;119(Pt 18):3856-65
76. McGuire JK, Li Q, Parks WC. Matrilysin
(matrix metalloproteinase-7) mediates
E-cadherin ectodomain shedding in injured
lung epithelium. Am J Pathol 2003;162(6):
1831-43
77. Lochter A, Sternlicht MD, Werb Z, et al.
The significance of matrix
metalloproteinases during early stages of
tumor progression. Ann N Y Acad Sci
1998;857:180-93
78. Noel A, Boulay A, Kebers F, et al.
Demonstration in vivo that
stromelysin-3 functions through its
proteolytic activity. Oncogene 2000;19(12):
1605-12
79. Stallings-Mann ML, Waldmann J, Zhang Y,
et al. Matrix metalloproteinase induction of
Rac1b, a key effector of lung cancer
progression. Sci Transl Med 2012;4(142):
142ra95
•• Transgenic expression of Rac1b in mouse
lung epithelial cells stimulated EMT and
spontaneous tumor development by
MMP-induced mechanisms. Rac1b was
also found to be expressed in early-stage
human lung adenocarcinomas.
80. Bar-Sagi D, Hall A. Ras and Rho GTPases:
a family reunion. Cell 2000;103(2):227-38
81. Boettner B, Van Aelst L. The role of Rho
GTPases in disease development. Gene
2002;286(2):155-74
82. Price LS, Collard JG. Regulation of the
cytoskeleton by Rho-family GTPases:
implications for tumour cell invasion. Semin
Cancer Biol 2001;11(2):167-73
83. Sahai E, Marshall CJ. RHO-GTPases and
cancer. Nat Rev Cancer 2002;2(2):133-42
84. Jordan P, Brazao R, Boavida MG, et al.
Cloning of a novel human Rac1b splice
variant with increased expression in
colorectal tumors. Oncogene 1999;18(48):
6835-9
85. Schnelzer A, Prechtel D, Knaus U, et al.
Rac1 in human breast cancer:
overexpression, mutation analysis, and
characterization of a new isoform, Rac1b.
Oncogene 2000;19(26):3013-20
86. Fiegen D, Haeusler LC, Blumenstein L,
et al. Alternative splicing of Rac1 generates
Rac1b, a self-activating GTPase. J Biol
Chem 2004;279(6):4743-9
87. Matos P, Collard JG, Jordan P.
Tumor-related alternatively spliced Rac1b is
not regulated by Rho-GDP dissociation
inhibitors and exhibits selective downstream
signaling. J Biol Chem 2003;278(50):
50442-8
88. Orlichenko L, Geyer R, Yanagisawa M,
et al. The 19-amino acid insertion in the
tumor-associated splice isoform Rac1b
confers specific binding to p120 catenin.
J Biol Chem 2010;285(25):19153-61
89. Silva AL, Carmo F, Bugalho MJ. RAC1b
overexpression in papillary thyroid
carcinoma: a role to unravel. Eur J
Endocrinol 2013;168(6):795-804
90. Pelisch F, Khauv D, Risso G, et al.
Involvement of hnRNP A1 in the matrix
metalloprotease-3-dependent regulation of
Rac1 pre-mRNA splicing. J Cell Biochem
2012;113(7):2319-29
91. Nelson CM, Khauv D, Bissell MJ,
Radisky DC. Change in cell shape is
required for matrix
metalloproteinase-induced epithelial-
mesenchymal transition of mammary
epithelial cells. J Cell Biochem 2008;105(1):
25-33
92. Lee K, Chen QK, Lui C, et al. Matrix
compliance regulates Rac1b localization,
NADPH oxidase assembly, and
epithelial-mesenchymal transition. Mol Biol
Cell 2012;23(20):4097-108
93. Coussens LM, Fingleton B, Matrisian LM.
Matrix metalloproteinase inhibitors and
cancer: trials and tribulations. Science 2002;
295(5564):2387-92
94. Thomson S, Buck E, Petti F, et al.
Epithelial to mesenchymal transition is a
determinant of sensitivity of non-small-cell
lung carcinoma cell lines and xenografts to
epidermal growth factor receptor inhibition.
Cancer Res 2005;65(20):9455-62
95. Witta SE, Gemmill RM, Hirsch FR, et al.
Restoring E-cadherin expression increases
sensitivity to epidermal growth factor
receptor inhibitors in lung cancer cell lines.
Cancer Res 2006;66(2):944-50
96. Yauch RL, Januario T, Eberhard DA, et al.
Epithelial versus mesenchymal phenotype
determines in vitro sensitivity and predicts
clinical activity of erlotinib in lung cancer
patients. Clin Cancer Res 2005;11(24 Pt 1):
8686-98
The importance of matrix metalloproteinase-3 in respiratory disorders Review
informahealthcare.com 421
Exp
ert R
evie
w o
f R
espi
rato
ry M
edic
ine
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Ron
a W
illia
mso
n on
07/
16/1
4Fo
r pe
rson
al u
se o
nly.





























