Embed Size (px)
Citation preview

Full Paper
322
The Influence of Silkworm Species on CellularInteractions with Novel PVA/SilkSericin Hydrogels
Khoon S. Lim, Joydip Kundu, April Reeves, Laura A. Poole-Warren,Subhas C. Kundu, Penny J. Martens*
Sericin peptides and PVA are chemically modified with methacrylate groups to produce acovalent PVA/sericin hydrogel. Preservation of the sericin bioactivity following methacryl-ation is confirmed, and PVA/sericin hydrogels are fabricated for both B. mori and A. mylittasericin. Cell adhesion studies confirm the preservationof sericin bioactivity post incorporation in PVA gels.PVA/A. mylitta gels are observed to facilitate celladhesion to a significantly greater degree thanPVA/B. mori gels. Overall, the incorporation of sericindoes not alter the physical properties of the PVA hydro-gels but does result in significantly improved cellularinteraction, particularly from A. mylitta gels.
1. Introduction
Hydrogels are soft-crosslinked polymers with high water
content and good macromolecular diffusivity. Although
these characteristics are similar to the extracellular matrix
(ECM), our group and others have recognized the critical
need for incorporation of biological molecules into syn-
thetic hydrogels to augment biofunctionality. Of particular
importance for tissue engineering applications are
improvements in cell adhesive and cell signaling proper-
ties.[1–3] ECM proteins such as collagen[4,5] and its derivative
gelatin,[6,7] laminin[8,9] and glycosaminoglycans such as
K. S. Lim, A. Reeves, Prof. L. A. Poole-Warren, Dr. P. J. MartensGraduate School of Biomedical Engineering, University of NewSouth Wales, Sydney 2052, AustraliaE-mail: [email protected]. Kundu, Prof. S. C. KunduDepartment of Biotechnology, Indian Institute of Technology,Kharagpur 721302, India
K.S.Lim and J. Kundu contributed equally to this paper.
Macromol. Biosci. 2012, 12, 322–332
� 2012 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim wileyonline
chondroitin sulfate[1] and heparin,[3] are the most com-
monly studied biological modifiers for synthetic hydrogels.
More recently, the silk protein sericin has shown promise
for enhancing cell interactions with hydrogels that have
otherwise low interactivity.[2,10,11] Sericin is a protein
secreted from the middle silk gland of a mature silkworm
larva and acts as the glue for adhesion of fibroin based fibers
during cocoon formation.[12,13] The glue-like protein is
composed of random coil and b-sheet secondary structures
with a high abundance of hydrophilic amino acids that
confers water solubility.[10,12,14–16] It has been previously
reported that sericin from the domesticated silkworm
Bombyxmorihas antioxidant properties as well as being UV
protective, cryoprotective and chemoprotective.[13,17–22] In
terms of cellular interactions, there is some evidence that
B. Mori sericin promotes cell adhesion and cell survival
of fibroblasts,[20,23] epithelial cells[24] and osteoblasts,[11]
although the mechanism for this is unknown. As there are
no mammalian proteins or peptides possessing multi-
functional characteristics such as those of sericin, it is of
interest to copolymerize sericin with synthetic polymers to
library.com DOI: 10.1002/mabi.201100292

The Influence of Silkworm Species on Cellular Interactions with . . .
www.mbs-journal.de
create biosynthetic hydrogels, and to study whether the
bioactivity of sericin is transmitted to the gel. Other
advantages are that the source of sericin is sustainable and
the likelihood of viral and prion transfer tends to be lower
for insect proteins compared with mammalian proteins.
While the majority of research on silk proteins has
centered on proteins derived from B. mori, there is a
growing interest in other silkworm species. Sericin
extracted from the wild-type silkworm Antheraea mylitta,
appears to be significantly different to that of its
domesticated cousin. B. mori sericin consists of proteins
with major molecular weight fractions at 150, 250 and
400 kDa while A. mylitta sericin has fraction at 70, 200 and
>200 kDa.[12,15,16] Moreover, antibody cross-reactivity
between A. mylitta sericin and B. mori sericin is variable
suggesting that the two sericins are biochemically differ-
ent.[12] A. mylitta sericin has also been reported to have
lower tyrosine and serine contents than B. mori sericin.[25]
However there has been little research directly comparing
the bio-functionality of the two species which will be
addressed in this study.
The key challenges for incorporating proteins such as
sericin in hydrogels are in the formation of stable, covalent
linkage of the protein to the synthetic polymer and in
maintenance of the function of the biological molecule after
integration. Another key issue revolves around using the
minimum biological information required to achieve the
desired cellular functions. By reducing the amount of
protein used and achieving efficient incorporation, the
physical and mechanical properties of the base polymer are
minimally perturbed and likely to remain stable following
fabrication and use in specific applications.
Previous studies have produced sericin-based gels via
simply blending the sericin into gels,[20] by ethanol
treatment to induce gelation, or crystallinity of seri-
cin[10,26,27] or have integrated sericin with synthetic
hydrogels using chemical crosslinkers such as glutaralde-
hyde.[2,17,28] The problems with such approaches relate to
the stability of gels in the former instances and the
likelihood for chemical toxicity and regulatory concerns in
the latter.
The present study proposes an approach to produce a
copolymerized network of sericin and poly(vinyl alcohol)
(PVA) to form a dimensionally stable hydrogel with
improved biological performance. Several studies have
shown that PVA can be functionalized with acrylate based
side groups,[1,3,29] and photopolymerized to form cross-
linked hydrogels in the presence of a non-toxic, water
soluble initiator.[30] This research will study the feasibility
of chemically modifying sericin proteins using equivalent
approaches to those used for functionalizing PVA. This will
be extended to examine the impact of this modification on
selected biological functions of sericin. Finally, the bioac-
tivity of functionalized sericin after incorporation into PVA
www.MaterialsViews.com
Macromol. Biosci. 20
� 2012 WILEY-VCH Verlag Gmb
hydrogels will be examined based on cell responses to this
co-hydrogel.
2. Experimental Section
2.1. Macromer Synthesis
2.1.1. Isolation and Characterization of Sericin from
Silkworm Cocoons
A. mylitta and B. mori silk cocoons were collected from Jhargram
Tropical Tasar Farms and Debra Sericulture Farm, West Midnapore,
West Bengal, India, respectively. Silk sericin peptides were
extracted from the cocoons according to a method modified from
Sofia et al.[31] In brief, A. mylitta and B. mori cocoons were cut into
small pieces then boiled in excess sodium carbonate (Na2CO3)
solution (0.02 M) for 1 and 4 h, respectively. The supernatant was
centrifuged at 1000 rpm for 10 min to remove fibers and impurities,
then dialyzed against deionized water for 48 h in a 10 kDa
molecular weight cut-off tubing. The solution was then lyophilized
to obtain dried sericin. The extracted sericin was characterized by
sodium dodecylsulfate polyacrylamide gel electrophoresis (SDS-
PAGE) to confirm the presence and molecular weights of the
peptides. The peptides were separated by using an 8% SDS-PAGE gel
at a constant voltage of 80 V for approximately 3 h. Peptide bands
were visualized using Coomassie Brilliant Blue R-250 (Sigma) and a
methanol/water 1:1 destaining solution that contained 20% acetic
acid.
2.1.2. Synthesis of PVA Methacrylate (PVA-MA)
PVA-MA was prepared by reacting PVA (Sigma-Aldrich, 13–23 kDa,
98% hydrolyzed) with 2-isocyanatoethyl methacrylate (ICEMA)
(Sigma-Aldrich, 98% purity) in dimethyl sulfoxide (DMSO) accord-
ing to a method described by Bryant et al.[1] In a typical experiment,
a 10 wt% solution of PVA (10 g) in DMSO (100 mL) was heated to
60 8C under nitrogen atmosphere until the PVA was completely
dissolved. ICEMA (0.405 cm3) was then added with vigorous stirring
and the solution was left to react for 4 h. To stop the reaction, the
solution was precipitated in toluene. The precipitated polymer was
redissolved in water and then purified by ultrafiltration through a
10 kDa molecular weight cut-off membrane. Lastly, the purified
solution was sterilized by filtration through a 0.22mm sterile filter,
and then lyophilized to obtain sterile dried PVA-MA.
2.1.3. Synthesis of Sericin Methacrylate (Sericin-MA)
Sericin was methacrylated using a similar method to PVA but with
slight modifications.[1] In brief, sericin (1 g) was dissolved in lithium
chloride (LiCl) in DMSO solution (100 cm3, 1 M) to form 1 wt%
solution. The solution was then purged with N2 gas for 30 min,
followed by the addition of ICEMA (0.0283 cm3), then left to react for
5 h under N2 atmosphere. The reaction was stopped by precipitat-
ing the solution in ethanol, and then centrifuged at 1000 rpm for
10 min to collect the precipitate. Purification was done by re-
dissolving the sericin-MA in water then dialyzed against deionized
water for 48 h in a 10 kDa molecular weight cut-off tubing. The
purified sericin-MA solution was filtered through a 0.22mm sterile
filter, and then lyophilized to obtain sterile dried sericin-MA.
12, 12, 322–332
H & Co. KGaA, Weinheim323

324
www.mbs-journal.de
K. S. Lim, J. Kundu, A. Reeves, L. A. Poole-Warren, S. C. Kundu, P. J. Martens
2.2. Macromer Characterization
2.2.1. Macromer Characterization Using NMR
Both PVA-MA and sericin-MA were dissolved in deuterium oxide
(D2O) then analyzed using 1H NMR (300 MHz Bruker Advance DPX-
300 spectrometer) to quantify the amount of methacrylate groups
attached. To calculate percent methacrylation for PVA, the area of
methacrylate vinyl proton peaks, d¼6.1 (s, H1) and 5.8 (s, H2) was
compared to the area of the protons in the PVA backbone, d¼ 4.0
(s, H3).[3,32] Similarly, percent methacrylation of sericin is calculated
by comparing the area of the methacrylate vinyl proton peaks to
the area of the protons associated with the serine and glycine
groups, d¼3.6–4.0 (m, Ser bCH2 and Gly aCH2), in the sericin
backbone.[33] The number of crosslinkers per chain was then
calculated from
Crossl
% Cel
inker per chain ¼ % methacrylation �MWpolymer
MWRU(1)
where, MW is molecular weight and RU is the repeating unit of the
polymer.
2.2.2. L929 Cell Growth Inhibition Study
Samples (PVA, PVA-MA, sericin and sericin-MA) were prepared at
concentrations of 4 mg �mL�1 (in 0.9% saline) and diluted to
1 mg �mL�1 with Eagle’s minimum essential medium (EMEM)
containing 10% fetal bovine serum (FBS) and 1% penicillin/
streptomycin (PS). Murine dermal fibroblasts (L929) at a concen-
tration of 5�104 cells �mL�1 were seeded onto 35-mm diameter
tissue culture dishes. After 24 h of incubation at 37 8C in a 5% CO2
humidified atmosphere, the media was discarded and 1 mL of
sample solution was added to the cells. Following an additional 48 h
of incubation, the cells were trypsinized, and counted with a cell
viability analyzer (Vi-cell XR, Beckman Coulter). The percentage cell
growth inhibition was calculated according to
l growth inhibition ¼ 1 � Number of cells in sample dish
Number of cells in media control dish
(2)
2.2.3. Antioxidant Properties of Sericin
The antioxidant properties of sericin-MA compared to the non-
methacrylated sericin isolated from both B. mori and A. mylitta
cocoons was determined following a previously described proto-
col.[15] In brief, human keratinocytes (HaCaT cells) were seeded in
tissue culture dishes (35 mm diameter) at a density of
1�106 cells �mL�1 and allowed to attach and grow for 24 h. The
cells were pre-treated with sericin and sericin-MA (50 and
100 ng �mL�1) for 24 h and then subjected to hydrogen peroxide
(H2O2, 0.8�10�3M) induced oxidative stress. The pre-treated and
stressed cells were then washed and harvested by trypsinization.
The cells were resuspended in sterile PBS and homogenized then
centrifuged for 10 min at 5 000 rpm to collect the supernatant of
each sample for biochemical assays. HaCaT cell damage caused
by H2O2 was quantitatively assessed from the ratio of catalase,
lactate dehydrogenase (LDH) and malondialdehyde (MDA) released
Macromol. Biosci. 20
� 2012 WILEY-VCH Verlag Gmb
from damaged cells to the undamaged cells after the induction of
oxidative stress. Catalase is an enzyme secreted when cells are
oxidative damaged, and was determined by measuring the
reduction in absorbance of NADH at 240 nm for 5 min following
the breakdown of hydrogen peroxide. LDH activity corresponding
to the cell membrane integrity was expressed as change in
absorbance of the reaction mixture at 340 nm per min following the
breakdown of pyruvate by the enzyme on addition of NADH. MDA
concentration in the medium, which reflects the degree of lipid
peroxidation, was quantified by measuring the absorbance at
535 nm following reaction with thiobarbituric acid reagent
(0.5 cm3, 1:1 v/v, mixture of 0.67% thiobarbituric acid and acetic
acid).
2.2.4. L929 Cell Adhesion and Proliferation Studies on
Sericin Coatings
24 well polystyrene plates (well area of 2 cm2), were coated with
sericin and sericin-MA. In brief, sterile filtered sericin and sericin-
MA solutions were added to the wells then allowed to dry on a hot
plate kept at 37 8C in a sterile laminar hood for 2 h. All the coatings
were at a final concentration of 25mg � cm�2. Tissue culture plastics
(TCP), i.e. non-coated wells, were used as controls in this
experiment. L929 fibroblasts suspended in EMEM containing
10% FBS and 1% PS were seeded on the coatings and controls at
a concentration of 6000 cells �mL�1, which corresponds to a density
of 30 cells �mm�2. At the time points of interest (4 h, 1 d and 4 d), the
media and non-adherent cells were removed from the well, and
1 mL of fresh media were re-applied to each well. Each well was
then photographed (3 pictures per well) using a phase contrast
microscope at 10� magnification. ImageJ was used to count the
attached cells.
2.3. Fabrication and Characterization of PVA/Sericin
Hydrogels
2.3.1. Fabrication of PVA/Sericin Hydrogels
Dried PVA-MA was dissolved in water at 80 8C.[3] Upon complete
dissolution, the solution was cooled to room temperature and the
photoinitiator, 2-hydroxy-1-[4-hydroxyethoxy(phenyl)]-2-methyl-
1-propanone (Irgacure 2959, Ciba Specialty Chemicals), was added
to the polymer solution (0.05 wt%). The samples were UV-irradiated
(Greenspot, peaks 310–365 nm, 30 mW � cm�2) for 3 min.[3,32,34] To
prepare PVA/sericin hydrogels, both PVA-MA and sericin-MA were
dissolved separately then mixed at room temperature prior to
addition of initiator. The total content of macromer in the hydrogels
was 20 wt%.
2.3.2. Swelling Study and Mass Loss Analysis
Hydrogel discs (10 mm diameter, 1 mm thick) were fabricated for
the swelling and mass loss studies.[3,32,34,35] The initial wet mass of
each sample (minitial;t0) was measured, and three samples were
immediately lyophilized to obtain their dry weights (mdry;t0). This
enabled the calculation of the actual starting% macromer for all
samples,[3]
12, 12, 3
H & Co
% macromer ¼mdry;t0
minitial;t0
� 100 (3)
22–332
. KGaA, Weinheim www.MaterialsViews.com

The Influence of Silkworm Species on Cellular Interactions with . . .
www.mbs-journal.de
The initial dry mass of the remaining samples (mi,dry) was then
calculated using the actual % macromer (Equation 3) and each
disc’s individual starting wet weight (minitial).
www.M
mi;dry ¼ minitial � % macromer (4)
se samples were then submerged in a sink of phosphate-
Thebuffered saline solution (pH¼7.4), and incubated in an orbital
shaker incubator set at 37 8C. Samples were removed from the
incubator after 1 d, blotted dry and weighed (mswollen). The swollen
samples were then freeze-dried and weighed again (mdry). The mass
swelling ratio (q) and mass loss/sol fraction were calculated as
follows:[3,32]
q ¼ mswollen
mdry(5)
Mass loss% ¼mi;dry �mdry
mi;dry
� �� 100% (6)
The sol fraction of the hydrogels is equal to the mass loss at 1 d, as
it has been shown that any polymer chains not attached to the
network will diffuse out of the sample in this timeframe.[34,35]
2.3.3. Sericin Release Studies
PVA/sericin hydrogels were prepared (10 mm diameter, 1 mm
thick), immersed in 2 mL phosphate buffer saline (PBS) and
incubated at 37 8C. 0.5 mL of supernatant was withdrawn after
1 d of incubation and stored frozen. The released sericin was
quantified using the microbicinchoninic acid (microBCA) assay
(BCA kit for protein determination, Pierce). In brief, the supernatant
containing sericin was mixed with the BCA reagents, then
measured for absorbance at 562 nm. The mass of sericin in the
supernatant was determined by using sericin standard curves
constructed for the two different species, respectively. The sericin
mass loss percentage was calculated as follows:
Sericin mass loss% ¼ msericin in supernatant
msericin in hydrogel
� �� 100% (7)
where msericin in hydrogel is the amount of sericin originally
incorporated into the hydrogel.
2.3.4. L929 Cell Adhesion and Proliferation Studies on
PVA/Sericin Hydrogels
Hydrogels (7.5 mm diameter, 1 mm thick) were prepared under
sterile conditions, and then soaked in media to extract the sol
fraction in the network. After 1 d, the media was discarded and
hollow metal fences were placed on top of the hydrogels to prevent
the hydrogels from floating and to ensure the cells were seeded only
on the hydrogel surface. Cells were seeded onto the hydrogels
through the metal fences (12.57 mm2) at a density of
80 cells �mm�2. Media was added through the side path of the
metal fences to surround the hydrogels. All samples were prepared
in triplicate. Live-dead assay was done to determine the amount of
the adhered cells after 4 h, 1 d and 3 d of incubation. In brief, the gels
were washed with Dulbecco’s phosphate buffered saline (DPBS)
then stained with propidium iodide (PI) and Calcein-AM (Cal-AM)
aterialsViews.com
Macromol. Biosci. 20
� 2012 WILEY-VCH Verlag Gmb
both at 1mg �mL�1. After 10 min incubation with the stains, the gels
were again washed with DPBS then images were captured using
the fluorescent microscope. ImageJ was used to count the attached
live cells. In a separate experiment, cells were seeded onto TCP
through the metal fences, and visualization suggested that
adhesion and morphology of cells was similar to those on PVA/
sericin hydrogels.
2.4. Statistical Analysis
All results were analyzed using an ANOVA model with replication
and combination of fixed and random variables. The models were
constructed using Minitab statistical software (Minitab Inc.,
version 15). Samples in a study (n¼1) were all prepared in
triplicates, and all studies were repeated 3 times (n¼ 3). A p<0.05
was considered as statistically significant.
3. Results and Discussion
3.1. Macromer Synthesis
3.1.1. Isolation of Sericin from Silkworm Cocoons
The molecular weights of the isolated silk sericin peptides
from both species were measured by 8% SDS-PAGE analysis,
and were observed to range from 20 to 200 kDa (results not
shown), and is consistent with the literature.[12,14,16,19,36]
Current processes used to extract sericin from silkworm
cocoons involve a range of treatments including heat
(121 8C), acid (citric acid), alkali (Na2CO3) and urea. Recently,
it was shown that these extraction processes induce sericin
degradation or hydroxylation, which can further influence
the molecular weight of the sericin products.[21,37] How-
ever, numerous studies showed that these degraded sericin
peptides retain their beneficial biological properties, such
as moisture absorption, antioxidation, tyrosinase activity
inhibition, anticancer activity and protecting cells from
death caused by serum deprivation.[19,38] It was also
reported that alkali degraded B. mori sericin are suitable
for cell culture and promoted collagen production from
fibroblasts.[20] Hence, it has been well established that the
degraded sericin remained bioactive, and it is expected that
incorporating these degraded sericin peptides into PVA
hydrogels will improve the gel’s bioactivity.
3.2. Macromer Characterization
3.2.1. PVA and Sericin Methacrylation
PVA and both species of sericin peptides were all
successfully functionalized with methacrylate groups as
shown by 1H NMR (Figure 1). The methacrylate groups will
allow the polymers to be photopolymerized into cross-
linked hydrogels. For PVA-MA, ICEMA reacted with the
12, 12, 322–332
H & Co. KGaA, Weinheim325
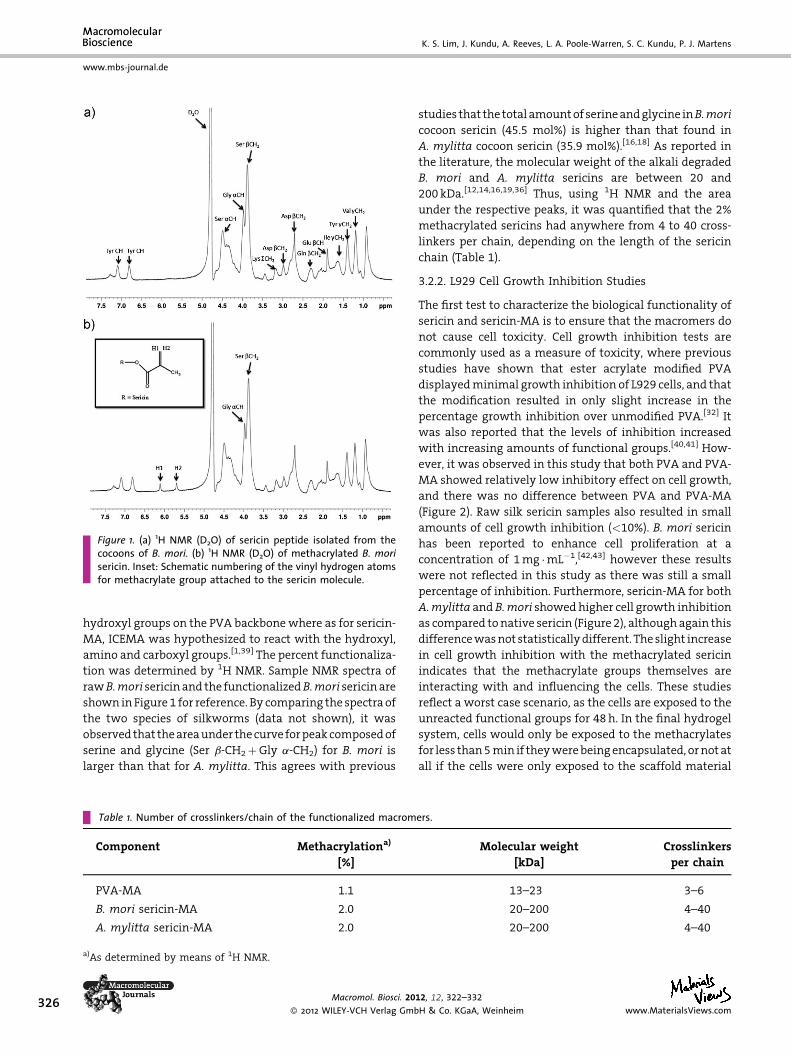
Figure 1. (a) 1H NMR (D2O) of sericin peptide isolated from thecocoons of B. mori. (b) 1H NMR (D2O) of methacrylated B. morisericin. Inset: Schematic numbering of the vinyl hydrogen atomsfor methacrylate group attached to the sericin molecule.
326
www.mbs-journal.de
K. S. Lim, J. Kundu, A. Reeves, L. A. Poole-Warren, S. C. Kundu, P. J. Martens
hydroxyl groups on the PVA backbone where as for sericin-
MA, ICEMA was hypothesized to react with the hydroxyl,
amino and carboxyl groups.[1,39] The percent functionaliza-
tion was determined by 1H NMR. Sample NMR spectra of
rawB.mori sericin and the functionalizedB.mori sericin are
shown in Figure 1 for reference. By comparing the spectra of
the two species of silkworms (data not shown), it was
observed that the area under the curve for peak composed of
serine and glycine (Ser b-CH2þGly a-CH2) for B. mori is
larger than that for A. mylitta. This agrees with previous
Table 1. Number of crosslinkers/chain of the functionalized macrom
Component Methacrylationa)
[%]
PVA-MA 1.1
B. mori sericin-MA 2.0
A. mylitta sericin-MA 2.0
a)As determined by means of 1H NMR.
Macromol. Biosci. 20
� 2012 WILEY-VCH Verlag Gmb
studies that the total amount of serine and glycine inB.mori
cocoon sericin (45.5 mol%) is higher than that found in
A. mylitta cocoon sericin (35.9 mol%).[16,18] As reported in
the literature, the molecular weight of the alkali degraded
B. mori and A. mylitta sericins are between 20 and
200 kDa.[12,14,16,19,36] Thus, using 1H NMR and the area
under the respective peaks, it was quantified that the 2%
methacrylated sericins had anywhere from 4 to 40 cross-
linkers per chain, depending on the length of the sericin
chain (Table 1).
3.2.2. L929 Cell Growth Inhibition Studies
The first test to characterize the biological functionality of
sericin and sericin-MA is to ensure that the macromers do
not cause cell toxicity. Cell growth inhibition tests are
commonly used as a measure of toxicity, where previous
studies have shown that ester acrylate modified PVA
displayed minimal growth inhibition of L929 cells, and that
the modification resulted in only slight increase in the
percentage growth inhibition over unmodified PVA.[32] It
was also reported that the levels of inhibition increased
with increasing amounts of functional groups.[40,41] How-
ever, it was observed in this study that both PVA and PVA-
MA showed relatively low inhibitory effect on cell growth,
and there was no difference between PVA and PVA-MA
(Figure 2). Raw silk sericin samples also resulted in small
amounts of cell growth inhibition (<10%). B. mori sericin
has been reported to enhance cell proliferation at a
concentration of 1 mg �mL�1,[42,43] however these results
were not reflected in this study as there was still a small
percentage of inhibition. Furthermore, sericin-MA for both
A.mylitta and B.mori showed higher cell growth inhibition
as compared to native sericin (Figure 2), although again this
difference was not statistically different. The slight increase
in cell growth inhibition with the methacrylated sericin
indicates that the methacrylate groups themselves are
interacting with and influencing the cells. These studies
reflect a worst case scenario, as the cells are exposed to the
unreacted functional groups for 48 h. In the final hydrogel
system, cells would only be exposed to the methacrylates
for less than 5 min if they were being encapsulated, or not at
all if the cells were only exposed to the scaffold material
ers.
Molecular weight
[kDa]
Crosslinkers
per chain
13–23 3–6
20–200 4–40
20–200 4–40
12, 12, 322–332
H & Co. KGaA, Weinheim www.MaterialsViews.com
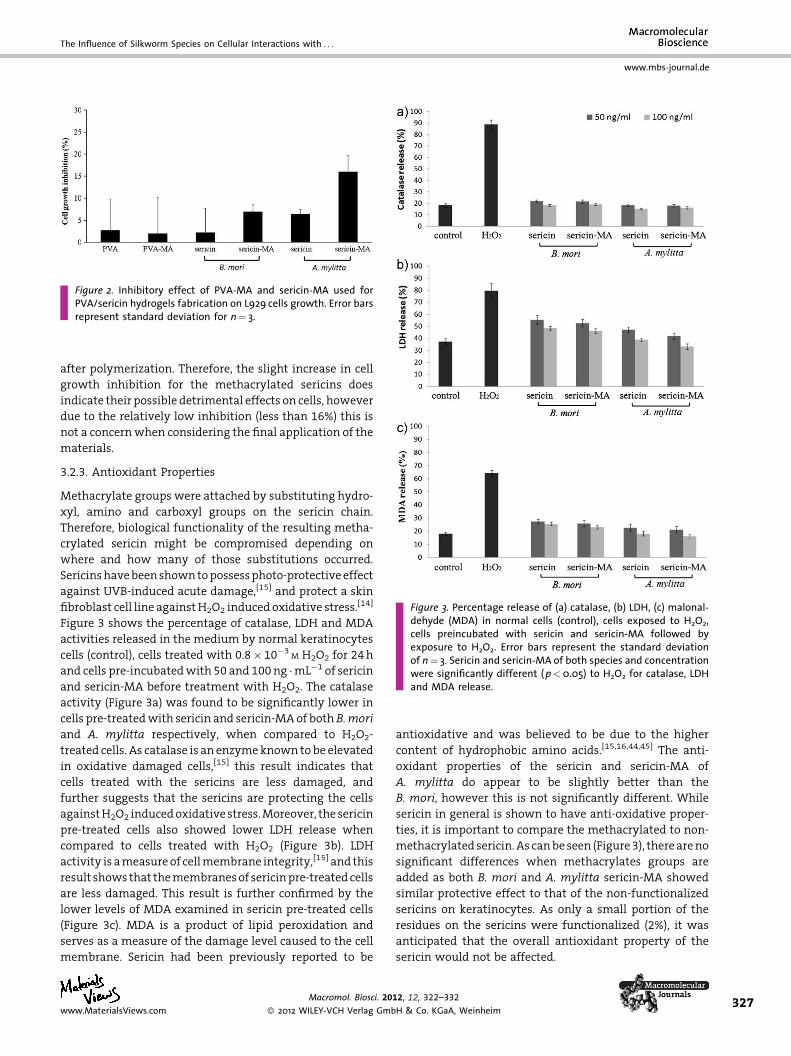
Figure 3. Percentage release of (a) catalase, (b) LDH, (c) malonal-dehyde (MDA) in normal cells (control), cells exposed to H2O2,cells preincubated with sericin and sericin-MA followed byexposure to H2O2. Error bars represent the standard deviationof n¼ 3. Sericin and sericin-MA of both species and concentrationwere significantly different (p<0.05) to H2O2 for catalase, LDHand MDA release.
Figure 2. Inhibitory effect of PVA-MA and sericin-MA used forPVA/sericin hydrogels fabrication on L929 cells growth. Error barsrepresent standard deviation for n¼ 3.
The Influence of Silkworm Species on Cellular Interactions with . . .
www.mbs-journal.de
after polymerization. Therefore, the slight increase in cell
growth inhibition for the methacrylated sericins does
indicate their possible detrimental effects on cells, however
due to the relatively low inhibition (less than 16%) this is
not a concern when considering the final application of the
materials.
3.2.3. Antioxidant Properties
Methacrylate groups were attached by substituting hydro-
xyl, amino and carboxyl groups on the sericin chain.
Therefore, biological functionality of the resulting metha-
crylated sericin might be compromised depending on
where and how many of those substitutions occurred.
Sericins have been shown to possess photo-protective effect
against UVB-induced acute damage,[15] and protect a skin
fibroblast cell line against H2O2 induced oxidative stress.[14]
Figure 3 shows the percentage of catalase, LDH and MDA
activities released in the medium by normal keratinocytes
cells (control), cells treated with 0.8� 10�3M H2O2 for 24 h
and cells pre-incubated with 50 and 100 ng �mL�1 of sericin
and sericin-MA before treatment with H2O2. The catalase
activity (Figure 3a) was found to be significantly lower in
cells pre-treated with sericin and sericin-MA of both B.mori
and A. mylitta respectively, when compared to H2O2-
treated cells. As catalase is an enzyme known to be elevated
in oxidative damaged cells,[15] this result indicates that
cells treated with the sericins are less damaged, and
further suggests that the sericins are protecting the cells
against H2O2 induced oxidative stress. Moreover, the sericin
pre-treated cells also showed lower LDH release when
compared to cells treated with H2O2 (Figure 3b). LDH
activity is a measure of cell membrane integrity,[15] and this
result shows that the membranes of sericin pre-treated cells
are less damaged. This result is further confirmed by the
lower levels of MDA examined in sericin pre-treated cells
(Figure 3c). MDA is a product of lipid peroxidation and
serves as a measure of the damage level caused to the cell
membrane. Sericin had been previously reported to be
www.MaterialsViews.com
Macromol. Biosci. 20
� 2012 WILEY-VCH Verlag Gmb
antioxidative and was believed to be due to the higher
content of hydrophobic amino acids.[15,16,44,45] The anti-
oxidant properties of the sericin and sericin-MA of
A. mylitta do appear to be slightly better than the
B. mori, however this is not significantly different. While
sericin in general is shown to have anti-oxidative proper-
ties, it is important to compare the methacrylated to non-
methacrylated sericin. As can be seen (Figure 3), there are no
significant differences when methacrylates groups are
added as both B. mori and A. mylitta sericin-MA showed
similar protective effect to that of the non-functionalized
sericins on keratinocytes. As only a small portion of the
residues on the sericins were functionalized (2%), it was
anticipated that the overall antioxidant property of the
sericin would not be affected.
12, 12, 322–332
H & Co. KGaA, Weinheim327
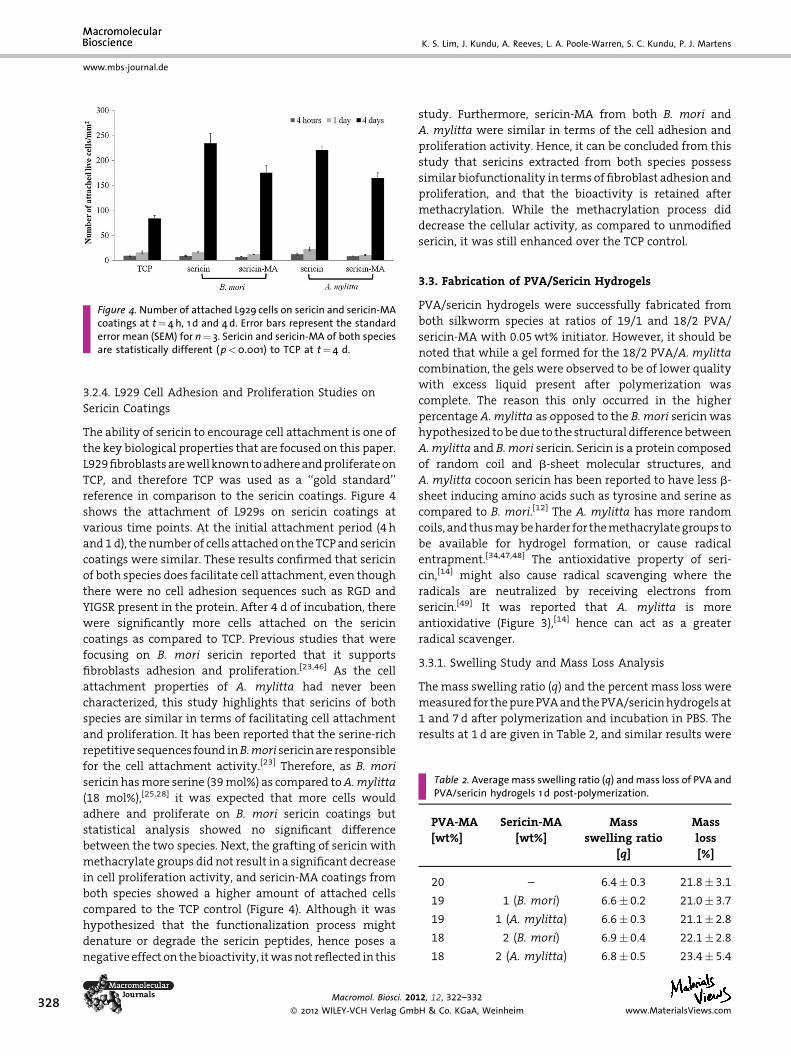
Figure 4. Number of attached L929 cells on sericin and sericin-MAcoatings at t¼4 h, 1 d and 4d. Error bars represent the standarderror mean (SEM) for n¼ 3. Sericin and sericin-MA of both speciesare statistically different (p<0.001) to TCP at t¼4 d.
Table 2. Averagemass swelling ratio (q) andmass loss of PVA andPVA/sericin hydrogels 1 d post-polymerization.
PVA-MA
[wt%]
Sericin-MA
[wt%]
Mass
swelling ratio
[q]
Mass
loss
[%]
20 – 6.4� 0.3 21.8� 3.1
19 1 (B. mori) 6.6� 0.2 21.0� 3.7
19 1 (A. mylitta) 6.6� 0.3 21.1� 2.8
18 2 (B. mori) 6.9� 0.4 22.1� 2.8
18 2 (A. mylitta) 6.8� 0.5 23.4� 5.4
328
www.mbs-journal.de
K. S. Lim, J. Kundu, A. Reeves, L. A. Poole-Warren, S. C. Kundu, P. J. Martens
3.2.4. L929 Cell Adhesion and Proliferation Studies on
Sericin Coatings
The ability of sericin to encourage cell attachment is one of
the key biological properties that are focused on this paper.
L929 fibroblasts are well known to adhere and proliferate on
TCP, and therefore TCP was used as a ‘‘gold standard’’
reference in comparison to the sericin coatings. Figure 4
shows the attachment of L929s on sericin coatings at
various time points. At the initial attachment period (4 h
and 1 d), the number of cells attached on the TCP and sericin
coatings were similar. These results confirmed that sericin
of both species does facilitate cell attachment, even though
there were no cell adhesion sequences such as RGD and
YIGSR present in the protein. After 4 d of incubation, there
were significantly more cells attached on the sericin
coatings as compared to TCP. Previous studies that were
focusing on B. mori sericin reported that it supports
fibroblasts adhesion and proliferation.[23,46] As the cell
attachment properties of A. mylitta had never been
characterized, this study highlights that sericins of both
species are similar in terms of facilitating cell attachment
and proliferation. It has been reported that the serine-rich
repetitive sequences found inB.mori sericin are responsible
for the cell attachment activity.[23] Therefore, as B. mori
sericin has more serine (39 mol%) as compared toA.mylitta
(18 mol%),[25,28] it was expected that more cells would
adhere and proliferate on B. mori sericin coatings but
statistical analysis showed no significant difference
between the two species. Next, the grafting of sericin with
methacrylate groups did not result in a significant decrease
in cell proliferation activity, and sericin-MA coatings from
both species showed a higher amount of attached cells
compared to the TCP control (Figure 4). Although it was
hypothesized that the functionalization process might
denature or degrade the sericin peptides, hence poses a
negative effect on the bioactivity, it was not reflected in this
Macromol. Biosci. 20
� 2012 WILEY-VCH Verlag Gmb
study. Furthermore, sericin-MA from both B. mori and
A. mylitta were similar in terms of the cell adhesion and
proliferation activity. Hence, it can be concluded from this
study that sericins extracted from both species possess
similar biofunctionality in terms of fibroblast adhesion and
proliferation, and that the bioactivity is retained after
methacrylation. While the methacrylation process did
decrease the cellular activity, as compared to unmodified
sericin, it was still enhanced over the TCP control.
3.3. Fabrication of PVA/Sericin Hydrogels
PVA/sericin hydrogels were successfully fabricated from
both silkworm species at ratios of 19/1 and 18/2 PVA/
sericin-MA with 0.05 wt% initiator. However, it should be
noted that while a gel formed for the 18/2 PVA/A. mylitta
combination, the gels were observed to be of lower quality
with excess liquid present after polymerization was
complete. The reason this only occurred in the higher
percentage A. mylitta as opposed to the B. mori sericin was
hypothesized to be due to the structural difference between
A. mylitta and B. mori sericin. Sericin is a protein composed
of random coil and b-sheet molecular structures, and
A. mylitta cocoon sericin has been reported to have less b-
sheet inducing amino acids such as tyrosine and serine as
compared to B. mori.[12] The A. mylitta has more random
coils, and thus may be harder for the methacrylate groups to
be available for hydrogel formation, or cause radical
entrapment.[34,47,48] The antioxidative property of seri-
cin,[14] might also cause radical scavenging where the
radicals are neutralized by receiving electrons from
sericin.[49] It was reported that A. mylitta is more
antioxidative (Figure 3),[14] hence can act as a greater
radical scavenger.
3.3.1. Swelling Study and Mass Loss Analysis
The mass swelling ratio (q) and the percent mass loss were
measured for the pure PVA and the PVA/sericin hydrogels at
1 and 7 d after polymerization and incubation in PBS. The
results at 1 d are given in Table 2, and similar results were
12, 12, 322–332
H & Co. KGaA, Weinheim www.MaterialsViews.com
The Influence of Silkworm Species on Cellular Interactions with . . .
www.mbs-journal.de
obtained at 7 d (data not shown). It has been previously
shown that any polymer chains not incorporated in the PVA
gels will be released with the first 24 h,[34,35] and no further
changes in swelling or mass loss occur, and these results
demonstrated similar findings. Statistical analysis showed
that there was no significant difference between the mass
swelling ratio (q) of the pure PVA gels and either of the 19/1
PVA/sericin gels. However, the values of q for 18/2 PVA/
sericin gels from both species were statistically higher than
that of the pure PVA gels. Previous studies have reported
that incorporation of ionic biological polymers such as
heparin and chondroitin sulfate into PVA hydrogels has
increased the swelling capacity,[1,3,50] and this study also
demonstrates a trend towards increased swelling with the
incorporation of sericin. Sericin proteins have been reported
to be negatively charged at neutral pH,[26,50] and therefore it
would be expected that the more negatively charged chains
that are incorporated the more the swelling will increase.
All the mass loss values obtained (Table 2) were
comparable to each other, and to previous studies done
on photopolymerized methacrylate hydrogels.[3,30,34,35]
Statistical analysis showed that there were no significant
differences in the mass loss for PVA and PVA/sericin
hydrogels, which indicates that the sericin peptides did not
disrupt the polymerization process. Although the photo-
polymerization process with this PVA macromer is not
100% efficient, it is known that the polymerization kinetics
can be controlled through a variety of techniques. Previous
studies done in our lab had shown that using other
functional groups that are more reactive (ester acrylates) or
increasing the amount of methacrylates grafted on the PVA
backbone can increase the polymerization efficiency.[35]
3.3.2. Sericin Release Studies
Another indication of the quality of the gel formation is the
measurement of the amount of sericin that is lost from
Figure 5. Sericin release (%) from fabricated PVA/sericin hydrogelsafter 1 d of incubation. Three gels were prepared for each exper-iment (n¼ 1) and a total of three repeats of each experiment wereperformed (n¼ 3). As sericin release varied throughout the threeexperiments, all nine data points were shown.
www.MaterialsViews.com
Macromol. Biosci. 20
� 2012 WILEY-VCH Verlag Gmb
the gel. As shown in Figure 5, there is a large amount of
sericin that was lost from all of the gels in the first 24 h,
although in all cases a significant portion of the sericin still
remained within the gel. This is consistent with previous
findings that all unincorporated molecules will diffuse out
of the hydrogel during this time frame.[34,35] The amount of
sericin released from the gels ranged between 20 and 60% of
the original amount of sericin incorporated. The release and
the quantification of the amount of sericin was variable
throughout the three experiments, therefore all of the data
is shown in Figure 5 for completeness. The 18/2 PVA/
A. mylitta gels had the greatest variability in the results,
although were within the same range as the other gels. It
was hypothesized that increasing the concentration of
sericin may perturb the crosslinking process and thus result
in an increase in ‘‘unbound’’ sericin able to be released. The
mechanism underlying this perturbation is likely to relate
to the antioxidative radical scavenging properties of sericin.
This is supported by the observation that the higher
variability of release in A. mylitta gels correlated with the
A. mylitta sericin being more anti-oxidative. The two
silkworm species are biochemically different, and therefore
incomparable biological behaviors can be expected. This
increase in variability, along with the visual differences in
these gels, supports the hypothesis that the sericin is acting
in some way to decrease the effectiveness of the
polymerization, whether through radical scavenging or
another method. Increasing the percent methacrylation of
sericin would support formation of more highly crosslinked
networks of PVA and sericin, which could reduce the
variation in sericin release. The disadvantages associated
Figure 6. Number of attached L929 cells on PVA and PVA/sericinhydrogels at t¼4 h, 1 d, and 3 d. Error bars represent the standarderrormean (SEM) for n¼ 3. The numbers of cells attached on PVA/sericin hydrogels are significantly different to 20% PVA hydrogels(p<0.001). PVA/sericin hydrogels of A. mylitta has significantlyhigher cell attachment compared to B. mori (p¼0.001). In termsof sericin concentration, cell attachment on 18/2 PVA/sericinhydrogels is significantly higher to 19/1 PVA/sericin for B. mori(p<0.05) but not for A. mylitta (p>0.05). For noting, in aseparate experiment, cells were grown on TCP by the samemethods and the number of attached L929 cells were quantifiedto be 68� 3 (t¼ 4h), 98� 4 (t¼ 1 d) and 436� 20 (t¼ 3 d).
12, 12, 322–332
H & Co. KGaA, Weinheim329
330
www.mbs-journal.de
K. S. Lim, J. Kundu, A. Reeves, L. A. Poole-Warren, S. C. Kundu, P. J. Martens
with increasing the sericin functional group density relate
to the potential for disruption of biological function with
higher methacrylate substitution. This would need to be
studied in future research.
3.3.3. Cell Adhesion Studies
The cell adhesion study was used to evaluate the biological
functionality of the PVA/sericin hydrogels. The aim was to
determine if the sericin still had bio-functionality after
chemical modification and incorporation into the gels, and
also if the sericins were accessible to cells exposed to the
surface of the gels. It is known that cells do not adhere to
pure PVA hydrogels due to its highly hydrophilic nat-
ure.[51,52] Therefore, pure PVA hydrogels were used as a
control, and as expected, L929 cells did not adhere and
proliferate on them (Figure 6). Previous studies have
reported that by incorporating biological polymers such
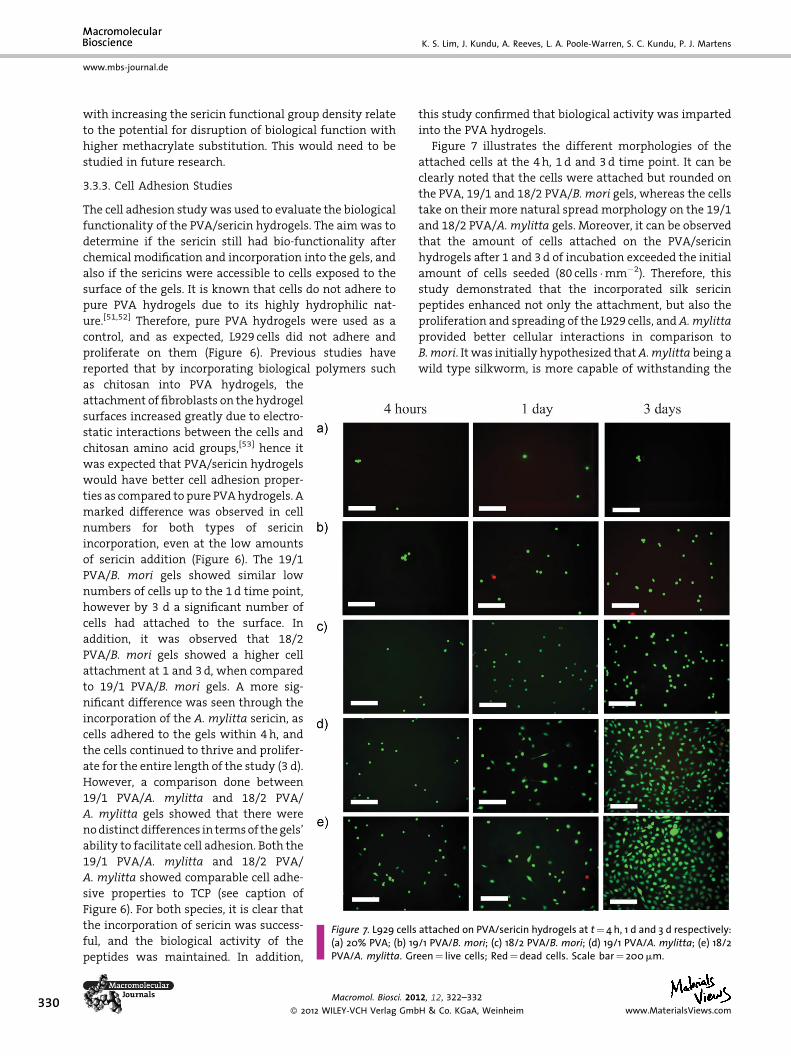
Figure 7. L929 cells attached on PVA/sericin hydrogels at t¼4 h, 1 d and 3 d respectively:(a) 20% PVA; (b) 19/1 PVA/B. mori; (c) 18/2 PVA/B. mori; (d) 19/1 PVA/A. mylitta; (e) 18/2PVA/A. mylitta. Green¼ live cells; Red¼dead cells. Scale bar¼ 200mm.
as chitosan into PVA hydrogels, the
attachment of fibroblasts on the hydrogel
surfaces increased greatly due to electro-
static interactions between the cells and
chitosan amino acid groups,[53] hence it
was expected that PVA/sericin hydrogels
would have better cell adhesion proper-
ties as compared to pure PVA hydrogels. A
marked difference was observed in cell
numbers for both types of sericin
incorporation, even at the low amounts
of sericin addition (Figure 6). The 19/1
PVA/B. mori gels showed similar low
numbers of cells up to the 1 d time point,
however by 3 d a significant number of
cells had attached to the surface. In
addition, it was observed that 18/2
PVA/B. mori gels showed a higher cell
attachment at 1 and 3 d, when compared
to 19/1 PVA/B. mori gels. A more sig-
nificant difference was seen through the
incorporation of the A. mylitta sericin, as
cells adhered to the gels within 4 h, and
the cells continued to thrive and prolifer-
ate for the entire length of the study (3 d).
However, a comparison done between
19/1 PVA/A. mylitta and 18/2 PVA/
A. mylitta gels showed that there were
no distinct differences in terms of the gels’
ability to facilitate cell adhesion. Both the
19/1 PVA/A. mylitta and 18/2 PVA/
A. mylitta showed comparable cell adhe-
sive properties to TCP (see caption of
Figure 6). For both species, it is clear that
the incorporation of sericin was success-
ful, and the biological activity of the
peptides was maintained. In addition,
Macromol. Biosci. 20
� 2012 WILEY-VCH Verlag Gmb
this study confirmed that biological activity was imparted
into the PVA hydrogels.
Figure 7 illustrates the different morphologies of the
attached cells at the 4 h, 1 d and 3 d time point. It can be
clearly noted that the cells were attached but rounded on
the PVA, 19/1 and 18/2 PVA/B. mori gels, whereas the cells
take on their more natural spread morphology on the 19/1
and 18/2 PVA/A. mylitta gels. Moreover, it can be observed
that the amount of cells attached on the PVA/sericin
hydrogels after 1 and 3 d of incubation exceeded the initial
amount of cells seeded (80 cells �mm�2). Therefore, this
study demonstrated that the incorporated silk sericin
peptides enhanced not only the attachment, but also the
proliferation and spreading of the L929 cells, and A. mylitta
provided better cellular interactions in comparison to
B.mori. It was initially hypothesized that A.mylitta being a
wild type silkworm, is more capable of withstanding the
12, 12, 322–332
H & Co. KGaA, Weinheim www.MaterialsViews.com

The Influence of Silkworm Species on Cellular Interactions with . . .
www.mbs-journal.de
harsh conditions of the extraction and functionalization
processes used to synthesize sericin-MA.[14,15] In this case,
theA.mylitta sericin would therefore be less denatured and
degraded than the B. mori sericin, and retained more of its
structure, biological and chemical characteristics of the
natural sericin. However, the cell adhesion and prolifera-
tion study done on sericin coatings showed no difference
between both species, before and after methacrylation
(Figure 4), this finding implies that the resistive nature ofA.
mylitta may not be the answer to the vast difference
observed. As such, it may be due to the way sericin is bound
in the hydrogel network and presented to cells. The
different amino acid composition and structural conforma-
tion of both sericins may affect the radicals pathway during
the crosslinking process, and cause dissimilarity in sericin
distribution in the hydrogels. The interaction between
sericin-MA and PVA-MA during the hydrogel fabrication is
yet to be explored and is beyond the scope of this paper. As
the exact nature of the biofunctionality of the sericin
proteins or peptides is not known, the precise reason for the
differences between the two types of sericins is also difficult
to determine. While none of the known cell adhesion
sequences are found in the sericin (e.g., RGD, YIGSR), they
remain to be fully sequenced. It is hypothesized that
the sericin may contain other, lesser known adhesion
sequences such as the serine-rich repetitive sequence found
in B. mori,[23] or that the sericins are inducing attached
fibroblasts to secrete type 1 collagen that is known to
enhance cell proliferation. The characterization of the
structure, chemistry and denaturation of the sericin
proteins or peptides, is outside the scope of this paper;
however, it is clear that sericin retained its bioactivity, i.e.
antioxidative and cell adhesion properties, after modifica-
tion, and these attributes were successfully transmitted
into PVA hydrogels. Moreover, there are very distinct
differences between the two species and the exact nature of
those differences needs to be explored.
4. Conclusion
In conclusion, isolated cocoon sericin peptides from B. mori
and A. mylitta were chemically modified and successfully
copolymerized with PVA to form PVA/sericin hydrogels.
Both the modified and unmodified sericins were character-
ized to be similar in terms of bio-functionality. The
incorporation of sericin did not significantly affect the
quality of the gels as compared to the pure PVA gels. In
addition, while not all incorporated sericin-MA molecules
were crosslinked into the hydrogel network, a significant
portion of the peptides remained after polymerization and
incubation. There were distinct differences between the
properties of sericin obtained from the two silkworm
species. These results demonstrated that the A. mylitta
www.MaterialsViews.com
Macromol. Biosci. 20
� 2012 WILEY-VCH Verlag Gmb
sericin appeared to have a higher radical scavenging
capability, and was also more biologically active and
available after incorporation into the hydrogels as com-
pared to B. mori sericin. This work demonstrates that
PVA/sericin hydrogels have the potential to be used in
biomedical device applications, especially as tissue engi-
neering scaffolds, and that the addition of sericin might be
an easy method of changing the bio-functionality of
pure synthetic hydrogels without interfering with the
base network structure. This novel PVA/sericin hydrogel
may be applied in encapsulation systems for various cell
types where cell survival and differentiated cell function
combined with immunoprotection of encapsulated cells
are required. Specifically, such encapsulation systems may
be used for insulin producing islet cells for treatment of
diabetes, where the incorporated sericin provides antiox-
idative capacity and UV protection to cells during the photo-
encapsulation process. Furthermore, the presence of sericin
in the gel may also protect encapsulated cells in vivo by
scavenging harmful reactive oxygen species.
Acknowledgements: The authors kindly thank Ms. Cathy Liu forher work on the mass loss and swelling studies, and Dr. Ross Odellfor his help with statistical analysis. This work was supported bythe Australian Government under the Australia-India StrategicResearch Fund (BF010049) and Indo Australia Biotechnology Fund,DBT (BT/PR9552/ICD/16/755/2006), Government of India, NewDelhi, and is a joint effort from the laboratories of Prof. S. C. Kunduat the Indian Institute of Technology, Kharagpur and Prof. Poole-Warren and Dr. Martens at the University of New South Wales.
Received: July 22, 2011; Revised: September 28, 2011; Publishedonline: January 16, 2012; DOI: 10.1002/mabi.201100292
Keywords: biomaterials; hydrogels; photopolymerization; poly-(vinyl alcohol); silk sericin
[1] S. J. Bryant, K. A. Davis-Arehart, N. Luo, R. K. Shoemaker, J. A.Arthur, K. S. Anseth, Macromolecules 2004, 37, 6726.
[2] B. B. Mandal, B. Ghosh, S. C. Kundu, Int. J. Biol. Macromol.2011, 49, 125.
[3] A. Nilasaroya, L. A. Poole-Warren, J. M. Whitelock,P. Jo Martens, Biomaterials 2008, 29, 4658.
[4] E. Farrell, F. J. O’Brien, P. Doyle, J. Fischer, I. Yannas, B. A.Harley, B. O’Connell, P. J. Prendergast, V. A. Campbell, TissueEng. 2006, 12, 459.
[5] H. J. Lee, J. S. Lee, T. Chansakul, C. Yu, J. H. Elisseeff, S. M. Yu,Biomaterials 2006, 27, 5268.
[6] J. W. Nichol, S. T. Koshy, H. Bae, C. M. Hwang, S. Yamanlar,A. Khademhosseini, Biomaterials 2010, 31, 5536.
[7] J. Zimmermann, K. Bittner, B. Stark, R. Mulhaupt, Biomaterials2002, 23, 2127.
[8] S. P. Massia, S. S. Rao, J. A. Hubbell, J. Biol. Chem. 1993, 268,8053.
12, 12, 322–332
H & Co. KGaA, Weinheim331

332
www.mbs-journal.de
K. S. Lim, J. Kundu, A. Reeves, L. A. Poole-Warren, S. C. Kundu, P. J. Martens
[9] S. Hou, Q. Xu, W. Tian, F. Cui, Q. Cai, J. Ma, I.-S. Lee, J. Neurosci.Methods 2005, 148, 60.
[10] H. Teramoto, K. Nakajima, C. Takabayashi, Biosci. Biotechnol.Biochem. 2005, 69, 845.
[11] A. Motta, B. Barbato, C. Foss, P. Torricelli, C. Migliaresi,J. Bioact. Compat. Polym. 2011, 26, 130.
[12] R. Dash, S. K. Ghosh, D. L. Kaplan, S. C. Kundu, Comp. Biochem.Physiol. Part B: Biochem. Mol. Biol. 2007, 147, 129.
[13] T. Khire, J. Kundu, S. C. Kundu, V. K. Yadavalli, Soft Matter2010, 6, 2066.
[14] R. Dash, C. Acharya, P. Bindu, S. Kundu, BMB Rep. 2008, 41,236.
[15] R. Dash, M. Mandal, S. Ghosh, S. Kundu, Mol. Cell Biochem.2008, 311, 111.
[16] R. Dash, S. Mukherjee, S. C. Kundu, Int. J. Biol. Macromol. 2006,38, 255.
[17] Y. Q. Zhang, Biotechnol. Adv. 2002, 20, 91.[18] M. Mondal, K. Trivedy, S. Nirmal Kumar, Caspian J. Env. Sci.
2007, 5, 63.[19] P. Aramwit, S. Damrongsakkul, S. Kanokpanont, T. Srichana,
Biotechnol. Appl. Biochem. 2010, 55, 91.[20] P. Aramwit, S. Kanokpanont, T. Nakpheng, T. Srichana, Int. J.
Mol. Sci. 2010, 11, 2200.[21] Y. Q. Zhang, M. L. Tao, W. D. Shen, Y. Z. Zhou, Y. Ding, Y. Ma,
W. L. Zhou, Biomaterials 2004, 25, 3751.[22] K. Tsujimoto, H. Takagi, M. Takahashi, H. Yamada,
S. Nakamori, J. Biochem. 2001, 129, 979.[23] K. Tsubouchi, Y. Igarashi, Y. Takasu, H. Yamada, Biosci. Bio-
technol. Biochem. 2005, 69, 403.[24] N. Nagai, T. Murao, Y. Ito, N. Okamoto, M. Sasaki, Biol. Pharm.
Bull. 2009, 32, 933.[25] S. C. Kundu, B. C. Dash, R. Dash, D. L. Kaplan, Prog. Polym. Sci.
2008, 33, 998.[26] H. Oh, J. Lee, A. Kim, C. Ki, J. Kim, Y. Park, K. Lee, Fibers Polym.
2007, 8, 470.[27] B. C. Dash, B. B. Mandal, S. C. Kundu, J. Biotechnol. 2009, 144,
321.[28] B. B. Mandal, A. S. Priya, S. C. Kundu, Acta Biomater. 2009, 5,
3007.[29] A. Muhlebach, B. Muller, C. Pharisa, M. Hofmann, B. Seiferling,
D. Guerry, J. Polym. Sci. Part A: Polym. Chem. 1997, 35, 3603.[30] F. Cavalieri, F. Miano, P. D’Antona, G. Paradossi, Biomacro-
molecules 2004, 5, 2439.
Macromol. Biosci. 20
� 2012 WILEY-VCH Verlag Gmb
[31] S. Sofia, M. B. McCarthy, G. Gronowicz, D. L. Kaplan, J. Biomed.Mater. Res. 2001, 54, 139.
[32] D. Mawad, P. J. Martens, R. A. Odell, L. A. Poole-Warren,Biomaterials 2007, 28, 947.
[33] Y. Tamada, M. Sano, K. Niwa, T. Imai, G. Yoshino, J. Biomater.Sci. Polym. Ed. 2004, 15, 971.
[34] P. Martens, K. S. Anseth, Polymer 2000, 41, 7715.[35] P. Martens, J. Blundo, A. Nilasaroya, R. A. Odell, J. Cooper-
White, L. A. Poole-Warren, Chem. Mater. 2007, 19, 2641.[36] Y. Takasu, H. Yamada, K. Tsubouchi, Biosci. Biotechnol.
Biochem. 2002, 66, 2715.[37] P. Vaithanomsat, V. Kitpreechavanich, Sep. Purif. Technol.
2008, 59, 129.[38] M. Takahashi, K. Tsujimoto, Y. Kato, H. Yamada, H. Takagi,
S. Nakamori, Biotechnol. Lett. 2005, 27, 893.[39] T. Furuzuno, K. Ishihara, N. Nakabayashi, Y. Tamada, Bioma-
terials 2000, 21, 327.[40] H. R. Mellor, J. Nolan, L. Pickering, M. R. Wormald, F. M. Platt,
R. A. Dwek, G. W. J. Fleet, T. D. Butters, Biochem. J. 2002, 366,225.
[41] E. Yoshii, J. Biomed. Mater. Res. 1997, 37, 517.[42] P. Aramwit, S. Kanokpanont, W. De-Eknamkul, T. Srichana,
J. Biosci. Bioeng. 2009, 107, 556.[43] S. Terada, T. Nishimura, M. Sasaki, H. Yamada, M. Miki,
Cytotechnology 2002, 40, 3.[44] N. Kato, S. Sato, A. Yamanaka, H. Yamada, N. Fuwa,
M. Nomura, Biosci. Biotechnol. Biochem. 1998, 62, 145.[45] J. B. Fan, L. P. Wu, L. S. Chen, X. Y. Mao, F. Z. Ren, J. Food.
Biochem. 2009, 33, 74.[46] N. Minoura, S. I. Aiba, Y. Gotoh, M. Tsukada, J. Biomed. Mater.
Res. 1995, 29, 1215.[47] H. Teramoto, A. Kakazu, K. Yamauchi, T. Asakura, Macromol-
ecules 2007, 40, 1562.[48] C. Vepari, D. L. Kaplan, Prog. Polym. Sci. 2007, 32, 991.[49] M. Valko, D. Leibfritz, J. Moncol, M. T. D. Cronin, M. Mazur,
J. Telser, Int. J. Biochem. Cell Biol. 2007, 39, 44.[50] G. Capar, S. S. Aygun, M. R. Gecit, J. Membr. Sci. 2009, 342, 179.[51] C. R. Nuttelman, D. J. Mortisen, S. M. Henry, K. S. Anseth,
J. Biomed. Mater. Res. 2001, 57, 217.[52] T. Hayami, K. Matsumura, M. Kusunoki, H. Nishikawa,
S. Hontsu, Mater. Lett. 2007, 61, 2667.[53] T. Koyano, N. Minoura, M. Nagura, K.-I. Kobayashi, J. Biomed.
Mater. Res. 1998, 39, 486.
12, 12, 322–332
H & Co. KGaA, Weinheim www.MaterialsViews.com





















![Structural and thermal studies of [PVA–LiAc] : TiO 2 polymer nanocomposite system](https://img.pdfslide.net/doc/110x75/634fcbbf48cb0bd7320f9f1e/structural-and-thermal-studies-of-pvaliac-tio-2-polymer-nanocomposite-system.jpg)







