Embed Size (px)
Citation preview

ANIMAL BEHAVIOUR, 2002, 64, 629–643doi:10.1006/anbe.2002.3082, available online at http://www.idealibrary.com on
The interplay between nutrient balancing and toxin dilution inforaging by a generalist insect herbivore
M. S. SINGER*, E. A. BERNAYS† & Y. CARRIEvRE‡
**Interdisciplinary Program in Insect Science, Center for Insect Science, University of Arizona†Center for Insect Science, University of Arizona
‡Department of Entomology, University of Arizona
(Received 18 April 2001; initial acceptance 31 July 2001;final acceptance 19 February 2002; MS. number: A9045R )
Food mixing by herbivores is thought to balance nutrient intake and possibly dilute secondarymetabolites characteristic of different host plant species. Most empirical work on insect herbivores hasfocused on nutrient balancing in laboratory settings. In this study, we characterize food mixingbehaviour of the caterpillar Grammia geneura (Strecker) (Lepidoptera: Arctiidae) in nature and use theobserved patterns to design ecologically relevant experiments that reveal the relative importance of theseprocesses in food-switching behaviour. Our design involved both choice and no-choice experiments withchemically defined diets in which primary nutrients and secondary metabolites were manipulated intandem. We analysed two stages in the process of food-switching behaviour: leaving food and acceptingnew food. In nature, an individual’s rate of leaving host plants was positively associated with itsprobability of rejecting plant species most recently eaten, but not related to its probability of acceptingdifferent host plant species. Furthermore, an individual’s leaving rate was negatively related to its averagefeeding bout duration. This relationship resulted partly from variation in the response of individuals tonutrient imbalance and partly from shortened feeding bouts prior to switching, suggesting that a declinein feeding excitation preceded searching for food that differed from that most recently eaten. Laboratoryexperiments with synthetic diets indicated the importance of secondary metabolites in the decline infeeding excitation prior to switching. Preference for new food depended strongly on secondarymetabolites in a manner consistent with toxin dilution. This is the first experimental evidence for theprocess of toxin dilution in caterpillars, and for the combined influence of nutrients and secondarymetabolites on their foraging patterns in nature.
2002 The Association for the Study of Animal Behaviour. Published by Elsevier Science Ltd. All rights reserved.
Correspondence and present address: M. S. Singer, Department ofEntomology, University of Arizona, Tucson, AZ 85721, U.S.A. (email:[email protected]). E. A. Bernays is at the Center for InsectScience, University of Arizona, Tucson, AZ 85721, U.S.A.
Optimal foraging theory argues that diet selection bygeneralist herbivores should be greatly influenced bylimitations in food availability and quality (Stephens &Krebs 1986). For generalist insect herbivores, smallresource demands relative to resource size generallyensures sufficient food availability for individuals. Hence,food quality is of utmost importance because it severelylimits individual performance, and presumably fitness.The food quality of individual plants or plant parts variesin acceptability or suitability due to physical differences(e.g. leaf toughness), and variation in the composition ofprimary nutrients (Slansky & Scriber 1985) and secondarymetabolic compounds (Bernays & Chapman 1994). For
0003–3472/02/$35.00/0 2002 The Association for the Study of An629
food mixing insects (i.e. those in which individuals eatmultiple plant species), host plant switching may func-tion to balance nutrient intake on the one hand, whileminimizing toxicity from specific secondary metabolitesin host plants on the other.
The nutrient balance hypothesis proposes that foodmixing allows individuals to balance intake of differentnutrients (Pulliam 1975; Westoby 1978; Rapport 1980;Waldbauer et al. 1984; Simpson et al. 1995). Physiologicaland behavioural experiments clearly demonstrate thatfood mixing can function to balance nutrient intake,thereby enhancing growth and development (Waldbauer& Friedman 1991; Simpson & Raubenheimer 1996).Among insects, food selection on the basis of nutrientbalancing has been demonstrated in grasshoppers(Simpson & Raubenheimer 1993), aphids (Abisgold et al.1994) and caterpillars (Waldbauer et al. 1984; Simmondset al. 1992).
imal Behaviour. Published by Elsevier Science Ltd. All rights reserved.

630 ANIMAL BEHAVIOUR, 64, 4
There is less experimental support for the toxin dilu-tion hypothesis: food mixing allows individuals to avoidingesting toxic doses of the particular secondary metabo-lites characteristic of specific foods (Freeland & Janzen1974; Freeland & Saladin 1989). Despite widespread evi-dence that secondary metabolites are often especiallytoxic to polyphagous insects (Bernays & Chapman 1994;Karban & Baldwin 1997), studies of herbivorous insectshave yet to claim any strong support for the toxindilution hypothesis. Indeed, it may be difficult to separateherbivore responses to secondary metabolites from thoseto other phytochemicals (including primary nutrients)because interactions among such chemicals directly influ-ence herbivore physiology (Slansky 1992; Hagele &Rowell-Rahier 1999; Simpson & Raubenheimer 2001).
None the less, there have been few attempts to testmechanistic predictions of these hypotheses with evi-dence from the detailed foraging behaviour of individualsobserved in nature and the laboratory. So far, these effortshave been confined to grasshoppers (Bernays et al. 1992,1994; Raubenheimer & Bernays 1993; Chambers et al.1996) with one exception (Dethier 1988). The relativelylarge amount of experimental work conducted exclu-sively in the laboratory has shown that both macro-nutrients and secondary metabolites alone and incombination influence the feeding behaviour of food-mixing insects (reviewed in Bernays & Chapman 1994,2000). However, because food-mixers may experience awide variety of combinations of nutrients and secondarymetabolites in natural host plants, field studies con-ducted in conjunction with simplified laboratory experi-ments are critical for understanding the relative roles ofnutrients and secondary metabolites in the mechanismand function of foraging in its natural context. Here, weattempt such a study with the food mixing caterpillarGrammia geneura (Strecker) (Lepidoptera: Arctiidae). First,we identify quantitative parameters of host plant switch-ing by caterpillars in nature. Second, we examinecomponents of switching behaviour by caterpillars intheir natural habitat and those offered synthetic diets inthe laboratory. Third, we use laboratory experiments withsynthetic diets to determine the influence of primarynutrients and secondary metabolites on the process ofswitching.
We designed analyses of observations and experimentsto address the following behavioural predictions of thenutrient balance and toxin dilution hypotheses. Thenutrient balance hypothesis predicts that food switchingshould involve leaving nutritionally unbalanced foodand accepting new food that is nutritionally complemen-tary to that recently eaten. By contrast, the toxin dilutionhypothesis predicts that switching should involve leavingfood after excessive exposure to toxic or deterrent effectsof particular secondary metabolites and accepting newfood lacking these secondary metabolites. We reasonedthat the avoidance of potential toxins by a food mixingherbivore would be maladaptive if switching occurred inresponse to intoxication (i.e. ‘passive response’ modelin Glendinning & Slansky 1995). Therefore, rather thanlook at toxicity per se, we hypothesized that food mixingherbivores would use secondary metabolites as signals of
particular food types (i.e. plant species in nature), andthat food switching would be initiated when such com-pounds deterred individuals from further feeding on aparticular food type (more akin to ‘active mechanisms’ inGlendinning & Slansky 1995). The increased acceptanceof novel food types would result from the absence ofsecondary metabolites recently experienced or possiblythe presence of different ones.
METHODSStudy System
Grammia geneura inhabits arid savanna and grassland ofthe southwestern U.S.A., where its larvae graze on at least80 species of (mostly herbaceous) flowering plant fromat least 50 taxonomically disparate families (Singer2000). Many of its host genera (e.g. Astragalus, Plantago,Euphorbia, various composites) are commonly avoided byother generalist herbivores (e.g. livestock) because ofdeterrence or toxicity. All insects in this study were (orwere recently descended from stock) collected in south-eastern Arizona, where the larval stages coincide withboth winter/spring and summer rainy periods. Individualcaterpillars move among small, mostly annual herbs inmixed-species patches (typically >10 spp.) in whichindividual plants are often contiguous or nearly so.Caterpillars of this species are most commonly found atground level and rarely climb more than approximately1 m from the ground (M. Singer, personal observation).Most feeding occurs diurnally, when larvae are mostactive (M. Singer, personal observation). A particularlyunusual feature of the life history is that adult femalesoviposit on the ground, rather than on host plants,forcing larvae to perform all host plant selection (M.Singer, personal observation). Short-term observationsand analysis of plant fragments in the faeces of lateinstars revealed that individual caterpillars eat multiplehost plant species (Singer 2000).
Pattern of Food Mixing
Field observationsTo quantify host plant switching by individual caterpil-
lars, we first observed a total of 43 caterpillars in nature atseveral different times and locations. Observations of5–180 min were made during both spring (near Oracle,Pinal Co., Arizona, U.S.A., April 1995) and summer (nearArivaca, Pima Co., Arizona, August 1993, 1995). Thesedata are included in only a single analysis here (seebelow).
We used relatively long-term observations (7 h) innature to measure two key components of food switch-ing: leaving host plants and accepting new plants. Wedefine switching as the following sequence of events: (1)feeding on one food item, (2) walking away from it, (3)contacting a second food item with mouthparts, and (4)eating the second food. Leaving refers to steps 1–2, whileaccepting new food involves steps 3–4. Food mixing refersto switches between different plant species. We observed11 final-instar larvae continuously for 7 h between 0900

631SINGER ET AL.: NUTRIENT BALANCE AND TOXIN DILUTION
and 1700 hours, when most larval activity occurs (AshCreek, Rincon Mtns, Pima Co., Arizona, 6 March–17 April1996). The data from one individual was not included inthe analyses because it rarely fed. All observation dayswere mostly or entirely clear and sunny. Air and groundtemperatures (in the shade) in foraging areas weremeasured at several times of day during observations.Each observer tracked a single, focal caterpillar through-out an observation session. Focal caterpillars were col-lected for rearing following the observation period tomake sure they were unparasitized, final-instar larvae.
For all observations, times at which various behavioursinitiated and terminated were recorded with digitalwatches. Behaviour was classified in the following way:‘feeding’ was defined as rhythmic movement of the headwhile it was in contact with the plant surface (ingestionof leaf tissue was usually visible); ‘walking’ was defined asany locomotion exclusive of feeding; ‘resting’ was definedas a lack of locomotion or feeding but included in-stantaneous movements like defecation and twitching;‘tasting’ was defined as a brief period (<1 s) of contactbetween a caterpillar’s mouthparts and plant tissue.Tasting followed by walking was scored as a ‘rejection’.Durations of feeding, resting and walking were measuredto the nearest second, while tastes and rejections wererecorded as instantaneous events. Feeding events werecomposed of discrete bouts. These could not be groupedas meals because there was no clear interbout criterion todefine meals (Simpson 1982), so bouts rather than mealswere used in all analyses. All plants contacted weredetermined to species when possible (>95%). We providea list of all host plant species eaten by focal caterpillars(Table 1).
Table 1. Plant species eaten by caterpillars observed in nature(1993–1996)
Plant species Family
Amaranthus sp. AmaranthaceaeAmbrosia confertiflora AsteraceaeBidens leptocephala AsteraceaeCirsium neomexicanum AsteraceaeErigeron sp. AsteraceaeMachaeranthera gracilis AsteraceaeMicroseris linearifolia AsteraceaeStephanomeria pauciflora AsteraceaeAmsinckia intermedia BoraginaceaePectocarya platycarpa BoraginaceaePlagiobothrys arizonicus BoraginaceaeSalsola tragus ChenopodiaceaeAcalypha neomexicana EuphorbiaceaeEuphorbia heterophylla EuphorbiaceaeEuphorbia prostrata EuphorbiaceaeAstragalus nothoxys FabaceaeLotus humistratus FabaceaeLupinus concinnus FabaceaeSenna leptocarpa FabaceaeErodium cicutarium GeraniaceaePhacelia distans HydrophyllaceaeRhynchosida physocalyx MalvaceaeSida abutiloides MalvaceaeSphaeralcea angustifolia MalvaceaeAllionia incarnata NyctaginaceaeOenothera primiveris OnagraceaePlantago patagonica PlantaginaceaeBromus rubens PoaceaeEriogonum polycladon PolygonaceaeEriogonum wrightii PolygonaceaeRumex sp. Polygonaceae
Analysis of host plant mixing
We used linear regression to analyse the relationshipbetween critical parameters of switching behaviour innature: leaving rate (explanatory variable) and the prob-ability of either rejecting a plant species most recentlyeaten [p(Rs)] or accepting a plant species that differedfrom that most recently eaten [p(Ad)] (response variables).We calculated the leaving rate of individuals (per h) as thenumber of times an insect left its host plant divided bythe total observation duration. To obtain values for theresponse variables, we imposed several criteria on theincluded data (7 h observations, N=10 insects). First, weonly considered contacts with plants during the walkingphase immediately following a feeding event of 60 s ormore to ensure that the plant previously eaten wasacceptable. Second, we excluded all contacts with grassspecies. These are generally far less acceptable than forbsto G. geneura (Singer & Stireman 2001). The probability ofrejecting plant species most recently eaten [p(Rs)] wascalculated for each individual as:
p(Rs)=Rs/Cs
where Rs is total number of rejections of plant speciesmost recently eaten, and Cs is the total number ofcontacts with plant species most recently eaten. The
probability of accepting different forb species [p(Ad)] wascalculated for each individual as:
p(Ad)=Fd/Cd
where Fd is the total number of feeding events on differ-ent forb species, and Cd is the total number of contactswith different forb species than that most recently eaten.
We analysed the association between leaving, switch-ing and mixing rates (number per h), and average feedingbout durations of individuals (mean of median boutdurations for each food) because we expected rejection offood (manifested by locomotion) to result from negativephysiological feedbacks from food (manifested by short-ened feeding bouts). We looked at this association amongindividual insects because our observations suggestedthat certain individuals left hosts frequently in associ-ation with brief feeding bouts, while others left hostsinfrequently and fed for relatively long periods at a time.For field insects observed for 7 h, we used a simple linearregression to analyse the relationship between leavingrate, switching rate and mixing rate (response variables inseparate analyses), and average feeding bout duration ofindividuals (explanatory variable in each analysis).
Upon finding that switching rates were most stronglynegatively associated with average feeding bout duration(see Results), we tested the prediction that feeding bouts

632 ANIMAL BEHAVIOUR, 64, 4
associated with switching were shorter than other feedingbouts. We considered feeding bouts immediately before(bout 1) and after leaving (bout 2) for each field insectthat moved between individual plants (N=30 insects). Weused one-way analysis of variance (ANOVA) models toanalyse feeding bout durations associated with foodswitching (bout 1 and bout 2 were response variables inseparate analyses), with the result of leaving (nonswitchesversus switches: either between different individualplants of the same species or between different plantspecies) as the explanatory variable. These analyses againused the average bout duration of individuals (mean ofthe median bout duration for each switch type; N=54averaged feeding bouts).
Process of Food Mixing: Experiments withSynthetic Food
DietsFor both the choice and no-choice tests, we prepared
synthetic diets of varied primary nutrient concentrations(digestible carbohydrate and protein) and secondarymetabolites (citral and coumarin) in an otherwise con-stant, chemically defined substrate (Table 2). We choseone suboptimal diet with a relatively low concentrationof protein relative to digestible carbohydrate, one dietnear the optimal balance of the nutrients according toprevious studies of other caterpillars in the superfamilyNoctuoidea (mean relative protein:carbohydrate (PC)ratio=57:43, Simpson & Raubenheimer 1993), andanother suboptimal diet with a relatively high concen-tration of protein relative to digestible carbohydrate.Specifically, the ratios were: 83:17 P:C, 60:40 P:C, and17:83 P:C. We estimated that P:C ratios of host plantfoliage span a similar range, based on previously docu-mented nutrient concentrations (Slansky & Scriber 1985).Hereafter, the first diet will be termed the protein-biaseddiet (P), the second will be the balanced diet (PC), and thethird will be the carbohydrate-biased diet (C). The sec-ondary metabolites were intended to represent signalcompounds (but not necessarily toxins) that the caterpil-lars would perceive as different, and might use to distin-guish among host plant species in nature. Coumarin
(cou) and citral (cit) are volatile and likely to stimulateolfactory receptors as well as gustatory receptors. Becauseolfactory inputs are usually more specific than gustatorysensation in insects (Chapman 1998), we thought thecaterpillars would be likely to discriminate between thesechemicals. The broad taxonomic range of plants used byG. geneura suggested that coumarin and citral would beecologically relevant. We conservatively used relativelylow concentrations (0.25% dry weight of diet) of thesechemicals to minimize their possible toxicity or deter-rence to inexperienced insects. Coumarin does stimulatea gustatory deterrent cell at 5 mM in G. geneura (Bernays& Chapman 2001); the effect of citral on the taste systemis unknown but appears not to be phagostimulatory fromour observations reported here. We offered the followingdiets to insects in the behavioural experiments describedbelow: Pcou, Ccou, PCcou, Pcit, Ccit, PCcit.
Experimental conditions
Caterpillars (different sets of full siblings in each experi-ment) were reared until the second or third day of thefinal stadium on a standard, wheat germ-based syntheticdiet (no added coumarin or citral) under controlled con-ditions of 28:25 �C, 16:8 h light:dark cycle, then givenchemically defined food for a conditioning period(defined below for each experiment) prior to obser-vational experiments. We used this conditioning periodto allow time for postingestive effects of the conditioningfoods to influence foraging. About 1 h prior to obser-vations, we replaced the conditioning food with fresh testfood. Cages were transparent, plastic boxes (11�11�4 cm) with two screened, ventilation holes (1.5-cmdiameter) on opposite sides. The food pieces provided(one or two) were approximately 2�2�2 cm in size.
During the observations in choice and no-choice tests(always 0900–1500 hours), we continuously recorded thecaterpillars’ activities using The Observer software pro-gram (Noldus 1991). Specifically, we recorded periods offeeding, walking and resting, as well as the instantaneousbehaviour of tasting, classified as in field observations.During each of the observation sessions, two observerseach monitored nine individual insects in a room illumi-nated by fluorescent lights, and held at 28 �C (�1 �C). Alltreatments were represented during each session, butwere haphazardly arranged among the two observers,who were blind to the treatment groups of individualinsects.
Table 2. Recipes for artificial diets used in choice and no-choiceexperiments
DietProtein source(% dry weight)
Carbohydrate source(% dry weight)
PC Casein (24.84) Sucrose (16.89)P Casein (34.77) Sucrose (6.95)C Casein (6.95) Sucrose (34.77)
All diets contained constant proportions of cellulose (53.60%),essential micronutrients, minerals and vitamins: salt mix (2.38%),linoleic acid (0.49%), cholesterol (0.49%), ascorbic acid (0.29%),vitamin mix (0.06%) and choline chloride (0.74%) in a 3.2%solution of agar and distilled water. Coumarin or citral (0.25%) wasalso added.
Choice experiment
To test our predictions of the nutrient balance andtoxin dilution hypotheses, we analysed the food-switching behaviour of final-instar larvae continuouslyobserved over a 6-h period. Caterpillars were caged indi-vidually with two pieces of food placed approximately4 cm apart (approximately the full length of a singlecaterpillar), so that switching would cause caterpillars tomove between them. Insects were confined with theirconditioning food for 12 h at 25 �C prior to observations.

633SINGER ET AL.: NUTRIENT BALANCE AND TOXIN DILUTION
In this experiment, the conditioning food choices andtest food choices for individual caterpillars were the same.We assumed that foraging patterns would be establishedduring the conditioning period and that observationswould thus reveal stable patterns.
The experimental design involved six treatmentgroups, each replicated 12 times (three in each obser-vation session). Treatments included (1) Ccou/Pcit, (2)PCcou/PCcit, (3) PCcou/PCcou, (4) PCcit/PCcit, (5) Pcou/Ccou and (6) Pcit/Ccit. Although these treatments do notrepresent the full range of diet combinations that arepossible, they are necessary and sufficient to test theinfluence of nutrients, secondary metabolites and theirinteraction on food switching. Additional combinationsof diet types (e.g. Ccit/Pcou) could not be tested simul-taneously with those above because too few insects wereavailable. The use of synthetic diets with both manipu-lated nutrients and secondary metabolites mimics theircombined presence in natural host plants, allowing theoccurrence of possible nutrient–secondary metaboliteinteractions. According to the nutrient balance hypoth-esis, insects should reject and leave food in the P versus Ctreatments more readily than in the PC versus PC treat-ments. In addition, switches to foods with complemen-tary nutrients (P to C, C to P) should result in increasedphagostimulation (measured as increased feeding boutduration) for the second food over the first. The toxindilution hypothesis predicts that rejecting and leavingshould follow excessive exposure to either cou or cit.Accordingly, switches to foods with different secondarymetabolites (cit to cou, cou to cit) should result inincreased feeding bout duration of the second foodrelative to the first.
No-choice experiment
To test the predictions of both hypotheses regardingthe acceptability of foods following a longer period ofexposure to foods that are either nutritionally deficient orpotentially toxic due to secondary metabolites, we specifi-cally addressed the readiness of insects to accept and eatnew foods. We used a C diet as the conditioning foodhere because its nutritive bias probably reflects that ofhost plant foliage in nature (i.e. nitrogen limited). Similarresponses to citral and coumarin foods in the choice test(see Results) suggested that it was reasonable to chooseCcit or Ccou as a conditioning food. We confined final-instar larvae (as above) with the Ccit diet only for 20 h at25 �C. About 1 h prior to observations, we replaced thepartially consumed Ccit diet with a new piece of Ccit dietto equalize food quality and availability. During theobservations, we monitored the first feeding bout on Ccit(the conditioning food), then during the quiescent periodfollowing this bout, we removed the Ccit food andreplaced it with the test food (Ccit again or a novel food).We carefully placed the new food approximately 1–3 cmin front of each caterpillar so that it would be likely toencounter the food soon after activity resumed. By thismethod we ensured that caterpillars encountered the newfood without disrupting their normal activity cycle.
Insects responded either by tasting the new food andwalking away (rejection) or by feeding (acceptance). Intwo separate analyses, we used the proportion of insectsthat accepted new food upon first contact, and thefeeding bout durations of those that accepted new food asthe respective response variables.
This experiment involved four treatment groups of newfoods: Ccit (i.e. same food=control), Ccou (different sec-ondary metabolite), Pcit (complementary nutrient bias),and Pcou (different secondary metabolite and comp-lementary nutrient bias). Each treatment was replicated17 times (except Ccou, N=19), and was represented dur-ing each observation session. As in the previous experi-ment, observers were blind to the treatment groups ofindividual insects. According to the nutrient balancehypothesis, Pcou and Pcit, the nutritionally complemen-tary foods to Ccit, should be more acceptable than thenutritionally similar foods, Ccou and Ccit. The toxindilution hypothesis, however, predicts that foods withnovel secondary metabolites, Ccou and Pcou, shouldbe more acceptable than their nutritionally identicalcounterparts, Ccit and Pcit.
Analysis of experimental results
As with insects feeding on plants, we compared therelationship between leaving rate and average feedingbout duration among insects in different treatmentsof the choice test. We used analysis of covariance(ANCOVA) to explain interindividual variation in leavingrate (response variable) with average feeding bout dur-ation of individuals as the covariate, and diet treatmentas the explanatory variable. To qualitatively contrast thenutrient balance and toxin dilution hypotheses, insectsoffered P versus C foods were analysed separately fromthose offered PC versus PC foods.
We investigated the possibility that feeding boutsassociated with switches were shorter than other feedingbouts for two reasons. First, field insects had shortenedfeeding bouts immediately prior to switching betweenindividual plants (see Results). In the choice experiment,we examined whether this reduction in feeding was aresponse to nutrients, secondary metabolites, or both.Second, we specifically hypothesized that P versus Ctreatments would show reduced feeding bouts in associ-ation with switches more than PC treatments becausethere was a negative association between the leaving rateand average feeding bout duration of individuals offeredP versus C foods in the choice test with synthetic foods(see Results). To determine the influence of primarynutrients and secondary metabolites on switching behav-iour, we considered feeding bouts immediately before(bout 1) and after leaving (bout 2) for each insect thatswitched between food pieces in relation to its feedingbouts that were not part of switches (bout 3). For theseanalyses, we also specified that bout 3 feeds preceded (forcomparison with bout 1) or followed (for comparisonwith bout 2) bouts of walking. We used two-way ANOVAmodels (bout 1 minus bout 3 and bout 2 minus bout3 were response variables in separate analyses), with
634 ANIMAL BEHAVIOUR, 64, 4
nutrient content, ‘N’ (same: PC versus PC; or different: Pversus C), secondary metabolites, ‘T’, (same or different),and their interaction as explanatory variables. Theseanalyses used the median feeding bout duration ofindividuals for each food type.
To determine whether insects were most stimulated bynew food that either was nutritionally complementary orcontained a different secondary metabolite, we comparedthe difference between feeding bouts 1 and 2 amongdifferent switch types in the choice test. Evidence foreither hypothesis would be an increase in the duration ofbout 2 relative to bout 1 when switches were betweenfoods with complementary nutritive bias or differentsecondary metabolites, respectively. The value of bout 2minus bout 1 was the response variable in a two-wayANOVA with N and T switch types (same as above) andtheir interaction as the factors. The same feeding boutwas never counted more than once in determining anindividual’s median value of bout 2 minus bout 1. Thusin certain cases (e.g. an individual switched in successioncausing bout 2 of the first switch to be bout 1 of thesecond switch), the second occurrence of a feeding boutwas excluded from the analysis.
To assess the acceptability of different food types in thechoice experiment, we compared across food types(PCcou, PCcit, Pcou, Pcit, Ccou, Ccit) the duration of onlythose feeding bouts not associated with switches. In aone-way ANOVA, the average feeding bout duration ofindividuals (mean of median bout duration for each foodtype) was the response variable.
To test the predictions of each hypothesis in theno-choice test, we compared both the outcome of the firstencounter with test food (feed or reject) and, for thoseindividuals that fed, the duration of the first feeding bouton each test food (Ccit, Ccou, Pcit, Pcou). We analysedthe outcome of the first encounter across all treatmentswith a G test and Williams’s correction (Sokal & Rohlf1995). We then used G tests on subsets of the data aspairwise comparisons to evaluate specific hypotheses(unplanned tests of the homogeneity of replicates testedfor goodness of fit, Sokal & Rohlf 1995). To analyse theduration of the first and second feeding bouts on testfoods (response variables in separate analyses), we used atwo-way ANOVA with the same switch categories as inthe choice test, ‘N’ and ‘T’ and their interaction, asfactors. We included only feeding bouts lasting longerthan 1 s in the analysis of bout 1 and bout 2.
When necessary, the variables were transformed in allanalyses (log[X+1] for rates and bout durations; arcsinesquare-root for proportions) to improve normality andhomogeneity of variance of the data (Zar 1984). Wereport an estimate of power for nonsignificant statisticaltests. Such estimates were obtained using an alpha levelof 0.05 and assuming the effect size and variance esti-mated from sampling data were close to the true param-eters (SAS Institute 2000). Under these assumptions, apower value reflects the likelihood of detecting an effectgiven the sample size that was used. All analyses wereperformed with the software package JMP (SAS Institute2000), except G tests, which followed Sokal & Rohlf(1995).
RESULTS
16000
30
Time of day (hours)
(b)
Rel
ativ
e %
0800 1000 1200 1400
5
10
15
20
25
Feeding boutsSwitches
160020
30(a)
Tem
per
atu
re (
°C)
0800 1000 1200 1400
22
24
26
28
AirGround
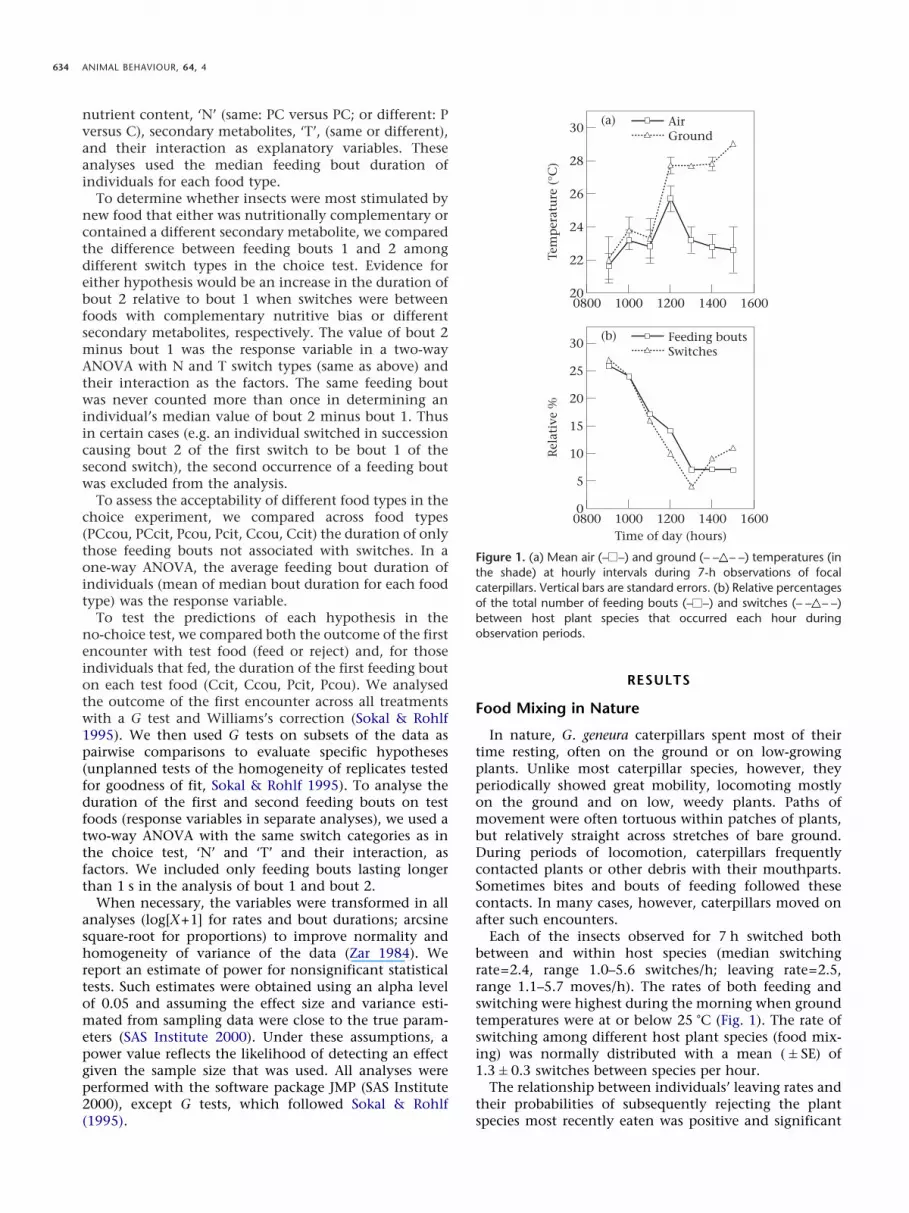
Figure 1. (a) Mean air (– –) and ground (– – – –) temperatures (inthe shade) at hourly intervals during 7-h observations of focalcaterpillars. Vertical bars are standard errors. (b) Relative percentagesof the total number of feeding bouts (– –) and switches (– – – –)between host plant species that occurred each hour duringobservation periods.
Food Mixing in Nature
In nature, G. geneura caterpillars spent most of theirtime resting, often on the ground or on low-growingplants. Unlike most caterpillar species, however, theyperiodically showed great mobility, locomoting mostlyon the ground and on low, weedy plants. Paths ofmovement were often tortuous within patches of plants,but relatively straight across stretches of bare ground.During periods of locomotion, caterpillars frequentlycontacted plants or other debris with their mouthparts.Sometimes bites and bouts of feeding followed thesecontacts. In many cases, however, caterpillars moved onafter such encounters.
Each of the insects observed for 7 h switched bothbetween and within host species (median switchingrate=2.4, range 1.0–5.6 switches/h; leaving rate=2.5,range 1.1–5.7 moves/h). The rates of both feeding andswitching were highest during the morning when groundtemperatures were at or below 25 �C (Fig. 1). The rate ofswitching among different host plant species (food mix-ing) was normally distributed with a mean (�SE) of1.3�0.3 switches between species per hour.
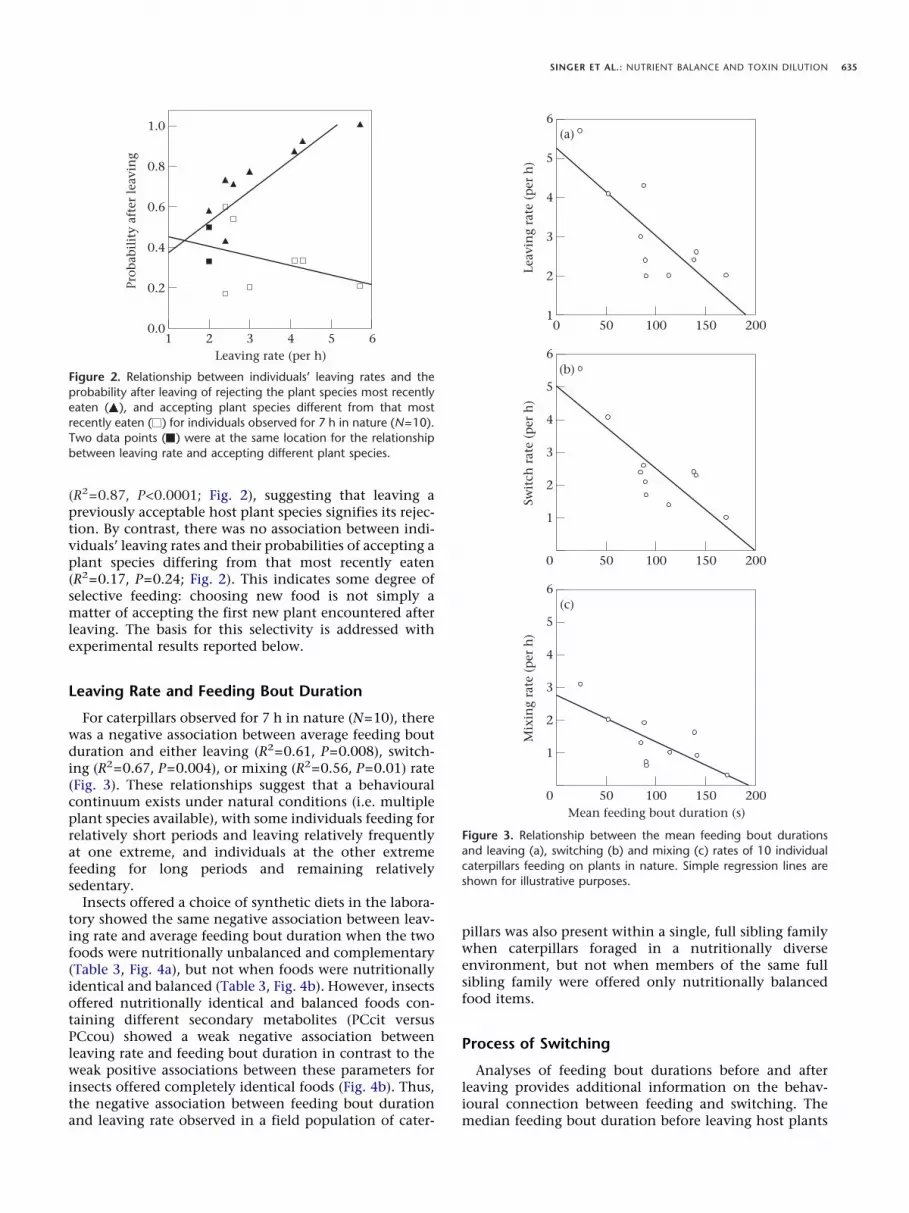
The relationship between individuals’ leaving rates andtheir probabilities of subsequently rejecting the plantspecies most recently eaten was positive and significant
635SINGER ET AL.: NUTRIENT BALANCE AND TOXIN DILUTION
(R2=0.87, P<0.0001; Fig. 2), suggesting that leaving apreviously acceptable host plant species signifies its rejec-tion. By contrast, there was no association between indi-viduals’ leaving rates and their probabilities of accepting aplant species differing from that most recently eaten(R2=0.17, P=0.24; Fig. 2). This indicates some degree ofselective feeding: choosing new food is not simply amatter of accepting the first new plant encountered afterleaving. The basis for this selectivity is addressed withexperimental results reported below.
Leaving Rate and Feeding Bout Duration
For caterpillars observed for 7 h in nature (N=10), therewas a negative association between average feeding boutduration and either leaving (R2=0.61, P=0.008), switch-ing (R2=0.67, P=0.004), or mixing (R2=0.56, P=0.01) rate(Fig. 3). These relationships suggest that a behaviouralcontinuum exists under natural conditions (i.e. multipleplant species available), with some individuals feeding forrelatively short periods and leaving relatively frequentlyat one extreme, and individuals at the other extremefeeding for long periods and remaining relativelysedentary.
Insects offered a choice of synthetic diets in the labora-tory showed the same negative association between leav-ing rate and average feeding bout duration when the twofoods were nutritionally unbalanced and complementary(Table 3, Fig. 4a), but not when foods were nutritionallyidentical and balanced (Table 3, Fig. 4b). However, insectsoffered nutritionally identical and balanced foods con-taining different secondary metabolites (PCcit versusPCcou) showed a weak negative association betweenleaving rate and feeding bout duration in contrast to theweak positive associations between these parameters forinsects offered completely identical foods (Fig. 4b). Thus,the negative association between feeding bout durationand leaving rate observed in a field population of cater-
pillars was also present within a single, full sibling familywhen caterpillars foraged in a nutritionally diverseenvironment, but not when members of the same fullsibling family were offered only nutritionally balancedfood items.
60.0
1.0
Leaving rate (per h)
Prob
abil
ity
afte
r le
avin
g
1 2 3 4
0.2
0.4
0.6
0.8
5
Figure 2. Relationship between individuals’ leaving rates and theprobability after leaving of rejecting the plant species most recentlyeaten ( ), and accepting plant species different from that mostrecently eaten ( ) for individuals observed for 7 h in nature (N=10).Two data points ( ) were at the same location for the relationshipbetween leaving rate and accepting different plant species.
200 0
6
Mean feeding bout duration (s)
Mix
ing
rate
(p
er h
)
50 100 150
2
3
4
5
1
(c)
200 0
6
Swit
ch r
ate
(per
h)
50 100 150
2
3
4
5
1
(b)
200 1
6
Leav
ing
rate
(p
er h
)
0 50 100 150
2
3
4
5
(a)
Figure 3. Relationship between the mean feeding bout durationsand leaving (a), switching (b) and mixing (c) rates of 10 individualcaterpillars feeding on plants in nature. Simple regression lines areshown for illustrative purposes.
Process of Switching
Analyses of feeding bout durations before and afterleaving provides additional information on the behav-ioural connection between feeding and switching. Themedian feeding bout duration before leaving host plants

636 ANIMAL BEHAVIOUR, 64, 4
Table 3. Analysis of covariance to explain interindividual variation in the rate of leaving food in the choice test withsynthetic diets
Source of variation df SS F P (power)
P versus C treatmentsFeeding bout duration 1 0.594 16.59 0.0003Diet 2 0.016 0.22 0.8050 (0.29)Feeding bout duration*diet 2 0.012 0.17 0.8413 (0.07)Error 30 1.075
PC versus PC treatmentsFeeding bout duration 1 0.015 0.73 0.3994 (0.13)Diet 2 0.075 1.85 0.0331Feeding bout duration*diet 2 0.072 1.76 0.1889 (0.34)Error 30 0.610
P: 85% protein; C: 85% carbohydrate; PC: 60:40% protein:carbohydrate.Adjusted R2 (P versus C diets=0.32; PC versus PC diets=0.21).
(field insects: all switching individuals pooled, N=30)differed according to whether or not the plant subse-quently eaten was the same individual, a different indi-vidual of the same species, or a different species (ANOVA:F2,51=6.42, P=0.003) (bout 1, Fig. 5). Specifically, shorterfeeding bouts preceded switches to different individualplants of the same (same versus different individual ofsame species contrast: F1,51=9.19, P=0.004) or differentspecies (same versus different species contrast: F1,51=8.97,P=0.004). The median feeding bout following leaving(bout 2, Fig. 5) did not differ according to switch type(ANOVA: F2,52=0.64, P=0.53, power=0.15). Therefore,shortened feeding bout durations were associated withthe initiation of host plant switching in nature.
The responses of insects offered a choice of syntheticfoods show that the reduced feeding bout duration priorto switching was associated with effects of secondarymetabolites but not primary nutrients (Table 4). Themean duration of bout 1 was shorter than the comparablemean nonswitching bout in treatments containing alter-native secondary metabolites (‘different’), but not inthose containing only a single one (‘same’) (Fig. 6). Thiswas true regardless of the nutrient content of food choices(P versus C, PC versus PC). Thus, insects respondeddifferently to food before switching in situations wherethey had experienced a single secondary metabolite ver-sus a combination of two during the conditioning period.Comparisons analogous to those done for field data(above) revealed no effects of nutrients or secondarymetabolites on the duration of the feeding bout on thesecond food (bout 2) relative to nonswitching bouts (datanot shown).
Comparison of the difference between feeding boutsafter and before switches (bout 2�bout 1) shows thatfeeding bout duration on new food increased when sec-ondary metabolites were different (Fig. 7b, Table 5) butnot when nutrient composition was complementary (Fig.7a, Table 5). Feeding bout 2 increased relative to bout 1 inswitches between foods with different secondary metabo-lites (Fig. 7b; PC versus PC, N=8: 5 cou to cit switches;3 cit to cou switches; P versus C, N=6: 3 Pcit to Ccou
switches; 3 Ccou to Pcit switches), but not in switchesbetween foods with the same secondary metabolites (PCversus PC, N=13 and P versus C, N=21). However, feedingbout 2 did not increase relative to bout 1 in switchesbetween nutritionally complementary foods (cit versuscit or cou versus cou, N=21: 11 P to C switches; 10 C to Pswitches; Pcit versus Ccou, N=6) compared to switchesbetween PC foods (Fig. 7a; cit versus cit or cou versus cou,N=13; cit versus cou, N=6).
In an analysis of all feeding bouts not associated withswitches, bout durations differed across food types in thechoice test (ANOVA on median feeding bout duration ofindividuals per food type: F5,88=3.20, P=0.01; Fig. 8).Longer feeding bouts on PC and P relative to C foodsindicate the protein-rich foods are the most phago-stimulatory (PC versus P contrast, F1,88=1.20, P=0.28,power=0.19; PC versus C contrast, F1,88=13.32, P=0.0004; P versus C contrast, F1,88=5.10, P=0.026). Similarfeeding bout durations on coumarin and citral foods ofeach nutritive type indicate similar acceptabilities ofthese two secondary metabolites (cou versus cit contrast,F1,88=0.040, P=0.84, power=0.05).
Responses of insects in the no-choice test confirm thatcaterpillars switching to new foods most readily acceptedand increased feeding on those with secondary metabo-lites that differed from the ones recently experienced. Theduration of final, conditioning feeding bouts on Ccit didnot differ across treatments prior to tests (ANOVA:F3,58=0.77, P=0.52, power=0.21). However, the decisionto feed or reject test foods differed according to diettreatment (G test: �2
3=17.28, P<0.001). Further compari-sons (Fig. 9) revealed differences for Ccit versus Pcit(G4,1=6.46, P<0.05) and Pcit versus Pcou (G4,1=11.54,P<0.05), but not for Ccit versus Ccou (G4,1=1.53, NS) andCcou versus Pcou (G4,1=0, NS). The first feeding bout onnew foods also differed greatly among treatments (Table6, Fig. 10a). Predictions of the nutrient balance hypoth-esis alone were not supported: the first feeding bouts on Cfoods were longer (opposite to predicted) than those onP foods (Fig. 10a). By contrast, predictions of the toxindilution hypothesis were supported: the first feeding

637SINGER ET AL.: NUTRIENT BALANCE AND TOXIN DILUTION
bouts on cou foods were longer than those on cit foods(Fig. 10a). There was no significant interaction betweennutrient content and secondary metabolites (Table 6),although the statistical power was relatively low (0.08).
The second feeding bout on new foods differed acrossdiet treatments as well (Table 6, Fig. 10b). The predictionof the nutrient balance remained unsupported. However,the effect of secondary metabolites was no longer consist-ent with the toxin dilution hypothesis. The appearance ofthe data (Fig. 10b) suggests an interaction between nutri-ents and secondary metabolites, yet it was not statisticallysignificant (Table 6, power=0.33) .
DISCUSSION
5000
4
Mean feeding bout duration (s)
(b)
1
3
2
PCcou versus PCcit
100 200 300 400
PCcou versus PCcouPCcit versus PCcit
4000
4(a)
1
3
2
Pcit versus Ccou
100 200 300
Pcou versus CcouPcit versus Ccit
Leav
ing
rate
(p
er h
)
Figure 4. Relationship between the mean feeding bout duration andleaving rate of individual caterpillars feeding on synthetic diets in thechoice test (diet P: 83% protein; diet C: 83% carbohydrate; diet PC:60:40% protein:carbohydrate; secondary metabolites: cou: cou-marin; cit: citral). These parameters were negatively associated forinsects with access to nutritionally unbalanced, complementary foodtypes (P versus C foods: (a)), but showed no association for insectsoffered two pieces of nutritionally balanced, identical foods (PCversus PC foods: (b)). Simple regression lines are shown forillustrative purposes. The continuous lines in (a) and (b) correspondto the Pcit versus Ccou and PCcit versus PCcit choice situations,respectively.
0
400
Food 2 type relative to food 1
Mea
n f
eed
ing
bou
t d
ura
tion
(s)
SP
100
200
300
NP NS
Bout 1Bout 2
Figure 5. Mean durations of final-instar feeding bouts before (bout1) and after (bout 2) leaving, categorized by the type of switchmade. The abbreviation ‘SP’ refers to leaving and returning to thesame individual plant, ‘NP’ refers to a switch to a different individualplant of the same species, and ‘NS’ refers to a switch to a plant of adifferent species. Vertical bars are standard errors.
Pattern of Food Mixing
The relatively long-term behavioural observations ofcaterpillars in nature showed a positive association
between two important components of food switching:(1) the rate of leaving food and (2) the probability ofsubsequently rejecting the food type most recently eaten.This association not only suggests that leaving foodgenerally signifies its rejection, but also a mechanisticconnection between leaving rate and the strength ofrejection. Individuals observed to leave host plants at thehighest rates nearly always rejected host plant speciesmost recently eaten upon subsequent encounter. Thetype of mechanism responsible is not known, but couldinclude physiological (e.g. detoxification enzymaticactivity) or neurological (e.g. behavioural habituation)processes differentially induced among individuals. Thatis, differential induction of detoxification enzymes orhabituation would cause individuals to differ in theirtolerance of host species, with the least tolerant individ-uals frequently leaving plants and rejecting the same typeupon subsequent encounter. Alternatively, this patterncould be explained by fixed differences (e.g. geneticvariation) among individuals in the tendency to mixfoods.
Conditional differences in foraging pattern imply thatthe process of food mixing behaviour depends upon bothfood characteristics and caterpillar experience. Field cat-erpillars (with a choice of host plants) and caterpillars inthe choice test given two different synthetic foods thatwere nutritionally unbalanced showed a negative associ-ation between leaving rate and average feeding boutduration (Figs 3, 4a). For field insects, this pattern is mostsimply (but perhaps not only) explained by the findingthat shortened feeding bouts preceded food switching(Fig. 5). So, individual caterpillars with high switching(and leaving) rates would have relatively many shortenedfeeding bouts, causing a reduction in average feedingbout duration. Thus when food choices were chemicallyheterogeneous, the process of food switching began withshortened feeding bouts, signifying rejection of the firstfood.

638 ANIMAL BEHAVIOUR, 64, 4
Table 4. Two-way analysis of variance showing the effects of primary nutrients (‘nutritive differences’ between foodchoices), secondary metabolites (‘toxin differences’ between food choices) and their interaction on the differencebetween the average feeding bout duration prior to switching (bout 1) and the average duration of feeding boutsnot associated with switching (bout 3) in the choice experiment
Source of variation df SS F P (power)
Nutritive differences 1 157.82 0.0039 0.9507 (0.05)Toxin differences 1 179 305.31 4.43 0.0448Nutrients*toxins 1 20 570.66 0.51 0.4821 (0.11)Error 27 1 093 340.8
Adjusted R2=0.09.
–300
200
Toxin
Bou
t 1
– bo
ut
3 (s
)
Same
–100
0
100
–200
Different
Figure 6. The mean difference between feeding bout 1 (boutimmediately before switching) and bout 3 (nonswitch bout), cat-egorized by the type of switch made (same versus different toxin foreach nutritive choice: : P versus C; : PC versus PC). Diet categoryabbreviations as in Fig. 4. Negative values indicate that the feedingbout prior to switches is reduced. Vertical bars are standard errors.
100
350
Bout 1
200
250
300
150
Bout 2
(b)
100
350
200
250
300
150
(a)
Mea
n f
eed
ing
bou
t d
ura
tion
(s)
Figure 7. Mean duration of feeding bouts 1 and 2 of switches in thechoice experiment with synthetic diets. Switches between foods thatcontained identical, balanced ( : PC versus PC) or complementary( : P versus C) nutrient bias address the nutrient balance hypothesis(a). Switches between foods that contained the same ( ) or different( ) secondary metabolites address the toxin dilution hypothesis (b).Diet category abbreviations as in Fig. 4. Vertical bars are standarderrors.
For laboratory insects, however, this pattern cannot beexplained as such because feeding bouts associated withswitches were not shortened for insects in two of the Pversus C treatments. Rather, nutritive variation amongfood choices provoked a variable response among the fullsibling individuals, ranging from leaving food relativelyoften in association with generally shorter feeding boutsto rarely leaving and taking longer feeds. This patternmay be explained by several possibilities. First, individ-uals may vary in behavioural response to nutrient imbal-ance according to differences in nutritional requirements(e.g. due to size or sex). Another possibility is that indi-viduals vary behaviourally per se, perhaps as a bet-hedging strategy (discussed below) or because there are avariety of equally successful ways to meet nutritionalrequirements. Regardless of the ultimate reason, it islikely that the more sedentary individuals compensatedfor nutritionally unbalanced food by feeding in longerbouts, thereby causing the negative association betweenleaving rate and average feeding bout duration.
The degree to which individual caterpillars (includingthose from the same full sibling family) varied in rates ofswitching and leaving in response to a choice of foodtypes is striking. In nature, such behavioural variationwould tend to make some individuals relative host plantspecialists and others relative generalists at any given
time. At an evolutionary level, the variance among indi-viduals in degree of food mixing is consistent with astrategy of diversified bet hedging (Philippi & Seger 1989;Carriere et al. 1995). Simply stated, this is a theoreticallydistinctive form of risk spreading wherein a genotypemay maximize its geometric mean fitness by producingphenotypically variable offspring, each phenotype suitedto a different environment. This strategy, like other formsof bet hedging, is particularly successful when theenvironment varies unpredictably over time, as is the casefor the herbaceous plant community in the habitat of

639SINGER ET AL.: NUTRIENT BALANCE AND TOXIN DILUTION
G. geneura (Singer 2000). While the mechanistic basis forthis variation cannot be evaluated without studies ofbehavioural repeatability or heritability, the variationitself is at least partly due to different leaving rates ofindividual caterpillars in response to nutritional vari-ation. This demonstration of individual variation was theonly influence of primary nutrients on food-switchingbehaviour.
0
400
Food type
Mea
n f
eed
ing
bou
t d
ura
tion
(s)
PCcou
100
200
300
PCcit Pcou Pcit Ccou Ccit
Figure 8. Mean durations of nonswitch bouts on different food typesin the choice experiment. Vertical bars are standard errors.
0.0
1.0
Test food
Freq
uen
cy o
f ac
cep
tan
ce
Ccit
0.2
0.5
0.8
Ccou Pcit Pcou
aa
b
a
Figure 9. Proportion of insects previously conditioned on Ccit thataccepted the test food upon first contact in the no-choice test.Diet category abbreviations as in Fig. 4. Letters denote significantdifferences derived from G tests (see text for details) .
0
250
Test foodCcit
50
150
Ccou Pcit Pcou
100
200
(b) Second test bout
0
250
Ccit
50
150
Ccou Pcit Pcou
100
200
(a) First test bout
Mea
n f
eed
ing
bou
t d
ura
tion
(s)
Figure 10. Mean durations of the first (a) and second (b) feedingbouts on test foods for insects previously conditioned on Ccit in theno-choice experiment. Diet category abbreviations as in Fig. 4.Vertical bars are standard errors.
Process of Switching
Laboratory experiments with synthetic diets revealedhow effects of secondary metabolites predominatedthroughout the process of switching between differentfood types, the essence of food mixing behaviour. Effectsof secondary metabolites but not primary nutrientsreduced feeding prior to switches, apparently inducingcaterpillars to leave food. Secondary metabolites furtherdetermined the acceptability of food subsequentlyencountered in a manner consistent with toxin dilution.
The reduced feeding bout duration immediately priorto switching foods in both field and laboratory settings
indicates reduced feeding excitation (Simpson 1995). Thisappears to be a response to secondary metabolites in thepresent study. In the laboratory choice experiment, feed-ing bouts prior to switches were reduced when foodchoices contained different secondary metabolites (asin nature), consistent with toxin dilution operating viaseveral possible processes. First, insects confined to foodchoices with a single secondary metabolite (unlike fieldinsects) would have ingested relatively greater amounts ofit than insects that switched between foods with differentsecondary metabolites. As a result of increased ingestion,the former could have relatively increased activity ofdetoxification enzymes (Glendinning & Slansky 1995;Snyder & Glendinning 1996). This metabolic increasemight curtail the potentially toxic accumulation ofingested secondary metabolites, allowing the retention ofa relatively high level of feeding excitation. Indeed, therewas no reduction in feeding prior to switching for insectswith a choice between foods with the same secondarymetabolite in this experiment. Second, the presence ofalternative foods may be detected: feeding insects mightsmell or become distracted by odours of different second-ary metabolites from nearby food, curtail feeding, thenswitch. If no alternative odours are detectable, feedingexcitation may remain constant until other processes

640 ANIMAL BEHAVIOUR, 64, 4
Table 5. Two-way analysis of variance showing the effects of primary nutrients (‘nutritive differences’ between foodchoices), secondary metabolites (‘toxin differences’ between food choices) and their interaction on the differencebetween the average duration of feeding bouts immediately prior to switching (bout 1) and the average durationof feeding bouts immediately after switching (bout 2) in the choice experiment
Source of variation df SS F P (power)
Nutritive differences 1 9403.46 0.24 0.6318 (0.08)Toxin differences 1 169 742.97 4.24 0.0492Nutrients*toxins 1 9538.52 0.24 0.6294 (0.08)Error 27 1 080 652.1
Adjusted R2=0.07.
Table 6. Analysis of variance showing the effects of primary nutrients (‘nutritive differences’ between conditioningand test foods), secondary metabolites (‘toxin differences’ between conditioning and test foods) and theirinteraction on the duration of the first (bout 1) and second (bout 2) feeding bouts on test food in the no-choiceexperiment
Source of variation df SS F P (power)
Bout 1Nutritive differences 1 2.98 16.12 0.0002Toxin differences 1 1.33 7.23 0.0093Nutrients*toxins 1 0.056 0.30 0.5854 (0.08)Error 58 10.70
Bout 2Nutritive differences 1 3.25 8.75 0.0021Toxin differences 1 0.63 1.69 0.6986 (0.07)Nutrients*toxins 1 0.79 2.14 0.1305 (0.33)Error 52 20.06
Adjusted R2 (bout 1=0.23, bout 2=0.13).
(e.g. intrinsic activity rhythm: Bernays & Singer 1998)cause switching. Third, insects might habituate to par-ticular secondary metabolites via a reduction in chemo-sensory sensitivity, possibly related to detoxificationactivity (Glendinning & Slansky 1995). Fourth, insectscould have learned whether more than one food type(based on secondary metabolites) was present, resultingin neurally driven changes in response to foods. Theseprocesses are not mutually exclusive.
Feeding stimulation from new foods was consistentwith toxin dilution but not nutrient balancing. Switchesin the choice test showed that new foods with a differentsecondary metabolite were more phagostimulatory thanfoods with the same one (Fig. 7b). Evidence for this comesfrom the increase in feeding duration from bout 1 to bout2 when the second food contained a different secondarymetabolite, but no such increase when foods in bout 1and 2 contained the same secondary metabolite. Whenswitches occurred between nutritionally complementaryfoods, there was no sign of increased phagostimulationby the second food relative to the first (Fig. 7a). Thisexperiment does not rule out the possibility of feedingpreference based on nutritive need occurring over alonger time scale (e.g. >4 h as reported for Spodopteralittoralis: Simpson et al. 1988). However, such a processwould not explain the high frequency of host plantswitching by G. geneura in nature.
The results of the no-choice experiment generallymatched the prediction that caterpillars exposed to food
containing one secondary metabolite would more readilyaccept (Fig. 9) and initially eat more (Fig. 10a) food witha new secondary metabolite relative to food with thesame secondary metabolite. For food acceptance, thisprediction was upheld only for insects tested with theprotein-biased foods. The effect may have been absent forthe carbohydrate-biased diet because of the extremelyhigh sucrose concentrations in the food, likely to over-whelm differences in chemosensory response to thecoumarin and citral. Sucrose is a potent phagostimulantfor G. geneura (Bernays et al. 2000). Because insects can-not taste protein directly, the secondary metabolites werelikely to have dominated the flavour of the protein-biasedfoods, causing a large difference in the acceptability ofPcit and Pcou. Initial feeding bout durations for thoseinsects that initiated feeding revealed no interactionbetween primary nutrients and secondary metabolites,consistent with other work showing no effect of inter-actions between dietary nutrient content and secondarymetabolites on deterrency (Glendinning & Slansky 1994).Together, these results suggest that caterpillars in natureare likely to reject, or feed only briefly, on host plantscontaining secondary metabolites recently ingested inlarge amounts. The relatively high probability of cater-pillars rejecting host plant species most recently eatensupports this idea (Fig. 2).
Changes in feeding duration between the first twobouts on test food in the no-choice test suggest theimportance of postingestive feedbacks from secondary

641SINGER ET AL.: NUTRIENT BALANCE AND TOXIN DILUTION
metabolites and nutrients. The mean feeding durationson Ccou and Ccit foods reversed in the second boutrelative to the first (Fig. 10a, b), indicating possible postingestive effects of secondary metabolites. Relatively longbouts on Ccit may have been possible because detoxifi-cation enzyme activity induced over the previous 20 hreduced the accumulation of citral, thus allowinga maximal amount of compensatory feeding on theprotein-deficient food. The decrease in feeding on Ccou isconsistent with short-term sensitization to coumarin orthe accumulation of quantities that provided some nega-tive postingestive feedback. The difference in bout dur-ation between Pcit and Pcou foods was maintained in thesecond bout, perhaps because many of the insects in theP treatments fed so little (or not at all) upon first contact.Therefore, these insects received little chemosensory orpostingestive information that could be used to modifysubsequent feeding. Some of the insects in the P treat-ments (especially those with the novel secondary com-pound), however, fed for longer periods and probablygained positive, postingestive feedbacks from nutrients.This explanation is suggested by the positive associationbetween the duration of feeding bouts 1 and 2 for indi-vidual caterpillars in the P treatments (Spearman rankcorrelation: rS=0.38, N=33, P<0.03), but not in the Ctreatments (rS= �0.03, N=36, NS).
Results of the no-choice test did not support themechanistic prediction that nutritionally complementaryfoods should be most phagostimulatory followingexposure to nutritionally unbalanced food (Simpson& Raubenheimer 1996). After 20 h of exposure tocarbohydrate-biased food, caterpillar feeding bout dur-ations on new, protein-biased foods were considerablyshorter than those on new, carbohydrate-biased foods(Fig. 10a). These feeding bouts were also much shorterthan those on the same protein-biased foods in the choicetest (Fig. 8). The comparison of nonswitch feeding boutsacross foods in the choice test (longer bouts on PC and Pthan on C foods) dispel the possibility that final instarsrejected P foods in the no-choice test because their nutri-ent intake targets (sensu Simpson & Raubenheimer 1993)were more carbohydrate biased than anticipated. Rather,this difference probably indicates that feeding bout dur-ations in the choice test show the ‘normal’ feeding ofundeprived, nutritionally homeostatic insects.
A possible problem in using synthetic diets like theones used here is their simplicity relative to natural hostplants. The surface concentrations of amino acids andsugars in plants, which directly elicit gustatory responsesof insects, may not accurately represent the internalconcentrations of digestible protein and carbohydrate(Bernays & Chapman 1994). While the lack of added freeamino acids in the synthetic diets used here may beunrealistic, several important parameters of foragingbehaviour were similar between insect responses to thesediets and natural host plants. We do not think these dietsdiminished our chance of detecting nutrient balancing,as experiments with similar diets have demonstratedfeeding decisions based on nutrient balancing over simi-lar time scales in grasshoppers (Simpson & Abisgold 1985;Chambers et al. 1997) and in the noctuid caterpillars
Helicoverpa (=Heliothis) zea (Cohen et al. 1988; Friedmanet al. 1991) and Spodoptera littoralis (Simmonds et al.1992). However, the low concentrations of singlesecondary metabolites used in our synthetic dietsmay have reduced our chances of detecting behaviourconsistent with toxin dilution, making this study a ratherconservative test of its relative importance.
We have presented experimental evidence for the inter-play between processes that balance nutrient intake andthose that reduce ingestion of particular secondarymetabolites in food mixing behaviour, along with fieldobservations consistent with these experimental results.Food mixing in herbivorous insects may frequentlyinvolve both nutrient balancing and toxin dilution, astheory generally predicts. It would be informative toknow if these processes vary in relative importanceamong different food mixing species. One might predict,for example, that species feeding on a mixture of grasses(e.g. Locusta migratoria, Orthoptera) would not oftenencounter the problem of toxicity from secondarymetabolites, which are typically not at noxious concen-trations in grass tissue. However, they would face greatdifficulty in ingesting an optimal nutrient balance due totypically high carbohydrate:nutrient ratios in grass tissue.Indeed L. migratoria routinely chooses a balanced nutrientintake from various combinations of nutritionally un-balanced but complementary foods (Behmer et al. 2001).By contrast, species like G. geneura that feed on forbs,characteristically defended with potently deterrent ortoxic secondary metabolites, would be expected to face theproblem of toxicity to a great degree. Further work thatrelates physiological processes and behaviour observed inthe laboratory to behavioural patterns in nature will benecessary to determine more generally the relative roles ofnutrient balancing and toxin dilution in food mixing.
Acknowledgments
We thank R. F. Chapman for help in designing experi-ments and helping us run them blind, and R. A.Abernathy, J. L. Spencer and E. M. Wintermute for assist-ance with observations. A. Telang kindly offered recipesfor synthetic diets. R. F. Chapman, N. Moran, A. Mira andan anonymous referee critically reviewed earlier drafts ofthis manuscript. We especially thank F. Slansky for aconstructively critical, thorough review of the manu-script. This work was supported by a fellowship to M.S.S.from The Center for Insect Science, University of Arizona,and by the NSF-funded Research Training Group inthe Analysis of Biological Diversification, University ofArizona.
References
Abisgold, J. D., Simpson, S. J. & Douglas, A. E. 1994. Nutrientregulation in the pea aphid Acyrthosiphon pisum: applicationof a novel geometric framework to sugar and amino acidconsumption. Physiological Entomology, 19, 95–102.
Behmer, S. T., Raubenheimer, D. & Simpson, S. J. 2001.Frequency-dependent food selection in locusts: a geometricanalysis of the role of nutrient balancing. Animal Behaviour, 61,995–1005.

642 ANIMAL BEHAVIOUR, 64, 4
Bernays, E. A. & Chapman, R. F. 1994. Host-plant Selection byPhytophagous Insects. New York: Chapman & Hall.
Bernays, E. A. & Chapman, R. F. 2000. Plant secondary compoundsand grasshoppers: beyond plant defenses. Journal of ChemicalEcology, 26, 1773–1794.
Bernays, E. A. & Chapman, R. F. 2001. Taste cell responses in thepolyphagous arctiid, Grammia geneura: towards a general patternfor caterpillars. Journal of Insect Physiology, 47, 1029–1043.
Bernays, E. A. & Singer, M. S. 1998. A rhythm underlying feedingbehaviour in a highly polyphagous caterpillar. PhysiologicalEntomology, 23, 295–302.
Bernays, E. A., Bright, K., Howard, J. J., Raubenheimer, D. &Champagne, D. 1992. Variety is the spice of life: frequentswitching between foods in the polyphagous grasshopperTaeniopoda eques Burmeister (Orthoptera: Acrididae). AnimalBehaviour, 44, 721–731.
Bernays, E. A., Bright, K. L., Gonzalez, N. & Angel, J. 1994. Dietarymixing in a generalist herbivore: tests of two hypotheses. Ecology,75, 1997–2006.
Bernays, E. A., Chapman, R. F. & Singer, M. S. 2000. Sensitivity tochemically diverse phagostimulants in a single gustatory neuronof a polyphagous caterpillar. Journal of Comparative Physiology A,186, 13–19.
Carriere, Y., Roff, D. A. & Deland, J. P. 1995. The joint evolution ofdiapause and insecticide resistance: a test of an optimality model.Ecology, 76, 1497–1505.
Chambers, P., Sword, G., Angel, J. E., Behmer, S. & Bernays,E. A. 1996. Foraging by generalist grasshoppers: two differentstrategies. Animal Behaviour, 52, 155–165.
Chambers, P. G., Raubenheimer, D. & Simpson, S. J. 1997. Theselection of nutritionally balanced foods by Locusta migratoria:the interaction between food nutrients and added flavours.Physiological Entomology, 22, 199–206.
Chapman, R. F. 1998. The Insects: Structure and Function. 4th edn.Cambridge: Cambridge University Press.
Cohen, R. W., Waldbauer, G. P. & Friedman, S. 1988. Natural dietsand self-selection: Heliothis zea larvae and maize. EntomologiaExperimentalis et Applicata, 46, 161–171.
Dethier, V. G. 1988. The feeding behaviour of a polyphagouscaterpillar (Diacrisia virginica) in its natural habitat. CanadianJournal of Zoology, 66, 1280–1288.
Freeland, W. J. & Janzen, D. H. 1974. Strategies in herbivory bymammals: the role of plant secondary compounds. AmericanNaturalist, 108, 269–289.
Freeland, W. J. & Saladin, L. R. 1989. Choice of mixed dietsby herbivores: the idiosyncratic effects of plant secondarycompounds. Biochemical Systematics and Ecology, 17, 493–497.
Friedman, S., Waldbauer, G. P., Eertmoed, J. E., Naeem, M. &Ghent, A. W. 1991. Blood trehalose levels have a role in thecontrol of dietary self-selection by Heliothis zea larvae. Journal ofInsect Physiology, 37, 919–928.
Glendinning, J. I. & Slansky, F., Jr. 1994. Interactions ofallelochemicals with dietary constituents: effects on deterrency.Physiological Entomology, 19, 173–186.
Glendinning, J. I. & Slansky, F., Jr. 1995. Consumption of a toxicfood by caterpillars increases with dietary exposure: support for arole of induced detoxification enzymes. Journal of ComparativePhysiology A, 176, 337–345.
Hagele, B. F. & Rowell-Rahier, M. 1999. Dietary mixing in threegeneralist herbivores: nutrient complementation or toxin dilution?Oecologia, 119, 521–533.
Karban, R. & Baldwin, I. T. 1997. Induced Responses to Herbivory.Chicago: University of Chicago Press.
Noldus, L. P. J. J. 1991. The Observer: a software system forcollection and analysis of observational data. Behavioural ResearchMethods, Instruments and Computers, 23, 415–429.
Philippi, T. & Seger, J. 1989. Hedging one’s evolutionary bets,revisited. Trends in Ecology and Evolutionary Biology, 4, 41–44.
Pulliam, H. R. 1975. Diet optimization with nutrient constraints.American Naturalist, 109, 765–768.
Rapport, D. J. 1980. Optimal foraging for complementaryresources. American Naturalist, 116, 324–346.
Raubenheimer, D. & Bernays, E. A. 1993. Patterns of feeding in thepolyphagous grasshopper Taeniopoda eques: a field study. AnimalBehaviour, 45, 153–167.
SAS Institute. 2000. JMP Version 4. Cary, North Carolina: SASInstitute.
Simmonds, M. S. J., Simpson, S. J. & Blaney, W. M. 1992. Dietaryselection behaviour in Spodoptera littoralis: the effects of condi-tioning diet and conditioning period on neural responsivenessand selection behaviour. Journal of Experimental Biology, 162,73–90.
Simpson, S. J. 1982. Patterns in feeding: a behavioural analysis usingLocusta migratoria nymphs. Physiological Entomology, 4, 325–336.
Simpson, S. J. 1995. Regulation of a meal: chewing insects. In:Regulatory Mechanisms in Insect Feeding (Ed. by R. F. Chapman &G. de Boer), pp. 137–156. New York: Chapman & Hall.
Simpson, S. J. & Abisgold, J. D. 1985. Compensation by locustsfor changes in dietary nutrients: behavioural mechanisms.Physiological Entomology, 10, 443–452.
Simpson, S. J. & Raubenheimer, D. 1993. A multi-level analysisof feeding behaviour: the geometry of nutritional decisions.Philosophical Transactions of the Royal Society of London, Series B,342, 381–402.
Simpson, S. J. & Raubenheimer, D. 1996. Feeding behaviour,sensory physiology and nutrient feedback: a unifying model.Entomologia Experimentalis et Applicata, 80, 55–64.
Simpson, S. J. & Raubenheimer, D. 2001. The geometric analysis ofnutrient–allelochemical interactions: a case study using locusts.Ecology, 82, 422–439.
Simpson, S. J., Simmonds, M. S. J. & Blaney, W. M. 1988.A comparison of dietary selection behaviour in larval Locustamigratoria and Spodoptera littoralis. Physiological Entomology, 13,225–238.
Simpson, S. J., Raubenheimer, D. & Chambers, P. G. 1995. Themechanisms of nutritional homeostasis. In: Regulatory Mechanismsin Insect Feeding (Ed. by R. F. Chapman & G. de Boer), pp. 251–278. New York: Chapman & Hall.
Singer, M. S. 2000. Ecological maintenance of food mixing in thewoolly bear caterpillar Grammia geneura (Strecker) (Lepidoptera:Arctiidae). Ph.D. thesis, University of Arizona, Tucson.
Singer, M. S. & Stireman, J. O., III. 2001. How foraging tacticsdetermine host-plant use by a polyphagous caterpillar. Oecologia,129, 98–105.
Slansky, F., Jr. 1992. Allelochemical-nutrient interactions inherbivore nutritional ecology. In: Herbivores: Their Interactions withSecondary Metabolites. Vol. 2 (Ed. by G. A. Rosenthal & M. R.Berenbaum), pp. 135–174. New York: Academic Press.
Slansky, F., Jr & Scriber, J. M. 1985. Food consumption andutilization. In: Comprehensive Insect Physiology, Biochemistry andPharmacology. Vol. 4 (Ed. by G. A. Kerkut & L. I. Gilbert),pp. 87–163. Oxford: Pergamon.
Snyder, M. J. & Glendinning, J. I. 1996. Causal connectionbetween detoxification enzyme activity and consumption of atoxic plant compound. Journal of Comparative Physiology A, 179,255–261.
Sokal, R. R. & Rohlf, F. J. 1995. Biometry. New York: W. H. Freeman.Stephens, D. W. & Krebs, J. R. 1986. Foraging Theory. Princeton,
New Jersey: Princeton University Press.Waldbauer, G. P. & Friedman, S. 1991. Self-selection of optimal
diets by insects. Annual Review of Entomology, 36, 43–63.

643SINGER ET AL.: NUTRIENT BALANCE AND TOXIN DILUTION
Waldbauer, G. P., Cohen, R. W. & Friedman, S. 1984. Self-selection of an optimal nutrient mix from defined diets by larvae ofthe corn earworm, Heliothis zea (Boddie). Physiological Zoology,57, 590–597.
Westoby, M. 1978. What are the biological bases of varied diets?American Naturalist, 112, 627–631.
Zar, J. H. 1984. Biostatistical Analysis. 2nd edn. Englewood Cliffs,New Jersey: Prentice Hall.





























