Embed Size (px)
Citation preview

This page intentionally left blank

The Mango, 2nd Edition
Botany, Production and Uses
Edited by
Richard E. Litz
Tropical Research and Education Center
and
Center for Tropical AgricultureUniversity of Florida
18905 SW 280 StreetHomestead, FL
33031-3314USA

CABI is a trading name of CAB InternationalCABI Head Offi ce CABI North American Offi ceNosworthy Way 875 Massachusetts AvenueWallingford 7th FloorOxfordshire OX10 8DE Cambridge, MA 02139UK USA
Tel: +44 (0)1491 832111 Tel: +1 617 395 4056Fax: +44 (0)1491 833508 Fax: +1 617 354 6875E-mail: [email protected] E-mail: [email protected]: www.cabi.org
© CAB International 2009. All rights reserved. No part of this publication may be reproduced in any form or by any means, electronically, mechanically, by photocopying, recording or otherwise, withoutthe prior permission of the copyright owners.
A catalogue record for this book is available from the British Library, London, UK.
Library of Congress Cataloging-in-Publication Data
The mango : botany, production and uses/edited by Richard E. Litz. -- 2nd ed. p. cm.
Includees bibliographical references and index.ISBN 978-1-84593-489-7 (alk. paper)1. Mango. I. Litz, Richard E.SB379.M2M35 2009634’.44--dc22 2008029843
ISBN-13: 978 1 84593 489 7
Typeset by AMA Dataset Ltd, Preston, UK.Printed and bound in the UK by the MPG Books Group, Bodmin.
The paper used for the text pages in this book is FSC certifi ed. The FSC (Forest Stewardship Council) is an international network to promote responsible management of the world's forests.

Contents
Contributors vii
Preface ix
Acknowledgements xi
1. Introduction: Botany and Importance 1S.K. Mukherjee and R.E. Litz
2. Taxonomy and Systematics 19J.M. Bompard
3. Important Mango Cultivars and their Descriptors 42R.J. Knight, Jr, R.J. Campbell and I. Maguire
4. Breeding and Genetics 67C.P.A. Iyer and R.J. Schnell
5. Reproductive Physiology 97T.L. Davenport
6. Ecophysiology 170B. Schaffer, L. Urban, P. Lu and A.W. Whiley
7. Fruit Diseases 210D. Prusky, I. Kobiler, I. Miyara and N. Alkan
8. Foliar, Floral and Soilborne Diseases 231R.C. Ploetz and S. Freeman
9. Physiological Disorders 303V. Galán Saúco
v

vi Contents
10. Pests 317J.E. Peña, M. Aluja and M. Wysoki
11. Crop Production: Propagation 367S. Ram and R.E. Litz
12. Crop Production: Mineral Nutrition 404I.S.E. Bally
13. Crop Production: Management 432J.H. Crane, S. Salazar-García, T.-S. Lin, A.C. de Queiroz Pinto and Z.-H. Shü
14. Postharvest Physiology 484J.K. Brecht and E.M. Yahia
15. Postharvest Technology and Quarantine Treatments 529G.I. Johnson and P.J. Hofman
16. World Mango Trade and the Economics of Mango Production 606E.A. Evans and O.J. Mendoza
17. Fruit Processing 628L.C. Raymundo, M.T. Ombico and T.M. de Villa
18. Biotechnology 641R.E. Litz, M.A. Gómez-Lim and U. Lavi
Index 671
The colour plate section can be found following page 372.

Contributors
N. Alkan, Department of Technology and Storage of Agricultural Products, Agricultural Research Organization (ARO), The Volcani Center, PO Box 6, Bet Dagan 50250, Israel.
M. Aluja, Instituto de Ecología AC, Km 2.5 Antigua Carretera a Coatepec No. 357, Congregación El Haya, Apartado Postal 63, CP 9100 Xalapa, Veracruz, Mexico. E-mail: [email protected]
I.S.E. Bally, Horticulture and Forestry Sciences, Department of Primary Industries and Fisheries, 28 Peters Street (PO Box 1054), Mareeba, QLD 48890, Australia. E-mail: [email protected]
J.M. Bompard, Les Mazes, 34160 Montaud, France. E-mail: [email protected]. Brecht, 1143 Fifi eld Hall, Department of Horticultural Sciences, University of Florida, Gaines-
ville, FL 32611-0690, USA. E-mail: [email protected] .eduR.J. Campbell, Fairchild Tropical Botanic Garden, 10901 Old Cutler Rd, Miami, FL 33156-4296,
USA. E-mail: [email protected]. Crane, Tropical Research and Education Center, University of Florida, 18905 SW 280 Street,
Homestead, FL 33031-3314, USA. E-mail: [email protected] .eduT.L. Davenport, Tropical Research and Education Center, University of Florida, 18905 SW 280 Street,
Homestead, FL 33031-3314, USA. E-mail: [email protected] .eduE.A. Evans, Tropical Research and Education Center, University of Florida, 18905 SW 280 Street,
Homestead, FL 33031-3314, USA. E-mail: [email protected] .eduS. Freeman, Department of Plant Pathology and Weed Research, Agricultural Research Organization
(ARO), The Volcani Center, Bet Dagan 50250, Israel. E-mail: [email protected]. Galán Saúco, Departamento de Fruticultura Tropical, Instituto Canario de Investigaciones Agrarias,
Apartado Correos 60, 38200 La Laguna, Tenerife, Canary Islands, Spain. E-mail: [email protected]. Gómez-Lim, Centro de Investigacion y de Estudios Avanzados del IPN (CINVESTAV), Apar-
tado Postal 629, Irapuato GTO, Mexico 36500. E-mail: [email protected]. Hofman, Department of Primary Industries and Fisheries, PO Box 5083, SCMS Nambour, QLD
4560, Australia. E-mail: [email protected]. Iyer, Indian Institute of Horticultural Research, Hessaraghatta Lake Post, Bangalore 560089,
India. E-mail: [email protected]. Johnson, Horticulture 4 Development, PO Box 412, Jamison, ACT 2614, Australia. E-mail: greg.
[email protected]. Knight, Jr, Tropical Research and Education Center, University of Florida, 18905 SW 280 Street,
Homestead, FL 33031-3314, USA. E-mail: [email protected] .edu
vii

viii Contributors
I. Kobiler, Department of Technology and Storage of Agricultural Products, Agricultural Research Organiza-tion (ARO), The Volcani Center, PO Box 6, Bet Dagan 50250, Israel. E-mail: [email protected]
U. Lavi, Department of Horticulture, Agricultural Research Organization (ARO), The Volcani Center, PO Box 6, Bet Dagan 50250, Israel. E-mail: [email protected]
T.-S. Lin, 111 Room, No. 4 Hall, Department of Horticulture, National Taiwan University, No. 1 Sec. 4 Roosevelt Road, 106 Taipei, Taiwan. E-mail: [email protected]
R.E. Litz, Tropical Research and Education Center, University of Florida, 18905 SW 280 Street, Home-stead, FL 33031-3314, USA. E-mail: relitz@ufl .edu
P. Lu, EWL Sciences, PO Box 39443, Winnellie, NT 0821, Australia. E-mail: ping.lu@ewlsciences. com.au
I. Maguire, Tropical Research and Education Center, University of Florida, 18905 SW 280 Street, Homestead, FL 33031-3314, USA. E-mail: imaguire@ufl .edu
O.J. Mendoza, Tropical Research and Education Center, University of Florida, 18905 SW 280 Street, Homestead, FL 33031-3314, USA.
I. Miyara, Department of Technology and Storage of Agricultural Products, Agricultural Research Organization (ARO), The Volcani Center, PO Box 6, Bet Dagan 50250, Israel.
S.K. Mukherjee (deceased), Department of Agriculture, Calcutta University, 35 Ballygunge Circu-lar Road, Calcutta 700 019, India.
M.T. Ombico, Fruit and Vegetable Laboratory, Food Science Cluster, College of Agriculture, Univer-sity of the Philippines Los Baños, Laguna, 4031, Philippines.
J.E. Peña, Tropical Research and Education Center, University of Florida, 18905 SW 280 Street, Home-stead, FL 33031-3314, USA. E-mail: [email protected] .edu
R.C. Ploetz, Tropical Research and Education Center, University of Florida, 18905 SW 280 Street, Homestead, FL 33031-3314, USA. E-mail: [email protected] .edu
D. Prusky, Department of Technology and Storage of Agricultural Products, Agricultural Research Organiza-tion (ARO), The Volcani Center, PO Box 6, Bet Dagan 50250, Israel. E-mail: [email protected]
A.C. de Queiroz Pinto, Private Consultant Tropical Fruits, SHCGN 706 Bloco P Casa 13, 70740-716, Brasilia-DF, Brazil. E-mail: [email protected]
S. Ram (deceased), Department of Horticulture, GB Pant University of Agriculture and Technology, Pantnagar 263 145, India.
L.C. Raymundo, Fruit and Vegetable Laboratory, Food Science Cluster, College of Agriculture, Uni-versity of the Philippines Los Baños, Laguna, 4031, Philippines. E-mail: [email protected]
S. Salazar-García, Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, Campo Experimental Santiago Ixcuintla, Km. 6 Carret. Intnal. Tepic-Mazatlán, Apartado Postal 100, San-tiago Ixcuintla, Nayarit 63300, Mexico. E-mail: [email protected]
B. Schaffer, Tropical Research and Education Center, University of Florida, 18905 SW 280 Street, Homestead, FL 33031-3314, USA. E-mail: [email protected] .edu
R.J. Schnell, United States Department of Agriculture (USDA) Agriculture Research Service (ARS), Subtropical Horticultural Research Unit/National Germplasm Repository, 13601 Old Cutler Road, Miami, FL 33158, USA. E-mail: [email protected]
Z.-H. Shü, Department of Biological Science and Technology, Meiho Institute of Technology, 23 Ping Kuang Road, Neipu, Pingtung, 91202, Taiwan. E-mail: [email protected]
L. Urban, Directeur de l’UR Génétique et écophysiologie de la qualité des agrumes (GEQA), Centre INRA de Corse, 20230 San Giuliano, France. E-mail: [email protected]
T.M. de Villa, Fruit and Vegetable Laboratory, Food Science Cluster, College of Agriculture, Univer-sity of the Philippines Los Baños, Laguna, 4031, Philippines.
A.W. Whiley, Sunshine Horticultural Services Pty Ltd, 287 Dulong Road, Nambour, QLD 4560, Australia. E-mail: [email protected]
M. Wysoki, Department of Entomology, Institute of Plant Protection, The Volcani Center, Bet Dagan 50250, Israel. E-mail: [email protected]
E.M. Yahia, Facultad de Química, Universidad Autónoma de Querétaro, Querétaro, 76190 Qro, Mexico. E-mail: [email protected]

ix
Preface
The fi rst edition of The Mango: Botany, Production and Uses appeared in 1997, and went into an unprecedented second printing in the following year. Despite the worldwide importance of the mango, this was the fi rst book that was devoted solely to this fruit crop species since the publication of The Mango by Gangolly et al. in 1957 and The Mango: Botany, Cultivation and Utilization by L.B. Singh in 1960. The appearance of The Mangoes: their Botany, Nomenclature, Hor-ticulture and Utilization by Kostermans and Bompard in 1993 had provided a much-needed taxonomic and systematic revision of mango and the related Mangifera species; the Kostermans and Bompard book also stimulated interest in the Mangifera spp. germplasm for breeding and rootstock development.
The Mango: Botany, Production and Uses (Litz, 1997) provided a fresh per-spective of the mango. The authors represented several countries, including India, Australia, Israel, the UK, France, USA, Mexico, Pakistan and South Africa, and refl ected the expansion of mango production outside its tradi-tional areas of cultivation during the mid-20th century and the development of new technologies in these new lands. The worldview of the fi rst edition was unique, and the authors were at the forefront of the advance of science in support of mango production. I wish to particularly acknowledge L.A. Milne (South Africa), R.V. Mosqueda-Vazquez (Mexico), S.K. Mukherjee (India) and S. Ram (India), who contributed to the fi rst edition, and who have passed away since then.
Since 1997, other mango books have appeared: El Cultivo del Mango by V. Galán Saúco in 1999 (Spain), Mango Cultivation edited by R.P. Srivastava in 1998 (India), A Cultura da Mangueira edited by P.J. de Carvalho Genu and A.C. de Queiroz Pinto in 2002 (Brazil) and El Mango by E. Yahia Kazuz, J. de J. Ornelas Paz and R. Ariza Flores in 2006 (Mexico). These books have gener-ally targeted audiences in specifi c mango-producing countries. Drs Galán Saúco, Pinto and Yahia are also contributors to the second edition of TheMango: Botany, Production and Uses.

x Preface
Much has happened in the decade following the appearance of the fi rst edition of The Mango: Botany, Production and Uses. China has emerged as the second largest producer of mango fruit; India’s production is now less than half of the world total. Fresh mangoes are now consumed worldwide and are available year-round in the European Union (EU), North America and Japan. The availability of fruit of a range of mango cultivars is increasing. Mango products, including fruit nectars, leather, dried fruit slices, preserves, yogurt, etc. have become widely popular outside the tropics.
The authorship of the second edition of The Mango: Botany, Production and Uses represents the USA, Mexico, Brazil, Australia, the Philippines, Tai-wan, India, Israel, France and Spain, and includes leading authorities in each fi eld. The subject matter of this book ranges from the most basic to the applied, and is designed to be a compendium that will remain highly relevant for researchers and growers for many years.
I would like to express my appreciation and thanks to all of the authors for their persistence during the 3-year gestation period. I would like to express my gratitude to Ian Maguire of the Tropical Research and Education Center of the University of Florida for his photographic assistance. Financial assistance provided by Dr Yungcong Li, also of the Tropical Research and Education Center, for reproduction of colour plates is gratefully acknowl-edged. Special thanks to Pamela A. Moon, Guillermo Padilla and Irene Perea who tolerated me while I worked on this project.
Richard E. Litz
References
de Carvalho Genu, P.J. and de Queiroz Pinto, A.C. (eds) (2002) A Cultura da Mangueira.EMBRAPA Informacao Tecnologica, Brasilia DF.
Galán Saúco, V. (1999) El Cultivo del Mango. Mundi-Prensa, Madrid.Gangolly, S.R., Singh, R., Katyal, S.L. and Singh, D. (1957) The Mango. Indian Council
for Agricultural Research, New Delhi, India.Kostermans, A.J.G.H. and Bompard, J.M. (1993) The Mangoes: their Botany, Nomencla-
ture, Horticulture and Utilization. Academic Press, London.Litz, R.E. (ed.) (1997) The Mango: Botany, Production and Uses. CAB International,
Wallingford, UK.Singh, L.B. (1960) The Mango: Botany, Cultivation and Utilization. Leonard Hill, London.Srivastava, R.P. (1998) Mango Cultivation. International Book Distributing Co, Lucknow,
India.Yahia Kazuz, E., de J. Ornelas Paz, J. and Ariza Flores, R. (2006) El Mango. Editorial Trillas,
S.A. de C.V., Mexico.

xi
Acknowledgements
The assistance provided by Ian Maguire is gratefully acknowledged. Artwork for the covers was provided by Ian. Special thanks to Campbell, Penelope and Anna.

This page intentionally left blank

© CAB International 2009. The Mango, 2nd Edition: Botany, Production and Uses(ed. R.E. Litz) 1
1 Introduction: Botany and Importance
S.K. Mukherjee1 and R.E. Litz2
1Calcutta University, Calcutta, India2University of Florida, Florida, USA
1.1 Introduction 11.2 Description of Mango 2
The tree 2Flowers 2The fruit 3The seeds and polyembryony 4
1.3 History of Cultivation 5Origin of Mangifera indica 5Domestication of mango 9Distribution 10
1.4 Germplasm Conservation 11Genetic erosion 11Collection and documentation of Mangifera germplasm 12Relevance of germplasm resources to mango improvement 12
1.5 Importance of Mango 12Cultivars 12
1.6 Production and Uses 14
1.1 Introduction
Mango has become a major fruit crop of the tropics and subtropics, particu-larly in Asia, where the mango has always been the most important fruit crop and where it has been considered the ‘king of fruits’ (Purseglove, 1972). A generation ago, the Green Revolution culminated, creating surpluses of sta-ple and horticultural crops in many developing countries. The Green Revo-lution was the result of nearly a century of effort of applying Mendelian genetics to crop improvement (i.e. conventional breeding) together with the optimization of agronomic and horticultural practices and the successful management of insect pests and diseases. However, improvement of tree

S.K. Mukherjee and R.E. Litz2
crops has lagged far behind fi eld crops for several reasons: their heterogene-ity, polyploidy, lengthy juvenile period, time required for evaluation of trees in the fi eld, and the relatively high cost of maintaining tree plantings. For the most part, fruit cultivars continue to be ancient selections, many of which have serious problems, including alternate bearing, lack of disease resistance, low yields, etc. The rapid growth of mango production in recent years has been due to its expansion into new growing regions of the New World, China and parts of Africa; the planting of regular bearing selections; and the adop-tion of modern fi eld practices, which include irrigation management, control of fl owering, etc. Agricultural practices are currently undergoing another revolution, as integrated pest and disease management replaces the earlier reliance on agrichemicals, and emerging fi elds within biotechnology begin to impact cultivar development.
1.2 Description of Mango
The tree
The mango tree is believed to have evolved as a canopy layer or emergent species of the tropical rainforest of South and South-east Asia (Kaur et al., 1980; Bompard, Chapter 2, this volume). Mature trees can attain a height of 40 m or more, and can survive for several hundred years. Mango trees that have been domesticated by selection from openly pollinated seedling popu-lations show variation in tree architecture (i.e. shape and size). The tree is an arborescent evergreen. Leaves are simple and alternate, with petioles that range in length from 1 to 12.5 cm. Leaf morphology is highly variable, de-pending on the cultivar: leaves can be lanceolate, oblong, ovate and interme-diate types involving these forms. Leaf length ranges from 12 to 38 cm and width can be between 2–13 cm. Young leaves are copper-coloured, changing gradually to light and then dark green with age. The leaves are spirally arranged in whorls and are produced in fl ushes. The canopy is normally oval, elongated or dome shaped. The juvenile period of seedling trees can range from 3 to 7 years. The root system consists of a long, vigorous taproot and abundant surface feeder roots.
Flowers
Mango fl owers are borne on terminal pyramidal panicles, and are glabrous or pubescent; the infl orescence is rigid and erect, up to 30 cm long, and is widely branched, usually tertiary, although the fi nal branch is always cymose. The infl orescence is usually densely fl owered with hundreds of small fl ow-ers, which are 5–10 mm in diameter. The fl owers are either monoecious or polygamous, and both monoecious and polygamous fl owers are borne within a single infl orescence (Plate 1). The pistil aborts in male fl owers. The ratio of monoecious to polygamous fl owers is strongly infl uenced by

Introduction: Botany and Importance 3
environmental and cultural factors. The fl owers have four or fi ve sepals and petals that are ovate to ovoid to lanceolate and also thinly pubescent. The fl oral disc also is four- or fi ve-lobed, fl eshy and large and located above the base of the petals. There are fi ve large, fl eshy stamens, only one or two of them being fertile; the remaining stamens are sterile staminodes that are sur-mounted by a small gland. In addition, two or three smaller fi laments arise from the lobes of the nectaries. The stamens are central. The ovule is anatro-pous and pendulous. It is believed that the fl owers are cross-pollinated by fl ies (see Davenport, Chapter 5, this volume).
Mukherjee (1951a, 1953) investigated the pollen morphology of mango and 12 other Mangifera species. Their pollen grains were tricolpate of almost the same size. Mondal et al. (1982, cited in Kostermans and Bom-pard, 1993) attempted to correlate pollen morphology with taxonomic relationships of 17 Mangifera species based upon different characteristics of the exine and sporoderm. They demonstrated that all of the species of section II (subgenus Limus) possess coarse exine; whereas there was no clear correlation with pollen type in species within section I (subgenus Mangifera).
The fruit
DescriptionThe mango fruit is a large, fl eshy drupe, containing an edible mesocarp of varying thickness. The mesocarp is resinous and highly variable with respect to shape, size, colour, presence of fi bre and fl avour. The fl avour ranges from turpentine to sweet. The exocarp is thick and glandular. There is a character-istic beak that develops laterally on the proximal end of the fruit. A sinus is always present above the beak. Fruit shape varies, including elongate, oblong and ovate or intermediate forms involving two of these shapes. Fruit length can range from 2.5 to > 30 cm, depending on the cultivar. The endo-carp is woody, thick and fi brous; the fi bres in the mesocarp arise from the endocarp.
The mango fruit is climacteric (see Brecht and Yahia, Chapter 14, this volume), and increased ethylene production occurs during ripening. Chloro-phyll, carotenes, anthocyanins and xanthophylls are all present in the fruit. The skin is generally a mixture of green, red and yellow pigments, although fruit colour at maturity is genotype dependent. During ripening the chloro-plasts in the peel become chromoplasts, which contain yellow and red pig-ments (Krishnamurthy and Subramanyam, 1970; Akamine and Goo, 1973; Salunkhe and Desai, 1984; Mitra and Baldwin, 1997). Peel colour obviously is cultivar dependent (see Knight et al., Chapter 3, this volume). Fruit of ‘Bom-bay Green’ is green; ‘Carabao’, ‘Manila’, ‘Mulgoa’ and ‘Arumanis’ are greenish-yellow; ‘Dashehari’ and ‘Alphonso’ are yellow; and ‘Haden’, ‘Keitt’ and ‘Tommy Atkins’ have a red blush. The red blush is due to the presence of anthocyanins (Lizada, 1991). The pulp carotenoids in ripe fruit also vary with respect to cultivar (Mitra and Baldwin, 1997).

S.K. Mukherjee and R.E. Litz4
FlavourFlavour of the mango mesocarp is a function of carbohydrates, organic acids, lactones, monoterpene hydrocarbons and fatty acids (Mitra and Baldwin, 1997). During fruit maturation, starch that accumulates in the chloroplasts is hydrolysed to sucrose, glucose and fructose (Medlicott et al., 1986; Selvaraj et al., 1989; S. Kumar et al., 1994); sucrose is present in slightly higher concen-trations than either fructose or glucose. Organic acid content decreases dur-ing ripening (Krishnamurthy and Subramanyam, 1970). The dominant organic acid is citric acid, but glycolic acid, malic acid, tartaric acid and oxalic acids are also present (Sarker and Muhsi, 1981; Medlicott and Thompson, 1985). The peach-like fl avour of mangoes is attributed to the presence of lac-tones (Lakshminarayana, 1980; Wilson et al., 1990).
NutritionMango fruit contain amino acids, carbohydrates, fatty acids, minerals, organic acids, proteins and vitamins. During the ripening process, the fruit are ini-tially acidic, astringent and rich in ascorbic acid (vitamin C). Ripe mangoes contain moderate levels of vitamin C, but are fairly rich in provitamin A and vitamins B1 and B2. Perry and Zilva (1932) determined the vitamin A, C and D content of the fruit of three Indian mango cultivars, and found that the pulp of mangoes is a concentrated source of vitamin C. The pulp of mango fruit contains as much vitamin A as butter, although vitamin D is not present in a signifi cant quantity. Fruit acidity is primarily due to the presence of malic and citric acids. In addition, oxalic, malonic, succinic, pyruvic, adipic, galac-turonic, glucuronic, tartaric, glycolic and mucic acids are also present (Jain et al., 1959; Fang, 1965). Acidity is cultivar related; for example, immature Florida cultivars have low acidity (0.5–1.0%) in comparison with ‘Alphonso’ (3%). During ripening, acidity decreases to 0.1–0.2%. Following fruit set, starch accumulates in the mesocarp. Free sugars, including glucose, fructose and sucrose, generally increase during ripening; however, the sucrose content increases three- to fourfold due to the hydrolysis of starch. Sucrose is the principal sugar of ripe mangoes. The sucrose content of ripe fruit of three Indian cultivars, ‘Alphonso’, ‘Pairie’ and ‘Totapuri’, ranges from 11 to 20% representing 15 to 20% of the total soluble solids (Popenoe, 1932).
The seeds and polyembryony
Mango seeds are solitary, large and fl at, ovoid oblong and surrounded by the fi brous endocarp at maturity. The testa and tegumen are thin and papery. Embryos are dicotyledonous. Seeds of monoembryonic mango types contain a single zygotic embryo, whose cotyledons can be unequal in size or lobed in shape. The seeds of polyembryonic mango types contain one or more embryos (Plate 2); usually one embryo is zygotic, whereas the remaining embryos are derived directly from the nucellus, a maternal tissue. Nucellar embryos apparently lack a suspensor. Polyembryony has also been reported in Mangifera casturi, M. laurina and M. odorata (Bompard, 1993). Certain

Introduction: Botany and Importance 5
polyembryonic cultivars reportedly can produce seeds with adventitious nucellar embryos only, for example ‘Strawberry’ (Juliano, 1934), ‘Carabao’ and ‘Pico’ (Juliano and Cuevas, 1932) and ‘Olour’ and ‘Cambodiana’ (Maheshwari et al., 1955). Early studies suggested that polyembryony appeared to be a polygenic trait (Juliano, 1934; Sturrock, 1968), segregating as a recessive character in the progeny of controlled crosses. Recent studies, however, have demonstrated that the polyembryony trait is inherited as a dominant character (Aron et al., 1998). Several studies have shown that nucel-lar seedlings can be distinguished from the single zygotic seedling of poly-embryonic seeds by isozymes (Schnell and Knight, 1992; Degani et al., 1993) and DNA markers, for example single sequence repeats (SSRs) (Eiadthong et al., 1999a), amplifi ed fragment length polymorphisms (AFLPs) (Kashkush et al., 2001) and inter-simple-sequence-repeats (ISSRs) (Gonzalez et al., 2002). Mango seeds are considered to be recalcitrant, and cannot survive for more than a few days or weeks at ambient temperatures (Parisot, 1988). This important characteristic of mango seeds would have inhibited the long distance dis-persal of mango by seed until recent times.
1.3 History of Cultivation
Origin of Mangifera indica
The largest number of Mangifera species occurs in the Malay Peninsula, the Indonesian archipelago, Thailand, Indochina and the Philippines (Mukher-jee, 1985; Bompard, 1989; see Bompard, Chapter 2, this volume). The most recent classifi cation of Mangifera species was based upon fl oral morphology (Kostermans and Bompard, 1993) and included 69 species, most of which are included in two subgenera Mangifera and Limus with another 11 species occupying an uncertain position (Table 1.1). Eiadthong et al. (1999b) described the phylogenetic relationships among Mangifera species using genomic restriction fragment length polymorphisms (RFLPs) and amplifi cation of chloroplast DNA (cpDNA), and suggested that the Mangifera species should be classifi ed using molecular data. In the next few years, it is likely that molecular biology will have a major impact on phylogenetic studies involving mango and its relatives.
Mangifera species with a single fertile stamen are distributed in north-eastern India, Myanmar, Thailand and the Malay Peninsula. Many of the mango relatives have small fruits with thin, acidic fl esh, large seeds, abun-dant fi bre and astringent resinous substances that are localized near the skin. In addition to M. indica, edible fruit is produced by at least 26 other species in the genus, primarily species found in South-east Asia (Gruezo, 1992). Mangifera caesia, known as ‘binjai’ or ‘kemang’ in South-east Asia, is culti-vated in Java, where it bears fruit in the mango off-season (Bompard, 1992a). Mangifera foetida is less commonly cultivated due to its highly astringent fruit; however, the fruit is widely used for pickling and as a substitute for tamarind (Bompard, 1992b). Mangifera kemang and M. altissima are consumed

S.K. M
ukherjee and R.E. Litz
6
Table 1.1. Classifi cation of Mangifera species according to Kostermans and Bompard (1993).
Genus Subgenus Section Species
Mangifera Mangifera Marchandora Pierre M. gebede MiqEuantherae Pierre M. caloneura Kurz
M. cochinchinensis EnglerM. pentandra Hooker f.
Rawa Kosterm. M. andamanica King M. minutifolia Evard.M. gracilepes M. nicobarica Kosterm.M. griffi thii Hooker f. M. paludosa Kosterm.M. merrillii Mukherji M. parvifolia Boerl. & Koorders M. microphylla Griff. ex Hooker f.
Mangifera Ding Hou M. altissima Blanco. M. mucronulata Bl.M. applanata Kosterm. M. oblongifolia Hooker f.M. austro-indica Kosterm. M. orophila Kosterm.M. austro-yunnanensis Hu M. pedicellata Kosterm.M. casturi Kosterm. M. pseudo-indica Kosterm.M. collina Kosterm. M. quadrifi da JackM. dewildei Kosterm. M. rigida Bl.M. dongnaiensis Pierre M. rubropetala Kosterm.M. fl ava Evard. M. rufocostata Kosterm.M. indica L. M. similis Bl.M. lalijiwa Kosterm. M. sulauesiana Kosterm.M. laurina Bl. M. sumbawaensis Kosterm.M. linearifolia (Mukherji) Kosterm. M. sylvatica Roxb.M. longipetiolata King M. swintonioides Kosterm.M. magnifi ca Kochummen M. timorensis Bl.M. minor Bl. M. torquenda Kosterm.M. monandra Merr. M. zeylanica (Bl.) Hooker f.

Introduction: Botany and Im
portance7
Limus (Marchand) Kosterm.
M. blommesteinii Kosterm. M. leschenaultii MarchandM. caesia Jack M. macrocarpa Bl.M. decandra Ding Hou M. odorata Griff.M. foetida Lour. M. pajang Kosterm.M. kemanga Bl. M. superba Hooker f.M. lagenifera Griff.
Species of uncertain position
M. acutigemma Kosterm. M. persiciformis Wu & MingM. bompardii Kosterm. M. subsessifolia Kosterm.M. bullata Kosterm. M. taipa Buch.-HamiltonM. campospermoides Kosterm.
M. transversalis Kosterm.
M. hiemalis Liang Jian Ying M. utana UtanaM. maingayii Hooker f.

S.K. Mukherjee and R.E. Litz8
as fresh fruit or used green as a salad (Angeles, 1992; Bompard, 1992a). Mangifera pajang has the largest fruit in the genus, and is an attractive fruit. Mangifera odorata is grown in the Philippines and Indonesia, and has occa-sionally been used as a rootstock for mango (Ochse, 1931; Bompard, 1992c). Mangifera odorata is widely grown in the humid lowlands of South-east Asia in areas that are unsuitable for mango as a mango substitute. Mangifera lau-rina and M. pentandra are appreciated as salad ingredients (Bompard, 1992d). In addition, M. griffi thii, M. minor, M. monandra, M. quadrifi da and M. similis have palatable fruit that are considered to have great potential (Gruezo, 1992). All mango cultivars belong to the species M. indica.
According to De Candolle (1884), ‘It is impossible to doubt that it (the mango) is a native of south Asia or of the Malay Archipelago, when we see the multitude of varieties cultivated in those countries, the number of ancient names, in particular a Sanskrit name, its abundance in the gardens of Bengal, of Deccan peninsula, and of Ceylon even in Rheede’s time (i.e. 1683).’ Although the centre of origin and diversity of the genus Mangifera is now fi rmly established as being in South-east Asia, the origin of M. indica has been a matter of speculation for many years. The fossil record provides few clues, as only a single fossil bearing the imprint of a leaf of M. pentandra has ever been found (Seward, 1912). Mangifera indica is believed to have fi rst appeared during the Quatenary period (Mukherjee, 1951b). Blume (1850) considered that mango might have originated from several related species, primarily located in the Malay archipelago.
On the basis of ancient accounts of travellers and the written historical record, it was believed for many years that mango must have originated in India and spread outwards from there to South-east Asia and thence to the New World and Africa. Because north-eastern India is at the northernmost edge of the distribution of the Mangifera species, Hooker (1876) suggested that mango might have been naturalized in India. The historical record pro-vides a sometimes confl icting account of the distribution of mango. Miquel (1859) did not record it as being wild in the Indonesian archipelago. Accord-ing to Rumphius (1741), the mango was introduced into certain islands of the Indonesian archipelago within recent times; however, the mango was in cul-tivation in Java at least as early as ad 900–1100, when the temple at Borobo-dur was built and faced with carvings of the Buddha in contemplation under a mango tree (Plate 3). Based upon taxonomic and recent molecular evidence, it is now apparent that the mango probably evolved within a large area including north-western Myanmar, Bangladesh and north-eastern India (see Bompard, Chapter 2, this volume).
Polyembryonic and monoembryonic M. indicaWithin M. indica, there are two distinct types that can be distinguished on the basis of their mode of reproduction and their respective centres of diversity: a subtropical group with monoembryonic seed (Indian type) and a tropical group with polyembryonic seed (South-east Asian). A few polyembryonic cultivars occur along the west coast of India; however, they may have been introduced into Goa from South-east Asia, perhaps by the Portuguese from

Introduction: Botany and Importance 9
their colonies of Malacca in the Malay Peninsula or Timor in the Indonesian archipelago. Kumar et al. (2001) estimated the genetic relatedness among ten polyembryonic and monoembryonic cultivars from the west coast of south-ern India using genomic and chloroplast DNA RFLP analysis. The cultivars could be grouped on the basis of embryo type (i.e. monoembryonic and poly-embryonic) and had distinctly different genetic backgrounds. They con-cluded that polyembryonic mangoes could not have originated in India, and must have been introduced, probably from South-east Asia.
Domestication of mango
Historical recordIt is probable that mango cultivation originated in India, where De Candolle (1884) estimated that mango cultivation appeared to have begun at least 4000 years ago. In the early period of domestication, mango trees probably yielded small fruit with thin fl esh. Such fruit can be found today in north-eastern India and in the Andaman Islands (Anonymous, 1992). Folk selections of superior seedlings over many hundreds of years would have resulted in larger fruit with thicker fl esh. Mukherjee (1950a, b) described many of these primitive selections from Orissa in north-eastern India; they demonstrated great variation in fruit shape and size.
The mango is a very important cultural and religious symbol of India. Buddhist pilgrims Fa-Hien and Sung-Yun mentioned in their travel notes that the Gautama Buddha was presented with a mango grove by Amradarika (c.500 bc) as a place for meditation (Popenoe, 1932). According to Burns and Prayag (1921), a mango tree is depicted in friezes on the stupa of Bharut, which was constructed c.100 bc. Other travellers to India, including the Chi-nese Hwen T’sung (ad 632–645), the Arabs Ibn Hankal (ad 902–968) and Ibn Batuta (ad 1325–1349) and the Portuguese Lurdovei de Varthema (ad 1503–1508), all described the mango. The Indian subcontinent was the birthplace of some of the earliest highly developed civilizations, and over the centuries, India exerted strong cultural, religious and commercial infl uence over South and South-east Asia. In successive waves, Hinduism, Buddhism and Islam were introduced into South-east Asia from India. To this day, many com-monly used words in Indonesia are derived from both Sanskrit and Tamil. One of the most widely used words for mango in Malaysia and Java (Indone-sia) is ‘mangga’, which is derived from the Tamil ‘manga’. Traders and monks from India possibly introduced superior selections of mango into South-east Asia; however, vegetative propagation was unknown in India until after the arrival of the Portuguese in Goa in the 15th century. Moreover, the most im-portant mango selections of Thailand, Cambodia, Vietnam, Malaysia, Indo-nesia and the Philippines historically have all been of the polyembryonic type, and have traditionally been seed propagated. Until the establishment of Portuguese enclaves on the coast of India beginning in the late 15th century, mango cultivars did not exist in India, as there was no known method for vegetatively propagating superior selections (see Iyer and Schnell,

S.K. Mukherjee and R.E. Litz10
Chapter 4, this volume). However, under the Moghul emperor Akbar (1556–1605), the best selections of seedling mangoes were propagated by approach grafting and were planted in large orchards. The ‘Lakh Bagh’, a mango orchard of 100,000 trees, was planted near Darbhanga in Bihar. Perhaps noth-ing more eloquently attests to the importance of this fruit and the esteem in which it was held than this vast mango orchard. The Ain-i-Akbari, an ency-clopedic work that was written during the reign of Akbar, contains a lengthy account of the mango, and includes information about the quality of the fruit and varietal characteristics. There was evidently a strong body of informa-tion about mango cultivation that had accumulated up to that time. Most of the mango cultivars of India had their origin in those years, and have been maintained under cultivation for over 400 years by vegetative propagation. ‘Alphonso’, ‘Dashehari’, ‘Langra’, ‘Rani Pasand’, ‘Safdar Pasand’ and other mango cultivars were selected during that time. Relics of orchards from the time of Akbar are found in different parts of India, and it has been suggested that they could still provide valuable material for selection of superior mango cultivars.
Distribution
Spreading from the centres of domesticationThe global spread of mangoes and their cultivation outside their original centres of domestication probably did not occur until the beginning of the European voyages of discovery and colonialization in the 15th and 16th centuries. Because mango seeds are recalcitrant, and cannot survive for more than a few days or weeks, mango germplasm in the early days must have been transported as ripe fruit, seedlings or, later on, as grafted plants. It is believed that the Portuguese transported the mango from their colonies in India to their African colonies, although Purseglove (1972) suggested that it might also have been introduced to Africa via Persia and Arabia in the 10th century by Arab traders. The Portuguese later introduced the mango into Brazil from their African colonies of Mozambique and Angola. Spaniards, who encountered a mango-growing civilization in the Philippines after Magellan’s passage across the Pacifi c Ocean, introduced polyembryonic mango types to their New World colonies through the Pacifi c trading ports of Mexico and Panama. The most important, traditional mango cultivar in Mexico remains the ‘Manila’, refl ecting its Philippine origin. ‘Carabao’ and ‘Manila’ are probably identical. The mango was introduced to the West Indies in the mid- to late 18th century, probably from Brazil. The fi rst introductions of mango into Florida (USA) occurred in 1861, and involved the ‘No. 11’ polyembryonic seedling from Cuba. Seven years later, another polyembry-onic selection, ‘Peach’ was introduced into the state (Knight and Schnell, 1993). Many of the early introductions into Florida proved to be unproduc-tive, although ‘Mulgoba’ was planted on a small commercial scale (this culti-var is referred to as ‘Mulgoa’ in India, ‘Mulgoba’ in the USA and ‘Malgoa’ in Malaysia).

Introduction: Botany and Importance 11
Secondary ‘centres of diversity’In 1910, a seedling of ‘Mulgoba’ came into production in Florida. Its fruit had a highly attractive red blush, and appeared to bear more heavily than its parent(s) (Wolfe, 1962). This selection was named ‘Haden’. Although ‘Haden’ was not superior with respect to fruit quality in comparison to the imported germplasm from India, its genetic base was much wider. During the 20th century, more introductions of mango germplasm into Florida occurred from South-east Asia (the Philippines, Cambodia), India and elsewhere. It was at one time believed that these introductions of mango germplasm created a secondary centre of diversity of the species (Knight and Schnell, 1993). ‘Eldon’, ‘Glenn’, ‘Lippens’, ‘Osteen’, ‘Parvin’, ‘Smith’, ‘Springfels’, ‘Tommy Atkins’ and ‘Zill’ are progeny of ‘Haden’. ‘Saigon’ seedlings were selections made from ‘Cambodiana’, a polyembryonic introduction from Indochina. From ‘Saigon’ seedlings, ‘Alice’, ‘Herman’ and ‘Florigon’ were selected. Based upon more recent genetic analysis involving microsatellite markers, it is now estimated that the majority of Florida cultivars are descended from only four monoembryonic Indian mango cultivar accessions, i.e., ‘Mulgoba’, ‘Sandersha’, ‘Amini’ and ‘Bombay’, together with the polyembryonic ‘Tur-pentine’ from the West Indies (Schnell et al., 2006). The Florida mango culti-vars have been found to be highly adaptable to many agroecological areas and bear regularly, whereas many of the outstanding Indian cultivars have been unproductive outside their centre of domestication, and are alternate bearing. These selections also have a highly attractive red blush at maturity, fi rm fl esh, a high fl esh to seed ratio and a regular bearing habit. Some of the Florida cultivars, for example ‘Tommy Atkins’, ‘Keitt’, etc. are also moder-ately resistant to anthracnose, the most important production and posthar-vest problem of mango in many areas. In the latter half of the 20th century, plantings of Florida cultivars have been established in many countries and now form the basis of international trade of mangoes.
Current distributionThe mango is cultivated commercially throughout the tropics and in many subtropical areas. It is grown at the equator and at a latitude of 35–37 in southern Spain. According to Knight and Schnell (1993), ‘The process that began in Florida – introduction of superior germplasm from abroad followed by selection of improved cultivars adapted to local conditions – is now underway in many areas.’
1.4 Germplasm Conservation
Genetic erosion
The Mangifera species have their centre of diversity and origin in South-east Asia, a region that has experienced great economic development in recent years. Vast wooded areas have been completely or partially deforested either for expanding agriculture or for removal of tropical hardwoods for export.

S.K. Mukherjee and R.E. Litz12
This has caused great genetic erosion within many species and genera. The Mangifera species, like many other tropical fruit trees, are canopy and emer-gent trees of the tropical rainforest (Kaur et al., 1980). These trees are widely scattered in the tropical rainforest, fl ower erratically and reproduce from large seeds that deteriorate rapidly. As such, they are particularly vulnerable and in danger of extinction.
Collection and documentation of Mangifera germplasm
The International Plant Genetic Resources Institute (IPGRI), formerly known as the International Board for Plant Genetic Resources (IBPGR), commis-sioned an ecogeographical study of known Mangifera genetic resources (Muk-herjee, 1985). Based upon this documentation, a joint IBPGR-International Union for the Conservation of Nature (IUCN)-World Wildlife Fund (WWF) project was initiated to collect wild mangoes on the island of Borneo and in the Malay Peninsula (Bompard, 1989), the regions that held the highest con-centrations of Mangifera species. Kostermans and Bompard (1993), in the lat-est revision of the taxonomy of Mangifera, recognized 69 species, many of which were collected during the course of this project (Table 1.1). Because of the loss of natural habitat, the establishment of in situ and ex situ germplasm collections of Mangifera species was considered to be imperative.
Relevance of germplasm resources to mango improvement
The genetic improvement of mango hitherto has depended on the utilization of the genetic variability found within a single species, M. indica. According to Mukherjee (1985), ‘A concerted sampling strategy should be devised for exsitu samples to meet urgent needs for use in research for improvement of the crop through breeding or as rootstocks. Sources of resistance to mango mal-formation, anthracnose, powdery mildew, gall midge are urgently needed.’
1.5 Importance of Mango
Cultivars
A partial list of the principal mango cultivars has been provided in Table 1.2. This list includes many cultivars that were identifi ed in a survey of world mango production compiled by Watson and Winston (1984). The distribution of mango cultivars outside their centres of domestication can be attributed primarily to three historical events: (i) the movement of Indian varieties (monoembryonic) along the trade routes of the Portuguese to Africa and South America; (ii) the spread of South-east Asian varieties (polyembryonic) across the Pacifi c Ocean to Central and South America by the Spaniards; and (iii) the identifi cation of improved mango cultivars initially in Florida and

Introduction: Botany and Importance 13
Table 1.2. Most important mango cultivars in major producing countries.
Continent Country Cultivars
Africa Cote d’Ivoire ‘Amelie’, ‘Kent’Egypt ‘Alphonso’, ‘Bullock’s Heart’, ‘Hindi be Sennara’,
‘Langra’, ‘Mabrouka’, ‘Pairie’, ‘Taimour’, ‘Zebda’Kenya ‘Boubo’, ‘Ngowe’, ‘Batawi’Mali ‘Amelie’, ‘Kent’South Africa ‘Fascell’, ‘Haden’, ‘Keitt’, ‘Kent’, ‘Sensation’, ‘Tommy
Atkins’, ‘Zill’Asia Bangladesh ‘Aswina’, ‘Fazli’, ‘Gopal Bhog’, ‘Himsagar’, ‘Khirsapati’,
‘Langra’China ‘Gui Fei’, ‘Tainong No. 1’, ‘Keitt’, ‘Sensation,’ ‘Zill’, ‘Zihua’,
‘Jin Huang’India ‘Alphonso’, ‘Banganapalli’, ‘Bombay’, ‘Bombay Green’,
‘Chausa’, ‘Dashehari’, ‘Fazli’, ‘Fernandian’, ‘Himsagar’, ‘Kesar’, ‘Kishen Bhog’, ‘Langra’, ‘Mallika’, ‘Mankurad’, ‘Mulgoa’, ‘Neelum’, ‘Pairi’, ‘Samar Behisht’, ‘Suvarnarekha’, ‘Totapuri’, ‘Vanraj’, ‘Zardalu’
Indonesia ‘Arumanis’, ‘Dodol’, ‘Gedong’, ‘Golek’, 'Madu’, 'Manalagi’Israel ‘Haden’, ‘Tommy Atkins’, ‘Keitt’, ‘Maya’, ‘Nimrod’, ‘Kent’,
‘Palmer’Malaysia ‘Apple Rumani’, ‘Arumanis’, ‘Golek’, ‘Kuala Selangor 2’,
‘Malgoa’Myanmar ‘Aug Din’, ‘Ma Chit Su’, ‘Sein Ta Lone’, 'Shwe Hin Tha’Pakistan ‘Anwar Ratol’, ‘Began Pali’, ‘Chausa’, ‘Dashehari’,
‘Gulab Khas’, ‘Langra’, ‘Siroli’, ‘Sindhri’, ‘Suvarnarekha’, ‘Zafran’
The Philippines ‘Carabao’, ‘Manila Super’, ‘Pico’Taiwan ‘Irwin’, ‘Jin-hwung’, ‘Keitt’, ‘Tommy Atkins’, ‘Tainong
No. 1’, ‘Tsar-swain’Thailand ‘Nam Doc Mai’, ‘Ngar Charn’, ‘Ok Rong’, ‘Keow Savoey’,
‘Pimsen Mum’Australia ‘Calypso’, ‘Kensington Pride’North and
Central America
Costa Rica ‘Haden’, ‘Irwin’, ‘Keitt’, ‘Mora’, ‘Tommy Atkins’Dominican Republic
‘Haden’, ‘Keitt’, ‘Kent’, ‘Tommy Atkins’
Guatemala ‘Haden’, ‘Keitt’, ‘Kent’, ‘Tommy Atkins’Haiti ‘Francine’, ‘Madame Francis’Mexico ‘Ataulfo’, ‘Haden’, ‘Keitt’, ‘Kent’, ‘Manila’, ‘Palmer’,
‘Sensation’, ‘Tommy Atkins’, ‘Van Dyke’USA ‘Keitt’, ‘Kent’, ‘Tommy Atkins’
SouthAmerica
Brazil ‘Bourbon’, ‘Coite’, ‘Coquinho’, ‘Coracao’, ‘Espada’, ‘Haden’, ‘Itamaraca’, ‘Keitt’, ‘Mamao’, ‘Palmer’, ‘Rosa’, ‘Tommy Atkins’, ‘Uba’, ‘Van Dyke’
Colombia ‘Vallenato’Ecuador ‘Haden’, ‘Keitt’, ‘Kent’, ‘Tommy Atkins’Peru ‘Haden’, ‘Keitt’, ‘Kent’, ‘Tommy Atkins’Venezuela ‘Haden’, ‘Keitt’, ‘Kent’, ‘Tommy Atkins’

S.K. Mukherjee and R.E. Litz14
later in other new mango-producing areas, as a result of open and controlled pollination among local and introduced mango germplasm from India and South-east Asia.
Further information about many of the mango cultivars, including their fruit characters, is available in Knight et al. (Chapter 3, this volume), and in publications by Burns and Prayag (1921) for mangoes of Maharashtra, Naik and Gangolly (1950) for south Indian mangoes, Singh and Singh (1956) for Uttar Pradesh mangoes, Mukherjee (1948) for Bengal mangoes and Camp-bell (1992) for Florida mangoes.
Because many clonally propagated mango cultivars have unique local and/or regional names, there is considerable confusion in nomenclature. The Indian Agricultural Research Institute (IARI), New Delhi, has been recog-nized by the International Society for Horticultural Science (ISHS) as the International Registration Authority for Mango, whose mission is to consoli-date superfl uous names of mango cultivars. The potential for molecular, for example randomly amplifi ed polymorphic DNA (RAPD), markers, to resolve much of this confusion has been demonstrated by Schnell and Knight (1992), Degani et al. (1993), Schnell et al. (1995), Eiadthong et al. (1999a), Kashkush et al. (2001) and Gonzalez et al. (2002) (see Bompard, Chapter 2 and Iyer and Schnell, Chapter 4, this volume).
There is little variation among seedlings derived from polyembryonic mangoes. None the less, a certain amount of variability does occur, probably as a result of somatic mutation. Thus, in Indonesia there are several ‘Aru-manis’ selections that are denoted numerically, for example ‘Arumanis 1’, ‘Arumanis 2’, etc. In addition, although Philippine mango cultivars are dis-tinguished by different names, for example ‘Carabao’, ‘Manila’, ‘Philippine’, etc., the differences among them are quite subtle.
1.6 Production and Uses
The mango is the most important fruit of Asia, and currently ranks fi fth in total production (in metric tonnes) among major fruit crops worldwide, after Musa (bananas and plantains) (105,815,354 t), Citrus (all types) (105,440,168 t), grapes (65,584,233 t) and apples (59,444,377 t) (FAOSTAT, 2006). According to the Food and Agriculture Organization of the United Nations (FAO) database (FAOSTAT, 2006), world mango production has increased from 16,903,407 t in 1990 to 28,221,510 t in 2005. Much of this new production has occurred outside the traditional centres of mango culture of South and South-east Asia. In 1990, India produced approximately 51% of the world’s mangoes, but by 2005, India’s share had declined to approximately 38%, despite the substantial increase in mango production since 1990 (from 8,645,405 to 10,800,000 t between 1990 and 2005). The current leading producing nations after India include (in metric tonnes) China (3,450,000), Thailand (1,800,000), Pakistan (1,673,900), Mexico (1,600,000), Indonesia (1,478,204), Brazil (1,000,000) and the Philippines (950,000). Although world production has increased by 67% between 1990 and 2005, mango exports have increased almost sixfold

Introduction: Botany and Importance 15
from 158,030 to 907,782 t, with total export value estimated to be US$583,763,000 (FAOSTAT, 2006). The major exporting countries are (in met-ric tonnes) Mexico (212,505), India (156,222) and Brazil (111,181). As a result, mangoes are widely available as fresh fruit and as processed products (i.e. dried fruit, dairy products, juice, pickles, etc.).
Mangoes are an important component of the diet in many less developed countries in the subtropics and tropics. In regions of the world that have experienced low living standards and serious nutritional defi ciencies, their attractiveness and fl avour have also enhanced the quality of life. Surplus production has increasingly been processed and fruit of certain cultivars is destined for export as fresh fruit. Approximately 1% of mango production is utilized for processing for juice, nectars, preserves (including chutney), fruit leather, dried fruit slices, frozen pulp and as a fl avouring for baked goods, ice cream, yoghurt, etc. (see Raymundo et al., Chapter 17, this volume). No part of the fruit is wasted. In India and the subcontinent, the seed is used for extraction of starch ‘amchur’, and the peels (skin) have been used as a source of anacardic acid. Mango wood is a low quality timber, and the bark of the tree is an important source of tannins for curing leather.
References
Akamine, E.K. and Goo, T. (1973) Respiration and ethylene production during ontogeny of fruit. Journal of the American Society for Horticultural Science 98, 381–383.
Angeles, D.E. (1992) Mangifera altissima Blanco. In: Verheij, E.W.M. and Coronel, R.E. (eds) Plant Resources of South-east Asia No.2: Edible Fruits and Nuts. Pudoc-DLO, Wageningen, the Netherlands, pp. 206–207.
Anonymous (1992) Annual Report. Central Institute of Horticulture for Northern Plains, Lucknow, India.
Aron, Y., Czosnek, H., Gazit, S. and Degani, C. (1998) Polyembryony in mango (Mangifera indica L.) is controlled by a single dominant gene. HortScience 33, 1241–1242.
Blume, C.L. (1850) Museum Botanicum Lugduno – Batavum 1, 190–193.Bompard, J.M. (1989) Wild Mangifera species in Kalimantan (Indonesia) and in Malay-
sia. Final Report. International Board for Plant Genetic Resources, Rome.Bompard, J.M. (1992a) Mangifera caesia Jack. and Mangifera kemanga Blume. In: Ver-
heij, E.W.M. and Coronel, R.E. (eds) Plant Resources of South-east Asia No.2: Edible Fruits and Nuts. Pudoc-DLO, Wageningen, the Netherlands, pp. 207–209.
Bompard, J.M. (1992b) Mangifera foetida Lour. and Mangifera pajang Kostermans. In: Verheij, E.W.M. and Coronel, R.E. (eds) Plant Resources of South-east Asia No.2: Edible Fruits and Nuts. Pudoc-DLO, Wageningen, the Netherlands, pp. 209–211.
Bompard, J.M. (1992c) Mangifera laurina Blume and Mangifera pentandra Hooker f. In: Verheij, E.W.M. and Coronel, R.E. (eds) Plant Resources of South-east Asia No.2: Edible Fruits and Nuts. Pudoc-DLO, Wageningen, the Netherlands, pp. 216–218.
Bompard, J.M. (1992d) Mangifera odorata Griffi th. In: Verheij, E.W.M. and Coronel, R.E. (eds) Plant Resources of South-east Asia No.2: Edible Fruits and Nuts. Pudoc-DLO, Wageningen, the Netherlands, pp. 218–221.
Bompard, J.M. (1993) The genus Mangifera re-discovered: the potential of wild species to mango cultivation. Acta Horticulturae 341, 69–77.

S.K. Mukherjee and R.E. Litz16
Burns, W. and Prayag, S.M. (1921) The Book of Mango. Bulletin 103, Department of Agriculture, Bombay, India.
Campbell, R.J. (1992) A Guide to Mangoes in Florida. Fairchild Tropical Garden, Miami, Florida.
De Candolle, A.P. (1884) Origin of Cultivated Plants. Hafner, London.Degani, C., Cohen, M., Reuveni, O., El-Batsri, R. and Gazit, S. (1993) Frequency and
characteristics of zygotic seedlings from polyembryonic mango cultivars, deter-mined using isozymes as genetic markers. Acta Horticulturae 341, 78–85.
Eiadthong, W., Yonemoni, K., Kanzaki, S., Sugiura, A., Utsunomiya, N. and Subhadrab-andhu, S. (1999a) Amplifi ed fragment length polymorphism (AFLP) analysis for studying the genetic relationships among Mangifera species in Thailand. Journal of the American Society for Horticultural Science 125, 160–164.
Eiadthong, W., Yonemoni, K., Sugiura, A., Utsunomiya, N. and Subhadrabandhu, S. (1999b) Analysis of phylogenetic relationship in Mangifera by restriction site analy-sis of an amplifi ed region of cpDNA. Scientia Horticulturae 80, 145–155.
Fang, T.T. (1965) Chromatographic fractionation of nonnitrogenous organic acids of mango and guava fruits by silica gel column. Memoirs of the College of Agriculture, National Taiwan Museum 8, 236.
FAOSTAT (2006) Available at: http://faostat.fao.org/site/340/default.aspx (accessed 22 October 2006).
Gonzalez, A., Coulson, M. and Brettell, R. (2002) Development of DNA markers (ISSRs) in mango. Acta Horticulturae 575, 139–143.
Gruezo, W.S. (1992) Mangifera L. In: Verheij, E.W.M. and Coronel, R.E. (eds) Plant Re-sources of South-east Asia No.2: Edible Fruits and Nuts. Pudoc-DLO, Wageningen, the Netherlands, pp. 203–206.
Hooker, J.D. (1876) The Flora of British India 2. Reeve, London.Jain, N.L., Krishnamurthy, G.V. and Lal, G. (1959) Nonvolatile organic acids in unripe
pickling mangoes and salted mango slices by paper chromatography. Food Science8, 115–117.
Juliano, J.B. (1934) Origin of embryos in the strawberry mango. Philippine Journal of Science 54, 553–563.
Juliano, J.B. and Cuevas, N.L. (1932) Floral morphology of the mango (Mangifera indicaL.) with special reference to the Pico variety from the Philippines. Philippine Agri-culturist 21, 449–472.
Kashkush, K., Jinggui, F., Tomer, E., Hillel, J. and Lavi, U. (2001) Cultivar identifi cation and genetic map of mango (Mangifera indica). Euphytica 122, 129–136.
Kaur, A., Ha, C.O., Jong, K., Sands, V.E., Chan, H.T., Soepadmo, E. and Ashton, P.S. (1980) Apomixis may be widespread among trees of the climax rain forest. Nature271, 440–442.
Knight, R.J., Jr and Schnell, R.A. (1993) Mango (Mangifera indica L.) introduction and evaluation in Florida and its impact on the world industry. Acta Horticulturae 341, 125–135.
Kostermans, A.J.G.H. and Bompard, J.M. (1993) The Mangoes: Botany, Nomenclature, Horticulture, Cultivation and Utilization. Academic Press, London.
Krishnamurthy, S. and Subramanyam, H. (1970) Respiratory climacteric and chemical changes in the mango fruit Mangifera indica L. Journal of the American Society for Horticultural Science 95, 333–337.
Kumar, N.V.H., Narayanaswamy, P., Prasod, D.T., Mukunda, G.K. and Sondhu, S.N. (2001) Estimation of genetic diversity of commercial mango (Mangifera indica L.)cultivars using RAPD markers. Journal of Horticultural Science and Biotechnology 76, 529–533.

Introduction: Botany and Importance 17
Kumar, S., Das, D.K., Singh, A.K. and Prasad, U.S. (1994) Sucrose metabolism during maturation and ripening of mango cultivars. Plant Physiology and Biochemistry 21, 27–32.
Lakshminarayana, S. (1980) Mango. In: Nagy, S. and Shaw, P.E. (eds) Tropical and Subtropical Fruits. AVI Publishing Co., Westport, Connecticut, USA, pp. 184–257.
Lizada, M.C. (1991) Post harvest physiology of mango – a review. Acta Horticulturae 291, 437–449.
Maheshwari, P., Sachar, R.C. and Chopra, R.N. (1955) Embryological studies in mango (Mangifera indica L.). In: Proceedings of the 42nd Indian Scientifi c Congress, Baro-da, India, p. 233.
Medlicott, A.P. and Thompson, A.K. (1985) Analysis of sugars and organic acids in ripening mango fruit (Mangifera indica L. var. Keitt) by high performance liquid chromatography. Journal of the Science of Food and Agriculture 36, 56–566.
Medlicott, A.P., Bhogol, M. and Reynolds, S.B. (1986) Changes in peel pigmentation during ripening of mango fruit (Mangifera indica var. Tommy Atkins). Annals of Applied Biology 109, 651–656.
Miquel, F.A.G. (1859) Flora van Nederlandsch Indie 1, 627–634.Mitra, S.K. and Baldwin, E.A. (1997) Mango. In: Mitra, S.K. (ed.) Postharvest Physiology
and Storage of Tropical and Subtropical Fruits. CAB International, Wallingford, UK, pp. 85–122.
Mukherjee, S.K. (1948) The varieties of mango (M. indica L.) and their classifi cation. Bulletin of the Botanical Society of Bengal 2, 101–133.
Mukherjee, S.K. (1950a) Wild mangoes of India. Science and Culture 15, 469–471.Mukherjee, S.K. (1950b) Mango. Its allopolyploid nature. Nature 150, 196–197.Mukherjee, S.K. (1951a) Pollen analysis in Mangifera in relation to fruit set and taxonomy.
Journal of the Indian Botanical Society 30, 49–55.Mukherjee, S.K. (1951b) Origin of mango. Indian Journal of Genetics and Plant Breeding
11, 49–56.Mukherjee, S.K. (1953) Origin, distribution and phylogenetic affi nities of the species of
Mangifera L. Journal of the Linnean Society, Botany 55, 65–83.Naik, K.C. and Gangolly, S.R. (1950) A Monograph on Classifi cation and Nomenclature
of South Indian Mangoes. Government Press, Madras, India.Ochse, J.J. (1931) Fruits and Fruiticulture in the Dutch East Indies. G. Kolff, Batavia,
(Jakarta), Indonesia.Parisot, E. (1988) Etude de la croissance rhythmique chez de jeunes manguiers (Mangifera
indica L.). Description, germination et conservation de graines polyembryonnees de manguier. Fruits 43, 97–105.
Perry, E.O.V. and Zilva, S.S. (1932) Preliminary Report on Vitamin Content of the Mango.Empire Marketing Board, London.
Popenoe, W. (1932) Manual of Tropical and Subtropical Fruits. Macmillan Co., New York.
Purseglove, J.W. (1972) Mangoes west of India. Acta Horticulturae 24, 107–174.Rumphius, G.E. (1741–1750) Herbarium Amboinense. Vol. 1–6. Den Haag, Amsterdam.Salunkhe, D.K. and Desai, B.B. (1984) Mango. In: Postharvest Biotechnology of Fruits,
Vol. 1. CRC Press, Boca Raton, Florida, pp. 77–94.Sarker, S. and Muhsi, A.A. (1981) A study on the content and interconversions of or-
ganic acids of mango (Mangifera indica L.) at various stages of fruit development. Bangladesh Journal of Agricultural Science 8, 69–75.
Schnell, R.J. and Knight, R.J., Jr (1992) Frequency of zygotic seedlings from fi ve polyem-bryonic mango rootstocks. HortScience 27, 174–176.

S.K. Mukherjee and R.E. Litz18
Schnell, R.J., Ronning, C.M. and Knight, R.J., Jr (1995) Identifi cation of cultivars and validation of genetic relationships in Mangifera indica L. using RAPD markers. Theoretical and Applied Genetics 90, 269–274.
Schnell, R.J., Brown, J.S., Olano, C.T., Meerow, A.W., Campbell, R.J. and Kuhn, D.N. (2006) Mango genetic diversity analysis and pedigree inferences for Florida culti-vars using microsatellite markers. Journal of the American Society for Horticultural Science 13, 214–224.
Selvaraj, Y., Kumar, R. and Pal, D.K. (1989) Changes in sugars, organics, amino acids, lipids, lipid constituents and aroma characteristics or ripening mango (Mangiferaindica L.) fruit. Journal of Food Science and Technology 26, 306–311.
Seward, A.C. (1912) Dictyledonous leaves from Assam. Records of the Geological Sur-vey of India 42, 100.
Singh, L.B. and Singh, R.N. (1956) A Monograph on the Mangoes of UP. Superintendent of Printing, Uttar Pradesh Government, Lucknow, India.
Sturrock, T.T. (1968) Genetics of mango polyembryony. Proceedings of the Florida State Horticultural Society 81, 311–314.
Watson, B.J. and Winston, E.C. (1984) Plant genetic improvement. In: Proceedings of the First Australian Mango Research Workshop. Commonwealth Scientifi c and Indus-trial Research Organization (CSIRO), Canberra, pp. 104–138.
Wilson, C.W., Shaw, P.E. and Knight, R.J., Jr (1990) Importance of some lactones and 2,5-dimethyl-4-hydroxy-3-(2H)-furanone to mango (Mangifera indica L.) aroma. Journal of Agricultural Food Chemistry 38, 1556–1559.
Wolfe, H.S. (1962) The mango in Florida – 1887 to 1962. Proceedings of the Florida State Horticultural Society 75, 387–391.

© CAB International 2009. The Mango, 2nd Edition: Botany, Production and Uses(ed. R.E. Litz) 19
2 Taxonomy and Systematics
J.M. BompardLes Mazes, Montaud, France
2.1 Introduction 192.2 The Genus Mangifera L. 20
Distribution 20Ecology and habitat 20
2.3 Taxonomy and Systematics 22Taxonomic history 22
2.4 Phytogeography 28Species distribution 28Subgenera and section distribution 29
2.5 Interspecifi c Molecular Characterization 302.6 Region of Origin of the Genus 312.7 Origin of the Common Mango 32
The common mango in South-east Asia 322.8 Conclusion 35
Potential contribution of wild species to mango cultivation 35Source of rootstock 35Hybridization 36Potential of wild species 36
2.1 Introduction
The genus Mangifera is one of the 73 genera (c.850 species) belonging to the family of Anacardiaceae, in the order of Sapindales. Anacardiaceae is a fam-ily of mainly tropical species, with a few representatives in temperate regions. Malesia, which is the phytogeographic region extending from the Malay Peninsula south of the Kangar-Pattani line to the Bismarck Archipelago east of New Guinea (Whitmore, 1975) contains more species in the Anacardiaceae than any other area. Within Malesia occurrence is mainly in Western Malesia (Ding Hou, 1978b).

J.M. Bompard20
Apart from mango, several other cultivated fruit trees belong to the fam-ily, for example the ambarella or Otaheite apple (Spondias dulcis Forst.) prob-ably from Melanesia, and the yellow and purple mombins (Spondias mombin L. and S. purpurea L., respectively) from tropical America, the Bouea species from IndoMalesia, dragon plums (Dracontomelum spp.) from IndoMalesia and the Pacifi c region, kaffi r plum (Harpephyllum caffrum Bernh. ex K. Krause) and the marula plum (Sclerocarya caffra Sond.) of southern Africa. The cashew (Anacardium occidentale L.) is from tropical America and the pistachio (Pistaciavera L.) from Iran and Central Asia. Anacardiaceous species also yield other valuable products: wood (several genera), gums and resins (Pistacia spp.), varnishes (Rhus spp. and Melanorrhoea spp., ‘lacquer trees’) and tanning materials (Rhus spp. and Schinopsis spp.). It is also a family well known for the dermal irritation produced by some of its members, such as the poison ivies and oaks (Rhus spp.) in North America, rengas (Gluta spp.) in South-east Asia and other species including some Mangifera species whose resinous sap may induce a mild to strong allergic reaction.
2.2 The Genus Mangifera L.
Distribution
The range of natural distribution of the 69 Mangifera species is mainly restricted to tropical Asia, and extends as far north as 27° latitude and as far east as the Carolines Islands. Wild mangoes occur in India, Sri Lanka, Ban-gladesh, Myanmar, Sikkim, Thailand, Kampuchea, Vietnam, Laos, southern China, Malaysia, Singapore, Indonesia, Brunei, the Philippines, Papua New Guinea and the Solomon and Carolines Islands. The highest species diver-sity, c.29 species, occurs in western Malesia, especially in peninsular Malay-sia and in Borneo and Sumatra, which represent the heart of the distribution range of the genus (Fig. 2.1).
Ecology and habitat
The majority of Mangifera species occur as a rule as scattered individuals in tropical lowland rainforests on well-drained soils. The species are distrib-uted mostly below 300 m, but can occur up to c.1000 m above sea level, on well-drained soils (44 species), in periodically inundated areas (ten species) and in certain types of swamp forest (i.e. M. gedebe, M. griffi thii and M. parvi-folia). Three species are mainly found in sub-montane forests above 1000 m and occasionally up to 1700 m above sea level (M. bompardii, M. dongnaiensis and M. orophila). There are also species that are adapted to seasonally dry climates in deciduous or semi-deciduous forests (e.g. M. caloneura, M. collina, M. timorensis and M. zeylanica). A few species occur north of the Tropic of Cancer, for example M. austro-yunnanensis and M. persiciformis in China, M.

Taxonomy and Systematics 21
sylvatica Roxb. in Sikkim and southern China, at altitudes of 600–1900 m above sea level; apparently wild M. indica can also be found outside the tropics.
Wild mangoes are large trees, 30–40 m (occasionally 54 m) in height, with tall columnar boles. Several species are exploited for their timber. The major-ity of wild mangoes occur as scattered individuals at very low densities in lowland forests on well-drained soils. Some of these are very rare; there are normally one to three trees above 40 cm in diameter/10 ha. Only a few spe-cies (M. gedebe, M. griffi thii and M. parvifolia) are gregarious in certain types of swamp forest. Most species are evergreen although a few are deciduous in the rainforests following a dry period, and stand bare for a short time before fl ushes of new leaves appear. A deciduous habit that is not linked to a sea-sonal climate also occurs in other genera of Anacardiaceae (Ding Hou, 1978b).
In the rainforest of western Malesia, Mangifera species fl ower and fruit very irregularly. As with many other genera in the region, mast or general fruiting at intervals of 3–8 years is the dominant pattern. In mast years, the ground beneath the trees can be covered with mangoes, whose strong smell attracts many animals. Isolated fl owering may occur after a dry period and is generally followed by a poor fruit crop. The occurrence of fl owering of a few species, for example the ‘lanjut’ (M. lagenifera) is only once every 5–10 years. There seem to be clear reproductive barriers between species in the wild, although limited hybridization among cultivated species has been reported (see section 2.8, Conclusion, this chapter).
30° N
4
2
5
4
3
613
27
2728
95
7 4
3
5
2
20° N
10° N
0° N
10° N
30° N
20° N
10° N
0° N
10° N
80° 100 120 160
80° 100 120 160140
Fig. 2.1. Distribution of Mangifera species in the range of the genus. Numbers shown indicate the number of wild species in each area: Sri Lanka, India and Sikkim, Andaman and Nicobar Islands, Myanmar, Thailand, Indochina, China, peninsular Malaysia, Sumatra, Borneo, Java, Lesser Sunda Islands, Sulawesi, Moluccas, the Philippines, New Guinea, and Solomon Islands (the Caroline Islands not represented).

J.M. Bompard22
These widely scattered towering tree species, often with an inaccessible crown, are undercollected and poorly represented in herbarium collections (Bompard, 1995). Because of their irregular fl owering, the fl owers and fruits of a few species are still unknown. Collecting plant material is consequently very diffi cult, and plant explorations are still yielding new records or new species. Many species have been recently recorded for the fi rst time, even from peninsular Malaysia, a country that has already been rather well combed by botanists, having one of the highest collecting indices in the Malesian region. Other species still await to be discovered. Sadly some species of very limited range may already have been lost to posterity by deforestation.
Our very meagre knowledge of the wild mangoes is due to the fact that identifi cation at the species level from leaves only is often diffi cult because of intraspecifi c variation in vegetative characters. Moreover, many of the origi-nal species were based on very poor specimens. Consequently, frequent mis-identifi cation of herbarium material has resulted in much confusion, requiring a critical revision of all the specimens in these collections. It is not uncommon that the same species has been described from different places under differ-ent names. For instance, M. inocarpoides described by Merrill and Perry from New Guinea in 1941, M. camptosperma and M. reba (recorded by Pierre in South Vietnam in 1897) are now recognized to be a single species M. gedebe Miquel, a species initially named in 1861 from a specimen collected in Suma-tra. Mangifera longipes Griffi th is now treated as M. laurina Blume, because this name takes precedence as it was validly published 4 years earlier.
After thorough study of herbarium collections and fi eld collections, a number of species have been newly described. Sixty-nine species are now recorded, including 13 species of uncertain affi nities, in contrast with the 49 species recognized by Mukherjee (1949). As more collections are made, there will doubtless be further taxonomic adjustments made to the genus Mangifera.
2.3 Taxonomy and Systematics
Taxonomic history
Subdivision of the genusAn historical review of the subdivisions of the genus Mangifera shows that two major groups have been rather consistently recognized in taxonomic treatments. Hooker (1862) was the fi rst to recognize two sections based on the characters of the fl ower disc: section I with a disc broader than the ovary, and section II with a disc stalk-like or wanting. These sections were later named by Marchand (1869) Amba, an Indian name for the common mango, and Limus, a Sundanese name for M. foetida in West Java, respectively. He also added a section Manga for M. leschenaultii, which in fact belongs to the section Limus.
In his monograph of the Anacardiaceae, Engler (1883) maintained Hook-er’s sections, and subdivided group A (Hooker’s section I) into two groups,

Taxonomy and Systematics 23
one group with four or fi ve petals and the other group with four petals. He considered the following sequence of morphological characters to be impor-tant for classifi cation: (i) texture of the leaves; (ii) number of fertile stamens; (iii) prominence of veins; (iv) pilosity of infl orescences; and (v) leaf shape.
Pierre (1897) further divided the genus Mangifera into fi ve sections based on fl ower characters, i.e. number of stamens, the attachment of stamens to the disk, and the style. Two of these fi ve sections – namely section I Euan-therae, with a short thick fl ower disc and 4–12 fertile stamens, and section V Marchandora then consisting of M. camptosperma (currently considered a syn-onym of M. gedebe) are still maintained as they form clear-cut sections.
In his monograph, Mukherjee (1949) recognized two unnamed sections, conserving Hooker’s subdivision. Ding Hou (1978a) adopted the same method in his revision of the Malesian Anacardiaceae recognizing only Hooker’s two original sections and providing them with proper names and synonyms: section Mangifera (section I Hooker, section Amba Marchand, group A Engler, sections Euantherae and Marchandora Pierre) and section Limus (section II Hooker, sections Limus and Manga Marchand, group B Engler, and sections Eudiscus and Microdiscus Pierre).
Most recent classifi cation of the genusThe taxonomic classifi cation referred to herein follows that proposed by Kos-termans and Bompard (1993). This treatise includes the results of collections and surveys carried out between 1986 to 1998 in Borneo and peninsular Malaysia, which were initiated and sponsored by the International Institute for Genetic Resources (now Biodiversity International) and the World Wide Fund for Nature.1 It was published under the auspices of the International Board for Plant Genetic Resources (now International Plant Genetics Resources Institute) and the Linnean Society of London.
The most recent treatment of Mangifera refl ects the current status of what is still fragmentary knowledge. It can provide a basis for further studies involving all aspects of the wild relatives of mango, but particularly their potential in mango breeding. Determining phylogenetic affi nities based upon molecular markers could change our thinking about relationships among Mangifera species and among the cultivated forms of M. indica (see Interspe-cifi c Molecular Characterization section, this chapter).
The morphological characters used for identifi cation have been placed in the following sequence of importance:
Shape of the fl oral disc (see section Subdivision of the genus).1. Number of fertile stamens.2. Seed labyrinthine or not.3. Shape of secondary branches of the infl orescences: open or lax panicle, 4.
fl owers glomerulate or sub-glomerulate, the ramifi cations racemoid or spike like.
Pubescence of the infl orescence.5. Shape, number and attachment of the nerves (ridges or fi ngers) at the 6.
inner surface of the petals.

J.M. Bompard24
Shape and size of the petals.7. Flowers tetra- or pentamerous (not a very constant character and often 8.
overlapping).Reticulation of the leaves, especially of the lower surface.9. Shape of the leaf (only fully grown leaves of sterile branches can be used).10. Texture of the leaves.11. Deciduous or non-deciduous trees.12. Colour of the fl owers.13. Shape, colour and smoothness of the fruit.14. Number and size of the stone fi bres.15.
Kostermans (Kostermans and Bompard, 1993) raised the sections to the rank of subgenus, i.e. subgenus Limus (Marchand) Kosterm., having a disc narrower than the base of the ovary, stalk-like or even lacking and subgenus Mangifera (Ding Hou) Kosterm., having a disc broader than the base of the ovary, cushion-like, often divided in four or fi ve lobes.
SUBGENUS LIMUS (MARCHAND) KOSTERM. Mangifera species of the subgenus Limus are quite distinctive and show only remote affi nity with the common mango. This taxon is more primitive than the subgenus Mangifera and may be ances-tral to it, although the two subgenera may have originated from two different ancestors. The subgenus Limus consists of 11 species, which are native to the rainforests of western Malesia (peninsular Thailand, Malay Peninsula, Suma-tra, West Java and Borneo), with the exception of M. foetida, which extends to the east, possibly as far as New Guinea, and M. odorata which is only known in cultivation.
Kostermans divided the subgenus Limus into two sections: (i) section Deciduae for deciduous trees (i.e. M. caesia, M. kemanga, M. pajang, M. superba and possibly M. blommesteinii, M. decandra and M. lagenifera); and (ii) section Perennes for non-deciduous species (i.e. M. foetida, M. leschenaultii, M. macro-carpa and M. odorata) (Kostermans and Bompard, 1993). In deciduous trees, the bracts enclosing the buds leave a characteristic collar of dense, narrow scars, which persist on old twigs and are especially prominent in M. caesia and M. kemanga.
Mangifera lagenifera and M. decandra have ten stamens, fi ve of which are fertile. The other nine species have only one (and rarely two) fertile stamen(s) and two to four staminodes. The two species with fi ve fertile stamens (M.decandra and M. lagenifera) and M. superba, M. caesia, M. kemanga and M. blom-mesteinii, whose leaves are apically aggregated into rosettes at the end of mas-sive twigs are particularly distinctive. The fruits of these species are broadly ellipsoid or pear shaped, not compressed, and have dirty whitish or pinkish mesocarp and lanceolate, and fi brous, non-ligneous leathery endocarp.
Mangifera subsessilifolia shows some affi nity with M. lagenifera and M.blommesteinii; however, it has been placed among the species of uncertain taxonomic position due to a lack of complete study material. This is not a very rare species, but fl owering and fruiting seem to occur at intervals of, or > 5 years, similar in this respect to M. lagenifera, which can be found growing

Taxonomy and Systematics 25
in old orchards in peninsular Malaysia. The fl owers and fruits of M. sub-sessilifolia are still unknown.
Mangifera foetida, M. odorata, M. caesia and M. kemanga are widely cultivated in the humid lowlands of the Malay Peninsula, Sumatra, Borneo, Java and Bali. They have also been introduced elsewhere in South-east Asia; M. caesia, M. foe-tida and M. odorata are grown in the southern part of the Philippines, M. foetida is grown in Myanmar, and M. odorata is found in Indochina. They have been described in general reviews of tropical fruit (Ochse and Bakhuizen, 1931; Ochse et al., 1961; Molesworth, 1967; Verheij and Coronel, 1991).
Mangifera pajang, an endemic and commonly cultivated species in Bor-neo, is hardly known outside its native island. This deciduous tree has very stout twigs, with leaves more or less aggregate at the apices. The globose fruits, up to 20 cm in diameter, are the largest known fruits in the genus. The rough, potato-brown rind (0.5–1 cm thick) can be peeled off like that of a banana. Its bright, deep yellow, thick and fi brous fl esh is sweet with a dis-tinctive taste (Kostermans, 1965; Bompard, 1991a). In orchards in Borneo where M. foetida and M. pajang are both cultivated, forms with leaves and fruits having intermediate characters are occasionally found.
Mangifera caesia, M. foetida, M. pajang and especially M. odorata are impor-tant in tropical humid regions where the common mango cannot be grown satisfactorily. Mangifera pajang has potential as an ornamental tree, having brilliant rose-red blossoms (Philipps et al., 1982).
SUBGENUS MANGIFERA. The subgenus Mangifera contains most of the species (47), and is divided into four sections: (i) section Marchandora Pierre; (ii) section Euan-therae Pierre; (iii) section Rawa Kosterm.; and (iv) section Mangifera Ding Hou.
Section Marchandora Pierre. This section has only one species, M. gedebe Miquel (syn. M. camptosperma Pierre, M. inocarpoides Merr. and Perry, M. reba Pierre). The labyrinthine seed is unique to this species, wherein the inner integu-ments penetrate the cotyledons and form numerous irregular folds. The fl at, discus-like fruit has only a very thin mesocarp. Mangifera gedebe grows in inundated places along rivers or lakes. The seed fl oats in water and is dis-persed during periods of high water, and this may explain its wide distribu-tion, from Myanmar through Malesia to New Guinea and the Bougainville Island.
Section Euantherae Pierre. The three species in this section (M. caloneura Kurz (syn. M. duperreana Pierre), M. cochinchinensis Engler and M. pentandra Hook. f.) appear to be the most primitive among the species of the subgenus Mangifera. The fl owers are characterized by the presence of fi ve fertile sta-mens. The three species are mainly confi ned to Myanmar, Thailand, Indo-china and the north of the Malay Peninsula. The region is in the transition zone from the humid tropical rainforest to monsoon forest, and these species show an adaptation to low rainfall. Mangifera cochinchinensis, which occurs in south-eastern Thailand and in Vietnam, has small oblong fruits with a thin seed; the fruits are much relished by local people in southern Vietnam, although they are very acidic. Mangifera caloneura and M. pentandra are closely

J.M. Bompard26
related, and can be mistaken for M. indica. However, their leaves are more leathery, have a more conspicuously dense reticulation, and the panicles are much more hirsute than the common mango. Mangifera caloneura occurs from Myanmar through Thailand to Indochina, in lowland evergreen forests, as well as in semi-deciduous forests. It is cultivated for its acidic-sweet fruit, and has been planted along the streets of Vientiane and Ho Chi Minh City (Saigon). Mangifera pentandra, apparently native to the northern Malay Pen-insula close to the Kra isthmus transition zone, is found in old orchards, in scattered locations, especially in Kedah and possibly also in peninsular Thai-land. It is also grown in the Anambas Islands and in Sabah, where it might have been introduced in early times. It is a prolifi c bearer, with small man-goes, c.8 cm length, and ripening green or yellow. The pale orange, watery pulp has a sweet taste and few fi bres.
Section Rawa Kosterm. This group, consisting of nine species, is not well delimitated. Most species have thick twigs and rather coriaceous leaves seated on protruding pedestals. The small, hardly fl attened ovoid or ellip-soid fruits that are black or partly red at maturity in several species are also characteristic. ‘Rawa’ is the Malay word for marsh, indicating that these spe-cies usually are found in periodically or permanently inundated areas. The fi ve species that occur in west Malesia (M. gracilipes, M. griffi thii, M. micro-phylla, M. paludosa and M. parvifolia) grow primarily in the swamps of south peninsular Malaysia, in central coastal areas of east Sumatra and western Borneo, and occasionally in peripheral uplands. It has also been reported from the Andaman Islands and from Thailand (Sreekumar et al., 1996; Eiad-thong et al., 2000a).
Mangifera andamanica and M. nicobarica are endemics from the Andaman and Nicobar Islands, respectively. Mangifera merrillii is a rare species endemic to the Philippines and M. minutifolia is known solely from a single collection from southern Vietnam. Mangifera griffi thii and M. microphylla are the only cultivated species within section Rawa. The former species is considered to be representative of the section, and is cultivated along the eastern coast of peninsular Malaysia and in western Borneo, and rarely in Sumatra. The fruits are small (3–5 cm long) and oblong or ovoid; the skin is rose-red, turning purplish black at maturity. The rind is thin and easily removed from the orange-yellow pulp, which is juicy and pleasantly sweet. Different forms are recognized by local people, according to the size and taste of fruits. Mangiferamicrophylla is a related, but less well-known species, having thinner leaves and a rather similar fruit.
Section Mangifera Ding Hou. With more than 30 species, section Mangifera is by far the largest. The common mango and the related M. laurina belong here. Species within the section have the same distribution range as the genus. The section may be divided into three groups based on fl oral structure and organ number variation: (i) those having pentamerous fl owers; (ii) those having tetramerous fl owers; and (iii) an intermediate group of species hav-ing both pentamerous and tetramerous fl owers. Within these three groups, it is possible to distinguish species with either puberulous or glabrous panicles.

Taxonomy and Systematics 27
Only characteristics of representative species within each group, especially those found in cultivation, are described below.
Pentamerous fl owers (14 species): Three species, M. laurina, M. minor and M. sylvatica, show affi nity with the common mango. Mangifera laurina is a species of the lowland forests of Malesia, where it is also under cultivation in old orchards. It can be distinguished from the common mango by having lax and widely pyramidal, glabrous or sparingly puberulous panicles. The fl ow-ers are smaller and are not glomerulate; the petals have a different shape, texture and colour. The fruit resemble those of a small common mango, with orange-yellow pulp, which is almost liquid at maturity. It is generally con-sumed when unripe. Several forms are in cultivation; however these are now becoming rare. Mangifera laurina is well suited to the humid tropical lowlands, fruiting well in areas where the common mango cannot be grown satisfacto-rily; moreover, it appears to be highly resistant to anthracnose (Bompard, 1991b).
Mangifera minor occurs east of Wallace’s line, from Sulawesi to New Guinea (east Malesia) and to the Carolines Islands in the east. It is adapted to a wide range of ecological conditions, growing equally well in dry savannahs and in tropical rainforests up to 1300 m. The fruit is obliquely oblong, 5–10 cm long, much narrowed, the tip obtuse, with a distinct beak and sinus. It is found in cultivation, although the yellowish fruit pulp is acidic and scant. Mangifera sylvatica is found from Sikkim (up to 1200 m) to northern Myan-mar and Thailand, and apparently also in Yunnan up to 1900 m. The fruit is obliquely ovate, 8–10 cm long, much compressed distally forming a hook, has scanty whitish-yellow pulp which is almost fi breless. Other species are occasionally found in cultivation, for example M. rufocostata, which is esteemed by the Banjarese people of South Kalimantan for its very sour fruits that are used to prepare a spicy condiment with chilli.
Tetramerous fl owers (15 species): Mangifera altissima is apparently endemic to the Philippines, where it occurs mainly at low elevations in the forests from northern Luzon to Mindoro (Brown, 1950). The fruit is mango shaped, ovoid or ellipsoid, slightly compressed, up to 8 cm length, green or some-what yellow when ripe, with whitish, sweetish-acidic fl esh. It is commonly found in dooryards, and thrives in regions with distinct wet and dry seasons (Angeles, 1991).
Mangifera torquenda occurs wild in west Malesia, and is cultivated in south Sumatra and in Borneo, where it is common in the forests and orchards of eastern Kalimantan. The sub-globose fruit, c.7.5 cm long and 6.5 cm in diameter, is yellow-green with darker spots at maturity, and has a thin rind. The pale yellow pulp has a rather pleasant sweet-acid, slightly resinous taste and a light turpentine smell. Short fi bres are attached to the seed. It is closely related to M. longipetiolata.
Mangifera magnifi ca is a common species in the rainforests of western Malesia, occasionally cultivated in central Sumatra and in West Kalimantan, where it has a special importance in the myths of Land Dayak peoples. The fruit is ovoid oblong, up to 12 cm long, 10 cm in diameter, only slightly compressed, greyish green with brown spots. The pulp is whitish, soft at

J.M. Bompard28
maturity, sweetish acid. Sweeter forms are reported in central Kalimantan (J.J. Afriastini, personal communication). The stone is unique in the genus in that it lacks fi bres adhering to it.
Mangifera quadrifi da is found from peninsular Malaysia to the Moluccas. The fruit is ellipsoid-globose, 6–8 cm long, green covered with black dots turn-ing completely black at maturity, and has a pale yellow, sweet-acid pulp. Another form is recognized by its more coriaceous leaves, smaller fruits, c.4 cm long, having dark yellow pulp, purplish around the stone, and a sweet, palat-able taste, somewhat like prunes. Both forms are cultivated in old orchards.
Tetra- and pentamerous fl owers (four species, and also M. indica): Mangifera casturi is related to M. quadrifi da, from which it can be distinguished by leaf and fruit characters. It has never been collected in the wild, and is a favourite among the Banjarese people in south Kalimantan. The fruits are small, a little compressed and up to 6 cm in length, becoming completely black at matu-rity. The orange pulp is very sweet and palatable, and resembles ‘honey mango’ or ‘mangga madu’ grown in East Java. Although M. casturi bears heavily, it has a strong- to alternate-bearing habit. It is an excellent fruit for the humid tropical lowlands, and appears to be resistant to anthracnose. Sev-eral differently named forms exist; these have polyembryonic seeds. Mangiferarubropetala is also only known in cultivation, and may be a primitive race of M. indica.
SPECIES OF UNCERTAIN TAXONOMIC POSITION. There is a group of 11 disparate spe-cies of uncertain taxonomic position that cannot be placed with certainty due to the absence of adequate material. There are three species only known in China.
2.4 Phytogeography
Species distribution
An examination of the present distribution of the genus shows that the larg-est number of Mangifera species in either subgenera is found in western Malesia on the Sunda shelf. A decreasing number of species occurs towards the genus boundary east of Wallace’s line in east Malesia, and in its northern and western range of distribution. While peninsular Malaysia and the islands of Sumatra and Borneo have the highest diversity of species, the number of species becomes gradually lower in east Malesia, especially in the Lesser Sunda Islands, Moluccas and New Guinea. This is explained by the geologic and paleogeographic features of the Malesian region which spans two large partly submerged continental shelves, the Asiatic shelf (Sunda Shelf linking the Malay Peninsula with the islands of Sumatra, Java, Borneo and Palawan) and the Australasian shelf (Sahul Shelf linking the Aru islands and New Guinea with Australia). During the last glaciation period (c.22,500–11,000 bp) the shelves were regions of land uncovered by the lowering of sea level, and present day peninsular Malaysia, Sumatra and Borneo were connected by

Taxonomy and Systematics 29
land bridges during the late period of maximal sea lowering. During the cool periods of glacial maxima, the Malesian forest was reduced in extent, but there is no evidence that it was reduced to isolated island forests. The Sunda-land and Papuasian rainforest blocks are therefore comparable to refugia in terms of species richness and the high degree of endemism (Whitmore, 1981). Mangifera has undergone major species development in west Malesia, which has remained relatively stable over a long period of time. The current vegeta-tion of west Malesia probably differs very little from that at the end of the Tertiary (van Steenis, 1950). A lower number of Mangifera species is found in Java and the Philippines, regions less often connected with Asia during the Pleistocene.
Only three species occur in New Guinea, which is largely covered with rainforest. These include M. minor, M. mucronulata and the widely distributed M. gedebe. Mangifera foetida also occurs, but may have been introduced. Mangifera minor occurs from Celebes and the Philippines to the Solomon Islands; M. mucronulata is found in the Moluccas, New Guinea and the Solomon Islands. The distribution of these species suggests a late immigration of a Laurasian genus from Sundaland via the Philippines, Sulawesi and the Moluccas into New Guinea, which is supported by the geological history of the region. No Mangifera species have ever been recorded from northern Australia.
Very few species are found in peninsular India and Sikkim. From present-day distribution, there is little evidence of migration of species into the sub-continent of India after its collision with Eurasia in the middle Eocene (Audley-Charles et al., 1981). According to Mehrotra et al. (1998), fossil leaves described as Eomangiferophyllum damalgiriensis Mehr. from the Upper Palaeo-cene in north-eastern India are an analogue of the modern genus Mangifera.
Mangifera sylvatica occurs along the northern limit of the range of Mangifera, with more or less discontinuity, from Sikkim to northern Thailand and to the southern part of Yunnan, where it is reported in mountains up to 1900 m above sea level (Anonymous, 1980). The few species that grow in southern China are very poorly known: M. austro-yunnanensis from western Yunnan, M. persiciformis from south-eastern Yunnan and southern Guizhou at latitudes up to 26°N and M. hiemalis, the ‘winter mango’ from Guangxi near the northern border Vietnam. In the revised Flora of China (Min and Bar-fod, 2008), M. austro-yunnanensis is considered to be conspecifi c with M.indica, M. hiemalis is treated as a synonym of M. persiciformis, and M. laurina is recorded from the lowland forests of south Yunnan.
Subgenera and section distribution
The species distribution is especially meaningful when the ranges of the spe-cies of each subgenus and section are considered separately.
Subgenus LimusAll species of the subgenus Limus are restricted to the Malesian area (M. foetida and M. macrocarpa occurring in peninsular Thailand), whereas all the species

J.M. Bompard30
with fi ve fertile stamens, considered the most primitive condition, are con-fi ned to west Malesia (M. decandra in Sumatra and Borneo; M. lagenifera in the two latter areas and in peninsular Malaysia). Only M. caesia, M. foetida and closely related M. leschenaultii occur in east Malesia.
Subgenus MangiferaIn the subgenus Mangifera, M. gedebe is the only species belonging to the sec-tion Marchandora, and has the widest range within the genus, extending from Myanmar through Malesia to New Guinea and Bougainville Island. Section Euantherae is centred in the region from Myanmar to Vietnam. Mangiferapentandra is only known from peninsular Malaysia, the Anambas Islands and Borneo. Section Rawa is mainly in western Malaysia and shows notable diversifi cation in the swamps and peripheral uplands in the south of penin-sular Malaysia, east central Sumatra (notably the Riau province) and west Borneo. During the glacial period this area, termed the ‘Riouw pocket’ (Cor-ner, 1978), formed a vast plain connecting the Malay Peninsula, Sumatra and Borneo, and is believed to have been fi lled with swamps. Mangifera merrillii is an endemic of the Philippines, M. minutifolia is an endemic of Vietnam, M.andamanica and M. nicobarica are endemics of the Andamans and Nicobar Islands. None of the species of section Mangifera occurring in mainland South-east Asia, north of the isthmus of Kra, are found in eastern Malesia; however, it would be interesting to assess the genetic relatedness of M. syl-vatica and M. minor, and also M. laurina, which may prove to be phylogeneti-cally very closely related.
2.5 Interspecifi c Molecular Characterization
Molecular biology techniques now make it possible to assess genetic related-ness in a more precise way. Published data support some of the groupings based on anatomical characters (Kostermans and Bompard, 1993) but not entirely.
RAPD (random amplifi cation of polymorphic DNA) markers were fi rst used in mango by Schnell and Knight (1993) and Schnell et al. (1995). Nine Mangifera species were analysed and compared to the traditional taxonomic groupings. The unweighted pair group method of arithmetic averages (UPGMA) cluster analysis for the subgenus Limus was not supportive of the separation between sections Perennes and Deciduae, which, admittedly, has a weak taxonomic basis. It confi rmed the relatedness between M. foetida and M. pajang. The UPGMA cluster analysis of the subgenus Mangifera supported the current taxonomy based on fl ower morphology. It showed the related-ness between M. quadrifi da and M. torquenda (both placed in the group of species with tetramerous fl owers), but also with M. casturi, although the lat-ter species has tetra- and pentamerous fl owers. One of the most signifi cant results was the evidence for the existence of interspecifi c hybridization within the studied species of the section Mangifera (see also Yonemori et al., 2002).
Phylogenetic relationships among 14 Mangifera species of Thailand were analysed by comparing amplifi ed fragment length polymorphism (AFLP)

Taxonomy and Systematics 31
markers (Eiadthong et al., 2000b), and by comparing sequences of the inter-nal transcribed spacer (ITS) region of nuclear ribosomal DNA (nrDNA) (Yonemori et al., 2002). They demonstrated that the common mango was closely related to M. laurina, M. sylvatica and M. oblongifolia of subgenus Mangifera to which M. indica belongs. A close relationship between M. indica and M. sylvatica has been corroborated by Nishiyama et al. (2006), who com-pared signal intensity of genomic in situ hybridization (GISH) on somatic metaphase chromosomes of M. indica, using labelled DNA of eight wild Mangifera species.
Furthermore, Eiadthong et al. (2000b) and Yonemori et al. (2002) have demonstrated that M. odorata, M. foetida and M. macrocarpa (of subgenus Limus) were related to M. indica. It is not surprising in the case of M. odorata whose hybrid origin (M. foetida × M. indica) has now been established, but this calls into question the position of the section Perennes.
Results of molecular studies do not permit a comprehensive view of the phylogenetic relationships among the genera. So far, they are rather support-ive of the groupings based on phenotype within the subgenus Mangifera (notably sections Rawa and Euantherae), but not for the subgenus Limus which will need to be redescribed, and likely restricted to the group of species related to M. caesia (M. kemanga, M. lagenifera, M. superba and possibly M.decandra). More studies will be needed to infer phylogenetic relationships within the section Mangifera. Keeping in mind the frequent misidentifi cations in collections and botanic gardens, herbarium specimens of studied material must be deposited in the national herbaria so that its taxonomic position can be ascertained in case of doubt.
2.6 Region of Origin of the Genus
Based on morphological, phytogeographical and fossil evidence, Mukherjee (1953) argued that:
although the highest number of species of both sections is concentrated in the Malay Peninsula [19 were then recorded], the centre of origin of the genus cannot be restricted to that area alone, as both the phylogenetically older species, i.e. with pentacyclic fl owers (M. duperreana, now reduced to M.caloneura, and M. lagenifera), occur in Siam and Indochina, and the former is absent from Malay Peninsula.
He concluded that the genus had its origin somewhere in the Myanmar–Thailand–Indochina area or in the Malayan area. Careful identifi cation of the greatest part of herbarium materials available has allowed a more accurate delimitation of the distribution ranges of the Mangifera species, notably of the subgenus Limus, and has revealed, among other things, that M. lagenifera does not occur north of Kra isthmus contrary to Mukherjee’s assertions. Fur-thermore, the ten-stamen species, M. decandra, which was described by Ding Hou in 1972 and hence was unknown to Mukherjee, is confi ned to Borneo and Sumatra, and to date has not been recorded from peninsular Malaysia.

J.M. Bompard32
Without overemphasizing the present great species diversity of subgen-era in the Malay Peninsula, Borneo and Sumatra, the available evidence points to a Sundaic origin for the genus. This, however, must not minimize the particular importance of the region stretching from Myanmar to Indo-china as another centre of diversifi cation, as attested by the range of species belonging to the section Euantherae. Unfortunately, many of the species of this region remain poorly known, and it can be expected that plant collecting in the region will yield interesting new fi ndings. The speciation that occurred in this region with a likely radiation centre today traced by the range of the section Euantherae, is of special signifi cance as it has given rise to the com-mon mango.
2.7 Origin of the Common Mango
The common mango apparently originated in regions on the western border of the secondary centre of diversifi cation mentioned above. Truly wild mango trees have been recorded in Bangladesh (Chittagong Hill tract, c.23°N), north-eastern India (‘undoubtedly indigenous in the evergreen tracts of valley of Assam’ according to Kanjilal et al., 1937), and in Myanmar where it was reported as ‘not unfrequent in the tropical and lower mixed forests all over Burma from Arracan and Pegu down to Tenasserim’ (Kurz, 1877). It would be desirable to assess its affi nity with the species of the section Euantherae, as well as with species of other sections of the subgenus Mangifera that occur in the same area and region. It is also believed to be wild ‘in the sub-Himalayan tract, in deep gorges of the Baraitch and Gonda hills in Oudh, and the outer hills in Kamaon and Garhwal’ (Brandis, 1874). The common mango has been grown and disseminated for such a long time in India that semi-wild trees can be found in the forests throughout the subcontinent. The fruits of wild trees are said to be small and of poor quality. Watt (1891) mentioned two so-called ‘almost unaltered wild varieties’ existed under cultivation in Tirhoot, ‘one originating from Kangra, a very variable one, and the other from Sikkim which was evidently the progenitor of the varieties cultivated in Malda’.
The common mango in South-east Asia
The Linnean binomial (Mangifera indica) indicates in this instance the place where the common mango was selected and improved, and not necessarily its place of its origin. It has been traditionally accepted that mango was domesti-cated several millennia ago in India (see Mukherjee and Litz, Chapter 1, this volume); however, it cannot be excluded that domestication occurred inde-pendently in several areas, possibly in the south-western and south-eastern regions of its centre of origin, or later differentiated in those two regions. This hypothesis would account for the differences that exist between the local polyembryonic cultivars of Myanmar, Thailand, Indochina and Indonesia, and the monoembryonic Indian cultivars. Note that polyembryony occurs

Taxonomy and Systematics 33
also in the cultivated M. casturi, M. laurina and M. odorata. Aron et al. (1998) have demonstrated that polyembryony in mango is under the control of a single dominant gene.
According to Juliano (1937), Bijhouwer suggested that there were two main centres of domestication of mango, ‘one in India with monoembryonic mangoes, the other in the Saigon area, Indonesia and the Philippines with polyembryonic mangoes’. The ‘Saigon’ area must in fact be extended to southern Vietnam, other parts of Indochina, Thailand and Myanmar, which were recognized by Valmayor (1962) as homes of polyembryonic mangoes. Notwithstanding, the origin of polyembryonic mangoes is probably better placed in Myanmar, and possibly the eastern part of Assam. According to Brandis (1874), ‘in Burma, the mango is not generally grafted, and seeds of a good kind, as a rule, produce fruit of a similar description’. There are only a few polyembryonic mango cultivars in India. They are restricted to the south-western coastal region, and geographically isolated from the polyembryonic mangoes of Myanmar and South-east Asia. Analysis of genetic relatedness using RAPD markers among polyembryonic and monoembryonic cultivars grown in the west coast of southern India suggest that the polyembryonic types are unlikely to have originated from India and might have been intro-duced from South-east Asia (Ravishankar et al., 2004).
Indian Buddhist monks might have introduced the common polyembry-onic mango to South-east Asia, fi rst along land trade routes through Myan-mar, where they might have found better races, and from there into insular South-east Asia. It is well established that some local names of the common mango currently used in parts of Indonesia are of Sanskrit origin (‘ampelam’ and its cognates), and are sometimes used to designate M. laurina, which is a truly native species. Vernacular names do not always travel with a plant, and even if they did so in the case of the common mango, it is very unlikely that these introductions were the fi rst ones and that they came obligatorily from India. In the absence of a comprehensive classifi cation of the innumerable South-east Asian cultivated forms of the common and wild mangoes, includ-ing the countless primitive races, we have to rely on linguistics and the rich history and prehistory of this region.
Vernacular namesThe different local names of the common mango in Indonesia (‘pauh’, ‘ampe-lam’ and its variants, and ‘mangga’) bear evidence of a long history of con-tacts with mainland Asia and India, and point to possible introduction at different times from different places. In some parts of Indonesia, the vernacu-lar names ‘paoh’ or ‘pauh’ refer either to primitive races of the common mango, or to native species, as a rule the ones most closely resembling the common mango, for example: ‘pauh asal’ (= native mango) for M. pentandra in peninsular Malaysia; ‘pahohutan‘ or ‘pahutan’ (= forest mango) for M.altissima in the Philippines; and ‘pao pong’ (= forest mango) for M. minor in Flores, Lesser Sunda Islands. ‘Pau’ is a word belonging to Austronesian lan-guages, nowadays spoken over a very wide area from Madagascar to the Easter Islands by people who originate from mainland Asia. These languages

J.M. Bompard34
are still spoken by certain minority populations in Vietnam, Cambodia and the Mergui Archipelago off the coast of Myanmar (Bellwood et al., 1995). In Cambodia, which was occupied by the Chams from about the 3rd to the 15th century ad, ‘pa:uh’ is a Chamic word. ‘Sva:y’, used by the Khmers (as in ‘sva:y srok’ meaning mango of the village (M. indica), and ‘sva:y prey’, wild mango (i.e. M. caloneura) as attested in pre-Angkorian Khmer inscriptions dating from the 6th to the 8th century ad (Pou and Martin, 1981)) is of Austro-Asiatic origin. ‘Sva:y’ has cognates in south Vietnam (‘xoay’) and in Asian languages spoken by aboriginal people in peninsular Malaysia. ‘Wai’, another cognate, is a vernacular name of M. minor in several parts of New Guinea. Pawley and Ross (1995) proposed ‘wai’ and ‘pau(q)’ as the reconstructed Proto-Oceanic terms referring, respectively, to a generic name for mango, and a species that is probably M. indica.
Nowadays, these two words are generic terms for mango fruits that rather closely resemble M. indica. In the same way, ‘thayet’ which is the com-mon vernacular name referring to M. indica in Myanmar (‘sinnin thayet’ and ‘taw-thayet’ for M. caloneura and M. sylvatica, respectively), or ‘mamuang’ in Thai languages are probably generic names.
Obviously, linguistic evidence alone provided by these vernacular names is not suffi cient to prove the time and place of an introduction. None the less, it is signifi cant that in mainland South-east Asia none of the vernacular names of the common mango exhibits signs of an Indian infl uence, moreover, cog-nates of these names are also applied to primitive races in some parts of insular South-east Asia.
Evidence of early trade in South-east AsiaThe history of plant domestication in mainland South-east Asia has undoubt-edly involved introduction of plants by people migrating from the mainland into insular South-east Asia. In more recent times, there is evidence of con-tacts and sea trade since at least the fi rst centuries ad between mainland and insular South-east Asia to indicate that there have been numerous opportuni-ties for introduction of the common mango from different places at different times prior to the 4th century (before the Indianization of early South-east Asian states) into present-day Malaysia and Indonesia.
Recent studies based on archaeological evidence stress the long unrecog-nized importance of South-east Asian trade (emanating from South-east Asia) between ports established along the Java Sea, those of mainland Asia, and India, back to the 1st century ad, and possibly earlier (Walker and San-toso, 1984). Trade routes connected the developing population centres of the mainland, such as the earliest known South-east Asian political entity, Funan, an advanced agrarian society located on the southern Vietnam coast, which became infl uenced by the Indians and reached the zenith of its commercial prosperity in the middle of the 3rd century (Hall, 1985). Increasingly, king-doms organized according to the Indian concept of royalty were established in the Indonesian archipelago, for example Kutai in East Kalimantan (4th century) and Central Java (8th to 9th century), the latter being famous for the Buddhist temple at Borobudur, where sculptures depict the mango tree.

Taxonomy and Systematics 35
It is highly probable that the eventual introductions of superior cultivars of polyembryonic mangoes from the south-west coast of India, ‘between the 6th and 14th century, the height of classical South-east Asian civilization and also the golden age of early south Indian civilization’ (Hall, 1985), were not the fi rst ones.
During the 16th and 17th centuries, the Portuguese and Spaniards con-tributed to the widest distribution of superior varieties in the archipelago, espe-cially to the east. The name mango itself derives from the Tamil ‘man-kay’ or ‘man-ga’ (see Mukherjee and Litz, Chapter 1, this volume), which the Portu-guese adapted as ‘manga’ and ‘mangueira’ when they colonized west India.
Superior Philippine cultivars originated through introduction of culti-vars from Indonesia, for example ‘Dodol’ into Mindanao, and from Indo-china, for example ‘Carabao’ and ‘Pico’ in Luzon, the Visayas and northern Mindanao (Wester, 1920; Bondad et al., 1984). However, these introductions dating from the fi rst half of the 17th century were also preceded by the intro-duction of primitive races of the common mango as well as other species into the Sulu Archipelago and Mindanao through contacts with north Borneo, as attested by their local names quoted by Wester (1920), that is mampalam (M. indica, and possibly also M. laurina), baonoh (M. caesia) and wannih (M.odorata).
The South-east Asian M. indica germplasm includes many races that defy classifi cation. Natural cross-pollination has undoubtedly occurred with native species, such as M. laurina, which was also brought into cultivation in several areas before the introduction of M. indica.
2.8 Conclusion
Potential contribution of wild species to mango cultivation
To date, the improvement and breeding of common mango has depended on the use of genetic variability within a single species, M. indica. Mukherjee (1957) observed that ‘similarity in chromosome number and pollen morphol-ogy in different species suggests close compatibility during hybridization and stock-scion relationship if other species are used as stock for the com-mon mango’. Biotechnology opens new perspectives for mango improve-ment (Litz, 2004). As noted by Litz et al. (Chapter 18, this volume), the transformation of mango with genes from other species could address a number of plant breeding objectives.
Source of rootstock
Grafting experiments between M. indica and other species are reported in the literature, for example budding of M. indica on M. foetida and M. odorata in Java (Ochse and Bakhuizen, 1931), M. odorata on M. indica in the Philippines (Wester, 1920), and M. indica on M. zeylanica in Sri Lanka (Gunaratman, 1946).

J.M. Bompard36
Mangifera indica ‘Madu’ in Java, and M. laurina in Sabah have been used as rootstocks for M. casturi. Trials of grafted M. caesia on M. indica (Wester, 1920) and M. indica on M. kemanga or M. caesia (Ochse and Bakhuizen, 1931) were unsuccessful, as these two species have distinct bark features and only remote affi nity with the common mango. Better compatibility can be expected using species more closely related to the common mango within the subgenus Mangifera. In West Kalimantan, M. laurina is occasionally used as a rootstock for the common mango on periodically inundated riverbanks. It has been tried as a rootstock by the Department of Agriculture in Sabah (Lamb, 1987). Campbell (2004) reported that M. casturi, M. griffi thii, M. laurina, M. odorata, M. pentandra and M. zeylanica grafted on M. indica had a high percentage of success.
Several species that can grow in permanently inundated areas (i.e. M.gedebe, M. quadrifi da, M. griffi thii and other species of the section Rawa) repre-sent a potential source of rootstock for the development of mango cultivation on poorly drained soils or in areas liable to prolonged fl ood. Other species may be a source of dwarfi ng rootstocks.
Hybridization
From our observations in Borneo, natural interspecifi c hybridization involv-ing various cultivated Mangifera species can occasionally occur. Suspected hybrids were observed between wild M. gedebe and cultivated M. laurina in the lakes area along the Mahakam River in East Kalimantan, where impor-tant populations of M. gedebe occur; between cultivated M. foetida and M. pajang, two species showing close affi nity, in different areas of Kalimantan where both species are grown together; and between closely related M. caesia and M. kemanga in cultivation. A hybrid origin has been suggested for M. odorata (M. indica × M. foetida), which is unknown in the wild (Ding Hou, 1978a). Based on AFLP analysis, Teo et al. (2002) and Kiew et al. (2003) have con-fi rmed that M. odorata is a hybrid between M. foetida and M. indica. The index of similarity showed that M. odorata is closer to M. foetida (76% similarity) than it is to M. indica (66%). Yamanaka et al. (2006) showed a high genetic similarity among 11 landraces of M. odorata from the Malaysian Agricultural Research and Development Institute (MARDI) gene bank. Higher variability can be expected from Sumatra and Java samples.
Existing information about experimental interspecifi c hybridization is scarce. According to Mukherjee et al. (1968), successful crosses between M. odorata and M. zeylanica were made in India.
Potential of wild species
There is little doubt that wild mangoes are potentially valuable in breeding programmes. Some species have important horticultural implications as they demonstrate many desirable characteristics (Bompard, 1993). Fairchild (1948)

Taxonomy and Systematics 37
noted that crosses between the common mango and related fi ve-stamen spe-cies of the section Euantherae might produce hybrids with better pollinating quality. Mangifera pentandra, which is grown in peninsular Malaysia and Sabah, is a prolifi c bearer, due to its high proportion of hermaphrodite to male fl owers.
Stress resistanceIn the Malesian rainforests, wild mangoes thrive well under an ever-humid climate, without a prolonged dry season, i.e. is in areas with an annual rain-fall > 4000 mm and no monthly mean < 100 mm and where the common mango cannot be grown satisfactorily. Species, occurring in subtropical areas, including primitive races of the common mango, or in high altitude tropical forests, should be evaluated for cold tolerance, opening up the possibilities for mango production in subtropical and Mediterranean areas. Mangifera lau-rina and other species related to the common mango that grow in the rainfor-est (e.g. M. minor in New Guinea) are apparently immune to anthracnose. Sharma and Choudhury (1976) also observed that trees of an unknown wild race found in the Tripura State (north-eastern India) were free from mango malformation.
Potential new fruitsExtensive, yet largely unrecorded variability also exists among the non-indica species under cultivation. Sadly, this gene pool is barely represented in exist-ing collections, and is rapidly vanishing. An increasing number of horticul-turists are demonstrating a keen interest in the wild relatives of the mango. It is hoped that local peoples who have contributed to the recognition and maintenance of these species can benefi t from future innovative mango breeding.
Since early times, local peoples have planted seeds collected from trees that were observed to produce better quality fruits in the forests around their settlements. In areas now completely devoid of lowland primary forest, espe-cially in Sumatra and Borneo, the only wild relatives still found are those which have been integrated into indigenous agroforests which represent gene banks for an amazing diversity of fruit trees. A tenuous but constant selection pressure over many centuries has resulted in improved selections of several species. Today, some of these selections hold economic importance for their intrinsic characteristics. In Malesia, forms of M. odorata and M. foetida with sweeter and less fi brous fl esh have been identifi ed. The ‘wani’, a form of M. caesia from Bali and Borneo, has green-skinned fruit with milky white soft fl esh and a sweet taste quite different from the fruit of common forms of M.caesia. In addition, there are many interesting selections of M. casturi, M. grif-fi thii and M. torquenda.
Further improvement of these wild mangoes is especially desirable owing to their local economic importance in the wet tropical regions. Use of vegetative propagation methods must be encouraged. With proper selection, there is every reason to believe that other Mangifera species can become valu-able commercial fruits.

J.M. Bompard38
Acknowledgement
Thanks are due to Dr Dawn Frame who assisted in correcting the text.
Note
1Surveys were carried out in Kalimantan in cooperation with the Indonesian Institute of Science (LIPI) and the Indonesian Commission on Germplasm, and in Malaysia with the Forest Research Institute of Malaysia (FRIM).
References
Angeles, D.E. (1991) Mangifera altissima Blanco. In: Verheij, E.W.M. and Coronel, R.E. (eds) Plant Resources of South-east Asia No.2: Edible Fruits and Nuts. Pudoc-DLO, Wageningen, the Netherlands, pp. 206–207.
Anonymous (1980) Mangifera L. In: Cheng, M. and Ming, T.L. (eds) Flora Reipublicae Popularis Sinicae. Vol. 45(1). Science Press, Beijing, Peoples Republic of China, pp. 73–78.
Aron, Y., Czosnek, H., Gazit, S. and Degani, C. (1998) Polyembryony in mango (Mangifera indica L.) is controlled by a single dominant gene. HortScience 33, 1241–1242.
Audley-Charles, M.G., Hurley, A.M. and Smith, A.G. (1981) Continental movements in the mesozoic and cenozoic. In: Whitmore, T.C. (ed.) Wallace’s Line and Plate Tec-tonics. Clarendon Press, Oxford, UK, pp. 9–23.
Bellwood, P., Fox, J.J. and Tryon, D. (eds) (1995) The Austronesians, Historical and Com-parative Perspectives. The Australian National University, Canberra.
Bompard, J.M. (1991a) Mangifera caesia Jack and Mangifera kemanga Blume, Mangiferafoetida Lour. and Mangifera pajang Kostermans. In: Verheij, E.W.M. and Coronel, R.E. (eds) Plant Resources of South-east Asia No.2: Edible Fruits and Nuts. Pudoc-DLO, Wageningen, the Netherlands, pp. 207–211.
Bompard, J.M. (1991b) Mangifera laurina Blume and Mangifera pentandra Hooker f.; Mangifera odorata. In: Verheij, E.W.M. and Coronel, R.E. (eds) Plant Resources of South-east Asia No.2: Edible Fruits and Nuts. Pudoc-DLO, Wageningen, the Neth-erlands, pp. 216–220.
Bompard, J.M. (1993) The genus Mangifera re-discovered: the contribution of wild spe-cies to mango cultivation. Acta Horticulturae 341, 69–77.
Bompard, J.M. (1995) Surveying Mangifera in the tropical rain forests of South-east Asia. In: Guarino, L., Ramanatha Rao, V. and Reid, R. (eds) Collecting Plant Genetic Diver-sity. Technical Guidelines. CAB International, Wallingford, UK, pp. 627–637.
Bondad, N.D., Rivera, F.N., Agcopra, D.B. and Minh Tam Aurin (1984) Philippine man-goes and their relationship to South-east Asian cultivars. Philippine Geographical Journal 28, 59–71.
Brandis, D.D. (1874) Forest Flora of North West and Central India. Allen, London, pp. 125–127.
Brown, W.H. (1950) Useful Plants of the Philippines. Vol. 2. Bureau of Science, Manila, the Philippines, pp. 336–340.
Campbell, R.J. (2004) Graft compatibility between Mangifera species and Mangiferaindica L. ‘turpentine’ rootstocks and their subsequent horticultural traits. Acta Horti-culturae 645, 311–313.

Taxonomy and Systematics 39
Corner, E.J.H. (1978) The Freshwater Swamp-forest of South Johore and Singapore. Bo-tanic Gardens, Parks and Recreation Department, Singapore.
Ding Hou (1972) A new species of Mangifera. Reinwardtia 8, 323–327.Ding Hou (1978a) Flora Malesiana Praecursores 56 (Anacardiaceae). Blumea 24,
1–41.Ding Hou (1978b) Anacardiaceae. 4. Mangifera. In: van Steenis, C.G.G.J. (ed.) Flora
Malesiana I. Vol. 8. Rijksherbarium, Leiden, the Netherlands, pp. 423–440.Eiadthong, W., Yonemori, K., Sugiura, A., Utsunomiya, N. and Subhadrabandhu, S. (2000a)
Records of Mangifera species in Thailand. Acta Horticulturae 509, 213–224.Eiadthong, W., Yonemori, K., Sugiura, A., Utsunomiya, N. and Subhadrabandhu, S.
(2000b) Amplifi ed fragments length polymorphism analysis for studying genetic relationships among Mangifera species in Thailand. Journal of the American Society for Horticultural Science 125, 160–164.
Engler, A. (1883) Anacardiaceae. In: de Candolle, A.P. (ed.) Monographiae Phanerog-amarum. Vol. 4. Masson, Paris, pp. 195–215.
Fairchild, D. (1948) The mango relatives of Cochin China; those with fi ve-stamen fl ow-ers. Proceedings of the Florida State Horticultural Society 61, 250–255.
Gunaratman, S.C. (1946) The cultivation of mango in the dry zone of Ceylon (part II). Tropical Agriculturist 102, 23–30.
Hall, K.R. (1985) Maritime Trade and State Development in Early South-east Asia. Allen and Unwin, Sydney.
Hooker, J.D. (1862) Mangifera. In: Bentham, G. and Hooker, J.D. (eds) Genera Plantarum.Vol. I. Reeve and Co., London, p. 420.
Juliano, J.B. (1937) Embryos of Carabao mango (Mangifera indica Linn.). PhilippinesAgriculturist 25, 749–760.
Kanjilal, U.N., Kanjilal, P.C. and Das, A. (1937) Mangifera. Flora of Assam 1(2), 335–336.Kiew, R., Teo, L.L. and Gan, Y.Y. (2003) Assessment of the hybrid status of some Malesian
plants using Amplifi ed Fragment Length Polymorphism. Telopea 10, 225–233.Kostermans, A.J.G.H. (1965) New and critical Malesian plants VII. Reinwardtia 7,
19–46.Kostermans, A.J.G.H. and Bompard, J.M. (1993) The Mangoes. Botany, Nomenclature,
Horticulture, and Utilization. Academic Press, London.Kurz, S. (1877) Forest Flora of British Burma. Vol. 1. Burma Government Printers, Cal-
cutta, pp. 304–305.Lamb, A. (1987) The potential of some wild and semi-wild fruit trees in Sabah and the
progress made by the Department of Agriculture, Sabah in establishing a germplasm pool. Paper presented in Forest Research Institute Malaysia, Kepong, Malaysia.
Litz, R.E. (2004) Biotechnology and mango improvement. Acta Horticulturae 645, 85–92.Marchand, L. (1869) Révision du Groupe des Anacardiacées. Baillière, Paris.Mehrotra, R.C., Dilcher, D.L. and Awasthi, N. (1998) A palaeocene Mangifera-like leaf
fossil from India. Phytomorphology 48, 91–100.Min, T. and Barfod, A. (2008) Anacardiaceae, Mangifera. In: Wu, Z.Y., Raven, P.H. and
Hong, D.Y. (eds) Flora of China. Vol. 11. Missouri Botanical Garden Press, St Louis, Missouri and Science Press, Beijing, pp. 338–339. Available at: http://fl ora.huh.harvard.edu/china/mss/volume11/index.htm#alphabetical_list (accessed 5 August 2008).
Molesworth, A.B. (1967) Malayan Fruits. An Introduction to the Cultivated Species.Donald Moore Press, Singapore.
Mukherjee, S.K. (1949) A monograph of the genus Mangifera L. Lloydia 12, 73–136.Mukherjee, S.K. (1953) Origin, distribution and phylogenetics affi nities of the species of
Mangifera L. Journal of the Linnean Society, Botany 55, 65–83.

J.M. Bompard40
Mukherjee, S.K. (1957) Cytology of some Malayan species of Mangifera. Cytologia 22, 239–241.
Mukherjee, S.K., Singh, R.N., Majumder, P.K. and Sharma, P.K. (1968) Present position regarding breeding of mango (Mangifera indica L.) in India. Euphytica 17, 462–467.
Nishiyama, K., Choia, Y.A., Honshoa, C., Eiadthong, W. and Yonemori, K. (2006) Ap-plication of genomic in situ hybridization for phylogenetic study between Mangifera indica L. and eight wild species of Mangifera. Scientia Horticulturae110, 114–117.
Ochse, J.J. and Bakhuizen, R.C. (1931) Fruits and Fruitculture in the Dutch East Indies.G. Kolff, Batavia (Jakarta), Indonesia.
Ochse, J.J., Soulé, M.J, Dijkman, M.J. and Wehlburg, C. (1961) Tropical and Subtropical Agriculture. Vol. 2. MacMillan, New York.
Pawley, A. and Ross, M. (1995) The prehistory of Oceanic languages: a current view. In: Bellwood, P., Fox, J.J. and Tyron, D. (eds) The Austronesians. The Australian Na-tional University, Canberra, pp. 39–74.
Philipps, A., Philipps, S.M. and Philipps, C. (1982) Some ornamental plants of Sabah. Nature Malaysiana 7(4), 20–27.
Pierre, L. (1897) Flore Forestière de la Cochinchine. Vol.1, fasc. 23. Doin, Paris.Pou, S. and Martin, M. (1981) Les noms des plantes dans l’épigraphie ancienne khmère.
Asie du Sud Est et Monde Insulindien 12, 3–70.Ravishankar, K.V., Chandrashekara, P., Sreedhara, S.A., Dinesh, M.R., Lalitha, A. and
Saiprasad, G.V.S. (2004) Diverse genetic bases of Indian polyembryonic and mo-noembryonic mango (Mangifera indica L) cultivars. Current Science 87, 870–871.
Schnell, R.J. and Knight, R.J., Jr (1993) Genetic relationships among Mangifera spp. based on RAPD markers. Acta Horticulturae 341, 86–92.
Schnell, R.J., Ronning, C.M. and Knight, R.J. (1995) Identifi cation of cultivars and valida-tion of genetic relationships in Mangifera indica L. using RAPD markers. Theoreti-cal and Applied Genetics 90, 269–274.
Sharma, D.K. and Choudhury, S.S. (1976) Occurrence of an unknown wild race of Mangifera in Tripura. Current Science 45, 305–306.
Sreekumar, P.V., Veenakumari, K. and Padhye, P.M. (1996) Mangifera griffi thii (Anacar-diaceae): an addition to the Indian mangoes, from Andaman Islands, India. Ma-layan Nature Journal 50, 85–87.
Teo, L.L., Kiew, R., Set, O., Lee, S.K. and Gan, Y.Y. (2002) Hybrid status of kuwini, Mangifera odorata (Anacardiaceae) verifi ed by amplifi ed fragment polymorphism. Molecular Ecology 11, 1465–1469.
Valmayor, R.V. (1962) The Mango, its Botany and Production. University of the Philip-pines, Laguna, the Philippines.
van Steenis, C.G.G.J. (1950) The delimitation of Malaysia and its main plant geographi-cal divisions. In: van Steenis, C.G.G.J. (ed.) Flora Malesiana. Series I. Sijthoff and Noordhoff Publishers, Groningen, the Netherlands, pp. 70–75.
Verheij, E.W.M. and Coronel, R.E. (eds) (1991) Plant Resources of South-east Asia No.2: Edible Fruits and Nuts. Pudoc-DLO, Wageningen, the Netherlands.
Walker, M.J. and Santoso, S. (1984) Romano-Indian pottery in Indonesia. In: van de Velde, P. (ed.) Prehistoric Indonesia, a Reader. Foris, Dordrecht, the Netherlands, pp. 376–383.
Watt, G. (1891) A Dictionary of the Economic Products of India. Vol. 5. Offi ce of the Superintendent, Government Printing, Calcutta; W.H. Allen, London, pp. 146–157.
Wester, P.J. (1920) The Mango. Bureau of Agriculture Bulletin No. 18. Bureau of Agricul-ture, Manila, the Philippines.
Whitmore, T.C. (1975) Tropical Rain Forests of the Far East. Clarendon, Oxford, UK.

Taxonomy and Systematics 41
Whitmore, T.C. (1981) Paleoclimate and vegetation history. In: Whitmore, T.C. (ed.) Wal-lace’s Line and Plate Tectonics. Clarendon, Oxford, UK, pp. 36–42.
Yamanaka, N., Hasran, M., Xu, D.H., Tsunematsu, H., Idris, S. and Ban, T. (2006) Ge-netic relationship and diversity of four Mangifera species revealed through AFLP analysis. Genetic Resources and Crop Evolution 53, 949–954.
Yonemori, K., Honsho, C., Kanzaki, S., Eiadthong, W. and Sugiura, A. (2002) Phyloge-netic relationships of Mangifera species revealed by ITS sequences of nuclear ribo-somal DNA and a possibility of their hybrid origin. Plant Systematics and Evolution231, 59–75.

© CAB International 2009. The Mango, 2nd Edition: Botany, Production and Uses42 (ed. R.E. Litz)
3 Important Mango Cultivars and their Descriptors
R.J. Knight, Jr,¹ R.J. Campbell² and I. Maguire¹1University of Florida, Florida, USA
²Fairchild Tropical Botanic Garden, Florida, USA
3.1 Introduction 433.2 Criteria for Cultivar Description 443.3 Mango Cultivars 45
‘Alfa’ (Brazil) 45‘Alphonso’ (India) 45‘Amelie’ (West Africa) 46‘Arumanis’ (Indonesia) 46‘Ataulfo’ (Mexico) 46‘B74’ (‘Calypso’™) (Australia) 47‘Banganpalli’ (India) 47‘Beta’ (Brazil) 47‘Bombay Green’ (India) 47‘Cambodiana’ (Vietnam) 48‘Carabao’ (Philippines) 48‘Chausa’ (India) 48‘Cogshall’ (Florida, USA) 49‘Coração de Boi’ (Brazil) 49‘Dasheheri’ (India) 49‘Espada’ (Brazil) 49‘Ewais’ (Egypt) 50‘Excellent Succari’ (Egypt) 50‘Extrema’ (Brazil) 50‘Fajri’ (India) 50‘Fernandin’ (India) 51‘Genovea’ (Egypt) 51‘Glenn’ (Florida, USA) 51‘Golek’ (Indonesia) 51‘Haden’ (Florida, USA) 52‘Himsagar’ (India) 52‘Hindi Besennara’ (Egypt) 52‘Hindi Khassa’ (Egypt) 52

Mango Cultivars and Descriptors 43
‘Irwin’ (Florida, USA) 53‘Julie’ (West Indies) 53‘Keitt’ (Florida, USA) 53‘Kensington’ (Australia) 54‘Kent’ (Florida, USA) 54‘Khanefy’ (Egypt) 54‘Kyo Savoy’ (Thailand) 55‘Langra’ (India) 55‘Mabrouka’ (Egypt) 55‘Madame Francis’ (Haiti) 55‘Mallika’ (India) 56‘Manila’ (Mexico) 56‘Manzanillo’ (Mexico) 56‘Mesk’ (Egypt) 56‘Mulgoa’ (India to Florida, USA) 57‘Nabeel’ (Egypt) 57‘Nam Doc Mai’ (Thailand) 57‘Neelum’ (India) 58‘Nuwun Chan’ (Thailand) 58‘Okrung’ (Thailand) 58‘Osteen’ (Florida, USA) 59‘Pairi’ (India) 59‘Palmer’ (Florida, USA) 59‘Rosa’ (Brazil) 59‘Sensation’ (Florida, USA) 60‘Suvarnarekha’ (India) 60‘Tahar’ (Israel) 60‘Taimour’ (Egypt) 61‘Tommy Atkins’ (Florida, USA) 61‘Totapuri’ (India) 61‘Turpentine’ (West Indies) 62‘Vallenato’ (Colombia) 62‘Van Dyke’ (Florida, USA) 62‘White Succari’ (Egypt) 62‘Zebda’ (Egypt) 63
3.4 Conclusion 63
3.1 Introduction
The mango (Mangifera indica L.) has traditionally been grown in an area that extends southwards and eastwards from India through Myanmar and Vietnam to Indonesia. It probably is not indigenous to the Philippines, where it has long been cultivated (Valmayor, 1962), but Mangifera species are endemic there (Bon-dad, 1982). This crop is best adapted to a warm tropical monsoon climate, with a pronounced dry season followed by rains. Fruit of the best quality is usually produced in such areas, but specifi c races are known to fruit in humid regions. For example, some Mangifera species bear dependably on the island of Borneo, where most standard cultivars do not mature normal crops of fruit. Numerous Mangifera species closely related to the common mango are indigenous to

R.J. Knight et al.44
Borneo and nearby parts of Malaysia and Indonesia, and this region is the prob-able centre of diversity for this genus (see Bompard, Chapter 2 this volume).
Most crops long cultivated over an extended time and area show consider-able diversity, refl ecting different selection criteria in different regions of cul-ture as well as genetic responses to varied environmental infl uences. Certainly this is true of the mango. Indian cultivars differ markedly from those grown in South-east Asian countries and in Egypt. An additional factor that has promoted genetic diversity in the group recently has been the widespread introduction of this crop into new areas of cultivation, many in the western hemisphere, over the last 500 years. In this manner, genetically diverse germ-plasm has been brought from widely dispersed areas of the original range of the species and grown in mixed plantings where, through the cross-pollination natural to the species, new genetic combinations have been made and selected under many varying conditions of microclimate. In Florida, since the late 18th century, enough such importations and genetic recombinations have occurred to qualify the southern part of this state as a secondary centre of diversity for the crop. A new group of mango clones designated the Florida cultivars has been exported to Brazil, Israel, Australia and other places where the process of increasing diversity under new and varying cultural and environmental con-ditions continues (Knight and Schnell, 1994; Schnell et al., 2006).
Some Florida cultivars, most notably ‘Haden’, have been important in aid-ing the establishment of a modern mango industry in other parts of the world (Knight and Schnell, 1994), and the phenomenon fi rst observed in Florida is now occurring elsewhere; we are presented with the prospect of the importation of cultivars of outstanding merit from their countries of origin to be grown, fi rst experimentally and then commercially, in new regions. For this reason it is important to become familiar with the characteristics of a group of cultivars that currently are known in the commerce and/or horticulture of one or more coun-tries, and that may have potential for expanded culture or use in breeding.
3.2 Criteria for Cultivar Description
In the recent past, efforts to assemble lists of mango descriptors produced two publications that cover the subject (Mukherjee, 1985; IBPGR (now Inter-national Plant Genetic Resources Institute, IPGRI), 1989) and provide people who manage collections with morphological criteria to identify cultivars. The Descriptor List used by IPGRI documents passport data (identifying the acces-sion and information recorded by collectors), characterization (recording char-acters considered to be highly heritable which can easily be seen in the fi eld and are expressed in all environments) and preliminary evaluation, which records a limited number of additional traits thought desirable by a consensus of users of the crop. Plant data are important in preliminary evaluation, and include for the tree, habit and height of the mature tree; for the leaf, shape, length and width, and colour of the young leaf; and for the infl orescence, position, shape, density of fl owers, length, colour, hairiness, presence or absence of leafy bracts, and percentage of fl owers in an average infl orescence. (Some research indicates

Mango Cultivars and Descriptors 45
that both leafy bracts and number of perfect fl owers are infl uenced by local conditions and vary in their expression with differing environments.)
Additional plant data used in initial evaluation include those for the fl ower, diameter in millimetres, type (pentamerous, tetramerous or both), nature of disc (swollen, broader or narrower than ovary, reduced or absent) and number of stamens; fruit, length, width and thickness, weight, shape, skin colour (which may be compared with reference cultivars), skin thickness, skin texture, ratio of pulp to skin and stone, texture of pulp, adherence of skin to pulp, fi bre in pulp and its quantity and length, and stem insertion; and stone, length, weight, veins and pattern of venation, presence or absence of fi bres and their length.
Additional plant data for leaves, infl orescence and fruit have been col-lected and some of these, notably season (maturity period), productivity, eat-ing quality and attractiveness are quite important. Unfortunately, from the viewpoint of those who expect to apply these criteria outside the Indian sub-continent, reference cultivars are for the most part Indian and many are not readily available outside India. Other important characters that have been evaluated or proposed for evaluation include susceptibility to stress (drought, wind, fl ooding), susceptibility to specifi c diseases and pests, molecular markers, cytological characters and identifi ed genes. Because of the extreme comprehensiveness of this list and the limited availability of many of the proposed descriptor evaluations at this time, we have tried to utilize such information as is available to make the comparison, identifi cation and evalu-ation of specifi c well-known cultivars a practical possibility.
3.3 Mango Cultivars
A list of mango cultivars that are of interest in areas other than their places of origin, with descriptions intended to help differentiate them, follows (see Plates 4–40). Spelling and name variants in some cases represent efforts to transliterate from other orthographies to the Roman alphabet, and in others refl ect regional differences in usage.
‘Alfa’ (Brazil)
A monoembryonic cultivar developed by EMBRAPA Cerrado, Brazil, from crossing ‘Mallika’ × ‘Van Dyke’. The tree is semi-dwarf in habit and high-yielding, resistant to Oidium mangiferae and malformation, and moderately resistant to anthracnose (Colletotrichum gloeosporioides); the fruit is large (435 g), pink-red, fi rm, medium fi brous and of good quality (16% total soluble solids (TSS), 0.23% acidity) (Pinto et al., 2004).
‘Alphonso’ (India)
Also known as ‘Appus’, ‘Badami’, ‘Gundu’, Haphus’, ‘Kagdi’, ‘Khader’ and ‘Khader Pasand’. The tree is moderately large, with broadly rounded, dense

R.J. Knight et al.46
canopy; the fruit (Plate 4) is yellow, ovate-oblique, averaging 6 cm long by 5 cm broad, weighing 225–325 g (mean 226 g); the skin is thin; the fl esh is fi rm to soft, low in fi bre, yellow, sweet with characteristic aroma and with a very pleasant taste preferred by many who know this cultivar, bringing pre-mium prices on Indian and international markets. Seed is monoembryonic in a large, woody stone; the quality is excellent; ripening fruit in late to midsea-son. Bearing is irregular, medium to heavy in India, but light and irregular in Florida (Prasad, 1977; R.J. Knight, Jr, personal communication, 1995).
‘Amelie’ (West Africa)
Also known as ‘Gouverneur’ in the Caribbean. The tree is tall with a rounded, dense canopy; the fruit is green to orange-yellow with the advance of the season, rounded, 10–15 cm long by approximately 10 cm broad by approximately 7.8 cm thick and weighing 300–600 g (average 360 g). The skin is thick and separated with diffi culty; the fl esh is soft, juicy, melting, without fi bre, a deep orange colour, sweet and perfumed, free from turpentine, and provides the best of mango tastes. Seed is monoembryonic in a medium-sized, elongate, narrow stone that adheres to the fl esh, having a few short, pliable fi bres that are not objectionable; the quality is excellent; the season early. The fruit closely resem-bles that of ‘Julie’. ‘Amelie’ is exported to France, along with ‘Kent’, from Burkina Faso, Ivory Coast and Mali. ‘Amelie’ is increasing in popularity on the French market, chiefl y in Paris and the surrounding area. It brings lower prices than cultivars with blushed fruit because the consumer is not always aware when it is ripe (Naville, 1985, 1986; R. LePrette, personal communication, 1996).
‘Arumanis’ (Indonesia)
Also referred to as ‘Harumanis’. The tree is vigorous and tall with a slightly open canopy. The fruit (Plate 5) is greenish yellow with large, light-yellow dots, elongate oblong with rounded base, 11–14 cm long by 6.6–7.5 cm broad by 4.75–6.5 cm thick, weighing 200–350 g. The skin is thick, tough and easily separated, the fl esh fi rm and juicy with little fi bre, lemon yellow, sweet, slightly insipid with a strong aroma, of poor to fair quality. Seed is polyem-bryonic in a thick, woody stone; this cultivar ripens midseason and bears regularly. Relatively easy to propagate by graftage, scionwood survives well; widely planted in humid parts of the world where many better-quality culti-vars fail to fruit (R.J. Knight, Jr, personal communication, 1995).
‘Ataulfo’ (Mexico)
A polyembryonic cultivar sold in North American markets under the name ‘Ataulfo’ and as ‘Champagne’™. Originated in Tapachula, Chiapas, Mexico reportedly from seed brought from Costa Rica in about 1930. The tree is

Mango Cultivars and Descriptors 47
vigorous and upright, a mid-range producer with production averages of 10–20 t/ha possible. The tree is not highly adaptive to different climatic/edaphic conditions. It is moderately resistant to anthracnose disease. The fruit (Plate 6) is small (200–300 g), elongate, of good quality, sweet with slight acidity, yellow, fi rm, standing shipping stress well, and ripens from early to midseason (Campbell et al., 2002; Magallanes-Cedeño, 2004).
‘B74’ (‘Calypso’™) (Australia)
A monoembryonic cultivar that originated from the controlled cross of ‘Sensa-tion’ × ‘Kensington’. The tree is upright, with low to moderate vigour and is highly productive, with good tolerance of fl ower and fruit diseases; the fruit (Plate 7) is moderately large (457.4 ± 38.1 g), ovate (10.12 ± 0.27 cm long by 9.13 ± 0.28 cm wide), fi bre-free and fi rm, bright yellow overlaid with red blush, with extended shelf life and potential for shipment to overseas markets; ripens late in the season; patented (Whiley, 2001; Whiley and Hofman, 2006).
‘Banganpalli’ (India)
Also called ‘Beneshan’ and ‘Chappatai’. The tree is medium sized with a rounded canopy; the fruit is primrose-yellow, ovate-oblique, large and the skin smooth, thin and shiny, fl esh fi rm to meaty with juice moderately abun-dant, without fi bre, maize-yellow, with pleasant aroma and sweet taste. Seed is monoembryonic, in an oblong stone covered with sparse fi bres; quality good; ripens midseason and bears heavily (Singh, 1960).
‘Beta’ (Brazil)
A cultivar developed by EMBRAPA Cerrado, Brazil, from crossing ‘Amra-pali’ × ‘Winters’ (M20222 United States Department of Agriculture (USDA)). The tree is moderately vigorous and free of malformation, high-yielding but irregular, moderately resistant to anthracnose and Oidium; the fruit is small (310 g), yellow, fi rm with low fi bre, of excellent quality (24.8% TSS, 0.16% acidity) (Pinto et al., 2004).
‘Bombay Green’ (India)
Also called ‘Bhojpuri’, ‘Bombai’, ‘Hiralal Bombai’, ‘Kali Bombai’, ‘Laile Alipur’, ‘Malda’, ‘Sarauli’ and ‘Sheeri-Dhan’. The tree is tall and erect; the fruit (Plate 8) is apple green with ochre blush at the base and on some exposed parts, dots abundant, with brown specks in the middle, ovate with beak almost missing, medium sized, with tough, thick, non-adhering smooth skin; the fl esh is cadmium-orange, fi rm and juicy with scanty fi bre just under the skin,

R.J. Knight et al.48
very sweet with pleasant aroma, of very good quality; seed is monoembryonic in a full, thick, medium-sized stone. This cultivar ripens early in the season and is a medium bearer. ‘Bombay Yellow’ is said to be practically identical to this cultivar but for a slight difference in fruit colour. The present ‘Bombay Green’ is said to be a degenerate form of the original one (Singh, 1960). In Jamaica it is sometimes called ‘Peter’, which suggests a confusion with ‘Pairi’, but the Jamaican ‘Peter’ is without the bright red blush normal to ‘Pairi’.
‘Cambodiana’ (Vietnam)
Also known as ‘Xoai Voi’. The tree is moderately vigorous, with a dense, rounded canopy; the fruit (Plate 9) is greenish yellow, unblushed with a few small white dots, oblong to ovate, 9–11.5 cm long by 6.5–7.5 cm broad by 5–6 cm thick, weighing 220–340 g; the skin is thin, tender and adherent; the fl esh contains little fi bre, is tender and melting, lemon yellow, sweet and mildly subacid with a pleasant aroma; the seed is polyembryonic in a thick, woody stone. Ripens early in the season. Brought to Florida in 1902, where it gave rise to the ‘Saigon’ landrace (Campbell, 1992).
‘Carabao’ (Philippines)
The tree is vigorous, forming a large and dense canopy; the fruit (Plate 10) is greenish to bright yellow, brushed with a few small green dots, long and slender, with base rounded to slightly fl attened, 11–13 cm long by 6.5–7 cm broad by 6–6.5 cm thick, weighing 270–440 g; the skin is thick, medium tough and easily separated; the fl esh is without fi bre, tender and melting, lemon yellow, spicy and sweet with a mild aroma, of good to excellent quality; seed is polyembryonic in a thin, papery stone. Ripens early in the season (Camp-bell, 1992). This is a heavy bearer that may alternate; however, it can be induced to fruit by potassium nitrate treatment in the tropics (Bondad and Linsangan, 1979). It is highly resistant to bacterial black spot (Xanthomonascampestris pv. mangiferaeindicae) in Queensland (Mayers et al., 1988). It was introduced to Florida in 1909. ‘Carabao’ is important in commerce between the Philippines and Japan and is increasingly imported into the USA.
‘Chausa’ (India)
Also called ‘Samar Bahisht Chausa’ and ’Khajari’. The tree is tall and spread-ing; the fruit is canary yellow to raw sienna when fully ripe, with numerous obscure medium-sized dots with minute specks inside them, oblong with prominent beak, obtuse to rounded, medium sized; the skin is thin and some-what adhering, pulp raw sienna, soft and juicy with scanty fi ne, long fi bres near the skin; the fruit is very sweet with a luscious, delightful aroma, of excellent quality; seed is monoembryonic in a thick, medium-sized oblong

Mango Cultivars and Descriptors 49
stone with fi ne, short fi bres all over the surface and a tuft of long fi bres on the ventral edge. Ripens late in the season and is a light bearer (Singh, 1960).
‘Cogshall’ (Florida, USA)
A monoembryonic cultivar that originated on Pine Island in Lee County. The tree is relatively small, forming a rounded canopy, moderately susceptible to anthracnose and consistently productive; the fruit is medium to large, aver-aging about 350 g, yellow with a bright crimson blush, oblong (11–14 cm long by 7.5–8.5 cm broad by 6.2–8 cm thick) of excellent quality, rich and sweet in taste, with tender skin and soft fl esh. Ripens early to midseason over about 4 weeks, a season longer than some cultivars. It is recommended for the home garden, not commercial planting, in Florida but is now grown commercially on Mauritius and marketed in France. Seedling of ‘Haden’ (Campbell and Campbell, 1995; Schnell et al., 2006).
‘Coração de Boi’ (Brazil)
The tree is vigorous, precocious and productive; the fruit is greenish yellow and intense red on the side exposed to the sun, cordiform, medium sized, pulp yel-low and fi brous. The seed is polyembryonic. There are two seasons in São Paulo, January–February and September–December. This is one of the best-known commercial cultivars in São Paulo state (Sampaio, 1980; A.C. Pinto, personal communication, 1996; L.C. Donadio, personal communication, 1996).
‘Dasheheri’ (India)
Also known as ‘Dasheri’ and ‘Aman Dusehri’. The tree is of medium height and moderate vigour, spreading, with a rounded, medium-dense canopy; the fruit is primrose to canary yellow with abundant light-yellow dots, oblong to oblong-oblique with base rounded to obliquely rounded, medium sized, skin smooth, medium thick, tough and non-adhering; the fl esh is yellow, fi rm, with almost no fi bre, scanty juice and a delightful aroma, very sweet taste, of excellent quality; seed is monoembryonic in a thick, medium-sized stone. Rip-ens midseason and is heavy bearing; fruit keeps well (Singh, 1960).
‘Espada’ (Brazil)
The tree is tall and develops rapidly, with a dense canopy, very productive; the fruit is intense green or greenish yellow, oblong-elongate with a concave base, medium sized, with smooth, thick skin; the fl esh has much fi bre, is egg-yellow, with a strong aroma of turpentine. The quality is considered good for fresh consumption. The polyembryonic seed is in an oblong stone, covered

R.J. Knight et al.50
with soft fi bres and many nerves. There are two seasons per year in São Paulo, January–February and November–December (Sampaio, 1980; A.C. Pinto, personal communication, 1996).
‘Ewais’ (Egypt)
A polyembryonic cultivar of major commercial importance. The tree is vigor-ous, the fruit small (275 g), yellow with no blush, with small, light-brown slightly corky dots, ovate-oblong in shape (11.7 cm long by 7.2 cm wide by 6.3 cm thick), with adherent skin of intermediate thickness, relatively free of disease; fl esh orange, juicy but susceptible to jelly seed, with no objectionable fi bre, sweet and agreeable in taste, of very good quality. The stone is large (38.5 g). Fruit ripens midseason (Knight and Sanford, 1998). In warm subtrop-ics this cultivar has shown a tendency for fl owering in the warm season, with fruit ripening during the cool winter. It has good anthracnose tolerance.
‘Excellent Succari’ (Egypt)
A polyembryonic mango of minor commercial importance. The tree is vigor-ous, ripening fruit in late midseason. The fruit is small (280 g), green with a yellow overlay and small, yellow smooth dots, ovate-oblong in shape (11 cm long by 7 cm wide by 6.4 cm thick), with non-adherent skin of intermediate thickness quite free of surface disease; the fl esh is orange, melting (without jelly seed) and juicy with no objectionable fi bres, a delightfully sweet taste and excellent quality; stone large (36.6 g) (Knight and Sanford, 1998). It has moderate to good anthracnose tolerance in the warm subtropics.
‘Extrema’ (Brazil)
The tree is upright, vigorous and productive. The fruit is yellow with green-ish areas, ovate-reniform, weighing 350–400 g, with smooth and thin skin, and yellow, watery fl esh with almost no fi bres with a moderately resinous, agreeable taste. The quality is considered good for fresh consumption and processing. The polyembryonic seed is in a large, fi brous stone. Ripens early in the season (Sampaio, 1980; A.C. Pinto, personal communication, 1996).
‘Fajri’ (India)
Also spelled ‘Fazli’. The tree is of medium size and moderately vigorous, with rounded, open canopy. The fruit is light chrome yellow with small, dark-coloured fairly sparse dots, obliquely oval with base slightly rounded and beak distinct to slightly prominent, large (averaging 14.3 cm long by 9.8 cm broad, weighing 500 g on average) with a medium-thick skin that is

Mango Cultivars and Descriptors 51
smooth with some inclination to be warty, and fi rm to soft, fi breless fl esh of a light cadmium yellow with a pleasant aroma and a sweet taste, having juice that may be scanty to moderately abundant, of good to very good quality. The seed is monoembryonic in a large, oblong stone that is covered with a sparse, short and soft fi bre. Ripens midseason to late (Gangolly et al., 1957; N. Balasundaram, India, personal communication, 1990).
‘Fernandin’ (India)
The tree is moderately vigorous with a dense, rounded canopy; the fruit is bright yellow with an attractive bright-red blush, ovate-oblique, averaging 12.2 cm long by 8.5 cm broad, weighing 450 g; the skin is rough and warty, thick and adherent, fl esh bright yellow, moderately to abundantly juicy, thick, with no objectionable fi bre, with delightful to piquant aroma and sweet to very sweet, delicious taste, of superior quality; seed is monoembryonic; season late (Gangolly et al., 1957; Singh, 1960).
‘Genovea’ (Egypt)
A polyembryonic cultivar of minor commercial importance. The fruit is small (234.5 g), green with a yellow overlay and medium-sized smooth yellow dots, ovate-oblong in shape (11 cm long by 6 cm wide by 5.6 cm thick), a thin adherent skin relatively free of surface disease; fl esh orange, fi rm (no jelly seed) and juicy with no objectionable fi bres, a sweet agreeable taste of ac-ceptable quality; stone large (53 g) (Knight and Sanford, 1998).
‘Glenn’ (Florida, USA)
The tree is moderately vigorous, small to medium with dense, rounded can-opy of compact growth; the fruit (Plate 11) is bright yellow with orange-red blush, with numerous small yellow and white dots, oval to oblong with a rounded base, 9.5–12.5 cm long by 7.5–8.5 cm broad by 7–8 cm thick, weigh-ing 400–620 g; the skin is thin, tough and easily separated, fl esh soft and juicy, with little fi bre, deep yellow, rich and spicy with a strong, pleasant aroma, of excellent quality; seed is monoembryonic in a thick, woody stone. Ripens early in the season and is a regular bearer. This is a seedling of ‘Haden’ (Campbell, 1992; Schnell et al., 2006).
‘Golek’ (Indonesia)
The tree is moderately vigorous with an upright, open canopy; the fruit (Plate 12) is greenish yellow with an orange overlay and prominent white dots, oblong with rounded base, 9.5–12.5 cm long by 6–8 cm broad by 5.5–6.5 cm thick, weighing 200–365 g; the skin is thin, tough and easily separated;

R.J. Knight et al.52
the fl esh is soft and juicy with abundant fi bre (not objectionable), deep yel-low, sweet, insipid with a mild aroma, of poor to fair quality; the seed is polyembryonic in large, woody stone with abundant fi ne fi bre. Ripens mid-season (R.J. Knight, Jr, personal communication, 1995).
‘Haden’ (Florida, USA)
The tree is vigorous, with a large, spreading canopy; the fruit (Plate 13) is bright yellow with a deep crimson or red blush and numerous large yellow dots, oval with a rounded base, 10.5–14 cm long by 9–10.5 cm broad by 8.5–9.5 cm thick, weighing 510–680 g; the skin is thick, tough and adherent; the fl esh is fi rm and juicy with abundant fi bre, deep yellow, rich and sweet with a pleasant aroma, of good to excellent quality; the seed is monoembyonic in a medium-thick woody stone. Ripens early to midseason and bearing is sometimes irregular. This is a seedling of ‘Mulgoba’ × ‘Turpentine’ and is the fi rst of the Florida mango cultivars, introduced in 1910 and since grown in many other countries. It is the seed parent of numerous other cultivars (Campbell, 1992; Knight and Schnell, 1994; Schnell et al., 2006).
‘Himsagar’ (India)
The tree is vigorous, tall, with a dense, spreading canopy; the fruit (Plate 14) is greenish yellow to bright yellow with no blush, with light-yellow dots, ovate with a fl attened base, 12–15 cm long by 8.5–9.5 cm broad by 7.5–8.5 cm thick, weighing 465–585 g; the skin is thin, tough and easily separated; the fl esh is fi rm and juicy with no fi bre, orange, rich and sweet with a mild aroma, of good to excellent quality; the seed is monoembryonic in a thick, woody stone. This is a late midseason cultivar that bears well (R.J. Knight, Jr, per-sonal communication, 1995).
‘Hindi Besennara’ (Egypt)
A polyembronic cultivar of major commercial importance. The tree is of me-dium vigour, ripening fruit early to midseason. The fruit (Plate 15) is of small to medium size (319 g), green with orange overlay, with small white corky dots, oblong-cylindrical in shape (15.4 cm long by 6.7 cm wide by 6.5 cm thick) with thick, non-adherent skin relatively free of surface disease; the fl esh is orange, yielding and juicy with no objectionable fi bres, pleasantly sweet in taste, of very good quality; the stone is large (47.2 g) (Knight and Sanford, 1998).
‘Hindi Khassa’ (Egypt)
A polyembryonic cultivar of major commercial importance. The tree is vig-orous, ripening fruit in late midseason. The fruit is of medium size (461 g),

Mango Cultivars and Descriptors 53
yellow with no blush, with intermediate-sized smooth, light-yellow dots, oblong-cylindrical in shape (16 cm long by 6.6 cm wide by 6.9 cm thick), with thick, adherent skin relatively free of surface disease; fl esh is orange, fi rm and juicy with no objectionable fi bres, of mediocre taste and a quality not suitable for export; stone is large (55 g) (Knight and Sanford, 1998).
‘Irwin’ (Florida, USA)
The tree is small to medium, moderately vigorous, with open canopy. The fruit (Plate 16) is bright yellow with a crimson or bright red blush, numerous large white dots, ovate with rounded base, 11.5–13 cm long by 8–9 cm broad by 6.5–7.5 cm thick, weighing 340–450 g; the skin is medium-thick, tender and adherent; the fl esh is soft, tender, melting and juicy without fi bre, lemon yellow, sweet and mild with a pleasant aroma, of good quality; the seed is monoembryonic in a thin, papery stone. The stone may be seedless following cool weather at fl ower-ing time. This is an early, regular and heavy bearer. The fruit is usually soft with a short postharvest life, but it is often exported from tropical America to Europe. It is a seedling of ‘Lippens’ × ‘Haden’ (Campbell, 1992; Schnell et al., 2006).
‘Julie’ (West Indies)
Also called ‘St Julienne’. The tree is compact (dwarf), with a dense canopy; the fruit (Plate 17) is greenish yellow with a light pink to maroon blush and numerous small white dots, rounded with fl attened apex, pronouncedly compressed laterally, 7–9.5 cm long by 4–7.5 cm broad by 2–5.5 cm thick, weighing 200–325 g with a thin, tender skin and soft, melting, juicy, orange fl esh with scanty fi bre, of a rich, spicy fl avour with a strong, pleasant aroma, of good quality; seed is monoembryonic in a thin, papery stone. This cultivar ripens midseason and is a regular producer of small crops. The fruit is often severely infected with anthracnose disease, but its unique taste is preferred by many West Indians, and it is exported to the London market (C.W. Campbell, personal communication, 1996).
‘Keitt’ (Florida, USA)
The tree is medium sized, moderately vigorous, upright with open canopy; the fruit (Plate 18) is greenish yellow, with a pink or red blush, numerous small white or yellow dots, oval, with rounded base, 13–15 cm long by 9–11 cm broad by 8.5–10 cm thick, weighing 510–2000 g; the skin is thick, tough and adherent; the fl esh is fi rm and juicy, with little fi bre, lemon yellow, sweet and mild with a pleasant aroma, of good to excellent quality; the seed is monoembryonic in a thick and woody stone. This cultivar ripens late in the season. It is a seedling of ‘Brooks’. After ‘Tommy Atkins’ it is the most com-mercially important cultivar in the export mango industry of the western

R.J. Knight et al.54
hemisphere. It is resistant to anthracnose disease, packing and shipping stress and is heavily productive (Campbell, 1992; Schnell et al., 2006). It is highly sus-ceptible to bacterial black spot in Queensland (Mayers et al., 1988).
‘Kensington’ (Australia)
Also known as ‘Kensington Pride’ and ‘Bowen’. ‘Kensington’ has a large, vig-orous tree with spreading canopy; the fruit (Plate 19) is yellow with an orange-red blush on the shoulder, round ovate with a fl attened base and a slight beak, 10.5–13 cm long by 8.5–9.6 cm broad by 7.5–8.5 cm thick, weighing 350–750 g; the skin is thick, tender and adherent; the fl esh is soft and juicy, with moderate to little fi bre, sweet with a characteristic fl avour that makes it the most popu-lar cultivar in Australian markets, of excellent quality; seed is polyembryonic in a moderately thick, woody stone. This cultivar ripens midseason and it bears well. It is unusually susceptible locally, in Florida, to damage by red-banded thrips (Selenothrips rubricinctus (Giard.)), and may be killed by this pest without adequate countermeasures (R.J. Campbell, personal communication, 1994; R.J. Knight, Jr, personal communication, 1995). It is moderately suscep-tible to anthracnose and bacterial spot (Mayers et al., 1988).
‘Kent’ (Florida, USA)
The tree is large and vigorous with a dense, upright canopy; the fruit (Plate 20) is greenish yellow with a red or crimson blush, numerous small yellow dots, oval, with rounded base, 11–13 cm long by 9.5–11 cm broad by 9.9.5 cm thick, weighing 600–750 g; the skin is thick, tough and adherent; the fl esh is fi rm, tender, melting and juicy with little fi bre, deep yellow to orange-yellow, sweet with a rich fl avour and pleasant aroma, of excellent quality; the seed is monoembryonic in a thick, woody stone. Fruit ripens late midseason to late and bearing may be alternate. It is a seedling of ‘Haden’ × ‘Brooks’, which is a seedling of ‘Totapuri’ (‘Sandersha’) (Schnell et al., 2006). ‘Kent’ is not commonly commercial in Florida because it is prone to storage disease, but is a successful commercial cultivar in drier parts of Mexico, Central and South America and West Africa (Campbell, 1992). It is highly susceptible to bacterial black spot in Queensland (Mayers et al., 1988).
‘Khanefy’ (Egypt)
A cultivar of minor commercial importance. The fruit is large (475 g), green with a yellow overlay and large, brown, smooth dots, ovate in shape (10.7 cm long by 8.3 cm wide by 8.6 cm thick), with an adherent skin quite free of sur-face disease; the fl esh is yellow, often with jelly seed, juicy, with no objection-able fi bres and a bland fl avour unacceptable to many Western palates. The stone is moderately large (53 g) (Knight and Sanford, 1998).

Mango Cultivars and Descriptors 55
‘Kyo Savoy’ (Thailand)
The tree is large, vigorous, with an open canopy made up of long branches; the fruit is green when harvested (before the ripening process begins) turn-ing to greenish yellow, oblong, 11.5–12.5 cm long by 5.5–6.5 cm broad by 5–6 cm thick, weighing 230–340 g; the skin is thin, tender and adherent; the fl esh is medium fi rm, tender and not very juicy with no fi bre, pale yellow, very sweet with an insipid taste and a mild, pleasant aroma, of fair to good quality; the seed is highly polyembryonic in a medium-thin stone. This is a regular producer (C.W. Campbell, personal communication, 1995). The fruit is often consumed green.
‘Langra’ (India)
Also called ‘Darbhanga’, ‘David Ford’, ‘Hadialaziz’, ‘Hajipur Langra’, ‘Har-doi Langra’, ‘Lan Garhi’, ‘Langra Faquirwala’, ‘Sylhet’ and ‘Tikari’. The tree is moderately vigorous, forming a dense canopy; the fruit is greenish yellow with medium to big dark-green dots, ovalish to oblong, 8–10.5 cm long by 6.5–7.5 cm broad by 6–7 cm thick, weighing 235–375 g; the skin is medium smooth, thick; the fl esh is fi rm to soft, fi breless, lemon yellow, very sweet with a strong, pleasant aroma, juice moderately abundant; seed is monoem-bryonic in a medium-sized, fl attened stone covered with dense, short and soft fi bre; quality is very good. Fruit ripens early to midseason (Gangolly et al., 1957; R.J. Knight, Jr, personal communication, 1995).
‘Mabrouka’ (Egypt)
A major commercial cultivar considered to have been originally introduced from India. The tree is moderately vigorous; the fruit (Plate 21) is large (481 g), yellow with an orange to red blush and small, light-yellow, smooth dots; ovate-oblong in shape (13.7 cm long by 8.9 cm wide by 8.2 cm thick) with a thick, non-adherent skin relatively free of surface disease; the fl esh is yellow, fi rm and juicy with no objectionable fi bre, moderately agreeable in taste, of acceptable quality. The monoembryonic seed is in a moderately large (51 g) stone. Fruit ripens late midseason, ships well and has been marketed in Poland (Singh, 1960; Knight and Sanford, 1998).
‘Madame Francis’ (Haiti)
The tree is moderately vigorous, medium sized, forming an open canopy; the fruit (Plate 22) is greenish to bright yellow, with no blush and a few large rus-set dots, oblong, sigmoid with rounded base, 15–17 cm long by 8.5–11 cm broad by 5.5–7.5 cm thick, weighing 370–520 g; the skin is thin, tender and adherent; the fl esh is soft and juicy with medium fi bre, orange, rich spicy and sweet with a pleasant aroma, of fair to good quality; seed is polyembryonic

R.J. Knight et al.56
in a thin, papery stone. This cultivar ripens early to midseason and bears well. Shipped to North American markets from Haiti nearly 10 months of the year (R.J. Campbell, personal communication, 2007).
‘Mallika’ (India)
The tree is a moderately vigorous dwarf with a dense canopy; the fruit (Plate 23) is bright yellow with no blush and numerous small, light-yellow dots, oblong with rounded base, 10–12 cm long by 6.5–7.5 cm broad by 5–5.5 cm thick, weighing 280–450 g; the skin is thick, tough and easily separating; the fl esh is soft, tender and juicy with little fi bre, deep yellow to orange, rich, strongly aromatic and sweet, of excellent quality; seed is monoembryonic in a medium-thick and woody stone. This cultivar ripens midseason and is an irregular producer. This cultivar came from crossing ‘Neelum’ and ‘Dashe-hari’ (Singh et al., 1972; Campbell, 1992).
‘Manila’ (Mexico)
The tree is large, vigorous, with an upright, open canopy; the fruit (Plate 24) is bright yellow, sometimes with a light-pink blush, a few small reddish dots, long and slender with rounded base and bluntly pointed apex sometimes with a small beak, 12.5–14 cm long by 5.5–6 cm broad by 5–5.5 cm thick, weighing 180–260 g; the skin is thin, medium tough and easily separating; the fl esh is medium fi rm and juicy, with little to abundant fi bre, deep yellow, sweet, rich and spicy in taste with a pleasant aroma, of good to very good quality; seed is polyembryonic in a medium-thick and woody stone. This cultivar ripens early midseason and crops fairly dependably. For a long time ‘Manila’ has been the most popular mango in Mexico.
‘Manzanillo’ (Mexico)
The tree is large, of medium vigour with an upright canopy; the fruit is yel-lowish orange with 75% of the surface blushed an intense dark red with numerous dots, oval with moderately fl attened base, averaging 12 cm long by 10 cm broad by 7.5 cm thick, and 660 g in weight; the fl esh is low in fi bre, slightly subacid and very palatable, quality high; seed is monoembryonic in a relatively small stone. This cultivar ripens early in the season but spread over a 60-day harvest period. It bears heavily without pronounced alterna-tion and the fruit stores and ships well (Núñez-Elisea, 1984).
‘Mesk’ (Egypt)
A major commercial cultivar. The tree is vigorous, the fruit small to medium sized (312.5 g), yellow with a red blush, with small, corky yellow dots;

Mango Cultivars and Descriptors 57
ovate-oblong (11.3 cm long by 7.4 cm wide by 6.5 cm thick) with adherent skin intermediate in thickness and fairly free of surface disease; the fl esh is orange, frequently jelly-seeded, with no objectionable fi bres and a sweet, agreeable taste of very good quality. The polyembryonic seed is in a moder-ately large (52.5 g) stone. Fruit ripens late in the season (Knight and Sanford, 1998).
‘Mulgoa’ (India to Florida, USA)
Also spelled ‘Mugoba’ and ‘Mulgova’. The tree is large, vigorous with open, spreading canopy; the fruit (Plate 25) is bright yellow with a pink blush and numerous large white dots, oval to ovate with fl attened base, 8.5–10.5 cm long by 6.5–7.5 cm broad by 5–6 cm thick, weighing 340–450 g; the skin is thick, medium tough and adherent; the fl esh is soft, tender, melting and juicy, with little fi bre, lemon yellow, rich spicy and sweet with strong, pleas-ant aroma, of good to excellent quality; seed is monoembryonic in a thick, woody stone. This cultivar ripens midseason to late and is a shy, irregular bearer. Introduced to Florida in 1889 and called ‘Mulgoba’, this is the seed parent of ‘Haden’, fi rst of a series of cultivars known as the Florida group. A question exists whether the cultivar known in Florida is identical with the Indian cultivar or is a seedling rootstock that survived after the scion was killed by cold. In either case its superior quality ensured its retention and propagation (Campbell, 1992). Literature serves to compound the nomencla-tural confusion, as illustrated by Gangolly et al. (1957) whose ‘Mulgoa’ fruit, yellow overall and roundish oblique with a deeply depressed stem insertion, does not resemble the cultivar introduced to Florida. Singh (1960), on the other hand, portrays a rounded, lightly blushed greenish yellow fruit that closely resembles the Florida mango. Furthermore, vegetative propagation of selected chance seedlings has resulted in a variety of clonal types carried under this name in India (Ratnam and Chellapa, no date, post-1954).
‘Nabeel’ (Egypt)
A minor commercial cultivar. The fruit is large (495 g), green with small yel-low dots that are smooth; ovate-oblong (14 cm long by 9 cm wide by 7 cm thick), with adherent skin relatively free of surface disease; the fl esh is orange, fi rm and juicy, without objectionable fi bre, with a passable but not out-standingly pleasing taste and acceptable quality. The seed is polyembryonic in a large (56.6 g) stone (Knight and Sanford, 1998).
‘Nam Doc Mai’ (Thailand)
The tree is vigorous, medium sized with upright, dense canopy; the fruit (Plate 26) is greenish to bright yellow with a slight pink blush and numerous

R.J. Knight et al.58
small green dots, long and slender, sigmoid in shape with a rounded base, 17–19 cm long by 7.5–8.5 cm broad by 6.5–7.5 cm thick, weighing 340–580 g; the skin is medium thick, tender and easily separated from the fl esh which is soft, tender and juicy with no fi bre, lemon yellow, rich, spicy and very sweet with pleasant aroma, of excellent quality; seed is polyembryonic in a thin, papery stone. This cultivar ripens early midseason, fruits regularly and may have multiple crops in one season (Campbell, 1992). It is highly resistant to foliar infection, and resistant to fruit infection by bacterial black spot in Queensland (Mayers et al., 1988).
‘Neelum’ (India)
The tree is moderately vigorous with a small, compact canopy; the fruit is bright yellow with no blush and numerous small white dots, oval with fl at-tened or slightly rounded base, 9.5–11 cm long by 7.5–8.5 cm broad by 6–6.5 cm thick, weighing 230–300 g; the skin is thick, tender and easily sepa-rating; the fl esh is soft, melting and juicy with no fi bre, deep yellow, mild and sweet with a delightfully pleasant aroma, of good to excellent quality; seed is monoembryonic in a medium-thick, woody stone. This cultivar is a late, heavy bearer (Campbell, 1992).
‘Nuwun Chan’ (Thailand)
The tree is moderately vigorous, small, upright with a dense canopy; the fruit (Plate 27) is greenish yellow with a pink to red blush, numerous small green dots, long and slender with a fl attened base, 16–18 cm long by 7–8 cm broad by 6–6.5 cm thick, weighing 340–500 g; the skin is thick, tough and easily separating; the fl esh is soft, melting, juicy with little fi bre, pale yellow, mild and sweet with a faint, pleasant aroma, of good eating quality; seed is poly-embryonic in a thick, woody stone. This cultivar is an early, regular bearer. Fruit is often eaten green (Campbell, 1992).
‘Okrung’ (Thailand)
The tree is moderately vigorous, medium sized and upright, forming a dense canopy; the fruit (Plate 28) is green to greenish yellow with no blush and numerous small white dots, oblong and sigmoid with a rounded base, 11–13 cm long by 5–6 cm broad by 4.5–5.5 cm thick, weighing 160–240 g; the skin is thick, tough and medium adherent; the fl esh is soft and juicy with abundant fi bre, yellow or greenish, mild, somewhat insipid and very sweet with a pleasant aroma, of good quality; seed is polyembryonic in a thick, woody stone. This cultivar ripens midseason, is a heavy producer and some-times bears more than one crop/year (Campbell, 1992).

Mango Cultivars and Descriptors 59
‘Osteen’ (Florida, USA)
The tree is vigorous, medium sized, forming a dense canopy; the fruit (Plate 29) is yellow-orange with a purple or lavender blush and numerous small white dots, oblong with rounded base, 12–15.5 cm long by 9–10.5 cm broad by 8.6–9.5 cm thick, weighing 500–760 g; the skin is thick, tough and easily sepa-rating; the fl esh is fi rm and juicy, with little fi bre, lemon yellow, mild and sweet with a pleasant aroma, of good quality; seed is monoembryonic in a thick and woody stone. This cultivar ripens late midseason to late and is a regular producer. It is a ‘Haden’ seedling (Campbell, 1992; Schnell et al., 2006).
‘Pairi’ (India)
Also written ‘Pairie’, ‘Paheri’ and ‘Pirie’; synonyms are said to be ‘Peter’, ‘Peter Pasand’, ‘Grape’, ‘Gohabunder’, ‘Nadusalai’, ‘Rasjuri’ and ‘Yerra Goa’. The tree is moderately vigorous, forming a dense, rounded canopy; the fruit (Plate 30) is medium sized, green to yellow with a bright red blush, roundish, skin smooth, thick, fl esh golden yellow, slightly juicy, fi breless, with a deli-cious subacid taste, of excellent quality; the thick stone covered with short, bristly fi bre encloses monoembryonic seed (Popenoe, 1927; Singh, 1960). This cultivar has long been popular as a dooryard fruit tree in Hawaii.
‘Palmer’ (Florida, USA)
The tree is moderately vigorous, forming a large, upright, tight canopy; the fruit (Plate 31) is yellow-orange with a dark-red to crimson blush and a few small white dots, oblong with rounded base, 12–15 cm long by 8.5–10 cm broad by 6.5–7.5 cm thick, weighing 510–850 g; the skin is medium thick, tough and adherent; the fl esh is fi rm and melting with little fi bre, orange-yellow, mild and aromatic, of good quality; seed is monoembryonic in a medium-thick woody stone. This is a late midseason cultivar and is a regular bearer. It is a seedling of ‘Haden’ (Schnell et al., 2006). In Florida it is of minor commercial importance (Campbell, 1992). It is grown in Israel and is the seed parent of ‘Naomi’. It is attracting increased attention in the western hemi-sphere export market as a result of its superior eating quality.
‘Rosa’ (Brazil)
The tree is medium sized, of slow growth with a rounded canopy; the fruit (Plate 32) is yellow to rose-red on the side exposed to sun, oblong-cordiform and medium sized; the skin is thick and smooth; the fl esh is fi rm and mod-erately juicy, fi brous, golden yellow, moderately sweet with a turpentine aroma, of ordinary quality, susceptible to anthracnose disease; the seed is polyembryonic in a small, oblong stone. This cultivar ripens midseason to

R.J. Knight et al.60
late. It is one of the most important commercial cultivars in the Federal Dis-trict of Brazil, used for juice as well as fresh consumption, and is one of the most well-known cultivars in Brazil (Sampaio, 1980; L.C. Donadio, personal communication, 1996; A. Pinto, personal communication, 1996).
‘Sensation’ (Florida, USA)
The tree is vigorous, with a moderately open, symmetrical canopy; the fruit (Plate 33) is dark yellow with a prominent dark-red to purple blush that cov-ers most of its surface, oval with rounded base and rounded apex, 9–11.5 cm long by 7–8 cm broad by 6.5–7 cm thick, weighing 280–340 g; the skin is medium thick, tough and easily separating; the fl esh is fi rm and medium juicy, fi breless, deep yellow, mild and sweet with a weak, pleasant aroma, of fair to good quality; seed is monoembryonic in a thick, woody stone. This cultivar ripens midseason to late (Campbell, 1992). It is a seedling of ‘Haden’ × ‘Brooks’ (Schnell et al., 2006), and the seed parent of ‘B74’. It alter-nates severely, and in ‘on’ years the fruit may be clustered so heavily that it becomes diseased before maturity, thus ‘Sensation’ is not of commercial importance. It is highly resistant to bacterial black spot in Queensland (May-ers et al., 1988), but often has severe internal breakdown (browning, water soaking) (A.W. Whiley, personal communication, 1996).
‘Suvarnarekha’ (India)
Also called ‘Swarnarekha’ and ‘Sundri’. The tree is moderately vigorous and tall, with a rounded, dense, spreading canopy; the fruit is light cadmium yel-low with a blush of jasper red and abundant small, light-coloured dots, ovate oblong with a base slightly fl attened, of medium size, 11 cm long by 8.2 cm broad, weighing 400 g; the skin is medium thick, easily separated, fl esh soft, fi breless, primrose yellow with a pleasant aroma, sweet taste and abundant juice, of medium to good quality; seed is monoembryonic in an oblong-oval stone covered with soft, short fi bre. Ripens early in the season early and is heavy bearing (Gangolly et al., 1957).
‘Tahar’ (Israel)
The tree is vigorous, medium sized, with an upright, dense canopy; the fruit is bright yellow with a dark-red blush and numerous small white dots, ovate with fl attened base, 11.5–13 cm long by 8.9–9.5 cm broad by 7.5–8 cm thick, weighing 360–520 g; the skin is thick, tough and easily separating; the fl esh is soft and juicy with little fi bre, deep yellow, mild, aromatic and slightly insipid with a strong odour not appreciated by many, of fair to good quality; seed is monoembryonic in a medium-thick woody stone. This cultivar ripens in late midseason and bears well in Israel (Campbell, 1992).

Mango Cultivars and Descriptors 61
‘Taimour’ (Egypt)
A major commercial cultivar. The tree is vigorous; the fruit (Plate 34) is large (500 g), ripening late in the season, dark green with large light-brown dots, smooth in texture, ovate-oblong (12.8 cm long by 8.4 cm wide by 8 cm thick), with non-adherent skin of intermediate thickness, quite free from surface disease; fl esh is orange, fi rm (free of jelly seed) and juicy with no objection-able fi bre, of a delightfully rich, sweet taste and excellent quality. The seed is polyembryonic in a medium-sized (50.8 g) stone (Knight and Sanford, 1998).
‘Tommy Atkins’ (Florida, USA)
The tree is vigorous, with a dense, rounded canopy; the fruit (Plate 35) is orange-yellow, with a crimson or dark-red blush and numerous small, white dots, oval to oblong, with broadly rounded base, 12–14.5 cm long by 10–13 cm broad by 8.5–10 cm thick, weighing 450–700 g; the skin is thick, tough and adherent; the fl esh is fi rm and medium juicy; with a medium amount of fi bre, lemon to deep yellow, mild and sweet with a strong pleasant aroma, of fair to good quality; seed is monoembryonic in a thick, woody stone. This cultivar ripens early to midseason. It is a ‘Haden’ seedling (Schnell et al., 2006). ‘Tommy Atkins’ is the most important commercial cultivar in the western hemisphere export mango market; it is highly resistant to anthracnose dis-ease and handling and shipping stress, and a consistent, heavy producer (Campbell, 1992). ‘Jelly seed’ (internal breakdown) is a serious problem in the moist subtropics and tropics outside Florida, where the mango is grown on calcareous, well-drained soil (A.W. Whiley, personal communication, 1996).
‘Totapuri’ (India)
Also called ‘Bangalora’, ‘Collector’, ‘Kallamai’, ‘Killi’ (‘Gilli’), ‘Mukku’, ‘Sand-ersha’ and ‘Thevadiyamuthi’. The tree is of medium size, vigorous, spread-ing with an open canopy; the fruit (Plate 36) is greenish yellow with a pink blush and a few small, white dots, oblong, base rounded, apex rounded to bluntly pointed with a large beak, 17.5–20 cm long by 9–11.5 cm broad by 8.5–10.5 cm thick, weighing 800–1100 g; the skin is thick, tough and adherent; the fl esh is fi rm and medium juicy with a weak, somewhat repugnant aroma, of poor to fair quality; seed is monoembryonic in a thin, papery stone. This cultivar ripens late midseason, is productive and regular bearing. Fruit cracks when exposed to heavy rains at ripening time. ‘Totapuri’ was imported to Florida twice, as ‘Sandersha’ in 1901 and as ‘Totapuri’ in the early 1960s. It is the seed parent of ‘Anderson’ and ‘Brooks’, which is itself the parent of ‘Kent’. It is called ‘Totapuri’ in Bangalore, and ‘Bangalora’ in much of the rest of India (Gangolly et al., 1957; Campbell, 1992).

R.J. Knight et al.62
‘Turpentine’ (West Indies)
The tree is vigorous, with a large, spreading, rounded canopy; the fruit (Plate 37) is bright yellow with a few large white dots, occasionally with a pink blush, oval with a fl attened base, 7.5–8.5 cm long by 6.5–7.5 cm broad by 6–6.5 cm thick, weighing 140–200 g; the skin is thick, tough and easily sepa-rating; the fl esh is fi rm and juicy, with abundant coarse fi bre, lemon yellow, rich, aromatic, spicy, resinous and sweet with a strong, pleasant aroma, of poor to fair quality; seed is polyembryonic in a thick, woody stone. This cul-tivar ripens early midseason to late midseason and is a heavy producer but may alternate. It is commonly used as a grafting stock (Campbell, 1992).
‘Vallenato’ (Colombia)
The tree is vigorous, with an upright, dense canopy; the fruit (Plate 38) is bright yellow, with a crimson blush, oblong with fl attened base, 8–9 cm long by 7–8 cm broad by 6–7 cm thick, weighing 195–340 g; the skin is thin, tough and adherent; the fl esh is fi rm, juicy with abundant fi ne fi bre (not objection-able), pale yellow, mild and sweet with a strong, pleasant aroma, of good to excellent quality; seed is monoembryonic. This cultivar ripens in early mid-season (R.J. Campbell, personal communication, 1995).
‘Van Dyke’ (Florida, USA)
The tree is moderately vigorous, with a large, open canopy; the fruit (Plate 39) is bright yellow with a bright red or crimson blush, oval with rounded base, 9–11.5 cm long by 7.5–9.5 cm broad by 7–8 cm thick, weighing 250–520 g; the skin is thick, tough and easily separating; the fl esh is quite fi rm, melting and juicy with little fi bre, orange-yellow, rich, spicy and sweet with a strong, pleasant aroma, of good to excellent quality, but susceptible to inter-nal breakdown; seed is monoembryonic in a medium-thick, woody stone. This cultivar ripens in late midseason and is a regular, heavy producer. It is a seedling of ‘Haden’ (Campbell, 1992; Schnell et al., 2006).
‘White Succari’ (Egypt)
A cultivar of major importance. The tree is vigorous; the fruit is medium large (410 g), greenish yellow with yellow overlay and small, brown dots of smooth texture, ovate-oblong in shape (11.25 cm long by 8.3 cm wide by 8.0 cm thick), with thin adherent skin reasonably free of surface disease; fl esh is orange, yielding and juicy with no objectionable fi bres, of an agree-able sweet taste and very good quality. The seed is polyembryonic in a moderate to large-sized stone (49 g), the season early (Knight and Sanford, 1998).

Mango Cultivars and Descriptors 63
‘Zebda’ (Egypt)
A cultivar of major importance. The tree is vigorous and regularly produc-tive; the fruit (Plate 40) is large (660 g), green with no overlay and small, brown dots of smooth texture, oblong-cylindrical in shape (14.6 cm long by 9.7 cm wide by 8.3 cm thick), with non-adherent skin quite free of surface disease; fl esh is deep orange, fi rm and juicy, with no objectionable fi bre and a mild, sweet taste, of acceptable quality. The seed is polyembryonic in a moderately small (52 g) stone. This cultivar is of late-midseason maturity (Knight and Sanford, 1998). It is highly tolerant of anthracnose and resistant to malformation (R.C. Ploetz, personal communication, 2007).
3.4 Conclusion
The mango fruit’s nutritional value, aesthetic and gustatory appeal have assured its growing importance in non-traditional markets since the late 1950s, as it has been introduced to consumers previously unacquainted with it. Furthermore, the migration of large populations from South-east Asia and other regions where this fruit is a traditional crop to metropolitan centres where it has not been well known has created a permanent demand for it in these new markets. An additional factor permitting market expansion has been the growing mango production in areas previously unimportant in world commerce such as Mexico, Brazil, Australia, West Africa, Israel, Flor-ida and the Canary Islands. The fact that most new markets are remote from areas of production has necessitated selection of cultivars for fresh market sale that are dependably productive and resistant to harvest, handling and shipping stress, with relatively long shelf life, for example ‘Tommy Atkins’, ‘Keitt’ and ‘Madame Francis’. The fruit quality of mango cultivars well suited to packing and shipping has been a secondary consideration, and is gener-ally not so high as that of cultivars acknowledged to be superior for eating. Economic factors obviously must dictate what is grown for the fresh market.
The commercial market for processed mango products permits other cultivars to be utilized, and these may vary with the product that is mar-keted. Cultivars chosen for purée or juice preparation are likely to be quite different from those used for manufacture of chutney or other products requiring pulp that maintains its integrity after it is cooked. ‘Totapuri’ (‘Sand-ersha’) or ‘Turpentine’, for example, considered mediocre for fresh consump-tion, can be used to prepare excellent chutneys, as can many ‘criollo’ types in the West Indies. ‘Tommy Atkins’ makes outstandingly good dried fruit sec-tions, sweet and aromatic, even though its fresh-fruit quality is generally conceded not to be high. Mango butter and mango leather are other products that are appreciated by many who know them (see Raymundo et al., Chapter 17, this volume; Campbell and Campbell, 1983; Campbell and Smith, 1987). As more fruit that is wholesome, but not of export quality, becomes available in areas of increasing production, it is likely that processed mango products will become more common.

R.J. Knight et al.64
Despite the recognized high quality of many well-known mango cultivars, considerable cultivar improvement is still needed in most regions of culture before anything approaching perfection is likely to be achieved (Table 3.1). For any given area, cultivars that combine adequate resistance to disease and packing and shipping stress, regular heavy production, high quality, and attractive appearance throughout a long bearing season are all requisites. Production of seedlings from controlled crossing of different parents having desired characters, followed by vigorous selection and evaluation of the resultant selections, can produce such improved cultivars. Pursuit of com-mon goals, including the cooperative exchange and testing of elite germ-plasm in different regions of production, can accelerate progress towards this objective (Lavi et al., 1989, 1993; Knight, 1993). Such interregional and inter-national activities are to be encouraged because of their potential for advanc-ing mango production and utilization in the world.
Acknowledgements
For their help in reviewing portions of this chapter and/or contributing vast quantities of information on mango cultivar descriptors and attributes, the authors are profoundly grateful to the following: Dr N. Balasundaram, Head, Sugarcane Breeding Institute, Regional Centre, Karnal, India 132001; the late Dr Carl W. Campbell, Tropical Research and Education Center, 18905 SW 280
Table 3.1. Ratings of selected mango cultivars grown in Florida (Source: Knight, 1993).
Cultivar Shapea Sizeb Firmnessc Colourd Anthracnosee Fibref Tasteg Yieldh Scorei
‘Alphonso’ 3 5 7 2 3 7 9 1h x‘Boribo’ 3 8 8 4 7 9 5 6 x‘Carabao’ 5 6 7 3 5 9 8 6h x‘Haden’ 3 9 8 8 5 7 7 3h x‘Keitt’ 4 10 9 6 8 9 8 8 ///‘Kensington’ 3 8 7 7 7 8 7 6 /‘Langra’ 2 6 8 3 5 8 8 3h x‘Pope’ 3 9 5 7 2 8 8 1 x‘Tommy Atkins’
3 9 9 9 9 6 6 7 ///
‘Van Dyke’ 3 7 10 9 7 8 7 6 ///
aRatings of 1 (round) to 5 (long) indicate fruit shape, not its desirability.bRatings below 6 justify discard; those of 7 and above show size only, not merit.cRatings of 1–10 where 1 = least and 10 = most.dRatings of 1–10 where 1 = least and 10 = most.eRatings of 1–10 where 1 = most and 10 = least susceptible.fRatings of 1–10 where 1 = most and 10 = least.gRatings of 1–10 where 1 = worst and 10 = best.hTrends markedly towards alternate bearing.iOne or more checks (/) show overall value; (x) indicates no commercial acceptability.

Mango Cultivars and Descriptors 65
Street, Homestead, Florida 33031 USA; Luis C. Donadio, Universidade Estad-ual Paulista, Rodavia Carlos Tonanni KM5, Jaboticabal, 14870, São Paulo, Brazil; Dr Shmuel Gazit, Department of Horticulture, Hebrew University of Jerusalem, Rehovot 76100, Israel; Mr Remy LePrette, Directeur, Interfel, 155 Rue F.G. Poissonierre, Paris 75009, France; Dr Alberto C. Pinto, Lider de Pro-jeto/CPAC, EMBRAPA, Caixa Postal 08 223, CEP 73301-970, Brasilia, DF, Brazil; Dr Eli Tomer, Department of Fruit Trees, the Volcani Centre, Institute of Horticulture, PO Box 6, Bet Dagan 50-250, Israel; Dr Anthony W. Whiley, Maroochy Horticultural Research Centre, PO Box 5093 SCMC, Nambour, Queensland 4560, Australia.
References
Bondad, N.D. (1982) Mango and its relatives in the Philippines. Philippine Geographic Journal 26, 88–100.
Bondad, N.D. and Linsangan, E. (1979) Flowering in mango induced with potassium nitrate. HortScience 14, 527–528.
Campbell, B.A. and Campbell, C.W. (1983) Preservation of tropical fruits by drying. Proceedings of the Florida State Horticultural Society 96, 229–231.
Campbell, B.A. and Smith, J. (1987) An overview of tropical fruit uses in Florida. Pro-ceedings of the Florida State Horticultural Society 100, 408–411.
Campbell, C.W. and Campbell, R.J. (1995) ‘Cogshall’, a mango for the home garden. Proceedings of the Florida State Horticultural Society 108, 369–370.
Campbell, R.J. (ed.) (1992) A Guide to Mangos in Florida. Fairchild Tropical Garden, Miami, Florida, USA.
Campbell, R.J., Ledesma, N. and Campbell, C.W. (2002) Tropical Mangos: How to Grow the World’s Most Delicious Fruit. Fairchild Tropical Garden, Miami, Florida, USA.
Gangolly, S.R., Singh, R., Katyal, S.L. and Singh, D. (1957) The Mango. Indian Council of Agricultural Research, New Delhi, India.
International Board for Plant Genetic Resources (IBPGR) (1989) Descriptors for Mango. International Board for Plant Genetic Resources, Rome.
Knight, R.J., Jr (1993) Evaluating important fruit characters in mango germplasm. Fruit Varieties Journal 47, 25–30.
Knight, R.J. and Sanford, R.L. (1998) Mango Cultivar Evaluation. Publication No. 42. (USAID (United States Agency for International Development) Project No. 263–0240). ATUT (Agricultural Technology Utilization and Transfer) /Ronco, Giza, Egypt.
Knight, R.J. and Schnell, R.J. (1994) Mango introduction in Florida and the ‘Haden’ cultivar’s signifi cance to the modern industry. Economic Botany 48, 139–145.
Lavi, U., Tomer, E. and Gait, S. (1989) Inheritance of agriculturally important traits in mango. Euphytica 54, 5–10.
Lavi, U., Sharon, D., Tomer, E., Adato, A. and Gazit, S. (1993) Conventional and modern breeding of mango cultivars and rootstocks. Acta Horticlturae 341, 146–151.
Magallanes-Cedeño, R. (2004) Area-wide assessment of the ‘Ataulfo’ mango cultivation in the Soconusco region of Chiapas, Mexico. Acta Horticulturae 645, 361–363.
Mayers, P.E., Whiley, A.W., Hutton, D.G. and Saranah, J.B. (1988) Integrated control of bacterial black spot (Xanthomonas campestris pv. mangiferaeindicae) of mango. 1. Evaluation of 23 cultivars of mango for foliar and fruit resistance to bacterial black spot under orchard conditions at Childers, south east Queensland. Maroochy Hor-ticultural Research Report 5, 100–101.

R.J. Knight et al.66
Mukherjee, S.K. (1985) Systematic and Ecogeographic Studies of Crop Gene Pools. 1.Mangifera L. International Board for Plant Genetic Resources, Rome.
Naville, R. (1985) Les fruits tropicaux et subtropicaux sur le marche français en 1984. Fruits 40, 345–356.
Naville, R. (1986) Les importations françaises de fruits tropicaux et subtropicaux en 1985. Fruits 41, 409–420.
Núñez-Elisea, R. (1984) ‘Manzanillo-Núñez’: a new Mexican mango cultivar. Proceed-ings of the Florida State Horticultural Society 97, 360–363.
Pinto, A.C.Q., Andrade, S.R.M., Ramos, U.H.V. and Cordeiro, M.C.R. (2004) Intervari-etal hybridization in mango (Mangifi ca indica L.): techniques, main results and their limitations. Acta Horticulturae 645, 327–330.
Popenoe, W. (1927) Manual of Tropical and Subtropical Fruits. Macmillan, New York.Prasad, A. (1977) Bearing behaviour and fruit quality of south Indian varieties of mango
in northern India. Indian Journal of Horticulture 34, 372–376.Ratnam, L.V. and Chellappa, T. (no date, post-1954) Mulgoas of Hyderabad. In: Rao,
B.U. (ed.) The Mango a Souvenir. Department of Agriculture, Neo Silver Jubilee Press, Hyderabad, India.
Sampaio, J.M.M. (1980) Características gerais de algumas cultivares e tipos de manguei-ras no Brasil. In: Donadio, L.C. (ed.) Anais do I˚ Simposio Brasileiro sobre a Cultura da Mangueira. Departamento de Fitotecnia, Faculdade de Ciencias Agrarias e Vet-erinarias, Universidade Estadual de Jaboticabal, São Paulo, Brazil, pp. 35–50.
Schnell, R.J., Brown, J.S., Campbell, R.J., Kuhn, D.N., Meerow, A.W. and Olano, C.T. (2006) Mango genetic diversity analysis and pedigree inferences for Florida culti-vars using microsatellite markers. Journal of the American Society for Horticultural Science 131, 214–224.
Singh, L.B. (1960) The Mango: Botany Cultivation and Utilization. Leonard Hill, London.Singh, R.N., Majumder, P.K., Sharma, D.K. and Mukherkjee, S.K. (1972) Some promis-
ing mango hybrids. Acta Horticulturae 24, 117–119.Valmayor, R. (1962) The Mango: its Botany and Production. University of the Philip-
pines, Laguna, the Philippines.Whiley, A.W. (2001) Mango (Mangifera indica) ‘B74’. Plant Varieties Journal 14, 45–46.Whiley, A.W. and Hofman, P.J. (2006) ‘Calypso’™ Best Practices Manual. (On CD).
Horticulture Ltd, Nambour, Australia.

© CAB International 2009. The Mango, 2nd Edition: Botany, Production and Uses(ed. R.E. Litz) 67
4 Breeding and Genetics
C.P.A. Iyer1 and R.J. Schnell21Indian Institute of Horticultural Research, Bangalore, India
2USDA ARS, National Germplasm Repository, Miami, Florida, USA
4.1 Introduction 684.2 Origin of Cultivars and Distribution 68
Mangifera species and mango 68History of cultivation 69Impact of Florida mangoes 70
4.3 Reproductive Mechanisms 70Polyembryony 70Floral biology and pollination 71Incompatibility 72Cytology 72
4.4 Inheritance of Characters 73Dwarfness, regular bearing and precocity 73Flesh colour 74Skin colour 74Flowering time 74Beak 74Disease resistance 74Other horticultural traits 75
4.5 Breeding Objectives 75General objectives 75Specifi c objectives 76
4.6 Methods of Breeding 78Selection from open-pollinated seedlings 78Controlled pollination 79
4.7 Handling of Hybrid Populations and Selection 80Criteria for initial selection 80Pre-selection 81Potential for marker assisted selection (MAS) 81Molecular markers 81
4.8 Minimizing Problems in Breeding 83Heavy fruit drop 83

C.P.A. Iyer and R.J. Schnell68
Long juvenile phase 83Polyembryony 84
4.9 Achievements of Conventional Breeding 84India 85Other countries 86
4.10 Mutations 87Somatic mutations 87Induced mutations 88
4.11 Breeding Potential of Wild Species 884.12 Conclusions 89
4.1 Introduction
Mango has been considered to be a diffi cult plant species to improve in breeding programmes because of certain inherent characteristics including: (i) a long juvenile phase; (ii) a high level of heterozygosity resulting in unpre-dictable outcomes in hybridization; (iii) only one seed per fruit; (iv) heavy fruit drop leading to low retention of crossed fruits; (v) polyembryony in many cultivars; and (vi) the large area required for a meaningful assessment of hybrids. Despite these drawbacks, mango breeding can be successful because of its wide range of genetic variation and the ease with which a selected hybrid can be vegetatively propagated. Barring a few hybrid variet-ies resulting from planned hybridization programmes, which are now gain-ing increased attention, almost all known cultivars have resulted from the selection of chance seedlings from natural cross-pollinations. However, in Florida, a number of cultivars have resulted from the screening of seedlings from known mother plants. Most of the present-day mango cultivars were selected on the Indian subcontinent; these selections were made based mainly on fruit quality, with very little emphasis on modern horticultural and industrial requirements. These requirements include precocity, dwarf-ness, heavy and regular bearing, absence of physiological disorders, resis-tance to disease and pests and good shipping qualities. With decreasing land availability and the rising cost of labour, tree architecture requirements have also changed. The need for new cultivars to meet these requisites pinpoints the importance of planned hybridization rather than merely depending on chance seedlings. Current knowledge of hybridization techniques, inheri-tance patterns, management of hybrid populations and the development of genetic markers have greatly reduced the uncertainty in mango breeding.
4.2 Origin of Cultivars and Distribution
Mangifera species and mango
Almost all the commercial cultivars belong to Mangifera indica. However, a few commercial varieties of South-east Asia belong to other species, i.e. M. altissima, M. caesia, M. foetida, M. griffi thi, M. odorata, M. pentadra, M. sylvatica,

Breeding and Genetics 69
M. zeylanica, M. laurina, M. lagenifera, M. cochinchinensis, etc. The monoem-bryonic mango (M. indica) originated in north-eastern India (Assam), the Indo-Myanmar border region and Bangladesh (Chittagong Hill tract), where it is still found as a wild tree, with very small fruits. It may also occur in the lower Himalayan tract, near Nepal, Bhutan and Sikkim. Polyembryonic mangoes are considered to have originated in South-east Asia. Wild man-goes, representing different Mangifera species, can be found in tropical Asia, particularly north-eastern India, Sri Lanka, Myanmar, Thailand, Indo-China, southern China, Malaysia, Indonesia, Papua New Guinea, the Philippines and as far as the Solomon and Caroline Islands in the east. There are more than 60 species worldwide. The highest specifi c diversity is found in the heart of the distribution area of the genus Mangifera; the Malay Peninsula, Borneo and Sumatra (Bompard, 1993).
History of cultivation
Mango has been cultivated in India for at least 4000 years and over 1000 vari-eties are recognized there (Mukherjee, 1953). Almost all of them are selec-tions made from naturally occurring, open-pollinated seedlings. However, based on random amplifi cation of polymorphic DNA (RAPD) analysis, Rav-ishankar et al. (2004) felt that the mono- and polyembryonic types of Indian mango cultivars have a different genetic base, and that the polyembryonic types might have been introduced from South-east Asia and are unlikely to have originated in India. Mango culture gradually spread to tropical and subtropical countries throughout the world, where selections were made that were adapted to particular growing conditions. Thus, selection by man has played the most signifi cant role in the development of new mango culti-vars. The explorers who tasted the mango in the regions of its origin were enchanted with its aromatic qualities, ambrosial fl avour and creamy, smooth and silky texture, and introduced the fruit to other tropical regions. The spread of Hinduism and Buddhism assisted in the distribution of mangoes in South-east Asia. The Chinese traveller Hwen T’sang who visited India in the fi rst half of the 7th century ad returned to China with the mango. The mango was known in Baghdad in the 7th century.
The Persians or Omanis may have taken mangoes to East Africa around the 10th century ad. The fruit was introduced to East and West Africa in the early 16th century by the Portuguese and thence into Brazil. After being established in Brazil, the mango was carried to the West Indies, being fi rst planted in Barbados by about 1742 and later in the Dominican Republic and Jamaica (about 1782). The mango was introduced into Mexico from the Phil-ippines by the Spanish and also from the West Indies (Morton, 1987). Duval et al. (2006) developed microsatellite markers for studying the genetic vari-ability of Caribbean mangoes and concluded that there were two routes of mango to the French West Indies, namely, cultivars grown in Central Amer-ica (Mexico) and South America (Colombia) introduced from South-east Asia and also from former French colonies in the Indian Ocean. As the mango

C.P.A. Iyer and R.J. Schnell70
adapted to new locales, new cultivars were selected based on local adapta-tion and fruit preferences.
Impact of Florida mangoes
The fi rst recorded successful introduction of mango into Florida (USA) was made in 1861 (Knight, 1980). The earliest introductions were from the West Indies and India, followed by the introduction of several hundred accessions in the 20th century from South-east Asia, India and from other mango-growing areas of the world (Florida Mango Forum, 1951). The introduction of mangoes into Florida and subsequent development of a Florida group of mangoes has been reviewed by Knight and Schnell (1994). The Florida mango cultivars are unique in that they are hybrids between Indian cultivars (primarily mono-embryonic) and the South-east Asian cultivars (primarily polyembryonic) selected under south Florida conditions. The mango breeding system favours out-crossing. Therefore, the proximity of numerous genotypes of disparate geographical origins led to the production of many new seedlings by inter-pollination in Florida (Knight and Schnell, 1993). Florida selections are therefore not the result of a formal breeding programme. Early Florida selec-tions were made by growers and enthusiasts and historical information is often anecdotal. The Florida Mango Forum, established in 1938 for the advancement of mango production, documented historical information on the parentage of Florida cultivars in their proceedings. In addition to the United States Department of Agriculture (USDA) Germplasm Resources In-formation Network (GRIN) database, several sources compile information on Florida mango selections and introduction of accessions to Florida ( Ruehle and Ledin, 1956; Singh, 1960; Campbell and Campbell, 1993; Schnell et al., 2006). With the exception of South-east Asia, Australia and some African countries, which cultivate mostly locally selected varieties, the majority of countries produce cultivars developed in Florida, i.e. ‘Haden’, ‘Tommy Atkins’ and ‘Keitt’ (Galan Sauco, 1997). These Florida selections are now widely grown commercial cultivars affording production stability across many environments (see Mukherjee and Litz, Chapter 1, this volume).
4.3 Reproductive Mechanisms
Polyembryony
Nucellar embryosMangoes can be classifi ed into two groups, monoembryonic and polyembry-onic, based on their mode of reproduction from seeds. In general, monoem-bryonic seeds are found in the sub-tropical group (Indian type) and the polyembryonic seeds in the tropical group (South-east Asian). Monoembry-onic mango seeds each contain a single zygotic embryo, and hence only one seedling per seed that is of probable hybrid origin. Polyembryonic mango

Breeding and Genetics 71
seeds can contain one or more embryos, one of which is usually, but not always zygotic. Adventitious embryos develop from the nucellus, a maternal tissue surrounding the embryo sac, and consequently the seedlings of poly-embryonic mangoes are genetically identical to the maternal parent. Adven-titious embryos can also originate by direct budding from the cotyledons and hypocotyls of other nucellar embryos (Juliano, 1934). According to Mahesh-wari and Rangaswamy (1958), the nucellar cells destined to form adventi-tious embryos are recognizable by their dense cytoplasm and starchy contents. They gradually push into the embryo sac cavity where they divide and differentiate into embryos.
Inheritance of polyembryonyPolyembryony is genetically determined. Leroy (1947) considered that adven-tive embryony probably refl ects the effect of one or more recessive genes. This view was supported by Sturrock (1968), whose study of the progenies of monoembryonic mango hybridized with polyembryonic cultivars indicated that monoembryony was possibly a dominant trait. In contrast, Aron et al. (1998) and Brettell et al. (2004) observed that polyembryony in mango is con-trolled by a single dominant gene. Schnell et al. (2006) reported that 58 of the Florida cultivars had been classifi ed with 50 being monoembryonic and eight polyembryonic. Information from the Florida cultivars parentage analysis using 25 microsatellite markers supported the fi ndings of Aron et al. (1998) where polyembryony was found to be dominant. ‘Haden’ is a cross of the monoembryonic ‘Mulgoba’ and the polyembryonic ‘Turpentine’. If we assume that a single dominant gene controls this trait, all of the Indian cultivars in Florida must be homozygous recessive and the ‘Turpentine’ parent of ‘Haden’ must have been heterozygous. The evidence suggests that ‘Haden’ inherited the recessive allele from ‘Turpentine’, as all identifi ed progeny of ‘Haden’ are monoembryonic with the exception of ‘Winters’. The most probable pollen parent of ‘Winters’ is ‘Ono’, a polyembryonic cultivar from Hawaii. The fre-quency of this dominant allele is low in the Florida population and absent from the Indian cultivars in Florida. In view of these interesting fi ndings, and since a thorough knowledge of inheritance of polyembryony is essential for speculating the origin of M. indica, more work on these lines is warranted.
Floral biology and pollination
The mango infl orescence is primarily terminal, although axillary and multi-ple panicles may also arise from axillary buds. Both perfect (hermaphrodite) and staminate (male) fl owers occur in the same infl orescence. The total num-ber of fl owers in a panicle may vary from 1000 to 6000, depending on the cultivar (Mukherjee, 1953). Initial fruit set in mango is directly related to the proportion of perfect fl owers, although the fi nal fruit set does not necessarily depend on this ratio (Iyer et al., 1989). It appears that the proportion of per-fect fl owers in a cultivar becomes critical for optimum fruit set only when the proportion drops to 1%.

C.P.A. Iyer and R.J. Schnell72
Flowers begin to open early in the morning and anthesis has generally been completed by noon. The greatest number of fl owers opens between 9 and 10 a.m. Although the receptivity of the stigma continues for 72 h after anthesis, it is most receptive during the fi rst 6 h; however, there are reports that the stigma can become receptive even before anthesis has occurred (Singh, 1960). The minimum time required for pollen grains to germinate is 1.5 h (Sen et al., 1946; Singh, 1954; Spencer and Kennard, 1955). Singh and Singh (1952) observed 98% pollen viability after 11 months in storage at 7°C and 25% relative humidity (RH), and 65.7% viability after 24 months of stor-age at 0°C and 25% RH.
Mango is cross-pollinated, which is carried out by insects such as the common housefl y, honeybees and thrips, and possibly by other insects al-though to a lesser extent. Pollination by wind and gravity has been sug-gested to occur in mango (Popenoe, 1917; Maheshwari, 1934; Malik, 1951). In nature, > 50% of fl owers do not receive any pollen. Some workers had sug-gested that self-pollination in certain cultivars can also occur quite frequently (Dijkman and Soule, 1951). Studies by Issarakraisila and Considine (1994) have shown that for polyembryonic ‘Kensington’, a night temperature of < 10°C results in pollen grains with low viability (< 50%). The optimum temperature for normal meiosis is between 15 and 33°C with 70–85% viability.
Incompatibility
Although the existence of self-sterility in mango was suspected several years ago (Ruehle and Lynch, 1948, cited in Sharma and Singh, 1970; Dijkman and Soule, 1951), the prevalence of self-incompatibility was clearly established in monoembryonic ‘Dashehari’ by Singh et al. (1962). Subsequently, detailed studies indicated that the four popular monoembryonic cultivars of northern India (i.e. ‘Dashehari’, ‘Langra’, ‘Chausa’ and ‘Bombay Green’) were self- incompatible (Mukherjee et al., 1968; Sharma and Singh, 1970). Embryo-logical studies have shown that although fertilization takes place after self-pollination, degeneration of endosperm occurs 15 days after pollination involving self-incompatible parents (Mukherjee et al., 1968). The self- incompatibility system operating in mango appears to be of the sporophytic type. Instances of cross-incompatibility among certain mango cultivars have also been reported (Ram et al., 1976), necessitating the identifi cation of suitable pollinizers for mango.
Using an approach involving isozyme analysis, Dag et al. (2006) have initiated studies in many commercial mango cultivars in Israel and con-cluded that self-pollination is not a yield-limiting factor in monoembryonic ‘Maya’ and the practice of planting ‘Maya’ in solid blocks is sound. They had obtained similar results earlier with monoembryonic ‘Tommy Atkins’ (Dag et al., 1997).

Breeding and Genetics 73
Cytology
Chromosome numberInformation on the cytology of mango is quite limited. Only a few Mangifera species (i.e. M. indica, M. caloneura, M. sylvatica, M. foetida, M. caesia, M. odorata and M. zeylancia) have been studied, and were found to have chromosome numbers of 2n = 2x = 40 and n = x = 20 (Mukherjee, 1950, 1957; Roy and Visweswariya, 1951). Chromosome numbers and ploidy status of other species are yet to be studied. The only exception to this chromosome number that has been reported to date (Roy and Visweswariya, 1951) involves ‘Vallikolamban’, which was reported to be tetraploid (2n = 4x = 80), although subsequent stud-ies have indicated that it is only a diploid (Majumder and Sharma, 1990).
PolyploidyMango has been referred to as an allopolyploid. Due to the presence of secondary associations at metaphase of meiosis, Mukherjee (1950) suggested that the basic chromosome number of Mangifera is n = 8. In addition, the high number of somatic chromosomes and the correspondingly high number of nucleolar chromosomes led him to conclude that mango is an allopolyploid. However, the evidence used to arrive at this conclusion is not unequivocal. In fact, the molecular marker evidence is antithetical to this conclusion. Results from Duval et al. (2005), Viruel et al. (2005) and Schnell et al. (2005, 2006) using microsatellite markers all indicate that M. indica is diploid.
Although many wild Mangifera species are potentially valuable for crop improvement, they are yet to be exploited. Mukherjee (1963) felt that the different Mangifera species could intercross easily, based on the success ob-tained with interspecifi c crosses between M. zeylanica and M. odorata.
4.4 Inheritance of Characters
High heterozygosity in the cultivars that are used in hybridization and the inadequate number of hybrid progenies realized has made accurate genetic analysis in mango very diffi cult. However, based on limited data, some indi-cations are available which would be useful in selecting parents in breeding programmes designed with specifi c objectives. In studies of the distribution of different traits in seedlings derived from open-pollination (where the pol-len parent is unknown), Lavi et al. (1989) observed: (i) there is no maternal effect on juvenile period and fertility; (ii) there is a slight effect of the female parent on fruit taste and size; (iii) there is a maternal parent effect on harvest season and fruit colour.
Dwarfness, regular bearing and precocity
An analysis based on observations of more than 1000 hybrids, involving sev-eral combinations, has revealed that dwarfness, regular bearing and precocity

C.P.A. Iyer and R.J. Schnell74
are controlled by recessive genes (Sharma and Majumder, 1988a). Regularity of bearing appears to be linked with precocity. Characters contributing to biennial bearing are dominant over those governing regular bearing habit.
Flesh colour
Sharma (1987) considered that additive gene action may be involved in the inheritance of fl esh colour; however, studies involving monoembryonic ‘Alphonso’ and ‘Neelum’ have indicated that light yellow is dominant over orange-yellow (Iyer, 1991).
Skin colour
With regard to skin colour of fruit, Sharma (1987) observed that when red cultivars were crossed with green cultivars, the F1 seedlings exhibited vari-ous gradations of red. Iyer and Subramanyam (1987) also found a wide array of colours in the hybrids when the coloured monoembryonic ‘Janardhan Pasand’ was crossed with green-fruited cultivars, indicating that colour is mediated by a number of loci.
Flowering time
The fl owering response of mango cultivars in subtropical and tropical envi-ronments differs greatly (see Davenport, Chapter 5 this volume). Trees can be stimulated to fl ower under certain conditions in tropical environments using ethephon; however, this is ineffective in subtropical environments. Schnell and Knight (1998) investigated the repeatability of fl owering using eight cultivars over six harvest cycles (years), collecting data weekly. Three characters were evaluated: days to bloom (DTB) (from 1 November in each year), days in bloom (DIB), and days in bloom and fruit (DIBF). Signifi cant differences were detected for all three characters for both years and cultivars. Signifi cant differences were not detected for replicate trees within cultivars. Repeatability (R) of the fl ower phenology characters was high (R = 0.73, 0.88 and 0.77 for DTB, DIB and DIBF, respectively). This indicates that much of the variation is heritable and useful for extending fl owering times in subtropical environments.
Beak
The presence of a beak on mango fruit appears to be a dominant character since most of the hybrid plants had this feature when monoembryonic ‘Ban-galora’ (‘Totapuri’) was used as one of the parents in controlled crosses (Iyer and Subramanyam, 1979). Bunch bearing was found to be a dominant char-acter, as indicated in many crosses (Sharma et al., 1972) involving bunch-bearing types with single-fruited cultivars.

Breeding and Genetics 75
Disease resistance
Bacterial canker (Xanthomonas campestris pv. mangiferaeindicae) resistance appears to have cytoplasmic inheritance. Whenever ‘Neelum’, a susceptible cultivar, is used as the female parent, susceptibility is transmitted to all the hybrids, irrespective of the male parent (Sharma and Majumder, 1988a). It has been suggested that internal breakdown (spongy tissue) is mediated by recessive genes (Iyer, 1991). Susceptibility to ‘mango malformation’ appears to be dominant, since crosses involving resistant ‘Bhadauran’ did not yield any resistant hybrids (Sharma and Majumder, 1988a).
Other horticultural traits
The genetics of inheritance of various horticultural traits appears to be unclear. More knowledge will be forthcoming only when large-scale con-trolled hybridization experiments are undertaken at different mango research centres. However, the information now available for some of the characters is very useful in deciding which parental genotypes ought to be used in hybridization programmes.
Brettell et al. (2004) subjected the large number of mango hybrids obtained from the Australian National Mango Breeding Programme to a biometrical analysis. Their data indicated that many of the important fruit quality aspects, including fruit weight, fruit shape, ground skin colour, fruit width and pulp depth have high heritabilities and can therefore be readily selected in a breed-ing programme. Of particular interest is the observation that a high frequency of hybrids with a red or burgundy blush can be recovered from crosses where one parent has an intense red blush. Similarly, while the unique fl avour com-pounds associated with ‘Kensington Pride’ are also found in nearly 50% of the hybrids involving ‘Kensington Pride’, leaf fragrance was not found to be a reliable predictor of fruit fl avour.
4.5 Breeding Objectives
General objectives
Breeding objectives vary from region to region, depending on the specifi c trait(s) for which improvement is sought. However, they can be broadly gen-eralized to consist of the development of cultivars with: (i) regular bearing; (ii) dwarf tree habit; (iii) precocity; (iv) attractive, good sized (300–500 g), good quality fruits (appealing fl avour and fi rm fl esh without fi bres); (v) resistance to major diseases and pests; (vi) freedom from physiological dis-orders; and (vii) good shipping qualities and shelf life. While it would be hard to combine all these characteristics within a relatively short time, espe-cially resistance to all major diseases and pests, all of these characteristics are basic for commercial success.

C.P.A. Iyer and R.J. Schnell76
With regard to the improvement of rootstocks by breeding, the main desirable features are: (i) polyembryony; (ii) dwarfi ng; (iii) tolerance of adverse soil conditions (high pH, calcareous soil, etc.); and (iv) good scion-compatibility.
Specifi c objectives
In addition to improving general characters such as yield and quality, breed-ing has also been undertaken for certain specifi c purposes.
DwarfnessBecause of the obvious benefi ts of comparatively dwarf trees for orchard man-agement and fruit quality, attempts have been focused on obtaining hybrids with a dwarf tree framework. Breeding for dwarfness is important in mango, since a consistent dwarfi ng effect of any rootstock has not been established to date. The Indian cultivars that could be useful as a source for dwarfness include ‘Kerla Dwarf’, ‘Janardan Pasand’, ‘Manjeera’, ‘Amrapali’, ‘Creeping’ (Iyer and Subramanyam, 1986) and ‘Nileswar Dwarf’ (Singh, 1990).
Regular bearingThe causes of irregular bearing vary from region to region. In general, the main reason for alternate bearing, particularly in subtropical India, is the lack of initiation of vegetative growth soon after fruiting. However, two cul-tivars, ‘Neelum’ and ‘Bangalora’ (‘Totapuri’), which are regular bearers, have been extensively used as either of the parents in a hybridization programme to transfer the regular bearing habit to hybrids. ‘Neelum’ has been observed to be a good combiner and has contributed to the evolution of many regular-bearing Indian hybrid cultivars. However, ‘Bangalora’ is not a suitable par-ent since the hybrids possess very prominent beaks and their fruit quality is invariably poor. The regular bearing Florida cultivars (i.e. ‘Tommy Atkins’, ‘Keitt’, etc.) also have potential as parents.
Fruit colourMost of the commercial cultivars in South-east Asia possess green skin. Efforts are underway to produce new hybrid cultivars that retain the good qualities of these fruits together with attractive skin colour, so that they will occupy a better position in international trade. Since good skin colour has been shown to be transmissible to hybrids from suitably coloured parental cultivars, a number of cultivars with coloured skin are being used for hybrid-ization. In general, the attractively coloured Florida cultivars have been found to be suitable parents. In addition, there are several Indian cultivars (e.g. ‘Janardan Pasand’, ‘Suvarnarekha’, etc.) that would be suitable for use as parents for this purpose.
In Florida, the skin colour of the mango is an important factor and red skin is considered essential for mangoes shipped to northern markets. In the past, the evaluation of mango colour has been subjective and based on visual

Breeding and Genetics 77
ratings. Large errors are associated with these types of ratings, which makes evaluation based on fruit colour diffi cult. Ayala-Silva et al. (2005) used a colo-rimeter to quantify fruit colour, quality and differentiation among cultivars. Mango colour was measured with a Minolta Chroma Meter CR-400 (Osaka, Japan) portable tristimulus colorimeter and fruit chromaticity was recorded in Commission Internationale de l’Eclairage (CIE) L*, a* and b* colour space coordinates. In this system of colour representation the values L*, a* and b* describe a uniform three-dimensional CIE colour space. If the L*, a* and b* coordinates are known, then the colour is not only described, but also located in quantifi able space. Maternal half-sib families (MHS) of the mango culti-vars, ‘Keitt’, ‘Tommy Atkins’, ‘Tyler Premier’, ‘Mamita’, ‘White Alfonso’ and ‘Sandersha’ were evaluated along with two parental check clones, ‘Tommy Atkins’ and ‘Keitt’. Signifi cant differences were found for each of the L*, a* and b* colour space coordinates. Further work is underway to estimate the herita-bility of these traits to estimate their usefulness for breeding and selection.
Disease resistanceMANGO MALFORMATION. Although no breeding work has been reported that spe-cifi cally addresses disease or pest resistance/tolerance, cultivars are known to show varying degrees of susceptibility to biotic stress (see Ploetz and Freeman, Chapter 8, this volume ). Mango malformation, caused by Fusarium subglutin-ans, is a very serious disease that has threatened the very survival of the mango industry in many subtropical mango-growing regions. As there are no reliable cultural and chemical control measures available, breeding for resistance/tolerance has been attempted using ‘Bhadauran’ as the resistant parent; however, all of the F1 hybrids were susceptible to the disease (Sharma and Majumder, 1988a). In this respect, the observations of Ram et al. (1987) are very encouraging. Out of 102 cultivars screened, three of them, namely, ‘Bhydayam Dula’, ‘Samar Bahist Rampur’ and ‘Mian Sahib’, were free of malformation and could be tried as one of the parents in hybridization.
BACTERIAL CANKER. Bacterial canker is a serious problem with many cultivars. The only cultivar possessing true resistance to canker is ‘Bombay Green’ (Prakash and Srivastava, 1987) and hence could be a potential gene donor.
ANTHRACNOSE. Anthracnose, caused by Colletotrichum gloeosporioides Penz., is the most widespread disease in all mango-growing countries, manifesting itself in blossom blight, peduncle blight, leaf spot, twig blight, wither tip, fruit russetting and fruit rot. ‘Tommy Atkins’ is moderately tolerant of anthracnose and coupled with its other desirable qualities (i.e., regular bearing, fruit colour, etc.) should be a good parent in breeding programmes. In addition, ‘Parish’ and ‘Fairchild’ have been reported to be relatively resistant (Yee, 1958).
POWDERY MILDEW. Powdery mildew caused by Oidium mangiferae Berthet, has been reported to cause heavy loss of crops in years when RH is very high and accompanied by cool nights during fl owering. Cultivar differences with respect to susceptibility are recognized, and ‘Pairi’ (‘Raspuri’) is highly susceptible.

C.P.A. Iyer and R.J. Schnell78
Gupta (1976) has listed those cultivars that are most tolerant of this disease – ‘Neelum’, ‘Zardalu’, ‘Bangalora’, ‘Totapuri-Khurd’ and ‘Janardan Pasand’ – and hence could be valuable in breeding programmes.
PEST RESISTANCE. Considerable variation is also known to occur among mango cultivars with respect to their susceptibility to attack and injury by insect pests. Although no resistant genotypes have been reported for the mango hopper (Idiocerus spp.), the insect has been observed to avoid colonizing open plant types where free movement of wind is possible, an observation that could be useful in selection. Although complete resistance is not known to either fruit fl y (Bactrocera spp.) or seed weevil (Stenochetus mangiferae), variation in the degree of susceptibility has been reported (Iyer, 1991). Rossetto et al. (2006) observed that resistance to fruit fl y is compatible with fruit quality and pro-ductivity and advocated that resistance to fruit fl y should be one of the objec-tives of all mango breeding programmes. Their results also indicated that the main factors for resistance of mangoes to fruit fl ies lie in the fruit peel and not in the fruit pulp.
4.6 Methods of Breeding
Selection from open-pollinated seedlings
In India, almost all cultivars are selections that were made from naturally occur-ring open-pollinated seedlings. All of the Florida cultivars were selected from open-pollinated seedling progenies; none has come from a controlled breeding programme. Among the 64 Florida cultivars evaluated in the parentage analysis by Schnell et al. (2006), the genetic background was found to be based on as few as four Indian cultivars and the polyembryonic cultivar ‘Turpentine’. Two Indian cultivars, ‘Mulgoba’ and ‘Sandersha’, are in the background of most Florida types with ‘Amini’, ‘Bombay’, ‘Cambodiana’, ‘Long’, ‘Julie’ and ‘Nam Doc Mai’ making lesser contributions. In the parentage analysis ‘Turpentine 10’ was iden-tifi ed as a most probable paternal parent for ‘Haden’. The polyembryonic seed-ling races of Cuba and Florida were considered the same by Popenoe (1920) who called them the West Indian race (commonly known as ‘Turpentine’ in Florida). ‘Haden’ was reported as the maternal parent for ten cultivars included in the analysis, but based on the parentage analysis, 31 cultivars were found to have ‘Haden’ as one of the most likely parents. Likewise, the other important early Florida selection ‘Brooks’ is the parent of seven cultivars. ‘Haden, ‘Brooks’ and seedlings of ‘Haden’ and ‘Brooks’ have contributed disproportionately to the Florida group. In Florida, modern selection and breeding programmes for mango have focused on cultivars with exceptional production, red skin, disease resistance and extended shelf life. Methodology for crop improvement consists of collecting seeds from selected maternal parents with desired characteristics and growing them in close proximity to desirable male parents. Seedlings are screened by leaf aroma and horticultural traits, leading to a fi eld population of thousands of candidate seedlings (Campbell and Zill, 2006).

Breeding and Genetics 79
In the Canary Islands, Spain, breeding mainly involves selection of open-pollinated seedlings of ‘Lippens’. Lavi et al. (2004) indicated that mother trees should not be chosen entirely on the basis of their phenotypes and trees with inferior performance could also be included since progeny performance is quite unpredictable. They observed that most of the variance components of the agriculturally interesting traits are non-additive (Lavi et al., 1998) and most of these traits result from dominant and epistatic interactions.
The Israel mango-breeding programme therefore relies on open-pollination involving many mango cultivars from various parts of the world and screen-ing approximately 100 seedlings from each mother tree. The seedlings are grown on their own roots in the nursery for about a year and then planted in the fi eld at spacings of 2 × 6 m. Fruiting occurs after 3–6 years and fi rst selec-tion is carried out based on fi eld and laboratory data. Fruit characteristics at this stage are good skin colour, fruit weight of 400–600 g and high fruit qual-ity (good taste, absence of fi bres and small seed). Where a long harvest sea-son is desired both early and late harvest seedlings are selected and a general idea about shelf life of these seedlings is obtained. The second selection is carried out under commercial conditions by several experienced farmers using grafted plants. Plants that successfully pass this stage are planted in semi-commercial plots for a fi nal assessment before recommendation to farm-ers. The two selection stages are aimed at shortening the breeding programme and minimizing both the false negatives (loss of interesting seedlings which were not identifi ed) and the false positives (wrong identifi cation of interest-ing seedlings which should actually be rejected). New mango cultivars have also been selections made from open-pollinated seedlings. ‘Maya’ and ‘Nim-rod’ are seedlings of the same mother tree (Oppenheimer, 1967); ‘Tahar’ is a seedling of ‘Irwin’ (Slor and Gazit, 1982) and ‘Naomi’ is a seedling of ‘Palmer’ (Tomer et al., 1993). The other promising selections include ‘Shelly’ (late sea-son), ‘Tango’ (early season), ‘Selection 20/1’ (large fruit with aborted seeds) and ‘Selection 1/5’ (shiny red colour).
The South African mango-breeding programme also places a major emphasis on open-pollinated seedling selection. The screening of seedlings, besides undergoing tests for various plant and fruit characters, also includes shipping quality tests in which fruits are packed in 4 kg crates and stored at 11°C for 28 days to simulate shipping conditions. After storage the fruits are allowed to ripen at room temperature (25 ± 2°C) and screened thoroughly. Promising seedlings are again fi eld-tested with ‘Sabre’ as rootstock with a row of 40 trees along with ten trees of commercial varieties. Two cultivars, ‘Joa’ (‘Palmer’ seedling) and ‘Chene’ (‘Kent’ seedling), were released from the breeding programme in 1996. Another high yielding selection ‘A2-CD28’ (‘Fascell’ seedling) is a midseason clone with an attractive pink blush (Human et al., 2006) and has been recommended for Plant Breeders’ Rights in 2005. The other promising selections are ‘Osteen’ (‘Haden’ seedling) and ‘Neldica’ (‘Palmer’ seedling).
The 'R2E2' cultivar developed in Australia is a seedling progeny of the Florida cultivar 'Kent', and is now the second most popular mango grown in Australia.

C.P.A. Iyer and R.J. Schnell80
Controlled pollination
Hand pollinationThe traditional, cumbersome method involving the continued pollination of fl owers on a panicle over several days when the fl owers are open has now been replaced in India with more effi cient methods. The current method involves the pollination of a limited number of fl owers per panicle (maxi-mum of ten), utilizing a larger number of panicles since it is very rare that a panicle bears more than one fruit to maturity. Using this method, fruit set as high as 3.85% can be achieved compared to the 0.23–1.57% effi ciency involv-ing other methods (Mukherjee et al., 1968; Singh et al., 1980).
CagingThe enclosure of two desirable parents of synchronous fl owering in a screen house with pollinating insects provides a more practical method of hybridiza-tion. This method has been used in Israel (Degani et al., 1993), Brazil (Pinto and Byrne, 1993) and South Africa (Cilliers et al., 1996). A standardized caging technique for mango breeding was previously used in India following the discovery of self-incompatibility in some of the most popular commercial culti-vars (Sharma and Singh, 1970). This procedure involves the planting of self-in-compatible (female) and male parents in specially prepared breeding plots, and are enclosed in an insect-proof cage into which freshly reared housefl ies, or any other suitable pollinator, are introduced to effect cross-pollination (Sharma et al., 1972).
Polycross matingA polycross is simply the use of a number of advanced selections or current commercial clones planted in a design that maximizes the chance for cross-pollination. The polycross design has been extensively used in sugarcane breeding where small fl ower size and low numbers of seedlings per cross make controlled pollinations diffi cult. At USDA Subtropical Horti-culture Research Station (SHRS) in Miami the clones ‘Haden’, ‘Tommy Atkins’, ‘Kent’, ‘Keitt’ and ‘Nam Doc Mai’ have been used to produce new seedlings for selection. Five clones of each genotype were planted, with at least one plant of each genotype next to all other genotypes. Over 1000 seed-lings from known mother trees are planted as maternal half-sib families. The pollen parent of superior selections is determined using microsatellite markers.
4.7 Handling of Hybrid Populations and Selection
Criteria for initial selection
Primary selection from the hybrid progeny is based on: (i) precocity; (ii) fruit size and shape; (iii) skin colour; (iv) fruit characteristics (high pulp to stone ratio and freedom from fi bre and physiological disorders); and (v) fruit qual-ity. Following this preliminary evaluation, selected hybrids are retained for

Breeding and Genetics 81
further screening. It is important to graft the hybrids onto proper rootstocks as early as possible, as grafted plants are precocious. At least ten grafted plants of each selected hybrid are used in the fi nal selection, which is based on yield, regularity in bearing and response to diseases and pests, in addition to other desirable fruit characters. At least 3 consecutive years’ performance data should be collected before deciding on their suitability for release as new cultivars.
Pre-selection
Trees have a long juvenile phase, and the development of pre-selection meth-ods is important for discarding inferior seedlings at a very early stage, obvi-ating the need for maintaining a large number of seedlings for long periods. This can save time, land and labour. Leaf fl avour has been reported to be directly correlated with fruit fl avour (Majumder et al., 1972; Whiley et al., 1993). Emergence of new growth fl ushes, simultaneously with fruiting or immediately after harvest, is indicative of regular bearing (Sharma et al., 1972). A higher phloem to xylem ratio, associated with dwarfi ng, has been used effectively as a pre-selection criterion. Genotypes in which the ratio exceeds 1.0 are least vigorous, those with a ratio between 0.6 and 1.0 are of medium vigour and those with a ratio of less than 0.6 are most vigorous (Kurian and Iyer, 1992). In addition, higher levels of phenolics in the apical bud is associated with reduced vigour and dwarfi ng (Iyer, 1991). Although Majumder et al. (1981) indicated that low stomatal density is an indicator of dwarfness this has not been confi rmed by other workers (Iyer, 1991). Regular bearing mango cultivars have low polyphenol oxidase (PPO) activity (cate-cholase and cresolase) compared to alternate bearers (Sharma, 2003). Sharma et al. (2000) observed that a strong positive correlation existed between the incidence of fl oral malformation and both enzyme activity (catecholase and cresolase) and phenolic content and speculated that PPO activity can be used as a biochemical index for screening mango germplasm against malforma-tion disease.
Potential for marker assisted selection (MAS)
More than 65 microsatellite markers have been developed for mango and these are easily used to verify parentage using a software package such as cervus (Marshall et al., 1998). When caging trees or using the polycross mat-ing design it is possible to identify the male parent from a set of potential male parents. This has been useful in cacao breeding where mistakes in pol-lination have lead to the estimation of unreliable breeding values for parental clones. The development of linkage maps and identifi cation of quantitative trait loci (QTL) for productivity and quality traits has led to a very successful MAS in cacao (Schnell et al., 2007). This could serve as a model for future mango breeding and selection efforts.

C.P.A. Iyer and R.J. Schnell82
Molecular markers
Molecular markers can be used for estimating genetic relationships among clones, for parentage analysis and for the development of a saturated linkage map. Isozymes were the fi rst markers to be used for fi ngerprinting mango cultivars, to determine self- versus cross-pollination and to estimate genetic relationships (Degani et al., 1990; Knight and Schnell, 1994). RAPD markers were also used to fi ngerprint cultivars and estimate genetic relationships in mango (Schnell et al., 1995). A group of ‘Haden’ seedlings and a random group of seedlings were evaluated using 11 RAPD primers. This study sup-ported the ‘Haden’ parentage of ‘Eldon’, ‘Lippens’, ‘Tommy Atkins’ and ‘Zill’; however, the parentage of ‘Glenn’ and ‘Osteen’ was questioned. Adato et al. (1995) used DNA fi ngerprinting (DFP) to evaluate genetic relationships between 26 mango cultivars and 14 rootstocks. They provided a pedigree that further confi rmed the relationship between many of the ‘Haden’ seed-lings. Lopez-Valenzuela et al. (1997) used RAPD markers to estimate genetic diversity among 15 rootstock cultivars using 13 markers, and identifi ed a specifi c RAPD band associated only with the polyembryonic types. Eiad-thong et al. (1999) utilized anchored simple sequence repeat markers to anal-yse 22 mango cultivars; they were able to distinguish genotypes, but were unable to fi nd markers unique to either monoembryonic or polyembryonic types, or for the Thai cultivars selected for green harvest (crispy mango) from the cultivars selected for ripe fruit production. Kashkush et al. (2001) utilized amplifi ed fragment length polymorphisms (AFLP) to estimate genetic rela-tionships between 16 cultivars and seven rootstock cultivars. They also anal-ysed 29 progeny from a cross of ‘Tommy-Atkins’ and ‘Keitt’ and produced a crude linkage map that identifi ed 13 of the 20 linkage groups.
Viruel et al. (2005) developed the fi rst reported set of 16 microsatellite markers for mango, of which 14 produced the expected one or two amplifi -cation products per genotype. These 14 microsatellites were used to evaluate 28 mango genotypes that included 14 Florida cultivars. Discrimination of all 28 genotypes was possible and the average number of alleles per locus was 5.3. Previously known pedigree information for the ‘Haden’ family of man-goes was confi rmed and was in agreement with previously published RAPD and DFP analyses (Adato et al., 1995; Schnell et al., 1995) with one exception. Viruel’s clone of ‘Zill’ was not resolved as a seedling of ‘Haden’. Schnell et al. (2005) developed a second set of 15 microsatellite markers and analysed 59 Florida cultivars and four related species. Two of the microsatellites were monomorphic among the Florida cultivars; the other 13 had an average num-ber of alleles per locus of 4.2 with polymorphism information content (PIC) values varying from 0.21 to 0.63.
Schnell et al. (2006) used 25 microsatellite loci to estimate genetic diversity among 203 unique mangoes (M. indica L.), two M. griffi thii Hook. f. and three M. odorata Griff. accessions maintained at the National Germplasm Repository (NGR) and by Fairchild Tropical Botanic Garden (FTBG) in Miami, Florida. The 25 microsatellite loci had an average of 6.96 alleles per locus and an average PIC value of 0.552 for the M. indica population. The total

Breeding and Genetics 83
propagation error in the collection (i.e. plants that had been incorrectly labelled or grafted) was estimated to be 6.13%. When compared by origin, the Florida cultivars were more closely related to Indian than to South-east Asian cultivars. Unbiased gene diversity (Hnb) of 0.600 and 0.582 was found for Indian and South-east Asian cultivars, respectively, and both were higher than Hnb among Florida cultivars (0.538). When compared by horticultural type, Hnb was higher among the polyembryonic types (0.596) than in the monoembryonic types (0.571).
To date 63 microsatellite markers have been developed for mango (Duval et al., 2005; Honsho et al., 2005; Schnell et al., 2005; Viruel et al., 2005). This number is more than adequate for genetic diversity studies and for par-entage analysis as has been demonstrated by Schnell et al. (2006); however, it is not enough to develop a saturated linkage map for the 20 linkage groups of mango. Developing an additional 200 microsatellite or single nucleotide polymorphic markers is a major objective of the USDA Agriculture Research Service (ARS) programme in Miami over the next 2 years. Three experimen-tal populations have been developed and planted in the fi eld as mapping populations. The fi rst population is an F2 population derived from self-polli-nation of ‘Tommy Atkins’ consisting of 168 seedlings that was planted in the fi eld in 1995. The second population is an F2 population derived from self-pollination of ‘Haden’. A total of 224 seedlings from a single isolated ‘Haden’ tree have been in the fi eld for 3 years. Phenotypic data collection is in prog-ress for both of these populations. The development of a saturated linkage map and the identifi cation of QTL for important traits are objectives for the USDA-ARS programme in Miami for the next 5 years.
4.8 Minimizing Problems in Breeding
Heavy fruit drop
Heavy fruit drop ultimately results in few hybrid fruits, despite the large number of fl owers used for cross-pollination. While many recommendations are available to minimize mango fruit drop with growth regulators, these have not been very useful in breeding programmes where the number of fl owers remaining in a panicle is very low. Iyer and Subramanyam (1972) suggested that embryo culture could be used to rescue hybrid embryos, and Sahijram et al. (2005) developed in-vitro techniques to rescue immature mango embryos from controlled crosses and recovered hybrid plants.
Long juvenile phase
Normally, mango seedlings require 3–10 years to fl ower, thereby prolonging the breeding programme. Grafting individual hybrids on the proper rootstocks at the earliest possible stage and growing them in a location where climatic stress (particularly cold weather) prevails, induces precocious

C.P.A. Iyer and R.J. Schnell84
fl owering. Iyer (1991) has reported signifi cant differences between seedlings on their own roots and grafted plants of the same genotype with respect to fruit size, quality and even colour in the early years. However, different results have been reported in a recent study of the effect of rootstocks on the performance of seedling scions with respect to ten horticultural traits (Lahav et al., 1995). No difference of practical importance was found between the original seedlings and their grafted duplicates.
Singh (1969) has suggested that young mango seedlings can be induced to fl ower and fruit if they are grafted onto comparable shoots of a bearing tree (a few days before fl owering). The scions are defoliated and girdled. Using this technique, it has been reported that the fruit characteristics of F1 hybrids can be determined within 2 years and F2 hybrids within 4 years, thus eliminating at least 10 years from the period required to raise and evaluate F2 populations (Singh, 1963).
Ethephon (Chacko et al., 1974) and paclobutrazol (Anonymous, 1984) have fl ower-inducing properties in mango and could also be utilized for shortening the juvenile phase. However, they must be used with caution since chemical induction of fl owering can alter fruit size and this could lead to errors in judgement when making selections within the hybrid progeny.
Polyembryony
Seeds of polyembryonic mango cultivars characteristically contain several nucellar embryos, and may also contain a zygotic embryo. While nucellar seedlings are preferred as rootstocks for mango because of their uniformity, the breeder, on the other hand, is generally interested in sexual seedlings for the selection of improved rootstocks. Until recently, crosses involving poly-embryonic cultivars as the maternal parents were generally not performed, since reliable methods for identifying zygotic seedlings were not available. The use of polymorphic enzyme systems (isozymes) (Degani et al., 1990, 1992) to identify zygotic seedlings (Schnell and Knight, 1992; Truscott, 1992; Degani et al., 1993) is based on the fact that nucellar seedlings should have the same isozyme alleles as the maternal parent. A variation at a locus coding for an enzyme indicates that the plant has originated by sexual reproduction. Zygotic seedlings arising from self-pollination are distinguished from nucel-lar seedlings by being homozygous at one or more loci at which the female parent is heterozygous. Statistically, when three or four heterozygous loci are examined, up to 88 or 94%, respectively, of the selfed zygotic seedlings are identifi able (Moore and Castle, 1988). Cross-pollination by another cultivar of the same genotype is equivalent to self-pollination. Zygotic seedlings from cross-pollination are distinguishable from those resulting from self-pollination if they express an allele not carried by the female parent.
The frequency of occurrence of zygotic seedlings varies among the poly-embryonic mango cultivars, i.e. 22% with ‘13-1’ (Degani et al., 1993), 20 and 24% with ‘Turpentine’ (Degani et al., 1993 and Schnell and Knight, 1992, respectively), 2 and 4% with ‘Sabre’ (Truscott, 1992 and Schnell and Knight,

Breeding and Genetics 85
1992, respectively) and 36 and 64% with ‘Madu’ and ‘Golek’, respectively (Schnell and Knight, 1992).
4.9 Achievements of Conventional Breeding
Despite the many problems associated with mango breeding for cultivar devel-opment, many useful hybrids have been released. The earliest attempts were probably made in the West Indies to combine the good qualities of the Indian mango with the indigenous types by controlled pollination (Brooks, 1912).
India
Intervarietal hybridization in India has resulted in the release of many culti-vars. The work at Sabour initially yielded two promising hybrids: ‘Mahmood Bahar’ and ‘Probashanker’, both combinations of ‘Bombay’ and ‘Kalapady’ (Roy et al., 1956). Subsequently, four more hybrids have been developed. These are: ‘Sundar Langra’ (‘Sardar Pasand’ × ‘Langra’) having ‘Langra’ quality and regular bearing habit; ‘Alfazli’ (‘Alphonso’ × ‘Fazli’) with ‘Fazli’ quality and early ripening; ‘Sabri’ (‘Gulabkhas’ × ‘Bombai’) having ‘Bombai’ fruit shape and colour of ‘Gulabkhas’ with regular bearing habit; and ‘Jawa-har’ (‘Gulabkhas’ × ‘Mahmood Bahar’) having high pulp and early bearing habit (Hoda and Ramkumar, 1993). Developed in Kodur, the hybrid ‘Swarna-jehangir’, combining the high quality of ‘Jehangir’ and the attractive colour of ‘Chinnaswarnarekha’, is a prolifi c bearer and is the best of all hybrids devel-oped at this centre. The other hybrids released from Kodur are ‘Neeludin’ (‘Neelum’ × ‘Himayuddin’), ‘Neelgoa’ (‘Neelum’ × ‘Yerra Mulgoa’) and ‘Neeleshan’ (‘Neelum’ × ‘Baneshan’).
Two excellent, regular-bearing hybrids, ‘Mallika’ and ‘Amrapali’, were developed and released by the Indian Agricultural Research Institute (IARI), New Delhi (Singh et al., 1972). ‘Mallika’ is a hybrid between ‘Neelum’ and ‘Dashehari’ with a high total soluble solids (TSS) content, a higher percentage of pulp, fi breless fl esh and a fruit size of about 300 g. ‘Amrapali’ (‘Dashehari’ × ‘Nee-lum’) is precocious, distinctly dwarf and hence amenable to high-density planting, a regular bearer with excellent quality and is also very rich in vitamin A. Recently, two more cultivars, ‘Arunima’ (‘Amrapali’ × ‘Sensation’) and ‘Pusa Surya’ (a selection from ‘Eldon’) have been released from the IARI. A promising mango hybrid ‘Ambika’, a cross between ‘Amrapali’ and ‘Janardhan Pasand’, having a yellow colour with red blush, fi rm fl esh and scanty fi bre was released from the Central Institute of Sub-Tropical Horticulture, Lucknow.
Four hybrid cultivars were released from the Indian Institute of Horticul-tural Research in Bangalore: ‘Arka Aruna’ (‘Banganapalli’ × ‘Alphonso’), ‘Arka Puneet’ (‘Alphonso’ × ‘Banganapalli’), ‘Arka Anmol’ (‘Alphonso’ × ‘Janardhan Pasand’) and ‘Arka Neelkiran’ (‘Alphonso’ × ‘Neelum’). ‘Arka Aruna’ is dwarf, and large fruited with a high percentage of pulp and high TSS content. It is ideal for homesteads. ‘Arka Puneet’ is very similar to

C.P.A. Iyer and R.J. Schnell86
‘Alphonso’ but free of ‘spongy tissue’, has a good shelf life and is not suscep-tible to fruit fl y attack. ‘Arka Anmol’ is a heavy bearer with good keeping quality (Iyer and Subramanyam, 1993). ‘Arka Neelkiran’ is free of spongy tissue and has excellent skin colour.
‘Ratna’ is a cross between ‘Alphonso’ and ‘Neelum’ that was carried out at the Fruit Research Station, Vengurla, Maharashtra; it has a larger fruit size, fruit quality similar to ‘Alphonso’ and is free of ‘spongy tissue’ (Salvi and Gunjate, 1988). A parthenocarpic mango cultivar, ‘Sindhu’, has been devel-oped at this station as a result of back-crossing ‘Ratna’ with ‘Alphonso’ (Gun-jate and Burondkar, 1993).
Two hybrid cultivars were released from the Fruit Research Station in Sangareddy, Andhra Pradesh. ‘Au-Rumani’ (‘Rumani’ × ‘Mulgoa’) is a regular and prolifi c bearer with fi breless fl esh. ‘Manjira’ (‘Rumani’ × ‘Neelum’) is a dwarf, regular and prolifi c bearer with good quality fruits.
The Paria Research Station in Gujarat developed three mango hybrids, ‘Neelphonso’ (‘Neelum’ × ‘Alphonso’), ‘Neeleshan Gujarat’ (‘Neelum’ × ‘Bane-shan’) and ‘Neeleshwar’ (‘Neelum’ × ‘Dashehari’). These hybrids are supe-rior in TSS, total sugars and vitamin C, in addition to their dwarfi ng habit, with respect to their parents (Sachan et al., 1988).
Other countries
USAMango hybridization was reported from Hawaii in the 1920s, but no out-standing problem appears to have been addressed or solved (Pope, 1929). A number of crosses have been reported in Florida (Young and Ledin, 1954; Sturrock, 1969), but all of the Florida cultivars are chance seedlings and none came from controlled pollinations.
IsraelThere is an extensive breeding programme in Israel aimed at producing higher yielding cultivars with good quality, attractive fruit and with longer harvest periods. Several hundred seedlings from open and controlled polli-nations have been evaluated, and 14 of them have been identifi ed as being of interest (Lavi et al., 1993). The rootstock breeding programme is aimed at developing rootstocks resistant to or tolerant of soil stresses, i.e. calcareous soils, saline irrigation water and heavy non-aerated soils that predominate in the mango-growing regions of Israel. Several interesting mono embryonic and polyembryonic rootstocks have been selected (Lavi et al., 1993), but none has performed better than ‘13-1’, the currently preferred rootstock in Israel (Gazit and Kadman, 1980).
AustraliaA breeding programme to develop a new cultivar which retains the charac-teristic fl avour of ‘Kensington’, but with improved productivity, greater dis-ease resistance, enhanced skin colour and better postharvest performance,

Breeding and Genetics 87
was initiated in Queensland, Australia. These features are found in many Florida cultivars (i.e. ‘Irwin’, ‘Sensation’ and ‘Tommy Atkins’) which are being used as maternal parents in crosses with ‘Kensington’ (Whiley et al., 1993). Promising hybrids have been identifi ed in crosses involving ‘Sensa-tion’, for example ‘Calypso’™ (see Knight et al., Chapter 3, this volume). ‘Calypso’™ has increased shelf life, firmer fruit, extra blush for cosmetic appeal, a higher flesh-to-seed ratio and consistent yields of high-quality fruit. The Australian mango breeding programme was strengthened since 1994 by launching a major effort involving various organizations located in different agro-climatic zones in hybrid production, as well as regional testing.
BrazilBreeding has been initiated in the tropical savannah of Brazil to develop cul-tivars that are dwarf and with good quality fruit. Hybridizations have involved local, Indian and Florida cultivars. ‘Amrapali’ and ‘Imperial’ were good male parents to confer dwarfi ng in the progeny (Pinto and Byrne, 1993). Out of 2088 seedlings in the fi eld, 209 seedlings were selected in the fi rst year and 42 of these were later identifi ed as promising, from which four have been released as new cultivars (Pinto et al., 2004). These four are: ‘Alfa’ (‘Mal-lika’ × ‘Van Dyke’), which is semi-dwarf, high yielding and regular bearing; ‘Beta’ (‘Amrapali’ × ‘Winter’), high yielding and moderately resistant to anthracnose and Oidium; ‘Roxa’ (‘Amrapali’ × ‘Tommy Atkins’), with excel-lent fruit quality; and ‘Lita’ (‘Amrapali’ × ‘Tommy Atkins’), high yielding with excellent fruit quality.
South AfricaThe South African breeding programme at the Citrus and Subtropical Fruit Research Institute (CSFRI) is based on introductions, open-pollination and mass selection. Four new cultivars have been released: ‘Heidi’, ‘Neldawn’, ‘Neldica’ and ‘Ceriese’. In addition, 12 promising selections have been iden-tifi ed for further evaluation (Marais, 1992).
4.10 Mutations
Somatic mutations
Asexual propagation enables the preservation of accumulated mutations (macro and micro), which would normally be eliminated during sexual propogation. In many fruit crops, bud mutations and chimeras occur rather frequently and can provide an additional source of variability for selection. However, such reported instances are relatively few in mango. Roy and Visweswariya (1951) observed mutants of ‘Puthi’ in which the number of palisade cell layers differed from the original cultivar. Naik (1948) observed signifi cant variation among trees of the same clone with respect to fruit shape, size, colour and quality, which was ascribed to bud mutations. ‘Davis Haden’, a sport of ‘Haden’, is larger than ‘Haden’ and its season of maturity is about

C.P.A. Iyer and R.J. Schnell88
a month earlier (Young and Ledin, 1954). ‘Rosica’ from Peru, is a bud mutant of ‘Rosado de lca’. Unlike its parent, ‘Rosica’ is high yielding and regular bearing, and does not produce seedless fruits (Medina, 1977).
Oppenheimer (1956), after a survey of many orchards in India, reported wide variability in the performance of trees of the same clone within a single orchard. Mukherjee et al. (1983) conducted a survey of mangoes in eastern India and identifi ed some superior clones. Singh and Chadha (1981), in a study of orchards of ‘Dashehari’, located four clones which were superior in performance. Singh et al. (1985) isolated two high-yielding clones from orchards of ‘Langra’. Within ‘Kensington’, strains have also been identifi ed that show improved resistance to bacterial black spot (Whiley et al., 1993).
Roy (1950) observed a mutant of ‘Alphonso’ with respect to fruit shape, and suspected it to be a mericlinal chimera. Pandey (1998) has described seven clones of ‘Alphonso’: ‘Alphonso Behat’ and ‘Alphonso Bihar’ from Bihar, ‘Alphonso Batli’, ‘Alphonso Black’ and ‘Alphonso Bombay’ from Maharashtra, ‘Alphonso Punjab’ from Punjab and ‘Alphonso White’ or ‘Bili Ishada’ from the North Canara district of Karnataka. Rajput et al. (1996) assembled several ‘Dashehari’ variants and after 14 years of observation, reported that the clone ‘Dashehari 51’ was superior with respect to yield and regular bearing. Other somatic mutants include: ‘Cardozo Mankurad’ with large fruits of attractive colour and high yields from ‘Mankurad’ of Goa; dwarf selections from the ‘Rumani’ and ‘Bangalora’ (Ramaswamy, 1989); development of ‘Paiyur’, a dwarf selection from ‘Neelum’ (Vijaya Kumar et al., 1991); ‘Rati Bangana-palli’ and ‘Nuzuvid’ from ‘Banganapalli’ (Anonymous, 1999); and ‘MA-1’, regular bearing and high yielding with resistance to ‘spongy tissue’ from ‘Alphonso’ (Mukunda, 2003).
In Thailand, Chaikiattiyos et al. (2000) selected clone ‘SKoo7’ (now known as ‘Kaew Sisaket’) from 320 ‘Kaew’ plants; ‘SKoo7’ has higher yield and superior quality. Jintanawongse et al. (1999) also made superior selections for yield and fruit quality from ‘Nam Dok Mai’, ‘Khiew Sawoey’, ‘Rad’ and ‘Nang Klang Wau’ and DNA fi ngerprints of all these clones were made for comparison with the parental clone.
For these studies, it is important to conduct a replicated cultivar evalua-tion trial against standard commercial cultivars to establish that these varia-tions are stable and not due to environmental responses. The use of genetic markers should be explored to confi rm that the new clones are genetically distinct from the original cultivar.
Induced mutations
Mutation induction using ionizing radiation was attempted by Siddiqui et al. (1966). Siddiqui (1985) irradiated dormant buds of ‘Langra’ with high doses of rays, and grafted them onto 1-year-old seedlings. A bud graft exposed to 3.0 kR bore fruits which were heavier, larger and had a more cream-yellow pulp than the control. This variability was stable over three seasons. Sharma and Majumder (1988b) irradiated bud sticks, topworked them onto 10-year-old

Breeding and Genetics 89
seedlings, and found that dosages above 5 kR are lethal for mango and that the lethal dose required for 50% mortality (LD50) lies between 2 and 4 kR. Effective dosages of the chemical mutagens, ethane methyl sulfonate (EMS) and N-nitroso methyl urea (NMU), were 1.5 and 0.05%, respectively. The spectrum of mutations induced by physical and chemical mutagens was observed to be more or less the same, indicating the high sensitivity of certain loci. The mutants included dwarfness, changes in shape and serration of leaves and in TSS content in ‘Dashehari’. As in other perennial crops, muta-genesis techniques that can allow useful traits to be targeted, as well as isolating mutated sectors from a chimera, are essential.
4.11 Breeding Potential of Wild Species
Bompard (1993; see Bompard, Chapter 2, this volume) has made a compre-hensive study of the wild Mangifera species and enumerated their potential use in breeding. Mangifera laurina, which has subglabrous and laxly fl owered panicles and is well adapted to areas with perpetual wet climates, is resistant to anthracnose. Mangifera orophila from Malaysia and M. dongnaiensis from Vietnam are both restricted to mountain forests 1000–1700 m above sea level and their hybrids with mango could extend cultivation into temperate zones.
Mangifera magnifi ca is completely free of fi bres; M. rufocostata and M.swintonioides have an off-season bearing habit; M. pajang (endemic to Borneo) and some strains of M. foetida have good quality fruits. Similarly ‘Wani’ from Bali and Borneo, the best variety of M. caesia, has a distinctive taste. Mangiferacasturi from South Kalimantan is a prolifi c bearer with small, black, sweet fruits having good potential. Mangifera altissima is reportedly resistant to mango pests, such as hoppers, tip borers and seed borers (Angeles, 1991). Sharma and Choudhury (1976) observed that wild Mangifera trees identifi ed in Tripura State (north-eastern India) were free of mango malformation. The wild species could also contribute to higher productivity. Fairchild (1948) observed that crosses between fi ve-stamened mango and the Indian mango (only one fertile stamen) could produce hybrids having better pollinating quality. The interspecifi c compatibility of these species with M. indica must be verifi ed before they can be utilized in hybridization programmes (as sug-gested by Bompard, 1993; Kostermans and Bompard, 1993; see Bompard, Chapter 2, this volume).
4.12 Conclusions
Until recently, all mango cultivars arose as chance seedlings or as seedling selections from known mother trees. Enthusiasm for controlled hybridiza-tion by means of hand pollination waned because of the tedious nature of the task and heavy fruit drop, resulting in only very few hybrids. This low hybrid population was inadequate for selection and hence not many outstanding hybrids were obtained. However, improvements in pollinating techniques

C.P.A. Iyer and R.J. Schnell90
and more rapid screening of hybrid populations have enabled the release of many hybrid mango cultivars of commercial value. Because of the world market’s demand for mangoes with specifi c qualities, the synthesis of new cultivars has become imperative. Rapid strides in molecular biology and in other aspects of biotechnology have opened up new approaches in plant breeding. The development of polymerase chain reaction (PCR)-based genetic markers, specifi cally microsatellites, and their application to classical breeding offer tremendous potential for mango improvement. The develop-ment of a saturated linkage map and the identifi cation of QTL for important traits will allow the implementation of a MAS programme. The introduction of specifi c genes for disease resistance from cultivars and wild species into popular cultivars should soon be a reality. Without resorting to these new technologies, mango breeding will continue to be a slow process.
References
Adato, A., Sharon, D. and Lavi, U. (1995) Application of DNA fi ngerprints for identifi ca-tion and genetic analyses of mango (Mangifera indica) genotypes. Journal of the American Society for Horticultural Science 120, 259–264.
Angeles, D.E. (1991) Mangifera altissima. In: Verheij, E.W.M. and Coronel, R.E. (eds) Plant Resources of South-East Asia No.2: Edible Fruits and Nuts. Pudoc-DLO, Wageningen, the Netherlands, pp. 206–207.
Anonymous (1984) Paclobutrazol, Plant Growth Regulators for Fruits. ICI, Fernhurst, West Sussex, UK.
Anonymous (1999) Improvement and management of horticultural crops. Department of Agricultural Research and Education (DARE)/Indian Council of Agricultural Re-search (ICAR) Annual Report 1989–99. ICAR, New Delhi, India, pp. 44–60.
Aron, Y., Czosnek, H. and Gazit, S. (1998) Polyembryony in mango is controlled by a single dominant gene. HortScience 33, 1241–1242.
Ayala-Silva, T., Schnell, R.J., Meerow, A.W., Winterstein, M., Cervantes, C. and Brown, J.S. (2005) Determination of color and fruit traits of half-sib families of mango (Mangifera indica L.). Proceedings of the Florida State Horticultural Society 118, 253–257.
Bompard, J.M. (1993) The genus Mangifera rediscovered: the potential contribution of wild species to mango cultivation. Acta Horticulturae 341, 69–77.
Brettell, R.I.S., Johnson, P.R., Kulkarni, V.J., Muller, W. and Bally, I.S.E. (2004) Inheri-tance of fruit characters in hybrid mangoes produced through controlled pollina-tion. Acta Horticulturae 645, 319–326.
Brooks, A.J. (1912) Artifi cial cross fertilization of the mango. West Indies Bulletin 12, 567–569.
Campbell, R.J. and Campbell, C.W. (1993) Commercial Florida mango cultivars. Acta Horticulturae 341, 55–59.
Campbell, R.J. and Zill, G. (2006) Mango selection and breeding for alternative markets and uses. (Abstract). The Eighth International Mango Symposium, 5–10 February, Sun City, South Africa, p. 19.
Chacko, E.K., Kohli, R.R., Doreswamy, R. and Randhawa, G.S. (1974) Effect of 2-chloroethyl phosphonic acid on fl ower induction in juvenile mango seedlings. Physiologia Plantarum 32, 188–190.

Breeding and Genetics 91
Chaikiattiyos, S., Kurubunjerdjit, R., Akkaravessapong, P., Rattananukul, S., Chueychum, P. and Anuput, P. (2000) Improvement and evaluation of the selected ‘Kaew Sisaket’ mango in Thailand. Acta Horticulturae 509, 185–192.
Cilliers, B., Human, C.F., Snyman, J.C. and Carstens, K. (1996) Strategies, progress and results from the South African mango breeding programme. Acta Horticulturae445, 241–244.
Dag, A., Eisenstein, D., Degani, C., El-Batsri, R., Zelig, M., Ziv, G. and Gazit, S. (1997) ‘Tommy Atkins’ mango as pollenizer for ‘Lily’. Acta Horticulturae 455, 209–216.
Dag, A., Degani, C. and Gazit, S. (2006) Gene fl ow in mango orchards and its impact on yield. (Abstract). The Eighth International Mango Symposium, 5–10 February, Sun City, South Africa, p. 23.
Degani, C., El-Batsri, R. and Gazit, S. (1990) Enzyme polymorphism in mango. Journalof the American Society for Horticultural Science 115, 844–847.
Degani, C., Cohen, M., El-Batsri, R. and Gazit, S. (1992) PGI isozyme diversity and its genetic control in mango. HortScience 27, 252–254.
Degani, C., Cohen, M., Reuvani, O., El-Batsri, R. and Gazit, S. (1993) Frequency and characteristic of zygotic seedlings from polyembryonic mango cultivars, deter-mined using isozymes as genetic markers. Acta Horticulturae 341, 78–85.
Dijkman, N.J. and Soule, M.J., Jr (1951) A tentative method of mango selection. Pro-ceedings of the Florida State Horticultural Society 64, 257.
Duval, M., Bunel, F.J., Sitbon, C. and Risterucci, A.M. (2005) Development of microsatellite markers for mango (Mangifera indica L.). Molecular Ecology Notes 5, 823–826.
Duval, M., Bunel, J., Sitbon, C., Risterucci, A.M., Calcabre, C. and Le Bellec, F. (2006) Genetic diversity of Caribbean mangoes using microsatellite markers. (Abstract). The Eighth International Mango Symposium, 5–10 February, Sun City, South Africa, p. 29.
Eiadthong, W., Yonemori, K., Sugiura, A., Utsunomiya, N. and Subhadrabandhu, S. (1999) Identifi cation of mango cultivars of Thailand and evaluation of their genetic variation using the amplifi ed fragments by simple sequence repeat-(SSR-) anchored primers. Scientia Horticulturae 82, 57–66.
Fairchild, D. (1948) The mango relative of Cochin China: those with fi ve stamen fl owers. Proceedings of the Florida State Horticultural Society 61, 250–255.
Florida Mango Forum (1951) Mango Studies. Florida Mango Forum, Stuart, Florida.Galan Sauco, V. (1997) Mango world production (outside Israel, Egypt and India). Acta
Horticulturae 455, 15–22.Gazit, S. and Kadman, A. (1980) ‘13-1’ mango rootstock selection. HortScience 15, 669.Gunjate, R.T. and Burondkar, M.M. (1993) Parthenocarpic mango developed through
hybridization. Acta Horticulturae 341, 107–111.Gupta, J.H. (1976) Reaction of mango varieties to powdery mildew (Oidium mangiferae
Berthet) in Uttar Pradesh. Progressive Horticulture 8, 63–64.Hoda, M.N. and Ramkumar (1993) Improvement of mango. In: Proceedings of the Na-
tional Seminar on Irregular Bearing in Mango – Problems and Strategies (12–13 July 1991). Kamal Printing Press, Muzafarpur, Bihar, India, pp. 34–35.
Honsho, C., Nishiyama, K., Eiadthong, W. and Yonemori, K. (2005) Isolation and char-acterisation of new microsatellite markers in mango. Molecular Ecology Notes 5, 152–154.
Human, C.F., Rheeder, S. and Sippel, A.D. (2006) New cultivars and hybrid selections from the mango breeding programme of the Agricultural Research Council in South Africa. (Abstract). The Eighth International Mango Symposium, 5–10 February, Sun City, South Africa, p. 27.
Issarakraisila, M. and Considine, J.A. (1994) Effects of temperature on pollen viability in mango cv. ‘Kensington’. Annals of Botany 73, 231–240.

C.P.A. Iyer and R.J. Schnell92
Iyer, C.P.A. (1991) Recent advances in varietal improvement in mango. Acta Horticultu-rae 291, 109–132.
Iyer, C.P.A. and Subramanyam, M.D. (1972) Possible role of embryo culture in mango breeding. Indian Journal of Horticulture 29, 135–136.
Iyer, C.P.A. and Subramanyam, M.D. (1979) Improvement of mango by selection and hybridization. Annual Report of the Indian Institute of Horticultural Research. In-dian Institute of Horticultural Research, Bangalore, India, p. 16.
Iyer, C.P.A. and Subramanyam, M.D. (1986) Creeping, a promising genotype for intro-duction of dwarfness in mango. Indian Journal of Horticulture 43, 221–223.
Iyer, C.P.A. and Subramanyam, M.D. (1987) Improvement of mango by selection and hybridization. Annual Report of the Indian Institute of Horticultural Research. In-dian Institute of Horticultural Research, Bangalore, India, p. 11.
Iyer, C.P.A. and Subramanyam, M.D. (1993) Improvement of mango. In: Chadha, K.L. and Pareek, O.P. (eds) Advances in Horticulture. Vol.1. Malhotra Publishing, New Delhi, India, pp. 267–278.
Iyer, C.P.A., Subbaiah, M.C., Subramanyam, M.D. and Prasada Rao, G.S. (1989) Screen-ing of germplasm and correlation among certain characters in mango. Acta Horti-culturae 231, 83–90.
Jintanawongse, S., Chunwongse, J., Chumpong, S. and Hiranpradit, H. (1999) Improve-ment of existing commercial mango cultivars in Thailand. (Abstract). The Sixth In-ternational Mango Symposium, 6–9 April, Pattaya, Thailand.
Juliano, J.B. (1934) Origin of embryos in the strawberry mango. Philippine Journal of Science 54, 553–563.
Kashkush, K., Jinggui, F., Tomer, E., Hillel, J. and Lavi, U. (2001) Cultivar identifi cation and genetic map of mango (Mangifera indica). Euphytica 122, 129–136.
Knight, R.J., Jr (1980) Origin and world importance of tropical and sub-tropical fruits. In: Nagy, S. and Shaw, P.E. (eds) Tropical and Sub-tropical fruits. Composition, Proper-ties and Uses. AVI Publishing, Westport, Conneticut, pp. 1–20.
Knight, R.J. and Schnell, R.J. (1993) Mango (Mangifera indica) introduction, and evaluation in Florida and its impact on the world industry. Acta Horticulturae 341, 125–135.
Knight, R.J. and Schnell, R.J. (1994) Mango (Mangifera indica L.) introduction and eval-uation in Florida and its impact on the world industry. Acta Horticulturae 341, 125–135.
Kostermans, A.J.G.H. and Bompard, J.M. (1993) The Mango: their Botany, Nomencla-ture, Horticulture and Utilization. Academic Press, London.
Kurian, R.M. and Iyer, C.P.A. (1992) Stem anatomical characters in relation to tree vigour in mango. Scientia Horticulturae 50, 245–248.
Lahav, E., Tomer, E., Gazit, S. and Lavi, U. (1995) Performance of avocado (Persea americana Mill.) and mango (Mangifera indica L.) seedlings compared with their grafted trees. Journal of the American Society for Horticultural Science 120, 265–269.
Lavi, U., Tomer, E. and Gazit, S. (1989) Inheritance of agriculturally important traits in mango. Euphytica 44, 5–10.
Lavi, U., Sharon, D., Tomer, E., Adato, A. and Gazit, S. (1993) Conventional and modern breeding of mango cultivars and rootstocks. Acta Horticulturae 341, 145–151.
Lavi, U., Tomer, E. and Hillel, J. (1998) Components of genetic variation and genetic correlations of mango traits. Scientia Horticulturae 75, 11–25.
Lavi, U., Kashkush, K., Saada, D., Shats, H., Ravid, U. and Tomer, E. (2004) Mango breeding and the potential of modern biology. Acta Horticulturae 645, 51–59.
Leroy, J.F. (1947) La polyembryonie chez les Citrus son interet dans la culture et lame-lioration. Revue Internationale de Botanique Appliquee Paris 27, 483–495.

Breeding and Genetics 93
Lopez-Valenzuela, J.A., Martinez, O. and Paredes-Lopez, O. (1997) Geographic differ-entiation and embryo type identifi cation in Mangifera indica L. cultivars using RAPD markers. HortScience 32, 1105–1108.
Maheshwari, P. (1934) The Indian mango. Current Science 3, 97–98.Maheshwari, P. and Rangaswamy, N.S. (1958) Polyembryony and in vitro culture of
embryos of Citrus and Mangifera. Indian Journal of Horticulture 15, 275–282.Majumder, P.K. and Sharma, D.K. (1990) Mango. In: Bose, T.K. and Mitra, S.K. (eds)
Fruits: Tropical and Subtropical. Naya Prokash, Calcutta, pp. 1–62.Majumder, P.K., Singh, R.N., Sharma, D.K. and Mukerjee, S.K. (1972) Preliminary stud-
ies on inheritance in Mangifera indica L. Acta Horticulturae 24, 101–106.Majumder, P.K., Sharma, D.K. and Singh, R.N. (1981) Breeding for dwarfness in mango
(Mangifera indica L.). National Symposium on Tropical and Subtropical Fruit Crops,Horticultural Society of India, Bangalore, p. 3.
Malik, P.C. (1951) Morphological and biology of the mango fl ower. Indian Journal of Horticulture 14, 1–23.
Marais, Z. (1992) Mango evaluation for breeding. Citrus and Subtropical Fruit Research Institute (CSFRI) Information Bulletin 234, 7.
Marshall, T.C., Slate, J., Kruuk, L. and Pemberton, J.M. (1998) Statistical confi dence for likelihood-based paternity inference in natural populations. Molecular Ecology 7, 639–655.
Medina, J.P. (1977) ‘Rosica’ – a new mango variety selected in Ica, Peru. Fruit Varieties Journal 31, 88–89.
Moore, G.A. and Castle, W.S. (1988) Morphological and isozymic analysis of open-pollinated citrus rootstock population. Journal of Heredity 79, 59–63.
Morton, J.F. (1987) Fruits of Warm Climates. Creative Resource Systems, Winterville, North Carolina, pp. 221–237.
Mukherjee, S.K. (1950) Mango; its allopolyploid nature. Nature 166, 196–197.Mukherjee, S.K. (1953) The mango – its botany, cultivation, uses and future improve-
ments, especially as observed in India. Economic Botany 7, 130–162.Mukherjee, S.K. (1957) Cytology of some Malayan species of Mangifera. Cytologia 22,
239–241.Mukherjee, S.K. (1963) Cytology and breeding of mango. Punjab Horticultural Journal
3, 107–115.Mukherjee, S.K., Singh, R.N., Majumder, P.K. and Sharma, D.K. (1968) Present position
regarding breeding of mango (Mangifera indica L.) in India. Euphytica 17, 462–467.
Mukherjee, S.K., Chakraborty, S., Sadhukhan, S.K. and Saha, P. (1983) Survey of man-goes of West Bengal. Indian Journal of Horticulture 40, 7–13.
Mukunda, G.K. (2003) Studies on the performance of certain clones of mango cv. Al-phonso. PhD thesis, University of Agricultural Sciences, Bangalore, India.
Naik, K.C. (1948) Improvement of the mango (Mangifera indica L.) by selection and hybridization. Indian Journal of Agricultural Science 18, 35–41.
Oppenheimer, C. (1956) Study tour report on subtropical fruit growing and research in India and Ceylon. Special Bulletin No.3 State of Israel Ministry of Agriculture. Ag-ricultural Research Station, Rehovot, Israel.
Oppenheimer, C. (1967) Nimrod – a new mango variety selected in Israel. Proceedings of the Florida State Horticultural Society 80, 358–359.
Pandey, S.N. (1998) Mango cultivars. In: Srivastava, R.P. (ed.) Mango Cultivation. Inter-national Book Distributing Co., Lucknow, India, pp. 39–99.
Pinto, A.C.Q. and Byrne, D.H. (1993) Mango hybridization studies in tropical savannah (‘Cerrados’) of Brazil Central region. Acta Horticulturae 341, 98–106.

C.P.A. Iyer and R.J. Schnell94
Pinto, A.C.Q., Andrade, S.R.M., Ramos, V.H.V. and Cordeiro, M.C.R. (2004) Intervarietal hybridization in mango: techniques, main results and their limitations. Acta Horti-culturae 645, 327–331.
Pope, W.T. (1929) Mango Culture in Hawaii. Hawaii Agricultural Experiment Station Bulletin 58. Hawaii Agricultural Experiment Station, Hawaii.
Popenoe, W. (1917) The Pollination of the Mango. US Department of Agriculture (USDA) Bulletin 542. USDA, Washington DC.
Popenoe, W. (1920) The mango. In: Manual of Tropical and Subtropical Fruits. The Mac-millan Co., New York, p. 135.
Prakash, O. and Srivastava, K.C. (1987) Mango Diseases and their Management – aWorld Review. Today and Tomorrows Printers, New Delhi, India.
Rajput, M.S., Chadha, K.L. and Negi, S.S. (1996) Dashehari – 51, a regular bearing and high yielding clone of mango cv. Dashehari. (Abstract). The Fifth International Mango Symposium, 1–6 September, Tel Aviv, Israel, p. 42.
Ram, N., Kamalwanshi, R.S. and Sachan, I.P. (1987) Studies on mango malformation. Indian Journal of Mycology and Plant Pathology 17, 29–33.
Ram, S., Bist, L.D., Lakhanpal, S.C. and Jamwal, I.S. (1976) Search of suitable pollinizer for mango cultivars. Acta Horticulturae 57, 253–263.
Ramaswamy, N. (1989) Survey and isolation of ‘plus trees’ of mango. Acta Horticulturae231, 93–96.
Ravishankar, K.V., Chandrasekhar, P., Sreedhar, S.A., Dinesh, M.R., Anand, L. and Saip-rasad, G.V.S. (2004) Diverse genetic bases of Indian polyembryonic and monoem-bryonic mango cultivars. Current Science 87, 870–871.
Rossetto, C.J., Bortoletto, N., Carvalho, C.R.L., Walder, J.M.M., Nogueira, N.L., Arthus, V. and Lopes, L.A. (2006) Mango resistance to fruit fl ies: 1. Varietal selection and mechanism of resistance. (Abstract). The Eighth International Mango Symposium, 5–10 February, Sun City, South Africa, p. 22.
Roy, B. (1950) A mango chimera. Current Science 19, 93.Roy, B. and Visweswariya, S.S. (1951) Cytogenetics of mango and banana. Report of
Maharashtra Association for Cultivation of Science, Pune, India.Roy, R.S., Mallik, P.C. and Sinha, R.P. (1956) Mango breeding in Bihar, India. Proceed-
ings of the American Society for Horticultural Science 68, 259–264.Ruehle, G.D. and Ledin, R.B. (1956) Mango Growing in Florida. Florida Agricultural
Experiment Station Bulletin 574. Florida Agricultural Experiment Station, Florida.Sachan, S.C.P., Katrodia, J.S., Chundawat, B.S. and Patel, M.N. (1988) New mango hy-
brids from Gujarat. Acta Horticulturae 231, 103–105.Sahijram, L., Bollamma, K.T., Naren, A., Soneji, J.R., Dinesh, M.R. and Halesh, G.K.
(2005) In vitro hybrid embryo rescue in mango (Mangifera indica L.) breeding. In-dian Journal of Horticulture 62, 235–237.
Salvi, M.J. and Gunjate, R.T. (1988) Mango breeding work in Konkan region of Maha-rashtra state. Acta Horticulturae 231, 100–102.
Schnell, R.J. and Knight, R.J., Jr (1992) Frequency of zygotic seedlings from fi ve polyem-bryonic mango rootstocks. HortScience 27, 174–176.
Schnell, R.J. and Knight, R.J., Jr (1998) Phenology of fl owering among different mango cultivars. Proceedings of the Florida State Horticultural Society 111, 320–321.
Schnell, R.J., Ronning, C.M. and Knight, R.J., Jr (1995) Identifi cation of cultivars and validation of genetic relationships in Mangifera indica L. using RAPD markers. Theoretical and Applied Genetics 90, 269–271.
Schnell, R.J., Olano, C.T., Quintanilla, W.E. and Meerow, A.W. (2005) Isolation and characterization of 15 microsatellite loci from mango (Mangifera indica L.) and

Breeding and Genetics 95
cross-species amplifi cation in closely related taxa. Molecular Ecology Notes 5, 625–627.
Schnell, R.J., Brown, J.S., Olano, C.T., Meerow, A.W., Campbell, R.J. and Kuhn, D.N. (2006) Mango genetic diversity analysis and pedigree inferences for Florida culti-vars using microsatellite markers. Journal of the American Society for Horticultural Science 131, 214–224.
Schnell, R.J., Brown, J.S., Kuhn, D.N., Cervantes-Martinez, C., Borrone, J.W., Olano, C.T., Motamayor, J.C., Phillips, W., Johnson, E., Monteverde-Penso, E.J., Amores, F. and Lopes, U. (2007) Current challenges of tropical tree crop improvement: integrating ge-nomics into an applied cacao breeding program. Acta Horticulturae 738, 129–144.
Sen, P.K., Mallik, P.C. and Ganguly, B.D. (1946) Hybridization of the mango. IndianJournal of Horticulture 4, 4–15.
Sharma, D.K. (1987) Mango breeding. Acta Horticulturae 196, 61–67.Sharma, D.K. and Choudhury, S.S. (1976) Occurrences of an unknown wild race of
Mangifera in Tripura. Current Science 45, 305–306.Sharma, D.K. and Majumder, P.K. (1988a) Further studies on inheritance in mango. Acta
Horticulturae 231, 106–111.Sharma, D.K. and Majumder, P.K. (1988b) Induction of variability in mango through
physical and chemical mutagens. Acta Horticulturae 231, 112–116.Sharma, D.K. and Singh, R.N. (1970) Self incompatibility in mango (Mangifera indica
L.). Horticultural Research 10, 108–115.Sharma, D.K. Majumder, P.K. and Singh, R.N. (1972) Inheritance pattern in mango
(Mangifera indica L.). In: Proceedings of the Symposium on Recent Advances in Horticulture. Uttar Pradesh Institute of Agricultural Sciences, Kanpur, Uttar Pradesh, India, pp. 66–68.
Sharma, R.R. (2003) Catecholase and cresolase activity as biochemical index for screen-ing mango seedlings at nursery stage for bearing behaviour in their future reproduc-tive life. Plant Genetic Resources (PGR) Newsletter 133, 31–34.
Sharma, R.R., Singh, C.N., Chbonkar, P., Goswami, A.M. and Singh, S.K. (2000) Poly-phenol oxidase activity as an index for screening mango (Mangifera indica) germ-plasm against malformation. Plant Genetic Resources (PGR) Newsletter 124, 41–43.
Siddiqui, S.H. (1985) Induced somatic mutation in mango. Mangifera indica L. cv. Langra. Pakistan Journal of Botany 17, 75–79.
Siddiqui, S.H., Mujeeb, K.A. and Vati, S.M. (1966) Evolution of new varieties of mango (Mangifera indica L.) through induced somatic mutations by ionizing radiations. In: Proceedings of the First Agricultural Symposium. Atomic Energy Commission, Dac-ca, Bangladesh, pp. 34–37.
Singh, H. and Chadha, K.L. (1981) Improvement of Dashehari by clonal selection. (Ab-stract). National Symposium on Tropical and Subtropical Fruit Crops. Horticultural Society of India, Bangalore, p. 5.
Singh, L.B. (1960) The Mango – Botany, Cultivation and Utilisation. Leonard Hill, London.Singh, L.B. (1963) A new technique for inducing early fruiting in mango hybrids based
on movement of hormones. Proceedings of the Florida State Horticultural Society75, 410–412.
Singh, L.B. (1969) Mango. In: Ferwerda, F.P. and Wit, F. (eds) Outlines of Perennial Crop Breeding in the Tropics. Veenen and Zonen, Wageningen, the Netherlands, pp. 309–327.
Singh, R.N. (1954) Studies on fl oral biology and subsequent developments of fruit in the mango varieties, Dashehari and Langra. Indian Journal of Horticulture 11, 69–88.
Singh, R.N. (1990) Mango. Series No. 3. Indian Council of Agricultural Research (ICAR), New Delhi, India.

C.P.A. Iyer and R.J. Schnell96
Singh, R.N., Majumder, P.K. and Sharma, D.K. (1962) Self-incompatibility in mango var. Dashehari. Current Science 31, 209.
Singh, R.N., Majumder, P.K., Sharma, D.K. and Mukherjee, S.K. (1972) Some promising mango hybrids. Acta Horticulturae 24, 117–119.
Singh, R.N., Sharma, D.K. and Majumder, P.K. (1980) An effi cient technique of mango hybridization. Scientia Horticulturae 12, 299–301.
Singh, R.N., Gorakh, S., Rao, O.P. and Mishra, J.S. (1985) Improvement of Banarsi Langra through clonal selection. Progressive Horticulture 17, 273–277.
Singh, S.N. and Singh, S.P. (1952) Studies on the storage and longevity of some fruits and vegetables. Journal of Agriculture and Animal Husbandry Uttar Pradesh 2, 3–11.
Slor, E. and Gazit, S. (1982) ‘Tahar’, a new mango cultivar. Alon Honotea 36, 807. (In Hebrew)
Spencer, J.L. and Kennard, W.C. (1955) Studies on mango fruit set in Puerto Rico. Tropi-cal Agriculture 32, 323–330.
Sturrock, D. (1969) Final report on some mango hybrids – 1969. Proceedings of the Florida State Horticultural Society 82, 318–321.
Sturrock, T.T. (1968) Genetics of mango polyembryony. Proceedings of the Florida State Horticultural Society 81, 311–314.
Tomer, E., Lavi, U., Degani, C. and Gazit, S. (1993) ‘Noami’: a new mango cultivar. HortScience 28, 755–756.
Truscott, M. (1992) Biochemical screening of polyploidy mango seedlings. Citrus and Subtropical Fruit Research Institute (CSFRI) Information Bulletin 237, 17–18.
Vijaya Kumar, M., Ramaswamy, N. and Rajagopalan, R. (1991) Exploiting natural vari-ability in mango. In: Proceedings of the National Seminar on Irregular Bearing in Mango – Problems and Strategy, Pusa, Bihar, India. Rajendra Agricultural Univer-sity, Sabour, Bihar, India, pp. 55–56.
Viruel, M.A., Escribano, P., Barbieri, M., Ferri, M. and Hormaza, J.I. (2005) Fingerprint-ing, embryo type and geographic differentiation in mango (Mangifera indica L., Anacardiaceae) with microsatellites. Molecular Breeding 15, 383–393.
Whiley, A.W., Mayers, P.E., Saranah, J. and Bartley, J.P. (1993) Breeding mangoes for Australian conditions. Acta Horticulturae 341, 136–145.
Yee, W. (1958) The Mango in Hawaii. Hawaii Agricultural Experiment Station Series Circular No. 388. Hawaii Agricultural Experiment Station, Hawaii.
Young, T.W. and Ledin, R.B. (1954) Mango breeding. Proceedings of the Florida State Horticultural Society 67, 241–244.

© CAB International 2009. The Mango, 2nd Edition: Botany, Production and Uses(ed. R.E. Litz) 97
5 Reproductive Physiology
T.L. DavenportUniversity of Florida, Florida, USA
5.1 Introduction 985.2 Phenology 995.3 Shoot Development 100
Vegetative shoots 102Reproductive shoots 104
5.4 Flowering Mechanisms 105Shoot initiation 105Induction 106Florigenic promoter (FP) or stimulus 108Vegetative promoter (VP) 110
5.5 Environmental Infl uence on Vegetative and Reproductive Development 111Temperature 111Water relations 113Effect of N on fl owering 114Photoperiod 116
5.6 Hormonal Infl uence on Flowering 116Ethylene 116Auxin 117Cytokinins 118Gibberellins 119Plant growth retardants 121
5.7 Photoassimilate Infl uence on Flowering 1235.8 Horticultural Manipulation of Flowering 1235.9 Conceptual Flowering Models 124
Carbohydrate-regulated fl owering models 124Hormone-regulated fl owering models 127
5.10 Floral Management 1335.11 Floral Biology 134
Sex ratio 134Environmental determinants of sex ratio 134Physiological determinants of sex ratio 135

T.L. Davenport98
Anthesis and dehiscence 136Pollen 136Pollination 137
5.12 Fruit Development 1395.13 Stenospermocarpy 1395.14 Fruit Set and Retention 140
Sex ratio 140Mineral nutrients 141Hormonal control 141
5.15 Alternate Bearing 1455.16 Conclusions 145
5.1 Introduction
Flowering and fruit set are the most critical of all events occurring after estab-lishment of a tree crop. Given favourable growth conditions, the timing and intensity of fl owering greatly determine when and how much fruit are pro-duced. Many important details about fl owering are becoming clearer, espe-cially in herbaceous plants, at the physiological, biochemical and molecular levels (see reviews by Searle, 1965; Zeevaart, 1976, 2006; Bernier et al., 1981, 1993; Halevy, 1985–1986; Bernier, 1988; Kinet, 1993; Boss et al., 2004; Komeda, 2004; Putterill et al., 2004; Corbesier and Coupland, 2005).
Cool temperatures in the subtropics stimulate mango fl owering and age of the last vegetative fl ush has an important bearing on its ability to fl ower in marginally cool or warm temperatures of the tropics (van der Meulen et al., 1971; Davenport, 2000, 2003). Consequently, mango fl owering can be en-hanced during its normal season or manipulated to occur at other times of the year in the tropics. For example, potassium nitrate (KNO3) can stimulate out-of-season fl owering in mangoes in tropical latitudes (Barba, 1974; Núñez-Elisea, 1985; Davenport, 1993; Protacio, 2000), although this treatment has not always been dependable. Various aspects of mango fl owering and/or fruit set have been reviewed (Singh, 1958a, 1979; L.B. Singh, 1960, 1977; Chacko, 1986, 1991; Chadha and Pal, 1986; Davenport, 1993, 2000, 2003; Dav-enport and Núñez-Elisea, 1997; Singh et al., 2005), and M.J. Soule (1950) pub-lished an extensive annotated bibliography of the older literature related to mango reproduction.
Understanding mango fl owering is essential to effi ciently utilize man-agement systems that extend the fl owering and crop production seasons. Recent studies of mango fl owering have resulted in conceptual models that help explain the physiological basis of fl owering (Chacko, 1991; Cull, 1991; Kulkarni, 1991, 2004; Whiley et al., 1991; Davenport and Núñez-Elisea, 1997; Davenport, 2000, 2003). Control of fl owering allows growers to harvest their crops at the most profi table times. Increasing the season of availability improves competitiveness in the international marketplace, and promotes the most effi cient use of resources as costs of inputs continue to rise.
This chapter addresses the physiology of mango fl owering, early fruit set and retention. Cultivar names and type of embryony (i.e. monoembryonic or

Reproductive Physiology 99
polyembryonic) are purposefully left out to focus on the physiological aspects of reproduction regardless of whether they are tropically or subtrop-ically adapted or from Indian or South-east Asian origin. Cultivars are selected for their productivity in specifi c environments. Transfer of a cultivar to a different environment often results in some alteration in performance. It is reasonable to assume that the underlying mechanisms by which all culti-vars respond to their environment within the framework of their genetic limi-tations are similar. The concepts described herein, therefore, apply to all cultivars, regardless of origin.
5.2 Phenology
Growth of mango and other tropical trees is not continuous (Nakasone et al., 1955; Halle et al., 1978; Verheij, 1986; Davenport, 1993, 2000, 2003). Apical buds spend most of the time in rest. Growth occurs as intermittent, ephem-eral fl ushes of shoots from apical or lateral buds (Naik and Mohan Rao, 1942; Singh, 1958a, b). Stems are quiescent or resting terminal vegetative structures on branches from which shoot growth occurs. Shoots are elongating vegeta-tive or reproductive structures that emerge from apical or lateral buds of stems. Vegetative shoots develop a prescribed number of nodes during growth before entering a resting state as a stem. Depending on environment, periods of stem rest are generally short in young plants but usually last sev-eral months between episodes of growth in mature trees. Vegetative growth generally occurs up to three or four times a year on individual branches, depending upon cultivar and growth conditions.
Development of the vegetative shoot from initiation of growth to full elongation requires 3–6 weeks, depending on the cultivar and climatic condi-tions (Whiley et al., 1991). During this period, 10–20 new leaves are generally produced before returning to a resting state. These rhythmic episodes of extension growth are recorded on each branch as segments consisting of compressed internodes interspersed with long internodes, that is articulate growth (Tomlinson and Gill, 1973). Davenport (1992, 2003, 2006) referred to regions of compressed internodes as intercalations and the entire segment of long internodes terminating in an intercalation as an intercalary unit. The number of intercalations between each branching point indicates the number of vegetative growth episodes or fl ushes that have occurred between each fl owering fl ush.
Flushes of vegetative growth occur on groups of stems borne on scaffold-ing branches in isolated sections of tree canopy. Flushing stems are usually connected at some common branch point within the tree limbs. Asynchro-nous fl ushes of growth at various times in random portions of a tree canopy may appear to be continuous growth but are simply fl ushes occurring in various parts of the total canopy over time. Flowering fl ushes generally occur after extended periods of stem rest in the low-latitude tropics or during cool winter months in the high-latitude tropics and subtropics. Like vegetative fl ushes, reproductive fl ushes are usually asynchronous in tropical climates

T.L. Davenport100
(Verheij, 1986). In the subtropics, however, trees exposed to cold tempera-tures (3–10°C) display synchronized fl owering fl ushes throughout the tree canopy approximately 1 month later. Subsequent vegetative fl ushes also tend to be synchronous for one or two growth cycles depending upon the number of retained fruit. Less intense, cool weather (10–18°C), however, results in asynchronous reproductive fl ushes in responsive stems as is typical of trees growing in the tropics. The timing of fl owering fl ushes of cultivars in various locations has been reviewed by L.B. Singh (1960), Chadha and Pal (1986) and Pandey (1989). Variations in fl owering patterns occur in all cultivars depend-ing on their age and whether they are growing in dry or humid tropics or subtropics (L.B. Singh, 1960).
5.3 Shoot Development
Flushes of vegetative extension growth of mango stems terminate with for-mation of determinate panicles. Several weeks to a few months after separa-tion of the last fl ower or fruit from these panicles are required for the central axis of the panicle or rachis to dry and mechanically separate from the sup-porting stem, depending on the longevity of attached fruit. Five to ten lateral vegetative shoots typically develop from axillary buds located at the termi-nal intercalation positioned in a compact whorl surrounding the panicle scar of each stem (see Fig. 1 of Reece et al., 1949). These lateral shoots become the branch points of stems. These branching shoots form 10–15 leaves before the apical buds return to a resting state to establish them as individual stems. Ini-tiation of these lateral vegetative shoots may occur 2–3 months after desicca-tion of panicles which fail to set fruit. Fruit-bearing stems do not initiate new lateral shoots until several months after separation of fruit and rachis from the stem (Kulkarni and Rameshwar, 1989). Such delayed vegetative growth can reduce the potential for new shoots to fl ower during the next fl owering sea-son (Singh and Khan, 1939; L.B. Singh, 1960, 1972; Monselise and Gold-schmidt, 1982). The apical bud of stems is at rest for most of the year in mature trees. Stems on centennial trees typically produce only one vegetative fl ush during the year (N. Golez, personal communication, the Philippines, 1989).
The apical resting bud of each newly established lateral stem (intercalary unit) is surrounded by a compact whorl of 10–12 leaves with short inter-nodes (intercalation) (Fig. 5.1). Protective bud scales are green but may be brown at the tips due to desiccation (Sen and Mallik, 1941; Mustard and Lynch, 1946; Singh, 1958b; Ravishankar et al., 1979). Resting buds possess a number of pre-formed nodes, each of which contains a leaf bract or leaf pri-mordium and a lateral meristem (Fig. 5.2; see Figs 7–12 in Chaikiattiyos et al., 1994). The outermost, proximally located dried leaf bracts (bud scales) pro-tect the more distal interior leaf bracts, leaf primordia and lateral meristems from mechanical damage and desiccation. Leaf bracts are vestigial non-developed leaves. Scales abort upon evocation of new shoots. Proximally located bracts in apical buds fail to further develop beyond some enlarge-ment and also abort with elongation of shoots.

Reproductive Physiology 101
If apical buds are initiated during vegetatively inductive conditions, bracts develop as small leaves and the leaf primordia develop as the full-sized leaves of vegetative shoots. Additional leaves result from nodes formed by renewed activity of the apical meristem. The number of leaves (nodes) is dependent upon the mean temperature during initiation, and increases as temperatures rise (Whiley et al., 1989). The lateral meristems of the apical bud develop as axillary buds at the base of petioles in the elongating vegeta-tive shoot, each bearing protective bracts, leaf primordia and lateral mer-istems (Fig. 5.3; see Fig. 1 of Reece et al., 1949).
In contrast, if shoot growth is initiated under fl oral inductive conditions, the leaf bracts and primordia fail to fully develop, but the lateral meristems
Fig. 5.1. Apical bud of resting mango stem.
Leaf primordia (bracts)
Apical dome(includes meristem)
Lateral meristematicprimordia
Fig. 5.2. Stylized cross-section of apical bud showing positions of apical meristem, lateral meristems and leaf primordia.

T.L. Davenport102
begin to elongate and branch at each node forming secondary, tertiary and quaternary lateral meristems. Each branch point in the lateral infl orescence from the panicle axis to the fl oral pedicels bears a fl oral bract (i.e. partially developed vestigial leaf primordium) (Fig. 5.4). The distal half of the panicle structure is derived from newly formed nodes laid down by cell divisions in the apical meristem prior to returning to a resting state. Mixed shoots, bear-ing both leaves and infl orescences at each node, result from development of both the primary leaf primordia and the lateral meristems, which form the infl orescences in the same nodes as leaves.
Vegetative shoot induction, thus, involves stimulating development of leaf primordia from resting buds while repressing development of lateral meristems. Leaf primordia then follow a predetermined cascade of genetic signals resulting in leaf development at each node. Because all shoots emerge from resting buds, a vegetatively induced event does not involve simply inhibition of fl owering. The putative inductive signal directing differentia-tion of leaf primordia onto leaves upon initiation is termed a vegetative pro-moter (VP) rather than a fl oral inhibitor.
Shoots bearing only infl orescences (generative shoots) result from induc-tive development of lateral meristems and suppression of leaf primordial development. A predetermined cascade of fl owering gene signals is activated in lateral meristems resulting in lateral cymose infl orescences terminating with fl owers. A distinct fl origenic promoter (FP) may be responsible for spe-cifi c activation of the lateral meristems of mango. Mixed shoot induction results in combined development of leaf primordia and lateral meristems.
Vegetative shoots
Vegetative shoots bear only leaves (Fig. 5.5). The anatomy of mango vege-tative shoot development has been described (Singh, 1958b; Chaikiattiyos
Stem
Fig. 5.3. Axillary bud of resting mango stem. Leaf petioles (arrows).

Reproductive Physiology 103
et al., 1994). Vegetative shoots may arise either from axillary buds, if no apical bud exists due to fl owering in the previous fl ush, or from the apical bud when present. The latter is considered extension growth or addition of an intercalary unit on the existing stem, but the developmental events during shoot formation from either apical or lateral buds are basically the same. Cells in the leaf primordia of initiating buds begin to form individual leaves in the proximal portion of the vegetative shoot. Soon thereafter, the apical meristem activates to form more nodes bearing leaf primordia and lateral meristems. These newly formed leaf primordia develop as the distal portion of the vegetative shoot if environmental conditions remain vegetatively inductive (Núñez-Elisea et al., 1996). Newly elongating vegetative shoots are green in most cultivars but may be bronze or red in others. Fully expanded
1°Pedicel
1°Pedicel
1° Bract 2° Bract
3° Bract
2° Bract
1° Bract
2°Pedicel 1°
Pedicel
1
1°Pedicel
1° Bract
4°
2°3°
5°
º1°
Axis
Axis
Fig. 5.4. Diagram and photos of mango infl orescence depicting the panicle axis and primary (1°), secondary (2°) and succeeding levels of pedicel and cymose fl oral archi-tecture. Vestigial leaf promorida (fl oral bracts) are depicted at the base of each level of pedicel architecture.

T.L. Davenport104
leaves are a shade of red, depending upon cultivar and cultural conditions and are thin and limp from lack of lignifi cation. The apical buds of vegetative shoots generally become quiescent before completion of the limp, red-leaf stage (Núñez-Elisea and Davenport, 1995). Internodes are compressed at the apex, and leaf development is arrested thereby forming a bud with protec-tive outer scales, inner leaf primordia, lateral meristems and the apical mer-istem. Fully expanded leaves become light green and stiff as they become lignifi ed and suberized. Vegetative shoots are mature when leaves become dark green, which occurs when they are c.2 or 3 months old.
Reproductive shoots
Two types of reproductive shoots typically occur in mango. Generative shoots display only fl owers and have fl oral bracts or non-developed leaves at the base of each lateral infl orescence (Fig. 5.5). Terminal infl orescences, i.e. panicles or thyrsoids (Weberling, 1989), develop from dormant apical buds. The anatomy of panicle development has been described (Juliano and Cuevas, 1932; Musahib-ud-din, 1946; Mustard and Lynch, 1946; Singh,
VEGETATIVE GENERATIVE MIXED CHIMERIC V/F TRANSITION F/V TRANSITION
GENERATIVE CHIMERIC V/F TRANSITION VEGETATIVE MIXED F/V TRANSITION
Fig. 5.5. Stylized diagrams and photomontage of shoot types found in mango. Transition shoots shift from vegetative to fl oral (V/F) or fl oral to vegetative (F/V). Arrow ( )represents individual leaves; fl oral diagram ( ) represents lateral infl orescences.

Reproductive Physiology 105
1958b; L.B. Singh, 1960; Sturrock, 1966; Ravishankar et al., 1979; Scholefi eld, 1982; Scholefi eld et al., 1986). The complexes of primary to quaternary branch-ing lateral structures of the infl orescence each terminate with three cymose fl owers. The terminal fl ower opens fi rst, followed by two subtending lateral fl owers. These complexes form the lateral infl orescence structures emerging from the central axis of the panicle. The central axis extension also terminates in a similar fashion. Morphological stages of fl oral buds and panicle develop-ment were described by Shu (1981) and Oosthuyse (1991a). Reece et al. (1949) described the development of infl orescences initiated in lateral buds when the terminal bud is missing. There are more nodes in dormant apical buds and their bracts are more developed than in axillary buds; however, fl oral evocation is indistinguishable.
Generative shoot development in apical buds initially involves swelling of the lateral meristems and their bud scales. Each axillary meristem devel-ops as an infl orescence on a primary peduncle. The apical meristem then forms new lateral meristems and leaf primordia for the distal portion of pan-icle development if fl oral inductive conditions persist (Núñez-Elisea et al., 1996). Panicles may be open or compact, depending upon internode elonga-tion, which is cultivar dependent (L.B. Singh, 1960), but the architecture gen-erally conforms to that in Fig. 5.5. Mixed shoots develop under weak fl oral inductive conditions (i.e. in the low-latitude tropics). Both leaves and pri-mary pedunculate infl orescences develop from the same nodes (Fig. 5.5). Leaf primordia and lateral meristems develop as leaf and fl oral structures, respectively.
5.4 Flowering Mechanisms
Mango stems undergo varying periods of rest between episodes of growth, depending on tree age and environmental infl uences. Resting mango buds must, therefore, respond to two distinctly different signals for shoots to occur. The fi rst signal initiates growth of the shoot and the second determines if it will be vegetative or reproductive. The signals that regulate initiation of shoot growth in resting buds differ from the inductive signals that regulate shoot type.
Shoot initiation
Initiation is the onset of shoot development, regardless of the type of shoot evoked. It involves cell division and elongation of cells in leaf primordia (vegetative shoots), lateral meristems (generative shoots) or both (mixed shoots) in the nodes of the resting buds, and is followed by cell divisions in the apical meristem to form more nodes. Shoot initiation is stimulated by pruning, defoliation and irrigation during dry conditions, or transition from the dry to rainy season in the tropics. Application of nitrogen (N)-containing fertilizers, exposure to ethylene, or a shift from cool to warm temperatures

T.L. Davenport106
also stimulates shoot initiation. Reece et al. (1946, 1949), Mustard and Lynch (1946), Núñez-Elisea and Davenport (1992b), Núñez-Elisea et al. (1996) and Davenport et al. (2006a) observed that the vegetative or reproductive fate of mango buds remains undetermined until after shoot growth is initiated. Reece et al. (1949) proposed that a putative signal that triggers initiation of shoot development is separate and different from the inductive signal, which determines the fate of the shoot. Removal of apical buds by pruning stimu-lates initiation of axillary shoots (Singh and Singh, 1956; Núñez-Elisea and Davenport, 1992b; Núñez-Elisea et al., 1996; Davenport et al., 2006a). Defolia-tion of the apical whorl of fi ve to ten leaves also stimulates shoot initiation in dormant apical buds (Núñez-Elisea et al., 1991; Núñez-Elisea and Daven-port, 1995). The fate of shoots that emerge in response to these initiation stim-uli, however, is determined by other factors that are prevalent at the time of initiation. Tip pruning, for example, during warm summer months results in initiation of vegetative shoots from axillary buds, whereas pruning during cool winter months usually results in initiation of axillary infl orescences.
Induction
Induction in mango is the temporary commitment of buds to evoke a par-ticular developmental pathway (i.e. vegetative shoot, generative shoot or mixed shoot) when growth is initiated. Initiation of herbaceous plant fl ower-ing refers to the onset of fl oral bud growth in actively growing vegetative shoots after the fl oral inductive event (Bernier et al., 1981, 1993; Halevy, 1985–1986; Bernier, 1988; Huala and Sussex, 1993; Kinet, 1993). The inductive sig-nal is formed in leaves, but the responsive buds are in continuous vegetative growth at the time of fl oral induction in herbaceous plants and fl oral initia-tion follows; whereas mango buds are in rest. Although the mango bud must be initiated to grow, that growth is induced according to forces already present.
Whereas the fl oral inductive signal in mango may be present prior to bud initiation, it must be present at the time of initiation for fl owering to occur (Kulkarni, 1988a; Núñez-Elisea and Davenport, 1995; Núñez-Elisea et al., 1996; Davenport and Núñez-Elisea, 1997; Davenport et al., 2006a). The induc-tive signal can be shifted from fl oral (F) to vegetative (V) or vegetative to fl oral, forming F/V or V/F transition shoots, by altering temperatures dur-ing early shoot development (Batten and McConchie, 1995; Núñez-Elisea et al., 1996) (Fig. 5.5). This shift in morphogenic responses during shoot devel-opment demonstrates the plasticity and temporal nature of induction, indi-cating that cells of the apical meristem do not become irreversibly determined under inductive conditions. These results demonstrate that, rather than being irreversibly committed to a vegetative or reproductive fate at the onset of shoot initiation, the mango apical meristem provides progenitor cells, some of which differentiate into specifi c target cells at each node in the apex. The apical meristem, therefore, may not be directly involved in the fl owering process.

Reproductive Physiology 107
Target cells within leaf primordia and lateral meristems are competent to respond to inductive signals; for example when initiated to grow under veg-etatively inductive conditions, individual leaf primordia develop as leaves and subtending lateral meristems associated with each developing leaf develop as dormant axillary buds with protective bracts. These axillary buds may develop in subsequent fl ushes as vegetative shoots when initiated in vegetatively inductive conditions or as axillary infl orescences under fl oral inductive conditions. Under strongly fl oral-inductive conditions, leaf pri-mordia fail to develop beyond the bract stage, become dormant, and lateral meristems develop. Each lateral meristem forms nodes consisting of leaf pri-mordia and meristems that are infl uenced by the putative fl oral-inductive stimulus, which suppresses development of newly formed leaf primordia. Subsequently formed meristems form pedunculate structures that terminate in cymose infl orescences borne on each tertiary peduncle (Fig. 5.4). Forma-tion of the primary, secondary, tertiary and quaternary peduncles, as well as pedicels of infl orescences are always accompanied by a subtending, aborted bract or vestigial leaf at each node (Fig. 5.4). Such development is attributed to a sequence of gene expression (Coen et al., 1990; Coen and Meyerowitz, 1991; Weigel et al., 1992; Coen and Carpenter, 1993; Lumsden, 1993; Yanofsky, 1995). Shoot initiation during weakly fl oral-inductive conditions activates growth of leaf primordia to develop leaves and the lateral meristems to pro-duce peduncles bearing lateral infl orescences in each node of mixed shoots. The bases of each pedicel branch within each lateral infl orescence also bear a vestigial leaf.
Upon termination of cell divisions in the apical meristem at the end of a fl ushing period, no more nodes are formed. The apical bud of vegetative shoots becomes quiescent, and the resting leaf primordia, bracts and lateral meristems are poised to resume growth at a later date. When reproductive or mixed shoots become quiescent, the lateral meristems ultimately develop determinant cymose infl orescences. The most distally located meristem is possibly the determinant extension of the central axis forming the terminal cymose fl oral group.
Chimeric shoots (Fig. 5.5) can occur in mango trees when shoot initiation occurs during fl oral inductive conditions. They display infl orescences on one side of the longitudinally bisected shoot and leaves on the other. The shoot axis is red on the fl oral side of red fruiting cultivars (typical of panicles) and green on the vegetative side (typical of vegetative shoots). This difference in the two sides extends to the apical bud, which bears an undeveloped infl orescence on the fl oral side and leaf bracts on the vegetative side. The explanation for this spatial differentiation is that target nodes on each side of the apical bud respond to the different inductive signals at the same time. The apical meristem is not implicated except to form more nodes for the lat-eral inductive responses on each side in the second portion of growth. Differ-ences in inductive signals on each side of an existing shoot probably cause the differential response. This phenomenon indicates that the fate of nodes on each side of the shoot cannot be attributed to a single mother cell in the apical meristem. The inductive response must involve cells formed in later

T.L. Davenport108
cell divisions and would be determined by their location within nodes of the bud.
Florigenic promoter (FP) or stimulus
Early fl owering work provided evidence for the presence of a graft transmis-sible fl oral stimulus (i.e. fl origen) that was induced in leaves and was trans-located to buds to stimulate fl oral development (Chailakhyan, 1936; Zeevaart and Boyer, 1987). Florigen was functionally conserved across plant species (Lang, 1965, 1984; Zeevaart, 1976; Lang et al., 1977). Floral induction in most plants involves sensing of some environmental cue (i.e. daylength, water stress or vernalizing temperature) in some organ (e.g. leaves). A putative fl o-ral stimulus or alteration in the ratio of fl origenic to anti-fl origenic compo-nents may be translocated to target cells in meristems (Bernier et al., 1981). Photoassimilate movement from leaves in phloem facilitates its transport to buds where it can interact to initiate fl owering (King and Zeevaart, 1973). Until recently, a fl oral stimulus could not be identifi ed. Alternative hypoth-eses were proposed that nutrient diversion to the meristems could be involved (Sachs and Hackett, 1983) or that fl oral induction might be con-trolled by multiple factors, including the putative fl oral stimulus, photoas-similates and phytohormones (Bernier et al., 1993).
Molecular biology of fl owering in the facultative, long-day, model plant, Arabidopsis thaliana (reviewed in Zeevaart, 2006 and Aksenova et al., 2006), has provided insight into the nature of the fl oral stimulus (FP). A network of four interacting genetic signalling pathways may result in fl owering in response to photoperiodic, vernalization, gibberellin and autonomous envi-ronmental cues (Perilleux et al., 1994; Mouradov et al., 2002; Perilleux and Bernier, 2002; Boss et al., 2004; Komeda, 2004; Putterill et al., 2004; Corbesier and Coupland, 2005). The photoperiodic pathway involves activation of the CONSTANS (CO) gene that encodes a zinc-fi nger protein, which in turn induces expression of the FLOWERING LOCUS T (FT) gene in the phloem tissue of leaves. FT is the terminal, integrating gene of the four path-ways regulating fl owering in Arabidopsis. Its transcribed mRNA was initially thought to be the FP that is transported in phloem to buds (Huang et al., 2005); however, evidence indicates that the translated protein product of FT is translocated to Arabidopsis buds (Corbesier et al., 2007). Analogous proteins encoded by Hd3a, an ortholog of FT in rice (Tamaki et al., 2007), and the aspen ortholog, PtFT1, which along with CO regulates the timing of fl owering and growth cessation of Populus trichocarpa (Bohlenius et al., 2006), appear to be the FP. In the buds, the protein product of FT is thought to combine with the bZIP transcription factor (FD) protein to activate transcription of fl oral iden-tity genes (i.e. APETALA1) to begin fl oral expression (Abe et al., 2005; Wigge et al., 2005). Similar mechanisms are likely to exist in mango.
Zhang et al. (2005) and Davenport et al. (2006b) isolated a CONSTANS-like gene (MiCOL) from mango leaf DNA. CO is a circadian expression gene interacting with the photoperiodic pathway in Arabidopsis (Putterill et al.,

Reproductive Physiology 109
2004), and is central to activation of the FT gene in Arabidopsis during long days. Its role in mango fl owering is unclear. The mango ortholog has 79%, 76% and 62% homology with two apple CO genes, MdCOL2 and MdCOL1, and the Arabidopsis CO gene (AtCO), respectively. Isolation of the FT or homologous gene responsible for synthesis of the FP has been unsuccessful.
Studies with mango indicate that a FP is synthesized in leaves during exposure to cool, fl oral-inductive temperatures and moves to buds to induce fl owering (Reece et al., 1946, 1949; Singh and Singh, 1956; L.B. Singh, 1959, 1962, 1977; R.N. Singh, 1961; Sen et al., 1972; Núñez-Elisea and Davenport, 1989, 1992b; Davenport and Núñez-Elisea, 1990; Davenport et al., 1995, 2006a). Unlike receptor sites in buds of Thlaspi arvense (Metzger, 1988) and other plants requiring vernalization for fl oral induction (Zeevaart, 1976; Bernier et al., 1981), mango leaves appear to be where the putative fl oral stim-ulus is produced. Complete defoliation of girdled branches during inductive conditions results in vegetative shoots instead of generative shoots (Reece et al., 1949; Sen et al., 1972; Núñez-Elisea and Davenport, 1989, 1992b; Núñez-Elisea et al., 1996; Davenport et al., 2006a). It appears to be transported over long distances from leafy branches to defoliated branches (Sen et al., 1972; Núñez-Elisea et al., 1996).
The putative, temperature-regulated FP is short-lived in situ (Núñez-Elisea and Davenport, 1989, 1992b; Davenport et al., 1995; Núñez-Elisea et al., 1996). Leafl ess cuttings from trees during cool, fl oral inductive conditions produce infl orescences when stimulated to grow within 7 days of transfer to warm, non-inductive conditions; the infl uence of the removed leaves lasts for 13 days when cuttings are stored at cool temperatures (Davenport et al., 2001a). The same cuttings produce only vegetative shoots in both storage conditions after the initial loss of reproductive shoot production. There are more leaves on mango stems than are necessary for fl oral induction in cool temperatures. Stems bearing as little as one-quarter of a cross-sectioned leaf induce 95% generative shoots (Davenport et al., 2006a); the remaining shoots are vegetative. Half of a leaf or more resulted in 100% generative shoots. Thus, the limiting amount of leaf necessary for fl oral induction is less than a quarter of a leaf per stem. Davenport et al. (2006a) demonstrated the quanti-tative movement of mango FP from half to fi ve leaves on a donor stem to fi ve leafl ess receiver stems located as far as 100 cm from the donor stem in isolated branches during exposure to cool, fl oral inductive temperatures. The FP moves with photoassimilates in phloem from donor leaves to buds in the receiver stems.
The mango fl oral stimulus is graft transmissible (L.B. Singh, 1959, 1962; Kulkarni, 1986, 1988b, 1991). Flowering of seedling stems is stimulated by grafting onto mature trees or by grafting mature stems onto juvenile plants (L.B. Singh, 1959, 1962). Some mango cultivars selected in the tropics can fl ower at higher temperatures than others and are not restricted to winter fl owering (Kulkarni, 1991). Transfer of the FP from tropical to subtropical selec-tions was accomplished using reciprocal grafts between the two cultivar types (Kulkarni, 1986, 1988b, 1991). Subtropical cultivars that seldom fl ower in warm temperatures fl ower in the ‘off’ season using these techniques. Three conditions

T.L. Davenport110
were essential for summer fl owering to occur in the low-temperature-requiring cultivars (receptors) when grafted to the summer fl owering type (donors): (i) the summer-fl owering donor cultivar stocks or scions were in a fl owering cycle; (ii) buds on the receptor scions or stocks of grafted plants had initi-ated shoot growth during this cycle; and (iii) receptor stocks or scions had been completely defoliated for transfer and/or expression of the fl oral stimu-lus. The presence of any leaves on the receptor plants resulted in vegetative shoots.
Girdling experiments to isolate treated mango branches from the rest of the tree suggest that the FP is translocated via phloem to apical buds (King and Zeevaart, 1973; Bernier et al., 1981; Núñez-Elisea and Davenport, 1989, 1992b; Núñez-Elisea et al., 1996; Davenport et al., 2006a). Shading experiments to reduce photosynthate loading into the phloem also support this (Kulkarni, 1991). Reduced fl owering responses were observed in isolated leafy branches that were provided with 90% and complete shading, which stopped photosyn-thate production entirely, mimicked defoliation during cool, fl oral inductive conditions, resulting in a vegetative growth response (R. Núñez-Elisea, T.L. Davenport and B. Schaffer, Florida, 1991, unpublished results).
Vegetative promoter (VP)
An independently regulated VP probably contributes to induction of vegeta-tive shoots as opposed to a fl oral inhibitor or expression of a default vegeta-tive status in the absence of suffi cient FP at the time of shoot initiation. Grafting studies (L.B. Singh, 1959, 1962; Kulkarni, 1986, 1988b, 1991, 2004) demonstrated that complete removal of leaves from receptor stems is required to express fl owering of those receptors when they are grafted to fl owering donor stems. Kulkarni (1986, 1988b, 1991, 2004) considered that a putative fl oral inhibitor in leaves of the non-induced receptor stems might antagonize the infl uence of the fl oral stimulus from donor leaves. Others have noted a relationship between leaf age and the ability of shoots to be reproductive (Singh et al., 1962a; Scholefi eld et al., 1986). KNO3-stimulated early fl owering in the tropics is successful only on stems that are at least 4 (Davenport, 2003) to 7 months old (Astudillo and Bondad, 1978; Bondad and Apostol, 1979; Núñez-Elisea, 1985). Young stems often produce vegetative shoots when ini-tiated under conditions that are fl oral inductive for more mature stems (Núñez-Elisea and Davenport, 1995; Davenport, 2003). The putative VP appears to be most active in leaves of young stems and slowly dissipates over time to allow expression of the FP when shoots are initiated to grow in warm conditions.
The VP may be a gibberellin or closely associated with the gibberellin synthesis pathway as indicated by enhanced fl owering responses of trees to plant growth retardants. Mangoes growing in wet and humid, low-latitude tropics tend to produce frequent vegetative fl ushes and fl ower sporadically, perhaps due to higher levels of the VP in the young stems combined with low levels of the putative FP when shoot initiation occurs. Paclobutrazol

Reproductive Physiology 111
(PBZ) reduces the time in rest necessary to allow fl oral induction during warm temperature conditions by c.1 month (Davenport, 2003), thus increas-ing the potential to produce reproductive shoots in younger stems when ini-tiated to grow. PBZ and uniconazole, triazole compounds that inhibit kaurene oxidase in the gibberellin-synthesis pathway (Dalziel and Lawrence, 1984; Rademacher, 1991), stimulate production of fl owering shoots during weakly inductive conditions (Burondkar and Gunjate, 1991, 1993; Tongumpai et al., 1991a; Voon et al., 1991; Nartvaranant et al., 2000; Yeshitela et al., 2004a). Application of PBZ to mango trees bearing 1-month-old stems produced infl orescences when bud break was initiated 3 months later by foliar applica-tion of KNO3 (Davenport, 2003).
Vegetative or reproductive induction at the time of shoot initiation is governed by the ratio of the putative fl oral promotive to inhibitory compo-nents (Lang et al., 1977; Lang, 1984; Kulkarni, 1988a; see Bernier et al., 1981 for additional references). The mango fl oral inhibitor should be viewed as an age-dependent VP. The presence of an age-regulated VP in mango leaves, which moves with the temperature-regulated FP and photoassimilates in phloem, may explain the induction of specifi c receptors by this promoter in targeted leaf primordia to cause development of leaves in vegetative or mixed shoots. A gradual decrease in the level or infl uence of the VP may cause vegetative shoots to develop when initiation occurs on 2-month-old stems, and generative or mixed shoots when initiation occurs in stems from 4- to 7-month-old stems, given the constantly warm daily temperatures maintaining a low level of FP in both situations.
5.5 Environmental Infl uence on Vegetative and Reproductive Development
The effects of temperature and water relations on determinating vegetative and reproductive growth of mango have been addressed (Davenport and Núñez-Elisea, 1997; Davenport, 2000; Kulkarni, 2004; Bangerth, 2006). This section focuses on the impacts of temperature, plant water relations, mineral nutrition and photoperiod on shoot initiation and induction.
Temperature
The developmental fate of mango buds is strongly infl uenced by tempera-ture (Davenport and Núñez-Elisea, 1997). Cool night temperatures < 15°C in combination with day temperatures < 20°C typically induce fl owering if shoot initiation occurs when plants are exposed to these conditions (Ou, 1980, 1982; Wolstenholme and Mullins, 1982a, b; Shu and Sheen, 1987; Whi-ley et al., 1988, 1989, 1991; Núñez-Elisea et al., 1993; Núñez-Elisea, 1994; Núñez-Elisea and Davenport, 1994a, b). The physiological and molecular basis for temperature perception in leaves with respect to fl oral induction is not understood (Samach and Wigge, 2005). Whiley et al. (1988, 1989, 1991)

T.L. Davenport112
described the vegetative growth and fl owering responses of several mo-noembryonic and polyembryonic cultivars to four temperature regimes rang-ing from vegetatively inductive (30°C day/25°C night) to fl oral inductive (15°C day/10°C night). The effect of temperature on marcotted, container-grown plants that were tip pruned or defoliated in order to stimulate shoot initia-tion was also studied (Davenport, 1987; Núñez-Elisea et al., 1991, 1993, 1996; Núñez-Elisea and Davenport, 1994b). Mango trees develop vegetative shoots when shoot initiation occurs in warm temperatures (30°C day/25°C night), whereas infl orescences develop when shoots initiate growth in cool tempera-ture conditions (18°C day/10°C night; or 15°C day/10°C night) (Whiley et al. 1989; Núñez-Elisea and Davenport, 1991b, 1995; Núñez-Elisea et al., 1993, 1996; Batten and McConchie, 1995). Bangerth et al. (2004) reported changes in the major phytohormones in stems of containerized mango trees during exposure to cool, fl oral inductive temperatures. The minimum leaf age and time of exposure to a low temperature regime (18°C day/10°C night) required by stems for fl oral induction was examined (Núñez-Elisea and Davenport, 1995). Leaves are competent to respond to cool temperatures at 7 weeks, forming a small percentage of generative shoots. As they age, higher propor-tions of generative shoots are induced and warmer temperatures can stimu-late fl oral induction. The response to temperature is moderated by age of the previous fl ush. Stems that are 4–5 months beyond the limp, red-leaf stage of development will be induced to form generative shoots if initiated to grow at 25–30°C (Davenport, 2003).
Whiley et al. (1988, 1989, 1991) observed that at least 17 weeks are required for initiation of reproductive shoots on non-clipped stems of trees maintained at 15°C day/10°C night. In similar experiments with different cultivars with-out previous clipping of distal leaves to stimulate initiation, infl orescences were observed after 5 weeks at 15°C day/10°C night (Chaikiattiyos et al., 1994). Although inductive conditions were present in each of these studies, shoot initiation was delayed by the presence of distal leaves. The earlier ini-tiation of infl orescence development in tip-pruned or tip-defoliated stems compared to intact ones demonstrates that the fl oral stimulus may be pres-ent, but the buds are not induced until initiation occurs. It demonstrates the importance of stimulating initiation of stems by tip defoliation or pruning at the onset of incubation in controlled environment conditions so that the inductive response can be observed within a reasonable length of time. The variable delays in shoot initiation in these studies occurred because the experimental protocols depended on the plants’ internal initiation cycle to initiate shoots. This cycle slows down when plants are exposed to lower tem-peratures (Whiley et al., 1988, 1989, 1991).
Floral or vegetative induction occurs when shoots are initiated. Resting buds of plants that are exposed to cool temperatures (18°C day/10°C night) for > 3 weeks and then transferred to a warm temperature (30°C day/25°C night) before initiation, produce only vegetative shoots (Núñez-Elisea et al., 1996). Thus, the stems do not ‘remember’ that they had been exposed to fl oral inductive conditions while still in rest. They responded to warm conditions present when shoot initiation occurred.

Reproductive Physiology 113
This response to temperature conditions at the time of shoot initiation extends to the formation of transition shoots if conditions change during early shoot development. First reported by Naik and Mohan Rao (1943), transition shoots are an unusual transition in expression of shoot type during a single growth fl ush (Kulkarni, 1988b; Núñez-Elisea and Davenport, 1989, 1992b; Batten and McConchie, 1995). The transition typically occurs near the middle of the extending shoot. Resting buds possess preformed nodes, each of which contains a primordial leaf or bract and a lateral meristem. The api-cal meristem initiates cell division at the same time or soon after the nodal target tissues begin development (Mustard and Lynch, 1946; L.B. Singh, 1960; Núñez-Elisea et al., 1996). Vegetative or infl orescence development in the pre-formed primordia is underway before the apical meristem begins to pro-duce differentiating cells. Transfer from a warm, vegetatively inductive con-dition to a cool, fl oral inductive environment at early bud break results in formation of V/F transition shoots (Fig. 5.5). Transfer from cool to warm con-ditions at the same stage of bud break results in formation of F/V transition shoots (Batten and McConchie, 1995; Núñez-Elisea et al., 1996).
The fl owering response to temperature occurs in mangoes growing in subtropical latitudes where cool temperature is the dominant induction fac-tor. Many cultivars fl ower erratically in the low-latitude tropics, providing continuously warm temperatures with high soil and atmospheric moisture. Under such conditions, the age of stems is the dominant inductive factor (Buell, 1954; Nakasone et al., 1955; Ravishankar et al., 1979; Ou and Yen, 1985; Issarakraisila et al., 1992), and occasional cool night temperatures in the upper latitude tropics have a positive moderating effect (Davenport, 2003).
Water relations
In the absence of cool temperatures, mango trees in the tropics may fl ower in response to irrigation or rain following periods of water stress lasting 6–12 weeks or more (Pongsomboon, 1991). Plant water stress has been presumed to provide the stimulus for fl owering (reviewed in Whiley, 1993; Chaikiatti-yos et al., 1994; Schaffer et al., 1994; Davenport and Núñez-Elisea, 1997); how-ever, most of these studies have failed to substantiate prolonged tree water defi cit as a successful agent for fl oral induction.
Experiments with container-grown trees fail to produce infl orescences after 8 weeks of water defi cit (Wolstenholme and Hofmeyr, 1985). Under glasshouse conditions (27°C day/22°C night; relative humidity (RH) ≥ 90%), container-grown, monoembryonic cultivars were water stressed through defi cit irrigation for 14 days, resulting in an average leaf xylem water poten-tial of −3.9 MPa (Davenport, 1992; Núñez-Elisea and Davenport, 1992a, 1994b). Following resumption of irrigation, all trees grew vegetatively. Sim-ilarly, only vegetative growth was obtained when container-grown trees were deprived of irrigation for 36 days during summer, although leaf xylem water potentials of −3.78 MPa were attained (Núñez-Elisea and Davenport, 1994b). Water stress imposed on plants during the cool autumn months

T.L. Davenport114
(night temperatures < 15°C) do not increase the proportion of apical buds forming infl orescences, but expedited shoot initiation after rewatering (Núñez-Elisea and Davenport, 1994b). These results demonstrated that cool temperatures provide inductive conditions, whereas relief of water stress accelerated shoot initiation under cool, inductive temperatures. Flowering was delayed when container-grown monoembryonic mangoes were water-stressed at 18°C day/15°C night (Chaikiattiyos et al., 1994). Water-stressed trees held at 29°C day/25°C night did not fl ower.
Mango trees growing in the low-latitude tropics may fl ower after an extended period of mild water stress (Harris, 1901; Collins, 1903; Kinman, 1918; Gangolly et al., 1957; Gangolly, 1960; L.B. Singh, 1960). Pongsomboon et al. (1991) observed fl owering in fi eld-grown trees in the tropics following 6 weeks of withholding water. The primary impact of water stress appears to be prevention of shoot initiation during stress. The accumulating age of stems is greater in water-stressed trees than in trees maintained under well-watered conditions that promote frequent vegetative fl ushes (Davenport, 1992, 1993; Schaffer et al., 1994). This delay in fl ushing may provide more time for accu-mulation of a putative FP (Schaffer et al., 1994) or reduction in the level of a putative VP (Davenport and Núñez-Elisea, 1997; Davenport, 2000). Some cultivars appear to be better adapted to such delays in growth and perform better in dry environments in the tropics.
Effect of N on fl owering
Subsequent to the discovery of ethephon to stimulate mango fl owering (Gon-zalez, 1923; Alcala and San Pedro, 1935), Barba (1974), Bueno and Valmayor (1974), Astudillo and Bondad (1978), Bondad et al. (1978), Bondad and Apos-tol (1979), Pantastico and Manuel (1978) and Bondad and Linsangan (1979) reported that KNO3 could be used for the same purpose. This has been exploited in the low- and mid-latitude tropics (Mosqueda-Vázquez and de los Santos de la Rosa, 1981; Mosqueda-Vázquez and Avila-Resendiz, 1985; Núñez-Elisea, 1985, 1986; Ou and Yen, 1985; Winston and Wright, 1986; Tongumpai et al., 1989; Goguey, 1993; Ravishankar et al., 1993; Sergent et al., 1996; Yeshitela et al., 2004b, 2005). The nitrate (NO3
–) anion is the active component of KNO3 (Bueno and Valmayor, 1974), and ammonium nitrate (NH4NO3) is twice as effective as KNO3 (Núñez-Elisea, 1988; Núñez-Elisea and Caldeira, 1988). In the low- and mid-latitude tropics, receptive trees respond by developing visible reproductive buds within 2 weeks after application. The effective spray concentration is 1–10% KNO3, depending on the age of the trees and climate. Two to four per cent KNO3 or calcium nitrate (Ca(NO3)2) and 1–2% NH4NO3 are effective for stimulating fl owering in most conditions. The physiological and temporal timing of application is important. Old trees, non-vigorous trees, and trees in which vegetative fl ushes have been discouraged by low water potentials produce the best response to NO3
– induction (N. Golez, personal communication, the Philip-pines, 1989).

Reproductive Physiology 115
Chemical bud forcing is most effective in the tropics where distinct wet and dry seasons prevail. The response to chemical bud forcing by NO3
− and ethephon diminishes at latitudes > 22° N or S (Mosqueda-Vázquez and de los Santos de la Rosa, 1981). Their effect may involve the decline of night temperatures from ≥ 20°C around the equator to ≤ 10°C between 22° and 27° N or S latitude during winter months or by late summer vegetative fl ushes. Trees in the wet or dry subtropics at 25° N or S have not responded to treat-ments (Davenport, 1993).
Stems must be suffi ciently mature, dark green with a minimum age of 4 months since the previous limp, red-leaf stage in easily induced cultivars and 5 months for more recalcitrant cultivars to obtain a reproductive shoot response in the low-latitude tropics (Davenport, 2003). Bueno and Valmayor (1974) indicated that leaves must be brittle when hand-crushed. Núñez-Elisea (1986, 1988) reported that stems must be at least 6 months old. Trees that experience autumn dry periods become responsive to treatments as early as October (northern hemisphere). Groups of stems within tree cano-pies are produced through asynchronous fl ushes of growth, and vary in age; only a few are responsive to the fi rst inductive spray. Subsequent biweekly applications cause fl owering in canopy sectors as they reach the age-depen-dent requirement for initiation. Early and out-of-season fl owering and fruit-ing can thereby be achieved.
KNO3 may be fl oral inductive in mango (Barba, 1974); however, trees in the upper latitude tropics typically fl ush vegetatively rather than produce bloom when either KNO3 or NH4NO3 is sprayed between June and Septem-ber (N. Golez, personal communication, the Philippines, 1989). The warm, rainy season producing frequent fl ushes of growth during this period is con-ducive to a vegetative response to the sprays. These results indicate that KNO3 and NH4NO3 stimulate shoot initiation but do not determine bud morphogenesis. In buds released after KNO3 or NH4NO3 treatments, the ratio of leaf-generated FP to VP and not NO3
– causes initiating buds to become reproductive. Kulkarni (1988b, 2004) suggested that the fl oral stimulus is present in stems when buds are forced in response to KNO3 and suggested that KNO3 may also sensitize buds to the fl oral stimulus. Davenport (2003), T.L. Davenport and J. Oleo (2006, unpublished data) and F. Ramirez and T.L. Davenport (submitted for publication) observed 100% vegetative shoots when 4% KNO3 was foliar applied to 2-month-old stems; whereas, applica-tion of the same spray treatment to 4.5-month-old stems on trees in the same orchards resulted in 100% reproductive shoots.
Trees with high leaf N levels rarely fl ower in the tropics. Lack of fl ower-ing is always due to frequent vegetative fl ushes of growth, especially during the rainy season. Mango trees must have leaf N levels of 1.4% or less in order to suppress frequent fl ushes of vegetative growth (Davenport, 2003). Leaf N levels of < 1.1% suppress frequent fl ushes but also provide insuffi cient nutri-tion to support good cropping. Thus, 1.1–1.4% N levels in leaves appear to be optimum for good commercial production and control of fl owering time in a managed orchard. The application of KNO3 to the foliage of the resting stems 4–5 months after the limp, red-leaf stage will cause a fl owering response.

T.L. Davenport116
Photoperiod
Flowering in most trees does not appear to be under photoperiodic control (Kozlowski et al., 1991). Mango cultivation is concentrated between 27° N and 27° S where the shortest annual photoperiod is c.10.5 h and the longest photoperiod is c.13.5 h. Cultivars in the upper-latitude tropics and subtropics fl ower during the winter when photoperiods are short; however, trees in the low-latitude tropics, where a 12-h photoperiod is nearly constant, can fl ower at any time of the year. Furthermore, fl owering on spring-initiated shoots in the subtropics occurs during summer (Schaffer et al., 1994). Studies have failed to demonstrate a correlation between 8-h photoperiods and fl owering (Maiti, 1971; Maiti and Sen, 1978; Maiti et al., 1978). Núñez-Elisea and Daven-port (1995) studied the effects of 11-, 12-, 13- and 24-h photoperiods at 18°C day/10°C night, or 11- and 13-h photoperiods at 30°C day/25°C night on fl owering of container-grown trees. Photoperiod had no effect on the fate of buds, and the promotive effect of cool temperatures on fl owering was inde-pendent of photoperiod. Photoperiods of 11-, 12- or 13-h with 18°C day/10°C night, caused fl owering in trees within 40 days. The 24-h photoperiod with 12-h thermoperiods of 18°C and 10°C caused fl owering of trees within 35 days. Photoperiods of 11- or 13-h at 30°C day/25°C night resulted in vegeta-tive growth only. With warm temperatures, vegetative shoots were produced in 17 days. These results confi rm that fl oral induction is caused by cool tem-peratures and not by short photoperiods and that warm temperature, not a long photoperiod, caused vegetative induction.
5.6 Hormonal Infl uence on Flowering
FP is a protein product of the FT gene in Arabidopsis (Corbesier et al., 2007) and the Hd3a gene in rice (Tamaki et al., 2007) and moves in phloem from leaves to buds; there is little evidence that phytohormones are directly involved as the FP. Phytohormones appear to be responsible for shoot initia-tion in conditions that are fl oral inductive.
Ethylene
Smudging has been utilized to stimulate mango fl owering in the Philippines. Only branches that attain suffi cient age respond to smudging by forming reproductive shoots (Acala and San Pedro, 1935; Bueno and Valmayor, 1974). Rodriguez (1932), investigating smoke-induced fl owering of pineapple, pro-posed that ethylene, generated by burning material, may stimulate fl ower-ing. Dutcher (1972) confi rmed that smoke from smudge fi res contained ethylene. Smudging and the use of ethephon in 1968 by F. Manuel (Barba, 1974) and others (Bondad, 1972, 1976) to promote mango fl owering sug-gested that endogenous ethylene is integral for fl oral induction (Barba, 1974; Bondad, 1976; Chadha and Pal, 1986). Ethephon effectively promotes fl owering

Reproductive Physiology 117
of mangoes under specifi c conditions in the low-latitude tropics (Davenport and Núñez-Elisea, 1997).
The involvement of endogenous ethylene in fl owering is supported by observations that indirectly link it to symptoms of ethylene production. Extrusion of latex from terminal buds occurs at the time of infl orescence ini-tiation, and epinasty of mature leaves near the apex during expansion of the panicle has been observed (Davenport and Núñez-Elisea, 1990, 1991). Both are symptoms of plants exposed to high ethylene levels (Abeles, 1973). Indi-rect support also comes from reports that KNO3-stimulated fl owering of mango is mediated by increased levels of endogenous ethylene (Thuck-Thye, 1978; Lopez et al., 1984). Mosqueda-Vázquez and Avila-Resendiz (1985) reported that the effi cacy of KNO3 was negated by cobalt chloride (CoCl2) and silver nitrate (AgNO3), which inhibit the synthesis and action of ethyl-ene, respectively, when sprayed 1–4 h after KNO3. Saidha et al. (1983) reported a gradual increase in endogenous leaf ethylene production as the season of fl oral initiation approached. Ethylene production by stems producing repro-ductive shoots was up to fi vefold that of resting stems.
Inconsistent (Pandey et al., 1973; Sen et al., 1973; Winston and Wright, 1986) or non-responsive results with ethephon (Pandey and Narwadkar, 1984; Ou and Yen, 1985; Pandey, 1989) or smudging (Sen and Roy, 1935), especially during warm, non-inductive conditions, have been reported. Dav-enport and Núñez-Elisea (1990, 1991) reported elevated ethylene production in mango stems in response to ethephon sprays without an accompanying fl oral response. Experiments were conducted during fl oral-inductive and non-inductive periods. Unlike Saidha et al. (1983), they observed no increase in ethylene production rates prior to or during panicle development.
The effect of ethylene on fl owering is unresolved. It is likely that ethylene stimulates shoot initiation by inhibiting auxin transport from leaves to buds and stems (Morgan and Gausman, 1966; Beyer and Morgan, 1971; Riov and Goren, 1979, 1980; Ramina et al., 1986). This may increase the ratio of cytoki-nin to auxin in buds and stimulate shoot initiation (Davenport, 2000). Other factors (i.e. cool temperatures or aged leaves) may be responsible for fl oral induction (Ona and de Guzman, 1982; Davenport, 1993).
Auxin
Although auxin may have a critical role in fl oral induction of mango (Chadha and Pal, 1986; Hegele et al., 2006), there is little supporting evidence. The application (L.B. Singh, 1961; Singh and Singh, 1963; Bakr et al., 1981; Pandey and Narwadkar, 1984) and analysis of auxin in leaves (Paulas and Shanmu-gavelu, 1989; Sivagami et al., 1989), stems (Chen, 1987) and shoots (Chacko et al., 1972b) have been reported in relation to mango fl owering. These studies are inconclusive due to inconsistencies in purifi cation and analytical methodolo-gies (Davenport and Núñez-Elisea, 1997).
Auxin may indirectly stimulate root-produced cytokinins through initia-tion of new root growth. Auxin is transported basipetally from growing

T.L. Davenport118
shoots and leaves to roots (Goldsmith, 1968; Cane and Wilkins, 1970; Wilkins and Cane, 1970; Goldsmith and Ray, 1973; Lomax et al., 1995) and stimulates root initiation (Hassig, 1974; Wightman et al., 1980). The effi cacy of various auxins for stimulating adventitious rooting of mango marcots and cuttings was reviewed by Davenport and Núñez-Elisea (1997).
Auxin inhibits shoot initiation (Davies, 1995) and confers apical domi-nance by preventing axillary bud break. Leaf-produced auxin and petiolar auxin transport capacity declines as leaves age (Veen, 1969; Veen and Jacobs, 1969; Davenport et al., 1980). The interaction of decreasing auxin and accu-mulating cytokinins in resting buds may explain the cyclic nature of shoot initiation. The ratio of cytokinin to auxin levels in buds regulates shoot initiation (Skoog and Miller, 1957; Bangerth, 1994; Cline et al., 1997; Beveridge et al., 2003).
Cytokinins
Relationships between mango fl owering and the endogenous levels of cyto-kinins in leaves (Paulas and Shanmugavelu, 1989; Kurian et al., 1992), stem tips (Agrawal et al., 1980) and xylem sap (Chen, 1987) and the effect of cyto-kinin applications on bud break and shoot development have been reported. Chen (1985) described precocious fl owering of mango shoots in response to early October application of 6-benzylaminopurine (BA). Flowering was observed 1 month following application and 3 months later on non-treated trees. Núñez-Elisea et al. (1990) reported numerous reproductive shoots per stem in response to the synthetic cytokinin, thidiazuron, during cool, fl oral inductive conditions; however, numerous vegetative shoots per stem were initiated when thidiazuron was applied during warm, vegetatively induc-tive conditions. Early bud break was not achieved following foliar applica-tion of Promalin (commercial formulation of BA and gibberellins A4+A7) (Oosthuyse, 1991b), BA (A.K. Singh and Rajput, 1990) or kinetin (Singh and Singh, 1974).
Chen (1987) reported the lowest levels of putative trans zeatin and its riboside were translocated from roots during the vegetative shoot growth and resting stages, whereas the highest levels occurred during early fl ower-ing and full bloom. Paulas and Shanmugavelu (1989) observed no signifi cant difference in cytokinin levels of the fourth and fi fth leaves during resting bud and fl owering. Cytokinin levels in mango stem buds increased during expo-sure to cool, fl oral inductive temperatures (Bangerth et al., 2004). Agrawal et al. (1980) described 11 cytokinin-like substances isolated from stem tips of an alternate-bearing cultivar in ‘on’ and ‘off’ years. Kurian et al. (1992) reported a link between PBZ applications and reduction in cytokinins in mango leaves with treatments, perhaps caused by reduction in feeder root development and formation of thick, blunt roots (Bausher and Yelenosky, 1987; Peng et al., 1991; Burrows et al., 1992; Yelenosky et al., 1993). Concurrent with this response was suppression of bud initiation and reduced internode lengths for c.2 years.

Reproductive Physiology 119
The role of cytokinins in fl owering is unresolved due to sampling of dif-ferent organs at non-comparable times or conditions. The elevated cytokinin levels found prior to and during fl owering and the fl owering response to applied BA led to the conclusion that cytokinins are involved in fl owering of mango (Chen, 1985, 1987; Bangerth, 2006); however, such responses can be explained if cytokinins are involved in stimulation of bud break (i.e. shoot initiation) during fl oral inductive conditions.
A well-documented role for cytokinins in higher plants, especially evi-dent in vitro, is bud organogenesis (Skoog and Miller, 1957; Miller, 1963; Takahashi, 1986; Salisbury and Ross, 1992; Davies, 1995; Haberer and Kieber, 2002). The primary cytokinins in higher plants are trans zeatin, dihydrozeatin, isopentenyl adenine and their ribosides. They are translocated from roots and accumulate in resting buds (Hendry et al., 1982a, b) or can possibly be synthe-sized in nearby tissues as regulated by auxin (Nordstrom et al., 2004; Tanakaet al., 2006). Their rate of accumulation may relate to periodic root fl ushes that alternate with shoot fl ushes (Krishnamurthi et al., 1960; Bevington and Castle, 1986; Cull, 1987, 1991; Parisot, 1988; Williamson and Coston, 1989).
Gibberellins
Gibberellins are tetracyclic diterpenoid compounds that vary in biological activity according to the type and location of substituted side groups on a basic ent-gibberellane skeleton. The number of known gibberellins is > 100 (Pearce et al., 1994). Reproductive shoot initiation is suppressed in many woody angiosperms by gibberellic acid (GA3) (Pharis and King, 1985). GA3 inhibits mango fl owering (older literature reviewed in Davenport and Núñez-Elisea, 1997; Núñez-Elisea and Davenport, 1998).
GA3 inhibition of mango fl owering is correlated with the applied con-centration (Kachru et al., 1971, 1972) and may cause buds to develop vegeta-tively under fl oral-inductive conditions. Núñez-Elisea and Davenport (1991a, 1998) reported a delay in initiation of axillary shoots when GA3 was foliar applied to deblossomed stems during cool, fl oral inductive temperatures. Higher concentrations caused longer delays in shoot initiation. GA3 did not inhibit fl oral induction, so long as cool, inductive temperatures were present during axillary shoot initiation. Late initiating buds, which grew during warm, spring temperatures, however, formed vegetative shoots. Similar delays in reproductive shoot initiation in response to GA3 application was reported by Shawky et al. (1978) and Turnbull et al. (1996). Multiple applica-tions, even at lower rates, are more effective than a single application (Tomer, 1984; Turnbull et al., 1996; Davenport and Smith, 1997). GA3 treatment has been recommended in the Canary Islands to delay fl owering until the danger of frost has passed (Galán-Saúco, 1990). In the subtropics of Australia, it is used to prevent fl owering in newly planted trees during the spring so that the full growing period can be utilized for vegetative growth, thereby hastening orchard establishment (A.W. Whiley, personal communication, Queensland, 1996).

T.L. Davenport120
Response to GA3 varies among cultivars, growing conditions and timing of application (Tomer, 1984; Oosthuyse, 1995a; Turnbull et al., 1996; Sánchez-Sánchez et al., 2004). GA3 can delay shoot initiation beyond the fl oral induc-tive window, resulting in a vegetative fl ush when shoots develop in warm weather (Kachru et al., 1971, 1972; Núñez-Elisea and Davenport, 1991a, 1998; S. Gazit, personal communication, Israel, 1993). The variable response to GA3 may be related to levels of active gibberellin in buds at the time of applica-tion, inconsistent uptake or differential sensitivity of buds, depending on their position (apical versus axillary) or age (Núñez-Elisea and Davenport, 1991a, 1998). Effi cacy is related to the timing of application; immediately prior to normal shoot initiation appears to be most effective (Davenport and Smith, 1997).
Reports of endogenous gibberellins in mango tissues, especially in buds, are diffi cult to interpret with respect to a regulatory role in bud break or fl owering. Problems include sampling of tissues other than apical buds, i.e. whole stems (Tongumpai et al., 1991b), leaves (Paulus and Shanmugavelu, 1989; Sivagami et al., 1989) and xylem sap (Chen, 1987), or at times when developing shoots may contribute to the overall result (Chen, 1987). Pal and Ram (1978) tentatively identifi ed the presence of gibberellins A1, A3, A4, A5, A6, A7 and A9. Chen (1987) identifi ed gibberellins A1/3, A4/7, A5, A17, A20 and A29. The estimated levels of gibberellins in apical buds for 6 months prior to the fl owering season were reported to be higher in the ‘off’ year than in the ‘on’ year of an alternate-bearing cultivar (Pal and Ram, 1978). Chen (1987) reported the highest levels of gibberellins in xylem sap during leaf differen-tiation and lower concentrations during rest, panicle emergence and full fl owering. Tongumpai et al. (1991b) observed increasing levels of gibberellins in whole stems over the 16 weeks prior to vegetative shoot emergence and decreasing levels over the same period prior to panicle development. Gib-berellins A1, epi-A1, A3, A19, A20 and an unidentifi ed gibberellin in buds and leaves from shoot and stem tips of different ages have been quantifi ed (Dav-enport et al., 2001b). The detected gibberellins are members of the early 13-hydroxylation pathway of gibberellin synthesis (Takahashi, 1986; Pearce et al., 1994). Gibberellins A3 and A19 were the most abundant gibberellins in apical stem buds. The concentration of GA3 increased within buds with increasing age of stems, although concentrations of other GAs were variable. The concentration of GA3 did not change signifi cantly with age in leaves, whereas that of most of the other GAs declined. Davenport et al. (2001b) concluded that elevated GA3 levels in buds may enhance or maintain the synthesis or activity of endogenous auxin to maintain low cytokinin/auxin ratios and enhance inhibition of shoot initiation (Jacobs and Case 1965; Scott et al., 1967; Pharis et al., 1972; Ross et al., 1983; Law 1987; Law and Hamilton, 1989).
The roles of gibberellins and other phytohormones in shoot initiation and induction is unclear. Endogenous levels in buds and leaves must be cor-related with physiological events in individual stems. Experimental approaches should include examination of resting buds up to both vegetative and repro-ductive shoot initiation to avoid misinterpretation of results. Experiments

Reproductive Physiology 121
should utilize plants grown under defi ned conditions with specifi c environ-mental controls for evaluation of cause and effect. Finally, extraction and purifi cation protocols should include quantifi able internal standards and use of sensitive unambiguous analytical techniques.
Plant growth retardants
Plant growth retardants have been evaluated to stimulate early or more intense fl owering, especially in the ‘off’ year of alternate-bearing cultivars (Davenport and Núñez-Elisea, 1997). They are in three main classes: (i) the gibberellin transport inhibitor, daminozide (N-dimethylamino-succinamic acid), known as alar or B-Nine; (ii) the onium type, chloremquat chloride (2-chloroethyl trimethylammonium chloride), known as cycocel and CCC; and (iii) the steroid-synthesis-inhibiting triazoles, for example PBZ (PP-333), known as Cultar®, and uniconazole, known as XE-1019 or Sumagic (Rademacher, 1991, 2000a). The latter two classes of compounds inhibit ent-kaurene syn-thetase, an enzyme in the gibberellin synthesis pathway (Nickell, 1983; Dalziel and Lawrence, 1984; Rademacher, 1991, 2000a). Applying daminoz-ide results in increased gibberellin levels, perhaps due to the inability to dis-tribute it properly (Rademacher, 1991). Plant responses may depend upon whether target tissues are near the site of gibberellin synthesis or suffi ciently removed from it to be affected by the inhibited translocation.
Daminozide and cycocelThe effi cacy of daminozide and cycocel for increasing fl owering in the ‘off’ sea-son of alternate-bearing cultivars has been studied (Maiti et al., 1972; Mukhopad-hyaya, 1978; Rath and Das, 1979; Suryanarayana, 1980; Rath et al., 1982; Ou and Yen, 1985), together with their ability to stimulate early fl owering (Suryanarayana and Rao, 1977; Chen, 1985; Kurian and Iyer, 1993a, b). Enhanced, inconsistent fl owering occurs in response to these compounds, especially cycocel.
TriazolesPBZ is being used (except in the USA where it has not been cleared for use) to stimulate enhanced or early fl owering. It is best applied to the soil due to its low solubility, long residual activity and lack of effi cient foliar uptake (Rademacher, 2000b). PBZ applied as a soil drench (1–20 g active ingredient (ai)/tree) reduces internode lengths and causes earlier and enhanced fl ower-ing in mango trees (Hasdiseve and Tongumpai, 1986; Haw, 1986; Hongsb-hanich, 1986). Depending on climate, residual activity lasts for c.2 years (Kulkarni, 1988a). These results have been confi rmed in different locations in the tropics (Davenport and Núñez-Elisea, 1997; Yeshitela et al., 2004a, b). Nartvaranant et al. (2000) recommended soil application of PBZ at 1–1.5 g ai/m of canopy diameter to achieve fl owering in 90–120 days if the trees are stimulated to fl ush. Davenport (2003) observed that such treatments allowed a reduction of c.1 month in the time required for stem rest before stimulating them to initiate reproductive shoots using KNO3. PBZ also reduces alternate

T.L. Davenport122
bearing of some cultivars (Hillier and Rudge, 1991; Burondkar and Gunjate, 1993; Rao, 1997; Rao et al., 1997; Rao and Srihari, 1998; Vijayalakshmi and Srinivasan, 1999). Cultivars that tend to fl ower with minimal inductive impe-tus are more responsive and can be induced to fl ower out-of-season using PBZ (Tongumpai et al., 1989). Núñez-Elisea et al. (1993) demonstrated that application of PBZ and uniconazole advanced bud break of containerized trees in controlled environment chambers, but cool temperatures were neces-sary to induce fl owering. Initiated shoots were induced to be vegetative in warm temperatures. The greater proportion of purely reproductive panicles in treated plants (compared with controls) suggests that triazoles impact the level of a putative VP, probably a gibberellin. Whiley (1993) suggested a sec-ondary mechanism for the fl oral promotive action of PBZ on mangoes, not-ing inconsistent responses in the literature between cultivars, environments and application times.
Application of PBZ reduces the number of panicles, despite increased fruit set (Goguey, 1990). Davenport (1987, 1994) observed neither growth inhibition nor enhanced or early fl owering in response to root drenches or bark banding with uniconazole (1–5 g ai/tree) in trees growing in alkaline, calcareous soil. He reported that new shoot growth was stunted with extremely short internodes when trees were severely pruned soon after or as long as 3 years after treatment. Yield was severely reduced due to the lack of normal growth fl ushes. The growth stunting effect continued for 7 years after pruning. Davenport (1994) warned that use of triazole plant growth retar-dants for control of tree growth, fl owering or yield must be done with con-siderable caution, especially if severe pruning of the trees is anticipated. Residual uniconazole or PBZ applied as a soil drench or bark band is appar-ently retained in high concentrations in main scaffolding branches. In Cen-tral and South America, growers utilize PBZ annually to stimulate early fl owering. A test tree should be severely pruned to determine if the trees are affected by PBZ to anticipate the orchard response to later severe pruning.
Certain gibberellins (i.e. GA1) are necessary for shoot elongation. Inhibi-tion of bud break and shoot elongation in response to application of the growth retardants cycocel (Maiti et al., 1972) and triazoles (Kulkarni, 1988a; Burondkar and Gunjate, 1991, 1993; Tongumpai et al., 1991a; Kurian et al., 1992; Winston, 1992; Kurian and Iyer, 1993a, b; Núñez-Elisea et al., 1993; Wer-ner, 1993) have been reported. Elongation of panicles is inhibited, especially by high levels of triazoles (Kulkarni, 1988b; Winston, 1992; Davenport, 1994; Salomon and Reuveni, 1994). Infl orescences in treated trees may become compact, improving opportunities for disease and insect attack (Winston, 1992). Kurian et al. (1992) associated reduced cytokinin levels in leaves with inhibition of shoot initiation in plants treated with soil drenches of PBZ. Ele-vated, concentration-dependent levels of phenolic compounds were also found in resting apical buds of PBZ-treated trees (Kurian et al., 1994). They suggested that low cytokinin activity and high phenolic levels in buds con-tributed to inhibition of shoot initiation.
The combined impact of the gibberellin synthesis-inhibiting triazoles on shoot initiation, induction, and elongation implies that several different

Reproductive Physiology 123
gibberellins regulate specifi c activities in mango plants. This is supported by the inhibitory effect of GA3 on shoot initiation in contrast with early initia-tion of fl owering in triazole-treated trees. Compression of reproductive and vegetative shoot internodes may involve inhibition of GA1 synthesis. Stimu-lation of fl owering instead of vegetative growth during early initiation in triazole-treated plants in marginal or non-fl oral inductive conditions, sug-gests that the putative VP, a gibberellin other than GA3 or GA1, is reduced when gibberellin synthesis is inhibited.
5.7 Photoassimilate Infl uence on Flowering
Flowering may be regulated by C:N ratios with high levels being conducive to fl owering (Kraus and Kraybill, 1918). Photoassimilates reaching the apical bud from leaves was central to several theories of fl oral induction (Sachs, 1977; Bernier and Sachs, 1979; Bernier et al., 1981, 1993; Bernier, 1988) includ-ing mango (Mallik, 1951; L.B. Singh, 1960; Chacko and Ananthanarayanan, 1982; Rameshwar, 1989) and other species (Allsopp, 1965; Sachs, 1977; Mishra and Dhillon, 1978; Ramina et al., 1979; Bernier et al., 1981; Sachs and Hackett, 1983). The theory of photoassimilate diversion to the apical bud (Sachs et al., 1979) is the basis for the carbohydrate-regulated fl owering models (see below). Sugars are utilized during panicle development (Ravishankar and Mohan Rao, 1982). Starch reserves and C:N ratios have been correlated with fl owering (Mishra and Dhillon, 1978; Suryanarayana, 1978a, b, c; Chacko and Ananthanarayanan, 1982; Whiley et al., 1988, 1989, 1991; Robert and Wolsten-holme, 1992; Shivashankara and Mathai, 1995), and the subject has been reviewed (L.B. Singh, 1960, 1972; Singh, 1979; Chacko, 1986, 1991; Chadha and Pal, 1986; Pandey, 1989). Starch accumulation during extended periods of canopy rest prior to fl owering provides supportive evidence, but there is little consensus regarding the role of carbohydrates and N in fl owering.
Photoassimilates may be necessary for fl oral induction. If a fl origenic promoting gene product is synthesized in leaves in small amounts, it must be able to move to those buds via phloem. Due to the requirement for high sol-ute concentrations to motivate phloem fl ow, the low concentration of the FP could not cause fl uid movement through sieve tubes of the phloem on its own. The much higher concentrations of photoassimilated sugars carried by water loading into the phloem in leaves passively transports the FP towards the various sinks, including respiring buds, where they are utilized for fl oral induction.
5.8 Horticultural Manipulation of Flowering
Mango fl owering can be stimulated by trunk or branch girdling, defoliation and deblossoming (Pandey, 1989). Responses vary with cultivar and envi-ronment. Trunk girdling of mango trees to promote fl owering is inconsis-tently effective (Kinman, 1918; Gaskins, 1963; Winston and Wright, 1986) and

T.L. Davenport124
can be detrimental to trees, especially if done in subsequent years. It has been shown to increase fl owering in the ‘off’ year of alternate-bearing cultivars; however, it either has no effect or is only marginally benefi cial in the ‘on’ year (Mallik, 1951; Rath and Das, 1977, 1979; Rath et al., 1982; Rameshwar, 1989). Girdling in late summer or early autumn usually results in less vegetative fl ushing prior to fl owering, which is enhanced in trees exposed to marginally inductive conditions. Tree response is dependent on the width of the girdle. Narrow cuts result in either a short-term or no response; whereas, girdles that are too wide can kill trees if they do not close within a reasonable time. Gir-dling cuts phloem transport, starves roots of photoassimilates and interrupts auxin transport to roots (Morris and Thomas, 1978; Hegele et al., 2004). These are detrimental to root development and can alter the bud cytokinin:auxin ratio due to reduced cytokinin translocation from roots. This results in delayed shoot initiation, which can impact the level of the age-dependent, putative VP when shoot initiation occurs. The delay in fl ushing, therefore, enhances fl owering. Defoliation of trees stimulates fl ushing, possibly by altering the cytokinin:auxin ratio in buds because leaves are the primary source of auxin.
Bloom delay is useful where recurring temperatures < 15°C stimulate fl owering, but continued low temperatures hamper pollination, fertilization and early fruit development (Young and Sauls, 1979; Wolstenholme and Mul-lins, 1982a, b; Whiley et al., 1988; Galán-Saúco et al., 1991). Low temperatures cause production of seedless, underdeveloped fruit. Deblossoming stimu-lates growth of dormant axillary buds, which produce infl orescences if initia-tion occurs under conditions conducive to fl oral morphogenesis (Singh et al., 1974). Late blooms can also be promoted with ethephon (Chadha and Pal, 1986; Galán-Saúco et al., 1993) and cycloheximide (Pal and Chadha, 1982; Shu, 1993). Sprays of these compounds cause abscission of apical panicles, thereby releasing dormant axillary buds that will produce infl orescences under cool, fl oral-inductive conditions of the subtropics.
5.9 Conceptual Flowering Models
Several conceptual models have been proposed that attempt to explain the physiological basis of mango fl owering. Each model should be viewed as a collection of integrated ideas, which require rigorous testing for validity within the context of the models. A useful model should explain how fl ower-ing and vegetative growth is regulated in all cultivars and races in both humid and dry climates in the tropics and subtropics. It should also be sup-ported by the preponderance of research evidence. The fl owering models are either carbohydrate-regulated or hormone-regulated.
Carbohydrate-regulated fl owering models
Cull (1987, 1991) presented a holistic approach for tree crop research and management to maximize sustainable fruit production. This concept is based

Reproductive Physiology 125
on the axiom of genotype/environment adaptability expressed through the annual phenological cycle and is an alternative to the traditional reductive-based approach to crop research and development. He proposed that pro-ductive cultivars follow ‘normal’ phenological patterns from year to year due to gene expression in specifi c environments. A signifi cant departure from this pattern results in reduced or total crop failure. Annual variations in climatic conditions that alter tree phenology can be countered by strategic applications of nutrients, water, plant growth regulators and canopy manip-ulation. The model does not attempt to explain the intricacies of shoot ini-tiation or induction, but takes a broader approach in detailing temporal relationships between reproductive and vegetative growth that lead to reli-able cropping.
The fundamental principle underlying this model is that yield is the product of photoassimilate (carbohydrate) accumulation and subsequent redistribution during the annual growth cycle. Accumulated photoassimi-lates would drive critical growth events that require higher levels of resources than are available from current photoassimilate supplies. Cultivars that pro-ceed with balanced reproductive, vegetative and rest phases are more likely to have suffi cient carbon resources to meet periods of critical demand and therefore will sustain high yields. The model illustrates fl oral initiation as occurring after a 2- to 3-month rest period during autumn/winter when a critical threshold level of carbohydrate is reached in buds together with a putative fl oral stimulus. Bud break during cool weather results in a high per-centage of fl owering stems (> 90%; Searle et al., 1995) with fruit set and reten-tion suppressing vegetative fl ushing on individual fruiting stems until after they have matured and harvested. Shortly after harvest, vegetative buds are released and a fl ush of growth occurs during the summer, which is followed by a period of strong root growth. The regenerated canopy becomes a source for rebuilding photoassimilate reserves that are stored in the roots, bark and resting stems. In the tropics, growth events are less orderly, and cultivar and management skills are of greater importance. The pre-fl owering rest period is usually achieved by drought as temperatures remain above the critical threshold for shoot growth (15°C) (Whiley et al., 1989). Other practices used with some success to enforce canopy quiescence are girdling and the applica-tion of growth retardants.
The principles of phenological modelling have been advanced into work-ing pheno/physiological models for avocado (Whiley, 1994) and mango (Searle et al., 1995). The advantage of this approach is that the annual pro-gression of growth cycles with associated physiological changes is studied concurrently, adding a further dimension to our understanding of tree growth and development. Information gathered in this way provides opportunities to identify and assess critical yield-limiting events, which in the case of man-goes largely relates to the success or failure of fl owering.
Chacko (1991) proposed a fl owering model driven by assimilate supply and diversion to apical meristems (Fig. 5.6). Environmental conditions, such as water stress, cool temperatures, high evaporative demand, fl ooding, girdling and other events that inhibit vegetative growth result in a shift in
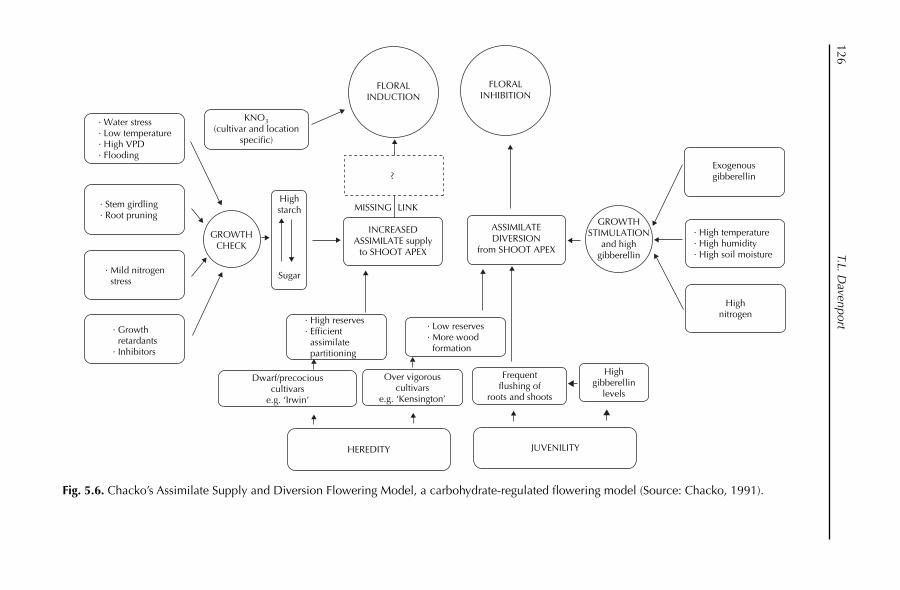
T.L. D
avenport126
FLORALINDUCTION
KNO3(cultivar and location
specific)
Highstarch MISSING
INCREASEDASSIMILATE supply
to SHOOT APEX
ASSIMILATEDIVERSION
from SHOOT APEX
GROWTHSTIMULATION
and highgibberellin
Exogenousgibberellin
LINK
?
GROWTHCHECK
Sugar
· Water stress· Low temperature· High VPD· Flooding
· High temperature· High humidity· High soil moisture
Highnitrogen
Highgibberellin
levels
JUVENILITYHEREDITY
Over vigorouscultivars
e.g. ‘Kensington’
Dwarf/precociouscultivars
e.g. ‘Irwin’
Frequentflushing of
roots and shoots
· Stem girdling· Root pruning
· High reserves· Efficient assimilate partitioning
· Low reserves· More wood formation
· Mild nitrogen stress
· Growth retardants· Inhibitors
FLORALINHIBITION
Fig. 5.6. Chacko’s Assimilate Supply and Diversion Flowering Model, a carbohydrate-regulated fl owering model (Source: Chacko, 1991).

Reproductive Physiology 127
carbohydrate partitioning and a diversion of soluble assimilates to stem api-ces. The elevated carbohydrate status in buds, together with a fl oral stimulus, results in fl oral induction. Vigorously growing cultivars and juvenile plants have low starch reserves (Whiley et al., 1988, 1989, 1991) and a diversion of soluble assimilates from stem apices results in fl oral inhibition. Conditions that promote vegetative growth, i.e. high temperature and moisture, high gibberellins and N, also lead to fl oral inhibition. Experiments involving chemical girdling of trees are based on this model (Blaikie et al., 1999).
Hormone-regulated fl owering models
Tri-factor Hypothesis of FloweringExtensive work on movement of the putative fl oral stimulus across grafts from donor to receptor stems (Kulkarni, 1986, 1988b) and the inhibitory infl u-ence of fruit on subsequent fl owering (Kulkarni and Rameshwar, 1989) form the basis of a fl owering model proposed by Kulkarni (1991): the Tri-factor Hypothesis of Flowering in mango (Kulkarni, 2004). This theory (Fig. 5.7) proposes an interactive role for a putative, cyclically produced fl oral stimu-lus in leaves, a fl oral inhibitor in leaves and fruits, and bud activity during the fl oral cycle. During dormancy following a vegetative cycle, genetic and
Flowering promotersynthesized in theleaves in the floral
cycle
Flowering inhibitorand vegetative
promotersynthesized in the
leaves and possiblyother organs
Bud activityin synchrony with
the floral cycle
Pure panicles Mixed leafy panicles Vegetative flush
Genetic and Environmental Factors
Fig. 5.7. Kulkarni’s Tri-factor Hypothesis of Flowering in mango, a hormone-regulated fl owering model (Source: Kulkarni, 2004).

T.L. Davenport128
environmental factors determine the level of synthesis of the putative fl oral stimulus. Flowering occurs only if certain correlative factors are present, for example if the receptor bud becomes active. If fruits are or have been recently present on the stem, vegetative growth will result. Presence of the putative fl oral inhibitor in leaves interferes with expression of the fl oral stimulus resulting in vegetative growth. The level of the fl oral stimulus determines the response: high levels give rise to normal panicles, intermediate levels give rise to mixed panicles and low levels result in vegetative growth.
Comprehensive Conceptual Flowering ModelThis is a model of fl owering involving the various classes of phytohormones (Davenport, 1992, 1993, 2000, 2003; Davenport and Núñez-Elisea, 1997) (Fig. 5.8) based on many lines of experimental evidence as well as on research of other tropical and subtropical fruit crops with similar phenological cycles (Menzel, 1983; Davenport, 1990, 1992; Menzel et al., 1990; Menzel and Simp-son, 1994; Davenport and Stern, 2005). Focusing on events occurring in indi-vidual buds, it is applicable to monoembryonic and polyembryonic cultivars in the tropics and subtropics and attempts to explain the physiological basis for the annual progression of the phenological cycle.
SHOOT FORMATION. Two distinct events must occur for vegetative or repro-ductive growth to occur in resting apical or lateral buds of mango: (i) the
Mango flowering model
PHOTOASSIMILATES FRUIT
MIXED SHOOT GENERATIVE SHOOT
CHILLING TEMP.OTHER FACTORS?
WATER STRESS
PROMOTERIN LEAVESINDUCTION
VEGETATIVE SHOOTAUXINGIBBERELLINS
SHOOT INITIATION
ROOT INITIATION
ROOTS CYTOKININS
GIRDLING
CHILLING TEMP.
STORAGE CARBOHYDRATES
PRUNINGDEFOLIATIONNITRATE SPRAYETHYLENE
FREQUENTVEGETATIVE
GROWTH
AUXINGIBBERELLINS A3
Ax
GA1GA3GAx
Fig. 5.8. Davenport’s Comprehensive Conceptual Hormone-regulated Flowering Model (Source: Davenport and Núñez-Elisea, 1997; Davenport, 2000). Single lines indicate promotive impact. Double lines indicate inhibitory impact.

Reproductive Physiology 129
bud(s) must be initiated to grow (shoot initiation); and (ii) at the time of ini-tiation, shoot development (i.e. vegetative, mixed, or generative) is deter-mined (induction). Although conditions for fl oral induction may be present prior to shoot initiation, determination of that inductive condition in buds is not made until initiation occurs. Initiation and induction events are regulated by different signals and each may be manipulated by different stimuli. Removing the apical whorl of leaves or tip pruning physiologically mature stems stimulates shoot initiation in apical or lateral buds, respectively. If containerized plants are maintained in warm temperatures (30°C day/25°C night) following initiation, vegetative shoot growth is induced. If they are kept under cool conditions (18°C day/10°C night), initiating shoots are induced to be generative. In either of the two temperature regimes without pruning, they do not initiate shoots until the natural fl ushing event occurs much later. They become vegetative or reproductive according to the tem-perature at the time of shoot initiation. If transferred from cool to warm tem-peratures before shoot initiation, new shoot growth is induced to be vegetative. Induction is therefore determined at the time of shoot initiation, and plants rapidly lose their fl oral inductive potential when removed from the cool envi-ronment. Determination of shoot type can be reversed during morphogenesis by transferring containerized trees from warm-to-cool or cool-to-warm con-ditions (Batten and McConchie, 1995; Núñez-Elisea et al., 1996).
INITIATION CYCLE. The cyclic initiation of vegetative or reproductive shoots is common to all mango cultivars. Developing vegetative shoots are rich sources of auxins and gibberellins, which may be inhibitors in an internal cycle that regulates shoot initiation. Auxins are actively transported basipe-tally to roots from production sites in stems (Goldsmith, 1968; Cane and Wilkins, 1970; Wilkins and Cane, 1970; Goldsmith and Ray, 1973), and they stimulate adventitious root growth in mango and other crops (Hassig, 1974; Wightman et al., 1980; Sadhu and Bose, 1988; Rajan and Ram, 1989; Núñez-Elisea et al., 1992). Elevated auxin synthesis in periodically fl ushing shoots is likely to form a concentrated pulse of auxin, which inhibits recurring bud break and moves basipetally to the roots. This pulse of elevated auxin may stimulate initiation of new root fl ushes following each vegetative fl ush. Alteration of root and shoot growth occurs in mango (Krishnamurthi et al., 1960; Cull, 1987, 1991; Parisot, 1988) and other tropical and subtropical trees (Bevington and Castle, 1986; Williamson and Coston, 1989; Ploetz et al., 1991, 1993). Timing of the root fl ush may depend on the distance from stems to roots, the physiological condition of the transport path, and environmental conditions (i.e. temperature or water relations).
New roots that develop following growth stimulation are a primary source of cytokinins (Davies, 1995). Cytokinins are transported passively to stems via the xylem sap in all plants and are active in bud break (Went, 1943; Kende and Sitton, 1967; Sitton et al., 1967; Itai et al., 1973; Haberer and Kieber, 2002). Cytokinins stimulate shoot initiation in mango (Chen, 1985; Núñez-Elisea et al., 1990) and other plants (Oslund and Davenport, 1987; Belding and Young, 1989; Williamson and Coston, 1989; Davenport, 1990; Davies, 1995;

T.L. Davenport130
Henny, 1995). Auxin inhibits shoot initiation (Davies, 1995) and confers api-cal dominance by preventing axillary bud break. Leaf-produced auxin and petiolar auxin transport capacity declines as leaves age (Davenport et al., 1980). Auxin and cytokinins may therefore be involved in the periodic cycle of bud break.
A critical balance of these two phytohormones, possibly modulated by GA3, may regulate shoot initiation. During a rest period, the inhibitory action of auxin transported to buds decreases with time; whereas, cytokinin lev-els in buds increase (Chen, 1987). When a critical cytokinin/auxin ratio is achieved, the buds are stimulated to grow, thereby resetting the cycle with initiation of new shoots. The interaction of auxin and cytokinin to control bud break in plants is a concept that is supported by molecular studies (see review by Nordstrom et al., 2004). Moreover, initiation of shoot growth occurs following pruning, defoliation or the application of thidiazuron (Núñez-Elisea et al., 1990). Vigorous cultivars (Whiley et al., 1989) and young, small trees under vegetatively promotive conditions fl ush frequently with only short periods of rest; however, this cycle slows with age. Old centennial trees fl ush infrequently (N. Golez, personal communication, the Philippines, 1989).
Foliar or soil-applied NO3− stimulates initiation of reproductive shoots
only if applied after resting stems have attained an age to overcome any veg-etatively inductive infl uence. In contrast, high N in soils leads to high N lev-els in leaves resulting in frequent vegetative fl ushes. The mechanism whereby NO3
− stimulates shoot initiation is unknown.Seeds are rich sources of auxin and gibberellins, which contribute to the
strong inhibition of bud break commonly observed on fruit-bearing mango stems. The longer that fruit are attached to stems, the longer the postharvest inhibition may last in the stem (Kulkarni and Rameshwar, 1989; Kulkarni, 1991).
Water stress inhibits shoot initiation by its direct impact on cell division and elongation possibly by interfering with translocation of cytokinins from roots. There is little evidence that water stress is directly involved in induc-tive processes. During water stress, roots continue to grow and produce cyto-kinins (Itai and Vaadia, 1965; Itai et al., 1968; Wu et al., 1994). Reduced xylem fl ux due to limited soil hydration, and transpiration due to increased sto-matal resistance during water stress may reduce the amount of cytokinins reaching stems. After rewatering, the increased levels of cytokinins in roots may translocate to and accumulate in buds. Auxin synthesis and transport from leaves are reduced during water stress (Davenport et al., 1980) and may require several days for correction after rewatering. This rapid shift in the cytokinin/auxin ratio of buds may explain the shooting response that occurs soon after relief of water stress. GA3 may act with auxin to inhibit shoot ini-tiation (Davenport et al., 2001b). Early fl owering in plants treated with PBZ may be a response to lowered gibberellin levels, thus lowering the level of initiation inhibitor.
This model could explain why sectors of tree canopies fl ush in the trop-ics. Mango trees fl ush often and synchronously throughout the canopy when they are young. With advancing age, the frequency of fl ushing is reduced

Reproductive Physiology 131
and synchrony is lost, resulting in sporadic fl ushes of vegetative or reproduc-tive growth in sections of the canopy. As the distance between stems and roots increases, the time required for transport of the putative pulses of ele-vated auxin levels to roots, formed during a vegetative fl ush, is increased. Groups of stems exhibiting simultaneous fl ushing ultimately connect to a common branch. Dye trace studies indicate that water transport remains in strict phylotaxic alignment from secondary roots to the canopy, even in large trees (T.L. Davenport, unpublished results, Florida, 1991). Unless disturbed by girdling or by pruning of branches or roots, specifi c branches in the can-opy communicate only with those roots in phylotaxic alignment with them. The hormone transport time may vary among sections of the canopy as the tree grows. This generates individual initiation cycles in sections of the can-opy that are separately maintained unless resynchronized with the rest of the tree following a canopy-wide environmental trigger.
Synchronization of growth throughout trees occurs following exposure to low temperature, water stress, light pruning of the entire tree and any condition that would increase the postulated cytokinin/auxin ratio in buds throughout the canopy. An increased ratio may occur by inhibiting auxin transport from leaves to buds, or increasing cytokinin translocation from roots to stems. Winter in the subtropics would reduce auxin transport; whereas, water stress in the tropics may impact the availability of cytokinins from roots and auxin from leaves. The intensity of the initiation response (i.e. synchronization of fl ushes in the canopy) may be regulated by decreased auxin transport at low temperatures, the base level of which may be deter-mined by the age of individual stems. Passage of a strong, extended cold front during subtropical winters produces synchronized fl owering. Milder winters with weak cold fronts result in asynchronous fl owering in sections of trees. The oldest sectors of canopies fl ower fi rst, followed by sectors bearing sequentially younger fl ushes in subsequent cold fronts. Vegetative fl ushes occur when night temperatures are > 18°C for signifi cant periods between cold fronts.
INDUCTION SWITCH. Floral or vegetative induction is possibly governed by the interactive ratio of a FP that is up-regulated in low temperatures to an age-regulated VP in leaves at the time of shoot initiation. High FP:VP ratios would be conducive to induction of generative shoots, low ratios conducive to veg-etative shoots and an intermediate ratio of the two would be conducive to mixed shoots. Regardless of the endogenous levels of the two components perceived in buds at the time of initiation, fl owering and vegetative growth responses can best be explained by the ratio of the two.
Although the putative FP seems to be up-regulated during leaf exposure to cool temperatures (< 18°C), there appears to be a basal level present at all times in leaves exposed to higher temperatures. Flowering of mango occurs in low-latitude tropics lacking cool night temperatures when stems become suffi ciently aged so that the ratio of the basal level of resident FP to decreas-ing VP increases to a critical threshold to provide fl oral induction when shoots are initiated. This could explain how fl owering on non-synchronized

T.L. Davenport132
branches may occur at any time of the year in trees growing in low-latitude tropics. High proportions of mixed shoots are commonly found in these con-ditions, indicating the marginally fl oral-inductive ratios present under these conditions. In contrast, fl owering in younger stems having higher levels of VP is observed only when initiation occurs in cool, fl oral-inductive tempera-tures. More fl owering occurs throughout the canopy when stems are exposed to cool temperatures, attributable to the higher ratio of up-regulated FP to resident VP.
Genetic differences in base levels of the putative FP and/or VP or the receptors of these components could explain the range in fl owering responses in tropical and subtropical cultivars and why a cultivar grown in an environ-ment different from that in which it was selected is less productive. Cultivars selected in the subtropics usually fl ower as well in the low-latitude tropics as those selected in the tropics. Cool temperatures in the subtropics sometimes cause earlier fl owering in tropical cultivars than those selected in the sub-tropics. Kulkarni (1991) demonstrated that several multi-fl owering cultivars can induce fl owering in receptor graft plants and cause a range of the fl ower-ing response of the receivers to donors. Some cultivars may produce higher base levels of putative FP than others. These are the same cultivars that read-ily fl ower under warm temperatures and fl ower early during cool winter months. The Comprehensive Conceptual Flowering Model suggests that fl owering can occur at any time in any cultivar regardless of origin so long as stems are suffi ciently old to reduce the VP level to below the critical FP/VP ratio when initiation occurs.
Although the putative FP, perhaps a product of an ortholog of the Arabi-dopsis FT gene, has not been identifi ed, the VP may be a gibberellin. Triazoles and other plant growth retardants that inhibit gibberellin synthesis, promote strong and out-of-season fl owering under conditions that would normally be marginally or non-fl oral inductive.
PHOTOASSIMILATES. Photoassimilates produced by leaves provide carbohy-drates essential for development of roots and other plant organs, including fruit. They are either used immediately by the nearest sinks (Finazzo et al., 1994) or are stored in locations throughout the tree to be used when demand for carbon resources exceeds the existing photosynthetic supply (Whiley et al., 1988, 1989, 1991). A direct role for carbohydrates in shoot initiation or induction is not part of this model, although they facilitate mass fl ow in phloem from leaves to passively carry the FP to buds.
ALTERNATE BEARING. High levels of auxin and gibberellins produced in seeds possibly inhibit shoot initiation on fruit-bearing stems for weeks or months following fruit removal. Rapid production of new shoots following light pruning of fruit-bearing stems after harvest indicates that residual levels of auxin and gibberellins linger only in the rachis and last intercalary unit. If fruit are not set on the lingering rachis, there is less inhibition. Heavy fruit set in 1 year impacts the timing of subsequent shoot initiation on the large num-ber of fruit-bearing branches. Substantial delays in subsequent vegetative

Reproductive Physiology 133
fl ushes until close to the normal fl owering period impact the fl owering abil-ity of young shoots. This may explain the occurrence of chronic alternate bearing in some cultivars.
5.10 Floral Management
The Comprehensive Conceptual Flowering Model is consistent with growth and development patterns of mango trees in the tropics and subtropics. It provides a reasonable explanation for the various events in the phenologi-cal model of Cull, predicts what will happen under a defi ned set of cir-cumstances and is being used to develop strategies for consistent mango fl owering. Flowering can be obtained at any time of the year in a fl owering management programme (Davenport, 2003).
The fl owering management programme begins each season with tip pruning of the entire canopy of orchard trees. Tip pruning can be done imme-diately after harvest to move production forward in the following year or c.1–2 months after harvest, depending upon cultivar, in order to achieve har-vest at the same time as the previous year. If suffi cient soil water is available at the time of pruning, a vegetative fl ush will occur on all pruned stems c.1 month later. The number of new shoots that will mature to become stems will be fi ve- to eightfold greater than the original number of pre-pruned stems due to initiation of many lateral vegetative shoots on each stem. This increase in terminal stem number in the canopy will be refl ected in a concomitant increase in yield. The frequent fl ushes that can cause an early second fl ush of vegetative growth tend to be suppressed.
The new stems must not fl ush a second time until at least 5 months after pruning (Davenport, 2006). If they fl ush within 3–4 months after pruning, they will be induced to be vegetative. Pre-prune leaf N levels in the stems must be 1.1–1.4% in order to suppress a second fl ush of vegetative growth during the rainy season. Mild water stress after the post-prune fl ush during the dry season will suppress a second, undesired vegetative fl ush when leaf N levels are above the optimum range. Pruning near the end of the dry sea-son in non-irrigated or furrow-irrigated trees should be avoided. Transition from dry to wet season 2–3 months after pruning causes a rain-stimulated vegetative fl ush prior to achieving suffi cient age of stems from the last fl ush. Test sprays of 4% KNO3 on two to three representative trees should be applied 5 months (for easily induced cultivars) and 6 months (for more dif-fi cult to fl ower cultivars) after pruning. If no developing shoots occur within 2 weeks, the spray is repeated. A fl owering response is usually evident after the second application. The other trees that were pruned on or near the same date can then receive the foliar spray and will respond by synchronized fl ow-ering. Although Davenport (2003) described the appropriate timing of PBZ in a fl owering management programme, it is not recommended because fl owering can be achieved without it. For orchard trees to be amenable to tip pruning, effi cient spray application of KNO3 and easy harvesting, they should be no taller than 4 m. Pruning to rejuvenate large mango trees and

T.L. Davenport134
properly shape trees for the annual fl owering management programme is recommended (Davenport, 2006).
5.11 Floral Biology
Detailed descriptions of generative shoots were reviewed in Davenport and Núñez-Elisea (1997). Juliano and Cuevas (1932) described ovary devel-opment.
Sex ratio
Sex ratio (i.e. the proportion of perfect to staminate fl owers) is a variable component within panicles, trees and among cultivars. This ratio varies with cultivar, but is usually < 50% (Davenport and Núñez-Elisea, 1997). Most per-fect and staminate fl owers are borne in the proximal portion of panicles due to their architecture (Musahib-ud-din and Dinsa, 1946; Cobin, 1950). The variability in the perfect/staminate fl ower ratio may be governed by physi-ological and environmental conditions. Most studies indicate that although the total number of fl owers is substantially less in the distal half of panicles, there is a greater proportion of perfect fl owers in this region (Davenport and Núñez-Elisea, 1997); however, this condition may be reversed in some culti-vars (Hussein et al., 1989).
Perfect fl owers tend to form in the terminals of individual infl orescences while staminate fl owers are displayed in the earlier forming fl owers located closer to the panicle axis. When panicles begin to elongate in the lower infl o-rescences, only staminate fl owers form and the perfect fl owers form at the terminus of each lateral infl orescence. As more distally located lateral infl o-rescences begin elongation and anthesis, they too fi rst display staminate fl owers before perfect fl owers. These infl orescences are progressively shorter than previously formed proximal infl orescences, and there are fewer stami-nate fl owers. The fi nal vertical spike of the panicle is composed almost exclu-sively of perfect fl owers. Flowers abscise soon after anthesis, thereby shifting the sex ratio. The sex ratio should include sex type of all fl owers in each panicle; however, sex ratios are normally determined at some arbitrary moment during panicle development. Thus, the sex ratio is naturally vari-able, increasing from extremely low to extremely high values so that timing of observations during panicle development is critical.
Environmental determinants of sex ratio
Tropical cultivars yield poorly in the subtropics due to a small proportion of perfect fl owers on infl orescences (Singh and Singh, 1959; Singh et al., 1965; Singh, 1971). Cool weather during infl orescence development contributes to fewer perfect fl owers (Naik and Mohan Rao, 1943; Singh et al., 1965, 1966).

Reproductive Physiology 135
Infl orescences that emerge during the middle and end of the fl owering sea-son produce two and seven times more perfect fl owers, respectively, than the early breaking infl orescences (Majumder and Mukherjee, 1961; Singh et al., 1966). This response correlates with higher temperatures later in the fl ower-ing season. In controlled-environment studies, low temperatures (15°C day/10°C night) reduced the proportion of perfect fl owers, particularly in tropical, polyembryonic cultivars relative to subtropical, monoembryonic cultivars (Sukhvibul et al., 1999).
Physiological determinants of sex ratio
Endogenous factors affect the ratio of perfect to staminate fl owers. Bajwa et al. (1956), Majumder and Mukherjee (1961) and Joubert et al. (1993) reported that lateral infl orescences on mixed shoots carried higher proportions of per-fect fl owers. Infl orescences on older trees produce higher proportions of per-fect fl owers than those on young trees (Naik and Mohan Rao, 1943; Majumder and Mukherjee, 1961; Chacko and Randhawa, 1971; Pandey, 1989). This also occurs in infl orescences borne on grafted compared with seedling trees (Musahib-ud-din and Dinsa, 1946). The effect of tree maturity or rootstocks on sex ratio of fl owers is not understood. Panicles carried within the canopy of some cultivars (Majumder and Mukherjee, 1961; Singh et al., 1966) or on particular sides of the canopy (Mukherjee, 1953; Majumder and Mukherjee, 1961) have been reported to have higher proportions of perfect fl owers.
Application of some hormones and growth regulators alters the sex ratio of infl orescences. GA3, applied at concentrations of 50–100 mg/l just prior to infl orescence shoot initiation, substantially reduces the proportion of perfect fl owers (Maiti, 1973), as do combination sprays of urea (0, 3 and 6%) and GA3 (0, 15 and 30 mg/l) (Rajput and Singh, 1989). Soil-applied PBZ (10 g ai/tree) signifi cantly increases the ratio of perfect/staminate fl owers (Kurian and Iyer, 1993a). Increases in fl oral ratio also occur with daminozide, whereas maleic hydrazide either had no effect or lowered the ratio (Singh et al., 1965; Subhad-rabandu, 1986). Foliar application of 50 mg/l BA with 2% calcium ion (Ca2+) increased the proportion of perfect fl owers (Singh and Rajput, 1990). Naphtha-lene acetic acid (NAA) at concentrations of 50, 100 and 200 mg/l increased the perfect/staminate fl ower ratio (Mallik et al., 1959; Singh et al., 1965).
Other factors infl uencing sex ratios of infl orescences include stem age and mineral nutrients. Gunjate et al. (1983), Desai et al. (1986) and Hussein et al. (1989) reported that infl orescences from stems that grew at different times during the previous summer/autumn period had signifi cantly differ-ent perfect/staminate fl ower ratios. Singh and Dhillon (1987) found that boron (B) levels affect sex ratio.
Sex ratios can be manipulated with growth regulators, but has no com-mercial advantage (A.W. Whiley, personal communication, Queensland, 1996). Increases in fruit yield resulting from chemically increased perfect/staminate fl ower ratios have not been observed, suggesting that perfect fl ower numbers are not the primary limitation to crop performance (Schaffer

T.L. Davenport136
et al., 1994). If only one or two fruits were set on each terminal, the tree would carry an unusually heavy crop. It is unlikely that reduced perfect fl ower numbers due to cool temperatures during infl orescence development is directly responsible for poor fruit set and yields. Pollen viability, growth and ovule fertilization are probably the main factors contributing to low fruit set under these conditions.
Anthesis and dehiscence
Floral anthesis generally occurs at night in polyembryonic cultivars (Wagle, 1929; Torres, 1931; Galang and Lazo, 1937; Pimentel et al., 1984) and at night or early morning in monoembryonic types (Popenoe, 1917; Musahib-ud-din and Dinsa, 1946; Singh, 1954a; Randhawa and Damodaran, 1961a, b). Subsequent dehiscence of the four-lobed anthers occurs during the daylight morning hours revealing pale blue pollen grains (Torres, 1931; Galang and Lazo, 1937; Mallik, 1957; L.B. Singh, 1960). Anthesis and anther dehiscence are delayed by low temperatures or overcast days (Singh, 1954a; De Wet and Robbertse, 1986a). Dehiscence is also delayed by high RH, and pollination occurs primarily around midday (Mallik, 1957; Randhawa and Damodaran, 1961a, b).
Stigmas are receptive from c.18 h prior to anthesis to at least 72 h after anthesis with optimum receptivity within 3 h from anthesis (Popenoe, 1917; Wagle, 1929; Sen et al., 1946; Singh, 1954a; Spencer and Kennard, 1956; Rand-hawa and Damodaran, 1961a, b; Gunjate et al., 1983; Pimentel et al., 1984; Robbertse et al., 1994). Receptive stigmas are shiny and white-green, whereas non-receptive stigmas are desiccated and yellow-brown. Pollen germination generally occurs within 90 min of deposition, although the percentage ger-mination of pollen deposited on stigmas is relatively poor (Singh, 1954a; S.N. Singh, 1961). Pollination is initiated by the formation of two unusual protu-berances that meet to form a bridge or ponticulus connecting the dorsal side of the ovule with the ovary wall in line with the base of the style (Joel and Eisenstein, 1980). The ponticulus may guide the elongating pollen tube to the ovule. Ovule fertilization occurs 48–72 h after pollination (S.N. Singh, 1961; Ram et al., 1976). Both zygote cell and endosperm nuclear division appear to rest for about 2 weeks following pollination despite cell division and growth of the ovary (Sharma and Singh, 1972; Ram et al., 1976). A description of embryo development is presented by U.R. Singh (1961). Application of B may improve stigma receptivity, pollen tube germination and growth and ovule fertilization (De Wet and Robbertse, 1986b; Robbertse et al., 1988; De Wet et al., 1989) as well as fruit development (Chen, 1979; Robbertse et al., 1990); however, Jutamanee et al. (2002) could not verify the effect of B.
Pollen
Pollen grains are 20–45 m long and are oblong when dry and more spherical when hydrated (Popenoe, 1917; Jivanna Rao, 1923; Bijhouwer, 1937;

Reproductive Physiology 137
Mukherjee, 1950; Singh, 1954a; Randhawa and Damodaran, 1961b; S.N. Singh, 1961). There are generally three equilateral, tapering furrows along the longitudinal sides of dry pollen that give hydrated grains a roughly tri-angular shape when viewed on end (Popenoe, 1917; Singh, 1954a; S.N. Singh, 1961; U.R. Singh and A.P. Singh, 1973). Each furrow has a germpore in its centre (Mukherjee, 1950; S.N. Singh, 1961). Anthers produce c.250–650 pollen grains with a mean of 410 grains per anther (Popenoe, 1917, 1920; Spencer and Kennard, 1955).
In vitro germination of mango pollen has been reported (Popenoe, 1917; Spencer and Kennard, 1955; Young, 1958; Randhawa and Damodaran, 1961b; S.N. Singh, 1961). Germination on stigmas was c.10% less than that on artifi cial media, that is 78% across cultivars (Spencer and Kennard, 1955), although lower rates of germination have been reported (Mukherjee, 1950). Mango pol-len is most viable soon after anther dehiscence and rapidly degrades (Sen et al., 1946; Spencer and Kennard, 1955; Mallik, 1957; S.N. Singh, 1963). Although the initial percentage of viable pollen is generally ≥ 90% during warm weather (Popenoe, 1917; Mukherjee, 1949a, b; Singh, 1954a; S.N. Singh, 1961), cool tem-peratures early in the fl owering season result in abnormal, non-viable pollen grains (Popenoe, 1917; U.R. Singh and A.P. Singh, 1973; Shu et al., 1989; Gazit et al., 1992; Issarakraisila et al., 1992). The pre-vacuolate stage of meiosis during microsporogenisis is the most sensitive period to temperatures < 10°C (Issara-kraisila and Considine, 1994). Germination and pollen tube growth are reduced by cool temperatures (S.N. Singh, 1961; Mullins, 1986; Robbertse et al., 1988; Whiley et al., 1988; De Wet et al., 1989) and completely inhibited at tempera-tures < 15°C (Popenoe, 1917; Young, 1955; Sukhvibul et al., 2000).
Pollination
Pollination is a major yield-limiting constraint, due to the large number of fl owers on trees and low fruit set. Unlike polyembryonic cultivars, which produce nucellar embryos, pollination is necessary for fruit set with mo-noembryonic cultivars (Popenoe, 1917; Young, 1942; Spencer and Kennard, 1955; Gunjate et al., 1983; Pimentel et al., 1984; Robbertse et al., 1994). Pollen compatibility within and between cultivars has been widely investigated. Complete or partial self-incompatibility has been reported (Mukherjee et al., 1961; Singh et al., 1962b; Sharma and Singh, 1970, 1972; Ram et al., 1976; De Wet et al., 1989; Robbertse et al., 1993). Incompatibility is evident by degen-eration of embryonic and nucellar tissues and excessive loss of fruitlets. Cross incompatibility between some cultivars has also been noted (Saha and Chhonkar, 1972; Ram et al., 1976; Robbertse et al., 1993).
WindEarly investigators concluded that the species is wind pollinated (Hartless, 1914). Initially wet pollen dries to a powdery consistency on anthers soon after anthesis in dry conditions (Pimentel et al., 1984), whence it is likely to be liberated in moving air or via gravity to adjacent stigmas on the same and

T.L. Davenport138
nearby fl owers (Naik and Mohan Rao, 1943; Mallik, 1957). Singh (1954a) and S.N. Singh (1961) suggested, however, that the amount of pollen moving in air streams was too low for wind to be a pollination vector. They did not report the location of pollen-collecting slides or take into account the close proximity of fl owers within infl orescences or numbers of open fl owers in the canopy. Panicles bagged to exclude pollinating insects were reported to set fruit (Free and Williams, 1976), which were retained to maturity, thereby con-fi rming that mango pollen can be transferred by air movement or gravity (Bijhouwer, 1937; Mallik 1957). The tacit assumption that open-pollinated fl owers are exclusively crossed is likely to be incorrect, although mango may favour cross-pollination.
InsectPopenoe (1917) reasoned that pollen transfer occurs primarily within fl owers by insects. Panicles bagged to exclude insect visitation generally result in less fruit set than on panicles in the open (Popenoe, 1917; Musahib-ud-din and Dinsa, 1946; Mallik, 1957; Free and Williams, 1976; Jiron and Hedstrom, 1985). Insects working mango fl owers include Diptera, Hymenoptera, Lepidoptera and Coleoptera (Popenoe, 1917; Simao and Maranhao, 1959; Randhawa and Damodaran, 1961b; McGregor, 1974; Anderson et al., 1982; Jiron and Hed-strom, 1985). Flies of various genera are common on mango fl owers (Pope-noe, 1917; Burns and Prayag, 1921; Bijhouwer, 1937; Singh, 1954a; Spencer and Kennard, 1955; Eardley and Mansell, 1993). Polistes wasps are observed on mango fl owers but are considered to be ineffectual for pollen transfer (Spencer and Kennard, 1955; Free and Williams, 1976; Wolfenbarger, 1977). Honeybees (Hymenoptera) are occasional visitors (Young, 1942; Simao and Maranhao, 1959; Smith, 1960; Morton, 1964; Jiron and Hedstrom, 1985; Mac-Millan, 1991; Du Toit and Swart, 1993, 1994; Eardley and Mansell, 1993, 1994), but only if other more inviting fl owers are not present (Spencer and Kennard, 1955; Free and Williams, 1976; McGregor, 1976). They are assumed to be the most effective pollinators of mango and may be more effective if hives are placed in orchards during fl owering (Du Toit and Swart, 1993, 1994). Ander-son et al. (1982) recorded actual pollen transfer on mango fl owers by insects and found, in order of importance, the most effi cient pollinators to be wasps, bees, large ants and large fl ies.
With few exceptions (Mallik, 1957), pollen deposition rates are generally low (Naik and Mohan Rao, 1943; Mukherjee, 1951). Differences in pollination rates can be attributed to environmental conditions during fl owering, differ-ing attraction of insects to specifi c cultivars, proximity of more attractive fl owering species or a combination of the above. Young (1942) observed that insects visit only 10–12% of available fl owers. Depending on weather condi-tions, insect activity on mango fl owers is usually continuous from early morning to late afternoon, but nocturnal activity of some species has also been reported (Jiron and Hedstrom, 1985).
The role of insects in cross-pollination is not understood. Anderson et al. (1982) observed various insects carrying pollen to and from fl owers and noted pollination subsequent to those visits; however, they made no distinction

Reproductive Physiology 139
between actual pollen depositions by visiting insects and pollen transferred by other means. Wester (1920) considered that pollination is facilitated by wind and to a lesser extent by insects, and this conclusion is probably correct in most environments. Self-pollination within fl owers by insects while the pollen is still damp is likely to occur. Use of isozyme (Degani et al., 1990; Rob-bertse et al., 1993) and microsatellite DNA markers (Adato et al., 1995; Schnell et al., 2005) to discern ratios of cross- versus self-pollinated fruitlets and off-spring is the most accurate procedure to confi rm self- and cross-pollination. Initial fruit set of pollinated fl owers is inconsequential since most of these fruitlets abscise before reaching maturity (Lynch and Mustard, 1950; Singh, 1954a; Randhawa and Demodaran, 1961a).
5.12 Fruit Development
Fruit growth is correlated with several growth regulating substances. Enlarge-ment is sigmoidal reaching a constant size c.2–3 weeks before maturity (Singh, 1954a; Randhawa and Damodaran, 1961a, b; Ram, 1983; Prakash and Ram, 1984). The highest rates of fruit growth have been correlated with peak levels of putative endogenous auxins found in seeds (Chacko et al., 1970a, b; Singh and Singh, 1974; Chen, 1981; Ram, 1983; Prakash and Ram, 1984). Baghel et al. (1987a, b) reported increased fruit mass with a combination spray of NAA and urea to pre-anthesis panicles. Free and bound gibberellins, especially in seeds (Ogawa, 1963; Ram, 1983), peak similarly to putative aux-ins during fruit development (Chacko et al., 1970c, 1972a; Ram and Pal, 1979; Chen, 1981). Cytokinins tentatively identifi ed as zeatin and zeatin riboside and other active fractions appear to vary in concentration in seeds and peri-carp with two distinct peaks of activity. No particular relationship to growth was evident (Ram, 1983; Ram et al., 1983). Seeds produced more cytokinin activity than did the pericarp. In contrast, Chen (1983) found only one peak of cytokinin activity in seeds and pulp occurring at about half full size of both seed and pericarp. Seed tissues contain the highest cytokinin activity. Levels of endogenous auxin, gibberellins and cytokinins in leaves during fruit set were compared to production in leaves during other periods with-out conclusive results (Paulas and Shanmugavelu, 1989).
5.13 Stenospermocarpy
Abscission of non-fertilized and fertilized fl owers is normal. Fruitlet abscis-sion from pea size on is often associated with embryo abortion (Chandler, 1958; U.R. Singh, 1961; Singh, 1964; Lakshminarayana and Aguilar, 1975; Ram et al., 1976) and is referred to as stenospermocarpy (Soule, 1985). Steno-spermocarpy in mango is unusual (Chacko and Singh, 1969a) but occurs regularly in some cultivars (Núñez-Elisea and Davenport, 1983; Whiley et al., 1988). Stenospermocarpic fruitlets have slower growth rates than seeded fruit, generally become misshapen and fail to reach full size.

T.L. Davenport140
Stenospermocarpy in some cultivars has been correlated with low tem-peratures during fl owering and early fruit set in the subtropics (Lakshmina-rayana and Aguilar, 1975; Young and Sauls, 1979; Whiley et al., 1988; Schaffer et al., 1994). Núñez-Elisea and Davenport (1983) reported that stenospermo-carpic fruit often occur distal to seeded fruitlets within panicles and sug-gested that embryo abortion is associated with high temperatures when these latter fruit set. Secondary spring fl owering of some monoembryonic culti-vars under high temperatures has resulted in high proportions of steno-spermocarpic fruit (E.K. Chacko, personal communication, Australia, 1995). Application of auxins, gibberellins and cytokinins produce seedless fruit in some cultivars, suggesting that the abscission zone is protected by these hor-mones despite the loss of the endogenous supply from the aborted seed (Ven-kataratnam, 1949; Chacko and Singh, 1969a, b; Kulkarni and Rameshwar, 1978).
5.14 Fruit Set and Retention
Fruit set and retention of mango was recently reviewed by Singh et al. (2005). Abscission of fl owers and fruitlets is accomplished by rapid formation of a separation layer in the abscission zone in the pedicel-peduncle junction (Bar-nell, 1939). U.R. Singh (1961) described formation of the abscission zone dur-ing fl oral ontogeny and of the separation layer during abscission of fl owers and fruitlets. The majority of panicles lose all fruitlets (Núñez-Elisea and Davenport, 1983). The pattern of fruitlet abscission is asymptotic with the greatest losses occurring during the fi rst weeks after anthesis (Núñez-Elisea and Davenport, 1983; Prakash and Ram, 1984; Searle et al., 1995). Except for the tendency to retain fruit in the distal portion of panicles, abscission of fl owers and fruitlets is random. It can involve fruitlets regardless of size or location.
Of the 8–13% of perfect fl owers setting fruit, < 1% reach maturity (Bijhou-wer, 1937; Sen, 1939; Naik and Mohan Rao, 1943; Mukherjee, 1949b; U.R. Singh, 1960; Randhawa and Damodaran, 1961a; Singh, 1978; Gunjate et al., 1983; Prakash and Ram, 1984). Generally, most fruit are set on the most distal spike portion of panicles (Chadha and Singh, 1963; Núñez-Elisea and Daven-port, 1983). Fruit loss has been associated with embryo abortion, resulting in blackened or shrivelled embryos (Singh, 1954a, 1964; Chandler, 1958; U.R. Singh, 1961; Sharma and Singh, 1972; Ram et al., 1976) after the fruit is sepa-rated from the tree (Núñez-Elisea and Davenport, 1983).
Sex ratio
The perfect/staminate fl oral ratio in panicles may infl uence fruit set and pro-ductivity (Naik and Mohan Rao, 1943; Singh, 1954b; Singh and Singh, 1959; U.R. Singh, 1960). Mallik (1957) noted that more perfect fl owers are formed in ‘on’ than ‘off’ years of alternate-bearing cultivars. Other studies, however,

Reproductive Physiology 141
have demonstrated that the number of perfect fl owers does not correlate with subsequent yield (Randhawa and Damodaran, 1961a) so long as the proportion of perfect fl owers is not < 4% (Singh, 1964, 1971). Most fruit are borne in the distal portion of panicles (Shawky et al., 1977), which may be correlated with the high ratio of perfect to staminate fl owers there. Schole-fi eld and Oag (1984) estimated that one mature fruit is harvested for each 169 perfect fl owers in the distal half of the panicle; whereas 592 perfect fl owers are required to produce one fruit in the proximal half. Therefore, intrinsic factors other than sex ratio regulate fruit set.
Mineral nutrients
Boron is one of seven micronutrients required for normal plant growth. The physiological function of B is unknown (Hu and Brown, 1994), although it is essential for fl oral development, pollen germination, pollen tube growth, embryo development and growth of organs (i.e. fruit) (Vasil, 1963; Agarwala et al., 1981; Dell and Huang, 1997; Shorrocks, 1997). Defi cient soils are com-monly found in mango-producing areas of Australia, Thailand, Central and South America and Africa where symptoms are common (Aitken et al., 1987; Singh et al., 2005). Boron applications to defi cient mango trees increase normal fruit set (Robbertse et al., 1990; Raja et al., 2005). Fruitlet abscission in mangoes has also been attributed to zinc (Zn) defi ciency (Jiron and Hedstrom, 1985).
Hormonal control
AuxinResearch demonstrating improved fruit set and retention following applica-tion of several auxin analogues to pre-anthesis panicles or to panicles bearing fruitlets of various sizes has been reviewed (Davenport and Núñez-Elisea, 1997; Singh et al., 2005). NAA is the most effective auxin analogue for improv-ing fruit retention (Prakash and Ram, 1986; Khan et al., 1993). Initial fruit set was substantially increased when sprays of 200 mg/l indole acetic acid (IAA) were applied to developing panicles (Singh et al., 1965). A 300–400% increase in fruit set resulted when NAA (40 or 50 mg/l) was sprayed at the pre-anthesis stage (Ram, 1983; Singh and Ram, 1983; Prakash and Ram, 1986). Chen (1981) reported no effect on fruit retention when 5 mg/l of either naphthaleneacet-amide or β-naphthoxoyacetic acid were applied three times at 2-week inter-vals to panicles in which fruit had reached 4 mm in diameter.
Despite increased fruit retention of mango using exogenous applications of auxins, few studies have examined endogenous auxins in fruit as related to retention (Chacko et al., 1970a, b; Ram et al., 1983; Prakash and Ram, 1984). Singh and Singh (1974) were unable to detect signifi cant differences in endog-enous auxins or inhibitors when comparing alternate and regular bearing cultivars. Chen (1981) observed lower levels of auxin-like substances in

T.L. Davenport142
mesocarp and calyx tissues of abscised fruits than those of intact fruits. Simi-lar decreases in auxin and gibberellins with an increase in abscisic acid as fruitlets abscised were reported by Bains et al. (1999). The interaction of auxin in fruit and abscission zones to maintain mango fruit retention is not clear.
Continuous auxin synthesis and basipetal transport to the abscission zone is critical for maintenance of plant organs, including fruit (Crane, 1964; Nitsch, 1965; Morgan et al., 1977; Davenport et al., 1980; Roberts and Osborne, 1981). Increased mango fruit set and retention in response to exogenously applied auxins confi rms this requirement; however, other hormonal factors also appear to be involved. Developing seeds are rich sources of all the known classes of phytohormones, including auxins (Crane, 1964; Nitsch, 1965; Chacko et al., 1970a, b, c; Chen, 1981). Hence, exogenous enrichment of auxin in the presence of other seed-produced phytohormones facilitates increased fruit retention. In contrast, NAA (10 and 20 mg/l) spray-applied to bagged, self-pollinated fl owers, does not result in development of stenospermocarpic fruits beyond the marble size (Venkataratnam, 1949; Chacko and Singh, 1969a, b). Similarly, applications of 250 or 500 mg/l GA3 or 250 mg/l BA alone to panicles does not promote production of stenospermocarpic fruits (Chacko and Singh, 1969a, b). Supplying exogenous β-naphthoxyacetic acid (10 mg/l), BA (250 mg/l) and GA3 (250 and 500 mg/l) together in multiple sprays until half grown, however, resulted in retention of several seedless fruit to maturity. Chen (1983) and Oosthuyse (1995b) observed that gibberel-lin, cytokinin and auxin reduce fruit drop of open-pollinated fruitlets of some cultivars. Thus, although auxin is important for maintaining the abscission zone, the presence of other phytohormones appears to be important for fruit-let development (Chacko et al., 1970a, b; Ram, 1983; Ram et al., 1983).
CytokininsAlthough cytokinins are not generally thought to be associated directly with abscission, Ram (1983) and Ram et al. (1983) concluded that low cytokinin levels during fruit development might contribute to fruit loss. Chen (1983) observed a correlation of low cytokinin levels in stenospermocarpic fruits with abscission at the marble stage of growth. Application of 250 mg/l BA to bagged panicles does not promote production of seedless fruits (Chacko and Singh, 1969a, b). The synthetic cytokinin, N-(2-chloro-4-pyridyl)-N’-phenylurea (CPPU) also does not improve fruit set when applied alone at a rate of 10 mg/l to post-anthesis panicles (Oosthuyse, 1995b). The role of cytokinins in separation events remains inconclusive.
GibberellinsGibberellins do not appear to be directly linked to the onset of abscission (Chacko et al., 1970c, 1972a; Ram and Pal, 1979; Chen, 1981; Ram, 1983). Spray applications of GA3 to pre- and post-anthesis panicles to increase fruit set and retention have been inconsistent. Increased yield (Teaotia et al., 1967; Singh and Ram, 1983; Rajput and Singh, 1989) and production of seedless fruit (Kulkarni and Rameshwar, 1978) have been reported from these treatments, but Chacko and Singh (1969a, b) observed no such effects. Chen (1983) and

Reproductive Physiology 143
Oosthuyse (1995b) investigated the effects of several foliar applications of GA3 starting at the 4 mm diameter stage, but were unable to improve fruit set.
Several classes of gibberellin-synthesis inhibitors have been tested for reducing fruit drop. The growth retardants, daminozide and cycocel, increased fruit set when applied to post-anthesis panicles (Singh and Ram, 1983). The authors suggested that increased fruit retention might have been mediated through increased cytokinin-like activity of the growth retardants. Although initial fruit set was promoted by PBZ, yield was not improved (Kurian and Iyer, 1993a). It is not clear whether the contrasting results of increased yield (Kurian and Iyer, 1993b) were due to reduced fruit loss or more intense fl ow-ering in response to treatment. Goguey (1990) reported increased fruit set and retention using soil-applied PBZ at 5 g ai/tree. Spray application of uni-conazole, a more biologically active triazole (500–2000 mg/l), reportedly increased fruit set and yield (Galila and El-Masry, 1991). It is diffi cult to resolve the contradictory results demonstrating enhancement of fruit reten-tion by GA3 and inhibitors of its synthesis.
InhibitorsAbscisic acid (ABA) is possibly involved in fruitlet abscission. Although cor-relations exist between certain inhibitors and abscission of mango fruitlets, no clear cause and effect relationships have been established. Fruit drop was correlated with levels of an acidic inhibitor, possibly ABA (Chacko et al., 1970b, 1972a; Singh and Singh, 1974; Ram, 1983; Prakash and Ram, 1984). Chen (1981) reported similar changes in putative ABA with maximum levels occurring during early fruit drop and with advancing age of fruits. Putative ABA levels in abscised and retained fruits were compared and were highest in the calyx and mesocarp of abscised fruitlets.
EthyleneEthylene has the greatest immediate impact on fl ower and fruitlet abscission. Van Lelyveld and Nel (1982) reported higher levels of ethylene in abscised fruitlets compared with those retained on trees. Núñez-Elisea and Davenport (1983, 1984, 1986) examined the dynamics of ethylene production in intact and excised fruitlets from onset to separation. Increased production began in explants about 26 h postharvest and increased logarithmically until fruit sep-aration. Abscission of the fruitlets began 48 h after the onset of enhanced ethylene production. Similar results with avocado fruitlet abscission experi-ments (Davenport and Manners, 1982) indicate that the onset of ethylene production in intact fruitlets is spontaneous in individual fruitlets followed by abscission 48 h later. The pericarp provided the bulk of ethylene for induc-tion of abscission processes; the pedicel produced no ethylene. There was reduced fruit drop in response to inhibitors of ethylene production and action (Singh and Ram, 1983; Naqvi et al., 1990, 1992). Whereas increased peroxi-dase (Van Lelyveld, 1978) and polyphenol oxidase activities have been reported in abscissed mango fruitlets (Van Lelyveld and Nel, 1982), Núñez-Elisea and Davenport (1984) observed no changes in peroxidase activity or protein levels prior to separation of fruitlets.

T.L. Davenport144
Abscission of stenospermocarpic fruits has been associated with small increments in ethylene production (Núñez-Elisea and Davenport, 1983). Sen-sitivity to low levels of endogenous ethylene may refl ect the absence of seed-produced auxins. Protection of the abscission zone depends on a constant supply of auxin, and ethylene production levels in tissues correlate with endogenous auxin levels (Roberts and Osborne, 1981).
Despite their roles in cell division, cell enlargement and maintenance of the abscission zone in developing fruit, a specifi c recommendation for exog-enous application of plant growth regulators, either alone or in combination, to improve yield of mango has not been adopted. Phytohormones have little residual effect on fruit development, and multiple applications of products to counteract the short-term responses are prohibitively expensive.
PhotoassimilatesWolstenholme and Whiley (1995) discussed the ecophysiology of the mango as a basis for preharvest management. They proposed that the adaptive sur-vival strategies of the mango explain its notoriously poor cropping perfor-mance. Mechanisms that impart tolerance to heat, drought and fl ood stresses, which the tree has developed for survival in harsh environments, have come at considerable carbon cost with the resultant diversion of photoassimilate resources away from fruiting.
There is abundant evidence that heavy cropping in tree crops exhausts stored reserves (Jones et al., 1975; Kaiser and Wolstenholme, 1994; Whiley et al., 1996) and that current photosynthate is often unable to satisfy the demands of fruit set and fruit growth after heavy and prolonged fl owering (Chacko et al., 1982). There are signifi cant genotypic differences in photoassimilation rates between low- and high-yielding cultivars growing in both the tropics and the subtropics of Australia (Chacko et al., 1995; Searle et al., 1995). At each location, photoassimilation rates were considerably greater on the higher-yielding cultivar, and this difference was maintained from fl owering through to fruit maturation. 14C studies during the fruit set and abscission period also demonstrated strong discrimination in the movement of assimilates, which was dominated by randomly located fruit on panicles of the low-yielding cultivar (Chacko et al., 1995). In contrast, assimilate discrimination to fruitlets was less severe in the high-yielding cultivar with a more even distribution of photoassimilates. It was concluded that the availability and distribution of photoassimilates during the fruit set and establishment stages was largely responsible for the yield differences between the cultivars.
Supporting evidence for the role of photoassimilates in fruit set and retention also comes from enrichment studies (Schaffer et al., 1999). Container-grown plants that fl owered in the open were transferred to controlled-environment glasshouse rooms immediately after the completion of anthesis. Temperatures were 28°C day/20°C night while the atmospheric carbon diox-ide (CO2) concentrations were 350 or 600 mol/mol. Photoassimilation of trees in the CO2-enriched rooms was approximately 60% greater than those held at partial pressures of 350 mol/mol CO2. Fruit retention and fi nal yield were signifi cantly higher on those trees grown at the partial pressure of

Reproductive Physiology 145
600 μmol/mol CO2. Higher levels of available assimilates during the fruiting cycle appear to benefi t fruit retention and yield.
5.15 Alternate Bearing
Alternate bearing of certain mango cultivars has plagued growers (Hartless, 1914; Rao, 1997). Lack of production in the ‘off’ year is usually a result of lack of fl oral initiation (R.N. Singh, 1959), and has been reviewed (Mukherjee, 1953; Gangolly et al., 1957; L.B. Singh, 1960, 1972; Chadha and Pal, 1986; Pan-dey, 1989). Shoot initiation and induction described in this chapter perhaps offers a clearer understanding of this phenomenon.
5.16 Conclusions
This chapter has provided a comprehensive review of investigations of vari-ous factors potentially involved in mango fl owering, fruit set and retention. Many of the reports cited are contradictory. Such variable results refl ect: (i) the different experimental approaches utilized, especially in fi eld experi-ments; (ii) the range of environments in which experiments have been con-ducted; and (iii) differences in the responses of cultivars to treatments. As a consequence, it is diffi cult to draw unambiguous conclusions with respect to the role of specifi c factors on fl owering, fruit set and retention. More research is clearly needed in these areas, particularly in controlled environments. For example, although KNO3 is utilized with great success to stimulate fl owering in tropical conditions, confusion remains as to whether it effects initiation or fl oral induction. Future studies on fl owering research should include results of the proportion of stems that remain in rest and those that produce vegeta-tive shoots as well as the proportion of reproductive shoots. Providing this information allows an analysis of the impact of treatments on initiation and inductive events.
This chapter also presents several hypothetical models for shoot devel-opment and fl owering. Each of the proposed models consists of a series of hypotheses that invite further study to test their validity. Future research should challenge these models so that fl owering and crop yield can be better understood in both the tropics and the subtropics.
References
Abe, M., Kobayashi, Y., Yamamoto, S., Daimon, Y., Yamaguchi, A., Ikeda, Y., Ichinoki, H., Notaguchi, M., Goto, K. and Araki, T. (2005) FD, a bZIP protein mediating signals from the fl oral pathway integrator FT at the shoot apex. Science 309, 1052–1056.
Abeles, F.B. (1973) Ethylene in Plant Biology. Academic Press, New York, 302 pp.Acala, P.E. and San Pedro, A. (1935) Bud differentiation in smudged mango trees. Philip-
pine Agriculture 24, 27–48.

T.L. Davenport146
Adato, A., Sharon, D., Lavi, U., Hillel, J. and Gazit, S. (1995) Application of DNA fi nger-prints for identifi cation and genetic analyses of mango (Mangifera indica) geno-types. Journal of the American Society for Horticultural Science 120, 259–264.
Agarwala, S.C., Sharma, P.N., Chatterjee, C. and Sharma, P.C. (1981) Development and enzymatic changes during pollen development of boron defi cient maize. Journal of Plant Nutrition 3, 329–336.
Agrawal, A., Ram, S. and Garg, G.K. (1980) Endogenous cytokinins of mango (Mangiferaindica L.) shoot tips and their signifi cance in fl owering. Indian Journal of Experi-mental Biology 18, 504–509.
Aitken, R.L., Jeffery, A.J. and Compton, B.L. (1987) Evaluation of selected extractants for boron in some Queensland soils. Australian Journal of Soil Research 25, 263–273.
Aksenova, N.P., Milyaeva, E.L. and Romanov, G.A. (2006) Florigen goes molecular: seventy years of the hormonal theory of fl owering regulation. Russian Journal of Plant Physiology 53, 401–406.
Alcala, P.E. and San Pedro, A. (1935) Bud differentiation in smudged mango trees. Phil-ippine Agriculturist 24, 27–40.
Allsopp, A. (1965) The signifi cance for development of water supply, osmotic relations and nutrition. In: Lang, A. (ed.) Encyclopedia of Plant Physiology. XVII. Springer-Verlag, Berlin, pp. 504–552.
Anderson, D.L., Sedgley, M., Short, J.R.T. and Allwood, A.J. (1982) Insect pollination of mango in northern Australia. Australian Journal of Agriculture Research 33, 541–548.
Astudillo, E.O. and Bondad, N.D. (1978) Potassium nitrate-induced fl owering of ‘Ca-rabao’ mango shoots at different stages of maturity. Philippine Journal of Crop Science 3, 147–152.
Baghel, B.S., Sharma, R.K. and Nair, P.K.R. (1987a) Effi cacy of prefl owering spray of urea and NAA on physical standards of mango (Mangifera indica L.). Progressive Horti-culture 19, 231–234.
Baghel, B.S., Sharma, R.K. and Nair, P.K.R. (1987b) Infl uence of prefl owering spray of urea and NAA on fruit retention of mango (Mangifera indica L.). Progressive Horti-culture 19, 200–202.
Bains, K.S., Bajwa, G.S. and Singh, Z. (1999) Effects of indole-3-acetic acid, gibberellic acid, and abscisic acid on abscission of mango fruitlets. Tropical Agriculture 76, 88–92.
Bajwa, B.S., Bakshi, J.C. and Kocher, T.S. (1956) A note on the fl oral biology of Mangiferaindica L. var. Dashaheri. Indian Journal of Horticulture 13, 206–209.
Bakr, E.I., Abdalla, K.M., Meligi, M.A. and Ismail, I.A. (1981) Floral differentiation in mango as affected by growth regulators, ringing and defoliation. Egyptian Journal of Horticulture 8, 161–166.
Bangerth, F. (1994) Response of cytokinin concentration in the xylem exudate of bean (Phaseolus vulgaris L.) plants to decapitation and auxin treatment, and relationship to apical dominance. Planta 194, 439–442.
Bangerth, F. (2006) Flower induction in perennial fruit trees: still an enigma? Acta Hor-ticulturae 727, 177–195.
Bangerth, F., Naphrom, D., Sruamsiri, P., Hegele, M., Boonplod, N. and Manochai, P. (2004) Hormonal changes in various tissues of mango trees during fl ower induction following cold temperature. Acta Horticulturae 645, 453–457.
Barba, R.C. (1974) Induction of fl owering of the mango by chemical spray. Proceedings of the Crop Science Society of the Philippines 5, 154–160.
Barnell, E. (1939) Studies in tropical fruits. V. Some anatomical aspects of fruit fall in two tropical arboreal plants. Annals of Botany 3, 77–89.

Reproductive Physiology 147
Batten, D.J. and McConchie, C.A. (1995) Floral induction in growing buds of lychee (Litchi chinensis) and mango (Mangifera indica). Australian Journal of Plant Physi-ology 22, 783–791.
Bausher, M.G. and Yelenosky, G. (1987) Morphological changes in citrus associated with relatively high concentrations of paclobutrazol. Journal of Plant Growth Regu-lation 5, 139–147.
Belding, R.D. and Young, E. (1989) Shoot and root temperature effects on axillary cyto-kinin levels during budbreak in young apple trees. HortScience 24, 115–117.
Bernier, G. (1988) The control of fl oral evocation and morphogenesis. Annual Review of Plant Physiology 39, 175–219.
Bernier, G. and Sachs, R.M. (1979) Photosynthesis and fl owering. In: Marcelle, R., Clysters, H. and van Pouche, M. (eds) Photosynthesis and Plant Development. Dr. W. Junk Publishers, The Hague, pp. 137–150.
Bernier, G., Kinet, J.M. and Sachs, R.M. (1981) The Physiology of Flowering. Vols 1 and 2. CRC Press, Boca Raton, Florida.
Bernier, G., Havelange, A., Houssa, C., Petitjean, A. and Lejeune, P. (1993) Physiologi-cal signals that induce fl owering. The Plant Cell 5, 1147–1155.
Beveridge, C.A., Weller, J.L., Singer, S.R. and Hofer, J.M.I. (2003) Axillary meristem de-velopment. Budding relationships between networks controlling fl owering, branch-ing, and photoperiod responsiveness. Plant Physiology 131, 927–934.
Bevington, K.B. and Castle, W.S. (1986) Annual root growth pattern of young citrus trees in relation to shoot growth, soil temperature, and soil water content. Journal of the American Society for Horticultural Science 110, 840–845.
Beyer, E.M., Jr and Morgan, P.W. (1971) Abscission: the role of ethylene modifi cation of auxin transport. Plant Physiology 48, 208–212.
Bijhouwer, A.P.C. (1937) Een Bijdrage tot de Kennis Omtrent het Bloeien en het Vruchtdragende, Vermogen van den Mangga (Mangifera indica L.). H. Veenman and Zonen, Wageningen, the Netherlands, 106 pp.
Blaikie, S.J., Leonardi, J., Chacko, E.K., Muller, W.J. and Scott, N.S. (1999) Effect of cinc-turing and chemical treatments on growth, fl owering and yield of mango. Austra-lian Journal of Experimental Agriculture 39, 761–770.
Bohlenius, H., Huang, T., Charbonnel-Campaa, L., Brunner, A.M., Jansson, S., Strauss, S.H. and Nilsson, O. (2006) CO/FT regulatory module controls timing of fl owering and seasonal growth cessation in trees. Science 312, 1040–1043.
Bondad, N.D. (1972) Effects of 2-chloroethane-phosphonic acid on some Philippine horticultural crops. Philippine Journal of Geography 16, 31–35.
Bondad, N.D. (1976) Response of some tropical and subtropical fruits to pre- and post-harvest applications of ethephon. Economic Botany 30, 67–80.
Bondad, N.D. and Apostol, C.J. (1979) Induction of fl owering and fruiting in immature mango shoots with KNO3. Current Science 48, 591–593.
Bondad, N.D. and Linsangan, E. (1979) Flowering in mango induced with potassium nitrate. HortScience 14, 527–528.
Bondad, N.D., Blanco, A.E. and Mercado, E.L. (1978) Foliar sprays of potassium nitrate for fl ower induction in mango (Mangifera indica cultivar Pahutan) shoots. Philip-pines Journal of Crop Science 3, 251–256.
Boss, P.K., Bastow, R.M., Milne, J.S. and Dean, C. (2004) Multiple pathways in the deci-sion to fl ower: enabling, promoting, and resetting. The Plant Cell 16, S18–S31.
Buell, E.P. (1954) Flowering and fruiting habits of the mango in wet zone. Tropical Agri-culturist 11, 280–284.
Bueno, P.B. and Valmayor, R.V. (1974) Potassium nitrate: key to mango fl owering. Agri-culture Los Banos 13, 4–16.

T.L. Davenport148
Burns, W. and Prayag, S.H. (1921) The Book of the Mango. Bombay Department of Agriculture Bulletin 103. Bombay Department of Agriculture, Bombay, 98 pp.
Burondkar, M.M. and Gunjate, R.T. (1991) Regulation of shoot growth and fl owering in ‘Alphonso’ mango with paclobutrazol. Acta Horticulture 291, 79–84.
Burondkar, M.M. and Gunjate, R.T. (1993) Control of vegetative growth and induction of regular and early cropping in ‘Alphonso’ mango with paclobutrazol. Acta Horti-culture 341, 206–215.
Burrows, G.E., Boag, T.S. and Stewert, W.P. (1992) Changes in leaf, stem, and root anat-omy of chrysanthemum cv. Lillian Hoek following paclobutrazol application. Jour-nal of Plant Growth Regulation 11, 189–194.
Cane, A.R. and Wilkins, M.B. (1970) Auxin transport in roots. Journal of Experimental Botany 21(66), 212–218.
Chacko, E.K. (1986) Physiology of vegetative and reproductive growth in mango (Mangifera indica L.) trees. In: Proceedings of the First Australian Mango Research Workshop. Commonwealth Scientifi c and Industrial Research Organization (CSIRO), Melbourne,pp. 54–70.
Chacko, E.K. (1991) Mango fl owering – still an enigma. Acta Horticulture 291, 12–21.Chacko, E.K. and Ananthanarayanan, T.V. (1982) Accumulation of reserve substances
in Mangifera indica L. during fl ower initiation. Zeitschrift für Pfl anzenphysiologie106, 281–205.
Chacko, E.K. and Randhawa, G.S. (1971) Towards an understanding of the factors affect-ing fl owering in mango. Andhra Agriculture Journal 18, 226–236.
Chacko, E.K. and Singh, R.N. (1969a) Growth regulators induced parthenocarpy in mango (Mangifera indica L.). Current Science 38, 249–250.
Chacko, E.K. and Singh, R.N. (1969b) Induction of parthenocarpy in mango (Mangiferaindica L.) using plant growth regulators. HortScience 4, 121–123.
Chacko, E.K., Singh, R.N. and Kachru, R.B. (1970a) Gibberellin-like substances in de-veloping fruits of the mango (Mangifera indica L.). Journal of Horticultural Science45, 371–378.
Chacko, E.K., Singh, R.N. and Kachru, R.B. (1970b) Physiology of fl owering and fruit growth in mango (Mangifera indica L.): partition, bioassay and characterization of naturally occurring auxins and inhibitors in immature fruits. Indian Journal of Experimental Biology 8, 135–138.
Chacko, E.K., Kachru, R.B. and Singh, R.N. (1970c) Changes in the level of acidic and neutral growth promoters during fruit development in Dashehari mango (Mangiferaindica L.). Journal of Horticulture Science 45, 341–349.
Chacko, E.K., Kohli, R.R. and Randhawa, G.S. (1972a) Studies on the effect of 2-chloroethyl phosphonic acid (Ethrel) on mango. I. Flower induction in an ‘off’ year in Langra trees. Indian Journal of Horticulture 29, 1–4.
Chacko, E.K., Singh, R.N. and Kachru, R.B. (1972b) Studies on the physiology of fl ower-ing and fruit growth in mango (Mangifera indica L.). VII. Naturally occurring auxins and inhibitors in the shoots of fl owering (on) and vegetative (off) mango trees. In-dian Journal of Horticulture 29, 115–125.
Chacko, E.K., Reddy, Y.T.N. and Ananthanarayanan, T.V. (1982) Studies on the relation-ship between leaf number and area and fruit development in mango (Mangiferaindica L.). Journal of Horticultural Science 57, 483–489.
Chacko, E.K., Lu, P. and Kohli, R.R. (1995) Photoassimilate production and distribution in relation to productivity in mango (Mangifera indica L.). In: Mango 2000 – MarketingSeminar and Production Workshop Proceedings. Department of Primary Industries, Brisbane, pp. 109–124.
Chadha, K.L. and Pal, R.N. (1986) Mangifera indica. In: Halevy, A.C. (ed.) CRC Hand-book of Flowering. Vol. 5. CRC Press, Boca Raton, Florida, pp. 211–230.

Reproductive Physiology 149
Chadha, K.L. and Singh, K.K. (1963) Studies on fruit drop in mango. I. Fruit set, its reten-tion and factors affecting it. Indian Journal of Horticulture 20, 172–185.
Chaikiattiyos, S., Menzel, C.M. and Rasmussen, T.S. (1994) Floral induction in tropical fruit trees: effects of temperature and water-supply. Journal of Horticultural Science69, 397–415.
Chailakhyan, M.K. (1936) New facts in support of the hormonal theory of plant develop-ment. C. R. Acad. Sci. URSS 13, 79–83.
Chandler, W.H. (1958) Evergreen Orchards, 2nd edn. Lea Febiger, Philadelphia, Penn-sylvania, 535 pp.
Chen, W.S. (1979) Physiological studies of fruiting in mango trees. 1. Effect of calcium and boron nutrition on fruiting. National Science Council Monthly (Republic of China) 7, 1220–1230.
Chen, W.S. (1981) Physiological studies of fruiting in mango trees. II. Effect of endoge-nous growth substances on fruiting. Proceedings National Science Council Part B, Life Sciences (Republic of China) 5, 49–55.
Chen, W.S. (1983) Cytokinins of the developing mango fruit. Plant Physiology 71, 356–361.Chen, W.S. (1985) Flower induction in mango (Mangifera indica L.) with plant growth
substances. Proceedings National Science Council Part B, Life Sciences (Republic of China) 9, 9–12.
Chen, W.S. (1987) Endogenous growth substances in relation to shoot growth and fl ow-er bud development of mango. Journal of the American Society for Horticulture Science 112, 360–363.
Cline, M., Wessel, T. and Iwamura, H. (1997) Cytokinin/auxin control of apical domi-nance in Ipomoea nil. Plant and Cell Physiology 38, 659–667.
Cobin, M. (1950) Mango selection, propagation and culture. University of Florida Agricultural Experiment Station Annual Report. University of Florida, Gainesville, Florida, pp. 257–259.
Coen, E.S. and Carpenter, R. (1993) The metamorphosis of fl owers. The Plant Cell 5, 1175–1181.
Coen, E.S. and Meyerowitz, E.M. (1991) The war of the whorls: genetics interaction controlling fl ower development. Nature 353, 31–37.
Coen, E.S., Romero, J.M., Doyle, S., Elliott, R., Murphy, G. and Carpenter, R. (1990) Floricaula: a homeotic gene required for fl ower development in Antirrhinum ma-jus. Cell 63, 1311–1322.
Collins, G.N. (1903) The Mangoes in Puerto Rico. United States Department of Agricul-ture (USDA) Bulletin No. 28. USDA, Washington, DC.
Corbesier, L. and Coupland, G. (2005) Photoperiodic fl owering of Arabidopsis: integrat-ing genetic and physiological approaches to characterization of the fl oral stimulus. Plant, Cell and Environment 28, 54–66.
Corbesier, L., Vincent, C., Jang, S., Fornara, F., Fan, Q., Searle, I., Giakountis, A., Farrona, S., Gissot, L., Turnbull, C.G.N. and Coupland, G. (2007) FT protein movement con-tributes to long-distance signaling in fl oral induction of Arabidopsis. Science 316, 1030–1033.
Crane, J.C. (1964) Growth substances in fruit setting and development. Annual Review of Plant Physiology 15, 303–326.
Cull, B.W. (1987) A whole plant approach to productivity research for mango. In: Prense-ley, R.T. and Tucker, G. (eds) Mangoes – A Review. Commonwealth Science Coun-cil, London, pp. 19–28.
Cull, B.W. (1991) Mango crop management. Acta Horticulturae 291, 154–173.Dalziel, J. and Lawrence, D.K. (1984) Biochemical and biological effects of kaurene
oxidase inhibitors, such as paclobutrazol. British Plant Growth Regulator Group Monograph 4, 1–14.

T.L. Davenport150
Davenport, T.L. (1987) Opportunities for tropical fruit crop growth regulation. In: Pro-ceedings of the 14th Annual Meeting of the Plant Growth Regulation Society of America, Honolulu, Hawaii, pp. 406–409.
Davenport, T.L. (1990) Citrus fl owering. Horticultural Reviews 12, 349–408.Davenport, T.L. (1992) Benefi cial effects of water stress. In: Davenport, T.L. and Har-
rington, H.M. (eds) Plant Stress in the Tropical Environment, Proceedings of a Workshop held in Kailua-Kona, Hawaii. USDA/Co-operative States Research Ser-vice/Caribbean Basin Administrative Group/Pacifi c Basin Administrative Group, Gainesville, Florida, pp. 16–20.
Davenport, T.L. (1993) Floral manipulation in mangos. In: Chia, L.E. and Evans, D.O. (eds) Proceedings of the Conference on Mango in Hawaii. Cooperative Extension Service, University of Hawaii, Honolulu, pp. 54–60.
Davenport, T.L. (1994) Potential problems with use of uniconazole on mango (Mangiferaindica). PGRSA Quarterly (published by Plant Growth Regulation Society of Amer-ica) 22, 143–153.
Davenport, T.L. (2000) Processes infl uencing fl oral initiation and bloom: the role of phytohormones in a conceptual fl owering model. HortTechnology 10, 733–739.
Davenport, T.L. (2003) Management of fl owering in three tropical and subtropical fruit tree species. HortScience 38, 1331–1335.
Davenport, T.L. (2006) Pruning strategies to maximize tropical mango production from the time of planting to restoration of old orchards. HortScience 41, 544–548.
Davenport, T.L. and Manners, M.M. (1982) Nucellar senescence and ethylene produc-tion as they relate to avocado fruitlet abscission. Journal of Experimental Botany 33, 815–826.
Davenport, T.L. and Núñez-Elisea, R. (1990) Ethylene and other endogenous factors pos-sibly involved in mango fl owering. Acta Horticulturae 275, 441–448.
Davenport, T.L. and Núñez-Elisea, R. (1991) Is endogenous ethylene involved in fl ower-ing of mango? Acta Horticulturae 291, 85–94.
Davenport, T.L. and Núñez-Elisea, R. (1997) Reproductive physiology. In: Litz, R.E. (ed.) The Mango: Botany, Production and Uses. CAB International, Wallingford, UK, pp. 69–146.
Davenport, T.L. and Smith, S.M. (1997) Evaluation of potential treatments to stimulate mango fl owering in sub-tropical latitudes. PGRSA Quarterly (published by Plant Growth Regulation Society of America) 25, 114–129.
Davenport, T.L. and Stern, R.A. (2005) Flowering. In: Menzel, C.M. and Waite, G.K. (eds) Litchi and Longan: Botany, Cultivation and Uses. CAB International, Walling-ford, UK, pp. 87–113.
Davenport, T.L., Morgan, P.W. and Jordan, W.R. (1980) Reduction of auxin transport capacity with age and internal water defi cits in cotton petioles. Plant Physiology65, 1023–1025.
Davenport, T.L., Ramos, L. and Alani, A. (1995) Evidence for a transmissible fl origenic promoter in mango. (Abstract). In: Proceedings of the 22nd Annual Meeting of the Plant Growth Regulation Society of America, Minneapolis, Minnesota, p. 159.
Davenport, T.L., Kulkarni, V. and White, T.L. (2001a) Longevity of the fl origenic pro-moter in mango. (Abstract). In: Proceedings of the 28th Annual Meeting of the Plant Growth Regulation Society of America, Miami Beach, Florida, p. 53.
Davenport, T.L., Pearce, D.W. and Rood, S.B. (2001b) Correlation of endogenous gibberellic acid with initiation of mango shoot growth. Journal of Plant Growth Regulation 20, 308–315.
Davenport, T.L., Ying, Z., Kulkarni, V. and White, T.L. (2006a) Evidence for a translocat-able fl origenic promoter in mango. Scientia Horticulturae 110, 150–159.

Reproductive Physiology 151
Davenport, T.L., Zhang, T. and Ying, Z. (2006b) Isolation of genes potentially regulating mango fl owering. (Abstract). In: Proceedings of the 33rd Annual Meeting of the Plant Growth Regulation Society of America, Quebec City, Quebec, pp. 109–110.
Davies, P.J. (ed.) (1995) Plant Hormones – Physiology, Biochemistry and Molecular Biol-ogy. Kluwer Academic, Boston, 833 pp.
Degani, C., El-Batsri, R. and Gazit, S. (1990) Enzyme polymorphism in mango. Journalof the American Society for Horticultural Science 115, 844–847.
Dell, B. and Huang, L. (1997) Physiological response of plants to low boron. Plant and Soil 193, 103–120.
Desai, A.G., Limaye, V.P. and Gunjate, R.T. (1986) Floral biology of Alphonso, Goamankur and Keser varieties of mango. Journal of Maharashtra Agriculture University 56, 3815.
De Wet, E. and Robbertse, P.J. (1986a) Pollen studies and stigma receptivity of Mangiferaindica L. cvs Haden and Sensation. South African Mango Growers’ Association Yearbook 6, 27–32.
De Wet, E. and Robbertse, P.J. (1986b) A preliminary study of the pollen of Mangifera indica L. cv. Haden in South Africa. South African Journal of Plant and Soil 3, 87–89.
De Wet, E., Robbertse, P.J. and Goeneveld, M.T. (1989) The infl uence of temperature and boron on pollen germination in Mangifera indica L. South African Journal of Plant Soil 6, 228–234.
Dutcher, R.D. (1972) Induction of early fl owering in ‘Carabao’ mango in the Philippines by smudging and ethephon application. HortScience 7, 343.
Du Toit, A.P. and Swart, D.J. (1993) Pollination of mango in the Letsitele Valley during the 1992 fl owering season. First report. South African Mango Growers’ Association Yearbook 13, 129–130.
Du Toit, A.P. and Swart, D.J. (1994) Pollination of mango in the Letsitele Valley during the 1993 fl owering season. Second report. South African Mango Growers’ Associa-tion Yearbook 14, 67–68.
Eardley, C.D. and Mansell, M.M. (1993) Preliminary report on the natural occurrence of insect pollinators in a mango orchard. South African Mango Growers’ Association Yearbook 13, 127–128.
Eardley, C.D. and Mansell, M.W. (1994) Report on the natural occurrence of insect pol-linators in a mango orchard. South African Mango Growers’ Association Yearbook14, 65–66.
Finazzo, S.F., Davenport, T.L. and Schaffer, B. (1994) Partitioning of photoassimilates in avocado (Persea americana Mill.) during fl owering and fruit set. Tree Physiology14, 153–164.
Free, J.B. and Williams, I.H. (1976) Insect pollination of Anacardium occidentalis L., Mangifera indica L., Blighiaa sopida Koenig and Persea americana Mill. Tropical Agriculture 53, 125–139.
Galang, F.G. and Lazo, F.D. (1937) The setting of ‘Carabao’ mango fruits as affected by certain sprays. Philippine Journal of Agriculture 8, 187–211.
Galán-Saúco, V. (1990) Los Frutales Tropicales en los Subtropicos. I. Aguacate, Mango, Litchi y Longan. Ediciones Mundi-Prensa, Madrid, 133 pp.
Galán-Saúco, V., Fernández, D.G. and López, R.T. (1991) Effects of ethephon on mango fl owering. Acta Horticulturae 291, 43–50.
Galán-Saúco, V., Fernández, D.G., Hernández, P. and García, V. (1993) Comparison of manual and ethephon-induced deblossoming of mango cv. Keitt in the Canary Is-lands. Acta Horticulturae 341, 248–255.
Galila, A.S. and El-Masry, H.M. (1991) Effect of Ethrel and S-3307 D (a new plant growth retardant) on fl owering, malformation, fruit set and yield of Ewais mango cultivar. Annals of Agricultural Science (Cairo) 36, 645–650.

T.L. Davenport152
Gangolly, S.R. (1960) Some problems in mango research need an urgent solution. Pun-jab Fruit Journal 23, 39–46.
Gangolly, S.R., Singh, R., Katyal, S.L. and Singh, D. (1957) The Mango. Indian Council of Agricultural Research (ICAR), New Delhi, 530 pp.
Gaskins, M.H. (1963) Girdling mango seedlings for inducing early fruit bearing. Pro-ceedings of the Florida State Horticultural Society 50, 360–363.
Gazit, S., Adato, I. and Roizman-Bustan, Y. (1992) Factors responsible for seasonal changes in successful pollination rate of mango fl owers in Israel. (Abstract). IV In-ternational Mango Symposium, Miami Beach, Florida, p. 36.
Goguey, T. (1990) Effets d’applications répétées de CULTAR (paclobutrazol) sur Mangiferaindica L. var. Valencia. Fruits 45, 599–607.
Goguey, T. (1993) Study of the effects of three fl ower-inducing substances on ‘Kent’ and ‘Zill’ mango. Acta Horticulturae 341, 216–224.
Goldsmith, M.H.M. (1968) The transport of auxin. Annual Review of Plant Physiology19, 347–360.
Goldsmith, M.H.M. and Ray, P.M. (1973) Intracellular localization of the active process in polar transport of auxin. Planta 111, 297–314.
Gunjate, R.T., Jorwekar, D.P. and Lad, B.L. (1983) Pollination, fruit set and fruit drop in Alphonso mango. Journal of Maharashtra Agricultural University 8, 168–170.
Gonzalez, L.G. (1923) The smudging of mango trees and its effects. Philippine Agricul-ture 12, 15–27.
Haberer, G. and Kieber, J.J. (2002) Cytokinins. New insights into a classic phytohor-mone. Plant Physiology 128, 354–362.
Halevy, A.H. (ed.) (1985–1986) CRC Handbook of Flowering. Vols 1–5. CRC Press, Boca Raton, Florida.
Halle, F., Oldeman, R.A.A. and Tomlinson, P.B. (1978) Tropical Trees and Forests: an Architectural Analysis. Springer-Verlag, New York, 437 pp.
Harris, W. (1901) Historical notes on economic plants of Jamaica. Bulletin of the Botany Department 8, 161–178.
Hartless, A.C. (1914) Mango crops and some factors infl uencing them. Agricultural Jour-nal of India 9, 141–159.
Hasdiseve, C. and Tongumpai, P. (1986) Effects of paclobutrazol on vegetative growth, fl owering, and fruiting of mango ‘Nam Doc Mai Twai #4’. In: Proceedings of the National Conference. Kasetsart University 24, Bangkok, Thailand, pp. 295–302.
Hassig, B.E. (1974) Origins of adventitious roots. New Zealand Journal of Forest Science4, 229–310.
Haw, K.C. (1986) The potential use of PP 333 on fruits in Malaysia with particular refer-ence to mangoes. In: Anon. Cultar, Tropical Crops Workshop, 20–22 January 1986, Imperial Chemical Industries (ICI), Fernhurst, Haslemere, Surrey, UK.
Hegele, M., Boonplod, N., Bangerth, F., Naphrom, D., Chattrakul, A., Sruamsiri, P. and Manochai, P. (2004) Changes in photosynthesis, IAA export from leaves and cyto-kinins in the xylem sap after girdling of young mango trees in combination with different growth regulators and their possible signifi cance for fl ower induction. Acta Horticulturae 645, 417–424.
Hegele, M., Bangerth, F., Naphrom, D., Sruamsiri, P. and Manochai, P. (2006) Control of fl ower induction in tropical/subtropical fruit trees by phytohormones using the ex-ample of longan and mango. Acta Horticulturae 727, 217–276.
Hendry, N.S., Van Staden, J. and Allan, P. (1982a) Cytokinins in citrus. I. Fluctuations in the leaves during seasonal and developmental changes. Scientia Horticulturae 16, 9–16.
Hendry, N.S., Van Staden, J. and Allan, P. (1982b) Cytokinins in citrus. II. Fluctuations during growth in juvenile and adult plants. Scientia Horticulturae 17, 247–256.

Reproductive Physiology 153
Henny, R.J. (1995) Thidiazuron increases basal bud and shoot development in Spathip-hyllum ‘Petite’. Plant Growth Regulation Society of America Quarterly 23, 13–16.
Hillier, G.R. and Rudge, T.G. (1991) Promotion of regular fruit cropping in mango with Cultar. Acta Horticulturae 291, 51–59.
Hongsbhanich, N. (1986) Effects of paclobutrazol on vegetative growth, fl owering and fruit setting in mango ‘Nam Dok Mai Twai #4’. In: Anon. Cultar, Tropical Crops Workshop, January 20–22, 1986, Imperial Chemical Industries (ICI), Fernhurst, Haslemere, Surrey, UK.
Hu, H. and Brown, P.H. (1994) Localization of boron in cell walls of squash and to-bacco and its association with pectin. Plant Physiology 105, 681–689.
Huala, E. and Sussex, I.M. (1993) Determination and cell interactions in reproductive meristems. The Plant Cell 5, 1157–1165.
Huang, T., Bohlenius, H., Erikson, S., Parcy, F. and Nilsson, O. (2005) The mRNA of the Arabidopsis gene FT moves from leaf to shoot apex and induces fl owering. Science309, 1694–1696.
Hussein, M.A., Mahmoud, H.M., Ahmed Amen, K.I. and Abo-El-Ez, A.T. (1989) Com-parative studies on sex distribution of some mango varieties Mangifera indica L. under Assuit conditions. Assuit Journal of Agricultural Science 20, 79–82.
Issarakraisila, M. and Considine, J.A. (1994) Effects of temperature on microsporogene-sis and pollen viability in mango cv. Kensington. Annals of Botany 73, 231–234.
Issarakraisila, M., Considine, J.A. and Turner, D.W. (1992) Seasonal effects on fl oral biol-ogy and fruit set of mangoes in a warm temperature region of Western Australia. Acta Horticulturae 321, 626.
Itai, C. and Vaadia, Y. (1965) Kinetin-like activity in root exudate of water stressed sun-fl ower plants. Physiologiea Plantarum 18, 941–944.
Itai, C., Richmond, A. and Vaadia, Y. (1968) The role of root cytokinins during water and salinity stress. Israel Journal of Botany 17, 187–195.
Itai, C., Ben-Zioni, A. and Ordin, L. (1973) Correlative changes in endogenous hormone levels and shoot growth induced by short heat treatments to the root. Physiologie Plantarum 29, 355–360.
Jacobs, W.P. and Case, D.B. (1965) Auxin transport, gibberellin, and apical dominance. Science 148, 1729–1731.
Jiron, L.F. and Hedstrom, I. (1985) Pollination ecology of mango (Mangifera indica L.) (Anacardiaceae) in the neotropic region. Turrialba 35, 269–277.
Jivanna Rao, P.S. (1923) Pollen sterility in relation to vegetative propagation. Journal of Madras Agricultural Student University 11, 419–426.
Joel, D.M. and Eisenstein, D. (1980) A bridge between the ovule and ovary wall in Mangifera indica L. (Anacardiaceae). Acta Botanica Neerlandica 29, 203–206.
Jones, W.W., Embleton, T.W. and Coggins, C.W., Jr (1975) Starch content of roots of ‘Kinnow’ mandarin trees bearing fruits in alternate years. HortScience 10, 514.
Joubert, J.P., Robbertse, P.J., Coetzer, L.A. and Wishart, D.L. (1993) Infl orescence char-acteristics and fl ower sex ratio studies of container-grown mango trees. South African Mango Growers’ Association Yearbook 13, 27–33.
Juliano, J.B. and Cuevas, N.L. (1932) Floral morphology of the mango (Mangifera indicaLinn.) with special reference to the Pico variety from the Philippines. PhilippineAgriculturist 21, 449–472.
Jutamanee, K., Eoomkham, S., Pichakum, A., Krisanapook, K. and Phavaphutanon, L. (2002) Effects of calcium, boron and sorbitol on pollination and fruit set in mango cv. Namdokmai. Acta Horticulturae 575, 829–834.
Kachru, R.B., Singh, R.N. and Chacko, E.K. (1971) Inhibition of fl owering in mango (Mangifera indica L.) by gibberellic acid. HortScience 6, 140–141.

T.L. Davenport154
Kachru, R.B., Singh, R.N. and Chacko, E.K. (1972) Inhibition of fl owering in Mangiferaindica L. by gibberellic acid. Acta Horticulturae 24, 206–209.
Kaiser, C. and Wolstenholme, B.N. (1994) Aspects of delayed harvest of ‘Hass’ avocado (Persea americana Mill.) fruit in a cool subtropical climate. II. Fruit size, yield, phe-nology and whole-tree starch cycling. Journal of Horticultural Science 69, 447–457.
Kende, H. and Sitton, D. (1967) The physiological signifi cance of kinetin and gibberellin like root hormones. Annals of New York Academy of Sciences 144, 235–243.
Khan, M.A., Malik, A.B., Makhdoom, M.L. and Haq, A. (1993) Investigations on the ef-fi ciency of exogenous synthetic growth regulators on fruit drop in mango (Mangiferaindica Linn.). Egyptian Journal of Horticulture 20, 1–14.
Kinet, J.M. (1993) Environmental, chemical, and genetic control of fl owering. Horticul-tural Reviews 15, 279–334.
King, R.W. and Zeevaart, J.A.D. (1973) Floral stimulus movement in Perilla and fl ower inhibition by noninduced leaves. Plant Physiology 51, 727–738.
Kinman, C.F. (1918) The Mango in Puerto Rico. Agricultural Experiment Station Bulletin No. 24. Agricultural Experiment Station, Puerto Rico, 30 pp.
Komeda, Y. (2004) Genetic regulation of time to fl ower in Arabidopsis thaliana. AnnualReview of Plant Biology 55, 521–535.
Kozlowski, T.T., Kramer, P.J. and Pallardy, S.G. (1991) The Physiological Ecology of Woody Plants. Academic Press, New York, 657 pp.
Kraus, E.J. and Kraybill, H.R. (1918) Vegetation and Reproduction with Reference to the Tomato. Bulletin of the Oregon Agricultural Experiment Station Number 149. Ore-gon State University, Corvallis, Oregon.
Krishnamurthi, S., Randhawa, G.S. and Sivaraman Nair, P.C. (1960) Growth studies in the mango (Mangifera indica L.) under Delhi (subtropical) conditions. Indian Journal of Horticulture 17, 107–118.
Kulkarni, V.J. (1986) Graft-induced off-season fl owering and fruiting in the mango (Mangifera indica L.). Journal of Horticultural Science 61, 141–145.
Kulkarni, V.J. (1988a) Chemical control of tree vigour and the promotion of fl owering and fruiting in mango (Mangifera indica L.) using paclobutrazol. Journal of Horti-cultural Science 63, 557–566.
Kulkarni, V.J. (1988b) Further studies on graft-induced off-season fl owering and fruiting in mango (Mangifera indica L.). Journal of Horticultural Science 63, 361–367.
Kulkarni, V.J. (1991) Physiology of fl owering in mango studied by grafting. Acta Horti-culturae 291, 95–104.
Kulkarni, V.J. (2004) The tri-factor hypothesis of fl owering in mango. Acta Horticulturae645, 61–70.
Kulkarni, V.J. and Rameshwar, A. (1978) Natural and gibberellic acid induced partheno-carpy in mango. Current Science 47, 353–355.
Kulkarni, V.J. and Rameshwar, A. (1989) Effect of deblossoming and defruiting on off-season fl owering and fruiting in mango (Mangifera indica L.). Scientia Horticulturae39, 143–148.
Kurian, R.M. and Iyer, C.P.A. (1993a) Chemical regulation of tree size in mango (Mangifera indica L.) cv. Alphonso. II. Effects of growth retardants on fl owering and fruit set. Journal of Horticultural Science 68, 355–360.
Kurian, R.M. and Iyer, C.P.A. (1993b) Chemical regulation of tree size in mango (Mangifera indica L.) cv. Alphonso. III. Effects of growth retardants on yield and quality of fruits. Journal of Horticultural Science 68, 361–364.
Kurian, R.M., Murti, G.S.R. and Iyer, C.P.A. (1992) Changes in cytokinin levels in mango (Mangifera indica. L.) leaf extracts following soil drenches with paclobutrazol. Gar-tenbauwissenschaft 57, 84–86.

Reproductive Physiology 155
Kurian, R.M., Iyer, C.P.A. and Murti, G.S.R. (1994) Total phenols of stem apical bud in relation to tree vigour in mango (Mangifera indica L.). Gartenbauwissenschaft 59, 268–270.
Lakshminarayana, S. and Aguilar, P.H. (1975) Effect of orchard heating in reducing par-thenocarpic fruits in ‘Haden’ mango. Proceedings of the Horticultural Society 88,502–505.
Lang, A. (1965) Physiology of fl ower initiation. In: Ruhland, W. (ed.) Handbuch der Pfl anzenphysiologie XV/1. Springer Verlag, Berlin, pp. 1380–1536.
Lang, A. (1984) Die photoperiodische regulation von forderung und hemmung der blutenbildung. Bericht Deutsch Botanik Gesellschaft 97, 293–314.
Lang, A., Chailakhyan, M.K. and Frolova, I.A. (1977) Promotion and inhibition of fl ower formation in a day neutral plant in grafts with a short-day plant and a long-day plant. Proceedings of the National Academy of Sciences USA 74, 2412–2416.
Law, D.M. (1987) Gibberellin-enhanced indole-3-acetic acid biosynthesis: D-tryptophan as the precursor of indole-3-acetic acid. Physiologia Plantarum 70, 626–632.
Law, D.M. and Hamilton, R.H. (1989) Reduction in the free indole-3-acetic acid levels in Alaska pea by the gibberellin biosynthsis inhibitor, uniconazol. Physiologia Plan-tarum 76, 535–538.
Lomax, T.L., Muday, G.K. and Rubery, P.H. (1995) Auxin transport. In: Davies, P.J. (ed.) Plant Hormones. Kluwer Academic Publishers, Dordrecht, pp. 509–530.
Lopez, M.R., Mosqueda-Vázquez, R. and Pimienta, B.E. (1984) El nitrato de potasio como promotor de etileno endogeno y la induccion fl oral del mango (Mangiferaindica) cv. Manila. Resumes. X Congreso Nacional de Fitogenetica Instituto. Tecno-logico Agropecuario No. 20 de Aguascalientes, p. 110.
Lumsden, P.J. (1993) Mechanisms of signal transduction. In: Jorden, B.R. (ed.) The Mo-lecular Biology of Flowering. CAB International, Wallingford, UK, pp. 21–45.
Lynch, S.J. and Mustard, M.J. (1950) Mangoes in Florida. Florida Department of Agricul-ture, Florida, 82 pp.
MacMillan, H.E. (1991) Tropical Planting and Gardening, 6th edn. McMillan, London. 767 pp.
Maiti, S.K. (1971) Effect of photoperiod on growth and fl owering of mango (Mangiferaindica L.). Indian Agriculture 15, 213–216.
Maiti, S.C. (1973) Effect of gibberellic acid (GA3) sprays on sex expression of mango (Mangifera indica L.). Science and Culture 39, 150–151.
Maiti, S.C. and Sen, P.K. (1978) Studies on photoperiodic response of biennial and regu-lar bearing mangoes. Udyanika 2, 11–16.
Maiti, S.C., Basu, R.N. and Sen, P.K. (1972) Chemical control of growth and fl owering in Mangifera indica L. Acta Horticulturae 24, 192–195.
Maiti, S.K., Maiti, S.C. and Sen, P.K. (1978) Effects of short day with and without ringing on growth and fl owering of mango (Mangifera indica L.). Indian Agriculture 22, 159–164.
Majumder, P.K. and Mukherjee, S.K. (1961) Studies on the variability of sex-expression in mango (Mangifera indica L.). Indian Journal of Horticulture 18, 12–19.
Mallik, P.C. (1951) Inducing fl owering in mango by ringing the bark. Indian Journal of Horticulture 8, 1–10.
Mallik, P.C. (1957) Morphology and biology of the mango fl ower. Indian Journal of Hor-ticulture 14, 1–23.
Mallik, P.C., Sahay, R.K. and Singh, D.L. (1959) Effect of hormones on the sex ratio in the mango. Current Science 28, 410.
McGregor, S.E. (1974) Insect pollination of tropical crops. In: Compte-Rendue du III Symposium International sur la Pollinisation. International Bee Research Associa-tion, Prague, pp. 47–55.

T.L. Davenport156
McGregor, S.E. (1976) Flowering and fruiting of plants. Pollinating agents and their com-parative value. Avocado. In: Insect Pollination of Cultivated Crop Plants. Agriculture Handbook No. 496. United States Department of Agriculture (USDA), Washington, DC, pp. 7–15, 19–23, 93–98.
Menzel, C.M. (1983) The control of fl oral initiation in lychee: a review. Scientia Horti-culturae 21, 201–215.
Menzel, C.M. and Simpson, D.R. (1994) Lychee. In: Schaffer, B. and Anderson, P.C. (eds) Handbook of Environmental Physiology of Fruit Crops. Vol. II, Sub-Tropical and Tropical Crops. CRC Press, Boca Raton, Florida, pp. 123–145.
Menzel, C.M., Watson, B.J. and Simpson, D.R. (1990) Longan. In: Bose, T.K. and Mitra, S.K. (eds) Fruits: Tropical and Subtropical. Naya Prokash, Calcutta, pp. 522–546.
Metzger, J.D. (1988) Localization of the site of perception of thermoinductive tempera-tures in Thlaspi arvense. Plant Physiology 88, 424–428.
Miller, C.O. (1963) Kinin and kinin-like compounds. In: Linskens, H.F. and Tracey, M.V. (eds) Modern Methods of Plant Analysis. Vol. 6. Springer, Berlin, pp. 194–202.
Mishra, K.A. and Dhillon, B.S. (1978) Carbohydrates and mineral composition of leaves in relation to fruit-bud differentiation in ‘Langra’ mango. Indian Journal of Agricul-tural Sciences 47, 46–50.
Monselise, S.P. and Goldschmidt, E.E. (1982) Alternate bearing in fruit trees. Horticul-tural Reviews 4, 128–173.
Morgan, P.W. and Gausman, H.W. (1966) Effects of ethylene on auxin transport. PlantPhysiology 41, 45–52.
Morgan, P.W., Jordan, W.R., Davenport, T.L. and Durham, J.L. (1977) Abscission re-sponses to moisture stress, auxin transport inhibitors, and ethephon. Plant Physiol-ogy 59, 710–712.
Morris, D.A. and Thomas, T. (1978) A microautoradiographic study of auxin transport in the stem of intact pea seedlings (Pisum sativum L.). Journal of Experimental Biology 29, 147–157.
Morton, J.F. (1964) Honeybee plants of south Florida. Proceedings of the Florida State Horticultural Society 77, 415–436.
Mosqueda-Vázquez, R. and de los Santos de la Rosa, F. (1981) Aspersiones de nitrato de potasio para adelantar e inducir la fl oracion del mango cv. Manila en Mexico. Pro-ceedings of the Tropical Region of the American Society for Horticultural Science25, 311–316.
Mosqueda-Vázquez, R. and Avila-Resendiz, C. (1985) Induccion fl oral del mango con aplicaciones de KNO3 y su inhibicion al aplicar AgNO3 o CoCl2. Horticultura Mex-icana 1, 93–101.
Mouradov, A., Cremer, F. and Coupland, G. (2002) Control of fl owering time: interacting pathways as a basis for diversity. The Plant Cell 14, S111–S130.
Mukherjee, S.K. (1949a) The mango and its relatives. Science and Culture 15, 5–9.Mukherjee, S.K. (1949b) A monograph on the genus Mangifera L. Lloydia 12, 73–136.Mukherjee, S.K. (1950) Cytological investigations of the mango (Mangifera indica L.)
and the allied Indian species. Proceedings of the National Institute of Science in India 16, 4.
Mukherjee, S.K. (1951) Pollen analysis in Mangifera in relation to fruit-set and taxono-my. Journal of the Indian Botanical Society 30, 49–55.
Mukherjee, S.K. (1953) The mango – its botany, cultivation, uses and future improve-ment, especially as observed in India. Economic Botany 7, 130–162.
Mukherjee, S.K., Majumdar, P.K. and Chatterjee, S.S. (1961) An improved technique of mango hybridization. Indian Journal of Horticulture 18, 302–304.
Mukhopadhyaya, A.K. (1978) A note on the effect of growth retardants and L-methionine on fl owering of mango (Mangifera indica L.). Horticulture Abstracts 48, 254.

Reproductive Physiology 157
Mullins, P.D.F. (1986) Effect of varying climatic regimes on fl ower behavior and pollina-tion in ‘Sensation’ and ‘Haden’ mango trees. South African Mango Growers’ Asso-ciation Research Report 6, 3–42.
Musahib-ud-din (1946) Fruit bud differentiation in mangoes in the Punjab. Punjab Fruit Journal 10, 30–31.
Musahib-ud-din and Dinsa, H.S. (1946) The fl oral count and fruit set studies in some of the North Indian mango varieties. Punjab Fruit Journal 10, 35–42.
Mustard, M.J. and Lynch, S.J. (1946) Flower-bud formation and development in Mangiferaindica. Botanical Gazette 108, 136–140.
Naik, K.C. and Mohan Rao, M. (1942) Some factors governing fruit bud formation in mangoes (Mangifera indica L.). Madras Agriculture Journal 30, 328–335.
Naik, K.C. and Mohan Rao, M. (1943) Studies on blossom biology and pollination in mangoes (Mangifera indica L.). Indian Journal of Horticulture 1, 107–119.
Nakasone, H.Y., Bowers, F.A.I. and Beaumont, J.H. (1955) Terminal growth and fl ower-ing behavior of the Pirie mango (Mangifera indica L.) in Hawaii. Proceedings of the American Society for Horticultural Science 66, 183–191.
Naqvi, S.S.M., Alam, S.M. and Mumtaz, S. (1990) Effect of cobalt and silver ions and naphthaleneacetic acid on fruit retention in mango (Mangifera indica L.). Austra-lian Journal of Experimental Agriculture 30, 433–435.
Naqvi, S.S.M., Alam, S.M. and Mumtaz, S. (1992) Infl uence of growth regulators, cobalt, and silver ions on fruit drop in mango (Mangifera indica L.). Pakistan Journal of Horticulture 24, 197–200.
Nartvaranant, P., Subhadrabandhu, S. and Tongumpai, P. (2000) Practical aspect in pro-ducing off-season mango in Thailand. Acta Horticulturae 509, 661–668.
Nickell, L.G. (ed.) (1983) Plant Growth Regulating Chemicals. Vols 1 and 2. CRC Press, Boca Raton, Florida.
Nitsch, J.P. (1965) Physiology of fl ower and fruit development. In: Ruhland, W. (ed.) Handbuch der Pfl anzenphysiologie XV/1. Springer Verlag, Berlin, pp. 1537–1646.
Nordstrom, A., Tarkowski, P., Tarkowska, D., Norbaek, R., Astot, C., Dolezal, K. and Sandberg, G. (2004) Auxin regulation of cytokinin biosynthesis in Arabidopsis thal-iana: a factor of potential importance for auxin-cytokinin-regulated development. Proceedings of the National Academy of Science USA 101, 8039–8044.
Núñez-Elisea, R. (1985) Flowering and fruit set of a monoembryonic and polyembry-onic mango as infl uenced by potassium nitrate sprays and shoot decapitation. Pro-ceedings of the Florida State Horticultural Society 98, 179–183.
Núñez-Elisea, R. (1986) Producción Temprana de Mango ‘Haden’ y ‘Manila’ con Asper-siones de Nitrato de Potasio. Folleto para productores No. 8. Centro de Investiga-ciones Agrícolas Pacífi co Centro (CIAPAC), Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP), Secretaría de Agricultura y Recursos Hidráulicos (SARH), Campo Agrícola Experimental Tecomán, Tecomán, Mexico.
Núñez-Elisea, R. (1988) Nitrato de Amonio: Nueva Alternativa para Adelantar la Flo-ración y Cosecha del Mango. Desplegable para productores No. 4. Secretaría de Agricultura y Recursos Hidráulicos (SARH)-Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP), Campo Experimental Tecomán, Tecomán, Colima, Mexico.
Núñez-Elisea, R. (1994) Environmental, developmental, and bioregulator control of fl owering in mango. PhD thesis, University of Florida, Gainesville, Florida, USA.
Núñez-Elisea, R. and Caldeira, M.L. (1988) Induction of fl owering in mango (Mangiferaindica L.) with ammonium nitrate sprays. HortScience 23, 833.
Núñez-Elisea, R. and Davenport, T.L. (1983) Abscission and ethylene production of mango (Mangifera indica L.) fruit cv Tommy Atkins. Proceedings of the Florida State Horticultural Society 96, 185–188.

T.L. Davenport158
Núñez-Elisea, R. and Davenport, T.L. (1984) Ethylene production by mango fruitlets and its role in fruit abscission. (Abstract). In: Proceedings of the 11th Annual Meeting of the Plant Growth Regulation Society of America, Boston, Massachusetts, p. 156.
Núñez-Elisea, R. and Davenport, T.L. (1986) Abscission of mango fruitlets as infl uenced by enhanced ethylene biosynthesis. Plant Physiology 82, 991–994.
Núñez-Elisea, R. and Davenport, T.L. (1989) Expression of an endogenous fl owering promoter in mango (Mangifera indica). (Abstract). In: Proceedings of the 16th An-nual Meeting of the Plant Growth Regulation Society of America, Arlington, Vir-ginia, pp. 245–247.
Núñez–Elisea, R. and Davenport, T.L. (1991a) Flowering of ‘Keitt’ mango in response to deblossoming and gibberellic acid. Proceedings of the Florida State Horticultural Society 104, 41–43.
Núñez-Elisea, R. and Davenport, T.L. (1991b) Effect of duration of low temperature treat-ment on fl owering of containerized ‘Tommy Atkins’ mango. (Abstract). In: Proceed-ings of the 18th Annual Meeting of the Plant Growth Regulation Society of America, Boston, Massachusetts, pp. 39–41.
Núñez-Elisea, R. and Davenport, T.L. (1992a) Infl uence of water defi cit on vegetative growth and fl owering of containerized mango. In: Davenport, T.L. and Harrington, H.M. (eds) Plant Stress in the Tropical Environment, Proceedings of a Workshop held in Kailua-Kona, Hawaii. USDA/Co-operative States Research Service/Carib-bean Basin Administrative Group/Pacifi c Basin Administrative Group, Gainesville, Florida, pp. 24–25.
Núñez-Elisea, R. and Davenport, T.L. (1992b) Requirement for mature leaves during fl oral induction and fl oral transition in developing shoots of mango. Acta Horticul-turae 296, 33–37.
Núñez-Elisea, R. and Davenport, T.L. (1994a) Effects of exogenous GA3 and temperature on mango infl orescence formation. (Abstract). In: Proceedings of the 21st Annual Meeting of the Plant Growth Regulation Society of America, Portland, Oregon, pp. 60–61.
Núñez-Elisea, R. and Davenport, T.L. (1994b) Flowering of mango trees in containers as infl uenced by seasonal temperature and water stress. Scientia Horticulturae 58, 57–66.
Núñez-Elisea, R. and Davenport, T.L. (1995) Effect of leaf age, duration of cool tempera-ture treatment, and photoperiod on bud dormancy release and fl oral initiation in mango. Scientia Horticulturae 62, 63–73.
Núñez-Elisea, R. and Davenport, T.L. (1998) Gibberellin and temperature effects on dormancy release and shoot morphogenesis of mango (Mangifera indica L.). Scien-tia Horticulturae 77, 11–21.
Núñez-Elisea, R., Caldeira, M.L. and Davenport, T.L. (1990) Thidiazuron effects on growth initiation and expression in mango (Mangifera indica L.). HortScience 25, 1167.
Núñez-Elisea, R., Davenport, T.L. and Caldeira, M.L. (1991) An experimental system to study mango fl owering using containerized trees propagated by air-layering. Pro-ceedings of the Florida State Horticultural Society 104, 39–41.
Núñez-Elisea, R., Caldeira, M.L., Ferreira, W. and Davenport, T.L. (1992) Adventitious rooting of ‘Tommy Atkins’ mango air layers induced with naphthaleneacetic acid. HortScience 27, 926.
Núñez-Elisea, R., Davenport, T.L. and Caldeira, M.L. (1993) Bud initiation and morpho-genesis in ‘Tommy Atkins’ mango as affected by temperature and triazole growth retardants. Acta Horticulturae 341, 192–198.
Núñez-Elisea, R., Davenport, T.L. and Caldeira, M.L. (1996) Control of bud morphogen-esis in mango (Mangifera indica L.) by girdling, defoliation and temperature modi-fi cation. Journal of Horticultural Science 71, 25–40.

Reproductive Physiology 159
Ogawa, Y. (1963) Studies on the conditions for gibberellin assay using rice seedling. Plant and Cell Physiology 4, 227–237.
Ona, M.L.D. and de Guzman, E.V. (1982) Floral gross morphology and histology of ‘Carabao’ mango (Mangifera indica L.) treated with a chemical fl owering inducer. Philippine Agriculturist 65, 201–203.
Oosthuyse, S.A. (1991a) Stages of development of the mango panicle. South African Mango Growers’ Association Yearbook 11, 59–61.
Oosthuyse, S.A. (1991b) Effect of Promalin on fl owering and lateral branching of young man-go trees in the nursery. South African Mango Growers’ Association Yearbook 11, 54.
Oosthuyse, S.A. (1995a) Effect of aqueous application of GA3 on fl owering of mango trees: why in certain instances is fl owering prevented, and in others fl owering is only delayed? In: Mango 2000 – Marketing Seminar and Production Workshop Proceedings. Department of Primary Industries, Brisbane, pp. 75–85.
Oosthuyse, S.A. (1995b) Effect of post-bloom aqueous spray application of GA3, NAA, and CPPU on fruit retention fruit size, and yield in ‘Tommy Atkins’ and ‘Heidi’ mango. In: Mango 2000 – Marketing Seminar and Production Workshop Proceed-ings. Department of Primary Industries, Brisbane, pp. 105–109.
Oslund, C.R. and Davenport, T.L. (1987) Seasonal enhancement of fl ower development in ‘Tahiti’ limes by marcottage. HortScience 22, 498–501.
Ou, S.K. (1980) Temperature effect on fl owering and fruiting in the ‘Irwin’ mango (Mangifera indica L.). Journal of Agricultural Research in China 29, 301–308.
Ou, S.K. (1982) Temperature effect on differential shoot development of mango during fl owering period. Journal of Agricultural Research in China 31, 209–212.
Ou, S.K. and Yen, C.R. (1985) Effect of growth regulators on the fl owering of ‘Irwin’ mango at high temperature. In: Forcing Culture of Fruit Trees. Taichung District Agricultural Improvement Station, Taichung, Taiwan, pp. 137–143.
Pal, R.N. and Chadha, K.L. (1982) Deblossoming mangoes with cycloheximide. Journalof Horticultural Science 57, 331–332.
Pal, S. and Ram, S. (1978) Endogenous gibberellins of mango shoot-tips and their sig-nifi cance in fl owering. Scientia Horticulturae 9, 369–379.
Pandey, R.M. (1989) Physiology of fl owering in mango. Acta Horticulturae 231, 361–380.Pandey, R.M. and Narwadkar, P.R. (1984) Studies on the induction of growth and fl ower-
ing in Dashehari mango. Indian Journal of Horticulture 41, 171–176.Pandey, R.M., Singh, R.N. and Sinha, G.C. (1973) Usefulness of ethrel in regulating
fl ower bearing in mango. Science and Culture 39, 148–150.Pantastico, E.B. and Manuel, F.C. (1978) The Philippines Recommends for Mango 1979.
Report for Philippines Council for Agriculture and Resources Research, the Philip-pines, p. 43.
Parisot, E. (1988) Study of the growth rhythm in young mango plants (Mangifera indicaL.). III. Growth and development of young mango plants. Fruits 43, 235–247.
Paulas, D. and Shanmugavelu, K.G. (1989) Physiological and biochemical changes in the leaf tissues from quiescent to fruiting stages of mango. Acta Horticulturae 231, 394–398.
Pearce, D.W., Koskioka, M. and Pharis, R.P. (1994) Review – chromatography of gib-berellins. Journal of Chromatography A 658, 91–122.
Peng, L.Z., Tang, J., Xu, E., Song, N.J. and Lei, T. (1991) Effect of paclobutrazol on the morphological and anatomical characteristics of the new organs of citrus trees. China Citrus 20, 7–10.
Perilleux, C. and Bernier, G. (2002) The control of fl owering: do genetical and physio-logical approaches converge? In: O’Neill, S.D. and Roberts, J.A. (eds) Plant Repro-duction, Annual Plant Reviews. Sheffi eld Academic Press, Sheffi eld, pp. 1–32.

T.L. Davenport160
Perilleux, C., Bernier, G. and Kinet, J.M. (1994) Circadian rhythms and the induction of fl owering in the long-day grass Lolium temulentum L. Plant Cell Environment 17, 755–761.
Pharis, R.P. and King, R.W. (1985) Gibberellins and reproductive development in seed plants. Annual Review of Plant Physiology 36, 517–568.
Pharis, R.P., Kuo, C.C. and Glenn, J.L. (1972) Gibberellin, a primary determinant in the expression of apical dominance, apical control and geotropic movement of conifer shoots. In: Carr, D.J. (ed.) Plant Growth Substances, 1970. Springer-Verlag, New York, pp. 441–448.
Pimentel, R.B., Coronel, R.E. and Espino, R.F.C. (1984) Floral biology and fruit set in mango (Mangifera indica L.) cultivars: ‘Carabao’, ‘Pico’ and ‘Kancha Mitha’. Philip-pine Journal of Crop Science 9, 47–51.
Ploetz, R.C., Ramos, J.L., Parrado, E.S. and Shepard, E.S. (1991) Shoot and root growth cycles of avocado in South Florida. Proceedings of the Florida State Horticultural Society 104, 21–24.
Ploetz, R.C., Ramos, J.L. and Parrado, J.L. (1993) Periodicity of shoot and root growth in grafted avocado. Tropical Agriculture 70, 248–251.
Pongsomboon, W. (1991) Effects of temperature and water stress on tree growth, fl ower-ing, fruit growth and retention of mango (Mangifera indica L.). PhD thesis, Kasetsart University, Bangkok, Thailand.
Pongsomboon, W., Stephenson, R.A., Whiley, A.W. and Subhadrabandu, S. (1991) Development of water stress in juvenile mango (Mangifera indica L.) trees. Acta Horticulturae 321, 496–503.
Popenoe, W. (1917) The Pollination of the Mango. US Department of Agriculture (USDA) Bulletin 542. USDA, Washington DC.
Popenoe, W. (1920) Manual of Tropical and Sub-tropical Fruits. Macmillan, New York, 474 pp.
Prakash, S. and Ram, S. (1984) Naturally occurring auxins and inhibitor and their role in fruit growth and drop of mango ‘Dashehari’. Scientia Horticulturae 2, 241–248.
Prakash, S. and Ram, S. (1986) Effect of various concentrations of auxins and their time of application on fruit retention in mango (Mangifera indica L.) cv. Chausa. Proges-sive Horticulture 18, 167–174.
Protacio, C.M. (2000) A model for potassium nitrate-induced fl owering in mango. Acta Horticulturae 509, 545–552.
Putterill, J., Laurie, R. and Macknight, R. (2004) It’s time to fl ower: the genetic control of fl owering time. Bioessays 26, 363–373.
Rademacher, W. (1991) Biochemical effects of plant growth retardants. In: Gausman, H.W. (ed.) Plant Growth Regulators. Marcel Dekker, New York, pp. 169–200.
Rademacher, W. (2000a) Growth retardants: effects on gibberellin biosynthesis and oth-er metabolic pathways. Annual Review of Plant Physiology and Plant Molecular Biology 51, 501–531.
Rademacher, W. (2000b) Growth retardants in agriculture and horticulture. In: Proceed-ings of the 27th Annual Meeting of the Plant Growth Regulation Society of America, Kailua-Kona, Hawaii, pp. 87–92.
Raja, M.E., Kumar, S.C.A. and Raju, S.Y. (2005) Boron defi ciency in mango (Mangiferaindica L.): a cause delineation study in acidic soils of Maharashtra, India. Soil Science and Plant Nutrition 51, 751–754.
Rajan, S. and Ram, S. (1989) Studies on the root regeneration in mango air-layers. Acta Horticulturae 231, 192–197.
Rajput, C.B.S. and Singh, J.N. (1989) Effects of urea and GA3 sprays on the growth, fl ow-ering and fruiting characters of mango. Acta Horticulturae 231, 301–305.

Reproductive Physiology 161
Ram, S. (1983) Hormonal control of fruit growth and fruit drop in mango cv Dashehari. Acta Horticulturae 134, 169–178.
Ram, S. and Pal, S. (1979) Studies on the naturally occurring gibberellins in mango (Mangifera indica L.) fruit. Journal of Horticultural Science 54, 209–215.
Ram, S., Bist, L.D., Lakhanpal, S.C. and Jamwal, I.S. (1976) Search of suitable pollinizers for mango cultivars. Acta Horticulturae 57, 253–263.
Ram, S., Sirohi, S.C. and Rathore, V.S. (1983) Naturally occurring cytokinins in mango (Mangifera indica L.) fruit. Australian Journal of Plant Physiology 10, 65–73.
Rameshwar, A. (1989) Mango fl owering – stress induced. Acta Horticulturae 231, 433–439.
Ramina, A., Hackett, W.P. and Sachs, R.M. (1979) Flowering in Bougainvillea – function of assimilate supply and nutrient diversion. Plant Physiology 64, 810–813.
Ramina, A., Masia, A. and Vizzoto, G. (1986) Ethylene and auxin transport and metabo-lism in peach fruit abscission. Journal of the American Society for Horticultural Science 111, 760–764.
Randhawa, G.S. and Damodaran, V.K. (1961a) Studies on fl oral biology and sex-ratio in mango (Mangifera indica L.) I. A review. Indian Journal of Horticulture 18, 9–35.
Randhawa, G.S. and Damodaran, V.K. (1961b) Studies on fl oral biology and sex ratio in mango (Mangifera indica L.) var. Chausa, Dashehari and Krishanbhog. III. Anthesis, dehiscence, receptivity of stigma, pollination, fruit set and fruit development. Indi-an Journal of Horticulture 18, 51–64.
Rao, M.M. (1997) Towards Making Mango a Regular Bearer. University of Agricultural Sciences Technology Series No. 5, Dharwad, India, 94 pp.
Rao, M.M. and Srihari, D. (1998) Approaches for managing the problems of biennial bearing in Alphonso mango trees. Journal of Maharashtra Agricultural University 23, 19–21.
Rao, M.M., Srihari, D. and Patil, V.S. (1997) Further studies on chemical induction of fl owering directly on fruited shoots in off phase Alphonso mango trees. Karnataka Journal of Agricultural Science 10, 598–601.
Rath, S. and Das, G.C. (1977) Effect of ringing, notching and defoliation in mango shoots of Langra on fl ower induction. Orissa Journal of Horticulture 5, 1.
Rath, S. and Das, G.C. (1979) Effect of ringing and growth retardants on growth and fl owering of mango. Scientia Horticulturae 10, 101–104.
Rath, S., Das, G.C. and Singh, R.L. (1982) Manipulation of fl owering in mango by forc-ing the dormant buds. Bangladesh Horticulture 10, 39–41.
Ravishankar, H. and Mohan Rao, M. (1982) Changes in carbohydrate fractions and min-erals in cultivar Alphonso mango (Mangifera indica L.) shoots. Journal of Maharash-tra Agricultural University 7, 143–145.
Ravishankar, H., Rao, M.M. and Bojappa, K.M. (1979) Fruit-bud differentiation in mango ‘Alphonso’ and ‘Totapuri’ under mild tropical rainy conditions. Scientia Horticulturae10, 95–99.
Ravishankar, H., Nalawadi, U.G. and Hulamani, N.C. (1993) Use of growth regulators to manipulate alternate bearing rhythm in mango (Mangifera indica L.). Karnataka Journal of Agricultural Science 6, 7–12.
Reece, P.C., Furr, J.R. and Cooper, W.C. (1946) The inhibiting effect of the terminal bud on fl ower formation in the axillary buds of the ‘Haden’ mango. American Journal of Botany 33, 209–210.
Reece, P.C., Furr, J.R. and Cooper, W.C. (1949) Further studies of fl oral induction in the Haden mango (Mangifera indica L.). American Journal of Botany 36, 734–740.
Riov, J. and Goren, R. (1979) Effect of ethylene on auxin transport and metabolism in midrib sections in relation to leaf abscission of woody plants. Plant, Cell and Envi-ronment 2, 83–89.

T.L. Davenport162
Riov, J. and Goren, R. (1980) Effect of ethylene on auxin transport and metabolism in midrib sections in relation to leaf abscission of woody plants. Israel Journal of Botany28, 60.
Robbertse, P.J., De Wet, E. and Coetzer, L.A. (1988) The infl uence of temperature and boron on pollen tube growth in mango. South African Mango Growers’ Association Yearbook 8, 4–6.
Robbertse, P.J., De Wet, E. and Coetzer, L.A. (1990) The infl uence of boron application on fruit set and yield of mangos. South African Mango Growers’ Association Year-book 10, 24–27.
Robbertse, P.J., Coetzer, L.A., Tomer, E. and Truscott, M. (1993) Sexual compatibility between different mango cultivars. South African Mango Growers’ Association Yearbook 13, 24–26.
Robbertse, P.J., Coetzer, L.A., Tomer, E. and Smith, M.F. (1994) Sexual compatibility between different mango cultivars. South African Mango Growers’ Association Yearbook 14, 18–20.
Robert, J.P. and Wolstenholme, B.N. (1992) Phenological cycles, carbohydrate, status, and CPPU spray trials for four mango cultivars – current research in the Nkwalini Valley. South African Mango Growers’ Association Yearbook 12, 9–13.
Roberts, J.A. and Osborne, D.J. (1981) Auxin and the control of ethylene production during the development and senescence of leaves and fruits. Journal of Experimen-tal Botany 32, 875–889.
Rodriguez, A.G. (1932) Infl uence of smoke and ethylene on the fruiting of pineapple (Ananas sativus Schult.). Journal of the Agricultural University of Puerto Rico16, 5–18.
Ross, S.D., Pharis, R.P. and Binder, W.D. (1983) Growth regulators and conifers: their physiology and potential uses in forestry. In: Nickell, L.G. (ed.) Plant Growth Regu-lating Chemicals, Vol. II. CRC Press, Boca Raton, Florida, pp. 35–78.
Sachs, R.M. (1977) Nutrient diversion: an hypothesis to explain the chemical control of fl owering. HortScience 12, 220–222.
Sachs, R.M. and Hackett, W.P. (1983) Source-sink relationships and fl owering. In: Meudt, W.J. (ed.) Strategies of Plant Reproduction. Allenheld, Osmun, Ottawa, pp. 263–272.
Sachs, R.M., Hackett, W.P., Ramina, A. and Maloof, C. (1979) Photosynthetic assimila-tion and nutrient diversion as controlling factors in fl ower initiation in Bougainvil-lea (San Diego Red) and Nicotiana tabacum cv. Wis. 38. In: Marcelle, R., Clysters, H. and Van Poucke, M. (eds) Photosynthesis and Plant Development. Dr W. Junk Publishers, The Hague, the Netherlands, pp. 95–102.
Sadhu, M.K. and Bose, S. (1988) Some endogenous factors affecting rooting of mango cuttings. Gartenbauwissenschaft 53, 137–141.
Saha, B.P. and Chhonkar, V.S. (1972) Studies on the fl oral biology, pollen viability, and intervarietal compatibility in mango (Mangifera indica L.). (Abstract). In: Program and Abstracts of the Third International Symposium Sub-tropical and Tropical Hor-ticulture, Horticultural Society of India, Bangalore, India, p. 5.
Saidha, T., Rao, V.N.M. and Santhanakrishnan, P. (1983) Internal leaf ethylene levels in relation to fl owering in mango (Mangifera indica L.). Indian Journal of Horticulture40, 139–145.
Salisbury, F.B. and Ross, C.W. (1992) Plant Physiology. Wadsworth, Belmont, California, 540 pp.
Salomon, E. and Reuveni, O. (1994) Effect of paclobutrazol treatment on the growth and fi rst fl owering of intact and autografted seedlings of mango. Scientia Horticulturae60, 81–87.
Samach, A. and Wigge, P.A. (2005) Ambient temperature perception in plants. CurrentOpinion in Plant Biology 8, 483–486.

Reproductive Physiology 163
Sánchez-Sánchez, E., Cabrera-Carbajal, F., Padilla-Valenzuela, I., Samaniego Russo, J.A. and Aboytia-Mendivil, R. (2004) Gibberellic acid effect on sprouting and nutri-tional balance of young trees of ‘Keitt’ mango at the Mayo Valley, Sonora. Acta Horticulturae 645, 447–452.
Schaffer, B., Whiley, A.W. and Crane, J.H. (1994) Mango. In: Schaffer, B. and Andersen, P.C. (eds) Handbook of Environmental Physiology of Fruit Crops Vol. II, Subtropical and Tropical Crops. CRC Press, Boca Raton, Florida, pp. 165–197.
Schaffer, B., Whiley, A.W. and Searle, C. (1999) Atmospheric CO2 enrichment, root re-striction, photosynthesis, and dry-matter partitioning in subtropical and tropical fruit crops. HortScience 34, 1033–1037.
Schnell, R.J., Olano, C.T., Quintanilla, W.E. and Meerow, A.W. (2005) Isolation and characterization of 15 microsatellite loci from mango (Mangifera indica L.) and cross-species amplifi cation in closely related taxa. Molecular Ecology Notes 5, 625–627.
Scholefi eld, P.B. (1982) Scanning electron microscope study of fl owers of avocado, li-tchi, macadamia and mango. Scientia Horticulturae 16, 263–272.
Scholefi eld, P.B. and Oag, D.R. (1984) Flowering and fruit set of six cultivars of mango. In: Proceedings of the First Australian Mango Research Workshop. Commonwealth Scientifi c and Industrial Research Organization (CSIRO), Melbourne, pp. 96–103.
Scholefi eld, P.B., Oag, D.R. and Sedgley, M. (1986) The relationship between vegetative and reproductive development in the mango in northern Australia. Australian Jour-nal of Agricultural Research 37, 425–433.
Scott, T.K., Case, D.B. and Jacobs, W.P. (1967) Auxin-gibberellin interaction in apical dominance. Plant Physiology 42, 1329–1333.
Searle, N.E. (1965) Physiology of fl owering. Annual Review of Plant Physiology 16, 97–118.
Searle, C., Whiley, A.W., Simpson, D.R. and Saranah, J.B. (1995) A preliminary pheno-physiological model for ‘Kensington’ mango in subtropical Australia. In: Mango2000 – Marketing Seminar and Production Workshop Proceedings. Department of Primary Industries, Brisbane, pp. 127–135.
Sen, P.K. (1939) Annual Report Fruit Research Station Sabour, India for 1938–1939. Fruit Research Station, Sabour, India, pp. 12–18.
Sen, P.K. and Mallik, P.C. (1941) The time of differentiation of the fl ower bud of the mango. Indian Journal of Agricultural Sciences 11, 74–79.
Sen, P.K. and Roy, R.S. (1935) Mineral nutrition of mango. Proceedings of the Indian Science Congress 111, 123.
Sen, P.K., Mallik, P.C. and Ganguly, B.D. (1946) Hybridization of the mango. IndianJournal of Horticulture 4, 4–15.
Sen, P.K., Maiti, S.K. and Maiti, S.C. (1972) Studies on inductions of axillary fl owering in Mangifera indica L. Acta Horticulturae 24, 185–188.
Sen, P.K., Bandopadhyay, M., Roy, S.S. and Basu, R.N. (1973) Use of ethrel in control-ling non-uniform bearing of mango (Mangifera indica L.). Indian Agriculture 17, 285–288.
Sergent, E., Ferrari, D. and Leal, F. (1996) Effects of potassium nitrate and paclobutrazol on fl owering induction and yield of mango (Mangifera indica L.) cv. Haden. Acta Horticulturae 455, 180–187.
Sharma, D.K. and Singh, R.N. (1970) Self-incompatibility in mango (Mangifera indicaL.). Horticultural Research 10, 108–118.
Sharma, D.K. and Singh, R.N. (1972) Investigations on self-incompatibility in Mangiferaindica L. Acta Horticulturae 24, 126–130.
Shawky, I., Zidan, Z. and Dahshan, D.I. (1977) Sex distribution, fruit set and fruiting of Zebda mango infl orescence. Annals of Agricultural Science (Cairo) 20, 159.

T.L. Davenport164
Shawky, I., Zidan, Z., El-Tomi, A. and Dahsan, D.I. (1978) Effect of GA3 sprays on time of blooming and fl owering malformation in Taimour mango. Egyptian Journal of Horticulture 5, 123–132.
Shivashankara, K.S. and Mathai, C.K. (1995) Physiological diversity among the poten-tially productive branches of regular and irregular bearing mango cultivars. Photo-synthetica 31, 135–140.
Shorrocks, V.M. (1997) The occurrence and correction of boron defi ciency. Plant and Soil 193, 121–148.
Shu, Z.H. (1981) Flower differentiation of mango (Mangifera indica). National Science Council Monograph 9, 865–870.
Shu, Z.H. (1993) Chemical pruning and induction of panicles in mango. Acta Horticul-turae 341, 199–205.
Shu, Z.H. and Sheen, T.F. (1987) Floral induction in axillary buds of mango (Mangiferaindica L.) as affected by temperature. Scientia Horticulturae 31, 81–87.
Shu, Z.H., Sheen, T.F. and Lee, K.C. (1989) Current researches on the unfruitfulness of mango in Taiwan. Acta Horticulturae 231, 68.
Simao, S. and Maranhao, Z.C. (1959) Os insectos como agentes polinizadoses de Mangueira. Anais Escola Superior de Agricultura, ‘Luiz de Oucitoz’, University of São Paulo 76, 300–304.
Singh, A.K. and Rajput, C.B.S. (1990) Effect of GA, BA, and calcium on vegetative growth and fl owering in mango (Mangifera indica L.). Research and Development Report7, 1–11.
Singh, L. and Khan, A.A. (1939) Relation of growth to fruit bearing in mangoes. IndianJournal of Agricultural Sciences 9, 835–867.
Singh, L.B. (1959) Movement of fl owering substances in the mango (Mangifera indica L.) leaves. Horticultural Advance 3, 20–27.
Singh, L.B. (1960) The Mango. Botany, Cultivation and Utilization. Leonard Hill, Lon-don, 439 pp.
Singh, L.B. (1961) Biennial bearing in mango – effect of gibberellic acid and maleic hydrazide. Horticultural Advance 5, 96–106.
Singh, L.B. (1962) A new technique for inducing early fruiting in mango hybrids based on the movement of hormones. Proceedings of the Horticultural Society 75, 410–412.
Singh, L.B. (1972) Biennial bearing in mango – retrospect and prospect. Acta Horticul-turae 24, 145–148.
Singh, L.B. (1977) Mango. In: Alvim, P. de T. and Kozlowski, T.T. (eds) Ecophysiology of Tropical Crops. Academic Press, New York, pp. 479–485.
Singh, L.B. and Singh, R.N. (1956) Floral induction in axillary buds of mango shoots. Proceedings of the American Society for Horticultural Science 68, 265–269.
Singh, L.B. and Singh, R.N. (1959) Shy-bearing habit of south Indian mango varieties and their sex ratio. Annual Report Fruit Research Station Saharanpur 1957–1859.Fruit Research Station, Saharanpur, India, pp. 54–57.
Singh, R. and Singh, R.N. (1974) Growth-promoting and growth-inhibiting substances in developing fruits of biennial and regular bearing varieties of mango (Mangifera in-dica L.). Indian Journal of Horticulture 31, 16–22.
Singh, R.N. (1954a) Studies on the fl oral biology and subsequent development of fruits in the mango (Mangifera indica L.) varieties Dashehari and Langra. Indian Journal of Horticulture 11, 69–88.
Singh, R.N. (1954b) The sex ratio and fruit set in three varieties of mango. Science 119, 389–390.
Singh, R.N. (1958a) Studies in the differentiation and development of fruit buds in man-go (Mangifera indica L.) I. Review of the literature. Horticultural Advance 2, 1–7.

Reproductive Physiology 165
Singh, R.N. (1958b) Studies in the differentiation and development of fruit buds in man-go (Mangifera indica L.) II. Morphological and histological changes. Horticultural Advance 2, 37–43.
Singh, R.N. (1959) Studies in the differentiation and development of fruit buds in mango (Mangifera indica L.) varieties III. Mango shoots and fruit bud differentiation. Horti-cultural Advance 3, 28–49.
Singh, R.N. (1961) Studies in the differentiation and development of fruit-buds in mango (Mangifera indica L.). V. Effects of defoliation, decapitation and deblossoming on fruit-bud differentiation. Indian Journal of Horticulture 18, 1–11.
Singh, R.N. (1964) Sex, pollination and post-fertilization problems in mango. World Crops 16, 24–26.
Singh, R.N. (1971) Biennial Bearing in Fruit-trees. Indian Council of Agricultural Re-search (ICAR) Technical Bulletin (Agriculture) Number 30. ICAR, New Delhi.
Singh, R.N. (1978) Mango. Indian Council of Agricultural Research (ICAR), New Delhi, 99 pp.
Singh, R.N. (1979) Mango. Indian Council of Agricultural Research (ICAR) Low-priced Books Series No. 3. ICAR, New Delhi.
Singh, R.N., Majumder, P.K. and Sharma, D.K. (1962a) Age of leaf as affecting fruit bud formation in mango (Mangifera indica L.) var. Neelum. Science and Culture 28, 284–285.
Singh, R.N., Majumdar, P.K. and Sharma, D.K. (1962b) Self-incompatibility in mango (Mangifera indica L.) var. Dashehari. Current Science 31, 209.
Singh, R.N., Majumdar, P.K. and Sharma, D.K. (1965) Studies on the bearing behavior of some south Indian varieties of mango (Mangifera indica L.) under north Indian conditions. Tropical Agriculture 42, 171–174.
Singh, R.N., Majumder, P.K. and Sharma, D.K. (1966) Sex-expression in mango (Mangifera indica L.) with reference to prevailing temperature. Proceedings of the American Society for Horticultural Science 89, 228–229.
Singh, R.N., Majumder, P.K., Sharma, D.K., Sinha, G.C. and Bose, P.C. (1974) Effect of de-blossoming on the productivity of mango. Scientia Horticulturae 2, 399–403.
Singh, R.S. and Ram, S. (1983) Studies on the use of plant growth substances for fruit retention in mango cv. Dashehari. Indian Journal of Horticulture 40, 188–194.
Singh, S. and Singh, B. (1963) Alternate bearing in mango. II. Regulation of growth and bearing with some plant growth regulators. Punjab Horticulture Journal 3, 137–147.
Singh, S.N. (1961) Studies on the morphology and viability of the pollen grains of man-go. Horticultural Advance 5, 121–144.
Singh, S.N. (1963) Studies on the morphology, viability and preservation of pollen grains of mango (Mangifera indica L.), litchi (Litchi chinensis Sonn.) and loquat (Eriobotrya japonica Lindl.). Agra University Journal of Research Science 12, 317–322.
Singh, U.R. (1960) Studies in the fruit-drop of mango (Mangifera indica L.). II. Nature and extent of fruit-drop. Horticultural Advance 4, 142–154.
Singh, U.R. (1961) Studies in the fruit-drop of mango. IV. Embryo development, its de-generation and studies of fruit-pedicel and abscission zone. Horticultural Advance5, 218–227.
Singh, U.R. and Singh, A.P. (1973) Studies on the morphology and viability of pollen grains of mango. Punjab Horticulture Journal 13, 237–246.
Singh, Z. and Dhillon, B.S. (1987) Effect of foliar application of boron on vegetative and panicle growth, sex expression, fruit retention and physicochemical characters of fruits of mango (Mangifera indica L.) cv Dusheri. Tropical Agriculture 64, 305–308.
Singh, Z., Malik, A.U. and Davenport, T.L. (2005) Fruit drop in mango. Horticultural Reviews 31, 111–154.

T.L. Davenport166
Sitton, D., Itai, C. and Kende, K. (1967) Decreased cytokinin production in the roots as a factor in shoot senescence. Planta 73, 296–300.
Sivagami, S., Vijayan, K.P. and Natarajaratnam, N. (1989) Effect of nutrients and growth regulating chemicals on biochemical aspects and hormonal balance with reference to apical dominance in mango. Acta Horticulturae 231, 476–482.
Skoog, F. and Miller, C.O. (1957) Chemical regulation of growth and organ formation in plant tissues cultured in vitro. Symposium of the Society of Experimental Biology54, 118–130.
Smith, F.C. (1960) Beekeeping in the Tropics. Longmans, London, 265 pp.Soule, J. (1985) Glossary for Horticultural Crops. Wiley, New York, 898 pp.Soule, M.J. (1950) A Bibliography of the Mango (Mangifera indica L.). Florida Mango
Forum and University of Miami, Coral Gables, Florida, 89 pp.Spencer, J.L. and Kennard, W.C. (1955) Studies on mango (Mangifera indica L.) fruit set
in Puerto Rico. Tropical Agriculture 32, 323–330.Spencer, J.L. and Kennard, W.C. (1956) Limited stigmatic receptivity may contribute to
low fruit-set in the mango. Proceedings of the American Society for Horticultural Science 67, 287–289.
Sturrock, T.T. (1966) The mango infl orescence. Proceedings of the Horticultural Society53, 366–369.
Subhadrabandu, S. (1986) Studies of plant growth regulator effects on tropical and sub-tropical tree fruits of Thailand. Acta Horticulturae 175, 291–297.
Sukhvibul, N., Whiley, A.W., Smith, M.K., Hetherington, S.E. and Vithanage, V. (1999) Effect of temperature on infl orescence development and sex expression of mono- and poly-embryonic mango (Mangifera indica L.) cultivars. Journal of Horticultural Science and Biotechnology 74, 64–68.
Sukhvibul, N., Hetherington, S.E., Whiley, A.W., Smith, M.K. and Vithanage, V. (2000) Effect of temperature on pollen germination, pollen tube growth and seed develop-ment in mango (Mangifera indica L.). Acta Horticulturae 509, 609–616.
Suryanarayana, V. (1978a) Amino acid changes in mango shoots in relation to fl owering. Plant Biochemistry Journal 5, 50–57.
Suryanarayana, V. (1978b) Seasonal changes in ribonucleic acid and protein contents in mango shoots in relation to fl owering. Plant Biochemistry 5, 9–13.
Suryanarayana, V. (1978c) Seasonal changes in sugars, starch, nitrogen and carbon-nitrogen ratio in relation to fl owering in mango. Plant Biochemistry Journal 5, 108–117.
Suryanarayana, V. (1980) Amino acid changes in mango shoots as affected by growth retardants in relation to fl owering. Plant Biochemistry Journal 7, 78–82.
Suryanarayana, V. and Rao, V.N.M. (1977) Ascorbic acid changes in shoots of mango cv. Mulgoa as affected by growth retardants in relation to fl owering. Indian Journal of Plant Physiology 20, 88–90.
Takahashi, N. (ed.) (1986) Chemistry of Plant Hormones. CRC Press, Boca Raton, Flori-da, 277 pp.
Tamaki, S., Matsuo, S., Wong, H.L., Yokoi, S. and Shimamoto, K. (2007) Hd3a protein is a mobile fl owering signal in rice. Science 316, 1033–1036.
Tanaka, M., Takei, K., Kojima, M., Sakakibara, H. and Mori, H. (2006) Auxin controls local cytokinin biosynthesis in the nodal stem in apical dominance. The Plant Jour-nal 45, 1028–1036.
Teaotia, S.S., Singh, R.N., Upadhyay, S.K. and Srivastava, V.S. (1967) Effect of growth substances on fruit retention in mango (Mangifera indica L.) varieties ‘Langra’ and ‘Dashehari’. International Symposium on Sub-tropical and Tropical Horticulture.New Delhi, pp. 224–229.

Reproductive Physiology 167
Thuck-Thye, L. (1978) Ethylene and the induction of fl owering by potassium nitrate in mango (Mangifera indica L.). MSc thesis, University of the Philippines, Los Banos, the Philippines.
Tomer, E. (1984) Inhibition of fl owering in mango by gibberellic acid. Scientia Horticul-turae 24, 299–303.
Tomlinson, P.B. and Gill, A.M. (1973) Growth habits of tropical trees: some guiding principles. In: Meggers, B.J., Ayensu, E.S. and Duckworth, W.D. (eds) Tropical For-est Ecosystems in Africa and South America: a Comparative Review. Smithsonian Institution, Washington, DC, pp. 124–143.
Tongumpai, P., Hongsbhanich, N. and Voon, C.H. (1989) ‘Cultar’ – for fl owering regula-tion of mango in Thailand. Acta Horticulturae 239, 375–378.
Tongumpai, P., Jutamanee, K. and Subhadrabandhu, S. (1991a) Effect of paclobutrazol on fl owering of mango cv. Khiew Sawoey. Acta Horticulturae 291, 67–70.
Tongumpai, P., Jutamanee, K., Sethapakdi, R. and Subhandrabandu, S. (1991b) Variation in level of gibberellin-like substances during vegetative growth and fl owering of mango cv. Khiew Sawoey. Acta Horticulturae 291, 105–107.
Torres, J.P. (1931) Some notes on Carabao mango fl owers. Philippine Journal of Agricul-ture 2, 395–398.
Turnbull, G.C.N., Anderson, K.L. and Winston, E.C. (1996) Infl uence of gibberellin treat-ment on fl owering and fruiting patterns in mango. Australian Journal of Experimen-tal Agriculture 36, 603–611.
van der Meulen, J.H.E., Smith, T., van den Boom, I.B., Kok, A., Schwartz, C. and Jacobs, J. (1971) Mango Growing in South Africa. Leafl et No. 48. Citrus and Subtropical Fruit Research Institute, Nelspruit, South Africa, 40 pp.
Van Lelyveld, L.J. (1978) Peroxidase activity and isozymes in abscised and normal man-go (Mangifera indica L.) fruits. Zeitschrift für Pfl anzenphysiologie 89, 453–456.
Van Lelyveld, L.J. and Nel, E. (1982) Ethylene concentration and polyphenol oxidase activity in mango (Mangifera indica L.) fruit abscission. Zeitschrift für Pfl anzenphys-iologie 107, 179–182.
Vasil, I.K. (1963) Effect of boron on pollen germination and pollen tube growth. In: Lin-skens, H.F. (ed.) Pollen Physiology and Fertilization. North Holland, Amsterdam, pp. 107–119.
Veen, H. (1969) Auxin transport, auxin metabolism and ageing. Acta Botanica Neer-landica 18, 447–453.
Veen, H. and Jacobs, W.P. (1969) Transport and metabolism of indole-3-acetic acid in coleus petiole segments of increasing age. Plant Physiology 44, 1157–1162.
Venkataratnam, L. (1949) Hormone induced set and parthenocarpy in mango. CurrentScience 18, 409.
Verheij, E.W.M. (1986) Towards a classifi cation of tropical tree fruits. Acta Horticulturae175, 137–150.
Vijayalakshmi, D. and Srinivasan, P.S. (1999) Morpho-physiological changes as infl u-enced by chemicals and growth regulators in alternate bearing mango cv. Alphonso. Madras Agricicultural Journal 86, 485–487.
Voon, C.H., Pitakpaivan, C. and Tan, S.J. (1991) Mango cropping manipulation with Cultar. Acta Horticulturae 291, 219–228.
Wagle, P.V. (1929) A preliminary study of the pollination of the Alphonso mango. Agri-culture Journal of India 24, 259–263.
Weberling, F. (1989) Morphology of Flowers and Infl orescences. Cambridge University Press, Cambridge, UK.
Weigel, D., Alvarez, J., Smyth, D.R., Yanofsky, M.F. and Meyerowitz, E.M. (1992) LEAFY controls fl oral meristem identity in Arabidopsis. Cell 69, 843–859.

T.L. Davenport168
Went, F.W. (1943) Effect of the root system on tomato stem growth. Plant Physiology 18, 51–64.
Werner, H. (1993) Infl uence of paclobutrazol on growth and leaf nutrient content of mango (cv. Blanco). Acta Horticulturae 341, 225–231.
Wester, P.J. (1920) The mango. Philippine Bureau of Agriculture Bulletin 18, 3–70.Whiley, A.W. (1993) Environmental effects on phenology and physiology of mango – a
review. Acta Horticulturae 341, 168–176.Whiley, A.W. (1994) Ecophysiological studies and tree manipulation for maximisation of
yield potential in avocado (Persea americana Mill.). PhD thesis, The University of Natal, Pietermaritzburg, South Africa.
Whiley, A.W., Saranah, J.B., Rasmussen, T.S., Winston, E.C. and Wolstenholme, B.N. (1988) Effect of temperature on ten mango cultivars with relevance to production in Australia. In: Proceedings of the Fourth Australasian Conference on Tree and Nut Crops, Lismore, New South Wales, Australia, pp. 176–185.
Whiley, A.W., Rasmussen, T.S., Saranah, J.B. and Wolstenholme, B.N. (1989) Effect of temperature on growth, dry matter production and starch accumulation in ten man-go (Mangifera indica L.) cultivars. Journal of Horticultural Science 64, 753–765.
Whiley, A.W., Rasmussen, T.S., Wolstenholme, B.N., Saranah, J.B. and Cull, B.W. (1991) Interpretation of growth responses of some mango cultivars grown under controlled temperatures. Acta Horticulturae 291, 22–31.
Whiley, A.W., Rasmussen, T.S., Saranah, J.B. and Wolstenholme, B.N. (1996) Delayed harvest effects on yield, fruit size and starch cycling in avocado (Persea americanaMill.) in two subtropical environments. II. The late-maturing cv. Hass. Scientia Hor-ticulturae 66, 35–49.
Wigge, P.A., Kim, M.C., Jaeger, K.E., Busch, W., Schmid, M., Lohmann, J.U. and Weigel, D. (2005) Integration of spatial and temporal information during fl oral induction in Arabidopsis. Science 309, 1056–1059.
Wightman, F., Schneider, E.A. and Thimann, K.V. (1980) Hormonal factors controlling the initiation and development of lateral roots. II. Effects of exogenous growth fac-tors on lateral root formation in pea roots. Physiologia Plantarum 49, 304–314.
Wilkins, M.B. and Cane, A.R. (1970) Auxin transport in roots. Journal of Experimental Botany 21, 195–211.
Williamson, J.G. and Coston, D.C. (1989) The relationship among root growth, shoot growth, and fruit growth of peach. Journal of the American Society for Horticultural Science 114, 180–183.
Winston, E.C. (1992) Evaluation of paclobutrazol on growth, fl owering and yield of man-go cv. Kensington Pride. Australian Journal of Experimental Agriculture 32, 97–104.
Winston, E.C. and Wright, R.M. (1986) Mango fl ower induction: ethephon, potassium nitrate and cincturing. In: Proceedings of the First Australian Mango Research Work-shop. Commonwealth Scientifi c and Industrial Research Organization (CSIRO), Melbourne, pp. 202–210.
Wolfenbarger, D.O. (1977) Comments on mango pollination. Proceedings of the Florida State Horticultural Society 90, 240–241.
Wolstenholme, B.N. and Hofmeyr, D. (1985) Effects of various fl oral induction treat-ments on container-grown mango trees. South African Mango Growers’ Association Research Yearbook 5, 36–38.
Wolstenholme, B.N. and Mullins, P.C. (1982a) Flowering, pollination and fruit set of mango – a discussion of limiting factors. The Citrus and Subtropical Fruit Journal October, 11–22.
Wolstenholme, B.N. and Mullins, P.C. (1982b) Flowering, pollination and fruit set of mango – a discussion of limiting factors. South African Mango Growers’ Association Yearbook 2, 5–11.

Reproductive Physiology 169
Wolstenholme, B.N. and Whiley, A.W. (1995) Ecophysiology of the mango tree as a basis for pre-harvest management. In: Mango 2000 – Marketing Seminar and Production Workshop Proceedings. Department of Primary Industries, Brisbane, pp. 10–17.
Wu, Y., Spollen, W.G., Sharp, R.E., Hetherington, P.R. and Fry, S.C. (1994) Root growth maintenance at low water potentials: increased activity of xyloglucan endo-transglycosylase and its possible regulation by abscisic acid. Plant Physiology 106, 607–615.
Yanofsky, M.F. (1995) Floral meristems to fl oral organs: genes controlling early events in Arabidopsis fl ower development. Annual Review of Plant Physiology and Plant Mo-lecular Biology 46, 167–188.
Yelenosky, G., Vu, C.V.J. and Bausher, M.G. (1993) Paclobutrazol-induced dwarfi ng of ‘Valencia’ orange trees. Proceedings of the Florida State Horticultural Society 106, 329–332.
Yeshitela, T., Robbertse, P.J. and Stassen, P.J.C. (2004a) Paclobutrazol suppressed vegeta-tive growth and improved yield as well as fruit quality of ‘Tommy Atkins’ mango (Mangifera indica) in Ethiopia. New Zealand Journal of Crop and Horticultural Sci-ence 32, 281–293.
Yeshitela, T., Robbertse, P.J. and Stassen, P.J.C. (2004b) Effects of various inductive peri-ods and chemicals on fl owering and vegetative growth of ‘Tommy Atkins’ and ‘Keitt’ mango (Mangifera indica) cultivars. New Zealand Journal of Crop and Horticultural Science 32, 209–215.
Yeshitela, T., Robbertse, P.J. and Stassen, P.J.C. (2005) Potassium nitrate and urea sprays affect fl owering and yields of ‘Tommy Atkins’ (Mangifera indica) mango in Ethiopia. South African Journal of Plant and Soil 22, 28–32.
Young, T.W. (1942) Investigations of the unfruitfulness of the Haden mango in Florida. Proceedings of the Florida State Horticultural Society 55, 106–110.
Young, T.W. (1955) Infl uence of temperature on growth of mango pollen. Proceedings of the Florida State Horticultural Society 68, 308–313.
Young, T.W. (1958) A technique for testing mango pollen viability on artifi cial medium. Horticultural Advances 2, 72–75.
Young, T.W. and Sauls, J.W. (1979) The Mango Industry in Florida. Bulletin 189 Florida Cooperative Extension Service. University of Florida, Gainesville, pp. 1–70.
Zeevaart, J. (1976) Physiology of fl ower formation. Annual Review of Plant Physiology 27, 321–348.
Zeevaart, J.A.D. (2006) Florigen coming of age after 70 years. The Plant Cell 18, 1783–1789.
Zeevaart, J.A.D. and Boyer, G.L. (1987) Photoperiodic induction and the fl oral stimulus in Perilla. In: Atherton, J.G. (ed.) Manipulation of Flowering. Butterworths, London, pp. 269–277.
Zhang, T., Ying, Z. and Davenport, T.L. (2005) Isolation and characterization of a Constans-like gene in mango. (Abstract). In: Plant Biology 2005. Annual Meeting of the Amer-ican Society of Plant Biologists. American Society of Plant Biologists, Seattle, Wash-ington, p. 151.

© CAB International 2009. The Mango, 2nd Edition: Botany, Production and Uses170 (ed. R.E. Litz)
6 Ecophysiology
B. Schaffer,1 L. Urban,2 P. Lu3 and A.W. Whiley4
1University of Florida, Florida, USA 2Centre INRA de Corse, San Giuliano, France
3EWL Sciences, PO Box 39443, Winnellie, Northern Territory, Australia 4Sunshine Coast Horticultural Services Pty Ltd, Nambour, Queensland, Australia
6.1 Introduction 1706.2 Photosynthesis 172
Introduction 172Light 177Leaf temperature 179Elevated atmospheric CO2 concentration 181Humidity 182Flooding 182Internal factors 184Photosynthetic contributions by fruit 187
6.3 Plant Water Relations 1886.4 Tree Growth and Development 190
Light 190Temperature 192Drought 193Flooding 194Wind 195Salinity 196Elevated atmospheric CO2 concentration 197
6.5 Crop Production 198Temperature limitations to crop production 198Light interception and orchard design 199
6.6 Conclusions 200
6.1 Introduction
The genetic composition of mango cultivars is the primary determinant of yield potential. However, actual yield, as well as tree growth and develop-ment, are mediated by several endogenous factors including previous fruit load, postharvest vegetative growth, prefl owering maturity of terminal shoots,

Ecophysiology 171
production and mobilization of carbohydrates, nutritional status, plant growth substances and carbon to nitrogen ratios (Schaffer et al., 1994). These factors are either directly or indirectly affected by environmental variables such as light, temperature and water availability. Environmental conditions outside the range for optimum growth may also impose stress which results in physiological changes that reduce growth or cause permanent damage to mango trees. For example, major climatic events (i.e. extended drought, fl oods, wind storms, heat waves and freezes) can cause severe damage to crops due to development of excessive stress. However, mediated stress and the release from stress imposed by normal seasonal changes provide condi-tions that result in the progression of cropping cycles due to phenological changes in the plants. An example of benefi cial stress in mango is the improved synchrony and reliability of fl owering in subtropical climates due to cool winter temperatures. Thus, understanding the impact of the environ-ment on tree physiology and growth, and the particular adaptive strategies developed through the processes of evolution, can provide a framework to manage the crop to maximize genetic yield potential (Schaffer et al., 1994).
Physiological responses of mango to environmental variables can be related to the evolutionary centre of origin of a specifi c cultivar. Mango culti-vars are classifi ed into two ecotypes based on embryony (see Mukherjee and Litz, Chapter 1, this volume). A race with a single zygotic seed, monoembry-onic types, evolved in the dry subtropical, monsoonal regions of the Indian subcontinent with very hot summers but cooler winters. The polyembryonic types, produced through nucellar embryony, largely evolved in the consis-tently hot, humid tropics of South-east Asia where the monsoonal pattern still predominates but the dry season is shorter than that of the Indian sub-continent (Mukherjee, 1972). Hybridization occurs freely within and between the two ecotypes and has led to a proliferation of cultivars of widely varying genetic composition. Differences in growth and fl owering responses to tem-perature have been observed between the two embryonic ecotypes (Whiley et al., 1989) and selection and breeding offer potential for increasing the crop-ping performance of this notoriously low-yielding and recalcitrant tree across a wider range of environmental conditions (Schaffer et al., 1994).
Inevitably, the many unique features of the mango tree represent its evo-lutionary response to an indigenous environment that is particularly hostile, with sustained extreme heat and high evaporative demand for much of the year. This chapter provides an overview of the impact of environmental fac-tors on physiology, growth and productivity of mango. Plant responses will be considered in the context of the evolutionary origin and adaptability of the mango tree. Responses to light, temperature and water are emphasized, while the effects of atmospheric carbon dioxide (CO2) concentration, wind and salinity are also discussed.
Photosynthesis and plant water relations are closely associated with environmental conditions and directly affect plant growth and productivity. The principles of leaf gas exchange and plant water relations are discussed to provide a theoretical basis for interpreting physiological responses to the environment. The impact of photosynthesis and tree water relations on

B. Schaffer et al.172
growth and development under different environmental conditions is dis-cussed. Pollination, fertilization, fl owering and fruit set, which are strongly infl uenced by environmental factors, have been addressed elsewhere (see Davenport, Chapter 5, this volume; Schaffer et al., 1994). In the fi nal sec-tion of this chapter, mango crop production is integrated with aspects of ecophysiology.
6.2 Photosynthesis
Introduction
The net CO2 assimilation rate (Anet) in C3 plants is a function of the carboxyla-tion rate (Vc), the oxygenation rate (Vo) and the rate of CO2 evolution in light that results from processes other than photorespiration, sometimes called ‘day respiration’ (Rd):
Anet = Vc – 0.5Vo – Rd (6.1)
Rd is usually inferred from measurements of leaf CO2 exchanges after 5 min in the dark (i.e. ‘night respiration’ Rn). However, it has been repeatedly shown that Rd is lower than Rn (see Atkin et al. (2000) for review), so that light is known to inhibit respiration, with a Rd/Rn value ranging from 30 to 100% (see Peisker and Apel (2001) for review). Urban et al. (2008) established the following linear regression for Rd of mango leaves: Rd = 0.35Rn – 0.21, which may be used to infer Rd from Rn for photosynthetic photon fl ux (Q) values above 170 mol photons/m2/s.
Currently, modelling of Anet often uses the Harley et al. (1992) version of the Farquhar et al. (1980) model. According to this model, Anet can be expressed as:
Anet = (1 – 0.5O/( Ci))min(Wc, Wj, Wp) – Rd (6.2)
where O represents the partial pressure of oxygen (O2) in the intercellular air spaces (Pa), the specifi city factor of ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco). Ci is the partial pressure of CO2 in the intercellular air spaces (Pa), Wc the carboxylation rate limited by the amount, activation state or kinetic properties of Rubisco ( mol CO2/m2/s), Wj the carboxylation rate limited by the rate of ribulose bisphosphate regeneration ( mol CO2/m2/s), and Wp the carboxylation rate limited by triose phosphate utilization in sucrose and starch synthesis ( mol CO2/m2/s).
Usually O is set as 21 kPa (21%). The variable , which characterizes the ratio of the affi nities of CO2 and O2 for ribulose-1,5-bisphosphate in the active site of Rubisco, can be calculated from the CO2 compensation point * (the CO2 concentration at which photosynthesis equilibrates with respiration):
= 0.5O/ * (6.3)
where = 2220 mol CO2/mol O2 at 25°C for ‘Cogshall’ mango leaves (Urban et al., 2008), which is lower than those given by Epron et al. (1995): 2100–2900
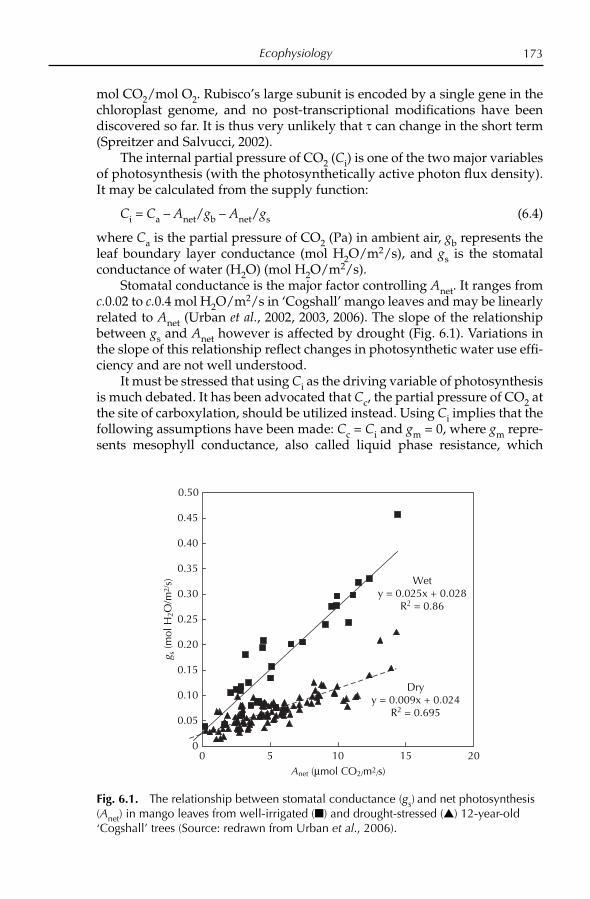
Ecophysiology 173
mol CO2/mol O2. Rubisco’s large subunit is encoded by a single gene in the chloroplast genome, and no post-transcriptional modifi cations have been discovered so far. It is thus very unlikely that can change in the short term (Spreitzer and Salvucci, 2002).
The internal partial pressure of CO2 (Ci) is one of the two major variables of photosynthesis (with the photosynthetically active photon fl ux density). It may be calculated from the supply function:
Ci = Ca – Anet/gb – Anet/gs (6.4)
where Ca is the partial pressure of CO2 (Pa) in ambient air, gb represents the leaf boundary layer conductance (mol H2O/m2/s), and gs is the stomatal conductance of water (H2O) (mol H2O/m2/s).
Stomatal conductance is the major factor controlling Anet. It ranges from c.0.02 to c.0.4 mol H2O/m2/s in ‘Cogshall’ mango leaves and may be linearly related to Anet (Urban et al., 2002, 2003, 2006). The slope of the relationship between gs and Anet however is affected by drought (Fig. 6.1). Variations in the slope of this relationship refl ect changes in photosynthetic water use effi -ciency and are not well understood.
It must be stressed that using Ci as the driving variable of photosynthesis is much debated. It has been advocated that Cc, the partial pressure of CO2 at the site of carboxylation, should be utilized instead. Using Ci implies that the following assumptions have been made: Cc = Ci and gm = 0, where gm repre-sents mesophyll conductance, also called liquid phase resistance, which
Dryy = 0.009x + 0.024
R2 = 0.695
0
0.05
0.10
0.15
0.20
0.25
0.30
0.35
0.40
0.45
0.50
0 5 10 15 20Anet (μmol CO2/m2/s)
g s (m
ol H
2O/m
2/s) Wet
y = 0.025x + 0.028R2 = 0.86
Fig. 6.1. The relationship between stomatal conductance (gs) and net photosynthesis (Anet) in mango leaves from well-irrigated (■) and drought-stressed (▲) 12-year-old ‘Cogshall’ trees (Source: redrawn from Urban et al., 2006).

B. Schaffer et al.174
encompasses diffusion from the intercellular leaf spaces to the carboxylation sites in the chloroplasts. There is a growing body of evidence that gm is not negligible in most species. The average value of gm in unstressed mango leaves (0.21 mol CO2/m2/s) (Urban et al., 2008), calculated using the method of Epron et al. (1995), is within the range of values for broadleaf species sur-veyed by Ethier and Livingston (2004) and Manter and Kerrigan (2004). The carboxylation rate (in Eqn 6.2) limited by the amount, activation state or kinetic properties of Rubisco (Wc) can be calculated as:
Wc = VcmaxCi/(Ci + Kc(1 + O/Ko)) (6.5)
where Vcmax represents the maximum rate of carboxylation ( mol CO2/m2/s), and Kc (Pa CO2) and Ko (Pa O2) are the Michaelis constants of Rubisco carboxylation and oxygenation, respectively. The Vcmax values of well-exposed mango leaves at a leaf temperature of 30°C are typically in the range of 80–100 mol CO2/m2/s (Urban et al., 2006). Specifi c values of Kc and Ko for mango leaves have not been estimated and are approximated using data from other species (i.e. cotton or tobacco).
The carboxylation rate limited by the rate of ribulose bisphosphate regen-eration (Wj) is controlled by the rate of electron fl ow J ( mol electrons/m2/s):
Wj = JCi/(4(Ci + O/ )) (6.6)
with
J = Q/(1 + 2 2Q2/Jmax2)0.5 (6.7)
where Q is the photosynthetically active photon fl ux density ( mol quanta/m2/s), represents leaf absorbance (no units), is the apparent effi ciency of light energy conversion (mol electrons/mol photons) and Jmax is the light-saturated rate of electron transport ( mol electrons/m2/s). Leaf absorbance of mango leaves, measured from 390–760 nm using an integrating sphere, was found to be close to 0.81 (Urban et al., 2008) and is in the normal range of
values of the literature (Bauerle et al., 2004). Leaf absorbance, which is pos-itively correlated with leaf chlorophyll content, may increase as a conse-quence of paclobutrazol treatments (Gonzalez and Blaikie, 2003). The apparent effi ciency of light energy conversion in mango reaches 0.32 mol electrons/
mol photons (Urban et al., 2004b), in the absence of photoinhibition or pho-todamage. This value corresponds to the mean value of operational (Sin-gaas et al., 2001). The Jmax values of well-exposed mango leaves at a leaf temperature of 30°C are typically in the 120–150 mol CO2/m2/s range. The Jmax as well as the Vcmax values are rather low when compared to values from other species and partly explain why maximal rates of leaf photosynthesis (Amax) are rather low, typically 12–15 mol CO2/m2/s.
The carboxylation rate limited by triose phosphate utilization during sucrose and starch synthesis (Wp in Equation 2), can be calculated by:
Wp = 3TPU + Vo/2 = 3TPU + Vc0.5Ci /(Ci ) (6.8)
where TPU is the rate of phosphate release in triose phosphate utilization during starch and sucrose production. The TPU is usually not included in

Ecophysiology 175
most studies on photosynthetic capacity because of methodological diffi cul-ties. However, Urban et al. (2003) found that TPU = 8–12 mol CO2/m2/s at a leaf temperature of 30°C in well-exposed ‘Cogshall’ mango leaves.
The variables Vcmax and Jmax are temperature dependent and their depen-dency is described by:
Parameter (Vcmax, Jmax, TPU) = exp(c – Ha/(RTl)) /(1+exp(( STl – Hd)/(RTl))) (6.9)
where c is a scaling factor, Ha (J/mol) the activation energy of the given parameter, R the gas constant (8.3143 J/°K/mol), Tl (°K) the leaf temperature,
S (J/mol) an entropy term and Hd (J/mol) the deactivation energy of the given parameter.
Similarly, the temperature dependency of Rd, , Kc and Ko is described by:
Parameter (Rd, , Kc, Ko) = exp(c – Ha/(RTl)) (6.10)
Proteins of the Calvin cycle and thylakoids represent the majority of leaf nitrogen (N). Therefore, photosynthetic capacity is strongly related to leaf N content expressed on an area basis (Na) (Field and Mooney, 1986; Evans, 1989; Kellomäki and Wang, 1997; Walcroft et al., 1997). To account for the relation-ship commonly observed between the parameters defi ning photosynthetic capacity (Vcmax, Jmax, TPU and Rd mainly) and Na (Field and Mooney, 1983; Harley et al., 1992) (Fig. 6.2), scaling factors c of Vcmax, Jmax, TPU and Rd may be related to Na, either linearly or slightly non-linearly.
In summary, leaf net photosynthesis depends on fi ve major classes of factors, either variables (external or internal factors) or parameters (more or less constant factors), provided that plants are not exposed to too extreme conditions; we may consider the internal factors as genetic factors. The fi ve classes of factors are:
The photosynthetically active photon fl ux density (1. Q), which is the major driving variable of photosynthesis. Gross photosynthesis is determined by Q while Ci determines the proportion of photorespiration, and thus net photosynthesis. One of the major environmental factors affecting Ci is water availability in the root zone through its effect on gs.
Leaf nitrogen concentration (2. Na), which is not a rate-determining factor of photosynthesis, unlike Q, but may be considered as a rate-limiting factor. In other words, Na sets the photosynthetic potential of a leaf (i.e. photosyn-thetic capacity). We shall see below which factors infl uence Na in mango leaves.
Leaf temperature infl uences leaf photosynthesis. Net photosynthesis is 3. positively correlated with leaf temperature in a normal range. Leaf tempera-ture (Tl) is not a driving variable of photosynthesis but it is the single most important rate-determining factor after Q. In addition, extreme temperatures may infl uence photosynthesis through their damaging effects. Kinetics of enzymes involved with photosynthetic reactions collectively comprise an additional set of factors that infl uence leaf net photosynthesis.
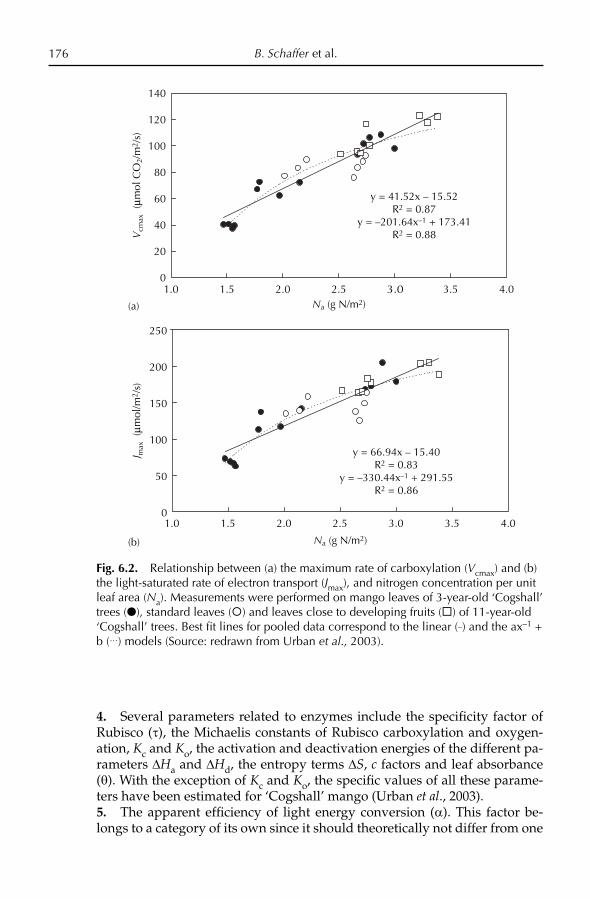
B. Schaffer et al.176
Several parameters related to enzymes include the specifi city factor of 4. Rubisco ( ), the Michaelis constants of Rubisco carboxylation and oxygen-ation, Kc and Ko, the activation and deactivation energies of the different pa-rameters Ha and Hd, the entropy terms S, c factors and leaf absorbance ( ). With the exception of Kc and Ko, the specifi c values of all these parame-ters have been estimated for ‘Cogshall’ mango (Urban et al., 2003).
The apparent effi ciency of light energy conversion (5. ). This factor be-longs to a category of its own since it should theoretically not differ from one
y = 41.52x – 15.52R2 = 0.87
y = –201.64x–1 + 173.41R2 = 0.88
0
20
40
60
80
100
120
140
(a)
(b)
1.0 1.5 2.0 2.5 3.0 3.5 4.0Na (g N/m2)
Vcm
ax (
μmol
CO
2/m
2 /s)
0
50
100
150
200
250
1.0 1.5 2.0 2.5 3.0 3.5 4.0
y = 66.94x – 15.40R2 = 0.83
y = –330.44x–1 + 291.55R2 = 0.86
J max
(μm
ol/m
2 /s)
Na (g N/m2)
Fig. 6.2. Relationship between (a) the maximum rate of carboxylation (Vcmax) and (b) the light-saturated rate of electron transport (Jmax), and nitrogen concentration per unit leaf area (Na). Measurements were performed on mango leaves of 3-year-old ‘Cogshall’ trees (●), standard leaves ( ) and leaves close to developing fruits ( ) of 11-year-old ‘Cogshall’ trees. Best fi t lines for pooled data correspond to the linear (_) and the ax–1 + b (…) models (Source: redrawn from Urban et al., 2003).

Ecophysiology 177
species to another and may be considered as a constant in the absence of photoinhibition and photodamage.
Light
Light exposurePlants allocate nitrogen resources within the canopy to enhance photosyn-thetic capacity at locations exposed to high incident light levels, thus maxi-mizing whole plant carbon gain (Field and Mooney, 1983; Hollinger, 1996; Carswell et al., 2000). For leaves of a given age and for a given nitrogen sup-ply, leaf N per unit leaf area appears to be strongly related with light expo-sure (DeJong and Doyle, 1985; Le Roux et al., 1999, 2001; Rosati et al., 1999, 2000). Photosynthetic light acclimation of leaves may result from changes in either leaf nitrogen concentration (Nm) or mass-to-area ratio (Ma) because Na = MaNm. Lynch and González (1993) observed a negative correlation between Nm and light exposure in the tropical fruit tree Borojoa patinoi, but such a behaviour is rare; positive correlations between Nm and light exposure are more commonly observed. In addition, photosynthetic light acclimation of leaves may result from changes in partitioning of total leaf N among the different pools of the photosynthetic machinery (Evans, 1989). In mango, light acclimation of photosynthesis results mainly from changes in Ma, and to a lesser extent from changes in allocation of total leaf N at low irradiance; whereas changes in Nm play only a minor role (Fig. 6.3). Light acclimation of mango leaves thus follows a pattern similar to peach leaves (Le Roux et al., 1999; Walcroft et al., 2002).
Light intensityPhotosynthesis of ‘Cogshall’ mango trees increases with increasing levels of light intensity to reach a maximum at Q = 1200 mol photons/m2/s (L. Urban, unpublished data). Whiley et al. (1999) measured Q at 1284 mol pho-tons/m2/s for fi eld-grown ‘Kensington Pride’ trees growing in subtropical Queensland, Australia, which is well below full sunlight (full sunlight ≥ 2000
mol photons/m2/s). Such a high threshold is a typical feature of sun plants. Individual leaves are rarely able to utilize full sunlight; whole trees consist of many leaves that shade each other, so that only a small fraction of a tree’s leaves are exposed to full sun at any given time of the day, while the rest of the leaves receive subsaturating photon fl uxes in the form of small patches of light that penetrate through gaps of the leaf canopy. Because the photosyn-thetic response of whole trees is the sum of the photosynthetic activity of all the leaves, only rarely is photosynthesis saturated with light at the whole-tree level.
While most leaves experience subsaturating light intensities, well-exposed leaves of the upper-crown may receive excessive quantities of light. Those leaves must dissipate the absorbed light energy in excess to prevent damage to the photosynthetic apparatus. Moderate decreases in maximal quantum effi ciency (i.e. quantum effi ciency of dark-adapted leaves Fv/FmPredawn) are
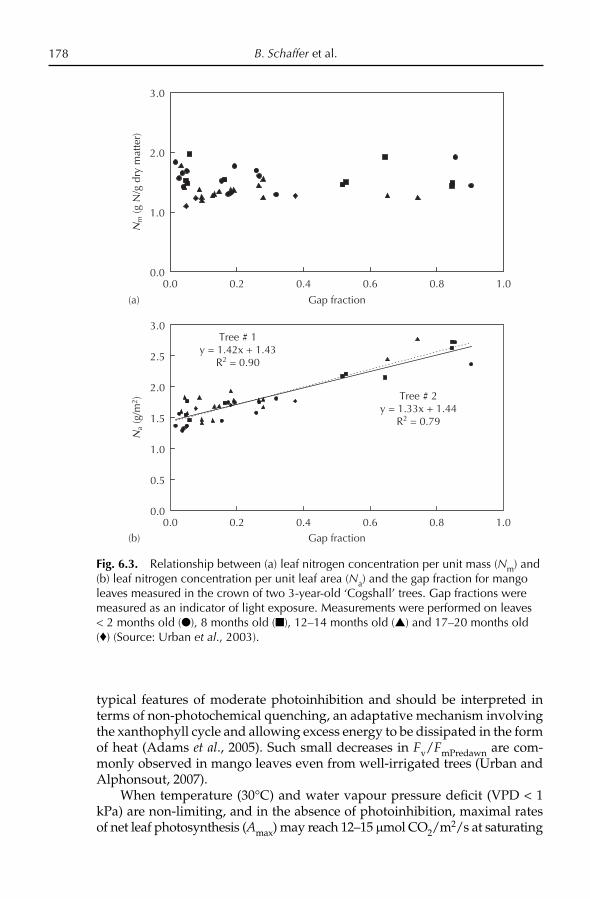
B. Schaffer et al.178
typical features of moderate photoinhibition and should be interpreted in terms of non-photochemical quenching, an adaptative mechanism involving the xanthophyll cycle and allowing excess energy to be dissipated in the form of heat (Adams et al., 2005). Such small decreases in Fv/FmPredawn are com-monly observed in mango leaves even from well-irrigated trees (Urban and Alphonsout, 2007).
When temperature (30°C) and water vapour pressure defi cit (VPD < 1 kPa) are non-limiting, and in the absence of photoinhibition, maximal rates of net leaf photosynthesis (Amax) may reach 12–15 mol CO2/m2/s at saturating
0.0
1.0
2.0
3.0
0.0 0.2 0.4 0.6 0.8 1.0
Gap fraction
Nm
(g N
/g d
ry m
atte
r)
Tree # 1y = 1.42x + 1.43
R2 = 0.90
0.0
0.5
1.0
1.5
2.0
2.5
3.0
0.0 0.2 0.4 0.6 0.8 1.0Gap fraction
Na
(g/m
2 ) Tree # 2y = 1.33x + 1.44
R2 = 0.79
(a)
(b)
Fig. 6.3. Relationship between (a) leaf nitrogen concentration per unit mass (Nm) and (b) leaf nitrogen concentration per unit leaf area (Na) and the gap fraction for mango leaves measured in the crown of two 3-year-old ‘Cogshall’ trees. Gap fractions were measured as an indicator of light exposure. Measurements were performed on leaves < 2 months old (●), 8 months old (■), 12–14 months old (▲) and 17–20 months old (♦) (Source: Urban et al., 2003).

Ecophysiology 179
Q, on ‘Cogshall’ trees. Whiley et al. (1999) measured Amax of 15.2 mol CO2/m2/s for ‘Kensington Pride’ trees growing in a subtropical climate in Queen-sland, Australia. However, values > 16 mol CO2/m2/s were observed on fi eld-grown trees of ‘Tommy Atkins’, ‘Haden’ and ‘Irwin’ on sunny days during the wet season in tropical regions of Australia (P. Lu, unpublished data). This is much higher than citrus (< 10 mol CO2/m2/s), but substan-tially lower than plum (approx. 26 mol CO2/m2/s). Whiley et al. (1999) esti-mated the light compensation point to be 29 mol photons/m2/s for leaves of non-stressed, fi eld-grown mango trees, which is much higher than that attributed to shade-tolerant species (< 10 mol photons/m2/s) (Harvey, 1979). The data show that mango trees are basically sun-adapted plants.
Leaf temperature
Effect of temperatures in a normal range on leaf photosynthetic capacityMedlyn et al. (2002) calculated optimal temperatures for Vcmax and Jmax to be 35–41°C and 30–38°C, respectively. Tree species native to cold climates had the lowest temperature optima for both Vcmax and Jmax. For ‘Cogshall’ mango trees, calculated temperature optima for Vcmax and Jmax are 44 and 45.5°C, respectively. They demonstrated that mango photosynthesis increases with temperature well above 40°C (Fig. 6.4). Although there is a clear lack of refer-ences for other tropical trees, it is tempting to attribute the temperature response of mango photosynthesis to its tropical origin.
Estimates of Ha, Hd and S are 7.0695, 17.0799 and 536 J/mol for Vcmax, and 3.8782, 10.2211 and 317 J/mol for Jmax, respectively. Both Hd and S are within the range of published values for Vcmax and Jmax (Dreyer et al., 2001). In addition, Ha for Vcmax is within the 60–80 kJ/mol range for many species, including crop species as well as deciduous and evergreen trees (Medlyn et al., 2002). For mango, Ha for Jmax is consistent with data published for evergreen species (Medlyn et al., 2002), which is consistent with the fact that mango leaves commonly are 2–4 years old before senescence and abscision. However, the Jmax/Vcmax at 25°C for mango is about 1.86, approximately 11% higher than the mean value calculated over the whole range of species stud-ied by Medlyn et al. (2002). Lower activation energies for Jmax than Vcmax result in a temperature-induced decrease in Jmax/Vcmax, confi rming previous observations by Walcroft et al. (1997) and Dreyer et al. (2001). The estimate of
Ha for Rd is 4.5710 J/mol. Ha is higher than the range of published values for Rd (Dreyer et al., 2001).
Chilling temperaturesFv/FmPredawn decreases in mango leaves with decreasing temperature, while chilling reduces quantum effi ciency (Whiley et al., 1999; Sukhvibul et al., 2000; Weng et al., 2006a, b). The decrease in Fv/FmPredawn may be interpreted to refl ect sustained engagement of zeaxanthin in photoprotective energy dis-sipation. Decreases in quantum effi ciency correspond to a decrease in the rate of electron fl ow. It may be argued that sustained zeaxanthin-dependent
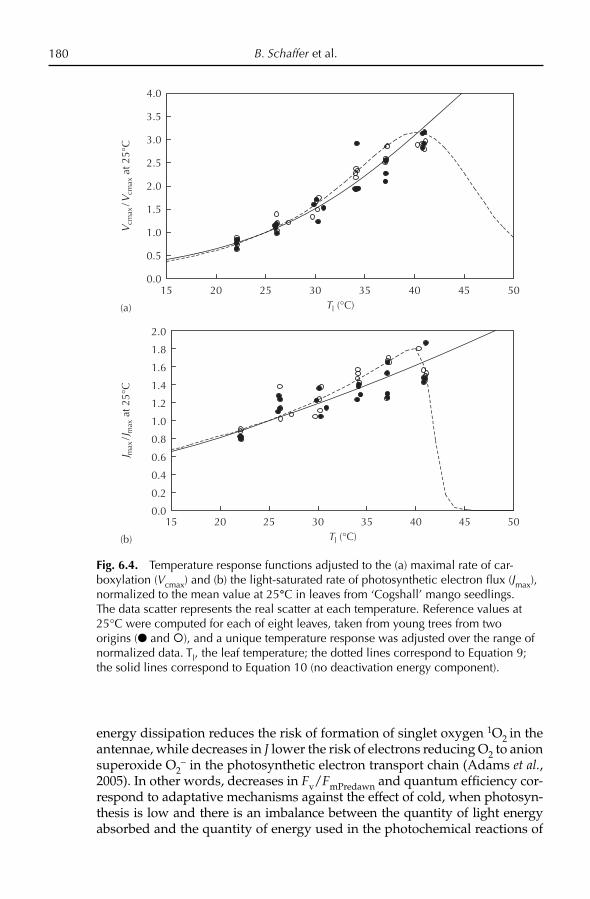
B. Schaffer et al.180
energy dissipation reduces the risk of formation of singlet oxygen 1O2 in the
antennae, while decreases in J lower the risk of electrons reducing O2 to anion superoxide O2
− in the photosynthetic electron transport chain (Adams et al., 2005). In other words, decreases in Fv/FmPredawn and quantum effi ciency cor-respond to adaptative mechanisms against the effect of cold, when photosyn-thesis is low and there is an imbalance between the quantity of light energy absorbed and the quantity of energy used in the photochemical reactions of
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
(a)
(b)
15 20 25 30 35 40 45 50
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
1.8
2.0
15 20 25 30 35 40 45 50Tl (°C)
J max
/Jm
ax a
t 25°
CV
cmax
/Vcm
ax a
t 25°
C
Tl (°C)
Fig. 6.4. Temperature response functions adjusted to the (a) maximal rate of car-boxylation (Vcmax) and (b) the light-saturated rate of photosynthetic electron fl ux (Jmax),normalized to the mean value at 25°C in leaves from ‘Cogshall’ mango seedlings. The data scatter represents the real scatter at each temperature. Reference values at 25°C were computed for each of eight leaves, taken from young trees from two origins (● and ), and a unique temperature response was adjusted over the range of normalized data. Tl, the leaf temperature; the dotted lines correspond to Equation 9; the solid lines correspond to Equation 10 (no deactivation energy component).

Ecophysiology 181
photosynthesis (Adams et al., 2005). Interestingly, Weng et al. (2006b) found that mango leaves transferred from warm and dark to chilling conditions showed only slight down-regulation of PSII effi ciency when compared to leaves moved from dim light to chilling conditions. Of course, long-term exposure to cold and very low temperatures (≤ 10°C) may eventually result in true photodamage, not just photoinhibition. Very low values of Fv/FmPredawn, decreases in chlorophyll content and slow recovery kinetics are all indicators of photodamage. Sukhvibul et al. (2000) observed that susceptibil-ity to cold-induced photodamage was more pronounced in polyembryonic cultivars than in monoembryonic cultivars, possibly refl ecting their different eco-evolutionary development.
Elevated atmospheric CO2 concentration
The CO2 concentration in the earth’s atmosphere has been increasing rapidly since the early 20th century and is continuing to rise, primarily due to burn-ing of fossil fuels (Houghton, 2005). Earth’s atmospheric CO2 concentration is currently about 370 mol CO2/mol (Houghton, 2005) and is projected to reach 600 mol CO2/mol by 2050. Elevated ambient CO2 levels will undoubt-edly affect cropping systems since atmospheric CO2 concentrations can sig-nifi cantly affect plant growth and productivity (Idso and Kimball, 1991; Houghton, 2005). There is little published information concerning the effects of elevated ambient CO2 levels on physiology, growth and production of tropical fruit trees, including mango.
Schaffer et al. (1997) exposed leaves of fi eld- and container-grown ‘Kens-ington’ (syn. ‘Kensington Pride’) trees to short durations (several minutes) of varying ambient CO2 concentrations. They found that under saturating light levels for photosynthesis, net photosynthesis increased as ambient CO2 con-centration increased up to 1200 mol CO2/mol. At ambient CO2 concentra-tions > 1200 mol CO2/mol, net photosynthesis stabilized, probably due to leaves reaching their maximum biochemical capacity to fi x carbon. Studies with ‘Cogshall’ mango trees indicated that when Ca increases, stomata close swiftly and Ci may become very unpredictable (L. Urban, unpublished data). Therefore, using Ci may be preferable to Ca for quantifying short-term effects of elevated CO2 concentations on Anet of mango. Saturating CO2 levels may often be reached at Ci = 800 mol CO2/mol air.
Long-term (6–12 months) exposure of ‘Kensington’ mango trees to an atmospheric CO2 concentration of 700 mol/mol resulted in higher net CO2 assimilation rates than in leaves of plants grown at atmospheric CO2 concen-trations of 350 mol/mol when net CO2 assimilation was measured at the same CO2 concentration as the growth environment. However, carboxylation effi ciency (the amount of CO2 fi xed per mole of ambient CO2) was lower for plants in the CO2-enriched environment compared to plants in the ambient (350 mol CO2/mol) environment (Schaffer et al., 1997). Although further studies are needed to determine the effects of long-term exposure to elevated CO2 concentrations on mango growth and productivity, it appears that

B. Schaffer et al.182
mango will benefi t from increases in atmospheric CO2 concentrations. How-ever, the effects of increased atmospheric CO2 concentrations associated with global warming on mango production may be offset by higher respiratory losses and increased assimilate partitioning to shoot growth in highly vegeta-tive cultivars (Schaffer et al., 1999). Therefore, responses of mango cultivars to elevated atmospheric CO2 concentrations need to be evaluated over a range of temperatures to ascertain their likely performance under changing atmospheric conditions.
Humidity
Although mango production occurs in the tropics and subtropics in areas of high and low relative humidity (RH) (Campbell, 1984), there are very few published reports on the effects of RH or VPD on physiology and tree growth.
In a study with container-grown ‘Kensington’ plants, Pongsomboon et al. (1992) reported that stomatal conductance was inversely correlated with VPD. Differences between cultivar responses to VPD have been observed in fi eld-grown ‘Irwin’ (monoembryonic) and ‘Kensington’ (polyembryonic) mango trees during the wet and dry season in tropical Australia. During the wet season and for well-irrigated trees during the dry season, both ‘Irwin’ and ‘Kensington’ showed decreasing stomatal conductance with increasing leaf-to-air vapour pressure defi cit (LAVPD) but ‘Kensington’ showed a more rapid decrease than ‘Irwin’ (Fig. 6.5) (P. Lu, unpublished data). It was also observed that daytime leaf xylem water potential was lower in ‘Irwin’ than in ‘Kensington’ while predawn water potentials were similar for both culti-vars (P. Lu, unpublished data). These results indicate that under similar soil water conditions, ‘Kensington’ tends to close stomata much more rapidly than ‘Irwin’ to conserve water under dry atmospheric conditions. This water conservation strategy is probably a refl ection of ‘Kensington’s’ adaptive responses to the hot and dry seasonal tropical environment under which it evolved (Wolstenholme and Whiley, 1995). Other studies in tropical Austra-lia revealed that polyembryonic ‘Nam Doc Mai’ behaved like ‘Irwin’ (P. Lu, unpublished data). However, ‘Nam Doc Mai’ in Thailand has comparatively low vigour compared to ‘Kensington’ when grown in the tropics.
Further research is required to determine if differences in photosynthetic or stomatal responses of mango to VPD are indeed based on embryonal char-acteristics. Clarifi cation of the reasons for variation would undoubtedly facil-itate breeding and selection of cultivars for dry and humid areas.
Flooding
The primary effect of fl ooding on plants is due to a reduction in soil oxygen concentration. Oxygen levels in the soil can decrease from 20% to < 5% within 1–2 days of fl ooding (Crane and Davies, 1988) and soils eventually become
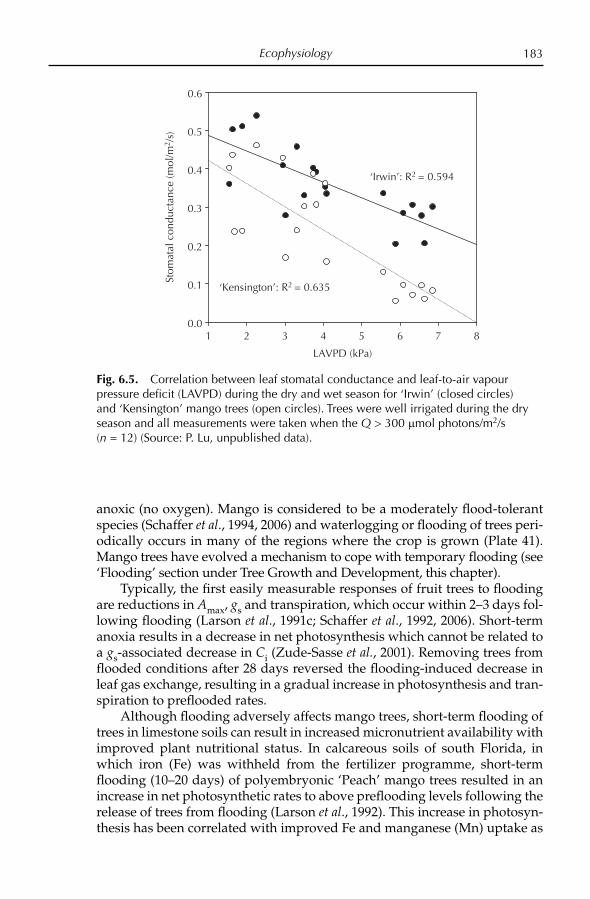
Ecophysiology 183
anoxic (no oxygen). Mango is considered to be a moderately fl ood-tolerant species (Schaffer et al., 1994, 2006) and waterlogging or fl ooding of trees peri-odically occurs in many of the regions where the crop is grown (Plate 41). Mango trees have evolved a mechanism to cope with temporary fl ooding (see ‘Flooding’ section under Tree Growth and Development, this chapter).
Typically, the fi rst easily measurable responses of fruit trees to fl ooding are reductions in Amax, gs and transpiration, which occur within 2–3 days fol-lowing fl ooding (Larson et al., 1991c; Schaffer et al., 1992, 2006). Short-term anoxia results in a decrease in net photosynthesis which cannot be related to a gs-associated decrease in Ci (Zude-Sasse et al., 2001). Removing trees from fl ooded conditions after 28 days reversed the fl ooding-induced decrease in leaf gas exchange, resulting in a gradual increase in photosynthesis and tran-spiration to prefl ooded rates.
Although fl ooding adversely affects mango trees, short-term fl ooding of trees in limestone soils can result in increased micronutrient availability with improved plant nutritional status. In calcareous soils of south Florida, in which iron (Fe) was withheld from the fertilizer programme, short-term fl ooding (10–20 days) of polyembryonic ‘Peach’ mango trees resulted in an increase in net photosynthetic rates to above prefl ooding levels following the release of trees from fl ooding (Larson et al., 1992). This increase in photosyn-thesis has been correlated with improved Fe and manganese (Mn) uptake as
1 2 3 4 5 6 7 80.0
0.1
0.2
0.3
0.4
0.5
0.6
‘Kensington’: R2 = 0.635
‘Irwin’: R2 = 0.594St
omat
al c
ondu
ctan
ce (m
ol/m
2 /s)
LAVPD (kPa)
Fig. 6.5. Correlation between leaf stomatal conductance and leaf-to-air vapour pressure defi cit (LAVPD) during the dry and wet season for ‘Irwin’ (closed circles) and ‘Kensington’ mango trees (open circles). Trees were well irrigated during the dry season and all measurements were taken when the Q > 300 mol photons/m2/s(n = 12) (Source: P. Lu, unpublished data).

B. Schaffer et al.184
a result of these elements becoming more soluble when calcareous soils are fl ooded (Larson et al., 1991b, 1992).
Internal factors
Leaf ageLeaf characteristics (i.e. photosynthetic capacity and the amount of N per unit area) are generally strongly infl uenced by leaf age, with maximum val-ues being observed when leaves have just completed full expansion (Con-stable and Rawson, 1980; Marshall and Biscoe, 1980; Dwyer and Stewart, 1986; Field, 1987; Wilson et al., 2000; Frak et al., 2001). Chlorophyll content is three to four times lower in young than in mature mango leaves (Zude and Ludders, 1997). Similarly, the concentration of Rubisco is lower in young than in mature, green leaves (Nii et al., 1995). In contrast to many other plant species, once mango leaves are mature the relationship between Na and irra-diance does not seem to be affected by leaf age (Urban et al., 2003). The Na values may remain high in old leaves experiencing high irradiance. This indicates that changes in Na in mango leaves are infl uenced by irradiance and not age, at least during the fi rst year.
Carbohydrate accumulation and source-sink balanceSource-sink imbalances can exert feedback down-regulation or repression of leaf photosynthesis through carbohydrate accumulation in leaves (Azcon-Bieto, 1983; Foyer, 1988; Koch, 1996; Schaffer et al., 1997; Whiley et al., 1999; Paul and Foyer, 2001; Paul and Pellny, 2003). Transient accumulations of car-bohydrates in leaves, as they have been observed during the diurnal period, may impair the rate of electron transport (Pammenter et al., 1993). Changes in photosynthetic capacity, not just assimilation rates, are more likely to be observed in association with lasting source-sink imbalances. One hypotheti-cal mechanism is that high levels of carbohydrates repress the expression of genes coding for several photosynthetic enzymes (Krapp and Stitt, 1995; Koch, 1996; Drake et al., 1997). Alternatively, carbohydrates may interact with hormonal signals to control gene expression (Thomas and Rodriguez, 1994). There is also some evidence that photosynthetic capacity is related to leaf carbohydrate status through the effect of the latter on phosphate availability (Riesmeier et al., 1993; Sun et al., 1999). In the long term, carbohydrate accu-mulation may eventually lead to cell death. High sugar concentration has been associated with senescence in leaves of several species (Noodén et al., 1997; Wingler et al., 1998; Quirino et al., 2001). Reduced energy utilization by CO2 assimilation, like the one resulting from carbohydrate accumulation, in combination with high energy capture is potentially dangerous and can result in over-reduction of the electron transport chain, photoinhibition and oxidative stress caused by photoreduction of oxygen to superoxide O2
− in the Mehler-ascorbate peroxidase reaction (Badger, 1985). Moreover, reactive sin-glet oxygen 1O2 can be formed through reaction of oxygen with triplet chlo-rophyll released by the breakdown of the chlorophyll-protein complexes in
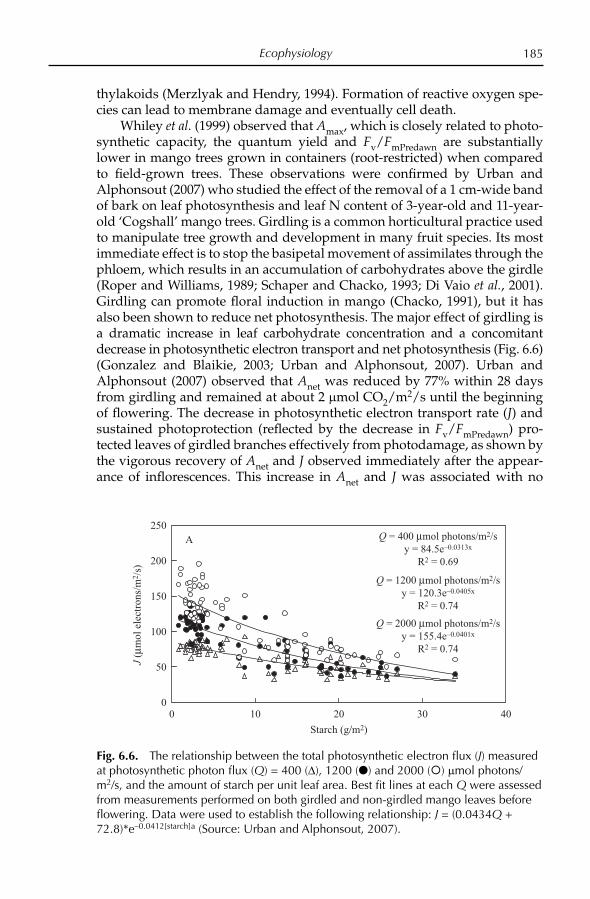
Ecophysiology 185
thylakoids (Merzlyak and Hendry, 1994). Formation of reactive oxygen spe-cies can lead to membrane damage and eventually cell death.
Whiley et al. (1999) observed that Amax, which is closely related to photo-synthetic capacity, the quantum yield and Fv/FmPredawn are substantially lower in mango trees grown in containers (root-restricted) when compared to fi eld-grown trees. These observations were confi rmed by Urban and Alphonsout (2007) who studied the effect of the removal of a 1 cm-wide band of bark on leaf photosynthesis and leaf N content of 3-year-old and 11-year-old ‘Cogshall’ mango trees. Girdling is a common horticultural practice used to manipulate tree growth and development in many fruit species. Its most immediate effect is to stop the basipetal movement of assimilates through the phloem, which results in an accumulation of carbohydrates above the girdle (Roper and Williams, 1989; Schaper and Chacko, 1993; Di Vaio et al., 2001). Girdling can promote fl oral induction in mango (Chacko, 1991), but it has also been shown to reduce net photosynthesis. The major effect of girdling is a dramatic increase in leaf carbohydrate concentration and a concomitant decrease in photosynthetic electron transport and net photosynthesis (Fig. 6.6) (Gonzalez and Blaikie, 2003; Urban and Alphonsout, 2007). Urban and Alphonsout (2007) observed that Anet was reduced by 77% within 28 days from girdling and remained at about 2 mol CO2/m2/s until the beginning of fl owering. The decrease in photosynthetic electron transport rate (J) and sustained photoprotection (refl ected by the decrease in Fv/FmPredawn) pro-tected leaves of girdled branches effectively from photodamage, as shown by the vigorous recovery of Anet and J observed immediately after the appear-ance of infl orescences. This increase in Anet and J was associated with no
Q = 2000 μmol photons/m2/sy = 155.4e–0.0401x
R2 = 0.74
0
50
100
150
200
250
0 10 20 30 40Starch (g/m2)
J(μ
mol
ele
ctro
ns/m
2 /s)
A
Q = 1200 μmol photons/m2/sy = 120.3e–0.0405x
R2 = 0.74
Q = 400 μmol photons/m2/sy = 84.5e–0.0313x
R2 = 0.69
Fig. 6.6. The relationship between the total photosynthetic electron fl ux (J) measured at photosynthetic photon fl ux (Q) = 400 ( ), 1200 (●) and 2000 ( ) mol photons/m2/s, and the amount of starch per unit leaf area. Best fi t lines at each Q were assessed from measurements performed on both girdled and non-girdled mango leaves before fl owering. Data were used to establish the following relationship: J = (0.0434Q + 72.8)*e–0.0412[starch]a (Source: Urban and Alphonsout, 2007).

B. Schaffer et al.186
decrease in leaf carbohydrate content during the fi rst month following the onset of fl owering, suggesting that the effect of carbohydrate accumulation on photosynthesis is mediated by sink activity. Apart from its negative effect on the carbon budget of mango trees, girdling appeared to be rather harm-less. However, leaf N concentration decreased, which indicates that there may indeed exist long-term negative effects of girdling on photosynthetic capacity. The width of bark (phloem) removed may be critical with respect to the intensity of the effect of girdling on the tree. Whiley et al. (2006) girdled the trunks of ‘B74’ (‘Calypso’™) mango trees in the Northern Territory of Australia in autumn (as soon as they had come out of the wet season). The girdles were no more than the thickness of a pruning saw (about 1 mm) and healed within 6 weeks. In the fi rst and third years after girdling, the trees had signifi cantly higher yields than non-girdled trees on which, coincidentally, fruit matured early, thus giving market advantage. There was no signifi cant difference in yield in the second year of treatment between girdled and non-girdled trees. The fi rst and third years had strong natural induction while the second year gave poor fl owering across all varieties in the district. Thus, dur-ing years of strong induction, this type of girdling most likely provided extra carbohydrate reserves to drive fl owering and support fruit set and retention while in the off-fl owering year there was suffi cient carbohydrate reserves to support reproductive activity. In contrast to observations with ‘Cogshall’ mango trees (Urban and Alphonsout, 2007), there was no evidence of long-term effects of narrow girdles on leaf N of ‘B74’ mango trees (Whiley et al., 2006). However, when wider girdles are made, tree recovery may take much longer leading to sustained physiological disruption.
Proximity of infl orescencesWhile the effects of water stress and high light, temperature and atmospheric CO2 concentration on photosynthesis are increasingly well described, very little is known about the effect of phenology, and especially of fl owering on photosynthesis of mango. There is some evidence that fl owering may have an effect on photosynthesis. Flowering-associated decreases in Anet and gs were observed in sweet cherry (Roper et al., 1988) and mango (Shivashankara and Mahai, 2000; Urban et al., 2004a). Lack of precise knowledge about the effect of fl owering on photosynthesis may impair our ability to adequately simulate photosynthesis, especially for tropical fruit trees for which fl ower-ing often extends over a long period of time. Mango fl owering can last for > 2 months. Therefore, its effect on photosynthesis should not be overlooked. Urban et al. (2004a) showed that the decrease in Anet in mango leaves close to infl orescences is not attributable to a gs-associated decrease in Ci or to an increase in Rd. Rd was lower in leaves close to infl orescences than in standard leaves. If any, the effect of Rd on Anet was a positive one. This study suggested strongly that the decrease in Anet was due to a decrease in the electron fl ow in photosystem II, but failed to provide direct evidence for it as well as the ele-ments for interpretation. Using a modelling approach, Urban et al. (2008) confi rmed that there is a decrease in the total light-driven photosynthetic electron fl ux in leaves close to infl orescences and showed that the decrease

Ecophysiology 187
in Anet is also attributable to an increase in photorespiration. The latter appears to be the consequence of a gm-associated decrease in Cc, while the former results from an increase in electron fl ow towards alternative sinks, a decrease in the amount of leaf N per unit leaf area, and, hypothetically, either a decrease in leaf N allocation to the bioenergetic pool of the photosynthetic machinery, inorganic phosphorus depletion in leaves, or feedback inhibition of photosynthesis. The latter hypothesis is least probable in the absence of carbohydrate accumulation in leaves close to infl orescences. Both of these hypotheses need to be tested to further our understanding of the inhibiting effect of infl orescences on photosynthesis of nearby leaves. Interestingly, net photosynthesis measured on leaves close to panicles bearing set fruits are intermediary between those measured on standard leaves and those mea-sured on leaves close to infl orescences, suggesting that changes in photosyn-thesis associated with fl owering are reversible. Urban et al. (2008) also showed that processes other than temperature or light acclimation, and acclimation to reduced sink activity, may cause leaf N concentration and photosynthetic capacity to vary in mango. Whiley (unpublished data) observed that the mango leaves immediately adjacent to infl orescences lose colour intensity as the infl orescence grows out. Although leaf N over this period was not mea-sured in mango, it has been measured in avocado (Whiley, 1994) which has a similar intense burst of fl owering in which a large biomass is produced in a short time. Leaf N declines rapidly in avocado leaves as the infl orescences break from buds and grow and then N stabilizes (at a lower concentration) by mid-bloom. This can be reversed by N applications during fl owering and the additional application of a growth retardant (paclobutrazol (PBZ)) giving leaf N and A a signifi cant boost. Similarly to avocado, it is likely that the reduction in A close to mango infl oresences is related to reduced leaf N.
Photosynthetic contributions by fruit
Fruit of many species have chlorophyll and photosynthetic activity, particu-larly during the early stages of growth (Jones, 1981; Whiley et al., 1991). How-ever, for most crops, respiratory losses from fruit exceed photosynthetic gains throughout ontogeny (Kriedemann, 1968; Whiley et al., 1991). An exception to this is blueberry (Vaccinium spp.) fruit in which there is a net photosyn-thetic gain from petal fall through to colour break, with an estimated 15% of the total fruit carbon requirement contributed from fruit photosynthesis (Birkhold et al., 1992). Studies with monoembryonic ‘Dashehari’ mangoes showed that when fruit were approximately 10 mm in diameter, the photo-synthetic rate of fruit was 2.7% that of leaves and declined to 1.2% of leaf photosynthesis at fruit maturity (Chauhan and Pandey, 1984). However, even this comparatively small carbon contribution may be important during the critical fruit set period when trees rely on stored carbohydrates and a rela-tively ineffi cient canopy to supply current photosynthates. Further studies with mangoes to establish optimum light regimes for fruit photosynthesis at different stages of ontogeny are warranted.

B. Schaffer et al.188
6.3 Plant Water Relations
In this section, theoretical concepts of plant water relations are briefl y out-lined to help interpret the effects of environmental factors on mango water relations (see also Nobel (1983) and Baker (1984)).
An important concept in plant water relations is water potential (Ψ), which is a measure of the free energy of water. For pure water, Ψ = 0. As sol-utes are added to water, its free energy decreases and becomes more nega-tive. Water moves along a gradient from higher to lower (more negative) .
can be expressed as:
= + p + m (6.11)
where is osmotic (or solute) potential which refers to the effect of solutes on the change in free energy of water; p is the hydrostatic or pressure poten-tial also referred to as the turgor pressure; and m is the matric potential, which is generally negligible in plant cells.
In plant cells, p is generally positive or equal to 0. However, in xylem tissue of transpiring plants, p is negative (under tension). The driving force for transpiration is the vapour pressure difference between the leaf (consid-ered to be water saturated) and the surrounding air. Thus, water moves from a greater to a lower (or more negative) p and hence along a decreasing gradient. The cohesive forces of the H2O molecules allows the xylem water to remain in a continuous column even though there is a negative p.
Plant water stress can be determined from Eqn 6.11 and from changes in . The components of , such as and p, can often be used to defi ne the
sources of water stress. The drought tolerance of mango highlights some unique aspects of physiology of this tree with respect to its water manage-ment. Typical mango environments in the tropics impose extreme water stress and high evaporative demand for prolonged periods. Adaptive strate-gies of mango trees include a deep root system (Sukonthasing et al., 1991), desiccation-tolerant surface feeder roots and drought avoidance mechanisms thought to be mediated by a comprehensive system of resin canals distrib-uted throughout the tree (Venning, 1948; Joel, 1980; Joel and Fahn, 1980a, b; Pongsomboon, 1991) and rapid stomatal closure. Plants with laticfers or resin ducts/canals have been reported to be drought tolerant due to extended maintenance of turgor following the withdrawal of water (Downton, 1981; Kramer, 1983; Kallarackal et al., 1990). While the mechanism of turgor main-tenance remains unresolved, it is believed that the latex or resin is probably involved in the modulation of plant water status (Kallarackal et al., 1990). The differentiation, structure and distribution of resin canals in mango has been described by Venning (1948), Joel (1980) and Joel and Fahn (1980a, b, c). Resin canals are present in trunks, shoots, leaves and fruit (exocarp) of mango in close association with the vascular tissues. The resin contains mainly ter-penes, but phenols and protein-carbohydrate mucilage are also present (Joel and Fahn, 1980c). In well-watered trees, the resin is under positive pressure and freely exudes from damaged or cut surfaces (Pongsomboon, 1991). In studies of the development of water defi cit in container-grown ‘Kensington’

Ecophysiology 189
mango trees, loss of turgor occurred in expanding leaves when leaf water potential ( l) reached –1.2 Mpa. In mature leaves turgor was not lost until l reached –1.75 MPa (Pongsomboon, 1991). Necrotic leaf areas appeared when
l reached approximately –3.2 MPa with permanent wilting developing at −3.45 MPa. This is high (less negative) compared with a l of –6.6, –5.0 MPa for orange (Citrus sinensis) and macadamia (Macadamia integrifolia), respec-tively (Fereres et al., 1979; Stephenson et al., 1989). Thus, mango leaves toler-ate less internal water stress than woody perennial fruit trees from more mesic environments. With mango, however, the permanent wilting point was reached 36 days after withholding water compared to 10 days for simi-larly sized macadamia trees (Stephenson et al., 1989). The higher critical threshold of l and the longer period of survival for ‘Kensington’ mango indicates that drought tolerance is based on more effective water regulation to prevent desiccation and on maintenance of leaf turgor rather than resis-tance by tissues to damage.
Reich and Borchert (1988) observed that stomatal regulation in mango signifi cantly reduced the rate of development of internal water defi cit when compared with four other tropical tree species. In water-withholding studies, the radial expansion of mango trunks continued when most of the other species were shrinking, indicating that mango trees could better tolerate drought conditions and maintain photoassimilation rates. This is consistent with the decrease in gs/Anet observed by Urban et al. (2006) as a consequence of drought (Fig. 6.1). A decrease in gs/Anet indicates that there is an increase in photosynthetic water use effi ciency.
In containerized ‘Kensington’ mango trees, there was a linear correlation between stomatal conductance and l during the development of water stress (Pongsomboon, 1991). In contrast, with avocado and macadamia, stomatal response was much more rapid with a curvilinear relationship between l and stomatal conductance, and stomatal closure reached at –1.2 and 3.0 MPa, respectively. The slower response of stomatal conductance to
l in mango trees appears to be related to a more effective mechanism for
the mediation of water defi cit development compared with avocado and macadamia.
Pongsomboon (1991) monitored leaf water potential, osmotic potential of resin ( r) and osmotic potential of the whole leaf tissue ( ) in container-grown ‘Kensington’ mango trees when water was withheld for a 45-day period. When tissues were fully hydrated, l and r were higher than . For 40 days of the drying cycle, l and r declined at a similar rate; however, declined to about –1.2 MPa within 4 days where it remained stable until 18 days into the drying cycle. There was a subsequent decline in for 28 days after withholding water when it stabilized at –2.0 MPa, remaining constant for another 12 days. Pongsomboon (1991) suggested that osmotic adjustment occurred, probably mediated through the resin as water defi cit developed. It appears, therefore, that the energy investment by the tree in a resin canal sys-tem is justifi ed by the vital drought-avoidance benefi ts conferred by main-taining turgor and preventing wilting under prolonged periods of water stress. Further investigations are required to substantiate these conclusions.

B. Schaffer et al.190
In a recent experiment of 2-year-old potted ‘Cogshall’ mango trees grafted on ‘Maison Rouge’ rootstock, midday minimum leaf water potential remained relatively constant, about –1.0 MPa, for the range of predawn leaf water potential (a surrogate of soil water potential) from 0 to –0.5 MPa, but when the predawn leaf water potential dropped below –0.5 MPa, leaf water potential fell rapidly to about –1.8 MPa (Gaelle Damour, Centre de coopéra-tion Internationale en Recherche Agronomique pour le Développement (CIRAD), personal communication). This ‘isohydric’ behaviour seemed to be associated with rapid stomatal closure in response to decrease in water avail-ability (Urban and Jannoyer, 2004).
Development of novel techniques to study xylem integrity have pro-vided some unique insights into stomatal function (Tyree and Sperry, 1988; Cochard et al., 2002). Xylem sap is transported under negative pressures in plants and therefore is susceptible to cavitation events that render xylem conduits non-conductive. Cavitation occurs when the negative sap pressure exceeds a threshold value defi ned by anatomical characteristics (Tyree and Sperry, 1988). Many species function very close to the point of embolism. Therefore, stomata control both plant water losses and sap pressure and thus may actively control the risk of xylem embolism (Jones and Sutherland, 1991). Xylem vulnerability to cavitation was studied in twigs of 13-year-old ‘Cogshall’ mango trees. Xylem vessels started to cavitate at a xylem water potential of about of –1.5 MPa and underwent a rapid and substantial loss (90%) of hydraulic conductivity when xylem water potential dropped from –2.0 to –3.0 MPa (Fig. 6.7) (H. Cochard, unpublished data). This result is con-sistent with the observation that leaves lose turgor when l reaches –1.75 MPa and permanent wilting appears when l is above –3.0 MPa (Pongsom-boon, 1991) when Fig. 6.7 predicts 90% of xylem hydraulic conductivity is lost. It also confi rms that mango, like many other trees, operates close to the point of embolism via effective stomatal control.
Direct measurement of xylem water potential using a pressure chamber is problematic due to the presence of latex (Castro Neto et al., 2004). It is more problematic in some cultivars, for example ‘Kensington’, than in others, such as ‘Irwin’. Alternative indicators of plant water status such as measurements of whole-tree water use (sap fl ow) and stem/branch shrinkage were found to be sensitive and integrated indicators of whole-tree water status in mango and were successfully used to schedule irrigation in a mango orchard in the seasonal dry-wet tropical region of northern Australia (Lu, 2002, 2006).
6.4 Tree Growth and Development
Light
In an orchard, light distribution within and between tree canopies can have a profound effect on growth and development of the fruit. We have previ-ously discussed the effect of light on photosynthesis and defi ned the opti-mum light levels required for mango leaves. When light levels fall below the
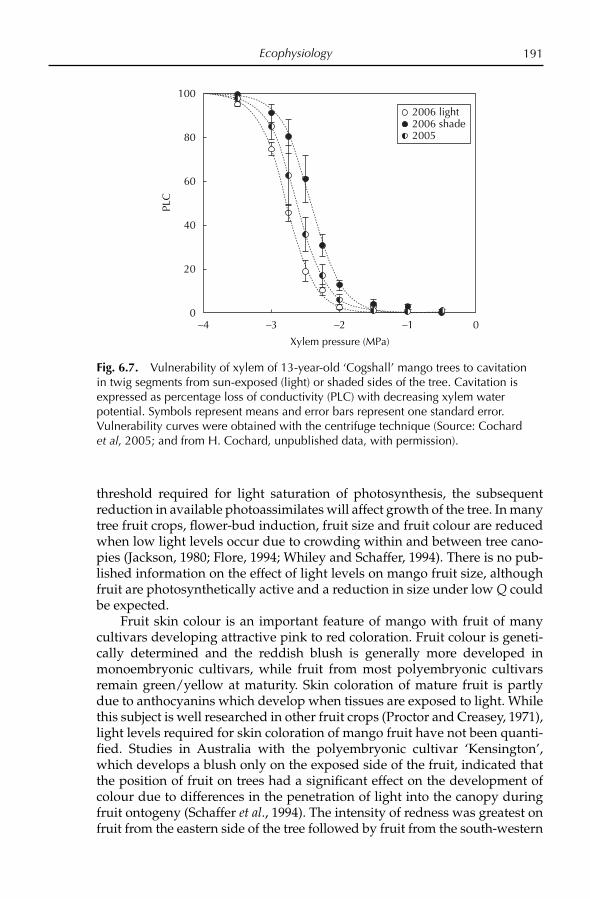
Ecophysiology 191
threshold required for light saturation of photosynthesis, the subsequent reduction in available photoassimilates will affect growth of the tree. In many tree fruit crops, fl ower-bud induction, fruit size and fruit colour are reduced when low light levels occur due to crowding within and between tree cano-pies (Jackson, 1980; Flore, 1994; Whiley and Schaffer, 1994). There is no pub-lished information on the effect of light levels on mango fruit size, although fruit are photosynthetically active and a reduction in size under low Q could be expected.
Fruit skin colour is an important feature of mango with fruit of many cultivars developing attractive pink to red coloration. Fruit colour is geneti-cally determined and the reddish blush is generally more developed in monoembryonic cultivars, while fruit from most polyembryonic cultivars remain green/yellow at maturity. Skin coloration of mature fruit is partly due to anthocyanins which develop when tissues are exposed to light. While this subject is well researched in other fruit crops (Proctor and Creasey, 1971), light levels required for skin coloration of mango fruit have not been quanti-fi ed. Studies in Australia with the polyembryonic cultivar ‘Kensington’, which develops a blush only on the exposed side of the fruit, indicated that the position of fruit on trees had a signifi cant effect on the development of colour due to differences in the penetration of light into the canopy during fruit ontogeny (Schaffer et al., 1994). The intensity of redness was greatest on fruit from the eastern side of the tree followed by fruit from the south-western
100
80
60
40
PLC
20
0–4 –3 –2
Xylem pressure (MPa)
–1 0
2006 light2006 shade2005
Fig. 6.7. Vulnerability of xylem of 13-year-old ‘Cogshall’ mango trees to cavitation in twig segments from sun-exposed (light) or shaded sides of the tree. Cavitation is expressed as percentage loss of conductivity (PLC) with decreasing xylem water potential. Symbols represent means and error bars represent one standard error. Vulnerability curves were obtained with the centrifuge technique (Source: Cochard et al, 2005; and from H. Cochard, unpublished data, with permission).

B. Schaffer et al.192
and northern sides of the tree. This information establishes an important con-cept with respect to the light regime but does not quantify the absolute light levels required for anthocyanin development. Further research is necessary to establish physiological parameters from which pruning and orchard man-agement strategies can be developed.
Temperature
Mango is a predominantly tropical species although the tree will usually grow and produce more successfully in frost-free subtropical latitudes with a marked dry season and high heat accumulation. Under optimum tempera-tures with non-limiting nutrients and water, the tree remains vegetative with growth fl ushes occurring at regular intervals. The large size and poor crop-ping of trees in the humid lowland tropics are well known, and there is a direct relationship between temperature and the frequency of vegetative fl ushes. Trees grown at 20°C days/15°C nights (20/15°C) required 20 weeks (mean of ten cultivars) to complete a growth/dormancy cycle while at 30/25°C the same cycle was completed in 6 weeks (Whiley et al., 1989). There are marked differences between cultivars with respect to their tendency towards vegetative growth. For instance, in controlled temperature studies over a 20-week period, at 30/25°C, ‘Irwin’ produced 2.0 growth fl ushes with approximately 45 days of dormancy between active growth periods while ‘Kensington’ produced 4.7 growth fl ushes with only 5 days of quiescence between fl ushes (Whiley et al., 1989). Dry matter accumulation over the 20 weeks was similar for the two cultivars; however, starch accumulation in the woody trunk tissues of ‘Irwin’ and ‘Kensington’ was 13 and 3.6% of dry mat-ter, respectively. The response differences between these two cultivars may be the contributing factor to their performance at tropical latitudes where temperature is non-limiting for growth. Under these conditions ‘Irwin’ has more reliable cropping than ‘Kensington’, suggesting that the genetically determined low-vigour trait is more sensitive to environmentally precipi-tated stresses that induce fl owering.
The number and size of leaves which develop on each growth fl ush are also infl uenced by temperature. Whiley et al. (1989) reported that on trees growing at 20/15°C an average of 7.1 leaves per fl ush were produced while at 30/25°C there were 13.6 leaves on each growth fl ush (data are mean values from ten cultivars). At 30/25°C the mean leaf size was 300% greater than those on trees growing at 20/15°C. Soil temperatures have also been reported to have a strong effect on the growth of mangoes. In studies with ‘Irwin’ grafted on ‘Turpentine’ rootstocks, episodic shoot growth occurred when soil temperatures were held at 27°C or 32°C for 120 days but an extended dormant period developed when soil temperatures were held at 21°C (Yusof et al., 1969). These results indicate that environmental control over shoot growth of mangoes may in part be related to soil temperatures.
From controlled temperature studies it has been calculated that the median daily temperature (mean of the maximum and minimum daily temperatures)

Ecophysiology 193
at which shoot growth ceases is approximately 15°C (mean value for ten cul-tivars) (Whiley et al., 1989). Subsequent studies (Issarakraisila et al., 1991) have confi rmed that 15°C is the critical minimum growth temperature for shoots of ‘Kensington’.
Stress-inducing temperatures which prevent shoot growth have been shown to promote fl oral induction in mangoes, but this is outside the scope of this discussion. For further information of the effects of temperature on pollination, fl oral initiation and fruit development, see Davenport, Chapter 5, this volume and Schaffer et al. (1994). We again emphasize that although mango is a ‘heat-loving’ crop well adapted to the hot, semi-arid subtropics and monsoonal tropics, in these environments it experiences extremes of heat, drought and evaporative demand that may cumulatively reduce potential production capacity.
Drought
Although mango is considered to be drought tolerant and may survive with-out rain or irrigation for > 8 months (Gandhi, 1955), water defi cits during the reproductive cycle can have severe effects on the retention and early growth of mango fruit. In studies with bearing, container-grown ‘Irwin’ trees, pre-dawn l levels were maintained at either less than –0.3 MPa (non-stressed) or –1.2 MPa (water stressed) for the fi rst 2 months after fruit set. For the fi rst 5 days following fruit set, all trees lost a similar percentage of fruit, but there-after fruit abscission was greater on water-stressed trees. After 1 month, drought-stressed trees had retained approximately 4% of their initial fruit set compared with approximately 8% on non-stressed trees. During the fi rst 30 days following fruit set, the rate of fruit growth for non-stressed trees was twice that of drought-stressed trees, and fi nal fruit size (measured 60 days after fruit set) of non-stressed trees was 20% greater than on water-stressed trees. In a separate study, another group of ‘Irwin’ trees was maintained stress-free (pre-dawn water potential above –0.3 MPa) during the fi rst month following fruit set with water-stress (–1.2 MPa) imposed on some trees dur-ing the second month of fruit development (Pongsomboon, 1991). There was no effect of drought on fruit retention but fi nal fruit size was 34% smaller for stressed compared with non-stressed trees.
In fi eld studies, Singh and Arora (1965) compared fruit drop of monoem-bryonic ‘Dashehari’ trees irrigated at 1-week intervals with trees irrigated at 3-week intervals. During the fi rst 6 weeks of fruit growth, weekly irrigation reduced fruit drop compared with the irrigation at 3-week intervals. During the latter stages of fruit development, these gains were lost as more fruit dropped from the weekly irrigated trees. In another study, fi eld-grown monoembryonic ‘Tommy Atkins’ trees were managed under different irriga-tion regimes from early fruit set until the start of the rainy season (approxi-mately 43 days) (Larson et al., 1989). Trees were irrigated on a 7- or 14-day schedule or received no irrigation. Pre-dawn l was –0.3 MPa for trees irri-gated on the 7-day schedule and decreased to –0.5 MPa for the non-irrigated

B. Schaffer et al.194
trees. Irrigation at 7-day intervals resulted in the greatest yield with the larg-est fruit, especially during the early harvest period (Larson et al., 1989).
Irrigation, therefore, particularly during the fi rst 4–6 weeks following fruit set, increases individual fruit size and yield. This is a critical period of fruit development since it is when cell division is most rapid and cell walls are developed. Even slight reductions in plant water status during this period can have adverse effects on fruit growth and retention (Pongsomboon, 1991). Although drought tolerance of the mango tree is well known, this comes at considerable cost to tree performance, particularly in areas with prolonged dry seasons that extend through fl owering and fruiting. Irrigation is there-fore one of the most powerful tools to alleviate non-lethal yet potentially yield-reducing drought stress.
Flooding
Studies with container-grown mango trees have reported variable responses with respect to tree survival. Larson et al. (1991c) observed that as many as 45% of trees died after their roots were submerged in water for 4–10 days, but in the surviving population no further mortality occurred when fl ooding was extended for up to 110 days. In other experiments, there was no tree mortality after container-grown mango trees were fl ooded from 1 to several months although tree growth was signifi cantly reduced (Larson et al., 1991c; B. Schaffer unpublished data, Homestead, Florida, 1993).
The ability of mango trees to survive prolonged fl ooding appears to be dependent on the development of hypertrophic (swollen) stem lenticels immediately above the water line (Plate 42). The initial stages of lenticel hypertrophy are characterized by the development of intercellular spaces in the phellem tissue and production of additional phellem tissue by increased phellogen activity. Later stages of hypertrophy are characterized by the development of intercellular spaces in the phellem tissue and cortex (Larson et al., 1991a). Observations vary among studies whether or not trees devel-oped hypertrophic lenticels or how quickly after fl ooding they formed. These anomalies have been attributed to environmental differences at the time of root submersion (Larson et al., 1991c). In trees that died as a result of fl ooding stress there was no lenticel hypertrophy; however, stem lenticels hypertro-phied within 4–10 days on mango trees that survived fl ooding (Larson et al., 1991a, c, 1993). Sealing hypertrophic lenticels of mango trees with silicone grease or petroleum jelly resulted in tree death within 3 days of fl ooding, thereby demonstrating their necessity for tree survival. The role of hypertro-phic lenticels in fl ood-tolerant species is not clear, although they are thought to eliminate potentially toxic metabolites such as ethanol, acetaldehyde and ethylene which result from anaerobic respiration in the roots (Chirkova and Gutman, 1972; Larson et al., 1993). They may also confer fl ood tolerance by enhancing O2 diffusion to the roots (Kozlowski, 1984).
In some instances, adventitious roots have developed above the water line when container-grown mango trees have been fl ooded for long periods

Ecophysiology 195
(Schaffer et al., 1994). It is likely that these roots facilitate the absorption and translocation of O2 to submerged roots and their development is a common morphological response of many woody plants to root anoxia. The develop-ment of adventitious roots has not been reported for fl ooded, fi eld-grown trees and they may only form on young trunks after extended fl ooding peri-ods, which usually do not occur under normal production conditions.
Vegetative growth of mango trees generally declines when trees become fl ooded for > 2–3 days. When trees in a limestone soil in containers were fl ooded for > 110 days, there was a 94% reduction in shoot extension growth, while fl ooding for approximately 10 days resulted in a 57% reduction in shoot extension growth (Larson et al., 1991c). In a subsequent study, the stem radial growth (a more sensitive indicator of tree growth than shoot extension growth) of mango trees decreased 2 weeks after roots were submerged. Flooding for > 14 days also signifi cantly reduced root dry weight, resulting in an increased shoot to root ratio (Larson et al., 1991c). These adverse effects of fl ooding on the growth of mango trees are expected as reduced net photo-synthesis and presumably higher root respiration limit the availability of carbon-based assimilates required for growth.
Wind
Most fruit trees benefi t from wind protection, particularly during the estab-lishment years when the disruption of physiological processes results in a signifi cant depression of growth in young trees. In addition, wind also causes abrasions to the skin of fruits, particularly when they are small, which develop into unsightly blemishes by the time they are fully grown thereby reducing quality and market value. However, the cost of windbreaks may not be offset by higher returns. In some mango-producing regions, winds are not suffi ciently strong to justify the cost of wind protection. Until recently, wind protection in South Africa was not recommended for mangoes due to the loss of potential cropping space by ‘living’ windbreaks, their potential to create frost pockets, and the likelihood of promoting the incidence of fl ower and fruit diseases through increased humidity (Van der Meulen et al., 1971); however, the value of windbreaks is well appreciated today in South Africa (B.N Wolstenholme, personal communication, Pietermaritzburg, South Africa, 1995).
In studies with ‘Kensington’ mangoes in Australia, where artifi cial wind-breaks were constructed to shelter trees from the prevailing summer south-easterly winds, a 600% increase in yield was recorded in the fi rst year following wind protection (Mayers et al., 1984). This signifi cant improvement in tree performance was a result of better growth of trees which set and held more fruit per panicle, suffered less damage to leaves (cuticle fracturing) and had reduced fruit loss from bacterial black spot (caused by Xanthomonas camp-estris pv. mangiferaeindicae) compared to the wind-exposed trees. These results indicate that wind can have a signifi cant effect on mango productivity from the reduction of both growth and yield through undisclosed physiological

B. Schaffer et al.196
mechanisms, and a decreased level of bacterial black spot infection. The pro-vision of windbreaks in orchards is expensive with decisions to be made on the use of either ‘living’ or artifi cial shelters. Requirements for wind protec-tion will vary depending on site circumstances, and all factors pertaining to crop performance will require careful consideration.
Salinity
Salt stress in mango trees produces symptoms similar to those described for other species (Harding et al., 1958; Ehlig, 1960; Kadman, 1964; Bingham et al., 1968). Mild symptoms of chloride toxicity are scorched leaf tips and margins and leaf curling, while in more severe cases growth ceases, leaves abscise and trees die. Necrotic areas develop on leaves of trees exposed to high sodium levels (Jindal et al., 1976; Kadman et al., 1976; Gazit and Kadman, 1980). Irri-gation water concentrations of 20–60 mM sodium chloride (NaCl) or sodium sulfate (Na2SO4) reduced leaf area and changed the branching structure of container-grown mango trees, suggesting that salinity resulted in reduced leaf cell elongation, and affected the activity of the terminal meristem (Schmutz and Lüdders, 1993). As the duration of the exposure to saline con-ditions increased, transpiration decreased exponentially (Schmutz and Lüd-ders, 1993). In a later study, Schmutz (2000) found that following a gradual increase in salinity of the nutrient solution applied to potted polyembryonic ‘13-1’ rootstocks (from 0 to 120 mM NaCl over 15 days), Amax signifi cantly declined despite there being no visible leaf symptoms of salt toxicity. The decline in Amax occurred within 6 days of beginning the salinity treatment, which was 15 mM NaCl for 3 days followed by 30 mM NaCl for 3 days. This indicates that photoassimilation in mango is quite sensitive to exposure of trees to salinity.
There is considerable variation in salinity stress of mango, both within and between populations of mono- or polyembryonic mango ecotypes. Based on the results of limited studies, there appears to be greater salt tolerance in polyembryonic than in monoembryonic populations (Jindal et al., 1975; Kad-man et al., 1976). In seedling populations from mono- and polyembryonic cultivars irrigated for 2 years with water containing approximately 10 mM chloride, most plants developed leaf scorching after 6 months which gradu-ally became more severe, culminating in degeneration and death. However, some seedlings which had no damage or only slight toxicity symptoms were mostly of the polyembryonic ‘13-1’ rootstock cutivar or related types (Kad-man et al., 1976). Leaf analyses revealed that the chloride concentration in tolerant seedlings (0.68–0.77%) was greater than in susceptible seedlings (0.43–0.55%). In addition, tolerant plants had lower leaf concentrations of potassium, calcium and magnesium than saline-sensitive seedlings, possibly a result of comparative nutrient dilution since vegetative growth was greater for saline-tolerant than for saline-sensitive seedlings. Kadman et al. (1976) also suggested that the mechanism of chloride tolerance in ‘13-1’ was based on greater physiological tolerance of chloride concentrations in leaf tissues,
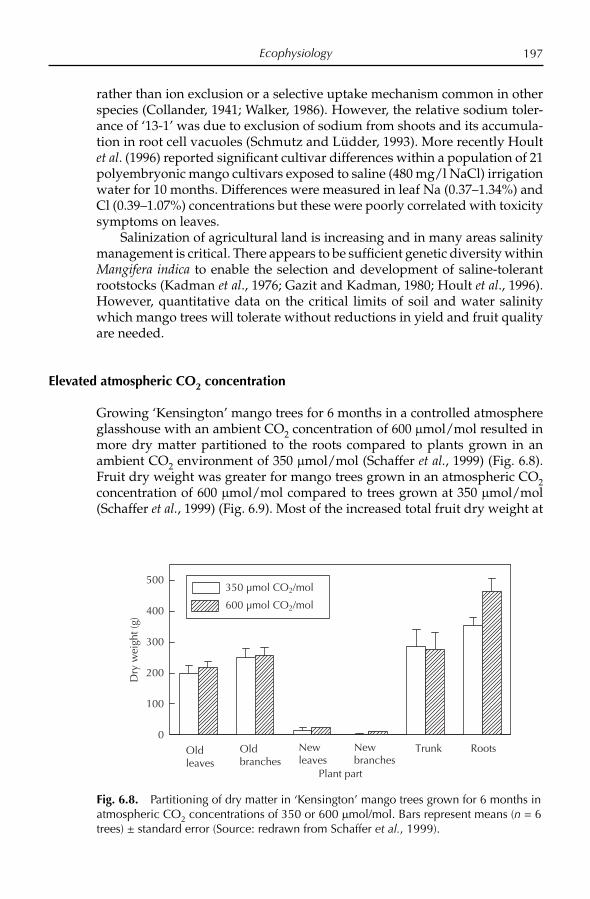
Ecophysiology 197
rather than ion exclusion or a selective uptake mechanism common in other species (Collander, 1941; Walker, 1986). However, the relative sodium toler-ance of ‘13-1’ was due to exclusion of sodium from shoots and its accumula-tion in root cell vacuoles (Schmutz and Lüdder, 1993). More recently Hoult et al. (1996) reported signifi cant cultivar differences within a population of 21 polyembryonic mango cultivars exposed to saline (480 mg/l NaCl) irrigation water for 10 months. Differences were measured in leaf Na (0.37–1.34%) and Cl (0.39–1.07%) concentrations but these were poorly correlated with toxicity symptoms on leaves.
Salinization of agricultural land is increasing and in many areas salinity management is critical. There appears to be suffi cient genetic diversity within Mangifera indica to enable the selection and development of saline-tolerant rootstocks (Kadman et al., 1976; Gazit and Kadman, 1980; Hoult et al., 1996). However, quantitative data on the critical limits of soil and water salinity which mango trees will tolerate without reductions in yield and fruit quality are needed.
Elevated atmospheric CO2 concentration
Growing ‘Kensington’ mango trees for 6 months in a controlled atmosphere glasshouse with an ambient CO2 concentration of 600 mol/mol resulted in more dry matter partitioned to the roots compared to plants grown in an ambient CO2 environment of 350 mol/mol (Schaffer et al., 1999) (Fig. 6.8). Fruit dry weight was greater for mango trees grown in an atmospheric CO2 concentration of 600 mol/mol compared to trees grown at 350 mol/mol (Schaffer et al., 1999) (Fig. 6.9). Most of the increased total fruit dry weight at
Oldleaves
Oldbranches
Newleaves
Plant part
Newbranches
Trunk Roots0
100
200
300
400
500
Dry
wei
ght (
g)
600 μmol CO2/mol
350 μmol CO2/mol
Fig. 6.8. Partitioning of dry matter in ‘Kensington’ mango trees grown for 6 months in atmospheric CO2 concentrations of 350 or 600 mol/mol. Bars represent means (n = 6 trees) ± standard error (Source: redrawn from Schaffer et al., 1999).
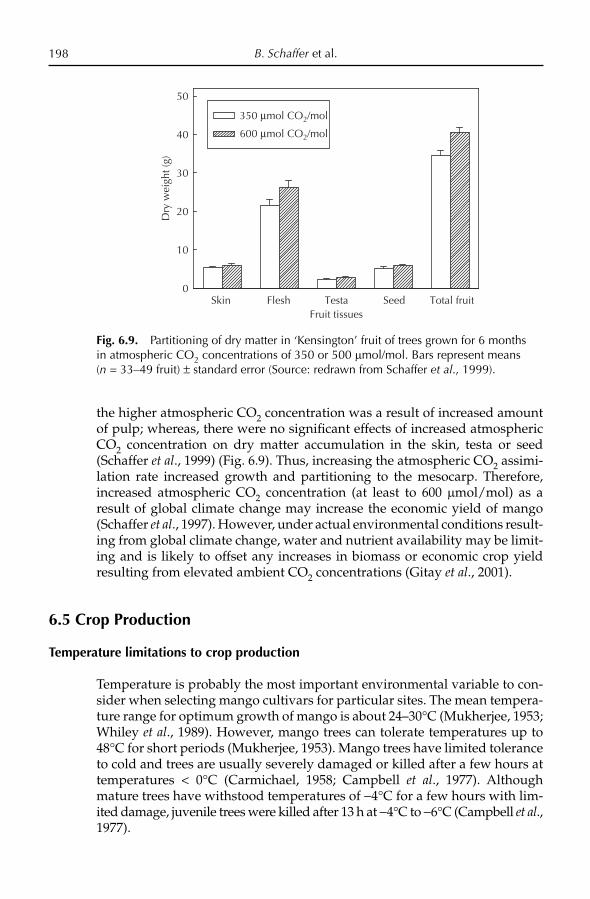
B. Schaffer et al.198
the higher atmospheric CO2 concentration was a result of increased amount of pulp; whereas, there were no signifi cant effects of increased atmospheric CO2 concentration on dry matter accumulation in the skin, testa or seed (Schaffer et al., 1999) (Fig. 6.9). Thus, increasing the atmospheric CO2 assimi-lation rate increased growth and partitioning to the mesocarp. Therefore, increased atmospheric CO2 concentration (at least to 600 mol/mol) as a result of global climate change may increase the economic yield of mango (Schaffer et al., 1997). However, under actual environmental conditions result-ing from global climate change, water and nutrient availability may be limit-ing and is likely to offset any increases in biomass or economic crop yield resulting from elevated ambient CO2 concentrations (Gitay et al., 2001).
6.5 Crop Production
Temperature limitations to crop production
Temperature is probably the most important environmental variable to con-sider when selecting mango cultivars for particular sites. The mean tempera-ture range for optimum growth of mango is about 24–30°C (Mukherjee, 1953; Whiley et al., 1989). However, mango trees can tolerate temperatures up to 48°C for short periods (Mukherjee, 1953). Mango trees have limited tolerance to cold and trees are usually severely damaged or killed after a few hours at temperatures < 0°C (Carmichael, 1958; Campbell et al., 1977). Although mature trees have withstood temperatures of −4°C for a few hours with lim-ited damage, juvenile trees were killed after 13 h at −4°C to −6°C (Campbell et al., 1977).
Fruit tissuesSkin Flesh Testa Seed Total fruit
Dry
wei
ght (
g)
0
10
20
30
40
50
600 μmol CO2/mol
350 μmol CO2/mol
Fig. 6.9. Partitioning of dry matter in ‘Kensington’ fruit of trees grown for 6 months in atmospheric CO2 concentrations of 350 or 500 mol/mol. Bars represent means (n = 33–49 fruit) ± standard error (Source: redrawn from Schaffer et al., 1999).

Ecophysiology 199
Monoembryonic mango cultivars tend to be more cold tolerant than polyembryonic cultivars, probably due their probable subtropical origin. There are also differences in fruit setting capacity between mono- and poly-embryonic cultivars at subtropical latitudes, where monoembryonic culti-vars crop more successfully when minimum temperatures fall below 12°C during fl owering (Searle et al., 1995). Differences in low temperature toler-ance have important implications for the selection of suitable cultivars for production under specifi c climatic conditions.
Light interception and orchard design
Light interception and utilization within tree canopies is a primary consider-ation in orchard design. Thus, tree spacing as well as pruning practices in orchards are primarily based on maximizing light for photosynthesis (see previous section on ‘Light’ under Photosynthesis, this chapter). The follow-ing equation describes the relationship between light interception, photosyn-thesis and yield in fruit tree orchards:
Biological Yield = (Light Available) (% Light Intercepted) (Photosynthe-sis) − Respiration (6.12)
where biological yield refers to dry matter production, including that of fruit (Lakso, 1994). The amount of light available is a function of climate and can-not be manipulated, and potential net photosynthetic effi ciency of a crop is inherent and cannot be altered without genetic manipulation. Thus, optimiz-ing biological yield is based on maximizing the percentage of light inter-cepted by the orchard canopy and minimizing stress so that photosynthetic potential is not compromised. With respect to light, the mango tree presents special problems due to long-lived leaves, dense canopies, and the potential for vigorous growth in some cultivars.
Maximizing light interception by the photosynthetic surface of an orchard is a function of tree spacing, canopy density and height. For some fruit tree trees, for example apple (Malus domestica) and citrus (Citrus spp.), the use of low vigour rootstocks and pruning provides opportunities for an array of management options. However, the lack of dwarfi ng rootstocks and the com-plications in pruning due to the fl oral morphology of mango (infl orescences are predominantly borne terminally on the most recently produced shoots) limit the effi cient harvest of light with respect to maintenance of productiv-ity. Improved productivity can be obtained by grafting, which either short-ens the juvenility phase and/or exerts some control over vigour. Compared to temperate fruit orchards, canopies of commercial mango orchards have a higher proportion of ‘shade’ to ‘sun’ leaves (Schaffer et al., 1994). Mango trees are rarely selectively pruned (Young and Sauls, 1981), although novel ideas of ‘heading back cuts’ after harvest have been investigated (Oosthuyse, 1992). In some areas, trees are mechanically topped and hedged to control tree size. Hedging encourages increased complexity (ramifi cation), resulting in a dense outer canopy. Where costs are not prohibitive, strategically timed, selective

B. Schaffer et al.200
pruning to increase the percentage of leaves exposed to > 60% of full sunlight will increase the photosynthetic effi ciency of the canopy with a potential improvement in yield. Schaffer and Gaye (1989) increased light interception of mango by removing 25% of the canopy. This resulted in higher chlorophyll concentrations in leaves of pruned canopies later in the year. Although net photosynthesis was not measured in that study, it is likely that the higher leaf chlorophyll concentrations in the pruned canopies resulted in higher photo-synthetic rates.
Light utilization of mango can be enhanced by pruning, but the timing of such treatments is critical. For example, in the subtropics, shoots produced following harvest generally fl ower 3–5 months after being exposed to induc-tive (cool) temperatures. Therefore, trees should be pruned immediately after harvest to improve light penetration and contain tree size. However, in the tropics there is a shorter period between the cessation of summer growth and fl owering and summer-grown shoots of many cultivars fail to induct that year (Scholefi eld et al., 1986; see Davenport, Chapter 5, this volume). There-fore, due to the removal of potential fl owering sites, summer pruning of mango trees in the tropics generally reduces yield in the following season.
Growth control of mango trees through the development of dwarfi ng rootstocks, low-vigour cultivars and pruning strategies to increase light pen-etration within tree canopies and by entire orchards, may improve yield per-formance of this crop. Improved information on light interception, critical leaf to fruit ratios and relationships between shoot maturity and fl oral induc-tion with respect to genotype/environmental interactions will enhance the development of improved cultural practices for mango production. It is per-tinent to emphasize that few evergreen fruit trees are as precocious as mango, or as large at maturity. Orchardists should take advantage of the precocity and of light interception principles by initial high-density planting, with hedgerows perhaps the best option. As trees become crowded, however, effi -ciency of light interception is compromised and remedial action before this occurs is required to maintain productivity.
6.6 Conclusions
Although much literature in the past stressed problems associated with low temperature on mango growth and production, sustained high temperatures with associated soil moisture stress during fruit ontogeny and high evapora-tive demand are perhaps the major reasons for relatively low yields in mango orchards worldwide, especially in the tropics. Although mango trees have a number of survival mechanisms that allow them to cope with stressful envi-ronments, these come at a considerable energy cost thereby potentially reduc-ing the availability of carbon-based inputs for fruiting. It is likely that annual assimilation gains and resource availability during critical developmental periods are inadequate for sustained high yields of quality fruit. These prob-lems can be alleviated by development of improved germplasm with adapta-tion to specifi c environments (Whiley et al., 2006). In the future, greater effort

Ecophysiology 201
is required for rootstock development and understanding the manipulative effect on whole tree physiology. Expansion of human populations and cli-mate change scenarios predict lesser and poorer quality water available for cropping systems across tropical and subtropical latitudes. Hence, greater salinity tolerance within the species should be identifi ed. Knowledge of mango physiology, particularly in relation to the tree’s responses to varying environmental conditions remains basic and must be expanded. A greater understanding of these principles together with their application will assist in the development of more productive cropping systems.
References
Adams, W.W. III, Zarter, C.R., Mueh, K.E., Amiard, V.S.E. and Demmig-Adams, B. (2005) Energy dissipation and photoinhibition: a continuum of photoprotection. In: Demming-Adams, B., Adams, W.W. III and Mattoo, A.K. (eds) Photoprotection, Photoinhibi-tion, Gene Regulation and Environment. Springer Verlag, Berlin, pp. 49–64.
Atkin, O.L.K., Millar, A.H., Gärdestrom, P. and Day, B. (2000) Photosynthesis, carbohy-drate metabolism and respiration in leaves of higher plants. In: Leegoood, R.C., Sharkey, T.D. and von Caemmerer, S. (eds) Photosynthesis, Physiology and Metabo-lism. Kluwer Academic, London, pp. 153–175.
Azcon-Bieto, J. (1983) Inhibition of photosynthesis by carbohydrates in wheat leaves. Plant Physiology 73, 681–686.
Badger, M.R. (1985) Photosynthetic oxygen exchange. Annual Review of Plant Physiology36, 27–53.
Baker, D.A. (1984) Water relations. In: Wilkins, M.G. (ed.) Advanced Plant Physiology. Pitman, London, pp. 297–316.
Bauerle, W.L., Weston, D.J., Bowden, J.D., Dudley, J.B. and Toler, B. (2004) Leaf absorp-tance of photosynthetically active radiation in relation to chlorophyll meter estimates among woody species. Scientia Horticulturae 10, 169–178.
Bingham, F.T., Fenn, L.B. and Oertli, J.J. (1968) A sand culture study of chloride toxicity to mature avocado trees. Proceedings of the Soil Science Society of America 32, 249–252.
Birkhold, K.T., Koch, K.E. and Darnell, R.L. (1992) Carbon and nitrogen economy of developing rabbiteye blueberry fruit. Journal of the American Society for Horticul-tural Science 117, 139–145.
Campbell, C.W. (1984) Tropical fruits and nuts. In: Martin, F.W. (ed.) CRC Handbook of Tropical Food Crops. CRC Press, Boca Raton, Florida, pp. 235–274.
Campbell, C.W., Knight, R.J. and Zareski, N.L. (1977) Freeze damage to tropical fruits in southern Florida in 1977. Proceedings of the Florida State Horticultural Society 90, 254–257.
Carmichael, W.W. (1958) Observations of cold damage to mangos in Dade County and the lower west coast. Proceedings of the Florida State Horticultural Society 71, 333–335.
Carswell, F.E., Meir, P., Wandell, E.V., Bonates, L.C.M., Kruijt, B., Barbosa, E.M., Nobre, A.D., Grace, J. and Jarvis, P.G. (2000) Photosynthetic capacity in a central Amazo-nian rain forest. Tree Physiology 20, 179–186.
Castro Neto, M.T., Reinhardt, D.H. and da S.Ledo, C.A. (2004) Determination of water potential on mango trees by pressure chamber. Acta Horticulturae 645, 425–427.
Chacko, E.K. (1991) Mango fl owering – still an enigma. Acta Horticulturae 291, 12–21.

B. Schaffer et al.202
Chauhan, P.S. and Pandey, R.M. (1984) Relative 14CO2 fi xation by leaves and fruits, and translocation of 14C-sucrose in mango. Scientia Horticulturae 22, 121–128.
Chirkova, T.V. and Gutman, T.S. (1972) Physiological role of branch lenticels in willow and poplar under conditions of root anaerobiosis. Soviet Plant Physiology 19, 289–295.
Cochard, H., Coll, L., Le Roux, X. and Améglio, T. (2002) Unraveling the effects of plant hydraulics on stomatal conductance during water stress in walnut. Plant Physiology128, 282–290.
Cochard, H., Damour, G., Bodet, C., Tharwat, I., Poirier, M. and Améglio, T. (2005) Evaluation of a new centrifuge technique for rapid generation of xylem vulnerabil-ity curves. Physiologia Plantarum 124, 410–418.
Collander, R. (1941) Selective absorption of cations by higher plants. Plant Physiology16, 691–720.
Constable, G.A. and Rawson, H.M. (1980) Effect of leaf position, expansion and age on photosynthesis, transpiration and water use effi ciency of cotton. Australian Journal of Plant Physiology 7, 89–100.
Crane, J.H. and Davies, F.S. (1988) Periodic and seasonal fl ooding effects on survival, growth, and stomatal conductance of young rabbiteye blueberry plants. Journal of the American Society for Horticultural Science 113, 488–493.
DeJong, T.M. and Doyle, J.F. (1985) Seasonal relationships between leaf nitrogen con-tent (photosynthetic capacity) and leaf canopy light exposure in peach (Prunus per-sica). Plant Cell and Environment 8, 701–706.
Di Vaio, C., Petito, A. and Buccheri, M. (2001) Effect of girdling on gas exchanges and leaf mineral content in the ‘Independence’ nectarine. Journal of Plant Nutrition 24, 1047–1060.
Downton, W.J.S. (1981) Water relations in Nerium oleander. Australian Journal of Plant Physiology 8, 329–334.
Drake, B.G., Gonzales-Meler, M. and Long, S.P. (1997) More effi cient plants: a conse-quence of rising atmospheric CO2? Annual Review of Plant Physiology and Plant Molecular Biology 48, 609–639.
Dreyer, E., Le Roux, X., Montpied, P., Daudet, F.A. and Masson, F. (2001) Temperature response of leaf photosynthetic capacity in seedlings from seven temperate species. Tree Physiology 21, 223–232.
Dwyer, L. and Stewart, D. (1986) Effect of leaf age and position on net photosynthetic rates in maize (Zea mays L.). Agricultural and Forest Meteorology 37, 29–46.
Ehlig, C.F. (1960) Effects of salinity on four varieties of table grapes grown in sand cul-ture. Proceedings of the American Society for Horticultural Science 76, 323–331.
Epron, D., Godard, C., Cornic, G. and Genty, B. (1995) Limitation of net CO2 assimila-tion rate by internal resistances to CO2 transfer in the leaves of two tree species (Fagus sylvatica L. and Castanea sativa Mill.). Plant Cell and Environment 18, 43–51.
Ethier, G.J. and Livingston, N.J. (2004) On the need to incorporate sensitivity to CO2transfer conductance into the Farquhar-von Caemmerer-Berry leaf photosynthesis model. Plant Cell and Environment 27, 137–153.
Evans, J.R. (1989) Photosynthesis and nitrogen relationships in leaves of C3 plants. Oecologia 78, 8–19.
Farquhar, G.D., von Caemmerer, S. and Berry, J.A. (1980) A biochemical model of pho-tosynthetic CO2 assimilation in leaves of C3 species. Planta 149, 78–90.
Fereres, E., Cruz-Romero, G., Hoffmann, G.J. and Rawlins, S.L. (1979) Recovery of or-ange trees following severe water stress. Journal of Applied Ecology 16, 833–842.
Field, C.B. (1987) Leaf-age effects on stomatal conductance. In: Zeiger, E., Farquhar, G.D. and Cowan, I. (eds) Stomatal Function. Stanford University Press, Palo Alto, Califor-nia, pp. 367–384.

Ecophysiology 203
Field, C.B. and Mooney, H. (1983) Leaf age and seasonal effects on light, water, and nitrogen use effi ciency in a California shrub. Oecologia 56, 348–355.
Field, C.B. and Mooney, H. (1986) The photosynthesis-nitrogen relationship in wild plants. In: Givnish, G.T. (ed.) On the Economy of Plant Form and Function. Cam-bridge University Press, Cambridge, pp. 25–55.
Flore, J.A. (1994) Stone fruit. In: Schaffer, B. and Andersen, P.C. (eds) Handbook of Envi-ronmental Physiology of Fruit Crops. Vol. 1, Temperate Crops. CRC Press, Boca Raton, Florida, pp. 233–270.
Foyer, C.H. (1988) Feedback inhibition of photosynthesis through source-sink regulation in leaves. Plant Physiology 26, 483–492.
Frak, E., Le Roux, X., Millard, P., Dreyer, E., Vandame, M. and Wendler, R. (2001) Chang-es in total leaf nitrogen and partitioning of leaf nitrogen drive photosynthetic ac-climation to light in fully developed walnut leaves. Plant Cell and Environment 24, 1279–1288.
Gandhi, S.R. (1955) The Mango in India. Farm Bulletin 6. Indian Council for Agricul-tural Research, New Delhi, 64 pp.
Gazit, S. and Kadman, A. (1980) 13-1 mango rootstock selection. HortScience 15, 669.
Gitay, H., Brown, S., Easterlin, W. and Jallow, B. (2001) Ecosystems and their goods and services. In: McCarthy, J.J., Osvaldo, F.C., Leary, N.A., Dokken, D.J. and Whilte, K.S. (eds) Climate Change 2001: Impacts, Adaptation and Vulnerability. Cambridge University Press, Cambridge, UK, pp. 237–242.
Gonzalez, A. and Blaikie, S.J. (2003) Seasonal variation of carbon assimilation in mango (cv. Kensington Pride): effect of fl owering treatments. Australian Journal of Agricul-tural Research 54, 309–321.
Harding, R.B., Miller, M.P. and Fireman, M. (1958) Absorption of salts by citrus leaves during sprinkling with water soluble surface irrigation. Proceedings of the Ameri-can Society for Horticultural Science 71, 248–256.
Harley, P.C., Thomas, R.B., Reynolds, J.F. and Strain, B.R. (1992) Modelling photosyn-thesis of cotton grown in elevated CO2. Plant Cell and Environment 15, 271–282.
Harvey, G.W. (1979) Photosynthetic performance of isolated leaf cells from sun and shade plants. Carnegie Institute Washington Yearbook 79, 161–164.
Hollinger, D.Y. (1996) Optimality and nitrogen allocation in a tree canopy. Tree Physiol-ogy 16, 627–634.
Houghton, J. (2005) Global warming. Reports on Progress in Physics 68, 1343–1403.Hoult, M.D., Donnelly, M.M. and Smith, M.W. (1996) Salt exclusion varies amongst
polyembryonic mango cultivar seedlings. Acta Horticulturae 455, 455–458.Idso, S.B. and Kimball, B.A. (1991) Downward regulation of photosynthesis and growth
at high CO2 levels. Plant Physiology 96, 990–992.Issarakraisila, M., Considine, J.A. and Turner, D.W. (1991) Pattern of vegetative and re-
productive growth of mango trees in a warm temperate region of Western Australia. Acta Horticulturae 291, 188–197.
Jackson, J.E. (1980) Light interception and utilization by orchard systems. Horticultural Reviews 2, 208–287.
Jindal, P.C., Singh, J.P. and Gupta, O.P. (1975) Screening of mango seedlings for salt tolerance. Haryana Journal of Horticultural Science 4, 112–115.
Jindal, P.C., Singh, J.P. and Gupta, O.P. (1976) Studies on salt tolerance in mango – injurious effects of salts on young mango seedlings. Progressive Horticulture 8, 65–71.
Joel, D.M. (1980) Resin ducts in the mango fruit: a defense system. Journal of Experi-mental Botany 31, 1707–1718.

B. Schaffer et al.204
Joel, D.M. and Fahn, A. (1980a) Ultrastructure of the resin ducts of the Mangifera indicaL. (Anacardiaceae). 1. Differentiation and senescence of the shoot ducts. Annals of Botany 46, 225–223.
Joel, D.M. and Fahn, A. (1980b) Ultrastructure of the resin ducts of the Mangifera indicaL. (Anacardiaceae). 2. Resin secretion in the primary stem ducts. Annals of Botany46, 779–783.
Joel, D.M. and Fahn, A. (1980c) Ultrastructure of the resin ducts of the Mangifera indicaL. (Anacardiaceae). 3. Secretion of the protein-polysaccharide mucilage in the fruit. Annals of Botany 46, 785–790.
Jones, H.G. (1981) Carbon dioxide exchange of developing apple (Malus pumila Mill.) fruits. Journal of Experimental Botany 32, 1203–1210.
Jones, H.G. and Sutherland, R.A. (1991) Stomatal control of xylem embolism. Plant Cell and Environment 14, 607–612.
Kadman, A. (1964) The uptake and accumulation of sodium in avocado seedlings. Pro-ceedings of the American Society for Horticultural Science 85, 179–182.
Kadman, A., Gazit, S. and Ziv, G. (1976) Selection of mango rootstocks for adverse water and soil conditions in arid areas. Acta Horticulturae 57, 81–88.
Kallarackal, J., Milburn, J.A. and Baker, D.A. (1990) Water relations of the banana. III. Effects of controlled water stress on water potential, transpiration, photosynthesis and leaf growth. Australian Journal of Plant Physiology 17, 79–90.
Kellomäki, S. and Wang, K.-Y. (1997) Photosynthetic responses of Scots pine to elevated CO2 and nitrogen supply: results of a branch-in-bag experiment. Tree Physiology17, 231–240.
Koch, K.E. (1996) Carbohydrated-modulated gene expression in plants. Annual Review of Plant Physiology and Plant Molecular Biology 47, 509–540.
Kozlowski, T.T. (1984) Responses of woody plants to fl ooding. In: Kozlowski, T.T. (ed.) Flooding and Plant Growth. Academic Press, New York, pp. 129–163.
Kramer, P.J. (1983) Water Relations of Plants. Academic Press, London.Krapp, A. and Stitt, M. (1995) An evaluation of direct and indirect mechanisms for the
‘sink-regulation’ of photosynthesis in spinach: changes in gas exchanges, carbohy-drates, metabolites, enzyme activities and steady-state transcript levels after cold-girdling source leaves. Planta 195, 313–323.
Kriedemann, P.E. (1968) Observations on gas exchange in the developing sultana berry. Australian Journal of Biological Science 21, 907–916.
Lakso, A.N. (1994) Apple. In: Schaffer, B. and Andersen, P.C. (eds) Handbook of Environ-mental Physiology of Fruit Crops. Vol. I. Temperate Crops. CRC Press, Boca Raton, Florida, pp. 3–43.
Larson, K.D., Schaffer, B. and Davies, F.S. (1989) Effect of irrigation on leaf water poten-tial, growth and yield of mango trees. Proceedings of the Florida State Horticultural Society 102, 226–228.
Larson, K.D., Davies, F.S. and Schaffer, B. (1991a) Floodwater temperature and lenticel hypertrophy of Mangifera indica L. American Journal of Botany 78, 1397–1403.
Larson, K.D., Graetz, D.A. and Schaffer, B. (1991b) Flood-induced chemical transforma-tions in calcareous agricultural soils of South Florida. Soil Science 152, 33–40.
Larson, K.D., Schaffer, B. and Davies, F.S. (1991c) Flooding, leaf gas exchange and growth of mango trees in containers. Journal of the American Society of Horticul-tural Science 116, 156–160.
Larson, K.D., Schaffer, B. and Davies, F.S. (1992) Flooding, mineral nutrition and gas exchange of mango trees. Scientia Horticulturae 52, 113–124.
Larson, K.D., Schaffer, B. and Davies, F.S. (1993) Floodwater oxygen content, lenticel hypertrophy, and ethylene evolution from mango (Mangifera indica L.) trees. Jour-nal of Experimental Botany 44, 665–671.

Ecophysiology 205
Le Roux, X., Sinoquet, H. and Vandame, M. (1999) Spatial distribution of leaf weight per area and leaf nitrogen concentration in relation to local radiation regime within an isolated tree crown. Tree Physiology 19, 181–188.
Le Roux, X., Walcroft, A.S., Daudet, F.A., Sinoquet, H., Chaves, M.M., Rodrigues, A. and Osorio, L. (2001) Photosynthetic light acclimation in peach leaves: importance of changes in mass:area ratio, nitrogen concentration, and leaf nitrogen partitioning. Tree Physiology 21, 377–386.
Lu, P. (2002) Measurement of whole-tree water use of some tropical and subtropical tree crops and its application in irrigation management. Acta Horticulturae 575, 781–789.
Lu, P. (2006) Mango water relations and irrigation schedule. In: Malik, A.U. (ed.) Pro-ceedings of the First International Conference on Mango and Date Palm: Culture and Export. University of Agriculture, Faisalabad, Pakistan, 20–23 June 2005, pp. 1–11.
Lynch, J. and González, A. (1993) Canopy nutrient allocation in relation to incident light in the tropical fruit tree Borojoa patinoi (Cuatr.). Journal of the American Society for Horticultural Science 118, 777–785.
Manter, D.K. and Kerrigan, J. (2004) A/Ci curve analysis across a range of woody plant species: infl uence of regression analysis parameters and mesophyll conductance. Journal of Experimental Botany 55, 2581–2588.
Marshall, B. and Biscoe, P.V. (1980) A model for C3 leaves describing the dependence of net photosynthesis on irradiance. II. Application to the analysis of fl ag leaf photo-synthesis. Journal of Experimental Botany 31, 41–48.
Mayers, P.E., Hutton, D.G. and Saranah, J. (1984) Integrated control of bacterial black spot of mangoes in south east Queensland. In: Chapman, G.R. (ed.) Proceedings of the First Australian Mango Research Workshop. Commonwealth Scientifi c and Industrial Research Organization (CSIRO), Melbourne, pp. 258–260.
Medlyn, B.E., Dreyer, E., Ellsworth, D., Forstreuter, M., Harley, P.C., Kirschbaum, M.U.F., Le Roux, X., Loustau, D., Montpied, P., Strassemeyer, J., Walcroft, A.S. and Wang, K. (2002) Temperature response of parameters of a biochemically-based model of photosynthesis: a review of experimental data. Plant Cell and Environment 25, 1169–1176.
Merzlyak, M.N. and Hendry, G.A.F. (1994) Free radical metabolism, pigment degrada-tion and lipid peroxidation in leaves during senescence. Proceedings of the Royal Society of Edinburgh 102B, 459–471.
Mukherjee, S.K. (1953) The mango – its botany, cultivation, uses, and future improve-ment, especially as observed in India. Economic Botany 7, 130–160.
Mukherjee, S.K. (1972) Origin of the mango (Mangifera indica). Economic Botany 26, 260–266.
Nii, N., Watanabe, T., Yamaguchi, K. and Nishimura, M. (1995) Changes of anatomical features, photosynthesis and ribulose-bisphosphate carboxylase-oxygenase content of mango leaves. Annals of Botany 76, 649–656.
Nobel, P.S. (1983) Biophysical Plant Physiology and Ecology. W.H. Freeman, New York.
Noodén, L.D., Guiamét, J.J. and John, J. (1997) Senescence mechanisms. Physiologia Plantarum 101, 746–753.
Oosthuyse, S.A. (1992) Ideas on pruning of mango trees. South African Mango Grower’s Association Yearbook 12, 1–8.
Pammenter, N.W., Loreto, F. and Sharkey, T.D. (1993) End product feedback effects on photosynthetic electron transport. Photosynthesis Research 35, 5–14.
Paul, M.J. and Foyer, C. (2001) Sink regulation of photosynthesis. Journal of Experimen-tal Botany 360, 1383–1400.

B. Schaffer et al.206
Paul, M.J. and Pellny, T.L. (2003) Carbon metabolite feedback regulation of leaf photo-synthesis and development. Journal of Experimental Botany 382, 539–547.
Peisker, M. and Apel, H. (2001) Inhibition by light of CO2 evolution from dark evolution: comparison of two gas exchange methods. Photosynthesis Research 70, 291–298.
Pongsomboon, W. (1991) Effects of temperature and water stress on tree growth, fl ower-ing, fruit growth and retention of mango (Mangifera indica L.). PhD thesis, Kasetsart University, Bangkok, Thailand.
Pongsomboon, W., Whiley, A.W., Stephenson, R.A. and Subhadrabandhu, S. (1992) Ef-fect of air temperatures on diurnal variation of water potential, conductance and CO2 assimilation of mango (Mangifera indica L.) leaves. Acta Horticulturae 321, 472–481.
Proctor, J.T.A. and Creasey, L.L. (1971) Effect of supplementary light on anthocyanin synthesis in McIntosh apples. Journal of the American Society for Horticultural Science 96, 523–526.
Quirino, B.F., Reiter, W.D. and Amasino, R.D. (2001) One of two tandem Arabidopsisgenes homologous to monosaccharide transporters is senescence-associated. PlantMolecular Biology 46, 447–457.
Reich, P.B. and Borchert, R. (1988) Changes with leaf age in stomatal function and water status of several tropical tree species. Biotropica 20, 60–69.
Riesmeier, J.W., Flugge, U.-I., Schulz, B., Heineke, D., Heldt, H.-W., Willmitzer, L. and Frommer, W.B. (1993) Antisense repression of the chloroplast triose phosphate translocator affects carbon partitioning in transgenic potato plants. Proceedings of the National Academy of Science of the USA 90, 6160–6164.
Roper, T.R. and Williams, L. (1989) Net CO2 assimilation and carbohydrate partitioning of grapevine leaves in response to trunk girdling and gibberellic acid application. Plant Physiology 89, 1136–1140.
Roper, T.R., Keller, J.D., Loescher, W.H. and Rom, C.R. (1988) Photosynthesis and car-bohydrate partitioning in sweet cherry: fruiting effects. Physiologia Plantarum 72, 42–47.
Rosati, A., Esparza, G., DeJong, T.M. and Pearcy, R.W. (1999) Infl uence of canopy light environment and nitrogen availability on leaf photosynthetic characteristics and photosynthetic nitrogen-use effi ciency of fi eld-grown nectarine trees. Tree Physiol-ogy 19, 173–180.
Rosati, A., Day, K.R. and DeJong, T.M. (2000) Distribution of leaf mass per unit leaf area and leaf nitrogen concentration determine partitioning of leaf nitrogen within cano-pies. Tree Physiology 20, 271–276.
Schaffer, B. and Gaye, G.O. (1989) Seasonal effects of pruning on light penetration, specifi c leaf density, and chlorophyll content of mango. Scientia Horticulturae 41, 55–61.
Schaffer, B., Andersen, P.C. and Ploetz, R.C. (1992) Responses of fruit crops to fl ooding. Horticultural Reviews 13, 257–313.
Schaffer, B., Whiley, A.W. and Crane, J.H. (1994) Mango. In: Schaffer, B. and Andersen, P.C. (eds) Handbook of Environmental Physiology of Fruit Crops. Vol. 2, Sub-tropicaland Tropical Crops. CRC Press, Boca Raton, Florida, pp. 165–197.
Schaffer, B., Whiley, A.W., Searle, C. and Nissen, R.J. (1997) Leaf gas exchange, dry mat-ter partitioning, and mineral element concentrations in mango (Mangifera indica L.) as infl uenced by elevated atmospheric CO2 concentration and root restriction. Journal of the American Society for Horticulture Science 122, 849–855.
Schaffer, B., Whiley, A.W. and Searle, C. (1999) Atmospheric CO2 enrichment, root re-striction, photosynthesis, and dry-matter partitioning in subtropical and tropical fruit crops. HortScience 34, 1033–1037.

Ecophysiology 207
Schaffer, B., Davies, F.S. and Crane, J.H. (2006) Responses of subtropical and tropical fruit trees to fl ooding in calcareous soil – a review. HortScience 41, 549–555.
Schaper, H. and Chacko, E.K. (1993) Effect of irradiance, leaf age, chlorophyll content and branch-girdling on gas exchange of cashew (Anacardium occidentale L.)leaves. Journal of Horticultural Science 68, 541–550.
Schmutz, U. (2000) Effect of salt stress (NaCl) on whole plant CO2-gas exchange in mango. Acta Horticulturae 509, 269–276.
Schmutz, U. and Lüdders, P. (1993) Physiology of saline stress in one mango (Mangiferaindica L.) rootstock. Acta Horticulturae 341, 160–167.
Scholefi eld, P.B., Oag, D.R. and Sedgley, M. (1986) The relationship between vegetative and reproductive development in the mango in northern Australia. Australian Journal of Agricultural Research 37, 425–433.
Searle, C., Whiley, A.W., Simpson, D.R. and Saranah, J.B. (1995) A preliminary phenol-physiological model for ‘Kensington’ in subtropical Australia. In: Mango 2000 – Marketing Seminar and Production Workshop Proceedings. Department of Primary Industries, Brisbane, pp. 124–127.
Shivashankara, K.S. and Mahai, C.K. (2000) Inhibition of photosynthesis by fl owering in mango (Mangifera indica L.). A study by gas exchange methods. Scientia Horticul-turae 83, 205–212.
Singaas, E.L., Ort, D.R. and deLucia, E.H. (2001) Variation in measured values of photo-synthetic quantum yield in ecophysiological studies. Oecologia 128, 15–23.
Singh, R.N. and Arora, K.S. (1965) Some factors affecting fruit drop in mango (Mangiferaindica L.). Indian Journal of Agricultural Science 35, 196–205.
Spreitzer, R.J. and Salvucci, M.E. (2002) Rubisco: structure, regulatory interactions, and possibilities for a better enzyme. Annual Review of Plant Physiology and Plant Molecular Biology 53, 449–475.
Stephenson, R.A., Ko, H.L. and Gallagher, E.C. (1989) Plant water relations of stressed, non-bearing macadamia trees. Scientia Horticulturae 39, 41–53.
Sukhvibul, N., Whiley, A.W., Smith, M.K. and Hetherington, S.E. (2000) Susceptibility of mango (Mangifera indica L.) to cold-induced photoinhibition and recovery at different temperatures. Australian Journal of Agricultural Research 51, 503–513.
Sukonthasing, S., Wongrakpanich, M. and Verheij, E.W.M. (1991) Mangifera indica L. In: Verheij, E.W.M. and Coronel, R.E. (eds) Plant Resources of South-East Asia, No. 2. Edible Fruits and Nuts. Pudoc, Wageningen, the Netherlands, pp. 211–216.
Sun, J., Okita, T.W. and Edwards, G.E. (1999) Modifi cation of carbon partitioning, pho-tosynthetic capacity, and O2 sensitivity in Arabidopsis plants with low ADP-glucose pyrophosphorylase activity. Plant Physiology 119, 267–276.
Thomas, B.R. and Rodriguez, R.L. (1994) Metabolite signals regulate gene expression and source/sink relations in cereal seedlings. Plant Physiology 54, 201–207.
Tyree, M.T. and Sperry, J.S. (1988) Do woody plants operate near the point of cata-strophic xylem dysfunction caused by dynamic water stress? Answers from a model. Plant Physiology 88, 574–580.
Urban, L. and Alphonsout, L. (2007) Girdling decreases photosynthetic electron fl uxes and induces sustained photoprotection in mango leaves. Tree Physiology 27, 345–352.
Urban, L. and Jannoyer, M. (2004) Functioning and role of stomata in mango leaves. Acta Horticulturae 645, 441–446.
Urban, L., Berteuil, F. and Léchaudel, M. (2002) A coupled photosynthesis and stomatal conductance model for mango leaves. Acta Horticulturae 584, 81–88.
Urban, L., Le Roux, X., Sinoquet, H., Jaffuel, S. and Jannoyer, M. (2003) A biochemical model of photosynthesis for mango leaves: evidence for an effect of the local source/sink balance on photosynthetic capacity. Tree Physiology 23, 289–300.

B. Schaffer et al.208
Urban, L., Lu, P. and Thibaud, R. (2004a) Inhibitory effect of fl owering on leaf photosyn-thesis in mango. Tree Physiology 24, 387–399.
Urban, L., Léchaudel, M. and Lu, P. (2004b) Interpreting the effect of fruit load and girdling on leaf net photosynthesis in mango. Journal of Experimental Botany 405, 2075–2085.
Urban, L., Montpied, P. and Normand, F. (2006) Season effects on leaf nitrogen partition-ing and photosynthetic water use effi ciency in mango. Journal of Plant Physiology163, 48–57.
Urban, L., Jégouzo, L., Damour, G., Vandame, M. and François, C. (2008) Interpreting the decrease in leaf photosynthesis during fl owering in mango. Tree Physiology 28, 1025–1036.
Van der Meulen, A., Smith, J.H.E., Kok, J.B., Schwartz, A. and Jacobs, C.J. (1971) Mango growing in South Africa. Citrus and Subtropical Fruit Research Institute Leafl et No. 48. Department of Agriculture and Technical, Pretoria.
Venning, F.D. (1948) The ontogeny of the laticiferous canals in the Anacardiacae. Amer-ican Journal of Botany 35, 637–644.
Walcroft, A.S., Whitehead, D., Silvester, W.B. and Kelliher, F.M. (1997) The response of photosynthetic model parameters to temperature and nitrogen concentration in Pi-nus radiata D. Don. Plant Cell and Environment 20, 1338–1348.
Walcroft, A., Le Roux, X., Diaz-Espejo, A., Dones, N. and Sinoquet, H. (2002) Spatial and temporal variability of photosynthetic capacity within a peach tree crown. Tree Physiology 22, 929–938.
Walker, R.R. (1986) Sodium exclusion and potassium-sodium selectivity in salt-treated trifoliate orange (Poncirus trifoliata) and Cleopatra mandarin (Citrus reticulata)plants. Australian Journal of Plant Physiology 13, 293–303.
Weng, J.H., Chen, Y.N. and Liao, T.S. (2006a) Relationships between chlorophyll fl uo-rescence parameters and photochemical refl ectance index of tree species adapated to different temperature regimes. Functional Plant Biology 33, 241–246.
Weng, J.H., Jhaung, L.H., Jiang, J.Y., Lai, G.M. and Liao, T.S. (2006b) Down-regulation of photosystem 2 effi ciency and spectral refl ectance in mango leaves under very low irradiance and varied chilling treatments. Photosynthetica 44, 248–254.
Whiley, A.W. (1994) Ecophysiological studies and tree manipulation for maximisation of yield potential in avocado (Persea americana Mill.). PhD thesis, University of Natal, Pietermaritzburg, South Africa, 174 pp.
Whiley, A.W. and Schaffer, B. (1994) Avocado. In: Schaffer, B. and Andersen, P.C. (eds) Handbook of Environmental Physiology of Fruit Crops, Vol. 2. Sub-Tropical and Tropical Crops. CRC Press, Boca Raton, Florida, pp. 3–35.
Whiley, A.W., Rasmussen, T.S., Saranah, J.B. and Wolstenholme, B.N. (1989) Effect of temperature on growth, dry matter production and starch accumulation in ten man-go (Mangifera indica L.) cultivars. Journal of Horticultural Science 64, 753–765.
Whiley, A.W., Schaffer, B. and Lara, S.P. (1991) Carbon dioxide exchange of developing avocado (Persea americana Mill.) fruit. Tree Physiology 11, 85–94.
Whiley, A.W., Searle, C., Schaffer, B. and Wolstenholme, B.N. (1999) Cool orchard tem-peratures or growing trees in containers can inhibit leaf gas exchanges of avocado and mango. Journal of the American Society for Horticultural Science 124, 46–51.
Whiley, A.W., Hofman, P.J., Marques, J.R., Stubbings, B. and Whiley, D.G. (2006) Devel-opment of best practice pre- and postharvest protocols for production of Calypso™ mango. Final Project Report, Horticulture Australia Ltd (FR02049), Sydney, Austra-lia, pp. 157.
Wilson, K.B., Baldocchi, D.D. and Hanson, P.J. (2000) Quantifying stomatal and non-stomatal limitations to carbon assimilation resulting from leaf aging and drought in mature deciduous tree species. Tree Physiology 20, 787–797.

Ecophysiology 209
Wingler, A., von Schaewen, R.C., Leegood, P.J. and Quick, W.P. (1998) Regulation of leaf senescence by cytokinin, sugars and light. Effects on NADH-dependent hy-droxypyruvate reductase. Planta 116, 329–335.
Wolstenholme, B.N. and Whiley, A.W. (1995) Ecophysiology of the mango tree as a basis for pre-harvest management. In: Mango 2000 – Marketing Seminar and Pro-duction Workshop, Department of Primary Industries, Brisbane, pp. 5–19.
Young, T.W. and Sauls, J.W. (1981) The Mango Industry in Florida. Florida Cooperative Extension Services Bulletin 189, University of Florida, Gainesville, Florida.
Yusof, I.M., Buchanan, D.W. and Gerber, J.F. (1969) The response of avocado and mango to soil temperature. Journal of the American Society for Horticultural Science 94, 619–621.
Zude, M. and Ludders, P. (1997) Vegetative growth cycles and comparison of chloro-phyll and phenol contents, gas exchange and water regime from young to old leaves. Journal of Applied Botany 71, 10–13.
Zude-Sasse, M., Hartmond, U., Ebert, G. and Ludders, P. (2001) Pyridine nucleotide charge reduces photosynthesis under short-term oxygen defi ciency. Journal of the American Society for Horticultural Science 126, 703–709.

© CAB International 2009. The Mango, 2nd Edition: Botany, Production and Uses210 (ed. R.E. Litz)
7 Fruit Diseases
D. Prusky, I. Kobiler, I. Miyara and N. AlkanAgricultural Research Organization, Bet Dagan, Israel
7.1 Introduction 2107.2 Anthracnose 211
The pathogenesis strategy 212Epidemiology – the disease cycle 213Mechanisms of fungal pathogenicity and fruit resistance 214Management 216
7.3 Alternaria Rot (Black Spot) 219Pathogenesis 219Mechanisms of fungal pathogenicity and fruit resistance 220Management 220
7.4 Stem-end Rots 221Pathogenesis 221Epidemiology 222Management 223
7.5 Other Minor Diseases 2247.6 Conclusions 224
7.1 Introduction
Postharvest diseases can reduce fruit quality and cause severe economic losses, due to decay resulting in completely unmarketable and blemished fruits that are often sold in less demanding local markets, where the prices are considerably lower than export prices. It is clear to the producer that quality at the time of harvest cannot be improved but merely maintained for a limited period of time. Harvesting fruits at the optimal stage, with respect to size and maturity, can, therefore, ensure peak quality and maximum shelf-life potential. Thus, managing total tree health can contribute to reducing postharvest losses. It is known that older and neglected orchards may become a profuse inoculum source for postharvest diseases. Furthermore, preharvest

Fruit Diseases 211
stress factors such as excess or shortage of water, fl uctuating or extreme envi-ronmental conditions and high nitrogen levels (Hawthorne, 1989) can render fruit more susceptible to postharvest diseases. After harvest, improper treat-ment of fruits through storage at non-optimal temperatures accelerates fruit deterioration as a result of enhancement of normal physiological processes such as respiration and ethylene production (Thompson et al., 2002). More-over, excessive temperatures in the fi eld during harvesting, in transit and in the packing house can render tropical fruit more susceptible to chilling injury and can contribute to the development of disease. Combinations of high tem-peratures and high relative humidity (RH) favour the growth of postharvest pathogens, and can contribute to the development of disease at the retail end (Banik et al., 1998). Proper, uninterrupted cooling, therefore, protects quality and extends the shelf life of produce. Precooling of products (Thompson etal., 2002), and application of top or liquid icing, vacuum, hydrocooling and forced air cooling (Thompson et al., 2002) are examples of effective alterna-tive methods that can be used to remove fi eld heat and to restrict pathogen growth. Effective cold-chain management is crucial to ensuring product integrity and preventing postharvest pathogens from spoiling produce in transit or shortly after arrival (Lizada, 1993). Furthermore, low-temperature storage conditions are generally not conducive to disease development, a fact that can be exploited to ensure quality and extend shelf life (Banik et al., 1998). However, improper cooling during shipping, or interrupted cooling will also promote microbial growth, resulting ultimately in product spoilage. In practice, several breaks in the cold chain might have signifi cant impacts on decay development.
Among the postharvest diseases of mango, anthracnose is the most prev-alent in humid growing regions. The incidence of this disease can reach almost 100% in fruit produced under wet or very humid conditions. Other common postharvest diseases of mango in humid areas are stem-end rot, caused by Lasiodiplodia theobromae (Pat.) Griffon & Maubl. (Sangchote, 1998) and Dothiorella dominicana Petrak. et Cif. (Johnson and Sangchote, 1994), and black mould rot, caused by Aspergillus niger van Tieghem. Mango black spot, caused by Alternaria alternata (Fr.:Fr.) Keissl. (Prusky et al., 1983), is prevalent in dry countries.
7.2 Anthracnose
Mango anthracnose is caused by Glomerella cingulata (Stoneman) Spauld. & H. Schrenk (anamorph: Colletotrichum gloeosporioides (Penz.) Penz. & Sacc. In Penz.), C. gloeosporioides Penz. var. minor J.H. Simmonds (Fitzell and Peak, 1984) and Colletotrichum acutatum J.H. Simmonds (Freeman et al., 1998). The pathogen also causes blossom blight, leaf blight and, in severe cases, tree dieback (Ploetz, 1994; Ploetz and Prakash, 1997). The form-genus Colletotri-chum Corda (form-order Melanconiales; form-class Coelomycetes; subdivi-sion Deuteromycotina) comprises imperfect fungal species which exist as Glomerella (subdivision Ascomycotina) in their sexual, teleomorphic or perfect

D. Prusky et al.212
state. These fungal pathogens occur worldwide, and the genus is synony-mous with anthracnose. Leaf anthracnose appears as irregular black necrotic spots on both sides of the mango leaves. Lesions often coalesce and form large necrotic areas, frequently along the leaf margins. Lesions develop pri-marily on young tissue, and conidia are formed in lesions of all ages. Under favourable conditions, the fungus can invade the twigs and cause dieback (Ploetz et al., 1996). Panicle anthracnose or blossom blight can affect both the infl orescence stalk and the individual fl owers; in the stalk, elongated dark grey to black lesions appear; the blighted fl owers are dry, and their colour ranges from brown to black. Fruits smaller than pea-size can be infected and abort; whereas, larger fruits that are aborted because of normal self-thinning or other physiological causes are usually mummifi ed. The resulting mummi-fi ed fruit are invaded saprophytically by C. gloeosporioides, and the fungus sporulates abundantly on them.
Although fi eld anthracnose causes considerable damage, the vast losses infl icted by postharvest anthracnose are of far greater economic importance. Postharvest anthracnose appears on the fruit surface, as rounded brown to black lesions with an indefi nite border. Lesions > 2 cm in diameter are fairly common. Lesions of various sizes can coalesce and cover extensive areas of the fruit, typically in a tear-drop pattern that develops from the basal towards the distal end of the fruit. Lesions are usually restricted to the peel, but in severe cases the fungus can invade the pulp. In advanced stages of the dis-ease, the fungus produces acervuli, and abundant orange to salmon-pink masses of conidia appear on the lesions.
The pathogenesis strategy
Colletotrichum gloeosporioides is regarded as a hemibiotrophic species that commences its invasion with a transient post-penetrative asymptomatic bio-trophy, characterized by a temporary confi nement with a localized mode of intracellular hemibiotrophy. This is succeeded by a phase of destructive necrotrophy that culminates in the appearance of disease symptoms and pro-duction of conidiomata by the pathogens (Prusky and Plumbley, 1992; Latunde-Dada et al., 1999). Quiescent species pass through a prolonged phase of pre-penetrative growth that is arrested in synchrony with the physiological state of the infected organ.
Following its landing on the fruit or host tissue, the ungerminated aseptate conidium differentiates to a melanized appressorium. In the case of Colletotrichum lindemuthianum, appressorial adhesion is mediated by the man-nose- and galactose-rich glycoproteins of the extracellular matrices, which are secreted during appressorial differentiation (Pain et al., 1996). Both the surface wax of the specifi c fruit and the hard surface of the tissue are recog-nized as host signals that selectively trigger germination and appresso-rium formation solely by the conidia of C. gloeosporioides (Podila et al., 1993; Liu and Kolattukudy, 1998). The melanin of the melanized appressoria pro-tects the fungus and behaves as a permeability barrier, and the appressoria

Fruit Diseases 213
contain osmolytes (i.e. glycerol) that are needed for generating the osmoti-cum from which high turgor pressure will drive the invasive forces of the penetrating hyphae through the small appressorial pores (0.5 m in Col-letotrichum sublineolum). The osmolyte is generated by the metabolism of stored glycogen, trehalose and lipids and, at an in vivo concentration of at least 3.3 M in mature-melanized appressoria (de Jong et al., 1997), this osmo-lyte is capable of generating turgor pressures as high as 8 MPa (Howard et al., 1991; Money and Howard, 1996). Melanized appressoria appear to be quite capable of a forcible, non-enzymatic penetration of an intact host surface. However, Dickman et al. (1982) suggested that attack on papaya by C. gloeo-sporioides depended on cutinase production by the pathogen. The germi-nated appressoria in Colletotrichum develop single infection hyphae that grow and extend into the waxy cuticle, reach the fi rst layers of pericarp cells, and then become quiescent for long periods of time (Coates et al., 1993; Prusky, 1996).
In recent years, the taxonomy of Colletotrichum has been clarifi ed by the adoption of molecular biological techniques that involve the use of poly-merase chain reaction amplifi cation, alignment of nucleotide sequences, and the construction of dendograms, phylogenetic trees and similarity matrices from the data generated. For example, Sherriff et al. (1994) and Sreenivasap-rasad et al. (1996) used homologies between the nucleotide sequences from amplifi ed non-transcribed regions (internally transcribed spacers 1 and 2 (ITS1, ITS2)) and the large subunits (domains 1 and 2) of ribosomal DNA extracted from a wide range of isolates to both justify and resolve the taxo-nomic status of Colletotrichum species. Bailey (1997) proposed the adoption of species aggregates based on these and other data.
Epidemiology – the disease cycle
Conidia are formed abundantly in the mango canopy (Fitzell and Peak, 1984) which, therefore, is considered to be the primary source of inoculum. In the fi eld, C. gloeosporioides produces conidia on lesions on leaves, twigs, panicles and mummifi ed fruits (Arauz, 2000). Conidia can be rain-splashed onto other leaves or fl owers, where they can cause secondary infections. Developing fruit can be infected, and some isolates can cause preharvest fruit loss (Gan-totti and Davis, 1993). In the case of postharvest anthracnose, developing fruits are infected in the fi eld, but the infections remain quiescent until the onset of ripening, which occurs after harvest. Once the climacteric period of the fruit starts, lesions begin to develop but there is no fruit-to-fruit infection. In this context Prusky and co-workers (Guetsky et al., 2005) suggested that the capability of C. gloeosporioides to cause early disease symptoms in unripe fruits depends on the activation of laccases by specifi c strains of the fungus. Details of these systems are discussed in the following section.
Colletotrichum gloeosporioides requires free water or RH > 95% to enable conidial germination and appressorium formation (Fitzell et al., 1984; Dodd et al., 1991). However, conidia can survive for 1–2 weeks under low RH and

D. Prusky et al.214
then germinate if exposed to 100% RH (Estrada et al., 1993). In general, infec-tion is favoured at temperatures ranging from 20 to 30°C.
Mechanisms of fungal pathogenicity and fruit resistance
Unripe mango fruits are reservoirs of extremely high concentrations of pre-formed antimicrobial compounds. This arsenal of constitutive resistance weapons may accumulate in the immature pericarp, but not in the mesocarp, at concentrations in fruit fresh weight of up to 220 g/g. They include mix-tures of 5-substituted resorcinols such as resorcinol-5-(12-heptadecadienyl) and resorcinol-5-(pentadecyl) (Droby et al., 1986, 1987; Prusky and Plumbley, 1992; Prusky, 1996) (Fig. 7.1). Faced by such a chemically adverse environ-ment, the fungus usually postpones its development until the concentrations of the compounds in the host decline. The capability to penetrate the host without encountering the arsenal of passive chemical resistance or triggering the weapons of active resistance is inherent in a number of fruit-infecting Colletotrichum species such as C. gloeosporioides, C. acutatum and A. alternata (Prusky and Keen, 1993). The conidia, melanized appressoria and penetra-tion pegs of these quiescent species have evolved mechanisms of physiologi-cal inactivity that enable them to pause until the host-ripening process and the decline of antifungal compounds starts, thus enabling the avoidance of host defences that exist within unripe, pre-climacteric fruits. Colletotrichum gloeosporioides is a fi eld-to-store pathogen whose conidia infect during the preharvest growth of fruits. In fruits such as mango and avocado, termina-tion of fungal quiescence is associated jointly with the climacteric production of ethylene during fruit ripening and the onset of dramatic reductions in the levels of preformed fungitoxic resorcinols (Droby et al., 1986, 1987). Flaishman
HO HOHO HO
(CH2)11
CH
CH
(CH2)3
CH3
(CH2)14
CH2
CH3
Resorcinol-5-(pentadecyl)Resorcinol-5-(12-heptadecenyl)
Fig. 7.1. Chemical structure of the 5-substituted resorcinols isolated from mango fruits.

Fruit Diseases 215
and Kolattukudy (1994) hypothesized that ethylene is involved in terminat-ing quiescence, by inducing appressoria formation and hyphal growth, which strongly suggests that Colletotrichum spp. must have coevolved with their hosts, to develop a mechanism that uses the host’s ripening hormone as a signal to reactivate the infection process. However, when Prusky and co-workers (Prusky et al., 1996) applied ethylene to unripe fruits they could not enhance the termination of quiescence as had been suggested, possibly indi-cating that ethylene is a signal that induces appressoria formation in vitro, but that does not enhance fungal growth in the fruit. Furthermore, since fun-gal infection and appressoria formation occur in the orchard, it is diffi cult to conceive how ethylene produced much later, during ripening and storage, could enhance appressoria formation on the fruit. Regardless of the mecha-nisms that may be involved in the onset and termination of quiescence, qui-escent infection appears to be a case of coevolution; it is advantageous to both pathogen and host in a natural ecosystem to allocate chemical defences to the immature fruit but not to the ripe fruit (Prusky and Keen, 1993).
Quiescence also may result from a localized host response that is often associated with an oxidative burst, i.e. production of reactive oxygen species (ROS). Localized generation of ROS during quiescence was found to be one of the earliest (within 2–3 h) detectable cytological defence responses to attempted penetration of unripe, resistant avocado fruits by C. gloeosporioides. Beno-Moualem and Prusky (2000) proposed that fungal infection of unripe fruits leads to production of ROS during quiescence, at the infection loci surrounding the germinated appressoria and their penetration hyphae. This localized ROS production would activate the synthesis of antifungal com-pounds or compounds that inhibit fungal metabolism (Ardi et al., 1998) at the infection loci, thereby enhancing and/or preserving the levels of antifungal compounds and, in turn, inhibiting fungal development and imposing quiescence.
Guetsky et al. (2005) suggested that initiation of quiescence could result from the fungal capability to induce laccase activity that modulates the metab-olism of preformed antifungal compounds and the activation of the quies-cent C. gloeosporioides that occurs during fruit ripening. Early activity of aggressive isolates in unripe fruits included increased laccase activities that resulted in early metabolism of preformed antifungal compounds leading to early appearance of symptoms.
When the levels of toxic compounds in the fruit peel decline, C. gloeospo-rioides can also enhance its colonization of ripening fruits dynamically by locally altering the pH of the fruit at the infection site to suit the increased expression of pathogenicity factors and the enzymatic arsenal (Yakoby et al., 2000; Prusky et al., 2001; Eshel et al., 2002; Prusky and Yakoby, 2003). In the pathogen C. gloeosporioides, the gene pelB encoding for the enzyme pectate lyase, a key factor for virulence, is expressed when the pH is > 6.0, a value at which decay is initiated in the tissue (Yakoby et al., 2000, 2001). Also in the case of A. alternata, another pathogen of mango fruits, the expression of the endoglucanase gene AaK1 is maximal at pH levels > 6.0 (i.e. values that are characteristic of decayed tissue). AaK1 is not expressed at the lower pH

D. Prusky et al.216
values at which the pathogen is quiescent (Eshel et al., 2002). Ambient alkal-ization by Colletotrichum is achieved by active secretion of ammonia, which is produced as a result of protease activity and deamination of amino acids. The pathogenicity of C. gloeosporioides and expression of the virulence factor PL-B both depend on raising the ambient pH (Drori et al., 2003). This modu-lation of environmental pH has been used as the basis for a new approach to disease control in mango fruits, and is discussed below.
Management
Control of postharvest anthracnose can be achieved by fi eld management, postharvest treatments or a combination of both. Management of mango anthracnose in the fi eld involves cultural and chemical practices, as well as cultivar selection.
Cultural controlSince the development of mango anthracnose is dependent on moisture or high RH, orchards ideally should be established in areas with a well-defi ned dry season, to allow for fruit development under conditions unfavourable for disease development. In the tropics, mango fl owering usually occurs dur-ing dry seasons, but anthracnose incidence of > 90% is common in fruits that develop during the rainy season (Arauz, 1999). In contrast, the incidence and severity of mango anthracnose can be close to zero in fruits that develop completely in the dry season, without the application of any other control measure (Arauz, 2000).
Considerable effort has been invested in understanding and managing mango fl owering. Flowering can be advanced by several weeks by applying potassium nitrate sprays to mature foliage (Núñez-Elisea, 1985). The growth retardant paclobutrazol, alone or followed by potassium nitrate sprays, can also be used to advance fl owering (Núñez-Elisea et al., 1993). Both treatments could contribute to the manipulation of the fl owering season to a less sensi-tive period. Field sanitation of the tree itself is diffi cult to practise. Elimina-tion of dry panicles and mummifi ed fruits is time consuming. Bagging results in reduced anthracnose severity, but it also reduces the red colour of the fruit, which could reduce consumer appeal (Hofman et al., 1997).
Resistant cultivarsAlthough all commercial mango cultivars are susceptible to anthracnose, some are less susceptible than others; ‘Tommy Atkins’ and ‘Keitt’ are less susceptible than ‘Irwin’, ‘Kent’, ‘Haden’ and ‘Edward’ (Campbell, 1992).
Chemical controlIn extreme situations, in which fruit develops entirely under disease-favouring conditions, seasonal applications of up to 25 sprays of protective and sys-temic fungicides have been reported (Dodd et al., 1997). However, few fungi-cides are approved in importing countries for use on mango. Therefore, the

Fruit Diseases 217
choice of fungicides depends on the intended destination of the fruit. Dithio-carbamate fungicides are highly effective for anthracnose control but can be used for only a few specifi c countries, whereas copper fungicides are recom-mended, but their effi cacy is lower (Arauz, 2000). Fungicides with erradicant activity for mango anthracnose include benzimidazoles and the imidazole prochloraz. Benomyl has been used under calendar-based schedules, usually in a mix with protectant fungicides, to delay the build-up of resistance in the pathogen population. Prochloraz has been used as a protectant or as an erad-icant spray (Estrada et al., 1996), but is mainly used only as a postharvest treatment.
Biological controlA number of microorganisms with in vitro or in vivo activity against C. gloeo-sporioides have been isolated (Jeffries and Koomen, 1992), but few examples had been used commercially in the fi eld until Korsten (2004) isolated Bacillusspp. from leaf and fruit surfaces, and effectively controlled anthracnose of mango. Postharvest control was achieved with semi-commercial preharvest sprays or postharvest packing house dips, sprays or ultra-low-volume appli-cations. Integrated treatments involving antagonists combined with quarter-strength or recommended dosages of fungicides such as prochloraz or sodium hypochlorite also effectively suppressed postharvest anthracnose of mango. Commercializing the antagonists in South Africa (Korsten, 2004) and in the Philippines proved to be diffi cult because of the limitations set by the local registration guidelines, and the effect of product formulation on antagonist performance in commercial applications.
Postharvest controlTraditionally, postharvest control of mango anthracnose has aimed at eradi-cation of quiescent infections on the fruit. Such eradication is achieved com-mercially by thermal and chemical treatments, or a combination of both (McMillan, 1987). Dipping fruit in hot water alone is moderately effi cient; temperatures of 50–55°C for 3–15 min have been recommended, with the higher temperatures corresponding to the shorter exposures. Fruit from Latin America entering the USA market must undergo a quarantine hot water treatment to eliminate fruit fl y (Ceratitis capitata and Anastrepha spp.) larvae; the fruit is immersed in water at 46°C for 90–120 min, depending on variety and fruit size. The effi cacy of the fruit fl y quarantine treatment varies from 60 to 85% for elimination of anthracnose infections (McGuire, 1991). Hot water treatments leave no chemical residue on the fruit and could be a good anthra-cnose control option for organically-produced mangoes or for mangoes tar-geted for markets in the USA, where no fungicides are currently approved for postharvest use. Temperature and time controls are critical, because fruits can easily be damaged by overexposure to heat. Several fungicides have been applied after harvest to control mango anthracnose. Benomyl, at rates vary-ing from 500 to 1000 g/ml, was used as a postharvest dip in the past, but its use is no longer permitted. Thiabendazole at 1000–2000 g/ml is also effec-tive, and there is interest in its registration for postharvest use with mango,

D. Prusky et al.218
as it is currently used on other fruits such as citrus. Prochloraz can be used
with fruit shipped to the European Union (EU), at rates up to 1000 g/ml; its effi cacy ranges from 65% under very high disease pressure to 94% under moderate disease pressure (Arauz, 2000). One advantage of benzimidazole fungicides (i.e. benomyl or thiabendazole) is that they are also effective in controlling stem-end rot caused by L. theobromae, which is the second most important postharvest disease of mango in tropical areas. Imidazoles such as prochloraz and imazalil are not effective against L. theobromae on mango (Estrada et al., 1996). The combination of hot water and fungicides is the most effective commercial postharvest treatment for the control of mango anthra-cnose; the rate of fungicide application and the duration of exposure to hot water are both lower, and effi cacy is higher, than either treatment used sepa-rately. The hot water and the fungicides can be applied sequentially or together. Irradiation of fruit to control anthracnose has been attempted: gamma irra-diation was not successful (Spalding and Reeder, 1986), but a short-wave infrared radiation treatment developed in South Africa resulted in anthra-cnose levels similar to those resulting from the commercial hot-water treat-ment, and was much faster and less expensive (Saaiman, 1996a).
Prusky and Keen (1993) suggested that it might be possible to prolong the period of fruit resistance and to delay the onset of anthracnose develop-ment until the fruit ripens. The climacteric rise occurs simultaneously with the production of ethylene and with changes in external and internal colour, fl avour, aroma and fi rmness, and with a reduction in fruit resistance to fun-gal attack (Prusky and Keen, 1993). Postharvest practices such as cold stor-age and controlled atmosphere storage maintain resistance to decay by delaying the ripening process, but there are two limitations to the potential benefi t of this approach in mango. First, mangoes, similarly to many other tropical and subtropical fruits, are sensitive to chilling and are injured at tem-peratures < 10–13°C, depending on the cultivar and the duration of expo-sure. Second, once the fruits are allowed to ripen under ambient conditions, disease develops normally. Nevertheless, some progress has been made towards anthracnose control through maintaining fruit resistance beyond the onset of ripening. In the last decade, a better understanding of the physi-ological basis of quiescent infections on tropical and subtropical climacteric fruits has been achieved, mainly through the work of D. Prusky and his co-workers (1993). The decline in antifungal compounds, which is brought about by oxidative processes, can be delayed so that it occurs closer to full ripeness. In avocadoes and mangoes, the concentrations of antifungal com-pounds were enhanced and, consequently, the decline in concentration was delayed, by exposure of fruit to an atmosphere containing 30% carbon diox-ide (CO2) for 24 h, or by treatment of fruit with the antioxidant compound butylated hydroxy anisole (Prusky, 1988; Kobiler et al., 1998). The delay resulted in less disease in ripe fruit.
Postharvest biological control of anthracnose in mangoes has been attempted. In an investigation with a strain of a Bacillus sp. that exhibited invitro activity against C. gloeosporioides, it was found that disease control invivo was obtained when fruits were inoculated with the bacterium 24 h prior

Fruit Diseases 219
to inoculation with the fungus, but not when fruit were inoculated with the pathogen fi rst (Korsten et al., 1992), which indicated that the quiescent phase of the fungus was not affected by the antagonist. Other approaches to disease control using biological methods included the use of a non-pathogenic strain of Colletotrichum magna that colonizes the fruit endophytically and prevents infection by C. gloeosporioides (Prusky et al., 1993); and the expression of an antifungal peptide in the yeast Saccharomyces, which controlled postharvest diseases caused by C. coccodes (Jones and Prusky, 2001).
7.3 Alternaria Rot (Black Spot)
Alternaria alternata (Fr.:Fr.) Keissl. causes black spot of mango. Conidiophores, arising singly or in small groups, are simple or branched, straight or bent, and sometimes geniculate, and pale- to mid-olivaceous or gold in colour, and smooth in texture. The conidia are oboclavata; they are borne in long chains in culture, and most have three to fi ve septa.
Pathogenesis
Germinated conidia penetrate mainly through wounds and specifi cally through lenticels of the fruits, and then become quiescent.
SymptomsAlternaria rot of mango has been increasingly reported as an important pathogen that causes blossom disease and postharvest fruit rot in ripening fruits in Australia, Egypt, India, Israel and South Africa. The symptoms are small, black circular spots that develop around the lenticels. Initially, the spots are concentrated around the stem end of the fruits, where there are large numbers of lenticels. The spots can grow and coalesce to become a sin-gle spot that covers a signifi cant part of the fruit surface. At fi rst, the decay is fi rm and does not penetrate the pulp more than 1–2 mm, but later the disease progresses into the fl esh, which darkens and becomes soft (Prusky et al., 1983). Symptoms of alternaria rot are more limited, darker and fi rmer than those of anthracnose. The former pathogen also attacks mango leaves, and symptoms can be observed throughout the year. The pathogen may also attack mango infl orescences, resulting in a signifi cant decrease in fruit set.
EpidemiologyThe main sources of inoculum are conidia released from infected leaves, twigs and infl orescences; however, Alternaria spores easily can be found in all the dry tissues of mango trees in the orchard. Conidia are transferred to the fruit by air currents and in dew runoff (Ploetz et al., 1994). Germination of conidia depends on the RH in the orchard during fruit growth. The area of quiescent Alternaria infection on mango fruit at harvest increased as the num-ber of hours of exposure to RH ≥ 80% increased over 320 h (Prusky et al.,

D. Prusky et al.220
1983). Regions with the highest potential for disease incidence are located close to the 85–90% RH isolines during the fruit growth period. The interme-diate regions lie between 75 and 85% RH, and the lowest potential risk is in the dry regions, where the prevailing RH is < 75% (Prusky et al., 1992).
Mechanisms of fungal pathogenicity and fruit resistance
As in the case of anthracnose, the mechanism of resistance against Alternaria is also related to the presence of high concentrations of preformed antimicro-bial compounds (Droby et al., 1986, 1987). Susceptibility was found to depend on the decline of the compound to subfungitoxic concentrations, which occurred faster in ‘Tommy Atkins’ than in ‘Keitt’.
Alternaria alternata can also alter the local fruit pH at the infection site dynamically, to match the increased expression of pathogenicity factors and the enzymatic arsenal (Eshel et al., 2002; Niem et al., 2007). In the case of A.alternata, the expression of the endoglucanase gene AaK1 was found to be maximal at pH levels > 6.0, which are characteristic of decayed tissue; it was not expressed at the lower pH values at which the pathogen was quiescent (Eshel et al., 2002). Alkalization of the ambient environment by Alternaria is achieved by active secretion of ammonia (Eshel et al., 2002), probably as a result of protease activity and deamination of amino acids. This modulation of environmental pH has been used as the basis of a new approach to disease control in mango fruits, and is discussed below.
Management
The losses caused by alternaria rot can be minimized by a regular fi eld-spraying programme and by postharvest fungicide treatments.
Field and postharvest chemical controlPreharvest treatments with dithiocarbamate fungicides inhibit the develop-ment of latent infection. Three sprays with the protectant fungicide maneb, starting 2 weeks after fruit set, are most effective for disease control (Prusky et al., 1983). However, since quiescent infections do not develop until after harvest and ripening, the application of a postharvest treatment by spraying the fruits on the packing line with 900 g/ml prochloraz is simpler and more effi cient than the preharvest fungicide treatment.
Control of alternaria rot is signifi cantly improved by a combination of physical and chemical treatments. The physical treatment includes a 15–20 s hot water spraying and brushing (HWB) treatment at temperatures between 50 and 55°C (Prusky et al., 1999). This new approach improved fruit quality at the same time as it reduced disease incidence. If this physical treatment is followed by a 250 g/ml prochloraz spray it can further improve disease control. Prusky et al. (1999) concluded that the type and strength of the post-harvest treatment should be varied according to the level of quiescent infection

Fruit Diseases 221
of A. alternata at the time of harvest. Although the fungicide prochloraz is very effective for the control of postharvest disease, a milder postharvest treat-ment, such as chlorine at 500 g/ml, can be applied to fruit in which a low incidence of quiescent infections is found at harvest (Prusky et al., 2002).
This postharvest physical-chemical treatment has been further improved in light of the recent fi nding that A. alternata pathogenicity may modulate the pH of the host environment to promote colonization (Eshel et al., 2002; Prusky and Yakoby, 2003; Prusky and Lichter, 2007). Application of a combination of HWB for 15–20 s, followed by spraying with 50 mM hydrochloric acid (HCl), effectively controlled alternaria rot in stored mango fruit. Similar HWB treat-ments followed by spraying with reduced concentrations of prochloraz at 45 g/ml in 50 mM HCl inhibited alternaria rot development better than treatment with HCl alone (Prusky et al., 2006). The enhancement of prochlo-raz activity in acidifi ed solutions was attributed to its enhanced solubility under acidic conditions, which resulted in an increase in the fungicidally active ingredient in the solution. This technology represents the latest devel-opment in the control of postharvest diseases, and provides a simple treat-ment for the control of diseases that alkalinize the host environment, including both alternaria rot and anthracnose.
7.4 Stem-end Rots
Stem-end rots of mango fruit present one of the most serious postharvest problems that affect this industry worldwide. They become more prominent as orchards become older. Losses increase when fruits are stored for pro-longed periods at low temperatures or when fruits ripen at temperatures > 28°C. The stem-end rot diseases are caused by a variety of fungal patho-gens including: D. dominicana (anamorph of Botryosphaeria dothidea), Dothi-orella mangiferae, L. theobromae (Botryodiplodia theobromae), Phomopsis mangiferae and Pestalotiopsis mangiferae, among which Botryosphaeria is the dominant pathogen (Darvas 1991; Johnson et al., 1992; Sangchote, 1998). Stem-end rot diseases cause heavy losses in mangoes during storage (Prusky et al., 1992; Mitra, 1997; Kobiler et al., 2001).
Pathogenesis
Johnson and co-workers (1993) have suggested that spores of the pathogen may germinate and penetrate into the host tissue through wounds, and remain as endophytes in the branches of mango trees.
SymptomsDepending on the fungus involved, a variety of symptoms may develop at the stem and as the fruit ripens. Infections by L. theobromae (B. theobromae), D.dominicana synonym Fusicoccum aesculi (anamorph of B. dothidea) and D.mangiferae, cause the formation of diffuse, water-soaked tissue that spreads

D. Prusky et al.222
from the stem end as fi ngerlike projections, which darken and coalesce into circumpedicular lesions or lobed margins (Johnson et al., 1992; Slippers et al., 2004, 2005). Necrosis remains beneath the cuticle and may penetrate through-out the fruit fl esh. Superfi cial mycelia may appear around the pedicel and ruptures of the skin or, in some cases, may penetrate through the epidermis. The ascomata of D. mangiferae are initially embedded, either separately or grouped in complex stromata, and they fi nally erupt through the epidermis and open. The spore wall is dark and thick, and becomes thinner towards the end. Conidiophores are hyaline, cylindrical, smooth and branched at the base. A watery fl uid may drain from the stem or from surface ruptures (Kor-sten, 2004). Diseased fruit could infect a whole box of fruits by direct contact, and thereby spread the pathogen in the box. In the case of injured fruit, lesions could appear at some distance from the stem. Stem-end rot diseases also have a major effect on the fl avour of the fruit.
The disease may also cause tip and branch dieback and cankers on mango trees (Ramos et al., 1991). Anamorph morphology is commonly used to iden-tify Botryosphaeria spp. (Jacobs and Rehner, 1998; Slippers et al., 2004), but the morphological distinctions among the anamorphs of some of the closely related species are not clear. Recent studies based on DNA sequence data have highlighted taxonomic groups and relationships in Botryosphaeria (Jacobs and Rehner, 1998; Slippers et al., 2004). These data, combined with morphological
characteristics, could clarify the current taxonomic confusion.Phomopsis mangiferae is a weak parasite of less economic importance than
the species above; it produces a dark, circumpeduncular lesion with defi ned edges that spreads relatively slowly but penetrates deeply into the fl esh. Fruiting bodies may develop on the affected surface. Phomopsis mangiferae can also be distinguished by a dark pinhead-size pycnidial fruiting body (Johnson et al., 1992). The lesion caused by Phomopsis may be similar to the stem-end symptoms cause by C. gloeosporioides and A. alternata. However, the lesions of the latter two diseases penetrate only to a depth of 10–20 mm.
Lesions of L. theobromae can be distinguished by their wrinkled black edges which have a velvety appearance. In the dark zone, pycnidial masses are formed (Johnson et al., 1992). In affected plants, twigs die back from the tips and into old wood, giving a scorched appearance to the limb with abun-dant gum secretions from branches, stems and the main trunk. Pestalotiopsismangiferae appears as silvery grey spots that vary in size, and are usually sharply outlined by a dark border. The fungus may appear as a member of the complex of stem-rot pathogens.
Epidemiology
The pathogens causing stem-end rots initiate inoculation in the orchards as the trees age. They are enabled to do so by mismanagement or neglect of orchards, and are affected by periods of rain and high RH at the beginning and end of the dry season (Johnson et al., 1991; Lonsdale, 1993; Johnson and Sangchote, 1994; González et al., 1999). Hyphae of the fungi colonize the

Fruit Diseases 223
fl oral parts of mango trees, develop endophytically in healthy tissue of all parts of mango plants, especially in the fruit pedicel, and remain quiescent until the fruit matures, at which stage the parasite is ready to infect through the stem end by developing in the xylem vessels of the maturing fruit (John-son et al., 1992). Pathogens can colonize fl ower parts, remain inactive pend-ing button abscission and then penetrate the stem end, as in the case of Diplodia natalensis in citrus (Nadel, 1944), of which no sexual stage has been found in nature. Fruits can be also infected at the stem by the soilborne patho-gen L. theobromae (anamorph of B. theobromae), if the fruits are placed on the ground (Johnson et al., 1992) after harvest. It is also possible that infection can result from transfer of spores by insects; decayed fruits produce volatiles, which are hypothesized to be attractants of an insect that could be a vector of the fungal spores (Nago and Matsumoto, 1994).
The range of pathogens that cause stem-end rot is infl uenced by tempera-ture, moisture stress and the nutrition status of the host. The initial systemic infection plays a crucial role in establishing blossom-blight infection. How-ever, secondary infection is apparently an even more important factor in devel-opment and incidence of soft stem-end rots. Secondary infections occur when rain washes spores away from various inoculum sources such as leaves and stems (Saaiman, 1996b). Endophytic Botryosphaeria spp. were found to be espe-cially prominent in trees continually exposed to water shortage and salt stress (Grobler et al., 2002), and Botryosphaeria spp. were found to move into develop-ing fruit, resulting in postharvest stem-end rot development (Lonsdale, 1993).
Management
Field and postharvest controlA variety of postharvest practices can be used to delay disease develop-ment. These include low-temperature storage and modifi ed- or controlled-atmosphere storage; however, the management of stem-end rots is far from being perfected.
Cultural controlJohnson et al. (1992) demonstrated that infection of mango fruit before har-vest occurred through endophytic colonization of pedicel tissue by Botry-osphaeria spp. present from previous growth fl ushes, and the possibility of pruning to promote growth fl ushing was tested as a means to reduce inocu-lum in the stem tissue from which new-season infl orescences emerged. Cooke et al. (1998) reported that the levels of endophytic organisms such as Botry-osphaeria spp. were reduced signifi cantly when a pruning programme was implemented in mango orchards as a preharvest control measure. Korsten (2006) found that prevention of water stress during fruit development and maturation, and avoidance of placing fruits on the ground suppressed dis-ease development. He also suggested that harvesting of immature fruit should be avoided; fruit should be cooled to 13°C immediately after harvest and chemical treatment, and stored in a well-ventilated place.

D. Prusky et al.224
Chemical controlPostharvest control of Botryosphaeria spp. was achieved by postharvest dip-ping, spraying or ultra-low-volume application of benomyl (where possible). Prochloraz or sodium hypochlorite also effectively suppressed postharvest stem-end rot of mango (Plan et al., 2002; Korsten, 2006). A combined treat-ment of wax and hot water (55 C) provide very effective control of most postharvest pathogens (Sangchote, 1998), but in some cases partial-vacuum infi ltration improved disease control, which suggests that control effi ciency may have been reduced because the fungicide did not reach the pathogen (Plan et al., 2002).
A treatment consisting of HWB and prochloraz followed by 2,4-dichloro-phenoxyacetic acid (2,4-D) diluted in wax, reduced side and stem-end decay by 50–70%, and improved fruit quality during prolonged storage (Kobiler et al., 2001). The best control was obtained by concentrations of 2,4-D ranging from 75 to 175 g/ml; this effi ciently controlled side rots caused by A. alternata and the stem-end rots caused by A. alternata, Phomopsis spp. and Lasiodiplodia spp. The combination of HWB, prochloraz application and 2,4-D treatment reduced the incidence of stem-end rot after 4 weeks of storage at 14°C and 7 days of shelf life at 20°C from 86 to 10% in ‘Tommy Atkins’ and from 63 to 12% in ‘Keith’ (Kobiler et al., 2001).
Biological controlBacillus licheniformis, on its own or alternated with copper oxychloride, has been evaluated as a preharvest spray treatment to control mango fruit dis-eases. Preharvest applications of B. licheniformis at 3-week intervals from fl owering until harvest controlled moderate levels of anthracnose, and of soft rot caused by Botryosphaeria, which suggests a potential treatment for com-mercial preharvest applications (Silimela and Korsten, 2006).
7.5 Other Minor Diseases
Other pathogens may attack mango fruits after harvest through occasional wounds, and cause severe diseases, such as black mould caused by Aspergillus spp. and transit rot caused by Rhizopus spp. Disease control begins in the fi eld, and is followed by postharvest sanitation, and avoidance of latex burn (stain) and mechanical injury. A hot water treatment consisting of 46°C for 60–120 min and fungicides can be used, depending on the cultivar (Spalding and Reeder, 1986). HWB at 55°C for 20 s shows good control (Prusky et al., 1999).
7.6 Conclusions
Long periods in transit, new marketing approaches for mangoes (e.g. ‘Ready to Eat’) and stringent international standards and requirements have raised the need for improved approaches to disease control, in order to preserve fruit quality. Integrated postharvest treatment has provided a more durable,

Fruit Diseases 225
consistent, sustainable and practical solution than previously available to enable producers to control postharvest diseases. Ensuring product quality and safety starts in the orchard, with proper handling conditions and is fol-lowed by optimal postharvest treatments; however, the combination of post-harvest treatments with the proper use of the cold-chain concept may provide the ultimate in favourable conditions for preserving fruit quality.
References
Arauz, L.F. (1999) Optimización económica del combate de enfermedades en cultivos: un ejemplo basado en la antracnosis del fruto del mango. (Abstract). In: XI Con-greso Nacional Agron. Recursos Naturales, V Congreso Nacional Entomol., IV Congreso Nacional Fitopatol., III Congreso Nacional Suelos I Congreso Nacional Ext. Agríc. Forestal. Vol. II. San José, Costa Rica, p. 61.
Arauz, L.F. (2000) Mango anthracnose: economic impact and current options for inte-grated management. Plant Disease 84, 600–611.
Ardi, R., Kobiler, I., Jacoby, B., Keen, N.T. and Prusky, D. (1998) Involvement of epicat-echin biosynthesis in the activation of the mechanism of resistance of avocado fruits to Colletotrichum gloeosporioides. Physiological and Molecular Plant Pathology53, 269–285.
Bailey, J.A. (1997) rDNA sequencing: an essential guide to the taxonomy and diagnosis of Colletotrichum. In: Dehne, H.-W., Adam, G., Diekmann, M., Frahm, J., Mauler-Machnik, A. and van Halteren, P. (eds) Diagnosis and Identifi cation of Plant Patho-gens, Vol. 11, Developments in Plant Pathology. Kluwer Academic, Dordrecht, the Netherlands, pp. 47–51.
Banik, A.K., Kaiser, S.A.K.M. and Dhua, R.S. (1998) Infl uence of temperature and hu-midity on the growth and development of Colletotrichum gloeosporioides and Diplodia natalensis causing postharvest fruit rot of mango. Advances in Plant Sci-ences 11, 51–57.
Beno-Moualem, D. and Prusky, D. (2000) Early events during quiescent infection devel-opment by Colletotrichum gloeosporioides in unripe avocado fruits. Phytopathology 90, 553–559.
Campbell, R.J. (ed.) (1992) A Guide to Mangoes in Florida. Fairchild Tropical Garden, Miami, Florida.
Coates, L.M., Muirhead, I.F., Irwin, J.A.G. and Gowanlock, D.H. (1993) Initial infection processes by Colletotrichum gloeosporioides on avocado fruit. Mycological Re-search 97, 1363–1370.
Cooke, A.W., van der Kruyssen, A. and Johnson, G.I. (1998) The effect of commercial pruning on colonization of mango by endophytic Dothiorella species. In: Champ, B.R., Highley, E. and Johnson, G.I. (eds) Post-harvest Handling of Tropical Fruit. Australian Centre for International Agricultural Research (ACIAR), Canberra, pp. 128–130.
Darvas, J.M. (1991) Dothiorella dominicana, a new mango pathogen in South Africa. Phytophylactica 23, 295–298.
de Jong, J.C., McCormark, B.J., Smirnoff, N. and Talbot, N.J. (1997) Glycerol generates turgor in rice blast. Nature 389, 244–245.
Dickman, M.B., Patil, S.S. and Kolattukudy, P.E. (1982) Purifi cation, characterization and role in infection of an extracellular cutinolytic enzyme from Colletotrichum gloeo-sporioides Penz. on Carica papaya L. Physiological Plant Pathology 20, 333−347.

D. Prusky et al.226
Dodd, J.C., Estrada, A.B., Matcham, J., Jeffries, P. and Jeger, M.J. (1991) The effect of climatic factors on Colletotrichum gloeosporioides, causal agent of mango anthra-cnose, in the Philippines. Plant Pathology 40, 568–575.
Dodd, J.C., Prusky, D. and Jeffries, P. (1997) Fruit diseases. In: Litz, R.E. (ed.) The Mango: Botany, Production and Uses. CAB International, Wallingford, UK, pp. 257–280.
Droby, S., Prusky, D., Jacoby, B. and Goldman, A. (1986) Presence of an antifungal com-pound and its relation in the latency of Alternaria alternata in unripe peel of mango fruits. Physiological and Molecular Plant Pathology 29, 173–183.
Droby, S., Prusky, D. and Jacoby, D. (1987) Induction of an antifungal agent in unripe mango fruits to demonstrate their involvement in latent infections of Alternaria al-ternata. Physiological and Molecular Plant Pathology 30, 285–292.
Drori, N., Kramer-Haimovich, H., Rollins, J., Dinoor, A., Okon, Y., Pines, O. and Prusky, D. (2003) A combination of external pH and nitrogen assimilation affect secretion of the virulence factor pectate lyase by C. gloeosporioides. Applied and Environ-mental Microbiology 69, 3258–3262.
Eshel, D., Miara, I., Ailing, T., Dinoor, A. and Prusky, D. (2002) pH regulates endogluca-nase expression and virulence of Alternaria alternata in persimmon fruits. Molecular Plant Microbe Interactions 15, 774–779.
Estrada, A.B., Dodd, J.C. and Jeffries, P. (1993) Effect of environment on the in vitro growth and development of Colletotrichum gloeosporioides isolates from the Phil-ippines. Acta Horticulturae 341, 360–370.
Estrada, A.B., Jeffries, P. and Dodd, J.C. (1996) Field evaluation of a predictive model to control anthracnose disease of mango in the Philippines. Plant Pathology 45, 294–301.
Fitzell, R.D. and Peak, C.M. (1984) The epidemiology of anthracnose disease of mango: inoculum sources, spore production and dispersal. Annals of Applied Biology 104, 53–59.
Fitzell, R.D., Peak, C.M. and Darnell, R.E. (1984) A model for estimating infection levels of anthracnose disease of mango. Annals of Applied Biology 104, 451–458.
Flaishman, M.A. and Kolattukudy, P.E. (1994) Timing of fungal invasion using host’s rip-ening hormone as a signal. Proceedings of the National Academy of Sciences USA91, 6579–6583.
Freeman, S., Katan, T. and Shabi, E. (1998) Characterization of Colletotrichum speciesresponsible for anthracnose diseases of various fruits. Plant Disease 82, 596–605.
Gantotti, B.V. and Davis, M.J. (1993) Pectic zymogram analysis for characterizing genetic diversity of the mango anthracnose pathogen. Acta Horticulturae 341, 353–359.
González, E., Umaña, G. and Arauz, L.F. (1999) Fluctuación poblacional de Botryodi-plodia theobromae Pat. en mango. Agronomía Costarricense 23, 21–29.
Grobler, L., Swart, S.H. and Korsten, L. (2002) Preliminary survey of a mango nursery and associated orchards to establish the status of endophytic Botryosphaeria spp. South African Mango Growers’ Association Yearbook 22, 104–106.
Guetsky, R., Kobiler, I., Wang, X., Perlman, N., Gollop, N., Avila-Quezada, G., Hadar, I. and Prusky, D. (2005) Metabolism of the fl avonoid epicatechin by laccase of Col-letotrichum gloeosporioides and its effect on pathogenicity on avocado fruits. Phy-topathology 95, 1341–1348.
Hawthorne, B.T. (1989) Effects of cultural practices on the incidence of storage rots in Cucurbita spp. New Zealand Journal of Crop and Horticultural Science 17, 49–54.
Hofman, P.J., Smith, L.G., Joyce, D.C., Johnson, G.I. and Meiburg, G.F. (1997) Bagging of mango (Mangifera indica cv. ‘Keitt’) fruit infl uences fruit quality and mineral composition. Postharvest Biology and Technology 12, 83–91.

Fruit Diseases 227
Howard, R.J., Ferrari, M.A., Roach, D.H. and Money, N.P. (1991) Penetration of hard substrates by a fungus employing enormous turgor pressures. Proceedings of the National Academy of Sciences USA 88, 11281–11284.
Jacobs, K.A. and Rehner, S.A. (1998) Comparison of cultural and morphological charac-ters and ITS sequences in anamorphs of Botryosphaeria and related taxa. Mycolo-gia 90, 601–610.
Jeffries, P. and Koomen, I. (1992) Strategies and prospects for biological control of dis-eases caused by Colletotrichum. In: Bailey, J.A. and Jeger, M.J. (eds) Colletotrichum: Biology, Pathology and Control. CAB International, Wallingford, UK, pp. 337–357.
Johnson, G.I. and Sangchote, S. (1994) Control of post-harvest diseases of tropical fruits: challenges for the 21st century. In: Champ, B.R., Highley, E. and Johnson, G.I. (eds) Post-harvest Handling of Tropical Fruit. Australian Centre for International Agricul-tural Research (ACIAR), Canberra, pp. 140–161.
Johnson, G.I., Mead, A.J., Cooke, A.W. and Dean, J.R. (1991) Mango stem end rot patho-gens – infection levels between fl owering and harvest. Annals of Applied Biology119, 465–473.
Johnson, G.I., Mead, A.J., Cooke, A.W. and Dean, J.R. (1992) Mango stem end rot patho-gens – fruit infection by endophytic colonization of the infl orescence and pedicel. Annals of Applied Biology 120, 225–234.
Johnson, G.I., Cooke, T. and Mead, A.J. (1993) Infection and quiescence of mango stem-end rot pathogens. Acta Horticulturae 341, 329–336.
Jones, R.W. and Prusky, D. (2001) Expression of an antifungal peptide in Saccharomy-ces: a new approach for biocontrol of the postharvest disease caused by Colletotri-chum coccodes. Phytopathology 92, 33–37.
Kobiler, I., Reved, R., Artez, L. and Prusky, D. (1998) Antifungal compounds regulating quiescent disease in mango. 1998. In: Johnson, G.I., Higley, E. and Joyce, D.C. (eds) Australian Centre for International Agricultural Research (ACIAR) Proceedings80. ACIAR, Canberra, pp. 109–114.
Kobiler, I., Shalom, Y., Roth, I., Akerman, M., Vinokour, Y., Fuchs, Y. and Prusky, D. (2001) Effect of 2,4-dichlorophenoxyacetic acid on the incidence of side and stem-end rots in mango fruits. Postharvest Biology and Technology 23, 23–32.
Korsten, L. (2004) Biological control in South Africa: can it provide a sustainable solu-tion for control of fruit diseases? South African Journal of Botany 70, 128–139.
Korsten, L. (2006) Advances in control of postharvest diseases in tropical fresh produce. International Journal of Postharvest Technology and Innovation 1, 48–61.
Korsten, L., Lonsdale, J.H., De Villiers, E.E. and De Jager, E.S. (1992) Pre-harvest con-trol of mango diseases. South African Mango Growers’ Association Yearbook 12, 72–78.
Latunde-Dada, A.O., O’Connell, R.J., Nash, C. and Lucas, J.A. (1999) Stomatal penetra-tion of cowpea (Vigna unguiculata) leaves by a Colletotrichum species causing la-tent anthracnose. Plant Pathology 48, 777–785.
Liu, Z.M. and Kolattukudy, P.E. (1998) Identifi cation of a gene product induced by hard-surface contact of Colletotrichum gloeosporioides conidia as a ubiquitin-conjugating enzyme by yeast complementation. Journal of Bacteriology 180, 3592–3597.
Lizada, C. (1993) Fruit handling systems in developing countries. In: Postharvest Han-dling of Tropical Fruits. Australian Centre for International Agricultural Research (ACIAR) Publication 50. ACIAR, Canberra, p. 102.
Lonsdale, J.H. (1993) Preliminary results on the mode of infection of Nattrassia mangifer-ae in mango. South African Mango Growers’ Association Yearbook 13, 97–99.
McGuire, R.G. (1991) Concomitant decay reductions when mangoes are treated with heat to control infestations of Caribbean fruit fl ies. Plant Disease 75, 946–949.

D. Prusky et al.228
McMillan, R.T., Jr (1987) Effectiveness of various postharvest treatments for mango de-cay control. Proceedings of the Florida State Horticultural Society 100, 7–9.
Mitra, S. (1997) Mango. In: Mitra, S.K. and Baldwin, E.A. (eds) Postharvest Physiology and Storage of Tropical and Subtropical Fruits. CAB International, Wallingford, UK, pp. 122–185.
Money, N.P. and Howard, R.J. (1996) Confi rmation of a link between fungal pigmenta-tion, turgor pressure and pathogenicity using a new method of turgor measurement. Fungal Genetics and Biology 20, 217–227.
Nadel, M. (1944) Anatomical study of the button of Shamouti oranges in relation to stem-end rot. Palestine Journal of Botany, Rehovot Series 4, 166–170.
Nago, H. and Matsumoto, M. (1994) An ecological role of volatiles produced by Lasio-diplodia theobromae. Bioscience Biotechnology Biochemistry 58, 1267–1272.
Niem, J., Miyara, I., Reuveni, M., Ettedy, Y., Flaishman, M. and Prusky, D. (2007) Core rot development in Red Delicious apples is affected by locule susceptibility to Al-ternaria alternata. Phytopathology 97, 1415–1421.
Núñez-Elisea, R. (1985) Flowering and fruit set of a monembryonic and a polyembry-onic mango as infl uenced by potassium nitrate sprays and shoot decapitation. Pro-ceedings of the Florida State Horticultural Society 98, 179–183.
Núñez-Elisea, R., Davenport, T.L. and Caldeira, M.L. (1993) Bud initiation and morpho-genesis in ‘Tommy Atkins’ mango as affected by temperature and triazole growth retardants. Acta Horticulturae 341, 192–198.
Pain, N.A., Green, J.R., Jones, G.L. and O’Connell, R.J. (1996) Composition and organi-sation of extracellular matrices around germ tubes and appressoria of Colletotri-chum lindemuthianum. Protoplasma 190, 119–130.
Plan, M.R.R., Joyce, D.C., Ogle, H.J. and Johnson, G.I. (2002) Mango stem-end rot (Botryosphaeria dothidea) disease control by partial-pressure infi ltration of fungi-cides. Australian Journal of Experimental Agriculture 42, 625–629.
Ploetz, R.C. (1994) Anthracnose. In: Ploetz, R.C., Zentmyer, G.A., Nishijima, W.T., Rohrbach, K.G. and Ohr, H.D. (eds) Compendium of Tropical Fruit Diseases. Amer-ican Phytopathological Society (APS) Press, St Paul, Minnesota, pp. 35–36.
Ploetz, R.C. and Prakash, O. (1997) Foliar, fl oral and soilborne diseases. In: Litz, R.E. (ed.) The Mango: Botany, Production and Uses. CAB International, Wallingford, UK, pp. 281–325.
Ploetz, R.C., Zentmyer, G.A., Nishijima, W.T., Rohrbach, K.G. and Ohr, H.D. (1994) Mango. In: Ploetz, R.C., Zentmyer, G.A., Nishijima, W.T., Rohrbach, K.G. and Ohr, H.D. (eds) Compendium of Tropical Fruit Diseases. American Phytopathological Society (APS) Press, St Paul, Minnesota, pp. 33–40.
Ploetz, R.C., Benscher, D., Vázquez, A., Colls, A., Nagel, J. and Schaffer, B. (1996) Man-go decline: research in Florida on an apparently wide-spread disease complex. Acta Horticulturae 455, 547–553.
Podila, G.K., Rogers, L.M. and Kolattukudy, P.E. (1993) Chemical signals from avocado surface wax trigger germination and appressorium formation in Colletotrichum gloeosporioides. Plant Physiology 103, 267–272.
Prusky, D. (1988) The use of antioxidants to delay the onset of anthracnose and stem end rot in avocado fruits after harvest. Plant Disease 72, 381–384.
Prusky, D. (1996) Pathogen quiescence in postharvest diseases. Annual Reviews of Phy-topathology 34, 413–434.
Prusky, D. and Keen, N.T. (1993) Involvement of preformed antifungal compounds in the resistance of subtropical fruits to fungal decay. Plant Disease 77, 114–119.
Prusky, D. and Lichter, A. (2007) Activation of quiescent infections by postharvest patho-gens during transition from the biotrophic to the necrotrophic stage. FEMS Micro-biological Letters 268, 1–8.

Fruit Diseases 229
Prusky, D. and Plumbley, R.A. (1992) Quiescent infections of Colletotrichum in tropical and subtropical fruits. In: Bailey, J.A. and Jeger, M.J. (eds) Colletotrichum: Biology, Pathology and Control. CAB International, Wallingford, UK, pp. 289–307.
Prusky, D. and Yakoby, N. (2003) Pathogenic fungi: leading or led by ambient pH? Molecular Plant Pathology 4, 509–516.
Prusky, D., Fuchs, Y. and Yanko, U. (1983) Assessment of latent infections as a basis for control of postharvest disease of mango. Plant Disease 67, 816–818.
Prusky, D., Plumbley, R. and Kobiler, I. (1991) Modulation of natural resistance of avo-cado fruits to Colletotrichum gloeosporioides by CO2. Physiological and Molecular Plant Pathology 39, 325–334.
Prusky, D., Gat, Z. and Burd, P. (1992) Effect of relative humidity during mango growth on the incidence of quiescent infections of Alternaria alternata. Plant Disease 77, 249–252.
Prusky, D., Freeman, S., Rodriguez, R.J. and Keen, N.T. (1993) A nonpathogenic mutant strain of Colletotrichum magna induces resistance to C. gloeosporioides in avocado fruits. Molecular Plant-Microbe Interactions 7, 326–333.
Prusky, D., Wattad, C. and Koliber, I. (1996) Effect of ethylene on the activation of qui-escent infections of Colletotrichum gloeosporioides in avocado fruits. MolecularPlant-Microbe Interactions 9, 864–868.
Prusky, D., Fuchs, I., Kobiler, I., Roth, I., Weksler, A., Shalom, Y., Falik, E., Zauberman, G., Pesis, E., Akerman, M., Yekutiely, O., Weisblum, A., Regev, R. and Artes, L. (1999) Effect of hot water brushing, prochloraz treatment and waxing on the inci-dence of black spot decay caused by Alternaria alternata in mango fruit. Postharvest Biology and Technology 15, 165–174.
Prusky, D., McEvoy, J.L., Leverentz, B. and Conway, L.S. (2001) Local modulation of host pH by Colletotrichum species as a mechanism to increase virulence. MolecularPlant Microbe Interactions 14, 1105–1113.
Prusky, D., Shalom, Y., Kobiler, I., Akerman, M. and Fuchs, Y. (2002) The level of quies-cent infection of Alternaria alternata in mango fruits at harvest determines the post-harvest treatment applied for the control of rots during storage. Postharvest Biology and Technology 25, 339–347.
Prusky, D., Kobiler, I., Akerman, M. and Miyara, I. (2006) Effect of acidic solutions and acidic prochloraz on the control of postharvest decay caused by Alternaria alter-nata in mango and persimmon fruits. Postharvest Biology and Technology 42, 134–141.
Ramos, L.J., Lara, S.P., McMillan, R.T. Jr and Narayanan, K.R. (1991) Tip dieback of mango (Mangifera indica) caused by Botryosphaeria ribis. Plant Disease 75, 315–318.
Saaiman, W.C. (1996a) Short wave infra-red radiation as an alternative to the hot water bath to control postharvest decay in mangoes. Acta Horticulturae 455, 773–779.
Saaiman, W.C. (1996b) Preliminary report on the time of infection of the soft brown rot pathogen Nattrassia mangiferae in mango. South African Mango Growers’ Associa-tion Yearbook 16, 55–57.
Sangchote, S. (1998) Fruit rots of mangosteen and their control. In: Champ, B.R., High-ley, E. and Johnson, G.I. (eds) Post-harvest Handling of Tropical Fruit. Australian Centre for International Agricultural Research (ACIAR), Canberra, pp. 81–86.
Sherriff, C., Whelan, M.J., Arnold, G.M., Lafay, J.-F., Brygoo, Y. and Bailey, J.A. (1994) Ribosomal DNA sequence analysis reveals new species groupings in the genus Col-letotrichum. Experimental Mycology 18, 121–138.
Silimela, M. and Korsten, L. (2006) Evaluation of pre-harvest Bacillus licheniformissprays to control mango fruit diseases. Crop Protection 26, 1471–1481.
Slippers, B., Crous, P.W., Denman, S., Coutinho, T.A., Wingfi eld, B.D. and Wingfi eld, M.J. (2004) Combined multiple gene genealogies and phenotypic characters differentiate

D. Prusky et al.230
several species previously identifi ed as Botryosphaeria dothidea. Mycologia 96, 83–101.
Slippers, B., Johnson, G.I., Crous, P.W., Coutinho, T.A., Wingfi eld, B.D. and Wingfi eld, M.J. (2005) Phylogenetic and morphological re-evaluation of the Botryosphaeriaspecies causing diseases of Mangifera indica. Mycologia 97, 99–110.
Spalding, D.H. and Reeder, W.F. (1986) Decay and acceptability of mangoes treated with combinations of hot water, imazalil, and g-radiation. Plant Disease 70, 1149–1151.
Sreenivasaprasad, S., Mills, P.R., Meehan, B.M. and Brown, A.E. (1996) Phylogeny and systematics of 18 Colletotrichum species based on ribosomal DNA spacer sequenc-es. Genome 39, 499–512.
Thompson, J.F., Mitchell, F.G. and Kasmire, R.F. (2002) Cooling horticultural commodi-ties. In: Kader, A.A. (ed.) Postharvest Technology of Horticultural Crops. University of California Press, Davis, California, pp. 97–112.
Yakoby, N., Kobiler, I., Dinoor, A. and Prusky D. (2000) pH regulation of pectate lyase secretion modulates the attack of Colletotrichum gloeosporioides on avocado fruits. Applied and Environmental Microbiology 66, 1026–1030.
Yakoby, N., Zhou, R., Kobiler, I., Dinoor, A. and Prusky, D. (2001) Development of Col-letotrichum gloeosporioides restriction enzyme-mediated integration mutants as biocontrol agents against anthracnose disease in avocado fruits. Phytopathology91, 143–148.

© CAB International 2009. The Mango, 2nd Edition: Botany, Production and Uses(ed. R.E. Litz) 231
8 Foliar, Floral and Soilborne Diseases
R.C. Ploetz1 and S. Freeman2
1University of Florida, Florida, USA 2Agricultural Research Organization, Bet Dagan, Israel
8.1 Introduction 2328.2 Foliar and Floral Diseases 232
Algal leaf spot 232Alternaria leafspot 233Anthracnose 235Apical necrosis 240Bacterial black spot (black canker) 241Black-banded disease 245Black mildew, sooty blotch and sooty mould 246Blossom blight 248Decline disorders 249Galls and scaly bark 256Grey leafspot 259Leaf blight 260Malformation 261Parasitic plants 270Phoma blight 270Phoma leafspot 270Pink disease 271Powdery mildew 271Scab 274Seca and sudden decline 274Stigmina leafspot 279
8.3 Soilborne Diseases 281Black root rot 281Nematode damage 281Phytophthora diseases 282Root rot and damping off 284Sclerotium rot 285Verticillium wilt 286White root disease 286
8.4 Conclusions 289

R.C. Ploetz and S. Freeman232
8.1 Introduction
Diseases affect all tissues and developmental phases of mango. Mango dis-eases have been reviewed in the fi rst edition of this book (Litz, 1997) and by Singh (1968), Cook (1975), Snowdon (1990), Ploetz et al. (1994) and Ploetz(2003). Lim and Khoo (1985), Prakash and Srivastava (1987) and Ridgeway (1989) have also reviewed this subject. This chapter focuses on foliar, fl oral and soilborne diseases of mango. Each is discussed with respect to signifi -cance, geographical distribution, history, symptoms, causal agent(s), epide-miology and management. These diseases are caused mainly by eukaryotes (Domain Eukaryota) among which the true fungi, Eumycota (Ascomycota and Basidiomycota), predominate. Other less important eukaryotes include the fungus-like oomycetes (Oomycota), nematodes (Metazoa), and parasitic plants and green algae (Plantae). With one exception, relatively minor pathogens of mango are prokaryotes in the Domain Eubacteria; all are Gram-negative -proteobacteria. None of these diseases is caused by other plant-pathogenic
eukaryotes (protozoa), prokaryotes ( - and -proteobacteria, Mollicutes and Firmicutes), or the nucleic acid-based pathogens, the viruses and viroids.
8.2 Foliar and Floral Diseases
Diseases above ground are the most important and conspicuous problems on mango. Since many of the pathogens that incite foliar diseases also affect panicles, diseases on each are combined in this section, as are a few diseases that affect the branches and trunks of mature trees.
Algal leaf spot
Algal leaf spot, also known as red rust, is caused by a parasitic green alga, Cephaleuros virescens Kunze (synonyms: Cephaleuros parasiticus Karst, Ce-phaleuros mycoidea Karst) (family Trentepohliaceae, division Chlorophyta) (Lim and Khoo, 1985). It is a common problem on mango and many other tropical and subtropical plants (Joubert and Rijkenberg, 1971). Although algal leaf spot can cause serious problems on tea Camellia sinensis (L.) O. Kuntz, black pepper, Piper nigrum L., and other important crops, it is usually debilitating on mango only in poorly managed orchards (Lim and Khoo, 1985). In the latter cases, abiotic or biotic stresses, such as mites, insects and other foliar diseases, can increase the severity of this disease.
Conspicuous symptoms of algal leafspot are orange to rust-coloured, velutinous patches on both leaf surfaces (Plate 43) (Lim and Khoo, 1985). They are initially 5–8 mm in diameter, but can merge to involve large, irregu-lar sections of the leaf. They later assume a dull, greyish green colour, and eventually become bleached patches. The alga can also affect twigs and branches, causing the bark to crack as the pathogen’s fi laments extend into the host cortical tissues. The orange tufts produced by C. virescens are the algal thallus located beneath the host cuticle. They produce erect cells, some
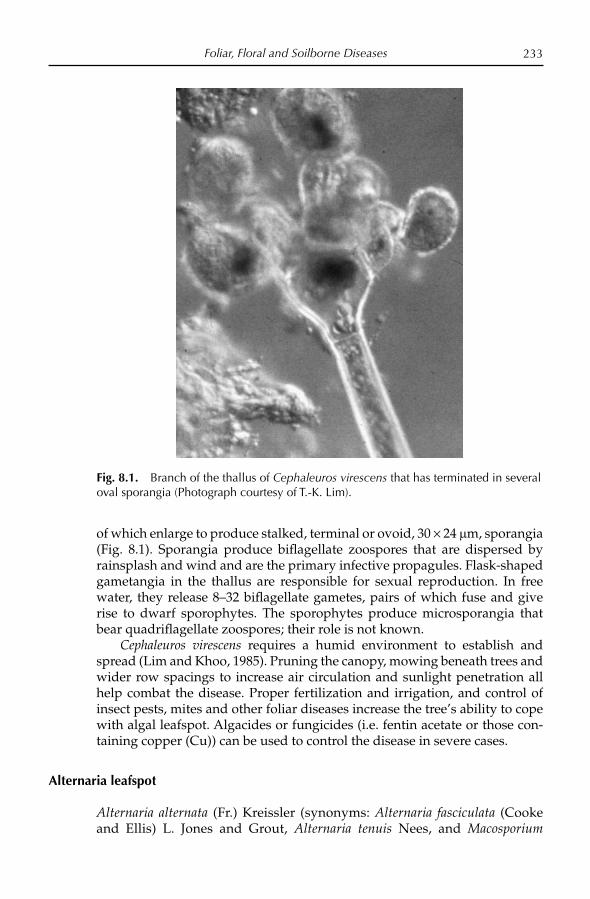
Foliar, Floral and Soilborne Diseases 233
of which enlarge to produce stalked, terminal or ovoid, 30 × 24 m, sporangia (Fig. 8.1). Sporangia produce bifl agellate zoospores that are dispersed by rainsplash and wind and are the primary infective propagules. Flask-shaped gametangia in the thallus are responsible for sexual reproduction. In free water, they release 8–32 bifl agellate gametes, pairs of which fuse and give rise to dwarf sporophytes. The sporophytes produce microsporangia that bear quadrifl agellate zoospores; their role is not known.
Cephaleuros virescens requires a humid environment to establish and spread (Lim and Khoo, 1985). Pruning the canopy, mowing beneath trees and wider row spacings to increase air circulation and sunlight penetration all help combat the disease. Proper fertilization and irrigation, and control of insect pests, mites and other foliar diseases increase the tree’s ability to cope with algal leafspot. Algacides or fungicides (i.e. fentin acetate or those con-taining copper (Cu)) can be used to control the disease in severe cases.
Alternaria leafspot
Alternaria alternata (Fr.) Kreissler (synonyms: Alternaria fasciculata (Cooke and Ellis) L. Jones and Grout, Alternaria tenuis Nees, and Macosporium
Fig. 8.1. Branch of the thallus of Cephaleuros virescens that has terminated in several oval sporangia (Photograph courtesy of T.-K. Lim).
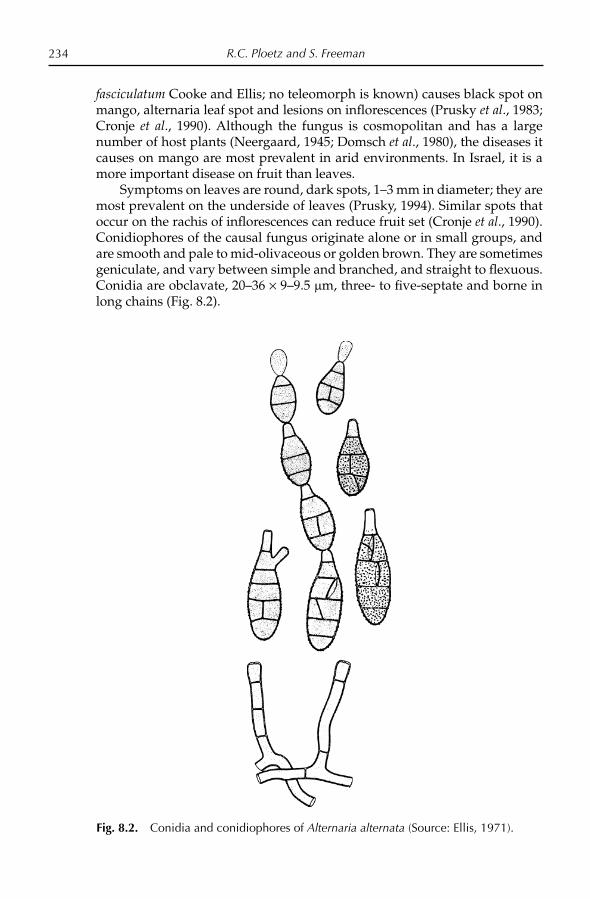
R.C. Ploetz and S. Freeman234
Fig. 8.2. Conidia and conidiophores of Alternaria alternata (Source: Ellis, 1971).
fasciculatum Cooke and Ellis; no teleomorph is known) causes black spot on mango, alternaria leaf spot and lesions on infl orescences (Prusky et al., 1983; Cronje et al., 1990). Although the fungus is cosmopolitan and has a large number of host plants (Neergaard, 1945; Domsch et al., 1980), the diseases it causes on mango are most prevalent in arid environments. In Israel, it is a more important disease on fruit than leaves.
Symptoms on leaves are round, dark spots, 1–3 mm in diameter; they are most prevalent on the underside of leaves (Prusky, 1994). Similar spots that occur on the rachis of infl orescences can reduce fruit set (Cronje et al., 1990). Conidiophores of the causal fungus originate alone or in small groups, and are smooth and pale to mid-olivaceous or golden brown. They are sometimes geniculate, and vary between simple and branched, and straight to fl exuous. Conidia are obclavate, 20–36 × 9–9.5 m, three- to fi ve-septate and borne in long chains (Fig. 8.2).

Foliar, Floral and Soilborne Diseases 235
Certain antifungal compounds are associated with the latency of A. alter-nata infections on fruit (Droby et al., 1986, 1987). Whether the same com-pounds are formed in leaves and infl orescences has not been reported. Infected leaves, twigs, infl orescences and leaf litter are signifi cant sources of inocu-lum for fruit infection. Conidia of A. alternata are dispersed by air currents (Prusky, 1994). Several different fungicides and combined treatments are effective against the diseases that are caused by this pathogen. Postharvest development of alternaria fruit rot during storage is usually prevented by a combination of hot water brushing in combination with prochloraz at 225 g/ml (Prusky et al., 2002).
The fi nding that A. alternata alkalinizes the host pH (Eshel et al., 2002) prompted investigations into modulating environmental pH with acid treat-ments to reduce fungal colonization. Spore germination and germ-tube elon-gation of A. alternata in vitro were inhibited by, respectively, 95 and 65% by exposure to 1.25 mM hydrochloric acid (HCl). Germination was completely inhibited by 2.5 mM HCl (Prusky et al., 2006). Acidifi ed solutions containing 45 g/ml prochloraz inhibited alternaria rot development better than treat-ment with HCl alone (Eshel et al., 2002). These results have not been extended to control foliar and fl oral diseases that are caused by this pathogen.
Anthracnose
Anthracnose is the most important disease of mango (Cook, 1975; Lim and Khoo, 1985; Ploetz, 2003), particularly on fruit in humid, high rainfall envi-ronments. It is usually replaced by, and is less important than, other diseases in dry production areas. Anthracnose can also be a serious problem on foli-age and fl owers. Crowded and moist conditions in nurseries can result in signifi cant damage to young leaves and, in extreme cases, new orchards have been devastated (Bose et al., 1973). Blossom blight, which has been attributed to one of the anthracnose agents but is also caused by other fungi, is covered separately below.
SymptomsOn panicles, necrotic fl owers abscise leaving persistent peduncles. Small, cir-cular dark spots also develop on pedicles and peduncles. Lesions may enlarge and coalesce to form large patches of necrotic, dark brown tissue. With suf-fi cient rainfall, salmon- to orange-coloured fructifi cations of the pathogen develop on the affected tissues. On leaves, lesions are dark brown and sur-rounded by chlorotic haloes, have irregular, rounded margins, and are not delimited by veins (Fig. 8.3). Lesions are 0.5–1.0 cm in diameter on mature leaves, but can expand on young leaves. Eventually, large, necrotic patches can develop that deteriorate and fall from the leaf giving it a tattered appear-ance. Although different mango cultivars are known to vary considerably in their resistance to anthracnose on fruit, reports on the foliar and fl oral resis-tance of different cultivars and whether resistances of the various organs are related have not been published.

R.C. Ploetz and S. Freeman236
AetiologyIn most production regions, anthracnose is caused by the ascomycete fungus, Colletotrichum gloeosporioides Penz. (teleomorph: Glomerella cingulata (Stonem.) Spauld. and Schrenk.) (Cook, 1975; Snowdon, 1990; Ploetz, 2003). In Austra-lia, Florida USA, India, Japan and Taiwan, Colletotrichum acutatum Simmonds (teleomorph: Glomerella acutata) plays a minor role (Fitzell, 1979; Prakash, 1990; Weng and Chuang, 1995; Taba et al., 2004; Tarnowski, unpublished). In Colombia, Colletotrichum boninense J. Moriwaki, Toy. Sato and T. Tsukiboshi has been reported (Afanador-Kafuri et al., 2003).
Colletotrichum spp. cause diseases on several subtropical and tropical hosts (Jeffries et al., 1990; Freeman et al., 1998). Cultures of the fungus on potato dextrose agar (PDA) are greyish white to dark grey and usually pro-duce an aerial mycelium ranging from a thick mat to sparse tufts (Holliday, 1980). Conidia are hyaline, unicellular and either cylindrical with obtuse ends or ellipsoidal with a rounded apex and a narrow, truncate base. They are 7–20 × 2.5–5 m, and are formed on hyaline to faintly brown conidiophores in
Fig. 8.3. Foliar symptoms of anthracnose (Photograph courtesy of S. Freeman).

Foliar, Floral and Soilborne Diseases 237
acervuli that are irregular in shape and about 500 m in diameter. Setae are 4–8 × 200 m, one- to four-septate, brown and slightly swollen at the base and tapered at the apex. Hyphopodia have been used to distinguish isolates of C.gloeosporioides and C. acutatum (Du et al., 2005), but provided ambiguous results in Florida (Palmateer et al., 2007).
Characterization and taxonomic identifi cation of Colletotrichum spp. has relied on morphology and host range (Freeman et al., 1998; Du et al., 2005). In general, C. gloeosporioides (Fig. 8.4) produces longer and narrower conidia than C. acutatum (Fig. 8.5), as well as circular vs lobed hyphopodia. However, variability in culture and host range has made these criteria unreliable for species identifi cation (Adaskaveg and Hartin, 1997; Freeman et al., 1998). Col-letotrichum gloeosporioides and C. acutatum are species complexes with a large number of hosts that are so broadly defi ned that the names are of ‘limited use to the taxonomist or plant health practitioner’ (Du et al., 2005). Several molec-ular tools have been implemented to differentiate within and among Col-letotrichum spp., including: species-specifi c polymerase chain reaction (PCR) primers, random amplifi ed polymorphic DNA (RAPD) and arbitrarily primed PCR (Freeman and Rodriguez, 1995; Afanador-Kafuri et al., 2003); A+T-rich DNA analyses (Freeman et al., 1993); sequence analyses of the inter-nal transcribed spacer (ITS) regions of ribosomal DNA (rDNA) (Sreenivasap-rasad et al., 1996; Freeman et al., 2001; Afanador-Kafuri et al., 2003) and MAT1-2 mating type sequences (Du et al., 2005).
(a)
(b)
(c)
(d)
(g)
(e)
(f )
10μ
50μ
Fig. 8.4. (a) Acervulus and emergent setae, (b) conidiophores, (c) conidia, (d) perithecium, (e) asci, (f) ascospores and (g) appressoria at hyphal termini of Glomerella cingulata (anamorph:Colletotrichum gloeosporioides) (Source: from Commonwealth Mycological Institute (CMI) description no. 315).
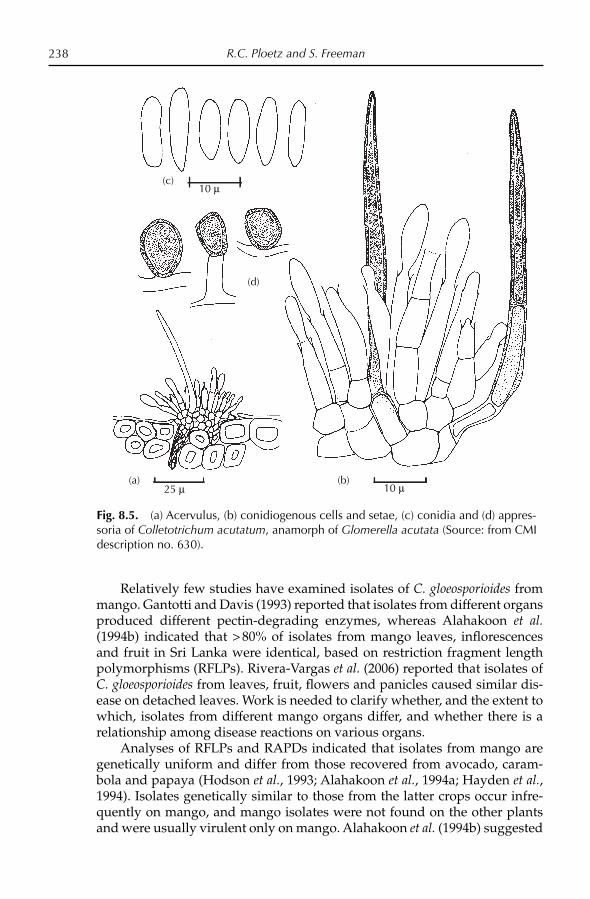
R.C. Ploetz and S. Freeman238
Relatively few studies have examined isolates of C. gloeosporioides from mango. Gantotti and Davis (1993) reported that isolates from different organs produced different pectin-degrading enzymes, whereas Alahakoon et al. (1994b) indicated that > 80% of isolates from mango leaves, infl orescences and fruit in Sri Lanka were identical, based on restriction fragment length polymorphisms (RFLPs). Rivera-Vargas et al. (2006) reported that isolates of C. gloeosporioides from leaves, fruit, fl owers and panicles caused similar dis-ease on detached leaves. Work is needed to clarify whether, and the extent to which, isolates from different mango organs differ, and whether there is a relationship among disease reactions on various organs.
Analyses of RFLPs and RAPDs indicated that isolates from mango are genetically uniform and differ from those recovered from avocado, caram-bola and papaya (Hodson et al., 1993; Alahakoon et al., 1994a; Hayden et al., 1994). Isolates genetically similar to those from the latter crops occur infre-quently on mango, and mango isolates were not found on the other plants and were usually virulent only on mango. Alahakoon et al. (1994b) suggested
(c)
(d)
10 μ
(a)25 μ
(b)10 μ
Fig. 8.5. (a) Acervulus, (b) conidiogenous cells and setae, (c) conidia and (d) appres-soria of Colletotrichum acutatum, anamorph of Glomerella acutata (Source: from CMI description no. 630).

Foliar, Floral and Soilborne Diseases 239
that the mango isolates comprise a C. gloeosporioides population that was dis-seminated worldwide from a single source, perhaps as an endophyte.
A recent study identifi ed Colletotrichum spp. that infect mango, passion-fruit (Passifl ora spp.) and tamarillo (Solanum betacea cav. Sendt.) in Colombia and assessed whether cross-infection between the host species occurred (Afanador-Kafuri et al., 2003). With species-specifi c PCR primers, most of the mango isolates were identifi ed as C. gloeosporioides. However, DNA of the passionfruit isolates and single isolates from tamarillo and mango were not amplifi ed by either C. acutatum- or C. gloeosporioides-specifi c primers; they were identifi ed later as C. boninense (Freeman, unpublished data). Further molecular analyses determined that the isolates of C. gloeosporioides from mango were heterogeneous, but that the population of C. boninense from pas-sionfruit, tamarillo and mango was uniform; it may not be host specifi c.
The origins and diversity of C. gloeosporioides on mango require more study. Furthermore, whether distinct populations of this diverse species occur on different mango cultivars and organs, and whether disease reac-tions on one mango organ could be used to predict those on another should be determined. These results would be relevant to host resistance and improvement of the crop, as well as the chemical and cultural control of this important disease.
Epidemiology and managementFitzell and Peak (1984) determined that conidia were the most important inoculum in Australia. They are produced on branch terminals, mummifi ed infl orescences, fl ower bracts and leaves. New leaf fl ushes are the most signifi -cant source of inoculum, and this was corroborated by Dodd et al. (1991) in the Philippines. Optimum production of conidia occurred between 25 and 30°C when free moisture was available, but also formed at 95–97% relative humid-ity (RH). The pathogen’s teleomorph played no apparent role in the spread of the disease (Fitzell and Peak, 1984). The threat posed by C. gloeosporioides to fruit production is greatest in areas with heavy rainfall. In general, this begins with the onset of fl owering (Lim and Khoo, 1985; Jeffries et al., 1990). During commercial production these diseases are managed with diverse chemicals. Registration of the various pesticides varies in different production areas (Jeffries et al., 1990).
Many of the most effective fungicides are systemic. They have selective modes of action, but are prone to lost or reduced effi cacy due to the develop-ment of resistance in the targeted pathogens. Resistance in C. gloeosporioides to benomyl is now common (Maymon et al., 2006), and there is inherent toler-ance in C. acutatum to this fungicide (Freeman et al., 1998; Peres et al., 2002). Newer strobilurin fungicides that are also susceptible to resistance problems must be used properly (no more than three applications per season and alter-nated with fungicides with different modes of action) to ensure that their effi cacy is not lost.
Although fungicide applications usually focus on reducing damage to fruit, disease control on foliage is indicated in some, and on infl orescences in most situations. Trees in nurseries usually require protection if they are

R.C. Ploetz and S. Freeman240
crowded or receive overhead irrigation. Since infected foliage and branch terminals represent important reservoirs of inoculum for blossom and fruit infection, fruit set and anthracnose control on fruit are enhanced if disease control is exercised prior to fl owering (Jeffries et al., 1990). Off-season control measures are benefi cial in production environments that receive signifi cant rainfall.
Apical necrosis
Apical necrosis was fi rst reported in Spain in 1991, and now occurs in Israel, Italy, Portugal and possibly Egypt (Cazorla et al., 1998, 2006; F.M. Cazorla, Málaga, 2007, personal communication). The disease can be quite damaging, and limits production when panicles are affected. Apical buds, leaves and panicles are susceptible (Fig. 8.6a–c), but fruit are not (Cazorla et al., 1998). Vegetative and fl oral apices are affected by a dark-brown to blackish necrosis (Fig. 8.6a, c). Necrosis on leaves begins as blackened, water-soaked lesions, 1–3 mm in diameter, that can coalesce and expand to cover large areas (Fig. 8.6c). Necrosis also extends from affected buds to petioles, through the leaf midrib, and can cover large areas (Fig. 8.6a). A milky bacterial exudate often develops on affected apical buds, but infrequently on petioles (Fig. 8.6b). Large portions of the canopy and high numbers of fl owers can be killed.
Fig. 8.6. Symptoms caused by the apical necrosis pathogen, Pseudomonas syringaepv. syringae. (a) Extensive necrosis on young stem, apical bud, petioles and leaves; (b) bacterial exudate on necrotic stem; and (c) death of a developing fl oral panicle and associated leaf necrosis (Photographs courtesy of F.M. Cazorla).
(a) (b) (c)

Foliar, Floral and Soilborne Diseases 241
The causal bacterium, Pseudomonas syringae pv. syringae van Hall, affects many perennial fruit crops (Hirano and Upper, 1983; Kennelly et al., 2007). It is an epiphyte and is generally not an aggressive pathogen. Disease usually develops after high populations of the bacterium develop in host tissues as a result of host predisposition. Strains from mango produce an antimetabolite toxin, mangotoxin, which plays a role in pathogen virulence and symptom development (Arrebola et al., 2007).
Cold, wet weather and host genotype are primary factors in the develop-ment of apical necrosis (Cazorla et al., 1998, 2006). ‘Tommy Atkins’, ‘Lippens’ and ‘Manzanillo’ are very susceptible whereas ‘Keitt’ and ‘Sensation’ are less so. Apical necrosis is managed in commercial orchards with Cu-containing pesticides, although there has been a recent increase in control failures and Cu resistance in Spain and Portugal (Cazorla et al., 2002, 2006). These out-breaks have been associated with several different Cu-resistance plasmids in the causal bacterium. Carzorla et al. (2006) determined that the plant resis-tance activator acibenzolar-S-methyl and the phosphonate derivative fosetyl-Al provided control comparable to Bordeaux mixture, and that the latter treat-ment might protect plants due to the protective fi lm it provides against wound entry for the pathogen.
Bacterial black spot (black canker)
Bacterial black spot is a destructive leaf, stem and fruit disease in many pro-duction areas (Gagnevin and Pruvost, 2001). In India, the disease is called bacterial canker or black canker due to the cankers it causes on the stems of some cultivars (Prakash et al., 1994). It can be the most important disease where fungal-induced diseases are well controlled (Gagnevin and Pruvost, 2001). Bacterial black spot has been identifi ed in Australia, Myanmar, the Comoros, India, Japan, Kenya, Malaysia, Mauritius, New Caledonia, Paki-stan, the Philippines, Réunion, Rodrigues, South Africa, Taiwan, Thailand and the United Arab Emirates (UAE) (Fukuda et al., 1990; Pruvost et al., 1992; Prakash et al., 1994; Gagnevin and Pruvost, 1995, 2001; Kishun, 1995; Ah-You et al., 2007b). Given the ease with which the pathogen is disseminated in propagation materials and its wide, confi rmed distribution, bacterial black spot may be more widely spread than is currently recognized.
SymptomsMango leaves, stems and fruit are affected (Manicom and Pruvost, 1994; Gagnevin and Pruvost, 2001). On leaves, water-soaked spots are initially 1–3 mm in diameter. As they enlarge, they become raised, black and angular, are limited by veins and surrounded by chlorotic haloes (Fig. 8.7). These lesions are larger and more conspicuously raised than those caused by other xan-thomonads that have been recovered from other species in the Anacardiaceae (Ah-You et al., 2007a). Lesions can merge to form large necrotic patches, and bacteria may ooze from lesions during wet conditions. Old lesions dry out, turn white or grey, and crack. Defoliation occurs in severe cases. Anthracnose
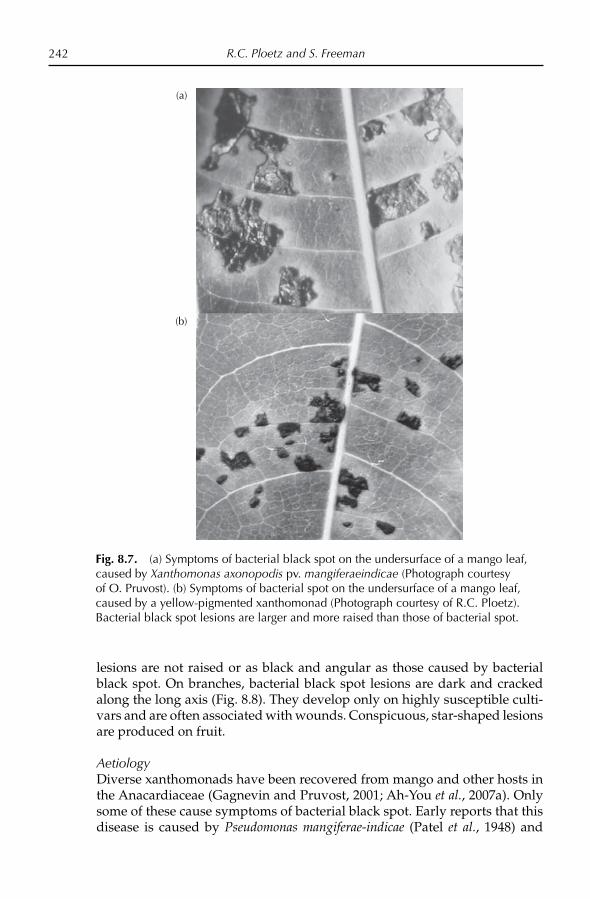
R.C. Ploetz and S. Freeman242
lesions are not raised or as black and angular as those caused by bacterial black spot. On branches, bacterial black spot lesions are dark and cracked along the long axis (Fig. 8.8). They develop only on highly susceptible culti-vars and are often associated with wounds. Conspicuous, star-shaped lesions are produced on fruit.
AetiologyDiverse xanthomonads have been recovered from mango and other hosts in the Anacardiaceae (Gagnevin and Pruvost, 2001; Ah-You et al., 2007a). Only some of these cause symptoms of bacterial black spot. Early reports that this disease is caused by Pseudomonas mangiferae-indicae (Patel et al., 1948) and
(a)
(b)
Fig. 8.7. (a) Symptoms of bacterial black spot on the undersurface of a mango leaf, caused by Xanthomonas axonopodis pv. mangiferaeindicae (Photograph courtesy of O. Pruvost). (b) Symptoms of bacterial spot on the undersurface of a mango leaf, caused by a yellow-pigmented xanthomonad (Photograph courtesy of R.C. Ploetz). Bacterial black spot lesions are larger and more raised than those of bacterial spot.

Foliar, Floral and Soilborne Diseases 243
Erwinia mangiferae (Steyn et al., 1974) are erroneous. The pathogen’s place-ment in Pseudomonas was probably due to its production of non-pigmented colonies in culture (P. syringae pv. syringae causes a different disease of mango, apical necrosis – see above). Cook (1975) indicated that E. mangiferae is a saprophyte that reached high populations in old lesions.
Pathological, cultural, biochemical, physiological, serological and genetic data indicate that strains of the pathogen from different production areas are diverse (Sanders et al., 1994; Gagnevin and Pruvost, 1995, 2001; Kishun, 1995; Pruvost et al., 2005). Genetic diversity is greatest among strains from South-east Asia, suggesting that this region of host diversity is also a centre of pathogen diversifi cation (Gagnevin and Pruvost, 1995).
The pathogen has a single fl agellum, is Gram-negative, rod shaped and 0.4–0.5 × 1.0–1.5 m (Manicom and Wallis, 1984). It is oxidase negative and does not reduce nitrates to nitrites. It cannot use asparagine as a sole carbon and nitrogen source, but is able to hydrolyse starch, esculin, gelatin and casein. On artifi cial media, colonies are cream coloured. (The latter trait is atypical for xanthomonads, which are usually yellow in culture.) Yellow-pigmented xanthomonads have been recovered from mango in Brazil, Réunion, South Africa and Florida USA. These strains cause non-raised leaf lesions (Fig. 8.7b), do not cause fruit or stem lesions, and should not be
Fig. 8.8. A twig canker caused by X. axonopodis pv. mangiferaeindicae that is associated with a wound (Photograph courtesy of O. Pruvost).

R.C. Ploetz and S. Freeman244
classifi ed as pv. mangiferaeindicae (Ah-You et al., 2007a). Three genetically and pathologically distinct groups were identifi ed from different geographic regions and hosts in the Anacardiaceae by Ah-You et al. (2007a). Group I strains were from the Old World, multiplied in mango and cashew (Anacar-dium occidentale L.), fell in amplifi ed fragment length polymorphism (AFLP) group 9.5 of Xanthomonas axonopodis, and contained strains that produced typical bacterial black spot symptoms on mango. These strains of Xanthomo-nas campestris pv. mangiferaeindicae sensu lato were redescribed as X. axonopo-dis pv. mangiferaeindicae sensu novo (s.n.) (Ah-You et al., 2007a). In contrast, Group II strains were from Brazil, multiplied in cashew, but not mango, and fell in AFLP group 9.6 of X. axonopodis. They are associated with symptoms on mango that differ from those of bacterial black spot, including brown, fl at leaf lesions, and black, depressed lesions on fruit of only a few cultivars that is sometimes associated with pulp rot. These strains were responsible for previous reports of bacterial black spot in Brazil (Gagnevin and Pruvost, 2001; Ah-You et al., 2007a). Group III strains are responsible for a unique syn-drome on Spondias dulcis and Spondias mombin in the French West Indies; they fell in AFLP group 9.4. Groups II and III were described, respectively, as X.axonopodis pv. anacardii and X. axonopodis pv. spondiae (Ah-You et al., 2007a).
Epidemiology and managementThe pathogen is disseminated by wind-driven rain (Gagnevin and Pruvost, 2001). Long-distance dissemination occurs via infected propagation material and, less frequently, in infected seed. True seed are not infected, but surface contamination is known. Insects may play a role in dissemination, but these interactions are little studied. The pathogen is an epiphytic colonist of leaves (Manicom, 1986; Pruvost et al., 1990), buds (Pruvost et al., 1993) and fruit (Pruvost and Luisetti, 1991b). Infection occurs through wounds and, less often, stomata on old leaves (young leaves are thought to be resistant due to their non-functional stomata) (Gagnevin and Pruvost, 2001). High RH (> 90%) and moderate temperatures (25–30°C) favour disease development (Kishun and Sohi, 1983; Pruvost and Luisetti, 1991b). There is a direct rela-tionship between the level of disease that develops on leaves and fruit (Man-icom, 1986; Pruvost et al., 1990). Pruvost and Luisetti (1991b) considered that leaf susceptibility was an important criterion when cultivars were selected for lower fruit susceptibility.
Resistance to bacterial black spot varies among mango cultivars, and resistant cultivars should be used where disease pressure is high (Manicom and Pruvost, 1994). When new orchards are established, pathogen-free plant-ing material should be utilized. Since the pathogen moves short distances in wind-blown aerosols (usually within orchards), the long-distance spread of the pathogen depends almost entirely upon the movement of infected plants (Manicom and Pruvost, 1994). Windbreaks can be used to reduce wounding and infected twigs (Fig. 8.8) should be pruned to reduce inoculum in the canopy.
Bacterial black spot can be quite diffi cult to control, particularly on sus-ceptible cultivars. Available chemicals may be only marginally effective

Foliar, Floral and Soilborne Diseases 245
under high disease pressure (Pruvost et al., 1989). During rainy weather, applications of Cu-based bactericides are recommended. The application schedules for these compounds focus on protecting fruit, and vary according to the length of time fruit are exposed to wet conditions (Manicom and Pru-vost, 1994). Although agricultural antibiotics (e.g. various formulations of streptomycin sulfate or nitrate) have been reported to be effective (Misra and Prakash, 1992; Viljoen and Kotzé, 1972), resistance that develops to these products after continuous use limits their long-term effectiveness against this disease. Biological control measures have not been widely researched. Pruvost and Luisetti (1991a) reported little success. In India, Kishun (1994) indicated that a strain of Bacillus coagulans from the phylloplane of mango was effective against strains of the pathogen, although control of bacterial black spot in the fi eld was not reported.
Black-banded disease
This is a relatively unimportant disease that affects mango leaves and branches (Reddy et al., 1961). The causal fungus, Rhinocladium corticola Mas-see (described as ‘corticolum’) (teleomorph: Peziotrichum corticolum (Massee) Subrumanian), was described on the bark of trees in Poona, India (Hughes, 1980). It produces repent, intricately branched, septate, olivaceous hyphae 5–7 m in diameter. Erect hyphae bear globose, olivaceous, densely and minutely tuberculate, 15–18 m conidia. Hughes (1980) questioned whether this was a species of Rhinocladium since it was quite different from other spe-cies in the genus. It forms a black, velvety mass of hyphae on affected sur-faces in conspicuous blotches or bands. The fungus is restricted to the outer portions of bark. Bordeaux mixture controls the disease, but is not required in most situations.
Black mildew, sooty blotch and sooty mould
Several ascomycetes produce dark-coloured, usually superfi cial growths on the surfaces of stems, leaves and fruit (Lim and Khoo, 1985). These range from thin, diffuse webs of dark hyphae to opaque, felty layers; in extreme cases, a thick crust of hyphae forms. The variations in appearance result, presumably, from the different species of fungi that are involved. These are usually not important problems in well-maintained orchards. However, layers of hyphae that the black mildew and sooty mould fungi form may be thick enough to block sunlight and inhibit photosynthesis. These blemishes also detract from the appearance and marketability of fruit.
Sooty moulds develop in the presence of aphids, mealybugs, scales and other sucking insects that produce honeydew (excreta) when feeding. Hon-eydew is a required food source for these fungi, and the problems they cause dissipate if the associated insects are controlled. In contrast, black mildews

R.C. Ploetz and S. Freeman246
and sooty blotch are not dependent on honeydew and grow directly on host surfaces.
AetiologyBlack mildew and sooty mould are similar in appearance, but their respec-tive causal agents are distinct (Lim and Khoo, 1985). The black or dark mil-dews are a group of mostly tropical obligate plant pathogens that produce two types of hyphopodia (Alexopoulos et al., 1996). Capitate hyphopodia are lobed appressoria from which infection haustoria are formed, whereas mucronate hyphopodia function as conidiogenous cells. Black mildew of mango is caused by Meliola mangiferae Earle (Sordariomycetes, Ascomycota) (Fig. 8.9).
In contrast, the fungi that cause sooty moulds are diverse saprophytes that require honeydew to colonize plant surfaces. In Malaysia, Lim and Khoo (1985) listed coelomycetes (Polychaeton), hyphomycetes (Tripospermum) and loculoascomycetes (Antennulariella, Chaetothyrium, Limacinula and Scorias), whereas the reported agents in India were hyphomycetes (Leptoxyphium,
(a)
(c)
(b)10 mm
10 μm
(d)
250 μm
1 mm
Fig. 8.9. (a–c) Signs of black mildew, and (d) microscopic features of the causal agent, Meliola mangiferae (Source: from CMI description no. 1355).

Foliar, Floral and Soilborne Diseases 247
Microxyphium and Tripospermum) and loculoascomycetes (Capnodium) (Butler and Bisby, 1931; Prakash, 1988). In Pakistan, 18 species in eight genera were associated with sooty mould, including the foliar pathogens Aspergillus, Alternaria, Botryodiplodia, Capnodium, Cladosporium, Curvularia, Fusarium andHelminthosporium spp. (Hamid and Jalaluddin, 2006). Since some of the sooty mould fungi may not sporulate on plants and because they are often found in combination with one another, it is usually diffi cult to identify the specifi c species that are involved.
Although ‘sooty blotch’ has been used as a synonym for ‘sooty mould’ (Singh, 1968), sooty blotch refers specifi cally to disease complexes on tem-perate and tropical plants that are not associated with honeydew and are caused by a diverse group of Dothidiomycetes (Ascomycota) (Johnson et al., 1997; Ploetz et al., 2000; Batzer et al., 2005). The specifi c agents that are associ-ated with sooty blotch on mango are not known, but resemble those from apple, carambola and pear (Plate 44).
Epidemiology and managementSooty moulds develop on honeydew that is produced on plant surfaces by aphids, mealybugs, scales and other sucking insects. They are managed by controlling the associated insects with oils and insecticides (Lim and Khoo, 1985). In Pakistan, spraying of fungicides (sulfur (S) and mancozeb) and insecticides (malathion, diazinon and coal tar at 1 kg/tree) separately reduced the incidence of sooty mould on foliage, whereas a mixture of fungicides and insecticides further decreased sooty mould incidence (Hamid and Jalalud-din, 2006). In India, sooty mould, caused by species of Microxyphium, Lep-toxyphium and Tripospermum, was best controlled with a spray of S and parathion-methyl (Prakash, 1991).
Sooty blotch management has not been investigated on mango; however, signs of these fungi have been removed from apple with various postharvest washes (Batzer et al., 2002), and managed in apple orchards with diverse contact and systemic fungicides (Williamson and Sutton, 2000).
Blossom blight
Blossom blight can reduce fruit set and production considerably since fl ow-ers and large areas of the panicle can be killed. When this disease was con-trolled with fungicides in the Philippines, a 55–80% increase in fruit set occurred (Pordesimo, 1982).
SymptomsBlossom blight starts as a wilt of the affected part of the infl orescence that is often curved, the ‘shepherd’s crook symptom’ (Fig. 8.10). The peduncle blackens and dies back from the tip. Internally, discoloration advances beyond the surface lesion. Large black lesions can develop lower on the peduncle, and once it is girdled the apex dies.

R.C. Ploetz and S. Freeman248
AetiologyThe cause of blossom blight is confused. Colletotrichum gloeosporioides has been reported most frequently as the responsible fungus (Fitzell et al., 1984; Jeffries et al., 1990), and A. alternata has also been reported to attack panicles and reduce fruit set (Cronje et al., 1990). Powdery mildew (see section below) also damages panicles, but its symptoms are distinct from those of blossom blight. In South Africa, symptoms caused by A. alternata and C. gloeosporioides are small, mainly superfi cial black spots, 1–2 × 2–5 mm, on the peduncle (Darvas, 1993; Lonsdale and Kotzé, 1993). Rather than blossom blight, Lonsdale and Kotzé (1993) indicated that these pathogens caused blossom spot. In con-trast, Lonsdale and Kotzé (1993) reported that Dothiorella mangiferae caused extensive, systemic damage, and Darvas (1993) indicated that Dothiorella domini-cana is the only fungus that caused typical symptoms of blossom blight. Crous et al. (2006) placed these fungi in a new genus, Neofusicoccum (see Decline disorders section below). Work is needed to determine the distribu-tion of Neofusicoccum-incited panicle disease, and the identity of the most important blossom blight pathogens worldwide.
Epidemiology and managementLittle is known about the epidemiology of Neofusicoccum-incited panicle dis-ease. Studies on the stem-end rot diseases have shown that the causal fungi are endophytes. The roles of internal and external sources of inoculum in the development of panicle disease are unknown. For optimal fruit set and
Fig. 8.10. Symptoms of blossom blight on panicles of mango. Note the blighted, curved terminals and almost complete lack of fruit set (Photograph courtesy of D. Benscher).

Foliar, Floral and Soilborne Diseases 249
development, blossom blight must be controlled. Once fl owering begins, early and frequent fungicide applications are necessary in many areas, depending on rainfall. Previously published infection models can be used to time applications appropriately (Fitzell et al., 1984; Dodd et al., 1991). Fitzell et al. (1984) investigated environmental conditions that were conducive to infection by C. gloeosporioides, and indicated that temperature and free mois-ture were important determinants of infection. They developed a model for scheduling fungicide application, which reduced the number of applications that were needed to control blossom blight. Presumably, systemic fungicides would be needed to control disease caused by endophytic agents.
Decline disorders
Several diseases of mango have been variously termed blight, canker, decline, gummosis, twig blight, tip dieback and stem bleeding. They have similar symptoms and aetiologies.
SymptomsThese widespread problems are not well understood. Symptoms include: (i) marginal scorching of leaf lamina; (ii) foliar symptoms of nutritional defi -ciencies, particularly of iron (Fe) and manganese (Mn); (iii) vascular discolor-ation (Fig. 8.11a); (iv) dieback of small branches basipetally from the terminal that may or may not progress to defoliation (Fig. 8.11b and c); (v) gummosis, an oozing of a clear or cloudy exudate either from terminal buds or from branches, scaffold limbs or trunks (Plate 45); and (vi) root degeneration (Lim and Khoo, 1985; Ploetz et al., 1996a; Ploetz and Prakash, 1997).
AetiologyDiverse biotic and abiotic factors may be primary causes of decline symp-toms or predisposing agents (McSorley et al., 1980; Kadman and Gazit, 1984; Schaffer et al., 1988; Ploetz and Prakash, 1997). Fungi are the most common agents. They are endophytes that also cause stem-end rots on mango fruit, and are usually secondary pathogens that cause disease on weakened, pre-disposed hosts (Johnson et al., 1992; Ploetz and Prakash, 1997; Slippers et al., 2005; Slippers and Wingfi eld, 2007). Several species cause all or some of the above symptoms when used to individually inoculate plants (Ploetz et al., 1996a). Their frequent association with one another in affected tissues may indicate that these symptoms usually develop, or develop more severely, after multiple infections.
Several of these pathogens are in the Botryosphaeriaceae (Dothidiomy-cetes, Ascomycota). The taxonomy and nomenclature of these fungi has been confused, and ‘phylogenetic understanding of major groups within Botry-osphaeria remains poor’ (Crous et al., 2006). With 28S rDNA sequence data, Crous et al. (2006) examined natural relationships among available members of the family. Ten lineages were distinguished, most of which contained ana-morphs with distinct morphological features. New relationships were revealed

R.C. Ploetz and S. Freeman250
in some of the lineages that necessitated the renaming of several taxa (Crous et al., 2006). These new names and holomorphs are used below when discuss-ing the taxa that occur on mango.
Lasiodiplodia theobromae (Pat.) Griffon and Maubl.) (synonyms: Botryodi-plodia theobromae Pat., Diplodia natalensis Pole-Evans, and Diplodia theobromae (Pat.) W. Nowell) is the most common and widespread cause of decline dis-eases of mango (Ploetz and Prakash, 1997), and affects many other host plants in the tropics (Punithalingam, 1976). Crous and Palm (1999) declared B. theo-bromae, a nomen dubium. Denman et al. (2000) reduced D. natalensis and L.theobromae to synonymy with D. theobromae. However, adopting this change was questioned by Burgess et al. (2006), who noted that fi ve species of Lasio-diplodia fell in a phylogenetic clade that had 100% bootstrap support; it was distinct from a clade that included species of Diplodia and Dothiorella. The teleomorph of L. theobromae, formerly Botryosphaeria rhodina (Cooke) Arx (synonym: Physalospora rhodina Cooke), is usually not found in nature. In the study of Crous et al. (2006), the genus Botryosphaeria was reserved for the type species Botryosphaeria dothidea (Moug.:Fr.) (anamorph: Fusicoccum aesculi Corda), which was in a different clade than L. theobromae. However, Crous etal. (2006) refrained from renaming B. rhodina until the poorly resolved clade in which it resided could be clarifi ed with work with additional isolates and analyses.
Lasiodiplodia theobromae attacks weakened trees that are predisposed by: high and low temperatures; drought; high RH; hardpan soils; sun scorch; and tar and tanglefoot (Muller, 1940; Das Gupta and Zacchariah, 1945; Alvarez-García and López-Gracía, 1971; Acuña and Waite, 1977; Ploetz et al., 1996a). It is often an endophyte, infects wounded plants, and is found in soil, on dead twigs, mummifi ed fruit and on organic debris beneath trees (Johnson et al., 1992).
(a) (b) (c)
Fig. 8.11. Among the symptoms that are associated with mango decline are: (a) internal/vascular discoloration and branch terminal death (tip dieback) that may not (b), or may be associated with defoliation (c) (Photographs courtesy of D. Benscher).
(a) (b) (c)

Foliar, Floral and Soilborne Diseases 251
Dieback caused by L. theobromae has been recognized as a signifi cant dis-ease in India since the 1940s. It was the most serious disease of mango in the Jaipur district (Verma and Singh, 1970), and affected 30–40% of the planta-tions in the Moradabad region of Uttar Pradesh (Prakash and Srivastava, 1987). Das Gupta and Zacchariah (1945) indicated that only L. theobromae caused dieback; Phoma sp. and two Fusarium spp. were not pathogenic. Lasio-diplodia theobromae caused a canker on mango in Indonesia (Muller, 1940) and Malaysia (Lim and Khoo, 1985), dieback in Egypt and the Sonsonate area of El Salvador (Acuña and Waite, 1977), and gummosis and dieback in Puerto Rico (Alvarez-García and López-Gracía, 1971).
Lasiodiplodia theobromae produces fl uffy, grey-black mycelium on oatmeal agar (OA) and PDA (Johnson, 1994b). Conidiomata may be simple or develop into aggregated stromatic bodies (Burgess et al., 2006). Cirri of conidia may ooze from ostioles. Conidia are initially hyaline, aseptate, granular, ovoid to ellipsoid and thick-walled (Fig. 8.12). Mature conidia are two celled, 17–33 × 10–15 m, and brown walled with numerous longitudinal striations. Paraphyses are usually present. The teleomorph was produced when indi-vidual isolates were cultured on caimito fruits (Chrysophyllum cainito L.) or papaya (Carica papaya L.) stems, suggesting that it was homothallic (Alvarez-García and López-Gracía, 1971).
One of the most important mango pathogens causes stem-end rot on fruit, dieback and blossom blight. Crous et al. (2006) refer to it as Neofusicoccum parvum (Pennycook and Samuels) Crous, Slippers and A.J.L. Phillips (formerlyFusicoccum parvum Pennycook and Samuels) (teleomorph: Botryosphaeria-like; formerly Botryosphaeria parva Pennycook and Samuels). Slippers et al. (2005) argued that D. dominicana Petro and Cif. may be synonymous with this fun-gus. Johnson (1992), an author of the Slippers et al. (2005) paper, had placed D. dominicana in synonymy with F. aesculi Corda (teleomorph B. dothidea (Moug.:Fr.) Ces. and De Not.). They indicated that this fungus had been mis-identifi ed as Botryosphaeria ribis Gross. and Duggar (anamorph: Fusicoccumsp.) and B. dothidea (anamorph: F. aesculi), due to overlapping host ranges and spore dimensions. They felt that the tip dieback fungus reported by Ramos et al. (1991) as B. ribis was probably N. parvum (= ‘B. parva’).
Neofusicoccum parvum produces cottony grey mycelium and discrete pyc-nidia or stromatic multilocular fruiting bodies on, respectively, PDA and OA (Johnson et al., 1991). Discrete, immersed pycnidia in a subcuticular pseudostroma are produced on mango. Conidia are fusiform to navicular, 14.7–25.5 (19) × 4.5–7 (5.2) m, hyaline and unicellular (Slippers et al., 2005). Sometimes, brown, biseptate conidia are observed. The teleomorph develops infrequently on OA, and has been found on mango twigs in tree litter in Australia (Johnson et al., 1991). On twigs, pseudothecia are subglobose to pyriform, 210 × 120 m, and form beneath the epidermis. Ascostromata are hemi-lenticular and up to 10 mm wide on OA. Asci are eight spored, bituni-cate and irregularly biseriate. Ascospores are hyaline, single celled, fusiform and 16–25 × 4.5–9.5 m.
Neofusicoccum mangiferum (Syd. and P. Syd.) Crous, Slippers and A.J.L. Phillips (basionym: Dothiorella mangiferae Syd. and P. Syd.; synonyms:
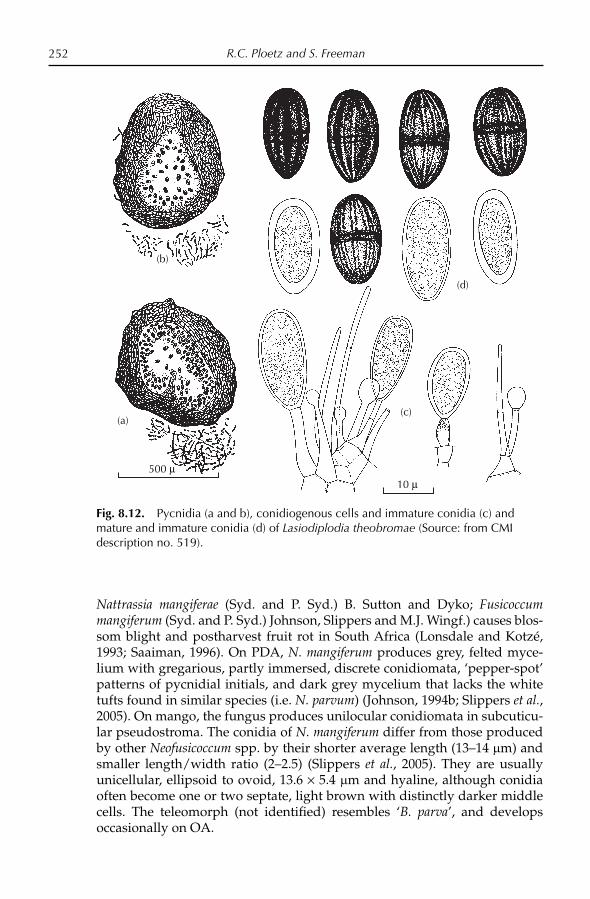
R.C. Ploetz and S. Freeman252
Nattrassia mangiferae (Syd. and P. Syd.) B. Sutton and Dyko; Fusicoccummangiferum (Syd. and P. Syd.) Johnson, Slippers and M.J. Wingf.) causes blos-som blight and postharvest fruit rot in South Africa (Lonsdale and Kotzé, 1993; Saaiman, 1996). On PDA, N. mangiferum produces grey, felted myce-lium with gregarious, partly immersed, discrete conidiomata, ‘pepper-spot’ patterns of pycnidial initials, and dark grey mycelium that lacks the white tufts found in similar species (i.e. N. parvum) (Johnson, 1994b; Slippers et al., 2005). On mango, the fungus produces unilocular conidiomata in subcuticu-lar pseudostroma. The conidia of N. mangiferum differ from those produced by other Neofusicoccum spp. by their shorter average length (13–14 m) and smaller length/width ratio (2–2.5) (Slippers et al., 2005). They are usually unicellular, ellipsoid to ovoid, 13.6 × 5.4 m and hyaline, although conidia often become one or two septate, light brown with distinctly darker middle cells. The teleomorph (not identifi ed) resembles ‘B. parva’, and develops occasionally on OA.
(b)
(d)
(c)(a)
500 μ10 μ
Fig. 8.12. Pycnidia (a and b), conidiogenous cells and immature conidia (c) and mature and immature conidia (d) of Lasiodiplodia theobromae (Source: from CMI description no. 519).

Foliar, Floral and Soilborne Diseases 253
A dieback disease of mango has been recognized in Niger (Reckhaus and Adamou, 1987). Neoscytalidium dimidiatum (Penz.) Crous and Slippers (basionym: Torula dimidiata Penz.; synonyms: Scytalidium dimidiatum (Penz.) B. Sutton and Dyko (Fig. 8.13); Fusicoccum dimidiatum (Penz.) D.F. Farr; Hendersonula toruloidea Natrass) causes sudden wilting of shoots to large branches, fi ring of leaves and trunk cankers from which a clear exudate orig-inates. Reckhaus and Adamou (1987) believed that water stress was a pri-mary, predisposing factor in the development of this disease.
Botryosphaeria dothidea (anamorph: F. aesculi) is an uncommon mango pathogen (Ploetz and Prakash, 1997; Slippers et al., 2005). It produces a fl uffy grey mycelium with discrete pycnidia on PDA or stromatic multilocular fruiting bodies on OA. Discrete, immersed pycnidia are produced on mango.
(e)
(d)
(c)
(a)
(b)
10 μ
50 μ
Fig. 8.13. (a) Vertical section of stroma, (b) part of pycnidial wall and conidiophores, (c) conidiophores, (d) conidia, and (e) the Scytalidium-like synanamorph of Neoscyta-lidium dimidiatum (Source: from CMI description no. 274).
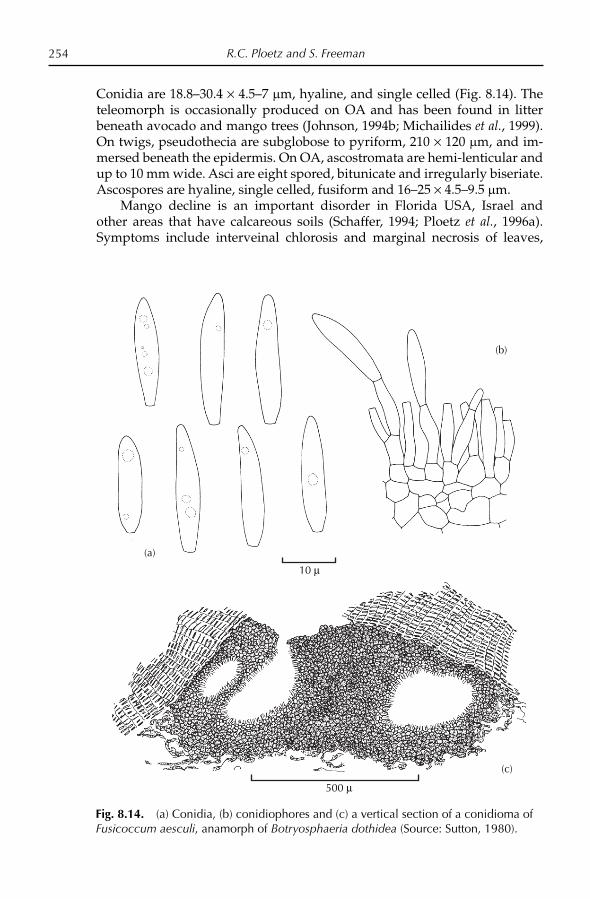
R.C. Ploetz and S. Freeman254
Conidia are 18.8–30.4 × 4.5–7 m, hyaline, and single celled (Fig. 8.14). The teleomorph is occasionally produced on OA and has been found in litter beneath avocado and mango trees (Johnson, 1994b; Michailides et al., 1999). On twigs, pseudothecia are subglobose to pyriform, 210 × 120 m, and im-mersed beneath the epidermis. On OA, ascostromata are hemi-lenticular and up to 10 mm wide. Asci are eight spored, bitunicate and irregularly biseriate. Ascospores are hyaline, single celled, fusiform and 16–25 × 4.5–9.5 m.
Mango decline is an important disorder in Florida USA, Israel and other areas that have calcareous soils (Schaffer, 1994; Ploetz et al., 1996a). Symptoms include interveinal chlorosis and marginal necrosis of leaves,
(a)
(c)
(b)
10 μ
500 μ
Fig. 8.14. (a) Conidia, (b) conidiophores and (c) a vertical section of a conidioma of Fusicoccum aesculi, anamorph of Botryosphaeria dothidea (Source: Sutton, 1980).

Foliar, Floral and Soilborne Diseases 255
dieback of young twigs that progresses to larger branches, reduced growth of secondary roots, gummosis and vascular discoloration. Several different factors have been associated with mango decline in Florida. Schaffer et al. (1988) used the Diagnosis and Recommendation Integrated System (DRIS) to assess the nutritional status of declining and healthy ‘Tommy Atkins’ trees. The nutrient imbalance index, an indication of the overall nutrient balance in trees, was greatest for declining trees. DRIS identifi ed Mn, Fe or both ele-ments as the most defi cient in declining trees, and in two of the three declin-ing orchards that they investigated, concentrations of these elements were below the critical range. Mineral defi ciencies may be predisposing factors in the development of mango decline, since pathogenic fungi are recovered from symptomatic trees (see below).
McSorley et al. (1980) detected the nematode Hemicriconemoides mangiferae Siddiqi at low, but consistent levels on declining mango trees. They sug-gested that it may have been responsible for the reduced root growth in affected trees, and could play a role in the development of the disorder.
Smith and Scudder (1951) reported that Diplodia sp. caused a dieback of mango. Additional species of fungi were examined by Ploetz et al. (1996a). Alternaria alternata, C. gloeosporioides, N. parvum (D. dominicana), L.theobromae and two Phomopsis spp. were recovered from trees with diverse decline symptoms, and caused one or more of these symptoms on ‘Keitt’ and ‘Tommy Atkins’. Colletotrichum gloeosporioides, N. parvum and L. theobromae were most damaging, and caused signifi cant bud necrosis, tip dieback, gummosis and vascular discoloration (Fig. 8.15); these symptoms were distinguishable only when C. gloeosporioides sporulated on inoculated branches.
In summary, several different fungi cause, or are associated with, decline symptoms worldwide; most are endophytes that have Botryosphaeria or Botryosphaeria-like teleomorphs (Botryosphaeriaceae). Stress and wound pre-disposition are usually associated with symptom development.
ManagementControlling decline disorders of mango is diffi cult. Techniques that could detect these pathogens in plants would be useful to identify pathogen-free propagation materials. The internal location and the diversity of fungi that are involved decrease the opportunities for controlling these disorders with fungicides (Peterson et al., 1991). Because signifi cant movement of some of these pathogens may occur via wind and rainsplash, strategic applications of broad-spectrum protectant fungicides may be effective at certain times of the year (Lonsdale and Kotzé, 1993), but have not been tested. In India, dieback was managed by pruning affected portions of the canopy and treating the wounded areas with a 5:5:50 Bordeaux mix (Prakash and Raoof, 1989). Man-agement of the controllable predisposing factors, such as drought stress, may also be benefi cial. A better understanding of the epidemiology of these dis-eases would assist these efforts. Pruning to force synchronous fl ushes of foliar growth might enable the avoidance of windows of infection for certain pathogens (Johnson, 1994b).

R.C. Ploetz and S. Freeman256
Galls and scaly bark
Gall and scaly bark disorders of mango are known in several producing regions. These diseases are usually minor problems.
SymptomsIn India, bark scaling develops as deep cracks along the entire rootstock por-tion of the plant, and cracks may penetrate the phloem and become necrotic (Prakash and Srivastava, 1987). These symptoms resemble those of a scaly bark disorder, ‘cuarteado’, in Colombia (Cook, 1975). In Hawaii, similar symptoms developed on mango seedlings (Cook et al., 1971). The bark from the soil line to the fi rst branches was rough and scaly, and xylem pegs, 5 mm long, were evident when the bark was removed around leaf scars and secondary branches.
In Mexico, a disorder known as ‘nanahuate’, ‘bolas’ or ‘buba of mango’, causes galls, 5–10 cm in diameter, which resemble a caulifl ower, are initially light green, but become dark brown as they die (Fig. 8.16) (Angulo and Villapudua, 1982). The galls remain attached to trees for many years, and severely affected branches die. Similar symptoms are found in Florida USA, and are associated with pruning injuries. Larger galls have also been noted in Puerto Rico, as well as the US Department of Agriculture (USDA) Agriculture Research Service (ARS) in Miami and University of Florida in Homestead (Fig. 8.17) (Ploetz et al., 1996b; R. Rodriguez, personal communication). Some of the latter galls are large, have rough, scaly exteri-ors, and are usually found on the main trunk or scaffold limbs of affected trees. Cracks may penetrate the phloem and become necrotic, but the branch death that is associated with galls in Mexico and India has not been observed.
(a) (b) (c)
Fig. 8.15. Decline symptoms induced on ‘Tommy Atkins’ plants artifi cially inoculated with isolates of: (a) C. gloeosporioides; (b) Neofusicoccum parvum; and (c) L. theobro-mae (Photographs courtesy of D. Benscher).
(a) (b) (c)

Foliar, Floral and Soilborne Diseases 257
AetiologyFusarium decemcellulare C. Brick (synonym: Fusarium rigidiuscula (Brick) Snyd. and Hans.) causes these diseases in Florida USA, Mexico and Venezuela (Malaguti and de Reyes, 1964; Angulo and Villapudua, 1982; Ploetz et al., 1996b). Colonies on PDA are dark carmine-red on the underside. The fungus produces microconidia in false heads or chains on branched and non-branched monophialides (Fig. 8.18). Large macroconidia, 92–55 × 7–5.5 m, are produced in slimy yellow sporodochia, c.1 mm in diameter. The fungus’s
Fig. 8.16. Galls of the ‘buba’ type in Haiti (Photograph courtesy of Carolyn Cohen, USDA, Animal and Plant Health Inspection Service (APHIS)).
Fig. 8.17. Large, 30-year-old gall on ‘Langra’ in the USDA-ARS mango germplasm repository in Miami (Photograph courtesy of R.C. Ploetz).
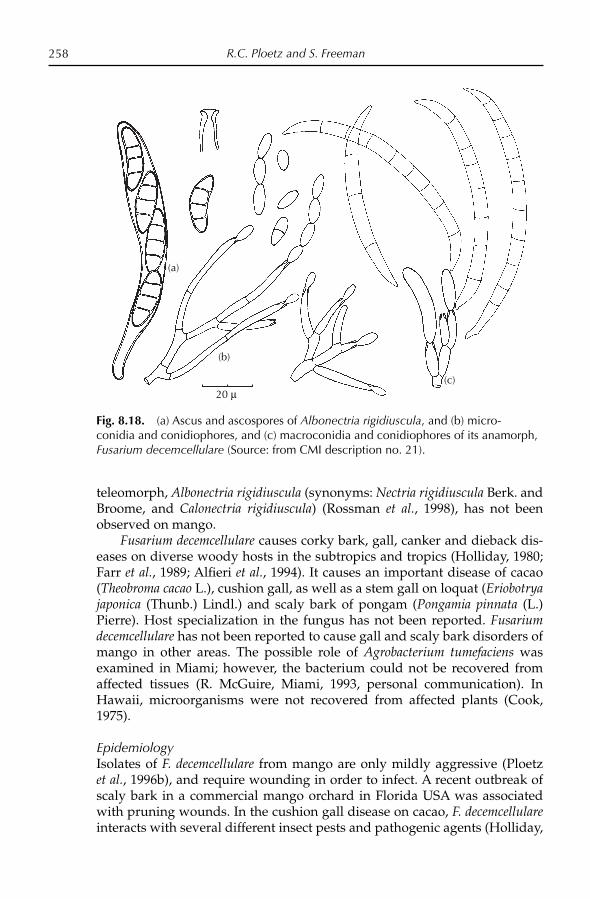
R.C. Ploetz and S. Freeman258
teleomorph, Albonectria rigidiuscula (synonyms: Nectria rigidiuscula Berk. and Broome, and Calonectria rigidiuscula) (Rossman et al., 1998), has not been observed on mango.
Fusarium decemcellulare causes corky bark, gall, canker and dieback dis-eases on diverse woody hosts in the subtropics and tropics (Holliday, 1980; Farr et al., 1989; Alfi eri et al., 1994). It causes an important disease of cacao (Theobroma cacao L.), cushion gall, as well as a stem gall on loquat (Eriobotryajaponica (Thunb.) Lindl.) and scaly bark of pongam (Pongamia pinnata (L.) Pierre). Host specialization in the fungus has not been reported. Fusariumdecemcellulare has not been reported to cause gall and scaly bark disorders of mango in other areas. The possible role of Agrobacterium tumefaciens was examined in Miami; however, the bacterium could not be recovered from affected tissues (R. McGuire, Miami, 1993, personal communication). In Hawaii, microorganisms were not recovered from affected plants (Cook, 1975).
EpidemiologyIsolates of F. decemcellulare from mango are only mildly aggressive (Ploetz et al., 1996b), and require wounding in order to infect. A recent outbreak of scaly bark in a commercial mango orchard in Florida USA was associated with pruning wounds. In the cushion gall disease on cacao, F. decemcellulare interacts with several different insect pests and pathogenic agents (Holliday,
(b)
(c)
(a)
20 μ
Fig. 8.18. (a) Ascus and ascospores of Albonectria rigidiuscula, and (b) micro-conidia and conidiophores, and (c) macroconidia and conidiophores of its anamorph, Fusarium decemcellulare (Source: from CMI description no. 21).

Foliar, Floral and Soilborne Diseases 259
1980; Ploetz, 2007). These insects may facilitate infection and disseminate the pathogen. Insect-F. decemcellulare interactions have not been investigated on mango.
ManagementNo pesticides have been identifi ed to control this problem. Measures that should be helpful include the removal and destruction of affected branches and trees in the orchard, disinfestation of pruning equipment to ensure that the pathogen is not spread during pruning operations, and the use of healthy planting material in new orchards.
Grey leafspot
Pestalotiopsis mangiferae (Henn.) Steyaert (synonym: Pestalolia mangiferae Henn.; no teleomorph of the fungus is known) causes grey leafspot and stem-end rot of mango fruit (Lim and Khoo, 1985; Johnson, 1994b). It is a weak pathogen that usually requires wounding in order to infect mango. Grey leafspot is usually unimportant and occurs mainly on unhealthy or poorly maintained trees.
Pestalotiopsis mangiferae produces abundant conidia in acervuli that develop in grey leafspot lesions and necrotic areas on fruit (Lim and Khoo, 1985). As lesions age, black columns of spores emanate through ruptures in the host epidermis. Conidia are produced that have three thick-walled, brownish, concolorous median cells and thin-walled, hyaline apical and basal cells; the apical cells bear three characteristic appendages (Fig. 8.19). Conidia are 20 × 5 m, fusiform and straight to slightly curved. Two other species of Pestalotiopsis that occur on mango produce larger conidia, Pestalo-tiopsis mangifolia Guba and Pestalotiopsis versicolor Speg. (synonyms: Pestaloti-opsis cliftoniae Tracy and Earle and Pestalotiopsis coccolobae Ellis and Everh.).
On leaves, symptoms are light grey spots, usually 2–20 mm in diameter (Lim and Khoo, 1985). These may coalesce to form large patches of necrotic tissue on leaves. Lesions are surrounded by dark, raised margins, and as they mature, raised black dots (which are columns of the pathogen’s conidia) are evident in lesion centres. Although most cultivars are susceptible, specifi c control measures are usually not required. Dithiocarbamate fungicides con-trol this disease.
Leaf blight
This disease has been reported in India and Nigeria (Hingorani et al., 1960; Cook, 1975; Okigbo, 2001; Okigbo and Osuinde, 2003), and the causal fun-gus, Macrophoma mangiferae Hingorani and Sharma (Ascomycota), has also been intercepted in shipments to the USA from Mexico (Systematic Mycol-ogy and Microbiology Laboratory, USDA-ARS, Beltsville). This is not a serious problem.

R.C. Ploetz and S. Freeman260
Macrophoma mangiferae produces subepidermal, globose pycnidia, 77–231 m in diameter. Hyaline conidiophores, 1.5–2 × 8–11 m, produce unicellular
conidia, 5.3–7 × 10.5–24.5 m. No teleomorph is known. Since the genus Mac-rophoma has been placed in synonymy with Sphaeropsis, this species should be redescribed.
Leaves, twigs and fruit are affected, especially when the latter are stored. Small, yellow spots gradually enlarge to become brown with wide purple margins. The lesions are initially circular, but develop an irregular appear-ance and may encompass large portions of the leaf surface. Pycnidia form most frequently on the underside of leaves. Elliptical stem lesions are infre-quent but can girdle stems. In India, the disease is most serious during the rainy season (Verma and Singh, 1996b). Macrophoma mangiferae survives in pycnidia that develop on bark of twigs of young mango plants, blighted leaves and as dormant mycelium in wood (Verma and Singh, 1996a). Okigbo (2001) reported that the fungus survived adverse conditions best in stems and branches.
Four applications of captan, Bordeaux mixture, captafol, carbendazim and thiophanate-methyl were effective on young plants (Verma and Singh, 1996a). The bacterium Bacillus subtilis NCIB 3610, isolated from soil under a mango tree, inhibited M. mangiferae on agar plates, and symptoms were reduced in the fi eld by the application of the antagonist (Okigbo and Osuinde, 2003).
(a)
(b)
50 μm
(c)
10 μm
Fig. 8.19. (a) Vertical section of an acervulus, (b) mature conidia, and (c) conidiog-enous cells and developing conidia of Pestalotiopsis mangiferae (Source: from CMI description no. 676).

Foliar, Floral and Soilborne Diseases 261
Malformation
Malformation is one of the most destructive mango diseases (Ploetz, 2001). Although trees are not killed, the vegetative phase of the disease impedes canopy development and the fl oral phase dramatically reduces fruit yield. Based on the incidence and severity of malformation in Egypt, an estimated US$15 million of fruit were lost in 1998 (Ploetz et al., 2002). Losses in more important producing countries (i.e. India) are undoubtedly much greater.
Malformation was fi rst reported in India in 1891 (Kumar and Beniwal, 1991), and has subsequently been observed in Brazil, Myanmar, Egypt, El Salvador, India, Israel, Malaysia, Mexico, Nicaragua, Oman, Pakistan, South Africa, Sudan, Spain, Swaziland, Uganda and the USA (Flechtmann et al., 1973; Crookes and Rijkenberg, 1985; Lim and Khoo, 1985; Kumar and Beni-wal, 1991; Ploetz, 2001; Kvas et al., 2007; S. Freeman, Bet Dagan, 2007, unpub-lished data; G.I. Johnson, Jamison, Australia, 2007, personal communication; J.F. Leslie, Manhattan, Kansas, 2007, personal communication). Since the pathogen is easily disseminated in infected germplasm and there are con-spicuous gaps in the disjunct distribution (note African and American records), malformation is probably more widely spread.
SymptomsMalformation affects vegetative and fl oral meristematic tissues (Fig. 8.20) (Ploetz, 2001). Vegetative malformation is most serious on seedlings and small plants in nurseries, especially where seedlings are grown beneath affected trees, a common practice in the Middle East (Ploetz et al., 2002; Youssef et al., 2007). Vegetative malformation also occurs on mature trees. Apical and axillary buds produce misshapen shoots with shortened inter-nodes and dwarfed leaves that are brittle and recurve towards the sup-porting stem (Fig. 8.20). Shoots may not expand fully, resulting in a bunched appearance (i.e. the ‘bunchy-top’ symptom of the disease).
Floral malformation is most important. Affected infl orescences usually do not set fruit or fruit are aborted. Primary or secondary axes on affected panicles are shortened, thickened and highly branched (Fig. 8.20). Malformed panicles produce up to three times the normal number of fl owers, range from one-half to two times the normal size, and have an increased proportion of male to perfect fl owers. Malformed panicles may also produce dwarfed and distorted leaves (exhibit phyllody).
AetiologyThe aetiology of malformation has been controversial for almost as long as the disease has been recognized (Ploetz, 2001). Suggested causes include mites (Narasimhan, 1954), nutritional problems (Prasad et al., 1965), physio-logical or hormonal imbalances (Dang and Daulta, 1982; Singh and Dhillon, 1989), viruses (Kauser, 1959) and unknown causes (Kumar and Beniwal, 1991). Summanwar et al. (1966) demonstrated that a fungus, identifi ed then as Fusarium moniliforme Sheld., was responsible for the fl oral phase of this disease. Varma et al. (1974) later showed that F. moniliforme also caused
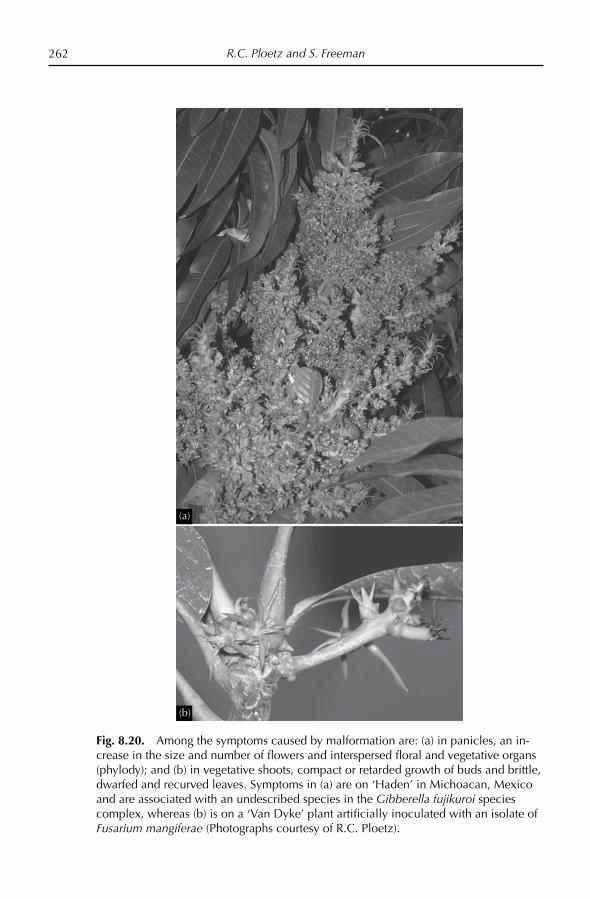
R.C. Ploetz and S. Freeman262
(a)
(b)
Fig. 8.20. Among the symptoms caused by malformation are: (a) in panicles, an in-crease in the size and number of fl owers and interspersed fl oral and vegetative organs (phylody); and (b) in vegetative shoots, compact or retarded growth of buds and brittle, dwarfed and recurved leaves. Symptoms in (a) are on ‘Haden’ in Michoacan, Mexico and are associated with an undescribed species in the Gibberella fujikuroi speciescomplex, whereas (b) is on a ‘Van Dyke’ plant artifi cially inoculated with an isolate of Fusarium mangiferae (Photographs courtesy of R.C. Ploetz).
(a)
(b)

Foliar, Floral and Soilborne Diseases 263
vegetative malformation. This pathogen has had several synonyms in the literature, including: Fusarium subglutinans (Wollenweb. and Reinking) Nelson, Toussoun and Marasas; F. moniliforme Sheldon var. subglutinans Wollenweb. and Reinking; and F. moniliforme Sheldon emend. Snyd and Hans. ‘Subglutinans’ sensu Snyd., Hans. and Oswald.
In 2002, 29 strains of the pathogen from Egypt, Florida USA, Israel, Malaysia and South Africa were redescribed as a new species in the Gibberella fujikuroi species complex (GFSC), Fusarium mangiferae Britz, Wingfi eld and Marasas (Steenkamp et al., 2000; Britz et al., 2002). Fusarium mangiferae resem-bles morphologically other members of the GFSC. It was established based on -tubulin and histone H3 DNA sequences, subtle morphological differ-ences, and because most of the examined strains had been shown in previous studies to cause malformation on artifi cially inoculated mango. The presence of F. mangiferae has been verifi ed in India (O’Donnell et al., 1998; Zheng and Ploetz, 2002), Oman (Kvas et al., 2007) and Spain (S. Freeman, Bet Dagan, 2007, unpublished results). Although a recent report from Pakistan mentions F. mangiferae, the identity of the pathogen there is not clear since the authors only discussed morphological characteristics of the pathogen (Iqbal et al., 2006).
Based on DNA sequence data (O’Donnell et al., 1998, 2000; Steenkamp et al., 1999, 2000), F. mangiferae is related to a lineage that includes Fusariumfujikuroi Nirenberg, Fusarium proliferatum (Matsushima) Nirenberg, and Fusarium sacchari (E. J. Butler) W. Gams (Marasas et al., 2006), and corresponds to the ‘Asian Clade’ of O’Donnell et al. (1998). Based on combined sequence data for fi ve genes, the closest known relative of F. mangiferae is an isolate from tropical rainforest soil in Papua New Guinea (Marasas et al., 2006).
Fusarium mangiferae produces white, fl occose mycelium on PDA with light- to dark-purple pigments in the agar (Leslie and Summerell, 2006). Cream-coloured sporodochia on carnation leaf agar (CLA) produce abun-dant thin-walled, long, slender and straight to slightly curved, three- to fi ve-septate macroconidia with curved apical cells and foot-shaped basal cells (Fig. 8.21). Single celled, rarely single septate, obovoid microconidia are produced in false heads on polyphialides with two to fi ve conidiogenous openings and on monophialides. Microconidial chains and chlamydospores are absent.
A second species, Fusarium sterilihyphosum Britz, Wingfi eld and Marasas, was described originally for isolates from a small area in South Africa (Britz et al., 2002). In subsequent work, it was detected in Brazil (Ploetz, 2003; K. O’Donnell, unpublished results), and was recently shown to cause malfor-mation in Brazil after artifi cial inoculation (Lima et al., 2006b). On PDA, colo-nies of F. sterilihyphosum produce white, fl occose mycelium with rose to light purple pigmentation in the agar (Leslie and Summerell, 2006). Uncommon, cream- to orange-coloured sporodochia are produced on CLA that produce rare, long, slender, three to fi ve-septate macroconidia (Fig. 8.22). On mono- and polyphialides, obovoid, oval to allantoid microconidia that are usually single celled are produced on false heads. Distinctive sterile coiled hyphae are produced by some isolates of this species.
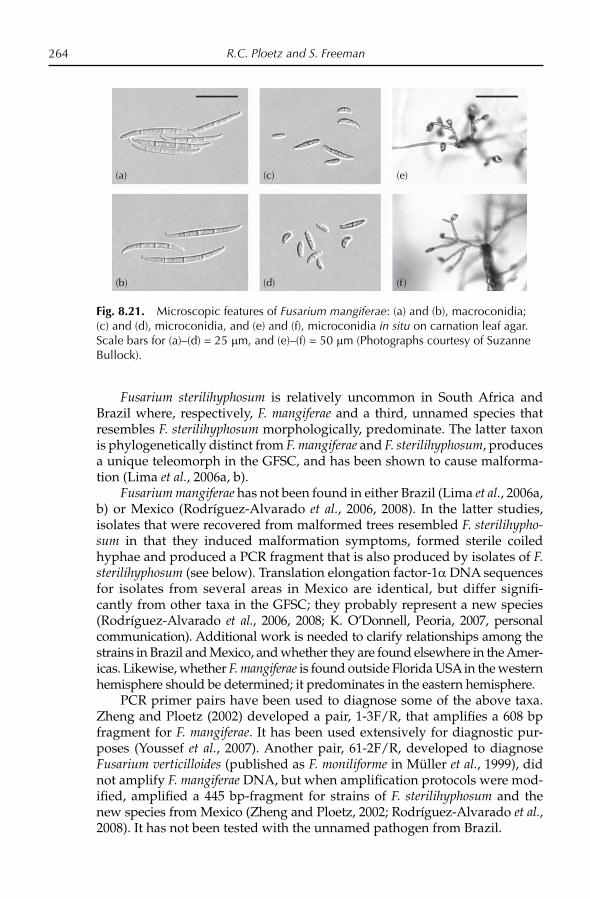
R.C. Ploetz and S. Freeman264
Fusarium sterilihyphosum is relatively uncommon in South Africa and Brazil where, respectively, F. mangiferae and a third, unnamed species that resembles F. sterilihyphosum morphologically, predominate. The latter taxon is phylogenetically distinct from F. mangiferae and F. sterilihyphosum, produces a unique teleomorph in the GFSC, and has been shown to cause malforma-tion (Lima et al., 2006a, b).
Fusarium mangiferae has not been found in either Brazil (Lima et al., 2006a, b) or Mexico (Rodríguez-Alvarado et al., 2006, 2008). In the latter studies, isolates that were recovered from malformed trees resembled F. sterilihypho-sum in that they induced malformation symptoms, formed sterile coiled hyphae and produced a PCR fragment that is also produced by isolates of F. sterilihyphosum (see below). Translation elongation factor-1 DNA sequences for isolates from several areas in Mexico are identical, but differ signifi -cantly from other taxa in the GFSC; they probably represent a new species (Rodríguez-Alvarado et al., 2006, 2008; K. O’Donnell, Peoria, 2007, personal communication). Additional work is needed to clarify relationships among the strains in Brazil and Mexico, and whether they are found elsewhere in the Amer-icas. Likewise, whether F. mangiferae is found outside Florida USA in the western hemisphere should be determined; it predominates in the eastern hemisphere.
PCR primer pairs have been used to diagnose some of the above taxa. Zheng and Ploetz (2002) developed a pair, 1-3F/R, that amplifi es a 608 bp fragment for F. mangiferae. It has been used extensively for diagnostic pur-poses (Youssef et al., 2007). Another pair, 61-2F/R, developed to diagnose Fusarium verticilloides (published as F. moniliforme in Müller et al., 1999), did not amplify F. mangiferae DNA, but when amplifi cation protocols were mod-ifi ed, amplifi ed a 445 bp-fragment for strains of F. sterilihyphosum and the new species from Mexico (Zheng and Ploetz, 2002; Rodríguez-Alvarado et al., 2008). It has not been tested with the unnamed pathogen from Brazil.
(a)
(b)
(c)
(d)
(e)
(f )
Fig. 8.21. Microscopic features of Fusarium mangiferae: (a) and (b), macroconidia; (c) and (d), microconidia, and (e) and (f), microconidia in situ on carnation leaf agar. Scale bars for (a)–(d) = 25 m, and (e)–(f) = 50 m (Photographs courtesy of Suzanne Bullock).
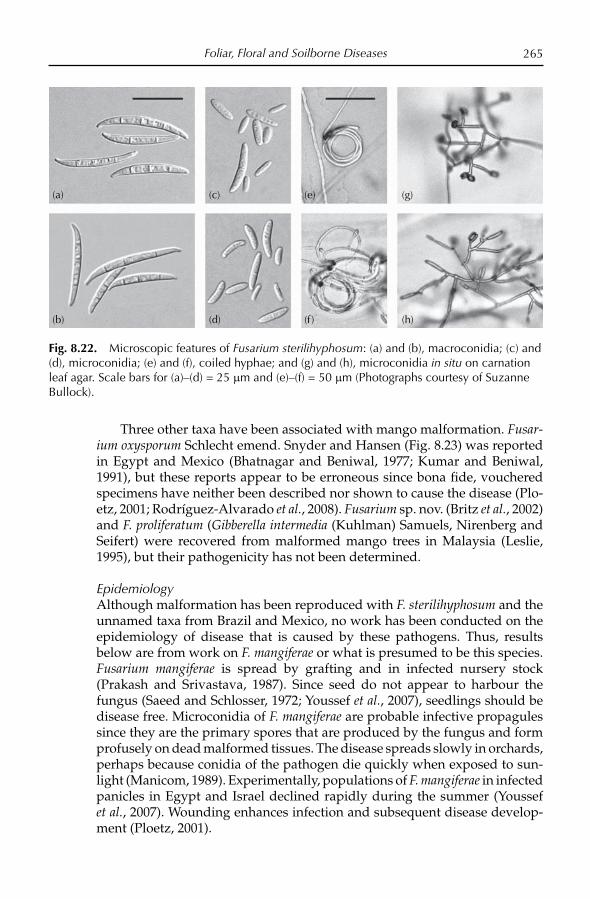
Foliar, Floral and Soilborne Diseases 265
Three other taxa have been associated with mango malformation. Fusar-ium oxysporum Schlecht emend. Snyder and Hansen (Fig. 8.23) was reported in Egypt and Mexico (Bhatnagar and Beniwal, 1977; Kumar and Beniwal, 1991), but these reports appear to be erroneous since bona fi de, vouchered specimens have neither been described nor shown to cause the disease (Plo-etz, 2001; Rodríguez-Alvarado et al., 2008). Fusarium sp. nov. (Britz et al., 2002) and F. proliferatum (Gibberella intermedia (Kuhlman) Samuels, Nirenberg and Seifert) were recovered from malformed mango trees in Malaysia (Leslie, 1995), but their pathogenicity has not been determined.
EpidemiologyAlthough malformation has been reproduced with F. sterilihyphosum and the unnamed taxa from Brazil and Mexico, no work has been conducted on the epidemiology of disease that is caused by these pathogens. Thus, results below are from work on F. mangiferae or what is presumed to be this species. Fusarium mangiferae is spread by grafting and in infected nursery stock (Prakash and Srivastava, 1987). Since seed do not appear to harbour the fungus (Saeed and Schlosser, 1972; Youssef et al., 2007), seedlings should be disease free. Microconidia of F. mangiferae are probable infective propagules since they are the primary spores that are produced by the fungus and form profusely on dead malformed tissues. The disease spreads slowly in orchards, perhaps because conidia of the pathogen die quickly when exposed to sun-light (Manicom, 1989). Experimentally, populations of F. mangiferae in infected panicles in Egypt and Israel declined rapidly during the summer (Youssef et al., 2007). Wounding enhances infection and subsequent disease develop-ment (Ploetz, 2001).
(a) (c) (e)
(b) (d) (f )
(g)
(h)
Fig. 8.22. Microscopic features of Fusarium sterilihyphosum: (a) and (b), macroconidia; (c) and (d), microconidia; (e) and (f), coiled hyphae; and (g) and (h), microconidia in situ on carnation leaf agar. Scale bars for (a)–(d) = 25 m and (e)–(f) = 50 m (Photographs courtesy of Suzanne Bullock).
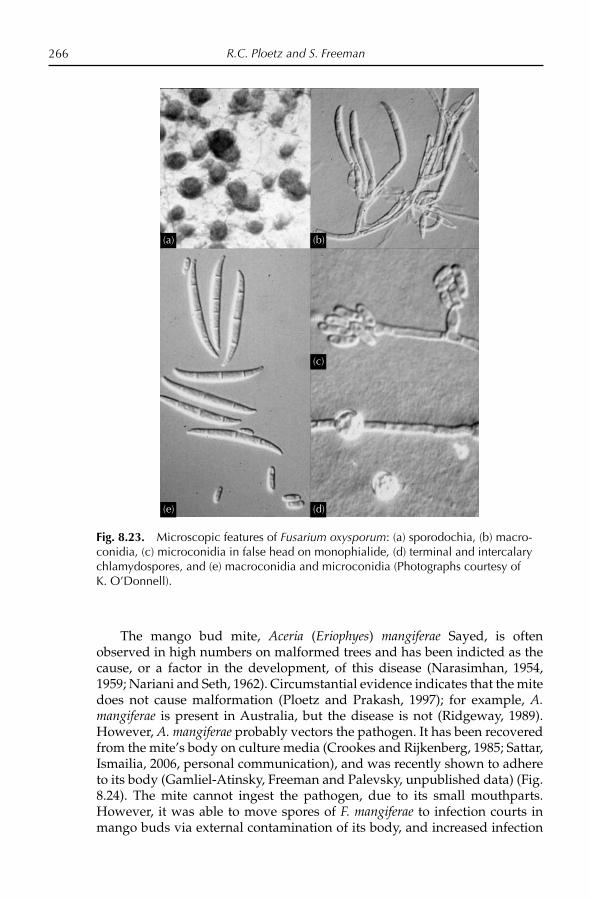
R.C. Ploetz and S. Freeman266
The mango bud mite, Aceria (Eriophyes) mangiferae Sayed, is often observed in high numbers on malformed trees and has been indicted as the cause, or a factor in the development, of this disease (Narasimhan, 1954, 1959; Nariani and Seth, 1962). Circumstantial evidence indicates that the mite does not cause malformation (Ploetz and Prakash, 1997); for example, A.mangiferae is present in Australia, but the disease is not (Ridgeway, 1989). However, A. mangiferae probably vectors the pathogen. It has been recovered from the mite’s body on culture media (Crookes and Rijkenberg, 1985; Sattar, Ismailia, 2006, personal communication), and was recently shown to adhere to its body (Gamliel-Atinsky, Freeman and Palevsky, unpublished data) (Fig. 8.24). The mite cannot ingest the pathogen, due to its small mouthparts. However, it was able to move spores of F. mangiferae to infection courts in mango buds via external contamination of its body, and increased infection
Fig. 8.23. Microscopic features of Fusarium oxysporum: (a) sporodochia, (b) macro-conidia, (c) microconidia in false head on monophialide, (d) terminal and intercalary chlamydospores, and (e) macroconidia and microconidia (Photographs courtesy of K. O’Donnell).
(a) (b)
(c)
(d)(e)

Foliar, Floral and Soilborne Diseases 267
of buds by the pathogen (Gamliel-Atinsky et al., 2007). Presumably, the mite’s feeding on buds facilitated infection. Aceria mangiferae does not appear to play a signifi cant role in disseminating the pathogen among trees. In Israel, mites were not found in traps that were designed to monitor their movement in a malformed mango orchard, although high numbers of F. mangiferae conidia were recorded in the air in this orchard (Gamliel-Atinsky et al., 2007).
The distribution of F. mangiferae in affected trees suggests that vegetative and fl oral buds are the primary sites of infection and that systemic coloniza-tion of older, subtending tissues does not occur. Freeman et al. (1999) trans-formed isolates of F. mangiferae from mango with the uidA reporter gene ( -glucuronidase), and used them to artifi cially inoculate mango. Their results verifi ed that bud and fl ower tissues of the host are primary infection sites, and that wounds provide points of entry for the pathogen. In Florida USA, F. mangiferae was restricted almost entirely to malformed fl oral and vegetative tissues (Ploetz, 1994). Levels of infection were highest in malformed fl owers and vegetative shoots (c.65–85%), were much lower or non-existent in asymp-tomatic tissues (0–11%), and were rare in branches (0–4%) even when they supported malformed fl owers or shoots. Remnant infections of F. mangiferae in scaffold branches and trunks were restricted almost exclusively to branch scars or dormant apices (Freeman, unpublished data). Reports of root infection by F. oxysporum (Kumar and Beniwal, 1991) or F. mangiferae (Abdel-Sattar, 1973; Kumar and Beniwal, 1991) causing malformation in seedling plants have not been corroborated. Although roots can be infected by
GFP-labelled microconida ofFusarium mangiferae
20.0 μm
Aceria mangiferae
Fig. 8.24. Body of the mango bud mite, Aceria mangiferae, to which green fl uorescent protein (gfp)-labelled microconidia of Fusarium mangiferae have adhered (Photograph courtesy of E. Gamiel-Atinsky).

R.C. Ploetz and S. Freeman268
F. mangiferae, these infections are not systemic and do not appear to result in symptom development (Youssef et al., 2007).
The localized and variable levels of infection by F. mangiferae that have been noted in diseased and non-symptomatic tissue (Ploetz, 1994; Youssef et al., 2007), suggest that there are thresholds of infection, whereby malfor-mation develops only after a suffi cient proportion of a host meristem is colo-nized by the pathogen. This hypothesis is supported by the long latent period that exists before symptoms develop in artifi cially inoculated plants and the hormonal perturbations that probably occur when meristematic tissues are infected with this pathogen (van Staden et al., 1989; van Staden and Nichol-son, 1989; Ploetz, 2001).
ManagementManagement of malformation can be diffi cult. New plantings should be established with pathogen-free nursery stock. Scion material should never be taken from an affected orchard, and any affected plants that are observed in the nursery should be removed and burned immediately. Nurseries should not be established in orchards that are affected by malformation. Once the disease is found in an orchard, control is possible, but time consuming. In these cases, cultural management has been most effective (Narisimhan, 1959; Singh et al., 1974; Manicom, 1989). All affected terminals and the subtending three nodes are cut from trees, removed from the fi eld and burned. Unfortu-nately, producers may be unwilling to devote the effort that is required to ensure that this approach succeeds. In addition, it may be diffi cult to impose this treatment on large trees.
A diverse array of pesticides, hormones and growth regulators has been tested against malformation, but these measures have been marginally effec-tive. Singh et al. (1994) obtained moderate control with sulfates of cobalt (Co), cadmium (Cd) and nickel (Ni) in India, but it is unlikely that these toxic com-pounds could be used safely. Darvas (1987) reduced the percentage of mal-formed infl orescences from 96% to 48% by injecting ‘Keitt’ trees with the fungicide fosetyl-Al. This reduction was signifi cant (P < 0.05), but the increase in fruit yield, 46–95 kg of fruit/tree, was not. Other fungicidal compounds have been generally less effective (Diekman et al., 1982; Chakrabarti and Ghosal, 1989). In general, the protected, internal location of the pathogen in affected trees makes it diffi cult to control this disease. When applied as foliar sprays or via continuous drip irrigation, prochloraz reduced the severity of malformation signifi cantly in Israel, but this was dependent on the timing of application (Freeman et al., unpublished data). Although disease was not completely controlled, this and other systemic fungicides might be useful in future integrated management programmes that would incorporate other measures such as removal of symptomatic terminals and use of tolerant cultivars.
Prakash and Srivistava (1987) indicated that ‘There is great variation in the susceptibility of existing varieties.’ Unfortunately, controlled inoculations have not been used to determine cultivar resistance, and these reports have come from non-replicated tests; cultivars listed as ‘resistant’ may have come

Foliar, Floral and Soilborne Diseases 269
from healthy nursery stock or may have escaped infection once planted in the fi eld (Ploetz, 2001). For example, Bastawros (1996) reported that two newly introduced cultivars in Egypt, ‘Kent’ and ‘Keitt’, were immune (0% disease); however, these cultivars are susceptible in Florida USA (Ploetz, unpublished data). Controlled inoculations with grafted plants of different genotypes and quantifi ed levels of virulent isolates of F. mangiferae have not been reported.
Recently, Ploetz (unpublished data) utilized previously described proto-cols (Freeman et al., 1999) to assess malformation development on grafted plants of diverse genotypes. Disease development was affected by: the length of time after inoculation and inoculated apical buds remained dormant after inoculation; the isolate of F. mangiferae that was used for inoculation; and mango genotype. Virulent isolates, patience (latent period ranged from 40 to 210 days), and suffi cient replication were needed to successfully conduct screenings for response to malformation. Future work is warranted to investi-gate attributes that are related, and might predict resistance, to this disease.
The symptoms of malformation suggest that a hormone imbalance occurs in affected tissues. Singh and Dhillon (1989) assayed levels of indole acetic acid (IAA), gibberellic acid (GA3) and zeatin in malformed and healthy mango seedlings. Whereas IAA and GA3 levels were, respectively, about ten and fi ve times lower in malformed plants, levels of zeatin were about fi ve times higher. Van Staden and colleagues (Nicholson and van Staden, 1988; van Staden and Nicholson, 1989; van Staden et al., 1989) examined specifi c cytokinins produced by mango and ‘F. moniliforme’ (presumably F. mangiferae). They determined that the cytokinin complements in healthy and malformed panicles differed qualitatively and quantitatively, and that the pathogen pro-duced some of the hormones and metabolites that were implicated in dis-ease development. However, it was impossible to assign unequivocal roles for production of hormones by the host and pathogen and the subsequent development of symptoms. For example, whether production of hormones by the pathogen directly caused the noted changes or whether hormone pro-duction by the host was somehow altered in the presence of the pathogen was not clear. Additional work would be needed to clarify these interactions, and whether F. sterilihyphosum and the unnamed pathogens in Brazil and Mexico also produce cytokinins or other hormones in affected plants.
Parasitic plants
The family Loranthaceae contains several parasitic plant species that affect mango. In Malaysia, Dendrophthoe (fomerly Loranthus) pentandra Linn. is the most important species (Lim and Khoo, 1985). Other Dendrophthoe spp., Elytranthe spp. and Viscum spp. are also known in Malaysia, but are less important. In India, Dendrophthoe falcata (formerly Loranthus longifl orus) is common, and other, less frequently encountered, species include Macrosolen cochinchinensis, Helicanthes elasticus and Elytranthe capitellata (Majumder and Sharma, 1990). These parasites are usually only important in neglected

R.C. Ploetz and S. Freeman270
orchards. Although they are photosynthetically self-suffi cient, the plants obtain water and minerals from the host plant via haustoria that penetrate and colonize the host vascular system. In severe cases, the removal of water and minerals from parasitized branches is suffi cient to reduce the vitality and yields of trees.
Since the appearance of these plants is distinct from the mango host, they are easily distinguished in infected trees (Lim and Khoo, 1985). The points at which the mango host is penetrated are usually characterized by swollen growths called burrs. Lim and Khoo (1985) and Majumder and Sharma (1990) indicated that affected portions of trees should be removed far enough below burrs to remove haustoria of the parasite. After affected tissues are removed, cut sur-faces can be treated with creosote or other wound dressings. These plants can also be treated with herbicides, such as 2,4-dichlorophenoxyacetic acid (2,4-D), but these are dosage sensitive treatments and pose a risk to the host plant.
Phoma blight
Phoma blight is widespread in India (Prakash and Singh, 1977). It occurs only on old leaves. Initially, lesions are minute and yellow-brown (Prakash and Singh, 1977). As they expand they darken to brown or cinnamon, become irregular, and may ultimately develop dark margins and dull-grey centres. In severe cases, necrotic patches as large as 13 cm in diameter may form that cause defoliation. The disease is caused by Phoma glomerata (Corda) Wollenw. and Hochapf (Prakash and Singh, 1977). It forms globose to obpyriform, light-coloured to car-bonaceous pycnidia that average 30–400 m in diameter. Pycnidia have one to three ostioles, form singly or in clusters, and have short necks. On PDA, pyc-nidia and conidia are abundant. Conidia are hyaline to dark coloured, ovoid to ellipsoid, unicellular or occasionally bicellular, and average 8.3 × 3.2 m.
Phoma leafspot
Another Phoma sp. causes a leafspot in India (Prakash and Singh, 1976b), and is referred to as phoma leafspot. On young leaves, Phoma sorghina (Sacc.) Boerema. Doren. and Vankest causes irregular to roughly circular, water-soaked spots, up to 2.5 mm in diameter. Lesions are brown with a yellow to brown margin. Lesions on midribs are elongated and more conspicuous, and may coalesce to up to 14 cm in diameter. They can be confused with those caused by anthracnose.
Pink disease
A basidiomycete, Erythricium salmonicolor (Berk. and Broome) Burdsall (syn-onyms: Corticium salmonicolor Berk. and Broome, and Phanerocbaete salmoni-color (Berk. and Broome) Jülich; anamorph: Necator decretus Massee) causes

Foliar, Floral and Soilborne Diseases 271
pink disease. Pink disease affects many economically important woody plants in the humid tropics, where it is one of the most destructive diseases of mango (Holliday, 1980). The disease is also known as cobweb, rubellosis and thread blight (Prakash and Srivistava, 1987). It has been most thor-oughly studied on rubber, Hevea brasiliensis, an important host in South-east Asia (Rao, 1975). On mango, pink disease can signifi cantly reduce tree vigour and yield, especially in 6- to 15-year-old trees (Lim and Khoo, 1985).
Symptoms fi rst appear as white, felty mycelial threads on twigs and branch crotches (Lim and Khoo, 1985). If favourable conditions persist, the mycelial threads coalesce to form a rough, pink crust on the bark surface. This stage usually takes 1 to several months to develop and coincides with the penetration of bark and internal colonization of the tree. Affected bark often cracks. As the fungus kills the vascular and cambial areas beneath the bark, branches above the colonized areas die, resulting in a sparse, patchy canopy.
Two types of sporulation occur (Holliday, 1980; Lim and Khoo, 1985). Erythricium salmonicolor produces a smooth, clammy, pinkish white hyme-nium over the pink crust it forms on bark. Basidiospores form on the hyme-nium and are borne on sterigmata on narrowly clavate to cylindrical basidia (Fig. 8.25). Basidiospores are hyaline, broadly ellipsoidal and 8–10 × 5–7 m. Conidia of N. decretus, which are produced in reddish-orange sporodochia, are hyaline, ellipsoidal, unicellular and 10–18 × 6–12 m. Although the infec-tion process has apparently not been studied in mango, basidiospores can infect rubber trees (Holliday, 1980). The anamorph and teleomorph are
(c)
(a)
(b) (f )
(e)
(d)
100 μ
20 μ
Fig. 8.25. (a) Conidium-bearing pustule, and (f) conidiogenous cells and conidia of Necator decretus, and (b) hymenium, (c) basal hyphae, (d) immature and mature basidia, and (e) basidiospores of its teleomorph, Erythricium salmonicolor (Source: from CMI description no. 511).

R.C. Ploetz and S. Freeman272
formed during wet conditions, and conidia and basidiospores are dispersed by rainsplash and wind.
Pink disease management relies on early detection and removal of affected tissues from orchards. When removal of syptomatic tissues is imprac-tical, control depends upon treatment with fungicides. Several protectant and systemic fungicides are effective (Lim, 1994). They should be used as soon as symptoms are evident, and as long as the disease is present. All cul-tivars of mango that have been tested in Malaysia are susceptible (Lim and Khoo, 1985).
Powdery mildew
Powdery mildew is a widespread and important disease of leaves, panicles and fruit. The disease can result in yield reductions as high as 90%, due mainly to its effect on fruit set and development (Schoeman et al., 1995).
SymptomsMango cultivars vary in their response to powdery mildew (Palti et al., 1974). On the most susceptible cultivars, virtually all foliar, fl oral and fruit parts of the plant are affected (Plate 46). Powdery growth can cover all tissues on panicles, resulting in a brown, shrivelled necrosis. Since fruit set and reten-tion can be affected, the disease can have a profound impact on yield. Foliage can also be damaged signifi cantly, and young leaves are most susceptible. White, powdery coatings of conidia develop on either side or both sides of leaves, depending on the cultivar. When damage occurs on the undersides of leaves it is often restricted to the mid-rib. In all cases, leaves become dis-torted, and affected areas turn purple and ultimately necrotic.
AetiologyPowdery mildew is caused by Oidium mangiferae Berthet, a host-specifi c fun-gus (Prakash and Srivistava, 1987; Ploetz and Prakash, 1997). It was fi rst described in Brazil (Berthet, 1914), and is now recognized in most mango-producing regions (Palti et al., 1974). Conidium and haustorium traits indi-cate that O. mangiferae belongs to the Erysiphe polygoni group (Johnson, 1994a). Although the pathogen was originally classifi ed as Erysiphe cichoracearum by Wagle (1928), Uppal et al. (1941) noted that it produced saccate and lobed appressoria, which are not characteristic of E. cichoracearum. The pathogen has no known teleomorph, a common trait for powdery mildew fungi in the tropics (Holliday, 1980). Conidia of O. mangiferae are unicellular, hyaline, elliptical to barrel-shaped and measure 33–43 × 18–28 m (Uppal et al., 1941; Palti et al., 1974). They are produced in large numbers on host surfaces, and impart a powdery appearance to affected tissues (Plate 46). The lengths of germ tubes vary depending upon RH, and they terminate in appressoria. Glob-ular haustoria form in host epidermal cells. Conidiophores are of the pseudoid-ium type, with two to four septa and a straight basal cell (Boesewinkel, 1980).

Foliar, Floral and Soilborne Diseases 273
EpidemiologyPowdery mildew is most severe during cool, dry weather. Conidia are dis-seminated by wind and are released on a diurnal basis (Schoeman et al.,1995). Peak spore release, between 11:00 to 16:00 h, was positively correlated with hourly temperature and negatively correlated with RH, vapour pres-sure defi cit and leaf wetness (all P < 0.01). Conidia germinate at temperatures ranging from 9 to 32°C (23°C is optimal), and at RH as low as 20% (Palti et al.,1974). Since germination occurs in such a wide range of RH, disease develop-ment is usually independent of this parameter. Infection can occur within 5–7 h, and conidia are produced within 5 days of infection. Disease develop-ment occurs within a very broad range of temperatures, 10–31°C. Gupta (1989) reported that dry weather encouraged disease development.
ManagementMango cultivars vary in their resistance to powdery mildew (Palti et al.,1974). ‘Zill’, ‘Kent’, ‘Alphonso’, ‘Seddek’ and ‘Nam Doc Mai’ are very sus-ceptible; ‘Haden’, ‘Glenn’, ‘Carrie’, ‘Zebda’, ‘Hindi be Sennara’, ‘Ewaise’ and ‘Keitt’ are moderately susceptible; and ‘Sensation’, ‘Tommy Atkins’ and ‘Kensington’ are slightly susceptible (Ploetz et al., 1994; Nofal and Haggag, 2006). In India, Tiwari et al. (2006) reported that ‘Baigan Phalli’, ‘Barbalia’, ‘Dabari’, ‘Dilpasand’, ‘Khirama’, ‘Nagarideeh’, ‘Oloor’ and ‘Totapari’ were highly resistant and ‘Amrapali’ was most susceptible.
Schoeman et al. (1995) recommended that the fi rst fungicide application to control this disease should occur when panicles begin to change colour. Assuming an effective period of 3 weeks for a given application, they con-cluded that applications should continue every third week until panicle sus-ceptibility decreased at the end of fruit set. Powdery mildew is easily controlled with S (Johnson, 1994a). Other fungicides are effective, but many have negative environmental impacts (Ray, 2003; Tavares et al., 2004). Foliar sprays of di-potassium hydrogen orthophosphate (K2HPO4) and potassium di-hydrogen orthophosphate (KH2PO4), systemic fungicides, and an alterna-tion of fertilizer and systemic fungicides inhibited powdery mildew on pan-icles (Reuveni et al., 1998; Nofal and Haggag, 2006). Treatments of the fertilizers with half or a quarter of the recommended rate of sterol-inhibitor fungicides and Kresoxym-methyl provided protection comparable with or superior to that of standard fungicides alone (Oosthuyse, 1998; Reuveni et al., 1998). Sulfur can burn fl owers and young fruit during warm, sunny condi-tions (Johnson, 1994a), and three fungicides used during bloom, dinocap, fenbuconazole and hexaconazole, can reduce pollen germination (Dag et al., 2001).
Scab
Elsinoë mangiferae Bitancourt and Jenkins (anamorph: Sphaceloma mangiferae Bitancourt and Jenkins) causes scab on mango (Bitancourt and Jenkins, 1943; Cook, 1975). The disease was fi rst recognized in Cuba and Florida USA in the

R.C. Ploetz and S. Freeman274
1940s and is now widespread in the western hemisphere. Scab is important in nurseries since young host tissues are most susceptible, and because moist environments aid infection (Ruehle and Ledin, 1955). Lesions, usually fi rst observed on the underside of leaves, are dark brown to black, and 1–2 mm in diameter. They may enlarge to 5 mm in diameter and become light grey with narrow, dark borders. Affected foliage develops a distorted appearance, and greyish blotches are produced on stems.
Elsinoë mangiferae produces brownish ascocarps, 30–48 × 80–160 m, in the host epidermis. Globular asci, 10–15 m in diameter, are dispersed in ascocarps, and contain one to eight hyaline ascospores. Ascospores are 4–6 × 10–13 m, three septate and constricted at the median septum; the sub-apical cell is longitudinally septate. Conidiophores of S. mangiferae are erect, sinuous, 2.5–3.5 × 12–35 m, and wider at the base. Conidia are brown, one or two celled, and 2–4 × 6–29 m.
Young host tissues are most susceptible. Rainy weather promotes sporu-lation of the fungus, but specifi c information is not available on the epidemi-ology of scab. Whether conidia and ascospores are infectious is not known.
Seca and sudden decline
This is a disease that is known by several different names in Brazil and the Middle East and is the only one that routinely kills mango trees. ‘Seca’ (dry-ing), ‘murcha’ (withering), branch blight and Recife sickness in Brazil, was fi rst recognized in Pernambuco in 1938, and is now also found in Bahia, Goias, the Federal District, Rio de Janiero and São Paulo (Ribeiro, 1997; Colo-simo et al., 2000; Silveira et al., 2006). It threatens neighbouring states due to its effi cient movement in infected propagation materials, on pruning equip-ment, and via a mobile beetle vector.
In 1998, a disease termed ‘sudden decline’ began to kill trees in Oman (Al Adawi et al., 2003, 2006), about the same time a similar problem (i.e. quick decline or sudden death) was observed in Pakistan (Malik et al., 2005). In many ways these diseases resembled seca. Circumstantial evidence sug-gested that the disease was introduced from Brazil by a producer with orchards in Oman and Pakistan (M. Deadman, Muscat, 2005, personal com-munication). By 2007, many mango-producing areas in Oman and Pakistan were affected and uncontrolled dissemination of infected germplasm may have spread the disease elsewhere in the region. Its spread into India should be investigated (A.W. Cooke, Indooropilly, 2007, personal communication).
SymptomsSymptoms include: discoloration of the vascular cambium; exudation of an amber-coloured gum from the trunk and branches, particularly from galler-ies of the putative beetle vector of the pathogen; wilting; rapid death of branches and entire trees without defoliation; and a scorched appearance of dead trees (Plate 47) (Junqueira et al., 2002; Al Adawi et al., 2006). On grafted trees, scions, rootstocks or both may be susceptible and exhibit vascular

Foliar, Floral and Soilborne Diseases 275
symptoms. In Oman, where susceptible Omani seedlings are used as root-stocks, the disease is frequently a problem of rootstocks (Al Adawi et al., 2006), whereas in Brazil, the disease is associated with the scion (P < 0.01) (Colosimo et al., 2000); rootstock cultivar had an insignifi cant impact on dis-ease development in the latter work (P > 0.05). When disease development begins in the canopy, symptoms may initiate in a branch or portion of a tree, but death of the entire plant usually follows. Where root/rootstock infection is involved, sudden death of the entire tree usually occurs.
AetiologyCeratocystis fi mbriata Ellis and Halst. sensu lato (s.l.) (anamorph: Thielaviopsis sp.) was reported in Brazil in the 1930s (Viegas, 1960; Ribiero, 1980; Silveira et al., 2006), and is recognized as the primary cause of seca. Diplodia recifi ensis Batista (= Lasiodiplodia theobromae?) was indicted as the cause of Recife sick-ness in Brazil (Batista, 1947), but it probably plays no role or a secondary role in the development of this disease (see below). In Oman, C. fi mbriata s.l. causes sudden decline, but L. theobromae and Ceratocystis omanensis Al Subhi, M.J. Wingf., M. van Wyk and Deadman are also associated with the disease (Al Adawi et al., 2006; van Wyk et al., 2007). The ease with which L. theobromae and the diffi culty with which C. fi mbriata s.l. are recovered from affected trees may have been responsible for previous assumptions that ‘D. recifi ensis’ caused Recife sickness in Brazil and L. theobromae caused sudden decline in Oman (Batista, 1947; Ploetz and Prakash, 1997; Al Adawi et al., 2003, 2006).
Ceratocystis contains many pathogens, particularly of trees (Kile, 1993). The wide host range of C. fi mbriata s.l. led Webster and Butler (1967) to hypothesize that it was a species complex, and DNA sequences have begun to delineate some of the host-specifi c, often morphologically indistinct, taxa in the species (van Wyk et al., 2007). A contemporary view is that C. fi mbriata sensu stricto (s.s.) specifi cally refers to the cause of black rot of sweet potato (Ipomoea batatas L.) on which it was fi rst described (Halsted and Fairchild, 1891). Other cryptic, monophyletic lineages of C. fi mbriata s.l. have been described as distinct species (Engelbrecht and Harrington, 2005; Johnson et al., 2005; van Wyk et al., 2005, 2007), and more will likely follow.
Two new Ceratocystis spp. have been described on mango in the Oman Gulf region. Ceratocystis omanensis belongs to the Ceratocystis moniliformis Hedgc. s.l. species complex (Al Subhi et al., 2006), which are typically not primary pathogens. Ceratocystis omanensis is a minor pathogen on mango. The primary sudden decline agent in Oman and Pakistan, C. fi mbriata s.l., represents a monophyletic lineage based on ITS, -tubulin and translation elongation factor (TEF) 1- DNA sequence comparisons, and it has unique morphological characteristics; it was renamed Ceratocystis manginecans M. van Wyk, A Al Adawi and M.J. Wingf. sp. nov. (van Wyk et al., 2007).
On 2% malt extract agar (MEA), colonies of C. manginecans are greyish olive and have a banana odour (van Wyk et al., 2007). Hyphae are smooth and segmented (Fig. 8.26). Ascomatal bases are globose, black and (153–)192–254(–281) m in diameter; ascomatal necks are dark brown, lighter towards the apices (514–)557–635(–673) m long, (25–)32–42(–48) m, wide at the
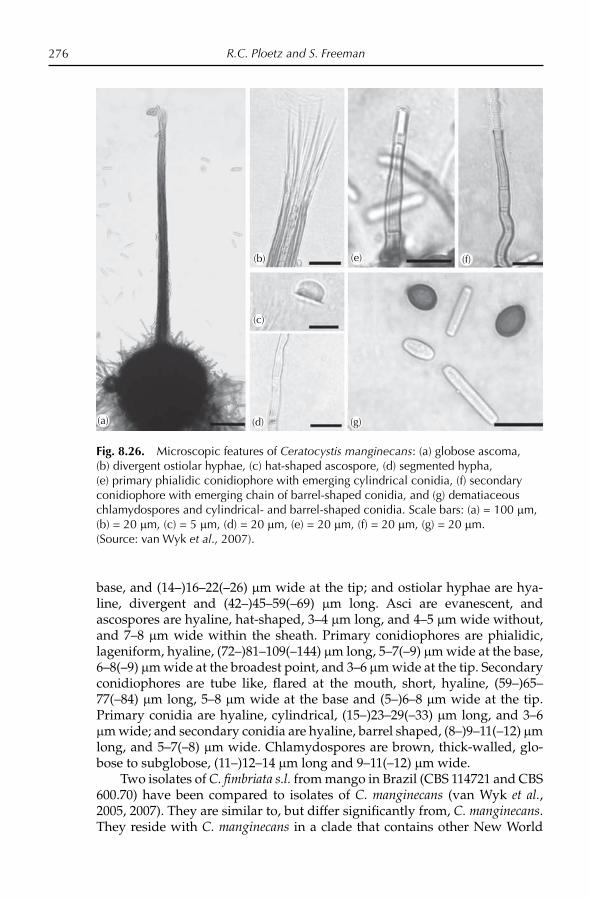
R.C. Ploetz and S. Freeman276
base, and (14–)16–22(–26) m wide at the tip; and ostiolar hyphae are hya-line, divergent and (42–)45–59(–69) m long. Asci are evanescent, and ascospores are hyaline, hat-shaped, 3–4 m long, and 4–5 m wide without, and 7–8 m wide within the sheath. Primary conidiophores are phialidic, lageniform, hyaline, (72–)81–109(–144) m long, 5–7(–9) m wide at the base, 6–8(–9) m wide at the broadest point, and 3–6 m wide at the tip. Secondary conidiophores are tube like, fl ared at the mouth, short, hyaline, (59–)65–77(–84) m long, 5–8 m wide at the base and (5–)6–8 m wide at the tip. Primary conidia are hyaline, cylindrical, (15–)23–29(–33) m long, and 3–6
m wide; and secondary conidia are hyaline, barrel shaped, (8–)9–11(–12) m long, and 5–7(–8) m wide. Chlamydospores are brown, thick-walled, glo-bose to subglobose, (11–)12–14 m long and 9–11(–12) m wide.
Two isolates of C. fi mbriata s.l. from mango in Brazil (CBS 114721 and CBS 600.70) have been compared to isolates of C. manginecans (van Wyk et al., 2005, 2007). They are similar to, but differ signifi cantly from, C. manginecans. They reside with C. manginecans in a clade that contains other New World
(a) (d) (g)
(c)
(e)(b) (f)
Fig. 8.26. Microscopic features of Ceratocystis manginecans: (a) globose ascoma, (b) divergent ostiolar hyphae, (c) hat-shaped ascospore, (d) segmented hypha, (e) primary phialidic conidiophore with emerging cylindrical conidia, (f) secondary conidiophore with emerging chain of barrel-shaped conidia, and (g) dematiaceous chlamydospores and cylindrical- and barrel-shaped conidia. Scale bars: (a) = 100 m,(b) = 20 m, (c) = 5 m, (d) = 20 m, (e) = 20 m, (f) = 20 m, (g) = 20 m.(Source: van Wyk et al., 2007).

Foliar, Floral and Soilborne Diseases 277
species, Ceratocystis cacaofunesta and Ceratocystis platani. Research is needed to: (i) examine additional isolates of C. fi mbriata s.l. from mango in Brazil; (ii) describe the putative, new species; (iii) determine whether C. manginecans is present in Brazil; and (iv) clarify pathogenic variation in the agent(s) in Oman and Brazil. At least two pathotypes of C. fi mbriata s.l. are evident in Brazil (Rossetto et al., 1996; Junqueira et al., 2002; Silveira et al., 2006).
Rossetto et al. (1996) evaluated 15 cultivars against two isolates of the pathogen in Brazil. Eight-year-old trees were inoculated c.40 cm beneath branch apices with IAC FITO 4905, which is pathogenic to ‘Jasmim’, and IAC FITO 334-1, which is not. ‘São Quirino’, ‘Irwin’, ‘Edwards’ and ‘Van Dyke’ were resistant, and ‘IAC 100 Bourbon’ was moderately resistant. ‘Glenn’, ‘Joe Welch’, ‘Zill’ and ‘Haden’ were susceptible, and ‘Kent’ responded as ‘Jas-mim’, resisting IAC FITO 334-1 and succumbing to IAC FITO 4905.
EpidemiologyGenotype has a profound impact on disease development, and severe epi-demics occur wherever susceptible rootstocks and/or scions are used. Greater disease develops when trees are stressed, although it is not clear whether this results from an increased attraction of the vector to stressed trees or reduced disease resistance in the host. The associated pathogens are moved easily in infected germplasm, the route by which the diseases have spread in Brazil and probably to Oman. Pruning implements also move the pathogen, and soil, once infested with chlamydospores of the pathogen, can be a long-term reser-voir of inoculum. Insect dissemination plays a particularly insidious role.
Beetles (Coleoptera: Scolytidae) are closely associated with seca in Brazil (Batista, 1947; Viegas, 1960; Piza, 1966; Ribiero, 1980). Batista (1947) indicated that Xyleborus affi nis was the sole vector of D. recifi ensis. In contrast, Ribiero (1980) reported that Hypocryphalus mangiferae Stebbing was the primary vec-tor of C. fi mbriata s.l. (Fig. 8.27). It produced galleries in the cambium of affected trees (Plate 47a), and was the only scolytid found on healthy, as well as diseased, trees. Hypocryphalus mangiferae is also associated with the dis-eases in Oman and Pakistan, where C. manginecans is recovered from the insect and trees are commonly found with insect probing damage before dis-ease develops (Al Adawi et al., 2006; van Wyk et al., 2007).
The interactions between H. mangiferae and the Ceratocystis pathogens of mango are incompletely understood. In olfactometer tests in Brazil, H. mangiferae was attracted to cultures of C. fi mbriata s.l., and larvae of the insect were raised to adulthood on the fungus (Ribiero, 1980). Several other species, many of which are in the genus Xyleborus, were also associated with seca, but because they were found only in diseased trees they appeared to be opportunistic feeders on C. fi mbriata s.l. rather than primary vectors. Although the sequence of events in Brazil and the Oman Gulf has not been studied, it is probable that H. mangiferae contaminates its body with these pathogens while feeding in diseased trees and subsequently disseminates the pathogen to healthy trees.
Hypocryphalus mangiferae is thought to be native to some of the same areas in southern Asia where mango evolved (Wood, 1982; Butani, 1993;

R.C. Ploetz and S. Freeman278
Atkinson and Peck, 1994; Mukherjee, 1997). Thus, the insect would have been introduced into Brazil and would have been a new encounter, rather than coevolved, relationship with C. fi mbriata s.l. (van Wyk et al., 2007). In contrast, if C. manginecans were introduced into Oman and Pakistan from Brazil, it may have then established a relationship with a native insect. Although the available information suggests that the H. mangiferae– Ceratocystis interactions on mango were recent, opportunistic encounters in the New World, additional work is needed.
ManagementGiven the ease with which these pathogens are moved and their destructive impact, preventing their dissemination to new areas must be a high priority. Pathogen-free propagation material should be used whenever new plantings are established and germplasm is moved. Clean pruning implements should be used in affected areas, and should be frequently disinfested with bleach, formalin or other disinfestants (Junqueira et al., 2002). Trees that have been killed by the disease must be removed and destroyed as they are signifi cant reservoirs of inoculum and infested vectors. Where partially resistant culti-vars are grown, the removal and burning of affected branches and treatment of the exposed branch stubs with Cu fungicides is recommended (Ribeiro et al., 1995; Ribeiro, 1997).
Managing these diseases with fungicides on susceptible cultivars would be a challenge. External applications of protectant or systemic fungicides would probably be ineffective, given the internal, protected location of the pathogen. Injecting fungicides, as is done to control Dutch elm disease, might be effective. However, this would probably not be allowed where concerns
Fig. 8.27. Hypocryphalus mangiferae, vector of the seca and sudden decline pathogens (Photograph courtesy of R.C. Ploetz).

Foliar, Floral and Soilborne Diseases 279
with pesticide contamination of fruit exist. Treatment of germplasm collec-tions and young, non-bearing trees might be the only situations in which fungicide injection would be possible.
Genetic resistance offers the best hope for managing these diseases. Var-ious levels of tolerance have been observed in Brazil and resistant clones have been developed. However, pathogenic variation in the causal fungus in Brazil has hindered progress (Rossetto et al., 1996; Junqueira et al., 2002; Sil-veira et al., 2006). Although disease responses of some genotypes vary in dif-ferent production areas in the country, ‘Manga D’agua’, ‘Pico’, ‘IAC 101’, ‘IAC 102’, ‘Edwards’, ‘Van Dyke’ and ‘Carabao’ resist two races of the patho-gen, and ‘Rosa’, ‘Sabina’, ‘Sao Quirino’, ‘Oliveira Neto’, ‘Jasmim’, ‘Sensation’, ‘Irwin’ and ‘Tommy Atkins’ are generally tolerant (Ribiero, 1997; Junqueira et al., 2002). ‘Kent’ and ‘Jasmim’ respond differentially (see above), and ‘Coquinho’, ‘Glenn’, ‘Joe Welch’, ‘Zill’ and ‘Haden’ are susceptible. Although ‘Espada’ is also reported to be tolerant, old trees are frequently attacked. In commercial orchards, the disease on ‘Espada’ is managed by grafting onto resistant rootstocks and pruning diseased branches. Colosimo et al. (2000) worked with other scion cutivars, although in a single location (and against a single pathotype?). They reported that ‘Oliveira’ was most resistant, ‘Car-lota’, ‘Imperial’, ‘Extrema’ and ‘Pahiri’ had intermediate resistance, and ‘Bourbon’ was most susceptible.
One must also recognize the impact of other diseases on different cultivars. Carvalho et al. (2004) described two new cultivars, ‘IAC 103 Espada Vermelha’ and ‘IAC 109 Votupa’, which had moderate resistance to seca. ‘IAC 103 Espada Vermelha’ also had moderate resistance to powdery mil-dew but was susceptible to anthracnose. Both cultivars were susceptible to malformation.
Stigmina leafspot
Stigmina leafspot is caused by Stigmina mangiferae (Koorders) Ellis (synonym: Cercospora mangiferae Koorders; a teleomorph for the fungus is not known). Lim and Khoo (1985) indicated that the disease occurs on a range of cultivars, and is most severe during rainy weather. Both young and old leaves are affected. Dark-brown spots, 1–2 mm in diameter, are formed initially by the fungus. These can enlarge and coalesce to 1 cm or larger, and are surrounded by conspicuous chlorotic haloes that aid diagnosis of this disease. The fungus produces large, olive-grey conidia, 30–60 × 3.5–5.0 m, usually on the lower leaf surface (Fig. 8.28). Conidia are wider at the base than the apex, are straight to curved, have three to seven septa, and are borne in subglobular, dark stromata that are 20–60 m in diameter.
Although the fungus sporulates sparsely on artifi cial media, it produces copious conidia on necrotic host tissue, especially in leaf litter. Thus, Lim and Khoo (1985) suggested that removing such debris from orchards and burning it would assist control efforts. Several different fungicides are effective (Lim and Khoo, 1985).
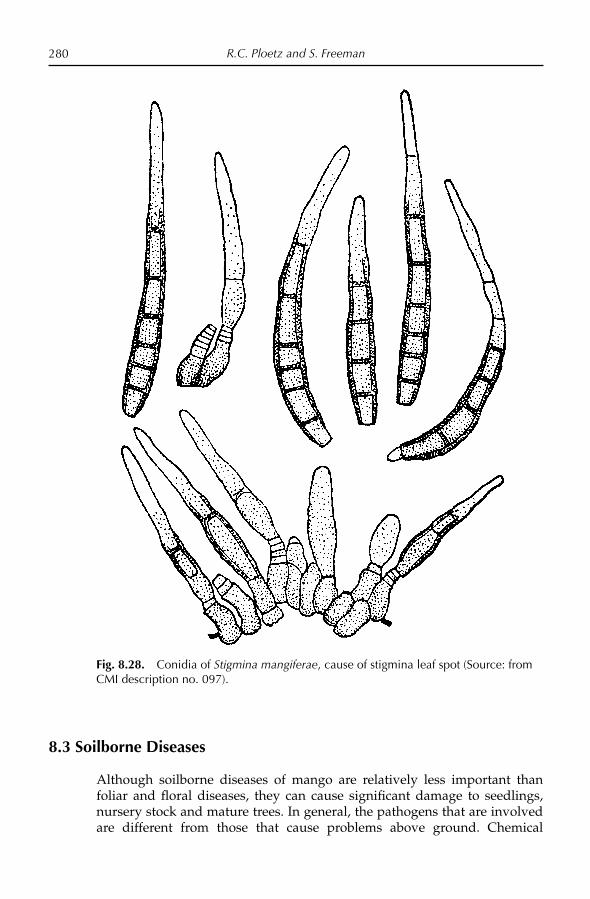
R.C. Ploetz and S. Freeman280
8.3 Soilborne Diseases
Although soilborne diseases of mango are relatively less important than foliar and fl oral diseases, they can cause signifi cant damage to seedlings, nursery stock and mature trees. In general, the pathogens that are involved are different from those that cause problems above ground. Chemical
Fig. 8.28. Conidia of Stigmina mangiferae, cause of stigmina leaf spot (Source: from CMI description no. 097).

Foliar, Floral and Soilborne Diseases 281
management is indicated rarely for these diseases; sanitation and other cultural measures are used most often.
Black root rot
Black root rot is reported to be an uncommon problem on young mango trees (Lim and Khoo, 1985). Canopies of affected plants wilt suddenly and subse-quently defoliate. Roots exhibit a water-soaked, blackened decay, and have an unpleasant, putrid odour. Although black root rot is associated with pro-longed fl ooding, its precise aetiology is not known. Several species of fungi have been recovered from affected plants, including Fusarium solani, F. oxyspo-rum and L. theobromae, but these were thought to be secondary colonizers of roots (Lim and Khoo, 1985). Although mango is generally considered to be fl ood intolerant, its fl ood tolerance is actually variable (Larson, 1991). Varia-tion in the responses of individual trees in orchards is evident after fl ooding, and when potted plants are fl ooded experimentally, they usually adapt by forming hypertrophied lenticels (intumescence) (Larson et al., 1993). Plants that do not adapt in this manner succumb fairly rapidly. Roots of the latter plants have symptoms of black root rot (R.C. Ploetz, Homestead, Florida, 1988, personal observations). Although fl ood tolerance is environmentally and biochemically complex (Larson et al., 1993), some of the fungi reported by Lim and Khoo (1985) may interact with fl ood-induced stress in this host to cause black root rot.
Nematode damage
Decline of mango trees due to nematodes has been reported in various regions (Khan et al., 1971, 2005; McSorley et al., 1980; Anita and Chaubey, 2003). Infestations occur in areas where warm temperatures and sandy, moist, well-drained soils predominate (Ploetz et al., 1994). Many nematode species have been recovered from mango roots, including: Helicotylenchusdihystera (Cobb) Sher, Quinisulcius acutus (Allen) Siddiqi, Rotylenchulus reniformis Linford and Oliveira, Criconemella sp., Pratylenchus brachyurus (Godfrey) Fil-ipjev and Schuurmans Stekhoven, Xiphinema sp., Meloidogyne sp., Praty-lenchus sp. and Hoplolaimus sp. However, only Hemicriconemoides mangiferae Siddiqi is pathogenic (Powers and McSorley, 1994). Although high popula-tions of R. reniformis are often found on mango trees, no correlation has been shown between their density and tree health.
Populations of H. mangiferae vary according to soil moisture and tem-perature (Khan et al., 1971). Soil moisture < 10% and > 30%, as well as tem-peratures < 15°C and > 35°C are detrimental to nematode survival and are likely to reduce populations. In addition, tree age appears to be a signifi cant factor, since H. mangiferae is found more frequently on old (> 10 years) than young trees (< 3 years). Management is diffi cult and may depend on pre-plant chemical applications plus cultural control measures (McSorley et al.,

R.C. Ploetz and S. Freeman282
1981). Phenamiphos and 1,2-dibromo-3-chloropropane reduce levels of H. mangiferae after planting; however, neither are registered for use in the USA. Anita and Chaubey (2004) reported that high organic matter content in the rhizosphere had a detrimental effect on nematode populations in the fi eld.
During orchard establishment, nematode-free nursery stock should be used. Since H. mangiferae is partially endoparasitic, it is moved easily to clean fi eld sites. The use of clean planting material in infested fi elds should also be avoided. If such land must be used, soil fumigation prior to planting should be conducted. Fruit yields may still be maintained if infected trees are well irrigated and fertilized.
Phytophthora diseases
Phytophthora palmivora (E.E. Butler) (Oomycota) causes diseases of mango in several areas. It caused wilt, crown rot, root rot and the death of nursery trees in Arizona USA, the Philippines and Thailand (Kueprakone et al., 1986; Matheron and Matejka, 1988; Tsao et al., 1994). Gumming and conspicuous bark lesions develop above ground on these plants, whereas root and crown rots are evident at or below the ground level. Crowded conditions and exces-sive irrigation and rainfall exacerbate these diseases. Sanitation, the use of less-crowded conditions and reduced irrigation would be benefi cial.
Damage has also been recorded on the trunks of fi eld-grown, mature trees in the Ivory Coast (Lourd and Keuli, 1975), and on fruit in Australia, Malaysia and West Africa (Turner, 1960 cited in Chee, 1969; Cooke, 2007). Mortality of trees is not observed, but substantial stem cracking and bleed-ing does occur. The symptoms that occur on fruit have not been recorded in Malaysia and West Africa, but on ‘Calypso’ fruit in Australia, a fi rm chocolate-brown decay is produced that has a sweet odour. Fruit isolates in Australia caused leaf blight and crown canker on mango seedlings (Fig. 8.29).
Phytophthora palmivora has coenocytic hyphae, up to 7 m in diameter, papil-late sporangia, 31–56.4 × 20.7–36.7 m, which germinate either directly with germ tubes or indirectly with motile zoospores (Fig. 8.30) (Waterhouse, 1970; Erwin and Ribiero, 1996). Zoospores are the primary infective propagule, and require free water for movement. Phytophthora palmivora is heterothallic. Antheridia are amphigynous and oogonia are spherical. Chlamydospores, c.35 m in diameter, are also formed.
Recently, a Phytophthora sp. was isolated in Spain from mango trees that were wilted, chlorotic and had sparse canopies and cracked bark (Zea-Bonilla et al., 2007). On V8 agar, sporangia were semi-papillate, obovoid and 51 (28–52) × 36 (22–37) m. Paragynous antheridia, spherical oogonia and oospores of 28 (19–32) m in diameter were produced homothalli-cally. Ribosomal DNA sequences (ITS1, 5.8S rDNA and ITS2) (GenBank Accession No. AM235209) were compared with those in GenBank; the closest match, 99% identity, was with Phytophthora citricola, corroborating the above morphological identifi cation (Fig. 8.31). An isolate deposited in
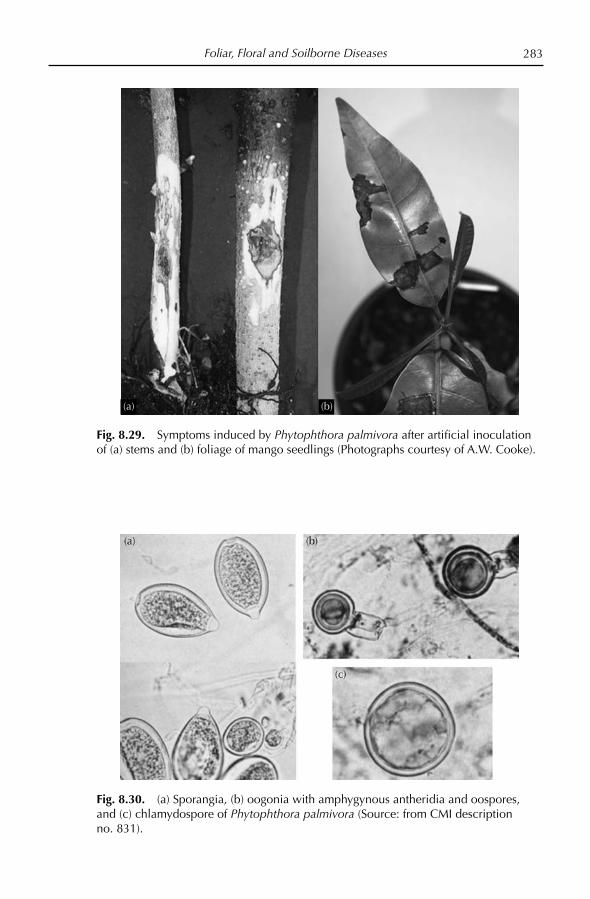
Foliar, Floral and Soilborne Diseases 283
Fig. 8.29. Symptoms induced by Phytophthora palmivora after artifi cial inoculation of (a) stems and (b) foliage of mango seedlings (Photographs courtesy of A.W. Cooke).
Fig. 8.30. (a) Sporangia, (b) oogonia with amphygynous antheridia and oospores, and (c) chlamydospore of Phytophthora palmivora (Source: from CMI description no. 831).
(a) (b)
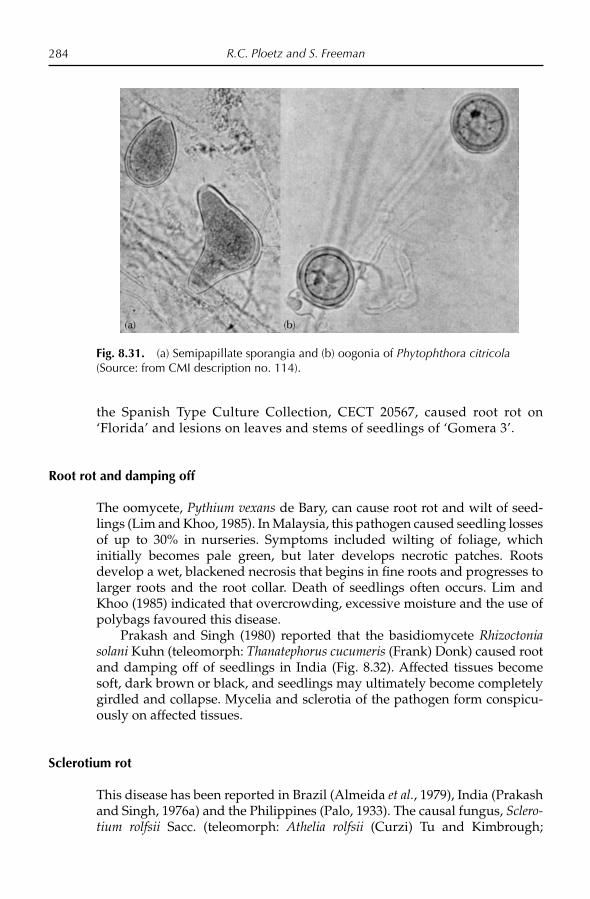
R.C. Ploetz and S. Freeman284
the Spanish Type Culture Collection, CECT 20567, caused root rot on ‘Florida’ and lesions on leaves and stems of seedlings of ‘Gomera 3’.
Root rot and damping off
The oomycete, Pythium vexans de Bary, can cause root rot and wilt of seed-lings (Lim and Khoo, 1985). In Malaysia, this pathogen caused seedling losses of up to 30% in nurseries. Symptoms included wilting of foliage, which initially becomes pale green, but later develops necrotic patches. Roots develop a wet, blackened necrosis that begins in fi ne roots and progresses to larger roots and the root collar. Death of seedlings often occurs. Lim and Khoo (1985) indicated that overcrowding, excessive moisture and the use of polybags favoured this disease.
Prakash and Singh (1980) reported that the basidiomycete Rhizoctoniasolani Kuhn (teleomorph: Thanatephorus cucumeris (Frank) Donk) caused root and damping off of seedlings in India (Fig. 8.32). Affected tissues become soft, dark brown or black, and seedlings may ultimately become completely girdled and collapse. Mycelia and sclerotia of the pathogen form conspicu-ously on affected tissues.
Sclerotium rot
This disease has been reported in Brazil (Almeida et al., 1979), India (Prakash and Singh, 1976a) and the Philippines (Palo, 1933). The causal fungus, Sclero-tium rolfsii Sacc. (teleomorph: Athelia rolfsii (Curzi) Tu and Kimbrough;
Fig. 8.31. (a) Semipapillate sporangia and (b) oogonia of Phytophthora citricola (Source: from CMI description no. 114).

Foliar, Floral and Soilborne Diseases 285
synonyms: Corticium rolfsii Curzi and Pellicularia rolftii E. West), produces globular, brown sclerotia, 1.0–2.6 mm in diameter. Sclerotia are resilient struc-tures that enable the pathogen to survive adverse environmental conditions. Symptoms begin with the formation of felty white tufts of mycelium of the pathogen around the base of seedlings. The fungus can girdle the entire stem to a height of 5 cm or more above the soil line. It eventually forms conspicu-ous sclerotia in high numbers. Ultimately, seedlings wilt and die. Seed may also rot prior to germination. The disease is controlled via sanitation and the disinfestation of seedbeds.
Verticillium wilt
Verticillium wilt of mango was fi rst reported in Florida USA (Marlatt et al., 1970). The disease was originally attributed to Verticillium albo-atrum Reinke and Berth., but this was before Verticillium dahliae Kleb. was recognized as a distinct species. Since the causal fungus described by Marlatt et al. (1970) formed microsclerotia, it is clear that V. dahliae was involved (Fig. 8.33).
Symptoms of the disease include a ‘fi ring’ and necrosis of leaves, usually in a portion of the canopy. Sectoral development of the disease often does not progress to other portions of the trees, which may recover. Killed leaves usu-ally remain attached to the tree, and the xylem of affected branches is dis-coloured brown (Fig. 8.34). Verticillium wilt is relatively uncommon, and is
(a)
(b)
(c)
20 μ
(d)
Fig. 8.32. (a) Sclerotial cells, (b) mycelium and (c) monilioid cells of Rhizoctonia solani, and (d) basidia and basidiospores of its teleomorph, Thanatephorus cucumeris(Source: from CMI description no. 406).
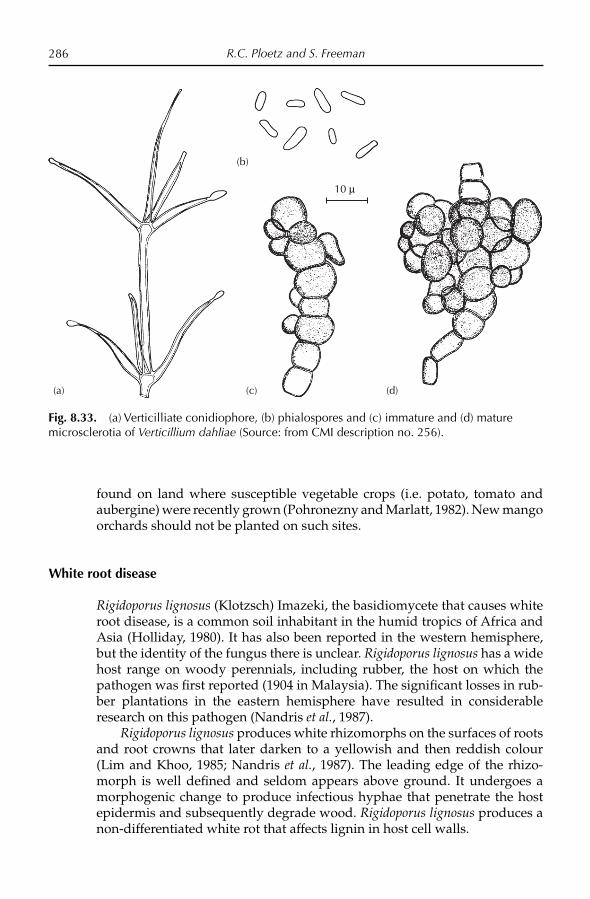
R.C. Ploetz and S. Freeman286
found on land where susceptible vegetable crops (i.e. potato, tomato and aubergine) were recently grown (Pohronezny and Marlatt, 1982). New mango orchards should not be planted on such sites.
White root disease
Rigidoporus lignosus (Klotzsch) Imazeki, the basidiomycete that causes white root disease, is a common soil inhabitant in the humid tropics of Africa and Asia (Holliday, 1980). It has also been reported in the western hemisphere, but the identity of the fungus there is unclear. Rigidoporus lignosus has a wide host range on woody perennials, including rubber, the host on which the pathogen was fi rst reported (1904 in Malaysia). The signifi cant losses in rub-ber plantations in the eastern hemisphere have resulted in considerable research on this pathogen (Nandris et al., 1987).
Rigidoporus lignosus produces white rhizomorphs on the surfaces of roots and root crowns that later darken to a yellowish and then reddish colour (Lim and Khoo, 1985; Nandris et al., 1987). The leading edge of the rhizo-morph is well defi ned and seldom appears above ground. It undergoes a morphogenic change to produce infectious hyphae that penetrate the host epidermis and subsequently degrade wood. Rigidoporus lignosus produces a non-differentiated white rot that affects lignin in host cell walls.
(a)
(b)
10 μ
(c) (d)
Fig. 8.33. (a) Verticilliate conidiophore, (b) phialospores and (c) immature and (d) mature microsclerotia of Verticillium dahliae (Source: from CMI description no. 256).

Foliar, Floral and Soilborne Diseases 287
The fungus is most damaging on mango if orchards are established in old rubber plantations or newly cleared jungle sites (Lim and Khoo, 1985). Previously colonized stumps and debris from rubber and other hosts are pri-mary sources of inoculum. Orange-yellow, bracket-like sporophores are pro-duced during the rainy season on the root collar, trunk or exposed roots (Fig. 8.35). Basidiospores are viable, but may play a secondary role in dis-seminating the disease. Rhizomorphs are more signifi cant epidemiologically, since they grow rapidly and can advance great distances in soil in the absence of woody substrates.
Fig. 8.34. Vascular discoloration caused by V. dahliae (Photograph courtesy of R.C. Ploetz).

R.C. Ploetz and S. Freeman288
White root disease is managed by eliminating or avoiding colonized woody debris when new orchards are established (Lim and Khoo, 1985). Unfortunately, this is diffi cult and often impractical. Alternative measures include: (i) treating planting holes with S to promote the growth of antago-nistic microorganisms; (ii) treating stumps after clearing operations with chemicals that discourage their colonization by basidiospores; and (iii) estab-lishment of leguminous cover crops that promote growth of the pathogen and the eventual exhaustion of its energy reserves.
8.4 Conclusions
As mango production continues to increase in different regions, so will the scope and types of disease problems. Although new fungicides and bacteri-cides will be developed in the future, it is probable that reliance on these tools will diminish. In recent years, the regulation of pesticides has increased while the number of new compounds that have been registered for use has decreased. Since it appears certain that this trend will continue, the problems posed by diseases must be solved increasingly with alternative disease control strategies.
10 μ (c)(d)
(a) (b)
20 μ
Fig. 8.35. (a) Upper and (b) lower surface of sporophore, (c) basidiospores, and (d) hyphae of Rigidoporus lignosus (Source: from CMI description no. 198).

Foliar, Floral and Soilborne Diseases 289
A willingness among producers to utilize new technologies will play a role in this process. New methods to detect important pathogens have been developed (Zheng and Ploetz, 2002), but more work is needed. Effective detection protocols, especially for those pathogens that can colonize host tis-sues without causing symptoms, could be used to interdict important, exotic pathogens and identify pathogen-free propagation materials. Detection pro-tocols could also indirectly assist disease control efforts through their use in epidemiological studies, research on disease resistance, or to clarify portions of disease cycles.
Acknowledgements
The authors thank Francisco Cazorla for comments on apical necrosis; Ber-nard Slippers and Mike Wingfi eld for comments on the decline section; Syl-via Fernandez, John Leslie, Christiano Lima, Kerry O’Donnell and Gerardo Rodriquez for information on malformation; Ali Obaid Al-Adawi and Mike Wingfi eld for comments on seca and sudden decline; and Robert Knight for translating Portuguese articles on seca. The following individuals are thanked for pictures: Francisco Cazorla, T.-K. Lim, Oliver Pruvost, Carolyn Cohen, Suzanne Bullock, Efrat Gamliel-Atinsky, Eric Palevsky and Tony Cooke. Kevin Hyde, editor of Fungal Diversity, is thanked for permission to use micrographs of Ceratocystis manginecans.
References
Abdel-Sattar, M.M.A. (1973) Histopathology of mango malformation. MSc. thesis, Al-Azhar University, Cairo.
Acuña, H.E. and Waite, B.H. (1977) La muerte regresiva del mango (Mangifera indica L.)en El Salvador. Proceedings of the American Society of Horticultural Sciences, Trop-ical Region 21, 15–16.
Adaskaveg, J.E. and Hartin, R.J. (1997) Characterization of Colletotrichum acutatumisolates causing anthracnose of almond and peach in California. Phytopathology87, 979–987.
Afanador-Kafuri, L., Minz, D., Maymon, M. and Freeman, S. (2003) Characterization of Colletotrichum isolates from tamarillo, passifl ora and mango in Colombia and identifi cation of a unique species from the genus. Phytopathology 93, 579–587.
Ah-You, N., Gagnevin, L., Chiroleu, F., Jouen, E., Rodrigues Neto, J. and Pruvost, O. (2007a) Pathological variations within Xanthomonas campestris pv. mangiferae-indicae support its separation into three distinct pathovars that can be distinguished by amplifi ed fragment length polymorphism. Phytopathology 97, 1568–1577.
Ah-You, N., Gagnevin, L., Pruvost, O., Myint, N.T. and Johnson, G.I. (2007b) First report in Myanmar of Xanthomonas axonopodis pv. mangiferaeindicae causing mango bacterial canker on Mangifera indica. Plant Disease 91, 1686.
Al Adawi, A.O., Deadman, M.L., Al Rawahi, A.K., Khan, A.J. and Al Maqbali, Y.M. (2003) Diplodia theobromae associated with sudden decline of mango in the Sultanate of Oman. Plant Pathology 52, 419.

R.C. Ploetz and S. Freeman290
Al Adawi, A.O., Deadman, M.L., Al Rawahi, A.K., Al Maqbali, Y.M., Al Jahwari, A.A., Al Saadi, B.A., Al Amri, I.S. and Wingfi eld, M.J. (2006) Aetiology and causal agents of mango sudden decline disease in the Sultanate of Oman. European Journal of Plant Pathology 116, 247–254.
Al Subhi, A.M., Al Adawi, A.O., Van Wyk, M., Deadman, M.L. and Wingfi eld, M.J. (2006) Ceratocystis omanensis, a new species from diseased mango trees in Oman. Mycological Research 110, 237–245.
Alahakoon, P.W., Brown, A.E. and Sreenivasaprasad, S. (1994a) Cross-infection potential of genetic groups of Colletotrichum gloeosporioides on tropical fruits. Physiological and Molecular Plant Pathology 44, 93–103.
Alahakoon, P.W., Brown, A.E. and Sreenivasaprasad, S. (1994b) Genetic characteriza-tion of Colletotrichum gloeosporioides isolates obtained from mango. InternationalJournal of Pest Management 40, 225–229.
Alexopoulos, C.J., Mims, C.W. and Blackwell, M. (1996) Introductory Mycology, 4th edn. John Wiley and Sons, New York.
Alfi eri, SA., Jr, Langdon, K.R., Kimbrough, J.W., El-Gholl, N.E. and Wehlburg, C. (1994) Diseases and Disorders of Plants in Florida. Bulletin No. 14. Florida Department of Agricultural and Consumer Services, Tallahassee, Florida.
Almeida, R.T. de, Landim, C.M.U. and Teixeira, L.M.S. (1979) Seedling blight (Sclero-tium rolfsii Sacc.) of cashew and mango in the state of Ceara, Brazil. Fitossanidade3, 40–41.
Alvarez-García, L.A. and López-Gracía, J. (1971) Gummosis, dieback, and fruit rot disease of mango (Mangifera indica L.) caused by Physalospora rhodina (B. and C.) Cke. in Puerto Rico. Journal of Agriculture University of Puerto Rico 55, 435–450.
Angulo, S.M. and Villapudua, J.R. (1982) Buba of mango (Mangifera indica L.) in the state of Sinaloa, Mexico. (Abstract). Phytopathology 72, 171.
Anita and Chaubey, A.K. (2003) Infl uence of soil temperature and moisture on popula-tion dynamics of ectoparasitic nematodes infesting Mangifera indica. Annals of Plant Protection Sciences 11, 181–183.
Anita and Chaubey, A.K. (2004) Impact of organic matter on the population dynamics of Hemicriconemoides mangiferae infesting Mangifera indica. Biological Memoirs 30,108–111.
Arrebola, E., Cazorla, F.M., Romero, D., Pérez-García, A. and de Vicente, A. (2007) A nonribosomal peptide synthetase gene (mgoA) of Pseudomonas syringae pv. syrin-gae is involved in mangotoxin biosynthesis and is required for full virulence. Mo-lecular Plant-Microbe Interactions 20, 500–509.
Atkinson, T.H. and Peck, S.B. (1994) Annotated checklist of the bark and ambrosia bee-tles (Coleoptera: Platypodidae and Scolytidae) of tropical Southern Florida. TheFlorida Entomologist 77, 313–329.
Bastawros, M.K. (1996) Mango malformation in Egypt. Acta Horticulturae 455, 566–574.Batista, A.C. (1947) Mal do recife (grave doenca da manggueira). Unpublished thesis,
Pernambuco College of Agriculture, Pernambuco, Brazil (Abstract in Review of Ap-plied Mycology (1948) 27, 77–78).
Batzer, J.C., Gleason, M.L., Weldon, B., Dixon, P.M. and Nutter, F.W., Jr (2002) Evalua-tion of postharvest removal of sooty blotch and fl yspeck on apples using sodium hypochlorite, hydrogen peroxide with peroxyacetic acid, and soap. Plant Disease86, 1325–1332.
Batzer, J.C., Gleason, M.L., Harrington, T.C. and Tiffany, L.H. (2005) Expansion of the sooty blotch and fl yspeck complex on apples based on analysis of ribosomal DNA gene sequences and morphology. Mycologia 97, 1268–1286.

Foliar, Floral and Soilborne Diseases 291
Berthet, J.A. (1914) Molestia cia mangueira. Bolm Agriculturae (São Paolo) 15, 818–819.
Bhatnagar, S.S. and Beniwal, S.P.S. (1977) Involvement of Fusarium oxysporum in causa-tion of mango malformation. Plant Disease Reporter 61, 894–898.
Bitancourt, A.A. and Jenkins, A.E. (1943) Scab of mango caused by Elsinoe. (Abstract). Phytopathology 33, 1.
Boesewinkel, H.J. (1980) The identity of mango mildew, Oidium mangiferae. Phyto-pathologische Zeitschrift 99, 126–130.
Bose, S.K., Sindhan, G.S. and Pandey, B.H. (1973) Studies on dieback disease of mango in the Tarai region of Kuaon. Progressive Horticulture 70, 557–584.
Britz, H., Steenkamp, E.T., Coutinho, T.A., Wingfi eld, B.D., Marasas, W.F.O. and Wing-fi eld, M.J. (2002) Two new species of Fusarium section Liseola associated with man-go malformation. Mycologia 94, 722–730.
Burgess, T.I., Barber, P.A., Mohali, S., Pegg, G., de Beer, W. and Wingfi eld, M.J. (2006) Three new Lasiodiplodia spp. from the tropics, recognized based on DNA sequence comparisons and morphology. Mycologia 98, 423–435.
Butani, D.K. (1993) Mango: Pest Problems. Periodical Expert Book. Agency D-42, Vivek Vihar, Delhi, India.
Butler, E.J. and Bisby, G.R. (1931) The Fungi of India. Scientifi c Monograph No.1. The Imperial Council of Agricultural Research in India, Calcutta.
Carvalho, C.R.L., Rossetto, C.J., Mantovani, D.M.B., Morgano, M.A., de Castro, J.V. and Bortoletto, N. (2004) Avaliaçã de cultivares de mangueira selectionadas pelo Insiti-tuto Agrontmico de Campinas comparadas a outras de importância comercial. Re-vista Brasil Fruticultura 26, 264–271.
Cazorla, F.M., Torés, J.A., Olalla, L., Pérez-García, A., Farré, J.M. and de Vicente, A. (1998) Bacterial apical necrosis of mango in southern Spain: a disease caused by Pseudomonas syringae pv. syringae. Phytopathology 88, 614–620.
Cazorla, F.M., Arrebola, E., Sesma, A., Pérez-García, A., Codina, J.C., Murillo, J. and de Vicente, A. (2002) Copper resistance in Pseudomonas syringae strains isolated from mango is encoded mainly by plasmids. Phytopathology 92, 909–916.
Cazorla, F.M., Arrebola, E., Olea, F., Velasco, L., Hermoso, J.M., Pérez-García, A., Torés, J.A., Farré, J.M. and deVicente, A. (2006) Field evaluation of treatments for the con-trol of the bacterial apical necrosis of mango (Mangifera indica) caused by Pseudomo-nas syringae pv. syringae. European Journal of Plant Pathology 116, 279–288.
Chakrabarti, D.K. and Ghosal, S. (1989) The disease cycle of mango malformation in-duced by Fusarium moniliforme var. subglutinans and the curative effects of mangiferin metal chelates. Journal of Phytopathology 125, 238–246.
Chee, K.H. (1969) Hosts of Phytophthora palmivora. Review of Applied Mycology 48,337–344.
Colosimo, E.A., Chalita, L.V.A.S. and Demetrio, C.G.B. (2000) Tests of proportional hazards and proportional odds models for grouped survival data. Biometrics 56, 1233–1240.
Cook, A.A. (1975) Diseases of Tropical and Subtropical Fruits and Nuts. Hafner Press, New York.
Cook, A.A., Milbrath, G.M. and Hamilton, R.A. (1971) Woody gall and scaly bark of Mangifera indica in Hawaii. (Abstract). Phytopathology 61, 1320.
Cooke, A.W. (2007) Phytophthora postharvest fruit rot in mango. Advisory poster pro-duced by the Queensland Government Department of Primary Industries and Fish-eries, Indooropilly, Queensland, Australia.
Cronje, C., Wehnwe, F.C. and Kotzé, J.M. (1990) Alternaria alternata as a lesion patho-gen of mango infl orescences in South Africa. Phytophylactica 22, 117–118.

R.C. Ploetz and S. Freeman292
Crookes, C.A. and Rijkenberg, F.H.J. (1985) A literature review of the distribution, symp-tomatology, cause and control of mango blossom malformation. South African Mango Grower’s Association Yearbook 5, 15–24.
Crous, P.W. and Palm, M.E. (1999) Reassessment of the anamorph genera of Botryoplo-dia, Dothiorella and Fusicoccum. Sydowia 52, 167–175.
Crous, P.W., Slippers, B., Wingfi eld, M.J., Rheeder, J., Marasas, W.F.O., Philips, A.J.L., Alves, A., Burgess, T., Barber, P. and Groenewald, J.Z. (2006) Phylogenetic lineages in the Botryosphaeriaceae. Studies in Mycology 55, 235–253.
Dag, A., Eisenstein, D. and Gazit, S. (2001) Effects of three fungicides used to control powdery mildew in mango on pollen germination and pollen-tube growth. Pro-ceedings of the InterAmerican Society for Tropical Horticulture 43, 123–126.
Dang, J.K. and Daulta, B.S. (1982) Mango malformation – a review. Pesticides 16, 5–11.Darvas, J.M. (1987) Control of mango blossom malformation with trunk injection. South
African Mango Grower’s Association Yearbook 7, 21–24.Darvas, J.M. (1993) Dothiorella dominicana, an important mango pathogen in South
Africa. Acta Horticulturae 341, 321–328.Das Gupta, S.N. and Zacchariah, H.T. (1945) Studies on the diseases of Mangifera in-
dica. Part V. On the die-back of mango trees. Journal of the Indian Botanical Society 24, 101–108. (Abstract in Review of Applied Mycology (1946) 25, 267–268).
Denman, S., Crous, P.W., Taylor, J.E., Kang, J.-C., Pascoe, I. and Wingfi eld, M.J. (2000) An overview of the taxonomic history of Botryosphaeria, and a re-evaluation of its anamorphs based on morphology and ITS and rDNA phylogeny. Studies in Mycol-ogy 45, 129–140.
Diekman, F., Manicom, B.Q. and Coetzee, K. (1982) An attempt to control blossom malformation of mangoes with chemical sprays. Subtropica 3, 15–16.
Dodd, J.C., Estrada, A.B., Matcham, J., Jefferies, P. and Jeger, M.J. (1991) The effect of climatic factors on Colletotrichum gloeosporioides, causal agent of mango anthra-cnose, in the Philippines. Plant Pathology 40, 568–575.
Domsch, K.H., Gams, W. and Anderson, T.-H. (1980) Compendium of Soil Fungi. Aca-demic Press, London.
Droby, S., Prusky, D., Jacoby, B. and Goldman, A. (1986) Presence of antifungal com-pounds in the peel of mango fruits and their relation to latent infections by Alter-naria alternata. Physiological and Molecular Plant Pathology 29, 173–183.
Droby, S., Prusky, D., Jacoby, B. and Goldman, A. (1987) Induction of antifungal resor-cinols in fl esh of unripe mango fruits and its relation to latent infection by Alter-naria alternata. Physiological and Molecular Plant Pathology 30, 285–292.
Du, M., Schardl, C.L., Nuckels, E.M. and Vaillancourt, L.J. (2005) Using mating-type gene sequences for improved phylogenetic resolution of Collectotrichum species complexes. Mycologia 97, 641–658.
Ellis, M.B. (1971) Dematiaceous Hypomycetes. Commonwealth Mycological Institute, Kew, Surrey, UK, 608 pp.
Engelbrecht, C.J. and Harrington, T.C. (2005) Intersterility, morphology and taxonomy of Ceratocystis fi mbriata on sweet potato, cacao and sycamore. Mycologia 97, 57–69.
Erwin, D.C. and Ribiero, O.K. (1996) Phytophthora Diseases Worldwide. American Phy-topathological Society (APS) Press, St Paul, Minnesota.
Eshel, D., Miyara, I., Ailinng, T., Dinoor, A. and Prusky, D. (2002) pH regulates endoglu-canase expression and virulence of Alternaria alternata in persimmon fruits. Mo-lecular Plant-Microbe Interactions 15, 774–779.
Farr, D.F., Bills, G.F., Chamuris, G.P. and Rossman, A.Y. (1989) Fungi on Plant and Plant Products in the United States. American Phytopathological Society (APS) Press, St Paul, Minnesota.

Foliar, Floral and Soilborne Diseases 293
Fitzell, R.D. (1979) Colletotrichum acutatum as a cause of anthracnose of mango in New South Wales. Plant Disease Reporter 63, 1067–1070.
Fitzell, R.D. and Peak, C.M. (1984) The epidemiology of anthracnose disease of mango: inoculum sources, spore production and dispersal. Annals of Applied Biology 104,53–59.
Fitzell, R.D., Peak, C.M. and Darnell, R.E. (1984) A model for estimating infection levels of anthracnose disease of mango. Annals of Applied Biology 104, 451–458.
Flechtmann, C.H.W., Kimati, H., Medcalf, J.C. and Ferré, J. (1973) Preliminary observa-tions on malformation in mango (Mangifera indica L.) infl orescences and the fun-gus, insects and mite found on them. Review of Plant Pathology 52, 162–163.
Freeman, S. and Rodriguez, R.J. (1995) Differentiation of Colletotrichum species respon-sible for anthracnose of strawberry by arbitrarily primed PCR. Mycological Research99, 501–504.
Freeman, S., Pham, M. and Rodriguez, R.J. (1993) Molecular genotyping of Colletotri-chum species based on arbitrarily primed PCR, A+T-rich DNA, and nuclear DNA analyses. Experimental Mycology 17, 309–322.
Freeman, S., Katan, T. and Shabi, E. (1998) Characterization of Colletotrichum speciesresponsible for anthracnose diseases of various fruits. Plant Disease 82, 596–605.
Freeman, S., Maimon, M. and Pinkas, Y. (1999) Use of GUS transformants of Fusarium subglutinans for determining etiology of mango malformation disease. Phytopathol-ogy 89, 456–461.
Freeman, S., Minz, D., Maymon, M. and Zveibil, A. (2001) Genetic diversity within Col-letotrichum acutatum sensu Simmonds. Phytopathology 91, 586–592.
Fukuda, T., Uehara, K., Azegami, K., Tabei, H. and Nishiyama, K. (1990) Bacterial can-ker of mango in Japan caused by Xanthomonas campestris pv. mangiferaeindicae. Annals of the Phytopathological Society of Japan 56, 474–480.
Gagnevin, L. and Pruvost, O. (1995) Assessment of genomic variability in Xanthomonascampestris pv. mangiferaeindicae. (Abstract). Phytopathology 85, 1163.
Gagnevin, L. and Pruvost, O. (2001) Epidemiology and control of mango bacterial black spot. Plant Disease 85, 928–935.
Gamliel-Atinsky, E., Sztejnberg, A., Maymon, M., Belausov, E., Palevsky, E. and Free-man, S. (2007) Interaction of the mango bud mite (Aceria mangiferae) with Fusari-um mangiferae, causal agent of mango malformation disease. Phytoparasitica 35, 198–199.
Gantotti, B.V. and Davis, M.J. (1993) Pectic zymogram analysis for characterizing genetic diversity of the mango anthracnose pathogen. Acta Horticulturae 341,353–359.
Gupta, J.H. (1989) Perpetuation and epidemiology of powdery mildew of mango. Acta Horticulturae 231, 528–533.
Halsted, B.D. and Fairchild, D.G. (1891) Sweet-potato black rot. Journal of Mycology 7, 1–11.
Hamid, M. and Jalaluddin, M. (2006) Recurring incidence of sooty mould of mango in Karachi and its control. International Journal of Biology and Biotechnology 3,561–565.
Hayden, H.L., Pegg, K.G., Aitken, E.A.B. and Irwin, J.A.G. (1994) Genetic relationships as assessed by molecular markers and cross-infection among strains of Colletotri-chum gloeosporioides. Australian Journal of Botany 42, 9–18.
Hingorani, M.K., Sharma, O.P. and Sohi, H.S. (1960) Studies on blight disease of mango caused by Macrophoma mangiferae. Indian Phytopathology 13, 137–143.
Hirano, S.S. and Upper, C.D. (1983) Ecology and epidemiology of foliar bacterial plant pathogens. Annual Review of Phytopathology 21, 243–269.

R.C. Ploetz and S. Freeman294
Hodson, A., Mill, P.R. and Brown, A.E. (1993) Ribosomal and mitochondrial DNA poly-morphisms in Colletotrichum gloeosporioides isolated from tropical fruits. Myco-logical Research 97, 329–335.
Holliday, P. (1980) Fungus Diseases of Tropical Crops. Cambridge University Press, Cambridge.
Hughes, S.J. (1980) New Zealand Fungi 29. Rhinocladium Sacc. et March. New Zealand Journal of Botany 18, 163–172.
Iqbal, Z., Mehboob-ur-Rahman, Dasti, A.A., Saleem, A. and Zafar, Y. (2006) RAPD anal-ysis of Fusarium isolates causing ‘Mango Malformation’ disease in Pakistan. World Journal of Microbiology and Biotechnology 22, 1161–1167.
Jeffries, P., Dodd, J.C., Jeger, M.J. and Plumbley, R.A. (1990) The biology and control of Colletotrichum species on tropical fruit crops. Plant Pathology 39, 343–366.
Johnson, E.M., Sutton, T.B. and Hodges, C.S. (1997) Etiology of apple sooty blotch dis-ease in North Carolina. Phytopathology 87, 88–95.
Johnson, G.I. (1992) Biology and control of stem-end rot pathogens of mango. PhD thesis, University of Queensland, Brisbane, Australia.
Johnson, G.I. (1994a) Powdery mildew. In: Ploetz, R.C., Zentmyer, G.A., Nishijima, W., Rohrbach, K. and Ohr, H.D. (eds) Compendium of Tropical Fruit Diseases. American Phytopathological Society (APS) Press, St Paul, Minnesota, pp. 38–39.
Johnson, G.I. (1994b) Stem-end rot. In: Ploetz, R.C., Zentmyer, G.A., Nishijima, W., Rohrbach, K. and Ohr, R.D. (eds) Compendium of Tropical Fruit Diseases. AmericanPhytopathological Society (APS) Press, St Paul, Minnesota, pp. 39–41.
Johnson, G.I., Cooke, A.W., Mead, A.J. and Wells, L.A. (1991) Stem end rot of mango in Australia: causes and control. Acta Horticulturae 219, 288–295.
Johnson, G.I., Mead, A.J., Cooke, A.W. and Dean, J.R. (1992) Mango stem end rot patho-gens – fruit infection by endophytic colonisation of the infl orescence and pedicel. Annals of Applied Biology 120, 225–234.
Johnson, J.A., Harrington, T.C. and Engelbrecht, C.J.B. (2005) Phylogeny and taxonomy of the North American clade of the Ceratocystis fi mbriata complex. Mycologia 97,1067–1092.
Joubert, J.J. and Rijkenberg, F.H.J. (1971) Parasitic green algae. Annual Review of Phyto-pathology 9, 45–64.
Junqueira, N.T.V., Pinto, A.C. de Q., da Cunha, M.M. and Ramos, V.H.V. (2002) Controle das doenças da mangueira. In: Zambolin, L., do Vale, F.X.R., Monteiro, A.J.A. and Costa, H. (eds) Controle de Doenças de Plantas Fruteiras. Vol. 1. Universidade Fed-eral de Viçosa, Viçosa, Brazil, pp. 323–403.
Kadman, A. and Gazit, S. (1984) The problem of iron defi ciency in mango trees and experiments to cure it in Israel. Journal of Plant Nutrition 7, 282–290.
Kauser, A.G. (1959) Malformation of infl orescence in mango. Punjab Fruit Journal 22,19–21.
Kennelly, M.M., Cazorla, F.M., de Vicente, A., Ramos, C. and Sundin, G.W. (2007)Pseudomonas syringae diseases of fruit trees: progress toward understanding and control. Plant Disease 91, 4–17.
Khan, A.M., Adhami, A. and Saxena, S.K. (1971) Population changes of some stylet-bearing nematodes associated with mango (Mangifera indica L.). Indian Journal of Nematology 1, 99–105.
Khan, A., Sayed, M. and Shaukat, S.S. (2005) Nematodes associated with mango in Sindh. International Journal of Biology and Biotechnology 2, 917–919.
Kile, G.A. (1993) Plant diseases caused by species of Ceratocystis sensu stricto and Chalara. In: Wingfi eld, M.J., Seifert, K.A. and Webber, J.F. (eds) Ceratocystis and

Foliar, Floral and Soilborne Diseases 295
Ophiostoma: Taxonomy, Ecology and Pathogenicity. American Phytopathological Society (APS) Press, St Paul, Minnesota, pp. 173–183.
Kishun, R. (1994) Evaluation of phylloplane micro-organisms from mango against Xan-thomonas campestris pv. mangiferaeindicae. Indian Phytopathology 47, 313.
Kishun, R. (1995) Detection and management of Xanthomonas campestris pv. mangiferaeindicae. In: Varma, J.P., Verma, A. and Kumar, D. (eds) Detection of Plant Pathogens and their Management. Angkar Publishers, New Delhi, India, pp. 173–182.
Kishun, R. and Sohi, H.S. (1983) Bacterial canker in mangoes. Indian Farmer’s Digest 14, 21–23.
Kueprakone, U., Saengkong, S., Pienpuck, K. and Choobumroong, W. (1986) Phytophtho-ra palmivora (Butl.) Butl., the causal organism of black rot of mango seedlings. ThaiAgricultural Research Journal 4, 67–73. (in Thai with an English abstract)
Kumar, J. and Beniwal, S.P.S. (1991) Mango malformation. In: Kumar, J., Chaube, H.S., Singh, U.S. and Mukhopadhyay, A.N. (eds) Plant Diseases of International Impor-tance. Vol. III: Diseases of Fruit Crops. Prentice Hall, Englewood Cliffs, New Jersey, pp. 357–393.
Kvas, M., Steenkamp, E.T., Al Adawi, A.O., Deadman, M.L., Al Jahwari, A.A., Marasas, W.F.O., Wingfi eld, B.D., Ploetz, R.C. and Wingfi eld, M.J. (2007) Fusarium mangifer-ae associated with mango malformation in the Sultanate of Oman. European Jour-nal of Plant Pathology 121, 195–199.
Larson, K.D. (1991) Physiological, anatomical and growth responses of mango (Mangiferaindica L.) trees to fl ooding. PhD thesis, University of Florida, Gainesville, Florida.
Larson, K.D., Schaffer, B. and Davies, F.S. (1993) Floodwater oxygen content, ethylene production and lenticel hypertrophy in fl ooded mango (Mangifera indica L.) trees. Journal of Experimental Botany 44, 665–671.
Leslie, J.F. (1995) Gibberella fujikuroi: available populations and variable traits. Cana-dian Journal of Botany 73, S282–S291.
Leslie, J.F. and Summerell, B.A. (2006) The Fusarium Lab Manual. Blackwell, Ames, Iowa. Lim, T.K. (1994) Pink disease. In: Ploetz, R.C., Zentmyer, G.A., Nishijima, W.T., Rohr-
bach, K.G. and Ohr, H.D. (eds) Compendium of Tropical Fruit Diseases. AmericanPhytopathological Society (APS) Press, St Paul, Minnesota, pp. 37–38.
Lim, T.K. and Khoo, K.C. (1985) Diseases and Disorders of Mango in Malaysia. Tropical Press, Kuala Lumpur, Malaysia.
Lima, C.S., Costa, S.S., Campos, M.A. and Pfenning, L.H. (2006a) A new Fusarium spe-cies associated with mango malformation in Brazil. In: XXXIX Annual Meeting of the Brazilian Phytopathological Society, 2006, Salvador. Fitopatologia Brasileira(S)31, 191.
Lima, C.S., Pfenning, L.H., Costa, S.S., Campos, M.A. and Leslie, J.F. (2006b) A new biological species of the Gibberella fujikuroi complex associated with mango mal-formation in Brazil. In: XXXIX Annual Meeting of the Brazilian Phytopathological Society, 2006, Salvador. Fitopatologia Brasileira (S) 31, 191.
Litz, R.E. (ed.) (1997) The Mango: Botany, Production and Uses. CAB International, Wallingford, UK.
Lonsdale, J.H. and Kotzé, J.M. (1993) Chemical control of mango blossom diseases and the effect on fruit set and yield. Plant Disease 77, 558–562.
Lourd, M. and Keuli, S.D. (1975) Note sur un chancre a Phytophthora du manguier en Cote d’lvoire. Fruits 30, 541–544.
Majumder, P.K. and Sharma, D.K. (1990) Mango. In: Bose, T.K. and Mitra, S.K. (eds) Fruits: Tropical and Subtropical. Naya Prokash, Calcutta, pp. 40–62.

R.C. Ploetz and S. Freeman296
Malaguti, G. and de Reyes, C. (1964) A gall disease of cacao and mango in Venezuela caused by Calonectria rigidiuscula. (Abstract). Phytopathology 54, 499.
Malik, M.T., Khan, S.M., Dasti, A.A. and Kazmi, M.R. (2005) First record of Ceratocystis pimbriata (sic) causal organism of mango sudden death in Pakistan. Pakistan Jour-nal of Phytopathology 17, 187–191.
Manicom, B.Q. (1986) Factors affecting bacterial black spot of mangoes caused by Xanthomonas campestris pv. mangiferaeindicae. Annals of Applied Biology 109,129–135.
Manicom, B.Q. (1989) Blossom malformation of mango. South African Mango Grower’s Association Yearbook 10, 11–12.
Manicom, B.Q. and Pruvost, O.P. (1994) Bacterial black spot. In: Ploetz, R.C., Zentmyer, G.A., Nishijima, W.T., Rohrbach, K.G. and Ohr, H.D. (eds) Compendium of Tropical Fruit Diseases. American Phytopathological Society (APS) Press, St Paul, Minnesota, pp. 41–42.
Manicom, B.Q. and Wallis, F.M. (1984) Further characterization of Xanthomonas camp-estris pv. mangiferaeindicae. International Journal of Systematic Bacteriology 34,77–79.
Marasas, W.F.O., Ploetz, R.C., Wingfi eld, M.J., Wingfi eld, B.D. and Steenkamp, E.T. (2006) Mango malformation disease and the associated Fusarium species. Phytopa-thology 96, 667–672.
Marlatt, R.B., Knight, R.J., Jr and Goldweber, S. (1970) Verticillium wilt of mango (Mangifera indica) in Florida. Plant Disease Reporter 54, 569–571.
Matheron, M.E. and Matejka, J.C. (1988) Phytophthora crown and root rot on nursery-grown mango trees delivered to Arizona. (Abstract). Phytopathology 78, 1572.
Maymon, M., Zveibil, A., Pivonia, S., Minz, D. and Freeman, S. (2006) Identifi cation and characterization of benomyl-resistant and -sensitive fi eld isolates of Colletotrichum gloeosporioides from statice (Limonium spp.). Phytopathology 96, 542–548.
McSorley, R., Parrado, J.L. and Goldweber, S. (1980) Observations on a mango decline in South Florida. Proceedings of the Florida State Horticultural Society 93, 132–133.
McSorley, R., Parrado, J.L. and Goldweber, S. (1981) Plant-parasitic nematodes associ-ated with mango and relationship to tree condition. Nematropica 11, 1–9.
Michailides, T.J., Morgan, D.P. and Felts, D. (1999) Other hosts of Botryosphaeria dothidea and their relation to Botryosphaeria pistachio blight. California Pistachio Production Reports. California Pistachio Commission, Fresno, California, USA.
Misra, A.K. and Prakash, O. (1992) Control of bacterial canker of mango by chemicals. Pesticides 18, 32–33.
Mukherjee, S.K. (1997) Introduction: botany and importance. In: Litz, R.E. (ed.) TheMango: Botany, Production and Uses. CAB International, Wallingford, UK, pp. 1–19.
Müller, E.M., Chelkowski, J. and Geiger, H.H. (1999) Species-specifi c PCR assays for the fungal pathogens Fusarium moniliforme and Fusarium subglutinans and their application to diagnose maize ear rot disease. Journal of Phytopathology 147, 497–508.
Muller, H.R.A. (1940) Overzicht van de belangrijkste mangga-ziekten in Nederlandsch Indie. (English abstract in Review of Applied Mycology (1940) 19, 355.)
Nandris, D., Nicole, M. and Geiger, J.P. (1987) Root rot diseases of rubber trees. PlantDisease 71, 298–306.
Narasimhan, M.J. (1954) Malformation of panicles in mango incited by spp. of Erio-phyes. Current Science 23, 297–298.
Narasimhan, M.J. (1959) Control of mango malformation disease. Current Science 28, 254–255.

Foliar, Floral and Soilborne Diseases 297
Nariani, T.K. and Seth, M.L. (1962) Role of eriophyid mites in causing malformation disease of mango. Indian Phytopathology 15, 231–234.
Neergaard, P. (1945) Danish Species of Alternaria and Stemphylium. Taxonomy, Parasiti-cism, Economical Signifi cance. Einar Munksgaard, Copenhagen, Denmark.
Nicholson, R.I.D. and van Staden, J. (1988) Cytokinins and mango fl ower malformation. I. Tentative identifi cation of the complement in healthy and malformed infl ores-cences. Journal of Plant Physiology 132, 720–724.
Nofal, M.A. and Haggag, W.M. (2006) Integrated management of powdery mildew of mango in Egypt. Crop Protection 25, 480–486.
O’Donnell, K., Cigelnik, E. and Nirenberg, H.I. (1998) Molecular systematics and phy-logeography of the Gibberella fujikuroi species complex. Mycologia 90, 465–493.
O’Donnell, K., Nirenberg, H.I., Aoki, T. and Cigelnik, E. (2000) A multigene phylogeny of the Gibberella fujikuroi species complex: detection of additional phylogeneti-cally distinct species. Mycoscience 41, 61–78.
Okigbo, R.N. (2001) Occurrence, pathogenicity and survival of Macrophoma mangifer-ae in leaves, branches and stems of mango (Mangifera indica L.). Plant Protection Science 37, 138–144.
Okigbo, R.N. and Osuinde, M.I. (2003) Fungal leaf spot diseases of mango (Mangiferaindica L.) in southeastern Nigeria and biological control with Bacillus subtilis. PlantProtection Science 39, 70–77.
Oosthuyse, S.A. (1998) Cost reduction of powdery mildew control in mango with mono potassium phosphate. South African Mango Growers’ Association Yearbook 18, 40–42.
Palmateer, A.J., Ploetz, R.C., van Santer, E. and Correll, J.C. (2007) First occurrence of anthra-cnose caused by Colletotrichum gloeosporioides on pitahaya. Plant Disease 91, 631.
Palo, M.A. (1933) Sclerotium seed rot and seedling rot of mango. Philippine Journal of Science 52, 237–261.
Palti, J., Pinkas, Y. and Chorin, M. (1974) Powdery mildew of mango. Plant Disease Re-porter 58, 45–49.
Patel, M.K., Kulkarni, Y.S. and Moniz, L. (1948) Pseudomonas mangiferae-indicae,pathogenic on mango. Indian Phytopathology 1, 147–152.
Peres, N.A.R., Souza, N.L., Zitko, S.E. and Timmer, L.W. (2002) Activity of benomyl for control of postbloom fruit drop of citrus caused by Colletotrichum acutatum. PlantDisease 86, 620–624.
Peterson, R.A., Johnson, G.I., Schipke, L.G. and Cooke, A.W. (1991) Chemical control of stem end rot. Acta Horticulturae 291, 304–307.
Piza, C. de T. (ed.) (1966) Anais do simposio sobre a seca da mangueira. (Abstract in Review of Applied Mycology (1967) 46, 378.)
Ploetz, R.C. (1994) Distribution and prevalence of Fusarium subglutinans in mango trees affected by malformation. Canadian Journal of Botany 72, 7–9.
Ploetz, R.C. (2001) Malformation: a unique and important disease of mango, Mangiferaindica L. In: Summerell, B.A., Leslie, J.F., Backhouse, D. and Bryden, W.L. (eds) Fusarium: Paul E. Nelson Memorial Symposium. American Phytopathological Soci-ety (APS) Press, St Paul, Minnesota, pp. 233–247.
Ploetz, R.C. (2003) Diseases of mango. In: Ploetz, R.C. (ed.) Diseases of Tropical Fruit Crops. CAB International, Wallingford, UK, pp. 327–363.
Ploetz, R.C. (2007) Cacao diseases: important threats to chocolate production world-wide. Phytopathology 97, 1634–1639.
Ploetz, R.C. and Prakash, O. (1997) Foliar, fl oral and soilborne diseases of mango. In: Litz, R.E. (ed.) The Mango: Botany, Production and Uses. CAB International, Wall-ingford, UK, pp. 281–325.

R.C. Ploetz and S. Freeman298
Ploetz, R.C., Zentmyer, G.A., Nishijima, W., Rohrbach, K. and Ohr, H.D. (eds) (1994) Compendium of Tropical Fruit Diseases. American Phytopathological Society (APS) Press, St Paul, Minnesota.
Ploetz, R.C., Benscher, D., Vasquez, A., Colls, A., Nagel, J. and Schaffer, B. (1996a) A re-evaluation of mango decline in Florida. Plant Disease 80, 664–668.
Ploetz, R., Vazquez, A. and Benscher, D. (1996b) First report of Fusarium decemcellulareas a pathogen of mango in the United States. Plant Disease 80, 1207.
Ploetz, R.C., Benscher, D., Dorey, A.J. and Vazquez, A. (2000) The epidemiology, control and cause of sooty blotch of carambola, Averrhoa carambola L., in South Florida. Fruits 55, 241–252.
Ploetz, R., Zheng, Q.I., Vazquez, A. and Abdel Sattar, M.A. (2002) Current status and impact of mango malformation in Egypt. International Journal of Pest Management48, 279–285.
Pohronezney, K. and Marlatt, R.B. (1982) Some Common Diseases of Mango in Florida.Plant Pathology Fact Sheet PP-23. University of Florida, Gainesville, Florida.
Pordesimo, A.N. (1982) Development of programmed spray application for the control of anthracnose. In: PCARRD Research Highlights 1982. Philippine Council for Ag-riculture, Forestry and Natural Resources Research and Development (PCARRD), Los Baños, the Philippines, pp. 38–39.
Powers, L.E. and McSorley, R. (1994) Nematodes. In: Ploetz, R.C., Zentmyer, G.A., Nishijima, W., Rohrbach, K. and Ohr, H.D. (eds) Compendium of Tropical Fruit Diseases. American Phytopathological Society (APS) Press, St Paul, Minnesota, pp. 42–43.
Prakash, O. (1988) Sooty mould disease of mango and its control. International Journal of Tropical Plant Disease 9, 277–280.
Prakash, O. (1990) Report of the plant pathologist. Annual Report of CIHNP (CMRS), Lucknow for 1989–1990.
Prakash, O. (1991) Sooty mould disease of mango and its control. International Journal of Tropical Plant Diseases 9, 277–280.
Prakash, O. and Raoof, M.A. (1989) Die back disease of mango (Mangifera indica),its distribution, incidence, cause and management. Fitopatologia Brasiliensis 14,207–215.
Prakash, O. and Singh, U.N. (1976a) Basal rot of mango seedling caused by Sclerotiumrolfsii. Indian Journal of Mycology and Plant Pathology 6, 75.
Prakash, O. and Singh, U.N. (1976b) New disease of mango. In: Proceedings of AICFRP Workshop, 24–28 May, Hyderabad, India.
Prakash, O. and Singh, U.N. (1977) Phoma blight, a new disease of mango. Plant Dis-ease Reporter 61, 419–421.
Prakash, O. and Singh, U.N. (1980) Root rot and damping off of mango seedlings caused by Rhizoctonia solani. Indian Journal of Mycology and Plant Pathology 10, 69.
Prakash, O. and Srivastava, K.C. (1987) Mango Diseases and their Management – a World Review. Tommorow’s Printer, New Delhi, India.
Prakash, O., Misra, A.K. and Raoof, M.A. (1994) Studies on mango bacterial canker disease. Biological Memoirs 20, 95–107.
Prasad, A., Singh, H. and Shukla, T.N. (1965) Present status of mango malformation disease. Indian Journal of Horticulture 22, 254–265.
Prusky, D. (1994) Alternaria rot (black spot). In: Ploetz, R.C., Zentmyer, G.A., Nishijima, W., Rohrbach, K. and Ohr, H.D. (eds) Compendium of Tropical Fruit Diseases. American Phytopathological Society (APS) Press, St Paul, Minnesota, pp. 34–35.
Prusky, D., Fuch, Y. and Yanko, U. (1983) Assessment of latent infections as a basis for control of post harvest disease of mango. Plant Disease 67, 816–818.

Foliar, Floral and Soilborne Diseases 299
Prusky, D., Shalom, Y., Kobiler, I., Akerman, M. and Fuchs, Y. (2002) The level of quies-cent infection of Alternaria alternata in mango fruits at harvest determines the post-harvest treatment applied for the control of rots during storage. Postharvest Biology and Technology 25, 339–347.
Prusky, D., Kobiler, I., Akerman, M. and Miyara, I. (2006) Effect of acidic solutions and acidic prochloraz on the control of postharvest decay caused by Alternaria alternatain mango and persimmon fruit. Postharvest Biology and Technology 42, 134–141.
Pruvost, O. and Luisetti, J. (1991a) Attempts to develop a biological control of bacterial black spot of mangoes. Acta Horticulturae 291, 324–337.
Pruvost, O. and Luisetti, J. (1991b) Effect of time of inoculation with Xanthomonascampestris pv. mangiferaeindicae on mango fruits susceptibility. Epiphytic survival of X. c. pv. mangiferaeindicae on mango fruits in relation to disease development. Journal of Phytopathology 133, 139–151.
Pruvost, O., Couteau, A. and Luisetti, J. (1989) Effi cacité de différentes formulations chemiques pour Jutter contre la maladie des taches noires de la mangue (Xan-thomonas campestris pv. mangiferaeindicae). Fruits 44, 343–350.
Pruvost, O., Couteau, A. and Luisetti, J. (1990) Development of bacterial black spot of mangoes and epiphytic populations of the pathogen (Xanthomonas campestris pv.mangiferaeindicae) under natural conditions in Reunion Island. Fruits 45, 125–140.
Pruvost, O., Couteau, A. and Luisetti, J. (1992) Pepper tree (Schinus terebenthifolius Radii), a new host plant for Xanthomonas campestris pv. mangiferaeindicae. Journal of Phytopathology 135, 289–298.
Pruvost, O., Couteau, A., Verniere, C. and Luisetti, J. (1993) Epiphytic survival of Xan-thomonas campestris pv. mangiferaeindicae on mango buds. Acta Horticulturae 341, 337–344.
Pruvost, O., Roumagnac, P., Gaube, C., Chiroleu, F. and Gagnevin, L. (2005) New me-dia for the semiselective isolation and enumeration of Xanthomonas campestris pv. mangiferaeindicae, the causal agent of mango bacterial black spot. Journal of Ap-plied Microbiology 99, 803–815.
Punithalingam, E. (1976) Botryodiplodia theobromae. Commonwealth Mycological In-stitute (CMI) Descriptions of Pathogenic Fungi and Bacteria No. 519. CMI, Kew, Surrey, UK.
Ramos, L.J., Lara, S.P., McMillan, R.T. and Narayanan, K.R. (1991) Tip dieback of mango (Mangifera indica) caused by Botyrosphaeria ribis. Plant Disease 75, 315–318.
Rao, B.S. (1975) Maladies of Hevea in Malaysia. Rubber Research Institute of Malaysia, Kuala Lumpur, Malaysia.
Ray, S.K. (2003) Effi cacy of different fungicides for management of powdery mildew of mango in West Bengal. Journal of Mycopathological Research 41, 153–155.
Reckhaus, P. and Adamou, I. (1987) Hendersonula dieback of mango in Niger. (Disease note). Plant Disease 71, 1045.
Reddy, G.S., Govinda, R.P. and Paparao, A. (1961) Black banded disease of mango. Andhra Agricultural Journal 8, 120–123.
Reuveni, M., Harpaz, M. and Reuveni, R. (1998) Integrated control of powdery mildew on fi eld-grown mango trees by foliar sprays of mono-potassium phosphate fertilizer, sterol inhibitor fungicides and the strobilurin Kresoxym-methyl. European Journal of Plant Pathology 104, 853–860.
Ribiero, I.J.A. (1980) Seca de mangueira. Agentes causais e estudio da molesta. Anais do I Simposio Brasiliero Sobre a Cultura de Mangueira, 24–28 Nov. 1980. SociedadBrasileira de Fruticultura, Jaboticobal, Brazil, pp. 123–130.
Ribiero, I.J.A. (1997) Doenças da mangueira (Mangifera indica L.). In: Kimati, H., Amo-rim, A., Bergamin-Filho, A., Camargo, L.E.A. and Rezende, J.A.M. (eds) Manual de

R.C. Ploetz and S. Freeman300
Fitopatologia. Vol. 2: Doenças das Plantas Cultivadas. Agronômica Ceres, São Paulo, Brazil, pp. 511–524.
Ribiero, I.J.A., Rosetto, C.J., Donadio, L.C., Sabino, J.C. Martins, A.L.M. and Gallo, P.B. (1995) Mango wilt. XIV. Selection of mango (Mangifera indica L.) rootstocks resis-tant to the mango wilt fungus Ceratocystis fi mbriata Ell. and Halst. Acta Horticultu-rae 370, 159–161.
Ridgeway, R. (ed.) (1989) Mango Pests and Disorders. Department of Primary Industries, Brisbane, Queensland, Australia.
Rivera-Vargas, L., Lugo-Noel, Y., McGovern, R.J., Sijo, T. and Davis, M.J. (2006) Occur-rence and distribution of Colletotrichum spp. on mango (Mangifera indica L.) in Puerto Rico and Florida, USA. Plant Pathology Journal 5, 191–198.
Rodríguez-Alvarado, G., Fernández-Pavía, S.P. and Ploetz, R.C. (2006) Fusarium sp. characterization causing mango malformation in Michoacan Mexico. Phytopathol-ogy 96, S99.
Rodríguez-Alvarado, G., Fernández-Pavía, S.P., Ploetz, R.C. and Valenzuela-Vázquez, M. (2008) A Fusarium sp., different from Fusarium oxysporum and F. mangiferae, is associated with mango malformation in Michoacán, Mexico. New Disease Reports. Plant Pathology 57, 781.
Rossetto, C.J., Ribeiro, I.J.A., Igue, T. and Gallo, P.B. (1996) Mango wilt: XV. Varietal re-sistance against two isolates of Ceratocystis fi mbriata. Bragantia 55, 117–121.
Rossman, A.Y., Samuels, G.J., Rogerson, C.T. and Lowen, R. (1998) Genera of Bionectri-aceae, Hypocreaceae and Nectriaceae (Hypocreales, Ascomycetes). Studies in My-cology 42, 1–248.
Ruehle, G.D. and Ledin, R.B. (1955) Mango Growing in Florida. Bulletin 574. Agricul-tural Experiment Station, University of Florida, Gainesville, Florida.
Saaiman, W.C. (1996) Biology and control of Nattrassia mangiferae. Acta Horticulturae455, 558–564.
Saeed, A. and Schlosser, E. (1972) Effect of some cultural practices on the incidence of mango malformation. Zeitschrift für Pfl anzenkrankheiten und Pfl anzenschutz 79, 349–351.
Sanders, G.M., Verschoor, J.A., van Wyngaard, S., Korsten, L. and Kotzé, J.M. (1994) Production of monoclonal antibodies against Xanthomonas campestris pv. mangifer-aeindicae and their use to investigate differences in virulence. Journal of Applied Bacteriology 77, 509–518.
Schaffer, B. (1994) Decline. In: Ploetz, R.C., Zentmyer, G.A., Nishijima, W., Rohrbach, K. and Ohr, H.D. (eds) Compendium of Tropical Fruit Diseases. American Phyto-pathological Society (APS) Press, St Paul, Minnesota, p. 43.
Schaffer, B., Larson, K.D., Snyder, G.H. and Sanchez, C.A. (1988) Identifi cation of mineral defi ciencies associated with mango decline by DRIS. HortScience 23, 617–619.
Schoeman, M.H., Manicom, B.Q. and Wingfi eld, M.J. (1995) Epidemiology of powdery mildew on mango blossoms. Plant Disease 79, 524–528.
Silveira, S.F., Harrington, T.C., Mussi-Dias, V., Engelbrecht, C.J.B., Alfenas, A.C. and Sil-va, C.R. (2006) Annona squamosa, a new host of Ceratocystis fi mbriata. Fitopatolo-gia Brazileira 31, 394–397.
Singh, L.B. (1968) The Mango: Botany, Cultivation, and Utilization. Leonard Hill, London.
Singh, R.N., Majumder, P.K., Sharma, D.K., Sinha, G.C. and Bose, P.C. (1974) Effect of de-blossoming on the productivity of mango. Scientia Horticulturae 2, 399–403.
Singh, Z. and Dhillon, B.S. (1989) Hormonal changes associated with vegetative malfor-mation of mango (Mangifera indica L.). Journal of Phytopathology 125, 193–197.

Foliar, Floral and Soilborne Diseases 301
Singh, Z., Singh, L., Arora, C.L. and Dhillon, B.S. (1994) Effect of cobalt, cadmium, and nickel as inhibitors of ethylene biosynthesis on fl oral malformation, yield, and fruit quality of mango. Journal of Plant Nutrition 17, 1659–1670.
Slippers, B. and Wingfi eld, M.J. (2007) Botryosphaeriaceae as endophytes and latent pathogens of woody plants: diversity, ecology and impact. Fungal Biology Reviews 21, 90–106.
Slippers, B., Johnson, G.I., Crous, P.W., Coutinho, T.A., Wingfi eld, B.D. and Wingfi eld, M.J. (2005) Phylogenetic and morphological re-evaluation of the Botryosphaeria species causing diseases of Mangifera indica. Mycologia 97, 99–110.
Smith, P.F. and Scudder, G.K. (1951) Some studies of mineral defi ciency symptoms in mango. Proceedings of the Florida State Horticultural Society 64, 243–248.
Snowdon, A.L. (1990) Color Atlas of Post-Harvest Diseases and Disorders of Fruits and Vegetables. Vol. 1. General Introduction and Fruits. CRC Press, Boca Raton, Florida.
Sreenivasaprasad, S., Mills, P.R., Meehan, B.M. and Brown, A.E. (1996) Phylogeny and systematics of 18 Colletotrichum species based on ribosomal DNA spacer sequenc-es. Genome 39, 499–512.
Steenkamp, E.T., Wingfi eld, B.D., Coutinho, T.A., Wingfi eld, M.J. and Marasas, W.F.O. (1999) Differentiation of Fusarium subglutinans f. sp. pini by histone gene sequence data. Applied and Environmental Microbiology 65, 3401–3406.
Steenkamp, E.T., Britz, H., Coutinho, T.A., Wingfi eld, B.D., Marasas, W.F.O., Wingfi eld, B.D., Marasas, W.F.O. and Wingfi eld, M.J. (2000) Molecular characterization of Fusarium subglutinans associated with mango malformation. Molecular PlantPathology 1, 187–193.
Steyn, P.L., Viljoen, N.M. and Kotzé, J.M. (1974) The causal organism of bacterial black-spot of mangoes. Phytopathology 64, 1400–1404.
Summanwar, A.S, Raychaudhuri, S.P. and Phatak, S.C. (1966) Association of the fungus Fusarium moniliforme Sheld. with the malformation in mango (Mangifera indica L.).Indian Phytopathology 19, 227–228.
Sutton, B.C. (1980) The Coelomycetes. Commonwealth Mycological Institute, Kew, Sur-rey, UK, 696 pp.
Taba, S., Takaesu, K., Ooshiro, A., Moromizato, Z. and Takushi, T. (2004) Anthracnose of mango caused by Colletotrichum acutatum. Japanese Journal of Tropical Agricul-ture 48, 57–61.
Tavares, S.C.C. de H., da Cruz, S.C., de Siqueira, J.G., Lima, M.L.C., Silva, P.C.G.C., de Neves, R.A.F. and Menezes, C.A. (2004) Alternative control to powdery mildew (Erysiphe poligoni d. c. Sin) on mango in submedio Sao Francisco river in Brazilian semi-arid region. Acta Horticulturae 645, 471–474.
Tiwari, R.K.S., Ashok, S., Rajput, M.L. and Bisen, R.K. (2006) Relative susceptibility of mango varieties to powdery mildew caused by Oidium mangiferae. Advances in Plant Sciences 19, 181–183.
Tsao, P.H., Luzaran, P.B., de los Santos, A.B., Portales, L.A., Gochango, A.M. and Gru-ber, L.C. (1994) Phytophthora crown and foot rot of mango detected in Philippine nurseries. (Disease note). Plant Disease 78, 100.
Uppal, B.N., Patel, M.K. and Kamat, M.N. (1941) Powdery mildew of mango. Journal of the University of Bombay 9, 12–16.
van Staden, J. and Nicholson, R.J.D. (1989) Cytokinins and mango fl ower malformation. The cytokinin complement produced by Fusarium moniliforme and the ability of the fungus to incorporate (8-14C) adenine into cytokinins. Physiological and Mo-lecular Plant Pathology 35, 423–431.

R.C. Ploetz and S. Freeman302
van Staden, J., Bayley, A.D. and Macrae, S. (1989) Cytokinins and mango fl ower malfor-mation. III. The metabolism of (3H) iso-pentenyladenine and (8-14C) zeatin by Fusarium moniliforme. Physiological and Molecular Plant Pathology 35, 433–438.
van Wyk, M., Al Adawi, A.O., Wingfi eld, B.D., Al Subhi, A.M., Deadman, M.L. and Wingfi eld, M.J. (2005) DNA based characterization of Ceratocystis fi mbriata iso-lates associated with mango decline in Oman. Australasian Plant Pathology 34, 587–590.
van Wyk, M., Al Adawi, A.O., Khan, I.A., Deadman, M.L., Wingfi eld, B.D., Ploetz, R.C. and Wingfi eld, M.J. (2007) Ceratocystis manginecans sp. nov., causal agent of the destructive mango die-back disease in Oman and Pakistan. Fungal Diversity 27, 213–230.
Varma, A., Lele, V.C., Raychauduri, S.P., Ram, A. and Sang, A. (1974) Mango malforma-tion: a fungal disease. Phytopathologische Zeitschrift 70, 254–257.
Verma, K.S. and Singh, J. (1996a) Perpetuation and management of Macrophomamangiferae causing blight of mango. Indian Journal of Mycology and Plant Pathol-ogy 26, 75–78.
Verma, K.S. and Singh, J. (1996b) Pathology and epidemiology of Macrophoma blight of mango. Indian Journal of Mycology and Plant Pathology 26, 98–100.
Verma, O.P. and Singh, R.D. (1970) Epidemiology of mango dieback caused by Botyo-diplodia theobromae Pat. Indian Journal of Agricultural Science 40, 813–818.
Viegas, A.P. (1960) Mango blight. Bragantia 19, 163–182. (Abstract in Review of Applied Mycology (1963) 42, 696.)
Viljoen, N.M. and Kotzé, J.M. (1972) Bacterial black spot of mango. Citrus Grower and Sub-Tropical Fruit Journal June, pp. 5–8.
Wagle, P.V. (1928) Studies in the shedding of mango fl owers and fruits. Part I, Memoirs of the Department of Agriculture. India Botanical Sciences 8, 219–249.
Waterhouse, G.M. (1970) The Genus Phytophthora de Bary, 2nd edn. Mycological Papers No. 122. Commonwealth Mycological Institute, Kew, Surrey, UK.
Webster, R.K. and Butler, E.E. (1967) A morphological and biological concept of the species Ceratocystis fi mbriata. Canadian Journal of Botany 45, 1457–1468.
Weng, F.J. and Chuang, T.Y. (1995) Grouping of mango anthracnose fungus in Taiwan. Plant Protection Bulletin 3, 295–309.
Williamson, S.M. and Sutton, T.B. (2000) Sooty blotch and fl yspeck of apple: etiology, biology, and control. Plant Disease 84, 714–724.
Wood, S.L. (1982) The bark and ambrosia beetles of North and Central America (Co-leoptera: Scolytidae), a taxonomic monograph. Great Basin Naturalist, Memoirs 6, 1–1356.
Youssef, S.A., Maymon, M., Zveibil, A., Klein-Gueta, D., Sztejnberg, A., Shalaby, A.A. and Freeman, S. (2007) Epidemiological aspects of mango malformation disease caused by Fusarium mangiferae and source of infection in seedlings cultivated in orchards in Egypt. Plant Pathology 56, 257–263.
Zea-Bonilla, T., Martín-Sánchez, P.M., Hermoso, J.M., Carmona, M.P., Segundo, E. and Pérez-Jimenéz, R.M. (2007) First report of Phytophthora citricola on Mangifera in-dica in Spain. New Disease Reports. Plant Pathology 56, 356.
Zheng, Q. and Ploetz, R. (2002) Genetic diversity in, and development of a PCR assay for identifying, the mango malformation pathogen. Plant Pathology 51, 208–216.

© CAB International 2009. The Mango, 2nd Edition: Botany, Production and Uses(ed. R.E. Litz) 303
9 Physiological Disorders
V. Galán SaúcoInstituto Canario de Investigaciones Agrarias, Tenerife (Canary Islands), Spain
9.1 Introduction 3039.2 Internal Fruit Breakdown 304
Symptoms 305Histology 305Altered physiology 306Causes 306Cultivar susceptibility 307Control 308
9.3 Lumpy Tissue 3099.4 Ricey Tissue 3109.5 Fruit Cracking 3109.6 Black Tip Disorder 3109.7 Lenticel Spot 3119.8 Other Fruit Disorders 3119.9 Stem Disorders 312
9.1 Introduction
Physiological disorders can be defi ned as ailments that have not been caused by infecting organisms. Unlike mango malformation, for example, which has been considered as a physiological disorder in the past (IBPGR, 1989) but has a phytopathological origin, true physiological disorders cannot be transmit-ted from plant to plant, mechanically or by insect bites. For the most part, they are a result of some form of physical damage or of an altered physiology of the tree or fruit. In the particular case of fruit disorders – the most preva-lent and important physiological disorders of mango – according to Subra-manyan et al. (1971), they are essentially the result of imbalances in metabolism induced by some factor or factors in the pre- or postharvest environment that leads to cell collapse and, typically, the appearance of waterlogged or

V. Galán Saúco304
brown areas on some part of the fruit. The complexity of events leading to the occurrence of physiological disorders makes it more diffi cult to pinpoint the causal factors than is the case of symptoms caused by pathogens or pests.
Physiological disorders occur relatively frequently in many fruit and vegetable species. Some well-known examples include bitter pit, water core and internal breakdown in apple (Atkinson et al., 1980) and in watermelon (Singh et al., 1975; Cirulli and Ciccarese, 1981), blossom end rot in tomato and pepper (Winsor and Adams, 1987), mesocarp discoloration and pulp spot in avocado (Van Lelyveld et al., 1984; Bower and Cutting, 1987) and yellow pulp in banana (Melin and Aubert, 1973).
Although physiological disorders of annual crops and temperate fruits have been studied extensively, tropical fruits have only recently been stud-ied in order to understand and control physiological disorders. This is largely due to the rapid expansion of these crops during the past few decades.
Preharvest factors that predispose mango fruit to physiological disorders include growing location, orchard condition, tree nutrition and condition at the time of harvest. Postharvest storage conditions, such as temperature, oxygen and carbon dioxide levels, packaging and surface coating treatments, are also potential contributing factors to the occurrence of these disorders (Subramanyan et al., 1971).
Evidence of specifi c metabolic changes that occur in mango fruits that express this type of disorder is scarce. Probably due to this lack of knowl-edge, disorders are usually named for the associated environmental factor, for example chilling injury, or by the altered appearance that the physiologi-cal disorder confers to the fruit. Clear examples of the latter are internal fruit breakdown, which is by far the most prevalent physiological disorder in mango, lenticel spot, fruit cracking and black tip disorder.
9.2 Internal Fruit Breakdown
Most opinion (Winston, 1984; Schaffer, 1994) is that several physiological dis-orders currently listed as separate disorders are in fact different degrees or aspects of the same problem, internal fruit breakdown (IFB). These include tip pulp, soft nose (i.e. waterlogging of the fl esh near the distal end), jelly seed (i.e. overripe fl esh around the seed, surrounded by fi rm fl esh), stem end breakdown (i.e. an open cavity in the pulp at the stem end), spongy tissue (i.e. areas of the fl esh that appear spongy and have a greyish black discolor-ation), soft centre and soft fl esh. This aggregation is still open to some debate. Raymond et al. (1998) asserted that temporal and spatial differences in symp-tom development within the fruit have been found between soft nose, jelly seed and stem end cavity, validating their return to separate disorder status, but they recognized that no major microscopic differences could be found. None the less, irrespective of the disparity of symptoms, all of these disor-ders have a common denominator: calcium imbalance. With this defi ning criterion, IFB would also include the physiopathy described in Malaysia as yeasty or insidious fruit rot, which shows a marked interdependence with

Physiological Disorders 305
calcium (Lim and Khoo, 1985) and also what Velasco Cárdenas (1974) calls ‘ablandamiento del pico’ (literally peak softening).
Cheema and Dani (1934) fi rst reported an IFB disorder, but perhaps the most comprehensive work published on physiological disorders of the mango fruit was that of Malo and Campbell (1978), who described the different degrees of tissue decomposition in fruit of ‘Tommy Atkins’. The comprehen-sive work of Wainwright and Burbage (1989) reviews the symptomatol-ogy, chemical changes and causes of IFB, illustrating its occurrence in many cultivars in virtually all the producing areas of the world. Losses from IFB vary geographically and also among cultivars, and can affect 100% of the harvested fruit (Subramanyan et al., 1971; Malo and Campbell, 1978; Bro-drick and Thord-Gray, 1982; Galán Saúco et al., 1984; Santos Filho et al., 2002; Cracknell Torres et al., 2003a).
Symptoms
Symptoms usually begin to appear early during fruit development and advance rapidly, eventually making the fruit inedible (see Plates 48–52). The process commences while the fruit is still hanging on the panicle, with an interruption occurring in the vascular tissues of the peduncle and endocarp, and usually followed by the formation of a cavity close to the funiculum (stem end breakdown). In advanced stages, the affected tissues around the cavity become grey or blackish. As the disorder progresses, the proximal end of the fruit becomes mushy to the touch (soft nose). In some cases, a yellow-ing of the skin between the apex and the stigmatic end of the fruit can also occur. The fi bre surrounding the endocarp may fully disintegrate and in severe cases an accumulation of a transparent, gelatinous substance devel-ops around the seed (jelly seed). The affected mesocarp matures more rap-idly than the healthy fl esh and acquires a characteristic deeper yellow colour; the affected tissue may be so extensive that greyish, watery tissues appear over the whole mesocarp (spongy tissue) (Malo and Campbell, 1978; Winston, 1984; Joshi and Roy, 1985; Wainwright and Burbage, 1989; Katrodia and Chuva, 1993). The absence of latex and the lack of fi rmness in the proxi-mal end of the fruit at harvest can be clear signs of the disorder (Chaplin, 1986), although the lack of latex alone has also been observed in black tip disorder (see below).
Histology
Histological differences have been found between fruits of different cultivars affected by IFB. For example, in the very susceptible ‘Tommy Atkins’ the xylem elements in the mesocarp and the mesocarp cells are all highly deteriorated. In the case of a less sensitive cultivar (i.e. ‘Lippens’) the affected mesocarp cells have intact cell walls, even though the tissue is obviously affected, showing a translucent mesocarp (Cracknell Torres and Galán Saúco, 2003).

V. Galán Saúco306
Altered physiology
A reduction in fi rmness, total soluble solids, total pectin and pectinase activ-ity was observed by these researchers in the affected mesocarp tissue, con-fi rming the results of Van Lelyveld and Smith (1979) and Roe and Bruemmer (1981), both of whom also observed greater activity of both pectinase and malic enzyme in affected pulp of ‘Alphonso’. On the other hand, the higher level of nitrogen, the lower levels of calcium, and the higher nitrogen to cal-cium ratio observed by Cracknell Torres and Galán Saúco (2003) in the affected tissue of ‘Tommy Atkins’ and ‘Lippens’ is in agreement with the studies of Young (1960) and Young and Miner (1961) but in confl ict with results of Krishnamurthy (1981), who found no differences among calcium levels between affected and unaffected mesocarp tissues. Other differences in chemical composition have been detected in affected and unaffected tis-sues (Subramanyan et al., 1971; Patkar et al., 1983; Nuevo et al., 1984; Gupta et al., 1985) and further research should be devoted to ascertain the true nature of this disorder.
Causes
Despite many attempts to isolate a pathogen from affected tissues, no patho-gen has yet been linked with IFB. In Malaysia, spongy tissue has been found in ‘Harumanis’ fruit which has been affected by the fruit-piercing moth Othreis spp. and in ‘Fan Siamese’ fruit, which has been associated with dam-age caused by phytotoxic sprays of some fungicides (Lim and Khoo, 1985). Postharvest vapour heat treatment at 46°C for 10 min has also been reported to induce IFB in ‘Carabao’ fruit (Esquerra et al., 1990).
Several environmental factors have been linked to IFB. For example, in India the disorder appears to be more prevalent in coastal areas (Subraman-yan et al., 1971). Gunjate et al. (1982) observed that it is associated with fruit that remains exposed to the sun following harvest. IFB has also been attributed to heat convection from the soil at air temperatures around 55°C (Katrodia and Rane, 1989). In several experiments with the cultivar ‘Alphonso’ in India, a positive correlation was observed between IFB inci-dence and relative density. Mangoes that were harvested with a relative den-sity between 1.00 and 1.20 showed no IFB, while fruits greater than 1.2 had a 30% incidence of IFB (Krishnamurthy, 1980). Fruit weight also seems to be an important factor at least in the case of some cultivars like ‘Alphonso’ in India (Subramanyan et al., 1971) and in Spain with ‘Sensation’ (Hermoso et al., 1996), with a positive correlation in both cases between fruit weight and incidence of IFB.
Cultural practices such as excessive irrigation have also been considered to be possible causes of IFB. Katrodia (1988) observed a higher incidence of IFB following periods of rainfall just prior to harvesting of ‘Alphonso’ in India; however, trials done elsewhere with ‘Tommy Atkins’ (Malo and Camp-bell, 1978) or ‘Sensation’ (Farré and Hermoso, 1993) showed no correlation

Physiological Disorders 307
between excessive irrigation and IFB. Furthermore, in Malaysia the incidence of IFB (insidious or yeasty fruit rot) has been reported to be higher during the drier months of the year (Lim and Khoo, 1985).
Most observations and fi eld studies associate IFB with calcium defi ciency, which is also the main cause of similar disorders in other fruits (Bangerth, 1979; Winsor and Adams, 1987). In general, low levels of leaf calcium at har-vest coincide with high IFB incidence. In Florida, IFB appears to occur more in acid and sandy soils with low calcium content than in basic, calcareous soils (Young, 1957; Schaffer, 1994), but in ‘Harumanis’ yeasty or insidious fruit rot occurs irrespective of various types of soils, ranging from acidic to alkaline. It has been suggested that the degree of IFB is aggravated in the presence of excess nitrogen fertilization (Young, 1960; Young and Miner, 1961; Young et al., 1962, 1965).
The direct relationship between IFB and calcium and nitrogen content has been confi rmed by analysis of affected and non-affected fruits of orchard-grown ‘Harumanis’ (Ahmad Tarmizi et al., 1993), and in soilless cul-tivation trials involving ‘Tommy Atkins’ (Cracknell Torres et al., 2003b). In the latter study, positive correlations were observed between mesocarpic nitrogen levels and IFB and between the nitrogen:calcium ratio and IFB, but there was a negative relationship between calcium and IFB.
Cultivar susceptibility
Although IFB has been observed in at least 65 cultivars in 23 countries (Galán Saúco, 1999), it is clear that varietal differences play a role in the intensity and expression of the symptoms, as these have been described in different degrees of intensity according to countries and cultivars.
According to Young (1957), cultivars of Indian origin and their offspring show a higher incidence of IFB, for example ‘Alphonso’ whose susceptibility has been well documented (Subramanyan et al., 1971; Gunjate et al., 1979, 1982; Krishnamurthy, 1980, 1981; Patkar et al., 1983; Gupta et al., 1985; Lim and Khoo, 1985; Katrodia and Rane, 1989; Lad et al., 1992). There is, however, confl icting evidence; some Indian cultivars, for example ‘Rajapuri’ (Katrodia, 1988), ‘Banganapalli’, ‘Kalapardy’, ‘Janradhan Pasand’ (Iyer and Subraman-yan, 1992), ‘Pairi’, ‘Kesar’ and ‘ Neelum’ (Lad et al., 1992), rarely demonstrate these disorders. All Florida cultivars are affected due to their Indian parent-age (Schaffer, 1994); ‘Haden’, ‘Kent’, ‘Keitt’, ‘Sensation’, ‘Tommy Atkins’ and ‘Van Dyke’ are repeatedly mentioned as particularly susceptible (De Larous-silhe, 1980; Galán Saúco and Fernández Galván, 1987; Campbell, 1992; Oost-huyse, 1993; Knight, 1997; Galán Saúco, 1999; Cracknell Torres et al., 2003a). IFB problems are also cited for the Malaysian cultivar ‘Harumanis’ or ‘Aru-manis’ (Lim and Khoo, 1985; Tengku Ab.Malik, 1992a, b; Ahmad Tarmizi et al., 1993).
IFB incidence in some cultivars is so low as to be virtually non-existent, particularly the polyembryonic and fi brous-fl eshed types like ‘Turpentine’ in Florida (Malo and Campbell, 1978), ‘Turpentine’, ‘Coquinho’ and ‘Espada’

V. Galán Saúco308
in Brazil (Ferreira, 1989), and ‘Gomera 1’ in the Canary Islands (Cracknell Tor-res et al., 2003a). There is at least one documented case of IFB in Australia involving fruit of the polyembryonic ‘Kamerunga White’ (Winston, 1984).
Cultivar-related differences with respect to IFB incidence have been described by Cracknell Torres et al. (2003a). They studied 28 cultivars and observed that ‘Edward’, ‘Gomera 1’, ‘Irwin’, ‘Valencia Pride’, ‘Mabroka’, ‘Ah Ping’ and ‘Heidi’ were almost free of this disorder. Modern Israeli cultivars, such as ‘Shelly’ and ‘Tango’ have also exhibited low frequency of IFB (Lavi etal., 1997a, b). Observations of the extent of spongy tissue in various hybrids and selfed progenies of ‘Alphonso’ clearly indicate that this disorder is genetically determined, and ‘Alphonso’ is apparently homozygous recessive for this character (Iyer and Subramanyan, 1992).
Control
Despite the close relationship that exists between nutrition and IFB, very few practical recommendations have been proposed for controlling this problem. There is general agreement with the observations of Young (1960) and Young and Miner (1961) that maintaining a low leaf content of nitrogen (<1.2%) and a calcium level ≥ 2.5% minimizes the amount of affected fruits. High nitrogen levels enhance vegetative growth and rapid fruit development. Calcium moves only in the xylem. Calcium concentration in organs such as fruits, which have a low rate of transpiration and which are preferably supplied via the phloem, can result in calcium levels falling below the critical level required for membrane integrity and cell wall stability (Marschner, 1995). The antagonistic effect of nitrogen fertilization, especially when ammonium salts are applied, and calcium uptake and accumulation on leaves and fruits, is well documented in many fruit species (Shear, 1975; Lewis et al., 1977; Lud-ders, 1979) and has been clearly demonstrated for mangoes (Young et al., 1962, 1965).
Lime application has also been recommended to increase the cation exchange percentage to values ≥7.0 (Ferreira, 1989). According to Schaffer (1994), IFB has been corrected in ‘Keitt’ by calcium application either to the soil as calcium carbonate (CaCO3) or as a foliar calcium nitrate (Ca (NO3)2) spray.
Cultural practices such as mulching have been cited as being benefi cial for reducing IFB. Mulching lowers the soil temperature and thereby miti-gates the refl ected or rising heat that eventually affects the fruit (Katrodia, 1988; Lad et al., 1992). The accompanying reduction in the transpiration stream of the tree and the consequent minor mobilization of calcium to the fruit (Bangerth, 1979) may also be important.
Certain rootstocks can improve calcium uptake and accumulation and provides an important new procedure for controlling IFB. Fieldwork was initiated in Malaysia at the end of the last century (Tengku Ab.Malik, 1996), but further studies remain unreported.
Early harvesting at the green-ripe stage has been recommended as a measure to reduce IFB (Young, 1957; Young and Miner, 1961; Galán Saúco

Physiological Disorders 309
et al., 1984; Winston, 1984), but for some cultivars this practice results in lower quality fruits. The elimination at harvesting of fruits that do not exude latex (Mead and Winston, 1991) and avoiding the exposure of fruit to direct sunlight during harvest (Gunjate et al., 1982) have been recommended as prac-tical methods to reduce the incidence of IFB in mangoes in the market. Accord-ing to Santos Filho et al. (2002), postharvest hydrothermal treatments should be avoided in fruits coming from orchards with a history of IFB incidence.
Preharvest dips of 0.5–2.0% of calcium chloride (CaCl2), applied from the second month after fruit set until harvest, can increase calcium content in the fruit, and has been reported to be an effective control measure for spongy tissue in fruit of ‘Alphonso’ (Gunjate et al., 1979). In contrast, foliar sprays with calcium have been reported to be ineffective for increasing leaf cal-cium content (McKenzie, 1995), and IFB has even been increased by applica-tion of calcium, which indicates that a nutrition imbalance within the fruit was induced (Oosthuyse, 1997).
An integrated approach to control insidious fruit rot incidence in ‘Haru-manis’ has been developed in Malaysia. This includes maintaining soil pH close to 6.0, late pruning, application of calcium sprays (2% CaCl2) to the fruits 4–6 weeks after fl owering, supplemental irrigation, monitoring nitro-gen and potassium fertilization and anticipated harvesting (Tengku Ab.Malik, 1992a, b). Galán Saúco (1999) suggested the following general inte-grated management measures: avoid planting sensitive cultivars (i.e. ‘Sensa-tion’, ‘Tommy Atkins’, etc.); use appropriate rootstocks (i.e. ‘Tangkai Panjang’ or others with high calcium absorption capacity); harvest at the green-ripe stage; use an appropriate fertilization programme that is rich in calcium and poor in nitrogen; avoid excessive irrigation close to fruit maturity; and mulch soil in regions with hot summers.
There are notable differences in IFB resistance among cultivars. Because of the widespread incidence of this problem, breeding for rootstocks and sci-ons for IFB resistance is critical for resolving this problem (Iyer and Degani, 1997). Iyer and Subramanyan (1992) have described the progress of breeding studies to overcome IFB in India, obtaining hybrids of ‘Alphonso’ by other Indian cultivars free from spongy tissue.
Biotechnological approaches also merit attention. Litz and Lavi (1997), based on work done in tomatoes to control jelly seed (Smith et al., 1990; Oeller et al., 1991), have suggested that control of IFB could be achieved by manipulating ripening using the antisense strategy, either by blocking ethyl-ene biosynthesis or with polygalacturonase, but no work has been reported in mango so far to test this suggestion.
9.3 Lumpy Tissue
This disorder, of unknown etiology, occurs frequently in Thailand, where it occurs mainly in the cultivars ‘Namta-an’, ‘Parg-grabong’ and ‘Pinsen Dang’, and in the Philippines in ‘Pico’. The disorder becomes apparent as fruit ripening begins, with indentations or scoring of the peel, which become

V. Galán Saúco310
increasingly marked as ripening progresses. Opened fruits display a meso-carp full of white lumps of intact, starch-fi lled cells (Lizada et al., 1984).
9.4 Ricey Tissue
This disorder is of unknown origin, but is common in ‘Carabao’ in the Philip-pines. No external symptoms are visible, but internal symptoms are found in ripening fruit and even at physiological maturity, and consist of small lesions. The lesions resemble grains of rice in size and aspect (hence the name) on the mesocarp, surrounded by cotton wool-like tissue. Except for the small changes in the texture of the affected area, no other organoleptic changes have been detected (Lizada et al., 1984).
9.5 Fruit Cracking
The only symptom of this disorder is that the fruit crack suddenly while still on the tree, the cracks forming clean, knifelike cuts breaking the skin and into the fl esh. Secondary infections by Colletotrichum gloeosporioides (the causal agent of anthracnose) or other fungal pathogens can occur. The cause of fruit cracking appears to be related to water tension differences in the skin during periods of high relative humidity and it occurs when trees are heavily irrigated after a prolonged dry period, and if heavy rains are intermixed with dry spells. Frequently almost all of the fruits on the tree are affected, although those near maturity are the most susceptible. The incidence of fruit cracking seems to be higher in cultivars with little or no fi bre. Control measures may include regular watering and mulching, but the ultimate solution may lie in breeding as in IFB (Lim and Khoo, 1985).
Another cause for fruit splitting is infection by bacterial black spot (Xanthomonas campestris), which is considered to be a major cause of fruit splitting. Consequently, prevention is the only solution to the problem by ensuring adequate wind protection, which is also benefi cial if the damage is caused by differences in water tension of the skin cells, and/or appropriate chemical sprays to prevent infection (Meurant et al., 1997).
9.6 Black Tip Disorder
This disorder is widespread in India (Prakash and Srivastava, 1987). Ram (1988) indicated that it was fi rst described in 1909. Three disorders frequently cited prior to 1958, namely taper tip, tip pulp and girdle necrosis, are in fact variants of black tip. The fi rst symptom of this disorder is the etiolation and yellowing of the distal end of fruit, which then turns black and hardens, causing the fruit to ripen prematurely and making it unmarketable (Plate 53). The vascular bundles in the pedicel may turn brown and decay. Affected fruits do not secrete latex at harvest.

Physiological Disorders 311
Outside India, the only known occurrence of black tip is from the Guang-dong province of China (Zhang et al., 1995). In both India and China, the disorder occurs in areas close to brick-making kilns, particularly where trees are more exposed to fume-laden winds coming from the brick works. Although not fully assessed, it seems that fl uorine is the causal agent of this disorder (Zhang et al., 1995), although a dry hot climate may enhance the effect of gases. Differences in susceptibility among Indian cultivars have been reported by Ram (1988), who also suggested the possibility of varietal selection for orchards in the vicinity of brick kilns. Majumder and Sharma (1985) recom-mend a minimum distance between kiln and trees of 1.6 km in the path of prevailing winds and 0.8 km on the other sides, as well as recommending prevention by spraying three times with borax (0.6%) and caustic soda (0.8%), for example prior to fl owering, during fl owering and at fruit set.
9.7 Lenticel Spot
This disorder is characterized by the development of small spots of corky tissue in the skin lenticels that darken as the fruit changes colour during ripening, which makes the fruit unmarketable. The causes are not entirely uncertain, but it is most often associated with incorrect postharvest practices, including low storage temperatures, high humidity of the storage atmos-phere, excessive immersion time in postharvest dips and excessive detergent in wash water (Oosthuyse, 1993; Meurant et al., 1997). Some growers link it to other causes, such as delayed harvesting or wet conditions during picking. Differences among cultivars with respect to the severity of this problem have been observed. It is very common in the late harvest of ‘Keitt’ in the subtrop-ics at the beginning of winter under cooler and wetter conditions.
9.8 Other Fruit Disorders
Spots and marks on green fruit can have several different causes: cold dam-age or chilling injury, usually seen as small raised brown or discoloured spots on the skin, or pitting surrounded by sunken areas affecting the epidermis and the endocarp (Lizada et al., 1984). Storage temperatures below 13°C have been identifi ed as major causes of chilling injury, and must be avoided (Meurant et al., 1997). High temperatures or sudden exposure to sunlight can cause sunburn, which is characterized by bleached or yellow patches on the skin, which may become leathery, yellow-brown to black, and lightly sunken. Trees should be well irrigated during fruit fi lling and harvested fruit must always be kept (even briefl y) in full shade.
Wind damage, resulting from prolonged rubbing of the fruit against leaves or dead twigs, requires adequate windbreaks and timely pruning of dead wood. If hail damage is likely to be a frequent problem, the whole orchard will need to be protected by nets. Damage caused during harvest, storage and transport, can only be reduced by careful handling. Sapburn,

V. Galán Saúco312
caused by prolonged contact with latex leaking onto the skin from a cut stem, can be minimized by suitable harvesting and packing operations. High levels of carbon dioxide (CO2) during storage provoke the development of off-fl avours and small internal lesions, as well as exudating brown tissues (Chaplin, 1986) and inhibition of ripening (Thompson, 1971).
A disorder named pulpa negra, literally ‘black fl esh’, which speaks quite clearly as to the symptoms, has been reported in Mexico, especially in ‘Haden’ (Mora et al., 1998). Although the problem was detected in fruits that had been stored at 13°C for more than 20 days, it is not entirely clear that low tempera-ture is the only cause as it also occurred in fruits reportedly stored only at room temperature.
9.9 Stem Disorders
While some slight cracking of the bark around the trunk may be normal, some cultivars, like ‘Manzanillo’ or ‘Gomera 4’, are prone to fi ssuring, with an unusually high degree of suberifi cation. This disorder is common in the Canary Islands, Spain, and it is more pronounced on the sides of the trunk that are wetted by sprinklers than on the less exposed sides (Fernández Galván and Galán Saúco, unpublished). This phenomenon of stem cracking has been also associated with bad orchard management, particularly with bad irrigation practices (Mora et al., 1998). Some rootstocks, including ‘Gomera 1’ in the Canary Islands, frequently exhibit gall-like growths along the branches, thought to be linked to climatic factors which facilitate swelling of lateral buds without being enough to provoke fl ushing and the corresponding shoot elongation.
References
Ahmad Tarmizi, S., Tengku Ab.Malik, T.B., Pauziah, M. and Zahrah, T. (1993) Incidence of insidious fruit rot as related to mineral nutrients in Harumanis mangoes. Malay-sian Agricultural Research and Development Institute (MARDI) Research Journal21, 43–49.
Atkinson, D., Jackson, J.E., Sharples, R.O. and Wallery, W.M. (1980) Mineral Nutrition of Fruit Trees. Butterworths, London.
Bangerth, F. (1979) Calcium related disorders of plants. Annual Review of Phytopathol-ogy 17, 97–122.
Bower, J.P. and Cutting, J.G.M. (1987) Some factors affecting pre-harvest quality in avo-cado fruit. South African Avocado Growers’ Association Yearbook 10, 143–146.
Brodrick, H.T. and Thord-Gray, R. (1982) Irradiation in perspective – the signifi cance for the mango industry. South African Mango Growers’ Association Research Report 2, 23–36.
Campbell, R.F. (1992) A Guide to Mangos in Florida. Fairchild Tropical Garden, Miami, Florida.
Chaplin, G.R. (1986) Postharvest physiology of mango fruit. In: Proceedings of the First Australian Mango Research Workshop. Commonwealth Scientifi c and Industrial Research Organization (CSIRO), Melbourne, Australia, pp. 261–270.

Physiological Disorders 313
Cheema, G.S. and Dani, P.G. (1934) Report on the export of mangoes to Europe in 1932 and 1933. Bulletin of the Department of Agriculture, Bombay 170, 1–31.
Cirulli, M. and Ciccarese, F. (1981) Effect of mineral fertilizers on the incidence of blossom-end-rot of watermelon. Phytopathology 71, 50–53.
Cracknell Torres, A. and Galán Saúco, V. (2003) The study of the problem of mango (Mangifera indica L.) internal breakdown. Acta Horticulturae 645, 167–174.
Cracknell Torres, A., Cid Ballarín, M.C., Socorro Monzón, A.R., Fernández Galván, D. and Galán Saúco, V. (2003a) Incidence of internal fruit breakdown in various man-go (Mangifera indica L.) cultivars. Acta Horticulturae 645, 315–318.
Cracknell Torres, A., Cid Ballarín, M.C., Socorro Monzón, A.R., Fernández Galván, D., Rosell García, P. and Galán Saúco, V. (2003b) Effects of nitrogen and calcium supply on the incidence of internal fruit breakdown in ‘Tommy Atkins’ mangoes (Mangifera indica L.) grown in a soilless system. Acta Horticulturae 645, 387–394.
De Laroussilhe, F. (1980) Le Manguier. Techniques Agricoles et Productions Tropicales.Maisonneuve et Larose, Paris, France.
Esquerra, E.B., Brena, S.R., Reyes, M.U. and Lizada, M.C.C. (1990) Physiological break-down in vapour heat-treated ‘Carabao’ mango. Acta Horticulturae 269, 425–434.
Farré, J.M. and Hermoso, J.M. (1993) Mulching and irrigation effects on growth, cropping and fruit quality of the mango cv. Sensation. Acta Horticulturae 341,295–302.
Ferreira, F.R. (1989) Colapso interno do fruto. Simposio Brasileiro Sobre A Cultura Da Mangueira 2. Faculdade de Ciencias Agrarias e Veterinarias; Sociedade Brasileira de Fruticultura, Jaboticabal, Brazil, pp. 149–155.
Galán Saúco, V. (1999) El Cultivo del Mango. Mundi-Prensa, Madrid, Spain.Galán Saúco, V. and Fernández Galván, D. (1987) El cultivo del mango en Canarias.
Cuadernos de Divulgación 1/87. Consejería de Agricultura Ganadería y Pesca, Gobierno de Canarias, Spain.
Galán Saúco, V., Fernández Galván, D. and Calvo, R. (1984) The incidence of soft-nose in mangos in the Canary Islands. Proceedings of the Florida State Horticultural Society 97, 358–360.
Gunjate, R.T., Tare, S.J., Rangwala, A.D. and Limaye, V.P. (1979) Effect of pre-harvest and post-harvest calcium treatments on calcium content and occurrence of spongy tissue in ‘Alphonso’ mango fruits. Indian Journal of Horticulture 37, 140–144.
Gunjate, R.T., Walimbe, B.P., Lad, B.L. and Limaye, V.P. (1982) Development of internal breakdown in ‘Alphonso’ mango by post-harvest exposure of fruits to sunlight. Science and Culture 48, 188–190.
Gupta, D.N., Lad, B.L., Chavan, A.S. and Salvi, M.J. (1985) Enzyme studies on spongy tissue: a physiological ripening disorder in ‘Alphonso’ mango. Journal of Maharash-tra Agricultural University 10, 280–282.
Hermoso, J.M., Guirado, E., Gómez, R., Castilla, A., Velasco, J.M. and Farré, J.M. (1996) Effects of nutrients and growth substances on internal breakdown of ‘Sensation’ mango fruits. Acta Horticulturae 455, 92–99.
International Board for Plant Genetic Resources (IBPGR) (1989) Descriptors for Mango.IBPGR, Rome.
Iyer, C.P.A. and Degani, C. (1997) Classical breeding and genetics. In: Litz, R.E. (ed.) The Mango, Botany, Production and Uses. CAB International, Wallingford, UK, pp. 49–68.
Iyer, C.P.A. and Subramanyan, M.D. (1992) Possibilities of overcoming physiological disorders in mango by breeding. Acta Horticulturae 317, 241–244.
Joshi, G.D. and Roy, S.K. (1985) Spongy tissue in mango, a physiological disorder. Indian Journal of Horticulture 29, 21–22.

V. Galán Saúco314
Katrodia, J.S. (1988) Spongy tissue in mango – causes and control measures. Acta Hor-ticulturae 23, 814–826.
Katrodia, J.S. and Chuva, H.P. (1993) Spongy tissue in mango. In: Chadha, K.L. and Pa-reek, O.P. (eds) Advances in Horticulture Vol. 4 – Fruit Crops: Part 4. Malhotra Publishing House, New Delhi, India, pp. 2069–2080.
Katrodia, J.S. and Rane, D.A. (1989) The pattern of distribution of spongy tissue in the affected ‘Alphonso’ fruits at different locations. Acta Horticulturae 231, 873–877.
Knight, R.J., Jr (1997) Important mango cultivars and their descriptors. In: Litz, R.E. (ed.) The Mango, Botany, Production and Uses. CAB International, Wallingford, UK, pp. 545–566.
Krishnamurthy, S. (1980) Internal breakdown during ripening of ‘Alphonso’ mango (Mangifera indica Linn.) in relation to specifi c gravity of the fruit. Journal of Food Science and Technology 17, 198–199.
Krishnamurthy, S. (1981) Chemical studies on internal fruit breakdown in ‘Alphonso’ mango (Mangifera indica Linn). Journal of Horticultural Science 56, 247–250.
Lad, B.L., Gunjate, R.T. and Salvi, M.J. (1992) Causes and control measures of spongy tissue disorder in ‘Alphonso’ mango fruit: an integrated approach. MaharashtraJournal of Horticulture 6, 25–32.
Lavi, U., Kaufman, D., Sharon, D. and Tomer, E. (1997a) ‘Tango’: a new mango cultivar. HortScience 32, 137.
Lavi, U., Kaufman, D., Sharon, D. and Tomer, E. (1997b) ‘Shelly’: a new mango cultivar. HortScience 32, 138.
Lewis, T.L., Martin, D., Cerny, J. and Ratkowsky, D.A. (1977) The effect of increasing the supply of nitrogen, phosphorous, calcium and potassium to the roots of Morton Worcester apple trees on leaf and fruit composition and on the incidence of bitter pit at harvest. Journal of Horticulture Science 57, 409–419.
Lim, T.K. and Khoo, K.C. (1985) Diseases and Disorders of Mango in Malaysia. Tropical Press SDN. BDH. Kuala Lumpur, Malaysia.
Litz, R.E. and Lavi, U. (1997) Biotechnology. In: Litz, R.E. (ed.) The Mango, Botany, Production and Uses. CAB International, Wallingford, UK, pp. 401–424.
Lizada, M.C.C., Kosiyachinda, S. and Mendoza, D.B. (1984) Physiological disorders of mango. In: Mendoza, D.B. and Willis, R.B.H. (eds) Mango. Association of South-east Asian nations (ASEAN) Food Handling Bureau, Kuala Lumpur, Malaysia, pp. 68–74.
Ludders, P. (1979) The effect of nitrogen nutrition on bitter pit in apple. Communications in Soil Science and Plant Analysis 6, 261–272.
Majumder, P.K. and Sharma, D.K. (1985) Mango. In: Bose, T.K. (ed.) Fruits of India. Tropical and Subtropical. Naya Prokash, Calcutta, India, pp. 69–153.
Malo, S.E. and Campbell, C.W. (1978) Studies on mango fruit breakdown in Florida. Pro-ceedings of the American Society for Horticultural Science Tropical Region 22, 1–15.
Marschner, H. (1995) Mineral Nutrition of Higher Plants. Academic Press. London.McKenzie, C.B. (1995) The effect of calcium and potassium sprays on ‘Sensation’ mango
leaf nutrients concentration and fruit quality. South Africa Mango Growers’ Asso-ciation Yearbook 15, 44–47.
Mead, A.J. and Winston, E.C. (1991) Description of the disorder ‘stem-end cavity’, possi-ble causes and suggestions for reducing its incidence in packing sheds. Acta Horti-culturae 291, 265–271.
Melin, P. and Aubert, B. (1973) Observations on a type of abnormal maturation of banana (yellow pulp) before harvesting. Fruits 28, 831–842.
Meurant, N., Holmes, R., MacLeod, N., Fullelove, G., Bally, I. and Kernot, I. (1997) Mango Information Kit. Agrilink Series. Queensland Horticulture Institute, Depart-ment of Primary Industries, Brisbane, Queensland, Australia.

Physiological Disorders 315
Mora, A.A., Vega, P.A. and Téliz, O.D. (1998) Enfermedades del mango. In: Téliz, D. (ed.) El Mango y su Manejo Integrado en Michoacán. GUIM (Grupo Interdisciplin-ario de Investigación en Mango). Colegio de Postgraduados, Montecillo, Texcoco, México, pp. 18–44.
Nuevo, P.A., Cua, A.U. and Lizada, M.C.C. (1984) Internal breakdown in ‘Carabao’ mango subjected to modifi ed atmospheres. III. Starch in the spongy tissue. Posthar-vest Research Notes 1, 34–35.
Oeller, P.W., Lu, M.W., Taylor, L.P., Pike, D.A. and Theologis, A. (1991) Reversible inhi-bition of tomato fruit senescense by antisense RNA. Science 254, 437–439.
Oosthuyse, S.A. (1993) Disorders of fi breless mangos grown in South Africa for export. South Africa Mango Growers’ Association Yearbook 13, 80–88.
Oosthuyse, S.A. (1997) The effect of calcium and magnesium chelate sprays at fl owering on fruit quality and physiological disorders in mango. South Africa Mango Growers’ Association Yearbook 17, 29–32.
Patkar, V.R., Gunjate, R.T. and Lad, B.L. (1983) Effect of spongy tissue on chemical com-position of ripe ‘Alphonso’ fruit. South Indian Horticulture 32, 83–85.
Prakash, O. and Srivastava, K.C. (1987) Mango Diseases and their Management. A World Review. Today and Tomorrow’s Publishers, New Delhi, India.
Ram, S. (1988) Factors associated with black tip and internal necrosis in mango and their control. Acta Horticulturae 231, 797–804.
Raymond, L., Schaffer, B., Brecht, J.K. and Crane, J.H. (1998) Internal breakdown in mango fruit: symptomatology and histology of jelly seed, soft nose and stem-end cavity. Postharvest Biology and Technology 13, 59–70.
Roe, B. and Bruemmer, J.H. (1981) Changes in pectic substances and enzymes during ripening and storage of ‘Keitt’ mangoes. Journal of Food Science 46, 186–189.
Santos Filho, H.P., Cavalcanti Cruz de Holanda Tavares, S., Pires de Matos, A., De Ol-iveira Costa, V.S., António, W. and Ferreira dos Santos, C. (2002) Doenças. Monito-ramento e controle. In: De Carvalho Genú, P.J. and De Queiroz Pinto, A.C.A. (eds) Cultura da Mangueira. Embrapa Informaçao Tecnológica, Brasilia, pp. 299–352.
Schaffer, B. (1994) Mango disorders caused by abiotic factors. In: Ploetz, R.C., Zent-myer, G.A., Nishijima, W.T., Rohrbach, K.G. and Ohr, H.D. (eds) Compendium of Tropical Fruit Diseases. APS Press, The American Phytopathological Society, Min-nesota, USA, pp. 43–44.
Shear, C.B. (1975) Calcium nutrition and quality in fruit crops. Communications in Soil Science and Plant Analysis 6, 233–244.
Singh, R., Kumar, J.C. and Nandpuri, K.S. (1975) A study on the infl uence of the struc-tural chemical constituents of the skin of watermelon (Citrullus lanatus Sch.) fruit on the incidence of its blossom-end-rot and cracking (physiological disorders). In-dian Journal of Horticulture 32, 98–101.
Smith, C.J.S.,Watson, C.F. and Morris, P.C. (1990) Inheritance and effect on ripening of antisense polygalacturonase genes in transgenic tomatoes. Plant Molecular Biology14, 369–374.
Subramanyan, H., Krishnamurthy, S., Subhadra, N.V., Dalal, V.B., Rahdhawa, G.S. and Chacko, E.K. (1971) Studies on internal breakdown, a physiological ripening disor-der in ‘Alphonso’ mangoes (Mangifera indica L.). Tropical Science 13, 203–210.
Tengku Ab.Malik, T.M. (1992a) Pattern of calcium accumulation in ‘Harumanis’ mango fruits at different stages of maturity. In: Proceedings of the International Radiation Protection Association (IRPA) Seminar (Agriculture Sector). Vol.1. Crops and Plants,Kuala Lumpur, Malaysia, pp. 399–400.
Tengku Ab.Malik, T.M. (1992b) Measures to control and prevent Insidious Fruit Rot (IFR) in ‘Harumanis’ mangoes. In: Proceedings of the International Radiation Protection

V. Galán Saúco316
Association (IRPA) Seminar (Agriculture Sector). Vol. 1. Crops and Plants, Kuala Lumpur, Malaysia, pp. 405–406.
Tengku Ab.Malik, T.M. (1996) Screening for mango rootstocks with high calcium uptake. In: Vijaysegaran, S., Pauziah, M., Mohamed, M.S. and Ahmad Tarmizi, S. (comps.) Proceedings of the International Conference on Tropical Fruits. Vol. III. Supplemen-tary, 23–26 July, Kuala Lumpur, Malaysia. Malaysian Agricultural Research and Development Institute (MARDI), Kuala Lumpur, Malaysia, pp. 189–194.
Thompson, A.K. (1971) The storage of mango fruits. Tropical Agriculture, 48, 63–70.Van Lelyveld, L.J. and Smith, J.H.E. (1979) Physiological factors in the maturation and
ripening of mango (Mangifera indica L.) fruit in relation to the jelly-seed physiolog-ical disorder. Journal of Horticultural Science 54, 283–287.
Van Lelyveld, L.J., Van Gerrish, C. and Dixos, R.A. (1984) Enzyme activities and poly-phenols related to mesocarp discolouration of avocado fruit. Phytochemistry 23, 1531–1534.
Velasco Cárdenas, J. (1974) El Mango en México. Descripción, Cultivo, Mejoramiento y Utilización. Comisión Nacional de Fruticultura, SAG, Mexico.
Wainwright, H. and Burbage, M.B. (1989) Physiological disorders in mango (Mangiferaindica L.) fruit. Journal of Horticultural Science 64, 125–135.
Winsor, G.W. and Adams, P. (1987) Diagnosis of Mineral Disorders in Plants. Vol. 3. Glasshouse Crops. Her Majesty’s Stationery Offi ce (HMSO), London.
Winston, E.C. (1984) Observations of internal mango fl esh breakdown: need for stan-dardization of terminology. In: Proceedings of the First Australian Mango Research Workshop. Commonwealth Scientifi c and Industrial Research Organization (CSIRO) Press, Melbourne, Australia, pp. 77–82.
Young, T.W. (1957) ‘Soft-nose’, a physiological disorder in mango fruits. Proceedings of the Florida State Horticultural Society 70, 280–283.
Young, T.W. (1960) Response of ‘Kent’ mangoes to nitrogen fertilization. Proceedings of the Florida State Horticultural Society 73, 334–336.
Young, T.W. and Miner, J.T. (1961) Relationship of nitrogen and calcium to ‘soft-nose’ disorder in mango fruits. Journal of the American Society for Horticultural Science78, 201–208.
Young, T.W., Koo, R.C.J. and Miner, J.T. (1962) Effects of nitrogen, potassium and calci-um fertilization on ‘Kent’ mangoes in deep, acid, and sandy soil. Proceedings of the Florida State Horticultural Society 75, 364–371.
Young, T.W., Koo, R.C.J. and Miner, J.T. (1965) Fertilizer trials with ‘Kent’ mangoes. Pro-ceedings of the Florida State Horticultural Society 78, 369–375.
Zhang, Ch., Huang, H. and Kuang, Y. (1995) A study of the cause of the mango black tip disorder. Scientia Horticulturae 64, 49–54.

© CAB International 2009. The Mango, 2nd Edition: Botany, Production and Uses(ed. R.E. Litz) 317
10 Pests
J.E. Peña,1 M. Aluja2 and M. Wysoki31University of Florida, Florida, USA
2Instituto de Ecología, AC, Xalapa, Veracruz, Mexico 3Institute of Plant Protection, Bet Dagan, Israel
10.1 Introduction 31710.2 Mango Fruit Pests 318
Fruit fl ies 318Fruit fl y control: brief overview 322Mango seed weevils 328Mango seed borer (Lepidoptera: Pyralidae) 331Fruitspotting bugs (Hemiptera; Coreidae) 332Thrips 333Blossom pests 334Pests of buds and leaves 338Pests of trunks, twigs and roots 344
10.3 Discussion 345
10.1 Introduction
Mango, like most fruit trees, is usually attacked by two or three key pests, several secondary pests and by a large number of occasional pests in local-ized areas where it is grown. Worldwide lists of pests of mango have been published by Laroussilhe (1980), Tandon and Verghese (1985) and Veeresh (1989). The pests of mango in India (Srivastava, 1998; Anonymous, 2006), Australia (Anonymous, 1989), Pakistan (Mohyuddin, 1981), Israel (Wysoki et al., 1993; Swirski et al., 2002), the USA (Peña, 1993), West Africa (Vannière et al., 2004), Brazil (Assis and Rabelo, 2005), Central America (Coto et al., 1995) and Puerto Rico (Martorell, 1975) have also been described. Some publications contain check lists of mango pests and most contain details of life histories and control of mango pests (Morin, 1967; Golez, 1991; Murray, 1991).
Of c.322 species of insects and mites that have been recorded as minor and major pests of mango, 127 (39%) are foliage feeders, 87 (27%) are fruit feeders, 36 (12%) feed on the infl orescence, 33 (10%) inhabit buds and 39

J.E. Peña et al.318
(12%) feed on branches, the trunk and roots. The four key pests (fruit fl ies, seed weevils, tree borers and mango hoppers) require annual control mea-sures. Secondary pests generally occur at sub-economic levels, but can become serious pests as a result of changes in cultural practices and cultivar or because of indiscriminate use of pesticides against a key pest. For exam-ple, Mohyuddin and Mahmood (1993) reported that scale insects became serious pests following non-judicious use of insecticides against fruit fl ies. Similarly, mites, Oligonychus spp., are secondary pests of mango, which can become serious because of human intervention. Occasional or incidental pests also can cause economic damage only in localized areas at certain times. The majority of pests reported here are of this category.
10.2 Mango Fruit Pests
With current world emphasis on quality fruit for local consumption and export, insects that blemish fruit by feeding, scratching or ovipositing in the pulp or seed can cause high losses. Only fruit fl ies, seed weevils and lepi-dopterous larvae actually penetrate the fruit pulp and seed. The feeding of other pests (e.g. Othreis materna (L.), Gonodonta pyrgo (Cram.), Gonodonta clo-tilda (Stoll) and Leptoglossus stigmai (Herbest)) often extends only into the pulp of ripening mangoes (Angeles and Requena, 1966).
Fruit fl ies
Most mango-producing countries are in fruit-fl y infested areas, and produc-ers suffer signifi cant direct economic losses (larval feeding renders fruit unmarketable) and indirect economic losses (quarantine restrictions hinder-ing exports), resulting from the presence of fruit fl ies (Hill, 1975; Umeya and Hirao, 1975; Anonymous, 1987; Yee, 1987; Singh, 1991; Aluja, 1993; Aluja et al., 1996; Vannière et al., 2004; Aluja and Mangan, 2008). Few insects have a greater impact on international marketing and world trade in agricultural produce than tephritid fruit fl ies (Hendrichs, 1996; Aluja and Mangan, 2008). Approximately 60 species of fruit fl ies are reported to attack mango and a related species, Mangifera foetida (White and Elson-Harris, 1992; Aluja et al., 1996; Malavasi and Zucchi, 2000; Clarke et al., 2001, 2005; Norrbom, 2004; Vayssières et al., 2005). Fruit fl ies attacking mango belong to the genera Anastrepha (c.12 species), Bactrocera (c.33 species), Ceratitis (eight species) and Dirioxa (two species) (White and Elson-Harris, 1992; Vayssières and Kala-bane, 2000; Lux et al., 2003; Norrbom, 2004; Vayssières et al., 2004, 2005; Sec-retariat of the Pacifi c Community, 2005). Some of these species are referred to as the ‘mango fruit fl y’: Anastrepha obliqua Macquart, Bactrocera frauenfeldi (Shiner), Ceratitis cosyra (Walker) (Aluja, 1993; Leblanc and Allwood, 1997; Lux et al., 2003; Steck, 2003). All Dacus species that attack mango have been recently placed in the genus Bactrocera (Drew) (Thomson, 2005).

Pests 319
The following discussion is mainly restricted to the biology, ecology and management of mango-infesting fruit fl ies. Regulatory issues, such as risk analysis and postharvest treatments are discussed by Johnson and Hofman in Chapter 15, this volume. Postharvest treatments specifi cally related to mangoes and fruit fl ies have been discussed by Sharp et al. (1988, 1989a, b), Hallman and Sharp (1990), Nascimento et al. (1992), Mangan and Hallman (1998), Shellie and Mangan (2002a, b) and Bustos et al. (2004). Broad regula-tory issues were recently reviewed by Follet and Neven (2006).
General reviews on biology, ecology and behaviour of economically important and non-pestiferous fruit fl ies, many of them infesting mangoes, were written or edited by Christenson and Foote (1960), Bateman (1972), Fletcher (1987), Robinson and Hooper (1989), Aluja (1994), Aluja and Norrbom (2000) and Malavasi and Zucchi (2000).
AnastrephaAnastrepha spp. are endemic to the western hemisphere and their range extends from the southern USA to northern Argentina and includes the Caribbean islands (Aluja, 1994) (Plate 54). Twelve Anastrepha species have been purportedly associated with mango (Norrbom, 2004). Of these, A. obli-qua, A. ludens (Loew) and A. suspensa (Loew) stand out as economically important pests of mangoes (Aluja et al., 1996; Norrbom, 2004). Anastrepha obliqua is reportedly the most common fruit fl y pest in the Americas (Jirón and Hedström, 1988; Nascimento et al., 1992). This species is the most com-mon fruit fl y pest of mangoes in Mexico, Costa Rica, Honduras and Guate-mala (Soto-Manitiú et al., 1987; Jirón and Hedström, 1991; Aluja et al., 1996; Camargo et al., 1996; Sponagel et al., 1996), but it also attacks mangoes in Cuba, Puerto Rico, Jamaica, El Salvador and Venezuela (Norrbom, 2004). In Mexico, A. ludens commonly attacks mangoes at higher elevations, while A. obliqua dominates at lower altitudes (Aluja et al., 1996). In Brazil and Ecuador, mangoes are mainly attacked by A. fraterculus (Wiedemann), but A. turpiniae Stone, A. serpentina Wiedemann, A. pseudoparallela (Loew) and A. zuelaniae Stone have been reported (Zucchi, 1988; Rebouças et al., 1996; Arias and Jines, 2004; Norrbom, 2004; Barbosa et al., 2005; A. Malavasi, January 2008, São Paulo, personal communication).
BIOLOGY OF ANASTREPHA FRUIT FLIES. Anastrepha fruit fl y biology today is based on basic studies carried out between 1900 and 1944 (Aluja, 1994, 1999). The basic life cycle is very similar among most pestiferous Anastrepha species. For example, egg incubation of A. ludens in mango requires 3.8 days, larval devel-opment requires 14.2 days and pupal development has been known to need 14.2 days at 27 ± 2°C (Leyva et al., 1991). Larvae pass through three instars before emerging from the fruit and burrowing into the ground to pupate (Aluja, 1994). Clutch size varies between one egg/clutch in A. obliqua and >100 eggs in A. grandis Mcquart (Aluja, 1994). Importantly, in species laying sev-eral eggs per clutch (e.g. A. ludens), clutch size is determined by females on a case-by-case basis and is greatly infl uenced by degree of ripeness and the concomitant degree of epicarp hardness (Díaz-Fleischer and Aluja, 2003;

J.E. Peña et al.320
Birke et al., 2006). Life expectancy varies greatly depending on the host on which the larvae developed and environmental conditions (for example see Toledo and Lara 1996, and Malavasi and Zucchi, 2000), but some adults can live for >150 days (Aluja et al., 2000).
Abundance of A. obliqua populations has been positively correlated with temperature and negatively correlated with relative humidity (RH) (Herrera and Viñas, 1977). However, studies by Celedonio-Hurtado et al. (1995) and Aluja et al. (1996) demonstrated the lack of a clear relationship between rain-fall and Anastrepha fl y captures in mango orchards in Mexico. They indicated that overall population fl uctuation patterns can vary greatly among orchards within a fairly small geographic region.
BactroceraBactrocera spp. are pests of mango in Africa and Australasia (Drew, 1989; Leblanc and Allwood, 1997; Leblanc et al., 1997; Tenakanai, 1997; Hancock et al., 2000; Hollingsworth et al., 2003; Clarke et al., 2005) (Plate 55). The common species reported on mango include B. tryoni (Frogatt), B. zonata (Saunders), B. dorsalis (Hendel), B. neohumeralis (Hardy), B. jarvisi (Tryon), B. papayae Drew and B. frauenfeldi (Schiner) (Umeya and Hirao, 1975; Drew and Hancock, 1994; Hollingsworth et al., 2003). Two species, B. phillippiensis Drew & Hancock and B. occipitalis Bezzi, have been recorded for Palau, Pacifi c Islands (Secretariat of the Pacifi c Community, 2005), and recently, a new spe-cies, B. invadens Drew, Tsuruta and White, was reported for West Africa (Kenya, Benin) (Lux et al., 2003; Vayssières et al., 2005). Bactrocera correcta (Bezzi), B. caryeae (Kapoor), B. curcubitae (Coquillett), B. diversa (Coquillett) and B. tau (Walker) have been reported in India (Australian Government, 2004).
BIOLOGY OF BACTROCERA FRUIT FLIES. The biology of dacine fruit fl ies was most recently reviewed by Fletcher (1987); additional details can be found in Christenson and Foote (1960), Bateman (1972), Robinson and Hooper (1989), White and Elson-Harris (1992) and Aluja and Norrbom (2000). As with most pestiferous fl ies, females within Bactrocera insert their eggs beneath the fruit skin, especially in ripening fruit; white banana-shaped eggs are usually deposited in clusters, hatching after 1.5–20 days (White and Elson-Harris, 1992; Messing, 1999). A single female can lay >1000 eggs over her lifetime (White and Elson-Harris, 1992). One generation requires c.37 days with a period of 19 days for egg to adult transformation; eggs hatch 38 h after ovi-position; larvae develop in 7–8 days and adults emerge in 10–11 days (Mess-ing, 1999). There are usually three larval instars. The larvae tunnel into the fruit, contaminating it with frass and providing entry for fungi and bacteria. Depending on factors such as temperature conditions and type of host, larval development can be completed in 7–8 days (Messing, 1999) but can take up to 2 weeks (White and Elson-Harris, 1992). When the infested fruit is imma-ture, the fruit ripens prematurely and is unfi t for marketing. Fully-grown larvae c.7 mm long drop to the ground and enter the soil where they pupate.

Pests 321
After emergence, the females require a protein source for egg maturation (White and Elson-Harris, 1992). Studies with B. dorsalis in India (Singh, 1991) indicated that pupal period was longest (18 days) at 15°C and shortest (6 days) at 35°C. Warm, humid weather is favourable for Bactrocera fruit fl ies and pest populations build up as mango ripening occurs. Bactrocera popula-tions decrease during dry periods.
Syed et al. (1970) reported that up to 30% of mango fruit were attacked by B. dorsalis in July and August. Mohyuddin and Mahmood (1993) reported that mango fruit are heavily attacked in Central Punjab during July and August, with up to 35% of the fruit being damaged by B. dorsalis and B. zonata. Vayssières et al. (2005) reported the presence of B. invadens after the fi rst sig-nifi cant rains in mid-April, reaching >900 males captured/trap/week. Trap captures peaked at 1800 fl ies/trap/week in mid-June when the presence of B. invadens was related to ripening of different mango cultivars. Ekesi et al. (2006) observed that B. invadens shared mango fruit with Ceratitis cosyra in Africa and suggested that B. invadens is predominantly a lowland pest.
CeratitisEight Ceratitis spp. have been reported to attack mango fruit. The Mediter-ranean fruit fl y C. capitata (Wiedemann) is a common polyphagous pest in mango-growing areas of Hawaii USA, Israel, Australia, Spain, Mexico, Réunion and Brazil and elsewhere in South America (Etienne, 1966; Morin, 1967; Galán-Saúco, 1990; Harris et al., 1993; Barbosa et al., 2005; Woods et al., 2005) (Plate 56). In Africa the most common species are C. cosyra (Walker), C. fasciventris (Bezzi), C. rosa (Karrsch), C. anonae (Graham) and less frequently C. capitata (Wiedemann) (Lux et al., 2003); whereas, C. catoirii Guer. occurs in Réunion (Etienne, 1968). Ceratitis quinaria and C. silvestrii are considered of economic importance in Benin (Vayssières et al., 2005). Ceratitis cosyra is broadly distributed across Africa and causes enormous damage, which can result in total loss of the crop. On average about 20–30% of mango produc-tion is lost due to this fl y species in various African countries (Lux et al., 2003).
BIOLOGY OF CERATITIS FRUIT FLIES. Flies within Ceratitis, particularly C. capitata, are quite cosmopolitan, and their basic life cycle varies greatly according to site (Papadopoulos et al., 1996). In Israel females seek suitable sites for ovipo-sition and puncture mango fruit early in the season, before the fruit has rip-ened. According to Wysoki et al. (1993) these ‘barren’ punctures damage the fruit, due to the leakage of resins from the fruit. The female can oviposit all over the fruit, with no preference for any part. Later, when fruit development is suitable for maggot development, the oviposition sites become light in colour and the tissue softens. The fully-grown maggots leave the fruit and pupate in the soil. The developmental period is c.3–4 weeks and 8–10 genera-tions/year can occur depending on temperature and other factors intrinsic to the fl y population (Hill, 1975).

J.E. Peña et al.322
Fruit fl y control: brief overview
The current trends in fruit fl y control call for coordinated, area-wide approaches (Hendrichs, 1996, 2001; Tan, 2000; Huang et al., 2006; Enkerlin, 2007; Orankanok et al., 2007) whose major objective is to overcome the often ineffective and environmentally unsustainable control schemes resulting from uncoordinated actions by individual producers. Aluja et al. (1996) pro-pose that since fruit fl ies that attack mango also attack other fruit crops in the same area, their management must be based on mango, wild hosts and other commercially grown host plants. Thus, to improve the effi ciency of fruit fl y management, host plant blooming and fruiting factors need to be elucidated. Hendrichs (1996) stated that when fruit growers pursue a concerted fruit fl y population management strategy over signifi cantly large areas, the number of fruit fl ies moving into orchards from neighbouring orchards is largely reduced. Aluja (1993, 1996) and Aluja et al. (1996) suggest a fruit fl y manage-ment scheme based on border trapping, enhancement of host-plant resistance through use of plant growth regulators, use of the sterile insect technique and bait stations and augmentative parasitoid releases (Sivinski, 1996; Sivin-ski et al., 1996; Malavasi and Zucchi, 2000; Montoya et al., 2000; Tan, 2000; Dyck et al., 2005; Mangan and Moreno, 2007). Aluja (1993) and Aluja and Liedo (1986) state that accomplishment of these goals depends on grower status (rich versus poor), access to technology, cost, scale (single orchard, regional level, national level), globalization of markets and local regulations restricting impact on the environment.
Monitoring and samplingMonitoring fruit fl ies attacking mango serves different purposes: (i) to apply a control or management tactic after the presence of the fruit fl y is noticed; and (ii) to verify if fruit fl y species will attack mango under natural condi-tions. In general, thresholds for adult fruit fl ies are quarantine-mediated (Beers et al., 1993). These thresholds vary from location to location, but de-pending on the fruit fl y species they are typically based on the capture of a single fruit fl y. In other fruit crops, a threshold of fi ve fl ies/trap is recom-mended resulting in a reduction from four chemical sprays to 1.5 sprays/season (Beers et al., 1993). Sampling for fruit fl ies in mango is mostly per-formed using adult traps, because eggs and young larvae are often diffi cult to see in the fruit and because the primary aim of management programmes is to prevent fruit damage.
In the case of pestiferous species within Anastrepha and some Bactrocera species, the most widely used traps since the early 1970s for monitoring and controlling populations are glass and plastic versions of the McPhail trap, which is baited with a mixture of protein (occasionally hydrolysed cotton seed together with borax, molasses or fermented juices) and water (Balock and Lopez, 1969; Jirón, 1995). More recently, human urine has been success-fully tested as bait for McPhail and McPhail-type traps for resource-poor farmers in tropical countries (Piñero et al., 2003; Aluja and Piñero, 2004). The McPhail trap has provided different results in mango orchards. Balock and

Pests 323
Lopez (1969) reported that high concentrations of McPhail traps reduced the build-up of fl y populations and protected mangoes from severe injury dur-ing certain periods of the year. However, the McPhail trap has several draw-backs. It is expensive, breaks easily, is cumbersome to service and, most importantly, is quite ineffi cient. Aluja et al. (1989), working in a mixed mango orchard in Chiapas, Mexico, found that only 31.1% of Anastrepha spp. fl ies landing on the McPhail trap were caught with many fl ies entering the trap but then escaping. Due to the low effi ciency of the McPhail trap it is being replaced with Multi-Lure® traps, which provide new trap designs. Dry synthetic-food-based lures have also been developed, i.e. BioLure® (Suterra LLC, Inc., Bend, Oregon) (Heath et al., 1995, 1997; Epsky et al., 1999) and Nu-Lure® (Advanced Pheromone Technologies) (Robacker and Warfi eld, 1993; Robacker et al., 1997; Robacker, 2001).
Fruit fl y presence has also been monitored in Australia using Dakpot® fruit fl y traps hung beneath the tree canopy (Anonymous, 1989). Methyl eugenol is considered the most powerful male lure for oriental fruit fl ies. Methyl eugenol was used for successful monitoring, control and erradication of B. dorsalis in Oahu Hawaii USA (Steiner and Lee, 1955), Rota Island (Steiner et al., 1965) and Okinawa, Kume, Miyako and Uaekama Islands (Japan) (Iwa-hashi, 1984). It has been used for monitoring B. umbrosa (F.) in the Philippines (Umeya and Hirao, 1975), and is used to lure B. invadens in Africa, which is unlike other African Dacini species that are attracted to Cue-lures (Lux et al., 2003; Anonymous, 2005). In Palau, Pacifi c Islands, two lures are used to attract mango fl ies: Bactrocera fraeunfeldi (Schiner) is attracted to Cue-lure, and B. occipitalis and B. philippinensis Drew and Hancock to methyl eugenol (Secretariat of the Pacifi c Community, 2005). Bactrocera dorsalis and B. umbrosa were monitored and controlled by mass trapping of males with methyl eugenol and infestations were brought to sub-economic levels in Pakistan (Mohyuddin and Mahmood, 1993). However, concern over the carcinogenic-ity of methyl eugenol (Waddell et al., 2004) calls for the development of other para-pheromones to attract Bactrocera fruit fl ies. Trimedlure is still consid-ered an important para-pheromone for the Mediterranean fruit fl y, with the exception of C. cosyra adults, which are attracted to terpinyl acetate and not to trimedlure (Steck, 2003). The attractiveness of mango compounds is currently being investigated. For example some of the volatiles emitted by ‘Tommy Atkins’ mangoes, i.e. terpenes (p-cymene and limonene), are attractive to C. capitata adults (Hernández-Sánchez et al., 2001).
Many questions linger with respect to the optimal trap number and time for trap placement in mango groves. In Naru, to produce mango free of B.frauenfeldi, 300–400 traps baited with methyl eugenol plus a toxicant were placed every square kilometre and trapping density was increased around mango plantings (Anonymous, 2002). Even though large numbers of traps can be utilized to increase detection sensitivity, the cumulative costs and logistical considerations do not make this option practical. Traps with spe-cifi c and effective lures that can detect the F1 generation at low trap densities (5–10 traps/km2) would fi t the description of a good detection and monitor-ing device (Tween, 1993).

J.E. Peña et al.324
Sampling for earlier fruit fl y stages can be used to demonstrate that the fruit is not susceptible to fruit fl y attack. For instance, the Caribbean fruit fl y, A. suspensa, may not attack green mango fruit (Peña et al., 2006b). Peña et al. (2006b) initiated research to determine if the Caribbean fruit fl y will attack green ‘Tommy Atkins’ mangoes and infest it under fi eld and forced labora-tory conditions. Through a sequential collection of fruit from fruit-fl y infested mango orchards, fruit were dissected for eggs and larvae. At the same time, fruit were stored and held for puparia emergence. In addition fruit were exposed in cages to wild fruit fl ies and traps were placed to verify the pres-ence of fruit fl ies. Estimating the time that ‘Tommy Atkins’ fruit remain immature and therefore non-hosts for fruit fl ies, may provide a window for mango exports in some fruit fl y-infested areas. Lux et al. (2003) also mention that small growers tend to harvest fruit before maturation as a strategy to evade fruit infestation.
Chemical controlMango plantations account for major insecticide use in the tropics (Cunning-ham, 1984). From the late 1960s to date, the conventional control of fruit fl ies was through toxic bait sprays that combine proteinaceous bait (e.g. hydroly-sed protein) with an insecticide (López et al., 1969; Soto-Manatiú et al., 1987; Mangan et al., 2006; Mangan and Moreno, 2007). For many years the insecti-cide of choice has been malathion (Peck and McQuate, 2000; Burns et al., 2001). Fruit fl ies are highly susceptible to any insecticide, and other com-pounds have also been widely used. For example, Singh (1991) reported that 5% aldrin dust, when mixed in soil, provided the highest residual toxicity to falling mature larvae (23.4% after 15 days), compared to BHC endosulfan and quinolphos. Vergherse et al. (2004), working on the control of B. dorsalis in India, used a rotation of fenthion (0.05%), deltamethrin (0.0028%), carbaryl (0.2%) and dimethoate (0.06%) to reduce the risk of resistance development. Yee (1987) concluded that weekly applications of malathion for 3 months can also provide effective control.
Since the late 1990s, there has been a concerted effort to fi nd environmen-tally friendly alternatives to malathion (Peck and McQuate, 2000). Cyromaz-ine, imidacloprid (organochlorinated compound), spinosad (bacteria-derived insecticide) and phototoxic dyes (Phloxine B) have been successfully tested against various fruit fl y species (Díaz-Fleischer et al., 1996; King and Hen-nessey, 1996; Peck and McQuate, 2000; Vargas et al., 2002; Liburd et al., 2004; McQuate et al., 2005). Despite their success, and as is typical with insecticides intensively applied on a large scale, resistance has already been documented in the case of spinosad (Wang et al., 2005; Hsu and Feng, 2006) or collateral damage (i.e. negative impact on natural enemies) (Stark et al., 2004). In Pakistan, application of pesticides caused reduction of fruit fl y infestations, but their use has created scale insect problems by eliminating their natural enemies (Mohyuddin and Mahmood, 1993). Such an effect had been reported by Ehler and Endicott (1984) with pests of olive, citrus and walnut. Another recent development with respect to chemical control of fruit fl ies has been the refi nement of the bait-station concept (Mangan and Moreno, 2007).

Pests 325
Without chemical control, the damage from C. capitata as a result of ‘bar-ren’ and fertile punctures can be as high as 60%. Control is achieved by aerial bait spraying, ground cover spraying and spot spraying of trees. In Spain, chemical control has been achieved by applying organophosphates and hydrolysed albumen (Khanzada and Naqvi, 1985), but this approach is under pressure because of environmental concerns. Decisions on when to apply insecticides are based on the appearance of the fi rst trapped males (trimedlure-baited traps are used). When applying insecticides directly, dimethoate (0.1%) and fenthion (0.15%) are used (Khanzada and Naqvi, 1985). Bait sprays are based on naziman (1:1 protein hydrolysate: malathion/4 l water; Wysoki et al., 1993). Removal of fallen fruit can also prevent build-up of Mediterranean fruit fl y populations.
Pestiferous Anastrepha spp. are susceptible to most insecticides (Shaw and Spisshakoff, 1958; Shaw, 1961). Bait sprays applied from the ground and from the air are successful, but they can cause environmental damage, surges of secondary pest populations and reductions in parasitoid populations (López et al., 1969; Soto-Manitiu et al., 1987). In Peru, control measures against Anastrepha in mango begin when McPhail trap catches average two adults/trap/week (Herrera and Viñas, 1977). In Mexico, control starts when the fruit is 85-days-old and is suspended 2 weeks before harvest (Cabrera et al., 1993). In Costa Rica, dipterex and malathion are sprayed weekly and reduce mango damage up to 40% (Soto-Manitiu et al., 1987). In Brazil, malathion with protein and sugarcane bagasse is used (Carvalho and De Queiroz, 2002). In Ecuador, Arias and Jines (2004) recommend a spray of malathion (1%) with protein (4%) once the fruit fl y population reaches 0.14 fruit fl ies/trap/day (FTD).
Biological controlPARASITOIDS. Classical biological control and repeated augmentative releases of mass-reared parasitoids have been used to suppress Anastrepha, Ceratitis and Bactrocera populations (Wharton, 1978; Sivinski, 1996; Sivinski et al., 1996, 1997, 2000; Montoya et al., 2000). In Florida USA, Mexico, Costa Rica, Brazil, Colombia and Peru, parasitoid species (i.e. Diachasmimorpha longicau-data (Ashmead), Fopius vandenboschi (Fullaway) and Aceratoneuromyia indica (Silvestri)) have been imported and released for the control of A. suspensa, A.ludens and A. fraterculus (Ovruski et al., 2000). Despite the widespread use of exotic parasitoids over the past 80–100 years, the current trend is to resort to native species as they pose less of an environmental threat to local fauna (García-Medel et al., 2007; Aluja et al., 2009).
Use of parasitoids with mango is hindered by the fact that fruit are very large and therefore provide larvae a refuge from parasitism (López et al., 1999). As a consequence, Aluja (1993) and Montoya et al. (2000) recommended that parasitoids should be released outside the mango orchards to attack fl y larvae in their much smaller native hosts and thereby signifi cantly reduce the size of populations entering mango orchards.
Several parasitoids, for example Opius fullawayi (= Diachasmimorpha fullawayi (Silvestri)), Diachasmimorpha kraussi, D. Fullaway, D. tryoni (Cam-eron), Opius bellus Gahan, Biosteres longicaudatus Ashmead (= D. longicaudata),

J.E. Peña et al.326
B. tryoni (Couron) (= D. tryoni (Cameron)) and Biosteres oophilus Fullaway (= Fopius arisanus (Sonan)), parasitize C. capitata (Beardsley, 1961; Wharton and Marsh, 1978). Bess et al. (1961) reported that the most important parasi-toids collected from C. capitata in Hawaii USA were F. vandenboschi, B. oophilus (= Opius oophilus) (= F. arisanus) and B. longicaudatus. In Brazil, mainly Doryc-tobracon areolatus (Szépligeti) (97%) and D. longicaudata (3%) parasitize fruit fl y larvae attacking mango (Carvalho and De Queiroz, 2002). In Kenya, Ghana, Tanzania, Uganda and Cote d’Ivoire, the most important parasitoids obtained from Ceratitis spp. emerging from mangoes were D. fullawayi, Fopius cauda-tus (Szépligeti), Psyttalia cosyrae (Wilkinson) and Tetrastichus giffardianus Sil-vestri (Lux et al., 2003). In Mexico and other parts of Latin America, the most common parasitoids attacking fruit fl ies that infest mangoes (A. obliqua, A.ludens, A. pseudoparallela, A. turpiniae) are D. areolatus, Doryctobracon brasilien-sis (Szépligeti), Doryctobracon crawfordi (Viereck), Doryctobracon fl uminensis (Lima) and Utetes anastrephae (Viereck) (López et al., 1999; Ovruski et al., 2000; Zucchi, 2000). In Pakistan, the parasitoids attacking B. zonata include Opiuslongicaudatus (= D. longicaudata), Dirhinus giffardii Silvestri, and Bracon sp.; O.longicaudatus (= D. longicaudata), D. giffardii and Spalangia grotiusi Girault were reported to attack B. dorsalis, albeit in small numbers (Syed et al., 1970).
Microbial controlUse of pathogens/disease agents (fungi, bacteria, nematodes) has been attempted with varying degrees of success. For example, Metarhizium anisopliae has been evaluated in small-scale mango orchards in Kenya using bait stations laced with the pathogen. Results do not show differences between use of pathogens and use of insecticides (malathion) (Lux et al., 2003). Lezama-Gutierrez et al. (2000) also evaluated isolates of M. anisopliae against larvae of A. ludens. They suggested that M. anisopliae can cause a 22–43% reduction in adult emergence, depending on the soil where the lar-vae pupariates. De la Rosa et al. (2002) evaluated the fungus Beauveria bassi-ana (Bals.) under laboratory conditions and concluded that highest mortality was achieved at the adult stage, while Dimbi et al. (2003) reported on the pathogenicity of M. anisopliae and B. bassiana on different species of Ceratitis. Poinar and Hislop (1981), Lindegren and Vail (1986) and Toledo et al. (2006) have investigated the use of various nematodes, Heterorhabditis bacteriophora, Heterorhabditis heliothidis (Khan, Brooks and Hirschmann) and Steinernema feltiae Filipjev, against Anastrepha, Bactrocera and Ceratitis. Finally, Robacker et al. (1996) and Toledo et al. (1999) have tested various strains/isolates of Bacillus thuringiensis (Berliner) against larvae of A. ludens, A. obliqua and A. serpentina. For additional details on microbial control of pestiferous fruit fl ies, we recommend the recent review by Dolinski and Lacey (2007).
PredatorsIn addition to parasitoids, pathogens and nematodes, ants have been used to control fruit fl ies in mango orchards. Peng and Christian (2006) used the weaver ant, Oecophylla smaragdina (Fabricius) for control of the Jarvis fruit fl y, B. jarvisi, in mango orchards in Australia. Van Mele et al. (2007 and references

Pests 327
therein) in Benin used an African weaver ant (Oecophylla longinoda). Aluja and colleagues (Aluja et al., 2005) investigated the potential of ants as possi-ble biological control agents in various tropical orchards and backyard gar-dens in which mango trees were growing with other fruit trees.
Cultural fruit fl y controlFruit bagging is one of the best solutions to prevent fruit fl y attack of mango and other tropical fruits (Aluja, 1996; Peña et al., 1999; Paderes and Orden, 2004). Success with mangoes can be quite high, but Bondad (1985) demon-strated that bagging materials do not always resist the effect of rain/wind. Therefore, while bagged mangoes tend to produce a greater amount of mar-ketable fruit than those not bagged, more research is needed to determine the type of bags to use for different mango varieties and the best time to bag fruit (Love et al., 2003).
Jirón (1995) reported that A. obliqua populations could be reduced by increasing planting distances in order to reduce RH and increase solar radia-tion within orchards. In India, cultural control practices include removal of fallen fruit and inter-tree ploughing and raking (followed by insecticide cover sprays). Such practices can reduce fruit fl y infestation between 77% and 100% (Verghese et al., 2004).
A cultural practice that can impinge on the success of management pro-grammes is the widespread use of potassium nitrate (KNO3) sprays to accel-erate and synchronize fl owering of mango trees. As a result, fruit harvests can be advanced and synchronized. Such a procedure can, under certain cir-cumstances, help control fruit fl ies, but can also exacerbate the problem. For example, in the case of the Mexican fruit fl y (A. ludens), a notorius pest of citrus that also attacks mangoes, advancing the mango harvest offers ideal conditions for adult pests to move from citrus groves to mango orchards. This would not occur if mango trees fl owered naturally since fruit ripen sev-eral months after the citrus harvest (Martin Aluja, personal observation).
Host resistanceYee (1987) reported that B. dorsalis does not attack all mango cultivars to the same extent. The most susceptible cultivars in Hawaii USA are ‘Hawaiian’, ‘Pirie’ and ‘Sandersha’. Singh (1991) indicated that the frequency of Bactro-cera injury to physiologically mature fruit of ‘Dashehari’ ranged from 3.6 to 10%, while in fully ripe fruit the frequency of injured fruit ranged from 10 to 25.9%. Highest damage was reported in fully ripe fruit of ‘Mallika’ followed by ‘Totapari’.
Susceptibility of different mango cultivars to attack by A. obliqua was measured by Carvalho et al. (1996) who observed that ‘Espada’ showed no infestation by A. obliqua, whereas ‘Carlota’ was highly infested. In this study, the survival of adults of A. obliqua was lower when the larvae were fed on ‘Espada’ compared to ‘Carlota’. Furthermore, ‘Espada’ had an adverse effect on the longevity of A. obliqua females, possibly due to the presence of toxic substances (Carvalho and De Queiroz, 2002) or absence of essential nutri-ents. Jirón and Soto-Manitiu (1987) also observed that susceptibility of

J.E. Peña et al.328
mangoes to A. obliqua differed among cultivars. ‘Rosinha’, ‘Coquinho’ and ‘Espada’ were resistant to A. obliqua attack, whereas ‘Smith’ and ‘Pope’ were highly susceptible. According to Joel (1980), mangoes contain resin ducts in the exocarp that confer protection against the vertical movement of the ovi-positor and larval movement. Other studies have shown that resistance is related to degree of maturity (Díaz-Fleischer and Aluja, 2003; Aluja and Mangan, 2008); immature mango fruit are less susceptible to A. suspensa than mature mangoes when infested artifi cially (Hennesey and Schnell, 2001). In Brazil, use of gibberellic acid (GA3) reduces the susceptibility of ‘Tommy Atkins’ mangoes to attack by C. capitata based on artifi cial delay of fruit maturity (de Macedo, 1988). Differences on attack by A. ludens to mango might be infl uenced by volatiles from green or yellow fruits (Garcia-Ramirez et al., 2004).
Quarantine treatmentsQuarantine treatments have been reviewed by Johnston and Hofman (Chap-ter 15, this volume). Several quarantine treatments have been developed for harvested mangoes: irradiation, hot water or hot water followed by immer-sion cooling are widely used (Sharp et al., 1988, 1989a, b, c; Hallman and Sharp, 1990; Nascimento et al., 1992; Mangan and Sharp, 1994; Mangan and Hallman, 1998; Shellie and Mangan, 2002a, b; Bustos et al., 2004; additional references in reviews by Mangan and Hallman, 1998 and Follet and Neven, 2006).
Mango seed weevils
The mango seed weevil, Sternochetus mangifereae (Fabricius), and the mango pulp weevil, Sternochetus frigidus (Fabricius) (Coleoptera: Curculionidae) are important pests of mango (Plates 57–59). Quarantine restrictions prevent the export of fresh weevil-infested mangoes into uninfested areas. The fl esh of ripe fruit is damaged when mango seed weevil adults emerge from the seed, and weevil-damaged seed may limit plant propagation in nurseries and orchards (Johnson, 1989). Early fruit drop may be caused by severe weevil infestations (Subramanian, 1925). The mango seed weevil occurs from India through South-east Asia to Australia, on tropical Pacifi c Islands, in parts of Africa, in the Caribbean region and in northern South America (Balock and Kozuma, 1964; Shukla and Tandon, 1985; Johnson, 1989; Schotman, 1989).
BiologySrivastava (1998) reports that S. mangiferae is a greyish brown weevil, 8 mm long and about 4 mm wide; its habits are nocturnal, usually feeding and ovi-positing at dusk. After emergence, adults enter diapause and the duration varies with the geographic range (Schotman, 1989). According to Shukla and Tandon (1985), all adults emerging in southern India during June enter dia-pause from July until late February in the following year. The beginning and the end of diapause seem to be associated with long-day and short-day

Pests 329
photoperiods, respectively (Balock and Kozuma, 1964). The mango seed weevil produces only a single generation each year. In Tamil Nadu, India, adults feed on leaves and tender mango shoots in March and April (Subra-manian, 1925). Shukla and Tandon (1985) report that females began oviposit-ing 3–4 days after mating when fruit reaches a marble-size. Oviposition varied from 3 to 5 weeks (Subramanian, 1925; Shukla and Tandon, 1985; Hansen et al., 1989). The female uses its snout to make a cavity in the fruit, lays a single egg and then covers it with a secretion (Pradhan, 1969).
According to Srivastava (1998), about six larvae can be found within a mango seed. Generally, only a single larva completes development within each fruit. Larval development and pupation occurs within the seed. Adults are formed 1 week later; however, adults generally emerge from the seed c.1–2 months after fruit drop (Balock and Kozuma, 1964). The weevils over-season under bark and stone walls, where they remain dormant until the next fl owering season (Van Dine, 1906; Balock and Kozuma, 1964; Shukla and Tandon, 1985).
Sternochetus frigidus attacks Mangifera indica, M. foetida and M. sylvatica. The weevil occurs in Bangladesh, Brunei, India, Indonesia, Myanmar, Paki-stan, Papua New Guinea, the Philippines, Singapore and Thailand (CAB International, 2003). Sternochetus frigidus lays eggs on mango fruit with a minimum diameter of 6 cm (De and Pande, 1988). Newly hatched larvae tun-nel directly through the fruit pulp. Pupation takes place in a brown cocoon within a chamber adjacent to the kernel. The weevils leave the ripe fruit through a hole in the peel. This pest is likely to survive storage and transpor-tation (Australian Government, 2004). Reproductively immature adult wee-vils overwinter inside seed or other protective places from May until February in India (De and Pande, 1988).
SamplingShukla et al. (1988) reported the intra-tree distribution of eggs of S. mangiferae on ‘Baganpalli’ mango; the highest number of eggs per fruit occurred on fruit in the lower region of the tree. With increasing tree height, egg deposition on fruit decreases. No statistical differences on fruit infestation were observed on north, south, east or west directional quadrats of the tree. Eggs were deposited in the lower region of the fruit rather than the pedicel. Weevils enter diapause in crevices in the tree trunk. Most of the weevils (87%) are at a height of 0–2 m in the trunk compared to 7% at 2–4 m and only 4% above 4 m. Emery (2002) considered that since both the mango pulp weevil (S. frigi-dus) and the mango seed weevil (S. mangiferae) infest fruit at an early stage, any fruit is a viable sample; however, as infested fruit all ripen precociously, the sensitivity of surveys is enhanced by seeking out nearly ripe and fallen fruit prior to harvest. If the survey coincides with the mango harvest, rejected or fallen fruit should be inspected. Fruit should be sampled by longitudinal dissection of fruit through the seed to expose the kernel. If the fruit is ripe, it should be struck along the longitudinal axis with a hammer, and the seed should be opened with pliers. The random sampling of 600 fruit from ran-domly selected properties in each area provides a 9% chance of detecting a

J.E. Peña et al.330
0.5% infestation of fruit. Sample size can be determined from the following formula:
Probability of > 1 infested fruit = 1 – probability of no infected fruit in total sample = 1 – (1 – 0.5%)600 = 1 – (1 – 0.005)600 = 1 – (0.995)600 = 95%
Economic damageFollett and Gabbard (2000) report that germination rates for infested seed of polyembryonic ‘Common’ are equal to those of uninfested seed. Germina-tion is signifi cantly reduced for infested seed of monoembryonic ‘Haden’ compared with uninfested control seed, although germination of infested seed was >70%. Direct feeding damage to the pulp was found in only 0.11% of 3602 mango fruit, which suggests that S. mangiferae is a less serious pest of mangoes than previously considered.
Cultural controlField sanitation, i.e. the removal of all fallen fruit and seed, is very labour intensive, and demands complete removal and disposal of fallen fruit. This procedure has been inconsistent in demonstrating pest control. In India, fi eld sanitation reduced infestation of the mango nut weevil, Sternochetus gravis(Fabricius), by only 22% (De and Pande, 1987). In Hawaii USA, fi eld sanita-tion failed to reduce infestation rates (Hansen and Amstrong, 1990).
Chemical controlVarious insecticides have been evaluated for controlling adult weevils, par-ticularly during oviposition (Balock and Kozuma, 1964; Shukla and Tandon, 1985). The most effective control was provided by the organophosphate fen-thion, which reduced infestation to <17%. In another fi eld test, the pyrethroid deltamethrin and the carbamate carbaryl were most effective, both resulting in <15% infestation rates. Spot application of diazinon on tree trunks was recommended based on cost, effi ciency and least environmental damage. Verghese et al. (2004) reported that commercially available azadirachtin was not effective for management of S. mangiferae in India.
Resistant cultivarsMango cultivars resistant to the mango weevil would be benefi cial. Potential mechanisms of resistance are seedless cultivars, those that form seed early or those that fruit off-season. Most cultivars grown in Hawaii and India are equally susceptible (Bagle and Prasad, 1984; Hansen et al., 1989), although ‘Itamaraca’ has shown some resistance (Balock and Kozuma, 1964).
Biological controlThe mango weevil has few natural enemies. Parasitoids are unknown, prob-ably because of the concealed nature of most life stages. Adults may be sus-ceptible to predation by ants, rodents, lizards and birds (Hansen, 1993). A baculovirus has been reported that affects the larvae of S. mangiferae (Shukla et al., 1984).

Pests 331
Mango fruit infested with seed weevil do not show any visible external symptoms and cause considerable quality control problems and economic loss to the mango-processing industry as well as restriction in export of fresh fruit. A non-destructive X-ray inspection method has been developed to detect weevil-infested fruit. X-ray radiographs of infested mangoes show dark areas in the seed corresponding to disintegrated kernel tissue as a con-sequence of feeding by developing grubs. Non-infested mangoes show a uni-formly light-grey area representing healthy kernel. There is a close agreement between fruit showing weevil infestation based on their X-ray images and physical examination of cut fruit, indicating the reliability of the technique. X-ray imaging has good potential for application in the processing industry and the export trade as a quality control measure (Thomas et al., 1995).
Mango seed borer (Lepidoptera: Pyralidae)
Distribution and biologyThe red banded mango caterpillar or mango seed borer, Deanolis sublimbalis Snellen, also referred to as Noorda albizonalis Hampson (Waterhouse, 1998), is an important pest of mangoes in the Philippines (Anonymous, 1984), India (Zaheruddeen and Sujatha, 1993), Vietnam (Nguyen et al., 1998; van Mele et al., 2001), China (Li et al., 1997), Thailand, Indonesia and Papua New Guinea (Cunningham, 1984). The oval white eggs are laid in groups at the fruit apex and take 3–4 days to hatch. The larvae develop through fi ve instars in 14–20 days and they pupate in cocoons in the soil. The period from egg to adult requires 28–40 days. The insect apparently prefers mango, but M. odorata and M. minor have also been recorded as hosts. Adults are generally nocturnal and spend most of the day under leaves on the tree. A shorter developmental period has been observed when larvae develop on pulp rather than in seed of ‘Carabao’ fruit, although those reared on the seed were larger and lived longer (Waterhouse, 1998).
DamageMango fruit in all stages of development are susceptible to attack (Water-house, 1998). The fi rst larval instars feed on tissues beneath the skin, and bore through the mango pulp to the seed, which is consumed. Up to 11 larvae have been recorded in a single fruit, but usually there is only one. Infested fruit split and rot, and fall to the ground (Anonymous, 1984). In the Guimaras Islands, the Philippines, Golez (1991) recorded 12.5% fruit infestation and in serious outbreak years, 40–50% yield reductions are possible. Waterhouse (1998) considered that since D. sublimbalis is capable of causing such levels of damage, it might be a more important pest of mangoes than has generally been realized. It may have been overlooked as a pest or has recently spread to new areas and has become evident as a pest there. Van Mele et al. (2001) suggested that damage caused by D. sublimbalis in the Mekong Delta has been wrongly attributed to fruit fl ies; however, Waterhouse (1998) states that soon after boring by D. sublimbalis, secondary infestations with fruit fl ies

J.E. Peña et al.332
(Bactrocera ferrugineous, B. frauenfeldi) occur, together with infections by bac-teria and fungi.
Biological controlAccording to Waterhouse (1998) no parasitoids were detected in Java, Indo-nesia. However, in the Guimaras Islands of the Philippines, the vespid wasp, Rychium attrisimum, preys on the larvae as they exit the fruit to pupate. Lar-vae are used to stock the wasps’ nests as food for their young. The egg para-sitoids Trichogramma chilonis Ishii and Trichogramma chilotreae attack the pest in Luzon (Golez, 1991). Leefmans and van der Vecht (1930) reported that an entomopathogenic fungus infected the larvae in Indonesia.
Monitoring and controlInfested fruit can be detected by the presence of a dark-brown ring and cat-erpillar frass at the point of entry (CAB International, 2003). Mango fruit become susceptible to the seed borer c.60 days post-induction, and insecti-cide applications should commence then. Further sprays at 75, 90 and 105 days post-induction are required to fully protect the fruit. The most effective chemicals are deltamethrin and cyfl uthrin (Golez, 1991). Infested fruit should be removed from trees before the larvae can leave them to attack neighbour-ing fruit; fruit should be wrapped in protective bags at 55–65 days after pol-lination, and fallen fruit should be destroyed (Anonymous, 1984).
Other Lepidoptera that can attack fruit have been reported in India and the Philippines (Australian Government, 2004). The pomegranate fruit borer, Deudores isocrates (Fabricius) (Lepidoptera: Lycaenidae), which is also a pest of loquat, lychee, guava and pear, could attack mango by laying single eggs on shoots; the emerging larva bore into the fruit (Srivastava, 1998). In the Philippines, the larvae of the cocoa tussock moth, Orgyia postica (Walker) (Lepidoptera: Lymantriidae), regularly attack leaves of cocoa, but its larva also attack mango fruit and panicles (Fasih et al., 1989).
Fruitspotting bugs (Hemiptera; Coreidae)
The yellowish green coreid bugs, Amblypelta lutescens lutescens (Distant) and Amblypelta nitida Stål occur along the coast of Queensland, Australia, and attack most of the tropical and subtropical fruit crops there (Waite and Huwer, 1998). They prefer to feed on young, green fruit, but A. l. lutescens also dam-ages the terminals of a number of hosts. In tropical north Queensland, A. l. lutescens is the dominant species and feeds on young fruit causing black lesions to develop and the fruit to fall. It also feeds on the terminals and leaf petioles, causing wilting and dieback (Cunningham, 1989). In the subtropical south, both species attack mango, but A. nitida confi nes its attention to the fruit, while A. l. lutescens also attacks fruit and terminal growth (G.K. Waite, 1995, unpublished results). The bugs breed in natural rainforest areas, and fl y into the orchards to feed on the fruit and terminals. Female bugs lay individual, opalescent green eggs under the leaves. There are fi ve nymphal instars and a generation takes c.40 days.

Pests 333
The main predators of fruitspotting bugs are spiders, particularly mem-bers of the family Thomisidae. Several species of egg parasitoids have been recorded. In north Queensland, Ooencyrtus sp. (Encyrtidae), Anastatus sp. (Eupelmidae) and Gryon sp. (Scelionidae) parasitized 37.5–91.6% of eggs of A. l. lutescens (Fay and Huwer, 1994). In south Queensland, Anastatus sp. and Gryon meridianum (Dodd) parasitize eggs of A. nitida and A. l. lutescens to a similar extent (Waite and Petzl, 1994).
Because fruitspotting bugs continuously migrate into orchards, more than one insecticide spray may be required to protect the young fruit. How-ever, the fruit are safe from attack once they are c.50 mm long, and two or three sprays of endosulfan at intervals of 2 weeks are generally suffi cient to protect them.
The coconut bug, Pseudotheraptus wayi Brown, was fi rst recorded on man-goes in South Africa in 1977, and now also attacks guavas, pecans, macada-mias, avocados and loquats. It causes damage similar to that of Amblypelta spp. (De Villiers, 1990).
Helopeltis spp. (Miridae) are minor pests of mango, cashew and cacao in the Philippines and in northern Australia, where they feed on fruit and cause superfi cial corky blemishes. Insecticides are used to control them, but in the Philippines, bagging is also effective (Anonymous, 1984).
Plant bugs within the Lygaeidae and Pyrrhocoridae, i.e. the Indian milk-weed bug, Spilostethus pandurus (Scopoli) and the red cotton bug, Dysdercus koenigri (Fabricius), can injure fruit, infl orescences and leaves of mangoes in India (Australian Government, 2004). However, D. koenigri is an important pest of cotton rather than mango (Schaefer and Ahmad, 2000), while S. pan-durus feeds preferentially on members of the Asclepediaceae (i.e. Calotropis) (Sweet, 2000).
A hymenopterous parasitoid complex attacking the eggs of the banana-spotting bug, A. l. lutescens, was fi rst reported in north Queensland. It includes an Anastatus sp. (Eupelmidae), Ooencyrtus sp. nov. (Encyrtidae) and a Gryon sp. (Scelionidae), and is similar to other complexes known to attack eggs of related coreids in Africa, Indonesia and Papua New Guinea. Parasitism ranged from 37.5–91.6% in eggs collected at three sites from orange jessa-mine, Murraya paniculata (L.) Jack. Anastatus sp. was the dominant parasitoid (Fay and Huwer, 1994).
Thrips
Grove et al. (2001) reported that in South Africa, the citrus thrips, Scirtothripsaurantii Faure and the red banded thrips Selenothrips rubrocinctus (Giard) are the only thrips that caused lesions on fruit (Plates 60–63). However, Grove et al. (2001) considered S. aurantii to have more economic importance than S.rubrocinctus. Grove and Pringle (2000), using a two-stage sampling system to determine population levels of S. aurantii, showed that S. aurantii has a clumped distribution, and recommended that 50 fruit per orchard should be examined in order to obtain accurate population estimates for pest management.

J.E. Peña et al.334
Blossom pests
Midges, caterpillars, hoppers, thrips and mites are the most important pests attacking mango infl orescences.
MidgesThe mango gall midge or mango blister midge, Erosomya mangiferae Felt, is a major pest, destroying fl owers and up to 70% of set fruit (Plate 64). It was fi rst described by Felt (1911) in St Vincent (West Indies). Barnes (1948) recog-nized nine gall midges from mango; two of these, Asynapta sp. and E. man-giferae, are from the West Indies. Butani (1979) reported fi ve cecidomyiid species on mango blossoms, including Erosomya indica (Grover and Prasad). Dasyneura mangiferae (Felt) was reported in Hawaii USA (Vannière et al., 2004). In recent times, Gagne and Etienne (2006) reported the species Gephy-raulus mangiferae (Felt), n.comb. infesting mango fl owers on the island of Guadeloupe, French West Indies. Male adults of E. mangiferae are 1.61 mm and females 1.32 mm long. Eggs are deposited in folds between sepals and petals of fl ower buds. The larval stage has four instars. Young larvae are cream coloured and late instar larvae are yellowish. Larval feeding prevents fl ower opening and consequently development of the fruit does not occur. Infested buds develop as long pointed galls, in which pupation occurs (Van-nière et al., 2004). Studies of population fl uctuation of Erosomya sp. have been conducted in India by Grover (1986a), who reported that emergence of adults was higher at 24°C and 60–82% RH compared to lower temperatures and RH. Abbas (1985) described systematic surveys to determine the percentage of infestation of E. indica, and showed that infestation follows a negative binomial infestation. The midge infests the newly emerged panicles by ovi-positing at bud burst stage, and the fi rst instar maggots bore into the grow-ing panicle. The second generation then infests very young fruit, which drop before the marble stage. Sampling of mango midges needs to include affected tissue, different trapping devices, pheromones, etc. On citrus, use of coloured sticky traps placed in the tree canopy provides a more effi cient method of sampling the citrus midge, Prodiplosis longifi la Cagné, than ground emergence traps and collection of larval samples (Peña and Duncan, 1992).
In a survey of parasitoids of cecidomyiid pests of mango in India, Grover (1986b) reported that Platygaster sp., Systasis sp. and Eupelmus sp. were asso-ciated with Dasineura sp., and Tetrastychus sp. was associated with E. indica. An external parasitoid, the pteromalid, Pirene sp., attacked Procystiphora mangiferae (Felt). Predators of the cecidomyiids include Formicai sp., Oeco-phila sp. and Camponotus sp.
Mango hoppersApproximately 18 species of leaf hoppers have been reported as pests of mango. Of these, Idioscopus clypealis Leth., Idioscopus niveosparsus Leth., Idiosco-pus magpurensis Pruthi and Amritodus atkinsoni Leth., are important (Virakta-math and Viraktamath, 1985; Viraktamath, 1997; Fletcher and Dangerfi eld, 2002) (Plate 65). The females deposit their eggs in panicles or midribs of tender

Pests 335
leaves. The adults and nymphs preferentially feed on young leaves and fl ow-ers or shoots, and excrete honeydew upon which sooty mould develops (Ahmed et al., 1981). This interferes with photosynthesis, adversely affecting plant growth and yield (Godase et al., 2004). Affected infl orescences turn brown, become dehydrated and fruit set does not occur.
There has been no systematic study of the biology of most of the leaf hoppers that attack mango; however, biology of A. atkinsoni, I. clypealis and I.niveosparus has been studied by Sohi and Sohi (1990). Both A. atkinsoni and I.niveosparsus are multivoltine. In A. atkinsoni, the egg, nymphal (fi ve instars) and adult stages require 7–9, 15–17 and 3–4 days, respectively (Patel et al., 1977). Development from egg to adult is normally complete in 25–30 days. There can be between one and six generations of A. atkinsoni in different areas of India. In Pakistan there are four to fi ve generations in the Central Punjab (Mohyuddin, unpublished data). In the Philippines, I. clypealis is reported to have one to four generations, whereas it has fi ve or six generations in India.
Idioscopus nagpurensis is univoltine. In Pakistan, it normally oviposits in mango infl orescences during March. Nymphs feed on infl orescences during March and April. From May to February of the following year, only aestivat-ing adults are found (Mohyuddin, unpublished data). Most of these species are quite fecund. Amritodus atkinsoni reportedly lays 200 eggs during its life-time (Rahman, 1940) and I. clypealis lays 100–190 eggs in the Punjab (Husain and Pruthi, 1924). Amritodus atkinsoni eggs are laid in the midribs of tender leaves, fl ower buds and infl orescences (Babu et al., 2002). Idioscopus niveospar-sus lays c.238 eggs in 9 weeks under laboratory conditions (Mohyuddin, unpublished data). Mohyuddin and Mahmood (1993) reported that A. atkin-soni and I. niveosparsus occur in upper portions of mango trees during differ-ent times of the year. Amritodus brevistilus and I. niveosparus populationsincrease from February to peak in March–April in Sri Lanka, while peaks of I. clypealis occur in March and September (Kudagamage et al., 2001). Idiosco-pus clypealis populations peak in south-eastern India during March and April (Tandon et al., 1983). Idioscopus nivesoparus and I. clypealis peaks coincided with major and minor fl owering periods while population peaks for A. bre-vistilus coincide with the occurrence of vegetative fl ushing (Kudagamage et al., 2001).
Azizur Rahman and Singh (2004) demonstrated that A. atkinsoni popula-tions on panicles of ‘Langra’ mango were negatively correlated with high RH; whereas no signifi cant relationships were observed with rainfall, sun-shine and wind velocity.
SAMPLING. Very few sampling studies have been reported for hoppers on mango. A sequential sampling plan for mango hoppers was recommended by Verghese et al. (1985) in India. Mohyuddin and Mahmood (1993) reported sampling by direct visual examination: A. atkinsoni and I. niveosparsus were found on upper portions of mango trees during different times of the year. They moved to the lower parts of the stems and the leaves during summer. Tandon et al. (1989) reported that distribution of I. niveosparsus was aggre-gated and was best explained by Iwao’s patchiness regression. To assess

J.E. Peña et al.336
damage, they recommended a sampling size of 59–98 panicles/tree. Verghese et al. (1985) developed a sequential sampling plan classifying infestations of adults and nymphs of I. clypealis as light, moderate and severe.
BIOLOGICAL CONTROL. Several natural enemies have been described from west and South-east Asia. Mohyuddin and Mahmood (1993) reported the egg par-asitoids, Gonatocentrus sp., Miurfens sp. nr. mangiferae Viggiani and Hayat, Centrodora sp. nr. scolypopae Valentine, Aprostocetus sp. and Quadrastichus sp., and the adult ectoparasitoid Epipyrops fuliginosa Tames in Pakistan. Fasih and Srivastava (1990) reported that Aprostocetus sp., Gonatocerus sp. and Polynema sp. parasitize eggs. Five species of predators, including Chrysopa lacciperda (Kimmins), Mallada boninensis (Okomote), Bochartia sp. and two unindenti-fi ed species (one each of Mantidae and Lygaeidae) prey on nymphs (Fasih and Srivastava, 1990). In India, Sadana and Kumari (1991) studied the effi -cacy of the lyssomanid spider, Lyssomanes sikkimensis on I. clypealis. Classical biological control of mango hoppers has not been attempted. Whitwell (1993) described four genera of parasitoids from Dominica, the most common being Aprostocetus sp., followed by Platygaster sp., Synopeas sp. and Zatropis sp. Peng and Christian (2005a, b) reported that the weaver ant, Oecophylla smarag-dina (Hymenoptera: Formicidae) is an effi cient biocontrol agent of I. nididulus in northern Australia. The entomopathogens, Verticillium lecanii (Zimmer-man) Viegas, Beauveria bassiana Balsamo (Vuillemin) and Isaria tax, infect I. clypealis in India (Kumar et al., 1993; Srivastava and Tandon, 1986) while the effectiveness of Metarhizium anisopliae var. anisopliae was tested under laboratory conditions against A. atkinsoni (Vyas et al., 1993).
CHEMICAL CONTROL. Several pesticides have been tried for controlling mango hoppers (Tandon and Lai, 1979; Yazdani and Mehto, 1980; Shah et al., 1983; Shukla and Prassad, 1984; Islam and Elegio, 1997; Kudagamage et al., 2001). Khanzada and Naqvi (1985) reported that six sprays of fenitrothion/year were effective for controlling mango hoppers in Pakistan. Nachiappan and Baskaran (1986) tested eight insecticides: phasalone, endosulfan, carbaryl, penthoate, fenitrothion, monocrotophos, quinalphos and phosphamidom. Endosulfan provided the best control when spraying was done 1 week after fl owering and then 14 days later. Mohyuddin and Mahmood (1993) reported that monitor applied at 5 m on tree trunks and leaves in May provided control of mango hoppers. Jhala et al. (1989) considered that sprays of carbaryl during the off-season maintained the hopper population at low-density levels.
Godase et al. (2004) demonstrated that sprays of 0.05% monocrotophos at the fi rst panicle emergence and a second spray 15 days later are essential to prevent yield loss. Kudagamage et al. (2001) found that imidacloprid (Admire SL 200) controlled mango hoppers if applied just after fl owering and again 10 days later.
LepidopteraThe lepidopteran fl ower feeders are the second most important infl orescence pests of mango. Geometrids, for example Chloropteryx glauciptera Hampson and

Pests 337
Oxydia vesulia (Cramer) infestation was reported in Dominica by Whitwell (1993). Infestations increase during the fl owering season, averaging three larvae/infl orescence (87% infestation) to c.100% infestation later in the fl ow-ering season. Eggs of the noctuidae Penicillaria jocosatrix Gueneé are laid pre-dominantly on or near the infl orescences or new leaves. The microlepidoptera complex attacking mango in Florida USA consists of Pococera attramentalis Lederer, Pleuroprucha insulsaria (Gueneé) (Plate 66), Platynota rostrana (Walker), Tallula spp. and Racheospila gerularia (Hübner) (Plate 67). Most of the damage to infl orescences (Plate 68) is caused by P. attramentalis and P. rostrana. Pococera attramentalis and P. attramentalis are also common pests of sorghum (Kring et al., 1987) and other tropical fruit trees. The larvae of both species feed on the axis of the infl orescence, petals and ovaries late in the fl owering season; dried fallen fl owers are webbed together and fastened to fl ower clusters to form nests (Patel et al., 1977). The Lepidoptera complex attacking mango fl owers in Australia consists of several species from the families Geometridae, Lymanthridae, Noctuidae, Pyralidae and Tortricidae. In Brazil, Barbosa (2005) and Barbosa et al. (2005) reported Pleuroprucha asth-enaria (Walker) (Lepidoptera: Geometridae) and Cryptoblabes gnidiella (Milliere) (Lepidoptera: Pyralidae) affecting mango infl orescences. The Pleuroprucha asthenaria life cycle from egg to adult is 17 days and the C. gnidiella life cycle is 36 days (Barbosa, 2005). Pleuroprucha asthenaria can cause premature ripen-ing of injured fruit while C. gnidiella infests infl orescences that are compacted from paclobutrazol applications (Barbosa, 2005).
According to Schreiner (1987), Dipel® reduced caterpillar damage, but careful monitoring or constant spraying was necessary to prevent signifi cant damage. In Brazil, the pesticides Bacillus thuringiensis, trichlorfon and lamb-dacyhalothrin provided 66–59% mortality (Barbosa, 2005). Control with pes-ticides is mostly unjustifi able in Florida USA and Australia, but regular monitoring is needed for early detection of population increases (Cunning-ham, 1984; Peña, 1993).
Classical biological control of lepidopteran insects attacking mango in Dominica was initiated with the introduction of the wasps, Aleiodes sp. and Euplectrus sp., and the fl y Blepharella lateralis Macquart. Populations of the pest were reduced to 25% of pre-release levels; parasitization rates were 20–99%, with Euplectrus sp. being the most abundant parasitoid (Nafus, 1991). The parasitoid Macrocentrus prob. delicatus attacks P. attramentalis; however, the parasitism rate is unknown (Peña, 1993). In Brazil, C. gnidiella is parasitized by Brachymeria pseudoovata Blanch (Hymenoptera: Chalcididae).
ThripsThe western fl ower thrips, Frankliniella occidentalis (Pergande) damages fl ow-ers and fruit in Israel (Wysoki et al., 1993). The developmental time of F. occi-dentalis from egg to egg at 25°C occurs between 14.8 and 16.65 days. The duration of development of F. occidentalis from egg to adult is closely related to environmental conditions, especially temperature. Frankliniella (possibly cubensis) is present in mango fl owers during the dry season in Costa Rica, requiring several applications of systemic insecticides (Jirón, 1993). In Florida,

J.E. Peña et al.338
the thrips complex consisting of Frankliniella bispinosa (Morgan) and F. kelliae (Sakimura) is the most frequently observed blossom pest on fl owers and causes damage by ovipositing in the panicle and feeding on the fl oral necta-ries and anthers, which may result in premature loss of pollen. The biology of F. bispinosa has been reviewed by Watson (1917), and Sakimura (1981) studied the taxonomy of F. kelliae. In South Africa, Grove et al. (2001) reported that Thrips acaciae Thybom, Thrips tenellus Trybom and S. aurantii were the most abundant species collected from mango fl owers.
SAMPLING. Thrips density is related to fl ower phenology and the prevalent dry season in Florida USA. Peña (1993) suggested that aerial trapping is superior to fl ower inspection, but because there is no method for determin-ing the true population size, the aerial trap method cannot be shown to be an unbiased estimator. In India, Verghese et al. (1985) determined that the lower mango canopy is better for sampling, and recommended a sample size of 55 panicles/tree for surveying. Verghese et al. (1988) indicated that distribution of Thrips palmi (Karny) on mango panicles is better explained by Iwao’s patchiness regression, which indicated an aggregated distribution and suggested that the lower canopy should be sampled. Sample sizes should be 55 panicles/tree for control and survey studies, with a 20% error of the mean and 92 panicles/tree to obtain a lower (5%) percentage error of the mean. In Florida, Peña et al. (2006a) showed that a cumulative number of 400–700 thrips/panicle during 4 weeks causes 33–50% yield reduction of ‘Keitt’ mangoes.
CHEMICAL CONTROL. The effi cacy of different pesticides (acetamiprid, fenpro-patrin, milbemectin and zeta-cypermethrin, novaluron) was tested by Peña et al. (2006a) against mango fl ower thrips. All treatments, except milbemec-tin, reduced thrips densities 5 days after application of the fi rst spray. Danitol and zeta-cypermethrin had the lowest numbers of thrips 12 and 20 days after the fi rst spray. All treatments had lower thrips densities compared to the control at 28, 35 and 43 days after the second application.
BIOLOGICAL CONTROL. Several parasitoids and predators, such as Ceranisusmenes (Walker) in Israel (Rubin and Kuslitizky, 1992) and the predators, Oriussp., Anystis agilis Banks and Hypoaspis aculifer (Canestrini) (Loomans et al., 1995), are candidates for biological control of F. occidentalis, while Dasyscapusparvipennis Gahan is considered to have good potential for the biological con-trol of fl ower thrips in Puerto Rico (Bartlet, 1938).
Pests of buds and leaves
Foliage feeders is one of the largest groups of injurious insects of mango. Pests of mango buds and foliage may cause damage by reducing the photo-synthetic area of the plant, thereby reducing the quantity of photosynthates translocated to the fruit. The most destructive mango leaf feeders are thrips,

Pests 339
midges, mites, scales, whitefl ies, mealybugs, weevils, ants, locusts and cater-pillars (Jeppson et al., 1975; Bhole et al., 1987; Jadhav and Dalvi, 1987; Tigvatt-nanont, 1988). Formation of leaf galls in India is caused by the Eurytomidae, Mangoma spinidorsum (Subba Rao, 1986), but there is little information on their importance as foliage pests.
ThripsThe Mediterranean mango thrips, Scirtothrips mangiferae Priesner, is a severe pest of mango in Israel, causing the young leaves to curl along the midrib, distorting their shape and leading to premature drop (Wysoki et al., 1993). The twigs of infested shoots are much shorter than those of uninfested ones. The population of the thrips is low during winter, increases in early spring and reaches its peak during summer (Wysoki et al., 1993). Yellow sticky traps can be used for monitoring thrips densities. Ganz et al. (1990) established that an average population of ten Mediterranean mango thrips per young shoot was the threshold above which chemical control is required. Effi cient control has been achieved by spray application of fl uvalinate or acephate (Ganz et al., 1990).
The red banded thrips, Selenothrips rubrocinctus (Giard), is an important pest of cacao in the Caribbean islands and attacks mango and avocado in Australia and Florida and Hawaii USA. The adults feed on the underside of leaves, causing necrosis and subsequent leaf drop. According to Hill (1975), S. rubrocinctus is only a pest in mango nurseries, and rarely damages mature trees. Its biology was reviewed by Moznette (1922). Adult thrips are dark bodied with a red band on the fi rst abdominal segment. The immature stages are light orange with abdominal segments one and two and the anal seg-ments bright red. The population of this species peaks during the dry season and declines during the rainy season. According to Yee (1987), the thrips are controlled by malathion (25% v/w).
The weaver ant, O. smaragdina (Hymenoptera: Formicidae) is considered an effective biological control of S. rubrocinctus in the Northern Territory of Australia (Peng and Christian, 2004).
MidgesMango leaves are attacked by different Cecidomyiidae species, especially in Asia, but also in the Caribbean region. Two genera, Procontarinia Kieffer and Cecconi and Erosomyia Felt, are particularly associated with mango and all known species have been reared from mango (USDA, 1981; Schreiner, 1991; Harris and Schreiner, 1992; Uechi et al., 2002; Gagne and Medina, 2004). Prasad (1971) described the biology of the main species attacking mango in India. A new species of gall midge, Procontarinia schreineri Harris, which attacks mango foliage in Guam, lays eggs on young mango leaves and larvae develop rapidly over c.5 days and induce blister galls. According to Harris and Schreiner (1992), the main factors affecting populations of this midge are rainfall and location. More galls are present during rainy periods, possibly because RH improves larval and pupal survival. No differences were observed in gall densities collected from lower and top portions of the tree. Askari and Radjabi (2003) observed overlapping generations of Procontarinia

J.E. Peña et al.340
matteiana in Iran related to the different leaf fl ushing patterns and found that the optimum pest temperatures were 10–26°C. Differences on susceptibility of mango cultivars to P. matteiana might indicate that susceptible cultivars should not be grown in areas infested by this gall midge (Jhala et al., 1987; Githure et al., 1998). Daneel et al. (2000) suggested that products to control P. matteiana should be applied after harvest, coinciding with the fi rst major fl ush and a second spray 6 weeks later. Austin (1984) and Sankaran and Mjeni (1989) have reported several platygastrid species parasitizing Procontarinia spp. and their prospects for biological control of the pest.
MitesThe mango bud mite, Aceria mangiferae (Sayed) (Acari: Eriophyidae), attacks buds and infl orescences (Keifer et al., 1982; Ochoa et al., 1994) (Plates 69–71). According to Jeppson et al. (1975) this mite stunts and brooms twigs, causing bud proliferation and appears to be responsible for necrosis of bud tissue cells (Varma et al., 1974). In Hawaii, Tegonotus mangiferae (Acari: Eriophyidae) feeds on the underside of leaves (Jeppson et al., 1975), while another species, Metaculus mangiferae (Attiah) (Acari: Eriophyidae) causes russeting of termi-nal leaves, buds and infl orescences. The latter is an important pest in Egypt, India, Palestine and Angola (Jeppson et al., 1975). The puncture wounds of several acarines (Acari: Tetranychidae) cause serious damage to leaves, which may dry and fall. The main pest in Mauritius, India, Egypt, Israel and Peru is Oligonychus mangiferus (Rahman and Sapra); in Israel, the spider mite Tet-ranychus cinnabarinus (Boisduval), which lives on the underside of the leaves, causes bronzing around the puncture wounds. The adult life span of O.mangiferus is 10.11 days for females and 4.21 days for males (Rai et al., 1988). The avocado red mites, Oligonychus yothersi McGregor and Oligonychus puni-cae (Hirst), feed on the upper surface of leaves and cause considerable sti-pling around the midrib at high population densities (Andrews and Poe, 1980). If the tetranychid mites are suffi ciently abundant, infested leaves may drop. There is little or no information on sampling techniques or for their economic thresholds.
Aceria mangiferae occurs wherever mango is grown (Denmark, 1983; Doreste, 1984). There has been controversy regarding a possible association between this mite and fl oral and foliar galls, i.e. mango malformation (Sayed, 1946; Narasimhan, 1959; Summanwar and Raychoudhury, 1968; Denmark, 1983; Ochoa et al., 1994). However, A. mangiferae does not cause mango mal-formation, but may be a carrier of Fusarium mangiferae, which is recognized as the causal agent of mango malformation (Varma et al., 1974; Freeman et al., 2004). Aceria mangiferae life history has been described by Abou-Awad (1981); it is completed in 15 days at 25–27°C.
DAMAGE. According to Keifer (cited in Jeppson et al., 1975), A. mangiferae infestation of buds results in arrested growth, with the stunted, short, young stems close together at the terminal branch. When the leaves fall, the overall effect is scanty growth of twiggy branches, with a few stubby, short branch-lets and discoloured buds. The mite appears to be responsible for necrosis of

Pests 341
bud tissue cells, which is initiated externally at the edge of the bud and pro-gresses toward the centre and internal areas of the bud (Peña et al., 2005).
SAMPLING. Peña et al. (2005) reported preliminary results of the distribution and sampling techniques for A. mangiferae. Taylor’s power law and Iwao’s patchiness regression were used to analyse spatial distributions of the mango bud mite in mango orchards. Taylor’s power law generally provided a better description of variance-mean relationships for the species than did Iwao’s patchiness regression. The species exhibited aggregated patterns of spatial distribution. More mites were found on apical buds than on lateral-latent buds. Sample size requirements for fi xed levels of precision were determined by using variance-mean relationships (Peña et al., 2005). For a given mean, and desired precision, different numbers of samples are required. For instance, for a 10% precision and at 0.5 mites/bud, c.220 samples are required, whereas for a 30% precision at 0.5 mites/bud, only 25 samples are required. Sampling small arthropods (i.e. mites) is operationally diffi cult and often time consuming. To ease this burden, presence-absence, or binomial, sam-pling was tested in place of complete counts for estimating or classifying densities of these organisms. Binomial sampling is based on defi ning the pro-portion of one or more individuals – the percent of incidence (the incidence P(I)) – and the density of animals (m) per sampling unit. At densities of three and ten mites/bud, 71% and 42% buds, respectively, are considered unin-fested. According to the equation, P(I) = 0.28 + 0.01 (mites/bud) an infesta-tion level of more than ten mites/bud or a P(I) > 38% could be used as the nominal threshold (Peña et al., 2005).
BIOLOGICAL CONTROL. The phytoseiid Amblyseius swirski Athias Henriot is associated with A. mangiferae (Abou-Awad, 1981). In Florida USA, several unidentifi ed phytoseiids occur on buds infested with A. mangiferae. Tenuipal-pid (Brevipalpus phoenicis), Tydeid and Tarsonemid (Tarsonemus confusus (Ewing)) mites also inhabit mango buds. Therefore, it is diffi cult to deter-mine the mite species that is the host prey (Peña, personal observation).
CHEMICAL CONTROL. Osman (1979) reported that applications of four full cov-erage sprays of dichlorvos were effective for controlling A. mangiferae in Egypt. Rai et al. (1966) cautioned that chemical control should be directed to apparently healthy and not malformed tissues. In Florida USA, agrimek plus citrus oil, fenproximate and fenpropathrin resulted in the lowest mite densi-ties 12 days after application. Agrimek plus citrus oil, and acequinocyl resulted in the lowest mite densities 26 days after treatment (Peña et al., 2005). In Brazil, Nascimento et al. (2002) recommended sulfur applications.
PLANT RESISTANCE. In Florida USA, the densities of A. mangiferae were mea-sured on 22 mango cultivars from December 1997 to June 1998. ‘Keenan’, an unknown cultivar, ‘cv. 9819’, ‘Brander’ and ‘Bombay Green’ had signifi cantly more mites than ‘Joellen’, ‘Duncan’, ‘Red Itamaraca’, ‘Smith’, ‘Wally’ and ‘Hindi’ (Peña et al., 2005).

J.E. Peña et al.342
ScalesARMORED SCALES. At least 26 species of diaspidids attack mangoes worldwide (Chua and Wood, 1990). In India, Aspidiotus destructor Signoret causes serious damage, while Parlatoria pergandii Comstock and Lepidosaphes gloverii (Pack-ard) damage 3-year-old plants (cited in Chua and Wood, 1990). Radionaspisindica (Marlatt) (= Leucaspis indica) encourages growth of black mould, which covers young branches (Dekle, 1976). Several diaspidids, e.g. Aulacaspismangiferae (tubercularis) Newstead, attack shoots and leaves; the oleander scale (Plate 72) in Florida USA (Miller and Davidson, 2005) and the mango scale in Ghana (van Halteren, 1970) cause similar damage. They are damag-ing not only because they feed on sap, but also because of the toxicity of their saliva (Singh, 1991). Scales inhabit both leaf surfaces and also are on fruit. Van Halteren (1970) concluded that A. mangiferae development is completed in 35–40 days for females and 23–28 days for males.
SOFT SCALES. Other species of Coccidae, Coccus viridis (Green), Coccus longu-lus, Ceroplastes actiniformis, Philephedra tuberculosa Nakahara and Gill and the mango shield scales Milviscutulus mangiferae (Green) and Viusonia stellifera (Westwood) in Asia, Africa, Australia, Israel and the Americas cause similar damage. These coccids are generally polyphagous, attacking different genera and species. They are mobile and injure mango because of the production of honeydew and the subsequent accumulation of sooty mould on the honey-dew (Escalante, 1974; Silva and Cavalcante, 1977). Most of these scales can be suppressed at sub-economic levels, either by application of selective pesti-cides (i.e. oils) or by biological control agents.
SAMPLING. Because of their small size, it is laborious to sample all stages of scales. Pheromones in tent-style traps have been used with other fruit crops, as well as monitoring crawler movement using sticky bands close to infested leaves. Either double-sided sticky tape, or tape coated with Vaseline® is effec-tive for trapping crawlers. Bands are removed and examined under a micro-scope to determine crawler numbers. Dark-coloured tape provides a better contrast to detect crawlers (Beers et al., 1993).
BIOLOGICAL CONTROL. In a survey of mango-producing areas in South Africa, Labuschagne (1993) determined that the predatory thrips Auleurodothrips fasciapennis Franklin and the parasitoid Aspidiotiphagus citrinus (Crawford) are the most important biocontrol agents of A. tubercularis. In South Africa, Joubert et al. (2000) obtained 46% parasitism of A. tubercularis using an unidentifi ed species of the parasitoid, Aphytis sp. Arias et al. (2004a) observed Coccidophilus spp. (Coleoptera: Coccinellidae) and Chrysopa spp. preying on A. tubercularis in Ecuador; the exotic predator Cybocephalus nipponicus (Coleoptera: Nitidulidae) was introduced to supplement predation of the former scale (Arias et al., 2004b). Several parasites have been recorded in Israel parasitizing the mango shield scale: Coccophagus lycimnia (Walker), C. eritraensis Compere, C. scutellaris (Dalman), C. bivittatus (Compere),

Pests 343
Microterys fl avus (Howard) and Metaphicus fl avus Howard. Usually no chem-ical control is required for this scale in Israel due to the activity of natural enemies (Kfi r and Rosen, 1980). Natural enemies for the control of the pink wax scale Ceroplastes rubens Maskell in Australia include the parasitic wasps Anicetus benefi cus Ishii and Yasumatsu, Aenasioidea varia Girault and Rhopal-encyrtoidea dubia Girault (Cunningham, 1984).
Whitefl ies and blackfl iesThe two fl y species of economic importance are the whitefl y, Aleurodicus dis-persus Russel and the blackfl y, Aleurocanthus woglumi Ashby. The whitefl ies suck cell sap from leaves, which wilt when whitefl y populations are high. High infestations can almost blacken entire trees, reducing photosynthetic effi ciency and causing defoliation (Angeles et al., 1971; Peña, 1993). A number of parasitoids, e.g. Encarsia opulenta (Silvestri) and Amitus hesperidus (Silves-tri), attack the immature stages and provide good control.
MealybugsMealybugs injure mango by sucking sap through their stylets, and excreting large amounts of honeydew onto fruit and leaves. Sooty mould fungus growth on the honeydew can render the fruit unmarketable, and reduce the photosynthetic effi ciency of leaves and cause leaf drop (CAB International, 2003). The margarodid mango mealybug Drossicha stebbingi (Green) is a seri-ous pest of mango in India and Pakistan (Prasad and Singh, 1976; Mohyud-din, 1981; Mohyuddin and Mahmood, 1993). It is univoltine. After mating, females enter the soil in June and die after laying eggs at a depth of up to 15 cm. These begin to hatch at the end of December or early January. The nymphs emerge from the soil and move to tender shoots where they settle. Prasad and Singh (1976) reported that the intensity of attack varied with respect to year and locality in India, probably because of soil and environ-mental conditions. Moderate rainfall (55–60 mm) during oviposition and dry conditions during hatching appear to favour development. Adults develop in April. They mate, and males die soon afterwards. The females enter the soil in May for oviposition, and the diapausing eggs remain in the soil until the end of December.
The pseudococcid fruit tree mealybug, Rastrococcus invadens Williams, is a serious pest of several crops, including mango in West Africa (Agounké et al., 1988). Mealybugs feed on leaves and fruit. Females have three moults and males have four. The entire life cycle can be completed in 31–84 days. The mealybugs weaken plants by puncturing the tissues and consuming sap, but the major damage is caused by the production of large amounts of honeydew upon which saprophitic fungi develop. The resultant thick black layer of sooty mould causes a drastic reduction in photosynthetic effi ciency, resulting in premature leaf drop. Rastrococcus invadens severely reduces fruit produc-tion in some areas of Africa (Moore and Cross, 1992).
SAMPLING. Boavida et al. (1992) devised sampling plans for R. invadens, but advised that the sampling strategy was only practical for estimating medium to high mealybug populations in the fi eld. Narasimham and Chacko (1991)

J.E. Peña et al.344
determined that densities of R. invadens Williams, Rastrococcus iceryodes (Green) and Rastrococcus mangiferae (Green) were signifi cantly higher on the abaxial than the adaxial surface. Mealybug density was also higher from ground level to 2 m compared with >2 m; they also observed that spatial distribution of R. iceryodes did not differ among internal and external cano-pies, whereas densities of R. invadens and R. mangiferae were higher in the external canopy. They also determined that there were statistical differences causing some inter-tree variation, but did not determine if the mealybugs followed a random or contagious distribution.
CONTROL. Various control methods, including banding tree trunks with vari-ous materials to prevent D. stebbingi nymphs from climbing (Lakra et al., 1980; Srivastava, 1981) and dusting chlorinated hydrocarbons on the soil (Srivastava, 1981), have been tried with little success. In Pakistan the mango mealybug was controlled by hoeing or ploughing the soil and conservation of the predator, Sumnius renardi Weise, by wrapping burlap around the trunks of the trees (Mohyuddin and Mahmood, 1993).
Pests of trunks, twigs and roots
Coleopterans and scalesStem-boring Coleoptera and scales as a group of injurious mango insects have not been studied in great detail (van Whervin, 1968; Woodruff, 1985). The wide host range of borers and overlapping borer species has compli-cated their study. Tunnelling of borer larvae in branches and trunks of mango and the slow feeding of some scale species in certain seasons and regions may cause serious reductions in yields and might also contribute to mango decline. However, borer occurrence and injury tends to be sporadic and below levels requiring direct action. Few natural enemies have been reported for suppression of borer populations in mango. Detailed coverage of stem-boring species is beyond the scope of this chapter; however, Hypocryphalusmangiferae (Stebbing), Apate monachus Fabricius and Batocera rubus L. are pests within this group.
Infestations of the mango scale, Radionaspis indica (Marlatt), and plumose scale, Morganella longispina, commonly occur on the trunk, branches and buds. Severe infestations include cracking of bark, exudation of sap and decline of the upper branches. Peña (1993) demonstrated that branches with both species of scale showed more decline symptoms than branches with low-scale density. Research on the bionomics and control of these scales is necessary to confi rm their role in mango decline symptomatology.
The scolytids, Hypocryphalus mangiferae (Stebbing) and Xylosandrus com-pactus (Eichoff) directly attack the main stem and branches (Silveira, 1960; Wysoki et al., 1993). Growth of fungal mycelia can extend terminally and basally from the beetle gallery in the mango tree and can kill the affected branches. The insects prefer trees that have been weakened by pathogens, wind, etc., but after a population has been established, the infestation spreads

Pests 345
to healthy trees. Hypocryphalus mangiferae has been associated with mango wilt disease in Brazil and Oman (van Wyk et al., 2007). Following initial trap-ping results from Berti Filho and Fletchman (1986), Rocha da Silva (2006) established that H. mangiferae is attracted to trees where the fungus is pres-ent. Scolytid beetles are attracted to mango trees in response to visual stimuli, to host-specifi c chemicals and to species-specifi c aggregation pheromones (Lindgreen et al., 1982). The evaluation of traps as tools for managing ambro-sia beetles on mangoes in Florida USA is necessary in order to reduce their damage in newly established groves.
TermitesMango orchards are becoming more common in dry and semi-arid areas with vast termite populations. Mango growing in infested areas often results in plant growth suppression as a result of reduced root establishment, inva-sion and pruning of roots (Rogers et al., 1999). For example, six termite spe-cies (Odontotermes indicus Thakur, O. lokanandi Chatterjee and Thakur, O.obesus (Rambur), O. giriensis (Roonwal and Chhotani), O. bhagwatii Chatter-jee and Thakur and Microtermes obesi Holm) were recorded from mango orchards in India (Srivastava and Singh, 2004). More species of termites were observed in winter than during the summer and rainy season. Veeresh et al. (1989) observed that O. wallonensis, O. horni and O. obesus constructed earthen sheeting on the stem of small mango trees. Singh (1960), cited by Srivastava (1998), reports Eutermes (Nasutitermes) costali, Calutermes (Cryptotermes) bre-bis, Heterotermes tenuis, Coptotermes gestroi, Neotermes (Kelatermes) bosei (Gard-neri) Synder, Microcereoutermes peroffi nis, Calotermes (Neotermes) greeni and Coptotermes curvignathus also affecting mango in India. According to Srivas-tava (1998), the most important termite species affecting mango are O. obesus and O. wallonensis; O. wallonensis nests in the root zone in Uttar Pradesh, India. The workers feed on roots, stems and branches.
Colonies of the subterranean termite, Coptotermes curvignathus Holmgren, were monitored in Malaysia using bait matrices containing 0.5% hexafl u-muron (Said Sajap et al., 2000). In Florida, urban dwellings infested with the Formosan termite, C. formosanus Shiraki, were treated with baits containing 0.5% weight/weight novifl umuron (Cabrera and Thoms, 2006). These baits might be useful when mango orchards are planned for areas infested with termites. In India, termite infestations are controlled with a combination of monthly irrigation, hoeing and application of neem cake (Singh and Singh, 2003).
10.3 Discussion
In general, most mango pests also occur on other fruit crop species. Fruit fl ies, scales, mites, thrips, lepidopteran fl ower feeders, mirids, weevils and beetles are mostly generalists, and some of their management schemes need to be implemented with this in mind. In the case of fruit fl ies, Aluja (1996) suggests surveying vegetation adjacent to infested mango orchards as

J.E. Peña et al.346
populations are sustained and multiplied in these locations and from them adult fl ies move into commercial orchards to attack ripening fruit (Aluja etal., 1996). Management of key pests (i.e. fruit fl ies, seed weevils, etc.) must be mandatory, in order to have an effect on a large region. The use of some mea-sures (i.e. quarantine, etc.) must involve neighbouring producing countries in order to have a positive effect on sanitation. The most progressive exam-ples in management of mango pests are in Australia, Mexico and Israel, while other producing countries, such as South Africa, Brazil and Ecuador, are tak-ing serious steps to reconcile the opposing forces of globalization of markets and sustainibility. For other areas where maximizing yields and blemish-free fruit is not a priority, the emphasis should be biological control. Management tactics that can be improved include the following:
Selective pesticides. Pesticides that are used in integrated pest man-1. agement programmes must have selective toxicity. The current trend is the development of chemicals that are highly effective for a limited group of in-sects. Díaz-Fleischer et al. (1996) suggested the use of cyromazine to reduce fertility of A. obliqua. Cunningham (1989) suggested that oils could be uti-lized for control of scales in mango; however, most of the recommendations are based on highly toxic or illegal, non-registered persistent chemicals (Singh, 1991; van Mele et al., 2001; de Bie, 2004). In South Africa, Joubert et al. (2004) tested kaolin, which is the active ingredient of Surround®, a non-toxic natural clay mineral, against the mango seed weevil, mango scale and citrus thrips. Surround® was effective against citrus thrips, mango weevil and co-conut bug, P. wayi, but caused outbreaks of mango scale and long-tailed mealybug, Pseudococcus longispinus. However, producers of export fruit rely on calendar-based chemical control when trees are heavily infested with mango scale (Joubert et al., 2004).Biological control. Biological control has great potential as a tactic for regulating pest populations in integrated pest management programmes in mango orchards. However, it will be diffi cult for biological control alone to reduce a pest from an economic to a completely non-economic status for pests attacking fruit. A combination of augmentative releases of parasi-toids and the use of sterile insects, at least from a theoretical perspective, has been considered to be more effective for fruit fl ies than either method applied alone (Barclay, 1987). Biological control should be highly effective for indirect pests. Indeed, numerous studies have been conducted in many mango-producing countries to promote the use of parasites and predators for this type of pest (Cunningham, 1989; Mohyuddin and Mahmood, 1993; Moore and Cross, 1993; Whitwell, 1993; Wysoki et al., 1993; Labuschagne et al., 1995).
Host plant resistance. Tolerance of mango to pests is mentioned for 2. Noorda sp. and Idioscopus sp. (Bagle and Prasad, 1984; Cunningham, 1989), while man-go resistance to Sternochetum mangiferae is mentioned by Hansen (1993). Carvalho et al. (1996) have also demonstrated the different degrees of suscepti-bility of mango cultivars to A. obliqua. Most of this research, however, needs

Pests 347
to be assessed further. Angeles (1991) reported that Mangifera altissima does not seem to be affected by leaf hoppers, tip borers and seed borers in the Philippines. There is little doubt that wild mangoes have potential in breed-ing. Determining the tolerance or insect resistance of mango cultivars and related species should be done in natural stands or in established germplasm collections. After the initial selection has been made, evidence for the pattern of resistance must be established and changes in the environment, whether geographic or temporal, should not disrupt or decrease the resistance to any great extent. Therefore, tests for resistance in mango to insects should in-clude provision for exposure to insects under varying conditions whenever possible.
Pheromones and trapping devices. Developments in the identifi cation 3. and synthesis of sex pheromones have resulted in their possible use for pest management in mango orchards (Chu et al., 1994; Khan et al., 2002, 2005; Sheikh et al., 2008). Food attractants, however, remain the most common monitoring tools. Trapping techniques can be utilized to reduce pesticide use by improving timing of sprays as a result of better monitoring of pest popu-lations. It remains uncertain if trapping techniques can be used to predict infestations by fruit-feeding pests and if they can be used for direct control (by mass trapping) over several years.
Cultural and physical control. Use of cultural and physical techniques 4. (i.e. pruning, bagging, etc.) depends on costs of control, availability of techni-cal assistance and market purposes. Paderes and Orden (2004) observed that bagging and pruning of ‘Manila’ in the Philippines is mostly infl uenced by the availability of technical assistance for growers.
Greatly increased regulation of pesticides, heightened public aware-ness of environmental contamination, pesticide resistance problems in pests and the high cost of chemical pest control has resulted in increasing reliance on integrated pest control as an important strategy in sustainable agriculture.
Acknowledgements
We are particularly indebted to Walther Enkerlin (Joint IAEA/FAO Division, Vienna), Andrea Birke-Biewendt, Alberto Anzures-Dadda and Larissa Guillén-Conde (Instituto de Ecología, AC) and Aldo Malavasi (private consultant) for their assistance and for providing many valuable references. M. Aluja acknowledges the fi nancial support of the Mexican Campaña Nacional Contra Moscas de la Fruta (Convenio SAGARPA-IICA-INECOL) and the Instituto de Ecología, AC. He also acknowledges support from CONACyT through a Sabbatical Year Fellowship (Ref. 79449) and thanks Benno Graf and Jörg Samietz (Forschungsanstalt Agroscope Changins-Wädenswil ACW), for pro-viding ideal working conditions during the fi nal phase of the publication process of this chapter.

J.E. Peña et al.348
References
Abbas, S. (1985) Studies on the mango infl orescence midge, Erosomyia indica Grover. Acta Horticulturae 231, 593–596.
Abou-Awad, B. (1981) Ecological and biological studies on the mango bud mite, Erio-phyes mangiferae (Sayed), with description of immature stages Eriophyoidea: Erio-phyidae. Acarologia 22, 145–150.
Agounké, D., Agricola, U. and Bokonon-Ganta, H. (1988) Rastrococcus invadens Wil-liams (Hemiptera: Pseudococcidae), a serious exotic pest of fruit trees and other plants in West Africa. Bulletin of Entomology Research 78, 695–702.
Ahmed, M.U., Ahmed, M., Baluch, M.A. and Naheed, R. (1981) Sooty mold on mango plants and its relationship with leafhoppers and climatic factors in Karachi-Pakistan during 1978–79. Pakistan Journal of Science and Industrial Research 24, 140–144.
Aluja, M. (1993) Manejo Integrado de la Mosca de la Fruta. Editorial Trillas, Mexico Distrito Federal, Mexico.
Aluja, M. (1994) Bionomics and management of Anastrepha. Annual Review of Ento-mology 39, 155–178.
Aluja, M. (1996) Future trends in fruit fl y management. In: McPheron, B. and Steck, G. (eds) Fruit Fly Pests: a World Assessment of their Biology and Management. St Lu-cie Press, Delray Beach, Florida, pp. 309–320.
Aluja, M. (1999) Fruit fl y (Diptera: Tephritidae) research in Latin America: myths, reali-ties and dreams. Anais da Sociedade Entomologica do Brasil 28, 565–594.
Aluja, M.R. and Liedo, P.F. (1986) Perspectives on future integrated management of fruit fl ies in Mexico. In: Mangel, M., Carey, J.R. and Plant, R.E. (eds) Pest Control: Op-erations and Systems Analysis in Fruit Fly Management. NATO ASI Series Vol. G11.Springer-Verlag, Berlin, pp. 9–42.
Aluja, M. and Mangan, R.L. (2008) Host status determination for pestiferous fruit fl y (Diptera: Tephritidae) species: conceptual, methodological and regulatory consid-erations. Annual Review of Entomology 53, 473–502.
Aluja, M. and Norrbom, A.L. (eds) (2000) Fruit Flies (Tephritidae): Phylogeny and Evolu-tion of Behavior. CRC Press, Boca Raton, Florida.
Aluja, M. and Piñero, J. (2004) Testing human urine as a low-tech bait for Anastrephaspp. (Diptera: Tephritidae) in small guava, mango, sapodilla and grapefruit orchards. Florida Entomologist 87, 41–50.
Aluja, M., Cabrera, M., Guillen, J., Celedonio, H. and Ayora, F. (1989) Behavior of Anas-trepha ludens, A. obliqua and A. serpentina (Diptera: Tephritidae) on a wild mango tree (Mangifera indica) harbouring three McPhail traps. Insect Science and its Ap-plication 10, 309–318.
Aluja, M., Celedonio-Hurtado, H., Liedo, P., Cabrera, M., Castillo, F., Guillen, J. and Rios, E. (1996) Seasonal population fl uctuations and ecological implications for management of Anastrepha fruit fl ies (Diptera: Tachinidae) in commercial mango orchards in southern Mexico. Journal of Economic Entomology 89, 654–667.
Aluja, M., Piñero, J., Jácome, I., Díaz-Fleischer, F. and Sivinski, J. (2000) Behavior of fl ies in the genus Anastrepha (Trypetinae: Toxotrypanini). In: Aluja, M. and Norrbom, A. (eds) Fruit Flies (Tephritidae): Phylogeny and Evolution of Behavior. CRC Press, Boca Raton, Florida, pp. 375–406.
Aluja, M., Sivinski, J., Rull, J. and Hodgson, P.J. (2005) Behavior and predation of fruit fl y larvae (Anastrepha spp.) (Diptera: Tephritidae) after exiting fruit in four types of habitats in tropical Veracruz, Mexico. Environmental Entomology 34, 1507–1516.
Aluja, M., Sivinski, J., Ovruski, S., Guillén, L., López, M., Cancino, J., Torres-Anaya, A., Gallegos-Chan, G. and Ruíz, L. (2009) Colonization and domestication of

Pests 349
seven species of native New World Hymenopterous larval-prepupal and pupal fruit fl y (Diptera: Tephritidae) parasitoids. Biocontrol Science and Technology (in press).
Andrews, K. and Poe, S. (1980) Spider mites of El Salvador, Central America (Acari: Tetranychidae). Florida Entomologist 63, 502–505.
Angeles, D.E. (1991) Mangifera atissima. In: Coronel, R.E. and Verheij, E.W.M. (eds)Plant Resources of South-east Asia No.2: Edible Fruits and Nuts. Pudoc-DLO, Wageningen, the Netherlands, pp. 206–207.
Angeles, N. and Requena, J. (1966) Moth injury of mangoes in Venezuela. Plant Protec-tion Bulletin FAO 14, 86–87.
Angeles, N., Dedordy, N., Paredes, P. and Requena, J. (1971) Citrus blackfl y in Venezu-ela. Agronomia Tropical 21, 71–75.
Anonymous (1984) Major insect pests and diseases of mango and their control. In: TheMango Technical committee. The Philippines recommends for Mango. Philippines Agriculture and Resources Research Foundation, 124 pp.
Anonymous (1987) Red-banded mango caterpillar Noorda albizonalis Hampsoni (Lepi-doptera: Pyralidae). Plant Quarantine Leafl et-Australia 51.
Anonymous (1989) Mango Pests and Disorders. Information Series QI89007. Queen-sland Department of Primary Industries, Brisbane, Australia.
Anonymous (2002) Fruit fl ies eradicated from Nauru. 3pp. Available at: http://www.pacifl y.org/Success_stories/Nauru_eradl.htm (accessed 10 September 2008).
Anonymous (2005) Malathion (CAS Number 121-75-5). Toxicology information about insecticides used for eradicating mosquitoes. Available at: http://www.atsdr.cdc.gov/consultations/malathion.html (accessed 20 July 2007).
Anonymous (2006) Importation of Fresh Mango Fruit (Mangifera indica) from India into the United States, a Qualitative Pest Risk Assessment. Animal and Plant Health In-spection Service, Plant Protection Quarantine, Raleigh, North Carolina.
Arias, M. and Jines, A. (2004) Manejo Integrado de Moscas de la Fruta en el Litoral Ecu-atoriano. Instituto Nacional de Investigaciones Agropecuarias – Programa de Mod-ernización de Servicios Agropecuarios, Guayaquil, Equador.
Arias, M., Jines, A., Bustos, P., Pluas, M. and Carrera, C. (2004a) Enemigos Naturales de Aulacaspis Tubercularis (Homoptera: Diaspididae) en Mango. Instituto Na-cional Autonomo de Investigaciones Agropecuarias (INIAP), Boliche, Guayas, Ecuador.
Arias, M., Jines, A., Maldonado, E. and Pluas, M. (2004b) Cybocephalus nipponicusEndrody-Younga (Coleoptera: Nitidulidae) un Predator Exótico Para el Control del Aulacaspis Tubercularis en Mango de Exportación. Instituto Nacional Autonomo de Investigaciones Agropecuarias (INIAP), Boliche, Guayas, Ecuador.
Askari, M. and Radjabi, G. (2003) Study on the biology and population fl uctuations of mango midge gall Procontarinia mattiana (Diptera: Cecidomyiidae) in Homozogan province. Applied Entomology and Phytopathology 70, 121–135.
Assis, E. and Rabelo, F. (2005) Pragas da Mangueira, Monitoramento, Nivel de Ação e Controle. Empresa Brasileira de Pesquisa Agropecuaria, Embrapa Semi-Arido, Min-isterio da Agricultura, Pecuaria e Abastecimiento, Petrolina, Brazil.
Austin, A.D. (1984) New species of Platygastridae (Hymenoptera) from India which parasitise pests of mango, particularly Procontarinia spp. (Diptera: Cecidomyiidae). Bulletin of Entomological Research 74, 549–557.
Australian Government (2004) Mangoes from India, Draft Revised Import Policy. De-partment of Agriculture, Fisheries and Forestry, Canberra, Australia.
Azizur Rahman, S.K. and Singh, G. (2004) Population dynamics of mango hopper (Am-ritodus atkinsoni) on ‘Langra’ mango (Mangifera indica) and its relationship with abiotic factors. Indian Journal of Agricultural Science 74, 566–569.

J.E. Peña et al.350
Babu, L.B., Maheswari, T.U. and Rao, N.V. (2002) Seasonal incidence and biology of the mango hopper Amritodus atkinsoni Lethier (Homoptera: Cicadellidae). Entomon27, 35–42.
Bagle, B. and Prasad, V. (1984) Varietal incidence and control of stone weevil Sternoche-tus mangiferae Fabricius. Indian Journal of Entomology 46, 389–392.
Balock, J.W. and Kozuma, T. (1964) Notes on the biology and economic importance of the mango weevil, Sternochetus mangiferae (Fabricius), in Hawaii (Coleoptera: Curculionidae). Proceedings of the Hawaiian Entomological Society 8, 353–364.
Balock, J.W. and Lopez, D.F. (1969) Trapping for control of the Mexican fruit fl y in mango and citrus groves. Journal of Economic Entomology 62, 53–56.
Barbosa, F.R. (2005) Arthropodes-pragas associados a cultura da mangueira no Brasil e seu controle. In: Menezes, E. and Barbosa, F.R. (eds) Pragas da Mangueira, Embra-pa Semi-arido. Ministerio da Agricultura, Pecuaria e Abastecimiento, Petrolina, Bra-zil, pp. 1–50.
Barbosa, F.R., de Gonçalves, M.E., Moreira, W.A., de Alencar, J.A., de Souza E.A., da Silva, C.S.B., Souza, A. and da Miranda, I. (2005) Arthrópodes-Praga e Predatores (Arthropoda) Associados à Cultura da Manguerira no Vale do São Francisco, Nor-deste do Brasil. Neotropical Entomology 34, 471–474.
Barclay, H.J. (1987) Models for pest control: complementary effects of periodic releases of sterile pests and parasitoids. Theoretical Population Biology 32, 76–89.
Barnes, H.F. (1948) Gall Midges of Economic Importance. Vol. 3: Gall Midges of Fruit.Crosby Lockwood and Sons, London.
Bartlet, K. (1938) The introduction and colonization in Puerto Rico of Dasyscapus parvi-pennis Gahan, a parasite of thrips. Agricultural Notes Puerto Rico Experiment Sta-tion US Department of Agriculture (USDA) 87, 6.
Bateman, M.A. (1972) The ecology of fruit fl ies. Annual Review of Entomology 17, 493–518.
Beardsley, J.W. (1961) A new review of the Hawaiian Braconidae (Hymenoptera). Pro-ceedings of the Hawaiian Entomology Society 27, 333–366.
Beers, E., Hull, L. and Jones, V. (1993) Sampling pest and benefi cial arthropods of apples. In: Pedigo, L. and Buntin, D. (eds) Handbook of Sampling Methods for Arthropods in Agriculture. Preface. CRC Press, Boca Raton, Florida, pp. 383–416.
Berti Filho, E. and Flechtman, C. (1986) A model of ethanol trap to collect scolytidae and platypodidae (Insecta, Coleoptera). Instituto de Pesquisas e Estudios Florestais34, 53–56.
Bess, H.A., van den Bosch, R. and Haramoto, F.H. (1961) Fruit fl y parasites and their ac-tivities in Hawaii. Proceedings of the Hawaiian Entomology Society 27, 367–378.
Bhole, S., Jadhav, V., Dumbre, R. and Dalvi, C. (1987) Seasonal incidence and chemical control of mango nursery pests in the Konkan region. Journal of the Maharashtra Agricultural University 12, 387–388.
Birke, A., Aluja, M., Greany, P., Bigurra, E., Pérez-Staples, D. and McDonald, R. (2006) Long aculeus and behavior of Anastrepha ludens renders gibberellic acid ineffective as an agent to reduce ‘Ruby Red’ grapefruit susceptibility to the attack of this pestiferous fruit fl y in commercial citrus orchards. Journal of Economic Entomology 99, 1184–1193.
Boavida, C., Neuenschwander, P. and Schulthess, F. (1992) Spatial distribution of Rastro-coccus invadens Williams (Homoptera: Pseudococcidae) in mango trees. Journal of Applied Entomology 114, 381–391.
Bondad, N.D. (1985) Mango bagging. Animal Husbandry and Agricultural Journal June Issue, 11–14.
Burns, R.E., Harris, D.L., Moreno, D.S. and Eger, J.E. (2001) Effi cacy of spinosad bait sprays to control Mediterranean and Caribbean fruit fl ies (Diptera: Tephritidae) in commercial citrus in Florida. Florida Entomologist 84, 672–678.

Pests 351
Bustos, M.E., Enkerlin, W., Reyes, J. and Toledo, J. (2004) Irradiation of mangoes as a postharvest quarantine treatment for fruit fl ies (Diptera: Tephritidae). Journal of Eco-nomic Entomology 97, 286–292.
Butani, D.K. (1979) Insects and Fruits. Periodical Expert Book Agency, Delhi, India.CAB International (2003) Crop Protection Compendium. CAB International, Walling-
ford, UK.Cabrera, B.J. and Thoms, E. (2006) Versatility of baits containing novifl umuron for con-
trol of structural infestations. Florida Entomologist 89, 20–31.Cabrera, H., Villanueva, J. and Ortega, D. (1993) Fruit fl y infestation timing on ‘Manila’
mango fruit at Veracruz, Mexico. (Abstract). IV International Mango Symposium,p. 118.
Camargo, C.A., Odell, E. and Jirón, L.F. (1996) Interspecifi c interactions and host prefer-ence of Anastrepha obliqua and Ceratitis capitata (Diptera: Tephritidae), two pests of mango in Central America. Florida Entomologist 79, 266–268.
Carvalho, P.J. and De Queiroz, A.C. (eds) (2002) A Cultura da Mangueira. CIP-Brasil, Brasilia.
Carvalho, R. da S., Nascimento, A., Morgante, J. and Fonseca, N. (1996) Susceptibility of different mango varieties to the attack of the fruit fl y, Anastrepha obliqua. In: McPheron, B. and Steck, G. (eds) Fruit Fly Pests: a World Assessment of their Biol-ogy and Management. St Lucie Press, Delray Beach, Florida, pp. 325–331.
Celedonio-Hurtado, H., Aluja, M. and Liedo, P. (1995) Adult population fl uctuations of Anastrepha species (Diptera: Tephritidae) in tropical orchard habitats of Chiapas, Mexico. Environmental Entomology 24, 861–869.
Christenson, L.D. and Foote, R.H. (1960) Biology of fruit fl ies. Annual Review of Ento-mology 5, 171–192.
Chu, Y., Lee, K.T. and Tseng, Y.H. (1994) Occurrence of melon and oriental fruit fl y in Republic of Naru. Plant Protection Bulletin 36, 131–140.
Chua, T.H. and Wood, B.J. (1990) Other tropical fruit trees and shrubs. In: Rosen, D. (ed.) Armored Scale Insects, their Biology, Natural Enemies and Control, 4B. El-sevier, New York, pp. 543–552.
Clarke, A.R., Allwood, A., Chinajariyawong, A., Drew, R.A.I., Hengsawad, C., Jirasurat, M., Kong-Krong, C., Kritsaneepaibon, S. and Vijaysegaran, S. (2001) Seasonal abun-dance and host use patterns of seven Bactrocera Macquart species (Diptera: Te-phritidae) in Thailand and Peninsular Malaysia. The Raffl es Bulletin of Zoology 49, 207–220.
Clarke, A.R., Armstrong, K.F., Carmichael, A.E., Milne, J.R., Raghu, S., Roderick, G.K. and Yeates, D.K. (2005) Invasive phytophagous pests arising through a recent tropi-cal evolutionary radiation: the Bactrocera dorsalis complex of fruit fl ies. AnnualReview of Entomology 50, 293–319.
Coto, D., Saunders, J.L., Vargas, C.L. and King, A.B.S. (1995) Plagas invertebradas de cultivos tropicales con enfasis en America central. Centro Agronómico Tropical de Investigación y Enseñanza. Serie Tecnica, Manual Técnico 12.
Cunningham, I.C. (1984) Mango insect pests. In: Australian Mango Research Workshop,Commonwealth Scientifi c and Industrial Research Organization (CSIRO), Mel-bourne, pp. 211–224.
Cunningham, I.C. (1989) Pests. In: Bagshaw, J. (ed.) Mango Pests and Disorders. Depart-ment of Primary Industries Information Series, Q189007, pp. 10–21.
Daneel, M.S., Jager, K. de, Steyn, W. and Husselman, J. (2000) Effi cacy of different in-secticides against gall fl y on mangoes. South African Mango Growers’ Association Yearbook 19, 20.
de Bie, C.A. (2004) The yield map of mango in Phrao, Thailand, as investigated through comparative performance evaluation. Scientia Horticulturae 102, 37–52.

J.E. Peña et al.352
De, K. and Pande, Y.D. (1987) Evaluation of certain non-insecticidal methods of reduc-ing infestation of the mango nut weevil, Sternochetus gravis (F.) in India. Tropical Pest Management 33, 27–28.
De, K. and Pande, Y.D. (1988) Bionomics and some behavioural aspects of the mango stone weevil, Sternochetus gravis (Fabricius) (Coleoptera: Curculionidae). Entomon13, 17–24.
De la Rosa, W., López, F.L. and Liedo, P. (2002) Beauveria bassiana as a pathogen of the Mexican fruit fl y (Diptera: Tephritidae) under laboratory conditions. Journal of Eco-nomic Entomology 95, 36–43.
De Macedo, F.P. (1998) Efeitos da aplicação de ácido giberélico em manga (Mangiferaindica L.) cultivar Tommy Atkins: aspectos horticulturais e entomológicos. Tesis de doctorado, Universidade de São Paulo, Brazil, 120 pp.
De Villiers, E.A. (1990) Coconut bugs in avocados. Farming in South Africa. Avocados H.5, 2pp.
Dekle, G.W. (1976) Florida armored scale insects. Arthropods of Florida and neighbor-ing land areas, Florida Department of Agriculture and Consumer Services, Gaines-ville, Florida, 3, 345 pp.
Denmark, H. (1983) Eriophyes mangiferae (Sayed) a pest of mango (Acarina: Eriophyidae).Entomology Circular Florida Department of Agriculture and Consumer Services 254.
Díaz-Fleischer, F. and Aluja, M. (2003) Clutch size in frugivorous insects as a function of host fi rmness: the case of the tephritid fl y Anastrepha ludens. Ecological Entomol-ogy 28, 268–277.
Díaz-Fleischer, F., Toledo, J., Enkerlin, W. and Fernandez, J. (1996) Cyromazine: effects on three species of Anastrepha. In: McPheron, B. and Steck, G. (eds) Fruit Fly Pests: a World Assessment of their Biology and Management. St Lucie Press, Delray Beach, Florida, pp. 336–337.
Dimbi, S., Maniania, N.K., Lux, S., Ekesi, S. and Mueke, J.K. (2003) Pathogenicity of Metar-hizium anisopliae (Metsch.) Sorokin and Beauveria bassiana (Balsamo) Vuillemin, to three adult fruit fl y species: Ceratitis capitata (Wiedemann), C. rosa var. fasciventrisKarsch and C. cosyra (Walker) (Diptera: Tephritidae). Mycopathologia 156, 375–382.
Dolinski, C. and Lacey, L.A. (2007) Microbial control of arthropod pests of tropical tree fruits. Neotropical Entomology 36, 161–179.
Doreste, E. (1984) Acarologia. Instituto Interamericano de Cooperacion para la Agricul-tura. San Jose, Costa Rica, 391 pp.
Drew, R.A.I. (1989) The tropical fruit fl ies (Diptera: Tephritidae: Dacinae) of the Austral-asian and Oceanian regions. Memoirs of the Queensland Museum 26, 521.
Drew, R.A.I. and Hancock, D.L. (1994) The Bactrocera dorsalis complex of fruit fl ies (Diptera: Tephritidae: Dacinae) in Asia. Bulletin of Entomological Research Supple-mental Series, No.2, 68 pp.
Dyck, V.A., Hendrichs, J. and Robinson, A.S. (eds) (2005) Sterile Insect Technique. Prin-ciples and Practice in Area-Wide Integrated Pest Management. Springer, Dordrecht, the Netherlands.
Ehler, I.E. and Endicott, P.C. (1984) Effect of malathion-bait sprays on biological control of insect pests of olive, citrus and walnut. Hilgardia 52, 1–47.
Ekesi, S., Nderitu, P.W. and Rwomushana, I. (2006) Field infestation, life history and demographic parameters of the fruit fl y Bactrocera invadens (Diptera: Tephritidae) in Africa. Bulletin of Entomological Research 96, 379–386.
Emery, R. (2002) Mango seed weevil and mango pulp weevil. Available at: http://www.agric.wa.gov.au/ento/Su...%20seed%20and%20pulp%20weevil.html (accessed 20 July 2007).
Enkerlin, W.R. (2007) Impact of fruit fl y control programmes using the sterile insect technique. In: Dyck, V.A., Hendrichs, J. and Robinson, A.S. (eds) Sterile Insect Tech-

Pests 353
nique. Principles and Practice in Area-Wide Integrated Pest Management. Springer, Dordrecht, the Netherlands, pp. 651–676.
Epsky, N., Hendrichs, J., Katsoyannos, B., Vasquez, A., Ros, J., Zumreolu, A., Bakri, A., Seewooruthum, S. and Heath, R. (1999) Field evaluation of female-targeted trappingsystems for Ceratitis capitata (Diptera: Tephritidae) in seven countries. Journal of Economic Entomology 92, 156–164.
Escalante, J.A. (1974) Insects of economic importance in Quillabamba, Cusco. Revista Peruana de Entomologia 17, 51–53.
Etienne, J. (1966) Lutte contre les mouches des fruits. Rapporte de Institute de Recher-ches Agronomiques Tropicales Réunion. Institute National de la Recherche Agronomique, Linne de Paradis, Reunion, France, pp. 112–113.
Etienne, J. (1968) Lutte contre les mouches des fruits. Rapporte de Institute de Recherches Agronomiques Tropicales Réunion. Institute National de la Recherche Agronomique, Linne de Paradis, Reunion, France, pp. 117–124.
Faih, H.A. and Huwer, R. (1994) Egg parasitoids collected from Amblypelta lutescens lutescens (Distant) (Hemiptera: Coreidae) in north Queensland. Journal of the Aus-tralian Entomology Society 32, 365–367.
Fasih, M., and Srivastava, R.P. (1990) Parasites and predators of insect pests of mango. Integrated Pest Control 32, 39–41.
Fasih, M., Srivastava, R.P., Abbas, S. and Sharma, S. (1989) Outbreak of Orgyia postica Walker (Lymantriidae: Lepidoptera), a new pest on mango in Uttar Pradesh. CurrentScience 58, 1258–1260.
Fay, H.A. and Huwer, R. (1994) Egg parasitoids collected from Amblypelta lutescens (Distant) (Hemiptera: Coreidae) in north Queensland. Journal of the Australian En-tomological Society 32, 365–367.
Fletcher, B.S. (1987) The biology of dacine fruit fl ies. Annual Review of Entomology 32, 115–144.Fletcher, M.J. and Dangerfi eld, P.C. (2002) Idioscopus clypealis (Lethierry), a second
new leafhopper pest of mango in Australia (Hemiptera: Cicadellidae: Idiocerinae). Australian Journal of Entomology 41, 35–38.
Follett, P. and Gabbard, Z. (2000) Effect of mango weevil (Coleoptera: Curculionidae) damage on mango seed viability in Hawaii. Journal of Economic Entomology 93, 1237–1240.
Follett, P. and Neven, L.G. (2006) Current trends in quarantine entomology. Annual Re-view of Entomology 51, 359–385.
Freeman, S., Klein-Gueta, D., Korolev, D. and Sztenjnberg, A. (2004) Epidemiology and survival of Fusarium mangiferae, the causal agent of mango malformation disease.Acta Horticulturae 645, 487–491.
Gagne, R.J. and Etienne, J. (2006) Gephyraulus mangiferae (Felt) N. Comb. (Diptera: Cecidomyiidae): a mango pest from India newly recorded from the western hemi-sphere. Proceedings of the Entomology Society of Washington 108, 930–937.
Gagne, R.J. and Medina, C.D. (2004) A new species of Procontarinia (Diptera: Cecid-omyiidae), an important new pest of mango in the Philippines. Proceedings of the Entomological Society of Washington 106, 19–25.
Galán-Saúco, V. (1990) Los Frutales Tropicales en los Subtropicos. MundiPrensa, Ma-drid, Spain, pp. 83–84.
Ganz, S., Hamieri, Y. and Nakach, Y. (1990) Control trials on the mango thrips. Annual Report of the Eden Experimental Station Israel. Institute of Plant Protection, Bet She’am, Israel.
García-Medel, D., Sivinski, J., Díaz-Fleischer, F., Ramirez-Romero, R. and Aluja, M. (2007) Foraging behavior by six fruit fl y parasitoids (Hymenoptera: Braconidae) re-leased as single- or multiple-species cohorts in fi eld cages: infl uence of fruit loca-tion and host density. Biological Control 43, 12–22.
García-Ramírez, M.J., Cibraián-Tovat, J., Arzufí-Barrera, R., López-Collado, J. and Soto-Hernández, M. (2004) Preferencia de Anastrepha ludens (Loew) (Diptera: Tephritidae)

J.E. Peña et al.354
por volátiles de frutos verdes o amarillos de mango y naranja. Agrociencia 38,423–430.
Githure, C.W., Schoeman, A.S. and McGeoch, M.A. (1998) Differential susceptibility of mango cultivars in South Africa to galling by the mango gall fl y, Procontarinia mat-teiana Kieffer & Cecconi (Diptera: Cecidoyiidae). African Entomology 6, 33–40.
Godase, S.K., Bhole, S.R., Shivpuje, P.R. and Patil, B.P. (2004) Assessment of yield loss in mango (Mangifera indica) due to mango hopper (Idioscopus niveoparvasus) (Ho-moptera: Cicadelidae). Indian Journal of Agricultural Sciences 74, 370–372.
Golez, H.G. (1991) Bionomics and control of the mango seed borer, Noorda albizonalis Hampson (Pyralidae: Lepidoptera). Acta Horticulturae 291, 418–424.
Grove, T. and Pringle, K. (2000) A sampling system for estimating population levels of the citrus thrips, Scirtothrips aurantii faure (Thysanoptera: Thripidae), mango or-chards. African Entomology 8, 223–226.
Grove, T., Giliomee, J.H. and Pringle, K. (2001) Thrips (Thysanoptera) species associated with mango trees in South Africa. African Entomology 9, 153–162.
Grover, P. (1986a) Integrated control of midge pests. Cecidologia Internationale 7, 1–28.Grover, P. (1986b) Population fl uctuation of Erosomya indica and Dasineura amaraman-
jarae and co-related extent of damage. Cecidologia Internationale 7,43–57. Hallman, G.J. and Sharp, J.L. (1990) Mortality of Caribbean fruit fl y (Diptera: Tephriti-
dae) larvae infesting mangoes subjected to hot-water treatment, then immersion cooling. Journal of Economic Entomology 83, 2320–2323.
Hancock, D.L., Hamacek, E.L., Lloyd, A.C. and Elson-Harris, M.M. (2000) The Distribu-tion and Host Plants of Fruit Flies (Diptera: Tephritidae) in Australia. Department of Primary Industries, Brisbane, Australia.
Hansen, J.D. (1993) Dynamics and control of the mango seed weevil. Acta Horticulturae341, 415–420.
Hansen, J.D. and Amstrong, J.W. (1990) The failure of fi eld sanitation to reduce infesta-tion by the mango nut weevil, Cryptorhynchus mangiferae (F.)(Coleoptera: Curcu-lionidae). Tropical Pest Management 36, 359–361.
Hansen, J.D., Armstrong, J.W. and Brown, S. (1989) The distribution of the mango wee-vil, Cryptorhynchus mangiferae (Coleoptera: Curculionidae), in Hawaii. Proceed-ings of the Hawaiian Entomological Society 29, 31–39.
Harris, E.J., Vargas, R.I. and Gilmore, J.E. (1993) Seasonality in occurrence and distribu-tion of Mediterranean fruit fl y (Diptera: Tephritidae) in upland and lowland areas of Kauai, Hawaii. Environmental Entomology 22, 404–410.
Harris, K.M. and Schreiner, I.H. (1992) A new species of the gall midge (Diptera: Ceci-domyiidae) attacking mango foliage in Guam, with observations on its pest status and biology. Bulletin of Entomological Research 82, 41–48.
Heath, R.R., Epsky, N.D., Guzman, A., Dueben, B.D., Manukian, A. and Meyer, W.L. (1995) Development of a dry plastic insect trap with food-based synthetic attractant for the Mediterranean and Mexican fruit fl ies (Diptera: Tephritidae). Journal of Eco-nomic Entomology 88, 1307–1315.
Heath, R.R., Epsky, N.D., Dueben, B.D., Rizo, J. and Jerónimo, F. (1997) Adding methyl substitute ammonia derivates to food-based synthetic attractant on capture of the Mediterranean and Mexican fruit fl ies (Diptera: Tephritidae). Journal of Economic Entomology 90, 1584–1589.
Hendrichs, J. (1996) Action programs against fruit fl ies of economic importance: session overview. In: McPheron, B. and Steck, G. (eds) Fruit Fly Pests: a World Assessment of their Biology and Management. St Lucie Press, Delray Beach, Florida, pp. 513–519.
Hendrichs, J. (2001) Use of the Sterile Insect Technique against Key Insect Pests. Food and Agriculture Organization (FAO)/International Atomic Energy Agency Division

Pests 355
of Nuclear techniques in Food and Agriculture. Available at: http://www.sustdev.org/journals/edition.02/download/sdi2_2_2.pdf (accessed 20 July 2007).
Hennesey, M. and Schnell, R.J. (2001) Resistance of immature mango fruits to Caribbe-an fruit fl y (Diptera: Tephritidae). Florida Entomologist 84, 318–319.
Hernández-Sánchez, G., Sanz-Barbosa, I., Casaña-Giner, V. and Primo-Yúfera, E. (2001) Attractiveness for Ceratitis capitata (Wiedemann) (Dipt., Tephritidae) of mango (Mangifera indica, cv. Tommy Atkins) airborne terpenes. Journal of Applied Ento-mology 125, 189–192.
Herrera, A.J. and Viñas, L.E. (1977) Fruit fl ies (Diptera: Tephritidae) on mangoes in Chu-lucanas, Piura. Revista Peruana de Entomoogia 20, 107–114.
Hill, D. (1975) Agricultural Insect Pests of the Tropics and their Control. Cambridge University Press, New York.
Hollingsworth, R.G., Drew, R., Allwood, A.J., Roming, M., Vagalo, M. and Tsatsia, F. (2003) Host plants and relative abundance of fruit fl y (Diptera: Tephritidae) species in the Solomon Islands. Australian Journal of Entomology 42, 95–108.
Hsu, J.-C. and Feng, H.-T. (2006) Development of resistance to Sinosad in oriental fruit fl y (Diptera: Tephritidae) in laboratory selection and cross-resistance. Journal of Economic Entomology 99, 931–936.
Huang, T.-Ch., Tze-chung, H., Cheng, E.Y., Kao, Ch., Hwang, Y. and Chiang, M. (2006) Area-wide Control of the Oriental Fruit Fly and Melon Fly in Taiwan. Available at: www.agnet.org/activities/sw/2006/954198798/paper-868546184.pdf (accessed 20 July 2007).
Husain, M.A. and Pruthi, H.S. (1924) A short note on the life history of the mango hop-per (Idiocerus spp.) in the Punjab. In: Report Proceedings V of the Entomology Meeting, Pusa, pp. 252–260.
Islam, M.S. and Elegio, D.T. (1997) Effect of time and frequency of insecticide spraying on the control of mango leafhoppers (Idioscopus spp.). Bangladesh Journal of Ento-mology 7, 93–99.
Iwahashi, O. (1984) The control project of the oriental fruit fl y in Okinawa. ChineseJournal of Entomology 4, 107–120. (in Chinese)
Jadhav, V. and Dalvi, C. (1987) Bionomics of mango nursery leaf eating caterpillar. Jour-nal of Maharashtra Agricultural University 12, 384–385.
Jeppson, L., Keiffer, H. and Baker, E. (1975) Mites Injurious to Economic Plants. Univer-sity of California Press, Berkeley, California.
Jhala, R.C., Patel, Z. and Shah, A. (1987) Studies on the relative occurrence of the leaf-gall midge (Procontarinia matteiana Keiffer and Cecconi) on different varieties of mango in south Gujarat, India. Tropical Pest Management 33, 277–279.
Jhala, R.C., Shah, A.H., Patel, Z.P. and Patel, R.L. (1989) Studies on population dynamics of mango hopper and scope of spraying in integrated pest management programme. Acta Horticulturae 231, 597–601.
Jirón, L.F. (1993) Plagas del follaje y de paniculas asociadas con el mango en Costa Rica. In: Memorias Ier Seminario Nacional de Mango. Punta Arenas, Chile, pp. 22–26.
Jirón, L.F. (1995) Opciones al uso de insecticidas en mango. In: Garcia, J., Fuentes, G. and Monge, J. (eds) Opciones al uso Unilateral de Plaguicidas en Costa Rica. Vol. 2. Editorial Euned, San José, Costa Rica, pp. 129–155.
Jirón, L.F. and Hedström, I. (1988) Occurrence of fruit fl ies of the genera Anastrepha and Ceratitis (Diptera: Tephritidae), and their host plant availability. Florida Entomolo-gist 71, 62–73.
Jirón, L.F. and Hedström, I. (1991) Population fl uctuations of economic species of Anas-trepha (Diptera: Tephritidae) related to mango fruiting phenology in Costa Rica.Florida Entomologist 74, 98–105.

J.E. Peña et al.356
Jirón, L.F. and Soto-Manitiu, J. (1987) Las Moscas de las frutas (Diptera: Tephritidae) en Costa Rica: situacion actual, analisis y comentario. Agronomia Costarricense 11, 255–261.
Joel, D.M. (1980) Resin ducts in the mango fruit: a defense system. Journal of Experimen-tal Botany 30, 1707–1718.
Johnson, P.J. (1989) Mango seed weevil (Sternochetus mangiferae F.): identifi cation, biology and management. University of Idaho Pip Tip 6, 3–7.
Joubert, P.H., Daneel, M.S. and Grove, T. (2000) Progress towards integrated pest man-agement (ipm) on mangoes in South Africa. Acta Horticulturae 509, 811–817.
Joubert, P., Grove, T., de Beer, M.S. and Steyn, W.P. (2004) Evaluation of kaolin (Surround WP) in an ipm program on mangoes in South Africa. Acta Horticulturae 645, 493–499.
Keifer, H.H., Baker, E.W., Kono, T., Delfi nado, M. and Styer, W. (1982) An illustrated guide to plant abnormalities caused by Eriophyid mites in North America. US De-partment of Agriculture, Agriculture Handbook No. 573, 74–75.
Kfi r, R. and Rosen, D. (1980) Parasites of soft scales (Homoptera: Coccidae) in Israel: an annotated list. Journal of the Entomological Society of South Africa 43, 113–128.
Khan, M.A., Ashfaq, M., Khaliq, A. and Ahmad, A. (2002) Effect of pheromone traps on the population capture of fruit fl y species and their infestation in mango orchards at Multan. Pakistan Entomologist 24, 153–157.
Khan, M.A., Ashfaq, M., Akram, W. and Lee, J.-J. (2005) Management of fruit fl ies (Dip-tera: Tephritidae) of the most perishable fruits. Entomological Research 35, 79–84.
Khanzada, A.G. and Naqvi, K.M. (1985) The optimum time for the control of mango hopper Idioscopus spp. (Hemiptera: Cicadellidae). Proceedings of the Entomologi-cal Society of Karachi 14/15, 149–156.
King, J.R. and Hennessey, M.K. (1996) Spinosad bait for the Caribbean fruit fl y (Diptera: Tephritidae). Florida Entomologist 79, 526–531.
Kring, T.J., Yearian, W.C. and Steward, V.B. (1987) Pleuroprucha insulsaria (Lepidoptera: Geometridae): a potential pest of grain sorghum. Journal of the Kansas Entomo-logical Society 60, 476–477.
Kudagamage, C., de Z. Rajapakse, H.L., Rajapakse, R.H. and Ratnasekara, D. (2001) The population dynamics and insecticidal control of three leaf hoppers, Amrytodus brevis-tilus Leth., Idioscopus niveosparus Leth., and Idioscopus clipealis Leth. (Homoptera: Cicadellidae) in mango. Journal of Entomological Research 25, 121–125.
Kumar, S., Rai, A., Patel, C. and Bhatl, R. (1993) Studies on spatial dsitribution of Amri-todes atckinsoni (Leth.) infesting mango in south Gujarat, India. (Abstract). IV Inter-national Mango Symposium, p. 122.
Labuschagne, T.I. (1993) Progress with integrated control of the mango scale Aulacaspis tubercularis Newstead. South African Mango Growers’ Association Yearbook 13, 134–135.
Labuschagne, T.I., Steyn, W.P. and De Beer, M.S. (1995) Voorkoms en moontlike alterna-tive beheermetodes van vrugtevliee opp mango’s. South African Mango Growers’ Association Yearbook 15, 102–105.
Lakra, R.K., Kharub, W. and Singh, Z. (1980) Pest management systems for the mango mealybug Drosicha mangifera Green – a polyphagous pest of fruit trees in Haryan. Indian Journal of Entomology 42, 153–165.
Laroussilhe, F. de (1980) Le Manguier. G.P. Maisonneuve and Larouse, Paris.Leblanc, L. and Allwood, A.J. (1997) Mango fruit fl y (Bactrocera frauenfeldi): why so many
in federated states in Micronesia? In: Allwood, A.J. and Drew, R.A.I. (eds) Manage-ment of Fruit Flies in the Pacifi c. Australian Centre for International Agricultural Research (ACIAR) Proceedings No. 76. ACIAR, Canberra, Australia, pp. 125–130.
Leblanc, L., Leweniqila, L., Tau, D., Tumukon, T., Kassim, A. and Hollingsworth, R. (1997) Can fruit fl ies be controlled in a village with a mixed orchard? Pacifi c Island

Pests 357
experiences. In: Allwood, A.J. and Drew, R.A.I. (eds) Management of Fruit Flies in the Pacifi c. Australian Centre for International Agricultural Research (ACIAR) Pro-ceedings No. 76. ACIAR, Canberra, Australia, pp.187–191.
Leefmans, S. and van der Vecht, J. (1930) The red-ringed mango caterpillar (in Dutch, English summary). Korte Mededeelingen van het lustituut voor Plantenziekten, 14 pp.
Leyva, J.L., Browning, H. and Gilstrap, F. (1991) Development of Anastrepha ludens (Dip-tera: Tephritidae) in several host fruit. Environmental Entomology 20, 1160–1165.
Lezama-Gutierrez, R., Trujillo, A., Molina-Ochoa, J., Rebolledo-Dominguez, O., Pesca-dor, A., Lopez-Edwards, M. and Aluja, M. (2000) Virulence of Metarrizium anisopliae(Deuteromycotina: Hypomycetes) on Anastrepha ludens (Diptera: Tephritidae): labo-ratory and fi eld trials. Journal of Economic Entomology 93, 1080–1084.
Li, L.Y., Wang, R. and Waterhouse, D.F. (1997) The distribution and importance of major insect pests and weeds of agriculture and plantation forests in southern China. Chinese Academy of Agricultural Sciences and ACIAR, 185 pp.
Liburd, O.E., Holler, T.C. and Moses, A.L. (2004) Toxicity of Imidacloprid-treated spheres to Caribbean fruit fl y, Anastrepha suspensa (Diptera: Tephritidae) and its parasitoid Diachasmimorpha longicaudata (Hymenoptera: Braconidae) in the laboratory. Journal of Economic Entomology 97, 525–529.
Lindegren, J. and Vail, P. (1986) Susceptibility of Mediterranean fruit fl y, melon fl y and oriental fruit fl y (Diptera: Tephritidae) to the entomogenous nematode Steinernemafeltiae in laboratory tests. Environmental Entomology 4, 465–468.
Lindgreen, S., Borden, J., Gray, D., Lee, P., Palmer, D. and Chong, L. (1982) Evaluation of two trap log techniques for ambrosia beetles (Coleoptera: Scolytidae) in timber processing areas. Journal of Economic Entomology 75, 577–586.
Loomans, A.J.M., van Lenteren, J.C., Tommasini, M.G., Maini, S. and Riudavets, J. (1995) Biological control of thrips pests. Wageningen Agricultural University Papers No. 95.
López, F., Chambers, D., Sanchez, M. and Kamasaki, H. (1969) Control of the Mexican fruit fl y by bait sprays concentrated at discrete locations. Journal of Economic Ento-mology 62, 1255–1257.
López, M., Aluja, M. and Sivinski, J. (1999) Hymenopterous larval-pupal and pupal parasitoids of Anastrepha fl ies (Diptera: Tephritidae) in Mexico. Biological Control15, 119–129.
Love, K., Bowen, R. and Paull, R. (2003) Increasing marketable production of exotic trop-ical fruit with protective covering. Western Region, Sustainable Agriculture Research and Education, Project Report, Farmer/Rancher, Sustainable Agriculture Research and Education, Utah State University, Captain Cook, Hawaii, USA.
Lux, S.A., Ekesi, S., Dimbi, S., Mohamed, S. and Billah, M. (2003) Mango-infesting fruit fl ies in Africa: perspectives and limitations of biological approaches to their man-agement. In: Neuenschwander, P., Borgemeister, C. and Langewald, J. (eds) Bio-logical Control in IPM Systems in Africa. CAB International, Wallingford, UK, pp. 277–293.
Malavasi, A. and Zucchi, R.A. (eds) (2000) Moscas-Das-Frutas de Importância Econômica no Brasil: Conhecimento Básico e Aplicado. Holos Editora, Ribeirão Preto, Brazil.
Mangan, R.L. and Hallman, G.H. (1998) Temperature treatments for quarantine security: new approaches for fresh commodities. In: Hallman, G.H. and Denlinger, D.L. (eds) Temperature Sensitivity in Insects and Application in Integrated Pest Manage-ment. Westview Press, Boulder, Colorado, pp. 201–234.
Mangan, R.L. and Moreno, D.S. (2007) Development of bait stations for fruit fl y popula-tion suppression. Journal of Economic Entomology 100, 440–450.
Mangan, R.L. and Sharp, J.L. (1994) Combination treatments. In: Sharp, J.L. and Hall-man, G.H. (eds) Quarantine Treatments for Pests of Food Plants. Westview Press, Boulder, Colorado, pp. 239–247.

J.E. Peña et al.358
Mangan, R.L., Moreno, D.S. and Thompson, G.D. (2006) Bait dilution, spinosad concen-tration, and effi cacy of GF-120 based fruit fl y sprays. Crop Protection 25, 125–133.
Martorell, L.F. (1975) Annotated Food Plant Catalog of the Insects of Puerto Rico. Agri-cultural Experiment Station, University of Puerto Rico, Río Piedras, Puerto Rico.
McQuate, G.T., Peck, S.L., Barr, P.G. and Sylva, C.D. (2005) Comparative evaluation of spinosad and phloxine B as toxicants in protein baits for suppression of three fruit fl y (Diptera: Tephritidae) species. Journal of Economic Entomology 98, 1170–1178.
Messing, R. (1999) Managing Fruit Flies on Farms in Hawaii. Cooperative Extension Service, College of Tropical Agriculture and Human Resources, University of Hawaii at Manoa, Hawaii.
Miller, D.R. and Davidson, J. (2005) Armored Scale Insect Pests of Trees and Shrubs (Hemiptera: Diaspididae). Comstock Publishing Associates, Ithaca, New York.
Mohyuddin, A.I. (1981) A review of biological control in Pakistan. In: Proceedings of the Second Pakistan Congress of Zoology, Tandojam, Pakistan, pp. 31–79.
Mohyuddin, A.I. and Mahmood, R. (1993) Integrated control of mango pests in Pakistan. Acta Horticulturae 341, 467–483.
Montoya, P., Liedo, P., Benrey, B., Cancino, J., Barrera, J.F., Sivinski, J. and Aluja, M. (2000) Biological control of Anastrepha spp. (Diptera: Tephritidae) in mango or-chards through augmentative releases of Diachasmimorpha longicaudata (Ash-mead) (Hymenoptera: Braconidae). Biological Control 18, 216–224.
Moore, D. and Cross, A.E. (1992) Competition between two primary parasitoids, Gyranu-soidea tebygi Noyes and Anagyrus mangicola Noyes, attacking the mealybug Ras-trococcus invadens Williams and the infl uence of a hyperparasitoid Chartocerus hyalipennis Hayat. Biocontrol Science and Technology 2, 225–234.
Moore, D. and Cross, A.E. (1993) Biological control of the fruit tree mealybug, Rastro-coccus invadens Williams; single or multiple introduction. Acta Horticulturae 341, 433–440.
Morin, C. (1967) Cultivo de Frutales Tropicales. Librerias ABC, Lima, Peru, pp. 225–229.Moznette, G. (1922) Insects injurious to the mango in Florida and how to combat them.
Farmers Bulletin 1257.Murray, R.C. (1991) Arthropod pests of mango in the Caribbean Islands. Review of Crop
Protection. University of the West Indies, Kingston, Jamaica.Nachiappan, R.M. and Baskaran, P. (1986) Field evaluation of certain insecticidal sprays
against mango leaf-hoppers. Pesticides 20, 41–44.Nafus, D. (1991) Biological control of Penicilllaria jocosatrix (Lepidoptera: Noctuidae)
on mango on Guam with notes on the biology of its parasitoids. Environmental Entomology 20, 1725–1731.
Narasinham, M. (1959) Control of mango malformation disease. Current Science28, 254.
Narasimham, U.A. and Chacko, M.J. (1991) The distribution of some Rastrococcus spp., (Homoptera: Pseudococcidae) on mango in India. Bulletin of Entomological Research81, 445–448.
Nascimento, A., Malavasi, A., Morgante, J. and Duarte, A.L. (1992) Hot-water immer-sion treatment for mango infested with Anastrepha fraterculus, A. obliqua, and Ceratitis capitata (Diptera: Tephritidae) in Brazil. Journal of Economic Entomology85, 456–460.
Nascimento do, A.S., da Silva Carvalho, da Costa Mendonca, M. and Braga, R. (2002) Pragas e seu controle. In: de Carvalho, P.J. and de Queiroz Pinto, A.C. (eds) A Cultura da Mangueira. Embrapa Informação Tecnológica, Brasília DF, Brazil, pp. 279–297.
Nguyen, T.N., Nguyen, V. and Le van, D. (1998) Insect pests on citrus, durian, longan and mango in selected provinces of the Meking Delta of Vietnam. In: Van Mele, P. and Nguyen, V.H. (eds) Proceedings of the First Symposium on Fruit Production in

Pests 359
the Mekong Delta Focusing on Integrated Pest Management. Cautho, Vietnam, pp. 71–76 (in Vietnamese).
Norrbom, A.L. (2004) Host Plant Database for Anastrepha and Toxotrypana (Diptera: Tephritidae: Toxotrypanini). Diptera Data Dissemination Disk (CD-ROM), North American Dipterist’s Society, Washington, DC.
Ochoa, R., Aguilar, H. and Vargas, C. (1994) Phytophagous Mites of Central America: an Illustrated Guide. Centro Agronomico de Investigacion y Enseñanza Technical Ser-vice 6, Turrialba, Costa Rica.
Orankanok, W., Chinvinijkul, S., Thanaphum, S., Sutantawong, M. and Enkerlin, W.R. (2007) Area-wide integrated control of oriental fruit fl y (Bactrocera dorsalis, Hen-del) and Guava fruit fl y (B. correcta, Bezzii) in Thailand using the sterile insect technique (SIT). In: Vreysen, M.J.B., Robinson, A.S. and Hendrichs, J. (eds) Area-wide Control of Insect Pests: From Research to Field Experimentation. Springer, Dordrecht, the Netherlands, pp. 517–526.
Osman, A.A. (1979) Notes on the control of the bud mite Aceria mangiferae (Sayed) in Egypt (Acarine: Eriophyidae). Bulletin of the Entomological Society of Egypt Eco-nomic Services 9, 119–126.
Ovruski, S., Aluja, M., Sivinski, J. and Wharton, R. (2000) Hymenopteran parasitoids on fruit infesting Tephritidae (Diptera) in Latin America and the southern United States: diversity, distribution, taxonomic status and their use in fruit fl y biological control. Integrated Pest Management Reviews 5, 81–107.
Paderes, A. and Orden, M.E. (2004) Factors Determinating the Adoption of Pruning and Bagging by Major Mango Producers in the Philippines. Available at: http://www.kmitl.ac.th/science/Journal/jan2004/MANGOAdopt.pdf (accessed 12 March 2005).
Papadopoulos, N.K., Carey, J.R., Katsoyannos, B.I. and Kouloussis, N.A. (1996) Over-wintering of the Mediterranean fruit fl y (Diptera: Tephritidae) in Northern Greece. Annals of the Entomological Society of America 89, 526–534.
Patel, R.K., Patel, S.R. and Shah, S. (1977) Biology of mango hopper, Amritodus atkin-soni (Leth.) (Jassidae: Hemiptera) in South Gujarat. Indian Journal of Entomology37, 150–153.
Peck, S.L. and McQuate, G.T. (2000) Field tests of environmentally friendly malathion replacements to suppress wild Mediterranean fruit fl y (Diptera: Tephritidae) popula-tions. Journal of Economic Entomology 93, 280–289.
Peña, J.E. (1993) Pests of mango in Florida. Acta Horticulturae 341, 395–406.Peña, J.E. and Duncan, R. (1992) Sampling methods for Prodiplosis longifi la (Diptera:
Cecidomyiidae) in limes. Environmental Entomology 21, 996–1001.Peña, J.E., Duncan, R., Vasquez, T. and Hennessey, M. (1999) Guava arthropod season-
ality and control of fruit fl ies in south Florida. Proceedings of the Florida State Horticultural Society 112, 206–209.
Peña, J.E., Palevsky, E., Otero-Colina, G., Ochoa, R. and Meister, C.W. (2005) Mango bud mite, Aceria mangiferae bionomics and control under Florida conditions. Pro-ceedings of the Florida State Horticultural Society 118, 228–234.
Peña, J.E., Duncan, R. and Meister, C. (2006a) Chemical control of fl owering thrips in-festing tropical fruits. Proceedings of the Florida State Horticultural Society 119, 21–24.
Peña, J.E., Gould, W., Hennessey, M.K., Hallman, G. and Crane, J.H. (2006b) Labora-tory and fi eld infestation studies on immature green ‘Tommy Atkins’ and ‘Keitt’ mangoes to determine host status to the Caribbean fruit fl y (Diptera: Tephritidae). Proceedings of the Florida State Horticultural Society 119, 16–20.
Peng, R.K. and Christian, K. (2004) The weaver ant, Oecophylla smaragdina (Hy-menoptera: Formicidae), an effective biological control agent of the red banded thrips

J.E. Peña et al.360
Selenothrips rubrocinctus (Thysanoptera: Thripidae) in mango crops in the Northern Territory of Australia. International Journal of Pest Management 50, 107–114.
Peng, R.K. and Christian, K. (2005a) The control effi cacy of the weaver ant, Oecophylla smaragdina (Hymenoptera: Formicidae), on the mango leafhopper, Idioscopus niti-dulus (Hemiptera: Cicadellidae) in mango orchards in the Northern Territory. Inter-national Journal of Pest Management 51, 297–304.
Peng, R. and Christian, K. (2005b) Integrated pest management in mango orchards in the Northern Territory Australia, using the weaver ant Oecophylla smaragdina (Hymenoptera: Formicidae) as a key element. International Journal of Pest Management 51, 149–155.
Peng, R.K. and Christian, K. (2006) Effective control of Jarvis’s fruit fl y Bactrocera jarvisi(Diptera: Tephritidae), by weaver ant, Oecophyla smaragdina (Hymenoptera: For-micidae), in mango orchards in the Northern Territory of Australia. InternationalJournal of Pest Management 52, 275–282.
Piñero, J., Aluja, M., Vázquez, A., Equihua, M. and Varón, J. (2003) Human urine and chicken feces as fruit fl y (Diptera: Tephritidae) attractants for resource-poor fruit growers. Journal of Economic Entomology 96, 334–340.
Poinar, G.O., Jr and Hislop, R.G. (1981) Mortality of Mediterranean fruit fl y adults (Cer-atitis capitata) from parasitic nematodes (Neoaplectana and Heterorhabditis spp.). Institute for Research and Cognitive Science, Medical Science. Microbiology, Para-sitology and Infectious Disease 9, 641.
Prasad, S.N. (1971) The Mango Midge Pests. Cecidological Society, Allahabad, India.Prasad, V. and Singh, R.K. (1976) Prevalence and control of mango mealy bug Drosicha
stebbingi (Green) in Bihar. Indian Journal of Entomology 38, 214–224.Rahman, K.A. (1940) Important insect pests of the mango and how to control them.
Punjab Fruit Journal 3, 6.Rai, B., Verma, S. and Kumar, K. (1966) Evaluation of pesticides for control of mango
bud mite, Aceria mangiferae Sayed (Acarina: Eriophyidae). Indian Journal of Ento-mology 28, 176–180.
Rai, B., Shah, A. and Patel, R. (1988) Biology of Oligonychus mangiferus (Tetranychidae: Acarina), a pest of mango in Gujarat. Gujarat Agricultural University Research Journal14, 5–10.
Rebouças, A., Bôas, I.V., Martins, J. and Magalhães, O. (1996) Manga, Tecnologia de Produção e Mercado. Universidade Estadual do Sudoeste da Bahia, Vitória da Conquista, Brazil.
Robacker, D.C. (2001) Roles of putrescine and 1-pyrroline in attractiveness of technical-grade putrescine to the Mexican fruit fl y (Diptera: Tephritidae). Florida Entomologist 84, 679–685.
Robacker, D.C. and Warfi eld, W.C. (1993) Attraction of both sexes of Mexican fruit fl y, Anastrepha ludens, to a mixture of ammonia, methylamine, and putrescine. Journalof Chemical Ecology 19, 2999–3016.
Robacker, D.C., Martínez, A.J., García, J.A., Díaz, M. and Romero, C. (1996) Toxicity of Bacillus thuringiensis to Mexican fruit fl y (Diptera: Tephritidae). Journal of Eco-nomic Entomology 89, 104–110.
Robacker, D.C., DeMilo, A.B. and Voaden, D.J. (1997) Mexican fruit fl y attractants: ef-fects of 1-pyrroline and other amines on attractiveness of a mixture of ammonia, methylamine, and putrescine. Journal of Chemical Ecology 23, 1263–1280.
Robinson, A.S. and Hooper, G. (eds) (1989) Fruit Flies. Their Biology, Natural Enemies and Control. Volumes A and B. Elsevier, Amsterdam, the Netherlands.
Rocha da Silva, C. (2006) Comportamento e comunicaçao quimica da broca da manguei-ra (Hypocryphalus mangiferae Stebb) (Coleoptera: Curculionidae: Scolytinae). Uni-versidade Estadual do Norte Fluminense, Camposdos Goytacazes, Rio do Janeiro, Theses, 59 pp.

Pests 361
Rogers, L., French, J. and Elgar, M. (1999) Suppression of plant growth on the mounds of the termite Coptotermes lacteus Froggatt (Isoptera: Rhinotermitidae). Insects Socieux46, 366–371.
Rubin, A. and Kuslitizky, W. (1992) First record of Ceranisus menes (Hymenoptera: Eulophidae) in Israel. Phytoparasitica 20, 123–124.
Sadana, G.L. and Kumari, M. (1991) Predatory potential of the lyssomanid spider, Lys-somanes sikkimeri Tikader on the mango hopper, Ideoscopus clipealis (Lethierry).Entomon 16, 283–285.
Said Sajap, A., Amit, S. and Welker, J. (2000) Evaluation of hexafl umuron for controlling the subterranean termite Coptotrermes curvignathus (Isoptera: Rhinotermitidae) in Malaysia. Journal of Economic Entomology 93, 429–433.
Sakimura, K. (1981) A review of Frankliniella bruneri Watson and description of F. kel-liae, n. sp. (Thysanoptera: Thripidae). Florida Entomologist 64, 483–490.
Sankaran, T. and Mjeni, A.M. (1989) Recent studies on the mango leaf-midge Procon-tarinia matteiana Kieffer and Cecconi (Diptera: Cecidomyiidae) and its parasites in India and on prospects for biological control of the pest in Oman. Acta Horticultu-rae 231, 587–592.
Sayed, M. (1946) Aceria mangiferae. Bulletin of the Society of Entomology 30, 7–10.Schaefer, C.W. and Ahmad, I. (2000) Cotton strainers and their relatives (Pyrrhocoroi-
dea: Pyrrhocoridae and Largidae). In: Schaefer, C.W. and Panizzi, A.R. (eds) Het-eroptera of Economic Importance. CRC Press, Boca Raton, Florida, pp. 271–307.
Schotman, L. (1989) FAO/RLAC Plant Quarantine Programme. Data sheet on mango seed weevil. FAO Accession XF8988239, 23 pp. http://www.fao.org/agris/search (accessed 1 September 2008).
Schreiner, I. (1987) Mango shoot caterpillar control on mango fl owers, 1985. Insecticide and Acaricide Tests 12, 94.
Schreiner, I. (1991) Procontarinia sp. nov.: a mango pest long misdiagnosed as anthra-cnose. In: Agricultural Development American Pacifi c, Crop Protection Conference Proceedings 1989, Hawaii Institute Tropical Agriculture and Human Resources, Research Extension Series 134, 53–55.
Secretariat of the Pacifi c Community (2005) Fruit Flies in Palau. Pest Advisory Leafl et No. 44, Palau, Pacifi c Islands.
Shah, A.H., Jhala, R.C., Patel, G.M. and Patel, S.H. (1983) Evaluation of effective dose of monocrotophos for the control of mango hopper Amritodus atkinsoni Lethierry (Cicadellidae: Homoptera) by injection method and its comparison with foliar application. Gujarat Agricultural University Research Journal 9, 14–18.
Sharp, J.L., Ouye, M.T., Thalman, R., Hart, W.G., Ingle, S.J. and Chew, V. (1988) Submer-sion of ‘Francis’ mango in hot water as a quarantine treatment for the West Indian fruit fl y and the Caribbean fruit fl y (Diptera: Tephritidae). Journal of Economic Ento-mology 81, 1431–1436.
Sharp, J.L., Ouye, M.T., Ingle, S.J., Hart, W.G., Enkerlin, E., Celedonio, H., Toledo, J., Stevens, L., Quintero, E., Reyes, J. and Schwarz, A. (1989a) Hot-water quarantine treatment for mangoes from the State of Chiapas, Mexico, infested with Mediterra-nean fruit fl y and Anastrepha serpentina (Wiedemann) (Diptera: Tephritidae). Jour-nal of Economic Entomology 82, 1663–1666.
Sharp, J.L., Ouye, M.T., Hart, W.G., Ingle, S.J., Hallman, G., Gould, W. and Chew, V. (1989b) Immersion of Florida mangoes in hot water as a quarantine treatment for Caribbean fruit fl y (Diptera: Tephritidae). Journal of Economic Entomology 82, 186–188.
Sharp, J.L., Ouye, M.T., Ingle, S.J. and Hart, W.G. (1989c) Hot water quarantine treat-ment for mangoes from Mexico infested with Mexican fruit fl y and West Indian fruit fl y (Diptera: Tephritidae). Journal of Economic Entomology 82, 1657–1662.

J.E. Peña et al.362
Shaw, J. (1961) Airplane applications of malathion bait spray for Mexican fruit fl y con-trol. Journal of Economic Entomology 54, 600–601.
Shaw, J. and Spisshakoff, L.M. (1958) Insecticidas para el control de la mosca Mexicana de la fruta en citricos y mango. Memorias Primer Congreso Naciona de Entomolo-gia Fitopatologia Escuela Nacional Agricola Chapingo. Escuela Nacional Agricola de Chapingo, Texcoco, Mexico, pp. 205–217.
Sheikh, A.D., Bashir, A. and Jilani, G. (2008) Incidence and Effects of Fruit Fly on Mango Quality in Punjab. Technology and Transfer Institute, Faisalabad, 3 pp.
Shellie, K.C. and Mangan, R.L. (2002a) Hot water immersion as a quarantine treatment for large mangoes: artifi cial versus cage infestation. Journal of the American Society for Horticultural Science 127, 430–434.
Shellie, K.C. and Mangan, R.L. (2002b) Cooling method and fruit weight: effi cacy of hot water quarantine treatment for Mexican fruit fl y (Diptera: Tephritidae) in mango. HortScience 37, 910–913.
Shukla, R.P. and Prassad, V.G. (1984) Evaluation of some insecticides against mango hopper, Idioscopus clypealis (Lethierry)(Homoptera: Cicadellidae). Indian Journal of Agricultural Science 54, 677–681.
Shukla, R.P. and Tandon, P.L. (1985) Bio-ecology and management of the mango weevil, Sternochetus mangiferae (Fabricius) (Coleoptera: Curculionidae). InternationalJournal of Tropical Agriculture 3, 293–303.
Shukla, R.P., Tandon, P.L. and Singh, S.J. (1984) Baculovirus – a new pathogen of mango nut weevil, Sternochetus mangiferae (Fabricius) (Coleoptera: Curculionidae). Cur-rent Science 53, 593–599.
Shukla, R.P., Tandon, P.L. and Suman, C.L. (1988) Intra-tree distribution of eggs and dia-pausing adults of the stone weevil. Acta Horticulturae 231, 566–570.
Silva, Q.M. and Cavalcante, R.D. (1977) Vinsonia stellifera (Westwood 1871), (Hom. Coccidae), on various plants in the state of Ceará, Brazil. Fitossanidade 2,25–26.
Silveira, da, R. (1960) Contribucao ao studo do Hypocryphalus mangiferae (Coleoptera: Scolytidae). PhD thesis, Escuela Agricultura de São Paulo, São Paulo, Brazil.
Singh, G. (1991) Loss assessment, ecology and management of mango fruit fl y, Dacussp. Acta Horticulturae 291, 425–436.
Singh, S.K. and Singh, G.B. (2003) Management of termite infestation in mango orchard by cultural practices and organic amendments. Indian Journal of Agricultural Re-search 37, 148–150.
Sivinski, J.M. (1996) The past and potential of biological control of fruit fl ies. In: McPher-on, B.A. and Steck, G.J. (eds) Fruit Fly Pests. A World Assessment of their Biology and Management. St Lucie Press, Delray Beach, Florida, pp. 365–375.
Sivinski, J.M., Calkins, C.O., Baranowski, R.M., Harris, D., Brambila, J., Diaz, J., Bums, R.E., Holler, T. and Dodson, D. (1996) Suppression of Caribbean fruit fl y (Anas-trepha suspensa (Loew) Diptera: Tephritidae) population through releases of the parasitoid Diachasmimorpha longicaudata (Ashmead) (Hymenoptera: Braconidae). Biological Control 6, 177–185.
Sivinski, J., Aluja, M. and López, M. (1997) Spatial and temporal distributions of parasi-toids of Mexican Anastrepha species (Diptera: Tephritidae) within the canopies of fruit trees. Annals of the Entomological Society of America 90, 604–618.
Sivinski, J., Piñero, J. and Aluja, M. (2000) The distributions of parasitoids (Hymenoptera) of Anastrepha fruit fl ies (Diptera: Tephritidae) along an altitudinal gradient in Vera-cruz, Mexico. Biological Control 18, 258–269.
Sohi, A.S. and Sohi, A.S. (1990) Mango leafhoppers (Homoptera: Cicadellidae) – a review. Journal of Insect Science 3, 1–12.

Pests 363
Soto-Manitiú, J., Jirón, L. and Hernández, R. (1987) Chemical control and ecological observations of fruit fl ies of the genus Anastrepha Schiner (Diptera: Tephritidae) on mango. Turrialba 37, 245–251.
Srivastava, R.P. (1981) Comparative effi cacy of various insecticidal dusts against mango mealybug eggs. Indian Journal of Entomology 43, 225–229.
Srivastava, R.P. (1998) Pests of mango. In: Srivastava, R.P. (ed.) Mango Cultivation. Inter-national Bood Distributing, Charbagh, Lucknow, India, pp.175–299.
Srivastava, R. and Tandon, P. (1986) Natural occurrence of two entomogenous fungi pathogenic to mango hopper, Idioscopus clypealis Leth. Indian Journal of Plant Pathology 4, 121–123.
Srivastava, S. and Singh, N.B. (2004) Survey of termites infesting mango orchards at four locations in Bareilly, Uttar Pradesh. Biological Memoirs 30, 48–51.
Stark, J.D., Vargas, R. and Miller, N. (2004) Toxicity of spinosad in protein bait to three economically important tephritid fruit fl y species (Diptera: Tephritidae) and their par-asitoids (Hymenoptera: Braconidae). Journal of Economic Entomology 97, 911–915.
Steck, G.J. (2003) Mango Fruit Fly, Marula Fruit Fly, Ceratitis cosyra (Walker) (Insecta: Dip-tera: Tephritidae). Available at: http://edis.ifas.ufl .edu/IN563 (accessed 20 July 2007).
Steiner, L.F. and Lee, R.K. (1955) Large area tests of male annihilation method for orien-tal fruitfl y control. Journal of Economic Entomology 48, 311–317.
Steiner, L.F., Mitchell, W.C., Harris, E., Kozuma, T. and Fujimoto, M. (1965) Oriental fruit fl y eradication by male annihilation. Journal of Economic Entomology 58, 961–964.
Subba Rao, B.R. (1986) Mangoma spinidorsum gen. et sp. n. (Hymneoptera: Eurytomi-dae) associated with leaf galls. Bulletin of Entomological Research 76, 389–392.
Subramanian, C.K. (1925) A note on the life history of Cryptorhynchus mangiferae Fab. Madras Agricultural Department Yearbook. Madras Agricultural Department, Ma-dras, India, pp. 29–36.
Summanwar, A. and Raychoudhury, S. (1968) The role of the eriophyid mite (Aceria mangiferae) in the causation of mango malformation. Indian Phytopathology 21, 463–464.
Sweet, M.H. (2000) Seed and chinch bugs (Lygaeoidea). In: Schaefer, C.W. and Panizzi, A. (eds) Heteroptera of Economic Importance. CRC Press, Boca Raton, Florida, pp. 143–264.
Swirski, E., Wysoki, M. and Izhar, Y. (2002) Subtropical pests of Israel. Fruit Board of Israel and Institute of Plant Protection, The Volcani Center, Tel Aviv, 284 pp.
Syed, R.A., Ghani, M.A. and Murtaza, M. (1970) Studies on trypetids and their natural enemies in West Pakistan. III. Dacus (Strumeta) zonatus (Saunders). Technical Bul-letin of the Commonwealth Institute of Biological Control 13, 1–16.
Tan, K.-H. (2000) Area-Wide Control of Fruit Flies and Other Insect Pests. Penerbit Uni-versiti Sains Malaysia, Pulau Pinang, Malaysia.
Tandon, P.L. and Lai, B. (1979) Studies on the chemical control of the mango hopper, Idioscopus clypealis Leth. (Cicadellidae: Homoptera). International Pest Control21, 6–9.
Tandon, P.L. and Verghese, A. (1985) World List of Insect, Mite and Other Pests of Man-go. Technical Document of the Indian Institute for Horticultural Research 5. Indian Institute for Horticultural Research, Bangalore, India.
Tandon, P.L., Lal, B. and Rao, G.S. (1983) Prediction of mango hopper Idioscopus cly-pealis (Leth.) population in relation to physical environmental factors. Journal of Entomology 8, 257–261.
Tandon, P.L., Verghese, A. and Prasada, G.S. (1989) Spatial distribution, sampling plan and appropriate transformation of the mango hopper, Idioscopus niveoparsus (Lethi-erry) (Homoptera: Cicadellidae). Giornale Italiano di Entomologia 4, 235–242.

J.E. Peña et al.364
Tenakanai, D. (1997) Fruit fl y fauna in Papua New Guinea. In: Allwood, A.J. and Drew, R.A.I. (eds) Management of Fruit Flies in the Pacifi c. Australian Centre for Interna-tional Agricultural Research (ACIAR) Proceedings No. 76. ACIAR, Canberra, Austra-lia, pp. 87–94.
Thomas, P., Kannan, A., Degwekar, V.H. and Ramamurthy, M.S. (1995) Non-destructive detection of seed weevil-infested mango fruits by X-ray imaging. Post Harvest Biol-ogy and Technology 5, 161–165.
Thomson, F.C. (ed.) (2005) BioSystematic Database of World Diptera, Version 7.5. Avail-able at: http://www.diptera.org/biosys.htm (accessed 5 November 2007).
Tigvattnanont, S. (1988) Biological and autoecological studies of the mango leaf-cutting weevil, Deporaus marginatus Pascos (Coleoptera: Attelabidae). Khon Kaen Agricul-tural Journal 16, 51–62.
Toledo, J. and Lara, J. (1996) Comparison of the biology of Anastrepha obliqua reared in mango (Mangifera indica L.) and in mombin (Spondias mombin) infested under fi eld conditions. In: McPheron, B. and Steck, G. (eds) Fruit Fly Pests: a World Assess-ment of their Biology and Management. St Lucie Press, Delray Beach, Florida, pp. 359–362.
Toledo, J., Liedo, P., Williams, T. and Ibarra, J. (1999) Toxicity of Bacillus thuringiensisβ-exotoxin to three species of fruit fl ies (Diptera: Tephritidae). Journal of Economic Entomology 92, 1052–1056.
Toledo, J., Rasgado, M.A., Ibarra, J.E., Gómez, A., Liedo, P. and Williams, T. (2006) Infection of Anastrepha ludens following soil applications of Heterorhabditisbacteriophora in a mango orchard. Entomologia Experimentalis et Applicata 119, 155–162.
Tween, G. (1993) Fruit fl y control and eradication program management: factors infl u-encing action criteria and program design. In: Aluja, M. and Liedo, P. (eds) Fruit Flies: Biology and Management. Springer, New York, pp. 308–310.
Uechi, N., Kawamura, F., Tokuda, M. and Yukawa, J. (2002) A mango pest, Procon-tarinia mangicola (shi) comb. Nov. (Diptera: Cecidomyiidae), recently found in Okinawa, Japan. Applied Entomology and Zoology 37, 589–593.
Umeya, K. and Hirao, J. (1975) Attraction of the jackfruitfl y, Dacus umbrosus F. (Diptera: Tephritidae) and lace wing, Chrysopa sp. (Neuroptera: Chrysopidae) to lure traps baited with methyl eugenol and cuelure in the Philippines. Applied Entomology and Zoology 10, 60–62.
United States Department of Agriculture (USDA) (1981) Plant Pest News 1.van Dine, D.L. (1906) The mango weevil (Cryptorhynchus mangiferae Falor.). Hawaii
Agricultural Experiment Station Bulletin 17, 1–11.van Halteren, P. (1970) Notes on the biology of the scale insect Aulacaspis mangiferae
Newst. (Diaspididae: Hemiptera) on mango. Ghana Journal of Agricultural Science3, 83–85.
van Mele, P., Cuc, T.T. and van Huis, A. (2001) Farmer’s knowledge, perceptions and practices in mango pest management in the Mekong Delta, Vietnam. InternationalJournal of Pest Management 47, 7–16.
van Mele, P., Vayssières, J.-F., van Tellingen, E. and Vrolijks, J. (2007) Effects of an African weaver ant, Oecophylla longinoda, in controlling mango fruit fl ies (Diptera: Tephritidae) in Benin. Journal of Economic Entomology 100, 695–701.
van Whervin, L.W. (1968) Citrus Weevils of Jamaica and Some of their Parasites. Techni-cal Bulletin No. 1. Citrus Research Unit, University of the West Indies, St Augus-tine.
van Wyk, M., Al Adawi, A., Khan, I., Deadman, M., Al Jahwari, A., Wingfi eld, B., Ploetz, R. and Wingfi eld, M. (2007) Ceratocystis manginecans sp. nov., causal agent of a

Pests 365
destructive mango wilt disease in Oman and Pakistan. Fungal Diversity 27, 213–230.
Van Zyl, E., Kotze, J.M. and Steyn, P.L. (1988) Isolation of Xanthomonas campestris pv. Mangifera indica from gall fl y induced lesions on mango leaves. Phytophylactica20, 89–90.
Vannière, H., Didier, C., Rey, J.Y., Diallo, T.M., Kéita, S. and Sangaré, M. (2004) La mangue en Afrique de l’Ouest francophone: les systèmes de production et les iti-néraires techniques. Fruits 59, 383–398.
Vargas, R.I., Miller, N.W. and Prokopy, R.J. (2002) Attraction and feeding responses of Med-iterranean fruit fl y and a natural enemy to protein baits laced with two novel toxins, phloxine B and spinosad. Entomologia Experimentalis et Applicata 102, 273–282.
Varma, A., Lele, V.C., Raychoudhuri, S.P., Ram, A. and Sang, A. (1974) Mango malforma-tion: a fungal disease. Phytopathologische Zeitschrift 79, 254–257.
Vayssières, J.-F. and Kalabane, S. (2000) Inventory and fl uctuations of the catches of Dip-tera Tephritidae associated with mangoes in Coastal Guinea. Fruits 55, 259–270.
Vayssières, J.-F., Sanogo, F. and Noussourou, M. (2004) Inventaire des espèces de mouches des fruits (Diptera: Tephritidae) inféodées au manguier au Mali et essais de lutte raisonnée. Fruits 59, 3–16.
Vayssières, J.-F., Goergen, G., Lokossou, O., Dossa, P. and Akponon, C. (2005) A new Bactrocera species in Benin among mango fruit fl y (Diptera: Tephritidae) species. Fruits 60, 371–377.
Veeresh, G.K. (1989) Pest problems in mango – world situation. Acta Horticulturae 231, 551–565.
Veeresh, G.K., Rajagopal, D. and Kumar, N.G. (1989) Management of termites in mango orchard. Acta Horticulturae 231, 633–638.
Verghese, A., Rao, G. and Prasada-Rao, G. (1985) Sequential plan for mango leaf hop-per, Idioscopus clypealis Lethierry. Entomology 10, 285–290.
Verghese, A., Tandon, P. and Prasada-Rao, G. (1988) Ecological studies relevant to the management of Thrips palmi Karny on mango in India. Tropical Pest Management34, 55–58.
Verghese, A., Tandon, P.L. and Stonehouse, J.M. (2003) Economic evaluation of inte-grated management of oriental fruit fl y Bactrocera dorsalis (Diptera: Tephritidae) in mango in India. Crop Protection 23, 61–63.
Verghese, A., Nagaraju, D.K., Jayanthi, P.D.K., Vasudev, V. and Madhura, H.S. (2004) Is azadirachtin useful in the management of the mango stone weevil (Sternoche-tus mangiferae Fabricius)? Journal of Food Agriculture and Environment 2, 213–216.
Viraktamath, C.A. (1997) A revision of the Idiocerine leafhopper genus Amritodus(Hemiptera:Cicadellidae) breeding on mango. Entomon 22, 111–117.
Viraktamath, S. and Viraktamath, C.A. (1985) New species of Busoniominus andIdioscopus (Homoptera: Cicadelidae: Idiocerinae) breeding on mango in south India.Entomon 10, 305–311.
Vyas, R.V., Patel, J.J., Godhani, P.H. and Yadav, D.N. (1993) Evaluation of green musca-rdine fungus (Metarrhizium anisopliae var. anisopliae) for control of mango hopper (Amritodus atkinsoni). Indian Journal of Agricultural Sciences 63, 602–603.
Waddell, W.J., Crooks, N.H. and Carmichael, P.L. (2004) Correlation of tumours with DNA adducts from methyl eugenol and tamoxifen in rats. Toxicological Sciences 79, 38–40.
Waite, G.K. and Huwer, R.K. (1998) Host plants and their role in the ecology of the fruitspotting bugs Amblypelta nitida Stål and Amblypelta lutescens (Hemiptera: Co-reidae). Australian Journal of Entomology 37, 340–349.

J.E. Peña et al.366
Waite, G.K. and Petzl, N. (1994) A search for egg parasites of Amblypelta spp. (Hemiptera: Coreidae) in south-east Queensland. In: Menzel, C. and Croswell, A. (eds) Ma-roochy Research Station Report No. 7, pp. 13–14.
Wang, X.G., Jarjees, E.A., McGraw, B.K., Bokonon-Ganta, A.H., Messing, R.H. and Johnson, M.W. (2005) Effects of spinosad-based fruit fl y bait GF-120 on tephritid fruit fl y and aphid parasitoids. Biological Control 35, 155–162.
Waterhouse, D.F. (1998) Biological control of insect pests: South-East Asian prospects.Australian Centre for International Agricultural Research Monograph No. 51, Can-berra, Australia, 548 pp.
Watson, J.R. (1917) Thysanoptera of Florida. The Florida Buggist 1, 53–77.Wharton, R.A. (1978) Classical biological control of fruit-infesting Tephritidae. In: Rob-
inson, A.S. and Hooper, G. (eds) Fruit Flies. Their Biology, Natural Enemies and Control. Vol. B. Elsevier, Amsterdam, the Netherlands, pp. 303–313.
Wharton, R.A. and Marsh, P.M. (1978) New world Opiinae (Hymenoptera: Braconidae) parasitic on Tephritidae (Diptera). Journal of Washington Academy of Science 68, 147–165.
White, I.M. and Elson-Harris, M. (1992) Fruit Flies of Economic Signifi cance. CAB Inter-national, Wallingford, UK.
Whitwell, A.C. (1993) The pest/predator/parasitoid complex on mango infl orescences in Dominica. Acta Horticulturae 341, 421–432.
Woodruff, R.E. (1985) Citrus weevils of Florida and the West Indies: preliminary report on systematics, biology and distribution (Coleoptera: Curculionidae). Florida Ento-mologist 68, 370–379.
Woods, B., Lacey, I.B., Brockway, C.A. and Johnstone, C.P. (2005) Hosts of Mediterra-nean fruit fl y Ceratitis capitata (Wiedemann) (Diptera: Tephritidae) from Broome and the Broome Peninsula, Western Australia. Australian Journal of Entomology 44, 437–441.
Wysoki, M., Ben-Dov, Y., Swirski, E. and Izhar, Y. (1993) The arthropod pests of mango in Israel. Acta Horticulturae 341, 452–466.
Yazdani, S.S. and Mehto, D.N. (1980) A note on the control of mango hopper, Amrito-dus atkinsoni Leth. at Dholi. Indian Journal of Entomology 42, 275–276.
Yee, W. (1987) The mango in Hawaii. Cooperative Extension Service, University of Hawaii Circular 388, 19–22.
Zaheruddeen, S.M. and Sujatha, A. (1993) Record of Deanolis albizonalis (Hampson) (Pyralidae: Odontinae) as mango fruit borer in Andhra Pradesh. Journal of the Bom-bay Natural History Society 90, 528.
Zucchi, R.A. (1988) Moscas-das-frutas (Diptera: Tephritidae) no Brasil: Taxonomia, Dis-tribuição Geográfi ca e Hospedeiros. Fundação Cargill, Campinas, Brazil.
Zucchi, R.A. (2000) Espécies de Anastrepha, sinonímias, plantas hospedeiras e para-sitóides. In: Malavasi, A. and Zucchi, R.A. (eds) Moscas-das-frutas de Importância Econômica no Brasil: Conhecimento Básico e Aplicado. Holos Editora, Ribeirão Preto, Brazil.

© CAB International 2009. The Mango, 2nd Edition: Botany, Production and Uses(ed. R.E. Litz) 367
11 Crop Production: Propagation
S. Ram1 and R.E. Litz2
1GB Pant University of Agriculture and Technology, Pantnagar, India2University of Florida, Florida, USA
11.1 Introduction 36711.2 Seed Propagation 369
Monoembryonic seed 369Polyembryonic seed 369
11.3 Vegetative Propagation 374Preparation of rootstock 376Attached methods 378Detached methods of grafting 380Effect of rootstock 385Top and double working 386Rooting 387Micropropagation 391
11.4 Comparative Performance of Trees Propagated by Different Methods 39111.5 Conclusions 392
11.1 Introduction
Mango reproduces naturally by seed, although this is rarely a horticultural practice, particularly for monoembryonic cultivars. Instead, vegetative prop-agation is utilized to preserve the unique phenotypes of superior selections, and has been based upon grafting and rooting methods, growing plants from nucellar seedlings of polyembryonic mangoes and micropropagation (Fig. 11.1). Grafting of mango can be either attached or detached. In the former, the scion is not severed from the mother plant until its union with the rootstock is complete, i.e. approach grafting, tongue, saddle and root grafting. In the latter, the scion is removed from the mother tree and then joined with the rootstock, and both are allowed to grow prior to cutting of the rootstock above the graft union. Detached methods include rind or crown grafting, cleft or wedge grafting, whip or splice grafting, side grafting, veneer grafting,
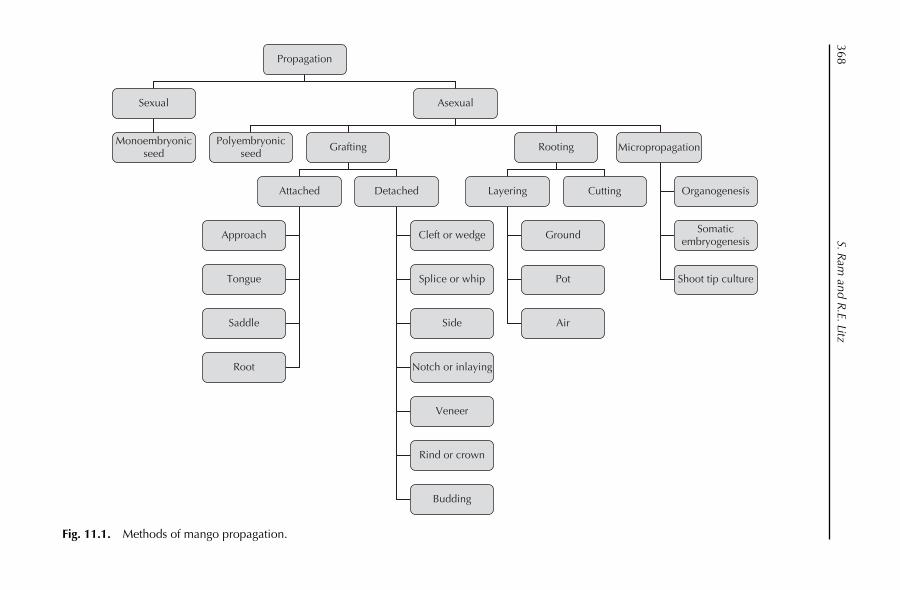
S. R
am and R
.E. Litz368Propagation
Sexual Asexual
Monoembryonicseed
Polyembryonicseed
Grafting Rooting Micropropagation
Attached Detached
Approach
Tongue
Saddle
Root
Cleft or wedge
Rind or crown
Veneer
Notch or inlaying
Side
Splice or whip
Budding
Layering Cutting
Ground
Air
Pot
Organogenesis
Shoot tip culture
Somaticembryogenesis
Fig. 11.1. Methods of mango propagation.

Crop Production: Propagation 369
notch or inlaying, ‘T’ or shield budding, forkert budding, modifi ed forkert budding, patch budding, modifi ed patch budding, chip budding, etc. Root-ing methods include layering and cutting techniques. Although in vitro meth-ods for regenerating mango have been reported via somatic embryogenesis, organogenesis and shoot tip culture (see Litz et al., Chapter 18, this volume), their practical application for propagation has not yet been demonstrated. The various methods of mango propagation are shown in Figs 11.2–11.8. Mango propagation has been discussed recently by Singh and Singh (1998), Galán-Saúco (1999) and Neto et al. (2002).
11.2 Seed Propagation
Monoembryonic seed
Seed propagation does not ensure true-to-type plant reproduction of monoembryonic mango selections; however, it was extensively used before vegetative methods for mango propagation were known (Singh, 1960). Large seedling orchards were planted during the medieval period of Indian history until vegetative propagation was introduced into India by the Portuguese in the late 15th century. Monoembryonic seeds contain only a single sexual embryo, and a single plant grows from a seed of a monoembryonic cultivar.
Polyembryonic seed
Polyembryony is common in mango cultivars that originated and are grown in the moist tropics. In Knight et al. (Chapter 3, this volume), there is a survey of the major polyembryonic and monoembryonic mango cultivars. Trees from nucellar seedlings are identical to the mother plant (see Mukherjee and Litz, Chapter 1, this volume). In polyembryonic seed, only one embryo is zygotic in origin; it either degenerates or produces weak and stunted seed-lings (Maheshwari et al., 1955; Sachar and Chopra, 1957; Singh, 1960; Stur-rock, 1968). Approximately three to eight seedlings normally originate from a single polyembryonic seed (Garner and Chaudhri, 1976), although 30 or more embryos have been recorded in a single polyembryonic mango seed (Juliano, 1934, 1937). Nucellar seedlings can be distinguished from the zygotic seedling on the basis of their greater vigour c.1 month after germination.
Polyembryonic mango cultivars have traditionally been seed propagated in South-east Asia and in some tropical Latin American countries. In many mango-growing areas, polyembryonic and monoembryonic mango selec-tions are grafted onto uniform, nucellar seedling rootstock. Zygotic and nucellar seedlings can be morphologically similar. Therefore, detection and separation of nucellar seedlings is important in breeding programmes involv-ing controlled pollinations. Different protein and DNA markers have been used. Degani et al. (1993) and Schnell and Knight (1993) demonstrated that isozymes and RAPD markers can be used to distinguish zygotic embryos from
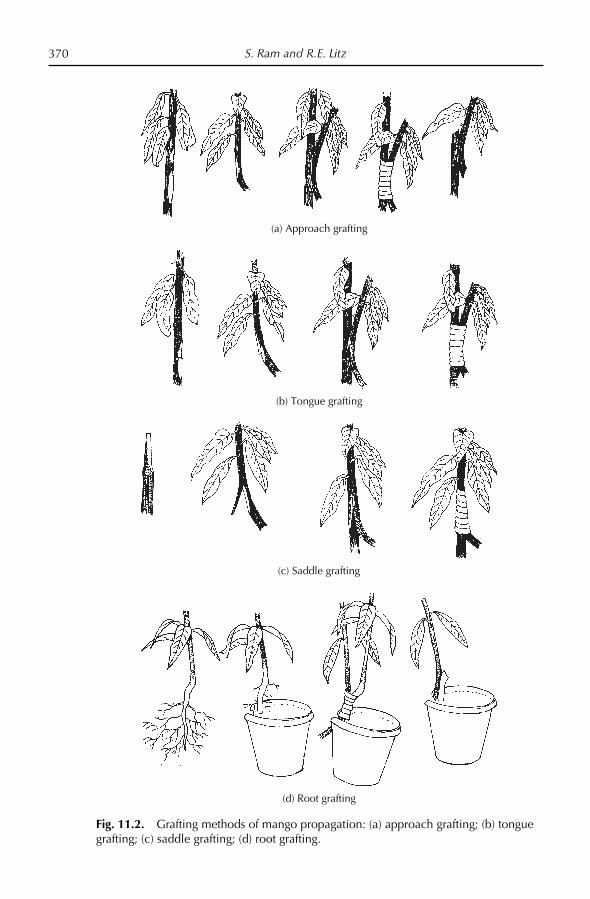
S. Ram and R.E. Litz370
(a) Approach grafting
(b) Tongue grafting
(c) Saddle grafting
(d) Root grafting
Fig. 11.2. Grafting methods of mango propagation: (a) approach grafting; (b) tongue grafting; (c) saddle grafting; (d) root grafting.

Crop Production: Propagation 371
(a) Cleft grafting
(b) Soft wood grafting
(c) Epicotyl grafting
(d) Splice (whip) grafting
Fig. 11.3. Grafting methods of mango propagation: (a) cleft grafting; (b) soft wood grafting; (c) epicotyl grafting; (d) splice or whip grafting.
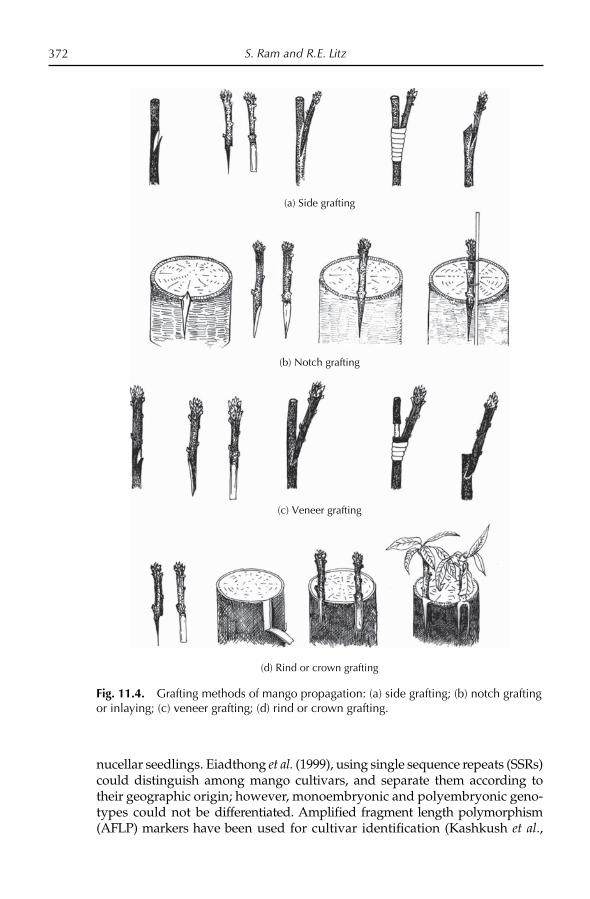
S. Ram and R.E. Litz372
nucellar seedlings. Eiadthong et al. (1999), using single sequence repeats (SSRs) could distinguish among mango cultivars, and separate them according to their geographic origin; however, monoembryonic and polyembryonic geno-types could not be differentiated. Amplifi ed fragment length polymorphism (AFLP) markers have been used for cultivar identifi cation (Kashkush et al.,
(a) Side grafting
(b) Notch grafting
(c) Veneer grafting
(d) Rind or crown grafting
Fig. 11.4. Grafting methods of mango propagation: (a) side grafting; (b) notch grafting or inlaying; (c) veneer grafting; (d) rind or crown grafting.

Plate 1. Mango flowers: polygamous (left) and monoecious (right) flowers. Monoecious flowers are functionally staminate.Plate 2. Longitudinal section through the micropylar end of the ovule of a polyembryonic mango cultivar. Arrows indicate the presence of globular stage nucellar embryos.Plate 3. A mango tree in a relief on the Buddhist temple at Borobodur, Indonesia. The temple is thought tohave been built between the end of the 7th and beginning of the 8th century AD.
1
2
3

Plate 4. ʻAlphonsoʼ.Plate 5. ʻArumanisʼ.Plate 6. ʻAtaulfoʼ.
4
5
76
8
Plate 7. ʻCalypsoʼ.Plate 8. ʻBombay Greenʼ.

Plate 9. ʻCambodianaʼ.Plate 10. ʻCarabaoʼ.Plate 11. ʻGlennʼ.
9 10
1112
1413
Plate 12. ʻGolekʼ.Plate 13. ʻHadenʼ.Plate 14. ʻHimsagarʼ.

Plate 15. ʻHindi Besennaraʼ.Plate 16. ʻIrwinʼ.
15
16
17
18
Plate 17. ʻJulieʼ.Plate 18. ʻKeittʼ.

Plate 19. ʻKensingtonʼ.Plate 20. ʻKentʼ.Plate 21. ʻMabroukaʼ.
19
20
21 22
23
Plate 22. ʻMadame Francisʼ.Plate 23. ʻMallikaʼ.

28
Plate 24. ʻManilaʼ.Plate 25. ʻMulgoaʼ.Plate 26. ʻNam Doc Maiʼ.
24 25
26
27
Plate 27. ʻNuwan Chanʼ.Plate 28. ʻOkrungʼ.

Plate 29. ʻOsteenʼ.Plate 30. ʻPairiʼ.Plate 31. ʻPalmerʼ.
29 30
31 32
33
Plate 32. ʻRosaʼ.Plate 33. ʻSensationʼ.

37
Plate 34. ʻTaimourʼ.Plate 35. ʻTommy Atkinsʼ.
34
35
36
Plate 36. ʻTotapuriʼ.Plate 37. ʻTurpentineʼ.

Plate 38. ʻVallenatoʼ.Plate 39. ʻVan Dykeʼ.Plate 40. ʻZebdaʼ.
38
39
40
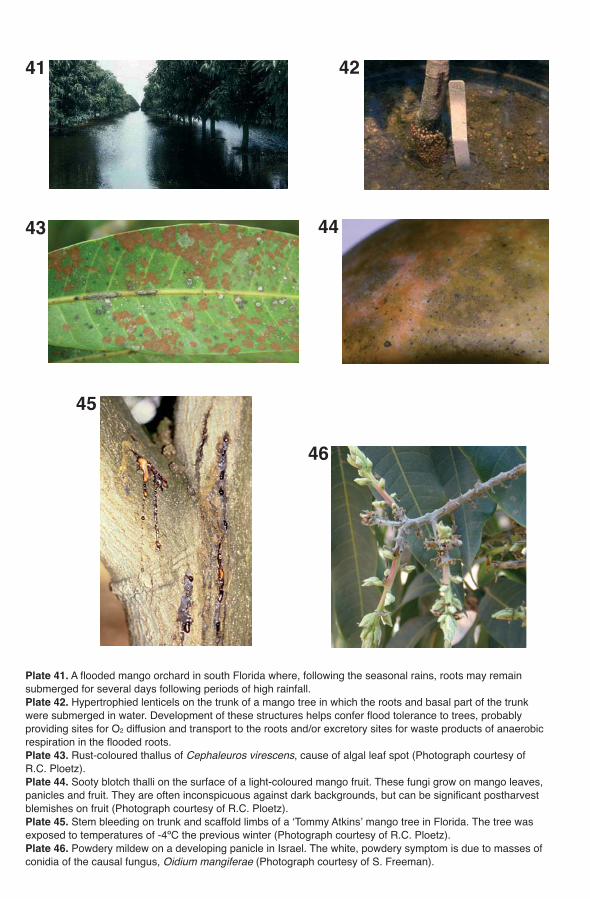
45
46
Plate 41. A flooded mango orchard in south Florida where, following the seasonal rains, roots may remain submerged for several days following periods of high rainfall.Plate 42. Hypertrophied lenticels on the trunk of a mango tree in which the roots and basal part of the trunkwere submerged in water. Development of these structures helps confer flood tolerance to trees, probably providing sites for O2 diffusion and transport to the roots and/or excretory sites for waste products of anaerobicrespiration in the flooded roots.Plate 43. Rust-coloured thallus of Cephaleuros virescens, cause of algal leaf spot (Photograph courtesy ofR.C. Ploetz).Plate 44. Sooty blotch thalli on the surface of a light-coloured mango fruit. These fungi grow on mango leaves,panicles and fruit. They are often inconspicuous against dark backgrounds, but can be significant postharvestblemishes on fruit (Photograph courtesy of R.C. Ploetz).Plate 45. Stem bleeding on trunk and scaffold limbs of a ʻTommy Atkinsʼ mango tree in Florida. The tree wasexposed to temperatures of -4ºC the previous winter (Photograph courtesy of R.C. Ploetz).Plate 46. Powdery mildew on a developing panicle in Israel. The white, powdery symptom is due to masses ofconidia of the causal fungus, Oidium mangiferae (Photograph courtesy of S. Freeman).
43 44
41 42

Plate 47. Sudden wilt in Oman and Pakistan is caused by a newly described ascomycete species, Ceratocystis manginecans. (a) Vascular streaking and galleries of the scolytid beetle vector, Hypocryphalusmangiferae, are typically found in affected trees, as is (b) unilateral death of portions of the canopy; ultimately trees are killed (Photographs courtesy of R.C. Ploetz).
47a
47b

53
Plate 48. Internal fruit breakdown: jelly seed (Photograph courtesy of A. Cracknell Torres).Plate 49. Internal fruit breakdown: soft nose (Photograph courtesy of A. Cracknell Torres).Plate 50. Internal fruit breakdown: stem end cavity (Photograph courtesy of A. Cracknell Torres).Plate 51. Internal fruit breakdown: spongy tissue (Photograph courtesy of A. Cracknell Torres).Plate 52. Internal fruit breakdown: necrotic tissue (Photograph courtesy of A. Cracknell Torres).Plate 53. Black tip disorder.
48 49
50 51
52

Plate 54. Anastrepha obliqua on mango leaf.Plate 55. Australian fruit fly Bactrocera sp.Plate 56. Mediterranean fruit fly Ceratitis capitata.
56
54
55
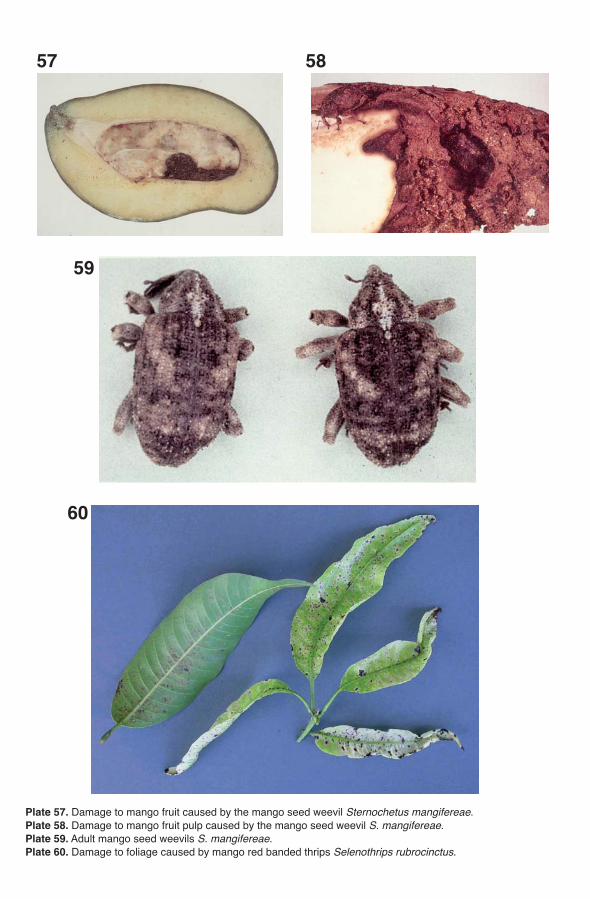
Plate 57. Damage to mango fruit caused by the mango seed weevil Sternochetus mangifereae.Plate 58. Damage to mango fruit pulp caused by the mango seed weevil S. mangifereae.Plate 59. Adult mango seed weevils S. mangifereae.Plate 60. Damage to foliage caused by mango red banded thrips Selenothrips rubrocinctus.
57
59
58
60

Plate 61. Mango inflorescence with thrips.Plate 62. The mango red banded thrips Selenothrips rubrocinctus.Plate 63. Adult mango red banded thrips S. rubrocinctus.Plate 64. The mango gall midge Erosomya mangiferae on a mango leaf.Plate 65. Leaf hopper Idioscopus sp. on a mango leaf.Plate 66. Pleuroprucha insullaria.
6162
6364
65 66
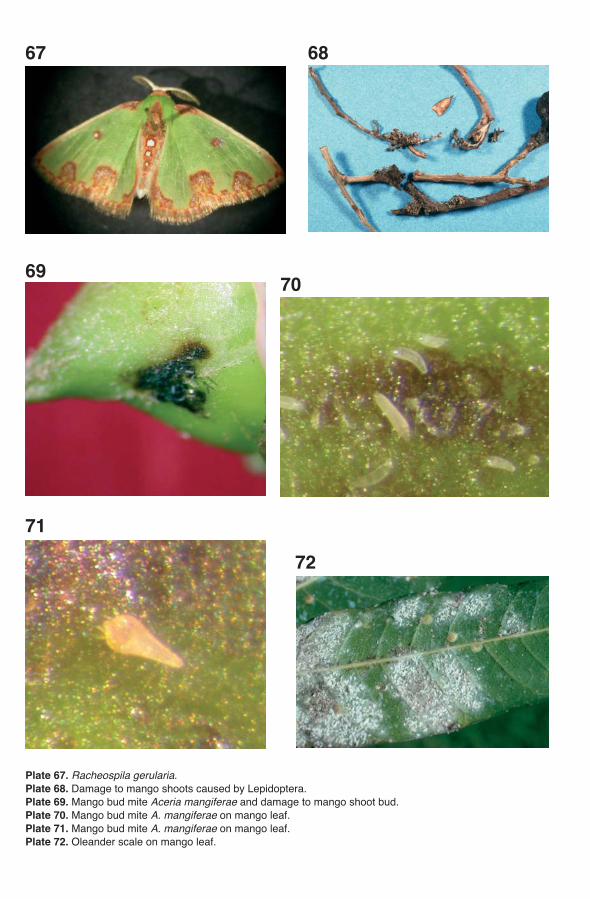
Plate 67. Racheospila gerularia.Plate 68. Damage to mango shoots caused by Lepidoptera.Plate 69. Mango bud mite Aceria mangiferae and damage to mango shoot bud.Plate 70. Mango bud mite A. mangiferae on mango leaf.Plate 71. Mango bud mite A. mangiferae on mango leaf.Plate 72. Oleander scale on mango leaf.
67 68
71
69
72
70

Plate 73. Effect of nitrogen (N) on fruit colour. (a) Excessive N suppresses mango fruit colour in the field. (b) Ripe ʻKeittʼ fruit grown on (front) low N (0 applied N) and (rear) grown on high N (417 g/tree).Plate 74. Effect of N on postharvest disease. Ripe ʻKeittʼ fruit grown on (left) high N (417 g/tree) and (right)low N (0 g/tree).
73a 73b
74

Plate 75. Magnesium (Mg) deficiency showing interveinal chlorosis.Plate 76. Zinc (Zn) deficiency showing pale-yellow leaves that have not fully expanded.Plate 77. Iron (Fe) deficiency, showing pasty pale-yellow leaf coloration.
75
77
76

Plate 78. Boron (B) deficiency symptoms showing (top left) stunted growth and sickle-shaped leaves; (top right) shot holes in leaves; (bottom left) bent tip of flower panicle; and (bottom right) gummosis fromcracking trunk.Plate 79. B toxicity symptoms: black staining leading to necrotic leaf margins
78
79
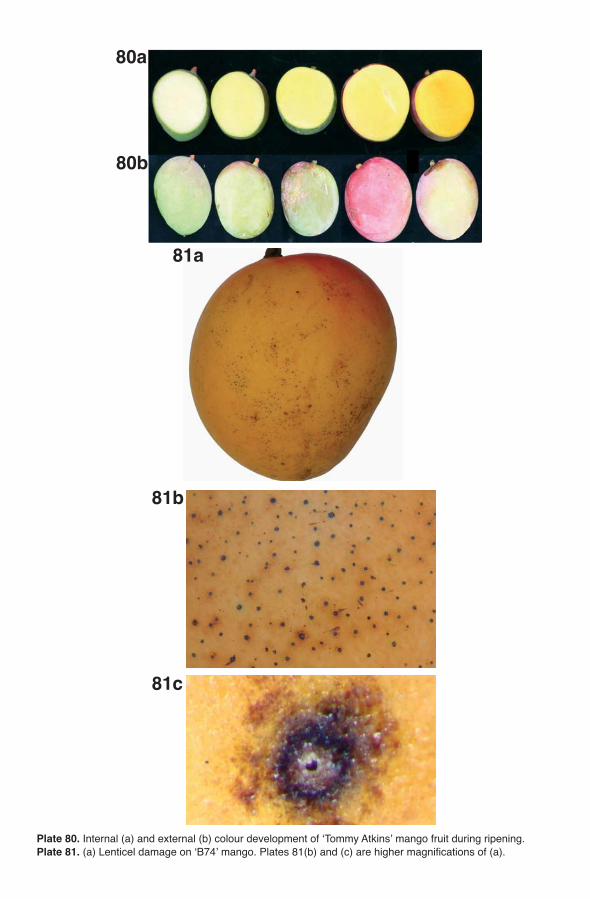
Plate 80. Internal (a) and external (b) colour development of ʻTommy Atkinsʼ mango fruit during ripening.Plate 81. (a) Lenticel damage on ʻB74ʼ mango. Plates 81(b) and (c) are higher magnifications of (a).
80a
81a
81b
81c
80b

Plate 82. Harvest aids used in Australia. Desapping using racks in the field based on placing the fruit onto a tarpaulin(a and b) or basket constantly sprayed with detergent and the fruit remaining wet with detergent for c.90 s.(c) A cherry picker is used to harvest high fruit.
82a
82b
82c

83
Plate 82. (d) Desappingusing racks in he field and (e) desapping conveyor used for desapping in the pack-house.Plate 83. Processed mango in the marketplace, including yoghurt, nectar, reconstituted drink (from powder),fruit leather, dried mango slices and preserved fruit.
82d
82e

Plate 84. Dried mango slices.Plate 85. (a) Dried green mango powder; (b) reconstituting green mango powder in water (green mango shake).
84
85a 85b
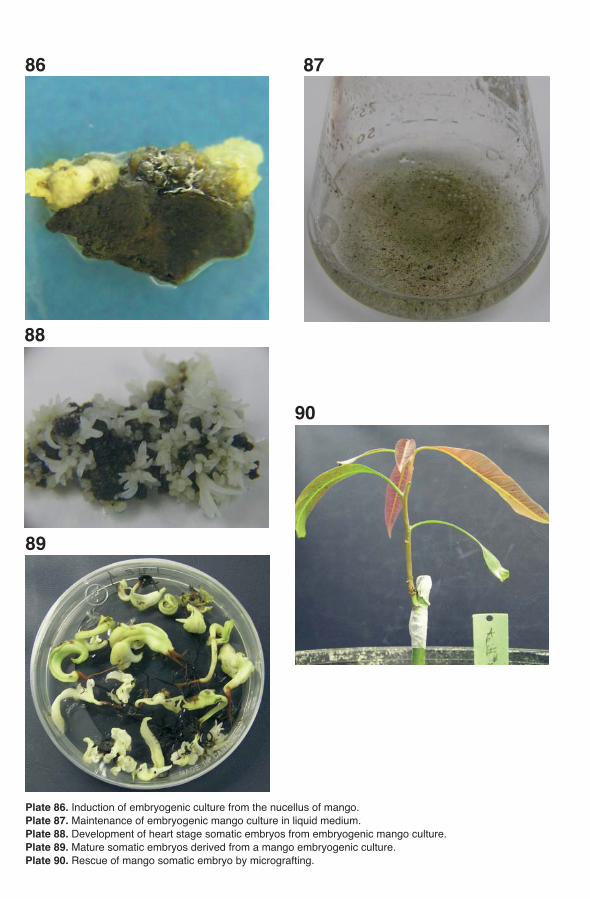
Plate 86. Induction of embryogenic culture from the nucellus of mango.Plate 87. Maintenance of embryogenic mango culture in liquid medium.Plate 88. Development of heart stage somatic embryos from embryogenic mango culture.Plate 89. Mature somatic embryos derived from a mango embryogenic culture.Plate 90. Rescue of mango somatic embryo by micrografting.
88
87
89
90
86

Crop Production: Propagation 373
(a) Shield budding
(b) Patch budding
(c) Modified patch budding
(d) Forkert budding
Fig. 11.5. Budding methods of mango propagation: (a) shield budding; (b) patch budding; (c) modifi ed patch budding; (d) forkert budding.

S. Ram and R.E. Litz374
2001) and Gonzalez et al. (2002) identifi ed a range of inter-simple-sequence-repeats (ISSR) primer sequences with potential for cultivar identifi cation.
11.3 Vegetative Propagation
Vegetative or asexual propagation preserves the genotype and phenotype of elite selections by means of grafting, rooting and micropropagation. Trees
(a) Modified forkert budding
(b) H-budding
(c) Chip budding
Fig. 11.6. Budding methods of mango propagation: (a) modifi ed forkert budding; (b) H-budding; (c) chip budding.

Crop Production: Propagation 375
propagated by grafting onto seedling rootstocks usually fl ower after 3 or 4 years in comparison to the 5–10 years for seedling trees and they are gener-ally smaller than seedling trees because they begin to bear fruit earlier. The oldest method for vegetatively propagating mango is probably the attached method, also known as inarching or approach grafting.
(a) Modified chip budding
(b) Flap budding
(c) Window budding
Fig. 11.7. Budding methods of mango propagation: (a) modifi ed chip budding; (b) fl ap budding; (c) window budding.
S. Ram and R.E. Litz376
Preparation of rootstock
Rootstocks are either zygotic or nucellar seedlings. Nucellar seedling root-stocks are clonal, and have many advantages over heterogeneous monoem-bryonic seedlings as rootstocks (see Crane et al., Chapter 13, this volume). Clonal rootstocks have generally been selected for specifi c soil types and
(a) Top working
(b) Double working
Fig. 11.8. Top working (a) and double working (b) of mango.

Crop Production: Propagation 377
stress tolerance, and behaviour of the scion cultivar on clonal rootstock is highly predictable. Monoembryonic seedlings are generally used as rootstocks in India. Polyembryonic ‘Turpentine’ and ‘Kensington Pride’ seedlings are used as rootstocks in Florida and Australia, respectively, whereas polyembryonic ‘Sabre’, ‘13-1’ and ‘4-9’ seedlings are used for rootstocks in Israel. Throughout South-east Asia, polyembryonic seedlings are used for rootstocks, e.g. poly-embryonic ‘Saing’ and ‘Thalapt’ in Myanmar (Grant and William, 1949). In Senegal, polyembryonic ‘Amelioree’, ‘De Cameron’ and ‘Madame Francis’ are used as rootstocks (Furon, 1966). In Brazil, seedlings of polyembryonic ‘Carabao’, ‘Mangua d’Agua’ and ‘Pico’ are used for rootstock; they are consid-ered to have resistance to Ceratocystis fi mbriata (Neto et al., 2002); the IAC series (IAC100-104) are used to control ‘seca de mangueira’ (Neto et al., 2002).
Freshly extracted mango seeds from ripe fruits germinate with higher frequency (76–91%) than those from overripe, fi rm or green fruits (Shant and Saproo, 1974). Seeds with large cotyledons from seedling trees germinate earlier, store better and seedling growth is more vigorous than seeds with small cotyledons (Naik, 1949; Simao, 1960). Naik (1941) noted differences in germination and vigour in monoembryonic seedling progenies of different parentages. Seedlings of ‘Chausa’ reportedly produced better height and girth than ‘Langra’ and seedling vigour was closely related to the weight of the seed stone (Giri and Chaudhery, 1966). Mango seeds are recalcitrant, and lose viability (Ledin and Ruehle, 1954; Ruehle and Ledin, 1955; Singh, 1960) within 30 days. Therefore, seed should be collected and sown within 1 week after collection. About 80% of seed germination occurs if they are sown within 1 month of extraction (Stephens, 1960; Sturrock, 1968; Chacko and Singh, 1971). Parisot (1988) recorded that seeds that are free of pulp could be stored for up to 84 days at 15°C on sterile cotton with deionized water. Husk-ing or decortication of the hard endocarp of the seed improves germination (Paul and Guneratnam, 1938; Simao, 1960; Chauran et al., 1979; Abdel-Galil, 1992). At the time of sowing, treatment with fungicides also improves germi-nation, particularly of dehusked seeds (Chauran et al., 1979). Seeds should be sown without injuring the cotyledons and preferably early in the season.
Seedbed preparationThe standard practice for seed germination varies. In India, seedbeds are used, whereas in most other countries, seeds are planted individually in pots or plastic bags. Nurseries are usually under partial shade to prevent exces-sive evaporation from the plant and soil and to protect seedlings from inclem-ent weather; shade is particularly useful in areas with dry, hot climates. Seedbeds can also be used in the open provided the soil is adequately moist. Highly sandy or loamy soils are generally unsuitable for mango seed germi-nation. Soil should be well drained with plenty of organic matter, and seeds should be sown c.15 cm apart in a special seedbed in which 25 cm top soil of the bed is excavated and the bed bottom is hardened with concrete or lined with an iron or polyethylene sheet to prevent elongation of the taproot and to encourage fi brous root development (Stephens, 1948). Germination can be expedited if seeds are soaked in 20–100 mg/l gibberellic acid (GA3) for 24–48 h

S. Ram and R.E. Litz378
or sprayed with 50–300 mg/l GA3; however, these seedlings are usually unsatisfactory for grafting. Seeds should be planted with convex edge up at a depth of 5–7 cm (Singh, 1960; Bakhshi, 1963) or with a small portion exposed above the ground level (Pope, 1929; Ruehle and Ledin, 1955; Bakhshi, 1963; Anonymous, 1975). Soil moisture should be maintained. Seeds germinate 2–3 weeks after planting (Ruehle and Ledin, 1955; Ahmed and Rashid, 1961; Singh and Jawanda, 1962). Seedlings that are 1-month-old with coppery leaves and with their stones attached survive transplanting better than older plants without much damage to their root system (Ruehle and Ledin, 1955). The taproot should be slightly pruned at the time of planting.
NurserySeedlings should be transplanted without disturbing the roots to nursery beds or pots at least 1 month before grafting. Either the crown of the seed-lings should be pruned or the distal half of the leaves should be removed to reduce transpiration. Seedbed-grown plants generally have better germina-tion rates and lower production costs than those grown directly in nursery beds. Seedlings should be actively growing at the time of grafting. Success in grafting depends on several factors, such as age and vigour of rootstock and scion, diseases, insect pests, environment and the skill of the grafter.
Attached methods
Approach grafting is expensive, cumbersome, requires a long time to pro-duce grafts and the success rate is often low. Other methods are now used because they are cheaper and easier.
Approach graftingA scion shoot, while still attached to the parent plant, can be grafted onto the seedling rootstock by making a long cut (5–7 cm) of similar length and depth on one side of both rootstock and scion through the cambium and slightly into the wood (Fig. 11.2a). The cut surfaces of rootstock and scion are pressed fi rmly together and secured with waxed string, raffi a or grafting tape. After union, the scion is severed below the graft union and the rootstock above the union. Several years ago, approach grafting was standard practice for propa-gating mango in many countries, but has been largely replaced by detached grafting methods, except for parts of India. For approach grafting, seedlings 45–60 cm long with a diameter of 1.25 cm are utilized (Asadullah and Khan, 1960; Singh, 1960). They are planted in pots or plastic bags containing soil mixture, and are kept beneath the mother tree. Alternatively, the ball of earth around the root system of the seedling grown for rootstock is tied in ‘jutties’ made of dried grass, paddy straw or plastic bags and planted under the mother tree or raised to the shoot height of the mother tree on a raised plat-form or directly suspended from branches of the mother tree. The seedling rootstock can be a few months- to 2-years-old (Naik, 1949; Garg, 1954; Ahmed, 1960; Singh, 1960). Juvenile seedlings whose leaves are copper coloured and

Crop Production: Propagation 379
with the seed still attached to the seedling are often used. The root system is covered with moist sphagnum moss or similar material and wrapped in polyethylene to prevent drying. Production costs are less with young seed-lings than with older material.
In the subtropics, approach grafting in spring can achieve 88–100% suc-cess with some cultivars (Asadullah and Khan, 1960; Majhail and Singh, 1962a, b; Talukdar and Ahmed, 1965). Majhail and Singh (1962b) observed that seedling size and age do not affect grafting success in spring, but that larger seedlings yielded better success from July to September. Approach grafting should be avoided during winter months when plants are dormant. In the tropics, mangoes can be approach grafted year-round (Ahmed, 1960; Singh, 1960). Approach grafting should be done when the trees are in active growth (Singh, 1960; Rao, 1967). In low rainfall areas, approach grafting should be done at the time of the onset of rains, while in regions of heavy rainfall, it should be done soon after the rainy season is over, provided tem-peratures are not <15°C, the critical temperature for the growth of mango trees (Whiley, 1993). Healthy trees should be used for scion wood. The suc-cess of approach grafting is cultivar dependent, for example ‘Langra’ scions establishes better than either ‘Dashehari’ or ‘Chausa’ (Asadullah and Khan, 1960). Better success has also been reported with seedlings of 1.3–1.6 cm diameter than with smaller shoots (Giri, 1966). Variations of approach graft-ing include multistock, infl orescence, top working and adjuvant grafting. At one time, south Indian nurserymen would approach graft a single large scion with two to fi ve rootstocks (Patwardhan and Deshmukh, 1931; Singh, 1960). Grafts with large and old scions having several branches become top heavy, and fail to thrive. New growth from infl orescences that are approach grafted is veg-etative. Approach grafting has also been used to top work old seedling trees with cultivars (Singh, 1960). Adjuvant grafting refers to approach grafting of diseased or damaged trees in order to rejuvenate them on new rootstocks.
Tongue graftingTongue grafting is practised with thicker rootstock than scion. The rootstock is fi rst cut as in approach grafting; a second cut is made into the wood of the stem approximately halfway down the fi rst cut in a sloping direction point-ing tongue upwards. A similar cut is made in the scion shoot. Both cuts are made in such a manner that one fi ts into the other tightly (Fig. 11.2b). Root-stock and scion are tied together with grafting tape and a graft union is com-plete in c.1–2 months. The scion is then severed from the mother tree and the rootstock is decapitated above the graft union. Tongue grafting is slightly more complicated than simple approach grafting; however, the success rate is better because three surfaces of the cambium layer are involved in the graft union (Burns and Prayag, 1921).
Saddle graftingWith saddle grafting, the rootstock is decapitated c.25 cm above ground level, and two upward sloping cuts are made on the sides of the rootstock to form a 5–7 cm long wedge on its cut end. On the stem of the scion shoot, a tongue

S. Ram and R.E. Litz380
is made and the outer surface of the tongue is not pared. The wedge of the rootstock is then fi tted into the groove of the tongue (Fig. 11.2c). Subsequently, the joint is secured with grafting tape and the scion shoot is separated from the mother tree after the union is complete (Singh, 1960). Saddle grafting is easier to perform than tongue grafting but is more diffi cult than approach grafting.
Root graftingRoot grafting is similar to bench grafting. A seedling plant (c.l year old) is potted in a U-shaped pot with a notched edge (7–8 cm deep and 5 cm wide) (Singh, 1960). The 7–10 cm taproot is projected through the notch and the pot itself contains the root. The seedling above the collar is staked, and placed in the shade for 1–1.5 months for establishment. Seedlings are approach grafted with the scion (Fig. 11.2d), and the graft is separated from the mother tree after the union is complete. Grafts are maintained in the shade and watered regularly. Naik (1941, 1949) reported 100% success with root grafting.
Detached methods of grafting
Cleft or wedge graftingSingh (1960) suggested that cleft grafting can be used with rootstocks of greater diameter than the scion and also used for top working (Fig. 11.3a). Rootstock of 5–7 cm or more in diameter should be used and cleft grafted after decapitating the stock c.45 cm above the ground. The decapitated root-stock is split to a depth of about 5 cm through the centre of the stem with a knife and a mallet. After the knife is removed, a hard wooden wedge is inserted to keep it open for the subsequent insertion of the scion. Scions (15–20 cm long) from a terminal shoot >3 months old is wedged securely (to a depth of 6–7 cm). The cleft of the scion is slipped into the split of the stock that has been forced open with the wooden wedge. The thicker edge of the cleft is placed towards the outer edge of the rootstock in such a way that the back of the rootstock and scion meet fi rmly. Another scion is also inserted diametrically opposite the fi rst one. The wooden wedge is removed and cut surfaces are sealed with grafting wax. The graft union requires c.2 months. Cleft or wedge grafting is appropriate for replacing the crown of young trees; however, with young seedling rootstock, this has also been used for large-scale propagation. In Brazil, Pinheiro et al. (1970) reported that cleft grafting was more successful (97%) than four other grafting methods tried. Cleft grafting is easier to use than veneer grafting and has a similar rate of success (Ram, 1993). Success can be improved when leaves are retained below the point where the rootstock is severed, when the grafting portion of the rootstock is a new fl ush and when the stem is pinkish green; however, the scion should be mature.
Soft wood graftingInsertion of the scion into the bronze-coloured stem of the rootstock has been referred to as soft wood grafting (Fig. 11.3b). Amin (1974, 1978) reported 100% success if leaves are not removed below the graft union. Seedling rootstocks

Crop Production: Propagation 381
of various ages can be used; however, success decreases with age, i.e. 80% with 1-year-old rootstocks and less with 6-month-old seedlings (Gaur, 1984; Reddy and Melanta, 1988; Kulwal and Tayde, 1989; Panickar and Desai, 1989; Gandhoke, 1993). This technique is easy and is effective in dry, hot weather and in areas with low precipitation.
Epicotyl or stone graftingEpicotyl grafting involves cleft grafting of scions onto 2-week-old seedling rootstock (Traub and Auchter, 1934). Predefoliation of the scion is not essen-tial, although it is widely practised (Fig. 11.3c). Moderate temperature and high relative humidity (RH) are important for success. A 2–3 cm long slant-ing cut is made in the epicotyl with a matching cut on the proximal portion of the scion in the splice method for grafting. For wedge grafting, a vertical cut 4–5 cm long is made in the decapitated epicotyl to receive the wedge-shaped scion. The scion and rootstock are tied together with grafting tape, and the small grafted plant is planted immediately and watered. The scions sprout within a month. The success rates of splice and wedge grafting meth-ods are reported to be >50% (Bhan et al., 1969; Gupta et al., 1988; Dhunaga et al., 1989; Gunjate, 1989; Jinturkar and Narwadkar, 1989; Kulwal and Tayde, 1989; Madalgeri et al., 1989; Narwadkar and Anserwadekar, 1989; Radhamony et al., 1989; Singh et al., 1989; Srivastava, 1989; Patil et al., 1991); under green-house conditions in the subtropics, success can be >90% (Ram, unpublished). Grafted plants are maintained under partial shade in a moderate and moist environment. In the subtropics, growth is initially slow and can require c.2 years to attain the size of a veneer graft; however, in the tropics, growth is much more rapid.
Splice or whip graftingThis is the easiest method for propagating mango and is widely used in the subtropics. The rootstock should be c.1–2 years old, the diameter of both rootstock and scion should be 0.6–1.0 cm. The rootstock is severed c.10–20 cm above the soil and a diagonal cut (3–4 cm long) is made at the distal end of the rootstock (Fig. 11.3d). A similar slanting cut is made on the proximal end of the scion and the cut surfaces of both the rootstock and the scion are bound together with grafting tape. The union (usually 90% successful) takes place in c.2 months (Ahmed, 1974). Torres (1949, 1960) used this method with 3- to 9-month-old seedling rootstock with a high success rate. Majumder and Rathore (1970) reported 50% success with splice grafting with 2-week-old seedlings and up to 60% success with 30-day-old seedlings; however, sur-vival of the grafts was poor.
Side graftingSide grafting, also known as the ‘Nakamura method’ (Fig. 11.4a), was for-merly popular (Burns and Prayag, 1921; Pope, 1929; Pope and Storey, 1933; Walters, 1932; Tanaka, 1939; Lynch and Mustard, 1950; Thrower, 1954; Ruehle and Ledin, 1955; Lynch and Nelson, 1956; Ahmed, 1960; Singh, 1960; Serpa, 1964; Rao, 1967). This method is supposed to be highly effective in mild

S. Ram and R.E. Litz382
weather in the absence of strong winds, intense heat and heavy rain (Rao, 1967), and success has also varied (50–100%) with different cultivars (Veera-raghavan, 1945). In India, side grafting is generally used in humid, coastal areas with 1.0–1.5 cm diameter rootstock. Scions should be c.10 cm long and defoliated c.1 week prior to grafting. The rootstock age has varied from 4 to 36 months with 72–90% success (Mulat, 1959; Kashyap et al., 1972; Faruque and Fakir, 1973; Kanwar and Bhajwa, 1974; Ben-ya’acov, 1976). The scion is inserted into the wedge and tied with grafting tape. The union is complete after 2–3 months. The top of the rootstock can be removed above the graft union after new scion growth occurs.
Notch grafting or inlayingThis method is used in the humid tropics with >8 cm diameter rootstock (Singh, 1960). The rootstock is decapitated c.45 cm above the ground level. V-shaped notches (4–5 cm) are made at equal distances, depending upon the thickness of the rootstock, from the periphery of the cut surface. The proxi-mal end of each scion is fi tted into each notch of the rootstock (Fig. 11.4b) and scions are secured to the rootstock with grafting wax. The union is normally complete in c.2–3 months (Singh, 1960).
Veneer graftingThis grafting technique was fi rst described by Lynch (1941), and has been widely adopted (Cooper and Furr, 1945; Ruehle and Ledin, 1955; Mukherjee and Majumder, 1961, 1962, 1964; Wolfe, 1963; Ahmed, 1964; Serpa, 1964; Jagirdar et al., 1968; Bhambota et al., 1971; Prasad et al., 1973; Ramirez Diaz, 1973; Gun-jate and Limaye, 1978; Ram and Bist, 1982; Pinto et al., 2004). Veneer grafting is usually more effective than other methods of grafting (Bhambota et al., 1971; Prasad et al., 1973), with approximately 90% success (Ram and Bist, 1982). For veneer grafting, 3- to 6-month-old 1.0–1.5 cm diameter scion shoots with lush green leaves should be defoliated 3–15 days prior to grafting (depending upon the season), leaving the petioles attached (Mukherjee and Majumder, 1961; Singh and Srivastava, 1979a, b; Ram and Bist, 1982; Rajan and Sinha, 1987; Bajpai et al., 1989). Prior defoliation may not be required under humid conditions (Gunjate et al., 1976) or when extremes of temperature and RH do not occur. The scion should be 10–15 cm long, although smaller scions (5–10 cm) have also been used (Ram and Bist, 1982). Older shoots can also be used (Mukherjee and Majumder, 1961; Jagirdar et al., 1968; Ram and Bist, 1982). Scions stored for 8 days at ambient temperature (25–35°C) in moist sphagnum moss covered with polyethylene can still be grafted with a 50% success rate during March through to July (Ram and Bist, 1982). Cultivar differences may be important.
The use of seedling rootstock at different ages (3 months to 2 years) has resulted in 40–100% success, depending upon the season, scion maturity, predefoliation period, storage condition of scion, etc. (Zill, 1951; Subra, 1954; Mukherjee and Majumder, 1961, 1962, 1964; Ahmed, 1964; Serpa, 1964; Jagirdar and Bhatti, 1968; Bhambota et al., 1971; Prasad et al., 1973; Ramirez Diaz, 1973; Gunjate and Limaye, 1978; Ram and Bist, 1982). Dipping or smearing

Crop Production: Propagation 383
growth regulator onto the cut surfaces of the scion and rootstock has been ineffective (Kulkarni et al., 1989). The rootstock is prepared for veneer grafting by making a slanting cut (5 cm long); an oblique cut is then made at the base of the fi rst cut, so that a piece of wood along with bark is removed (Fig. 11.4c).The base of the scion wood is then fi tted into the rootstock so that the cut surfaces with the cambium layers of scion and rootstock are in contact with each other. The rootstock and scion are secured together with grafting tape. The union takes place after 1.5–2.0 months. After scion growth begins, the rootstock is removed above the graft union. Some workers have recom-mended fi rst ringing and then removing the rootstock shoot 1–2 weeks later, while others have partially cut through the rootstock shoot before severing the shoot completely a few days later. After new leaves of the scion turn green, the rootstock may be removed completely together with grafting tape (Mukherjee and Majumder, 1961; Ram and Bist, 1982).
Rind or crown graftingRind or crown grafting has been used in the Americas and India. Its success depends upon several factors, such as season, rootstock, scion maturity, mod-erate temperature and high RH (Gangolly et al., 1957; Popenoe, 1959; Singh, 1960). The rootstock is decapitated 30 cm above the soil. A longitudinal cut (8 cm) is made in the bark from the top of the decapitated rootstock, down-wards, without injuring the wood below the bark. The bark is loosened from the wood with a wooden wedge or spatula and a freshly prepared 3-month-old scion is inserted after creating a wedge c.6 cm long at the proximal end of the scion. The fl ap of bark is then secured over the inserted scion with grafting tape. Several scions may be inserted into a single thick rootstock (Fig. 11.4d). High RH around the graft is important for success. The graft union occurs in c.1 month, but the grafting tape should be removed from the grafts only after the leaves of the scion turn green. Rind or crown grafting is a cumbersome method and is not often used in commercial nurseries; however, it can be useful for top working.
BuddingBudding involves the grafting of a single bud, with or without wood attached, with the rootstock. Budding methods are referred to by the shape of the bud, fl ap of the rootstock covering the bud before insertion, etc. The various meth-ods for budding are illustrated in Figs 11.5–11.7. The inserted bud eventually develops as the crown. Rootstock above the bud is severed after bud estab-lishment. Budding provides a stronger union and the graft union occurs ear-lier than with other grafting methods. Both rootstock and scion should be actively growing, and excision of the bud is easy. Stimulation of the bud prior to its excision by predefoliation, application of growth regulators and gir-dling can improve the effi ciency of the procedure. A bud is normally selected from a 3- to 4-month-old scion shoot, and is budded onto rootstock stems of 1.0–1.25 cm diameter c.25–30 cm above ground level where the rootstock is greyish green, except in cases of very juvenile rootstock. Generally 1- and 2-year-old rootstocks are better for grafting than younger ones. The bud is

S. Ram and R.E. Litz384
completely covered by grafting tape for a few weeks. Bud growth can be stimulated by removing grafting tape c.2 weeks after grafting.
Shield or ‘T’ buddingShield or ‘T’ budding of mango (Fig. 11.5a) occurs with varying success, ranging from 32–94% (Singh and Khan, 1943, 1946; Singh and Ram, 1946; Singh, 1946; Singh, 1960; Singh and Srivastava, 1961). In northern India and Pakistan, shield or ‘T’ budding is generally most effective from March to July (India) and March–April (Pakistan). Defoliating the scion shoot for 10–15 days before budding (Pope, 1929; Parsons, 1931; Khan, 1960; Rao, 1967; Teaotia and Maurya, 1970) and girdling of the scion shoot several weeks prior to budding can increase bud take (Anonymous, 1950). Scion cultivar differences also appear to be important, for example success is better with ‘Langra’ than with ‘Aman Dashehari’ and ‘Chausa’ (Ahmed and Chatha, 1960); however, success is higher with ‘Sindhri’ than with ‘Langra’ and ‘Banganpalli’ (Jagirdar and Ali, 1968). Marked differences in success have also been reported for dif-ferent agroecological zones in the same season: 45% in the dry hot plains of the Punjab and 90% in relatively cool areas (Bajwa and Ram, 1946).
Temperature, RH and storage media infl uence bud storage (Walters, 1932; Sundara Rao, 1956; Srivastava, 1963a, b; Rao, 1967). Storage of bud-wood in moist sphagnum moss for 10 days is generally satisfactory. Sealing the cut ends of budwood with melted wax and storage in a thermos fl ask with ice for 48 h does not adversely affect budding (Singh and Khan, 1943). Buds can be inserted in inverted ‘T’ cuts using 2- to 5-year-old rootstock (Higgins, 1910; Ruehle and Ledin, 1955). ‘T’ budding in situ is also successful (Singh and Khan, 1942, 1946; Bajwa and Ram, 1946; Khan, 1960) in March–April and August–October (Ullah and Ali, 1955). The bud should be secured with grafting tape. Soule (1971) described four stages in the formation of the bud union: (i) pre-callus with wound periderm up to 4 days after budding; (ii) callus proliferation 8 days after budding; (iii) cambial bridge between rootstock and scion and vascular tissue differentiation 12 days after budding; and (iv) the healed union after 2–4 weeks. If the buds are green up to 15 days after budding, the grafting tape should be removed and the rootstock should be girdled 10 cm above the union to stimulate the bud to sprout. The root-stock is fi nally removed above the budding point after a growth fl ush of the scion (Luthra and Sharma, 1946; Ruehle and Ledin, 1955; Hayes, 1957). Budded plants are transplanted only after at least one season’s growth.
Patch buddingPatch budding (Figs 11.5b and c) (Verma, 1942; Ullah and Ali, 1955; Singh, 1960; Moreuil, 1963; Teaotia, 1963; Jauhari and Singh, 1970; Teaotia and Maurya, 1970; Prasad and Singh, 1972; Prasad et al., 1973) is used as an alterna-tive to approach grafting (Garner and Chaudhri, 1976). Under subtropical north Indian conditions, March and May through to July is the optimum period for patch budding. In tropical Malaysia, patch budding is successful from December through to February. The scion is defoliated 2 weeks prior to budding. Rootstocks are severed above the budding point 1–2 weeks after budding.

Crop Production: Propagation 385
Patch budding is apparently superior to forkert budding with ‘Langra’ and ‘Dashehari’ (Teaotia, 1963).
Forkert buddingOver 90% grafting success has been reported with forkert budding (Fig. 11.5d) in tropical South-east Asia and Sri Lanka (Paul and Guneratnam, 1937a, b). In Maharashtra, India during July and August, success with this technique using 1-year-old rootstock was 60–70% (Gandhi, 1942), and 100% with ‘Langra’ and ‘Dashehari’ (Singh and Srivastava, 1962; Teaotia, 1963). A modifi ed forkert budding method (Fig. 11.6a) can be more effective than shield budding (Garner and Chaudhri, 1976). H-budding is another modifi -cation of the forkert method (Singh, 1960), and derives its name from an ‘H’-shaped cut made in the rootstock (Fig. 11.6b).
Chip buddingFor chip budding, 2- to 3-month-old seedlings are used (Fig. 11.6c). The bud is excised with a piece of wood attached to it. Graft union can occur 3–4 weeks after budding because rootstock tissues are partially undifferentiated and possess a broadly exposed cambium. The method is very effi cient, and is widely used (Lynch and Nelson, 1949, 1956; Soule, 1954; Subra, 1954; Ruehle and Ledin, 1955; Ahmed, 1960; Bhan et al., 1969; Rajput and Haribabu, 1971), often with modifi cations (Lynch and Mustard, 1950; Singh, 1960) (Fig. 11.7a).
Flap buddingThe excised bud shape is boat-shaped instead of rectangular (Fig. 11.7b). This procedure has been used in South-east Asia (Naik, 1949; Garner and Cha-udhri, 1976).
Window buddingThis technique is similar to fl ap budding except that a window is created in the fl ap for the bud so that fl ap does not press the bud while being tied (Fig. 11.7c). This method has been used in Queensland, Australia. The bud union occurs in c.4 weeks (Stephens, 1948).
Effect of rootstock
Relative advantages of polyembryonic rootstocksSwamy et al. (1972) described the results of a rootstock trial involving six polyembryonic rootstocks for ‘Baneshan’ and four for ‘Neelum’. Based on growth and fruit yield from 1942 to 1965, ‘Pahutan’ and ‘Goa’ rootstocks were recommended for ‘Neelum’ and ‘Olour’ rootstock was recommended for ‘Baneshan’. Oppenheimer (1960) reported the highest yield for ‘Haden’ mango occurred on polyembryonic ‘Sabre’ rootstock among three different rootstocks tried. George and Nair (1969) conducted a rootstock trial using polyembryonic ‘Chandrakaran’ and ‘Bappakai’ and monoembryonic ‘Puliyan’ rootstocks for

S. Ram and R.E. Litz386
‘Bennett’, ‘Alphonso’ and ‘Baneshan’. ‘Chandrakaran’ and ‘Bappakai’ grew more vigorously and produced higher yields during the fi rst 6 years of growth compared to monoembryonic ‘Puliyan’ rootstock. In south India, polyembryonic ‘Olour’ and ‘Bappakai’ and monoembryonic rootstocks were evaluated using ‘Neelum’ as a scion (Gowder and Irulappan, 1970). ‘Bap-pakai’ was recommended as a rootstock based on fruit yield and uniform growth of ‘Neelum’.
Performance of ‘Dashehari’ on 24 rootstocks (ten polyembryonic and 14 monoembryonic) was reported by Rajan and Pandey (1991a, b). Rootstocks strongly affected tree vigour with tree height ranging from c.4.0 to 6.0 m after 14 years of growth. ‘ST-9’ and ‘Latra’ rootstocks stimulated the most vigorous growth. ‘Ambalvi’, ‘Pahutan’, ‘Olour’, ‘Nakkare’, ‘Mylepelian’, ‘Rumani’, ‘Willard’, ‘Mundappa’, ‘Pahutan’, ‘Ambalvi’ and ‘Moovandan’ caused dwarfi ng. Stem swelling was recorded with ‘Mahmuda Vikarabad’ rootstock; however, the scion girth varied with rootstock. ‘Rumani’ and ‘Ambalvi’ rootstocks caused less scion girth (0.75–0.8 m) than the vigorous ‘ST-9’ and ‘Latra’ rootstocks (0.79–0.84 m). Crown spread was also greater with vigorous rootstocks than with dwarfi ng rootstocks. The canopy of ‘Dashehari’ trees on ‘ST-9’ and ‘Latra’ rootstock was 2.15 and 2.25 times greater than on ‘Rumani’, respectively. Fruit yield varied from 8 kg to 25 kg/tree. Dwarfi ng rootstock resulted in higher fruit yield per unit canopy volume without much change in fruit quality. Polyembryonic rootstocks ‘Ambalvi’, ‘Mylepelian’, ‘Olour’ and ‘Villaikolamban’ were compared with ‘Dashehari’ seedling rootstock using ‘Dashehari’ as the scion (Jauhari et al., 1972; Singh and Singh, 1976). The ‘Dashehari’ seedling rootstock was most vigorous and produced higher yields, whereas ‘Villaikolamban’ resulted in the least vigour and yield. ‘Villaikolamban’ caused dwarfi ng of ‘Alphonso’ and low yields (Anonymous, 1994), and has been recommended as a rootstock for high density ‘Alphonso’ orchards, based upon higher yield/m3 of canopy.
‘Neelum’ fruit quality on ‘Bappakai’ rootstock was better compared to ‘Neelum’ grown on ‘Olour’ and monoembryonic seedlings (Gowder and Iru-lappan, 1970). Larger ‘Neelum’ fruits having high total soluble solids devel-oped on ‘Pahutan’ rootstock in comparison with ‘Goa’, ‘Olour’ and ‘Salem’ rootstocks (Swamy et al., 1972). ‘Madu’ and ‘Gurih’ rootstocks delayed fruit-ing in Indonesia compared to ‘Gurung’, ‘Kopjor’, ‘Budadaja’, ‘Nanas’ and ‘Saigon’ (Kusumo et al., 1971). Polyembryonic rootstock ‘13-1’ has been dem-onstrated to tolerate calcareous soil containing 20% calcium carbonate and saline irrigation water containing >600 ppm chloride (Stoler, 1976; Kadman et al., 1978). Gazit and Kadman (1980) grew ‘Maya’ on ‘13-l’ rootstock on cal-careous soils with up to 20% lime and 250 ppm chloride. ‘Ann’ and ‘Gomera’ polyembryonic rootstocks also show salt tolerance (Galán-Saúco, 1993).
Top and double working
Top working is used to rejuvenate unproductive trees and for grafting on seedling trees (Fig. 11.8a). Two methods are used for top working mango. Branches or main limbs are cut back to 30 cm during winter months or with

Crop Production: Propagation 387
the onset of spring. The new shoots are budded or grafted in the following summer or autumn, usually by shield budding or veneer grafting. Singh (1990) suggested that half of a tree should be top worked in the fi rst year and the other half in the second year, although top working of a complete tree in a single year has been highly successful (Lynch, 1941; Ruehle and Ledin, 1955; Ahmed, 1960; Singh, 1960). Cutting back of limbs in October or early November and veneer grafting in March–April has been recommended in Florida (Carmichael, 1957/58; Miner, 1962); in Uttar Pradesh (India), top working on 8-year-old mango trees was more successful during June (Singh, 1960). After top working, trees come into bearing within 2 years (Furon and Plaud, 1972).
Grafting on an already grafted or budded tree is referred to as double working (Singh, 1960), and double-worked trees therefore consist of three genetically different parts, i.e. the rootstock, interstock and crown (Fig. 11.8b). In Florida, old plantings of ‘Haden’ have been double worked with ‘Tommy Atkins’ (Mitchell, 1971). Double working can also be used to repair trees affected by disease or cold injury and to develop a strong framework. ‘Kala-pady’ has reportedly been used as an interstock in order to dwarf ‘Langra’ (Sen, 1939). In one trial, 12 interstocks were grafted on two rootstocks for ‘Dashehari’; rootstock-interstock combinations signifi cantly affected tree height and vigour. Fruit yield was also infl uenced by rootstock in all the com-binations. ‘Kurukkan’ interstock on ‘ST-9’ rootstock yielded more fruit in comparison with ‘Ambalvi’ on the same rootstock. The maximum yield was obtained with ‘Nakkare’/‘Chandrakaran’ and ‘Ambalvi’/‘Chandrakaran’; whereas the lowest yield was obtained with ‘Ambalvi’/‘ST-9’. Iyer et al. (1990) reported varying success (20–73%) with double-worked ‘Langra’.
Rooting
Mango air layers have been induced to root either while they are still attached to the parent tree or by cutting shoots into pieces containing one to several buds. Several rooting methods have been successful with mango.
LayeringLayering is used chiefl y to propagate monoembryonic mango cultivars on their own roots. Rooting methods involve the initial training of the main stem. Shoots close to ground level are ringed and covered with soil, while other shoots are covered with soil mixture or wet sphagnum moss at the ringed portion and wrapped with polyethylene to maintain moisture (Singh, 1960; Majumder and Mukherjee, 1961; Mukherjee and Majumder, 1963). Auxins such as -naphthalene acetic acid (NAA) and indole butyric acid (IBA) are generally essential in order to induce rooting even after ringing.
Ground layeringVigorous, 1- or 2-year-old shoots are selected near the base of the parent tree. The greyish-green bark portion of the shoot (3–5 cm length) is ringed without

S. Ram and R.E. Litz388
injuring the wood. On the proximal end of the shoot just above the ring, growth regulators are applied and the ringed portion is buried in the soil. Depending upon the growth regulators and their concentration, ringed shoots (or layers) generate roots within 2–3 months. The rooted shoot can then be detached from the mother tree and planted.
Mound layering or stoolingThis technique involves the use of juvenile shoots or shoots with induced juvenility (through heading back and etiolation followed by use of growth regulators), and is more effi cient than simple layering (Majumder and Mukherjee, 1961). One-year-old mango seedlings should be decapitated 25–30 cm above ground level in February–March, and several new shoots will develop from the stem by May. When leaves of the new shoots turn green, the shoots are ringed at their proximal end and treated with 5000 ppm IBA in lanolin paste just above the ring. A soil mound is created around the base of ringed shoots. Rooting occurs after 1–2 months and rooted layers can be detached from the parent plant and established in individual pots. Each layer can generate between four and seven roots. Stooling has been success-ful with many cultivars using growth regulators and rooting cofactors in stool beds (Ram et al., 1981; Sirohi and Ram, unpublished data). Dwarf culti-vars root poorly, but can be stimulated to form thin and fi brous roots by applying auxin, for example IBA (15,000–30,000 ppm) and NAA (2500–10,000 ppm) mixtures in lanolin paste with or without rooting cofactors, such as caf-feic acid (0.046M) and catechol (0.046M). Caffeic acid appears to be highly synergistic to IBA.
Pot layeringWith pot layering, the ringed portion of the shoot is maintained in a soil mix-ture in special pots at a convenient height within the tree canopy. This can be done only during periods of high RH, but the success is low (c.15–20%) (Singh, 1960).
Air layeringAlthough air layering or marcottage is one of the oldest methods for propagat-ing mango, good success has been elusive (Grove, 1947; Garg, 1954; Rangacha-rlu and Rao, 1956; Rao and Rao, 1956; Choudhury, 1959; Jauhari and Nigam, 1962; Rao et al., 1963; Mukherjee and Majumder, 1963; Srivastava, 1963a, b; Mukherjee and Bid, 1965; Sen and Bose, 1966; Bid and Mukherjee, 1969; Basu et al., 1972; Prasad and Singh, 1972; Núñez-Elisea et al., 1992). Juvenility, high RH and moderate temperature are important factors for air layering mango (Singh, 1954; Rao et al., 1963; Rao, 1967; Chhonkar and Singh, 1972; Singh et al., 1979; Ram and Sirohi, 1982a). Cultivar effects on air layering are also pronounced; and vigorous cultivars root better than dwarf cultivars (Garg, 1954; Rao and Rao, 1956; Rao, 1967; Basu et al., 1972; Ram and Sirohi, 1982b).
Layering success can be improved with auxins, such as 2–3% indole ace-tic acid (IAA) and 0.5–2.0% NAA (Thakurta and Dutt, 1941; Singh and Teotia,

Crop Production: Propagation 389
1951; Srivastava, 1963a, b; Núñez-Elisea et al., 1992). Although Núñez-Elisea et al. (1992) used NAA alone, NAA and IBA mixtures not only improve root-ing but also accelerate the production of fi brous roots, which increases sur-vival. Etiolated layers just above the girdle require less auxin than non-etiolated branches. Root regeneration in air layers requires up to 90 days, but emer-gence of roots is fi rst noticeable a month after layering. Sen et al. (1961) and Mukjerjee and Bid (1965) observed that IBA depletes sugar in the bark and wood of the rooting zones. Ram and Sirohi (1982a) studied the interaction of rooting cofactors involved with air layering of mango. Application of 0.01–0.08M IBA in lanolin paste increased root numbers from 2.2 to 2.8 with ‘Dashehari’, but root number could be increased from 2.8 to 6.0 with 0.01–0.10M catechol. Survival was correlated with increased numbers of roots; however, IBA and catechol did not show any additive or synergistic effect on air layering. July was the best month for rooting of layers (>90% success) and winter months were the worst.
Rajan and Ram (1989) showed that 0.0573M NAA and 0.05 × 10 2M chlo-rogenic acid had a synergistic effect on rooting of layers and the number of roots formed during March when applied separately with 0.0735M IBA. In October, NAA and chlorogenic acid showed synergism with respect to num-ber of roots produced, but their effect on rooting was additive. Pyrogallol (0.06 × 10−2M) and 6-benzylaminopurine (0.22 × 10−3M) also had a synergistic effect on root regeneration during October and March. IBA increases endogenous root-ing cofactors and lowers rooting inhibitors, resulting in early rooting.
CuttingsThe potential of mango cuttings to form roots depends on maturity of the cutting, rooting medium, temperature, RH, etc. Girdling or etiolation of stems for a few weeks and use of growth regulators have improved rooting frequency (Thakurta and Dutt, 1941; Gardner and Piper, 1943; Van Overbeek and Gregory, 1945; Said and Shoushan, 1945; Khalifa et al., 1964; Dijkman, 1950; Ledin and Ruehle, 1954; Gowda, 1962; Ahmed, 1964; Sen and Bose, 1964; Mukherjee et al., 1966, 1967; Maurice, 1969; Sen et al., 1969; Ali and Nazir, 1970; Prasad and Pathak, 1970; Bid and Mukherjee, 1972; Bid et al., 1972). Rooting is improved with cuttings from the lower bole of the tree rather than from the upper bole. Mukherjee et al. (1966) reported that cut-tings from 4-year-old trees root more readily than those from 10-year-old trees, alhough Sen et al. (1968) had better success with older shoots. Hard-wood cuttings reportedly root better (Hussain, 1953; Benincasi, 1970) than semi-hardwood cuttings (Singh and Teaotia, 1951). Mango cuttings should be c.15 cm long and 1.2–1.8 cm girth with between three and fi ve buds (Garner and Chaudhri, 1976). Retention of one to two leaves at the apex of the cut-tings has helped rooting, and is attributed to the presence of rooting cofactors in the leaves.
In India and Pakistan, cuttings are generally made during the spring months (Hussain, 1953). Cuttings can be stored for a short time before apply-ing the rooting treatment. Khalifa et al. (1964) rinsed cuttings for 24 h with running tap water, which increased their shelf life. Longevity of cuttings can

S. Ram and R.E. Litz390
also be increased by inserting them in a solution consisting of 100 ppm IBA, 10 mg/litre vitamin B1, 2% ammonium sulphate ((NH4)2SO4) and 2% sucrose. Cuttings can be stored at 4°C for 20 days. Rooting of hardwood cuttings under mist can be improved with IBA and NAA (Lambourne, 1935; Cooper, 1936; Thakurta and Dutt, 1941; Cooper and Stoutemyer, 1945; Dijkman, 1950; Jauhari and Nigam, 1958; Sen and Bose, 1964; Mukherjee et al., 1965; Basu et al., 1966; Sen et al., 1969; Ali and Nazir, 1970; Rajan and Ram, 1983a, b). IBA increases reducing and non-reducing sugars, and stimulates a favourable carbon to nitrogen (C:N) ratio in ungirdled cuttings comparable to girdled cuttings without IBA. Application of NAA to the ring 1 week before cuttings are made and dipping their basal ends in an IBA solution at the time of detaching, increases rooting effi ciency (Van Overbeek and Gregory, 1945; Sen et al., 1969). Low pH potting mixtures (pH 4.5–7.0) reportedly are better for rooting than higher pH mixtures (Kains and McQuestan, 1955). Reddy and Majumder (1975) and Mitra and Bose (1986) used bottom heat at 30°C ± 2° and 35°C, respectively, to stimulate root formation. Reuveni et al. (1991) and Reuveni and Castoriano (1993) treated semi-hardwood cuttings of ‘Turpen-tine’, ‘Gomera’, ‘Sabre’ and ‘13-1’ with bottom heat at 20, 25, 30 and 35°C under mist, and observed most rooting occurred at 25 and 30°C. The percent-age of rooting in cultivars varied from 60–95% with between four and ten roots/cutting. Wounding of the proximal ends of cuttings from 7-month-old seedlings together with bottom heat increases the number of roots per cut-ting (Reddy, 1970).
The involvement of phytochrome in root regeneration of mango cuttings was proposed by Reddy et al. (1975), who observed 92% rooting with etio-lated and red-light-treated cuttings of 7-month-old seedlings, whereas cut-tings in a hot bed treated with 5000 ppm IBA and planted under normal light resulted in 41% rooting.
Reddy and Majumder (1975) achieved 100% rooting of cuttings from 7-month-old mango seedlings treated with 5000 ppm IBA combined with 2000 ppm each of umbelliferone, ruten or quercetin. Phenols and fl avonoids acted as rooting cofactors of IBA with juvenile cuttings. Sadhu et al. (1978) and Reddy and Majumder (1978) observed a synergistic effect of some phe-nolic compounds (e.g. p-hydroxybenzoic acid, p-coumaric acid and ferulic acid) used as a preplanting treatment with auxin for induction of rooting in hardwood mango cuttings. Synergism was more pronounced with IBA than IAA.
Basu et al. (1970) observed no signifi cant difference in the levels of endog-enous rooting substances (i.e. p-hydroxybenzoic acid, p-coumaric acid and abscisic acid) between juvenile and non-juvenile cuttings. Rajan and Ram (1983a, b) studied the levels of endogenous rooting hormones of mango cut-tings with bottom heat under mist and measured increased levels of rooting hormones and low levels of rooting inhibitors. Fayek et al. (1981) reported that shoots of 35-year-old ‘Madu’ contained more rooting inhibitors than shoots of 1-year-old plants. Using 15,000 ppm IBA and 10,000 ppm NAA, Rajan and Ram (1983a, b) obtained c.70% rooting with mature cuttings; about eight fi brous roots were regenerated as a result of each treatment, and all

Crop Production: Propagation 391
cuttings survived in the nursery. Sadhu (1979) and Sadhu and Bose (1980a, b) found that 10,000 ppm cycocel pretreatments of 10-year-old ‘Langra’ resulted in 41% rooting with 2.2 roots/cutting.
Micropropagation
Somatic embryogenesis from cultured nucellar explants of polyembryonic cultivars of Mangifera indica was fi rst reported by Litz et al. (1982, 1984). Sub-sequently, somatic embryogenesis was induced from the cultured nucellus of monoembryonic cultivars (Litz, 1984). DeWald et al. (1989a, b) optimized the protocols for large-scale production of embryogenic cultures in suspension and for somatic embryo maturation and germination. Shoot tip culture of ‘Turpentine’, ‘Gomera’ and ‘13-1’ seedlings has been described by Yang and Ludders (1993); the rate of multiplication was low, and performance of plants in the fi eld has not been tested. The current status of somatic embryogenesis and organogenesis of mango is reviewed by Litz et al., Chapter 18, this volume.
11.4 Comparative Performance of Trees Propagated by Different Methods
Ram and Sirohi (1989) compared the performance of ‘Dashehari’ propagated by cleft grafting, approach grafting, veneer grafting, epicotyl grafting, stool-ing and air layering. The trees were of uniform size at the time of planting. Approach-grafted plants did not establish in the fi eld as well as the other methods. During the fi rst 12 years, epicotyl grafts grew more rapidly than cleft-grafted, veneer-grafted, approach-grafted, stooled and air-layered trees, respectively (Ram, 1993). The development of the crown also followed the same pattern. The trees propagated by rooting did not develop an erect main stem to an adequate height (1.2 m), which hindered cultural operations under the tree. Trees propagated by approach grafting, veneer grafting and epicotyl grafting all produced branches close to the ground. Maximum yields were obtained in trees propagated by epicotyl and cleft grafting followed by veneer grafting, stooling, air layering and approach grafting, respectively. The architecture of cleft-grafted plants was much better than trees propa-gated by other methods; whereas, rooting methods produced twiggy, spread-ing and dwarf trees (Ram, 1993). After 18 years, cleft-grafted trees were superior with respect to architecture and yield, followed by epicotyl and veneer-grafted trees. Rajan and Pandey (1991a, b) conducted experiments with ‘Dashehari’ and ‘Langra’ that were propagated by different vegetative methods using monoembryonic seedling rootstock. Approach-grafted ‘Dashehari’ attained the greatest height followed by epicotyl-grafted, bud-ded and air-layered trees after 7 years. Budded ‘Langra’ grew most vigor-ously, followed by approach-grafted, veneer-grafted, epicotyl-grafted and air-layered trees. Similar growth of scion girth and crown spread was recorded with both cultivars. The type of shoot used for approach grafting

S. Ram and R.E. Litz392
affected tree growth of ‘Dashehari’. Grafting height should be minimized and close contact should be maximized to achieve faster growth when vigorous rootstocks are used.
11.5 Conclusions
Standard methods are widely utilized for propagating mango scion cultivars with increasing effi cacy. In many regions, including India and Mexico, scion cultivars are still being propagated on heterogeneous monoembryonic seed-ling rootstocks (see Crane et al., Chapter 13, this volume), despite the demon-strated advantages of clonal nucellar rootstocks. The potential of clonally propagated monoembryonic mango rootstock has not been properly investi-gated. The genetic heterogeneity of monoembryonic mangoes has been explored neither for stress tolerance nor for their effects on scion growth and development. Somatic embryogenesis could play an important role in such investigations as an alternative propagation method (see Litz et al., Chapter 18, this volume). Other Mangifera species also have interesting attributes, and should be screened for graft compatibility with mango (see Bompard, Chap-ter 2, this volume). The species that could be tested as rootstock for mango might extend mango cultivation to areas where abiotic and biotic stresses currently limit production and could provide a better source for dwarfi ng rootstock for high-density orchards. Mango species growing in swamps or seasonally inundated areas (i.e. Mangifera decandra, Mangifera gedebe, Mangiferainicarpoides, Mangifera griffi thii and Mangifera quadrifi da) represent a promis-ing source of rootstock for the development of mango cultivation on poorly drained soils and inundated lands. In West Kalimantan, Mangifera laurina is occasionally used as a rootstock for commercial mango cultivars on periodi-cally inundated riverbeds, and is now being tried at Sabah (Bompard, 1993). Mangifera zeylanica has been tried as a rootstock for several mango cultivars in Sri Lanka (Parsons, 1931). Interspecifi c hybridization of other Mangiferaspecies with the common mango could also increase the available genetic variability for rootstock development.
The potential for using species in other genera in the Anacardiaceae as rootstock has scarcely been explored. Burns and Prayag (1921) investigated the use of Semecarpus anacardium, Spondias mangifera (Spondias pinnate), Spondiasacuminata and Horigarna grahami as rootstocks for common mango without any success. Anacardium occidentale (cashew) seedlings have been reported to be graft compatible with mango scions and mango fruit on cashew rootstock were reportedly almost double the size of normal fruit with smaller seeds and free from fi bre (Garner and Chaudhri, 1976). Mango rootstock development should receive as much attention as breeding of scion cultivars.
Classical breeding and grafting studies should be greatly expanded to include the enormous genetic variability within the genus. Emerging bio-technological approaches also should not be overlooked as alternative prop-agation methods and as the means to engineer specifi c horticultural traits in candidate rootstocks.

Crop Production: Propagation 393
References
Abdel-Galil, H.A. (1992) Evaluation of certain techniques for germination of mango seed stones. Assiut Journal of Agricultural Sciences 23, 134–151.
Ahmed, S. (1960) Propagation of mangoes. Punjab Fruit Journal 23, 49–53.Ahmed, S. (1964) Propagation of mango by veneer grafting. West Pakistan Journal of
Agriculture Research 2, 32–44. Ahmed, S. (1974) Selection and evaluation of local mango cultivars and strains in North-
east Brazil. In: 6th Report of the UNDP/FAO/CEPED Project Brazil 71/555, Rome, Italy, pp. 1–3.
Ahmed, S. and Chatha, G.A. (1960) Some morphological and anatomical studies of scion buds and bud union zone in mangoes. Punjab Fruit Journal 23, 105–111.
Ahmed, S. and Rashid, A. (1961) Importance of seed in the fruit industry. AgriculturePakistan 12, 252–257.
Ali, N. and Nazir, M. (1970) Effect of auxins on carbohydrate and nitrogen contents of mango cuttings. Pakistan Journal of Science 22, 12–21.
Amin, R.S. (1974) A study on the establishment of mango orchard with wedge graft on ‘in-situ’ grown mango seedling in dry region of the Gujarat State. Haryana Horticul-tural Journal 3, 160–167.
Amin, R.S. (1978) In-situ soft wood grafting in mango. Indian Horticulture 23, 7–10. Anonymous (1950) Annual Report of the Florida Agricultural Experiment Station for the
Year ending June 30. University of Florida, Gainesville, Florida, p. 263. Anonymous (1975) The Philippines Recommends for Mango 1975. Philippine Council
for Agricultural Research, Manila, the Philippines. Anonymous (1994) Production Technology – Mango. Information Bulletin No. 12. Indian
Institute of Horticulture Research, Hessarghatta, Bangalore, India, p. 47.Asadullah, M. and Khan, M.D. (1960) Studies of various factors affecting success in
grafting by approach (inarching) in mangoes. Punjab Fruit Journal 23, 59–70.Bajpai, P.N., Singh, A.R., Yati, V. and Chaturvedi, O.P. (1989) Effect of cultivars and age
of rootstocks on the performance of veneer grafting in mango. Acta Horticulturae 231, 259–262.
Bajwa, B.S. and Ram, A. (1946) Budding in mango seedlings ‘in-situ’. Punjab Fruit Jour-nal 10, 10–13.
Bakhshi, I.C. (1963) Propagation studies with monoembryonic and polyembryonic vari-eties of mango. Punjab Horticultural Journal 3, 185–193.
Basu, R.N., Roychoudhury, N., Bose, T.K. and Sen, P.K. (1966) Rooting of mango cutting. Indian Agriculturist 10, 147–151.
Basu, R.N., Roychoudhary, N., Gosh, B. and Sen, P.K. (1970) Endogenous rooting fac-tors in the mango (Mangifera indica L.) seedling and mature shoots. Horticulture Science 2, 91–98.
Basu, R.N., Roychoudhury, N., Lahiri, B. and Sen, P.K. (1972) Biochemical changes during root formation in Mangifera indica L. air layers. Acta Horticulturae 24, 64–71.
Benincasi, R. (1970) Tentative di applicazione di tecniche di propagazione agamica in pi ante da frutto tropicali presso la stazione sperimentate di M’vuazi (Basso Congo) (Kinshasha). Revista di Agricoltura Subtropicale e Tropicale 64, 182–195.
Ben-ya’acov, A. (1976) Recovery of frost-destroyed mango trees by regrafting at the root shoot transition zone. Proceedings of the American Society for Horticulture Science Tropical Region 24, 151–156.
Bhambota, J.R., Rajput, M.S. and Sandhu, K.S. (1971) Veneer grafting a successful method of propagation. Punjab Horticultural Journal 11, 40–43.

S. Ram and R.E. Litz394
Bhan, K.C., Samaddar, R.N. and Yadav, P.C. (1969) Chip budding and stone-grafting of mangoes in India. Tropical Agriculture Trinidad 46, 247–253.
Bid, N.N. and Mukherjee, S.K. (1969) Varietal response to etiolation and growth-regulator treatment in air-layering of mango (Mangifera indica L.). Indian Journal of Agricul-tural Science 39, 1013–1019.
Bid, N.N. and Mukherjee, S.K. (1972) Studies into the effects of forced shoot etiolation and different media on the rootage of Mangifera indica L. cuttings. Acta Horticultu-rae 24, 77–81.
Bid, N.N., Mukherjee, S.K. and Majumder, P.K. (1972) Studies on different methods of propagation of Mangifera indica with respect to success, survival, growth and their clonal multiplication by stooling. Acta Horticulturae 24, 85–88.
Bompard, J.M. (1993) The genus Mangifera rediscovered. The potential contribution of wild species to mango cultivation. Acta Horticulturae 341, 69–77.
Burns, W. and Prayag, S.H. (1921) The Book of the Mango. Department of Agriculture Bulletin, Bombay, India.
Carmichael, W.W. (1957/58) Rejuvenation of old mango groves by hedging or top-working to new varieties. Proceedings of the Florida State Horticultural Society 70, 290–291.
Chacko, E.K. and Singh, R.N. (1971) Studies on the longevity of papaya, phalsa, guava and mango seeds. Proceedings of the International Seed Testing Association 36, 147–158.
Chauran, O.R., Manica, I., Pinheiro, R.V.R., Conde, A.R. and Chaves, J.R.P. (1979) The effect of storage time, seed treatment and fungicide on seed germination and growth of mango seedlings. Revista Ceres 26, 1–12.
Chhonkar, V.S. and Singh, R.K. (1972) Propagation of Mangifera indica L. by air layering. Acta Horticulturae 24, 89–92.
Choudhury, K.R. (1959) A short note on substitutes for moss as plastering material in air layering of mango. Science and Culture 25, 152–153.
Cooper, W.C. (1936) Transport of root-forming hormones in woody cuttings. Plant Phys-iology 11, 779.
Cooper, W.C. and Furr, J.R. (1945) The cinchona veneer graft method of propagating subtropical fruit trees. Proceedings of the Florida State Horticultural Society 58,176–180.
Cooper, W.C. and Stoutemyer, V.T. (1945) Suggestion for the use of growth substances in the propagation of tropical plants. Tropical Agriculture Trinidad 22, 21.
Degani, C., Cohen, M., Reuveni, O., El-Batsri, R. and Gazit, S. (1993) Frequency and characteristics of zygotic seedlings from polyembryonic mango cultivars, deter-mined using isozymes as genetic markers. Acta Horticulturae 341, 78–81.
DeWald, S.G., Litz, R.E. and Moore, G.A. (1989a) Optimizing somatic embyro pro-duction in mango. Journal of the American Society for Horticultural Science 114, 712–716.
DeWald, S.G., Litz, R.E. and Moore, G.A. (1989b) Maturation and germination of man-go somatic embryos. Journal of American Society for Horticultural Science 114,837–841.
Dhunaga, D.B., Arvindakshan, M. and Gopikumar, K. (1989) Standardization of stone grafting in mango. Acta Horticulturae 231, 170–174.
Dijkman, M.J. (1950) Rooting of ‘Haden’ mango (Mangifera indica L.) leaf bud cuttings. Science 111, 663–665.
Eiadthong, W., Yonemori, K., Sugiura, A., Utsunomiya, N. and Subhadrabandhu, S. (1999) Identifi cation of mango cultivars of Thailand and evaluation of their genetic variation using the amplifi ed fragments by simple sequence repeat-(SSR-) anchored primers. Scientia Horticulturae 82, 57–66.

Crop Production: Propagation 395
Faruque, A.H.M. and Fakir, M.M.A.S. (1973) Propagation of mango by different methods of grafting. Bangladesh Horticulture 1, 25–28.
Fayek, M.A., Salim, H.H., Ibrahi, F.A. and Menibary, M.A. (1981) Tentative identifi cation of the major inhibiting compound in mango shoots. Annals of Agricultural Science 16, 209–216.
Furon, V. (1966) La multiplication du manguier au Senegal. Fruits 21, 189–193.Furon, V. and Plaud, G. (1972) Topworking mango trees in Senegal. Fruits 27, 293–296. Galán-Saúco, V. (1993) The situation of mango culture in the world. Acta Horticulturae
341, 31–41.Galán-Saúco, V. (1999) El Cultivo del Mango. Ediciones Mundi-Prensa, Madrid, Spain. Gandhi, S.R. (1942) Recent advances in horticultural practices. Poona Agriculture Col-
lege Magazine 34, 86.Gandhoke, M.M.S. (1993) Flush grafting in mango – way to sure success. (Abstract).
Golden Jubilee Symposium Horticultural Research – Changing Scenario. Horticul-tural Society of India, Bangalore, India, p. 445.
Gangolly, S.R., Singh, R., Katyal, S.L. and Singh, D. (1957) The Mango. Indian Council of Agriculture Research, New Delhi, India.
Gardner, D.E. and Piper, R.B. (1943) Base of propagation of some sub-tropical fruits by cuttings from young seedlings. Proceedings of the Florida State Horticultural Soci-ety 56, 124–126.
Garg, M.L. (1954) Mango grafting made easy and cheap. Indian Journal of Horticulture 11, 145–146.
Garner, R.J. and Chaudhri, S.A. (1976) Mangifera indica – mango. In: Garner, R.J. and Chaudhri, S.A. (eds) The Propagation of Tropical Fruit Trees. Food and Agriculture Organization of the United Nations and the Commonwealth Agricultural Bureau, Wallingford, UK, pp. 403–474.
Gaur, N.V.S. (1984) Comparative evaluation of selected methods of mango propagation. Punjab Horticultural Journal 24, 1–6.
Gazit, S. and Kadman, A. (1980) 13-1 mango rootstock selection. HortScience 15,669.
George, P.V. and Nair, T.N.K. (1969) On the performance of mono- and polyembryonic rootstocks in mango grafts. Agriculture Research Journal Kerala 7, 7–9.
Giri, A. (1966) Transplanting success with varying stem girths of mango seedlings. Agri-culture Pakistan 17, 195–200.
Giri, A. and Chaudhery, M.Y. (1966) Relation of mango stone weight to its germination and seedling vigor. Pakistan Journal of Science 18, 148–150.
Gonzalez, A., Coulson, M. and Brettell, R. (2002) Development of DNA markers (ISSRs) in mango. Acta Horticulturae 575, 139–143.
Gowda, M.H.M. (1962) Some methods of propagation of mango. In: Fruit Nursery Prac-tices in India. Directorate of Extension, Ministry of Food and Agriculture, New Delhi, India, pp. 54–64.
Gowder, R.B. and Irulappan, I. (1970) Performance of Neelum variety of mango (Mangifera indica L.) on polyembryonic rootstocks. Madras Agriculture Journal 57, 29.
Grant, J.W. and William, A.N.P. (1949) Burma Fruits and their Cultivation. Burma Bul-letin 30. Department of Agriculture, Rangoon, Burma.
Grove, W.R. (1947) Wrapping air layers with rubber. Proceedings of the Florida State Horticultural Society 60, 184–187.
Gunjate, R.T. (1989) Standardization of stone grafting for the Konkan region. Acta Hor-ticulturae 231, 164–167.
Gunjate, R.T. and Limaye, Y.P. (1978) Veneer grafting of mango seedlings raised in earth-en pots. Journal of Maharashtra Agricultural University 3, 61–62.

S. Ram and R.E. Litz396
Gunjate, R.T., Uradya, V.S. and Limaye, V.P. (1976) Effect of season and defoliation of scion shoots on success in veneer grafting in ‘Alphonso’ mango. Journal of Maha-rashtra Agricultural University 3, 51–52.
Gupta, O.P., Jawanda, J.S. and Sharma, K.C. (I988) Stone grafting in mango under Jammu conditions. Progressive Horticulture 20, 11–14.
Hayes, W.B. (1957) Fruit Growing in India, 3rd edn. Kitabistan, Allahabad, India. Higgins, J.E. (1910) Shield budding the mango. Bulletin of the Hawaii Agriculture
Experiment Station 20, 1.Hussain, I. (1953) Some studies on the effect of hormones on the rooting of cuttings of
horticultural plants. PhD thesis, University of Punjab, Lahore, Pakistan. Iyer, C.P.A., Subramanyam, M.D. and Dinesh, M.R. (1990) A simple procedure for dou-
ble working in mango. Journal of Maharashtra Agricultural University 15, 244–245.
Jagirdar, S.A.P. and Ali, S.K. (1968) Effect on scion variety and methods of raising stock on bud-take in mango. West Pakistan Journal of Agricultural Research 6, 83–87.
Jagirdar, S.A.P. and Bhatti, M.S. (1968) Effect of type of scion wood and age of stock on the success of veneer grafting in mango (Mangifera indica). West Pakistan Journal of Agricultural Research 6, 88–97.
Jagirdar, S.A.P., Nizamani, M.H. and Shaikh, M.A. (1968) Varietal response of veneer grafting in mango. West Pakistan Journal of Agricultural Research 6, 86–89.
Jauhari, O.S. and Nigam, Y.N. (1958) Effect of hormones on rooting in cuttings of mango.Kanpur Agriculture College Journal 17, 1–3.
Jauhari, O.S. and Nigam, V.N. (1962) Air layering of mango. In: Fruit Nursery Practices in India. Directorate of Extension, Ministry of Food and Agriculture, New Delhi, India, pp. 77–81.
Jauhari, O.S. and Singh, R.D. (1970) Effect of bud activation, lopping and wrapping materials on budding in mango (Mangifera indica) variety Langra. Punjab Horticul-tural Journal 10, 198–202.
Jauhari, O.S., Teotia, S.S. and Upadhyay, S.K. (1972) Rootstock studies in Mangifera indica L. Acta Horticulturae 24, 107–109.
Jinturkar, S.P. and Narwadkar, P.R. (1989) Effect of environmental conditions on the success of epicotyl grafting in mango. Acta Horticulturae 231, 252–255.
Juliano, J.B. (1934) Origin of embryos in the strawberry mango. Philippine Journal of Science 54, 553–556.
Juliano, J.B. (1937) Embryos of carabao mango (Mangifera indica L.). Philippines Agri-culture 25, 749–760.
Kadman, A., Gazit, S. and Ziv, G. (1978) Experiments with the selection of mango root-stocks for the Southern Arava. Israel Journal of Botany 27, 34–35.
Kains, M.G. and McQuestan, L.M. (1955) Propagation of Plants. Orange Judd, New York.Kanwar, J.S. and Bhajwa, M.S. (1974) Propagation of mango by side grafting. Indian
Journal of Agricultural Science 44, 270–272.Kashkush, K., Jinggui, F., Tomer, E., Hillel, J. and Lavi, U. (2001) Cultivar identifi cation
and genetic map of mango (Mangifera indica). Euphytica 122, 129–136.Kashyap, R., Jyotishi, R.P. and Sharma, A.B. (1972) Techniques of side-grafting in mango.
Acta Horticulturae 24, 99–100.Khalifa, A.S., EI-Azzouni, M.M. and Wah, Y.A. (1964) Physiological studies on mango
cuttings. Indian Journal of Horticulture 21, 179–185.Khan, A.A. (1960) Mango budding ‘in-situ’. A new technique likely to revolutionise the
mango industry. Punjab Fruit Journal 23, 113–116.Kulkarni, V.I., Ratnam, K.K. and Ramkrishna, G. (1989) Propagation studies in mango.
Acta Horticulturae 231, 186–191.

Crop Production: Propagation 397
Kulwal, L.V. and Tayde, G.S. (1989) Studies on propagation of mango varieties by soft wood grafting under Akola condition. Acta Horticulturae 231, 256–258.
Kusumo, S., Soehendro, R. and Rijanto, B. (1971) Mango rootstock experiment at Poh-djentrek. Bulletin Hortikultura No.3. Tjaban Lembaga Penelitian Horticulture, Indonesia from Ringkasan. Publikasi dan Laporan Penelitian Pertanian 2, 333.
Lamboume, J. (1935) The etiolation shoot method of fruit propagation. Malaysian Agri-culture Journal 23, 514–527.
Ledin, R.B. and Ruehle, G.D. (1954) Mango selection, propagation and culture. AnnualReport of Florida Agricultural Experimental Station. University of Florida, Gainesville,Florida, pp. 291–292.
Litz, R.E. (1984) In vitro somatic embryogenesis from nucellar callus of monoembryonic mango. HortScience 19, 715–717.
Litz, R.E., Knight, R.J. and Gazit, S. (1982) Somatic embryos from cultured ovules of Mangifera indica L. Plant Cell Report 1, 264–266.
Litz, R.E., Knight, R.J, Jr and Gazit, S. (1984) In vitro somatic embryogenesis from Mangifera indica L. callus. Scientia Horticulturae 22, 233–240.
Luthra, J.C. and Sharma, M.M.L. (1946) Some studies on the conductability and histol-ogy of grafted mango shoots. Horticultural Botany 35, 4.
Lynch, S.J. (1941) Nursery Propagation and Topworking of Mangoes. Bulletin of Florida Agriculture Experimental Station 560. University of Florida, Gainesville, Florida, 4 pp.
Lynch, S.J. and Mustard, M.J. (1950) Mangoes in Florida. Bulletin of the Florida Depart-ment of Agriculture 135. University of Florida, Gainesville, Florida.
Lynch, S.J. and Nelson, R.O. (1949) Mango budding. Proceedings of the Florida State Horticultural Society 62, 207–209.
Lynch, S.J. and Nelson, R.O. (1956) Current methods of vegetative propagation of avo-cado, mango, lychee and guava in Florida. Ceiba 4, 315–337.
Madalgeri, M.B., Hulamani, N.C. and Patil, V.R. (1989) Response of mango varieties and hybrids to epicotyl grafting. Progressive Horticulture 21, 173–175.
Maheshwari, P., Sachar, R.C. and Chopra, R.N. (1955) Embryological studies in mango (Mangifera indica L.). In: Proceedings of Indian Science Congress, Baroda, Gujarat, India, p. 233.
Majhail, H.S. and Singh, K.K. (1962a) Inarching in mango. I. The effect of Alkathene wrapper, time of inarching and size of stock seedling. Punjab Horticultural Journal 2, 33–43.
Majhail, H.S. and Singh, K.K. (1962b) Inarching in mango II. The optimum period of grafting and age of stock seedling. Punjab Horticultural Journal 2, 109–113.
Majumder, P.K and Mukherjee, S.K. (1961) A note on the propagation of root-stocks in mango. Indian Journal of Horticulture 18, 167–168.
Majumder, P.K. and Rathore, D.S. (1970) Epicotyl grafting in mango (Mangifera indica L.). Current Science 39, 142.
Maurice, J. (1969) Section-grafting and rooting of tropical fruit trees. World Crops 21,265–267.
Miner, J.T. (1962) Top working mango trees in Florida. Proceedings of the Tropical Region of the American Society for Horticultural Science 6, 31–33.
Mitchell, E.F. (1971) Mango production and marketing practices – Florida. Proceedings of the Florida State Horticultural Society 84, 307–310.
Mitra, S.K. and Bose, T.K. (1986) Mango. In: Bose, T.K. (ed.) Propagation of Tropical and Subtropical Horticultural Crops. Naya Prokash, Calcutta, India, pp. 206–207.
Moreuil, C. (1963) Le manguier au Congo. Fruits 18, 295–301.Mukherjee, S.K. and Bid, N.N. (1965) Propagation of mango (Mangifera indica L.) II.
Effect of etiolation and growth regulator treatment on the success of air-layering. Indian Journal of Agricultural Science 35, 305–314.

S. Ram and R.E. Litz398
Mukherjee, S.K. and Majumder, P.K. (1961) ‘Veneer’ grafting in mango has its own advantages. Indian Horticulture 6, 3.
Mukherjee, S.K. and Majumder, P.K. (1962) ‘Veneer’ grafting in mango. In: Fruit Nursery Practices in India. Directorate of Extension, Ministry of Food and Agriculture, New Delhi, India, pp. 65–66.
Mukherjee, S.K. and Majumder, P.K. (1963) Standardization of root-stocks of mango. I. Studies on the propagation of clonal root-stocks by stooling and layering. IndianJournal of Horticulture 20, 204–209.
Mukherjee, S.K. and Majumder, P.K. (1964) Effect of different factors on the success of veneer grafting. Indian Journal of Horticulture 21, 46–50.
Mukherjee, S.K., Majumder, P.K., Bid, N.N. and Goswami, A.N. (1965) Clonal propaga-tion of mango (Mangifera indica L.) through cuttings. Current Science 34, 434–435.
Mukherjee, S.K., Majumder, P.K. and Goswami, A.M. (1966) Effect of position of the shoots on the rooting of mango cuttings. Science and Culture 32, 377–378.
Mukherjee, S.K., Majumder, P.K., Bid, N.N. and Goswami, A.M. (1967) Standardization of rootstocks of mango (Mangifera indica L.) II. Studies on the effects of source, invigoration and etiolation on the rooting of mango cuttings. Journal of Horticul-tural Science 42, 83–87.
Mulat, B. (1959) Mango grafting. Fruits 14, 219–223. Naik, K.C. (1941) Studies on the propagation of the mango. Mangifera indica L. Indian
Journal of Agriculture Science 11, 736–768.Naik, K.C. (1949) South Indian Fruits and their Culture. P. Varadachary and Company,
Madras, India.Narwadkar, P.R. and Anserwadekar, K.W. (1989) Effect of growth regulator on the suc-
cess of epicotyl grafting in mango. Acta Horticulturae 231, 175–178.Neto, M.T. de C., Fonseca, N., Filho, H.P.S. and Juior, A.T.C. (2002) Propagacao e padrao
da muda. In: Genu, P.J. de C. and Pinto, A.C. de Q. (eds) A Cultura da Mangueira.Embrapa Informação Tecnológica, Brasília, Brazil, pp. 117–136.
Núñez-Elisea, R., Caldeira, M.L., Ferreira, W. and Davenport, T.L. (1992) Adventitious rooting of ‘Tommy Atkins’ mango air layers induced with naphthaleneacetic acid. HortScience 27, 926.
Oppenheimer, C. (1960) The relationship between tree size and yield in mango and avocado. Horticulture Advance 4, 6–15.
Panickar, P. and Desai, A.G. (1989) Effect of age of scion mother tree, different fl ushes of rootstock and in-situ grafting on success and growth of soft wood grafts of ‘Alphonso’mango. Progressive Horticulture 21, 141–144.
Parisot, E. (1988) Study of the growth rhythm in young mango plants. 1. Description, germination and storage of polyembryonic mango seeds. Fruits 43, 97–105.
Parsons, T.H. (1931) The mango in Ceylon. Tropical Horticulturist 76, 119–211.Patil, A.A., Vadigeni, B.G. and Nalawadi, U.G. (1991) Response of mango varieties to
stone grafting. Current Research of University of Agricultural Sciences (Bangalore)20, 135–136.
Patwardhan, G.B. and Deshmukh, G.B. (1931) A Handbook of Horticultural Practices. Patwardhan, Poona, India.
Paul, W.R.C. and Guneratnam, S.C. (1937a) The propagation of mango in Jaffna. I. Trop-ical Agriculturist 88, 86–91.
Paul, W.R.C. and Guneratnam, S.C. (1937b) The propagation of mango in Jaffna. II. Tropical Agriculturist 88, 331–337.
Paul, W.R.C. and Guneratnam, S.C. (1938) Mango stocks. Tropical Agriculturist 90, 34–35.Pinheiro, R.V.R., Anderson, O. and Fortes, J.M. (1970) Comparacao de modalidades de enx-
ertia na propagacao da mangueira (Mangifera indica L.). Review Ceres 17, 264–273.

Crop Production: Propagation 399
Pinto, A.C.Q., Andrade, S.R.M., Amaro, A.A. and Gomes, U. (2004) Mango industry in Brazil. Acta Horticulturae 645, 37–50.
Pope, W.T. (1929) Mango Culture in Hawaii. Hawaii Agricultural Experiment Station Bulletin 58. Hawaii Agricultural Experiment Station, Hawaii, 58 pp.
Pope, W.T. and Storey, W.B. (1933) Grafting Tropical Fruit Trees in Hawaii. Hawaii Agricul-tural Experimental Station Circular 6. Hawaii Agricultural Experiment Station, Hawaii.
Popenoe, W. (1959) EI mango. Suelo Tico 11, 23–27.Prasad, A. and Pathak, R.A. (1970) Response of plant growth regulators to mango
(Mangifera indica L.). Tropical Agriculturist 126, 95–106.Prasad, A. and Singh, A.R. (1972) Mango propagation studies at Government Horticul-
tural Research Institute, Saharanpur, India. Acta Horticulturae 24, 93–98.Prasad, A., Singh, R.D. and Sirohi, R.S. (1973) Comparative study of veneer grafting and
patch budding in Mangifera indica L. cv Dashehari. Punjab Horticultural Journal 13, 30–34.
Radhamony, P.S., Gopikumar, K. and Valsalakumari, P.K. (1989) Varietal responses of scion to stone grafting in mango for commercial propagation. South Indian Horti-culture 37, 298–299.
Rajan, S. and Pandey, D. (1991a) Studies on propagational techniques in mango. Central Institute of Horticulture for Northern Plains Annual Report 1989–90. Central Insti-tute of Horticulture for Northern Plains, Lucknow, India, pp. 19–20.
Rajan, S. and Pandey, D. (1991b) Standardization of root stock on mango. Central Insti-tute of Horticulture for Northern Plains Annual Report 1989–90. Central Institute of Horticulture for Northern Plains, Lucknow, India, pp. 17–29.
Rajan, S. and Ram, S. (1983a) Some factors affecting root regeneration in mango cutting in mist and hot bed. Progressive Horticulture 15, 11–16.
Rajan, S. and Ram, S. (1983b) Some new concepts in rootage of mango. Progressive Horticulture 15, 155–156.
Rajan, S. and Ram, S. (1989) Studies on the root regeneration in mango air layers. Acta Horticulturae 231, 192–197.
Rajan, S. and Sinha, G.C. (1987) Use of aluminium foil for increasing veneer grafting success under adverse conditions. Progressive Horticulture 19, 141–142.
Rajput, C.B.S. and Haribabu, R.S. (1971) Recent techniques of mango propagation. World Crops 23, 146–148.
Ram, S. (1993) Factors affecting mango tree architecture. Acta Horticulturae 341, 177–191.Ram, S. and Bist, L.D. (1982) Studies on veneer grafting of mango in Tarai. Punjab Hor-
ticultural Journal 22, 64–71.Ram, S. and Sirohi, S.C. (1982a) Effect of growth regulators on air layering in mango.
(Abstract). National Seminar on Plant Propagation. Department of Horticulture BidhanChandra Krishi Vishwavidyalaya and Society for Advancement of Horticulture, Kalyani, West Bengal, pp. 72–73.
Ram, S. and Sirohi, S.C. (1982b) Performance of mango cultivar Dashehari trees propa-gated by different vegetative methods. (Abstract). National Seminar on Plant Propa-gation. Department of Horticulture Bidhan Chandra Krishi Vishwavidyalaya and Society for Advancement of Horticulture, Kalyani, West Bengal, pp. 93–94.
Ram, S. and Sirohi, S.C. (1989) Performance of Dashehari mango trees propagated by different vegetative methods. Acta Horticulturae 231, 210–215.
Ram, S., Sirohi, S.C. and Bist, L.D. (1981) Interaction of diphenolics with auxin in stool-ing of mango. Punjab Horticultural Joumal 21, 46–52.
Ramirez Diaz, J.M. (1973) The effect of cutting back root stock on the take and initial growth of the ‘Kent’ mango scion and its relationship with apical dominance. Agri-cultura Technica en Mexico 3, 245–252.

S. Ram and R.E. Litz400
Rangacharlu, V.S. and Rao, M.V. (1956) Air grafting of mango. (A new technique in the grafting of mango without the aid of soil or water). Andhra Agricultural Journal 1,105–109.
Rao, S.N., Swamy, G.S., Seeramulu, P. and Reddy, R.N. (1963) Growth regulators in propagation of mango by air layering. Punjab Horticultural Journal 3, 175–179.
Rao, V.N.M. (1967) Propagation practices. In: The Mango Handbook. Indian Council for Agricultural Research (ICAR), New Delhi, India, pp. 32–69.
Rao, V.N.M. and Rao, I.K.S. (1956) Mango grafting with the aid of plastic fi lm wrapper. South Indian Horticulture 4, 54–55.
Reddy, C.V. and Melanta, K.R. (1988) Effect of age of root stocks on the success of soft wood grafting of mango in container and in situ. South Indian Horticulture 36, 143–145.
Reddy, Y.N. (1970) Physiological and biochemical studies on the regeneration of mango (Mangifera indica L.) and guava (Psidium guajava L.) cuttings. PhD thesis, Indian Agricultural Research Institute, New Delhi, India.
Reddy, Y.N. and Majumder, P.K. (1975) Bottom heat – a new technique for rooting of hard wood cuttings of tropical fruits. Current Science 44, 444–445.
Reddy, Y.N. and Majumder, P.K. (1978) Synergism of phenols and fl avonoids with IBA in regeneration of mango and guava cuttings. Vatika 1, 37–44.
Reddy, Y.N., Majumder, P.K., Pandey, R.M. and Singh, R.N. (1975) Phytochrome medi-ated root generation in mung (Phaseolus aureus Roxb) and mango (Mangifera indicaL.) cuttings. Current Science 44, 509–510.
Reuveni, O. and Castoriano, M. (1993) Effect of bottom heat temperature on rooting of mango cuttings of different cultivars. Acta Horticulturae 341, 288–291.
Reuveni, O., Kuepper, W. and Wiebel, J. (1991) Rooting of mango cuttings under inter-mittent mist. Acta Horticulturae 291, 174–181.
Ruehle, G.D. and Ledin, R.B. (1955) Mango Growing in Florida. Bulletin of Florida Agri-culture Experimental Station 574. University of Florida, Gainesville, Florida, 90 pp.
Sachar, R.C. and Chopra, R.N. (1957) A study of the endosperm and embryo in Mangifera. Indian Journal of Agricultural Science 27, 219–228.
Sadhu, M.K. (1979) Effect of pretreatment of stock plants of mango with cycocel, ethrel and morphactin on the rooting of cutting and air layers. Scientia Horticulturae 10,363–368.
Sadhu, M.K. and Bose, S. (1980a) Effect of ethylene on rooting of cuttings and air layers of mango, guava and water-apple. Indian Journal of Horticulture 37, 335–337.
Sadhu, M.K. and Bose, S. (1980b) Note on promotion of rooting in diffi cult-to-root fruit-tree cuttings with 2-chloroethyl phosphonic acid and acetylene in the presence of auxins. Indian Journal of Agriculture Science 50, 448–450.
Sadhu, M.K., Bose, S. and Saha, L. (1978) Auxin synergist in rooting of mango cuttings. Scientia Horticulturae 9, 381–387.
Said, H. and Shoushan, A.A. (1945) Root formation of cuttings of plants which normally do not root. Nature 155, 791.
Schnell, R.J. and Knight, R.J. (1993) Genetic relationships among Mangifera spp. based upon RAPD markers. Acta Horticulturae 341, 86–92.
Sen, P.K. (1939) Annual Report of the Fruit Research Station, Sabour Bihar. Superinten-dent Government Printing, Patna, Bihar, India.
Sen, P.K. and Bose, T.K. (1964) Physiological studies on regeneration of roots. VI: Effects of growth substances on root formation in justicia, mulberry and mango stem cut-tings and concomitant changes in carbohydrate and nitrogenous substances in the rooting tissues. Indian Agriculturist 8, 102–119.
Sen, P.K. and Bose, T.K. (1966) Physiological studies on regeneration of roots. VII. Rela-tion of chemical composition to the regeneration of roots in airlayers of mango

Crop Production: Propagation 401
(Mangifera indica L.) and Litchi (Litchi chinensis Sam.) VIII. Role of leaves in rooting of stem cuttings. Indian Agriculturist 10, 75–86.
Sen, P.K., Bose, T.K. and Sasibhushan, T. (1961) Propagation of mango, Mangifera indicaL., by air-layering with growth substances. Indian Agriculturist 5, 167–172.
Sen, P.K., Basu, R.N., Bose, T.K. and Roychoudhary, N. (1968) Rooting of mango cut-tings under mist. Current Science 37, 144–146.
Sen, P.K., Basu, R.N., Bose, T.K. and Roychoudhary, N. (1969) Mist propagation of fruit tree cuttings. Plant Science 1, 216–219.
Serpa, D. (1964) Propagacion del mango. Faculty of Agronomy, Universidad Central de Venezuela 2, 24.
Shant, P.S. and Saproo, B.L. (1974) Effects of various stages of fruit maturity and different levels of pulp extraction on the germination of mango stones. Bangladesh Horticul-ture 2, 49–50.
Simao, S. (1960) Seeds of green mangoes for rootstock production. Review Agriculture Piracicaba 35, 183–188.
Singh, B. and Ram, A. (1946) Budding of mango seedlings in situ, a brief résumé of work done during the years 1943–45. Punjab Fruit Journal 10, 10.
Singh, J.R. and Srivastava, R.P. (1961) Budding as a commercial method of mango prop-agation. Gardening 3, 19–21.
Singh, J.R. and Srivastava, R.P. (1962) Studies in budding of mango. I. Indian Journal of Horticulture 19, 130–134.
Singh, K.K. and Jawanda, J.S. (1962) Some promising seedling mango trees of Punjab. Punjab Horticultural Journal 2, 133–138.
Singh, L. and Khan, A.A. (1942) Mango budding in situ. Punjab Fruit Journal 6, 195–206.Singh, L. and Khan, A.A. (1943) How to prolong the life of mango bud-wood. Punjab
Fruit Journal 7, 1264–1265.Singh, L. and Khan, A.A. (1946) Mango budding in situ, a new technique likely to revo-
lutionise the mango industry. Punjab Fruit Journal 10, 14.Singh, L.B. (1954) Propagation of mango by air layering for root-stocks. Proceedings of
the American Society of Horticultural Science 63, 128–130.Singh, L.B. (1960) The Mango. Leonard Hill (Books) Limited, London.Singh, M.P., Gill, S.S. and Khajuria, R.N. (1989) Standardization of propagation tech-
niques in mango. Acta Horticulturae 231, 179–181.Singh, N.P. and Srivastava, R.P. (1979a) Studies on the different aspects involved in
veneer grafting in mango. Progressive Horticulture 11, 67–74.Singh, N.P. and Srivastava, R.P. (1979b) Practical hints on veneer grafting in mango.
Indian Horitculture 23, 6–8.Singh, N.P., Srivastava, R.P., Singh, R. and Rajput, M.S. (1979) Seasonal effect on success in
different methods of mango propagation. Indian Journal of Horticulture 36, 135–139.Singh, R. and Singh, C.P. (1998) Planting and care, propagation, rootstock and irrigation
in mango crops. In: Srivastava, R.P. (ed.) Mango Cultivation. International Book Distributing Co., Lucknow, India, pp. 101–127.
Singh, R.N. (1990) Mango. Indian Council of Agricultural Research, New Delhi, India. Singh, S. (1946) Propagation of the mango in the Punjab. Punjab Fruit Journal 10, 8.Singh, S.N. and Teaotia, S.S. (1951) Effect of some hormones on rootage of mango. Sci-
ence and Culture 17, 207–210. Singh, U.R. and Singh, A.P. (1976) Rootstock studies in mango (Mangifera indica). Pro-
gressive Horticulture 8, 13–19. Soule, M.J., Jr (1954) Infl uence of type of graftage, age of plant, and certain environmen-
tal factors on the loss of water by young mango buddings. PhD thesis, University of Florida, Gainesville, Florida.

S. Ram and R.E. Litz402
Soule, M.J., Jr (1971) Anatomy of the bud union in mango (Mangifera indica L.). Journalof the American Society of Horticultural Science 96, 380–383.
Srivastava, R.P. (1963a) Studies in the response of plant growth regulators in air-layered shoots of mango (Mangifera indica L.). Journal of Science Research, Banaras Hindu University 11, 1–3.
Srivastava, R.P. (1963b) Propagation of mango and guava by transported buds. Scienceand Culture 29, 145–146.
Srivastava, R.P. (1989) Propagation of mango by newer techniques. Acta Horticulturae 231, 266–267.
Stephens, S.E. (1948) The mango in Queensland. Queensland Agricultural Journal 68,71–81; 146–153; 208–215.
Stephens, S.E. (1960) Mango growing in Queensland. Queensland Agriculture Journal 86, 761–766.
Stoler, S. (1976) Mango rootstock trial in the Jordan valley. Special Publication. Agricul-tural Research Organisation Israel 65, 95–98.
Sturrock, T.T. (1968) Nucellar embryos of the mango. Proceedings of the Florida State Horticultural Society 80, 350–354.
Subra, P. (1954) Greffage du manguier au Cameroun. Fruits 9, 498–501. Sundara Rao, Y.R. (1956) Preservation of bud wood. South Indian Horticulture 4, 3–12.Swamy, G.S., Rao, E.Y.R. and Raju, D.S. (1972) Polyembryonic rootstocks for mango.
Acta Horticulturae 24, 110–113. Talukdar, M.R. and Ahmed, S. (1965) Success of in arching done on three varieties of
mango on young stocks at Lyallpur. Pakistan Journal of Science 17, 72–74.Tanaka, T. (1939) A new method in mango propagation. Proceedings of the Agriculture
Society of Trinidad 39, 232–287. Teaotia, S.S. (1963) Budding in mango (Mangifera indica L.). Kanpur Agriculture
College Journal 23, 49–50.Teaotia, S.S. and Maurya, V.N.M. (1970) Studies on the propagation of mango by bud-
ding. Progressive Horticulture 2, 35–44.Thakurta, A.G. and Dutt, B.K. (1941) Vegetative propagation of mango from ‘gootee’
(marcottage) and cuttings by treatment of high concentration of auxin. Current Sci-ence 10, 297.
Thrower, L.B. (1954) Commercial horticulture in the Sudan. 2. Mango cultivation. Bulletin Ministry of Agriculture Sudan 11, 66.
Torres, J.P. (1949) Splice grafting of mango. Philippines Journal of Agriculture 14, 247–255.
Torres, J.P. (1960) Splice grafting of mango. Punjab Fruit Journal 23, 73–76.Traub, H.P. and Auchter, B.C. (1934) Propagation experiments with avocado, mango
and papaya. Proceedings of the American Society of Horticulture Science 30, 382–385.
Ullah, B. and Ali, S. (1955) Review of mango budding in situ in the Punjab. Pakistan Review of Agriculture 2, 74–76, 96.
Van Overbeek, J. and Gregory, L.E. (1945) Investigations on the fundamentals of plant propagation by means of cuttings. Second Annual Report of Director Institute of Tropical Agriculture, Mayaguez, Puerto Rico for the Fiscal Year of 1943–44. Insti-tute of Tropical Agriculture, Mayaguez, Puerto Rico, pp. 92–99.
Veeraraghavan, R. (1945) Side grafting in mangoes. Indian Journal of Horticulture 3, 45.Verma, S.R. (1942) A novel mango graft. Punjab Fruit Journal 6, 1183.Walters, E.A. (1932) Plant propagation. Tropical Agriculture, Trinidad 9, 35–39.Whiley, A.W. (1993) Environmental effects on phenology and physiology of mango. A
review. Acta Horticulturae 341, 168–176.

Crop Production: Propagation 403
Wolfe, B.S. (1963) The mango in Florida – 1887 to 1962. Proceedings of the Florida State Horticultural Society 75, 387–391.
Yang, Z. and Ludders, P. (1993) Effect of growth regulator and media on in vitro shoot tip culture of different cultivars of mango (Mangifera indica L.) rootstocks. Acta Horti-culturae 341, 240–247.
Zill, L. (1951) Top working of mango. In: Proceedings of the Florida Mango Forum.Florida Mango Forum, Florida, p. 147.

© CAB International 2009. The Mango, 2nd Edition: Botany, Production and Uses404 (ed. R.E. Litz)
12 Crop Production: Mineral Nutrition
I.S.E. BallyDepartment of Primary Industries and Fisheries, Queensland, Australia
12.1 Introduction 40512.2 Soils, Mineral Diagnosis and Sampling 40512.3 Tissue Mineral Diagnosis and Sampling 40612.4 Interpreting Soil and Leaf Analyses 40612.5 Nitrogen 407
Sources, uptake and translocation of N 409Nitrogen defi ciency and toxicity 409Effect of N on crop production and fruit quality 410Nitrogen management 411
12.6 Phosphorus 411Sources, uptake and translocation of P 411Phosphorus defi ciency and toxicity 412Effect of P on crop production and fruit quality 412
12.7 Potassium 412Sources, uptake and translocation of K 413Potassium defi ciency and toxicity 413Effects of K on crop production and fruit quality 414
12.8 Magnesium 414Sources, uptake and translocation of Mg 414Magnesium defi ciency and toxicity 414
12.9 Sulfur 415Sources, uptake and translocation of S 415Sulfur defi ciency and toxicity 415
12.10 Zinc 416Sources, uptake and translocation of Zn 416Zinc defi ciency and toxicity 416Effect of Zn on crop productivity 417
12.11 Manganese 417Role of Mn 417Sources, uptake and translocation of Mn 417Manganese defi ciency and toxicity 418

Crop Production: Mineral Nutrition 405
Effect of Mn on crop production 41812.12 Iron 418
Sources, uptake and translocation of Fe 418Iron defi ciency and toxicity 419
12.13 Calcium 419Sources, uptake and translocation of Ca 419Calcium defi ciency and toxicity 420Effects of Ca on crop production and fruit quality 420Calcium in leaves and fruit 421
12.14 Boron 422Sources, uptake and translocation of B 422Boron defi ciency and toxicity 423Effect of B on crop production and fruit quality 423
12.15 Conclusion 424
12.1 Introduction
Assessment of the mineral status of mango trees is not without its challenges. Like many other tropical woody perennial tree crop species, mangoes have complicated and variable phenological cycles that infl uence the trees’ uptake and translocation of minerals. Their extensive root system enables them to exploit unevenly distributed minerals throughout the soil profi le; these min-erals are often not assessed during routine soil analysis. The leaves, trunk, bark and roots act as mineral reserves that buffer many short-term mineral shortages (Robinson, 1986b). Soil and leaf mineral analyses used as short-term indicators of tree mineral status, tree productivity or fruit quality are therefore diffi cult and unreliable (Catchpoole and Bally, 1995).
12.2 Soils, Mineral Diagnosis and Sampling
Statistical relationships between the soil and tree mineral status have been diffi cult to establish because of the large mineral reserves in trees and the variable distribution of minerals in the soil profi le (Robinson, 1986b). How-ever, soil mineral analyses can be useful for determining the availability of the essential minerals, and if they are within the optimal range for mango. Soil analysis also provides valuable information on other soil properties that can infl uence mineral availability to the tree, i.e. pH, electrical conductivity (Ec) and concentrations of organic matter and clays. Regular soil analysis in conjunction with tissue analysis in mature cropping orchards can provide useful information on the changes in soil minerals over time and on the infl uences of fertilizer programmes on soil and tree status.
There are several publications that outline the techniques that can be uti-lized for obtaining representative soil samples for mineral analysis (Dowet al., 1991; Peverill et al., 1999; Anonymous, 2007). Before a new orchard is planted, the soil should be sampled from several depths and analysed to capture a full picture of the mineral status throughout the soil profi le. This

I.S.E. Bally406
will facilitate the adjustment of pH and deep placement of minerals before the trees are planted. In established orchards the sites of soil sampling will vary according to the age of the trees and the soil type. In sandy or duplex soil types where minerals are easily leached from the upper to the lower pro-fi le, deep (1–1.5 m) monitoring will indicate any mineral build-up and help avoid unnecessary fertilizer applications. In lighter sandy soils, surface sam-pling to depths of 10–15 cm can underestimate soil mineral concentrations as these layers are often leached and are low in clay colloids.
12.3 Tissue Mineral Diagnosis and Sampling
Leaf mineral analysis is commonly used to assess mango tree mineral status, and is useful for developing and monitoring tree fertilizer programmes. Leaves often display visual symptoms of toxic and defi cient concentrations of many minerals. Sampling mango leaves for mineral analysis should be done when the tree is at its most phenologically quiescent stage, i.e. when leaf mineral concentrations are most stable. One of the most stable periods in the mango phenological cycle is the dormant phase, which occurs after the completion of summer fl ushing and approximately 2 weeks before the emer-gence of fl ower panicles. The common practice of withholding irrigation water leading up to fl owering makes this period the most inactive of the year and an ideal time for leaf sampling. At other times of the year, leaf mineral concentrations sharply decrease when the tree is fl owering and fruiting and increase in the months following harvesting (Catchpoole and Bally, 1995; Oosthuyse, 2000b). Sampling at an inactive growth stage reduces variability between leaf samples and provides a stable reference point for annual com-parisons. Guides with respect to the most appropriate leaves for sampling have been variously reported (Kumar and Nauriyal, 1979; Chadha et al., 1980; Smith, 1992; Catchpoole and Bally, 1995), and generally concur that the most appropriate leaves to sample are the third or fourth leaf behind the apical bud, or the fi rst full-size leaf of the most recently matured dormant fl ush where leaves are fully expanded and hardened off.
Leaves recently sprayed with foliar nutrients or fungicides should be avoided or noted, as analyses of manganese (Mn), zinc (Zn), boron (B) and copper (Cu) are commonly affected by mineral residues on the outside of leaves or imbedded in the cuticle, and which are not available to the tree. Some publications recommend that sampled leaves should be washed with deionized water to reduce residues from sprays and dust from the operator’s hands (Reuter et al., 1986; Shu et al., 1992).
12.4 Interpreting Soil and Leaf Analyses
Several published soil and leaf mineral standards are available to assist in interpretating analysis results (Smith and Scudder, 1951; Young and Koo, 1969; Kumar and Nauriyal, 1977; Robinson, 1986a; Peverill et al., 1999; Stassen

Crop Production: Mineral Nutrition 407
et al., 1999) (Table 12.1). Most of these standards are in reasonable agreement with respect to the optimal ranges of individual minerals (Samra and Arora, 1997). Soil and leaf analyses should be interpreted by comparing with these standards and with previous soil and leaf analyses and past seasons’ crop-ping and fertilizer application history. The recommended optimal ranges of leaf mineral concentrations should be considered as general guides only, as high-yielding trees producing good quality fruit have been found to vary widely in leaf mineral composition (Catchpoole and Bally, 1995; Oosthuyse, 2000b; Medeiros et al., 2004).
Positive relationships between leaf minerals and tree productivity have been reported. Oosthuyse (1997) determined that leaf concentrations of nitro-gen (N), phosphorus (P), potassium (K), magnesium (Mg) and Zn infl uenced the number of fruit retained, and that leaf Zn and Mg also infl uenced fruit size. Rao and Mukherjee (1989) observed positive correlations between tree yields and leaf N and K from non-fruiting terminals in fi ve Indian mango cultivars with generally low N and K concentrations. Although these rela-tionships have been observed, there are many factors that can infl uence pro-ductivity. Therefore, predicting productivity based on leaf analysis alone is diffi cult. However, soil and leaf analyses are useful for identifying major mineral imbalances and long-term trends in tree nutrition, and can be used to adjust fertilizer programmes.
Leaf age can affect the mineral concentration in assays. Minerals that are mobile within the plant, for example N, P, K and Mg, generally decrease during leaf aging and the less mobile minerals, such as calcium (Ca), sulfur (S), B and Mn, accumulate in leaves with age (Chadha et al., 1980; Medeiros et al., 2004).
Mango leaves that have been sampled from fruiting terminals generally display increasing concentrations of N, K, Ca, Mn, iron (Fe), Cu and Zn dur-ing early fruit development and decreasing concentrations during late fruit development and maturation. These minerals, with the exception of Mg and P, also vary greatly among fruiting terminals (Oosthuyse, 1997, 2000b).
The Diagnosis and Recommendation Integrated System (DRIS) for inter-preting leaf mineral analysis, uses ratios of mineral concentrations rather than the absolute mineral concentration to identify limiting minerals in order of their effect on the tree (Beaufi ls, 1973). With mango, DRIS has been used with varying success. Raghupathi et al. (2004) observed that DRIS was unable to diagnose the nutrient imbalance of a particular nutrient in isolation; how-ever, others have used the technique more successfully. Schaffer et al. (1988) used DRIS to identify Mn and Fe as the most defi cient elements in orchards with tree decline, a disorder of unknown aetiology, and Hundal et al. (2005), Raj and Rao (2006) and Wadt et al. (2007) utilized DRIS to identify yield-limiting mineral combinations in mangoes grown in India and Brazil.
12.5 Nitrogen
Nitrogen is one of the most important elements for crop production, and has a signifi cant role in mango growth, yield and fruit quality. Nitrogen is an
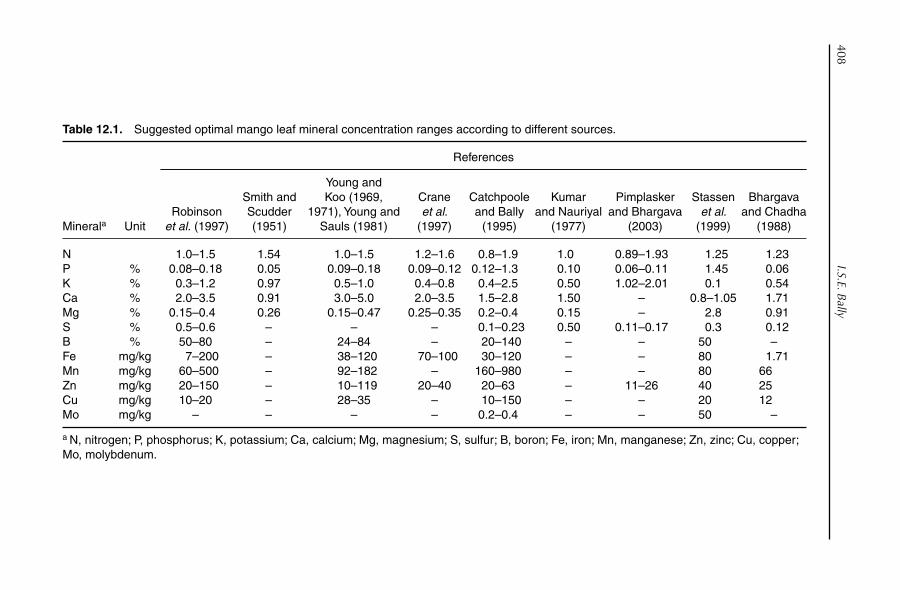
I.S.E. B
ally408
Table 12.1. Suggested optimal mango leaf mineral concentration ranges according to different sources.
Minerala
References
UnitRobinson
et al. (1997)
Smith and Scudder(1951)
Young and Koo (1969,
1971), Young and Sauls (1981)
Crane et al.
(1997)
Catchpooleand Bally
(1995)
Kumar and Nauriyal
(1977)
Pimplasker and Bhargava
(2003)
Stassenet al.
(1999)
Bhargava and Chadha
(1988)
N 1.0–1.5 1.54 1.0–1.5 1.2–1.6 0.8–1.9 1.0 0.89–1.93 1.25 1.23P % 0.08–0.18 0.05 0.09–0.18 0.09–0.12 0.12–1.3 0.10 0.06–0.11 1.45 0.06K % 0.3–1.2 0.97 0.5–1.0 0.4–0.8 0.4–2.5 0.50 1.02–2.01 0.1 0.54Ca % 2.0–3.5 0.91 3.0–5.0 2.0–3.5 1.5–2.8 1.50 – 0.8–1.05 1.71Mg % 0.15–0.4 0.26 0.15–0.47 0.25–0.35 0.2–0.4 0.15 – 2.8 0.91S % 0.5–0.6 – – – 0.1–0.23 0.50 0.11–0.17 0.3 0.12B % 50–80 – 24–84 – 20–140 – – 50 –Fe mg/kg 7–200 – 38–120 70–100 30–120 – – 80 1.71Mn mg/kg 60–500 – 92–182 – 160–980 – – 80 66Zn mg/kg 20–150 – 10–119 20–40 20–63 – 11–26 40 25Cu mg/kg 10–20 – 28–35 – 10–150 – – 20 12Mo mg/kg – – – – 0.2–0.4 – – 50 –
a N, nitrogen; P, phosphorus; K, potassium; Ca, calcium; Mg, magnesium; S, sulfur; B, boron; Fe, iron; Mn, manganese; Zn, zinc; Cu, copper; Mo, molybdenum.

Crop Production: Mineral Nutrition 409
essential component of many plant tissues, and occurs in chlorophyll, amino acids, proteins, enzymes and growth hormones and is a major driver of plant growth, having a direct effect on tree vigour (Marschner, 1995). Nitrogen at 100 g/tree/year has been shown to be suffi cient for mango tree growth (Kanwar et al., 1987), and N concentrations infl uence concentrations of other elements when N is either low or in excess (Sen et al., 1947).
Sources, uptake and translocation of N
Nitrogen in the soil occurs in many forms, but for the most part as large and complex organic molecules that comprise the organic matter. These mole-cules are too large for roots to absorb, and are broken down to nitrate (NO3
–) and ammonium (NH4
+). The concentration of N in soil is dependent on the concentrations of soil organic matter and mineral N. Nitrogen that is avail-able to the plant is determined by the processes of mineralization, immobili-zation, de-nitrifi cation, volatilization and leaching, which are infl uenced by temperature, moisture, pH and aeration. Nitrogen is easily leached from the soil by rain and irrigation, and consequently, soil N often limits tree growth, especially in sandy soils (Strong and Mason, 1999).
Common N fertilizers include: urea (CO(NH2)2, 46% N), potassium nitrate (KNO3, 13% N, 38% K), calcium nitrate (CaNO3, 15.5% N, 19% Ca), ammonium nitrate (NH4NO3, 35% N), sulfate of ammonia ((NH4)2SO4, 21% N, 23.6% S). Nitrogen is taken up by mango trees primarily through the roots as NO3
– and NH4+; NO3
– is the preferred source. Nitrogen can also be adsorbed through leaves as ammonia (NH3), urea and amino acids. Nitrate, after being adsorbed by the roots, is either reduced to NH4
+ in the roots or translocated to the leaves, stems or other tissues, where it is reduced to NH4
+ in a two-step process in which NO3
– is reduced to nitrite (NO2–) which is in turn reduced to
NH4+ (Marschner, 1995). Ammonium metabolism is complex, and several
pathways are necessary to produce amino acids, proteins, enzymes and hor-mones. Within the tree, N concentrations vary among tissues and are depen-dent on N availability in the soil and demand within the tree. If N is limiting, the plant can translocate N from older tissues to new growing tissues where it is required (Marschner, 1995).
Nitrogen defi ciency and toxicity
Nitrogen defi ciency symptoms appear in leaves as yellowing or chlorosis initially of the older leaves, slow growth and lack of vigour. Severe N defi -ciency can cause complete leaf yellowing, leaf and fruit abscission and death of twigs and branches (Sen et al., 1947; Smith and Scudder, 1951). Nitrogen toxicity is not often seen in mango; however, trees with high N concentra-tions have dark green leaves and excessive vegetative vigour, often at the expense of fl owering and cropping (Tiwari and Rajput, 1976). High N con-centrations are also associated with internal fruit disorders, e.g. jelly seed

I.S.E. Bally410
and internal breakdown in many cultivars (Tarmizi et al., 1993; Cracknell Torres et al., 2004; see Galán Saúco, Chapter 9, this volume).
Effect of N on crop production and fruit quality
Nitrogen has a major effect on mango tree vigour, stimulating both vegeta-tive and fl oral growth. Increases in shoot length, leaves per shoot and leaf area have been demonstrated by Singh et al. (1973), Tiwari and Rajput (1975), Syamal and Mishra (1989), Reddy et al. (2000) and Sergent et al. (2000).
Yeshitela et al. (2005) reported that N in combination with K, as KNO3 and urea, improved the percentage of terminal shoots that fl ower; however, excessive N can stimulate vegetative growth at the expense of fl owering, fruit set and fruit quality (Scholefi eld et al., 1986; Monselise and Goren, 1987; Silva et al., 2002). Nitrogen can cause increased fruit set and retention (Singhet al., 1973; Oosthuyse, 1997) and fruit weight and yield (Alvian, 1974; Tiwari and Rajput, 1975; Young and Koo, 1975; Alvian and Figueroa, 1977; Syamal and Mishra, 1989; Reddy et al., 2003). Kanwar et al. (1987) demonstrated that N at 100 g/tree/year-of-age was suffi cient for tree growth and Young and Koo (1975) reported that maximum yields were increased by N applications of 0.8–1.1 kg/tree/year. Many reports of the effect of N on increased mango fruit size and yields are based on data of N in combination with other ele-ments (i.e. P and K) and may be partly attributed to these combinations; however, N has a major infl uence on productivity in mango.
High N application rates that stimulate yield increases can also have negative effects on fruit quality. Nitrogen has been negatively associated with fruit colour in mango (Oosthuyse, 1993; McKenzie, 1994, 1995). Nguyen et al. (2004) demonstrated that high N applications during fruit growth inhibited the de-greening of ripening fruit, causing green skin at ripeness. Bally (2007) also reported a negative relationship between N and fruit colour, demonstrating that high leaf N concentrations reduce the percentage of yellow skin in ripe fruit, reduced the lightness and chroma (vividness) of the yellow colour, the percentage of skin covered with blush and the intensity of the blush colour (Plate 73). He also identifi ed a signifi cant exponential rela-tionship between the severity of sunburn and skin N concentrations; the effects of sunburn were reduced as N concentrations increased in the fruit skin.
Early indications of the negative effect of N on postharvest rots in mango were demonstrated by Weng and Chuang (1997), who observed that N posi-tively affected the germination rate, hyphal growth and appressoria forma-tion of Colletotrichum gloeosporioides. When Nguyen et al. (2004) investigated the effect of N on fruit quality, they found that 300 g/tree applied as a foliar spray signifi cantly increased the severity of anthracnose caused by C.gloeosporioides. These observations were confi rmed by Bally (2007), who found that the severity of fruit anthracnose during ripening increased with applied N (Plate 74). He also demonstrated that the N effect was because N enhanced the decline of antifungal resorcinol compounds in the fruit exocarp during

Crop Production: Mineral Nutrition 411
ripening. Nitrogen has been a recognized factor in determining internal fruit quality of mangoes, with imbalances of N and other minerals, principally Ca, being implicated as a major factor causing the various forms of internal breakdown (Young and Miner, 1961; Subramanyam et al., 1971; Malo and Campbell, 1978; Burdon et al., 1992; Tarmizi et al., 1993; Cracknell Torres et al., 2004; Torres et al., 2004; see Galán Saúco, Chapter 9, this volume).
Nitrogen management
Leaf N concentrations between 1 to 1.5% dry weight (DW) are generally con-sidered to be in the optimal range (Robinson et al., 1997). Optimum concen-trations for fruit skin N have not been published, although Catchpoole and Bally (1995) reported skin N concentrations of 0.069% DW in mature ‘Kens-ington Pride’ fruit. Leaf N concentrations vary throughout the year, and are infl uenced by the growth events during the phenological cycle. Nitrogen concentrations are generally greatest at the end of the summer vegetative fl ushing period and decrease during panicle growth, fl owering and fruiting (Catchpoole and Bally, 1995; Medeiros et al., 2004).
Because N is highly mobile within the tree and a primary driver of growth and fruit quality, the general practice of monitoring leaf and soil N annually is inadequate for assessing tree N status at all stages of tree phenol-ogy. A cheap, rapid test for N is needed to provide closer monitoring of changes in leaf N status and allow the rates and timing of supplementary N applications to be matched to tree and fruit demands. Bally and Still (per-sonal communication) have calibrated the Konica-Minolta Soil Plant Analy-sis Diagnostic (SPAD-502) meter to measure the chlorophyll content of leaves, which is directly related to their N status (Gonzalez et al., 2005).
12.6 Phosphorus
Phosphorus is important for cell division and growth and is a component of many essential plant molecules such as sugar-phosphates that are involved in respiration, photosynthesis and other metabolic pathways, nucleotides such as DNA and RNA, phospholipids in membranes and as pryophosphate (PPi) in ATP and other cellular energy metabolism molecules (Salisbury and Ross, 1992). Phosphorus is also involved in root tip elongation, fruit ripening and leaf expansion.
Sources, uptake and translocation of P
Phosphorus that is available for plants occurs in two forms in soil, primarily as the monovalent phosphate anion (H2OP4
–) and secondly as the divalent anion (HPO4
2–) in the soil water solution. The balance of these anions is dependent on soil pH, with HPO4
2– favoured in soils >pH 7 and H2OP4–

I.S.E. Bally412
favoured in soils <pH 7 and most readily available in soils with pH between 6 and 7. In higher pH soils, calcium phosphate compounds tie up P from plant availability, and in lower pH soils, P oxidizes with Fe, aluminium (Al) and Mn to become unavailable to plants. Other forms of soil P include liable P that is bound on clays and organic matter, which can become available to trees when dissolved in water.
Phosphorus is most rapidly taken up by the tree as HPO42– and slower as
H2OP4– and often enhanced by chemical association with N. After entry into
the roots, the anions are either converted to organic phosphates immediately or after transport to other tissues. Within the tree, P is easily translocated between tissues, with redistribution usually occurring from older leaves to younger actively growing tissues. Phosphorus concentrations within the mango tree are highest in the roots, wood and bark and lowest in the leaves (Vuuren and Stassen, 1997). Leaf P concentrations are generally at their low-est during fruit development and at their highest in the vegetative growth period (Medeiros et al., 2004).
Phosphorus defi ciency and toxicity
As P is a mobile element, defi ciencies are initially seen in the older leaves. Phosphorous defi ciency and toxicity are rare in mango trees, with toxicity symptoms occasionally seen in nursery stock where trees are grown in pot-ting media based on sugarcane mill/press mud. Symptoms appear as stunted seedling growth with bronzing of the leaves. Phosphorus defi ciency appears fi rst in older leaves as marginal necrosis with brown taints, tip necrosis, pre-mature abscission and stem dieback (Smith and Scudder, 1951; Singh and Saxena, 1994). Sen et al. (1947) describe defi ciency symptoms in leaves as developing a reddish-purple colour on the undersides of leaves that spreads over the entire leaf, eventually encompassing the veins, with leaves becom-ing thick and stiff. Phosphorus-defi cient trees have restricted root development that reduces tree and fruit size (Stassen et al., 1999).
Effect of P on crop production and fruit quality
There are several reports of P applied in combination with N and/or K and resulting in increased yields (Samra and Arora, 1997); however, these reports do not separate the P from the other elements. Reddy and Majmudar (1985) reported an association between higher concentrations of P in terminals and fl oral induction (Narwadkar and Pandey, 1989).
12.7 Potassium
Potassium in the cytoplasm is an activator of enzymes involved in photosyn-thesis, respiration and starch and protein synthesis (Bhandal and Malik, 1988;

Crop Production: Mineral Nutrition 413
Marschner, 1995). Potassium is important for cell growth due to its role in cell expansion and development of thick epidermal cell walls that increase the resistance of trees to pathogens and insect pests. Potassium is involved in tree water status by regulating water uptake by the roots and water loss through the leaf stomata (Salisbury and Ross, 1992).
Sources, uptake and translocation of K
Most of the K in soils (90–98%) is in the form of insoluble crystalline minerals that are unavailable to plants. Available K occurs in the soil solution as K+
ions and attached to cation exchange sites on clays (Gourley, 1999). Potas-sium moves readily between the exchange sites and the soil solution and is infl uenced by moisture and temperature. Heavy applications of Ca and Mg fertilizers compete with K for exchange sites, thereby reducing the availability and uptake of K (Lim and Khoo, 1985). Potassium concentrations are lower in soils with low cation exchange capacities, e.g. granites, sands, highly leached and acidic soils, and are higher in clay soils (Lim and Khoo, 1985; Gourley, 1999).
The main K fertilizers used in mango production are potassium chloride (KCl) also known as muriate of potash, potassium sulfate (K2SO4) and potas-sium nitrate (KNO3). Potassium sulfate is generally preferred to KCl because it has a neutral effect on pH and because mangoes are very sensitive to chlo-rides. Mango trees take up K as the K+ ion from the soil solution. Potassium is readily redistributed, typically from old leaves to young growing tissues. The highest concentrations of K occur in leaves, fruit, roots and bark (Vuuren and Stassen, 1997). Leaf K concentrations are generally at their highest at the end of the postharvest summer vegetative fl ushing period, decreasing with fl owering and fruiting (Catchpoole and Bally, 1995; Medeiros et al., 2004). Mango trees can absorb more K than is required for maximum tree growth; excessive uptake of K can cause imbalances with other minerals in the tree (Gourley, 1999).
Potassium defi ciency and toxicity
Potassium defi ciency fi rst appears in older mango leaves as the tree redistrib-utes K to young growing tissues. Sen et al. (1947) induced K defi ciency by growing mangoes in sand culture and observed brown necrosis on the leaf margins extending from the leaf tip towards the base. Smith and Scudder (1951) also generated K defi ciency symptoms in sand culture and described them as irregularly distributed yellow spots and necrotic areas along the margins of small, thin attenuated leaves which are very persistent. Lim and Khoo (1985) described the non-necrotic areas of the leaf as dull, yellowish green to light green; symptoms usually develop in the dry season or when irrigation has been stopped.

I.S.E. Bally414
Effects of K on crop production and fruit quality
Potassium nitrate applications just prior to and at the fl owering stage promote fl owering, increase fruit set and fruit retention (Sergent and Leal, 1989; Lyan-naz, 1994; Ferrari and Sergent, 1996; Oosthuyse, 1997; Rojas and Leal, 1997; Sergent et al., 1997; Saleh and El-Monem, 2003; Shinde et al., 2006). In the low and mid-latitude tropics, KNO3 is used to stimulate out-of-season fl owering; however, this effect is lost in higher latitudes (Davenport and Núñez- Elisea, 1997; see Davenport, Chapter 5, this volume). Shongwe and Roberts-Nkrumah (1997) suggested the KNO3 effect on fl owering is a result of lowering the tran-spiration rate and increasing water use effi ciency. Protacio (2000) suggests the KNO3 effect on fl owering is primarily due to N stimulation rather than K and he postulated that KNO3 overcomes the inhibitory effects of gibberellic acid (GA3) on starch accumulation by elevating the N concentrations over the N threshold to synchronize bud break from apices with an existing fl oral initial.
Potassium infl uences fruit quality of many species (Marschner, 1995); however, in mango there are only a few studies that link K nutrition with increased fruit quality. Shinde et al. (2006) observed that increased K fertiliza-tion increased fruit weight (5.15%), ascorbic acid (26.99%), organoleptic score for texture, fl avour, colour and shelf life and reduced physiological weight loss (22.79%) and spongy tissue (68.08%). Potassium in the form of monopotassium phosphate (KH2PO4) suppresses powdery mildew on mango (Reuveni et al., 1998; Oosthuyse, 2000a), but it is not clear if the effect is due to P or K.
12.8 Magnesium
Magnesium is a component of enzymes that transport P, develop green col-oration in chlorophyll, activate other enzymes and is involved in carbohydrate metabolism and nucleic synthesis (Marschner, 1995; Stassen et al., 1999).
Sources, uptake and translocation of Mg
Magnesium is present in soil as the minerals biotite, serpentine, olivine and hornblende, in organic matter, and as exchangeable Mg2+ in the soil solution. Mg2+ is the second most dominant cation after Ca2+ and successfully com-petes with K+ and Na+, replacing them on exchange sites (Aitken and Scott, 1999). Magnesium is a mobile element that is taken up by the plant as Mg2+. Magnesium concentrations in the tree can be depressed by high concentra-tions of other cations, such as Mn2+, Ca2+, K+, NH4
+ and Al3+, in the root environment (Aitken and Scott, 1999).
Magnesium defi ciency and toxicity
Magnesium defi ciency fi rst appears in older leaves, due to its high mobility within the tree, as pale green or yellow leaves with the inner vein areas of the

Crop Production: Mineral Nutrition 415
leaf lamina becoming mottled and necrotic while the leaf veins remain green. Young trees are stunted by Mg defi ciency. Smith and Scudder (1951) gener-ated symptoms of Mg defi ciency in sand culture and described them as a green wedge pattern formed by the lateral intrusion of a bronzed chlorosis along the leaf margin (Plate 75). Defi ciency symptoms are more likely to occur in textured, sandy or highly leached soils due to their low cation exchange capacity or when heavy applications of liming products, K fertil-izers or green manuring have been applied (Stassen et al., 1999). Magne-sium defi ciency has been associated with a browning skin discoloration in ‘Kensington Pride’ fruit.
12.9 Sulfur
Sulfur is a macro element required in large amounts by the tree. Sulfur is an essential component of the amino acids cystine and methionine that make up photosynthetic proteins, the vitamins thiamine and biotin and coenzyme-A used in the synthesis and breakdown of fatty acids (Salisbury and Ross, 1992).
Sources, uptake and translocation of S
Up to 70–90% of S in soil is present in organic matter (Marschner, 1995), and sulfates that are available for plants are mainly found in the surface layers and the soil solution with only small amounts adsorbed to soil colloids. As a result, S is very prone to leaching. High rainfall, light-textured and dry soils are adverse to S accumulation and uptake (Anonymous, 1988).
Supplementary sources of S include sulfates as minor components of fer-tilizers, i.e. superphosphate, sulfate of ammonia, potassium sulfate and gyp-sum as well as some fungicides and miticides. Organic-based sulfate compounds are not available to plants until they have been converted to the sulfate ion (SO4
2–) by bacteria in the mineralization process. The divalent sul-fate anions (SO4
2–) are actively taken up by the roots and translocated to shoots or growing tissues. Sulfur can also be adsorbed through the leaves as sulfur dioxide (SO2), usually from pollution from burning fossil fuels. Sulfur is not readily re-translocated from one tissue to another. The competitive effects of other anions such as phosphates and nitrates can reduce the uptake of S (Marschner, 1995).
Sulfur defi ciency and toxicity
Sulfur defi ciency in mango is uncommon as it is generally present in ade-quate quantities in most soils and is a component of many fertilizers, fungi-cides and miticides. Smith and Scudder (1951) describe S defi ciency in mango as lateral necrotic spots that occur on the vascular bundles and lamina on a very deep-green leaf and premature defoliation. In other species symptoms

I.S.E. Bally416
often appear to be similar to N defi ciency, but are distributed more evenly between young and old leaves, with the younger tissues developing symp-toms fi rst as S is not easily redistributed from older to younger tissues (Salisbury and Ross, 1992; Marschner, 1995).
Sulfur toxicity occurs when pollutants high in SO2 content are converted to bisulfate (HSO3–) when combined with water, and is further oxidized to sulfuric acid (H2SO4), often known as acid rain. Sulfur dioxide adsorbed by leaves reacts with water to form HSO3– that is toxic to the leaf, inhibiting photosynthesis and degrading chlorophyll (Marschner, 1995).
Klumpp et al. (2003) reported soil S concentrations of between 53.2 and 86.0 mg/kg DW in contaminated soils in the vicinity of a copper smelter in Brazil compared with 33–48 mg/kg DW in uncontaminated soils. Leaves from mango trees growing in the contaminated soils had S concentrations of 3.8 mg/g DW, double that of trees growing in uncontaminated soils (1.9 mg/g DW). At these concentrations no obvious leaf symptoms were visible, indicating that mango has a high tolerance of S in the soil and atmosphere. Information on mango leaf S concentrations is scarce; however, a wide range of concentrations are consid-ered to be optimal for mango growth (Table 12.1): 0.6–6.4 mg/g DW S (Marchalet al., 1991), 1.07–1.69 mg/g DW S (Pimplasker and Bhargava, 2003). Chaudhary and Nauriyal (1988) induced leaf defi ciency symptoms in 1-year-old seedlings in sand culture when leaf S concentrations were between 0.32% and 0.74% DW.
12.10 Zinc
Zinc is chemically bound to Fe and Mn to form components of chlorophyll, and is essential for photosynthesis (Weir and Cresswell, 1995). Zinc is essential for the synthesis of proteins and hormones, including auxins, and is required for the maintenance of biomembranes (Salisbury and Ross, 1992; Marschner, 1995).
Sources, uptake and translocation of Zn
Zinc is present in soil in a range of inorganic minerals and organic matter with the solubility of the inorganic forms decreasing as soil pH increases. Common Zn fertilizers include zinc oxide (ZnO, 80% Zn), zinc chelates (ZnNa2 EDTA and Zn(NH4)2 EDTA, 6.5–14% Zn), zinc sulfates (ZnSO4, 23% Zn). Zinc is taken up by the tree in the form of the zinc ion (Zn2+), and trans-located to regions of growth such as shoot tips. Zinc is not readily translo-cated to other tissues. Zinc is readily taken up through the leaves, and foliar Zn application is a common method of managing Zn in mango orchards.
Zinc defi ciency and toxicity
Zinc defi ciency is often referred to as ‘little leaf’ because leaves fail to reach full size. Symptoms fi rst appear in immature leaves in the coloured stage, as

Crop Production: Mineral Nutrition 417
a thickening of the leaf and failure to fully expand. Leaf expansion is often stunted on one side of the blade, causing it to form a sickle shape (Dilly et al., 1997) (Plate 76). Symptoms in fully mature leaves are prominent light-yellow or olive-green veins on the upper surface that are thick and brittle. Internode length is reduced, thereby causing a rosetting effect (Lim and Khoo, 1985; Agarwala et al., 1988; Marschner, 1995). Zinc and Fe defi ciency often occur together because of their association with calcareous high pH soils.
Effect of Zn on crop productivity
Singh and Rajput (1977) reported that foliar sprays of 2.0–8.0% ZnSO4 prior to fl owering increased the number of perfect fl owers in panicles and later increased fruit yield, fruit sugar, ascorbic acid and total soluble solids (TSS). Daulta et al. (1981) also observed that foliar Zn sprays increased fruit set and improved fruit quality, but had no effect on sex ratio. Kumar and Kumar (1989) demonstrated that 1% ZnSO4 sprays applied to ‘Dashehari’ trees improved postharvest fruit life by reducing weight loss, spoilage, increasing sugars, reducing acidity and slightly increasing vitamin A. Littlemore et al. (1991) observed that 1% ZnSO4 foliar sprays applied quarterly to ‘Kensing-ton Pride’ was suffi cient to maintain leaf Zn concentrations above critical concentrations and avoid symptoms of little leaf. Although leaf Zn concen-trations were improved and symptoms cured, no effect on tree yields was observed. Soil applications are often ineffective due to adverse soil conditions making it unavailable to trees.
12.11 Manganese
Role of Mn
Manganese is a trace element that is required in small amounts by the tree; it is important in the redox processes and is a cofactor, activating many enzymes that catalyse oxidation, reduction, decarboxylation and hydrolytic reactions. Manganese is an essential mineral for photosynthesis and the biosynthesis of proteins, carbohydrates and lipids (Marschner, 1995).
Sources, uptake and translocation of Mn
Manganese is found in soil in the insoluble form of manganese oxide (MnO2) which is reduced to the water soluble exchangeable ion (Mn2+) that is taken up by tree roots. Mn2+ is mainly found in the soil solution but can also be loosely bound to organic matter and clay colloids. Reduction of MnO2 to Mn2+ is carried out by organic reducing agents in aerobic soils and is favoured by reduced pH. As the soil pH increases, microbial oxidation of Mn2+ increases. Both reduction and oxidation processes occur in the soil simultaneously, but

I.S.E. Bally418
the balance of available Mn is governed by soil pH, with low pH favouring reduction and high pH favouring oxidation (Peverill et al., 1999).
Manganese defi ciency and toxicity
Manganese defi ciency in mango is initially expressed by light necrosis between the main veins of middle and younger leaves that later become necrotic. Buff-coloured spots appear on the primary leaf veins at the margins of the leaves and coalesce to form necrotic patches before leaf abscission. In severe Mn defi ciency, necrosis begins at the leaf tip, spreading to the base of the lamina (Smith and Scudder, 1951; Agarwala et al., 1988).
Effect of Mn on crop production
There are few reports on the effect of Mn on crop production or fruit quality. Schaffer (1994) suggested that mango decline was associated with trees that are defi cient in Mn and Fe.
12.12 Iron
Iron is a trace element that is required in small quantities, and is a component of several enzymes and molecules. It is a component of chlorophyll and leaf Fe concentrations directly impact photosynthesis. Iron-containing enzymes participate in oxidation processes that release energy from sugars and starches, the conversion of nitrates to ammonia and the biosynthesis of eth-ylene (Marschner, 1995). In proteins, Fe acts as an electron carrier by alternate oxidation and reduction between Fe2+ to Fe3+ (Salisbury and Ross, 1992).
Sources, uptake and translocation of Fe
Available Fe occurs in the soil solution in very low concentrations (~10–15 M) as the ionic forms Fe2+ and Fe3+. Fe2+ is far more soluble and more easily taken up than Fe3+; however, in well-aerated soil, Fe2+ is readily oxidized to Fe3+ and precipitated, becoming unavailable. Fe2+ can also be displaced by other cations or by alkaline soils (Salisbury and Ross, 1992). Plants partly overcome Fe unavailability by producing ligands that bind to Fe3+ to form soluble chelates that maintain Fe in solution. On the root surface the Fe3+ is reduced to Fe2+ which is immediately adsorbed by the roots. This process is often inhibited in high pH soils and in calcareous soils. Iron defi ciency is com-mon in mango (Schmidt, 1999). Iron is readily absorbed through leaves, which is a useful method for management of Fe when soil conditions are not conducive to uptake.

Crop Production: Mineral Nutrition 419
Iron defi ciency and toxicity
Iron defi ciency initially appears in young leaves as reduced concentrations of chlorophyll. Iron-defi cient leaves are pasty yellow/green with developing chlorosis of the whole leaf as severity increases (Plate 77). Leaves fail to develop to full size and necrosis of the leaf tips occurs in severe cases. Iron defi ciency is often seen on calcareous soils with >20% calcium carbonate (CaCO3), with poor aeration and drainage in the coolest part of the year (Marschner, 1995). Iron defi ciency reduces the ability of mango trees to photosynthesize, thereby stunting tree growth and reducing yields.
12.13 Calcium
A major role of Ca is membrane stability, which is achieved when Ca2+ binds to phosphate and carboxylate groups of phospholipids and proteins on the surface of the plasmalemma, and thereby preventing leaking of solutes to the cytoplasm (Kirkby and Pilbeam, 1984). Calcium protects cells from toxins, slowing the aging of plant tissues and promoting longer shelf life of many fruits. Calcium is important for pectin polymers that strengthen cell walls and provide defence from pathogens (Ferguson, 1984). Most Ca in trees is fi xed in cell walls and is not easily translocated. Calcium is essential for new root hair and leaf development.
Sources, uptake and translocation of Ca
Soil Ca occurs as exchangeable, non-exchangeable and soluble forms. Exchangeable Ca makes up 65–85% of the total exchange capacity of normal soils, and is weakly bonded, allowing rapid exchange with the soil solution, and is thus prone to leaching (Kirkby, 1979; Mengel and Kirkby, 1987). Non-exchangeable Ca is bound to minerals, for example feldspars, amphiboles, phosphates and carbonates. Soil Ca concentrations are often up to ten times higher than other cations such as K+ and Mg2+ and usually similar or slightly higher than sodium ions (Na+) (Mengel and Kirkby, 1987).
Calcium is taken up passively through limited unsuberized tips of actively growing roots and moves from the root cortex to the stele mainly through the apoplast or free space of the root tips. It then enters the xylem, moving upwards with the mass transpiration fl ow (Bangerth, 1979; Mengel and Kirkby, 1987; Ho and Adams, 1989; Marschner, 1995). Root Ca concentra-tions are lower than other cations such as K+ and Mg2+ because it is passively taken up and easily replaced from the exterior surface of the plasma mem-brane by other cations (Marschner, 1995). Root Ca concentrations can there-fore be infl uenced by the relative concentrations of other cations (K+, Mg2+, NH4
+) in the soil solution. The suppression of Ca uptake by high soil concen-trations of NH4
+ may account for the link between high N nutrition and many Ca-related disorders. On the other hand, high NO3
– can stimulate the uptake

I.S.E. Bally420
of cations by stimulating organic anion synthesis, which attracts Ca2+ (Kirkby, 1979; Mengel and Kirkby, 1987).
Calcium has low mobility in the phloem (Bangerth, 1979; Kirkby and Pilbeam, 1984), limiting translocation within the tree. Ca required by devel-oping organs is supplied via the xylem. Calcium defi ciencies are often caused by low availability, poor uptake and/or poor translocation within the plant. Root pressure can also contribute to the mass fl ow, but the extent to which root pressure contributes to Ca fl ow in mango is unclear.
The intensity of evaporation from any organ will directly infl uence the amount of Ca the organ receives (Marschner, 1995). Low transpiring plant organs, such as fruit, are more prone to Ca defi ciencies than actively transpir-ing leaves. This is especially the case if the organ is fast growing and requires high amounts of Ca (Mengel and Kirkby, 1987; Marschner, 1995). Limited Ca translocation occurs in the absence of mass fl ow through exchange adsorp-tion in the xylem vessels, similar to an exchange column (Mengel and Kirkby, 1987). As tissues grow, the cation exchange sites of the new cell walls act as a sink for the Ca in the xylem exchange column, drawing the Ca up the column (Clarkson, 1984). This exchange movement of Ca may be important for lower transpiring organs such as the shoot apex and fruit (Mengel and Kirkby, 1987; Marcelle et al., 1990). The translocation of Ca is induced by the polar move-ment of indole-3-acetic acid (IAA) that is produced in the shoot apex and rapidly growing organs (Banuelos et al., 1987; Ho and Adams, 1989).
Calcium defi ciency and toxicity
Calcium defi ciency symptoms refl ect the low mobility of Ca within the tree, fi rst appearing in young, actively growing tissues. Ca-defi cient plants generally display symptoms of membrane degeneration, which have been likened to senescence and fruit ripening (Fallahi et al., 1977; Bangerth, 1979). Many symp-toms of internal disorders in mango appear to be associated with premature ripening or cell degeneration (Burdon et al., 1991, 1992). Raymond et al. (1998a) observed that cell disruption and rupture of the cell walls were the fi rst micro-scopic indicators of soft nose, stem-end cavity and jelly seed. High Ca concen-trations reduce the binding sites for enzymatic degradation (Rhodes, 1980). No direct symptoms of Ca toxicity have been recorded in mango, but high concen-trations of Ca in soils can displace other minerals such as Mg, Zn, B, Cu and P.
Effects of Ca on crop production and fruit quality
In mango, fruit Ca has been implicated as a factor in fruit quality, for example with regards to internal physiological disorders, shelf life and disease resis-tance. Many symptoms of internal physiological disorders appear to be asso-ciated with low Ca-related membrane degeneration, i.e. premature ripening (Burdon et al., 1992; Raymond et al., 1998a). Links between internal break-down and Ca nutrition in mango were reported by Young (1957), who found

Crop Production: Mineral Nutrition 421
that increasing Ca fertilization generally resulted in decreased incidence of soft nose. Young and Miner (1961) and Shear (1975) suggested a link existed between Ca and N with respect to physiological disorders by demonstrating the negative effect of N on internal breakdown could be partly overcome by high Ca concentrations in the fruit. When Malo and Campbell (1978) were unable to alter internal breakdown in ‘Tommy Atkins’ by N or K fertilization over a 4-year period, the high Ca in the calcareous soils of Florida was suggested as a possible reason.
Recent investigations have monitored Ca concentrations in fruit as well as in soil. Subramanyam et al. (1971) observed that fruit with spongy-tissue disorder had Ca concentrations of 74 mg/100 g DW, compared with 85 mg/100 g DW for healthy fruit. Rane et al. (1976), Kadiyala (1995) and Tarmizi et al. (1993) also reported signifi cantly lower Ca concentrations in fruit with physiological disorders. Burdon et al. (1991, 1992) measured lower Ca concentrations (5.85 mg/100 g fresh weight (FW)) in the distal portions of ‘Kent’ fruit with soft-nose symptoms than in healthy fruit (8.02 mg/100 g FW), and concluded that physiological disorders are associated with Ca defi -ciency, but that Ca may be only one of many causal factors and not necessar-ily the primary cause. There have been other reports that increased Ca concentration is associated with physiological disorders; however, these have been attributed to mineral redistribution after tissue breakdown (Burdon et al., 1991; Raymond et al., 1998b).
Comparing fruit Ca data from different publications is diffi cult, as some quote Ca concentrations on a FW basis and others on a DW basis, and many authors do not specify whether they measured whole fruit or fruit pulp. None the less, evidence allows the conclusion that low Ca concentrations are a causal factor of physiological disorders in mango fruit.
There are mixed reports of the effect of fruit Ca on postharvest fruit breakdown in mango. Singh et al. (1987) reported that spray-to-runoff appli-cations of calcium chloride (CaCl2, 36% Ca) and calcium nitrate (Ca(NO3)2, 18% Ca) at rates between 5 and 20 ml/l a week before harvest, increased shelf life of ‘Alphonso’ mangoes by 10 days. However, the authors did not discuss whether the extended shelf life was due to delayed ripening or to reduced postharvest disease. When Duttaray et al. (1993) attempted to repeat this experiment to control stem-end rot caused by Diplodiia natalensis Pole Evans using 2500–5000 μl/l Ca as CaCl2 and 450–900 μl/l Ca as Ca(NO3)2, they sig-nifi cantly increased the incidence of stem-end rot. Simmons et al. (1997) used preharvest Ca sprays and manipulation of leaf:fruit ratios to increase fruit Ca, but were unable to signifi cantly reduce postharvest disease.
Calcium in leaves and fruit
Leaf concentrations of Ca increase with age, and are considered to be ade-quate if they are between 2–3.5% DW in acid soils and 3.0–5.0% in alkaline soils (Reuter and Robinson, 1997). Leaf Ca concentrations generally do not correlate well with fruit Ca concentrations (Bally, 2007).

I.S.E. Bally422
Calcium concentrations in mango fruit can vary between and within fruit. Gunjate et al. (1979) measured a gradient in Ca concentration from the stem end (122 mg/100 g DW) to the apex (110 mg/100 g DW) of ‘Alphonso’ fruit, and from the skin (142 mg/100 g DW) to the centre (130.4 mg/100 g DW) and to the seed (128.4 mg/100 g DW). Gradients of Ca concentrations from the inner-mesocarp to the outer-mesocarp, and from the stem end to the fruit apex have been reported (Burdon et al., 1991; Tarmizi et al., 1993; Shorter and Joyce, 1998). Variation in Ca concentrations within fruit tissues may be related to the position of the vascular tissue and the transpirational properties of the tissue that infl uence the diffusion path of Ca (Shorter and Joyce, 1998).
Concentrations of Ca on a DW basis increase rapidly during early fruit development (cell division stage) followed by a gradual reduction later (cell expansion stage); however, concentrations vary between reports. Gunjate et al. (1979) measured Ca concentrations in ‘Alphonso’ of 241 mg/100 g DW at fruit set decreasing to 68 mg/100 g DW at harvest. Chattopadhyay and Sarkar (1990) also observed a similar decline, but measured higher concen-trations of 600–700 mg/100 g DW Ca at fruit set and 350–450 mg/100 g DW Ca at harvest in four cultivars. More recently, Bally (2007) measured fruit mesocarp Ca between 0.22% and 0.33% 30 days after fruit set and between 0.08% and 0.14% at harvest (155 days after fruit set) in ‘Keitt’. Both Gunjate et al. (1979) and Bally (2007) also noted large fl uctuations in Ca concentra-tions in the fi rst half of fruit development that refl ected imbalances in supply and demand of developing fruit.
12.14 Boron
Boron binds as cis-diol borate complexes to mannose and certain other sug-ars in cell wall polysaccharides and also has a role in sugar movement in the tree. Boron also is important in nucleic acid synthesis (Salisbury and Ross, 1992), and is essential for pollen and fl ower development, pollen germina-tion and pollen tube growth (Stanley and Lichtenberg, 1963; Gupta et al., 1985) and is thus essential for mango fruit set. Boron also is important in the synthesis of proteins that translocate sugars (Gupta et al., 1985).
Sources, uptake and translocation of B
Most B in soil occurs as axenite, tourmaline, ulexite, colemenite and kermite; they are relatively insoluble and are released very slowly so that small amounts are available to plants (Gupta et al., 1985). Plant-available B is mainly associated with soil organic matter and soil solution and is dependent on soil physical and chemical properties, with the highest concentrations found in soils derived from marine sediments, and the lowest concentrations in light sandy soils derived from granites (Weir and Cresswell, 1995; Peverillet al., 1999). Low B concentrations are associated with low organic matter, high rainfall or irrigation, high pH, high Ca and dry soils (Gupta et al., 1985).

Crop Production: Mineral Nutrition 423
Supplementary forms of B include sodium borate as borax (Na2B4O7, 10% B) and as solubor (Na2B8O13·4H2O, 20% B), boric acid (HBO3-, 18% B), calcium borate (Ca3(BO3)2, 12% B) and calcium-sodium borate (CaNa3B5O10, 18% B).
Boron is primarily taken up by the roots as un-dissociated boric acid (B(OH)3) and transported in the xylem vessels by mass fl ow, accumulating in organs with the highest transpiration rates, that is leaves and growing shoots. There is only limited redistribution of B via the phloem (Raven, 1980); how-ever, this limited redistribution and xylem supply is usually suffi cient for normal tree health (Shelp et al., 1995).
Boron defi ciency and toxicity
Boron defi ciency is expressed in tissues that are rapidly expanding and have low transpirational rates, for example roots, fruits and shoots (Shelp et al., 1995). In mango, B defi ciency can result in poor fl owering, pollination and reduced fruit set. It is expressed in growing shoots by uneven cell division, causing leaves to grow lop-sided with a curved sickle shape and deformed lamina and margins (Plate 78). Leaves often have shot-holes that are sur-rounded by a light-green halo and ragged margins. Apical dominance can be lost with swelling of the internodes. The main raceme of panicles can develop a slight bend or kink towards the tip and in some cultivars the bark splits and oozes black gummy sap from the cracks, known as gummosis (Plate 78) (Nartvaranant et al., 2002). Agarwala et al. (1988) generated B defi ciency in 1-year-old seedlings using sand culture, and described mild defi ciency symp-toms, as mild chlorosis with a marked reduction in length and width of the leaves. In more severe cases, older leaves become chocolate-brown at the base, spreading to the tip before becoming completely chlorotic. Stems turned black, lost apical dominance and eventually stopped growing. Boron defi ciency can be aggravated by high N status of trees (Ram et al., 1989; Raja et al., 2005).
In mango fruit, B defi ciency causes cracks that split open; there is brown discoloration of the mesocarp. Lumpy, deformed fruit may also be a symp-tom of B defi ciency. Meneses et al. (1994) examined normal and deformed fruit using neutron capture radiography, and suggested that the deformities were due to B toxicity. Boron toxicity is common in mango with typical symp-toms appearing in leaves as dark spots on leaf margins that coalesce, eventu-ally leading to marginal leaf necrosis in more severe cases (Plate 79). Boron toxicity often occurs after excessive application of B fertilizers. Symptoms can be amended through leaching of fertilizer from the root zone, raising soil pH with applications of lime or stimulating growth through application of N; however, these measures may have other implications for crop production.
Effect of B on crop production and fruit quality
Boron defi ciency symptoms of trunk gummosis in ‘Kyo Savoy’ and ‘Nam Doc Mai’ in Thailand were successfully remedied by soil applications of 20 or

I.S.E. Bally424
25 g/m borax (11% B) during the summer wet season; however, the response time and effect on gummosis of the two cultivars differed (Nartvaranant et al., 2002). Foliar application of B solutions at the pre-fl owering stage increased yield and fruit quality in several studies. Dutta (2004) observed that 3000 ppm boric acid is optimal for maximizing yield and quality of ‘Him-sagar’ in West Bengal, India. Coetzer et al. (1991) reported that foliar applica-tion of B at fl owering raised leaf B concentrations to 60 mg/kg and increased yields from 14 to 22 kg/tree as well as improving fruit quality. When Loría-Meneses et al. (1992) applied boric acid to the skin of developing fruit they found it was not signifi cantly translocated into the mesocarp and remained in the surface layers of the skin around secretary glands. The response of mango trees to soil applications of B varies between cultivars. Rossetto et al. (2000) observed that ‘Winter’ was least sensitive, whereas, ‘Tommy Atkins’, ‘Haden’ and ‘Van Dyke’ in declining order were more sensitive.
12.15 Conclusion
Our understanding of mineral nutrition in mango is incomplete and lacks detailed knowledge of the effects of many minerals, as demonstrated above. A better understanding has generally come from observations of the effects of minerals on fruit quality rather than yield. Studying the effects of minerals on mango production and fruit quality is diffi cult because of their delayed response to applied minerals and large tree reserves. The effects of minerals on productivity and fruit quality can be greatly enhanced by also determin-ing their effects on other phenological events that contribute to productivity and fruit quality. More regular and targeted assessment of mango tree min-eral status will facilitate improved management of mango trees. This may require the determination of fruit mineral concentration standards for differ-ent stages of fruit development. Development and employment of low-cost, rapid and non-destructive techniques to measure minerals will enable mineral fl uxes within trees to be monitored closely.
References
Agarwala, S.C., Nautiyal, B.D., Chatterjee, C. and Sharma, C.P. (1988) Manganese, zinc and boron defi ciency in mango. Scientia Horticulturae 35, 99–107.
Aitken, R.L. and Scott, B.J. (1999) Magnesium. In: Peverill, K.I., Sparrow, L.A. and Reuter, D.J. (eds) Soil Analysis: an Interpretation Manual. Commonwealth Scientifi c and Industrial Research Organization (CSIRO), Melbourne, Australia, pp. 255–262.
Alvian, R.L. (1974) Mango fertilization on soils of the Maracay series. Agronomia Tropi-cal 22, 535–539.
Alvian, R.L. and Figueroa, M. (1977) Timing of nitrogen application in mangoes grown on Maracay soil in Aragua (Argenti Venezuela). Agronomia Tropical 27, 491–501.
Anonymous (1988) Sulfur. Consolidated Fertiliser Ltd (CFL), Brisbane, Australia.Anonymous (2007) A Guide to Soil Sampling. Spectrum Analytic. Available at: http://
www.spectrumanalytic.com/support/library/ff/soil_sampling_instructons.htm (accessed 16 March 2008).

Crop Production: Mineral Nutrition 425
Bally, I.S.E. (2007) The effect of preharvest nutrition and crop load on fruit quality and postharvest disease in mango (Mangifera indica L.). PhD dissertation, University of Queensland, Brisbane, Australia.
Bangerth, F. (1979) Calcium-related physiological disorders of plants. Annual Review of Phytopathology 17, 97–122.
Banuelos, G.S., Bangerth, F. and Marschner, H. (1987) Relationship between polar basipetal auxin transport and acropetal Ca2+ transport in tomato fruit. Physiologia Plantarum 71, 321–327.
Beaufi ls, E.R. (1973) Diagnosis and recommendation integrated system (DRIS): a gen-eral scheme of experimentation and calibration based on principles developed from research in plant nutrition. Soil Science Bulletin University of Natal 1, 131.
Bhandal, I.S. and Malik, C.P. (1988) Potassium estimation, uptake and its role in the physiology and metabolism of fl owering plants. International Review of Cytology 110, 205–245.
Bhargava, B.S. and Chadha, K.L. (1988) Leaf nutrient guide for fruit and plantation crops. Fertiliser News 33, 21–29.
Burdon, J.N., Moore, K.G. and Wainwright, H. (1991) Mineral distribution in mango fruit susceptible to the physiological disorder soft-nose. Scientia Horticulturae 48,329–336.
Burdon, J.N., Moore, K.G. and Wainwright, H. (1992) A preliminary examination of the physiological disorder ‘soft-nose’ in mango fruit. Acta Horticulturae 296, 15–22.
Catchpoole, D.W. and Bally, I.S.E. (1995) Nutrition of mango trees: a study of the rela-tionship between applied fertiliser, leaf elemental composition, and tree perfor-mance (fl owering and fruit yield). In: Proceedings of the Mango 2000 Marketing Seminar and Production Workshop. Queensland Department of Primary Industries and Fisheries, Townsville, Queensland, Australia, pp. 91–104.
Chadha, K.L., Samra, J.S. and Thakur, R.S. (1980) Standardisation of leaf-sampling technique for mineral composition of leaves of mango cultivar ‘Chausa’. ScientiaHorticulturae 13, 323–329.
Chattopadhyay, P.K. and Sarkar, S.K. (1990) Studies on the mineral composition of infl orescence and fruits at different stages of growth in four commercial culti-vars of mango in West Bengal. Haryana Journal of Horticultural Sciences 19, 268–272.
Chaudhary, S.K. and Nauriyal, J.P. (1988) Effect of defi ciency of calcium, magnesium and sulphur on the uptake of other nutrients in mango. Acta Horticulturae 231,296–300.
Clarkson, D.T. (1984) Calcium transport between tissue and its distribution in the plant. Plant, Cell and Environment 7, 449–465.
Coetzer, L.A., Robbertse, P.J. and de Wet, E. (1991) The infl uence of boron applications on fruit production and cold storage of mangoes. South African Mango Growers’ Association Yearbook 11, 29–31.
Cracknell Torres, A., Cid Ballarim, M.C., Socorro Monzon, A.R., Fernandez Galvan, D., Rosell Garcia, P. and Galan Sauco, V. (2004) Effects of nitrogen and calcium supply on the incidence of internal fruit breakdown in ‘Tommy Atkins’ mangoes (Mangiferaindica L.) grown in a soilless system. Acta Horticulturae 645, 387–401.
Crane, J.H., Bally, I.S.E., Tomer, E. and Mosqueada-Vazqez, R. (1997) Crop production. In: Litz, R.E. (ed.) The Mango: Botany, Production and Uses. CAB International, Wallingford, UK, pp. 203–256.
Daulta, B.S., Singh, H.K. and Chauhan, K.S. (1981) Effect of zinc and CCC sprays on fl owering fruiting and physio-chemical composition of fruit in mango (Mangiferaindica L.) cv. Dashehari. Haryana Journal of Horticultural Science 10, 161–165.

I.S.E. Bally426
Davenport, T.L. and Núñez-Elisea, R. (1997) Reproductive physiology of mango. In: Litz, R.E. (ed.) The Mango: Botany, Production and Uses. CAB International, Wallingford,UK, pp. 69–146.
Dilly, O., Romheld, V., Marschner, H. and Chen, Y. (1997) Nutritional disorder in mango trees on calcareous soil in Israel. Acta Horticulturae 448, 360.
Dow, A.I., Rushmore, F.A., Halvorson, A.R. and Tukey, R.B. (1991) Orchard Soil Sampling. Washington State University. Available at: http://cru.cahe.wsu.edu/CEPublications/eb1595/eb1595.html (accessed 16 March 2008).
Dutta, P. (2004) Effect of foliar boron application on panicle growth, fruit retention and physio-chemical characters of mango cv. Himsagar. Indian Journal of Horticulture 61, 265–266.
Duttaray, S.K., Singh, R.D.N., Kabir, J., Dhua, R.S. and Kaiser, S. (1993) Effect of pre-harvest treatment of calcium compounds on the incidence of Diplodia stem-end rot of mango in storage. Crop Research Hisar 63, 419–422.
Fallahi, E., Conway, W.S., Hickey, K.D. and Sams, C.E. (1977) The role of calcium and nitrogen in postharvest quality and disease resistance of apples. HortScience 32,831–835.
Ferguson, I.B. (1984) Calcium in plant senescence and fruit ripening. Plant, Cell and Environment 7, 477–489.
Gonzalez, A., Azam, G., Madddern, T. and Renfree, R. (2005) Evaluating the Use of the Spad-502 Meter to Manage Nitrogen in Trees (‘Kensington Pride’) in the Northern Territory. Department of Primary Industry, Fisheries and Mines – Technical Annual Report 323. Department of Primary Industry, Fisheries and Mines, Darwin, NorthernTerritory, Australia.
Gourley, C.J.P. (1999) Potassium. In: Peverill, K.I., Sparrow, L.A. and Reuter, D.J. (eds) Soil Analysis: an Interpretation Manual. Commonwealth Scientifi c and Industrial Research Organization (CSIRO), Melbourne, Australia, pp. 229–245.
Gunjate, R.T., Tare, S.J., Rangawala, A.D. and Limaye, V.P. (1979) Changes in calcium content in ‘Alphonso’ mango fruits and leaves from fruit-set to harvesting. IndianJournal of Horticulture 36, 383–386.
Gupta, U.C., Jame, Y.W., Campbell, C.A., Leyshon, A.J. and Nicholaichuk, W. (1985) Boron toxicity and defi ciency: a review. Canadian Journal of Soil Science 65, 318–409.
Ho, L.C. and Adams, P. (1989) Calcium defi ciency – a matter of inadequate transport to rapidly growing organs. Plants Today 2, 202–207.
Hundal, H.S., Singh, D. and Brar, J.S. (2005) Diagnosis and recommendation integrated system for monitoring nutrient status of mango trees in submountainous area of Punjab, India. Communications in Soil Science and Plant Analysis 36, 2085–2099.
Kadiyala, H.B. (1995) Composition and distribution of minerals in mango fruit during ripening and storage with reference to ‘internal breakdown’, a postharvest disorder. In: Proceedings of the International Symposium on Postharvest Science and Technology of Horticultural Crops. China Agricultural Scientech Press, Beijing, pp. 10–17.
Kanwar, J.S., Nijjar, G.S. and Kahlon, G.S. (1987) Effect of nitrogen, phosphorus and potassium fertiliser on the growth and productivity of ‘Dashehari’ mango (Mangiferaindica). Journal of Research of the Punjab Agricultural University 24, 411–422.
Kirkby, E.A. (1979) Maximising calcium uptake by plants. Communications in Soil Scienceand Plant Analysis 10, 89–113.
Kirkby, E.A. and Pilbeam, D.J. (1984) Calcium as a plant nutrient. Plant, Cell and Envi-ronment 7, 397–405.

Crop Production: Mineral Nutrition 427
Klumpp, A., Hintemann, T., Lima, J.S. and Kandeler, E. (2003) Bioindication of air pollution effects near a copper smelter in Brazil using mango trees and soil microbiological properties. Environmental Pollution 126, 313–321.
Kumar, O.V. and Kumar, G. (1989) Effect of pre-harvest foliar sprays of zinc on post-harvest changes in the quality of mango cv. Dashehari. Acta Horticulturae 231,763–770.
Kumar, S. and Nauriyal, J.P. (1977) Nutritional studies on mango – tentative leaf analysis standards. Indian Journal of Horticulture 34, 100–106.
Kumar, S. and Nauriyal, J.P. (1979) Foliar sampling technique in mango. Punjab Horti-cultural Journal 19, 10–15.
Lim, T.K. and Khoo, K.C. (1985) Diseases and Disorders of Mango in Malaysia. Tropical Press, Kuala Lumpur, Malaysia.
Littlemore, J., Winston, E.C., Howitt, C.J., Farrell, P.O. and Wiffen, D.C. (1991) Improved methods for zinc and boron applications to mango (Mangifera indica L.) cv. Kensington Pride in the Mareeba-Dimbulah district of North Queensland. Austra-lian Journal of Experimental Agriculture 31, 117–121.
Loría-Meneses, L.G., Jiménez-Dam, R., Gallardo-Badilla, M., Martini, F. and Theller, M. (1992) A study of the distribution and uptake of boron in mango fruits using neutron-capture-radiography. Comtes Rendus de l’Academie des Sciences, Paris 315, 255–264.
Malo, S.E. and Campbell, C.W. (1978) Studies on mango fruit breakdown in Florida. Proceedings of the Tropical Region of the American Society for Horticultural Science22, 1–19.
Marcelle, R., Simon, P. and Lennes, G. (1990) The effects of IAA and TIBA on calcium transport and accumulation in fruits. Acta Horticulturae 120, 193–198.
Marchal, J., Goguly, T. and Didier, C. (1991) A mineral assessment of the mango tree, variety Amelie. Fruits 46, 477–487.
Marschner, H. (1995) Mineral Nutrition of Higher Plants, 2nd edn. Academic Press, London.
McKenzie, C.B. (1994) The background skin colour of exported mango fruit in relation to tree nitrogen status. South African Mango Growers’ Association Yearbook 14,26–28.
McKenzie, C.B. (1995) The effect of calcium and potassium foliar and fruit sprays on ‘Sensation’ mango leaf nutrient concentrations and fruit quality. South African Mango Growers’ Association Yearbook 15, 44–47.
Medeiros, A.A., Amorim, J.A.R., Silva, D.J., Dantas, J.A. and Guerra, A.G. (2004) Min-eral composition of leaves and fruits of irrigated mango trees in Rio Grande do norte state Brazil. Acta Horticulturae 645, 403–408.
Meneses, L.G.L., Dam, R.J. and Martini, F. (1994) Neutron-capture radiography of the distribution of boron in normal and deformed mango fruits. Journal of Trace and Microprobe Techniques 12, 97–102.
Mengel, K. and Kirkby, E.A. (1987) Principles of Plant Nutrition, 4th edn. International Potash Institute, Worblaufen-Bern, Switzerland.
Monselise, S.P. and Goren, R. (1987) Preharvest growing conditions and postharvest behaviour of subtropical and temperate-zone fruits. HortScience 22, 1185–1189.
Nartvaranant, P., Subhadrabandhu, S. and Whiley, A.W. (2002) Effect of soil boron application on gummosis and leaf boron content of mango (Mangifera indica L.)cvs. Khieo Sawoei and Nam Dok Mai. Acta Horticulturae 575, 875–879.
Narwadkar, P.R. and Pandey, R.M. (1989) Studies on the phosphorus translocation in mango with special reference to alternate bearing. Acta Horticulturae 231, 440–447.
Nguyen, H., Hofman, P., Holmes, R., Bally, I., Stubbings, B. and McConchie, R. (2004) Effect of nitrogen on the skin colour and other quality attributes of ripe ‘Kensington

I.S.E. Bally428
Pride’ mango (Mangifera indica L.) fruit. Journal of Horticultural Science and Bio-technology 79, 204–210.
Oosthuyse, S.A. (1993) Effect of spray application of KNO3, urea and growth regulators on the yield of ‘Tommy Atkins’ mango. South African Mango Growers’ Association Yearbook 13, 58–62.
Oosthuyse, S.A. (1997) Effect of KNO3 sprays to fl owering mango trees on fruit reten-tion, fruit size, tree yield, and fruit quality. Acta Horticulturae 455, 359–366.
Oosthuyse, S.A. (2000a) Cost reduction of powdery mildew control in mango with mono potassium phosphate. Acta Horticulturae 509, 719–723.
Oosthuyse, S.A. (2000b) Variation of leaf nutrition status in relation to fruit growth in mango. Acta Horticulturae 509, 375–378.
Peverill, K.I., Sparrow, L.A. and Reuter, D.J. (eds) (1999) Soil Analysis: an Interpretation Manual. Commonwealth Scientifi c and Industrial Research Organization (CSIRO), Melbourne, Australia.
Pimplasker, M. and Bhargava, B.S. (2003) Leaf and soil nutrient norms in mango (Mangifera indica L.) grown in tribal belt of Southern Gujarat. Journal of the Indian Society of Soil Science 51, 268–272.
Protacio, C.M. (2000) A model for potassium nitrate-induced fl owering in mango. Acta Horticulturae 509, 545–552.
Raghupathi, H.B., Reddy, Y.T.N., Rein, M.K. and Bhargava, B.S. (2004) Diagnosis of nutrient imbalance in mango by DRIS and PCA approaches. Journal of Plant Nutri-tion 27, 1131–1148.
Raj, G.B. and Rao, A.P. (2006) Identifi cation of yield-limiting nutrients in mango through DRIS indices. Communications in Soil Science and Plant Analysis 37, 1761–1774.
Raja, M.E., Kumar, S.C.A. and Raju, S.Y. (2005) Boron defi ciency in mango (Mangiferaindica L.): a cause delineation study in acidic soils of Maharashtra, India. Soil Sci-ence and Plant Nutrition 51, 751–754.
Ram, S., Bist, L.D. and Sirohi, S.C. (1989) Internal fruit necrosis of mango and its control. Acta Horticulturae 231, 805–813.
Rane, D.A., Katordia, J.S. and Kulkarni, D.N. (1976) Problem of spongy tissue develop-ment in mango (Mangifera indica L.) cv. Alphonso, a review. Journal of Maharashtra Agricultural University 1, 89–94.
Rao, D.P. and Mukherjee, S.K. (1989) Nutrient status in leaf and soil of some cultivars of mango in relation to yield. Acta Horticulturae 231, 286–295.
Raven, J.A. (1980) Short- and long-distance transport of boric acid in plants. New Phy-tologist 84, 231–249.
Raymond, L., Schaffer, B., Brecht, J.K. and Crane, J.H. (1998a) Internal breakdown in mango fruit: symptomology and histology of jelly seed, soft nose and stem-end cav-ity. Postharvest Biology and Technology 13, 59–70.
Raymond, L., Schaffer, B., Brecht, J.K. and Hanlon, E.A. (1998b) Internal breakdown, mineral element concentration and weight of mango fruit. Journal of Plant Nutrition 21, 871–889.
Reddy, S.E. and Majmudar, A.M. (1985) Tracking phosphorus patterns in mango (Mangifera indica L.) and possible relations to fl oral induction. Nutrient Cycling in Agroecosystems 6, 225–234.
Reddy, Y.T.N., Kurian, R.M., Kohli, R.R. and Singh, G. (2000) Effect of nitrogen, phos-phorus and potassium on growth, yield and fruit quality of ‘Totapuri’ mango (Mangifera indica). Indian Journal of Agricultural Sciences 70, 475–478.
Reddy, Y.T.N., Kumar, R.M., Singh, G. and Raghupati, H.B. (2003) Long-term effects of nitrogen on growth, leaf-nutrient status and fruit yield of ‘Totapuri’ mango (Mangiferaindica). Indian Journal of Agricultural Sciences 73, 206–208.

Crop Production: Mineral Nutrition 429
Reuter, D.J. and Robinson, J.B. (eds) (1997) Plant Analysis: an Interpretation Manual, 2nd edn. Commonwealth Scientifi c and Industrial Research Organization (CSIRO), Melbourne, Australia.
Reuter, D.J., Robinson, J.B., Peverill, K.I. and Price, G.H. (1986) Guidelines for collecting, handling, and analysing plant materials. In: Reuter, D.J. and Robinson, J.B. (eds) Plant Analysis: an Interpretation Manual. Inkata, Melbourne, Australia, pp. 20–32.
Reuveni, M., Harpaz, M. and Reuveni, R. (1998) Integrated control of powdery mildew on fi eld-grown mango trees by foliar sprays of mono-potassium phosphate fertilizer, sterol inhibitor fungicides and the strobilurin Kresoxym-methyl. European Journal of Plant Pathology 104, 853–860.
Rhodes, K.J.C. (1980) The maturation and ripening of fruits. In: Thimann, K.V. (ed.) Senes-cence in Plants. CRC Press, Boca Raton, Florida, pp. 157–205.
Robinson, J.B. (1986a) Fruits, vines and nuts. In: Reuter, D.J. and Robinson, J.B. (eds) Plant Analysis: an Interpretation Manual. Inkata, Melbourne, Australia, pp. 120–147.
Robinson, J.B. (1986b) Tree mineral nutrition. Acta Horticulturae 175, 163–172.Robinson, J.B., Treeby, M.T. and Stephenson, R.A. (1997) Fruits, vines and nuts. In: Reuter,
D.J. and Robinson, J.B. (eds) Plant Analysis: an Interpretation Manual. Inkata, Mel-bourne, Australia, pp. 349–382.
Rojas, E. and Leal, F. (1997) Effects of pruning and potassium nitrate spray on fl oral and vegetative bud break of mango cv. Haden. Acta Horticulturae 455, 522–529.
Rossetto, C.J., Furlani, P.R., Bortoletto, N., Quaggio, J.A. and Igue, T. (2000) Differential response of mango varieties to boron. Acta Horticulturae 509, 259–264.
Saleh, M.M.S. and El-Monem, E. (2003) Improving the productivity of ‘Fagri Kalan’ man-go trees grown under sandy soil conditions using potassium, boron and sucrose as foliar spray. Annals of Agricultural Science (Cairo) 48, 747–756.
Salisbury, F.B. and Ross, C.W. (1992) Plant Physiology. Wadsworth, Belmont, California.
Samra, J.S. and Arora, Y.K. (1997) Mineral nutrition. In: Litz, R.E. (ed.) The Mango: Botany, Production and Uses. CAB International, Wallingford, UK, pp. 175–201.
Schaffer, B. (1994) Mango disorders caused by abiotic factors. In: Ploetz, R.C., Zentmyer, G.A., Nishijima, W.T., Rohrbach, K.G. and Ohr, H.D. (eds) Compendium of Tropical Fruit Diseases. American Phytopathological Society, St Paul, Minnesota, pp. 43–44.
Schaffer, B., Larson, K.D., Snyder, G.H. and Sanchez, C.A. (1988) Identifi cation of mineral defi ciencies associated with mango decline by DRIS. HortScience 23, 617–619.
Schmidt, W. (1999) Mechanisms and regulation of reduction-based iron uptake in plants. New Phytologist 141, 1–26.
Scholefi eld, P.B., Oag, D.R. and Sedgley, M. (1986) The relationship between vegetative and reproductive development in the mango in Northern Australia. Australian Jour-nal of Agricultural Research 37, 425–433.
Sen, P.K., Roy, P.K. and De, B.N. (1947) Hunger signs on mango. Indian Journal of Hor-ticulture. 5, 35–44.
Sergent, E. and Leal, F. (1989) Flowering induction in mango (Mangifera indica L.) with KNO3. Revista de la Facultad de Agronomia, Universidad Central de Venezuela 15, 17–32.
Sergent, E., Ferrari, D. and Leal, F. (1997) Effects of potassium nitrate and paclobutrazol on fl owering induction and yield of mango (Mangifera indica L.) cv. Haden. Acta Horticulturae 455, 180–187.
Sergent, E., Leal, F. and Anez, M. (2000) Potassium thiosulphate, urea and potassium nitrate applications on vegetative and fl oral growth in mango ‘Haden’. Acta Horti-culturae 509, 653–659.

I.S.E. Bally430
Shear, C.B. (1975) Calcium related disorders of fruits and vegetables. HortScience 10,316–365.
Shelp, B.J., Marentes, E., Kitheka, A.M. and Vivekanandan, P. (1995) Boron mobility in plants. Physiologia Plantarum 94, 356–361.
Shinde, A.K., Dabke, D.J., Jadhav, B.B., Kandalkar, M.P. and Burondkar, M.M. (2006) Effect of dose and source of potassium on yield and quality of ‘Alphonso’ mango (Mangifera indica). Indian Journal of Agricultural Science 76, 213–217.
Shongwe, V. and Roberts-Nkrumah, L.B. (1997) Physiological and growth responses of mango (Mangifera indica L.) to methanol and potassium nitrate application. Acta Horticulturae 455, 64–70.
Shorter, A.J. and Joyce, D.C. (1998) Effect of partial pressure infi ltration of calcium into ‘Kensington’ mango fruit. Australian Journal of Experimental Agriculture 38, 287–294.
Shu, Z., Sheen, T.F., Lin, S.L. and Lee, K.C. (1992) Effects of microelement-containing pesticides on nutrient concentrations of mango leaves. Acta Horticulturae 321,553–560.
Silva, D.J., Quaggio, J.A., Pinto, J.A.C., Pinto, A.C.Q. and Megalhaes, A.F.J. (2002) Nutrição e Adubação. In: A Cultura da Mangueira. Embrapa Informação Tecnológi-ca, Brasília, Brazil, pp. 191–221.
Simmons, S.L., Hofman, P.J., Whiley, A.W. and Hetherington, S.E. (1997) Effects of pre-harvest calcium sprays and fertilisers, leaf:fruit ratios and water stress on mango fruit quality. In: Coates, L.M., Hofman, P.J. and Johnson, G.I. (eds) Proceedings of the Disease Control and Storage Life Extension in Fruit. Australian Centre for Inter-national Agricultural Research (ACIAR), Canberra, Australia, pp. 19–26.
Singh, B.P., Singh, V.B., Shrivastava, P.K. and Singh, P.R. (1973) Effects of urea applica-tion on growth performance and mineral content of mangoes (Mangifera indica L.) variety Langra and Chausa. Balwamt Vidyapeeth Journal of Agricultural and Scien-tifi c Research 15, 54–58.
Singh, M.P. and Saxena, P.K. (1994) Defi ciency and excess feeding signs of macro-elements on mango (Mangifera indica L) cv. Dashehari. Recent Horticulture 1, 30–35.
Singh, R.N., Singh, G., Mishra, J.S. and Rao, O.P. (1987) Studies on the effect of pre- and post-harvest treatment of calcium nitrate and calcium chloride on the storage life of Amrapali mango. Progressive Horticulture 19, 1–9.
Singh, R.R. and Rajput, C.B.S. (1977) Effects of various concentrations of zinc on vegeta-tive growth characters, fl owering, fruiting and physico-chemical composition of fruits in mango (Mangifera indica L.). Haryana Journal of Horticultural Science 6, 10–14.
Smith, B.L. (1992) Time of leaf sampling and analysis norms for mangoes (cv Sensation). South African Mango Growers’ Association Yearbook 12, 54–56.
Smith, P.F. and Scudder, G.K. (1951) Some studies of mineral defi ciency symptoms in mango. Proceedings of the Florida State Horticultural Society 64, 243–248.
Stanley, R.G. and Lichtenberg, E.A. (1963) The effect of various boron compounds on invitro germination of pollen. Physiologia Plantarum 16, 337–346.
Stassen, P.J.C., Mostert, P.G. and Smith, B.L. (1999) Mango tree nutrition. A crop per-spective. Neltropika Bulletin 303, 41–51.
Strong, W.M. and Mason, M.G. (1999) Nitrogen. In: Peverill, K.I., Sparrow, L.A. and Reuter, D.J. (eds) Soil Analysis: an Interpretation Manual. Commonwealth Scientifi c and Industrial Research Organization (CSIRO), Melbourne, Australia, pp. 171–184.
Subramanyam, H., Krishnamurthy, S., Subhadra, N.V., Dalal, V.B., Randhawa, G. and Chacko, E.K. (1971) Studies on mineral breakdown, a physiological ripening disor-der in ‘Alphonso’ mangoes (Mangifera indica L.). Tropical Sciences 13, 203–211.

Crop Production: Mineral Nutrition 431
Syamal, M.M. and Mishra, K.A. (1989) Effect of NPK on growth, fl owering, fruiting and quality of mango. Acta Horticulturae 231, 276–281.
Tarmizi, A.S., Tengku, A.B.T.M., Pauziah, M. and Zahrah, T. (1993) Incidence of insidi-ous fruit rot as related to mineral nutrients in Harumanis mangoes. Journal of the Malaysian Agricultural Research and Development Institute 21, 43–49.
Tiwari, J.P. and Rajput, C.B.S. (1975) Effects of urea sprays on vegetative growth and fruit. Bangladesh Horticulture 3, 31–36.
Tiwari, J.P. and Rajput, C.B.S. (1976) Signifi cance of nitrogen on the growth, fl owering and fruiting of mango cultivars. Acta Horticulturae 57, 29–36.
Torres, M.D., Farre, J.M. and Hermoso, J.M. (2004) Nitrogen and calcium fertilisation on productivity and fruit quality of mango cv. Sensation. Acta Horticulturae 645,394–402.
Vuuren, B.P.H.J. and Stassen, P.J.C. (1997) Seasonal uptake of macro elements by young bearing ‘Sensation’ mango trees. Acta Horticulturae 455, 167–174.
Wadt, P.G.S., Silva, D.J., Maia, C.E., Tome, J.B., Pinto, P.A.D. and Machado, P. (2007) Modelling of functions in calculating DRIS indices. Pesquisa Agropecuaria Brasile-ira 42, 57–64.
Weir, R.G. and Cresswell, G.C. (1995) Plant Nutrient Disorders. 2 Tropical Fruit and Nut Crops. Inkata, Melbourne, Australia.
Weng, F.Y. and Chuang, T.Y. (1997) Nutrient requirement of conidial germination and characteristics of spore matrix for mango anthracnose fungus. Plant Pathology Bul-letin 6, 17–24.
Yeshitela, T., Robbertse, P.J. and Stassen, P.J.C. (2005) Potassium nitrate and urea sprays affect fl owering and yields of ‘Tommy Atkins’ (Mangifera indica) mango in Ethiopia. South African Journal of Plant and Soil 22, 28–32.
Young, T.W. (1957) ‘Soft nose’ a physiological disorder in mango fruits. Proceedings of the Florida State Horticultural Society 70, 280–283.
Young, T.W. and Koo, R.C.J. (1969) Mineral composition of Florida mango leaves. Pro-ceedings of the Florida State Horticultural Society 82, 324–328.
Young, T.W. and Koo, R.C. (1971) Variations in mineral content of Florida mango leaves. Journal of the Florida State Horticultural Society 84, 298–303.
Young, T.W. and Koo, R.C.J. (1975) Increasing yield of ‘Parvin’ and ‘Kent’ mangoes on Lakewood sands by increased nitrogen and potassium fertilisation. Proceedings of the Florida State Horticultural Society 87, 380–384.
Young, T.W. and Miner, J.T. (1961) Relationship of nitrogen and calcium to ‘soft nose’ disorder in mango fruits. Journal of the American Society for Horticultural Science 78, 201–208.
Young, T.W. and Sauls, J.W. (1981) The mango industry in Florida. Florida Cooperative Extension Service Bulletin 189, 1–70.

© CAB International 2009. The Mango, 2nd Edition: Botany, Production and Uses432 (ed. R.E. Litz)
13 Crop Production: Management
J.H. Crane,1 S. Salazar-García,2 T.-S. Lin,3
A.C. de Queiroz Pinto4 and Z.-H. Shü5
1University of Florida, Florida, USA 2Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, Santiago
Ixcuintla, Mexico 3National Taiwan University, Taipei, Taiwan
4Private Consultant on Tropical Fruits, Brasilia, Brazil5Meiho Institute of Technology, Pingtung, Taiwan
13.1 Introduction 43213.2 Production Areas and Yields 43413.3 Climate of Production Areas 43613.4 Soils and Soil Preparation 44013.5 Plant Propagation and Rootstocks 44313.6 Major Cultivars 44513.7 Plant Spacing 44913.8 Fertilizer Practices 45213.9 Irrigation Practices 46013.10 Vegetative Growth and Reproduction Manipulation 46313.11 Environmental Stress Management 46813.12 Harvesting Practices 47013.13 Conclusions 472
13.1 Introduction
Mangoes have been in cultivation for several thousand years (Mukherjee, 1953, 1972; Kostermans and Bompard, 1993), and crop production practices have continually improved (Singh, 1960, 1978; Crane et al., 1997). A detailed understanding of mango plant physiology and behaviour in relation to cli-matic and edaphic conditions, genetics and cultural practices dates back to the late 1950s (Mukherjee, 1953; Chadha and Pal, 1986; Chacko, 1989; Whiley, 1993; Schaffer et al., 1994; Kulkarni, 2004; Davenport, 2006). Recent reviews (Cull, 1991; Whiley, 1993; Schaffer et al., 1994; Davenport and Núñez-Elisea, 1997; Kulkarni, 2004; Davenport, 2006) on mango crop management and physiology illustrate that the best prospect for improving mango production must involve a holistic approach to cultural practices that considers the spe-cifi c climatic and edaphic environment, cultivar and tree phenology for a
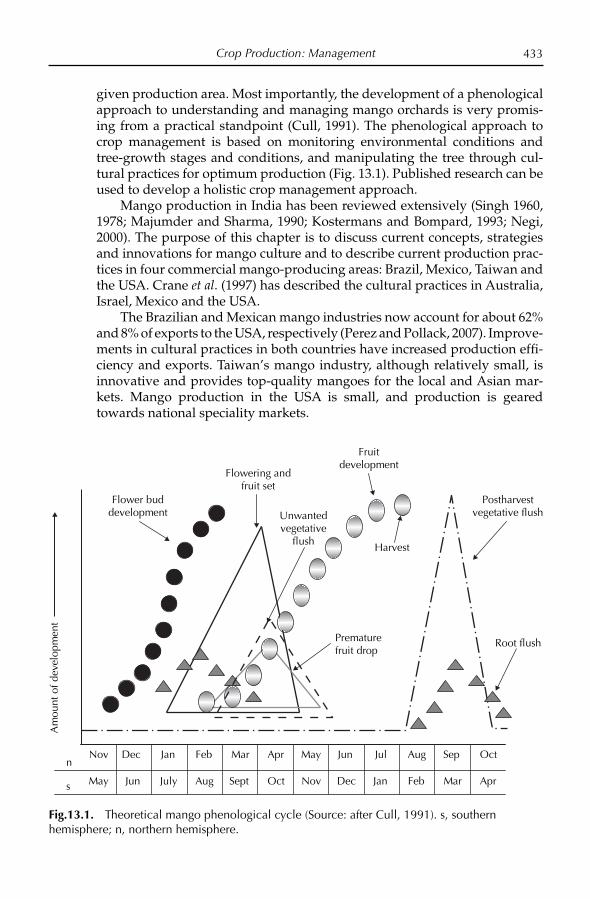
Crop Production: Management 433
given production area. Most importantly, the development of a phenological approach to understanding and managing mango orchards is very promis-ing from a practical standpoint (Cull, 1991). The phenological approach to crop management is based on monitoring environmental conditions and tree-growth stages and conditions, and manipulating the tree through cul-tural practices for optimum production (Fig. 13.1). Published research can be used to develop a holistic crop management approach.
Mango production in India has been reviewed extensively (Singh 1960, 1978; Majumder and Sharma, 1990; Kostermans and Bompard, 1993; Negi, 2000). The purpose of this chapter is to discuss current concepts, strategies and innovations for mango culture and to describe current production prac-tices in four commercial mango-producing areas: Brazil, Mexico, Taiwan and the USA. Crane et al. (1997) has described the cultural practices in Australia, Israel, Mexico and the USA.
The Brazilian and Mexican mango industries now account for about 62% and 8% of exports to the USA, respectively (Perez and Pollack, 2007). Improve-ments in cultural practices in both countries have increased production effi -ciency and exports. Taiwan’s mango industry, although relatively small, is innovative and provides top-quality mangoes for the local and Asian mar-kets. Mango production in the USA is small, and production is geared towards national speciality markets.
AprMarFebJanDecNovOctSeptAugJulyJunMay
OctSepAugJulJunMayAprMarFebJanDecNovn
s
Am
ount
of d
evel
opm
ent
Flower buddevelopment
Flowering andfruit set
Fruitdevelopment
Prematurefruit drop
Harvest
Root flush
Unwantedvegetative
flush
Postharvestvegetative flush
Fig.13.1. Theoretical mango phenological cycle (Source: after Cull, 1991). s, southern hemisphere; n, northern hemisphere.
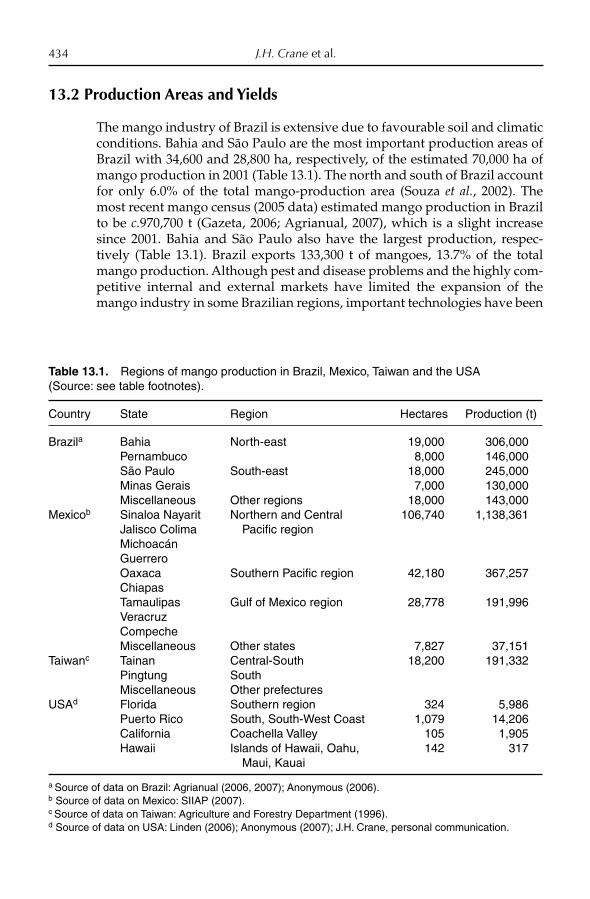
J.H. Crane et al.434
13.2 Production Areas and Yields
The mango industry of Brazil is extensive due to favourable soil and climatic conditions. Bahia and São Paulo are the most important production areas of Brazil with 34,600 and 28,800 ha, respectively, of the estimated 70,000 ha of mango production in 2001 (Table 13.1). The north and south of Brazil account for only 6.0% of the total mango-production area (Souza et al., 2002). The most recent mango census (2005 data) estimated mango production in Brazil to be c.970,700 t (Gazeta, 2006; Agrianual, 2007), which is a slight increase since 2001. Bahia and São Paulo also have the largest production, respec-tively (Table 13.1). Brazil exports 133,300 t of mangoes, 13.7% of the total mango production. Although pest and disease problems and the highly com-petitive internal and external markets have limited the expansion of the mango industry in some Brazilian regions, important technologies have been
Table 13.1. Regions of mango production in Brazil, Mexico, Taiwan and the USA (Source: see table footnotes).
Country State Region Hectares Production (t)
Brazila Bahia North-east 19,000 306,000Pernambuco 8,000 146,000São Paulo South-east 18,000 245,000Minas Gerais 7,000 130,000Miscellaneous Other regions 18,000 143,000
Mexicob Sinaloa Nayarit Northern and Central Pacifi c region
106,740 1,138,361Jalisco ColimaMichoacánGuerreroOaxaca Southern Pacifi c region 42,180 367,257ChiapasTamaulipas Gulf of Mexico region 28,778 191,996VeracruzCompecheMiscellaneous Other states 7,827 37,151
Taiwanc Tainan Central-South 18,200 191,332Pingtung SouthMiscellaneous Other prefectures
USAd Florida Southern region 324 5,986Puerto Rico South, South-West Coast 1,079 14,206California Coachella Valley 105 1,905Hawaii Islands of Hawaii, Oahu,
Maui, Kauai142 317
a Source of data on Brazil: Agrianual (2006, 2007); Anonymous (2006).b Source of data on Mexico: SIIAP (2007).c Source of data on Taiwan: Agriculture and Forestry Department (1996).d Source of data on USA: Linden (2006); Anonymous (2007); J.H. Crane, personal communication.

Crop Production: Management 435
developed to improve and support the mango industry (Santos Filho et al., 2002). This has resulted in an increase of 58% in mango exports from 1998 to 2005 (Souza et al., 2002; Agrianual, 2007).
There are c.181,525 ha of mangoes in 23 of Mexico’s 32 states (SIIAP, 2007); there are four large mango-producing regions (Table 13.1), ranging from 14°33N to 27°N latitude. They are distinguished by differences in cli-mate, soils and cultivars. There are 106,740 ha of mango in the Northern and Central Pacifi c region; the major cultivars are ‘Tommy Atkins’, ‘Ataulfo’, ‘Kent’, ‘Haden’ and ‘Keitt’. The Southern Pacifi c region has 42,180 ha and produces ‘Manila’ (‘Carabao’), ‘Ataulfo’, ‘Tommy Atkins’, ‘Haden’ and ‘Kent’. The leading cultivars of the Gulf of Mexico region (28,778 ha) are ‘Manila’ followed by ‘Tommy Atkins’. Mango production in Mexico was about 1.7 million t in 2006 and the states of Sinaloa, Guerrero, Nayarit, Oax-aca, Chiapas, Veracruz and Michoacán accounted for 92% of the crop (SIIAP, 2007). All these states except Veracruz (which abuts the Gulf of Mexico) are situated along the Pacifi c coastal and inland areas.
In 2006 Mexico exported 196,120 t to the USA valued at >US$132 million (EMEX, 2007). Mexico supplies 87.5% of the mangoes consumed in the USA and mango exports in 2006 were 17.9% higher than in the 2005. The major export cultivars are ‘Tommy Atkins’ (33%), ‘Kent’ (23%), ‘Ataulfo’ (19%), ‘Keitt’ (14%) and ‘Haden’ (11%). None the less, most mango production (c.85%) in Mexico is for the domestic market. The most important mango warehouses and distributors are in Mexico City, Guadalajara and Monterrey.
The 7-month mango season in Mexico is due to differences in latitude, phenological cycles, rainfall and soil moisture patterns and the use of growth regulators. Chiapas, along the South Pacifi c coast, is usually the fi rst to har-vest mangoes (‘Ataulfo’). Full bloom of ‘Ataulfo’ occurs from mid-November to mid-February and fruit is harvested from the end of January to the end of May. Mango harvesting continues northwards from Chiapas to the states of Oaxaca, Guerrero, Michoacán, Colima, Jalisco, Nayarit, Sinaloa and Sonora. Sinaloa is a major mango producer with the latest harvest season for ‘Manila’ (July), ‘Haden’ (July), ‘Tommy Atkins’ (July), ‘Kent’ (July–August) and ‘Keitt’ (August–September). The ‘Manila’ mango is produced mainly in Veracruz and Guerrero. Full bloom occurs in January–February and fruit is harvested in May–June.
The production area in Taiwan increased from 17,000 ha in 1987 to 21,000 ha in 1992 and stabilized at c.20,000 ha for c.10 years when the yield was 17 kg/tree; however, as production has increased to c.27 kg/tree, the plant-ing area has decreased to c.18,200 ha in 2006 (Table 13.1). There are six important mango-growing prefectures in Taiwan: Nantou, Taichung, Chiayi, Tainan, Kaohsiung and Pingtung from north to south in western and eastern Taiwan. Tainan and Pingtung are the most important production areas, com-prising about 90% of the total harvested area.
Mangoes are grown commercially in four areas of the USA: Florida (25–28°N), Puerto Rico (18–18°5´N), Hawaii (22–30°N) and California (33–33°5´N) (Table 13.1). Puerto Rico has the largest area (1079 ha) (Alvarado- Ortiz and Acin, 2004), followed by Florida (324 ha) (FASS, 2003; Anonymous,

J.H. Crane et al.436
2007; J.H. Crane, personal communication), Hawaii (142 ha) (NASS-Hawaii, 2006; Anonymous, 2007) and California (105 ha) (Linden, 2006; Anonymous, 2007). Mango production in the USA is estimated to be only 22,414 t with Puerto Rico accounting for >63% of the production (Alvarado-Ortiz and Acin, 2004; Linden, 2006; Anonymous, 2007). The four areas have different climates which present different production opportunities and challenges.
The main production areas of Florida are along the south-east coast in Miami-Dade County (25–27°N), the east (Palm Beach, Broward and Indian River Counties) and west (Lee County) coasts. The season in Florida is from May through to September. Puerto Rico is a Caribbean island (18°N, 67°W) (Espenshade, 1992; Bahr and Johnston, 1993c). Most of the commercial pro-duction is near Ponce on the south coast and Mayaguez in the south-west (23.9 ha). The main mango season in Puerto Rico is May through to Septem-ber, although some mangoes are harvested as late as mid-November. In Hawaii (19–22°N), production occurs on the islands of Hawaii, Oahu, Maui and Kauai. The season in Hawaii is from May to August, although some pro-duction occurs year-round. The California industry is on the western side of the Salton Sea in the Coachella Valley of southern California (33°N) (Scott, 1990). Mango production is from mid-August to October (Linden, 2006).
13.3 Climate of Production Areas
Mangoes are mainly cultivated in the tropics (25°N, 25°S) and subtropics (35°N, 35°S), although limited production also occurs in warm temperate/subtropical, i.e. Mediterranean-type areas, for example Israel, southern Spain and the Canary Islands and California. The ideal areas for mango production have a cool and/or dry period prior to fl owering followed by moderate soil moisture and moderately hot temperatures (30–33°C) (Chacko, 1986). The diversity of climates and soils in mango-production areas refl ects the adapt-ability of the species and improvements in cultural practices.
Temperature and availability of water are the most signifi cant environ-mental factors that infl uence commercial mango production by affecting the frequency, intensity, duration and time of vegetative growth and fl owering (Chacko, 1986, 1989; Whiley, 1993; Schaffer et al., 1994; Núñez-Elisea and Davenport, 1995; Davenport, 2006) and disease incidence (Johnson et al., 1989; Ploetz, 1994, 2003; Ploetz et al., 1994). Temperatures <15°C and >30°C inhibit pollen tube germination, thereby impeding fertilization and resulting in embryo abortion and fruit abscission.
Mango production in Brazil extends from 3°N, close to the Amazon region, where a humid, hot tropical climate predominates almost year-round to approximately 25°S in Paraná, which has a subtropical climate. Between these two extremes, there are various soil and climatic conditions where mango trees are grown intensively. The semi-arid tropical climate of the north-east region of Petrolina in Pernambuco (21°13´N and 47°50´W) is more appropriate for high-quality mango production than Votuporanga in São Paulo (21°13´S and 47°50´W), which has a subtropical climate (Table 13.2).
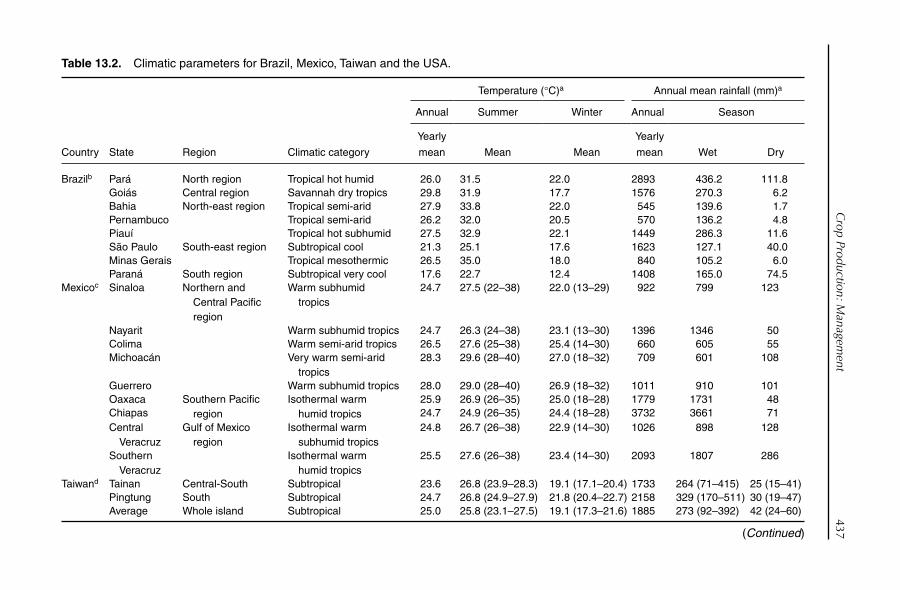
Crop Production: M
anagement
437Table 13.2. Climatic parameters for Brazil, Mexico, Taiwan and the USA.
Country State Region Climatic category
Temperature (°C)a Annual mean rainfall (mm)a
Annual Summer Winter Annual Season
Yearly
mean Mean Mean
Yearly
mean Wet Dry
Brazilb Pará North region Tropical hot humid 26.0 31.5 22.0 2893 436.2 111.8Goiás Central region Savannah dry tropics 29.8 31.9 17.7 1576 270.3 6.2Bahia North-east region Tropical semi-arid 27.9 33.8 22.0 545 139.6 1.7Pernambuco Tropical semi-arid 26.2 32.0 20.5 570 136.2 4.8Piauí Tropical hot subhumid 27.5 32.9 22.1 1449 286.3 11.6São Paulo South-east region Subtropical cool 21.3 25.1 17.6 1623 127.1 40.0Minas Gerais Tropical mesothermic 26.5 35.0 18.0 840 105.2 6.0Paraná South region Subtropical very cool 17.6 22.7 12.4 1408 165.0 74.5
Mexicoc Sinaloa Northern and Central Pacifi c region
Warm subhumid tropics
24.7 27.5 (22–38) 22.0 (13–29) 922 799 123
Nayarit Warm subhumid tropics 24.7 26.3 (24–38) 23.1 (13–30) 1396 1346 50Colima Warm semi-arid tropics 26.5 27.6 (25–38) 25.4 (14–30) 660 605 55Michoacán Very warm semi-arid
tropics28.3 29.6 (28–40) 27.0 (18–32) 709 601 108
Guerrero Warm subhumid tropics 28.0 29.0 (28–40) 26.9 (18–32) 1011 910 101Oaxaca Southern Pacifi c
regionIsothermal warm humid tropics
25.9 26.9 (26–35) 25.0 (18–28) 1779 1731 48Chiapas 24.7 24.9 (26–35) 24.4 (18–28) 3732 3661 71Central Veracruz
Gulf of Mexico region
Isothermal warm subhumid tropics
24.8 26.7 (26–38) 22.9 (14–30) 1026 898 128
Southern Veracruz
Isothermal warm humid tropics
25.5 27.6 (26–38) 23.4 (14–30) 2093 1807 286
Taiwand Tainan Central-South Subtropical 23.6 26.8 (23.9–28.3) 19.1 (17.1–20.4) 1733 264 (71–415) 25 (15–41)Pingtung South Subtropical 24.7 26.8 (24.9–27.9) 21.8 (20.4–22.7) 2158 329 (170–511) 30 (19–47)Average Whole island Subtropical 25.0 25.8 (23.1–27.5) 19.1 (17.3–21.6) 1885 273 (92–392) 42 (24–60)
(Continued)
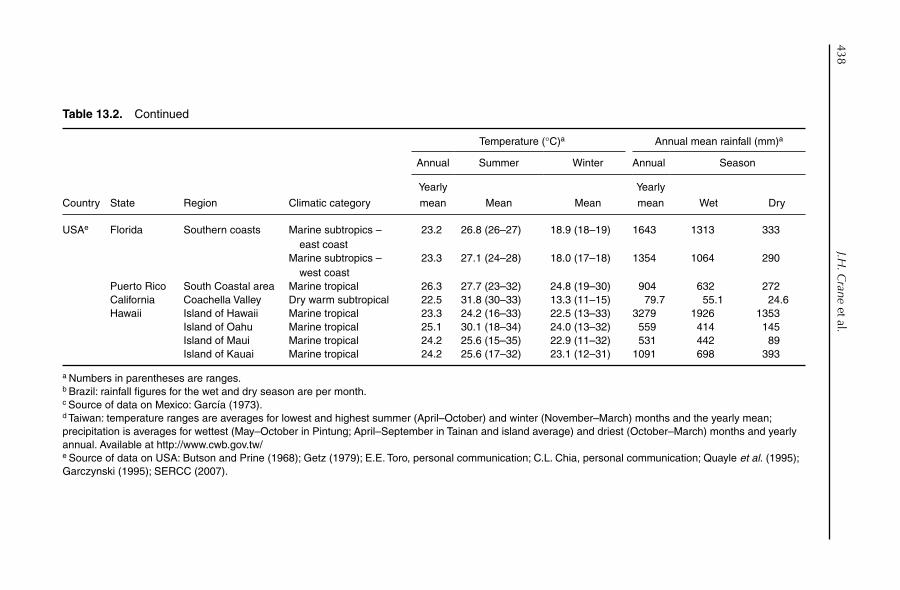
J.H
. Crane et al.
438
Table 13.2. Continued
Country State Region Climatic category
Temperature (°C)a Annual mean rainfall (mm)a
Annual Summer Winter Annual Season
Yearly
mean Mean Mean
Yearly
mean Wet Dry
USAe Florida Southern coasts Marine subtropics – east coast
23.2 26.8 (26–27) 18.9 (18–19) 1643 1313 333
Marine subtropics – west coast
23.3 27.1 (24–28) 18.0 (17–18) 1354 1064 290
Puerto Rico South Coastal area Marine tropical 26.3 27.7 (23–32) 24.8 (19–30) 904 632 272California Coachella Valley Dry warm subtropical 22.5 31.8 (30–33) 13.3 (11–15) 79.7 55.1 24.6Hawaii Island of Hawaii Marine tropical 23.3 24.2 (16–33) 22.5 (13–33) 3279 1926 1353
Island of Oahu Marine tropical 25.1 30.1 (18–34) 24.0 (13–32) 559 414 145Island of Maui Marine tropical 24.2 25.6 (15–35) 22.9 (11–32) 531 442 89Island of Kauai Marine tropical 24.2 25.6 (17–32) 23.1 (12–31) 1091 698 393
a Numbers in parentheses are ranges.b Brazil: rainfall fi gures for the wet and dry season are per month.c Source of data on Mexico: García (1973).d Taiwan: temperature ranges are averages for lowest and highest summer (April–October) and winter (November–March) months and the yearly mean; precipitation is averages for wettest (May–October in Pintung; April–September in Tainan and island average) and driest (October–March) months and yearly annual. Available at http://www.cwb.gov.tw/e Source of data on USA: Butson and Prine (1968); Getz (1979); E.E. Toro, personal communication; C.L. Chia, personal communication; Quayle et al. (1995); Garczynski (1995); SERCC (2007).

Crop Production: Management 439
Some cultivars, for example ‘Haden’ in north-eastern Brazil, have poor fruit set when temperatures are >35°C.
Solar radiation is very important for fruit development, since its dura-tion and intensity are directly related to photosynthesis and carbohydrate production (Mukherjee, 1953). Light incidence depends on the season (Allen et al., 1998). According to Lima Filho (2000), mango production in the semi-arid region of Brazil is where maximum solar radiation occurs in summer (October southern hemisphere; 528 cal/cm2/day) and the minimum in win-ter (June southern hemisphere; 363 cal/cm2/day), which corresponds to the fl owering and fruit development stages, respectively.
Mango production in Mexico is in the tropics. Most rainfall occurs dur-ing the summer and may be accompanied by hurricanes (Table 13.2). Climate in the Northern and Central Pacifi c region (17–27°N) ranges from warm sub-humid to warm/very warm semi-arid tropics with a 7-month dry season (García, 1973). In Colima and Michoacán, mangoes are irrigated due to low annual precipitation (semi-arid condition with a 6- to 8-month dry season; the mean annual temperature is 26.5–28.3°C). The Southern Pacifi c and Gulf of Mexico coastal regions have isothermal (< 5°C monthly temperature oscil-lation) warm, subhumid to humid climates with a 6–8-month dry season (García, 1973).
The Gulf of Mexico coastal region is affected by winds from the north (11–28 m/s) from October to April. These winds cause direct damage, such as limbs breaking and fl ower and fruit drop, and also increase plant respiration and transpiration rates that stress the trees. In the Pacifi c coastal regions hur-ricanes usually occur from August to October. Recent, unusually warm win-ters have resulted in undesirable late autumn or winter vegetative fl ushes and poor fl owering. Normal bloom occurs in late January and February; how-ever, late autumn or winter shoots delay anthesis during hot spring tempera-tures (April to early May) which result either in low fruit set or parthenocarpic fruit.
The primary production areas in Taiwan are from 22°N (Fungshan, Ping-tung) to 24°N (Nanto and Taichung prefectures). The average temperature of these areas is c.23°C, with a mean of 20°C during fl owering (December–March) and c.18.6°C during development (April–August) (Table 13.2). Flower induction in mango is not a problem in Taiwan because of its subtropical climate; however, fruit set can be affected by low temperature, rainfall, etc. A monsoon-type climate prevails in southern Taiwan with precipitation occurring mostly from May through to September. There is little rainfall (usually <50 mm) during fl ower bud formation and fl owering (autumn-winter) (Table 13.2).
The production areas of the USA have different climates. South Florida has a marine subtropical climate (Table 13.2). South-east Florida has a mean annual temperature of 23°C, a mean annual rainfall of 1643 mm and 62% relative humidity (RH) (Butson and Prine, 1968; Getz, 1979; Barrick and Black, 1980). Constant ocean-spawned winds of 4–9 km/h buffet the region from February through to October. The wet season (two-thirds annual rain-fall) occurs during the late spring into autumn (May–October, northern

J.H. Crane et al.440
hemisphere) and the dry season occurs during winter and early spring (November–April). Lowest temperatures (-4 to -6°C) occur from December through to February with a 70% probability of 0°C at least once each year (Bradley, 1975).
The Puerto Rican industry is mostly along the south and south-west coast at elevations between sea level and 50 m (Table 13.2) (Aponte-Morán et al., 1977). The La Cordillera central mountain range divides the island from north to south and has a major impact on the island’s climate, with annual rainfall of 1550 mm in the north and 904 mm in the south (Espenshade, 1992; SERCC, 2007). In the main production areas, the dry season is from Decem-ber to May, although July and August are also dry (Aponte-Morán et al., 1977). Mean maximum and minimum temperatures vary with altitude; how-ever, along the coastal production areas, 24–29°C is normal (Espenshade, 1992; SERCC, 2007). The lowest temperatures (18–20°C) occur along the coast during January and February.
The Hawaii mango industry is located mostly on the leeward coasts of Oahu, Hawaii and Maui islands; however, limited production occurs on the windward side of Kauai Island (Table 13.2). Each commercial planting has a distinct climatological niche that varies with altitude and location with respect to mountains and predominant north-east trade winds. Annual rain-fall is 531 mm on Maui, 559 mm on Oahu and 3279 mm near Hilo (east coast of Hawaii) (Table 13.2; C.L. Chia, personal communication). Temperatures, like rainfall, vary with elevation and location but generally range between 21 and 27°C (Bahr and Johnston, 1993b). Lower (13°C) and higher temperatures (32°C) occur occasionally.
The Coachela Valley of California is in the Salton Trough Desert area (Bahr and Johnston, 1993a) (Table 13.2). The valley ranges from 80 m below sea level to 488 m above sea level (Espenshade, 1992; Aslan et al., 1993) and the climate is arid. Temperatures in the valley vary considerably depending upon season, elevation and exposure. Some data reported herein represent the average for the area and may not accurately refl ect the particular micro-climate of the mango orchards in the valley (Garczynski, 1995). The mean annual temperature is 22.5°C; the average summer temperature is 31.8°C, and the average winter temperature is 13.3°C (Aslan et al., 1991; Garczynski, 1995). Winter temperatures range from 22.1°C to 4.5°C (Garczynski, 1995); however, Schacht (1992) reported a low of -4.4°C for 2 h. Summer tempera-ture extremes range from a low of 11°C to 50°C (Garczynski, 1995). The aver-age annual rainfall is 79.8 mm with two-thirds of this occurring during the winter (December–March) (Aslan et al., 1991; Garczynski, 1995). The valley is buffeted by wind and sandstorms during late spring, and these can cause severe damage (Schacht, 1992; Aslan et al., 1993).
13.4 Soils and Soil Preparation
Mangoes tolerate many soil types (Majumder and Sharma, 1990; Crane and Campbell, 1991; Kostermans and Bompard, 1993). Trees grow most vigorously

Crop Production: Management 441
in deep, fertile, moderately acid to neutral pH, loam-type soils. They tolerate infertile sands, volcanic ash and limestone-based soils, excessively drained and or periodically fl ooded soils and soils with acid (pH 4.5–7) to alkaline pH (pH 7–8.5). Mangoes are sensitive to saline and sodic soil conditions and proper irrigation practices and the use of salt-tolerant rootstocks is imperative for successful crop production in some areas (Schaffer et al., 1994).
Land preparation includes clearing virgin or existing orchard trees, disk-ing, destruction of subsurface hardpans (slip ploughing), mixing of upper and lower soil profi les, crushing superfi cial bedrock and mixing rock and soil layers, formation of land contours to facilitate drainage and/or fl ood irriga-tion, bedding and amending soils with organic matter and inorganic chemi-cals, for example hydrated lime (Ca(OH)2), dolomite and preplant fertilizers.
Mango is grown on a wide range of soil types, from latossols with a high percentage of sand in the north-east of Brazil to loamy oxysols in the south-east. Some areas in the north-east have shallow soils that need improved drainage. Soils of the central region are chemically poor and acid (pH 3.7–4.7) and require lime and/or gypsum application prior to orchard establishment. Soil analysis is used for determining the suitability of soil for production and potential fruit quality, especially in areas cultivated for export fruit. Typically soil analysis is conducted prior to land clearing and ploughing to determine nutrient levels and the need for lime and/or gypsum application. Soil sam-ples are taken from 0–20 cm and 20–40 cm depth of the soil profi le. Soil sam-ples are usually taken at a 0–30 cm and 30–60 cm soil depth for mature and established orchards. Ploughing, harrowing and lime and/or gypsum incor-poration are recommended at 30 cm depth and at least 30 days prior to rainy weather (Pinto and Ramos, 1998). Liming is very important for acid soils (<pH 4.0) in the central region of Brazil in order to increase soil pH to 6.0–6.5. It also improves the soil base saturation to 60–70% and results in better soil conditions for growth and production (Pinto, 2000). The amount of lime to be applied is based on a basis saturation equation (where hydrogen (H), aluminium (Al), calcium (Ca), magnesium (Mg) and potassium (K)):
Lime rate (t/ha): (T × 0.5) – S where T = (H + Al) + S and S = Ca + Mg + K
Gypsum applications are recommended for acid subsoils with >20% Al saturation and <0.5 cmol/dm3 Ca at soil depth of 60 cm (Andrade, 2004). This has been shown to signifi cantly reduce fruit physiological disorders, such as soft nose.
Production areas of Mexico vary in soil characteristics, and include fl at coastal areas and steep mountain slopes from sea level to >600 m above sea level. Orchards on sloped land commonly utilize individual tree terraces. Soil depth ranges from 30 cm to >3 m (alluvial soils). Shallow and eroded soils are common on hilly terrain. The presence of medium to large (10–50 kg) boulders can impede the use of machinery but they reduce erosion and increase soil-water storage capacity. In general, poor soil drainage is not a problem; however, areas with clay soils have drainage problems during peri-ods of heavy rain, especially if there is a shallow water table. A range of soil types is used for mango production in Mexico. In the Northern Pacifi c region,

J.H. Crane et al.442
mangoes are planted in cambisols (pH 6.0–7.0), luvisols (pH 6.5–7.5) and feozem (pH 5.0–6.0) soils with loamy textures (Anonymous, 1970, 1982). These soils are well drained, with 2–3% organic matter, moderate to high water holding capacity and cation exchange capacity (CEC) of 10–20+ meq/100 g soil. In the Central Pacifi c region mangoes are planted on cambi-sols, feozem and regosols with light textures (Anonymous, 1970, 1982). The regosols (pH 6.5–7.5) are well drained, with <2% organic matter content, low water holding capacity and CEC <10 meq/100 g soil. Soils planted to man-goes in the Southern Pacifi c region include nitosols, luvisols and feozem (Anonymous, 1970, 1982). The nitosols (pH 5.0–6.0) are well drained, with >3% organic matter content, low to moderate water holding capacity and low CEC (<10–20 meq/100 g soil). Soils in the Gulf of Mexico region include fl uvisols, cambisols and vertisols of loamy and clayey textures (Anonymous, 1970, 1982). The fl uvisols and vertisols are moderately and slowly drained soils, respectively. Soil pH for fl uvisols ranges from 5.5 to 7.5 with <2% organic matter content. Soil pH for the vertisols is alkaline (7.5–8.0) and the organic matter content is moderate (2.1–3%). The water holding capacity and CEC is low to moderate for the fl uvisols (<10–20 meq/100 g soil) and high for the vertisols (10–20+ meq/100 g soil).
Soil tillage is used in fl at lands before planting. Dimensions of planting holes range from 40–60 cm depth and 30–50 cm diameter in light-textured soils. Planting holes can be larger in heavy-textured or stony soils. On steep hills only planting holes are made and individual terraces are built up to hold soil, water, organic matter, fertilizers or soil amendments. Preplant soil analyses are rarely taken; however, chemical or organic fertilizers are com-monly applied at planting time. With rapid expansion of the mango industry, orchards have been planted in shallow soils (0.75–1.2 m depth) underlain with a hardpan. These soils are poorly drained and root growth and exten-sion are limited. Trees growing in such soils are prone to drought stress dur-ing dry periods, fl ooding stress after rains and nutritional defi ciencies. The weakened trees appear to be more susceptible to pathogens, for example Botryodiplodia theobromae, which causes cankers or stem dieback and eventu-ally tree death (Ponce-González and Salazar-García, 1992). Cultural practices are not available to ameliorate these problems and planting in such soils is not recommended.
Mangoes are grown in Taiwan on sandy loam, loamy sands, clay and coarse sandy soils. Soil pH ranges from 5.0 to >7.8. Most trees are grown on sloping land. Silt clay loam with pH 5.0–6.0 is mostly found in the Pingtung area, while the Tainan area has clay soils with pH 7.3–7.8. Acid soils are amended with Ca(OH)2 or dolomite and alkaline soils are amended with sulfur (S) or acid-based fertilizers.
The topography in the mango-production areas of Florida is fl at and ranges from sea level to c.6.1 m above sea level. Soils in Florida include vari-ous sands, muck and oolitic limestone. In the main production area (Miami-Dade County), mangoes are planted in extensively scarifi ed (crushed) and trenched oolitic limestone rock (Krome and Chekika very gravelly loam) (Colburn and Goldweber, 1961; Noble et al., 1996). The limestone soil is very

Crop Production: Management 443
permeable (1.5–5.1 cm/h), with high pH (7.4–8.4), low organic matter con-tent (3–10%), low water holding capacity (0.2–0.3 cm/cm of soil) and low CEC (16.0–37.0 meq/100 g soil) (Calhoun et al., 1974; Anonymous, 1989). In areas subject to fl ooding, crushed rock is formed into beds (0.6–1.0 m high and 1.0–1.5 m wide) before planting. The sandy soils in other Florida produc-tion areas are poorly to well drained with or without an undulation hard-pan 0.5–3.0 m down in the soil profi le (Henderson et al., 1984). The highly organic muck soils in Palm Beach County are poorly drained and underlain by dense limestone bedrock. Beds of varying dimensions are made in the sandy and muck areas to increase the proportion of the root system above fl ood levels. The sand and muck soils are characterized by an acid to alka-line pH (3.6–8.4) and low cation exchange capacities (Carlisle et al., 1978; Henderson et al., 1984).
In Puerto Rico, mangoes are grown on fl at and gently sloping land con-sisting of alluvial fans and terraces level with or slightly above the fl ood plain (Bierbolini et al., 1979). Some orchards in western Puerto Rico are located on moderately steep to very steep slopes (12–60% slope) and rounded hill tops that are somewhat eroded and superfi cial (Bierbolini et al., 1975). Soils in southern Puerto Rico are very deep, moderately well to well drained and consist of fi ne-textured sediment of sandy and clayey loam over gravelly fi ne-textured sediment. The pH ranges from moderately acid (pH 6) to alka-line (pH 8 or above). Soils have good to very good native fertility and good water holding capacity. Soils in the west are well drained and moderately acid (pH 5.0–6.0).
Mango production in Hawaii occurs on volcanic ash soils varying from recent to highly weathered (R. Yost, personal communication). They tend to be well drained; some soils must be slip ploughed to break up the hardpans in preparation for planting. The pH ranges from 5 to 8 and the fertility varies substantially. Mangoes in California are on lacustrine deposits consisting of fi ne-textured sediments that are highly stratifi ed sandy loam soils with clay lenses (S. Aslan, personal communication). The soil is alkaline (pH 7–8.4) and calcareous (Aslan et al., 1991). Land is slip ploughed to 1.5–1.8 m depth to break up and mix the stratifi ed soil layers.
13.5 Plant Propagation and Rootstocks
Mango production areas rely on traditional propagation, i.e. seedage, graft-ing and budding. Marcottage (air layering) methods have been tried for many years (Rajan and Ram, 1988; Majumder and Sharma, 1990) and although recently improved (Núñez-Elisea et al., 1989, 1991, 1992; Lambe et al., 1991; Núñez-Elisea and Davenport, 1995), it is still not used commercially. In vitro techniques hold great promise for rootstock propagation and cultivar improvement, and are currently being perfected and investigated (Litz, 1984, 1986, 2005; Litz et al., 1984; see Litz et al., Chapter 18, this volume).
The monoembryonic Florida cultivars in Brazil are vegetatively propa-gated by grafting and budding on polyembryonic rootstocks. Rootstock and

J.H. Crane et al.444
grafted mangoes are established in nurseries by sowing seeds in a soil sub-strate consisting of a mixture of organic matter and semi-sterilized soil plus 3 kg of superphosphate (3Ca(H2PO4)2) and 0.5 kg of potassium nitrate (KNO3) for each m3 of mixture (Pinto et al., 1981; Castro Neto et al., 2002). In the south-east, the indirect seeding technique is used: seed is sown in the soil to germi-nate and then transplanted to small softwood containers or ‘jacás’. Subsequently, seedlings are patch budded. In the central and north-east regions, seedlings are grown in substrate in plastic bags and subsequently whip or veneer grafted. The endocarp is removed prior to sowing to accelerate germination and improve uniformity of rootstock growth (Pinto and Genú, 1981).
The most important characteristics for mango rootstocks are vigour, high yield potential, compatibility with the scion, environmental adaptability and resistance to pests and diseases (Rosetto et al., 1996; Nascimento et al., 2002). Polyembryonic cultivars, that is ‘Espada’ and ‘Fiapo’ in the north-east and ‘Rosinha’ and ‘Coquinho’ in the south-east, are the most important root-stocks. The Agencia Paulista de Tecnologia Agropecuária (APTA) has developed root-rot-resistant rootstocks, including IAC-101, IAC-102 and IAC-106; however, they are not widely used.
There are no specifi c mango rootstocks in Mexico. Before the mid-20th century, seedling plants were used; however, uniform plantings did exist because polyembryonic ‘Manila’ was grown in the Gulf of Mexico region. Elsewhere, there were monoembryonic trees, resulting in a heterogeneous mix of trees. Since the late 1950s and early 1960s, polyembryonic criollo root-stocks have been used for rootstocks; however, due to their vigour, mono-embryonic rootstocks are still in use in some areas. Commercial production is done in partially shaded nurseries. A seedbed is prepared using sterilized substrate (loamy soil, composted organic matter or a mixture). The fi brous seedcoat is removed to expose the smooth, bean-shaped seed, which is soaked for about 10–30 min in 50 mg/l gibberellic acid (GA3) to reduce germination time. Seeds are placed on the seedbed (usually in May–July) with their hump up and watered every other day. After 3–5 weeks, vigorous and healthy seed-lings are transferred to black plastic bags 20 cm wide × 40 cm long fi lled with a sterilized mixture of sandy loam soil and organic matter (3:1). Rootstocks are veneer grafted when they are 4–6 months old. Grafted plants are ready for planting by May–July of the next year, during the rainy summer season.
Some experienced mango growers initially plant seedling trees in the orchard to ensure a vigorous tree. Grafting is then performed 3–6 months after planting. In some cases a low vigour interstock like ‘Esmeralda’ is top-worked onto the seedling rootstock and then grafted to the desired scion. The use of ‘Esmeralda’ interstocks has reduced tree canopy size by 28% and 33% for ‘Manila’ and ‘Ataulfo’, respectively (Mosqueda-Vázquez et al., 1996; Vázquez-Valdivia et al., 2000).
Mangoes in Taiwan are generally splice or veneer grafted; however, a combination of the two methods is used when the rootstock and scion are of different diameters. Polyembryonic ‘Tsar-swain’ seedlings are pre-ferred as clonal rootstocks although seedlings of monoembryonic ‘Irwin’ are sometimes used.

Crop Production: Management 445
The main commercial cultivars in the USA, ‘Keitt’, ‘Tommy Atkins’, ‘Van Dyke’, ‘Haden’ and ‘Brooks’, are monoembryonic. Orchards are planted with grafted or budded trees (Aponte-Morán et al., 1977; Chia et al., 1988; Crane and Campbell, 1991; Hamilton et al., 1992). The most common rootstocks in Florida are seedlings of polyembryonic ‘Turpentine’, ‘Number 11’ and ‘ Criollo’. In Hawaii, any mango seedlings are used. In California, ‘Turpen-tine’ or ‘Criollo’ are used. In Puerto Rico, polyembryonic ‘Mangotino’ and ‘Pasote’ seedlings are used in the Ponce region and ‘Mayaguezano’, ‘Turpen-tine’ and ‘Colombo Kidney’ in the Mayaguez region (Aponte-Morán et al., 1977; Toro, 1988; E.E. Toro, personal communication). Regardless of the seed source, roguing the zygotic seedlings is important for obtaining uniform tree size, growth characteristics and production (Schnell and Knight, 1991, 1992; Degani et al., 1993; Schnell et al., 1994). Seed is removed from mature fruit along with the husk, and the seed is planted in well-drained container media (Sauls and Campbell, 1980; Young and Sauls, 1989). Seedlings are large enough to graft or bud after 2–6 months. The most common vegetative prop-agation methods in Florida are veneer and cleft grafting (Sauls and Camp-bell, 1980; Crane and Campbell, 1991) and chip and ‘T’ or inverted ‘T’ budding. The trees are usually ready for fi eld planting in 6–12 months. Graft-ing may be done at any time of year when suitable rootstocks are available, but it is more successful during warm weather. In Puerto Rico the most common propagation method is cleft grafting (Aponte-Morán et al., 1977; Toro, 1988). Grafting is most successful during the spring and from September through to November.
Topworking of established trees is common. Trees are cut back to several major limbs or the trunk. In Florida, several sprouts emerging from the pruned limbs are allowed to develop to 1.3–7.6 cm diameter, and are veneer grafted with the new cultivar (Sauls and Campbell, 1980). In Puerto Rico, several sprouts are allowed to develop, and larger sprouts (2.5–7.6 cm diam-eter) are cleft grafted and smaller diameter sprouts (1.0–1.5 cm) are bark grafted (Toro, 1988).
13.6 Major Cultivars
For detailed information about cultivars, refer to Knight et al., Chapter 3, this volume.
Mango seeds were introduced by Portuguese and Spanish colonizers to Brazil during the 16th century. Subsequently, mango seeds were dispersed into the interior of the country. There are >120 local mango cultivars, many of which are the result of open pollinations among Indian and Philippine mango germplasm. There is a large amount of variability among the cultivars with respect to colour, shape and size of the fruit. Most of these cultivars are for the fresh fruit market (e.g. ‘Espada’, ‘Bourbon’, ‘Ubá’, ‘Rosa’, ‘Coité’, ‘Coquinho’ and ‘Lira’), or for juice, jellies and other value-added products (e.g. ‘Ubá’, ‘Coité’, ‘Mamão’) (Table 13.3). Some polyembryonic criollo culti-vars (e.g. ‘Espada’, ‘Coquinho’ ‘Comum’ and ‘Fiapo’) are used as rootstocks.
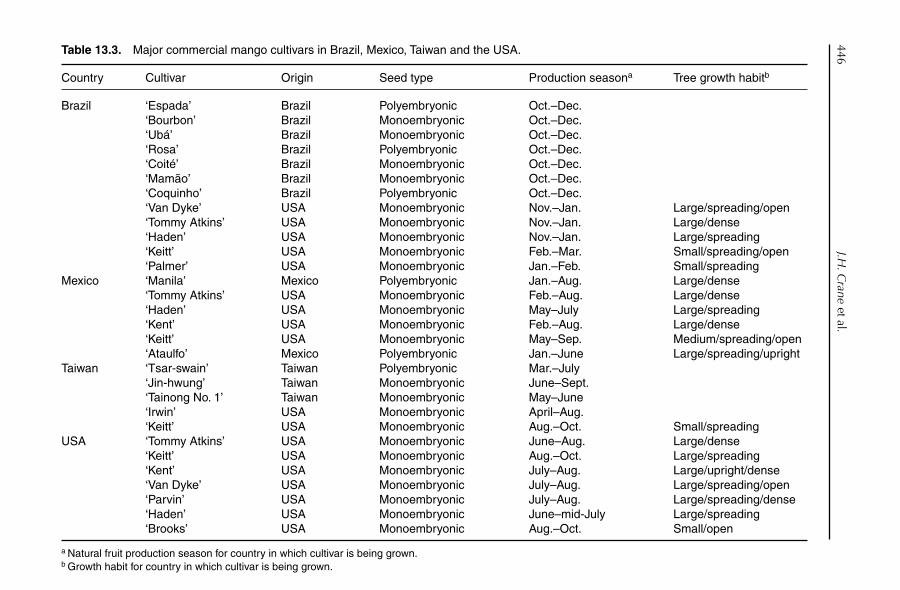
J.H
. Crane et al.
446Table 13.3. Major commercial mango cultivars in Brazil, Mexico, Taiwan and the USA.
Country Cultivar Origin Seed type Production seasona Tree growth habitb
Brazil ‘Espada’ Brazil Polyembryonic Oct.–Dec.‘Bourbon’ Brazil Monoembryonic Oct.–Dec.‘Ubá’ Brazil Monoembryonic Oct.–Dec.‘Rosa’ Brazil Polyembryonic Oct.–Dec.‘Coité’ Brazil Monoembryonic Oct.–Dec.‘Mamão’ Brazil Monoembryonic Oct.–Dec.‘Coquinho’ Brazil Polyembryonic Oct.–Dec.‘Van Dyke’ USA Monoembryonic Nov.–Jan. Large/spreading/open‘Tommy Atkins’ USA Monoembryonic Nov.–Jan. Large/dense‘Haden’ USA Monoembryonic Nov.–Jan. Large/spreading‘Keitt’ USA Monoembryonic Feb.–Mar. Small/spreading/open‘Palmer’ USA Monoembryonic Jan.–Feb. Small/spreading
Mexico ‘Manila’ Mexico Polyembryonic Jan.–Aug. Large/dense‘Tommy Atkins’ USA Monoembryonic Feb.–Aug. Large/dense‘Haden’ USA Monoembryonic May–July Large/spreading‘Kent’ USA Monoembryonic Feb.–Aug. Large/dense‘Keitt’ USA Monoembryonic May–Sep. Medium/spreading/open‘Ataulfo’ Mexico Polyembryonic Jan.–June Large/spreading/upright
Taiwan ‘Tsar-swain’ Taiwan Polyembryonic Mar.–July‘Jin-hwung’ Taiwan Monoembryonic June–Sept.‘Tainong No. 1’ Taiwan Monoembryonic May–June‘Irwin’ USA Monoembryonic April–Aug.‘Keitt’ USA Monoembryonic Aug.–Oct. Small/spreading
USA ‘Tommy Atkins’ USA Monoembryonic June–Aug. Large/dense‘Keitt’ USA Monoembryonic Aug.–Oct. Large/spreading‘Kent’ USA Monoembryonic July–Aug. Large/upright/dense‘Van Dyke’ USA Monoembryonic July–Aug. Large/spreading/open‘Parvin’ USA Monoembryonic July–Aug. Large/spreading/dense‘Haden’ USA Monoembryonic June–mid-July Large/spreading‘Brooks’ USA Monoembryonic Aug.–Oct. Small/open
a Natural fruit production season for country in which cultivar is being grown.b Growth habit for country in which cultivar is being grown.

Crop Production: Management 447
Florida selections (i.e. ‘Haden’, ‘Keitt’, ‘Tommy Atkins’, ‘Van Dyke’ and ‘Palmer’) are grown for the export trade. Most Florida cultivars produce from Decem-ber to January except ‘Palmer’, which is harvested between January and Feb-ruary and ‘Keitt’, which is harvested between February and March. ‘Tommy Atkins’ represents 80% of the commercial export volume in Brazil (Pinto, 2004); ‘Palmer’ is increasing in demand in the national market.
Mango production in Mexico relies on the following cultivars (from early to late beginning of harvest season): ‘Manila’, ‘Ataulfo’, ‘Kent’, ‘Haden’, ‘Tommy Atkins’ and ‘Keitt’ (Table 13.3). ‘Zill’, ‘Irwin’, ‘Sensation’, ‘Diplomático’, ‘Manililla’, ‘Oro’, ‘Ovo’ and criollos are also grown. Mango harvest seasons are no longer infl exible as they may be modifi ed by pruning, water manage-ment and growth regulators. The most widely planted cultivar in Mexico is ‘Manila’ (45,396 ha); no export fi gures have been reported. Although ‘Manila’ is grown all over the country it dominates in Veracruz (24,908 ha) and Guerrero (9198 ha). It is an alternate bearer and the harvest season is from May to June in Veracruz and July in Sinaloa (De los Santos and Mosqueda-Vázquez, 1988–89; C. Guzmán, personal communication). There are 22,890 ha of ‘Tommy Atkins’ and it is the major export cultivar to the USA (33% of total exports). Fruit is harvested from February (Michoacán) to August (Colima, Jalisco and Sinaloa) (V. Medina and C. Guzmán, personal commu-nications). ‘Kent’ is cultivated on 13,366 ha and accounts for 23% of exports to the USA. Sinaloa is the major producer of ‘Kent’ (9710 ha) and the begin-ning of harvest season is similar to ‘Tommy Atkins’ (Campbell, 1992); however, it can be harvested as late as September in Colima, Jalisco and Nayarit.
‘Ataulfo’ is from Chiapas and is now cultivated nationwide on >34,000 ha. The major producing states are Chiapas (south) and Nayarit (north) with 18,334 ha and 7156 ha, respectively. The harvest season is February–May in Chiapas and June–July in Sinaloa (V. Palacio and C. Guzmán, personal com-munications). In some years this cultivar is the third most important export to the USA, and accounted for 19% of exports in 2006 (EMEX, 2007). ‘Keitt’ (6828 ha) is the fourth most important export cultivar (14% exports). Most ‘Keitt’ orchards are located in Sinaloa (5198 ha). ‘Keitt’ is harvested from May (Nayarit) to September (Colima, Jalisco and Sinaloa). Large fruit size and fungal lesions sometimes cause marketing problems because harvest occurs during the rainy season. ‘Haden’ (27,768 ha) is still an important mango although exports to the USA have decreased to 11% (EMEX, 2007); the major producing states are Michoacán (15,573 ha), Guerrero (5053 ha) and Sinaloa (3518 ha). Fruit are harvested from February/March (Michoacán and Guerrero) to August/September in Colima, Jalisco and Michoacán. Alternate bearing and mango malformation are problems and ‘Haden’ orchards are being replaced by or topworked to cultivars like ‘Ataulfo’.
The mango was introduced into Taiwan from South-east Asia by the Dutch in the 16th century (Chen, 1991). The introductions were all polyem-bryonic and included ‘Tsar-swain’, ‘Pung-swain’, ‘Va-swain’ and ‘Kee-gway-swain’. ‘Tsar-swain’ is still very important (Table 13.3). From 1954 to 1973, 35 monoembryonic cultivars were introduced from the USA, and ‘Irwin’ and ‘Keitt’ have been major cultivars in Taiwan since 1960. ‘Jin-hwung’, which is

J.H. Crane et al.448
derived from a chance seedling of ‘White’ and ‘Keitt’, was selected in 1980. ‘Tainong No. 1’ and ‘Tainong No. 2’ are derived from controlled pollinations and were released in 1985 by the Fengshan Tropical Horticultural Experiment Station, Taiwan Agricultural Research Institute. Only ‘Tsar-swain’, ‘Jin-hwung’, ‘Irwin’ and ‘Keitt’ and ‘Tainong No. 1’ are commercially important today.
Fruit in the southern prefectures or in lower elevation orchards are har-vested earlier than the northern prefectures or orchards at higher elevations. ‘Tsar-swain’ comprises about 40% of total production; its season is from March to August, depending on the prefecture or location of the orchard. The fruit of ‘Tsar-swain’ is 154 g, 100 mm long and 64 mm wide, total soluble solids (TSS) value is 17° Brix, total titratable acidity 2.2% and seed/pulp weight ratio 0.84 g. ‘Irwin’ comprises 40% of the production and is harvested from Pingtung in April and in Tainan in August. ‘Jin-hwung’ comprises about 10% of production and is harvested from June to September. The fruit of ‘Jin-hwung’ is 965 g, 144 mm long and 99 mm wide, TSS value is 15° Brix, total titrateable acidity 2.3% and seed/pulp weight ratio 0.94 g. ‘Keitt’ makes up c.5% of production and is harvested from August to October. The fruit of ‘Tainong No. 1’ is 237 g, 99 mm long and 69 mm wide, TSS value is 20° Brix, total titrateable acidity 4.0% and seed/pulp weight ratio 0.85 g.
The major cultivars in Florida are ‘Keitt’ and ‘Tommy Atkins’, which account for c.70% and 20% of the hectarage, respectively (Table 13.3). Small commercial hectarages of ‘Van Dyke’, ‘Palmer’, ‘Irwin’, ‘Raposa’ and ‘Kent’ are also grown. In Puerto Rico, the major export cultivars are ‘Keitt’, which makes up c.60% of the hectarage, ‘Parvin’ (20%), ‘Irwin’ (10%), ‘Tommy Atkins’ (5%) and ‘Haden’ (< 5%). Other cultivars (i.e. ‘Davis-Haden’, ‘Palmer’, ‘Kent’, ‘Mayaguezano’, ‘Poste’ and ‘Cubano’) are grown on a small scale. The local cultivars are grown for the domestic market (Toro, 1988). The commer-cial hectarage of California is mostly ‘Keitt’, although other cultivars have been evaluated (Scott, 1990; Linden, 2006).
Immature ‘Keitt’ is the major early season cultivar (picked green) and mature ‘Tommy Atkins’ is the major early season cultivar in Florida (Table 13.3) (J.H. Crane, personal communication). Mature ‘Keitt’ and ‘Kent’ mangoes are the major late season cultivars. Six- to 8-year-old ‘Tommy Atkins’ trees produce 75–150 kg/tree and older trees may produce up to 300 kg/tree. Internal breakdown varies from year to year and may be aggravated by over-fertilization with nitrogen (N). Fruit are considered moderately resistant to anthracnose. The harvest season is June through to August. ‘Keitt’ trees are precocious and produce large crops regularly during July through to September.
‘Palmer’, ‘Irwin’ and ‘Van Dyke’ are grown commercially on a limited scale in Florida and Puerto Rico (except ‘Van Dyke’). In Florida, ‘Palmer’ is harvested from July to early September, ‘Irwin’ from June to early July, and ‘Van Dyke’ from July to August. In the recent past, ‘Bailey’s Marvel’, ‘Brooks’ and ‘Haden’ were grown commercially; however, their importance has declined due to natural disasters and susceptibility to anthracnose. ‘Haden’ is no longer grown commercially in Florida (Crane and Campbell, 1991; Campbell, 1992) but, ‘Glenn’, a seedling of ‘Haden’, has been recommended

Crop Production: Management 449
(Campbell and Campbell, 1996). ‘Harders’ is grown in Manoa, Oahu Island, Hawaii (Hamilton et al., 1992; Hamilton, 1993). Trees bear regularly and often produce off-season good-quality fruit in the late autumn and winter. ‘Rapoza’, a seedling of ‘Irwin’, was selected in Hawaii in 1984 (Hamilton et al., 1992); its harvest season is from mid-July to October.
13.7 Plant Spacing
Plant spacing and density are infl uenced by climate, soil type and depth, rootstock and scion vigour, growth habit and the targeted tree size. Cultural practices including tree-size control, fertilizer and irrigation availability, methods, rates, timing and frequencies; current technology and the necessity for orchard access by farm machinery also infl uence plant spacing and con-fi guration. The cost of borrowed capital, land, water, irrigation, orchard maintenance and net returns will dictate what cultivars are grown and how they are managed (see Evans and Mendoza, Chapter 16, this volume).
Initial plant spacing and density should be planned to maximize early yield and returns from young orchards and to maintain high yields from mature orchards. Trees in overcrowded orchards compete for water, nutri-ents and light and eventually lose production in the lower canopy. The effi -cacy of foliar sprays (nutrients, pesticides and growth regulators) is reduced and harvest is more diffi cult in crowded orchards.
Plant spacing among trees and rows has decreased in recent years to optimize returns on investments in land, equipment and orchard infrastruc-ture. This has been possible because of more insight into the physiology of mango trees, improvements in irrigation system design and effi ciency, and better fertilizer delivery systems and pruning (i.e. intense hand and/or mechanical pruning) and the use of plant growth regulators. Closer plant spacings require more tree-size control and expertise on the part of the pro-ducer. Various training systems are advocated for new trees to force complex branching and increase bearing surface volume and fruit production poten-tial (Fig. 13.2). Topping and hedging and/or hand pruning (selective prun-ing and/or heading back) is used to maintain mature tree size and fruit production and improve fruit colour (Fig. 13.3).
In Brazil, the traditional spacing of 10 m × 10 m in a rectangular or qua-dratic format, with a density of 100 plants/ha, has been replaced by plant-ings of 8 m (between rows) × 5 m (within rows) to 5 m × 5 m with higher plant densities, which vary from 250 to 400 plants/ha (Cunha and Castro Neto, 2000; Mouco et al., 2002). In general, growers use two types of pruning: for-mation and production pruning (Albuquerque et al., 2002). Formation prun-ing is used to establish the intial tree architecture and trees are pruned fi ve times for several years leaving c.243 branches prior to starting mango pro-duction (Fig. 13.2). Pruning systems include cleaning, skirting of the lower canopy, lateral, central and top-canopy pruning, and pruning to correct poor canopy development and maintain adequate canopy after production commences (Albuquerque et al., 2002).
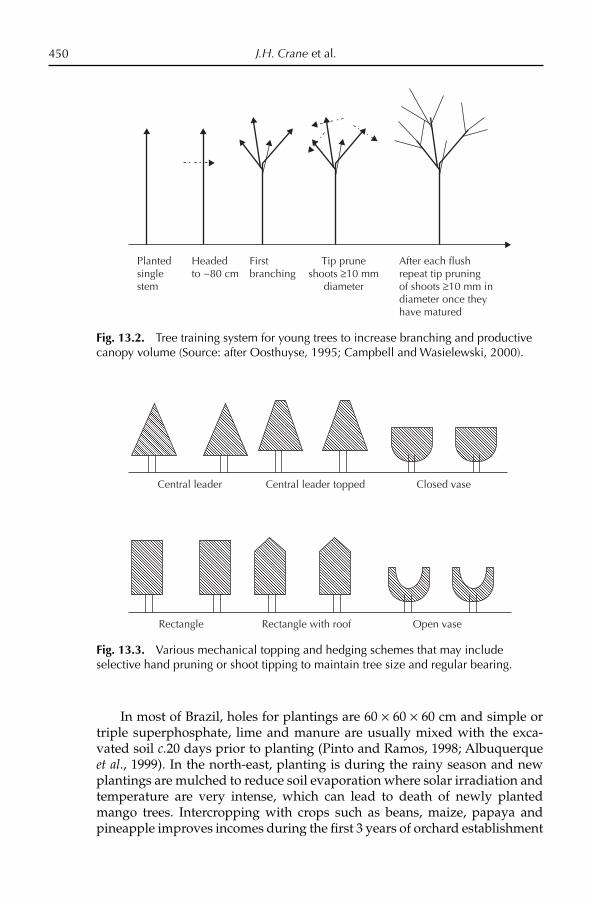
J.H. Crane et al.450
In most of Brazil, holes for plantings are 60 × 60 × 60 cm and simple or triple superphosphate, lime and manure are usually mixed with the exca-vated soil c.20 days prior to planting (Pinto and Ramos, 1998; Albuquerque et al., 1999). In the north-east, planting is during the rainy season and new plantings are mulched to reduce soil evaporation where solar irradiation and temperature are very intense, which can lead to death of newly planted mango trees. Intercropping with crops such as beans, maize, papaya and pineapple improves incomes during the fi rst 3 years of orchard establishment
Plantedsinglestem
Headedto ~80 cm
Firstbranching
Tip pruneshoots ≥10 mm
diameter
After each flushrepeat tip pruningof shoots ≥10 mm indiameter once theyhave matured
Fig. 13.2. Tree training system for young trees to increase branching and productive canopy volume (Source: after Oosthuyse, 1995; Campbell and Wasielewski, 2000).
Central leader
Rectangle Rectangle with roof Open vase
Central leader topped Closed vase
Fig. 13.3. Various mechanical topping and hedging schemes that may include selective hand pruning or shoot tipping to maintain tree size and regular bearing.

Crop Production: Management 451
(Mouco et al., 2002). Windbreaks, consisting of pine trees, elephant grass and three rows of banana plants, are often used during the fi rst 2 years of orchard establishment.
In Mexico, orchards were originally established at 10 m × 10 m to 16 m × 16 m, and trees were not pruned, which resulted in enormous and very pro-ductive trees; however, care and harvest from very large trees is problematic. Plant spacing is based on cultivar vigour (‘Ataulfo’ and ‘Manila’ are most vigorous), land slope (wider spaces as slope increases), farm machinery, cli-matic conditions and soil fertility (wider distances for warmer climates and more fertile soils) and water availability (Cruzaley-Sarabia et al., 2006). Irri-gated orchards may handle closer spacings if pruning is practised. Under rainfed conditions, soil moisture availability may have an important impact on tree size. For example, 10-year-old orchards may have a few big trees (c.69–100 trees/ha) or if trees are spaced more closely (300–600 trees/ha), tree size is reduced.
Currently, both square and rectangular planting patterns are common in Mexico although hexagonal planting systems are also used. For square designs, tree spacing ranges from 5 m (400 trees/ha) to 10 m (100 trees/ha). Rectangular planting systems have more options and the most common arrangements are 6 m × 5 m (333 trees/ha), 8 m × 5 m (250 trees/ha), 8 m × 6 m (205 trees/ha) and 10 m × 5 m (200 trees/ha) (Chávez-Contreras et al., 2001).
In Taiwan, plant spacing for monoembryonic cultivars (i.e. ‘Tsar-swain’) ranges from 6 m × 6 m to 10 m × 10 m (100–256 trees/ha). In contrast, plant spacing for polyembryonic cultivars ranges from 4 m × 5 m to 5 m × 6 m, depending on the topography and soil fertility. Orchards on sloped lands and infertile soils are less vigorous and are planted at higher densities than orchards on lowlands and fertile soils.
Recommended plant spacings in Hawaii and Florida are similar. The cooler climate of south-eastern California allows close spacing, i.e. 3 m in-row by 5.4 m between-rows (617 trees/ha). Traditional plant spacings in Florida were as high as 11 m in-rows and 11–14 m between-rows (64–82 trees/ha) (Young and Sauls, 1989). Plant spacings in Florida currently include 3.5 m × 6.1 m (468 trees/ha), 4.5 m × 6.1 m (364 trees/ha), 4.5 m × 7.6 m (292 trees/ha), 6.1 m × 6.1 m (268 trees/ha), 6.1 m × 7.6 m (215 trees/ha) and 7.6 m × 7.6 m (173 trees/ha) (Young and Sauls, 1989; Crane and Campbell, 1991). In Puerto Rico, plant spacings are 3.7 m × 5.5 m (491 trees/ha), 4.6 m × 9.1 m (238 trees/ha), 6.1 m × 9.1 m (180 trees/ha), 7.6 m × 7.6 m (173 trees/ha) and 9.1 m × 9.1 m (120 trees/ha) (Toro, 1988). Interplanting mango trees at moder-ate to wide plant spacings (i.e. 5.4–7.5 m in-row) with banana or plantain (Musa sp.), and papaya (Carica papaya L.) in Florida and plantains in Puerto Rico is widely practised. Mango trees have also been planted at close spacing (e.g. 3.0–4.5 m) and every other tree is removed if crowding becomes a prob-lem. Controlling tree size and maintaining crop productivity is important, otherwise, competition among trees will reduce yields and fruit quality (Toro, 1988; Crane and Campbell, 1991; Oosthuyse, 1995). Annual or biennial hand pruning and/or mechanical hedging and topping is necessary and should begin several years before trees begin to crowd and should continue after

J.H. Crane et al.452
trees grow to their desired size based on plant spacing. Mature trees are topped at 3.5–5 m, and hedged trees are cut on a slight angle (5–10°) to leave a 2.5–3.5 m row middle (J.H. Crane, personal communication). Trees can be maintained at in-row spacings of 2–3 m; however, this involves intense tree training and hand pruning, which most producers have been unwilling to adopt (Fig. 13.3) (Oothuyse, 1995; Oosthuyse and Jacobs, 1995; Stassen et al., 1999; Campbell and Wasielewski, 2000). Severe hedging is utilized to increase light penetration and re-establish inner productive canopy but this can result in little to no production in the following year. Some producers utilize a com-bination of mechanical pruning followed by selective pruning to open the inner canopy to light and air movement, improving fruit colour and reducing disease pressure.
13.8 Fertilizer Practices
Fertilizer practices are infl uenced by availability and cost of organic and inorganic materials, application costs, soil type and depth, irrigation prac-tices and rainfall, cultivar and production objectives. The response to fertil-izer is infl uenced by fertilizer source, rate, timing and method of application, tree-growth stage, climate, edaphic conditions, soil moisture status and cul-tivar vigour. The objective of fertilizer practices for young trees during years 1 and 2 is to establish the tree and maintain constant canopy growth and health while building a structurally strong tree. Objectives for bearing trees include maintenance of tree health, avoidance of alternate bearing and allow-ance for vegetative dormancy which results in fl ower induction and initiation.
Over-fertilization of bearing trees leads to continuous vegetative growth under some climatic conditions, reduced fl owering and fruit yields and increased occurrence of physiological disorders of the fruit (Schaffer et al., 1994; Coelho et al., 2002; Nguyen et al., 2004). Likewise, improper fertilization may lead to nutrient defi ciencies and/or toxicities which result in reduce tree growth, yields and fruit quality. Fertilizer practices will continue to be refi ned and developed as our understanding of the physiological needs and responses to essential plant nutrients increases and as concerns increase about the effect of excess and/or leached nutrients on the natural environment.
Fertilizer management is based on soil and leaf analysis in Brazil (Tables 13.4–13.6; Santos et al., 2002). The quantity and type of fertilizer application varies from the orchard establishment period (mostly vegetative plant growth) to the orchard production period. At the time of planting, 20–30 l/hole of cattle manure is usually mixed with the native soil. Subsequently, N, phosphorus (P) and potassium (K) are applied 2–3 times annually. During young tree establishment, P and K are recommended only if these elements are defi cient (Tables 13.5 and 13.6). These fertilizers are broadcast mechanically in the planting rows, and then incorporated into the top soil layer (Andrade, 2004; Sousa et al., 2004). The quantities of macronutrients applied at the time of planting and at various growth stages in south-eastern Brazil (mainly São Paulo) and the central regions are based on plant age and soil P and K content
Crop Production: Management 453
Table 13.4. Standard leaf nutrient content ranges for mature mango trees in Brazil, Mexico, Taiwan and Florida USA.
Mineral Unit
Range for mature treesa
Brazil Mexicob Taiwanc USAd
N % 1.2–1.4 1.2–1.5 1.4–1.7 1.0–1.5P % 0.08–0.16 0.07–0.13 0.1–0.15 0.09–0.18K % 0.5–1.6 0.6–0.7 0.9–1.2 0.5–1.0Ca % 2.0–3.5 2.3–3.4 1.0–1.8 3.0–5.0Mg % 0.25–0.5 0.14–0.19 0.2–0.35 0.15–0.47B ppm 50–100 NRe NR 24–54Fe ppm 50–200 97–114 60–120 38–120Mn ppm 50–100 191–802 30–200 92–182Zn ppm 20–40 14–20 20–100 101–119Cu ppm 10–50 5–12 5–20 28–35
a Recommended leaf nutrient levels based on research and the literature.b Source of data for Mexico: Guzmán-Estrada (2001, 2004, 2006). Range shown for all cultivars tested (see Table 13.8 for detail by cultivar). c Values are for ‘Irwin’ leaves. Source of data for Taiwan: Job (1989).d Source of data for USA: Young and Koo (1969, 1971); Young and Sauls (1989).e NR, not reported.
Table 13.5. Soil phosphate and potash corrective fertilization rates based on soil analysis in Brazil.(a) Rate of P2O5 (kg/ha) application.
Clay content (%)
Level of P availability in the soil
Low Medium Adequate
≤15 60 30 016–35 100 50 036–60 200 100 0>60 280 140 0
(b) Rate of K2O (kg/ha) application.
Soil K content (mg/dm3)
Level of K availability in the soil
Low Medium Adequate
Cation exchange capacity = <4.0 cmol/dm3 at pH 7 and <20% clay<15 50 – –16–40 – 25 –>40 – – 0Cation exchange capacity = >4.0 cmol/dm3 at pH 7 and >20% clay<25 100 – –25–80 – 50 –>80 – – 0
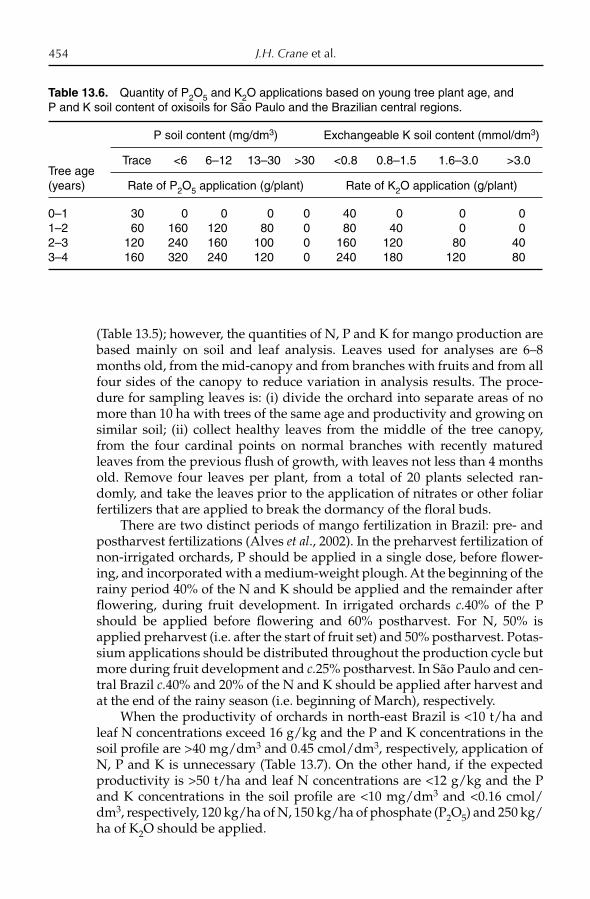
J.H. Crane et al.454
(Table 13.5); however, the quantities of N, P and K for mango production are based mainly on soil and leaf analysis. Leaves used for analyses are 6–8 months old, from the mid-canopy and from branches with fruits and from all four sides of the canopy to reduce variation in analysis results. The proce-dure for sampling leaves is: (i) divide the orchard into separate areas of no more than 10 ha with trees of the same age and productivity and growing on similar soil; (ii) collect healthy leaves from the middle of the tree canopy, from the four cardinal points on normal branches with recently matured leaves from the previous fl ush of growth, with leaves not less than 4 months old. Remove four leaves per plant, from a total of 20 plants selected ran-domly, and take the leaves prior to the application of nitrates or other foliar fertilizers that are applied to break the dormancy of the fl oral buds.
There are two distinct periods of mango fertilization in Brazil: pre- and postharvest fertilizations (Alves et al., 2002). In the preharvest fertilization of non-irrigated orchards, P should be applied in a single dose, before fl ower-ing, and incorporated with a medium-weight plough. At the beginning of the rainy period 40% of the N and K should be applied and the remainder after fl owering, during fruit development. In irrigated orchards c.40% of the P should be applied before fl owering and 60% postharvest. For N, 50% is applied preharvest (i.e. after the start of fruit set) and 50% postharvest. Potas-sium applications should be distributed throughout the production cycle but more during fruit development and c.25% postharvest. In São Paulo and cen-tral Brazil c.40% and 20% of the N and K should be applied after harvest and at the end of the rainy season (i.e. beginning of March), respectively.
When the productivity of orchards in north-east Brazil is <10 t/ha and leaf N concentrations exceed 16 g/kg and the P and K concentrations in the soil profi le are >40 mg/dm3 and 0.45 cmol/dm3, respectively, application of N, P and K is unnecessary (Table 13.7). On the other hand, if the expected productivity is >50 t/ha and leaf N concentrations are <12 g/kg and the P and K concentrations in the soil profi le are <10 mg/dm3 and <0.16 cmol/dm3, respectively, 120 kg/ha of N, 150 kg/ha of phosphate (P2O5) and 250 kg/ha of K2O should be applied.
Table 13.6. Quantity of P2O5 and K2O applications based on young tree plant age, and P and K soil content of oxisoils for São Paulo and the Brazilian central regions.
Tree age (years)
P soil content (mg/dm3) Exchangeable K soil content (mmol/dm3)
Trace <6 6–12 13–30 >30 <0.8 0.8–1.5 1.6–3.0 >3.0
Rate of P2O5 application (g/plant) Rate of K2O application (g/plant)
0–1 30 0 0 0 0 40 0 0 01–2 60 160 120 80 0 80 40 0 02–3 120 240 160 100 0 160 120 80 403–4 160 320 240 120 0 240 180 120 80
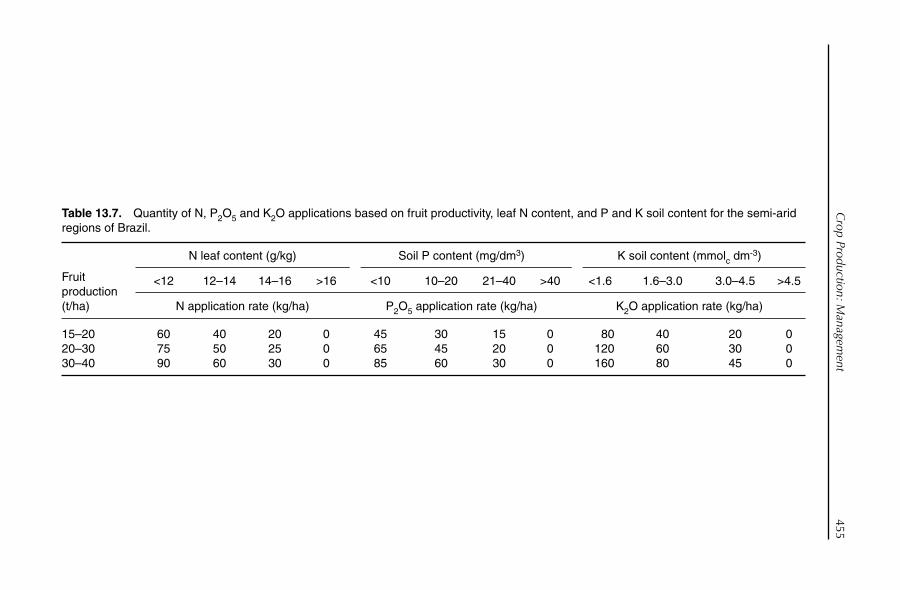
Crop Production: M
anagement
455
Table 13.7. Quantity of N, P2O5 and K2O applications based on fruit productivity, leaf N content, and P and K soil content for the semi-arid regions of Brazil.
Fruit production(t/ha)
N leaf content (g/kg) Soil P content (mg/dm3) K soil content (mmolc dm-3)
<12 12–14 14–16 >16 <10 10–20 21–40 >40 <1.6 1.6–3.0 3.0–4.5 >4.5
N application rate (kg/ha) P2O5 application rate (kg/ha) K2O application rate (kg/ha)
15–20 60 40 20 0 45 30 15 0 80 40 20 020–30 75 50 25 0 65 45 20 0 120 60 30 030–40 90 60 30 0 85 60 30 0 160 80 45 0
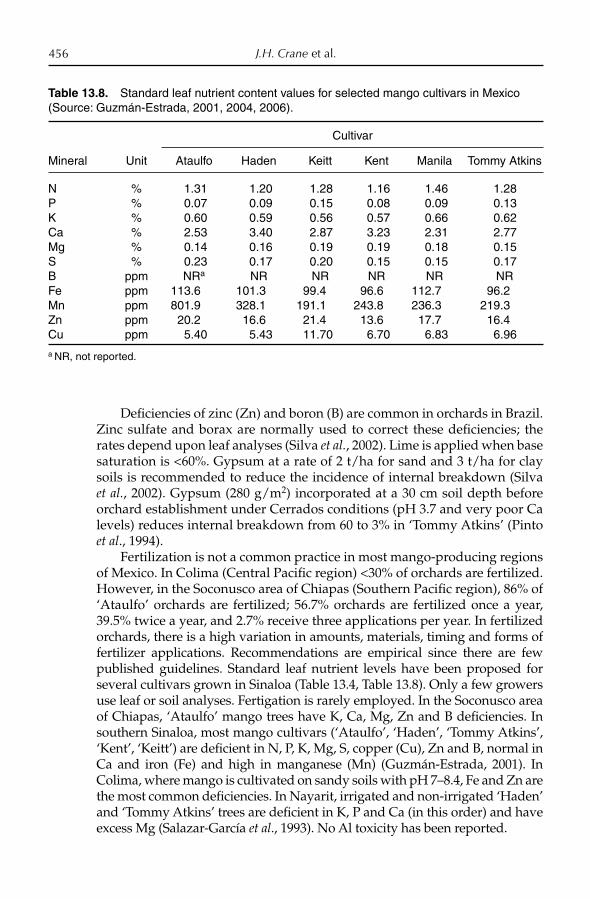
J.H. Crane et al.456
Defi ciencies of zinc (Zn) and boron (B) are common in orchards in Brazil. Zinc sulfate and borax are normally used to correct these defi ciencies; the rates depend upon leaf analyses (Silva et al., 2002). Lime is applied when base saturation is <60%. Gypsum at a rate of 2 t/ha for sand and 3 t/ha for clay soils is recommended to reduce the incidence of internal breakdown (Silva et al., 2002). Gypsum (280 g/m2) incorporated at a 30 cm soil depth before orchard establishment under Cerrados conditions (pH 3.7 and very poor Ca levels) reduces internal breakdown from 60 to 3% in ‘Tommy Atkins’ (Pinto et al., 1994).
Fertilization is not a common practice in most mango-producing regions of Mexico. In Colima (Central Pacifi c region) <30% of orchards are fertilized. However, in the Soconusco area of Chiapas (Southern Pacifi c region), 86% of ‘Ataulfo’ orchards are fertilized; 56.7% orchards are fertilized once a year, 39.5% twice a year, and 2.7% receive three applications per year. In fertilized orchards, there is a high variation in amounts, materials, timing and forms of fertilizer applications. Recommendations are empirical since there are few published guidelines. Standard leaf nutrient levels have been proposed for several cultivars grown in Sinaloa (Table 13.4, Table 13.8). Only a few growers use leaf or soil analyses. Fertigation is rarely employed. In the Soconusco area of Chiapas, ‘Ataulfo’ mango trees have K, Ca, Mg, Zn and B defi ciencies. In southern Sinaloa, most mango cultivars (‘Ataulfo’, ‘Haden’, ‘Tommy Atkins’, ‘Kent’, ‘Keitt’) are defi cient in N, P, K, Mg, S, copper (Cu), Zn and B, normal in Ca and iron (Fe) and high in manganese (Mn) (Guzmán-Estrada, 2001). In Colima, where mango is cultivated on sandy soils with pH 7–8.4, Fe and Zn are the most common defi ciencies. In Nayarit, irrigated and non-irrigated ‘Haden’ and ‘Tommy Atkins’ trees are defi cient in K, P and Ca (in this order) and have excess Mg (Salazar-García et al., 1993). No Al toxicity has been reported.
Table 13.8. Standard leaf nutrient content values for selected mango cultivars in Mexico (Source: Guzmán-Estrada, 2001, 2004, 2006).
Mineral Unit
Cultivar
Ataulfo Haden Keitt Kent Manila Tommy Atkins
N % 1.31 1.20 1.28 1.16 1.46 1.28P % 0.07 0.09 0.15 0.08 0.09 0.13K % 0.60 0.59 0.56 0.57 0.66 0.62Ca % 2.53 3.40 2.87 3.23 2.31 2.77Mg % 0.14 0.16 0.19 0.19 0.18 0.15S % 0.23 0.17 0.20 0.15 0.15 0.17B ppm NRa NR NR NR NR NRFe ppm 113.6 101.3 99.4 96.6 112.7 96.2Mn ppm 801.9 328.1 191.1 243.8 236.3 219.3Zn ppm 20.2 16.6 21.4 13.6 17.7 16.4Cu ppm 5.40 5.43 11.70 6.70 6.83 6.96
a NR, not reported.

Crop Production: Management 457
Except for N, visual symptoms of nutrient defi ciencies are uncommon in commercial mango orchards; however, fertilized trees have signifi cantly increased yield and fruit size. Nitrogen is most commonly applied, followed by P and K. Current fertilization practices vary with the mango-producing region, soil type, cultivar and tree age. The NPK recommendations for ‘Manila’ trees at 1–4 years, 5–10 years, 10–15 years, 16–20 years and >20 years old in the Gulf of Mexico region are: 0.2-0.1-0.1 kg/tree/year, 0.4-0.2-0.4 kg/tree/year, 0.6-0.3-0.6 kg/tree/year, 0.8-0.4-0.8 kg/tree/year and 1.0-0.5-1.0 kg/tree/year, respectively.
Young trees in the Southern Pacifi c region receive NPK at 0.4-0.2-0.2 kg/tree/year, respectively, from year 1 to year 5, and at 0.7-0.7-0.7 kg/tree/year thereafter. In Michoacán (Central Pacifi c region), mature ‘Haden’ and ‘Tommy Atkins’ trees receive NPK at 1.1-0.4-0.9 kg/tree/year (Chávez-Contreras et al., 2001). The NPK recommendation in the Northern Pacifi c region at 1–4 years, 5–10 years, and 10–15 years of age is 0.4-0.2-0.2 kg/tree/year, 1.3-0.55-0.85 kg/tree/year and 2.8-0.9-1.8 kg/tree/year, respectively. Although soil amendments are needed in regions with low or high pH soils (i.e. Chiapas and Colima) they are not used.
Micronutrients are not usually included in fertilization programmes; however, circumstantial evidence suggests there is an interaction between fl ower B defi ciency and extreme high temperature as a major factor causing stenocarpic fruit. Foliar B applications at either pre- or during full bloom have become a routine practice for ‘Ataulfo’ orchards. Zinc, Mn and Cu, are included with fungicide sprays during bloom and the rainy season.
General fertilizer recommendations for mangoes of different ages vary in Taiwan (Table 13.9). Fertilizers are applied in two equal rates: (i) after har-vest; and (ii) when fruit are at the marble stage of development. Fertilizer is applied by broadcasting, banding, side dressing and hole-application. Leaf analysis is used for nutrient diagnosis and fertilizer recommendations (Table 13.4). The rates of N, P and K increase with tree age (Table 13.9). Dolomite is commonly used to adjust soil acidity and as a Mg supplement. The applica-tion rates are 1 t/ha/year for sandy soils, 1.5 t/ha/year for loamy or silty soils and 2 t/ha/year for clay soils. Sulfur is applied as ammonium sulfate ((NH4)2SO4), magnesium sulfate (MgSO4) or calcium sulfate (CaSO4). Boron is applied at a rate of 50 g/tree or as a 0.3 % spray 2–3 times a year. No addi-tional micronutrient supplements are recommended.
Nutritional needs and fertilizer requirements under Florida conditions have been well studied, although similar studies have not been done in Puerto Rico (Aponte-Morán et al., 1977; Toro, 1988), Hawaii and California. The sandy and calcareous soils in Florida are very low in native nutrient content and cation exchange capacity. These soils require inorganic and organic fertilizers for optimum growth and production. The leaf mineral con-tent and defi ciency symptoms of young containerized ‘Haden’ and ‘Zill’ trees in sand culture have been described for N, P, K, Mg, Mn and S (Smith and Scudder, 1951). Obvious leaf symptoms of Ca, Cu, Zn and B were not observed during a 3-year investigation, although classical Zn defi ciency symptoms were observed in the fi eld.
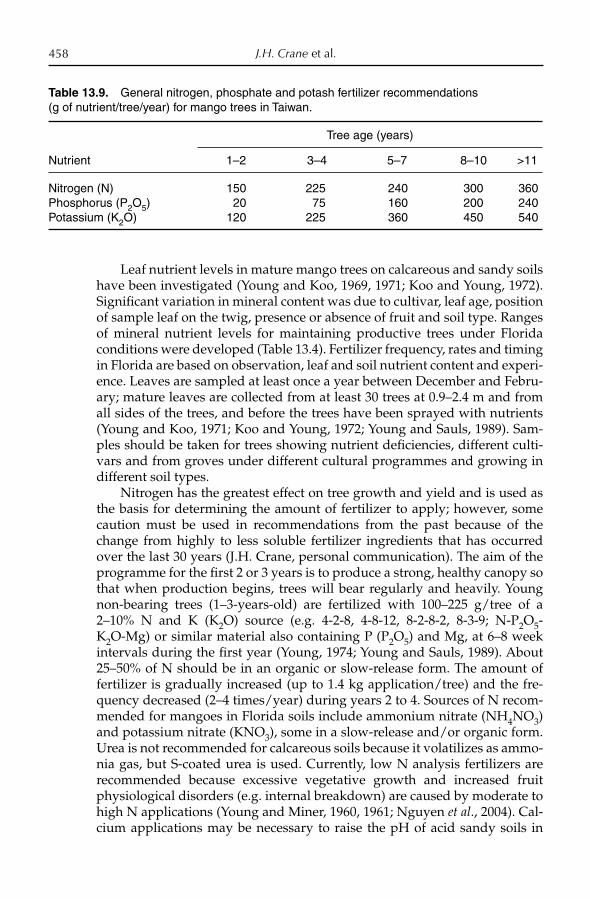
J.H. Crane et al.458
Leaf nutrient levels in mature mango trees on calcareous and sandy soils have been investigated (Young and Koo, 1969, 1971; Koo and Young, 1972). Signifi cant variation in mineral content was due to cultivar, leaf age, position of sample leaf on the twig, presence or absence of fruit and soil type. Ranges of mineral nutrient levels for maintaining productive trees under Florida conditions were developed (Table 13.4). Fertilizer frequency, rates and timing in Florida are based on observation, leaf and soil nutrient content and experi-ence. Leaves are sampled at least once a year between December and Febru-ary; mature leaves are collected from at least 30 trees at 0.9–2.4 m and from all sides of the trees, and before the trees have been sprayed with nutrients (Young and Koo, 1971; Koo and Young, 1972; Young and Sauls, 1989). Sam-ples should be taken for trees showing nutrient defi ciencies, different culti-vars and from groves under different cultural programmes and growing in different soil types.
Nitrogen has the greatest effect on tree growth and yield and is used as the basis for determining the amount of fertilizer to apply; however, some caution must be used in recommendations from the past because of the change from highly to less soluble fertilizer ingredients that has occurred over the last 30 years (J.H. Crane, personal communication). The aim of the programme for the fi rst 2 or 3 years is to produce a strong, healthy canopy so that when production begins, trees will bear regularly and heavily. Young non-bearing trees (1–3-years-old) are fertilized with 100–225 g/tree of a 2–10% N and K (K2O) source (e.g. 4-2-8, 4-8-12, 8-2-8-2, 8-3-9; N-P2O5-K2O-Mg) or similar material also containing P (P2O5) and Mg, at 6–8 week intervals during the fi rst year (Young, 1974; Young and Sauls, 1989). About 25–50% of N should be in an organic or slow-release form. The amount of fertilizer is gradually increased (up to 1.4 kg application/tree) and the fre-quency decreased (2–4 times/year) during years 2 to 4. Sources of N recom-mended for mangoes in Florida soils include ammonium nitrate (NH4NO3) and potassium nitrate (KNO3), some in a slow-release and/or organic form. Urea is not recommended for calcareous soils because it volatilizes as ammo-nia gas, but S-coated urea is used. Currently, low N analysis fertilizers are recommended because excessive vegetative growth and increased fruit physiological disorders (e.g. internal breakdown) are caused by moderate to high N applications (Young and Miner, 1960, 1961; Nguyen et al., 2004). Cal-cium applications may be necessary to raise the pH of acid sandy soils in
Table 13.9. General nitrogen, phosphate and potash fertilizer recommendations (g of nutrient/tree/year) for mango trees in Taiwan.
Nutrient
Tree age (years)
1–2 3–4 5–7 8–10 >11
Nitrogen (N) 150 225 240 300 360Phosphorus (P2O5) 20 75 160 200 240Potassium (K2O) 120 225 360 450 540

Crop Production: Management 459
other areas of Florida. Recommended Ca sources include dolomite, gypsum, calcium carbonate (CaCO3) and calcium nitrate (Ca(NO3)2).
Microelement application methods depend upon soil type, leaf analysis and other cultural practices (Young, 1974; Young and Sauls, 1989; Crane and Campbell, 1991). Foliar applications of Mn, Zn and Cu are made to trees growing in calcareous soils, because soil applications are ineffective due to high Ca content and pH. Soil applications of chelated Mn and Zn materials formulated for trees growing in calcareous soils are recommended. Foliar or soil applications of non-chelated Mn and Zn are appropriate for trees grow-ing in acid sands, although foliar applications may be more effi cacious and less expensive. Usually Cu is applied as a fungicide and additional applica-tions are unnecessary. Magnesium is frequently applied foliarly together with Mn and Zn. Typical sources of Mn include manganese sulfate (MnSO4), manganese nitrate (Mn(NO3)2) and manganese oxide (MnO), and of Zn include zinc sulfate (ZnSO4), zinc oxide (ZnO) and zinc nitrate (Zn(NO3)2). Recommended rates for foliar applications include 3.46–5.7 kg each of a Zn and an Mn sulfate per 937/l of water/ha. However, commercial microele-ment formulations containing Mn, Zn, Cu, molybdenum (Mo), S and B are used because of their convenience. Microelements are applied 2–4 times/year.
Iron is applied to soil as a drench with chelated materials formulated for calcareous soils; however, these are expensive (Young and Sauls, 1989). Typically, chelated iron as EDDHA (sodium ferric ethylenediamine di-(o-hydroxyphenylacetate)) and EDDTA (sodium ferric diethylnetriamine pentaacetate) can prevent and correct Fe defi ciency in calcareous and acid sandy soils, respectively. For young trees, 7.1–14.2 g/tree applied as a soil drench 2–4 times/year is recommended. Trees are irrigated for 1–2 days before applying a mixture of chelated iron and water to the soil around the base of the tree and then irrigated briefl y afterwards to ensure the material has moved into the upper soil profi le where most fi brous roots are located.
Foliar applications of ferrous sulphate (FeSO4) and other Fe compounds (including chelated materials) with or without adjuvants have been ineffec-tive for maintaining, preventing or correcting Fe defi ciency (Leonard and Stewart, 1953; Leonard and Calvert, 1971; Green et al., 1999). Studies with mild acids plus organosilicone adjuvant and FeSO4 show promise as an inexpensive foliar iron application method (Crane et al., 2007, 2008).
The fertilizer programme for fruit-producing trees involves maintenance of tree health and fruit production and quality. The early recommendations for mature mango trees growing in calcareous and acid sandy soils were 111–168 kg of N and K/ha/year (Young and Miner, 1960, 1961; Young et al., 1962, 1965; Young, 1974; Young and Koo, 1974); however, the recommended N rate has been reduced to 40–60% of earlier levels (J.H. Crane, personal communication) due to the slow-release component of modern mixed fertil-izer formulations. Nitrogen applications >225 kg/ha/year reduce fruit qual-ity, cause excessive vegetative growth and increase internal breakdown (Young and Miner, 1960, 1961; Raymond et al., 1998); leaf tissue analysis for N should dictate application (Davenport, 2006). Leaf N levels >1.5% can

J.H. Crane et al.460
result in little or no fl owering. Fertilizer mixtures containing 2–10% N and 4–16% K are used. Fertilizer rates may be reduced if leaf N and K levels are within acceptable ranges and tree performance is acceptable. Phosphorus applications may be reduced if leaf nutrient levels are within the acceptable range; fertilizer mixtures with 3–5% P are normal. Additional Ca applications, e.g. gypsum, dolomite, gypsum and Ca(NO3)2, on acid sandy soils increase Ca leaf levels and reduce the incidence of internal fruit breakdown (Young et al., 1962).
Microelements are applied to mature trees regardless of leaf analysis as a preventative measure. This is especially true for Fe and Zn, because correct-ing defi ciency of these elements is sometimes diffi cult and expensive. Rates for foliar sprays of Mg, Zn and Mn are similar to those for young trees with 2–4 applications/year. The rates for chelated iron for mature trees range from 14 to 113 g/tree 1–2 times/year.
Fertilizer may be applied by fertigation. Mixed fertilizers containing P are not recommended because phosphates can precipitate and clog the irri-gation system. Phosphate can be applied alone as dilute H3PO4. Iron can also be applied through microsprinklers or a drip system at increased frequency and reduced rates.
In Puerto Rico, fertilizers are also applied in dry form and through low-volume irrigation systems. Young non-bearing trees (1–3 years old) are fertil-ized with 454–1591 g/tree of a 12–15% N, 5–6% P (P2O5) and 8–10% K (K2O) source (e.g. 12-6-8, 12-6-10, 15-5-10; N-P2O5-K2O), split into two applications (December/February and April/May) (Toro, 1988). When trees begin to bear (after 3–4 years), 10-5-15, 12-6-16, or 10-5-20 is applied in split applications which increase in amount with tree age (3.4–6.8 kg/tree maximum). Some orchards are fertilized through the irrigation system at 7–15 day intervals (E.E. Toro, personal communication). In these plantings, (NH4)2SO4, urea, phosphoric acid (H3PO4) and KNO3 or potassium sulfate (K2SO4) are used. Micronutrients are applied either to the soil or to foliage in Puerto Rico; rec-ommendations for young trees are foliar applications at 21–30 day intervals during the growing season (Toro, 1988). Micronutrients are applied as needed in mature orchards and may be applied once during the spring and autumn. Sources of micronutrients are sulfate forms of Zn, Cu and Mn.
13.9 Irrigation Practices
Water requirements for mango production are not precisely known and cur-rent irrigation practices are based on experience and climatic and limited edaphic measurements. Irrigation practices are infl uenced by available tech-nology and cost, soil type and depth, rainfall amount and distribution, fertil-izer practices and production objectives. The response to irrigation is infl uenced by the rate, timing and method of application, tree-growth stage, climate, edaphic conditions and cultivar.
Irrigation of young trees assists tree establishment, prevents drought stress, and maintains constant canopy and root growth. Objectives for bearing

Crop Production: Management 461
trees are maintenance of tree health, avoidance of severe drought stress and enhancement of vegetative dormancy. Mangoes are drought tolerant (Schaffer et al., 1994); however, in some production areas under non-irrigated condi-tions, fruit production and quality may be reduced. Excessive irrigation can cause reduced tree growth and tree decline (Larson et al., 1989a, 1991d; J.H. Crane, personal communication). Mango production has been displaced onto marginal lands that possess low water and/or nutrient holding capacity and high pH. Saline water for irrigation is a concern. Irrigation methods include fl ood and furrow, high-pressure volume guns, high-volume under- and over-tree sprinkler and low-pressure microsprinkler and drip systems.
In north-eastern Brazil, under semi-arid tropical conditions, irrigation is essential throughout the hot and dry season (Albuquerque et al., 1999). Sev-eral irrigation systems are used, with c.41% of orchards using microsprinkler systems. About 21% use other types of irrigation (e.g. furrows, drip, basin, etc.) and 33% of orchards use no irrigation (Gomes et al., 2002). Mean produc-tivity of irrigated orchards may be as high as 25 t/ha compared with only 12 t/ha for non-irrigated orchards (Coelho et al., 2002). Irrigation is used on 14,500 ha (74% of the cultivated area) of commercial mango in the north-east. Microsprinkler irrigation is the most common method (30.3% of the irrigated area) (Gomes et al., 2002). Commercial mango orchards in the south-east, mainly in São Paulo, are not irrigated.
Fertigated orchards require well-trained people. Some nutrients can cause corrosion of irrigation pipes, and proper management is essential to prevent environmental damage. Proper selection of nutrients is critical due to problems of solubility and compatibility. Urea, (NH4)2SO4 and KNO3 are the primary N fertilizers; they are highly soluble and are compatible with most nutrients. However, the SO4
2– is incompatible with Ca and their mixture
causes precipitation and clogging of emitters. The oldest commercial orchards in Mexico were rainfed and established
in areas with deep soils and annual rainfall >900 mm (subhumid and humid tropics). However, growth of the mango industry has forced the planting of semi-arid tropical areas (annual rainfall 600–700 mm) (Colima and Micho-acán) (Table 13.2). Currently, 67% of orchards are rainfed (SIIAP, 2007) with annual rainfalls that fl uctuate from 900 to 3700 mm; 80% of the rainfall occurs in June–October. Irrespective of the amount of annual rainfall, since the late 1980s a signifi cant proportion of the new and old mango orchards (60,000 ha) have installed irrigation systems. This is to prevent water defi cits and to increase yield and fruit size and quality. Sources of irrigation water include rivers or deep wells and irrigation may be applied either by gravity or by electrical or diesel engines.
Irrigation management involves either furrow or pressurized systems. The furrow soil surface system (FSSS) can be simple, crossed, furrow-basin and fi sh spine. Low-pressure drip and microsprinkler systems are most com-mon, particularly the latter. Two microsprinklers (90–120 l/h) or 10–15 drip emitters (8–12 l/h) are used per mature tree in a low-pressure system. In semi-arid regions, mango potential evapotranspiration from October to June each year is 16,000 m3/ha. The FSSS system applies >25,000 m3/ha of water;

J.H. Crane et al.462
however, low-pressure systems can reduce water use to 5000 m3/ha with no negative effect on yield or fruit quality (L.M. Tapia, personal communica-tion). Water requirements in the semi-arid region of Michoacán are calculated by using evapotranspiration measured with a Class A pan and multiplied by the crop coeffi cient Kc, which for practical purposes is considered as 0.4 for vegetative growth and 0.8 for fruit growth and development (Chávez-Contreras et al., 2001). The FSSS water schedule for obtaining maximum yield and fruit quality in Michoacán is: September (pre-bloom), one 20 cm irrigation; October–December (bloom), two 10 cm irrigations spaced 20 days apart; January–April (fruit growth), eight 10 cm irrigations at 15–17 day inter-vals; May–July (vegetative growth), three 50 cm irrigations at 21–25 day inter-vals. The amount of water used for drip irrigation in mature mango trees during fruit growth and development is 1350 l/tree/week and for microsprin-klers it is 1560 l/tree/week. After the fi rst irrigation, growers water every 8 days in low-pressure irrigation systems and every 15–20 days in FSSS.
Soil-water content infl uences the frequency of fl owering, the number and length of panicles, yield and quality of mango fruits in Taiwan (Chang and Lu, 1995). Two types of irrigation system are used: (i) furrow fl ooding in the low lands; and (ii) microsprinklers, overhead sprinklers and drip (trickle) irrigation on sloped lands. Microsprinklers have become the most important irrigation system in recent years and fertigation is utilized by many produc-ers. Irrigation management is based on producer experience and observation of the soil, current weather and tree phenology.
In Florida, rainfall is not evenly distributed through the year, with the dry season occurring during the autumn–winter (October–April). During the wet spring–summer season (May–September), dry periods of 3 days or more can occur. Most orchards in Florida are irrigated only during prolonged dry periods. Several systems are common. High-volume overhead or under-tree sprinklers which run at high pressures (28,121–70,303 kg/m2) and distribute large amounts of water (0.51–0.89 cm/ha) and microsprinkler systems which run at low pressure (7030–28,121 kg/m2) and distribute lower volumes of water (37.9–1113.6 l/tree/ha). Some growers use both systems: the microsprin-kler system for irrigation and fertigation and the high-volume system for cold protection during freezing weather.
Well-established trees require little to no irrigation. During prolonged drought conditions (>30 days with no signifi cant rainfall), irrigation may be applied. Irrigation is not recommended during the cool autumn and winter months, to enhance the vegetative dormancy period induced by cool tem-peratures, to enhance synchronization of apical stems and to intensify the fl owering response after growth commences.
Currently, irrigation recommendations and practices are based on obser-vation and experience with c.6.35 cm water/application/ha. Preliminary research suggests that irrigation at 7-day intervals during the period of fruit development increases fruit size, earliness and yield (Larson et al., 1989b); however, this has not been implemented by growers. Most Florida producers rarely irrigate their mango orchards during the spring and summer as rain-fall during this period coincides with fruit development and is suffi cient for

Crop Production: Management 463
fruit production. In Puerto Rico, Hawaii and California, microirrigation is used. In Hawaii, where temperatures are consistently warm in the mango-growing areas, mature trees are more productive if irrigation is withheld for at least 2 months before fl owering (Chia et al., 1988). Drip and microsprinkler irrigation is used in California, but irrigation regimes have not been reported. In Puerto Rico, drip irrigation is more common than microsprinklers (E.E. Toro, personal communication). The drippers apply 3.79–7.75 l/tree/ha and trees are irrigated 2–3 times/week. When microsprinklers are used, one microsprinkler is placed between two trees in-row. Trees may be irrigated from 9–11 months of the year; water is withheld for 1–3 months prior to fl oral induction or fl owering.
13.10 Vegetative Growth and Reproduction Manipulation
Mango trees require new vegetative growth in order to produce fruit each year. The optimum temperature for vegetative growth is 24–30°C (Krishnamurthi et al., 1961; Shü and Sheen, 1987; Whiley et al., 1989) and when levels of essen-tial plant nutrients and water are not limiting, vigorous growth results. Mature leaves and a period of cessation of vegetative growth (i.e. mature api-cal and subtending meristems) are required for the transformation from veg-etative to reproductive growth (Núñez-Elisea and Davenport, 1992; Kulkarni, 2004; Davenport, 2006).
Canopy management and reproductive manipulation vary according to climatic conditions, cultivar and available technology (Davenport, 1993). Ultimate mango-tree size depends upon the climactic and edaphic condi-tions and cultural practices. Under optimum conditions, trees can reach heights and canopy diameters of 30 m or more (Kostermans and Bompard, 1993); however, it is diffi cult to protect large trees from insects, diseases and strong winds, and harvesting is diffi cult and costly. With the increase in costs for orchard establishment and maintenance, the number of trees per unit area has increased and tree size has decreased, whereas production of high quality fruit per unit area has increased. Growers in some production areas manipulate the period of fl owering and fruit production. Tree size can be controlled to maximize the number of trees per unit area and maintain productive tree canopy and yields.
The natural period of mango production in Brazil was originally from October to January. Production has increased from September through to April through the use of precocious and late-bearing cultivars along with fl oral induction techniques. There is potential that whole-year harvesting and mango supply can be achieved, although there are some months with low mango yields. There are three types of fl oral induction and their use depends upon phenology and season. In general, the steps are: (i) pruning of apical internodes after harvesting; (ii) application of paclobutrazol (PBZ) to stimulate fl owering by inhibiting gibberellin biosynthesis; (iii) spraying with K2SO4 (2–2.5%) to increase carbohydrate levels in the tree; (iv) drought stress to facilitate growth cessation; (v) applications of ethylene (optional); and

J.H. Crane et al.464
(vi) three applications of 3–4% KNO3 and Ca(NO3)2 to trigger fl owering (Albuquerque et al., 2002). This fl ower initiation protocol results in off-season fl owering and fruit production when mangoes are scarce and prices are high (between April and August) in the domestic market.
Tree-size control and pruning strategies appropriate for the various cul-tivars and diverse climatic regions in Mexico are under development. Researchers in conjuction with producers are investigating appropriate tech-nologies that take into account the current tree size, tree spacing and confi gu-ration in the orchards, the available technology, and the goal of the pruning programme. In young trees the central leader is pruned (5 mm above previ-ous growth scar) a year after planting to promote lateral branching and pre-cocious fl owering. Once new branches have matured the best-positioned 2–5 lateral branches around the main stem are selected. Trees are pruned for 2–3 years after every 2–3 vegetative fl ushes by heading back the wood of the last two growths (Guzmán-Estrada and Vázquez-Valdivia, 2006). Young tree training is important for ‘Ataulfo’ and ‘Keitt’ and results in strong canopies that will resist heavy fruit loads and strong winds. Tree training is unnecessary for ‘Manila’, ‘Haden’, ‘Tommy Atkins’ and ‘Kent’.
Usually mature trees are pruned every year after harvest to promote light penetration into the canopy and to remove weak, diseased, broken and poorly positioned branches. Mechanical topping and hedging is not widely practised. However, this pruning system was recently introduced in orchards in Nayarit to control the pink hibiscus mealybug (Maconellicoccus hirsutus Green). In Chiapas, 8-year-old trees and older are pruned by hand (machete) in alternate years to delay or avoid canopy crowding; one side of the canopy is pruned in one year and the other side is pruned the next year.
To rejuvenate old, very large mango trees and/or overcrowed orchards, trees are cut back to main limbs at 1–2 m above the soil level. This technique is common in the subhumid and semi-arid production areas of Chiapas and Veracruz. However, in the warm humid tropical areas of Chiapas and Veracruz this technique is not widely used because of the subsequent loss of fruit pro-duction and excessive regrowth (3–4 vegetative fl ushes/year) that occur. Selected trees can be removed to avoid overcrowding.
Climatic conditions are conducive to early bloom and harvest from the Chiapas to Michoacán production regions, where forced blooming and early fruit production is used by >80% of producers to obtain higher prices. Even when a signifi cant amount of advanced bloom is induced, blooming during the normal fl owering period may occur. The intensity of the normal bloom is infl uenced by the intensity of the fruit set by the advanced bloom. The most common method to advance bloom involves canopy sprays of KNO3 or NH4NO3 (Mosqueda-Vázquez and De los Santos, 1982; Núñez-Elisea, 1986, 1988; Guzmán-Estrada, 1991; Sandoval-Esquivez et al., 1993). The effect of these compounds is infl uenced by cultivar and environmental conditions (probably temperature). In the Gulf of Mexico region, one to two sprays of 2% KNO3 or 1% NH4NO3 solution are applied to ‘Manila’ trees any time from 15 October to 30 November to stimulate early fl owering. ‘Manila’ and ‘Ataulfo’ are sprayed with 2% KNO3 during the same period in the Southern

Crop Production: Management 465
Pacifi c region. In the Central Pacifi c region, ‘Haden’, ‘Manila’ and ‘Ataulfo’ trees are treated with one or two sprays of 2–4% KNO3 or 1–2% NH4NO3 at any time during the fi rst half of November. Similarly, ‘Haden’ and ‘Manila’ are treated during November with 8% KNO3 or 4% NH4NO3 in the Northern Pacifi c region. ‘Tommy Atkins’ does not respond to foliar nitrate treatments for promoting early fl owering. This is due to delayed fl oral initiation in this cultivar so that when nitrate treatments are applied the buds are not irrevers-ibly committed to fl owering (Pérez-Barraza et al., 2000). Consequently, veg-etative growth is produced in response to treatments that stimulate bud break (Pérez-Barraza et al., 2006a). However, application of PBZ promotes early bloom in ‘Tommy Atkins’ (Salazar-García and Vázquez-Valdivia, 1997).
Currently, soil applications of Cultar® (25% a.i.) close to the tree trunk is used for most cultivars in dosages that range from 1 ml/m canopy diameter for ‘Manila’ in Veracruz to 2–4 ml for ‘Tommy Atkins’, ‘Haden’ and ‘Ataulfo’ in Michoacán, applied at 1–2 year intervals. The response to PBZ treatment is enhanced by 30–45 days water stress and canopy sprays with nitrates (Chávez-Contreras et al., 2001). In Nayarit and Sinaloa late bloom and harvest are profi table because they are the last two production areas to be harvested in Mexico. Two canopy sprays of 50 mg/l GA3 (15 and 30 November) cause delayed bloom and shift 86% of the harvest to 1 month later (Pérez-Barraza et al., 2006b).
In Taiwan, cultivar, latitude, elevation and cultural practices are used for off-season production (Shü et al., 2000). Several strategies are utilized to stimulate fl owering and fruit set:
After harvest the last one or two vegetative fl ushes on each shoot are cut 1. back to control tree size. Subsequently, two fl ushes of healthy shoots are allowed to grow to serve as fruiting shoots for the next year. Weak or crowd-ed shoots are removed to facilitate ventilation and light penetration.
In general, fl owering is not a problem in subtropical Taiwan; however, 2. poor fruit set due to bad weather and lack of pollinators occurs occasionally. A recent programme to increase the population of pollinators, mainly the greenbottle fl ies (Chrysomyia megacephala Fabricius) in mango orchards has been very successful. Increased yield has been noted and the practice has been exploited commercially throughout the island.
Most of the mango fruit harvest goes to the domestic market within a 3. short time period, causing the price to decline rapidly. Off-season fruit pro-duction is thus very important to avoid this sharp drop in price.
Physical trunk damage by girdling and ringing and application of ethrel 4. is used to promote early fl owering of the early season cultivar ‘Tsar-swain’ (Liu, 1996). Foliar applications of KNO3 can promote early fl owering but only if applied to fl ower-bud-induced shoots; PBZ is not recommended for use on edible crops in Taiwan.
Panicle removal has been used to postpone fl owering and fruiting (Shü 5. and Sheen, 1987; Shü, 1993). Emerging terminal panicles are removed by hand. Chemical removal of terminal panicles with hydrogen cyanamide (CH2N2) or calcium cyanamide (CaCN2) causes leaf damage and is not as

J.H. Crane et al.466
effective as pruning (Hwang et al., 2004). Axillary panicle induction has some advantages, because fl owering can be timed to avoid frost or cold tempera-tures or a period of excessive rainfall during the normal fl owering period (Singh et al., 1974; Shen and Huang, 1980). Axillary panicle removal also reduces mango malformation and reduces alternate bearing (Majumder et al., 1976; Pal and Chadha, 1982).
Control of fl owering and tree size in the USA varies with respect to differ-ent climatic, edaphic and soil conditions as well as cultural practices (Daven-port, 1993). In Florida, cool temperatures during the winter months (December through to February) are usually adequate to arrest vegetative growth and induce fl ower bud differentiation. In some years, the duration of cool tempera-tures prohibits fl oral expression until late winter/early spring (February/March) and when continuous warm temperatures begin (March), profuse, syn-chronized fl owering occurs. In some years, warm and cool periods may occur for a few days to weeks during the winter and partial fl owering may occur 2–4 times during the winter. This prolongs the fl owering and harvest season.
Flowering and fruiting of trees are not actively manipulated in Florida. For young trees, vegetative growth is encouraged during the fi rst 2–3 years and panicles may be removed by hand or natural pathogens, i.e. powdery mildew or anthracnose are allowed to kill panicles and fl owers. Tipping and selective pruning of young trees is recommended to improve tree structure, control tree size and enhance early fruit production (Oosthuyse and Jacobs, 1995; Campbell and Wasielewski, 2000); however, selective pruning and mechanical topping and hedging are used to control mature-tree size. Selec-tive pruning usually involves removal of selected scaffold limbs to open up the tree canopy to light and to remove dead wood. Mature trees may or may not be allowed to grow together in the tree row to form hedgerows. Periodic mechanical topping at 3.5–5 m and hedging to leave a 2.5–3.5 m row middle is common (Crane and Campbell, 1991; J.H. Crane, personal communication). Limiting the between-row spread of the trees to 2.5–3 m improves light pen-etration into the tree canopy. In some orchards, hedging the inner sides of the canopy of adjacent rows every 2–4 years and/or topping every third or fourth row every 2–4 years is recommended. Trees are mechanically pruned immedi-ately after harvest. Timing the pruning to selected rows each year ensures that most of the planting will always be productive if continuous fl ushing of pruned parts of the canopy prohibits reproductive growth the following spring. Pruning trees shoots of 2–10 cm diameter immediately after harvest and then tip pruning 3–4 times to force multiple lateral growths has been advocated (Davenport, 2006). This strategy: (i) reduces or controls tree size; (ii) shapes trees to facilitate subsequent tip pruning; (iii) synchronizes the veg-etative fl ushing; and (iv) inhibits continuous vegetative fl ushing and prolongs the period of vegetative dormancy. After vegetative growth has ceased for 5 or more months, trees will synchronously fl ower when regrowth occurs.
In Puerto Rico, continuous vegetative growth of 1–2-year-old trees is promoted. During this time panicles may be removed by hand to promote vegetative growth, and this encourages rapid development of large trees.

Crop Production: Management 467
Temperatures are insuffi cient to inhibit vegetative growth and induce reli-able and consistent bud differentiation, causing erratic or poor fl owering and poorly synchronized fruit yields. The most common method for synchroniz-ing and enhancing the time of fl owering involves a combination of drought stress and timed application of KNO3. This method depends on cultivar and the desired time of fruit harvest. To slow (or stop) vegetative growth and to stress the trees, irrigation is withheld for 1–3 months prior to fl owering. The drought stress is prolonged until leaves become dark green and show slight signs of wilting. A 1–5% solution of KNO3 is applied to the foliage, which induces fl owering 3–4 weeks later. Regular irrigation is resumed when c.75% of the panicles have set fruit. The timing of the drought stress and KNO3 spray varies with cultivar and when fruit harvest is desired.
PBZ has been used to control vegetative growth after trees fl ush in response to pruning following harvest. A soil drench of PBZ (7–10 g/tree) is applied and trees are irrigated for 8–12 h. Irrigation is then withheld for 30–90 days until trees show signs of drought stress. A foliar application of 1–2% KNO3 is applied, and fl owering occurs 30–45 days later. The amount of tree training depends upon the cultivar and is practised on young trees when they are c.1–1.5 m high. ‘Keitt’ and ‘Palmer’ tend to have long branches of various lengths and benefi t from training to create a stronger limb structure and more compact tree. Trees are generally trained to a modifi ed central leader system, and some selective pruning of older trees is practised to open the canopy to more light and air movement. Mechanical topping and hedging are used to shape mature trees. Usually trees are topped to 3–4.5 m immedi-ately after harvest.
In Hawaii, seasons with heavy crops are commonly followed by light or no crops for 1–2 years (Nagao and Nishina, 1993). The small variation in warm temperatures and evenly distributed rainfall encourage vegetative growth, which reduces the potential for fruit production. This is less prob-lematic on the drier leeward sides of the islands. Diseases (i.e. anthracnose and powdery mildew) reduce production by infecting panicles and fl owers. Preliminary trials with 2 and 4% KNO3 applications during the winter (Feb-ruary) resulted in 66–84% fl owering 5 weeks after treatment (Nagao and Nishina, 1993); however, this procedure is not utilized commercially.
Young ‘Keitt’ mango trees are extensively trained by hand (usually twice a year) during the fi rst 4–5 years to improve the structural strength of scaf-fold limbs, increase the number of terminals and to control tree size in California. Panicles are removed by hand during the fi rst 4 years. Training is necessary because non-trained ‘Keitt’ trees tend to have a few very long scaf-fold limbs, little branching and are structurally weak. Bearing trees are pruned annually to maintain trees at 3.1–4.6 m. In California, cool tempera-tures during the winter months (November through to February) are ade-quate to arrest vegetative growth and induce bud differentiation. The duration of cool temperatures may inhibit fl oral expression until early spring (March and April), when warm temperatures allow profuse, synchronized fl owering. Warm periods during winter may stimulate early fl owering, which may be damaged by subsequent cold temperatures (Schacht, 1992).

J.H. Crane et al.468
Early season fl owering when cold temperatures and dry windy conditions prevail (December–February) results in poor fruit set and abnormal fruit. Therefore, pruning of mature trees just before April (early spring) delays fl owering and induces synchronous axillary fl owering after the danger of cold has passed.
13.11 Environmental Stress Management
Mango is an adaptable species that withstands a range of subtropical and tropical climates and soils. Physiological responses are related to the evolu-tionary history of mangoes, with monoembryonic cultivars better adapted to the subtropics and polyembryonic cultivars better adapted to the tropics.
In Brazil, the important commercial mango areas are in the tropical semi-arid climate of the north-east. Flooding and freezing rarely occur there, but wind and drought stress are very common and negatively affect growth of young mango trees and increase fruit drop. Windbreaks reduce wind stress; compact rows of elephant grass and/or rows of banana trees are utilized (Mouco et al., 2002). In general, 3–4 rows of banana trees are planted around or in perpendicular rows in the orchard against the main wind fl ow.
Drought stress, particularly in north-eastern Brazil, can suppress mango growth and production, and irrigation is used to ameliorate this problem. Drought stress has been used to increase endogenous ethylene concentra-tions and trigger fl oral induction 70–90 days after PBZ application. Drought stress should be avoided during fruit development. Coelho et al. (2002) reported the crop coeffi cient (Kc) for mango increased from 0.39 at fl owering to 0.85 during fruit development.
In Mexico, the Gulf of Mexico coastal region may experience strong, dry northerly winds (11–28 m/s) from October to April and cause limb breakage and increase fl ower and fruit drop by desiccation. To ameliorate this prob-lem, growers have planted east-west oriented bamboo (Bambusa vulgaris) and Australian pine (Casuarina equisetifolia) windbreaks. Bamboo is planted outside the Australian pine trees and two rows of pines are planted in a stag-gered arrangement. All coastal mango-producing regions in Mexico are potential targets of hurricanes during the summer rainy season, even in late October. Broken limbs are pruned, damaged trees are pruned back to sound wood, and some trees are replaced or top-worked.
Flooding for 2–3 weeks may occur during the summer rains. This is mainly a problem in low-lying areas of Chiapas and Nayarit that have loamy soils and a high water table. Drainage canals have been constructed in such areas to minimize the problem. Drought stress is common as 67% of Mexico’s mango orchards are not irrigated (SIIAP, 2007). Annual rainfall ranges from 900 to 3700 mm in Mexico and is concentrated in June to October. The dry season begins in October (close to the period of fl oral initiation) and contin-ues through the harvest period for early and some mid-season cultivars. Drought stress in areas of low rainfall causes intense fruit drop, which reduces yield and fruit size especially in heavily bearing trees. More that 60,000 ha of

Crop Production: Management 469
mangoes are irrigated in low rainfall areas. In Veracruz and Chiapas most orchards are adjacent to riverbanks and the deep soils provide root access to the water table. No data are available supporting the benefi t of irrigation in medium-high precipitation areas.
Most mango-producing regions in Mexico are frost free. The only report of frost damage to 2–4-year-old mango orchards was in Sonora where freez-ing events may occur every 6–8 years (E. Sánchez, personal communication). Low temperatures (≤10°C) during bloom reduce fruit set, especially in ‘Ataulfo’. Treatment with GA3 to delay the bloom has been suggested as a method to avoid low-temperature damage during fl owering but fl owering under high-temperature conditions is also detrimental to fruit set and crop yield (Pérez-Barraza et al., 2006b).
In Taiwan, damage from typhoons occurs periodically and trees are reset and either pruned to recover or replanted. In general, freezing temperatures are not a problem in the mango-production areas but cool temperatures can reduce fruit set. To avoid cool or cold temperatures during the normal fl ow-ering period, shoot tips may be pruned to delay and force fl owering from lateral buds. Flooding in production areas is uncommon, and drought stress is not an issue because most producers irrigate during dry periods.
The production areas of Hawaii and Puerto Rico are frost free; however, Florida and California may experience temperatures at and below freezing during the winter months (December through to February). In Florida, freez-ing temperatures (0 to −6°C) may occur for a few hours for 1–4 nights/year, although freezing temperatures for 13–15 h within a 24 h period have been reported (Johnson, 1970; Campbell et al., 1977). In California, temperatures as low as −6.6°C are common and frosts may occur 10–15 times/year during January and February (Aslan et al., 1993). Mango trees do not acclimate to cold temperatures (McKellar et al., 1983), although differences in cold toler-ance and recovery from cold damage have been observed (Carmichael, 1958). In Florida, high-volume overhead and under-tree irrigation is used to protect trees during freezing weather. At least 0.6 cm of water/ha is distributed. Overhead systems are designed for complete coverage (overlapping spray patterns) of the trees and under-tree systems are designed to spray 0.9–2.4 m into the tree canopy. Irrigation commences before freezing temperatures are reached (usually c.2–3°C) and continues until ice has melted. These systems are powered by diesel or gas engines as electrical power is unreliable during freezing weather conditions. In California, microsprinklers, wind machines and helicopters are used to raise the air temperature of plantings (Schacht, 1992).
Mango trees are relatively tolerant of wind stress (Schaffer et al., 1994; Crane and Balerdi, 2005); however, newly planted trees are commonly staked at planting in the calcareous soils of Florida to prevent damage to the bark and cambium caused by constant movement and rubbing against the rocky soil. Staking also stabilizes the tree against toppling during hurricanes. In contrast, tolerance to mature trees depends on tree size, with larger trees being more vulnerable to wind damage than pruned trees (Crane et al., 1993, 1994, 2001; Crane and Balerdi, 1996; NASS, 2006).

J.H. Crane et al.470
In the mid-20th century, windbreaks of C. equisetifolia were planted around many mango orchards in Florida. However, intrusion into the orchard, shading and damage to mango trees when trees toppled into orchards during hurricanes has stopped this practice. Some producers have topped remain-ing pine windbreaks at 4.9–6.7 m to reduce orchard shading and their potential to topple (Crane et al., 1993). In Hawaii, natural windbreaks are recommended for some areas where constant winds are a problem for establishing young trees (C.L. Chia, personal communication). In California, constant winds during spring may damage panicles and young fruit; man-made windscreens are used to protect trees (Scott, 1990; Schacht, 1992). The windscreens can be raised to prevent trapping of cold air within the plantings. Puerto Rico is affected by hurricanes but no specifi c ameliorating recommendations have been reported (Toro, 1988).
Flooding is not common in production areas of Puerto Rico, California and Hawaii; however, periodic fl ooding (1–21 days) is typical during the summer in Florida. Trees have been planted on beds of crushed limestone rock, 0.6–1.0 m high and 1.0–1.5 m wide, which allows part of the root system to be above water. Planting of orchards at sites at or below 2 m above sea level is not recommended. Mango trees are moderately fl ood tolerant (Larson et al., 1991c; Schaffer et al., 1994, 2006), although this is affected by fl oodwater tem-perature, oxygen content and anatomical adaptations of the rootstock (Lar-son et al., 1991a, 1993a, b). Periodic fl ooding of the limestone-based soils in Florida increases the availability of soil Mn and Fe (Larson et al., 1991b, 1991d, 1992), alleviating plant defi ciencies of these elements. Furthermore, rhizo-sphere anoxia increases reduction of Fe3+ to Fe2+ by nicotinamide adenine dinucleotide (NADH) in mango root tissue and Fe uptake by mango roots in oxygen-depleted media (Zude-Sasse and Schaffer, 2000).
13.12 Harvesting Practices
Harvesting is done by hand in Brazil, Mexico, Taiwan and the USA (Evans, 2007) and is one of the most expensive operations in mango production because fruit do not mature synchronously, and trees require multiple pickings. The technology to mechanize mango harvest is diffi cult because of differences in fruit colour, size and weight among cultivars, diffi culty in determining fruit maturity, requirement for multiple pickings, lack of tree-size-controlling rootstocks and tree training and moderate to large tree canopies.
Prior to harvesting in Brazil, excessive set fruit and injured and diseased fruit are removed, part of the rachis is removed to prevent scarring and bruis-ing of the fruit, and some leaves that shade fruit may be removed to allow for better peel colour development (Alves et al., 2002). Fruit in the lower part of the canopy are harvested from the ground; however, ladders are required for picking fruit high in the tree canopy. Peel damage due to latex exudation at the stem end of the fruit during harvest is a very common problem, and >50% of harvested fruit may be affected. This problem occurs mainly when fruit are picked high in the canopy with a picking pole by severing the fruit

Crop Production: Management 471
near the stem end of the pedicel. An improved picking pole that cuts the petiole c.2 cm above the pedicel reduces latex burn to <10%. Before trans-porting to the packing house, fruit containers are placed in the shade to avoid increasing the pulp temperature.
In Mexico, harvesting is done by hand when fruit have reached physio-logical maturity. Picking poles with bamboo baskets (Gulf of Mexico and Southern Pacifi c regions) or nylon net bags or cotton bags (Central and Northern Pacifi c regions) and a cutting blade at the distal end are used to reach fruit high in the canopy. Ladders may be used, although climbing trees is more common. Fruit is usually placed in 20–30 kg wooden or plastic boxes. The inner walls of the picking crates are covered by newspaper to absorb latex and decrease sap peel damage.
Several fruit maturity indexes exist for mango: pulp TSS content and/or acidity, pulp fi rmness, skin or pulp colour, calendar days or heat units from bloom to harvest. No index alone is successful because of differences among cultivars and signifi cant variability in fruit maturity within trees and orchards and among orchards. Pickers are trained to distinguish the following charac-teristics: (i) size, form and fruit colour; (ii) shoulder development (higher than the base of peduncle); (iii) cavity formation at the base of the peduncle; and (iv) increased lenticel size (Chávez-Contreras et al., 2001). The Associa-tion of Mango Packers and Exporters (EMEX; Empacadoras de Mango de Exportación) have established some minimal physical and chemical stan-dards to defi ne maturity for ‘Haden’, ‘Tommy Atkins’, ‘Kent’, ‘Keitt’ and ‘Ataulfo’. A photographic maturity index chart is used by growers, pickers, companies and packing houses. Mango fruit-peel damage by latex is common when tree sap pressure is high. To minimize this damage, irrigation is stopped at least 2 weeks prior to harvest; fruit are also picked with a long peduncle and are washed immediately after the harvest bin is full.
Mature-green mangoes are the fi rst fruit to be harvested and all the cri-ollo, Oro and Florida cultivars fi t this category. These mangoes are for the domestic market and usually sell for high prices. ‘Manila’ mangoes are picked when their colour changes from pale green to greyish green. ‘Ataulfo’ is harvested when the green peel shows a yellow colour break. The Florida mangoes are picked ‘mature-green’ and at colour break.
The harvest season in Taiwan begins in March with ‘Tsar-swain’ and ends in September or October with ‘Keitt’. Depending on the cultivar and market, fruits are harvested at 70–80% maturity. ‘Tsar-swain’ is harvested either at the green stage for pickles or at a mature ripe stage for domestic markets. ‘Irwin’ fruit are harvested at either 80% maturity for export or at >90% maturity for local markets. Green mature fruits may be triggered to ripen with calcium carbide, ethephon or ethrel at 30–40°C, depending on the cultivar. Mature fruits are stored at at 8–12°C for several days to several weeks. Fruit destined for export are disinfested using the vapour heat method; the fruit core temperature must reach and be held at 46.5°C for 30 min.
There are two markets for mango producers in the USA: the ‘green’ mar-ket for non-ripe fruit and the ‘tree-ripened’ market. The green and tree-ripened markets are speciality, niche markets where the green fruit are used as a

J.H. Crane et al.472
component of processed foods (e.g. chutneys, preserved pieces), whereas the tree-ripened fruit target the demand for ready-to-eat mangoes. Green mangoes are picked before the fruit is mature, whereas the tree-ripened mangoes are allowed to develop almost full colour and ripeness before being picked. Florida mangoes are more expensive than imports and the volume of fruit is very limited and cannot supply the demand of the national market. Usually multiple pickings are required to harvest the crop but this is infl uenced by market demand and prices, the number of blooms producing the crop and weather conditions.
Harvesting in the USA is by hand. A long picking pole with a canvas or nylon bag attached to a metal ring with a cutting blade at the distal end is commonly used in Florida (Aponte Morán et al., 1977; Crane and Campbell, 1991). Other picking aids such as ladders and mobile hydraulic lifts are also used. Time of harvesting depends upon cultivar, the intended market and market demand. In Florida, green mangoes may be picked after March, while fruit-picking time for the fresh market depends upon the cultivar reaching the mature stage desired (Crane and Campbell, 1991). In Puerto Rico, man-goes are harvested from March to November, depending upon cultivar, mar-ket price and the date when fl owering was induced. The mango season in California is restricted to September/November, and in Hawaii the main sea-son is May to August. Approximately 50% of mangoes produced in California are certifi ed organic (Linden, 2006).
13.13 Conclusions
Differences in mango culture are due to the climatic and edaphic conditions, available information and technology, and tradition in each production area. The interaction of climate and cultural practices, for example irrigation, fer-tilizer, pruning, etc., that are essential for optimizing crop yields and quality is not completely understood. None the less, horticultural systems can be developed and tested that can impact fruit production and quality. It is important that area- and, in many cases, site- and cultivar-specifi c cultural information and practices need to be developed to optimize production and fruit quality. This chapter has addressed the current state of mango culture in four different production areas, and has emphasized improvements that are essential for a prosperous industry.
Ackowledgements
The Mexican co-author acknowledges the following INIFAP researchers, based at several Research Stations (CE) and states: Ernesto Sánchez-Sánchez, CE-Valle del Yaqui (Sonora); Camerino Guzmán-Estrada, CE-Sur de Sinaloa (Sinaloa); R. Mosqueda-Vázquez (deceased) and Enrique N. Becerra-Leor, CE-Cotaxtla (Veracruz); Fulgencio M. Tucuch-Cauich, CE-EDZNA (Campeche);

Crop Production: Management 473
Rubén Cruzaley-Sarabia, CE-Iguala (Guerrero); Víctor Medina-Urrutia, CE-Tecomán (Colima); Víctor Palacio-Martínez, CE-Rosario Izapa (Chiapas).
References
Agrianual (2006) Gazeta, Santa Cruz, Rio Grande do Sul, Brazil.Agrianual (2007) Manga, mercado and perspectivas. Anuário da Agricultura Brasileira.
Instituto FNP, AGRA FNP Pesquisas Ltd, São Paulo, Brazil, pp. 378–386.Agriculture and Forestry Department (1996) Taiwan Agriculture Yearbook. Economic
Section, Agriculture and Forestry Department, Taiwan Provincial Government, Nantou, Taiwan.
Albuquerque, J.A.S. de, Mouco, M.A. do C., Medina, V.D., Santos, C.R. dos and Tavares, S.C.C. de H.O. (1999) Cultivo da Mangueira Irrigada no Semi-árido Brasileiro.Embrapa Semi-árido, Petrolina, Brazil.
Albuquerque, J.A.S. de, Mouco, M.A. do C., Medina V.D. and Vasconcelos, L.F.L. (2002) Sistemas de poda. In: Genu, P.J.C. and Pinto, A.C. de Q. (eds) A Cultura da Mangueira.Embrapa Informação Tecnológica, Brasília, Brazil, pp. 244–257.
Allen, R.G., Perrira, L.S., Raes, D. and Smith, M. (1998) Crop Evaporation, Guidelines for Computing Crop Water Requirements. Food and Agriculture Organization (FAO) Irrigation and Drainage Paper 56. FAO, Rome.
Alvarado-Ortiz, A.N. and Acin, N. (2004) Crop profi le for mangoes in Puerto Rico. Avail-able at: http://www.ipmcenters.org/cropprofi les/docs/PRmango.html (accessed 19 May 2008).
Alves, R.E., Filgueiras, H.A.C., Menezes, J.B., Assis, J.S. de, Lima, M.A.C. de, Amorim, T.B.F. and Martins, A.G. (2002) Colheita e pós-colheita. In: Genu, P.J.C. and Pinto, A.C. de Q. (eds) A Cultura da Mangueira. Embrapa Informação Tecnológica, Brasí-lia, Brazil, pp. 382–405.
Andrade, L.R.M. de (2004) Corretivos e fertilizantes para culturas perenes e semiper-enes. In: Sousa, D.M.G. and Lobato, E. (eds) Cerrado, Correção do Solo e Adubação.Embrapa Informação Tecnológica, Brasilia, Brazil, pp. 317–366.
Anonymous (1970) Clasifi cación del Suelo. Food and Agriculture Organization (FAO)/United Nations Educational, Scientifi c and Cultural Organization (UNESCO), México.
Anonymous (1982) Inventarios de Áreas Erosionadas, Rangos de Pendiente y Unidades de Suelo del Estado de Veracruz. Dirección General de Conservación del Suelo y Agua. Secretaría de Agricultura y Recursos Hidráulicos (SARH), México.
Anonymous (1989) Interim Report. South Dade Soil and Water Conservation District and Soil Conservation Service. United States Department of Agriculture (USDA), Soil Conservation Service, United States Government Printing Offi ce, Washington, DC.
Anonymous (2007) Importation of mangos from India. Federal Register 72, 10902–10907.
Aponte-Morán, C.E., Ayala-Almodóvar, A., Cibes-Viadé, H., Jackson, G., Liu, L.J., Lloréns, A., López-García, J., Orengo-Santiago, E., Pérez-Escolar, M.E., Pérez-López, A., Reyes-Soto, I., Toro-Toro, E. and Torres-Sepúlveda, A. (1977) Conjunto Tecno-logico para la Produccion de Mango. Publicacion 114. Universidad de Puerto Rico, Estacion Experimental Agricola, Río Peidras, Puerto Rico.
Aslan, S., Neja, R., Estrada, D., Dignon, W. and Mitchell, S. (1991) Soil, Water, and Cli-matic Considerations in Selecting Date Palm Planting Sites in the Coachella Valley.Coachella Valley Resource Conservation District, Coachella Valley Water District, Riverside County Farm Bureau, Coachella Valley Center, Indio, California, pp. 6–20.

J.H. Crane et al.474
Aslan, S., Bergan, M., Baetz, R., Slayback, R.D., Dyer, D. and Dignon, W. (1993) Ten-year Research Findings on Water-effi cient Ornamental Plants for the Coachella Val-ley. Desert Water Agency and Coachella Valley Resource Conservation District, Palm Springs, California, pp. 2–6.
Bahr, L.S. and Johnston, B. (eds) (1993a) California. Collier’s Encyclopedia. Vol. 5. P.F. Collier, New York.
Bahr, L.S. and Johnston, B. (eds) (1993b) Hawaii. Collier’s Encyclopedia. Vol. 11. P.F. Collier, New York.
Bahr, L.S. and Johnston, B. (eds) (1993c) Puerto Rico. Collier’s Encyclopedia. Vol. 19. P.F. Collier, New York.
Barrick, W.E. and Black, R.J. (1980) Florida Climate Data. Florida Energy Extension Ser-vice, EES-5. Florida Cooperative Extension Service, University of Florida, Institute of Food and Agricultural Sciences, Gainesville, Florida.
Bierbolini, R.E., Ramos, G.A., Mas, J.T., Torres, E.O., Trigo, J.E., Rivera, W.F., Brunet, J.E. and Rivera, L.H. (1975) Soil Survey of Magaguez Area of Western Puerto Rico.USDA-Soil Conservation Service and University of Puerto Rico, Cartographic Divi-sion, Washington, DC, pp. 1–311.
Bierbolini, R.E., Acevedo, G., Mas, J.T., Torres, E.O. and Rivera, L.H. (1979) Soil Survey of the Area of Southern Puerto Rico. United States Department of Agriculture (USDA), Soil Conservation Service and University of Puerto Rico, US Government Printing Offi ce, Washington, DC, pp. 1–87.
Bradley, J.T. (1975) Freeze Probabilities in Florida. Bulletin 777. Florida Cooperative Extension Service, University of Florida, Institute of Food and Agricultural Sciences, Gainesville, Florida.
Butson, K.D. and Prine, G.M. (1968) Weekly Rainfall Frequencies in Florida. Circular 5-187. Florida Cooperative Extension Service, University of Florida, Institute of Food and Agricultural Sciences, Gainesville, Florida.
Calhoun, F.G., Carlisle, V.W., Caldwell, R.E., Zelazny, L.W., Hammond, L.C. and Breland, H.L. (1974) Characterization Data for Selected Florida Soils. Soil Science Research Report Number 74. United States Department of Agriculture (USDA), Soil Conservation Service, Washington, DC.
Campbell, C.W. and Campbell, R.J. (1996) The ‘Glenn’ mango, an early-maturing culti-var. Proceedings of the Florida State Horticultural Society 109, 233–234.
Campbell, C.W., Knight, R.J. Jr and Zareski, N.L. (1977) Freeze damage to tropical fruits in southern Florida in 1977. Proceedings of the Florida State Horticulture Society 90, 254–257.
Campbell, R.J. (1992) A Guide to Mangos in Florida. Fairchild Tropical Garden, Miami, Florida.
Campbell, R.J. and Wasielewski, J. (2000) Mango tree training techniques for the hot tropics. Acta Horticulturae 509, 641–651.
Carlisle, V.W., Caldwell, R.E., Sodek III, F., Hammond, L.C., Calhoun, F.G., Granger, M.A. and Breland, H.L. (1978) Characterization Data for Selected Florida Soils. Soil Science Research Report Number 78-1. United States Department of Agriculture (USDA), Soil Conservation Service, Washington, DC.
Carmichael, W.W. (1958) Observations of cold damage to mangos in Dade County and the lower east coast. Proceedings of the Florida State Horticulture Society 71, 333–335.
Castro Neto, M.T., Fonseca, N., Santos Filho, H.P. and Cavalcante Junior, A.T. (2002) Propagação e padrão da muda. In: Genu, P.J.C. and Pinto, A.C. de Q. (eds) A Cultura da Mangueira. Embrapa Informação Tecnológica, Brasília, Brazil, pp. 118–136.
Chacko, E.K. (1986) Physiology of vegetative and reproductive growth in mango (Mangifera indica L.) trees. In: Proceedings of the First Australian Mango Research Workshop.

Crop Production: Management 475
Commonwealth Scientifi c and Industrial Research Organization (CSIRO), Melbourne, pp. 54–70.
Chacko, E.K. (1989) Mango fl owering – still an enigma. Acta Horticulturae 291, 12–21.Chadha, K.L. and Pal, R.N. (1986) Mangifera indica. In: CRC Handbook of Flowering,
Vol. 5. CRC Press, Boca Raton, Florida, pp. 211–230.Chang, M.-C. and Lu, C.-J. (1995) Effects of soil water content on the development, yield
and quality of mango (Mangifera indica). Research Bulletin Tainan, District Agricul-tural Improvement Station 32, 45–55.
Chávez-Contreras, X., Vega-Piña, A., Tapia-Vargas, L.M. and Miranda-Salcedo, M.A. (2001) Mango, su Manejo y Producción en el Trópico Seco de México. Libro Técnico 1. Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Aliment-ación (SAGARPA)-Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP), Campo Experimental Valle de Apatzingán. Michoacán, Mexico, pp. 108.
Chen, M.-S. (1991) Current status and prospect of mango production in Taiwan. In: Pro-ceedings of Symposium on Production Research of Fruit Crops, Chiayi City, Taiwan, pp. 317–332.
Chia, C.L., Hamilton, R.A. and Evans, D.O. (1988) Mango. Commodity Fact Sheet MAN-3(A). Hawaii Cooperative Extension Service, Hawaii Institute of Tropical Agriculture and Human Resources, University of Hawaii at Manoa, Honolulu, Hawaii.
Coelho, E.F., Oliveira, A.S. de, Netto, A.O.A., Teixeira, A.H. de C., Araújo, E.C.E. and Bassoi, L.H. (2002) Irrigação. In: Genu, P.J.C. and Pinto, A.C. de Q. (eds) A Cultura da Mangueira. Embrapa Informação Tecnológica, Brasília, Brazil, pp. 167–189.
Colburn, B. and Goldweber, S. (1961) Preparation of oolitic limestone soil for agricul-tural use. Proceedings of the Florida State Horticultural Society 74, 343–345.
Crane, J.H. and Balerdi, C.F. (1996) Effect of Hurricane Andrew on mango trees in Floridaand their recovery. Acta Horticulturae 455, 323–330.
Crane, J.H. and Balerdi, C.F. (2005) Preparing for and Recovery from Hurricane and Tropical Storm Damage to Tropical Fruit Groves in Florida. HS1022. Florida Coop-erative Extension Service, University of Florida, Institute of Food and Agricultural Sciences, Gainesville, Florida, pp. 1–8.
Crane, J.H. and Campbell, C.W. (1991) The Mango. Fact Sheet FC-2. Florida Coopera-tive Extension Service, University of Florida, Institute of Food and Agricultural Sci-ences, Gainesville, Florida.
Crane, J.H., Campbell, R.J. and Balerdi, C.F. (1993) Effect of Hurricane Andrew on tropical fruit trees. Proceedings of the Florida State Horticultural Society 106, 139–144.
Crane, J., Balerdi, C., Campbell, R., Campbell, C. and Goldweber, S. (1994) Managing fruit orchards to minimize hurricane damage. HortTechnology 4, 21–27.
Crane, J.H., Bally, I.S.E., Mosqueda-Vázquez, R.V. and Tomer, E. (1997) Crop produc-tion. In: Litz, R.E. (ed.) The Mango: Botany, Production, and Uses. CAB Inter national,Wallingford, UK, pp. 203–256.
Crane, J.H., Schaffer, B. and Campbell, R.J. (2001) Recovery from hurricanes and the long-term impacts on perennial tropical fruit crops in south Florida. HortScience 36, 1–6.
Crane, J., Schaffer, B., Li, Y., Evans, E., Montas, W. and Li, C. (2007) Effect of foliarly-applied acids and ferrous sulfate on iron nutrition of avocado trees. In: Proceedings of the VI Congreso Mundial Palta, Viña del Mar, Chile, pp. 1–13.
Crane, J.H., Schaffer, B., Li, Y.C., Evans, E.A., Montas, W. and Li, C. (2008) Effect of foliarly-applied acids and ferrous sulfate on leaf ferrous iron content and leaf greenness of lychee trees. Proceedings of the Florida State Horticultural Society 121 (in press).

J.H. Crane et al.476
Cruzaley-Sarabia, R., Ariza-Flores, R., Romero Gomezcaña, N.R., Noriega-Cantú, D.H. and Barrios-Ayala, A. (2006) Manual para Cultivar Mango en el Estado de Guerrero.Libro Técnico 3. Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación (SAGARPA)-Instituto Nacional de Investigaciones Forestales, Agríco-las y Pecuarias (INIFAP), Campo Experimental Iguala, Iguala, Guerrero, México, 96 pp.
Cull, B.W. (1991) Mango crop management. Acta Horticulturae 291, 154–173.Cunha, G.A.P. da and Castro Neto, M.T. (2000) Implantação do pomar. In: Manga
Produção, Aspectos Técnicos. Embrapa Informações Tecnológicas, Brasília, Brazil, pp. 29–30.
Davenport, T. (1993) Floral manipulation in mangos. In: Proceedings of a Conference on Mango in Hawaii. University of Hawaii at Manoa, College of Tropical Agriculture and Human Resources, Honolulu, Hawaii, pp. 54–60.
Davenport, T.L. (2006) Pruning strategies to maximize tropical mango production from the time of planting to restoration of old orchards. HortScience 41, 544–548.
Davenport, T.L. and Núñez-Elisea, R. (1997) Reproductive physiology. In: Litz, R.E. (ed.) The Mango: Botany, Production, and Uses, CAB International, Wallingford, UK, pp. 69–146.
Degani, C., Cohen, M., Reuveni, O., El-Batsri, R. and Gazit, S. (1993) Frequency and characteristics of zygotic seedlings from polyembryonic mango cultivars deter-mined using isozymes as genetic markers. Acta Horticulturae 341, 78–85.
De los Santos, R.F. and Mosqueda-Vázquez, R. (1988–89) Comparación de 21 cultiva-res y 12 selecciones Mexicanas de mango Mangifera indica L. en la zona central del estado de Veracruz. Revista Chapingo 62–63, 63–68.
Empacadoras de Mango de Exportación (EMEX) (2007) Cierre de Temporada 2006. Em-pacadoras de Mango de Exportación (EMEX, Asociación Civil). Zapopan, Jalisco, Mexico.
Espenshade, E.W. Jr (ed.) (1992) Goode’s World Atlas, 18th edn. Rand McNally, New York.
Evans, E. (2007) Mangoes: Estimated Production Costs in the Miami-Dade County, 2003–2004. Commodity Budgets. Agricultural Economics Extension. Available at: http://agecon-trec.ifas.ufl .edu/documents/mango0304.pdf (accessed 19 May 2008).
Florida Agricultural Statistics Service (FASS) (2003) Tropical Fruit, Production and Value, 2003. FASS, Orlando, Florida, pp. 1–2.
García, E. (1973) Modifi caciones al Sistema de Clasifi cación Climática de Köppen. Instituto de Geografía, Universidad Nacional Autonoma de Mexico, Mexico City.
Garczynski, C.J. (1995) National Weather Service, Station: 048892, 1951–1993.National Weather Service, University of California, Riverside, California.
Gazeta Grupo de Comunicações (2006) Anuário Brasileiro da Fruticultura. Gazeta SantaCruz, Rio Grande do Sul, Brazil.
Getz, R. (1979) Florida Daily Temperature Normals. Circular 464. Florida Cooperative Extension Service, University of Florida, Institute of Food and Agricultural Sciences, Gainesville, Florida.
Gomes, U., Tozetto, L.J. and Pinto, A.C. de Q. (2002) Censo Frutícola do Nordeste Brasil-eiro: Situação da Cultura de Manga. Boletim Técnico. CODEVASF, Brasília, Brazil, p. 6.
Green, J., Crane, J.H., Li, Y., Sanford, R. and Rodriguez, O. (1999) Preliminary results on the effectiveness of two organosilicone-based adjuvants plus iron to correct leaf iron chlorosis of containerized carambola (Averrhoa carambola) trees. Proceedings of the Florida State Horticultural Society 112, 176–178.

Crop Production: Management 477
Guzmán-Estrada, C. (1991) Aplicación foliar de agroquímicos para inducir la fl oración en Mangifera indica L. cv. Manila y Haden en el Sur de Sinaloa. Memoria del XXIV Congreso Nacional de la Ciencia del Suelo, Pachuca, Hidalgo, Mexico.
Guzmán-Estrada, C. (2001) Evaluación nutrimental en los huertos comerciales de man-go en el sur de Sinaloa. Horticultura Mexicana 3, 178.
Guzmán-Estrada, C. (2004) Evaluación y determinación del abastecimiento nutrimental en mango del sur de Sinaloa. Informes Anuales 2000, 2001–2003 y 2003–2004. Campo Experimental Sur de Sinaloa, Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP), Sur de Sinaloa, Mexico, 12 pp.
Guzmán-Estrada, C. (2006) Nutrient supply of mango in Southern Sinaloa, México. (Abstract). In: The Eighth International Mango Symposium, Sun City, Johannesburg, South Africa, 5–10 February.
Guzmán-Estrada, C. and Vázquez-Valdivia, V. (2006) La poda del mango. In: Vázquez-Valdivia, V. and Pérez-Barraza, M.H. (eds) El Cultivo del Mango: Principios y Tec-nología de Producción. Libro Técnico No. 1. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP), Centro de Investigación Regional Pacífi -co Centro (CIRPAC), Campo Experimental Santiago Ixcuintla, Santiago Ixcuintla, Mexico, pp. 87–120.
Hamilton, R.A. (1993) Origin and classifi cation of mango varieties in Hawaii. In: Pro-ceedings of a Conference on Mango in Hawaii. University of Hawaii at Manoa, College of Tropical Agriculture and Human Resources, Honolulu, pp. 28–33.
Hamilton, R.A., Chia, C.L. and Evans, D.O. (1992) Mango Cultivars in Hawaii. Informa-tion Text Series 042. Hawaii Cooperative Extension Service, Hawaii Institute of Tropical Agriculture and Human Resources, University of Hawaii at Manoa, Hono-lulu, Hawaii.
Henderson, W.G., Jr, Carter, L.J., Moore, A.L., Stein, R.A., Wettstein, C.A. and Yamataki, H. (1984) Soil Survey of Lee County, Florida. United States Department of Agricul-ture (USDA), Natural Resource Conservation Service, US Government Printing Offi ce, Washington, DC, pp. 1–185.
Hwang, S.F., Chen, D.L., Hu, W.S. and Shü, S.H. (2004) Axillary panicle induction by chemicals in mango tree (Mangifera indica L.). Acta Horticulturae 645, 177–182.
Job, J.-R. (1989) Fertilizer application rates, soil fertilities, yields and qualities of man-goes in Taiwan. Soil and fertilizer experiment report. Department of Agriculture and Forest, Taiwan Province, Taichung, pp. 201–229.
Johnson, G., Muirhead, I., Mayers, P. and Cooke, T. (1989) Diseases. In: Bagshaw, J., Brown, B., Cooke, T., Cunningham, I., Johnson, G., Mayers, P. and Muirhead, I. (eds) MangoPests and Disorders. Department of Primary Industries, Brisbane, Australia, pp. 1–9.
Johnson, W.O. (1970) Minimum Temperatures in the Agricultural Areas of Peninsular Florida, Summary of Seasons 1937–1967. Institute of Food and Agricultural Sci-ences (IFAS) Publication No. 9. IFAS, University of Florida, Gainesville, Florida.
Koo, R.J.C. and Young, T.W. (1972) Effects of age and position on mineral composition of mango leaves. Journal of the American Society for Horticultural Science 97, 792–794.
Kostermans, A.J.G.H. and Bompard, J. (1993) The Mangoes, their Botany, Nomencla-ture, Horticulture and Utilization. Academic Press, New York.
Krishnamurthi, S., Randhawa, G.S. and Sivaraman Nair, P.C. (1961) Growth studies in the mango (Mangifera indica L.) under Delhi (sub-tropical) conditions. Indian Jour-nal of Horticulture 18, 106–118.
Kulkarni, V.J. (2004) The tri-factor hypothesis of fl owering in mango. Acta Horticulturae 645, 61–70.
Lambe, A., Núñez-Elisea, R. and Davenport, T. (1991) New developments in marcotting mangos. Tropical Fruit World 1, 80–82.

J.H. Crane et al.478
Larson, K.D., Schaffer, B. and Davies, F.S. (1989a) Flooding, carbon assimilation and growth of mango trees. (Abstract). In: Program and Abstracts of the American Soci-ety for Horticultural Science, p. 126.
Larson, K.D., Schaffer, B. and Davies, F.S. (1989b) Effect of irrigation on leaf water potential, growth, and yield of mango trees. Proceedings of the Florida State Horti-culture Society 102, 226–228.
Larson, K.D., Davies, F.S. and Schaffer, B. (1991a) Floodwater temperature and stem lenticel hypertrophy in Mangifera indica (Anacardiaceae). American Journal of Botany 78, 1397–1403.
Larson, K.D., Graetz, D.A. and Schaffer, B. (1991b) Flood-induced chemical transforma-tions in calcareous agricultural soils of south Florida. Soil Science 152, 33–40.
Larson, K.D., Schaffer, B. and Davies, F.S. (1991c) Flooding, leaf gas exchange, and growth of mango in containers. Journal of the American Society for Horticultural Science 116, 156–160.
Larson, K.D., Schaffer, B. and Davies, F.S. (1991d) Mango responses to fl ooding in lime-stone soil. Proceedings of the Florida State Horticultural Society 104, 33–39.
Larson, K.D., Schaffer, B., Davies, F.S. and Sanchez, C.A. (1992) Flooding, mineral nutri-tion, and gas exchange of mango trees. Scientia Horticulturae 52, 113–124.
Larson, K.D., Schaffer, B. and Davies, F.S. (1993a) Floodwater oxygen content, ethylene production and lenticel hypertrophy in fl ooded mango (Mangifera indica L.) trees. Journal of Experimental Botany 22, 665–671.
Larson, K.D., Schaffer, B. and Davies, F.S. (1993b) Physiological, morphological and growth responses of mango trees to fl ooding. Acta Horticulturae 341, 152–159.
Leonard, C.D. and Calvert, D.V. (1971) Field tests with new iron chelates on citrus growing on calcareous soils. Proceedings of the Florida State Horticultural Society84, 24–31.
Leonard, C.D. and Stewart, I. (1953) Chelated iron as a corrective for lime-induced chlo-rosis in citrus. Proceedings of the Florida State Horticultural Society 66, 49–54.
Lima Filho, J.M.P. (2000) Determinação do potencial hídrico da mangueira utilizando-se a câmara de pressão. Anais do XVI Congresso Brasileiro de Fruticultura. (Resu-mos em CD ROM). Embrapa Agroindústria Tropical, Fortaleza, Brazil.
Linden, T. (2006) Organic Mangos Now Coming out of California. The Produce News. Available at: http://www.producenews.com/storydetail.cfm?ID=6216 (accessed 8 December 2007).
Litz, R.E. (1984) In vitro somatic embryogenisis from nucellar callus of monoembryonic mango. HortScience 19, 715–717.
Litz, R.E. (1986) Mango. In: Bajaj, Y.P.S. (ed.) Biotechnology in Agriculture and Forestry. Vol. 1: Trees. Springer-Verlag, Berlin, pp. 267–273.
Litz, R.E. (ed.) (2005) Biotechnology of Perennial Fruit and Nut Crops. CAB International, Wallingford, UK.
Litz, R.E., Knight, R.J., Jr and Gazit, S. (1984) In vitro somatic embryogenesis from Mangifera indica L. callus. Scientia Horticulturae 222, 233–240.
Liu, M.-F. (1996) Mangoes. Extension Bulletin. Agriculture and Forestry Department, Taiwan Provincial Government. Nantou, Taiwan.
Majumder, P.K. and Sharma, D.K. (1990) Mango. In: Bose, T.K. and Mitra, S.K. (eds) Fruits: Tropical and Subtropical. Naya Prokash, Calcutta, India, pp. 1–62.
Majumder, P.K., Sharma, D.K., Singh, M.P. and Singh, R.N. (1976) Improve productivity of malformed mango trees. Indian Horticulture 20, 7–8.
McKellar, M., Buchanan, D.W. and Campbell, C.W. (1983) Cold hardiness of two culti-vars of avocado and a mango. Proceedings of the Florida State Horticultural Society 96, 212–215.

Crop Production: Management 479
Mosqueda-Vázquez, R. and De los Santos, R.F. (1982) Aspersiones de nitrato de potasio para adelantar e inducir la fl oración del mango cv. Manila in Mexico. Proceedings of the Tropical Region, American Society for Horticultural Science 25, 311–331.
Mosqueda-Vázquez, R., Avila-Reséndiz, C., García, P.E., De los Santos, R.F. and Ireta-Ojeda, A. (1996) ‘Esmeralda’ un Clon Para Usarse Como Interinjerto y Reducir el Tamaño del Árbol de Mango Cultivar Manila. Technical Bulletin No. 12. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP)-Colegio de Postgraduados, Veracruz, México, 21 pp.
Mouco, M.A. do C., Albuquerque, J.A.S., Pinto, A.C. de Q., Castro Neto, M.T. de and Barbosa, F.R. (2002) Implantação do pomar. In: Genu, P.J.C. and Pinto, A.C. de Q. (eds) A Cultura da Mangueira. Embrapa Informação Tecnológica, Brasília, Brazil, pp. 138–143.
Mukherjee, S.K. (1953) The mango – its botany, cultivation, uses and future improve-ment, especially as observed in India. Economic Botany 7, 130–162.
Mukherjee, S.K. (1972) Origin of mango (Mangifera indica). Economic Botany 26, 260–264.
Nagao, M.A. and Nishina, M.S. (1993) Use of potassium nitrate on mango fl owering. In: Proceedings of a Conference on Mango in Hawaii. University of Hawaii at Manoa, College of Tropical Agriculture and Human Resources, Honolulu, Hawaii, pp. 61–66.
Nascimento, A.S. de, Carvalho, R. da S., Mendonça, M. da C. and Sobrinho, R.B. (2002) Pragas e seu controle In: Genu, P.J.C. and Pinto, A.C. de Q. (eds) A Cultura da Mangueira. Embrapa Tecnológica, Brasília, Brazil, pp. 277–297.
National Agricultural Statistics Service (NASS)-Hawaii (2006) Hawaii Tropical Specialty Fruits. NASS, United States Department of Agriculture (USDA), Hawaii Field Offi ce and Hawaii Department of Agriculture, Agricultural Development Division, Wash-ington, DC, pp. 1–4.
Negi, S.S. (2000) Mango production in India. Acta Horticulturae 509, 69–78.Nguyen, H., Hofman, P., Holmes, R., Bally, I., Stubbings, B. and McConchie, R. (2004)
Effect of nitrogen on the skin colour and other quality attributes of ripe ‘Kensington Pride’ mango (Mangifera indica L.) fruit. Journal of Horticultural Science and Bio-technology 79, 204–210.
Noble, C.V., Drew, R.W. and Slabaugh, J.D. (1996) Soil Survey of Dade County Area, Florida. United States Department of Agriculture (USDA) Natural Resource Conser-vation Service, US Government Printing Offi ce, Washington, DC, pp. 1–116.
Núñez-Elisea, R. (1986) Producción Temprana de Mango ‘Haden’ y ‘Manila’ con Asper-siones de Nitrato de Potasio. Folleto para productores No. 8. Centro de Investiga-ciones Agrícolas Pacífi co Centro (CIAPAC), Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP), Secretaría de Agricultura y Recursos Hidráulicos (SARH), Campo Agrícola Experimental Tecomán, Tecomán, Mexico.
Núñez-Elisea, R. (1988) Nitrato de Amonio: Nueva Alternativa para Adelantar la Flo-ración y Cosecha del Mango. Desplegable para productores No. 4. Secretaría de Agricultura y Recursos Hidráulicos (SARH)-Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP), Campo Experimental Tecomán, Tecomán, Colima, Mexico.
Núñez-Elisea, R. and Davenport, T.L. (1992) Requirement for mature leaves during fl oral induction and fl oral transition in developing shoots of mango. Acta Horticulturae296, 33–37.
Núñez-Elisea, R. and Davenport, T.L. (1995) Effect of leaf age, duration of cool tempera-ture treatment, and photoperiod on bud dormancy release and fruit initiation in mango. Scientia Horticulturae 62, 63–73.

J.H. Crane et al.480
Núñez-Elisea, R., Ferreira, W., Caldeira, M.L. and Davenport, T.L. (1989) Marcottage of mango: a useful tool for physiological studies of fl ower induction. (Abstract). In: Proceedings of the 86th Annual Meeting of the American Society for Hortcultural Science, Ames, Iowa, p. 115.
Núñez-Elisea, R., Davenport, T.L. and Caldiera, M.L. (1991) An experimental system to study mango fl owering using containerized trees propagated by air-layering. Pro-ceedings of the Florida State Horticultural Society 104, 39–41.
Núñez-Elisea, R., Ferreira, W., Caldeira, M.L. and Davenport, T.L. (1992) Adventitious rooting of ‘Tommy Atkins’ mango air layers induced with naphthaleneacetic acid. HortScience 27, 926.
Oosthuyse, S.A. (1995) Pruning of mango trees: an update. South African Mango Grow-ers’ Association Yearbook 15, 1–15.
Oosthuyse, S.A. and Jacobs, G. (1995) Relationship between branching frequency, and growth, cropping and structural strength of 2-year-old mango trees. Scientia Horti-culturae 64, 85–93.
Pal, R.N. and Chadha, K.L. (1982) Deblossoming mangoes with cycloheximide. Journalof Horticultural Science 57, 331–332.
Perez, A. and Pollack, S. (2007) Fruit and Tree Nuts Outlook. FTS-327. United States Department of Agriculture (USDA) Economic Research Service, Washington, DC, pp. 13–14.
Pérez-Barraza, M.H., Salazar-García, S. and Vázquez-Valdivia, V. (2000) Delayed infl o-rescence bud initiation, a clue for the lack of response of the ‘Tommy Atkins’ man-go to promoters of fl owering. Acta Horticulturae 509, 567–572.
Pérez-Barraza, M.H., Vázquez-Valdivia, V. and Salazar-García, S. (2006a) Defoliación de brotes apicales y su efecto sobre la diferenciación fl oral del mango ‘Tommy At-kins’. Fitotecnia Mexicana 29, 313–319.
Pérez-Barraza, M.H., Vázquez-Valdivia, V. and Salazar-García, S. (2006b) Manipulación de la fl oración y cosecha. In: Vázquez-Valdivia, V. and Pérez-Barraza, M.H. (eds) ElCultivo del Mango: Principios y Tecnología de Producción. Libro Técnico No. 1. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP), Centro de Investigación Regional Pacífi co Centro (CIRPAC), Campo Experimental Santiago Ixcuintla, Santiago Ixcuintla, Mexico, pp. 187–209.
Pinto, A.C. de Q. (2000) A teorática no Cultivo da Manga: Sinopse. Embrapa Cerrados, Planaltina, Brasilia, Brazil.
Pinto, A.C. de Q. (2004) Melhoramento genético da manga (Mangifera indica L.) no Brasil. In: Rozane, D.E., Darezzo, R.J., Aguiar, R.L., Aguilera, G.H.A. and Zam-bolim, L. (eds) Manga, Produção Integrada, Industrilalização e Comercialização.UFV, Viçosa, Brazil, pp. 17–78.
Pinto, A.C. de Q. and Genú, P.J. de C. (1981) Infl uência do adubo orgânico e de se-mentes sem endocarpo sobre a germinação e vigor de porta-enxertos de manguei-ra. Pesquisa Agropecuária Brasileira 16, 111–115.
Pinto, A.C. de Q. and Ramos, V.H.V. (1998) Formação do pomar. Guia Técnico do Produtor Rural 18, Embrapa Cerrados, Brasilia, Brazil, p. 2.
Pinto, A.C. de Q., Ramos, V.H.V., Junqueira, N.T.V., Lobato, E. and de Souza, D.M. (1994) Relação Ca/N nas folhas e seu efeito na produção e qualidade da manga ‘Tommy Atkins’ sob condições de Cerrados. (Abstract). Congresso Brasileiro de Fru-ticultura 13, Salvador, Bahia, Brazil, p. 763.
Ploetz, R.C. (1994) Mango disease caused by fungi. In: Ploetz, R.C., Zentmyer, G.A., Nishijima, W., Rohrbach, K. and Ohr, H.D. (eds) Compendium of Tropical Fruit Diseases. American Phytopathological Society (APS) Press, St Paul, Minnesota, pp. 34–40.

Crop Production: Management 481
Ploetz, R.C. (2003) Diseases of mango. In: Ploetz, R.C. (ed.) Diseases of Tropical Fruit Crops. CAB International, Wallingford, UK, pp. 327–363.
Ponce-González, F. and Salazar-García, S. (1992) Etiología del cancro del mango (Mangifera indica L.) cv. Manila en las Varas, Nayarit, México. Revista Mexicana de Fitopatologia 10, 153–155.
Quayle, R.G., Cram, R.S. and Burgin, M.G. (1995) Climatic Averages and Extremes for US Cities. Historical Climatology Series 6-3. US Department of Commerce, NationalOceanic and Atmospheric Administration (NOAA), Ashville, North Carolina, pp. 76–79.
Rajan, S. and Ram, S. (1988) Studies on the root regeneration in mango air-layers. Acta Horticulturae 231, 192–197.
Raymond, L., Schaffer, B., Brecht, J.K. and Hanlon, E.A. (1998) Internal breakdown, mineral element concentration, and weight of mango fruit. Journal of Plant Nutri-tion 21, 871–889.
Rossetto, C.J., Ribeiro, I.J.A., Gallo, P.B., Soares, N.B., Bortoletto, N. and Paulo, E.M. (1996) Mango breeding for resistance to diseases and pests. Acta Horticulturae455, 299–304.
Salazar-García, S. and Vázquez-Valdivia, V. (1997) Physiological persistence of paclobutrazol on the ‘Tommy Atkins’ mango (Mangifera indica L.) under rainfed conditions. Journal of Horticultural Science 72, 339–345.
Salazar-García, S., Gutiérrez-Camacho, G., Becerra-Bernal, E. and Gómez-Aguilar, R. (1993) Diagnóstico nutricional del mango en San Blas, Nayarit. Revista Fitotecnia Mexicana 16, 190–202.
Sandoval-Esquivez, A., Hernández, O.J., Montecillo, T.J.L. and Quilantan, C.J. (1993) Manual de Producción de Mango en la Costa de Chiapas. Publicación especial No. 1. Campo Experimental Rosario Izapa, Centro de Investigación Regional Pacífi co Sur (CIRPS), Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP), Secretaría de Agricultura y Recursos Hidráulicos (SARH), Tapachula, Chiapas, Mexico.
Santos Filho, H.P., Tavares, S.C.C. de H., Matos, A.P., Costa, V.S. de O., Moreira, W.A. and Santos, C.C.F dos (2002) Doenças, monitoramento e controle. In: Genu, P.J.C. and Pinto, A.C. de Q. (eds) A Cultura da Mangueira. Embrapa Informação Tecnológica, Brasília, Brazil, pp. 300–352.
Sauls, J.W. and Campbell, C.W. (1980) Mango Propagation. Fact Sheet FC-58. Florida Cooperative Extension Service, University of Florida, Institute of Food and Agricul-tural Sciences, Gainesville, Florida.
Schacht, H. (1992) Desert green. California Farmer 275, 10–11.Schaffer, B., Whiley, A.W. and Crane, J.H. (1994) Mango. In: Schaffer, B. and Andersen, P.C.
(eds) Handbook of Environmental Physiology of Fruit Crops, Vol. II, Sub-tropical andTropical Crops. CRC Press, Boca Raton, Florida, pp. 165–197.
Schaffer, B., Davies, F.S. and Crane, J.H. (2006) Responses of subtropical and tropical fruit trees to fl ooding in calcareous soil. HortScience 41, 549–555.
Schnell, R.J. and Knight, R.J., Jr (1991) Are polyembryonic mangos dependable sources of nucellar seedlings for rootstocks? Proceedings of the Florida State Horticultural Society 104, 44–47.
Schnell, R.J. and Knight, R.J., Jr (1992) Frequency of zygotic seedlings from fi ve polyem-bryonic mango rootstocks. HortScience 27, 174–176.
Schnell, R.J., Knight, R.J., Jr and Harkins, D.M. (1994) Eliminating zygotic seedlings in ‘Turpentine’ mango rootstock populations by visual roguing. HortScience 29, 319–320.
Scott, L. (1990) A Tropical Oasis. The Press-Enterprise, Riverside County, California.

J.H. Crane et al.482
Shen, T.-M. and Huang, P.-C. (1980) Effect of chemical treatments and infl orescence pinching on the regulation of fl owering date and fruit setting of mango tree (Mangifera indica L.). Journal of the Chinese Society for Horticultural Science 26,61–70.
Shü, Z.H. (1993) Chemical pruning and induction of panicles in mango. Acta Horticul-turae 341, 199–205.
Shü, Z.H. and Sheen, T.F. (1987) Floral induction in axillary buds of mango (Mangiferaindica L.) as affected by temperature. Scientia Horticulturae 31, 81–87.
Shü, Z.H., Yen, C.R., Ke, L.S., Lin, T.S., Shiesh, C.C., Wang, D.N. and Liu, M.F. (2000) Mango production in Taiwan. Acta Horticulturae 509, 87–94.
Silva, D.J., Quaggio, J.A., Pinto, P.A. da C., Pinto, A.C. de Q. and Magalhães, A.F. de J. (2002) Nutrição e adubação In: Genu, P.J.C. and Pinto, A.C. de Q. (eds) A Cultura da Mangueira. Embrapa Informação Tecnológica, Brasília, Brazil, pp. 192–221.
Singh, R.N., Majumder, P.K., Sharma, D.K., Sinha, G.C. and Bose, P.C. (1974) Effect of de-blossoming on the productivity of mango. Scientia Horticulturae 2, 399–403.
Singh, L.B. (1960) The Mango: Botany, Cultivation and Utilization. Leonard Hill, Lon-don.
Singh, R.N. (1978) Mango. Indian Council for Agricultural Research, New Delhi, India.Sistema Integral de Información Agroalimentaria y Pesquera (SIIAP) (2007) Anuario Es-
tadístico de la Producción Agrícola. SAGARPA-México. Available at: http://www.oeidrus-portal.gob.mx/aagricola_siap/icultivo/index.jsp (accessed 10 September 2007).
Smith, P.F. and Scudder, G.K., Jr (1951) Some studies of mineral defi ciency symptoms in mango. Proceedings of the Florida State Horticultural Society 64, 243–248.
Sousa, D.M.G. de, Lobato, E. and Rein, T.A. (2004) Adubação com fósforo. In: Sousa, D.MG. and Lobato, E. (eds) Cerrado, Correção do Solo e Adubação. Embrapa Infor-mação Tecnológica, Brasilia, Brazil, pp. 147–168.
Southeast Regional Climate Center (SERCC) (2007) Ponce 4E, Puerto Rico: Period of Record General Climate Summary – Precipitation. Available at: http://radar.meas.ncsu.edu/cgi-bin/sercc/cliMAIN.pl?pr7292 (accessed 22 August 2007).
Souza, J. da S., Almeida, C.O., Araújo, J.L.P. and Cardoso, C.E.L. (2002) Aspectos sócio-econômicos. In: Genu, P.J.C. and Pinto, A.C. de Q. (eds) A Cultura da Mangueira.Embrapa Informação Tecnológica, Brasília, Brazil, pp. 20–29.
Stassen, P.J.C., Grove, H.G. and Davie, S.J. (1999) Tree shaping strategies for higher density mango orchards. Journal of Applied Horticulture 1, 1–4.
Toro, E.E. (1988) Cultivo de Mangos en Puerto Rico. H-125. Universidad de Puerto Rico, Recinto de Mayaguez, Colegio de Ciencias Agricolas, Mayaguez, Puerto Rico, pp. 1–95.
Vázquez-Valdivia, V., Salazar-García, S. and Pérez-Barraza, M.H. (2000) Esmeralda in-terstocks reduce ‘Ataulfo’ mango tree size with no reduction in yield: results of the fi rst fi ve years. Acta Horticulturae 509, 291–296.
Whiley, A.W. (1993) Environmental effects on phenology and physiology of mango – a review. Acta Horticulturae 341, 168–176.
Whiley, A.W., Rasmussen, T.S., Saranah, J.B. and Wolstenholme, B.N. (1989) Effect of temperature on growth, dry matter production and starch accumulation in ten man-go (Mangifera indica L.) cultivars. Journal of Horticultural Science 64, 753–765.
Young, T.W. (1974) Fertilizing Mango Trees in Florida. Report SUB 68-1. University of Florida, Institute of Food and Agricultural Sciences, Agricultural Research and Edu-cation Center, Homestead, Florida.
Young, T.W. and Koo, R.C.J. (1969) Mineral composition of Florida mango leaves. Pro-ceedings of the Florida State Horticulture Society 82, 324–328.

Crop Production: Management 483
Young, T.W. and Koo, R.C.J. (1971) Variations in mineral content of Florida mango leaves. Proceedings of the Florida State Horticulture Society 84, 298–303.
Young, T.W. and Koo, R.C.J. (1974) Increasing yield of ‘Parvin’ and ‘Kent’ mango on Lakewood sand by increased nitrogen and potassium fertilization. Proceedings of the Florida State Horticulture Society 87, 380–384.
Young, T.W. and Miner, J.T. (1960) Response of ‘Kent’ mangos to nitrogen fertilization. Proceedings of the Florida State Horticulture Society 73, 334–336.
Young, T.W. and Miner, J.T. (1961) Relationship of nitrogen and calcium to ‘soft-nose’ disorder in mango fruits. Proceedings of the Florida State Horticulture Society 78, 201–208.
Young, T.W. and Sauls, J.W. (1989) The Mango Industry of Florida. Bulletin 189. Florida Cooperative Extension Service, University of Florida, Institute of Food and Agricul-tural Sciences, Gainesville, Florida.
Young, T.W., Koo, R.C.J. and Miner, J.T. (1962) Effects of nitrogen, potassium and calciumfertilization on ‘Kent’ mangos on deep, acid, sandy soil. Proceedings of the Florida State Horticulture Society 75, 364–371.
Young, T.W., Koo, R.C.J. and Miner, J.T. (1965) Fertilizer trials with ‘Kent’ mangos. Proceedings of the Florida State Horticulture Society 78, 369–375.
Zude-Sasse, M. and Schaffer, B. (2000) Infl uence of soil oxygen depletion on iron uptake and reduction in mango (Mangifera indica L.) root. Proceedings of the Florida State Horticultural Society 113, 1–4.

© CAB International 2009. The Mango, 2nd Edition: Botany, Production and Uses484 (ed. R.E. Litz)
14 Postharvest Physiology
J.K. Brecht1 and E.M. Yahia2
1University of Florida, Florida, USA2Universidad Autónoma de Querétaro, Querétaro, Mexico
14.1 Introduction 48414.2 Contribution of Mango Fruit to Human Nutrition and Health 48514.3 Mango Ripening Physiology 491
Climacteric behaviour 491Ethylene production and responses 493
14.4 Compositional Changes during Fruit Maturation and Ripening 494Organic acids 494Soluble sugars 495Structural polysaccharides 496Pigments and colour 498Phenolic compounds 502Flavour (taste, aroma) 504
14.5 Transpiration and Water Loss 50714.6 Physical Damage and Physiological Disorders 507
Chilling injury (CI) 507Heat injury 508
14.7 Modifi ed Atmospheres (MA) and Controlled Atmospheres (CA) 508Injuries associated with MA and CA 510Modifi ed atmosphere packaging (MAP) 511Semipermeable coatings 513Insecticidal CA 514
14.8 Manipulation of Mango Postharvest Physiology by Molecular Biology 51514.9 Conclusions 516
14.1 Introduction
Successful postharvest handling of mangoes requires knowledge of the post-harvest physiology of the fruit and how the fruit physiology determines the best handling practices to maintain and develop high fruit quality. For example, mango, like banana, tomato and avocado, is a climacteric fruit, which means

Postharvest Physiology 485
that it may be picked when mature but before ripening has commenced, and subsequently ripened postharvest. As mango fruit mature on the tree and begin to ripen, eating quality improves, but potential marketable life de-creases due to the diffi culty of controlling the ripening changes once they have been initiated, increased bruising susceptibility and increased decay. Susceptible mango cultivars tend to develop more internal breakdown (jelly seed, soft nose and stem-end cavity) the longer that harvesting is delayed (Raymond et al., 1998; see Galán Saúco, Chapter 9, this volume). As a tropical species, mangoes are subject to chilling injury (CI), which limits the use of refrigeration to maintain postharvest quality. Mangoes are also subject to other physiological disorders, physical damage and decay, the symptoms of which may make the fruit unmarketable (Yahia et al., 2006a).
Mangoes harvested at a mature but unripe stage of development (‘mature-green’) can be stored in the unripe state as long as the initiation of ethylene production and hence ripening is avoided. The initiation of ripening can be avoided by prompt cooling and storage at a low temperature at which ripen-ing does not occur or, more effectively, by changing the composition of the storage atmosphere so that the oxygen (O2) level is reduced and carbon diox-ide (CO2) level is raised. This latter approach is called either modifi ed atmo-sphere (MA) or controlled atmosphere (CA) storage, depending on the degree of control. These technologies slow fruit metabolism and specifi cally inhibit the initiation of ethylene production. With MA or CA transport or storage, mangoes can typically be maintained in a fi rm, green condition for several days longer than can be achieved with normal refrigerated air storage. How-ever, there are limits to the levels of O2 and CO2 that can be tolerated by mangoes and these limits are affected by several factors, including cultivar, maturity or ripeness stage, storage temperature and storage time (Yahia, 1998).
Mango postharvest physiology and technology have been described in previous reports, book chapters and reviews (Subramanyam et al., 1975; Lak-shminarayana, 1980; Ledger, 1986; Peacock, 1986; Lizada, 1991; Coates and Johnson, 1993; Johnson and Coates, 1993; Lizada, 1993; Heather, 1994; Jacobi et al., 1994; Johnson et al., 1997; Mitra and Baldwin, 1997; Tharanathan et al., 2006).
14.2 Contribution of Mango Fruit to Human Nutrition and Health
Consumers are becoming aware of the nutritional and health benefi ts of fresh fruits and vegetables. Mango fruit are a rich source of vitamin C (Table 14.1), although the content decreases during ripening (Thomas, 1975; Vinci et al., 1995). ‘Raspuri’ mango is rich in vitamin C (300 mg/100 g fresh fruit) during the early stages of development, but the concentration is less (39.1–69.5 mg/100 g) at maturity (Siddappa and Bhatia, 1954). The content of vitamin C was between 13 and 178 mg/100 g in the ripe fruit of 50 cultivars surveyed by Singh (1960). The vitamin C content in fully grown mango fruit of culti-vars in Puerto Rico ranged between 6 and 63 mg/100 g (Iguina de George
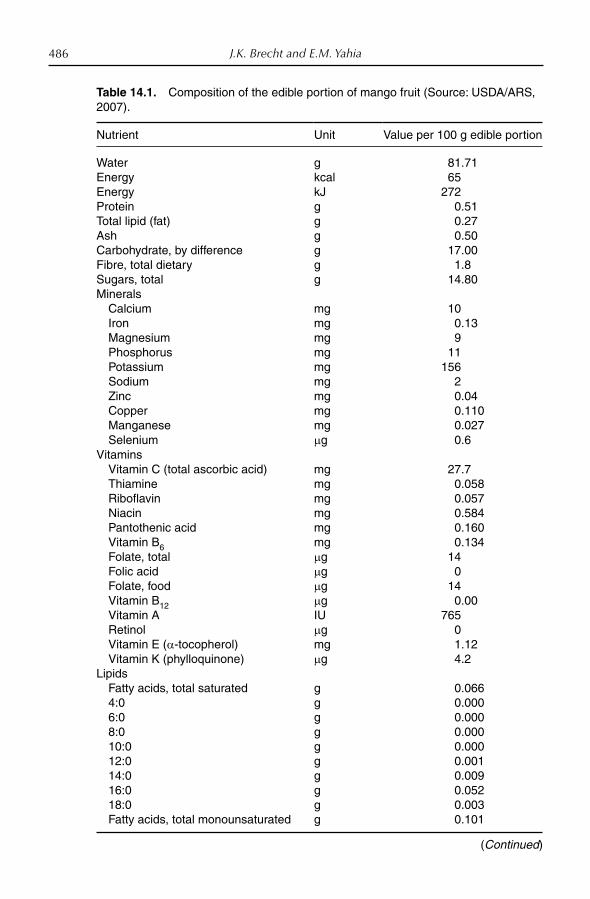
J.K. Brecht and E.M. Yahia486
Table 14.1. Composition of the edible portion of mango fruit (Source: USDA/ARS, 2007).
Nutrient Unit Value per 100 g edible portion
Water g 81.71Energy kcal 65Energy kJ 272Protein g 0.51Total lipid (fat) g 0.27Ash g 0.50Carbohydrate, by difference g 17.00Fibre, total dietary g 1.8Sugars, total g 14.80Minerals
Calcium mg 10Iron mg 0.13Magnesium mg 9Phosphorus mg 11Potassium mg 156Sodium mg 2Zinc mg 0.04Copper mg 0.110Manganese mg 0.027Selenium g 0.6
VitaminsVitamin C (total ascorbic acid) mg 27.7Thiamine mg 0.058Ribofl avin mg 0.057Niacin mg 0.584Pantothenic acid mg 0.160Vitamin B6 mg 0.134Folate, total g 14Folic acid g 0Folate, food g 14Vitamin B12 g 0.00Vitamin A IU 765Retinol g 0Vitamin E ( -tocopherol) mg 1.12Vitamin K (phylloquinone) g 4.2
LipidsFatty acids, total saturated g 0.0664:0 g 0.0006:0 g 0.0008:0 g 0.00010:0 g 0.00012:0 g 0.00114:0 g 0.00916:0 g 0.05218:0 g 0.003Fatty acids, total monounsaturated g 0.101
(Continued)

Postharvest Physiology 487
et al., 1969). Vitamin C content was 105.2, 65.7 and 17.3 mg/100 g in ‘Langra’, ‘Ashwini’ and ‘Fazli’ mangoes, respectively (Gofur et al., 1994), and decreased rapidly 5–7 weeks after fruit set, and when ripe fruit were stored at room temperature. Vitamin B1 (thiamine) in two mango cultivars was 35–60 g/100 g,
Table 14.1. Continued
Nutrient Unit Value per 100 g edible portion
16:1 undifferentiated g 0.04818:1 undifferentiated g 0.05420:1 g 0.00022:1 undifferentiated g 0.000Fatty acids, total polyunsaturated g 0.05118:2 undifferentiated g 0.01418:3 undifferentiated g 0.03718:4 g 0.00020:4 undifferentiated g 0.00020:5 n-3 g 0.00022:5 n-3 g 0.00022:6 n-3 g 0.000Cholesterol g 0
Amino acids Tryptophan g 0.008Threonine g 0.019Isoleucine g 0.018Leucine g 0.031Lysine g 0.041Methionine g 0.005Phenylalanine g 0.017Tyrosine g 0.010Valine g 0.026Arginine g 0.019Histidine g 0.012Alanine g 0.051Aspartic acid g 0.042Glutamic acid g 0.060Glycine g 0.021Proline g 0.018Serine g 0.022
OtherEthanol g 0.0Caffeine mg 0Theobromine mg 0-Carotene g 445-Carotene g 17-Cryptoxanthin g 11
Lycopene g 0Lutein + zeaxanthin g 0

J.K. Brecht and E.M. Yahia488
and vitamin B2 (ribofl avin) in three cultivars was 45–55 g/100 g (Stahl, 1935). Thiamine content of four Philippine cultivars was 57–600 g/100 g, and ribofl avin content of three cultivars was 37–730 g/100 g (Quinones et al., 1944). Folic acid in green mangoes was 36 mg/100 g (Gosh, 1960).
The mango fruit is a rich source of carotenoids, some of which function as provitamin A: -carotene (all-trans), -cryptoxthanin (all-trans and cis), zeaxanthin (all-trans), luteoxanthin isomers, violaxanthin (all-trans and cis) and neoxanthin (all-trans and cis) (Mercadante et al., 1997; Yahia et al., 2006b; Ornelas-Paz et al., 2007, 2008). Total carotenoid content rose from 12.3 to 38.0
g/g in ‘Keitt’ and from 17.0 to 51.2 g/g in ‘Tommy Atkins’ from the mature-green to the ripe stage (Mercadante and Rodriguez-Amaya, 1998), and ripen-ing alterations occurred principally in the major carotenoids, violaxantin and -carotene. With ‘Keitt’, all-trans- -carotene, all-trans-violaxanthin and 9-cis-violaxanthin increased from 1.7, 5.4 and 1.7 g/g, respectively, in the mature-green fruit to 6.7, 18.0 and 7.2 g/g in the ripe fruit (Mercadante and Rodriguez-Amaya, 1998). In ‘Tommy Atkins’ these carotenoids increased from 2.0, 6.9 and 3.3 g/g to 5.8, 22.4 and 14.5 g/g, respectively, during ripening. Geographic effects were reported to be substantial (Mercadante and Rodriguez-Amaya, 1998). Some of the cis and trans isomers of provitamin A reported in ‘Haden’ and ‘Tommy Atkins’ mangoes include 13-cis- -carotene (trace amounts), trans- -carotene (12.5–15.5 g/g) and trans- -cryptoxanthin (0.3–0.4 g/g) (Godoy and Rodriguez-Amaya, 1994). In processed mango juice, violaxanthin was not detected, auroxanthin appeared at an appre-ciable level, and -carotene was the principal carotenoid (Mercadante and Rodriguez-Amaya, 1998). The major carotenoid in ‘Bourbon’, ‘Haden’, ‘Extreme’, ‘Golden’ and ‘Tommy Atkins’ mangoes is -carotene (48–84% of the total), while epoxycarotenoids (violaxanthin, luteoxanthin and mutatox-anthin) constitute 13–49% of the total (Godoy and Rodriguez-Amaya, 1989). Mean vitamin A in these mangoes (retinol equivalents/100 g) ranges from 115.3 (‘Haden’) to 430.5 (‘Extreme’).
Children in Senegal with normal cytology had higher serum retinol and -carotene levels than those with abnormal cytology after massive oral doses
of vitamin A and consumption of mangoes (Carlier et al., 1992). Mango retinol is highly bioavailable (82% effi ciency) by estimating vitamin A and carotene reserves in the liver and plasma of rats (Yuyama et al., 1991). During mango fruit ripening, vitamin A increases – ripe mangoes are tenfold richer in caro-tene than partially ripe fruit, while unripe green mangoes contain only trace amounts (Modi and Reddy, 1967). Mevalonic acid, a precursor of carotenoids, increases progressively during mango ripening (Modi and Reddy, 1967). Vitamin A equivalents in 100 g of mango fruit are 1000 to 6000 IU (Singh, 1960). The -carotene content of the fruit of 30 mango cultivars in Puerto Rico ranged from 400 to 800 IU/100 g fresh fruit (Iguina de George et al., 1969). The development of -carotene in mangoes held at 16–21°C was lower than that at 20–28°C (Vazquez-Salinas and Lakshminarayana, 1985). Jungalwala and Cama (1963) identifi ed 16 different carotenoids in ‘Alphonso’ mangoes, and
-carotene accounted for 60% of the total. Of the oxycarotenoids, luteoxan-thin, violaxanthin and cis-violaxanthin were present in signifi cant amounts.
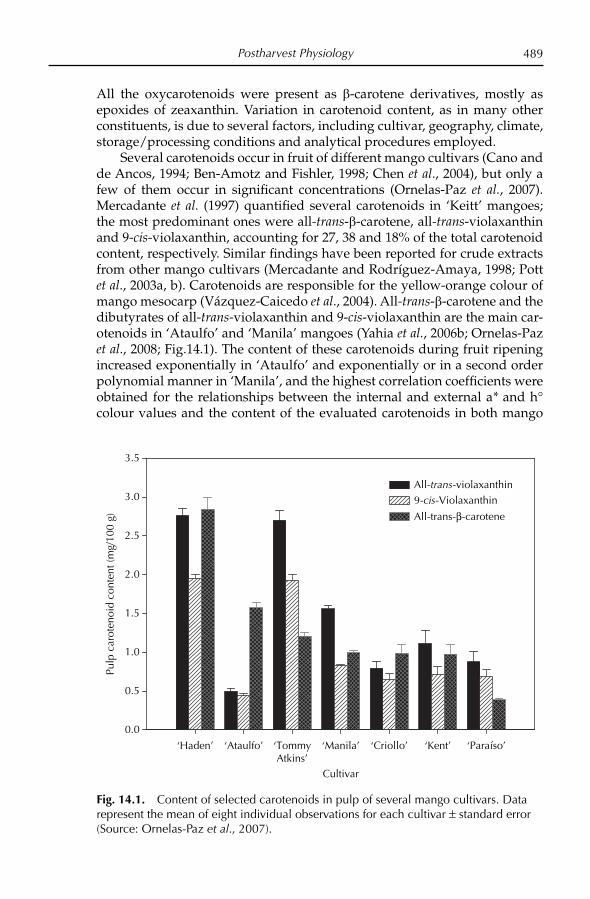
Postharvest Physiology 489
All the oxycarotenoids were present as -carotene derivatives, mostly as epoxides of zeaxanthin. Variation in carotenoid content, as in many other constituents, is due to several factors, including cultivar, geography, climate, storage/processing conditions and analytical procedures employed.
Several carotenoids occur in fruit of different mango cultivars (Cano and de Ancos, 1994; Ben-Amotz and Fishler, 1998; Chen et al., 2004), but only a few of them occur in signifi cant concentrations (Ornelas-Paz et al., 2007). Mercadante et al. (1997) quantifi ed several carotenoids in ‘Keitt’ mangoes; the most predominant ones were all-trans- -carotene, all-trans-violaxanthin and 9-cis-violaxanthin, accounting for 27, 38 and 18% of the total carotenoid content, respectively. Similar fi ndings have been reported for crude extracts from other mango cultivars (Mercadante and Rodríguez-Amaya, 1998; Pott et al., 2003a, b). Carotenoids are responsible for the yellow-orange colour of mango mesocarp (Vázquez-Caicedo et al., 2004). All-trans- -carotene and the dibutyrates of all-trans-violaxanthin and 9-cis-violaxanthin are the main car-otenoids in ‘Ataulfo’ and ‘Manila’ mangoes (Yahia et al., 2006b; Ornelas-Paz et al., 2008; Fig.14.1). The content of these carotenoids during fruit ripening increased exponentially in ‘Ataulfo’ and exponentially or in a second order polynomial manner in ‘Manila’, and the highest correlation coeffi cients were obtained for the relationships between the internal and external a* and h° colour values and the content of the evaluated carotenoids in both mango
Cultivar
‘Haden’ ‘Ataulfo’ ‘TommyAtkins’
‘Manila’ ‘Criollo’ ‘Kent’ ‘Paraíso’
Pulp
car
oten
oid
cont
ent (
mg/
100
g)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
All-trans-violaxanthin9-cis-Violaxanthin
All-trans-β-carotene
Fig. 14.1. Content of selected carotenoids in pulp of several mango cultivars. Data represent the mean of eight individual observations for each cultivar ± standard error (Source: Ornelas-Paz et al., 2007).
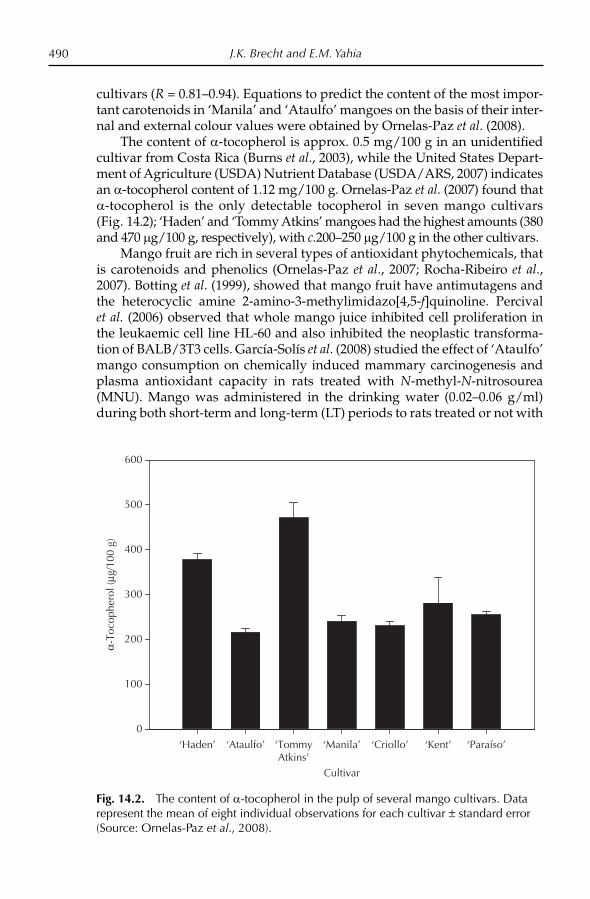
J.K. Brecht and E.M. Yahia490
cultivars (R = 0.81–0.94). Equations to predict the content of the most impor-tant carotenoids in ‘Manila’ and ‘Ataulfo’ mangoes on the basis of their inter-nal and external colour values were obtained by Ornelas-Paz et al. (2008).
The content of -tocopherol is approx. 0.5 mg/100 g in an unidentifi ed cultivar from Costa Rica (Burns et al., 2003), while the United States Depart-ment of Agriculture (USDA) Nutrient Database (USDA/ARS, 2007) indicates an -tocopherol content of 1.12 mg/100 g. Ornelas-Paz et al. (2007) found that
-tocopherol is the only detectable tocopherol in seven mango cultivars (Fig. 14.2); ‘Haden’ and ‘Tommy Atkins’ mangoes had the highest amounts (380 and 470 g/100 g, respectively), with c.200–250 g/100 g in the other cultivars.
Mango fruit are rich in several types of antioxidant phytochemicals, that is carotenoids and phenolics (Ornelas-Paz et al., 2007; Rocha-Ribeiro et al., 2007). Botting et al. (1999), showed that mango fruit have antimutagens and the heterocyclic amine 2-amino-3-methylimidazo[4,5-f]quinoline. Percival et al. (2006) observed that whole mango juice inhibited cell proliferation in the leukaemic cell line HL-60 and also inhibited the neoplastic transforma-tion of BALB/3T3 cells. García-Solís et al. (2008) studied the effect of ‘Ataulfo’ mango consumption on chemically induced mammary carcinogenesis and plasma antioxidant capacity in rats treated with N-methyl-N-nitrosourea (MNU). Mango was administered in the drinking water (0.02–0.06 g/ml) during both short-term and long-term (LT) periods to rats treated or not with
Cultivar
‘Haden’ ‘Ataulfo’ ‘TommyAtkins’
‘Manila’ ‘Criollo’ ‘Kent’ ‘Paraíso’
α-To
coph
erol
(μg/
100
g)
0
100
200
300
400
500
600
Fig. 14.2. The content of -tocopherol in the pulp of several mango cultivars. Data represent the mean of eight individual observations for each cultivar ± standard error (Source: Ornelas-Paz et al., 2008).

Postharvest Physiology 491
MNU. Rats treated with MNU showed no differences in mammary carcino-genesis or in plasma antioxidant capacity measured by both ferric reducing/antioxidant power (FRAP) and total oxyradical scavenging capacity assays. However, in animals not treated with MNU, but with an LT intake of mango, the plasma antioxidant capacity as measured by the FRAP assay tended to increase in a dose-dependent manner. This suggests that mango consump-tion by healthy subjects may increase antioxidants in plasma.
14.3 Mango Ripening Physiology
Ripening is part of the natural senescence of mango fruit. It is an irreversible process that contributes to organelle disruption and changes in chemical con-stituents, fl avour and texture. While ripening improves the eating quality of mango fruit, the postharvest life of the fruit is reduced. Natural senescence, and thus ripening, is aggravated and promoted by ethylene, mechanical injury and high temperature. This process can be delayed by lower tempera-ture, elimination of mechanical damage and reducing ethylene production (Yahia et al., 2006a). Ripening of mango is inhibited while fruit are attached to the tree, and respiration and ripening are stimulated upon detachment (Lakshminarayana, 1973). Burg and Burg (1962) reported that ethylene levels in the tissues of mature-green, attached mango fruit were relatively high (1.87 l/l) and suggested that ethylene was ineffective for promoting ripen-ing due to a ripening inhibitor supplied by the tree.
Changes associated with mango fruit ripening include: (i) fl esh colour from greenish yellow to yellow to orange in all cultivars (Plate 80a); (ii) skin colour from green to yellow in some cultivars (Plate 80b); (iii) chlorophyll decreases and carotenoid content increases; (iv) fl esh fi rmness decreases and juiciness increases; (v) starch is converted into sugars; (vi) total soluble solids (TSS) content increases; (vii) titratable acidity decreases; (viii) characteristic aroma volatiles increase; (ix) CO2 production rate increases from 40–50 to 160–200 mg/kg/h at 20°C; and (x) ethylene production rate increases from 0.1–0.2 to 1–3 l/kg/h at 20°C. Gowda and Huddar (2000) found the changes in eight mango selections during ripening included reductions in fruit weight, volume, length, thickness, fi rmness, pulp content, pulp:peel ratio, starch and vitamin C, and increases in TSS, pH, total sugars, sugar:acid ratio, pulp carotenoid content and peel colour.
Climacteric behaviour
Mango is a climacteric fruit, exhibiting a climacteric pattern of respiration and an increase in ethylene production during ripening (Cua and Lizada, 1990; Reddy and Srivastava, 1999; Lalel et al., 2003; Fig. 14.3). The initiation of ethylene production within the fruit triggers and coordinates the changes that occur during ripening. These changes include colour changes in the peel and fl esh, softening of the fl esh, and development of sweet fl avour and
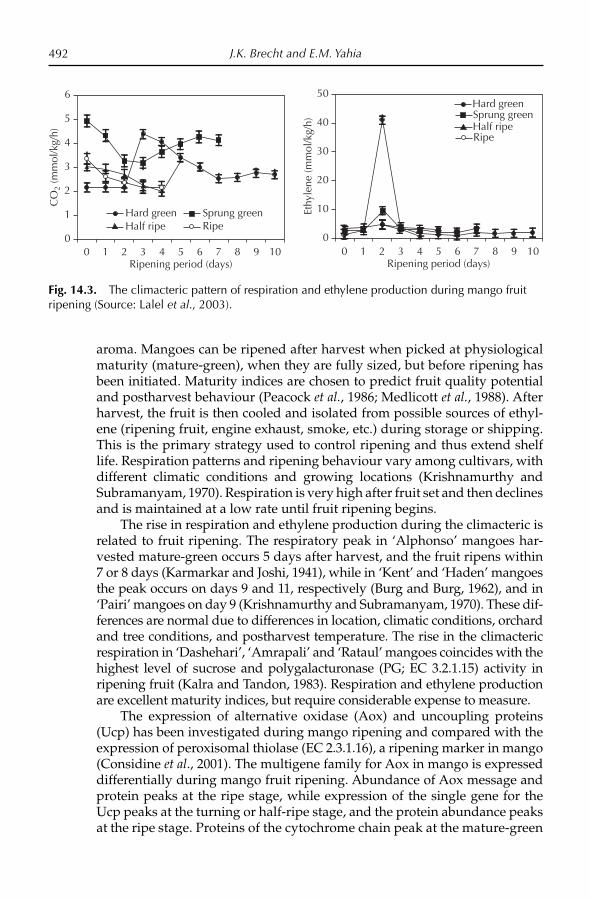
J.K. Brecht and E.M. Yahia492
aroma. Mangoes can be ripened after harvest when picked at physiological maturity (mature-green), when they are fully sized, but before ripening has been initiated. Maturity indices are chosen to predict fruit quality potential and postharvest behaviour (Peacock et al., 1986; Medlicott et al., 1988). After harvest, the fruit is then cooled and isolated from possible sources of ethyl-ene (ripening fruit, engine exhaust, smoke, etc.) during storage or shipping. This is the primary strategy used to control ripening and thus extend shelf life. Respiration patterns and ripening behaviour vary among cultivars, with different climatic conditions and growing locations (Krishnamurthy and Subramanyam, 1970). Respiration is very high after fruit set and then declines and is maintained at a low rate until fruit ripening begins.
The rise in respiration and ethylene production during the climacteric is related to fruit ripening. The respiratory peak in ‘Alphonso’ mangoes har-vested mature-green occurs 5 days after harvest, and the fruit ripens within 7 or 8 days (Karmarkar and Joshi, 1941), while in ‘Kent’ and ‘Haden’ mangoes the peak occurs on days 9 and 11, respectively (Burg and Burg, 1962), and in ‘Pairi’ mangoes on day 9 (Krishnamurthy and Subramanyam, 1970). These dif-ferences are normal due to differences in location, climatic conditions, orchard and tree conditions, and postharvest temperature. The rise in the climacteric respiration in ‘Dashehari’, ‘Amrapali’ and ‘Rataul’ mangoes coincides with the highest level of sucrose and polygalacturonase (PG; EC 3.2.1.15) activity in ripening fruit (Kalra and Tandon, 1983). Respiration and ethylene production are excellent maturity indices, but require considerable expense to measure.
The expression of alternative oxidase (Aox) and uncoupling proteins (Ucp) has been investigated during mango ripening and compared with the expression of peroxisomal thiolase (EC 2.3.1.16), a ripening marker in mango (Considine et al., 2001). The multigene family for Aox in mango is expressed differentially during mango fruit ripening. Abundance of Aox message and protein peaks at the ripe stage, while expression of the single gene for the Ucp peaks at the turning or half-ripe stage, and the protein abundance peaks at the ripe stage. Proteins of the cytochrome chain peak at the mature-green
00
1
2
3
4
5
6
1 2 3
Hard green Sprung greenRipeHalf ripe
Ripening period (days)
CO
2 (m
mol
/kg/
h)
Ethy
lene
(mm
ol/k
g/h)
4 5 6 7 8 9 10 00
10
20
30
40
50
1 2 3Ripening period (days)
4 5 6 7 8 9 10
Hard greenSprung green
RipeHalf ripe
Fig. 14.3. The climacteric pattern of respiration and ethylene production during mango fruit ripening (Source: Lalel et al., 2003).

Postharvest Physiology 493
stage, suggesting that increases in cytochrome chain components are impor-tant for facilitating the climacteric burst of respiration and that Aox and Ucp are important in postclimacteric senescence processes (Considine et al., 2001). Because both message and protein for the Aox and Ucp increase in a similar pattern, their expression is not controlled in a reciprocal manner but may be active simultaneously.
Fruit slicing affects respiration rate (Allong et al., 2001). Slicing of mature-green ‘Julia’ and ‘Graham’ mangoes increased respiration rate immediately after cutting, but it decreased signifi cantly within the fi rst 12 h of storage at 5 or 10°C, yet still remained at levels above that of the intact fruit throughout the storage period. The effect of slicing on half-ripe and fi rm-ripe fruit is an initial increase in respiration followed by a decline to levels of the intact fruit.
Ethylene production and responses
Mangoes have a moderate ethylene production peak of 1–3 l/kg/h during ripening at 20°C. Ethylene, applied directly or as ethrel, induces faster and more uniform fruit softening (Lakshminarayana, 1973; Barmore, 1974; Lak-shminarayana et al., 1974; Sornsrivichai and Waru-Aswapti, 1989). Ethylene treatment can be prior to shipping (Barmore and Mitchell, 1975). There is disagreement regarding the effect of ethylene treatment on quality (Chaplin, 1988), and this may be related to maturity when treated. Treatment of imma-ture fruit leads to softening, but the fruit have poor fl avour.
Mango fruit ripening is accompanied by increased ethylene production, which coordinates the ripening process. Mango expresses an autocatalytic increase in ethylene production during ripening (Mattoo and Modi, 1969b). Ethylene production starts before full ripeness is reached (Burg and Burg, 1962; Cua and Lizada, 1990). Ethylene production in unripe mango fruit is very low (<0.1 l/kg/h) (Burdon et al., 1996). Ethylene production decreases as the fruit matures, is then undetectable for a time and reappears upon ini-tiation of ripening (Akamine and Goo, 1973). ‘Kent’ and ‘Haden’ mango fruit have internal ethylene concentrations of c.0.01 l/l during the preclimacteric phase, increasing to c.0.08 l/l at the initiation of the climacteric, and up to 3.0 l/l at the climacteric peak. Burg and Burg (1962) reported that ethylene production rises when or before CO2 production rises in ripening mangoes, while Biale and Young (1981) included mangoes among fruits in which eth-ylene rises after CO2 production rises.
Only a small concentration of exogenous ethylene (≥0.005 l/l) is needed to initiate mango ripening (Wills et al., 2001). The small amount of ethylene in the fruit at harvest is suffi cient to initiate ripening (Burg and Burg, 1962). While fruit of ‘Amrapali’ and ‘Deshehari’ mangoes produce a measurable amount of ethylene during ripening (Reddy and Srivastava, 1999), ethylene production does not follow a climacteric pattern and two ethylene peaks (at the mature-green and full-ripe stages) were recorded. This is probably due to the way that ethylene was measured in the different fruit, and the lack of control exerted on maturity stages of fruit. In ‘Carabao’ mangoes, the peak of

J.K. Brecht and E.M. Yahia494
ethylene production occurs 110 days after fl ower initiation, and declines as fruit approached full maturity (Cua and Lizada, 1990). The content of 1-aminocyclopropane-1-carboxylic acid (ACC), the immediate precursor of ethylene, increases in different tissues (peel, outer and inner mesocarp) dur-ing ripening in both cultivars, ‘Amrapali’ and ‘Deshehari’, while ACC oxi-dase (ACO; EC 1.14.17.4), which catalyse the conversion of ACC to ethylene and ethylene production, decline (Reddy and Srivastava, 1999). Fruit peel has the highest levels of ethylene and ACO and less ACC accumulation than the outer and inner mesocarp at the mature-green stage. The inner mesocarp has less ACO activity and high ACC accumulation during the ripening pro-cess compared to peel; levels in the outer mesocarp are intermediate between those in the peel and inner mesocarp. Changes in the ability to convert ACC to ethylene in the peel are not related to changes in ripening parameters in the fruit pulp (Lederman et al., 1997). Mango seed also produces ethylene (Reddy and Srivastava, 1999). Fruit slicing has no measurable effect on ethyl-ene production in ‘Julia’ and ‘Graham’ mangoes (Allong et al., 2001).
Treatment of mango fruit with acetaldehyde or ethanol (0.1, 0.5 or 1% ethanol or acetaldehyde vapour) has concentration-dependent inhibitory effects on ethylene production (Burdon et al., 1996). Application of ACC to acetaldehyde- or ethanol-treated fruit discs indicates that acetaldehyde can completely eliminate increased ACO activity, whereas ethanol cannot (Bur-don et al., 1996). Accordingly, Burdon et al. (1996) suggested that acetalde-hyde can either inhibit ACO activity directly or prevent the increase in the enzyme, thereby providing a possible mechanism for retarding fruit ripening.
14.4 Compositional Changes during Fruit Maturation and Ripening
Several important metabolic changes occur during the maturation and ripen-ing of mangoes, and some of those are useful as maturity indices (Ketsa et al., 1991). The ripening changes are irreversible senescence processes that are related to degradation of organelles or changes in chemical constituents, and thus relate to the quality and postharvest life of the fruit. Natural senescence is aggravated and promoted by ethylene, mechanical injury and high tem-perature, and can be delayed by low temperature, elimination of mechanical damage and reduction of ethylene production.
Organic acids
Organic acids are important for respiratory activity and as fl avour constitu-ents. During maturation and ripening, mango fruit experience a substantial loss of organic acids. The predominant acids in mature mango fruit are citric, succinic, malic and tartaric acids; citric acid has the highest concentration and tartaric acid the lowest (Shashirekha and Patwardhan, 1976; Sarker and Muhsi, 1981; Medlicott and Thompson, 1985). Citric acid content increases steadily during fruit development in ‘Irwin’, reaching a maximum at the beginning of the endocarp-hardening period, and then decreases steadily

Postharvest Physiology 495
(Ito et al., 1997). In ‘Keitt’ the predominant organic acids are citric and malic acids, but tartaric, oxalic, ascorbic, and -ketoglutaric acids also are present, and the initial loss in acidity is due to a substantial loss in citric acid and a small loss in malic acid (Medlicott and Thompson, 1985). In ‘Badami’ man-goes, citric acid is the major organic acid, but malic and succinic acids are also present (Shashirekha and Patwardhan, 1976). In ‘Fazli’ mangoes, oxalic, citric, malic, pyruvic and succinic acids have been detected and tartaric acid has been detected in ‘Zardalu’ mangoes (Kumar et al., 1993). In general, citric and succinic acids decrease during ripening while malic acid shows different changes with different cultivars (Lizada, 1993).
Mango fruit contain organic acids involved in tricarboxylic acid cycle reactions (i.e. oxalic, succinic, pyruvic, oxaloacetic and -ketoglutaric acids). In ‘Pairi’ mangoes, maximum concentration of -oxoglutaric and pyruvic acids occur before the climacteric peak. Aspartic and glutamic acid concentrations increase for c.3 days after harvest and then decrease as the climacteric maxi-mum is reached (Krishnamurthy et al., 1971). Malic enzyme (EC 1.1.1.40), which catalyses the oxidative decarboxylation of L-malic to pyruvic acid, occurs in the three-quarter-ripe and ripe stages and the activity pattern during ripening is similar in ‘Alphonso’, ‘Banganpalli’, ‘Dasheri’, ‘Fazli’, ‘Langra’ and ‘Suvar-narekha’ (Selvaraj and Kumar, 1994). In ‘Alfonso’ (sic), the levels of malic dehy-drogenase (EC 1.1.1.37) and succinic dehydrogenase (EC 1.3.5.1) increase with the onset of ripening; whereas, the level of citrate synthase (EC 2.3.3.1) increases several-fold during maturation but decreases markedly during ripening (Baqui et al., 1974). The activity of malic enzyme increases during ripening, reaching a maximum immediately after the climacteric peak, and then declines (Dubery et al., 1984). The activity patterns of phosphoenol pyruvate carboxylase (PEPC; EC 4.1.1.49) and pyruvate decarboxylase (EC 4.1.1.1) during ripening vary among different cultivars, while malic enzyme activity increases during ripen-ing. PEPC activity is relatively high in ‘Alphonso’ and ‘Langara’, but low in ‘Dashehari’ and ‘Totapuri’ during ripening (Selvaraj and Kumar, 1994).
Soluble sugars
The increase in soluble sugars is a major change during mango fruit ripening, and sweetness is the most important compositional change related to mango fl avour. While starch content increases in chloroplasts during mango fruit development, it is almost completely hydrolysed to simple sugars during ripening (Medlicott et al., 1986; Selvaraj et al., 1989; Kumar et al., 1994; Ito et al., 1997). In ‘Alphonso’, starch content is 14% (by weight) at the immature stage and c.0.3% at the ripe stage. Similarly, starch is almost undetectable in ‘Irwin’ mangoes after ripening, whereas sucrose increases signifi cantly and fructose increases slightly (Ito et al., 1997). Starch content decreases slightly during ripening of ‘Haden’, but is insuffi cient to account for the observed increase in the level of sucrose (Castrillo et al., 1992).
Ripe mango contains up to 10–20% total sugars on a fresh weight (FW) basis, depending on the cultivar and the stage of ripeness. At the beginning

J.K. Brecht and E.M. Yahia496
of ripening, reducing sugars make up most of the sugar content, while there are more non-reducing (c.17%) than reducing (3%) sugars in completely ripe fruit. Sucrose contributes 57% of the total sugar in ripe ‘Keitt’ mangoes, with fructose and glucose making up 28% and 15%, respectively (Medlicott and Thompson, 1985). Although Krishnamurthy et al. (1971), Lakshminarayana (1973, 1975) and Shashirekha and Patwardhan (1976) reported a simultane-ous increase of glucose, fructose and sucrose during ripening, Vazquez-Salinas and Lakshminarayana (1985) observed a gradual reduction in glucose and fructose and a continuous increase of sucrose during ripening in ‘Haden’, ‘Irwin’, ‘Kent’ and ‘Keitt’. Medlicott and Thompson (1985) and Vazquez-Salinas and Lackshminarayana (1985) identifi ed the main reducing sugar as fruc-tose, while Selvaraj et al. (1989) reported that glucose is predominant. Con-fl icting reports on the relative concentrations of individual sugars in mango fruit during ripening is cultivar-dependent and due to different storage and handling conditions (Medlicott and Thompson, 1985).
Sucrose content increases during ripening as a result of starch hydrolysis from increased amylase (EC 3.2.1.1) activity (Mattoo and Modi, 1969a; Fuchs et al., 1980; Tandon and Kalra, 1983). The high activities of sucrose synthase (EC 2.4.1.13) and invertase (EC 3.2.1.26) in the mesocarp during ripening indicate active sucrose metabolism (Kumar et al., 1994). Hexoses and hexose phosphates can be formed from pyruvate by gluconeogenesis (Selvaraj and Kumar, 1994). The activity of glucose-6-phosphatase (EC 3.1.3.9) reportedly increases up to the three-quarter-ripe stage; whereas, fructose-1,6-diphosphatase (EC 3.1.3.11) activity increases as the fruit ripens from the three-quarter-ripe to full-ripe stage (Kumar and Selvaraj, 1990). The glycolytic enzyme hexokinase (6-phosphofructokinase; EC 2.7.1.11) has maximum activity at the ripe stage, while pyruvate kinase (EC 2.7.1.40) activity increases until the three-quarter-ripe stage and declines at ripening (Selvaraj and Kumar, 1994). The pattern of activity changes in hexokinase/phosphofructokinase and pyruvate kinase demonstrates that glycolysis is activated during mango fruit ripening.
Reducing sugars, mainly fructose, increase slightly during ripening, and sucrose synthase (EC 2.4.1.13) activity increases approximately ten times during the phase of rapid sucrose accumulation (Castrillo et al., 1992). This activity accounts for the maximum rate of sucrose synthesis. The proportion of sucrose phosphate synthase (EC 2.4.1.14) activity that is sensitive to inhibi-tion by inorganic phosphate changes during ripening (Castrillo et al., 1992). Maximum catalytic activity of sucrose synthase is constant throughout the ripening period and contributes signifi cantly to sucrose metabolism. The activities of neutral and acid invertases (EC 3.2.1.26) are very low in com-parison with the other enzymes of sucrose synthesis. Acid invertase activity increases and later decreases during ripening.
Structural polysaccharides
Pulp fi rmness is important for the evaluation of fruit maturity potential for transport and storage, and as a quality characteristic. Fruit softening and cell

Postharvest Physiology 497
wall changes are principal changes associated with fruit ripening. Fruit tex-ture changes are due to changes in cell walls and pectic substances in the middle lamella, and these are cultivar-related (Selvaraj and Kumar, 1989). Softening of mango fruit is characterized by increased solubility of cell wall pectins (Roe and Bruemmer, 1981; Tandon and Kalra, 1984; Lazan et al., 1986; Nasrijal, 1993). In general, water-soluble polysaccharides increase during ripening (Lazan et al., 1986; Brinson et al., 1988), but water- and alkali-soluble pectins decline in ‘Keitt’ mangoes, and ammonium oxalate-soluble pectins increase as the fruit become soft (Roe and Bruemmer, 1981). There is an over-all loss of galactosyl and deoxyhexosyl residues during mango fruit ripen-ing, the latter indicating degradation of the pectin component of the wall (Muda et al., 1995). The loss of galactose appears to be restricted to the chela-tor soluble fraction of the wall pectin, while loss of deoxyhexose seems to be more evenly distributed among the pectin.
Pectinesterase (PE; EC 3.1.1.11), which catalyses the deesterifi cation of methyl groups from acidic pectins, has been detected in ripening mangoes (Tahir and Malik, 1977; Roe and Bruemmer, 1981; Ali et al., 1990, 1995; Abu-Sarra and Abu-Goukh, 1992). Physiological maturity in ripened mangoes is associated with lower PE activity (van Lelyveld and Smith, 1979) and peel has higher PE activity than pulp (Ashraf et al., 1981). Endo-polygalacturonase (PG; EC 3.2.1.15), which is responsible for degrading the 1-4-linked galactur-onic acid residues, occurs in ripening fruit (Lazan et al., 1986, 1993; Abu-Sarra and Abu-Goukh, 1992). Enzymatic and/or non-enzymatic processes, in addi-tion to PG activity, are involved in the extensive softening of fruit (Mitcham and McDonald, 1992). Other cell wall hydrolases can be detected in ripening fruit, including cellulases (EC 3.2.1.4; Lazan et al., 1986; Abu-Sarra and Abu-Goukh, 1992), -galactosidase (EC 3.2.1.23; Ali et al., 1990, 1995; Lazan et al., 1993), galactanase (EC 3.2.1.145; Ali et al., 1990) and xylanase (EC 3.2.1.8; Ali et al., 1990).
Ripening in mangoes, as characterized by decreased tissue fi rmness, is initiated in inner mesocarp tissue close to the seed, and progresses outwards (Lazan et al., 1993). Pectin solubilization in inner and outer mesocarp tissues is comparable, but pectin solubilization begins earlier in the inner than in the outer mesocarp (Lazan et al., 1993). The outer mesocarp of ‘Keitt’ remains fi rm longer than ‘Tommy Atkins’, and the inner is softer than the outer meso-carp at each stage of ripening in both cultivars (Mitcham and McDonald, 1992). Cell wall neutral sugars, particularly arabinosyl, rhamnosyl and galac-tosyl residues, decrease with ripening in both cultivars. ‘Keitt’ has more loosely associated, chelator-soluble pectin, accumulates more soluble poly-uronides and retains more total pectin at the ripe stage than ‘Tommy Atkins’. Both cultivars have similar PG activity, which increases with ripening. The amount and molecular weight (MW) of cell wall hemicellulose decreases with ripening in both cultivars. Galactose is the only cell wall neutral sugar to show a signifi cant decrease during ripening of ‘Sensation’ mangoes (Sey-mour et al., 1990). Losses of neutral sugars can be due to hydrolysis of galac-tans and arabinogalactans by -galactosidase having galactanase activity.
-Galactosidase activity shows a parallel increase to tissue softening during

J.K. Brecht and E.M. Yahia498
ripening. The close correlations between changes in -galactosidase during rip-ening, and between changes in -galactosidase activity with tissue softening, and increased pectin solubility and degradation suggests that -galactosidase has an important role in cell-wall pectin modifi cation and mango fruit soft-ening during ripening (Ali et al., 1995).
Postharvest treatments, such as refrigeration, packaging, application of fruit coatings, etc., can retard mango fruit softening and activity of pectinases (Lazan et al., 1990; Nasrijal, 1993). Calcium (Ca) joins free carboxyl groups resulting from PE-catalysed hydrolysis of methyl ester bonds to form Ca-bridges between adjacent pectin molecules. Calcium application to ‘Haden’ mangoes by infi ltration or dipping extends their storage life by 1 week (Zam-brano and Manzano, 1995). Postharvest vacuum application of Ca to mango has also been tried (Tirmazi and Wills, 1981; Wills et al., 1988; van Eeden, 1992; Yuen et al., 1993). Vacuum (300 mm Hg) infi ltration of 1–4% calcium chloride (CaCl2) into ‘Amrapali’ and ‘Deashehari’ mangoes ripened at 25°C inhibits PG activity, while ethylene treatment (1 l/l) markedly increases its activity (Reddy and Srivastava, 1999). Pressure (115 kPa for 2 min) or vacuum infi ltration (32 kPa) with 1–8% CaCl2 delays ripening of ‘Kensington Pride’ mangoes by 12 or 8 days, respectively (Yuen et al., 1993). Yuen et al. (1993) reported that vacuum infi ltration of CaCl2 causes some peel injury, which can be reduced by: (i) increasing the temperature of the fruit fl esh or the CaCl2 solution during pressure infi ltration; (ii) packaging the fruit in sealed polyethylene during pressure infi ltration; and (iii) packaging the fruit in sealed polyethylene bags or cling or shrink wraps after CaCl2 treatment. Cal-cium chloride infi ltration of ‘Keitt’ mangoes reduces ethylene production, respiration rate and the incidence of storage decay (van Eeden, 1992).
Pigments and colour
Mango skin colour is important for its role in the perception of overall qual-ity (González-Aguilar et al., 2001) and can be important for determining the appropriate maturity for harvesting (Cocozza et al., 2004; Jha et al., 2007), pro-cessing (Mahayothee et al., 2004) and consumption (Cocozza et al., 2004; Jha et al., 2007). The loss of green colour is an obvious sign of fruit ripening in many mango cultivars. The development of the optimum skin colour usually defi nes mango quality. Some mango cultivars retain green colour in ripe fruit. Depending on the cultivar, skin colour can change from dark to olive-green; sometimes reddish, orange-yellow or yellowish hues appear from the base colour. Some cultivars develop a reddish blush, which has been attrib-uted to anthocyanins. Colour changes in mango fruit are due to the disap-pearance of chlorophyll and the appearance of other pigments (Fig. 14.4). Chloroplasts are transformed to chromoplasts containing yellow or red pig-ments (John et al., 1970; Lakshminarayana, 1980; Parikh et al., 1990; Lizada, 1993). Well-arranged grana and osmiophilic globules occur in chloroplasts of cells in the peel of unripe mangoes (Parikh et al., 1990), and lose integrity during ripening. Osmiophilic globules appear, indicating the transformation
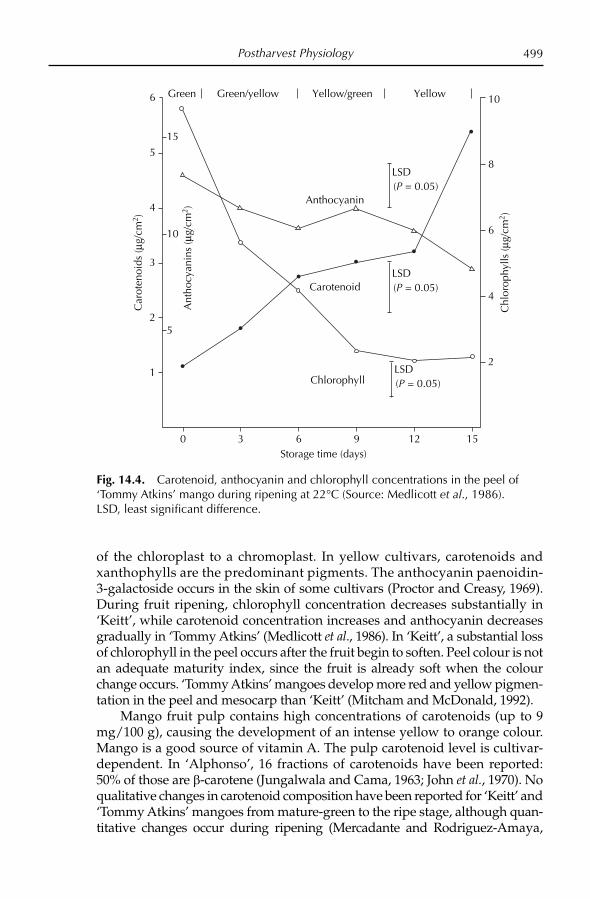
Postharvest Physiology 499
of the chloroplast to a chromoplast. In yellow cultivars, carotenoids and xanthophylls are the predominant pigments. The anthocyanin paenoidin-3-galactoside occurs in the skin of some cultivars (Proctor and Creasy, 1969). During fruit ripening, chlorophyll concentration decreases substantially in ‘Keitt’, while carotenoid concentration increases and anthocyanin decreases gradually in ‘Tommy Atkins’ (Medlicott et al., 1986). In ‘Keitt’, a substantial loss of chlorophyll in the peel occurs after the fruit begin to soften. Peel colour is not an adequate maturity index, since the fruit is already soft when the colour change occurs. ‘Tommy Atkins’ mangoes develop more red and yellow pigmen-tation in the peel and mesocarp than ‘Keitt’ (Mitcham and McDonald, 1992).
Mango fruit pulp contains high concentrations of carotenoids (up to 9 mg/100 g), causing the development of an intense yellow to orange colour. Mango is a good source of vitamin A. The pulp carotenoid level is cultivar-dependent. In ‘Alphonso’, 16 fractions of carotenoids have been reported: 50% of those are -carotene (Jungalwala and Cama, 1963; John et al., 1970). No qualitative changes in carotenoid composition have been reported for ‘Keitt’ and ‘Tommy Atkins’ mangoes from mature-green to the ripe stage, although quan-titative changes occur during ripening (Mercadante and Rodriguez-Amaya,
6
5
15
Green Green/yellow Yellow/green Yellow
Anthocyanin
LSD(P = 0.05)
LSD(P = 0.05)
LSD(P = 0.05)
Carotenoid
Chlorophyll
10
5
4
3
Car
oten
oids
(μg/
cm2 )
Chl
orop
hylls
(μg/
cm2 )
Ant
hocy
anin
s (μ
g/cm
2 )
2
1
10
8
6
4
2
0 3 6Storage time (days)
9 12 15
Fig. 14.4. Carotenoid, anthocyanin and chlorophyll concentrations in the peel of ‘Tommy Atkins’ mango during ripening at 22°C (Source: Medlicott et al., 1986).LSD, least signifi cant difference.
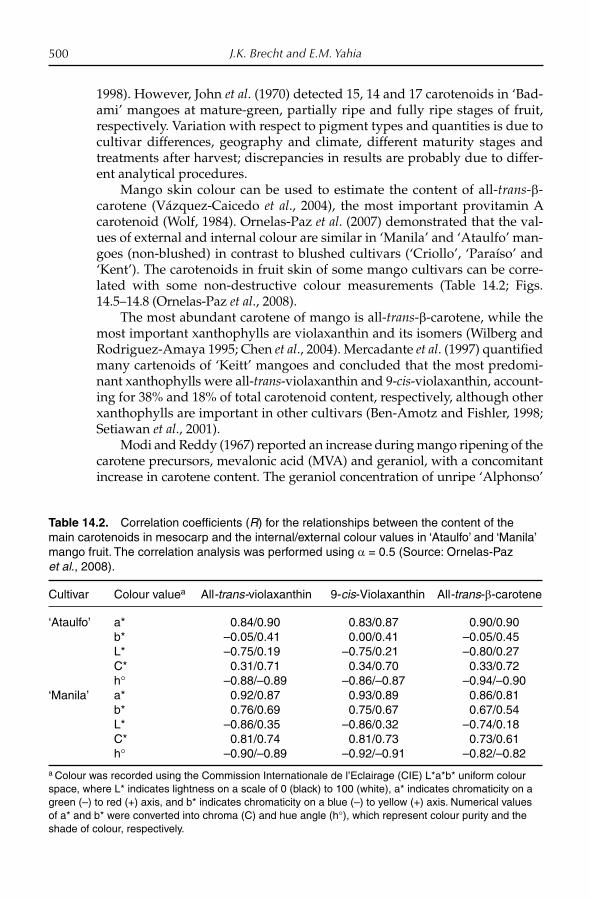
J.K. Brecht and E.M. Yahia500
1998). However, John et al. (1970) detected 15, 14 and 17 carotenoids in ‘Bad-ami’ mangoes at mature-green, partially ripe and fully ripe stages of fruit, respectively. Variation with respect to pigment types and quantities is due to cultivar differences, geography and climate, different maturity stages and treatments after harvest; discrepancies in results are probably due to differ-ent analytical procedures.
Mango skin colour can be used to estimate the content of all-trans- -carotene (Vázquez-Caicedo et al., 2004), the most important provitamin A carotenoid (Wolf, 1984). Ornelas-Paz et al. (2007) demonstrated that the val-ues of external and internal colour are similar in ‘Manila’ and ‘Ataulfo’ man-goes (non-blushed) in contrast to blushed cultivars (‘Criollo’, ‘Paraíso’ and ‘Kent’). The carotenoids in fruit skin of some mango cultivars can be corre-lated with some non-destructive colour measurements (Table 14.2; Figs. 14.5–14.8 (Ornelas-Paz et al., 2008).
The most abundant carotene of mango is all-trans- -carotene, while the most important xanthophylls are violaxanthin and its isomers (Wilberg and Rodriguez-Amaya 1995; Chen et al., 2004). Mercadante et al. (1997) quantifi ed many cartenoids of ‘Keitt’ mangoes and concluded that the most predomi-nant xanthophylls were all-trans-violaxanthin and 9-cis-violaxanthin, account-ing for 38% and 18% of total carotenoid content, respectively, although other xanthophylls are important in other cultivars (Ben-Amotz and Fishler, 1998; Setiawan et al., 2001).
Modi and Reddy (1967) reported an increase during mango ripening of the carotene precursors, mevalonic acid (MVA) and geraniol, with a concomitant increase in carotene content. The geraniol concentration of unripe ‘Alphonso’
Table 14.2. Correlation coeffi cients (R) for the relationships between the content of the main carotenoids in mesocarp and the internal/external colour values in ‘Ataulfo’ and ‘Manila’ mango fruit. The correlation analysis was performed using = 0.5 (Source: Ornelas-Paz et al., 2008).
Cultivar Colour valuea All-trans-violaxanthin 9-cis-Violaxanthin All-trans- -carotene
‘Ataulfo’ a* 0.84/0.90 0.83/0.87 0.90/0.90b* –0.05/0.41 0.00/0.41 –0.05/0.45L* –0.75/0.19 –0.75/0.21 –0.80/0.27C* 0.31/0.71 0.34/0.70 0.33/0.72h° –0.88/–0.89 –0.86/–0.87 –0.94/–0.90
‘Manila’ a* 0.92/0.87 0.93/0.89 0.86/0.81b* 0.76/0.69 0.75/0.67 0.67/0.54L* –0.86/0.35 –0.86/0.32 –0.74/0.18C* 0.81/0.74 0.81/0.73 0.73/0.61h° –0.90/–0.89 –0.92/–0.91 –0.82/–0.82
a Colour was recorded using the Commission Internationale de l’Eclairage (CIE) L*a*b* uniform colour space, where L* indicates lightness on a scale of 0 (black) to 100 (white), a* indicates chromaticity on a green (–) to red (+) axis, and b* indicates chromaticity on a blue (–) to yellow (+) axis. Numerical values of a* and b* were converted into chroma (C) and hue angle (h°), which represent colour purity and the shade of colour, respectively.
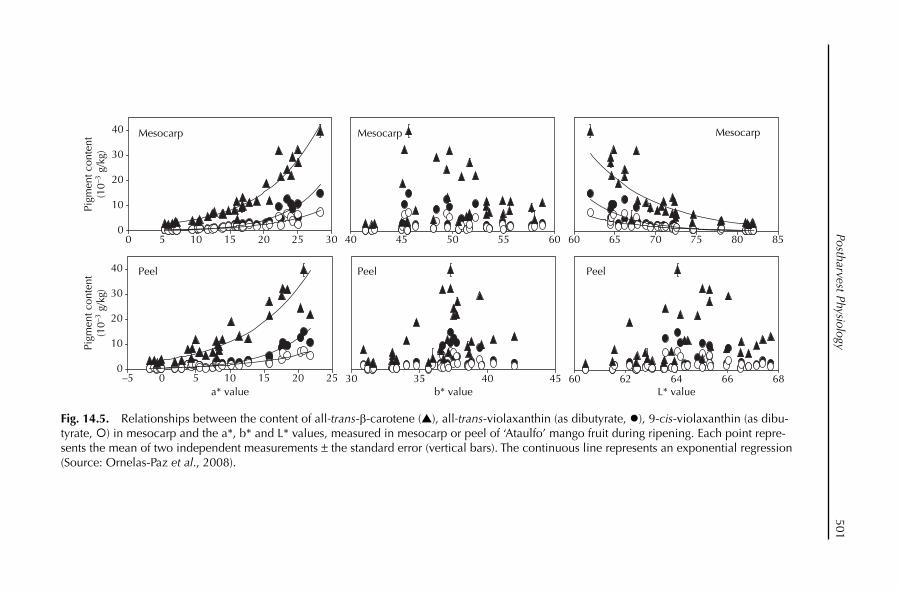
Postharvest Physiology501
30 35 40 45
0
10
20
30
40
0 5 10 15 20 25 30 40 45 50 55 60 60 65 70 75 80 85
0
10
20
30
40
–5 0 5 10 15 20 25
Pigm
ent c
onte
nt(1
0–3 g
/kg)
Pigm
ent c
onte
nt(1
0–3 g
/kg)
60 62 64 66 68
Mesocarp Mesocarp Mesocarp
Peel PeelPeel
a* value b* value L* value
Fig. 14.5. Relationships between the content of all-trans- -carotene (▲), all-trans-violaxanthin (as dibutyrate, ), 9-cis-violaxanthin (as dibu-tyrate, ) in mesocarp and the a*, b* and L* values, measured in mesocarp or peel of ‘Ataulfo’ mango fruit during ripening. Each point repre-sents the mean of two independent measurements ± the standard error (vertical bars). The continuous line represents an exponential regression (Source: Ornelas-Paz et al., 2008).
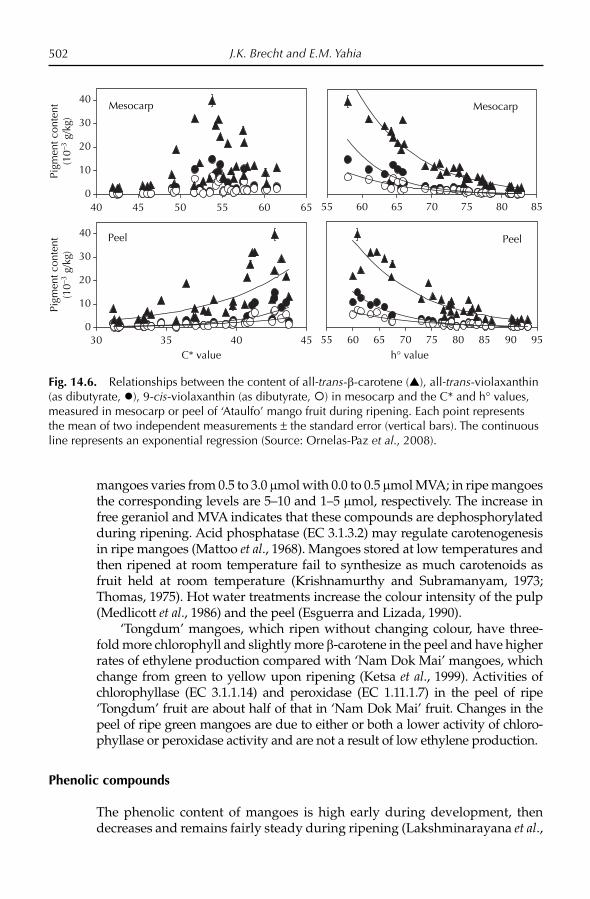
J.K. Brecht and E.M. Yahia502
mangoes varies from 0.5 to 3.0 mol with 0.0 to 0.5 mol MVA; in ripe mangoes the corresponding levels are 5–10 and 1–5 mol, respectively. The increase in free geraniol and MVA indicates that these compounds are dephosphorylated during ripening. Acid phosphatase (EC 3.1.3.2) may regulate carotenogenesis in ripe mangoes (Mattoo et al., 1968). Mangoes stored at low temperatures and then ripened at room temperature fail to synthesize as much carotenoids as fruit held at room temperature (Krishnamurthy and Subramanyam, 1973; Thomas, 1975). Hot water treatments increase the colour intensity of the pulp (Medlicott et al., 1986) and the peel (Esguerra and Lizada, 1990).
‘Tongdum’ mangoes, which ripen without changing colour, have three-fold more chlorophyll and slightly more -carotene in the peel and have higher rates of ethylene production compared with ‘Nam Dok Mai’ mangoes, which change from green to yellow upon ripening (Ketsa et al., 1999). Activities of chlorophyllase (EC 3.1.1.14) and peroxidase (EC 1.11.1.7) in the peel of ripe ‘Tongdum’ fruit are about half of that in ‘Nam Dok Mai’ fruit. Changes in the peel of ripe green mangoes are due to either or both a lower activity of chloro-phyllase or peroxidase activity and are not a result of low ethylene production.
Phenolic compounds
The phenolic content of mangoes is high early during development, then decreases and remains fairly steady during ripening (Lakshminarayana et al.,
0
10
20
30
40
40 45 50 55 60 65 55 60 65 70 75 80 85
0
10
20
30
40
30 35 40 45 55 60 65 70 75 80 85 90 95
MesocarpMesocarp
C* value
Peel Peel
h° value
Pigm
ent c
onte
nt(1
0–3 g
/kg)
Pigm
ent c
onte
nt(1
0–3 g
/kg)
Fig. 14.6. Relationships between the content of all-trans- -carotene (▲), all-trans-violaxanthin(as dibutyrate, ), 9-cis-violaxanthin (as dibutyrate, ) in mesocarp and the C* and h° values, measured in mesocarp or peel of ‘Ataulfo’ mango fruit during ripening. Each point represents the mean of two independent measurements ± the standard error (vertical bars). The continuous line represents an exponential regression (Source: Ornelas-Paz et al., 2008).
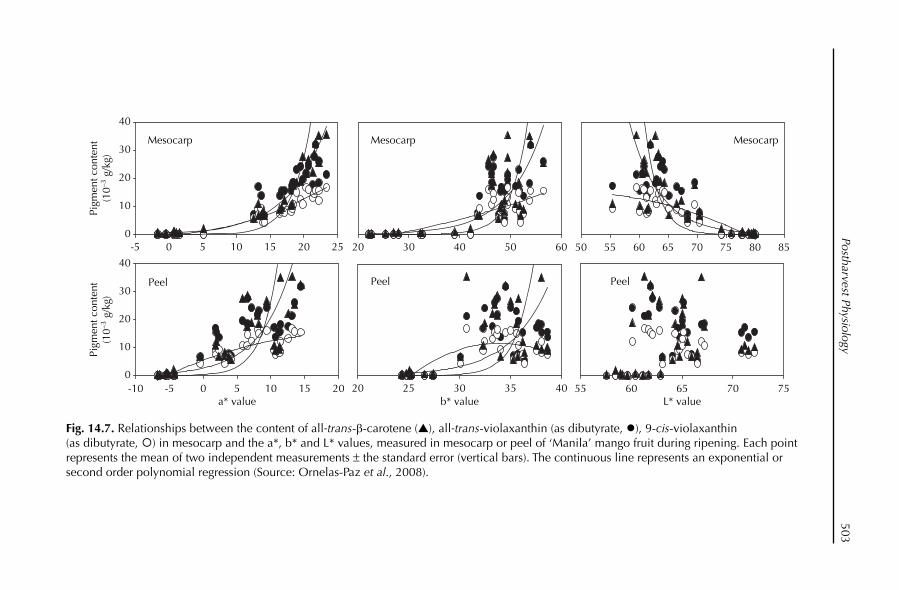
Postharvest Physiology503
0
10
20
30
40
-5 0 5 10 15 20 25 20 30 40 50 60 50 55 60 65 70 75 80 85
0
10
20
30
40
-10 -5 0 5 10 15 20 20 25 30 35 40 55 60 65 70 75
Mesocarp Mesocarp
Peel
a* value b* value L* value
Mesocarp
Peel Peel
Pigm
ent c
onte
nt(1
0–3 g
/kg)
Pigm
ent c
onte
nt(1
0–3 g
/kg)
Fig. 14.7. Relationships between the content of all-trans- -carotene (▲), all-trans-violaxanthin (as dibutyrate, ), 9-cis-violaxanthin(as dibutyrate, ) in mesocarp and the a*, b* and L* values, measured in mesocarp or peel of ‘Manila’ mango fruit during ripening. Each point represents the mean of two independent measurements ± the standard error (vertical bars). The continuous line represents an exponential or second order polynomial regression (Source: Ornelas-Paz et al., 2008).
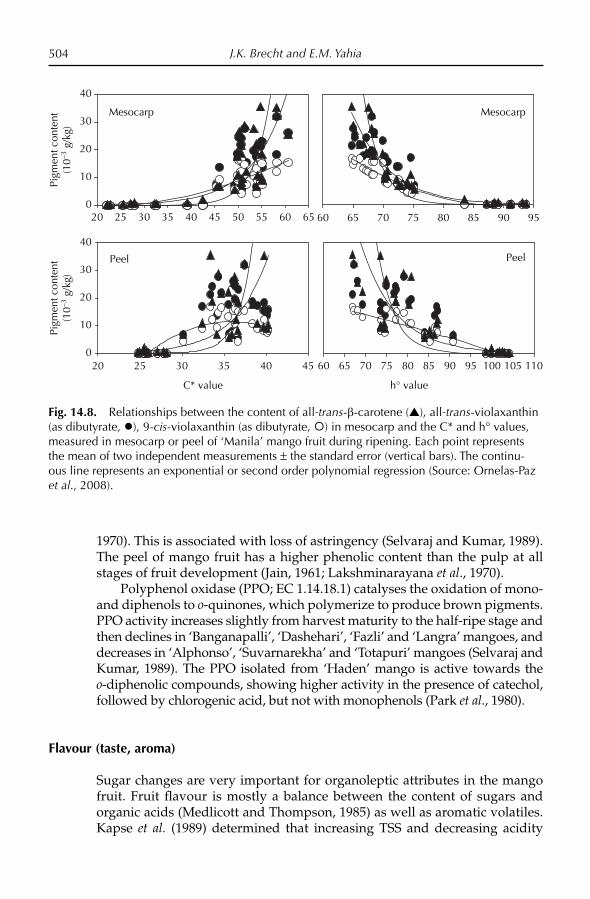
J.K. Brecht and E.M. Yahia504
1970). This is associated with loss of astringency (Selvaraj and Kumar, 1989). The peel of mango fruit has a higher phenolic content than the pulp at all stages of fruit development (Jain, 1961; Lakshminarayana et al., 1970).
Polyphenol oxidase (PPO; EC 1.14.18.1) catalyses the oxidation of mono- and diphenols to o-quinones, which polymerize to produce brown pigments. PPO activity increases slightly from harvest maturity to the half-ripe stage and then declines in ‘Banganapalli’, ‘Dashehari’, ‘Fazli’ and ‘Langra’ mangoes, and decreases in ‘Alphonso’, ‘Suvarnarekha’ and ‘Totapuri’ mangoes (Selvaraj and Kumar, 1989). The PPO isolated from ‘Haden’ mango is active towards the o-diphenolic compounds, showing higher activity in the presence of catechol, followed by chlorogenic acid, but not with monophenols (Park et al., 1980).
Flavour (taste, aroma)
Sugar changes are very important for organoleptic attributes in the mango fruit. Fruit fl avour is mostly a balance between the content of sugars and organic acids (Medlicott and Thompson, 1985) as well as aromatic volatiles. Kapse et al. (1989) determined that increasing TSS and decreasing acidity
C* value
60 65 70 75 80 85 90 95
0
10
20
30
40
20 25 30 35 40 45 60 65 70 75 80 85 90 95 100 105 110
Mesocarp
0
10
20
30
40
20 25 30 35 40 45 50 55 60 65
Mesocarp
Peel Peel
h° value
Pigm
ent c
onte
nt(1
0–3 g
/kg)
Pigm
ent c
onte
nt(1
0–3 g
/kg)
Fig. 14.8. Relationships between the content of all-trans- -carotene (▲), all-trans-violaxanthin(as dibutyrate, ), 9-cis-violaxanthin (as dibutyrate, ) in mesocarp and the C* and h° values, measured in mesocarp or peel of ‘Manila’ mango fruit during ripening. Each point represents the mean of two independent measurements ± the standard error (vertical bars). The continu-ous line represents an exponential or second order polynomial regression (Source: Ornelas-Paz et al., 2008).

Postharvest Physiology 505
increases fl avour ratings of mango fruit. Sucrose is the predominant sugar in ripe mango fruit (Tandon and Kalra, 1983; Medlicott and Thomson, 1985; Vazquez-Salinas and Lakshminarayana, 1985). The predominant acid in mango fruit is citric (Medlicott and Thompson, 1985; Lizada, 1993). Several factors affect sugar and acid contents in mango, including cultivar (Kapse et al., 1989; Kundu and Ghosh, 1992; Gowda et al., 1994), stage of maturity at harvest (Shashirekha and Patwardhan, 1976; Morga et al., 1979; Tandon and Kalra, 1983), postharvest treatments (Kumar et al., 1993) and storage conditions (Vazquez-Salinas and Lakshminarayana, 1985).
Ripe mangoes contain >300 volatiles (Pino et al., 2005), but not all of them are odour-active and thus do not contribute signifi cantly to aroma. Studies have identifi ed the volatiles of mango, but not their aromatic activity. The predominant volatiles in some cultivars are monoterpenes and sesquiter-penes (MacLeod and De Troconis, 1982; Engel and Tressl, 1983; Pino et al., 2005), as well as lactones and some fatty acids (MacLeod and Pieris, 1984; MacLeod and Snyder, 1985; Wilson et al., 1990). However, there is no indica-tion of the presence of a single fl avour impact component (Engel and Tressl, 1983). Some mango cultivars have a peach-like fl avour that may be related to the presence of lactones, which contribute to the fl avour of peaches (Prunuspersica) (Lakshminarayana, 1980; MacLeod et al., 1988, Wilson et al., 1990). MacLeod et al. (1988) detected four lactones in ‘Kensington Pride’ that are also the major volatiles of peach. Monoterpene hydrocarbons represent about 49% (w/w) of the total volatiles in ‘Kensington Pride’, with -terpinolene being the most abundant (26%) and 16 esters representing 33% (MacLeod et al., 1988). The esters, together with some of the lactones, contribute to the fl avour of ‘Kensington Pride’ mangoes.
Indian mangoes have a unique fl avour, which has been attributed to (Z)-ocimine (Engel and Tressl, 1983; Lizada, 1993). Pino et al. (1989) detected 83 volatiles in ‘Corazon’, ‘Bizcochuelo’ and ‘Super Haden’ mangoes, and total volatiles ranged between 39 mg/kg in ‘Bizcochuelo’ to 70 mg/kg in ‘Corazon’. The identifi ed volatiles include -cubebene, -maaliene, ethyl(Z)-9-hexade-canoate, ethyl(Z)-9,12-octadecanoate, ethyl(Z)(Z)(Z)-6,9,12-octadecanoate, cucarvone, 2-methylpropane-2-ol, 3-methylepentan-ol, thymol and carvacrol (Pino et al., 1989). MacLeod and Snyder (1985) listed the volatile components of several mango cultivars, including ‘Willard’ and ‘Parrot’ from Sri Lanka; levels of -terpinolene were similar to ‘Kensington Pride’.
Kostermans and Bompard (1993) considered that lack of fi bre was linked to an absence of aroma and fl at taste and smell, but some cultivars such as ‘Kensington Pride’ are low in fi bre and have a distinctive fl avour and aroma profi le, and a high level of -terpinolene (Bartley and Schwede, 1987; MacLeod et al., 1988). Lipid content of the pulp is correlated with the fl avour character-istics of some mango cultivars (Bandyopadhyay and Gholap, 1973a; Gholap and Bandyopadhyay, 1975b, 1976). The ripening of ‘Alphonso’ mangoes at ambient temperature is accompanied by a sharp increase in triglyceride con-tent, together with the development of a strong aroma and fl avour (Gholap and Bandyopadhyay, 1975a, 1976), but ripening at 10°C results in a bland aroma and fl avour (Bandyopadhyay and Gholap, 1973b). ‘Totapuri’ mangoes,

J.K. Brecht and E.M. Yahia506
a bland cultivar, showed no change in the development of aroma or in the pulp lipid content (Gholap and Bandyopadhyay, 1975b). During ripening at ambient temperature, palmitoleic acid content is higher than that of palmitic acid in ‘Alphonso’, whereas ripening at low temperature does not affect the proportions of these two fatty acids (Bandyopadhyay and Gholap, 1973b). The relative proportions of palmitoleic and palmitic acids in ‘Totapuri’ mango pulp are constant irrespective of the ripening conditions (Gholap and Ban-dyopadhyay, 1975b). Gholap and Bandyopadhyay (1976, 1980) suggested that the relative contents of palmitic and palmitoleic acids determine the fl a-vour quality of mango fruit.
The absence of lactones having coconut-like odour notes in ‘Totapuri’ mangoes may be signifi cant for differentiating its aroma characteristics from ‘Alphonso’, together with the presence of certain similar and dissimilar com-ponents (Bandyopadhyay, 1983). The aroma of green mangoes has been attributed to cis-ocimine in ‘Alphonso’ and -myrcene in ‘Batali’ mangoes (Gholap and Bandyopadhyay, 1976; Bandyopadhyay, 1983). Table 14.3 lists characteristic aromas of ‘Alphonso’ and ‘Totapuri’ mangoes and their possi-ble chemical identities.
In almost all fruits, aromatic volatiles are produced at later stages of rip-ening (Yahia, 1994). Tree-ripe ‘Tommy Atkins’ mangoes produce higher lev-els of all aroma volatiles except hexanal than do mature-green fruit (Bender et al., 2000a). Both mature-green and tree-ripe mangoes stored in 25 kPa CO2 tend to have lower terpene (especially p-cymene) and hexanal concentra-tions than those stored in 10 kPa CO2 and air-stored fruit. Acetaldehyde and ethanol levels tend to be higher in tree-ripe mangoes held in 25 kPa CO2 than in those from 10 kPa CO2 or air storage, especially at 8°C. Inhibition of volatile production by 25 kPa CO2 is greater in mature-green than in tree-ripe
Table 14.3. Characteristic aromas in ‘Alphonso’ and ‘Totapuri’ mangoes and their possible chemical causes (Source: Bandyopadhyay, 1983).
Aroma ‘Alphonso’ ‘Totapuri’
Fruit, estery Acetaldehyde, methyl acetate, ethyl acetate, n-butyl acetate
PropionaldehydeMethyl acetate
Green-mango-like cis-Ocimine -MyrceneCamphoraceous Not detected Detected, but not identifi edEarthy Caryophyllene-pinene Not detectedAlmond-like Benzaldehyde Not detectedBurnt-sugar-like Benzonitrile Not detectedSpicy Not detected -TerpineneSweet, sugar-like Detected, but not identifi ed Not detectedCoconut oil-like -Caprolactone, -octalactone,
-undecalactoneNot detected

Postharvest Physiology 507
mangoes, and at 8°C compared to 12°C for tree-ripe fruit. However, aroma volatile levels in tree-ripe mangoes from 25 kPa CO2 are equal to or greater than those in mature-green fruit treatments. Atmospheres that prolong mango shelf life by slowing ripening processes can allow tree-ripe mangoes to be stored or shipped without sacrifi cing their aroma quality.
Quality enhancement has been used to determine properties critical to fl a-vour acceptability of mangoes, and focus group interviews have been conducted to determine sensory attributes important to the purchase and consumption of mangoes (Malundo, 1996). Sugars and acids enhance perception of specifi c fl avour notes in mango, including aromatics (Malundo et al., 2001).
14.5 Transpiration and Water Loss
Water loss lowers fruit weight, resulting in shrivelling, and may further reduce quality by causing poor colour development and uneven ripening. Water is lost from mango fruit through stomata, lenticels and other openings. Relative humidity (RH) inside the fruit is 100% and water is lost when RH in the envi-ronment surrounding the fruit is <100%. Water loss is also greatly infl uenced by temperature. With constant RH and air movement, water loss increases signifi -cantly with any increase in temperature. Transpiration rate is infl uenced by cul-tivar and ripeness stage. It is correlated with skin thickness, morphological structure, epidermal cells and surface wax coating. For example, waxes usually develop on the epidermis of fruit in the later stages of development and thus it is common for fruit harvested early to shrivel faster compared with those har-vested at a more advanced stage of development (Yahia et al., 2006a).
14.6 Physical Damage and Physiological Disorders
Mangoes are susceptible to physical damage at every step of the postharvest handling chain (see Johnson and Hofman, Chapter 15, this volume) and reduction/elimination of mechanical injury is essential to reduce losses in quality and postharvest life. Mango fruit are susceptible to various physio-logical disorders that infl uence fruit quality (see Galán Saúco, Chapter 9, this volume). These disorders are either induced or inherent, and several of them become apparent during fruit ripening. Disorders, i.e. chilling injury (CI) and heat injury, may be induced after harvest. Inherent physiological disorders include ‘spongy stem end’ in ‘Kensington Pride’ (Brown et al., 1981), ‘soft nose’ in Florida mangoes (Young, 1957) and ‘internal breakdown’, ‘spongy tissue’ or ‘soft nose’ in Indian ‘Alphonso’ mangoes (Subramanyam et al., 1971).
Chilling injury (CI)
Low storage temperatures can injure mature-green mangoes if storage exceeds a day or so at or near 0°C to a few weeks at just below 12°C. This problem limits the use of low storage temperature to manage postharvest ripening

J.K. Brecht and E.M. Yahia508
and seriously affects the ability of handlers to store mangoes or transport them over long distances, because temperatures that are low enough to delay ripening, decay and senescence may damage the fruit. The symptoms of CI include greyish, scald-like discoloration on the skin, followed by pitting, uneven ripening, and poor fl avour, aroma and colour development (Hatton et al., 1965; Medlicott et al., 1990). The symptoms often are not apparent at the low temperature, but develop later, when the fruit are brought to warmer temperatures for ripening or are displayed for sale. Other symptoms in mango fruit held at room temperature for 1–2 days after low temperature storage were described as discoloured and with pitted areas on the surface (Srivastava, 1967; Kane, 1977) followed by increased susceptibility to micro-bial spoilage (Sadasivam et al., 1971; Subramanyam et al., 1975).
Chilling susceptibility varies with cultivar (Farooqui et al., 1985); ‘Haden’ and ‘Keitt’ are particularly susceptible. ‘Sensation’ developed more skin symptoms than ‘Sammar Bahisht’ mangoes (Farooqui et al., 1985). While CI has generally been reported to occur in mango fruit at temperatures below c.10–13°C (Mukherjee, 1958; Akamine, 1963; Hatton et al., 1965; Musa, 1974; Couey, 1986), some cultivars (i.e. ‘Dashehari’ and ‘Langara’) were reported to be safely stored at 7–8°C for up to 25 days (Mann and Singh, 1976). While most cultivars show injury at <10°C if fruit have just reached maturity, toler-ance of CI increases as fruit ripen (Medlicott et al., 1990; Mohammed and Brecht, 2002). Tolerance of ‘Keitt’ mango fruit of CI was induced by pre-storage heat treatments (McCollum et al., 1993).
Heat injury
Mango is highly tolerant of heat (Yahia et al., 2000; Jacobi et al., 2001b). Man-goes that are not stored in refrigerated conditions after harvest may be exposed to extremely high ambient temperatures in many production areas. This may lead to heat injury, especially if the fruit are exposed to >30°C for >10 days, but injury can also occur more rapidly at higher temperatures. The heat disinfestation treatments of mangoes that are required for insect quar-antine security may injure fruit that are not fully mature (Jacobi and Giles, 1997; Jacobi et al., 2001a).
External symptoms of heat injury include lenticel spotting and skin browning (‘scald’) with secondary disease development, while internal symptoms include mesocarp browning, tissue cavitation and ‘starch spots’ (Jacobi and Wong, 1992; Jacobi and Giles, 1997; Mitcham and McDonald, 1997; Jacobi et al., 2001a, b). Ripening of heat-injured mangoes may also be inhibited (Jacobi et al., 2001a, b).
14.7 Modifi ed Atmospheres (MA) and Controlled Atmospheres (CA)
Long-term marine shipping in MA and CA has been used for transit from several countries (Yahia, 1993). Research results are very contradictory due to

Postharvest Physiology 509
the different cultivars and maturity stages of mangoes used, different atmo-spheres implemented and lack of experimental controls. Optimum condition for prolonged shipping or storage is reported to be 3–5 kPa O2 plus 5–10 kPa CO2, which can delay ripening, but the benefi ts are not very signifi cant. Use of CA and MA would most likely be benefi cial in delaying fruit ripening during marine transport for 2 weeks or more.
Bender et al. (2000b) determined the tolerance of preclimacteric ‘Haden’ and ‘Tommy Atkins’ to reduced O2 levels for storage times in typical marine shipments. They reported that mangoes can tolerate 3 kPa O2 for 2–3 weeks at 12–15°C and that tolerance of low O2 decreases as mangoes ripen. All low O2 treatments reduced mature-green mango respiration; however, elevated ethanol production occurred in 2 and 3 kPa O2 storage, with the levels two to threefold higher in ‘Tommy Atkins’ than in ‘Haden’. ‘Haden’ fruit at the onset of the climacteric accumulated ethanol in 4 kPa O2 and produced 10–20 times more ethanol in 2 and 3 kPa O2 than preclimacteric fruit. There were no visible injury symptoms, but off-fl avour developed in mature-green fruit at 2 kPa O2 and in ripening-initiated fruit at 2 and 3 kPa O2. Ethanol production was not affected by storage in 25 kPa CO2. Ethylene production was reduced slightly by low O2; however, ‘Haden’ fruit also showed a residual inhibitory effect on ethylene production at 2 or 3 kPa O2 storage, while ‘Tommy Atkins’ fruit stored in 2 kPa O2 produced a burst of ethylene upon transfer to air at 20°C. Fruit fi rmness, total sugars and starch levels did not differ among treat-ments, but 2, 3 or 4 kPa O2 and 25 kPa CO2 maintained signifi cantly higher acidity than 5 kPa O2 or air. The epidermal ground colour responded differ-ently to low O2 and high CO2 in the two cultivars. Only 2 kPa O2 maintained ‘Haden’ colour better than air, while all low O2 levels maintained ‘Tommy Atkins’ colour better than air. High CO2 was more effective than low O2 in maintaining ‘Haden’ colour, but had about the same effect as low O2 on ‘Tommy Atkins’.
Properly selected atmospheres, which prolong mango shelf life by slow-ing ripening processes, can allow tree-ripe mangoes to be stored or shipped without sacrifi cing their superior aroma. Mature-green and tree-ripe ‘Tommy Atkins’ mangoes were stored for 21 days in air or in a CA (5 kPa O2 + 10 kPa or 25 kPa CO2) at 12°C (mature-green) and at either 8 or 12°C (tree-ripe) (Bender et al., 2000a). Tree-ripe mangoes produced much higher levels of all aroma volatiles except hexanal than mature-green fruit after ripening for 2 days. Both mature-green and tree-ripe mangoes stored in 25 kPa CO2 had lower terpene (especially p-cymene) and hexanal levels than those stored in 10 kPa CO2 and air-stored fruit. Acetaldehyde and ethanol levels were higher in tree-ripe mangoes from 25 kPa CO2 than in those from 10 kPa CO2 or air storage, especially at 8°C. Inhibition of volatile production by 25 kPa CO2 was greater in mature-green than in tree-ripe mangoes, and at 8°C com-pared to 12°C for tree-ripe fruit. Aroma volatile levels in tree-ripe mangoes from the 25 kPa CO2 treatment equalled or exceeded those in mature-green fruit treatments.
Mangoes have high tolerance of short-term elevated CO2 atmospheres (Yahia, 1998). Mangoes can tolerate CO2 atmospheres of up to 25 kPa for

J.K. Brecht and E.M. Yahia510
2 weeks at 12°C (Bender et al., 2000b). High (25 kPa) CO2 inhibits ethylene production, but increases ethanol production. Aroma volatiles are reduced following 25 kPa CO2 treatment, while 10 kPa CO2, low O2 atmospheres and storage temperature did not signifi cantly infl uence production of terpene hydrocarbons, which are characteristic of Florida-type mangoes. Mature-green ‘Tommy Atkins’ mangoes can be stored for 21 days in CA (5 kPa O2 + 10 kPa or 25 kPa CO2) at 12°C, while tree-ripe fruit can be stored for 21 days in the same atmospheres at either 8 or 12°C (Bender et al., 2000a).
The quality of ‘Keitt’ mangoes was evaluated during storage for 6 days at 20°C in an extremely low O2 (LO) CA (approximately 0.3 kPa) before stor-age in modifi ed atmosphere packaging (MAP) made from three, low-density polyethylene (LDPE) fi lms with different gas permeability characteristics (González-Aguilar et al., 1997). Both LO and MA treatments delayed the losses of colour, weight and fi rmness. Fruit maintained good appearance with a signifi cant delay of ripening. Mangoes are very tolerant of LO treat-ment; however, some MAP fruit developed a fermented taste after 10 and 20 days at 20°C. Short duration (6-day) storage of mangoes in LO did not other-wise have any deleterious effect on fruit quality during subsequent storage under MA or normal atmosphere. Properly selected atmospheres, which pro-long mango shelf life by slowing ripening, permit fruit to be shipped without sacrifi cing superior aroma.
Beaulieu and Lea (2003) studied ‘Keitt’ and ‘Palmer’ mangoes without heat treatment to assess volatile and quality changes in stored fresh-cut man-goes prepared from fi rm-ripe (FR) and soft-ripe (SR) fruit, and to assess what effect MAP may have on cut fruit physiology, overall quality and volatile retention or loss. Subjective appraisals of fresh-cut mangoes based on aroma and cut edge or tissue damage indicated that most SR cubes are unmarket-able by day 7 at 4°C. Both cultivars stored in MAP at 4°C had almost identical O2 consumption, which is independent of ripeness. The CO2 and O2 concen-trations measured for cubes stored in passive MAP indicated that the system is inadequate to prevent potential anaerobic respiration after 7 days storage.
Injuries associated with MA and CA
A 10 kPa CO2 atmosphere alleviates chilling symptoms in ‘Kensington Pride’ fruit, but higher concentrations are injurious; low O2 (5 kPa) has no signifi -cant effect (O’Hare and Prasad, 1993). Higher concentrations of CO2 (>10 kPa) are ineffective for alleviating CI at 7°C, and tend to cause tissue injury and high levels of ethanol in the pulp. Injury in ‘Kensington Pride’ caused by higher levels of CO2 appears to be more severe at lower temperatures (O’Hare and Prasad 1993; Bender et al., 1994, 1995), which could be a result of either compounding injury (chilling + CO2) or reduced sensitivity of ripe mango to CO2.
‘Rad’ mangoes develop internal browning and off-fl avour in atmo-spheres containing 6 and 8 kPa CO2 (Noomhorm and Tiasuwan, 1995). The presence of starchy mesocarp in ‘Carabao’ mangoes, which is characteristic

Postharvest Physiology 511
of internal breakdown, increases during storage in MA (Gautam and Lizada, 1984). Fruit stored for 4–5 days have severe symptoms, including air pockets in the mesocarp resulting in spongy tissue (Nuevo et al., 1984a, b). Paren-chyma cells of affected tissues have c.18 starch granules per cell, compared to c.2 starch granules in healthy adjacent cells. However, no difference in starch granule shape was detected between the spongy and healthy tissues. The spongy tissue, which usually occurs in the inner mesocarp near the seed and becomes evident during ripening, has almost ten times the starch content of healthy tissue in the same fruit. External symptoms of internal browning due to MA include failure of the peel to develop colour beyond the half-yellow stage.
‘Carabao’ mangoes stored in polyethylene bags (0.04 mm thickness) for 1 day at 25–31°C had a faint fermented odour that disappeared during sub-sequent ripening outside the bags (Gautam and Lizada, 1984). The fermented odour increases with time, and persists throughout ripening when the fruit are stored for 2–5 days. The respiratory quotient of this cultivar ranged from 0.59 at 21 kPa O2 to 6.03 at 2.4 kPa O2, which indicates a progressively anaer-obic metabolism (Sy and Mendoza, 1984). CO2 production decreases as O2 decreases from 21 to 3 kPa, but increases at <3 kPa O2. Fermentative decar-boxylation could explain the odour (Lakshminarayana and Subramanyam, 1970).
Pronounced decay appears after storage of ‘Rad’ mangoes for 20 days in atmospheres containing 4–6 kPa O2 with 4–8 kPa CO2 at 13°C and 94% RH, and severe incidence of decay appears after 25 days (Noomhorm and Tiasu-wan, 1995). Greater incidence of decay (stem-end rot and anthracnose) occurs in ‘Carabao’ mango stored in MA for 2–5 days at 25–31°C (Gautam and Lizada, 1984).
Modifi ed atmosphere packaging (MAP)
Modifi ed atmosphere packaging is used to create a benefi cial MA around a packaged product using semipermeable fi lm to restrict the movement of respiratory gases into and out of the package; at equilibrium, the respiration rate of fruit in MAP is equal to the diffusion of the respiratory gases through the fi lm. Fruit wrapped in 0.08 mm thick polyethylene bags, with and with-out perlite-potassium permanganate (KMnO4) and stored for 3 weeks at 10°C before treatment with ethylene developed normal colour, texture and fl avour (Esguerra et al., 1978). Individually sealed ‘Keitt’ in low-density (LDPE) and high-density (HDPE) polyethylene fi lms for 4 weeks at 20°C exhibited delayed ripening, reduced weight loss and did not develop any off-fl avours (Gonzalez et al., 1990). The LDPE had a thickness of 0.010 mm and permea-bilities of 700 cm3 O2/m2/h/atm and 0.257 g H2O/m2/h/atm. The HDPE fi lm had a thickness of 0.020 mm and permeabilities of 800 cm3 O2/m2/h/atm and 0.166 g H2O/m2/h/atm.
In a study to model MAP for mango, ‘Keitt’ fruit were individually vacuum packaged in LDPE fi lm (0.0245 mm thick, 25.0 g/m2) and stored at 7°C/80–90%

J.K. Brecht and E.M. Yahia512
RH, 12°C/75–85% RH, 17°C/70–80% RH, 22°C/65–75% RH or 25°C/65–75% RH (Yamashita et al., 1997). After mass transfer had reached steady state, respiration rates, moisture loss, permeability of peel and fi lm to water vapour, and composition of atmosphere around the fruit were determined for 33 days. Daily rates of weight loss increased from 4.1 g/kg of fruit at 7°C to 10.9 g/kg at 25°C. Respiration rates also increased with storage temperature for both packaged and unpackaged mangoes, and were 21, 38 and 43% less in packaged fruit at 12, 17 and 22°C, respectively. Permeability of peel was 600-fold greater than that of the plastic fi lm. The in-package CO2 levels increased and O2 decreased with time; concentration changes were greatest during the fi rst 10–15 days of storage and were more marked at the higher tempera-tures. Experimental and calculated values for CO2 levels differed by 29%, depending on temperature.
‘Tommy Atkins’ mangoes individually sealed in heat-shrinkable fi lms and stored for 2 weeks at 12.8°C and then ripened at 21°C had less weight loss, but did not show differences in fi rmness, skin colour development, decay development or time to fruit ripening, and had more off-fl avours than unwrapped fruit (Miller et al., 1983). Polyethylene fi lms used were: Clysar EH-60 fi lm of 0.01 mm nominal thickness, Clysar EHC-50 copolymer fi lm of 0.013 mm nominal thickness, and Clysar EHC-100 copolymer fi lm of 0.025 mm nominal thickness. Individual mature fruit of the same cultivar were later sealed in Clysar EHC-50 copolymer fi lm with 0.013 mm thickness, and Cryovac D955 with 0.015 mm thickness, and stored at 21°C and 85–90% RH (Miller et al., 1986). The O2 permeabilities of the fi lms were 620 cm3/24 h/m2/atm and 9833 cm3/24 h/m2/atm, respectively. Water permeability was 1.5 g/24 h/m2 and 2.0 g/24 h/m2 at 23°C, respectively. Fruit in MAP had less weight loss, but higher incidence of decay and off-fl avour at soft-ripeness than unsealed fruit. The authors concluded that there were no practical ben-efi ts from wrapping this fruit in these fi lms and storage at 21°C or even at lower temperatures. ‘Film wrapping mangoes at various stages of ripeness after harvest…will not improve the maintenance of mango quality during storage or ripening.’
‘Keitt’ mangoes were individually sealed in LDPE fi lms and in a heat-shrinkable copolymer (Cryovac D-955) fi lm with non-sealed mangoes as the control and stored for up to 5 weeks at 12°C, 17°C or 22°C (Yamashita et al., 1999). MAP reduced the rate constant of vitamin C degradation at all tem-peratures and vitamin C content of individually packaged mangoes was less affected by storage temperature than the control. Values for Q10 (the ratio of CO2 production to O2 consumption in respiration) were 1.3 and 1.0 for man-goes wrapped with the heat-shrinkable copolymer and the LDPE fi lms, respectively, and 2.8 for the non-sealed control.
The combined effect of hot benomyl (1000 ppm) at 55°C for 5 min and fi lm packaging in 0.01 mm PVC extended the storage life of mature-green ‘Nam Dok Mai’ mangoes stored at 13°C (Sornsrivichai et al., 1992). Fruit qual-ity was not affected by fi lm packaging after 4 weeks, but fruit showed infe-rior quality after 6 weeks. The inhibition of carotene pigmentation in the peel of this cultivar may be related to O2 concentration inside the package and not

Postharvest Physiology 513
to CO2 concentration (Yantarasri et al., 1994). At least 16 kPa O2 is essential for development of peel colour to the marketable stage (greenish).
‘Kensington Pride’ mangoes treated with heated benomyl (0.5 g/l at 51.5°C for 5 min) and sealed in polyethylene bags (0.04 mm thickness) for various durations at 20°C, had off-fl avour and lacked normal skin colour when ripened, but ripened satisfactorily in perforated bags (Chaplin et al., 1982). The postharvest life of these fruit was not consistently longer than the control. The CO2 concentration in the bags was >20 kPa while the O2 concen-tration was <5 kPa. The incidence of off-fl avours was reduced by including ethylene-absorbent blocks in the bags. The authors concluded that ‘mangoes cannot be stored satisfactorily at ambient temperature by such technique’; however, Stead and Chithambo (1980) reported that fruit ripening at 20–30°C was delayed 5 days by sealing in polyethylene bags (0.02 mm thickness) with ethylene-absorbent blocks without any abnormal fl avour.
‘Tommy Atkins’ and ‘Keitt’ mangoes were individually sealed in shrink-able Cryovac polyolefi n fi lms (0.015 or 0.019 mm thickness), either non- perforated (MD fi lm) or perforated with eight holes of 1.7 mm diameter/inch2 (MPY) or eight holes of 0.4 mm diameter/inch2 (SM60M) (Rodov et al., 1994). After 2–3 weeks at 14°C and an additional week at 17°C, mangoes packaged in perforated polyolefi n fi lms ripened normally. Optimum results were achieved when the fi lm with 0.4 mm perforations was combined with increased free volume inside the package by sealing the fruit within polysty-rene trays. After 3 weeks of storage and 1 week of shelf life, sealed ‘Keitt’ mangoes were inferior to the control; they were less ripe, but beyond 4 weeks (up to 6 weeks) sealed fruit had better quality scores because they were less overripe. Sealing did not reduce decay of fruit stored for long periods.
Non-perforated PVC fi lm packaging of ‘Nam Doc Mai’ mangoes was not suffi ciently permeable for O2 exchange to allow proper ripening (Yantarasri et al., 1995). Therefore, a ‘perforated MA’ was used in which fruit were wrapped in polystyrene trays (three fruit/pack) at 20°C with perforation area of ≥0.004 cm2. Fruit ripened normally with no off-fl avours. Colour devel-opment in the peel required a higher concentration of O2 than the fl esh. A fi lm of pore area ≥0.008 cm2 allowed fruit colour to develop after 3 weeks while a pore area of ≥0.39 cm2 allowed the fruit to colour within 2 weeks.
Semipermeable coatings
Some fruit coatings can create an internal MA within the fruit due to semi-permeable restriction of O2 and CO2 movement in and out of the fruit. Bald-win et al. (1999) tested two types of fruit coatings – polysaccharide-based and carnauba wax-based – for their effect on external and internal mango fruit atmospheres and quality factors during simulated commercial storage at 10 or 15°C with 90–99% RH, followed by simulated marketing conditions at 20°C and 56% RH. The coatings exhibited markedly different O2 permeabil-ity characteristics under laboratory conditions. Polysaccharide coatings were less permeable to respiratory gases (i.e. O2 and CO2) and more permeable to

J.K. Brecht and E.M. Yahia514
water vapour compared to carnauba wax. When applied to fruit under simu-lated commercial conditions, however, the differences between the coatings with regards to their permeability to respiratory gases were much reduced, most likely due to high RH during cold storage. Both coatings created a MA within the fruit, reduced decay and improved appearance by imparting a sub-tle shine; but only the polysaccharide coating delayed ripening and increased concentrations of fl avour volatiles. The carnauba wax coating signifi cantly reduced water loss compared to uncoated and polysaccharide-coating treat-ments.
‘Julie’ mangoes treated with 0.75% w/v aqueous solution of Pro-long semipermeable fruit coating (a mixture of sucrose esters of fatty acids and sodium salt of carboxy methyl cellulose) and stored at 25°C and 85–95% RH exhibited reduced weight loss, retarded ripening and increased storage life (6 days longer) without evidence of adverse effects on quality (Dhalla and Hanson, 1988). Treatment with 1.0% Pro-long could increase ethanol concen-tration in the pulp. Treatment with Pro-long (0.8–2.4%) also delayed ripening of ‘Haden’ (Carrillo-López et al., 1996).
Insecticidal CA
Mangoes are very tolerant of insecticidal atmospheres, and thus a potential commercial application is feasible, especially in combination with other treat-ments (i.e. heat). ‘Keitt’ mango tolerated as low as 0.2 kPa O2 and as high as 80 kPa CO2 for up to 5 days without any injury upon ripening, although fer-mentative odours could be noted while the fruit were under atmosphere stress (Yahia, 1993, 1994, 1995, 1997; Ortega and Yahia, 2000). Other mango cultivars were also evaluated and were very tolerant of extreme atmospheres (Yahia, 1998).
Storage of ‘Keitt’ mangoes in an insecticidal MA (0.03–0.26 kPa O2, 72–79 kPa CO2, balance nitrogen (N2)) and CA (0.2 kPa O2, balance N2; or 2 kPa O2 + 50 kPa CO2, balance N2) for up to 5 days at 20°C delayed fruit ripening as indicated by respiration, fl esh fi rmness and colour development (Yahia et al., 1989; Yahia, 1993; Yahia and Tiznado, 1993; Yahia and Vazquez, 1993). The activities of phosphofructokinase, alcohol dehydrogenase (EC 1.1.1.1) and pyruvate decarboxylase were enhanced but activity of pyruvate kinase, suc-cinate dehydrogenase and -ketoglutarate dehydrogenase (EC 1.2.4.2) was unaffected. Although these atmospheres caused changes in glycolysis and tricarboxylic acid cycle, there was no indication of injury and the fruit rip-ened normally in air. Sensory evaluation conducted after fruit ripening showed no off-fl avours, and there were no differences between fruit main-tained in the MA or CA and those maintained continuously in air. ‘Keitt’ mango is therefore very tolerant of insecticidal atmospheres, and 5 days exposure is suffi cient to control many insects (Rojas-Villegas et al., 1996).
Storage of ‘Keitt’ and ‘Tommy Atkins’ mangoes for 21 days at 12°C in atmospheres containing 25, 45, 50 or 70 kPa CO2 plus either 3 kPa O2 or air, induced ethanol production of 0.18–3.84 ml/kg/h after transfer to air at 20°C

Postharvest Physiology 515
for 5 days (Bender et al., 1994). Atmospheres containing 50 or 70 kPa CO2 caused fruit injury, and resulted in the highest ethanol production rates. Enclosure of ‘Haden’ and ‘Tommy Atkins’ mangoes in sealed 20 l jars with an initial atmosphere of 90 kPa CO2 in air or 97 kPa N2 + 3 kPa O2 for 24 h prior to storage delayed their ripening, and no injury was reported (Pesis et al., 1994).
14.8 Manipulation of Mango Postharvest Physiology by Molecular Biology
Mango fruit quality is compromised when harvest occurs before the fruit are fully mature since they are unable to achieve the full complement of fl avour and aroma during the postharvest handling period compared with fruit harvested at a fully mature or ripening-initiated stage of development. As a climacteric fruit, maturity in mango corresponds to attainment of ripening competence. The presence of ethylene is required for the progression and com-pletion of mango ripening. Thus, strategies for prolonging the postharvest life and maintaining postharvest quality of mango other than disease control are focused on reducing the effects of ethylene. This situation provides an excellent opportunity to utilize genetic transformation to improve mango postharvest quality by manipulating the role of ethylene (see Litz et al., Chap-ter 18, this volume). Cruz-Hernández et al. (1997) transformed ‘Hindi’ mango with mango ACO and ACC synthase (EC 4.4.1.14) in the antisense orienta-tion. A cDNA that codes for mango ACO was also isolated and characterized by Zainal et al. (1999). Suppression of mango ethylene biosynthesis should allow harvesting of advanced maturity fruit that contain high levels of sug-ars and possess enhanced capacity to produce ripe aroma volatiles after exposure to exogenous ethylene. Progression of ripening in such fruit can be easily halted at the most desirable and convenient time by simply removing exogenous ethylene.
A cDNA homologue of the ethylene receptor gene ETR-1, referred to as METR1, which codes for a polypeptide of 802 amino acids with a predicted 89 kDa MW has been isolated (Gutierrez-Martínez et al., 2001). Two or more ETR homologues exist in mango. The level of METR1 mRNA in the mesocarp increases transiently during wounding. Repression of genes involved in eth-ylene action in mango fruit should result in ethylene-insensitive fruit that are minimally affected by exposure to ethylene in the postharvest environment, resulting in better control of ripening and senescence to maintain mango postharvest quality.
Internal breakdown in mango fruit is essentially a problem of premature ripening; the longer harvest of susceptible varieties is delayed, the greater the incidence of internal breakdown. Using molecular approaches to reduce ethylene production and action in mature fruit could reduce internal break-down or premature ripening and lead to greatly improved quality. Another approach to mitigating internal breakdown would be to target genes involved in the maintenance of cell wall integrity. Vasanthaiah et al. (2006) demonstrated

J.K. Brecht and E.M. Yahia516
differential expression of several genes in tissue showing internal breakdown symptoms compared with healthy tissue. They suggested that oxidative stress may be one of the causes of the disorder.
Sane et al. (2005) have isolated and characterized an ethylene-dependent a-expansin gene, MiExpA1, which is correlated with softening in mango. Expression of MiExpA1 increases with the progression of ripening and treat-ment with 1-methyl cyclopropene inhibits both ripening/softening as well as MiExpA1 transcript and protein accumulation. Recently, a pectate lyase (EC 4.2.2.2) homologue from ripening mango (MiPel1) has been cloned (Chour-asia et al., 2006). A progressive increase in transcript accumulation was observed during ripening but expression was delayed signifi cantly in fruit in air without exogenous ethylene. The expression was specifi c to fruit and was triggered only during ripening. Increased transcript accumulation during ripening was associated with pectin solubilization. Pectate lyase may be closely associated with pectin degradation and have an important role in mango softening.
14.9 Conclusions
Mango fruit have the potential to develop extremely desirable texture, taste and aroma that make this fruit highly appreciated and desirable. Strategies used to extend mango shelf life are based on control of ripening, ethylene action and ethylene production. Therefore, fruit are usually harvested at the mature-green stage, prior to ripening initiation, and stored and transported at low temperatures at or near the threshold for induction of chilling injury. These practices result in poor quality, immature and chill-injured mangoes on the market. Successful handling of ripening-initiated mangoes is prob-lematic due to the fruit’s short shelf life and the increased incidence of inter-nal breakdown that accompanies delayed harvests makes international transport of ripening mangoes almost impossible. Consequently, the export market for fresh mangoes, which expanded rapidly in the 1990s, has not con-tinued its rapid expansion in recent years.
Future expansion of mango consumption will require an understanding of mango postharvest physiology in order to overcome the problems of CI, internal breakdown, and premature and uneven ripening. This may involve increased transport of tree-ripe mangoes in CA-equipped marine containers or in MAP. It may involve development of improved procedures for storage and ripening to offer preconditioned, ripening-initiated, ready-to-eat man-goes to consumers. It may also involve genetic transformation of mango to manipulate the progression and uniformity of ripening and softening.
References
Abu-Sarra, A.F. and Abu-Goukh, A.A. (1992) Changes in pectinesterase, polygalactu-ronase and cellulase activity during mango fruit ripening. Journal of Horticultural Science 67, 561–568.

Postharvest Physiology 517
Akamine, E.K. (1963) Haden mango storage. Hawaii Farm Science 12, 6. Akamine, E.K. and Goo, T. (1973) Respiration and ethylene production during ontogeny
of fruit. Journal of the American Society for Horticultural Science 98, 381–383.Ali, Z.M., Abu, S. and Lazan, H. (1990) Glycosidases in ripening mango. Transactions
of the Malaysian Society of Plant Physiology 1, 11–14.Ali, Z.M., Armugam, S. and Lazan, H. (1995) -Galactosidase and its signifi cance in
ripening mango fruit. Phytochemistry 38, 1109–1114.Allong, R., Wickham, L.D. and Mohammed, M. (2001) Effect of slicing on the rate of
respiration, ethylene production and ripening of mango fruit. Journal of Food Qual-ity 24, 405–419.
Ashraf, M., Khan, N., Ahmed, M. and Elahi, M. (1981) Studies on the pectinesterase activity and some chemical constituents of some Pakistani mango varieties during storage and ripening. Journal of Agricultural and Food Chemistry 29, 526–528.
Baldwin, E.A., Burns, J.K., Kazokas, W., Brecht, J.K., Hagenmaier, R.D., Bender, R.J. and Pesis, E. (1999) Effect of two edible coatings with different permeability character-istics on mango (Mangifera indica L.) ripening during storage. Postharvest Biology and Technology 17, 215–226.
Bandyopadhyay, C. (1983) Contribution of gas chromatography to food fl avour research. Pafai Journal 5, 26–30.
Bandyopadhyay, C. and Gholap, A.S. (1973a) Changes in fatty acids in ripening mango pulp (variety Alphonso). Journal of Agricultural and Food Chemistry 21, 496–497.
Bandyopadhyay, C. and Gholap, A.S. (1973b) Relationship of aroma and fl avour charac-ters of mango (Mangifera indica L.) to fatty acid composition. Journal of the Science of Food and Agriculture 24, 1497–1503.
Baqui, S.M., Mattoo, A.K. and Modi, V.V. (1974) Mitochondrial enzymes in mango fruit during ripening. Phytochemistry 13, 2049–2055.
Barmore, C.R. (1974) Ripening of mangos with ethylene and ethephon. Proceedings of the Florida State Horticultural Society 87, 331–334.
Barmore, C.R. and Mitchell, E.F. (1975) Ethylene pre-ripening of mangoes prior to ship-ment. Proceedings of the Florida State Horticultural Society 88, 469–471.
Bartley, J.P. and Schwede, A. (1987) Volatile fl avour components in the headspace of the Australian or ‘Bowen’ mango. Journal of Food Science 52, 353–360.
Beaulieu, J.C. and Lea, J.M. (2003) Volatile and quality changes in fresh-cut mangos prepared from fi rm-ripe and soft-ripe fruit, stored in clamshell containers and pas-sive MAP. Postharvest Biology and Technology 30, 15–28.
Ben-Amotz, A. and Fishler, R. (1998) Analysis of carotenoids with emphasis on 9-cis- -carotene in vegetables and fruits commonly consumed in Israel. Food Chemistry 62, 515–520.
Bender, R.J., Brecht, J.K. and Campbell, C.A. (1994) Response of ‘Kent’ and ‘Tommy Atkins’ mangoes to reduced O2 and elevated CO2. Proceedings of the Florida State Horticultural Society 107, 274–277.
Bender, R.J., Brecht, J.K. and Sargent, S.A. (1995) Inhibition of ethylene production in mango fruit by elevated CO2 and recovery during subsequent air storage. Proceed-ings of the Florida State Horticultural Society 108, 279–285.
Bender, R.J., Brecht, J.K., Baldwin, E.A. and Malundo, T.M.M. (2000a) Aroma volatiles of mature-green and tree-ripe ‘Tommy Atkins’ mangoes after controlled atmosphere vs. air storage. HortScience 35, 684–686.
Bender, R.J., Brecht, J.K., Sargent, S.A. and Huber, D.J. (2000b) Mango tolerance to re-duced oxygen levels in controlled atmosphere storage. Journal of the American Society for Horticultural Science 125, 707–713.

J.K. Brecht and E.M. Yahia518
Biale, J.B. and Young, R.E. (1981) The avocado pear. In: Hulme, A.C. (ed.) The Biochem-istry of Fruits and their Products. Vol. 2. Academic Press, New York, pp. 1–63.
Botting, K.J., Yong, M.M., Pearson, A.E., Harris, P.J. and Ferguson, L.R. (1999) Antimuta-gens in food plants eaten by Polynesians: micronutrients, phytochemicals and pro-tection against bacterial mutagenicity of heterocyclic amine 2-amino-3-methylimidazo[4,5-f]quinoline. Food and Chemical Toxicology 37, 95–103.
Brinson, K., Dey, P.M., John, M.A. and Pridham, J.B. (1988) Post-harvest changes in Mangifera indica mesocarp cell walls and cytoplasmic polysaccharides. Phy-tochemistry 27, 719–723.
Brown, B.I., Scott, K.J. and Chaplin, G.R. (1981) Spongy stem end in mango. Queen-sland Fruit and Vegetable News 52, 497.
Burdon, J., Dori, S., Marinansky, R. and Pesis, E. (1996) Acetaldehyde inhibition of ethyl-ene biosynthesis in mango fruit. Postharvest Biology and Technology 8, 153–161.
Burg, S.P. and Burg, E.A. (1962) Role of ethylene in fruit ripening. Plant Physiology 37, 179–189.
Burns, J., Fraser, P.D. and Bramley, P.M. (2003) Identifi cation and quantifi cation of caro-tenoids, tocopherols and chlorophylls in commonly consumed fruits and vegeta-bles. Phytochemistry 62, 939–947.
Cano, M.P. and de Ancos, B. (1994) Carotenoid and carotenoid ester composition in mango fruit as infl uenced by processing method. Journal of Agricultural and Food Chemistry 42, 2737–2742.
Carlier, C., Etchepare, M., Ceccon, J.F., Mourey, M.S. and Amedee-Manesme, O. (1992) Effi cacy of massive oral doses of retinyl palmitate and mango (Mangifera indica L.) consumption to correct an existing vitamin A defi ciency in Senegalese children. British Journal of Nutrition 68, 529–540.
Carrillo-López, A., Rojas-Villagas, R. and Yahia, E.M. (1996) Ripening and quality of mango fruit as affected by coating with ‘Semperfresh’. Acta Horticulturae 370, 206–216.
Castrillo, M., Kruger, N.J. and Whatley, F.R. (1992) Sucrose metabolism in mango fruit during ripening. Plant Science 84, 45–51.
Chaplin, G.R. (1988) Advances in postharvest physiology of mango. Acta Horticulturae231, 639–648.
Chaplin, G.R., Scott, K.L. and Brown, B.I. (1982) Effect of storing mangoes in polyethylene bags at ambient temperature. Singapore Journal of Primary Industries 10, 84–88.
Chen, J.P., Tai, C.Y. and Chen, B.H. (2004) Improved liquid chromatographic method for determination of carotenoids in Taiwanese mango (Mangifera indica L.). Journal of Chromatography 1054, 261–268.
Chourasia, A., Sane, V.A. and Nath, P. (2006) Differential expression of pectate lyase during ethylene-induced postharvest softening of mango (Mangifera indica var. Dashehari). Physiologia Plantarum 128, 546–555.
Coates, L.M. and Johnson, G.I. (1993) Effective disease control in heat-disinfected fruit. Postharvest News and Information 4, 35N–40N.
Cocozza, F.M., Jorge, J.T., Alves, R.E., Filgueiras, H.A.C., Garruti, D.S. and Pereira, M.E.C. (2004) Sensory and physical evaluations of cold stored ‘Tommy Atkins’ mangoes infl uenced by 1-MCP and modifi ed atmosphere packaging. Acta Horticul-turae 645, 655–661.
Considine, M.J., Daley, D.O. and Whelan, J. (2001) The expression of alternative oxi-dase and uncoupling protein during fruit ripening in mango. Plant Physiology 126,1619–1629.
Couey, M.H. (1986) Chilling injury in fruits of tropical and subtropical origin. Hort-Science 17, 162–165.

Postharvest Physiology 519
Cruz-Hernández, A., Gómez-Lim, M.A. and Litz, R.E. (1997) Transformation of mango somatic embryos. Acta Horticulturae 455, 292–298.
Cua, A.U. and Lizada, M.C.C. (1990) Ethylene production in the ‘Carabao’ mango (Mangifera indica L.) fruit during maturation and ripening. Acta Horticulturae 269, 169–179.
Dhalla, R. and Hanson, S.W. (1988) Effect of permeable coatings on the storage life of fruits. II. Pro-long treatment of mangoes (Mangifera indica L., cv Julia). InternationalJournal of Food Science and Technology 23, 107–112.
Dubery, I.A., Van Rensburg, L.J. and Schabort, J.C. (1984) Malic enzyme activity and related biochemical aspects during ripening of -irradiated mango fruit. Phytochem-istry 23, 1383–1386.
Engel, K.H. and Tressl, R. (1983) Studies on the volatile components of two mango vari-eties. Journal of Agricultural and Food Chemistry 31, 796–801.
Esguerra, E.B. and Lizada, M.C.C. (1990) The postharvest behavior and quality of ‘Cara-bao’ mangoes subjected to vapor heat treatment. ASEAN Food Journal 5, 6–12.
Esguerra, E.B., Mendoza, D.B., Jr and Pantastico, E.R.B. (1978) Regulation of fruit ripen-ing. II. Use of perlite-KMnO4 insert as an ethylene absorbent. Philippine Journal of Science 107, 23–31.
Farooqui, Q.A., Sattar, A., Daud, K. and Hussain, M. (1985) Studies on the postharvest chilling sensitivity of mango fruit (Mangifera indica L.). Proceedings of the Florida State Horticultural Society 98, 220–221.
Fuchs, Y., Pesis, E. and Zauberman, C. (1980) Changes in amylase activity, starch and sugar contents in mango fruit pulp. Scientia Horticulturae 13, 155–160.
García-Solís, P., Yahia, E.M. and Aceves, C. (2008) Study of the effect of ‘Ataulfo’ mango (Mangifera indica L.) intake on mammary carcinogenesis and antioxidant capacity in plasma of N-methyl-N-nitrosourea (MNU)-treated rats. Food Chemistry 111, 309–315.
Gautam, D.M. and Lizada, M.C.C. (1984) Internal breakdown in ‘Carabao’ mango sub-jected to modifi ed atmospheres. Storage duration and severity symptoms. Posthar-vest Research Notes 1, 28–30.
Gholap, A.S. and Bandyopadhyay, C. (1975a) Comparative assessment of aromatic prin-ciples in ripe ‘Alphonso’ and ‘Langra’ mangoes. Journal of Agricultural and Food Science 12, 262–263.
Gholap, A.S. and Bandyopadhyay, C. (1975b) Contribution of lipid to aroma of ripening mango. Journal of the American Oil Chemistry Society 52, 514–516.
Gholap, A.S. and Bandyopadhyay, C. (1976) Fatty acid composition as a quality index of ripe mango (Mangifera indica L.) pulp. Indian Food Packer 30, 63–64.
Gholap, A.S. and Bandyopadhyay, C. (1980) Fatty acid biogenesis in ripening mango (Mangifera indica cv. Alphonso). Journal of Agricultural and Food Chemistry 28, 839–841.
Godoy, H.T. and Rodriguez-Amaya, D.B. (1989) Carotenoid composition of commercial mangoes from Brazil. Lebensmittel-Wissenschaft und Technologie 22, 100–103.
Godoy, H.T. and Rodriguez-Amaya, D.B. (1994) Occurrence of cis-isomers of provitamin A in Brazilian fruits. Journal of Agricultural and Food Chemistry 42, 1306–1313.
Gofur, M.A., Shafi que, M. and Helali, O.H. (1994) Effect of various factors on the vita-min C (ascorbic acid) content of some mango varieties grown in Rajshahi region. Bangladesh Journal of Scientifi c and Industrial Research 29, 163–171.
Gonzalez, G., Yahia, E.M. and Higuera, I. (1990) Modifi ed atmosphere packaging (MAP) of mango and avocado fruit. Acta Horticulturae 269, 335–344.
González-Aguilar, G., Gardea, A., Martinez-Tellez, M.A., Baez, R. and Felix, L. (1997) Low oxygen treatment before storage in normal or modifi ed atmosphere packaging

J.K. Brecht and E.M. Yahia520
of mangoes to extend shelf life. Journal of Food Science and Technology 34, 399–404.
González-Aguilar, G.A., Buta, J.G. and Wang, C.Y. (2001) Methyl jasmonate reduces chilling injury symptoms and enhances colour development of ‘Kent’ mangoes. Journal of the Science of Food and Agriculture 81, 1244–1249.
Gosh, S. (1960) The content of folic acid and its conjugates in some common Indian fruits. Science and Culture 26, 287–292.
Gowda, I.N.D. and Huddar, A.G. (2000) Evaluation of mango hybrids for storage behav-iour and sensory qualities. Journal of Food Science and Technology (Mysore) 37, 620–623.
Gowda, I.N.D., Ramanjaneya, K.H., Iyer, C.P.A., Subramanyam, M.D. and Dinesh, M.R. (1994) Physico-chemical and processing quality of four new mango hybrids in comparison to two commercial cultivars. Journal of Food Science and Technology(Mysore) 31, 385–388.
Gutierrez-Martínez, P., López Gómez, R. and Gómez-Lim, M.A. (2001) Identifi cation of an ETR1-homologue from mango fruit expressing during fruit ripening and wound-ing. Journal of Plant Physiology 158, 101–108.
Hatton, T.T., Jr, Reeder, W.F. and Campbell, C.W. (1965) Ripening and Storage of Florida Mangoes. Marketing Research Report 725. Agricultural Research Service, United States Department of Agriculture (USDA), Washington, DC.
Heather, N.W. (1994) Quarantine disinfestation of tropical fruits: non-chemical options. In: Champ, B.C., Highley, E. and Johnson, G.I. (eds) Postharvest Handling of Tropi-cal Fruits. Australian Centre for International Agricultural Research (ACIAR) Pro-ceedings 50. ACIAR, Canberra, pp. 272–279.
Iguina de George, L.M., Collazo de Rivera, A.L., Benero, J.R. and Pennock, W. (1969) Provitamin A and vitamin C contents of several varieties of mango (Mangifera in-dica L.) grown in Puerto Rico. Journal of Agriculture of the University of Puerto Rico53, 100–105.
Ito, T., Sasaki, K. and Yoshida, Y. (1997) Changes in respiration rate, saccharide and or-ganic acid content during the development and ripening of mango fruit (Mangiferaindica L. ‘Irwin’) cultured in a plastic house. Journal of the Japanese Society for Horticultural Science 66, 629–635.
Jacobi, K.K. and Giles, J.E. (1997) Quality of ‘Kensington’ mango (Mangifera indicaLinn.) fruit following combined vapour heat disinfestation and hot water disease control treatments. Postharvest Biology and Technology 12, 285–292.
Jacobi, K.K. and Wong, L.S. (1992) Quality of ‘Kensigton’ mango (Mangifera indica L.) following hot water and vapour-heat treatments. Postharvest Biology and Technol-ogy 1, 349–359.
Jacobi, K.K., Coates, L.M. and Wong, L.S. (1994) Heat disinfestation of mangoes: effect on fruit quality and disease control. In: Champ, B.R., Haghley, E. and Johnson, G.I. (eds) Postharvest Handling of Tropical Fruits. Australian Centre for International Agricultural Research (ACIAR) Proceedings 50. ACIAR, Canberra, pp. 280–287.
Jacobi, K.K., MacRae, E.A. and Hetherington, S.E. (2001a) Effect of fruit maturity on the response of ‘Kensington’ mango fruit to heat treatment. Australian Journal of Ex-perimental Agriculture 41, 793–803.
Jacobi, K.K., MacRae, E.A. and Hetherington, S.E. (2001b) Postharvest heat disinfesta-tion treatments of mango fruit. Scientia Horticulturae 89, 171–193.
Jain, N.L. (1961) Chemistry and technology of mango. Review of Food Technology 3, 131–162.
Jha, S.N., Chopra, S. and Kingsly, A.R.P. (2007) Modeling of color values for non-destructive evaluation of maturity of mango. Journal of Food Engineering 78, 22–26.

Postharvest Physiology 521
John, J., Subbarayan, C. and Cama, H.R. (1970) Carotenoids in three stages of ripening of mango. Journal of Food Science 35, 262–265.
Johnson, G.I. and Coates, L.M. (1993) Postharvest diseases of mango. Postharvest News and Information 4, 27N–34N.
Johnson, G.I., Sharp, J.L., Milne, D.L. and Oosthuyse, S.A. (1997) Postharvest technol-ogy and quarantine treatments. In: Litz, R. (ed.) The Mango: Botany, Production and Uses. CAB International, Wallingford, UK, pp. 447–507.
Jungalwala, F.B. and Cama, H.R. (1963) Carotenoids in mango (Mangifera indica) fruit. Indian Journal of Chemistry 1, 36–40.
Kalra, S.K. and Tandon, D.K. (1983) Ripening behavior of ‘Dashehari’ mango in relation to harvest period. Scientia Horticulturae 19, 263–269.
Kane, O. (1977) Recherches sur l’évolution normale ou pathologique de mangues (Mangifera indica L.) conservées au froid dans l’air ou en atmosphère contrôlée. Thèse doct 3ème cycle. Université Pierre et Marie Curie, Paris, 124 pp.
Kapse, B.M., Rane, D.A. and Khedkar, D.M. (1989) Correlation between bio-chemical parameters and organoleptic evaluation in mango varieties. Acta Horticulturae231, 756–762.
Karmarkar, D.V. and Joshi, B.M. (1941) Respiration studies of the ‘Alphonso’ mango. Indian Journal of Agricultural Science 11, 993–1005.
Ketsa, S., Rattanamalee, S. and Babprasert, C. (1991) Growth, development, biochemi-cal changes and harvesting index of mango (Mangifera indica L.) cv. Tongdum. Kasetsart Journal (Natural Science) 25, 391–399.
Ketsa, S., Phakawatmongkol, W. and Subhadrabhandhu, S. (1999) Peel enzymatic activ-ity and colour changes in ripening mango fruit. Journal of Plant Physiology 154, 363–366.
Kostermans, A.J.G.H. and Bompard, J.M. (1993) The Mangoes: Botany, Nomenclature, Horticulture and Utilization. Academic Press, London, 249 pp.
Krishnamurthy, S. and Subramanyam, H. (1970) Respiratory climacteric and chemical changes in the mango fruit Mangifera indica L. Journal of the American Society for Horticultural Science 95, 333–337.
Krishnamurthy, S. and Subramanyam, H. (1973) Pre and postharvest physiology of the mango fruit. Tropical Science 15, 167–193.
Krishnamurthy, S., Patwardhan, M.V. and Subramanyan, H. (1971) Biochemical changes during ripening of the mango fruit. Phytochemistry 10, 2577–2581.
Kumar, R. and Selvaraj, Y. (1990) Fructose-1,6-biophosphatase in ripening mango (Mangifera indica L.) fruit. Indian Journal of Experimental Biology 28, 284–287.
Kumar, S., Das, D.K., Singh, A.K. and Prasad, U.S. (1993) Changes in non-volatile or-ganic acid composition and pH during maturation and ripening of two mango cul-tivars. Indian Journal of Plant Physiology 36, 107–111.
Kumar, S., Das, D.K., Singh, A.K. and Prasad, U.S. (1994) Sucrose metabolism during matu-ration and ripening of mango cultivars. Plant Physiology and Biochemistry 21, 27–32.
Kundu, S. and Ghosh, S.N. (1992) Studies on physico-chemical characteristics of mango cultivars grown in the laterite tract of West Bengal. Haryana Journal of Horticul-tural Sciences 21(3/4), 129–139.
Lakshminarayana, S. (1973) Respiration and ripening patterns in the life cycle of the mango fruit. Journal of Horticultural Science 48, 227–233.
Lakshminarayana, S. (1975) Relation of time of harvest on respiration, chemical con-stituents and storage life of mangoes. Proceedings of the Florida State Horticultural Society 88, 477–480.
Lakshminarayana, S. (1980) Mango. In: Nagy, S. and Shaw, P.E. (eds) Tropical and Sub-tropical Fruits. AVI Publishing Co., Westport, Connecticut, pp. 184–257.

J.K. Brecht and E.M. Yahia522
Lakshminarayana, S. and Subramanyam, H. (1970) Carbon dioxide injury and fermenta-tive decarboxylation in mango fruit at low temperature storage. Journal of Food Science and Technology 7, 148–152.
Lakshminarayana, S., Subhadra, N.V. and Subramanyam, H. (1970) Some aspects of the developmental physiology of the mango fruit. Journal of Horticultural Science 45, 133–142.
Lakshminarayana, S., Shety, M.S. and Krishnaprasad, C.A. (1974) Accelerated ripening of ‘Alphonso’ mangos by the application of ethrel. Tropical Science 17, 95–101.
Lalel, H.J.D., Singh, Z. and Tan, S.C. (2003) Maturity stage at harvest affects fruit ripen-ing, quality and biosynthesis of aroma volatile compounds in ‘Kensington Pride’ mango. Journal of Horticultural Science and Biotechnology 78, 225–233.
Lazan, H., Ali, Z.M., Lee, K.W. and Woon, J. (1986) The potential role of polygalactu-ronase in pectin degradation of softening of mango. ASEAN Food Journal 2, 93–98.
Lazan, H., Ali, Z.M. and Sani, H.A. (1990) Effect of Vaporgard on polygalacturonase, malic enzyme and ripening in mango. Acta Horticulturae 269, 359–366.
Lazan, H., Ali, Z.M., Soh, J. and Talkan, Z. (1993) The biochemical basis of differential ripening in mango. Acta Horticulturae 341, 500–509.
Lederman, I.E., Zauberman, G., Weksler, A., Rot, I. and Fuchs, Y. (1997) Ethylene-forming capacity during cold storage and chilling injury development in ‘Keitt’ mango fruit. Postharvest Biology and Technology 10, 107–112.
Ledger, S.N. (1986) Mango packaging. In: Proceedings of the First Australian Mango Research Workshop. Commonwealth Scientifi c and Industrial Research Organiza-tion (CSIRO), Melbourne, pp. 314–319.
Lizada, C. (1993) Mango. In: Seymour, G.B., Taylor, J.E. and Tucker, G.A. (eds) Biochem-istry of Fruit Ripening. Chapman and Hill, London, pp. 255–271.
Lizada, M.C. (1991) Postharvest physiology of the mango – a review. Acta Horticulturae219, 437–452.
MacLeod, A.J. and De Troconis, N.G. (1982) Volatile components of some mango fruit. Phytochemistry 21, 2523–2526.
MacLeod, A.J. and Pieris, N.M. (1984) Comparison of volatile components in some mango cultivars. Phytochemistry 23, 361–366.
MacLeod, A.J. and Snyder, C.H. (1985) Volatile components of two cultivars of mango from Florida. Journal of Agricultural and Food Chemistry 33, 380–384.
MacLeod, A.J., MacLeod, G. and Snyder, C.H. (1988) Volatile aroma constituents of mango (cv. Kensington). Phytochemistry 27, 2189–2193.
Mahayothee, B., Muhlbauer, W., Neidhart, S. and Carle, R. (2004) Infl uence of posthar-vest ripening process on appropriate maturity for drying mangoes ‘Nam Dokmai’ and ‘Kaew’. Acta Horticulturae 645, 241–248.
Malundo, T.M.M. (1996) Application of the quality enhancement (QE) approach to man-go (Mangifera indica L.) fl avor research. PhD dissertation, Department of Food Sci-ence and Technology, University of Georgia, Athens, Georgia, USA, 134 pp.
Malundo, T.M.M., Shewfelt, R.L., Ware, G.O. and Baldwin, E.A. (2001) Sugars and acids infl uence fl avor properties of mango (Mangifera indica). Journal of the American Society for Horticultural Science 126, 115–121.
Mann, S.S. and Singh, R.N. (1976) The cold storage life of ‘Deshari’ mangoes. Scientifi c Horticulture 5, 249–254.
Mattoo, A.K. and Modi, V.V. (1969a) Biochemical aspects of ripening and chilling injury in mango fruit. Proceedings of the International Conference on Tropical and Sub-tropical Fruit. Tropical Products Institute, London, pp. 111–115.
Mattoo, A.K. and Modi, V.V. (1969b) Ethylene and ripening of mangos. Plant Physiology44, 308–310.

Postharvest Physiology 523
Mattoo, A.K., Modi, V.V. and Reddy, V.V.R. (1968) Oxidation and carotenogenesis regu-lating factors in mangoes. Indian Journal of Biochemistry 5, 111–114.
McCollum, T.G., D’Aquino, S. and McDonald, R.E. (1993) Heat treatment inhibits man-go chilling injury. HortScience 28, 197–198.
Medlicott, A.P. and Thompson, A.K. (1985) Analysis of sugar and organic acids in ripen-ing mango fruit (Mangifera indica var. Keitt) by high performance liquid chromatog-raphy. Journal of the Science of Food and Agriculture 36, 561–566.
Medlicott, A.P., Bhogal, M. and Reynolds, S.B. (1986) Changes in peel pigmentation during ripening of mango fruit (Mangifera indica var. Tommy Atkins). Annals of Ap-plied Biology 109, 651–656.
Medlicott, A.P., Reynolds, S.B., New, S.W. and Thompson, A.K. (1988) Harvest maturity effects on mango fruit ripening. Tropical Agriculture 65, 153–157.
Medlicott, A.P., Sigrist, J.M.M. and Sy, O. (1990) Ripening of mangoes following low temperature storage. Journal of the American Society for Horticultural Science 115, 430–434.
Mercadante, A.Z. and Rodriguez-Amaya, D.B. (1998) Effects of ripening, cultivar differ-ences and processing on the carotenoid composition of mango. Journal of Agricul-tural and Food Chemistry 46, 128–130.
Mercadante, A.Z., Rodriguez-Amaya, D.B. and Britton, G. (1997) HPLC and mass spec-trometric analysis of carotenoids from mango. Journal of Agricultural and Food Chemistry 45, 120–123.
Miller, W.R., Hale, P.W., Spalding, D.H. and Davis, P. (1983) Quality and decay of mango fruit wrapped in heat-shrinkable fi lm. HortScience 18, 957–958.
Miller, W.R., Spalding, D.R. and Hale, P.W. (1986) Film wrapping mangoes at advancing stages of postharvest ripening. Tropical Science 26, 9–17.
Mitcham, E.J. and McDonald, R.E. (1992) Cell wall modifi cation during ripening of ‘Keitt’ and ‘Tommy Atkins’ mango fruit. Journal of the American Society for Horti-cultural Science 117, 919–924.
Mitcham, E.J. and McDonald, R.E. (1997) Effects of postharvest heat treatments on inner and outer tissue of mango fruit. Tropical Science 37, 193–205.
Mitra, S.K. and Baldwin, E.A. (1997) Mango. In: Mitra, S.K. (ed.) Postharvest Physiology and Storage of Tropical and Subtropical Fruits. CAB International, Wallingford, UK, pp. 85–122.
Modi, V.V. and Reddy, V.V.R. (1967) Carotenogenesis in ripening mangoes. Indian Jour-nal of Experimental Biology 5, 233–235.
Mohammed, M. and Brecht, J.K. (2002) Reduction of chilling injury in ‘Tommy Atkins’ mangoes during ripening. Scientia Horticulturae 95, 297–308.
Morga, N.S., Lustre, A.O., Tunac, M.M., Balagot, A.H. and Soriano, M.R. (1979) Physi-cochemical changes in Philippine ‘Carabao’ mangoes during ripening. Food Chem-istry 4, 225–234.
Muda, P., Seymour, G.B., Errington, N. and Tucker, G.A. (1995) Compositional changes in cell wall polymers during mango fruit ripening. Carbohydrate Polymers 26, 255–260.
Mukherjee, S.K. (1958) The origin of mango. Indian Journal of Horticulture 15, 129–134.Musa, S.K. (1974) Preliminary investigations on the storage and ripening of ‘Totapuri’
mangoes in the Sudan. Tropical Science 16, 65–73.Nasrijal, N.H. (1993) Effect of storage temperature on pectin depolymerization and soft-
ening of mango fruit. BSc thesis, Department of Botany, Universiti Kebangsaan, Bangi, Malaysia.
Noomhorm, A. and Tiasuwan, N. (1995) Controlled atmosphere storage of mango fruit, Mangifera indica L. cv. Rad. Journal of Food Processing and Preservation 19, 271–281.

J.K. Brecht and E.M. Yahia524
Nuevo, P.A., Cua, A.U. and Lizada, M.C.C. (1984a) Internal breakdown in ‘Carabao’ mango subjected to modifi ed atmospheres. III. Starch in the spongy tissue. Posthar-vest Research Notes 1, 63–64.
Nuevo, P.A., Pantastico, E.R.B. and Mendosa, D.B. (1984b) Gas diffusion factors in fruits I. Anatomical structure in mango fruit. Postharvest Research Notes 1, 1–5.
O’Hare, T.J. and Prasad, A. (1993) The effect of temperature and carbon dioxide on chill-ing symptoms in mango. Acta Horticulturae 343, 244–250.
Ornelas-Paz, J. de J., Yahia, E.M. and Gardea, A. (2007) Identifi cation and quantifi cation of xanthophyll esters, carotenes and tocopherols in the fruit of seven Mexican man-go cultivars by liquid chromatography-APcI+-time of fl ight mass spectrometry. Jour-nal of Agricultural and Food Chemistry 55, 6628–6635.
Ornelas-Paz, J. de J., Yahia, E.M. and Gardea, A.A. (2008) Changes in external and internal color during postharvest ripening of ‘Manila’ and ‘Ataulfo’ mango fruit and relationship with carotenoid content determined by liquid chromatography–APcI+-time-of-flight mass spectrometry. Postharvest Biology and Technology50, 145–152.
Ortega, D. and Yahia, E.M. (2000) Tolerance and quality of mango fruit exposed to con-trolled atmospheres at high temperatures. Postharvest Biology and Technology 20, 195–201.
Parikh, H.R., Nair, G.M. and Modi, V.V. (1990) Some structural changes during ripening of mangoes (Mangifera indica var. Alphonso) by abscisic acid treatment. Annals of Botany 65, 121–127.
Park, Y.K., Sato, H.H., Almeida, T.D. and Moretti, R.H. (1980) Polyphenoloxidase of mango (Mangifera indica var. Haden). Journal of Food Science 45, 1619–1621.
Peacock, B.C. (1986) Postharvest handling of mangoes. In: Proceedings of the First Australian Mango Research Workshop. Commonwealth Scientifi c and Industrial Research Organization (CSIRO), Melbourne, pp. 295–313.
Peacock, B.C., Murray, C., Kosiyachinda, S., Kasittrakakul, M. and Tansiriyakul, S. (1986) Infl uence of harvest maturity of mangoes on storage potential and ripe fruit quality. ASEAN Food Journal 2, 99–103.
Percival, S.S., Talcott, S.T., Chin, S.T., Mallak, A.C., Lounds-Singleton, A. and Pettit-Moore, J. (2006) Neoplastic transformation of BALB/3T3 cells and cell cycle of HL-60 cells are inhibited by mango (Mangifera indica L.) juice and mango juice extract. Journal of Nutrition 136, 1300–1304.
Pesis, E., Marinansky, R., Rot, I. and Weksler, A. (1994) Effect of postharvest application of acetaldehyde or a short period of anaerobiosis on fruit senescence. Proceedings of the Royal Society Edinburgh 102b, 473–478.
Pino, J., Rosado, A. and Sanchez, R. (1989) Volatile components of three cultivars of mango from Cuba. Die Nahrung 33, 709–715.
Pino, J., Mesa, J., Muñoz, Y., Martí, M.P. and Marbot, R. (2005) Volatile components from mango (Mangifera indica L.) cultivars. Journal of Agricultural and Food Chem-istry 53, 2213–2223.
Pott, I., Breithaupt, D.E. and Carle, R. (2003a) Detection of unusual carotenoid esters in fresh mango (Mangifera indica L. cv. ‘Kent’). Phytochemistry 64, 825–829.
Pott, I., Marx, M., Neidhart, S., Mühlbauer, W. and Carle, R. (2003b) Quantitative deter-mination of -carotene stereoisomers in fresh, dried, and solar-dried mango (Mangifera indica L.). Journal of Agricultural and Food Chemistry 51, 4527–4531.
Proctor, J.T.A. and Creasy, L.L. (1969) The anthocyanins of the mango fruit. Phytochem-istry 8, 2108.
Quinones, V.L., Guerrant, N.B. and Adams Dutcher, R. (1944) Vitamin content of some tropical fruits, their juices and nectars. Journal of Food Science 9, 415–417.

Postharvest Physiology 525
Raymond, L., Schaffer, B., Brecht, J.K. and Crane, J.H. (1998) Internal breakdown in mango fruit: symptomology and histology of jelly seed, soft nose and stem-end cav-ity. Postharvest Biology and Technology 13, 59–70.
Reddy, Y.V. and Srivastava, G.C. (1999) Ethylene biosynthesis and respiration in mango fruits during ripening. Indian Journal of Plant Physiology 4, 32–35.
Rocha-Ribeiro, S.M., Queiroz, J.H., Lopes, M., Campos, F.M. and Pinheiro, H.M. (2007) Antioxidant in mango (Mangifera indica L.) pulp. Plant Foods for Human Nutrition 62, 13–17.
Rodov, V., Ben Yehoshua, S., Fishman, S., Gotlieb, S., Fierman, T. and Fang, D.Q. (1994) Reducing decay and extending shelf life of bell peppers and mangoes by modifi ed atmosphere packaging. In: Champ, B.C., Highley, E. and Johnson, G.I. (eds) Post-harvest Handling of Tropical Fruits. Australian Centre for International Agricultural Research (ACIAR) Proceedings 50. ACIAR, Canberra, pp. 416–418.
Roe, B. and Bruemmer, J.H. (1981) Changes in pectic substances and enzymes during ripening and storage of ‘Keitt’ mangos. Journal of Food Science 46, 186–189.
Rojas-Villegas, A., Carrillo-Lopez, A., Silveira, M., Avena-Bustillos, R. and Yahia, E.M. (1996) Effects of insecticidal atmospheres on the mortality of fruits fl ies in mango. Acta Horticulturae 370, 89–92.
Sadasivam, R., Muthuswany, S., Sundaraj, J.S. and Vasudevan, V. (1971) Note on chilling injury in mango (Mangifera indica L.) fruits in refrigerated storage. Indian Journal of Agricultural Science 41, 715–716.
Sane, V.A., Chourasia, A. and Nath, P. (2005) Softening in mango (Mangifera indica cv. Dashehari) is correlated with the expression of an early ethylene responsive, rip-ening related expansin gene, MiExpA1. Postharvest Biology and Technology 38, 223–230.
Sarker, S. and Muhsi, A.A. (1981) A study on the content and interconversions of or-ganic acids of mango (Mangifera indica L.) at various stages of fruit development. Bangladesh Journal of Agricultural Science 8, 69–75.
Selvaraj, Y. and Kumar, R. (1989) Studies on fruit softening enzyme and polyphenoloxi-dase activity in ripening mango (Mangifera indica L.) fruit. Journal of Food Science and Technology 26, 218–222.
Selvaraj, Y. and Kumar, R. (1994) Enzymatic regulation in ripening mango fruit. IndianJournal of Horticulture 51, 316–323.
Selvaraj, Y., Kumar, R. and Pal, D.K. (1989) Changes in sugars, organics acids, amino acids, lipid constituents and aroma characteristics of ripening mango (Mangiferaindica L.) fruit. Journal of Food Science and Technology 26, 306–311.
Setiawan, B., Sulaeman, A., Giraud, D.W. and Driskell, J.A. (2001) Carotenoid content of selected Indonesian fruits. Journal of Food Composition and Analysis 14, 169–176.
Seymour, G.B., N’Diaye, M., Wainwright, H. and Tucker, G.A. (1990) Effect of cultivar and harvest maturity on ripening of mangoes during storage. Journal of Horticul-tural Science 65, 479–483.
Shashirekha, M.S. and Patwardhan, M.V. (1976) Changes in amino acids, sugars and nonvolatile organic acids in a ripening mango fruit (Mangifera indica L. Badami variety). Lebensmittel-Wissenschaft und Technologie 9, 369–370.
Siddappa, G.S. and Bhatia, B.S. (1954) Tender green mangoes as source of vitamin C. Indian Journal for Horticulture Science 11, 104–106.
Singh, L.B. (1960) The Mango. Leonard Hill, London.Sornsrivichai, J. and Waru-Aswapti, O. (1989) The effect of maturity and ripening tem-
perature on fruit quality of ethrel treated mangoes. (Abstract). In: Proceedings of the Third International Mango Symposium, Darwin, Northern Territory, Australia, p. 25.

J.K. Brecht and E.M. Yahia526
Sornsrivichai, S., Gomolmanee, S., Boonyakiat, D., Uthaibutra, J., Boon-Long, P. and Gemma, H. (1992) Seal packaging by plastic fi lm as a technique for limiting fungal decay of mangoes. Acta Horticulturae 296, 23–32.
Srivastava, H.C. (1967) Grading, storage and marketing. In: The Mango: a Handbook.Indian Council of Agricultural Research, New Delhi, India, pp. 99–109.
Stahl, A.L. (1935) Composition of Miscellaneous Tropical and Subtropical Florida Fruits.Bulletin 283. University of Florida Agricultural Experiment Station, Gainesville, Florida.
Stead, D.E. and Chithambo, G.S.G. (1980) Studies on the storage of tropical fruits in polyethylene bags. Luso: Journal of Science and Technology (Malawi) 1, 3–9.
Subramanyam, H., Krishnamurthy, S., Subhadra, N.V., Dalal, V.B., Randhawa, G.S. and Chacko, E.K. (1971) Studies on internal breakdown, a physiological ripening disor-der in ‘Alphonso’ mangoes (Mangifera indica L.). Tropical Science 13, 203–210.
Subramanyam, H., Krishnamurthy, S. and Parpia, H.A.B. (1975) Physiology and bio-chemistry of mango fruit. Advances in Food Research 21, 223–305.
Sy, D.A. and Mendoza, D.B., Jr (1984) Respiratory responses of ‘Carabao’ mango to dif-ferent levels of oxygen. Postharvest Research Notes 1, 3–4.
Tahir, M.A. and Malik, A.A. (1977) The activity of pectic enzymes (pectinesterase, polyga-lacturonase) in various mango products. Pakistan Journal of Research 29, 71–74.
Tandon, D.K. and Kalra, S.K. (1983) Changes in sugars, starch and amylase activity dur-ing development of mango fruit cv. Dashehari. Journal of Horticultural Science 58, 449–453.
Tandon, D.K. and Kalra, K. (1984) Pectin changes during the development of mango fruit cv. Dashehari. Journal of Horticultural Science 59, 283–286.
Tharanathan, R.N., Yashoda, H.M. and Prabha, T.N. (2006) Mango (Mangifera indica L.), the king of fruits – an overview. Food Reviews International 22, 95–123.
Thomas, P. (1975) Effect of postharvest temperatures on quality, carotenoids and ascor-bic acid contents in ‘Alphonso’ mangos on ripening. Journal of Food Science 40, 704–706.
Tirmazi, S.I.H. and Wills, R.B.H. (1981) Retardation of ripening of mangoes by posthar-vest application of calcium. Tropical Agriculture 58, 137–141.
United States Department of Agriculture (USDA)/Agricultural Research Service (ARS) (2007) USDA National Nutrient Database for Standard Reference, Release 20. Nutrient Data Laboratory Home Page. Available at: http://www.ars.usda.gov/nutri-entdata (accessed 28 August 2008).
van Eeden, S.J. (1992) Calcium infi ltration as a possible postharvest treatment to in-crease storage potential of mango fruit. South African Mango Growers’ Association Yearbook 12, 26–27.
van Lelyveld, L.J. and Smith, J.H.E. (1979) Physiological factor in maturation and ripen-ing of mango (Mangifera indica) fruit in relation to the jelly seed physiological dis-order. Journal of Horticultural Science 54, 283–287.
Vasanthaiah, H.K.N., Ravishankar, K.V., Shivashankara, K.S., Kodthalu, S., Anand, L., Narayanaswamy, P., Mukunda, G. and Prasad, T.G. (2006) Cloning and character-ization of differentially expressed genes of internal breakdown in mango fruit (Mangifera indica). Journal of Plant Physiology 163, 671–679.
Vázquez-Caicedo, A.L., Neidhart, S. and Carle, R. (2004) Postharvest ripening behav-iour of nine Thai mango cultivars and their suitability for industrial applications. Acta Horticulturae 645, 617–625.
Vazquez-Salinas, C. and Lakshminarayana, D. (1985) Compositional changes in mango fruit during ripening at different storage temperatures. Journal of Food Science 50, 1646–1648.

Postharvest Physiology 527
Vinci, G., Botre, F. and Mele, G. (1995) Ascorbic acid in exotic fruits: a liquid chromato-graphic investigation. Food Chemistry 53, 211–214.
Wilberg, V.C. and Rodriguez-Amaya, D.B. (1995) HPLC quantitation of major carote-noids of fresh and processed guava, mango and papaya. Lebensmittel-Wissenschaft und Technologie 28, 474–480.
Wills, R.B.H., Yuen, C.M.C., Sabri Laksmi, L.D.S. and Suyanti (1988) Effect of calcium infi ltration on delayed ripening of three mango cvs in Indonesia. ASEAN Food Jour-nal 4, 67–58.
Wills, R.B.H., Warton, M.A., Mussa, D.M.D.N. and Chew, L.P. (2001) Ripening of cli-macteric fruits initiated at low ethylene levels. Australian Journal of Experimental Agriculture 41, 89–92.
Wilson, C.W., Shaw, P.E. and Knight, R.J. (1990) Importance of some lactones and 2,5 dimethyl-4-hydroxy-3(2H)-furanone to mango (Mangifera indica L.) aroma. Journalof Agricultural and Food Chemistry 38, 1556–1559.
Wolf, G. (1984) Multiple functions of vitamin A. Physiological Reviews 64, 873–937.Yahia, E.M. (1993) Responses of some tropical fruits to insecticidal atmospheres. Acta
Horticulturae 343, 371–376.Yahia, E.M. (1994) The potential use of insecticidal atmospheres for mango, avocado,
and papaya fruits. In: Champ, B.C., Highley, E. and Johnson, G.I. (eds) Postharvest Handling of Tropical Fruits. Australian Centre for International Agricultural Re-search (ACIAR) Proceedings 50. ACIAR, Canberra, pp. 373–374.
Yahia, E.M. (1995) Application of differential scanning calorimetry in the study of avo-cado and mango fruit responses to hypoxia. In: Ait-Oubahou, A. and El-Otmani, M. (eds) Postharvest Physiology, Pathology and Technologies for Horticultural Crops: Recent Advances. Institut Agronomique and Vétérinaire Hassan II, Agadir, Moroc-co, pp. 206–209.
Yahia, E.M. (1997) Modifi ed/controlled atmospheres for mango. In: Kader, A.A. (ed.) Proceedings of CA´97, Vol. 3: Fruits Other Than Apples and Pears. University of California, Davis, California, pp. 110–116.
Yahia, E.M. (1998) Modifi ed and controlled atmospheres for tropical fruits. Horticultural Reviews, 22, 123–183.
Yahia, E.M. and Tiznado, M. (1993) Tolerance and responses of harvested mango to in-secticidal oxygen atmospheres. HortScience 28, 1031–1033.
Yahia, E.M. and Vazquez, L. (1993) Tolerance and responses of mango to insecticidal oxygen and carbon dioxide atmospheres. Food Science and Technology (Lebensmittel-Wissenschaft und Technologie) 26, 42–48.
Yahia, E.M., Medina, F. and Rivera, M. (1989) The tolerance of mango and papaya to atmospheres containing very high levels of CO2 and/or very low levels of O2 as a possible insect control treatment. In: Fellman, J.K. (ed.) Fifth Proceedings of Interna-tional CA Research Conference, Vol. 2 – Other Commodities and Storage Recom-mendations, Wenatchee, Washington, pp. 77–89.
Yahia, E.M., Villagomez, G. and Juarez, A. (2000) Tolerance and responses of different fruits and vegetables to hot air treatments at 43°C or 48°C and 50% RH for 160 minutes. In: Artes, F., Gil, M.I. and Conesa, M.A. (eds) Improving Postharvest Tech-nologies of Fruits, Vegetables and Ornamentals. Vol. 2. International Institute of Refrigeration, Paris, pp. 714–718.
Yahia, E.M., Ornelas-Paz, J. de J. and Ariza, F.R. (2006a) The Mango. Editorial Trillas, México City, Mexico, 224 pp. (in Spanish)
Yahia, E.M., Ornelas-Paz, J. de J. and Gardea, A. (2006b) Extraction, separation and partial identifi cation of ‘Ataulfo’ mango fruit carotenoids. Acta Horticulturae 712,333–338.

J.K. Brecht and E.M. Yahia528
Yamashita, F., Benassi, M.T. and Kieckbusch, T.G. (1997) Shelf life extension of individu-ally fi lm-wrapped mangoes. Tropical Science 37, 249–255.
Yamashita, F., Benassi, M.T. and Kieckbusch, T.G. (1999) Effect of modifi ed atmosphere packaging on the kinetics of vitamin C degradation in mangos. Brazilian Journal of Food Technology 2, 127–130.
Yantarasri, T., Uthaibutra, J., Sornrivichai, J., Kumpuan, W., Sardsud, V. and Kanathum, N. (1994) Modifi ed atmosphere packaging by perforated polymeric fi lm and its effect on physical properties of mango fruit. In: Champ, B.R., Highley, E. and Johnson, G.I. (eds) Postharvest Handling of Tropical Fruits. Australian Centre for International Agricultural Research (ACIAR) Proceedings 50. ACIAR, Canberra, pp. 438–440.
Yantarasri, T., Ben Yehoshua, S., Rodov, V., Kumpuan, W., Uthaibutra, J. and Sornsrivichal, J. (1995) Development of perforated modifi ed atmosphere package for mango. Acta Horticulturae 398, 81–91.
Young, T.W. (1957) ‘Soft nose’ a physiological disorder in mango fruit. Proceedings of the Florida State Horticultural Society 70, 280–283.
Yuen, C.M.C., Tan, S.C., Joyce, D. and Chettri, P. (1993) Effect of postharvest calcium and polymeric fi lms on ripening and peel injury in ‘Kensington Pride’ mangoes.ASEAN Food Journal 8, 110–113.
Yuyama, L.K.O., Duarte Favaro, R.M., Yuyama, K. and Vannucchi, H. (1991) Bioavail-ability of vitamin A from peach palm (Bactris gasipaes H.B.K.) and from mango (Mangifera indica L.) in rats. Nutrition Research 11, 1167–1175.
Zainal, Z., Tucker, G.A. and Lycett, G.W. (1999) Isolation and characterisation of a cDNA encoding 1-aminocyclopropane-1-carboxylate oxidase from mango (Mangifera indica L.). Asia-Pacifi c Journal of Molecular Biology and Biotechnology 7, 53–59.
Zambrano, J. and Manzano, J. (1995) Efecto de la aplicación de sales de calcio sobre la maduración de frutos de mango. Agronomía Tropical 45, 407–413.

© CAB International 2009. The Mango, 2nd Edition: Botany, Production and Uses(ed. R.E. Litz) 529
15 Postharvest Technology and Quarantine Treatments
G.I. Johnson1 and P.J. Hofman2
1Horticulture 4 Development, Jamison, Australian Capital Territory, Australia 2Department of Primary Industries and Fisheries, Nambour, Queensland, Australia
15.1 Introduction 53015.2 Considerations Infl uencing Postharvest Requirements 531
Market and consumer research 531Quality assurance (QA) and Good Agricultural Practice (GAP) 531
15.3 Preharvest Management 535Maturity 535Adjusting maturation time 538Skin colour and lenticel damage 538Storage life and physiological disorders 540Pests and diseases 542Weather conditions 544
15.4 Flavour and Aroma 54415.5 Harvesting and Transport to the Packhouse 544
Timing 544Sapburn 545Harvesting and desapping 546Transport to the packhouse 548
15.6 Packhouse Measures 548Delivery inspection and traceability 550Desapping and washing 550Disease control 550Brushing 554Grading and sizing 554Grade standards 555Disinfestation 557
15.7 Preparing Fruit for Market 566Surface coatings 566Packaging 567Inspection 568Palletizing 568Precooling 569

G.I. Johnson and P.J. Hofman530
Ethylene and ripening 56915.8 Pre- and Post-shipping Storage 571
Cool storage 571Controlled and modifi ed atmosphere storage 573
15.9 Transport 57515.10 Marketing 578
Networks and cooperatives 578Promotion and consumer education 578
15.11 Conclusions 579
15.1 Introduction
Postharvest handling of mangoes is the last phase (from the tree to mouth) of an agribusiness venture. To optimize productivity, profi table uses for all grades of fruit should be sought, and stable employment provided for key skilled staff. Sustainable land and water management, and compliance with health, safety and fi nancial obligations to employees are also necessary. Increasingly, Good Agricultural Practice (GAP) protocols need to be observed (GAP, 2003; FFTC-GAP, 2007).
This chapter reviews the technology and quarantine treatments that have been developed for the postharvest handling of mangoes. Peacock (1986), Led-ger (1986, 1991b) and Opara and Nguyen (1999) have previously reviewed postharvest handling of mangoes, while several others have reviewed spe-cifi c aspects of the topic (Johnson and Coates, 1993; Heather, 1994; Jacobiet al., 2001b; Singh et al., 2004; Yahia, 2006). Less than 10% of total world mango production is exported. Export markets for mangoes have expanded because of social changes and rising demand, increased international air cargo space for some sectors and promotion of export fruit production in developing countries (Procter and Cropley, 1994; FAOSTAT, 2008). Expan-sion of production to meet the supply requirements of export and distant domestic markets has been possible because of successful integrated man-agement strategies and disinfestation technologies to control diseases and insects (Johnson and Coates, 1993; Johnson and Heather, 1995; Ploetz, 2007), and increased land availability due to deforestation and diversifi cation away from rice. Market development has also been facilitated through harmoniza-tion of the rules of trade between nations and regions at global and near global levels (WTO, 2008), agreement on pest and disease risk management under the International Plant Protection Convention (IPPC), various bilateral and regional Free Trade Agreements (FTA, 2007), and the global expansion of supermarkets. Simultaneously, knowledge of and concerns about exotic pest risks to domestic fruit production, socio-political concerns about chemical residues on food, environmental management and labour conditions, and rising production and marketing costs, have impinged upon market access and stimulated international dialogue and research initiatives which address these concerns (Buchanan, 1994; Gullino and Kuijpers, 1994; Ploetz, 2003, 2007).

Postharvest Technology 531
15.2 Considerations Infl uencing Postharvest Requirements
Market and consumer research
Strategies and procedures for horticultural market research have been out-lined by Minnis (1993, 1994), Hall et al. (2001) and Kitinoja and Kader (2003). Increasingly, a supply chain approach is being taken (Johnson and Hofman, 2004). Hofman and Ledger (2006) proposed that the supply chain approach should be used to guide research and development and that there needs to be a champion in the supply chain with signifi cant infl uence and a desire for improving chain status and performance. Key features are identifi cation of market demand, through-put, price and profi t fl ows, seasonal fl uctuations in availability and demands, supply competitors (and their commodity statis-tics), importer-buyer requirements and relationships, options for value-adding, and consumer expectations and sales promotions. Point-of-sale transaction data from supermarkets and other sales outlets in target markets can be an invaluable source of intelligence and can be purchased from marketing infor-mation specialists. Detailed supply chain assessments can guide options for innovation and improvement (Johnson and Hofman, 2004).
The volume, grade and quality of product available for export requires analysis in relation to buyer specifi cations, retail customer and provedore preferences, technological and regulatory requirements for supplying the market, and the production, packaging, cooling and transportation proto-cols/options that are needed/available or specifi ed under agreed codes of practice (NRI, 2008a). Procter and Cropley (1994), Mahendra et al. (2002) and the Natural Resources Institute (NRI) (2008b) provide some perspectives on these issues.
Quality assurance (QA) and Good Agricultural Practice (GAP)
Postharvest handling assures timely delivery of a product that closely matches buyer specifi cations and complies with mandatory regulatory require-ments. Satisfying customers underpins quality assurance (QA) and obser-vance of GAP (Box 15.1), which aims to produce a product of the desired standard, compliant with regional or internationally agreed standards in production and handling, and encourage regular, larger and more frequent purchases, and brand loyalty. As export markets become increasingly com-petitive, responsive observance of QA and GAP certifi cation can be vital for maintaining and expanding market niche (Johnson and Coates, 1991; Askar and Treptow, 1993; Ledger and Premier, 2006). Piñeiro et al. (2004) provide generic guidelines for the development of quality and safety guidelines for fresh produce, and Ledger and Premier (2006) provide guidelines for a Mango Quality Plan.
Components of a postharvest handling system may be developed in commercial confi dence to enhance brand-name reputation and increase mar-ket share. Increasingly they are also agreed under contractual agreements

G.I. Johnson and P.J. Hofman532
with supermarkets or exporters. No regulations govern the pursuit of supe-rior but legal agribusiness practices that may enhance grower and seller rep-utations or provide additional advantages. Regulations govern chemical residues, pests, diseases, product and packaging specifi cations. The sections that follow need to be considered in developing the postharvest components of GAP protocols and dynamic QA systems.
Regulatory restrictions and quarantine treatmentsQuarantine treatments disinfest produce of target pests. They are a critical component of protocols designed to satisfy market-stipulated prohibitions against pest entry to countries, or regions within countries. Protocols stipulate procedures for monitoring, detecting, eliminating and handling pest-affected
Box 15.1. What is Good Agricultural Practice (GAP)?
GAP enhances trader-buyer relationships and the reputation of producers and traders for the suitability of produce and environmental care. A key feature of GAP is the role of independent certifying authorities (usually private sector) in accreditation and compliance monitoring, to provide additional assurance to buyers of certifi cation creditability. GAP can be applied to any farming system or scale. Key elements include: sustainable practices such as integrated pest management (IPM), responsible fertilizer use and care for the environment. It relies on four key principles (Wikipedia, 2007; NRI, 2008b):
Economic and effi cient production of adequate supplies of safe and nutri-•tious foods or other agricultural commodities.Compliance with sustainable practices which improve natural resource •availability.Assuring the economic and environmental sustainability of the farming •enterprise.Meeting social and cultural norms and expectations in production and •marketing.
GAP has gained global prominence because of the trend to making it a ‘com-pulsory standard’ for exporters to Europe. A private sector initiative, EurepGAP sets ‘voluntary standards’ for agricultural product certifi cation anywhere. It involves partnership between farmers and retailers wanting to establish and implement procedures and standards for certifying application and compliance in GAP. It is applied before the farm-gate (from planting to market dispatch), and is not nec-essarily a standard that is displayed to customers. EurepGAP involves an array of documentation including general regulations, control points, compliance cri-teria and checklists (EurepGAP, 2007).
In 2007, EurepGAP transformed into GLOBALGAP (GLOBALGAP, 2007) that could be tailored and adopted anywhere in the world. In parallel with the Eurep-GAP process, the South-east Asian countries have been working towards Association of South-east Asian Nations (ASEAN) GAP standards suited to fruit marketing within ASEAN (ASEAN-GAP, 2007; FFTC-GAP, 2007; Johnsonet al., 2008).

Postharvest Technology 533
produce. Frequently, a detailed pest risk analysis (PRA) is required, protocol effi cacy against pests of quarantine concern must be demonstrated, appli-cator integrity scrutinized and market-access requirements and rights ap-proved by regulatory and agricultural authorities in the importing and exporting country or region. Increasingly, systems approaches are being applied. These minimize the need for reliance on quarantine treatments by including PRA, production-based pest management systems, consideration of establishing pest-free areas or identifying commodities which are non-hosts of quarantine pests.
Restrictions or limitations on pesticide residues and other contaminants in marketed produce are also considerations in the choice of quarantine treat-ment and market access. Pesticide residue monitoring protocols are often required (Johnson and Heather, 1995; Sharp and Heather, 2002; McMaugh, 2005). By restricting or preventing market access, quarantine and pesticide residue restrictions can unintentionally operate as quasi-trade barriers, effec-tively reducing competition with domestic production of the same or alter-native products. However, this is decreasing because of better adherence to science-based decision making as the foundation for quarantine restrictions (WTO, 2008). Government authority web sites and export agencies can pro-vide information on quarantine and tariff barriers, pesticide use restrictions, inspection, packaging and labelling regulations (PPQ, 2007; IPPC, 2008) (see Disinfestation section under 15.6 Packhouse Measures, this chapter). Phyto-sanitary requirements for fresh mango exports to some markets are shown in Table 15.1. Marketing through reputable exporters, known to the importing country as suppliers of produce that comply with regulatory restrictions, can encourage producer and importer confi dence.
Limitations of the productMangoes ripen rapidly and have low tolerance of temperatures <10°C. Post-harvest handling procedures for mangoes aim to optimize quality and mini-mize premature ripening and fruit damage. Precise maintenance of fruit quality and the storage environment demands inputs at every stage from picking to the consumer (Ledger, 1991b; Milne, 1994; Ledger et al., 2002b).
Supply, logistics and transportMarket development cannot succeed without a reliable supply of suitable or market-compliant produce. Production seasons are usually short (2–8 weeks), cultivar appearance and fl avour diverse, maturation time variable, and orchards sometimes small and scattered. Practical solutions to these limita-tions (i.e. sourcing from a range of ecoclimates with differing maturation dates, or fl ower-induction technologies) are critical for industry develop-ment. Competitive air-freight rates and rapid road transport, combined with cool-chain handling and atmosphere management can make nearby export markets almost as accessible as distant domestic markets. Special perishable produce rates may be negotiated or subsidized by government, especially during industry establishment. Sea freight is necessary for moving large volumes when air freight is uneconomical. Out-turn problems often arise,

G.I. Johnson and P.J. Hofman534
especially during market development (Snowdon, 1994). Insurance should be arranged to cover losses. Specialist inputs may be required to identify causes of the out-turn problems and in the apportionment of liability amongst producer, shipper and marketer (Snowdon, 1990, 1994).
PersonnelQA systems and GAP protocols must encompass the human component of an organization or business (Bunt and Piccone, 1994; Rolle, 2006; Sonneveld, 2006; FFTC-GAP, 2007). Market agents, exporters, farm/packhouse suppli-ers, fi nance providers and transport personnel and companies need to be selected and worked with from the quality perspective. Human resource development, training and education have been of major signifi cance in the success of many industries.
Table 15.1. Typical phytosanitary requirements for mangoes for some countries.
CountryPhytosanitary requirementsa (usually for mangoes from individual countries or regions on a case-by-case basis)
Australia Approved treatment for fruit fl y, area free of pulp weevil (Sternochetus gravis (F.))
Canada No phytosanitary certifi cate requiredChina Phytosanitary certifi cate required. Field management measures for
specifi c pests of quarantine concern to China plus approved disinfestation treatment for fruit fl ies
EU Phytosanitary certifi cate requiredIndonesia Phytosanitary certifi cate required plus grown in area free of Queensland
and Mediterranean fruit fl yJapan Phytosanitary certifi cate required plus disinfestation schedule approved
for nominated mango cultivars and fruit fl y species and inspection of approved quantity of fruit (2–5%)
Korea Phytosanitary certifi cate required, combined with fi eld surveysMalaysia Must be free of seed weevil on inspectionNew Zealand Phytosanitary certifi cate required plus approved disinfestation schedule
for nominated fruit fl y speciesSaudi Arabia Phytosanitary certifi cate required. Require destructive test of 2% of
consignment for seed weevil, or fi eld survey verifi cation of block freedomSingapore No restrictionsUnited Arab emirates
Phytosanitary certifi cate required. Require destructive test of 2% of consignment for seed weevil, or fi eld survey verifi cation of block freedom
USA Phytosanitary certifi cate required plus disinfestation approved for nominated mango cultivars and fruit fl y species
a General requirements: Prior approval to import is required to access the market of many countries. Packhouse and disinfestation treatment/facility inspections may be required by exporting and importing regulatory authorities. Import permits may cover multiple importations but usually require renewal every 3–12 months. Phytosanitary certifi cates must be issued by a government authority. Fruit must be free of soil and debris and packed in clean, new containers. Timber packaging and pallets will be subject to additional requirements. Consignments found to contain quarantinable pests will be rejected, and either re-exported or destroyed.

Postharvest Technology 535
Ledger and Bagshaw (1994) refer to three general styles of management: (i) defect detection; (ii) defect prevention; and (iii) continuous improvement. They consider that quality management of horticulture has traditionally been the defect detection style, but it is now moving to the continuous improve-ment style via the implementation of QA, total quality management and supply-chain improvement systems. Key ingredients of successful imple-mentation of quality systems and supply chains are: (i) unqualifi ed commit-ment by the owner and senior management of the human, material and other resources needed to introduce and maintain the system; and (ii) employee understanding and active participation in the process (Ledger and Bagshaw, 1994).
Profi tability and sustainabilityHigher freight rates, tariffs and taxes, pesticide-use monitoring and quaran-tine and security clearance times can affect sales negotiations and profi t mar-gins, as can socio-cultural differences among retail buyer, importer, exporter and producer during marketing negotiations and exporter and buyer per-ceptions and consumer expectations of quality. Intended markets, retailers and regional trade fairs should be visited to make personal contacts and to assess suitability of the market and retailing facilities, conditions and pros-pects. Ongoing market monitoring is vital, with regular out-turn inspections by trained personnel, and contingency arrangements for product regrading at destination if required. Prompt, personal attention to client concerns or product problems can be essential for continuing success (Johnson, 1997; Vinning and Young, 2006).
15.3 Preharvest Management
The effects of production practices on fruit quality have been reviewed by Arpaia (1994), Hofman and Smith (1994), Hofman (1998) and Hewett (2006). The fruit characteristics infl uenced by preharvest factors include internal and external colour, shape, size, sweetness, vitality (the inherent capacity to maintain quality after harvest) (Hofman et al., 1997b), cleanliness and residue levels, and the occurrence of pest or disease infestations or biotic/abiotic damage. The main production factors infl uencing at-harvest quality apart from genotype include chemical treatment regimes and orchard hygiene, weather conditions before and at-harvest, irrigation, pruning systems and tree nutrition.
Maturity
Peacock (1986) considered that fruit maturity referred to its stage of ontog-eny, with fruit of different maturities being at different stages of ontogeny. Fully mature mango fruit are strictly those which have produced a fully developed seed and which have reached their full physiological potential in

G.I. Johnson and P.J. Hofman536
relation to size increase and dry matter accumulation within the constraints of the growth environment. When fruit size and dry matter concentration reach a plateau, climacteric fruit such as mango can undergo ripening, where colour, texture, fl avour and aroma may change (Watada et al., 1984). In these fruit, a sharp rise, followed by a decline in respiration also accompanies the transformation from not-ready-to eat (unripe), to edible (ripe), to senescent (overripe). Ripening signals the completion of seed ontogeny, and encour-ages dispersal of the seed by attracting vertebrate fructivores (Cipollini and Stiles, 1992). If fruit are not harvested, maturation and ripening occur on the tree. Ripe fruit fall to the ground, or are consumed by bats, primates, pha-langers, birds or humans, either on the tree or after detachment.
Softening and sweetening of fruit fl esh and colour changes can occur at any stage of ontogeny, even in pea-sized fruit (Oosthuyse, 1995). Fruit drop at any stage of development is preceded by these events, and the likelihood of their occurrence increases as fruit size and dry matter levels approach their maxima (Singh et al., 2004). Although the changes constitute some of the components of ripening, they can only be regarded as such if the fruit have attained physiological maturity, i.e. ‘the stage of development when a plant or plant part will continue ontogeny even if detached’ (Watada et al., 1984; Yashoda et al., 2006).
When fruit are removed from the tree several days before the onset of ripening, they are initially hard and green. The fruit progressively soften, change colour and develop aroma at a rate determined by cultivar, storage environment and at-harvest maturity. Ideally, fruit are picked, treated, packed and transported while hard-green, and arrive at retail markets at some pre-determined stage of colour development (usually more yellow or red, than green, and ‘sprung’, but still fi rm). The rate at which ripening will occur under particular storage conditions depends upon the stage of ontogeny at harvest. More mature fruit will ripen more rapidly than less mature fruit.
Accurately estimating when the fruit are ready for harvest is critical to consistently meet customer expectations. This is called horticultural matu-rity, and several criteria of horticultural maturity are possible. One is a legal minimum or buyer-specifi ed standard of maturity, which confi rms that the fruit would be acceptable for consumption or processing when ripe or ready to eat for green-eating types. Maturity estimation often relies on visual or calendar-based (days from fl owering) assessment or in some cases the appli-cation of a simple test (e.g. dry matter or fl esh colour assessment). In more exacting cases, more accurate estimates of horticultural maturity may be required to assess product suitability for more stringent or narrower quality specifi cations, as may be required for contract sales or sea-export consign-ments.
In both cases, easy to assess harvest indices relying on visual, chemo-sensory or fruit-age attributes are needed, and they must correlate with the commercially relevant fruit characteristics measured in prescribed tests. Pea-cock (1986), Harvey (1987) and Askar and Treptow (1993) reviewed methods for assessing fruit maturity. In mango, dry matter, fl esh colour, skin colour, fruit shape, Brix, specifi c gravity or days from fl owering or heat accumulation
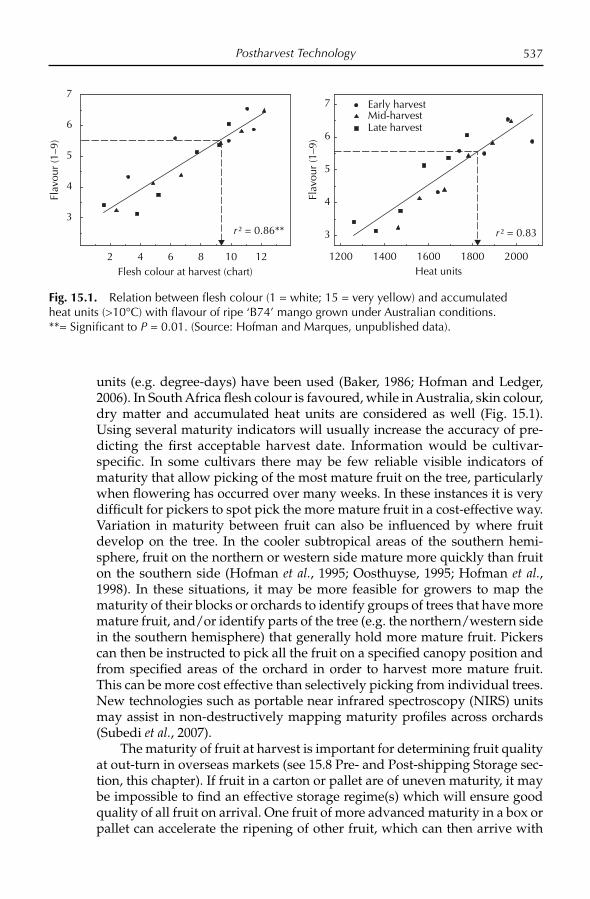
Postharvest Technology 537
units (e.g. degree-days) have been used (Baker, 1986; Hofman and Ledger, 2006). In South Africa fl esh colour is favoured, while in Australia, skin colour, dry matter and accumulated heat units are considered as well (Fig. 15.1). Using several maturity indicators will usually increase the accuracy of pre-dicting the fi rst acceptable harvest date. Information would be cultivar-specifi c. In some cultivars there may be few reliable visible indicators of maturity that allow picking of the most mature fruit on the tree, particularly when fl owering has occurred over many weeks. In these instances it is very diffi cult for pickers to spot pick the more mature fruit in a cost-effective way. Variation in maturity between fruit can also be infl uenced by where fruit develop on the tree. In the cooler subtropical areas of the southern hemi-sphere, fruit on the northern or western side mature more quickly than fruit on the southern side (Hofman et al., 1995; Oosthuyse, 1995; Hofman et al., 1998). In these situations, it may be more feasible for growers to map the maturity of their blocks or orchards to identify groups of trees that have more mature fruit, and/or identify parts of the tree (e.g. the northern/western side in the southern hemisphere) that generally hold more mature fruit. Pickers can then be instructed to pick all the fruit on a specifi ed canopy position and from specifi ed areas of the orchard in order to harvest more mature fruit. This can be more cost effective than selectively picking from individual trees. New technologies such as portable near infrared spectroscopy (NIRS) units may assist in non-destructively mapping maturity profi les across orchards (Subedi et al., 2007).
The maturity of fruit at harvest is important for determining fruit quality at out-turn in overseas markets (see 15.8 Pre- and Post-shipping Storage sec-tion, this chapter). If fruit in a carton or pallet are of uneven maturity, it may be impossible to fi nd an effective storage regime(s) which will ensure good quality of all fruit on arrival. One fruit of more advanced maturity in a box or pallet can accelerate the ripening of other fruit, which can then arrive with
Flesh colour at harvest (chart)2 4 6 8 10 12
Flav
our
(1–9
)
3
4
5
6
7
r ² = 0.86**
Heat units1200 1400 1600 1800 2000
Flav
our
(1–9
)
3
4
5
6
7 Early harvestMid-harvestLate harvest
r² = 0.83
Fig. 15.1. Relation between fl esh colour (1 = white; 15 = very yellow) and accumulated heat units (>10°C) with fl avour of ripe ‘B74’ mango grown under Australian conditions. **= Signifi cant to P = 0.01. (Source: Hofman and Marques, unpublished data).

G.I. Johnson and P.J. Hofman538
disease symptoms and with little or no shelf life. Variable maturity within treatment lots can also adversely affect product quality after heat disinfesta-tion (Jacobi et al., 1995).
On-farm record keeping and analysis of date of fl owering, seasonal prod-uct maturity, orchard management schedules, environmental data, transport regimes, market destinations and out-turn problems may enable some pre-diction of at-market quality and better selection of the appropriate destina-tion market. Recent research is focusing on non-destructive methods that could be used for checking fruit maturity in automated grading systems (Joyce et al., 1993; Subedi et al., 2007). Improvements in product quality and performance resulting from the effective use of such systems over several seasons, can provide considerable competitive advantages when developing buyer relationships or consumer ‘brand’/country-of-origin loyalty.
Adjusting maturation time
Flowering may be hastened or delayed by pruning, fl owering inducers or growth regulators (Davenport and Núñez-Elisea, 1997), to move the fruit-set period into a different time period, and bring forward or delay the harvest date to coincide with higher demand and market prices. Disadvantages could include higher at-harvest temperatures or greater risk of rainfall prior to harvest adversely affecting fruit quality.
Some cultivars and growing conditions are more favourable for manipu-lating fl owering date than others. In the Philippines, manipulation of ‘Cara-bao’ fl owering with potassium nitrate (KNO3) helps spread production and market supplies year-round (Bondad and Linsagan, 1979). The treatment also increases ‘Kent’ fl owering but does not alter timing (Goguey, 1993). By contrast, chemical manipulation of fl owering has not been effective on ‘Kens-ington Pride’ (Barba, 1974).
Evenness in fl owering within and between tree/block/farm lots can contribute to product uniformity and increased customer confi dence. Treat-ments that increase evenness of fl owering can have commercial benefi t in this regard also.
Skin colour and lenticel damage
Skin colour can affect sales, with markets preferring colour (green, yellow, orange, red blush) familiar to past purchase experience, known use and cul-tivar knowledge, or ethnic-group preferences. Fruit position on the tree af-fects red colour development, since sun exposure is important for anthocyanin development. Likewise, bagging of fruit can decrease red and green skin colour on ripe fruit (Hofman et al., 1997a). Nitrogen (N) can increase the pro-portion of the ripe fruit with green skin (Fig. 15.2) by retaining skin chloro-phyll (McKenzie, 1994; Nguyen, 2003; Nguyen et al., 2004; Bally, 2007). In cultivars susceptible to green skin at ripeness, N fertilization rates should be
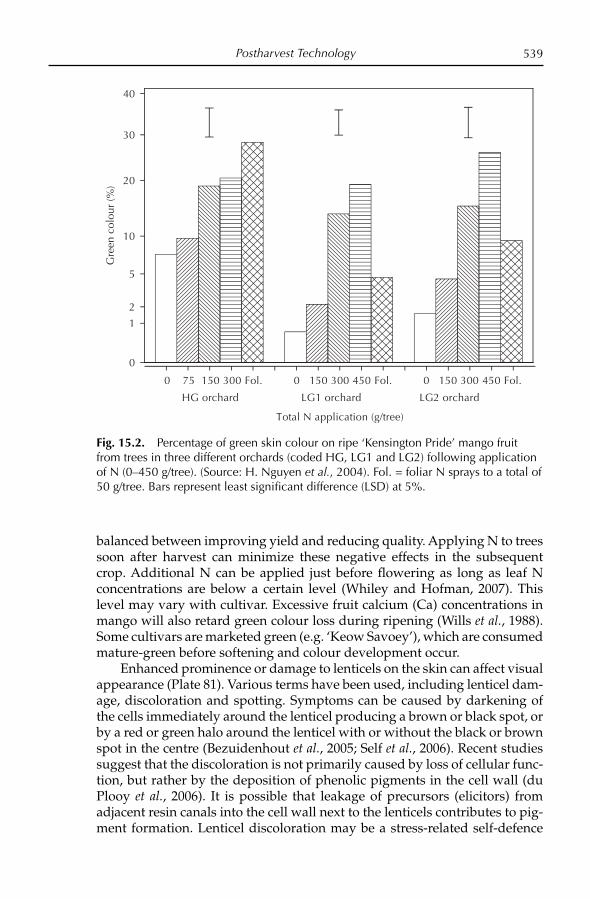
Postharvest Technology 539
balanced between improving yield and reducing quality. Applying N to trees soon after harvest can minimize these negative effects in the subsequent crop. Additional N can be applied just before fl owering as long as leaf N concentrations are below a certain level (Whiley and Hofman, 2007). This level may vary with cultivar. Excessive fruit calcium (Ca) concentrations in mango will also retard green colour loss during ripening (Wills et al., 1988). Some cultivars are marketed green (e.g. ‘Keow Savoey’), which are consumed mature-green before softening and colour development occur.
Enhanced prominence or damage to lenticels on the skin can affect visual appearance (Plate 81). Various terms have been used, including lenticel dam-age, discoloration and spotting. Symptoms can be caused by darkening of the cells immediately around the lenticel producing a brown or black spot, or by a red or green halo around the lenticel with or without the black or brown spot in the centre (Bezuidenhout et al., 2005; Self et al., 2006). Recent studies suggest that the discoloration is not primarily caused by loss of cellular func-tion, but rather by the deposition of phenolic pigments in the cell wall (du Plooy et al., 2006). It is possible that leakage of precursors (elicitors) from adjacent resin canals into the cell wall next to the lenticels contributes to pig-ment formation. Lenticel discoloration may be a stress-related self-defence
Total N application (g/tree)
0 75 150 300 Fol. 0 150 300 450 Fol. 0 150 300 450 Fol.
HG orchard LG1 orchard LG2 orchard
Gre
en c
olou
r (%
)
0
1
2
5
10
20
30
40
Fig. 15.2. Percentage of green skin colour on ripe ‘Kensington Pride’ mango fruit from trees in three different orchards (coded HG, LG1 and LG2) following application of N (0–450 g/tree). (Source: H. Nguyen et al., 2004). Fol. = foliar N sprays to a total of 50 g/tree. Bars represent least signifi cant difference (LSD) at 5%.
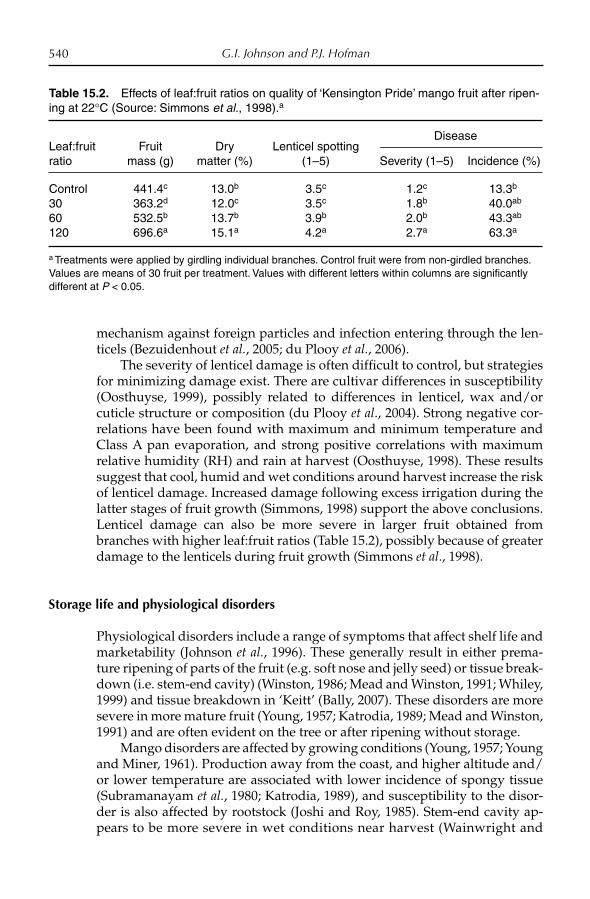
G.I. Johnson and P.J. Hofman540
mechanism against foreign particles and infection entering through the len-ticels (Bezuidenhout et al., 2005; du Plooy et al., 2006).
The severity of lenticel damage is often diffi cult to control, but strategies for minimizing damage exist. There are cultivar differences in susceptibility (Oosthuyse, 1999), possibly related to differences in lenticel, wax and/or cuticle structure or composition (du Plooy et al., 2004). Strong negative cor-relations have been found with maximum and minimum temperature and Class A pan evaporation, and strong positive correlations with maximum relative humidity (RH) and rain at harvest (Oosthuyse, 1998). These results suggest that cool, humid and wet conditions around harvest increase the risk of lenticel damage. Increased damage following excess irrigation during the latter stages of fruit growth (Simmons, 1998) support the above conclusions. Lenticel damage can also be more severe in larger fruit obtained from branches with higher leaf:fruit ratios (Table 15.2), possibly because of greater damage to the lenticels during fruit growth (Simmons et al., 1998).
Storage life and physiological disorders
Physiological disorders include a range of symptoms that affect shelf life and marketability (Johnson et al., 1996). These generally result in either prema-ture ripening of parts of the fruit (e.g. soft nose and jelly seed) or tissue break-down (i.e. stem-end cavity) (Winston, 1986; Mead and Winston, 1991; Whiley, 1999) and tissue breakdown in ‘Keitt’ (Bally, 2007). These disorders are more severe in more mature fruit (Young, 1957; Katrodia, 1989; Mead and Winston, 1991) and are often evident on the tree or after ripening without storage.
Mango disorders are affected by growing conditions (Young, 1957; Young and Miner, 1961). Production away from the coast, and higher altitude and/or lower temperature are associated with lower incidence of spongy tissue (Subramanayam et al., 1980; Katrodia, 1989), and susceptibility to the disor-der is also affected by rootstock (Joshi and Roy, 1985). Stem-end cavity ap-pears to be more severe in wet conditions near harvest (Wainwright and
Table 15.2. Effects of leaf:fruit ratios on quality of ‘Kensington Pride’ mango fruit after ripen-ing at 22°C (Source: Simmons et al., 1998).a
Leaf:fruit ratio
Fruit mass (g)
Dry matter (%)
Lenticel spotting (1–5)
Disease
Severity (1–5) Incidence (%)
Control 441.4c 13.0b 3.5c 1.2c 13.3b
30 363.2d 12.0c 3.5c 1.8b 40.0ab
60 532.5b 13.7b 3.9b 2.0b 43.3ab
120 696.6a 15.1a 4.2a 2.7a 63.3a
a Treatments were applied by girdling individual branches. Control fruit were from non-girdled branches. Values are means of 30 fruit per treatment. Values with different letters within columns are signifi cantly different at P < 0.05.

Postharvest Technology 541
Burbage, 1989; Mead and Winston, 1991). Incidence of spongy tissue has been reduced by mulches that decrease radiated and refl ected fi eld heat (Katrodia and Sheth, 1989). Severity of watery pulp breakdown in ‘Keitt’ is lower with higher crop loads from similar size trees (Bally, 2007), possibly because of smaller fruit size at high crop loads.
Several reports suggest links between low fruit Ca and mango disorders. High leaf Ca has been related to reduced soft nose (Young and Miner, 1961) and reduced stem-end cavity (Mead and Winston, 1991). Soil Ca applications can reduce stem-end cavity incidence (Whiley, 1999), but these responses are not always consistent. Applications of Ca to ‘Keitt’ from just before fl owering onwards did not increase leaf or fruit Ca during fruit growth or at commer-cial harvest (Bally, 2007). However, soil characteristics may also affect responses to soil Ca applications. In the sandy soils typical of Australian orchards, and even in the heavier clay subtropical soils with low cation exchange capacity, Ca can be rapidly removed from the top soil profi le, resulting in little long-term increase in soil solution Ca (Hofman and Mullen, 2005; Bally, 2007). Regular (two/week), small applications are required to consis-tently increase solution Ca (Hofman and Mullen, 2005). Other factors (e.g. vegetative vigour and water status) can infl uence fruit Ca uptake (Hofman and Smith, 1994).
Foliar Ca treatments have produced inconsistent effects, in some cases having little or no effect on fruit Ca concentrations or quality (Singh et al., 1987; Simmons et al., 1995), and in other instances reducing internal disor-ders (Chitarra et al., 2001). Postharvest dips have also had mixed results, with both positive (Wills et al., 1988; Singh et al., 2000) and nil or very small effects (Joyce et al., 2001) reported. As a result there is little commercial use of foliar or postharvest Ca treatments in mango. Future development of more labile Ca formulations may provide more consistent results.
Other nutrients have also been associated with fruit quality (Hofman and Smith, 1994). There are relatively few reports of potassium (K) and mag-nesium (Mg) effects on mango fruit quality. Higher Ca and Mg, and a ten-dency towards lower K in mango fruit, have been noted in fruit from orchards with no incidence of soft nose, compared with fruit from orchards with high incidence of the disorder (Burdon and Moore, 1991). Conversely, K levels are positively correlated with disease resistance; Karunanayake (2007) reported reductions in disease on mango fruit from trees receiving additional K. Application of triple the recommended level of K signifi cantly reduced stem end rot caused by Lasiodiplodia theobromae, while the severity of anthracnose was most reduced by application of the recommended level of K compared to nil and triple rate treatments.
Excess N can reduce storage life and quality, and excessive levels cause deterioration of avocado fruit quality (Wolstenholme, 2004) where its effect may be mediated through vegetative:reproductive balance and crop load (Hofman and Mullen, 2005). High N has been associated with increased dis-orders in mango (Young and Miner, 1961; Mead and Winston, 1991), possibly through the dilution effect of increased fruit size on Ca concentrations; how-ever, soil N applications later during fruit growth do not affect watery pulp

G.I. Johnson and P.J. Hofman542
breakdown in fruit ‘Keitt’ fruit (Bally, 2007). Heavy rain late during fruit development can release soil N previously unavailable to trees due to low soil moisture, potentially resulting in high fruit levels. Boron (B) defi ciency has been related to abnormal fruit development (Lahav and Whiley, 2002) and increased fruit storage disorders (Yogaratnam and Johnson, 1982; Smithet al., 1997). It may also be important in mango (Coetzer et al., 1991). The effect of larger fruit size and maturity on shelf and storage life (Seymour et al., 1990) can also be mediated through production factors infl uencing fruit set and leaf:fruit ratio. Fruit position in the canopy may also play a role here (see above).
Pests and diseases
Postharvest diseases and pests are reduced by various preharvest control measures including orchard hygiene, manipulation of fl owering, integrated management and the use of chemical and biological controls (Johnson et al., 1989a; Johnson, 1997; Fonseca et al., 2004b; Ploetz, 2004; Akem, 2006; Astridge and Baron, 2007a, b, c; Chin et al., 2007; Diedhiou et al., 2007). Prusky et al. (Chapter 7, this volume) and Ploetz and Freeman (see Chapter 8, this vol-ume) reviewed preharvest management of several postharvest diseases, while Dann et al. (2005, 2007) reviewed novel treatment options. Under the range of subtropical to tropical and dry to wet ecoclimates, combinations of treatments have been recommended to protect vegetative growth fl ushes, fl ower panicles and developing fruit from infections that lead to anthracnose, bacterial spot, powdery mildew, scab and stem-end-rot symptoms on fruit during development or after harvest (Poffl ey et al., 1999; Ledger, 2004; Sto-volt and Dirou, 2004; Akem, 2006).
Lonsdale (1993) found that monthly applications of copper oxychloride (CuCl2·3Cu(OH)2) in combination with mancozeb controlled most mango postharvest diseases. Copper (Cu) alone was less effective in controlling anthracnose. Copper sprays also provide protection against bacterial spot, while mancozeb can provide protection against scab (Poffl ey et al., 1999; Led-ger, 2004). Timmer and Zitko (1996) and Hardy et al. (2004) discussed the application of copper treatments to citrus for disease control. Formulations differ in their weather hardiness and indicated that retention of copper oxide (CuO) is superior to retention of copper chloride (CuCl2) or oxychloride, and application of Cu with the pH <7–6 can damage fruit and leaves. Lonsdale (1993) considered there was no disease control benefi t on mango by alternat-ing Cu with prochloraz sprays, but Ledger (2004) recommended their strate-gic application every 3–4 weeks in rotation with mancozeb and Cu when rainy conditions favoured anthracnose on developing fruit.
Azoxystrobin (Amistar®) and other strobilurin fungicides effectively control anthracnose, alternaria and powdery mildew on mango (Reuveniet al., 1998; Willingham et al., 1999; Reuveni, 2000; Sundravadana et al., 2006, 2007), Botryosphaera parva (Syn., Dothiorella dominicana) and Phomopsis sp. causing stem end rot (Everett et al., 2005), and Cercospora leaf spot (Ane-siadis et al., 2003) on other hosts. In Australia, no more than three strategic

Postharvest Technology 543
applications of azoxystrobin are recommended for fi eld control of anthra-cnose on mango in alternation with Cu, prochloraz and mancozeb schedules. Azoxystrobin should be appled as one or two applications at fl owering and/or early fruit set at no less than 14-day intervals, and again at 21 and 7 days before harvest (Stovolt and Dirou, 2004). Application of azoxystrobin to control anthracnose on mango leaves signifi cantly reduces subsequent fruit disease and boosts yield (Sunarharum, 2007).
Recent research has also focused on the potential of defence-boosting treatments, applied before or after harvest to reduce disease impacts on yield and shelf life (Terry and Joyce, 2004; Dann et al., 2007; Karunanayake, 2007). Anthracnose on mango fruit can be reduced by salicylic acid, its functional analogue benzothiadiazole (BTH) (= acibenzolar-S-methyl = Bion®), and ultraviolet (UV-C) irradiation with variable results, including phytotoxic effects (Zainuri et al., 2001; Zeng and Waibo, 2005; Zainuri, 2006; Zeng et al., 2006; Karunanayake, 2007).
Orchard hygiene, including reduction of inoculum by removal of old fruit and branches, and removing prunings from the orchard, can reduce anthracnose and stem end rots on fruit after harvest (Johnson, 1994; Ledger, 2004; Ploetz, 2004). Some fi eld diseases can disfi gure fruit. Following heavy rain close to harvest, bacterial spot can appear as discrete raised lesions, fol-lowed by fruit cracking or rupture and fruit drop. Black spot orchard man-agement practices (i.e. windbreaks, Cu sprays and use of resistant cultivars) can reduce the risk of damage on fruit (Johnson et al., 1996; Dodd et al., 1997; Ploetz and Prakash, 1997). In cool conditions, powdery mildew infection on young fruit can cause a ghosting symptom on mature fruit similar to that caused by powdery mildew on apples. Scab and sooty moulds can disfi gure fruit. Generally, spray programmes for anthracnose will control scab (Condé and Pitkethly, 2007), while sooty moulds are managed through integrated management of scale insects and by postharvest brushing (Johnson and Coates, 1993).
Tree nutrition can affect fruit disease incidence and severity. High Ca can reduce fruit diseases in many fruit crops (Hofman and Smith, 1994). Nitro-gen application before fl owering can increase mango fruit disease (H. Nguyenet al., 2004); applications up to 6 weeks after fl owering can increase anthra-cnose severity (Bally, 2007). Negative correlations occur between exocarp N percentage and antifungal resorcinol concentrations.
Poffl ey et al. (1999), Peña (2004) and Peña et al. (see Chapter 10, this vol-ume) discussed integrated pest management (IPM) for reducing pest dam-age and quarantine hazards associated with fruit. Preharvest control measures for fruit fl ies, seed weevils, scales and other skin defect-causing pests con-tribute signifi cantly to product quality improvement. IPM strategies can adequately control orchard pests while reducing reliance on pesticides (Cun-ningham, 1986, 1991a, b, c; Vijaysegaran, 1994; Peña, 2004; Peña et al., Chap-ter 10, this volume). Bagging of developing fruit can reduce or eliminate disease infection (Hofman et al., 1997a) and fruit fl y infestation (Kitagawa et al., 1992). However, for export market access, and regardless of effective fi eld control, postharvest disinfestation measures are often mandatory.

G.I. Johnson and P.J. Hofman544
Weather conditions
Rain before harvest, and high RH and temperatures can increase disease lev-els, fruit susceptibility to heat and brush damage, lenticel damage and reduce storage life (Dodd et al., 1991a; Estrada et al., 1993; Prusky et al., 1993a, b; Cooke and Johnson, 1994; Oosthuyse, 1998; Jacobi et al., 2001b). Disease risk predic-tion based on the monitoring of environmental variables to determine fungi-cide application frequency, can reduce pesticide residues (Fitzell and Peak, 1984; Fitzell et al., 1984; Peak et al., 1986; Dodd et al., 1991a, b, 1992; Prusky et al., 1993b). Irrigation frequency and water availability for tree growth can signifi -cantly impact postharvest diseases and disorders (Simmons, 1998).
15.4 Flavour and Aroma
Flavour is largely determined by sugars and volatiles in the ripe fruit, both of which increase in more mature fruit. The aroma produced by ripening and ripe mango can help attract customers, and provide some indication of fl a-vour development. In mango fruits, more than 280 different aroma volatile compounds have been reported (Singh et al., 2004). Variation in the constitu-ent aromatic compounds in mango cultivars results in aroma and fl avour diversity (MacLeod and Snyder, 1985; MacLeod et al., 1988; Torres et al., 2007). The high fruit levels of -terpinolene contribute to the characteristic fl avour of stronger-fl avoured cultivars such as ‘Kensington Pride’ (Bartley and Schwede, 1987; MacLeod et al., 1988). ‘Kensington Pride’ harvested at the green-sprung stage have higher concentrations of total aroma volatiles com-pared with fruit harvested at the hard-green or coloured stages (Lalel et al., 2003b). Most of the glycosidally bound aroma compounds increase in the pulp as the fruit matures, which contribute to improved fl avour. During the fi rst 7 days of ripening, -turpinolene is the major volatile compound, but in the later stages of ripening ethyl octanoate dominates (Lalel et al., 2003a).
15.5 Harvesting and Transport to the Packhouse
Harvesting of mango is determined according to attainment of acceptable/required maturity: (i) for arrival at market during the time of peak demand/highest price to maximize the chance of early sale; or (ii) to minimize loading wait at the shipping port. Fruit is generally picked into fi eld crates or bins, with or without the use of mechanical picking platforms.
Timing
Maturity is determined by assessing variables such as days from fl owering, accumulated heat units, fl esh dry matter percentage, fl esh colour or fruit shape/skin colour (see Maturity section under 15.3 Preharvest Management,

Postharvest Technology 545
this chapter). Fruit water potential fl uctuates diurnally, and can affect fruit quality. The water potential of fruit at harvest can affect susceptibility to han-dling, heat damage and product storage potential (Joyce and Patterson, 1994). In hot weather, fruit should be harvested in the coolest part of the day to reduce fruit overheating and energy requirements for postharvest cooling, and to minimize worker discomfort. Harvest during rain can reduce fruit quality (see Weather conditions section under 15.3 Preharvest Management, this chapter).
Sapburn
Severing the stem from the fruit causes relatively large volumes of latex to spurt or ooze from the cut stem. The sap is of low pH and high oil content and can burn the surface of the fruit (Bagshaw and Brown, 1989). The oil frac-tion contains terpinolene and resorcinols and is the fraction of the latex that causes the damage. Skin damage is particularly severe with ‘Kensington Pride’ (O’Hare, 1994), and less serious in Florida cultivars. In Pakistan, ‘Chausa’ is more susceptible than ‘Sindhri’ and ‘Dashehari’ (Maqbool et al., 2007). O’Hare (1994) observed that latex levels are lower and less phytotoxic in ‘Nam Doc Mai’, ‘Nang Klang Wun’, ‘Tong Dum’ and ‘Keow Savoey’ (0.16–0.48 ml/fruit), than in ‘Kensington Pride’ (1.67 ml/fruit). The oil compo-nent of the latex of Thai cultivars is much lower than that of ‘Kensington’ (O’Hare, 1994; Hassan, 2007). The concentrations and ratio of the two main resorcinols, 5-n-pentadecylresorcinol and 5-n-heptadecenylresorcinol, differ among cultivars (Hassan, 2007).
Factors affecting the potential of latex to cause sapburn are not well understood. It appears that both the terpene in the oil fraction of the sap, and adequate polyphenol oxidase (PPO)/peroxidise concentrations in the skin are required to develop sapburn; PPO and resorcinols in sap are less signifi -cant (Loveys et al., 1992; Robinson et al., 1993; John et al., 2002). Rain near harvest and high N in fruit result in more severe sapburn in ‘Kensington Pride’. However, negative relationships have been observed between exo-carp N concentration and alkyl resorcinols (Hassan, 2007). Sap from fruit harvested early in the day causes less sapburn than sap from fruit harvested later in the day, although early-harvested fruit exude more sap than late- harvested fruit (Maqbool et al., 2007).
Latex may provide protection against infestation by fruit fl y larvae (Joel, 1978, 1980) and may also contribute to disease tolerance. The 5-substituted resorcinols have antifungal properties (Cojocaru et al., 1985; Droby et al., 1986, 1987; Prusky and Plumbley, 1992). Karunanayake (2007) extracted a resorci-nol and a resorcinol derivative from the dichloromethane phase of Sri Lankan mango peel extracts that had antifungal properties. Differing relationships occur between resorcinol levels and relative susceptibilities of different mango cultivars to anthracnose (Karunanayake, 2007; Hassan et al., 2007). Strong positive relationships occur between resorcinol concentration in the peel and latex, and fruit resistance to artifi cially inoculated anthracnose.

G.I. Johnson and P.J. Hofman546
‘ Kensington Pride’ has more non-aqueous latex, higher concentrations of resor-cinols and greater tolerance of anthracnose than ‘Nam Doc Mai’ (Hassan, 2007). Less anthracnose occurs on fruit ripened with an intact stem, compared with de-sapped fruit. Hegnauer (1994) reviewed the phytochemistry of Mangifera.
Cultivars that are prone to sapburn can be harvested with 10–20 mm stems attached, and re-trimmed at the packhouse. Latex does not usually exude from longer stems because there is no continuity between the fruit and stem resin ducts (Joel, 1980), and the fruit lactifers are not severed; however, stems can break in transit to the packhouse, resulting in latex leakage and sapburn. Long stems left on the fruit to reduce sapburn has variable effects on disease depending on disease type and storage time. With shorter storage periods, anthracnose and stem-end-rot incidence and severity can be lower in fruit with stems attached due to higher levels of resorcinols (Hassan, 2007), but during longer storage stem-end-rot levels can increase due to the higher levels of inoculum associated with the retained stalk (Johnson et al., 1993).
Mango latex can cause skin disorders in humans (Keil et al., 1946; Oka et al., 2004). Bandyopadhyay et al. (1985) noted that resorcinol derivatives are allergens in the Anacardiaceae, and suggested that the 5-substituted resorci-nol in mango latex causes dermatitis. Both heptadec(adi)enyl resorcinols and pentadecylresorcinol can elicite an allergic reaction in sensitive patients (Okaet al., 2004). Harvesting and packhouse personnel must avoid contact with the latex of high-risk cultivars.
Harvesting and desapping
Rough handling at harvest can cause skin damage and internal fracturing or bruising. Using hooked sticks or shaking the tree to detach fruit causes skin damage and fl esh fracturing (Ledger, 1991a; Abu-Goukh and Mohamed, 2004) and sapburn. Mechanical damage during harvest also causes soft, darkened areas and bruises on fruit following hot water treatment. Mangoes should be handled as if they were eggs. Long-handled secateurs cut and grip the stem, allowing the fruit to be carefully lowered to the picking bin. Contact with soil and soilborne pathogens should be avoided (Johnson et al., 1993).
In cultivars where sapburn is a problem, latex should be drained from the fruit (desapping or bleeding) to minimize the incidence and severity of sapburn. Several systems have been assessed for reducing damage (Brown et al., 1986; Ledger, 1991b; Holmes et al., 1993; Lim and Kuppelweiser, 1993; O’Hare and Prasad, 1993; O’Hare, 1994; Shorter and Joyce, 1994). In Austra-lia, the main commercial practices (Plate 82) are:
Desapping in the fi eld with harvest aids using detergent. The basic de- ●
sign characteristics include detergent spraying onto a tarpaulin, a trough with the same detergent and a fi nal spray before fruit are placed in a fi eld bin. The fruit are either hooked from the tree in the direction of the harvest aid and onto the catching surface or the fruit are snapped directly off the tree and placed onto the tarpaulin. The fruit roll from the tarpaulin into

Postharvest Technology 547
the trough containing detergent and then into 300–400 kg fi eld bins. Alkaline detergents that deactivate damaging sap components are most effective; high concentrations of surfactant in the detergent are not re-quired. The crucial factors are that fruit should be exposed to detergent for at least 90 s, the detergent is either not recycled, or replaced before sap accumulation in the detergent causes other damage such as skin browning (Bally et al., 1997; O’Hare et al., 1999).Another design includes a motorized hydraulic ladder (cherry picker) ●
with the fruit desapped for 1–2 s before placing in a basket containing a spray of alkaline detergent. This system is particularly effective for tall trees, but care must be taken that fruit are covered by the detergent for at least 90 s. Picking fruit with long stems into small 18 kg crates and desapping in ●
the shed. The fruit are dipped into detergent before desapping and plac-ing on a long conveyor system that holds the fruit inverted and provides detergent/water sprays for a few minutes. The fruit are inverted for c.20 min before drying and packing.
In these systems, the detergent is not strongly alkaline, but the surfactant should be of suffi cient concentration to provide a protective coating around the fruit before desapping. Stem breakage must be minimized in the crates, as this can cause sapburn and quality loss (Holmes et al., 1993; Holmes and Bally, 1994). Latex must not spray or drip onto fruit being desapped. Workers who are sensitive to mango sap should wear hand protectants, aprons and footwear to minimize skin contact. The detergent must reduce sapburn and skin browning without causing other damage (i.e. lenticel spotting) (Fig. 15.3). Desapping in the fi eld by inverting fruit directly onto racks without detergents has been used for sensitive cultivars and when particular growing conditions have increased fruit susceptibility to lenticel spotting or other damage from detergents; however, labour costs are becoming prohibitive, requiring compromises between cost and quality. Desapping by inverting and placing on the ground signifi cantly increases the incidence and earlier appearance of stem end rot caused by soilborne L. theobromae, and is not recommended (Johnson et al., 1993).
Harvest aids have reduced in-shed desapping. Holmes et al. (1993) found 9–16% of fruit in fi eld crates were affected by sapburn when harvest aids were not used. Harvest aids provide the greatest reduction in total sapburn (from 69% to 15–18%). While harvest aids can signifi cantly reduce sapburn, inappropriate use can increase some forms of skin browning (Bally et al., 1997). Underhill and Dahler (1995) described four types of skin browning which produce symptoms distinct from the sapburn caused by the oil phase of latex. Several forms of skin browning involve tissue reactions with sap/detergent mixtures. Symptoms vary if latex enters fruit through micro-cracks in the cuticle or the lenticels (O’Hare et al., 1999). Holmes (2003) developed guidelines for the use of harvest aids.
Cost savings associated with the use of harvest aids can be lost if fruit-to-fruit or fruit-to-ground impact is not minimized during harvest. Rough

G.I. Johnson and P.J. Hofman548
harvesting can increase the incidence of bruising and internal fracturing, and lower wholesale returns. Thorough training of picking crews and supervi-sion of their performance is required to maintain good practice.
Transport to the packhouse
Harvested fruit should be transported to the packhouse as soon as possible, with no prolonged exposure to the sun. Rough handling and transport must be minimized. Roads/tracks from orchard to packhouse should be smooth, with transport vehicle tyres correctly infl ated, and special suspensions to reduce vibration and damage.
15.6 Packhouse Measures
Harvested fruit are transported to a central packhouse which provides shel-ter from rain and sun, and facilities for cleaning, treating, packing, cooling and storing fruit until consignment to market (Schoorl and Holt, 1982, 1985). Mechanized packhouse systems can offer labour savings and increased re-turns (Murray and George, 1994). When manual handling was reduced from fi ve to two steps, fruit appearance improved, disease losses were lowered, sizing accuracy improved, packing rate increased and space, labour and supervisory requirements were reduced (Murray and George, 1994).
A typical packhouse sequence is shown in Fig. 15.4. In packhouses that include a disinfestation facility, the sequence must be modifi ed to allow for
Detergent
Contro
lLO
C
Mango
Clea
r
Mango
Glo
Mango
Mag
ic
Mango
Plus
Mango
Was
h
Superc
once
ntrate
Lent
icel
spo
tting
(7da
ys a
fter h
arve
st)
1
2
3
Block 1Block 2Block 3
Fig. 15.3. Effect of detergents on lenticel spotting on skin of ‘B74’ mango (0 = no lenticel spotting; 4 = severe spotting). Fruit were obtained from three different blocks in two orchards, dipped in detergent for 2 min, then held at 20°C for 7 days (Source: Hofman and Ledger, 2006). The bar represents the least signifi cant difference (LSD) at 5%.

Postharvest Technology 549
1. Deliver 18. Air or sea freight
2. Weigh 17. Cool chain transport
3. Cold wash and brush 16. Palletizing
4. Hot water/fungicide bath 15. Outer boxing
5. Fungicide application 14. Cool/ethylene gas
6. Dry 13. Pack
7. Wax and brush (optional) 12. Disinfestation
8. Dry 11. Crates
9. Grade 10. Size
Processinggrade
Secondgrade
First grade(domestic/export)
Extra class(export)
Processing Local marketing
Fig. 15.4. Packhouse and marketing activities for mango. Waxes are not applied in some countries because of abnormal ripening and off-fl avour development. When disinfestation is not required, steps 11, 12 and 15 would be omitted. When fruit are heat disinfested, they may be packed into an inner box prior to cooling. For some markets accessible by air, the fruit may be treated with ethylene and stored until near ripe. The boxed fruit may then be packed into an outer carton prior to palletizing. Pre-ripening allows fruit to be rechecked prior to despatch, with fruit of unsatisfactory appearance (e.g. skin damage) redirected to domestic market or processing.

G.I. Johnson and P.J. Hofman550
sizing before disinfestation, with tray packing after treatment. Packhouse design and installation consultants can provide substantial savings by elimi-nating bottlenecks and minimizing product damage points.
Delivery inspection and traceability
Harvested fruit in fi eld crates should be treated, packed and cooled as soon as possible. Quality and contaminant management systems may require that a record system tracks the block or trees from which each bin of fruit is har-vested. This enables records to be kept of tree or block yield, quality perfor-mance and defect levels as well as labour performance rates and pesticide residues. Individual fruit in a tray can be traced back to the tree or block from which it was harvested. Block/tree-to-tray traceability systems and pack-out records allow problems (i.e. excessive or unapproved pesticide detections) to be traced to the site of the problem and relevant action taken. They can also be used to motivate producers or picking teams to deliver high quality produce.
At the packhouse, samples of fruit should be evaluated immediately for maturity, blemishes and disease and pest incidence and recorded in an appro-priately designed computerized system. Preharvest orchard inspections can reveal the defects that can be anticipated in the packhouse. Some degree of in-fi eld sorting can occur at the point of harvest, and soft or damaged fruit collected separately and discarded.
Desapping and washing
Unloading should avoid dropping, damaging and wounding of fruit. Fruit are normally unloaded from fi eld bins into bin dumps if desapping is unnec-essary or removed manually from the crates for desapping (see Harvesting and desapping section under 15.5 Harvesting and Transport to Packhouse, this chapter). Detergents and sanitizers are sometimes added to washing water. Their use requires careful consideration. Some may cause fruit dam-age, or promote early fruit disease expression (Korsten et al., 1993). Chlorine is added and carefully regulated to wash and/or rinse water in some pack-houses, but this is not essential. Quaternary ammonium disinfectants should not be added to wash water as their direct application to foodstuffs is gener-ally not permitted.
Disease control
Hot water and fungicide applicationHot water dips, or sprays over brushes, with or without fungicide, and fun-gicide sprays or dips, can eradicate quiescent fungal infections that have been established on and beneath the cuticle and within the pedicel prior to

Postharvest Technology 551
harvest (Johnson et al., 1989a, b, 1991, 1992; Kernot et al., 1999; Poffl ey et al., 1999; Plan et al., 2002). Suslow (2000) provides generic recommendations for the postharvest handling of produce. Postharvest disease treatment effi cacy varies with infection level, cultivar, ripening status and storage regime. Hot water treatment also cleans fruit, but can contribute to increased skin dam-age (Cooke and Johnson, 1994). Anthracnose caused by Colletotrichum gloeo-sporioides Penz. and Colletotrichum acutatum Simmonds, is controlled more readily than stem end rots (or soft brown rot) caused by anamorphs of Botry-osphaeria spp. (Fusicossum spp., Neofusicoccum spp., L. theobromae (Pat.) (Griff. and Maubl.)) (Johnson, 1994; Slippers et al., 2005; Crous et al., 2006) and Pho-mopsis mangiferae Ahmad, and alternaria rot caused by Alternaria alternata (Fr.) Keissler. The latter is generally only a problem in fruit from dry regions or in fruit from more humid areas during storage for 3 weeks or more (Prusky et al., 1980, 1993a, b, and Chapter 7, this volume; Johnson et al., 1990b).
Fruit are moved through a water bath for 5 min at 48–50°C for less mature fruit and hot-water-damage-susceptible cultivars (e.g. ‘Zill’ and ‘Irwin’) and at 50–55°C for mature fruit and less susceptible cultivars (Anonymous, 1994b). Treatment for 3 min may be adequate for control of anthracnose, while immersion for up to 7 min may enhance control of stem end rot (Muirhead and Grattidge, 1986; Sepiah, 1986; Johnson et al., 1989b). In large-scale facili-ties, dip tanks may range from 3000 to 5000 l, with fruit immersed and moved through the tank by a series of paddles. In tank construction, non-corrodible materials such as stainless steel and fi breglass are preferred, and the con-veyor system that contains the paddles should travel along the bottom of the tank to reduce damage to fruit that sink. Accuracy in temperature control, effi cacy of the heating unit and timing of fruit fl ow through the bath are critical. Temperature probe placement at pump inlet and outlet and thorough water circulation to ensure accurate temperature reading and to minimize hot spots are critical. Impurities (e.g. minerals, sediment and debris) in dip water can affect fungicide performance and stain or damage the fruit. In-line fi lters in the inlet and pump circulation systems should be installed and cleaned regularly.
Where acceptable, carbendazim can be added to the hot water at the rec-ommended rate, and topped up and replaced regularly, to provide improved control of stem end rot and anthracnose at lower temperatures (52°C). Beno-myl has been withdrawn for postharvest use, but much of the benomyl use information (from earlier research) is relevant for carbendazim (Johnson et al., 1997). Also, hot thiabendazole (TBZ) is generally as effective as hot benomyl for controlling stem end rot, but may provide inferior control of anthracnose (Coates et al., 1993). The active component of benomyl and TBZ in plants, carbendazim (MBC), is identical (Erwin, 1973; Muirhead, 1976); however, TBZ also contains sulfur (S) which affects its rate of breakdown and spec-trum of activity (D. Guest, personal communication, Melbourne, 1995). Beno-myl penetrates plant tissue more effectively than TBZ, carbendazim or thiophanate methyl (Eckert, 1983).
Dipping fruit in hot, dirty, latex-contaminated water can increase phyto-toxicity and lenticel damage. Hot fungicide dips lose effi cacy due to sap

G.I. Johnson and P.J. Hofman552
build-up in the dip tank and stripping out of fungicide (Wells and Littlemore, 1989). Ledger (2004) optimized dip:fruit ratio and dipping practices. Prochlo-raz provides good control of anthracnose and alternaria rot, but does not pro-vide control for stem end rot (Johnson et al., 1990b; Johnson and Coates, 1993). In South Africa, for local markets prochloraz is added at 405 ppm of active ingredient (ai) of a 45% emulsifi able concentrate (ec) formulation; for export markets, 810 ppm prochloraz is used. Fruit are immersed for 20 s. In Australia, prochloraz at 250 ppm is applied by overhead spray, and fruit require 15–20 s to pass through the prochloraz spray race on a roller conveyer system.
A maximum residue level (MRL) of 7.0 mg/kg for prochloraz for assorted tropical and subtropical fruits with an inedible peel is recommended (CODEX-MRL, 2008). This group MRL replaces individual fruit commodity MRLs, and takes into account the lower residues in the fl esh compared to the skin (Muller and Burt, 1989). Some registered use-rates for postharvest applica-tion of prochloraz are listed in Table 15.3.
Hot water sprays over brushes (55°C for 15–20 s) is an effective alterna-tive to hot water dips containing prochloraz for controlling alternaria rot (Prusky et al., 1999, 2006). Application of hot water spraying and brushing for 15–20 s (HWB) followed by a spray of 50 mM hydrochloric acid (HCl), alone or in combination with prochloraz, also improved control of alternaria rot (Prusky et al., 2006). These treatments have not been tested for anthracnose. Using 2,4-dichlorophenoxyacetic acid (2,4-D) diluted in wax after HWB and prochloraz reduces stem end rot (Kobiler et al., 2001). A hot water and beno-myl combination treatment followed by a prochloraz spray provides effec-tive control of anthracnose, stem end rot and alternaria rot during longer storage (Johnson et al., 1989a, 1990b). Similar benefi ts are now attributed to hot TBZ dip and cold prochloraz spray (Ledger, 2004).
When fungicides are used in the packhouse, spent dip suspensions and fungicide containers must be disposed using approved methods, often included with supplier recommendations. Carbendazim suspensions can be drained into a trench fi lled with stones, but runoff must be avoided. Car-bendazim and other benzimidazole fungicides are toxic to earthworms (Wright and Stringer, 1973).
Table 15.3. Registered uses of prochloraz for postharvest treatment of mango (Source: adapted from Lunn, 2004).
Country Forma Method Rate (kg ai/100 l)
Australia ec 30 s spray 0.025Brazil ec 2 min dip 0.05China ec/wp 1 min dip 0.05–0.1Colombia ec Not specifi ed 0.025Peru ec Not specifi ed 0.02–0.045South Africa ec 20 s dip 0.08
aai, active ingredient; ec, emulsifi able concentrate; wp, wettable powder.

Postharvest Technology 553
HeatPest disinfestation treatments involving heat provide some control of anthra-cnose, but do not adequately control mango fruit pathogens for export. Tem-perature and time combinations suitable for non-deleterious fruit disinfestation are sublethal to a signifi cant percentage of quiescent infections beneath the fruit cuticle and pedicel tissues (Coates and Johnson, 1993). In many regions, fruit skin temperatures frequently approach the mid-40°C range during pre-harvest development, a natural selection pressure favouring heat-tolerant fungal infection structures. For ‘Kensington Pride’, hot benomyl in combina-tion with either prochloraz or vapour heat at 46.5°C for 20 min controls stem end rot more effectively than hot benomyl alone and TBZ, alone or in combi-nation with vapour heat, during storage at 23°C for 15 days (Coates et al.,1993). Disease control in combination with heat disinfestation has been reviewed by Coates and Johnson (1993) and Jacobi et al. (1994, 2001a).
Future optionsHeat is an ideal disease control treatment, since it is environmentally safe and non-chemical. Its effectiveness would be enhanced if fruit tolerance could be increased by genetic manipulation or the development of pre-conditioning treatments. Pre-treatments to render quiescent structures of pathogens more susceptible to heat would also improve disease control. Measures to increase effi cacy could include other energy sources, chemicals, adjuvants, fumigants or microorganisms to damage or soften fungal wall structures.
Treatments to delay fruit ripening also limit or reduce disease losses. Storage quality would benefi t from the development of cultivars or pre-conditioning treatments to improve tolerance of fruit for cool storage or controlled atmo-sphere (CA) and modifi ed atmosphere (MA) storage (Brecht and Yahia, Chapter 14, this volume). With increasing concerns about the use of chemi-cals on food (Gullino and Kuijpers, 1994), and in view of current limitations on heat treatment and storage regime disease control effi cacy, non-deleteri-ous alternatives to synthetic fungicides are required. Alternatives to fungi-cides for controlling postharvest diseases have been reviewed by Johnson and Sangchote (1994) and Korsten (2006). Options include: (i) biological con-trol, i.e. the use of microorganisms to control pathogens (Wilson and Pusey, 1985; Jeffries and Koomen, 1992; Korsten et al., 1993, 1994; Korsten 2006); (ii) enhanced exploitation of naturally occurring antifungal compounds in fruit (Prusky et al., 1982; Johnson et al., 1998; Joyce et al., 1999; Zainuri et al., 2003; Hassan et al., 2007); (iii) application of fruit coatings such as chitosan with both MA and antifungal effects (El Ghaouth et al., 1992a, b; Wilson et al., 1994; El Ghaouth and Wilson, 1995); (iv) exposure to UV-C light (wavelength <280 nm) (Chalutz et al., 1992; Wilson et al., 1994). Zainuri (2006) reported some promise in the use of UV-C radiation for control of anthracnose, but fruit damage risks and treatment dose accuracy were critical; (v) containment of fruit in atmospheres containing high levels of carbon dioxide (CO2) for 24–48 h after harvest (fl ushing) (Prusky et al., 1992, 1993c); (vi) regulation of fruit ripening (Brady, 1994); and (vii) application of naturally occurring plant products (Fallik and Grinberg, 1992; Wagner and Flores, 1994). Many of these

G.I. Johnson and P.J. Hofman554
options may delay disease development by eliciting increases in antifungal compounds in the fruit (Prusky and Keen, 1993; Wilson et al., 1994; Zainuri, 2006).
What are the alternatives to synthetic fungicides for controlling mango dis-eases? Bacteria active against mango isolates of C. gloeosporioides, stem end and soft brown rot pathogens have been evaluated (Koomen et al., 1990; Korsten et al., 1991, 1992, 1993; Jeffries and Koomen, 1992; Coates et al., 1995; Korsten, 2006). Antifungal resorcinols in the peel of mango fruit interfere with the devel-opment of anthracnose and alternaria rot (Cojocaru et al., 1985; Droby et al., 1986, 1987; Prusky and Keen, 1993; Zainuri, 2006; Hassan, 2007), with higher levels present in some cultivars (Hassan, 2007; Karunanayake, 2007; Hassan et al., 2007). Lonsdale (1992, 1993) found that enclosure of ‘Keitt’ in high-density polyethyl-ene bags with 30% CO2 and 15% oxygen (O2) for 24 h at 11°C prior to storage, improved control of anthracnose. However, 24 h exposure to 20% CO2 signifi -cantly increased the incidence of soft brown rot (stem end rot) in ‘Keitt’ and ‘Kent’ compared to untreated fruit, especially in the absence of O2. UV irradia-tion of fruit for 10–30 s in combination with wax prior to storage is similar to hot water in reducing the incidence of soft brown rot compared to untreated fruit.
Brushing
Brushing on mango packing lines can occur after, or at the same time as, hot water and fungicide treatments. Hot water treatment washes sap away, and loosens superfi cial debris, scale insect carapaces and sooty mould, which are removed as the fruit pass over rotating brushes. Brushing also removes superfi cial deposits of fungicides that accumulate on fruit from orchard application of Cu fungicides (Lonsdale, 1993) and incorrect mixing or sedi-mentation of benzimidazole fungicides resulting from sap accumulation in dip tanks (Wells and Littlemore, 1989). Soft, non-damaging brushes should be used, washed every day and replaced seasonally.
For ‘Kensington Pride’ mangoes harvested after rain, skin marking, fruit shrivel and weight loss increase signifi cantly on fruit treated with a hot water and fungicide dip or a hot water and fungicide dip followed by treatment with prochloraz, when fruit brushing followed either or both treatments relative to untreated and untreated/brushed fruit. Prochloraz before brushing resulted in fruit quality similar to untreated or brushing only (Cooke and Johnson, 1994). Brushing can increase lenticel spotting (Oosthuyse, 1999). Therefore, the num-ber and type of brushes must remove foreign matter and polish the fruit, while not increasing risk of brush and lenticel damage, especially during wet weather and with heat treatments (see Weather conditions and Skin colour and lenticel damage sections, both under 15.3 Preharvest Management, this chapter).
Grading and sizing
The purposes of grading are to sort fruit into defi ned categories of unifor-mity and to divert out-of-grade fruit from the pack line to either a second

Postharvest Technology 555
grade, processing or reject line. Mangoes with defects outside acceptable lim-its as defi ned in a grade schedule or chart are manually removed and trans-ferred (by conveyer belt) to seconds or processing lines as appropriate. The purpose of sizing is to categorize fruit into size or weight groups for packing. Fruit must be sized prior to disinfestation with hot water or vapour heat to ensure consistent treatment responses. Typical systems include automatic graders that separate fruit by weight into groupings that correspond to pre-determined categories (Schoorl and Holt, 1982, 1985). Camera vision systems can separate for colour, defects and shape. Fruit usually accumulate in sepa-rate bins for packing into cartons or into bulk containers for processing or disinfestation. The fruit are packed manually into single-layer trays, with plastic or cardboard liners that have depressions designed to accommodate fruit of a particular size. The depressions provide some support for individ-ual fruit during packing, while the cardboard liners also provide some additional buffering against impact damage. The pattern of the depressions facilitates most effi cient utilization of carton space. Mango tray liners com-monly accommodate 12–25 fruit for 6.5 kg trays. Some tray liners may be inappropriate for sea export due to interference with vertical airfl ow.
Organic materials (i.e. paper, leaves or shredded wood) have been used to cushion individual fruit in cartons. These materials can harbour pathogens, for example Rhizopus stolonifer (Ehrenb. Fr. Lind.), which causes transit rot of mangoes and has been detected in shredded wood used in mango packaging. Shredded wood creates micro-wounds in the fruit skin, providing points of entry for hyphae growing on the wood. Losses are more severe when fruit have been removed from cold storage, allowing condensation to develop on the fruit and shredded wood ( Muirhead and Grattidge, 1986).
Grade standards
The International Standardisation Organisation (ISO) is a non-governmental organization (NGO), and is a network of national standards institutes (157 countries). ISO is the global leader for development and publication of stan-dards. ISO publishes a range of standards for fruit and vegetables, testing, crop and postharvest management procedures and food safety, system auditing and nutrient and water testing that are relevant to mango systems’ benchmarking and improvement. A range of standards may also be defi ned within GAP certifi cation protocols and nationally developed marketing arrangements. Agreed grade standards provide a reference point for produc-ers and traders in production and marketing (EurepGAP, 2007; ISO, 2008). The CODEX Alimentarius Commission also oversees the development of standards for fresh and processed fruit with the CODEX Committee on Fresh Fruit and Vegetables (CODEX, 2008a). CODEX standards for fresh fruit spec-ify provisions for quality, sizing, tolerances, presentation, marking and label-ling, and contaminants (CODEX, 2008a). There are CODEX standards for fresh mangoes, canned mangoes and mango chutney (CODEX, 2008b).

G.I. Johnson and P.J. Hofman556
Minimum requirements for grade standards specify that fruit intended for international trade should be intact, fi rm, fresh in appearance, sound, clean, free from black stains and bruising, free from damage caused by low temperatures and free from pests and pest damage. Fruit should be carefully picked at the stage of physiological development which will allow transport and handling and continuation of the ripening process so that fruit will ripen to consumer expectations. Class standards can depend on customer specifi -cations, and can be based on fruit size and appearance. Colour illustrations in Anonymous (1993) and Amesbury et al. (2002) are indicative of some of the quality standards that can be specifi ed for appearance, shape and colour, and tolerance levels for superfi cial skin defects. Similar charts are often available for individual cultivars and are produced during the development of QA systems for specifi c marketing groups and customers.
Packing-line QA inspectionsPacking-line control inspections are used to monitor grading effi cacy and packing-line damage. Packing-line inspection samples are taken soon after fruit pass points in the line at which defects are most likely to be overlooked and/or induced. For start and end-of-line pack-out checks, and out-turn inspections, randomly selected cartons are unpacked, and all fruit are checked for compliance with preset quality parameters. Most value is gained from quality control checks if records are kept and evaluated, with feedback/trou-ble shooting as necessary, to constantly improve the system (Ledger and Bag-shaw, 1994; Ledger and Premier, 2006). Computer analysis of such information provides a seasonal benchmarking record of QA improvement, and high-lights areas for attention in packing-line improvement and personnel train-ing. Record keeping is mandatory under GAP certifi cation systems.
Future optionsGreater automation of grading and packing will become necessary as pro-duction and labour costs increase, and as customers become more demanding (Hilton, 1994). Recent advances in computing have made possible high-speed sorting using visual systems for colour, shape and externally visible defects. Also, NIRS systems can now be used in-line to sort for fl esh characteristics that infl uence fl avour. In mango, percentage dry matter and fl esh colour are related to ripe fruit fl avour, and can be estimated using NIRS (Saranwonget al., 2004; Subedi et al., 2007). Estimation is suffi ciently accurate to allow acceptable separation into several categories for fi nal fl avour. NIRS may also be useful for predicting ripening behaviour and weight loss during ripening (Mahayothee et al., 2004). Given the infl uence of weight loss in chilling injury (CI) development during cold storage (Bower et al., 2003), NIRS may also be able to estimate potential for CI during cold storage.
There is interest in other non-destructive quality assessment for the pack-house. Joyce et al. (1993) noted that future innovation could lead to proton magnetic resonance imaging (MRI) technology suitable for packing-line applications to allow non-destructive detection of internal disorders and pest infestations. X-ray imaging may have potential for detecting seed weevil

Postharvest Technology 557
damage in mangoes (Thomas et al., 1995; Reyes et al., 2000), but recent inves-tigations suggest that neither X-ray imaging nor MRI is suffi ciently reliable for quarantine purposes, particularly where larvae are small (R.A. Jordan, personal communication, 2007). New methods of nuclear magnetic reso-nance (NMR) (Marigheto et al., 2008) may distinguish internal disorders such as jelly seed. Acoustic/ultrasonic methods can sort for fruit fi rmness (Miz-rach, 2008) and may help identify softening fruit with internal disorders and reduced storage life. Robotics in sorting and packing will be used increas-ingly where labour costs and availability are high.
Disinfestation
Disinfestation treatments, backed by integrated fi eld control programmes and/or area freedom stipulations under market access approvals, provide assurance to authorities of an importing country that the commodity will be free of target pests and not pose a quarantine threat (Johnson and Heather, 1995; Follett and Nevin, 2005). Market access application and approval ar-rangements for most countries operate under national legislation and regula-tions formulated under the framework of the IPPC and the World Trade Organisation (WTO) Agreement on the Application of Sanitary and Phyto-sanitary Measures (IPPC, 2008; SPS, 2008). Key aspects of quarantine regula-tion of crop pests are covered by International Standards for Phytosanitary Measures (ISPM), developed and agreed in consultation with signatory cou-ntries to the IPPC under the Food and Agriculture Organization (FAO) (ISPM, 2007a). ISPM (2007b) cover guidelines on the phytosanitary measures for regulated pests. Under the IPPC, a technical panel on phytosanitary mea-sures has oversight of international guidance on phytosanitary treatments including assessment and recommendations for use as international stan-dards. In addition to ISPM, a range of regional plant protection measures has been established. It has been agreed internationally (ISPM, 2007b) that phytosanitary treatments should fulfi l the following requirements:
Provide effective destruction, inactivation or removal of pests or render ●
them infertile or devitalized. Normally, stipulation of the destruction effi ca-cy level, with quantifi cation or statistical benchmarking, is required. When experimental data are unavailable or inadequate, other evidence is needed to support a claim of effi cacy, for example historical/practical experience.The technology and treatment regime used should be: (i) clearly delin- ●
eated, with evidence to confi rm adequate adherence to scientifi c meth-ods in generating data (experimental designs). Data in support of the treat-ment effi cacy should be verifi able, replicable and statistically valid, and preferably published in a peer-reviewed journal; (ii) appropriate for use in international or regional trade and research; and (iii) safe to apply, with no undesirable impacts on treated commodities or the environment.
Approvals for disinfestation treatments and market access are obtained on a country-by-country basis. A starting point for intending exporters is to
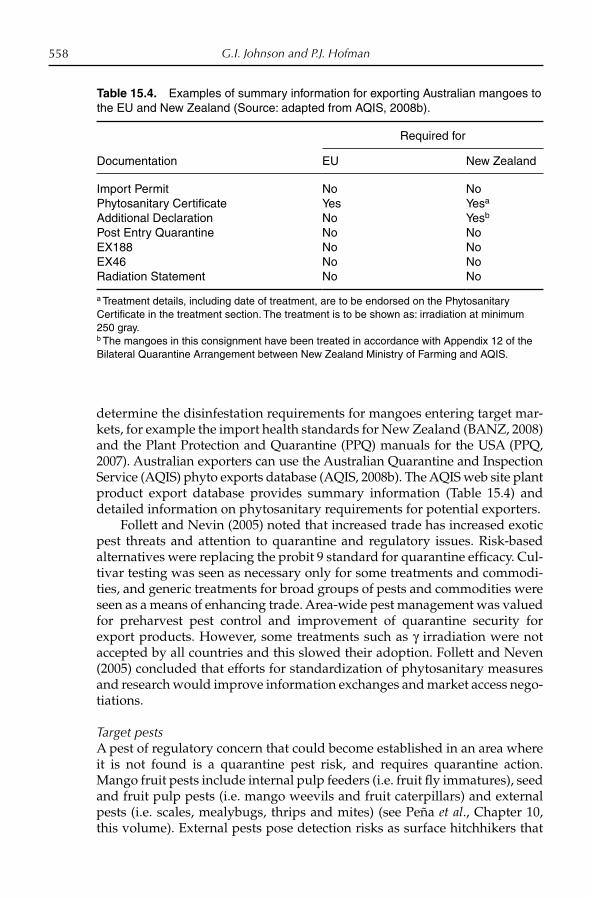
G.I. Johnson and P.J. Hofman558
determine the disinfestation requirements for mangoes entering target mar-kets, for example the import health standards for New Zealand (BANZ, 2008) and the Plant Protection and Quarantine (PPQ) manuals for the USA (PPQ, 2007). Australian exporters can use the Australian Quarantine and Inspection Service (AQIS) phyto exports database (AQIS, 2008b). The AQIS web site plant product export database provides summary information (Table 15.4) and detailed information on phytosanitary requirements for potential exporters.
Follett and Nevin (2005) noted that increased trade has increased exotic pest threats and attention to quarantine and regulatory issues. Risk-based alternatives were replacing the probit 9 standard for quarantine effi cacy. Cul-tivar testing was seen as necessary only for some treatments and commodi-ties, and generic treatments for broad groups of pests and commodities were seen as a means of enhancing trade. Area-wide pest management was valued for preharvest pest control and improvement of quarantine security for export products. However, some treatments such as γ irradiation were not accepted by all countries and this slowed their adoption. Follett and Neven (2005) concluded that efforts for standardization of phytosanitary measures and research would improve information exchanges and market access nego-tiations.
Target pestsA pest of regulatory concern that could become established in an area where it is not found is a quarantine pest risk, and requires quarantine action. Mango fruit pests include internal pulp feeders (i.e. fruit fl y immatures), seed and fruit pulp pests (i.e. mango weevils and fruit caterpillars) and external pests (i.e. scales, mealybugs, thrips and mites) (see Peña et al., Chapter 10, this volume). External pests pose detection risks as surface hitchhikers that
Table 15.4. Examples of summary information for exporting Australian mangoes to the EU and New Zealand (Source: adapted from AQIS, 2008b).
Documentation
Required for
EU New Zealand
Import Permit No NoPhytosanitary Certifi cate Yes Yesa
Additional Declaration No Yesb
Post Entry Quarantine No NoEX188 No NoEX46 No NoRadiation Statement No No
a Treatment details, including date of treatment, are to be endorsed on the Phytosanitary Certifi cate in the treatment section. The treatment is to be shown as: irradiation at minimum 250 gray.b The mangoes in this consignment have been treated in accordance with Appendix 12 of the Bilateral Quarantine Arrangement between New Zealand Ministry of Farming and AQIS.

Postharvest Technology 559
can be detected visually by inspectors. Such pests need to be controlled in the fi eld and removed before the fruits are exported. Internal pests, such as wee-vils, fruit fl y immatures or larvae of Lepidoptera, pose additional risks because of diffi culties of detection and their potential to damage fruit fl esh and/or mango seed. Immatures of mango seed weevil (Sternochetus mangiferae (F.)) occur in mango seed (but not fl esh) in most of Africa, Asia, Australia, the Pacifi c and the Carribbean (Waite, 2002), and are diffi cult to kill in situ with-out damaging the market quality of the treated mangoes. Orchard-control measures and surveying are discussed by Hansen (1991, 1993), Waite (2002) and Wittenberg (2007). The mango pulp weevil (Sternochetus gravis (F.), syn. Sternochetus frigidus (F.)) occurs in India, Bangladesh, part of the island of Palawan in the Philippines and a few other regions in South-east Asia (Waite, 2002; Astridge and Baron, 2007c; Catindig and Kong, 2007; Walker, 2007a). It causes severe damage to the fruit pulp only (de Jesus et al., 2007).
Follett and Gabbard (2000) concluded that mango seed weevil does not seriously affect mango yield or marketability. Nevertheless, the seed weevil is a major quarantine concern for countries which have not recorded it or claim area freedom, that is Middle Eastern countries and China (Waite, 2002). Seed and pulp-attacking Lepidoptera pests are quarantine risks in some countries (Waite, 2002; Walker, 2007b; Yarrow and Chandler, 2007). Entry of mangoes from countries having mango weevil and other Sternochetus spe-cies may be restricted or prohibited into countries free of these pests. Exten-sive surveying, sampling, implementation of fi eld control measures and/or area-freedom certifi cation and maintenance may be necessary for approval of market access (Johnson and Heather, 1995; Waite, 2002). Disinfestation treatments that ensure weevils are not able to reproduce may be acceptable when dosages for mortality damage fruit excessively.
Fruit fl y disinfestationTephritidae are the most important mango pests and occur wherever man-goes are grown (Waite, 2002). Eggs are oviposited below the peel. The wound provides an opening for microorganisms and scars the peel. Larvae feed and tunnel throughout the pulp. Fruit fl ies infest tropical and temperate fruits. It is the risk to temperate climate fruits and commodities produced in fl y-free areas that has prompted the development of quarantine restrictions and treatments for fruit fl y hosts.
At present, quarantine treatments against fruit fl ies are not required for fruit entering the European Union (EU), despite the large production of tem-perate fruit in fruit-fl y-free regions. Fly infestation has not been perceived as a threat because winter temperatures throughout much of the region effec-tively prevent establishment of the fl ies, despite geographical continuity with the distribution range of the Mediterranean fruit fl y (Ceratitis capitata (Wiedemann)). Canada does not require fruit fl y disinfestation of tropical produce for the same reason. Exotic fruit fl y pests could become established in southern Florida, Texas and California because of their subtropical climate. The USA requires that mangoes be disinfested by vapour heat, irradiation, hot water or hot air.

G.I. Johnson and P.J. Hofman560
In Queensland, Australia, fl y larvae infestation of mangoes in the mar-keting chain is rare, despite the widespread occurrence of endemic species of fruit fl ies (Bactrocera spp.). Preharvest control measures, and grading out of coloured fruit at the packhouse, effectively eliminate infestation of most com-mercial consignments; however, mangoes consigned to the Australian states in temperate regions free of the fl ies must be disinfested against fruit fl ies to help ensure area freedom of temperate-fruit-production areas (RSPM, 2004; Jessup et al., 2007). When effective fi eld control and grade-out of ripening fruit is in place, the mandatory disinfestation of mature-green mangoes entering the exclusion zone is probably unnecessary.
Potentially acceptable quarantine treatments that disinfest mangoes include vapour heat, hot air, hot water immersion, irradiation, quick-freezing, combination treatments and some miscellaneous treatments (Taylor et al., 2002; Ducamp Collin et al., 2007). The major constraints in the development of treatments have been the susceptibility of mangoes to heat, cold and irra-diation damage and O2 depletion, and the extensive research and negotiation required to obtain market access approvals to high-end markets (i.e. the USA, the EU and Japan). Treatments need to be verifi ed as non-damaging to a range of cultivars by fruit size by environments likely to be encountered (Jacobi and Gowanlock, 1995; ISPM, 2007b). Treatments that cannot be used because they lower fruit quality at dosages that kill pests are methyl bromide fumigation (Spalding et al., 1977) and cold temperature storage (Kane and Marcellin, 1978).
Vapour heatVapour heat treatment (VHT) involves heating air that is nearly saturated with moisture, and passing the air stream across the fruit (Jacobi et al., 2001b). When the temperature of the mango fruit is at or below the dew point of air, condensation occurs on the fruit surface and rapidly heats the fruit by con-ductive energy transfer. The core of the fruit next to the seed is heated to c. 45°C for the required time before cooling. Fruit have to be sorted for size before treatment because of different rates of attaining the required core temperature.
Vapour heat is used worldwide to disinfest mangoes of fruit fl ies. Jacobi et al. (2001b) list the VHT protocols approved for importation of mangoes into Japan from the Philippines, Taiwan, Thailand, Australia and Mexico. Conditions range from 43–47°C pulp core temperature for 10 min to 6 h; however, the most common treatment conditions are 46–47°C for 10–30 min. Melon fruit fl y (Bactrocera cucurbitae Coquillett) immatures in mangoes from Okinawa were killed at 44 ± 0.3°C core temperature for 3 h (Sunagawa et al.,1987). Taiwanese mangoes infested with melon fl y can be disinfested with vapour heat at 47.5°C until the centre pulp is >46.5°C for 45 min (Kuo et al.,1987).
A VHT schedule was approved against Queensland fruit fl y (Bactrocera tryoni), in ‘Kensington Pride’, ‘R2E2’, ‘Keitt’, ‘Palmer’ and ‘Kent’ from Aus-tralia for the Japanese market (AQIS, 2008b), which consists of a core tem-perature of 47°C for 15 min. The United States Department of Agriculture,

Postharvest Technology 561
Animal and Plant Health Inspection Service Plant Protection and Quarantine (USDA-APHIS PPQ) approved VHT as a quarantine treatment for Mexican fruit fl y (Anastrepha ludens (Loew)) and other Anastrepha species in ‘Manila’, and for mangoes from Taiwan infested with oriental fruit fl y (Anonymous, 1994a). Generic guidelines for use of VHT in treating commodities for the USA market are provided by the USDA-APHIS PPQ manual on vapour heat. Mangoes from Taiwan imported into Australia must be treated until the pulp temperature has been 46.5°C for 30 min (AQIS, 2008a).
Hot airHot or forced hot air systems also heat the air to 40–50°C, but at a lower RH. Relative humidity usually remains >50%, depending on ambient RH, but is never high enough to produce condensation. Heat is transferred to the fruit by convection, with no condensation of water on the skin (Gaffney and Arm-strong, 1990; Jacobi et al., 2001b). Relative humidity should be high enough to prevent fruit desiccation during treatment. Transfer of heat from the air to the skin is slow compared with VHT. Mangan and Ingle (1992) reported that a mean centre pulp temperature of >47°C killed all stages of West Indian fruit fl y, Anastrepha obliqua (Macquart), in Mexican mangoes, and Sharp (1992) found a centre pulp temperature of >46°C killed all stages of Caribbean fruit fl y, Anastrepha alletis (Loew), in Florida-grown mangoes.
Hot waterProvided that fruit are not damaged, hot water immersion is environmen-tally safe and effi cient for killing mango pests. Use of hot water to kill fruit fl y eggs and larvae intensifi ed in the USA when the Environmental Pro-tection Agency (EPA) removed ethylene dibromide from the market as a chemical fumigant because of health concerns (Anonymous, 1983). Sharp and Spalding (1984) showed that mangoes could be disinfested of Caribbean fruit fl y using hot water. The work led to more studies in Haiti and a disinfestation method for West Indian fruit fl y (Sharp et al., 1988), as well as Mediterranean fruit fl y and other Anastrepha spp. in Texas and Mexico (Sharp et al., 1989a, b), Puerto Rico (Segarra-Carmona et al., 1990) and Peru (Sharp and Picho-Martinez, 1990). Nascimento et al. (1992) developed a hot water treatment for fruit fl ies in mangoes in Brazil. Hot-water-treated mangoes may be imported into the USA from Mexico, Central America, South America and the West Indies (Anonymous, 1994a). Typical treatments include 46.1°C for 65 min for smaller fruit to 90 min for larger fruit (Jacobi et al., 2001b). Large commercial hot-water-treatment facilities have been constructed, certifi ed by the USDA-APHIS PPQ, and used in Mexico, Central and South America, and the West Indies. Generic guidelines for the use of hot water are provided by the USDA-APHIS PPQ manual for hot water treatment.
In Australia, Smith (1992) showed that immersing fi ve Australian mango cultivars in 48°C water for 30 min killed eggs and larvae of Bactrocera aquilo-nis; however, ‘Kensington Pride’ is more sensitive to hot water than to vapour heat, so the latter has been adopted for disinfestation of mangoes in Australia (Jacobi et al., 1994). Grové et al. (1997) found that treatment of several cultivars

G.I. Johnson and P.J. Hofman562
in hot water at 46.1°C for 90 min followed by refrigeration for 24 h did not damage fruit, although some cultivars showed severe lenticel damage. Refrigeration of ‘Tommy Atkins’ fruit immediately after treatment resulted in scald development. Weevils in ‘Alphonso’ mangoes from India were not killed when infested mangoes were immersed in water at 48–52°C for up to 90 min and 54–70°C for up to 5 min (Shukla and Tandon, 1985).
Compared with hot air treatments, hot water treatments can damage the skin, partly because of rapid heat transfer from the water to the skin com-pared with from the skin to the centre of the fruit. Damage includes skin scald-ing, lenticel damage, cavities, white starchy areas in the fl esh and delayed ripening (Jacobi et al., 2001b). Several factors infl uence damage severity after heat treatment, for example cultivar, temperature and duration (Jacobi et al., 2001b). Immature fruit have low heat tolerance, and small fruit are damaged by heat more readily than large fruit. Conditioning treatments (i.e. 37°C core temperature, for at least 12 h in air) can reduce injury, and preharvest condi-tions, especially rainfall before harvest, can increase skin damage (Esguerra and Lizada, 1990; Esguerra et al., 1990; Jacobi and Wong, 1992; Jacobi et al.,1994, 1995; Jacobi and Giles 1997). Better understanding of these infl uences could increase the commercial potential for hot water disinfestation.
Hot water dips could pose human health risks. Sivapalasingam et al. (2003) reported that an outbreak of Salmonella enterica that infected 72 patients from 13 USA states may have been due to contamination of hot-water-dipped mangoes from a single farm in Brazil. No outbreaks were reported among consumers in the EU of mangoes from the same farm, and the EU does not require hot water disinfestation.
IrradiationIrradiation involves γ rays (at <1000 Gy), X-rays, electrons and microwaves (Thomas, 1986; Velasco and Medina, 2004; Follett et al., 2007; Moreno et al., 2007). A 2005 FAO/International Atomic Energy Agency (IAEA) report indi-cated that >20 irradiation facilities have been planned, constructed or reno-vated in ten countries, some of which are mango exporters (Eustice, 2007). Radiation treatments have been developed for fruit fl ies in mangoes from Florida, Mexico, India and Australia. Von Windeguth (1986) treated mangoes with 76 Gy and disinfested them of Caribbean fruit fl y eggs and larvae. Third instar Mediterranean fruit fl y larvae in Mexican mangoes irradiated with 250 Gy did not emerge from pupae, and 60 Gy applied to third instar Mexican fruit fl y, and West Indian fruit fl y in Mexican mangoes prevented adult emer-gence (Bustos et al., 1992). Bustos et al. (2004) recommended a generic dose of 150 Gy for control of Mexican fruit fl y (A. ludens), the West Indian fruit fl y (A.obliqua), the sapote fruit fl y (Anastrepha serpentina) and the Mediterranean fruit fl y (C. capitata) in mango. ‘Kensington Pride’ mangoes infested with eggs and larvae of Queensland fruit fl y and Bactrocera jarvisi (Tryon) are disinfested with 74–101 Gy (Heather et al., 1991).
International guidelines for the use of irradiation as a phytosanitary measure are available (ISPM, 2003), and recently a fast track process has been proposed as an Annex to ISPM 28 (ISPM, 2008), which endorses irradiation

Postharvest Technology 563
at 70 Gy as a generic treatment to control Anastrepha spp. in fruit and vegeta-bles by extrapolating work on mango by Bustos et al. (2004). Heather (2004) provides generic guidelines for the development of irradiation protocols for disinfestation. Fruits are never exposed to radioactive materials (Anony-mous, 1986) and most modern treatment units use an electron beam process rather than a radioactive source for irradiation.
Irradiation can be used for controlling seed weevil and lepidopterous pests in fruit. Seo et al. (1974) reported that 206 and 329 Gy killed mango weevil in Hawaiian mango. Thomas (1975) showed that 500 Gy killed all mango weevil larvae and pupae and 750 Gy prevented adults from emerging from mangoes in Africa. A dose of 500 Gy, however, did not disinfest ‘Alphonso’ mangoes of seed weevil (Shukla and Tandon, 1985). Indian man-goes from approved packhouses must be irradiated with a minimum of 400 Gy at an approved and certifi ed irradiation treatment facility using Cobalt-60 (APEDA, 2007). A quarantine treatment of 300 Gy has been approved to ster-ilize mango seed weevil in mangoes exported from Hawaii to USA mainland markets (Follett, 2004). Follett and Lower (2000) demonstrated control of Cryptophlebia illepida (Butler), Cryptophlebia ombrodelta (Lower) and Cryp-tophlebia illepida (Lepidoptera: Tortricidae), and an irradiation quarantine dose of 250 Gy has been approved for Hawaiian mangoes. The treatment also controls fruit fl ies (Follett, 2004).
USA regulations covering irradation are described in the Code of Federal Regulations GPO Access (2008), revised annually (Wall, 2008), and this sum-marizes approved treatments for a range of pests (EPA, 2002) (Table 15.5),
Table 15.5. Minimum absorbed dose of gamma irradiation required by USDA for specifi c pests (Source: adapted from EPA, 2002).
Scientifi c name Common name Minimum absorbed dose (Gy)
Anastrepha ludens Mexican fruit fl y 70Anastrepha obliqua West Indian fruit fl y 100Anastrepha serpentina Sapote fruit fl y 100Anastrepha suspensa Caribbean fruit fl y 70Bactrocera cucurbitae Melon fruit fl y 150Bactrocera dorsalis Oriental fruit fl y 150Bactrocera jarvisi Jarvis fruit fl y 100Bactrocera tryoni Queensland fruit fl y 100Brevipalpus chilensis False red spider mite 300Ceratitis capitata Mediterranean fruit fl y 150Cryptophlebia illepida Koa seed worm 250Grapholita molesta Oriental fruit moth 200Sternochetus mangiferae Mango seed weevil 300All other fruit fl ies of the family Tephritidae which are not listed above
150
Plant pests of the class Insecta not listed above, except pupae and adults of the order Lepidoptera
400

G.I. Johnson and P.J. Hofman564
many of which can infest mangoes. The USDA-APHIS PPQ manual on irradia-tion provides generic guidelines. Irradiation was approved for the USA market as a phytosanitary treatment for all fresh fruits and vegetables from all coun-tries in 2002. Effects of -irradiation on mango fruit quality and disease control have been reported (Mitchell et al., 1992; Moreno et al., 2006; Reyes and Cisneros-Zevallos, 2007; Wall, 2008). Only marginal disease control was obtained with ‘Kensington Pride’ at the highest non-deleterious doses for mature-green fruit (300 Gy), with additive effects of disease control treatments and irradiation on disease reduction (Johnson et al., 1990a). Disease control may be more effective in cultivars with greater tolerance of irradiation (van der Linde and Thord-Gray, 1986; Johnson et al., 1990a). Other types of irradiation have been evaluated for mango disinfestation but none has been adequately suitable.
Quick freezingQuick freezing of mango, lowering the temperature to –17°C and holding at –6°C or below for 48 h is used to disinfest mangoes for processing (Anony-mous, 1994a; PPQ, 2007). The process is not approved for importing man-goes with seeds from most of the West Indies, French Guiana, all countries outside of North, Central and South America, Oceania, Hawaii, South-east Asia, the Philippines and the Republic of South Africa into continental USA because mango weevil could be present (Anonymous, 1994a; PPQ, 2007).
FumigationFumigation is an ideal methodology for ensuring effective control when the fumigant is effective and safe to use. Until 1994, New Zealand required fumi-gation of mangoes from Australia, the Cook Islands and the Philippines using 33, 29 or 22 g/m3 ethylene dibromide at 10–15, 15.5–19.5, or 20°C and above, respectively, at normal atmosphere pressure (NAP) to disinfest man-goes of fruit fl ies before entry. As part of the international phase-out of ozone-depleting substances, the process was banned in 1994 (Anonymous, 1992; N.W. Heather, personal communication, Brisbane, 1994) and most applica-tions as a fruit fumigant have ceased worldwide. Methyl bromide was phased out completely in the USA in 2005, but some emergency uses for quarantine applications may be permitted, e.g. to destroy a serious quarantine pest in an imported consignment or to meet offi cial requirements of an importing coun-try (EPA, 2008). Mangoes imported into Australia from countries where fruit fl ies occur must be fumigated with 16–35 g/m3 ethylene dibromide for 2 h at 21–26°C or above (Anonymous, 1985, 1988).
Phosphine is widely used as a fumigant of durable produce (grains and tobacco). It provides effective control of fruit fl y larvae and other pests in temperate fruits under experimental conditions (Horn and Horn, 2004). However, phosphine when mixed with water is highly explosive and the vapour is toxic to humans, so prospects for utilization are not strong.
Miscellaneous treatmentsCHEMICAL TREATMENTS. Postharvest chemical treatments using dimethoate are effective against Queensland fruit fl y with ‘Kensington Pride’ (Swaine et al.,

Postharvest Technology 565
1984). The treatment is required for Australian-grown mangoes entering all Australian states except Queensland and New South Wales, but is under review. The USA and the EU do not allow the use of chemicals to disinfest mangoes.
NATURAL PRODUCTS. The short shelf life of mango and the high level of insect mortality required obviates the use of natural products for disinfestation. Suhaila and Halim (1994) reported the potential of low toxicity, insecticidal compounds from edible plants that may be effective for topical application to harvested fruit. Extracts of black pepper (Piper nigrum) were particularly active in laboratory tests against vinegar fl y (Drosophila melanogaster (Meigen)).
ATMOSPHERES. CA and MA regimes could have potential for disinfesting man-goes, but there has been less interest in the technology because heat treatments and irradiation are faster (Ke and Kader, 1992; Yahia and Tiznado-Hernandez, 1993; Yahia and Vazquez-Moreno, 1993; Yahia, 1994; León et al., 2000). Treat-ments are limited to regimes which do not adversely affect ripe fruit quality. León et al. (2000) found that CA of 1% O2 and 30 or 50% CO2 disinfested ‘Manila’ mangoes of A. obliqua, but damage (as spongy tissue) was unaccept-ably high.
Shrink-wrapping has been ineffective as a quarantine treatment to disin-fest mangoes of fruit fl y immatures. Gould and Sharp (1990) reported that the time needed to disinfest Florida-grown mangoes infested with Caribbean fruit fl y eggs and larvae exceeded the shelf life of wrapped mangoes.
COMBINATION TREATMENTS. Serial applications of two or more treatments, which alone do not achieve quarantine security, have been used to disinfest mangoes. Seo et al. (1972) reported that eggs and larvae of Mediterranean fruit fl y, orien-tal fruit fl y and melon fl y were killed in mangoes immersed in water at 46.3°C for 120 min and then fumigated with ethylene dibromide. Lin et al. (1976) reported that all oriental fruit fl y and melon fl y larvae in Taiwan-grown man-goes were killed when fruit were immersed in 48–50°C water for 120 min, hydrocooled, dried and cooled, and then fumigated with ethylene dibromide.
Controlled Atmosphere/Temperature Treatment System (or CATTS) technology applies a short heat treatment in a low O2/high CO2 environ-ment, and controls quarantine insect pests while maintaining commodity quality (Mitcham, 2007; Neven, 2008). Trials using CATTS with mangoes have been conducted in Australia with promising results. Varith et al. (2007) evalu-ated a microwave-vapour heat treatment (MW-VHT) disinfestation technol-ogy for mangoes: the microwave component for pre-heating and the VHT component for the holding process. Temperatures of 46–55°C and holding times of 2–20 min effectively disinfested fruit of oriental fruit fl y eggs without effects on physico-chemical parameters, compared to untreated fruit. There was less heat damage compared with conventional VHT only fruit. MW-VHT shortened the process time by 90% compared with the conventional VHT.
PACKAGING. Some markets, for example Japan and the USA, require that fruit must be packed into insect-proof packages following disinfestation to preclude

G.I. Johnson and P.J. Hofman566
reinfestation during transportation or storage. The disinfestation facility feeds fruit into an insect-proof area within which waxing (optional), grading and packing occur.
15.7 Preparing Fruit for Market
Surface coatings
Surface coatings are used to improve fruit appearance and to alter gas per-meability to reduce moisture loss or retard ripening. Commercial use of sur-face coatings on mango fruit needs to be considered carefully because of the fi ne balance between benefi cial and undesirable effects on fruit quality. Neg-ative effects of coatings include reduction in chlorophyll loss (Fonseca et al., 2004a), anaerobic conditions and off-fl avours (Amarante and Banks, 2001) and skin damage, possibly due to cytotoxic reactions with other components in the coating formulation (Bower et al., 2003). Generally, coatings have less effect on delaying ripening during cold storage, compared with extending the shelf life at typical ripening temperatures (Amarante and Banks, 2001). Less signifi cant effects are observed in more mature and in ripening fruit. Coatings often delay skin colour change rather than softening, which increases the risk of soft, green fruit with less consumer appeal.
Coatings are generally emulsions of synthetic (e.g. polyethylene) or nat-ural (e.g. polysaccharides, carnauba, beeswax, etc.) origin. Surface coatings containing waxes, oils (e.g. carnauba, beeswax, etc.) and resins (e.g. shellac) have a greater effect on limiting water loss then reducing O2 and CO2 perme-ability, compared with those containing polysaccharides, (e.g. those based on cellulose) (Amarante and Banks, 2001). Formulations based on shellac result in a shinier appearance than those based on carnauba wax and polysaccharide-based waxes (Baldwin et al., 1999; Hoa and Ducamp, 2008).
Factors other than coating formulation can affect fruit gas permeability, i.e. cultivar, variations in skin permeability between fruit, inconsistency in coating thickness during application, interference from water during appli-cation causing coating cracking and coating thickness and evenness-of-spread over the fruit surface. The effect of coating on fruit quality can vary with holding conditions because of larger temperature effects on respiration rate than on coating permeability.
Shorter and Joyce (1994) found commercially formulated Avocado and Passionfruit Wax, a polyethylene and shellac emulsion, and Technimul 9122 Wax, a polyethylene-based emulsion, were acceptable with ‘Kensington Pride’ mango, while Peach Wax, a polyethylene-based emulsion and starch solution was unacceptable. With Peach Wax, deleterious modifi ed atmosphere effects on colour development, softening and fl avour were obtained (El Ghaouth et al., 1992b; Shorter and Joyce, 1994). Coating ‘Tommy Atkins’ mango with a carnauba-based coating and BeeCoat (based on beeswax) reduced water loss, shrinkage, chlorophyll breakdown, CI and decay after cold storage, and Bee-Coat also reduced red lenticel discoloration (Feygenberg et al., 2005). With

Postharvest Technology 567
‘Tommy Atkins’, polysaccharide and carnauba-based coatings modifi ed the atmosphere within the fruit and reduced decay, but only the polysaccharide-based coating delayed ripening (Baldwin et al., 1999). The carnauba-based coating signifi cantly reduced water loss compared with the polysaccharide-based coating treatments; carnauba-based coatings result in lower water per-meability and higher O2/CO2 permeability.
Coatings may reduce surface defects. Excessive water loss is associated with increased skin CI in avocado and mango, and carnauba-based coatings reduce CI in cold-stored mangoes (Bower et al., 2003). In this study, the carnauba-based coating contained numerous holes, which allowed respiration gas exchange (thereby preventing anaerobic respiration), while still providing effi cient control of water loss. Surface coatings may also reduce sapburn, skin browning and lenticel damage (Shorter and Joyce, 1994), but incorporating these potential benefi ts into commercial systems may be diffi cult.
Waxes should be applied by roller brushes in a specifi cally designed wax applicator or by very light hand application. Dipping fruit in a wax emulsion is not recommended. A uniform fl ow of fruit through the wax applicator must be maintained to prevent uneven wax application. Fruit should be dry before entering the wax applicator, otherwise foaming of water-emulsion waxes may occur. Brushes on the wax applicator need to be completely satu-rated with the wax mixture before any fruit passes over them. Complete cov-erage of the entire fruit surface is essential. Patchy application can be caused by insuffi cient wax, too few brushes following application (minimum of six brushes required), poor and/or inadequate drying facilities, and overload-ing of the unit. Brushes should be kept soft with regular washing with hot water.
Packaging
Packaging provides conveniently sized carriage units for product, protects individual fruit from contact rub and compression damage, and excludes dirt, pests and contaminants. McGregor (1987) and Hilton (1994) discussed key aspects of packaging for tropical produce. Packaging is also a marketing tool. Design and colours of symbols and text on carton exteriors portray a marketing image. Manufacturers of consumer products exploit packaging to great advantage (along with advertising) to increase both fi rst-time and repeat sales. Marketing and design professionals may be involved in the development and customer evaluation of product packaging. Cultural pref-erences need to considered, e.g. use or avoidance of red for some Asian mar-kets. Packaging is the external face of brand loyalty. Consistent product performance and quality is the core.
Some constraints to packaging may be specifi ed by market regulations, including carton dimensions and labelling requirements. Country of origin, cultivar, grower, packing shed, market agent, count (number per carton and weight range) and class may be required. The word ‘mangoes’ should be clearly visible (Anonymous, 1993). The information appears on the narrow

G.I. Johnson and P.J. Hofman568
sides of the cartons. Storage and product use information can also be printed on the cartons. Many QA systems require adequate labelling linked to appro-priate record keeping for plate-to-farm traceability. Clear labelling facilitates correct delivery, allows immediate buyer recognition of product profi le and ensures maintenance of accurate sales records. An exporting country may fi nd it of value to identify individual packers by barcoding or numbers stamped on cartons, so that sources of faulty packaging can be traced. Some countries also use date codes which enable exporters to determine the fresh-ness of the produce at the point of export and evaluate an importers’ capacity to achieve adequate turnover of the fruit without prolonged storage. It also provides invaluable feedback on the effi ciency of the total distribution chain.
Cartons used for export should be clean, strong, unbroken and new. The water absorption capacity of the material should be evaluated as excess absorption will lead to collapse on the pallet. The cartons’ strength will depend on the starch used by the manufacturer, the outer liner and the direc-tion and numbers of fl uting in the carton (Anonymous, 1994b). There is increasing pressure in the EU for recyclable packing material. Cartons that are recyclable should be marked with the appropriate international symbol. Returnable plastic crates are increasingly being used for domestic trade, but the return cost would make this less profi table for international trade.
Inspection
In some countries, independent inspectors check the fruit prior to palletizing to ensure that the relevant marketing, residue and phytosanitary standards have been met. Fruit for Japan is disinfested under the supervision of a Japa-nese inspector. Further inspections are usually made at the port of exit. Some exporting countries require a declaration by the grower to ensure that fruit will comply with the standards specifi ed by importing countries.
Palletizing
Handling mangoes on pallets allows convenient movement of large volumes of fruit. McGregor (1987) described critical features and arrangements for loading. The disadvantages of pallets for export are the cost, lower numbers of cartons per sea container and loss. Some domestic markets have pallet share systems. Relevant markets and transporters should be consulted con-cerning required pallet dimensions and appropriate access for fork-lift sys-tems. The correctly sized pallet, for example as designated by the ISO, which is designed to fi t snugly into a standard sea container, should also be used for the local market.
Precision stacking with each box fi tted exactly on top of the one below minimizes risk of damage. Collapsed or lopsided pallet stacks have usually been due to careless stacking and/or loose placement in the shipping con-tainer. Pallet slats should not block ventilation holes in the cartons. Cartons

Postharvest Technology 569
should be register-stacked so that ventilation is continuous. Link sheets, which bind the cartons together at intervals, should also be designed to ensure con-tinuous ventilation through the pallet. In the cold room, pallets should not be stacked against a wall or placed directly against each other (Boelema, 1987).
Precooling
Precooling removes fi eld heat from the product and lowers the temperature to that required for ripening, transportation or storage. Precooling also reduces the cooling demand on any in-transit cooling system. Precooling concepts and systems are described by Thompson et al. (2002). Forced-air cooling systems effi ciently and rapidly remove fi eld heat, and are preferred for bringing fruit to storage temperature. High RH systems are preferred as they reduce fruit water loss. Hydrocooling can increase the risk of infection by wound pathogens (i.e. Rhizopus spp.) and are less effective with large fruit. Kitinoja and Kader (2003) describe low-cost cooling facilities for use in devel-oping countries.
Ethylene and ripening
Induction of ripening is routinely employed with mangoes. There are effec-tive low technology methods involving calcium carbide (releases acetylene which mimics ethylene) or the leaves of particular trees (Lizada, 1994). More sophisticated systems include generation of ethylene from ethanol using catalytic conversion, pure ethylene gas, or a mixture of ethylene and an inert gas (CO2 or N2) to reduce the risk of explosion with 3–30% ethylene in air (Reid, 2002). A number of automatic ethylene control systems are available (PDS, 2008) to maintain ethylene concentrations within required limits.
Climacteric fruit have differing sensitivities to ethylene. ‘Kensington Pride’ mango is sensitive to concentrations as low as 0.01 l/l (O’Hare et al., 1994). Ripening is enhanced with concentrations up to 5–10 l/l, with very little ben-efi t at >50–100 l/l (Nguyen, 2003). There is more yellow colour on the ripe fruit when ripened at 20°C with 10 l/l ethylene for 3 days compared with no ethylene, resulting in a more attractive appearance. Also, diseases are gener-ally less in these fruit (Table 15.6), presumably because fruit ripen more quickly with less time for disease development. Good ethylene treatment can improve presentation appearance and increase saleable life (defi ned as the days from when the fruit reach at least 60% yellow skin colour to when the fruit had lost saleability because of disease) (Ledger et al., 2002a).
Ethylene can also reduce quality if not used appropriately. Ripening ‘Kensington Pride’ fruit at <18°C with ethylene can result in soft fruit with less yellow skin colour, most likely because ethylene stimulated softening to a greater extent than chlorophyll loss (Nguyen, 2003). Ripe fruit disease can also be greater. These effects can be aggravated with concentrations above 100 l/l (Fig. 15.5). ‘Kensington Pride’ fruit must be cooled to <24°C before
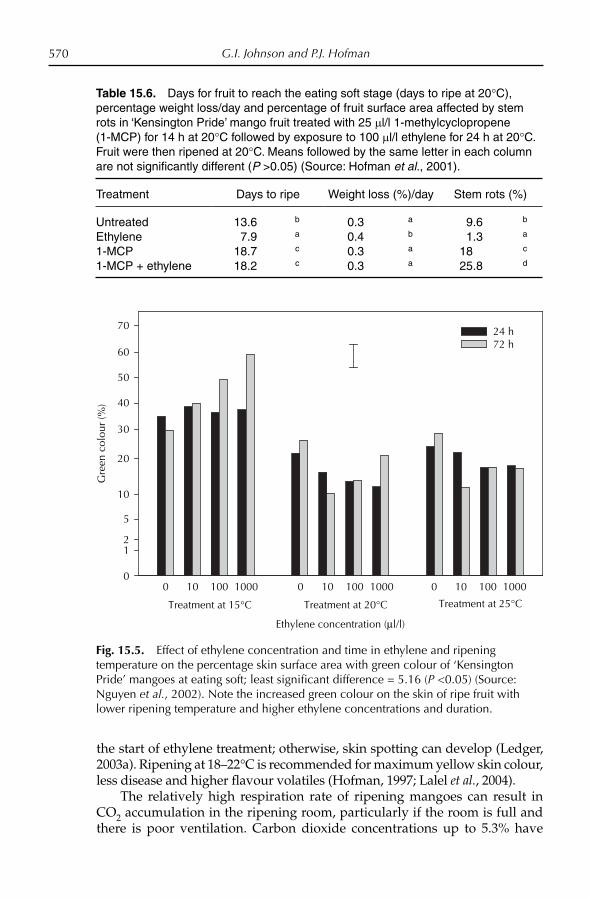
G.I. Johnson and P.J. Hofman570
the start of ethylene treatment; otherwise, skin spotting can develop (Ledger, 2003a). Ripening at 18–22°C is recommended for maximum yellow skin colour, less disease and higher fl avour volatiles (Hofman, 1997; Lalel et al., 2004).
The relatively high respiration rate of ripening mangoes can result in CO2 accumulation in the ripening room, particularly if the room is full and there is poor ventilation. Carbon dioxide concentrations up to 5.3% have
Table 15.6. Days for fruit to reach the eating soft stage (days to ripe at 20°C),percentage weight loss/day and percentage of fruit surface area affected by stem rots in ‘Kensington Pride’ mango fruit treated with 25 l/l 1-methylcyclopropene (1-MCP) for 14 h at 20°C followed by exposure to 100 l/l ethylene for 24 h at 20°C. Fruit were then ripened at 20°C. Means followed by the same letter in each column are not signifi cantly different (P >0.05) (Source: Hofman et al., 2001).
Treatment Days to ripe Weight loss (%)/day Stem rots (%)
Untreated 13.6 b 0.3 a 9.6 b
Ethylene 7.9 a 0.4 b 1.3 a
1-MCP 18.7 c 0.3 a 18 c
1-MCP + ethylene 18.2 c 0.3 a 25.8 d
Ethylene concentration (μl/l)
0 10 100 1000 0 10 100 1000 0 10 100 1000
Gre
en c
olou
r (%
)
0
12
5
10
20
30
40
50
60
70 24 h 72 h
Treatment at 15°C Treatment at 20°C Treatment at 25°C
Fig. 15.5. Effect of ethylene concentration and time in ethylene and ripening temperature on the percentage skin surface area with green colour of ‘Kensington Pride’ mangoes at eating soft; least signifi cant difference = 5.16 (P <0.05) (Source: Nguyen et al., 2002). Note the increased green colour on the skin of ripe fruit with lower ripening temperature and higher ethylene concentrations and duration.

Postharvest Technology 571
been recorded in ripening rooms (Ledger, 2007), which can cause more green colour and a dull appearance on the ripe fruit (Nguyen, 2003). Ripening room CO2 concentrations should be maintained at <1% with adequate ventilation to minimize fruit quality loss (Kernot et al., 1999; Ledger, 2007).
Accidental exposure of mangoes to ethylene and its analogues from adja-cent ripening rooms, exhaust fumes from internal compression engines or wound ethylene produced from damaged/ripening fruit can cause prema-ture ripening. Various systems can remove unwanted ethylene, for example oxidizing mechanisms such as potassium permanganate either in sachets or in ethylene scrubbing units in storage rooms, catalytic oxidizers or ozone-based systems (Reid, 2002). Smartfresh™ (active ingredient 1-methylcyclopropene; 1-MCP) is a relatively new approach for preventing undesirable ethylene effects. 1-MCP is a structural analogue of ethylene and irreversibly binds to the ethylene receptors in the plant, thus preventing ethylene-initiated ripen-ing. Ripening re-commences as additional ethylene-receptor sites are pro-duced in the fruit (Blankenship and Dole, 2003). Generally, Smartfresh™ treatment is applied in well-sealed cold rooms or plastic tents as soon as pos-sible after packing. 1-MCP concentrations of 250–200,000 l/l for 12 h are optimum for delaying ripening (Jiang and Joyce, 2000; Hofman et al., 2001; Adkins et al., 2002; Penchaiya et al., 2006), although most reports state 250–1000 l/l. 1-MCP treatment completely negated any effect of subsequent eth-ylene on ripening, and can almost double the days to eating soft compared with ethylene-treated fruit ripened at 20°C (Table 15.6) (Adkins et al., 2002). However, the 1-MCP effects were less in more mature fruits (Alves et al., 2004), and ethylene exposure before 1-MCP will negate any 1-MCP benefi t (Adkins et al., 2002). Any benefi cial effects of 1-MCP also appear to be less with longer-term storage (Hofman, unpublished results). 1-MCP treatment can cause more disease on ripe fruit, because the longer days to ripen allows more disease development (Hofman et al., 2001). Sourcing fruit from well-managed orchards can help minimize this effect (Adkins et al., 2005).
For some domestic markets, on-farm treatment of mangoes with ethyl-ene is used to ensure that fruit have more attractive colour when they are displayed at the wholesale market 48–72 h after dispatch from the farm. This practice improves returns as fruit can be delivered to retail outlets ready-to-eat. Ethylene induction of ripening is undesirable for more distant markets because fruit arrive at the market too ripe for sale, with greater risk of bruis-ing and disease.
15.8 Pre- and Post-shipping Storage
Cool storage
Cool storage is important when delivery time from harvest to the consumer exceeds the typical ripening time (5–10 days). The ideal storage temperature is dictated by the risk of CI, fruit ripening and disease development during stor-age, and storage time. CI is fi rst noted as greying of the skin, which intensifi es

G.I. Johnson and P.J. Hofman572
with lower temperatures and longer duration (Phakawatmongkol et al., 2004; Suresh et al., 2004). In more severe cases fl esh discoloration and abnormal ripening can occur. CI development can occur at regimes of 3–12°C (Sadasi-vam el al., 1971; Thomas and Oke, 1983; Chaplin et al., 1986 a, b, 1991a, b; Smillie et al., 1987; Thomas and Joshi, 1988; Medlicott et al., 1990b). Longer storage times require greater care with temperature selection, the quality of the fruit being stored and conditions before and after harvest. Storage should be for the minimum period necessary. The following factors affect the opti-mum storage temperatures and durations:
Genetic differences – cultivars differ in chilling sensitivity (Phakawat- ●
mongkol et al., 2004).Maturity – less mature fruit ripen more slowly at a given temperature, ●
and are more prone to CI and other storage-related disorders (Medlicott, 1985; Medlicott et al., 1987, 1990 a, b; Oosthuyse, 1993). Such fruit may not soften at all when exposed to temperatures that are suitable for stor-age of more mature fruit. In South Africa, adequately mature fruit can be stored at 8–10°C for 21–28 days (Oosthuyse, 1994). Placement in cold storage without delay and post-storage exposure to temperatures that promote ripening (e.g. 20°C) are important preconditions for success.Duration of storage – ‘Kensington Pride’ mangoes can be stored at 10 ● °C for 3 weeks or at 7°C for 2 weeks, after which skin colour development can be affected (McLaughlan and Wells, 1994). Generally, the shorter the storage time, the greater the tolerance to storage temperatures outside the 10–12°C range.Delays between harvest and cold storage, and ripeness stage – the longer ●
the delay between harvest and cold storage, the greater the risk of ripen-ing during storage. This applies particularly for more mature fruit. If prolonged, a delay may render refrigerated storage ineffective in pre-venting fruit from becoming soft during transit, despite the apparent ab-sence of softening on dispatch (Oosthuyse, 1994). Fruit should be picked, packed and placed in cold storage within 24 h. For fruit that have rip-ened, storage temperatures of less than 8°C can be used for up to 21 days without deterioration in quality during storage; however, the fruit will deteriorate rapidly after removal from storage (Van Straten and Oost-huyse, 1994). Some cultivars may be more sensitive to ripe storage, since ‘Kensington Pride’ fruit at the mid-climacteric stage will start to lose appearance after 3 days at 10°C because of increased disease and mild CI (H. Nguyen et al., 2004).Disease load and fruit tolerance of disease – certain mango cultivars are ●
very tolerant of postharvest pathogens (e.g. see Hassan, 2007). The con-ditions under which mangoes are grown may be unfavourable for infec-tion. In these situations, storage temperatures can be higher to reduce the risk of CI.
Development of CI in mango and other fruits is closely associated with antioxidant activity (Arafat, 2005; Kondo et al., 2005). Mango fruit held at 6°C for 10–20 days had lower antioxidant activity in the skin compared with fruit

Postharvest Technology 573
stored at 12°C. Application of several jasmonate derivatives before storage reduced CI at 6–7°C (González-Aguilar et al., 2000; Kondo et al., 2005), pos-sibly through an antioxidant mechanism. Other chemical treatments can also reduce CI. Polyamines occur naturally in fruit and decrease during storage under chill-inducing conditions, and application before storage can reduce CI (Nair et al., 2003). Salicylic acid appears to be involved in cell wall stability. Application of methyl salicylate, which breaks down to sali-cylic acid, signifi cantly reduced CI in ‘Zill’ mangoes stored at 7°C (Han et al., 2006). 2,4-D can also reduce mango CI, possibly through interaction with natural plant hormones and antioxidant levels in the fruit (Wang et al., 2008). Some of these treatments could have commercial application, but may have residue implications.
Decay is a major limitation to storage life. The incidence of postharvest decay on fruit that ripen after refrigerated storage is positively related to the duration of storage and the extent of ripening during storage (Oosthuyse, 1991, 1992, 1994). Disease development after post-storage exposure to ripen-ing temperatures can be reduced by minimizing the shipping period and by storing fruit at temperatures that inhibit softening and ground skin colour development. If CI occurs, disease develops earlier and will be more exten-sive (Oosthuyse, 1990).
Controlled and modifi ed atmosphere storage
Decreasing the O2 and/or increasing the CO2 concentration can have several advantages with respect to storage (i.e. reduced ethylene production, better fl avour retention, slowing softening and green skin colour loss and reduced CI) (Thompson, 1998; Yahia, 2006). However, if the O2 concentration is too low (dependent on cultivar, storage temperature, fruit maturity and ripeness stage) anaerobic respiration will commence, with associated production of ethanol and acetaldehydes, leading to off-fl avours and physiological disor-ders (Bender et al., 2000). Atmosphere modifi cation generally has less benefi t for tropical fruit compared with temperate fruit, but does have commercial potential for sea freight to distant markets. Atmosphere control can be active or passive, or combinations of the two. Surface coatings (see Surface coatings section under 15.7 Preparing Fruit for Market, this chapter) also provide modifi ed atmosphere inside the fruit.
With CA systems, O2 and CO2 concentrations are actively monitored and controlled by injecting N2 and CO2, or bleeding air into the container as required. In more passive systems, such as the MaXtrend® system (Maxtend, 2008) fruit respiration directly lowers O2 concentrations, and its concentra-tion is monitored and manipulated by venting as required. In some cases, the container is fl ushed with N2 at the start of storage to rapidly establish the desired atmospheres. Excess CO2 is absorbed with hydrated lime. In MA sys-tems, atmospheres are modifi ed by placing a semi-permeable membrane around the fruit (usually plastic fi lm), and relying on fruit respiration to modify the atmosphere.

G.I. Johnson and P.J. Hofman574
McLauchlan and Barker (1994) suggested 4% CO2 and 2–4% O2 for CA storage of ‘Kensington Pride’ mangoes at 13°C, and recommended further research on atmospheres <2% O2 and >10% CO2. Oxygen had the biggest effect on retarding skin colour and softening, with signifi cant retardation when decreasing from 4 to 2%. Subsequent research suggested that concen-trations of 1.5–2% may be more effective in retarding softening, although these concentrations may increase the risk of off-fl avours. ‘Tommy Atkins’ and ‘Haden’ can tolerate 2–3% O2 for 2–3 weeks at 12°C, but lower concentra-tions were not tested (Bender et al., 2000). In ‘Kensington Pride’ there was little additional capacity for CO2 concentrations between 6 and 10% to retard softening or loss of green colour (McLauchlan and Barker, 1994), although there may be some benefi t for storage of 1–2 weeks at concentrations >10% (Bender et al., 2000). For ‘Delta R2E2’ mangoes 3% O2 and 6% CO2 have been recommended (Lalel and Singh, 2006); however, this is a fi rm-fl eshed culti-var, which can perhaps tolerate higher O2 concentrations to improve vola-tiles, compared with the softer ‘Kensington Pride’. Longer storage times with CA could cause higher disease levels (Johnson et al., 1990b), higher acidity in the fl esh at eating soft (McLauchlan and Barker, 1994; Bender et al., 2000) and slower loss of green colour compared with non-stored fruit (Bender et al., 2000). For cultivars that normally have higher acidity, CA-stored fruit may need to be ripened for several more days to lower acidity.
Cold storage and CA can reduce volatiles production following ripening at room temperature. As the skin CI severity increases with decreasing stor-age temperature, total volatiles production appears to decrease (Singh et al., 2004). In ‘Kensington Pride’ and ‘R2E2’, CA storage signifi cantly reduces total concentrations of aroma volatile compounds compared with air-stored fruit, irrespective of storage period between 24–38 days (Singh et al., 2004; Lalel and Singh, 2006). Decreasing the O2 concentration from 3 to 1% at 6 or 8% CO2 or increasing CO2 concentration from 6 to 8% signifi cantly increased most of the monoterpenes, including terpinolene. Cold-stored fruit are known to have less aroma than those ripened without storage.
Fruit can tolerate short periods with <1% O2 or >20% CO2 (Yahia, 2006). This has been utilized for insect disinfestation (see Disinfestation section under 15.6 Packhouse Measures, this chapter). Mango can tolerate low O2 concentrations for 5 days at 20°C (Yahia, 1994). These short-term CA treat-ments may improve storage life or reduce CI during subsequent cold storage without atmosphere modifi cation; this has been noted with avocado (Truter and Eksteen, 1987; Pesis et al., 1994). Preliminary investigations suggested little benefi t of 20–60% CO2 for 1–8 days before cold storage of ‘Kensington Pride’ (Meiburg et al., 1998).
The optimum conditions for storage of each product to provide maximum storage life without quality loss must be determined taking into account cultivar, season and growing conditions. Measurement of chlorophyll fl uo-rescence has been used to monitor product performance under CA, with ad-justment of gas conditions to achieve the optimal storage-life/quality balance. Changes in chlorophyll characteristics and therefore chlorophyll fl uores-cence under CA occur before CI symptoms are obvious (DeEll and Toivonen,

Postharvest Technology 575
2003a). Thus monitoring changes in chlorophyll fl uorescence characteristics can provide advance warning of the potential for CI, and allow adjustment of storage conditions to minimize its development (DeEll and Toivonen, 2003b). This concept has now been marketed as ‘HarvestWatch’ (Harvest-Watch, 2008), and some preliminary success has been obtained with apples (Stephens and Tanner, 2005; DeLong et al., 2007).
Modifi ed atmosphere packaging (MAP) generally cannot reliably achieve the low O2 concentrations required to signifi cantly delay softening without damaging the fruit, but MAP can still have benefi cial effects relative to the costs of CA (Pesis et al., 2000; Rosa et al., 2001; Singh et al., 2001; Castro et al., 2005; Yahia, 2006). However, MAP can reduce quality if the cultivar/holding temperature/fi lm permeability/storage time combination is not optimal (Sornsrivichai et al., 1989), resulting in anaerobic conditions and off-fl avours. Excess moisture retention inside the bags can increase disease problems (Joyce and Patterson, 1994). Special fi lms have been developed with higher water vapour transmission rates (Pesis et al., 2000) or moisture absorption materials can be included. Ethylene absorption sachets can reduce chloro-phyll loss and red discoloration around the lenticels (Rosa et al., 2001).
MAP reduces weight loss (Singh and Janes, 2001; Bower et al., 2003), which maintains saleable weight, but may also reduce CI. There may be a direct relation between these, since weight loss can contribute to CI in avo-cado, and the reduction in CI obtained in mango through the use of wax coatings has been attributed to the same mechanism (Bower et al., 2003). Pesis et al. (2000) considered that lenticel discoloration is a symptom of mild CI, and noted that MAP reduced the red coloration around the lenticels in the blushed area and the green coloration around the lenticels in the green area of ‘Keitt’ mangoes. Less lenticel spotting occurs in ‘Kensington Pride’ man-goes stored under MAP (Yuen et al., 1993). It is not clear whether the reduc-tion in lenticel damage was due to CO2/O2 or humidity modifi cation.
Cultivar, fi lm type, number and mass of fruit per package, temperature, RH, time of storage, maturity of the fruit and production conditions are important for developing MAP systems (Brecht et al., 2003; Yahia, 2006). Important challenges are the differential effects of temperature on fruit respi-ration and fi lm permeability, resulting in differing gas concentrations around the fruit as temperature fl uctuates. Success with MAP depends on a consistent or at least predictable cold chain removal of the plastic fi lm before signifi cant temperature fl uctuations are likely to occur and using MAP fi lms that are unlikely to cause anaerobic conditions within the temperature range experi-enced in the cold chain. Brecht et al. (2003) suggested an approach to designing fl exible CA/MA systems to account for variations in the cold chain.
15.9 Transport
Transportation of tropical fruit and vegetables has been reviewed by McGregor (1987) and Thompson (2002). For local markets (<3 h access), transportation of fruit in non-refrigerated carriers is feasible, particularly if the fruit has

G.I. Johnson and P.J. Hofman576
been precooled and transported at night with few stops. Fruit must be shel-tered from direct sun and rain. For sea export, fruit must be cooled to the required vessel carrying temperature (or within 2°C thereof) and the cold chain must be maintained until the fruit is displayed for purchase.
Some retailers prefer fruit that are ready to eat within 1–2 days of receipt. In these situations, the ideal scenario is to ripen the product as close as possible to the retail outlet to minimize physical damage to the soft fruit. However, where end-market location and transport arrangements allow delivery to market within 3 days, ripening on-farm has advantages by reducing costs for growers, and extending ownership of the product. Transport time is a major consideration for determining optimum systems (Ledger, 2003b) (Table 15.7).
When the export dispatch facility is >1 h away from the packhouse, the following road transport recommendations apply:
The refrigerated truck should be clean and in good mechanical condi- ●
tion. The insulation and fl oor should be in a sound condition, the door seals must be intact and the doors must close very tightly.
Table 15.7. Ripening and transport recommendations for ‘Kensington Pride’ mango within Australia, to cater for the ‘ripe-for-tonight’ programme of major retail chains (Source: Ledger, 2003b).
System Aim Recommended handling conditions
System 1 – ripen at market
To deliver uniformly backward fruit to the market destination and then use ethylene to ripen fruit ready for retail sale. Temperature is managed through the chain to prevent mixed ripening and to avoid temperatures >22°C. This is the preferred system for Northern Territory and northern Western Australia growers send-ing >2000 km to market
Precool fruit to transport temperature within 12–15 h of packingTransport at 12–16°C for trips of 1–2 days and 12°C for longer tripsRipen at the market using 10 ppm ethylene for 2–3 days at 18–20°CContinue to hold fruit at 18–20°C until ready for saleStore at 10–12°C to slow ripening for a maximum of 3 days
System 2 – ripen on farm
To deliver fruit to the market destination ready for retail sale within 1–2 days. Fruit are ripened evenly using ethylene to colour stage 3 (30–50% yellow) before transport and temperature is managed through the chain to avoid high temperatures >22°C. Ripening on farm is not recommended for transport times >4 days
Precool to 18–20°C within 12–15 h of packingRipen using 10 ppm ethylene for 2–3 days at 18–20°CHold at 18–20°C until colour stage 3 (30–50% yellow)Transport at 12–16°C for trips of
1–2 days and 12°C for 3–4 day trips
Hold at market at 18–20°C until ready for saleStore at 10–12°C to slow ripening for a maximum of 3 days

Postharvest Technology 577
The refrigeration equipment must be correctly set on air delivery and ●
must be calibrated for each journey. Equipment needs to function reli-ably and receive regular servicing. Air should be delivered at the set point and fl uctuations should not exceed ±0.5°C from set point. Refriger-ated vehicles should be fi tted with temperature loggers monitoring the delivery air, and with a digital display on the outside of the box. Refrig-erated vehicles are not usually designed for, or capable of, lowering fruit temperatures so the fruit must be at the relevant shipping temperature when loading.Because of the shorter time involved, air-transported fruit may have less ●
stringent temperature requirements than sea-export fruit. Airlines carry-ing cargo may need to be consulted concerning the normal hold tem-peratures in their aircraft.Sea-export fruit should be held under refrigeration until loading. Sea ●
transport can be in refrigerated vessels, with entire refrigerated decks fi lled with pallets, or in sea containers, each of which is linked to a cen-tral ducted refrigeration system in refrigerated container vessels. Alter-natively, integral containers with their own individual cooling systems or integral CA containers may be used.
Close temperature monitoring on the vessels is essential. By monitoring delivery air temperatures (DAT) and return air temperatures (RAT), it is pos-sible to assess whether fruit is heating up due to respiration or inadequate precooling, and to take necessary steps (Anonymous, 1989; Eksteen, 1990). While most refrigerated container vessels monitor individual container air temperatures, including DAT and RAT, it is sometimes advisable to include additional temperature loggers which can measure air and fruit pulp tem-peratures for an entire journey.
The sea-freight component is generally the most time-consuming part of the whole fi eld-to-supermarket voyage (for example see Table 15.8). Essen-tial activities before and after transport can be signifi cant, for a relatively perishable product like mango. Minimizing time delays in each component of the distribution chain is important. To reduce product deterioration,
Table 15.8. Typical packing and shipping schedules for mangoes consigned by sea to the EU from South Africa.
Operation Days required
Picking and packing 1Precooling and accumulation of load 4Transport to port 2Port handling and accumulation of load 3Voyage time 17Discharge handling 1Transport and distribution 2Total 30

G.I. Johnson and P.J. Hofman578
producers and marketers should encourage training in perishable product handling and QA systems for personnel from trucking, sea-freight and air-freight companies who are responsible for loading, unloading and maintain-ing storage facilities.
Co-shipment or storage with fruit or fl owers that produce high levels of ethylene can cause unanticipated triggering of mango ripening. Co-shipment with papaya (Carica papaya) increases mango ripening (O’Hare et al., 1994). Conversely, co-shipment of carambolas (Averrhoa carambola) with mangoes caused ripening of the carambolas. The development of specialized packaging materials to eliminate extraneous ethylene may reduce the risk of unwanted ripening, although mixed transport should be avoided.
15.10 Marketing
Modern supermarket chains require large quantities of uniform produce that can be purchased on contract for delivery at a particular time to stores across a city or country. This allows the supermarket chain to promote the product at a special price. Mangoes are generally priced per fruit rather than by weight, although this is changing. Barcoding and/or Price Lookup Codes (PLU) on the labels of individual fruit for electronic checkout processing improves monitoring of purchase habits and stock control. The International Federation of Produce Standards (IFPS) (2008) provides a forum for stan-dardization of produce labelling and the PLUs are applicable internationally. Proctor and Cropley (1994) cautioned the need to ensure that label adhesives comply with food additive restrictions in the EU.
Networks and cooperatives
Marketing cooperatives or networks can assist individual producers to obtain critical mass in an industry, and fulfi l buyer expectations of large supply and seasonal spread of production (Glogoski, 1995; Griffi n, 1995; Higginbottom, 1995; M.C. Nguyen et al., 2004).
Promotion and consumer education
Mangoes are increasingly popular among affl uent consumers in the EU, North America and northern Asia. In the tropics, they are reminders of a non-urban living, which has become less common because of rapid indus-trialization and migration to the cities. Whether for domestic use or export, mangoes must compete in the fresh market with other equally attractive, nutritious, aromatic and tasty fruit. Mangoes must also increasingly compete with the snack food, beverage and entertainment industries.
Consumer education can encourage consumption and sales. Customers can be educated how to select and store mangoes and how to use both the

Postharvest Technology 579
fresh and the processed products in a variety of ways, thereby increasing total demand. Production of mango slices in take-away packs can tap domes-tic and export markets for ready-to-eat, healthy products and circumvent some disinfestation requirements (see Raymundo et al., Chapter 17, this vol-ume). Siriphanich (1994) has reviewed minimal processing of tropical fruit and noted the advantages of gaining market access and reducing transporta-tion costs.
15.11 Conclusions
Mango production has been based almost entirely on Mangifera indica, albeit a variable meld of thousands of cultivars which may be derived from inter-specifi c hybrids of a few closely related species (Kostermans and Bompard, 1993). Given its perishable nature, capitalizing more on the diversity of exist-ing germplasm to develop cultivars with superior storage traits linked to customer appeal could deliver major benefi ts.
Future improvements in postharvest technology and quarantine treat-ment will come from refi nement of preharvest management, for example reducing disease inoculum and increasing fruit resistance to disease, reduc-ing harvest costs and fruit damage, improving postharvest treatments and systems, and supply chain approaches to enhance fruit longevity and quality and reduce the risks of product damage. Improvements will also accrue from the provision of user-friendly information for supply chain personnel, but only if the information is utilized and implemented. Increases in throughput via the automation of harvesting and treatment systems for fruit will increase as production and marketing costs escalate. Labour saving and work effi -ciency will also become more critical. Innovative transport arrangements may become necessary as regional development places greater pressures on transport systems. International, collaborative joint-marketing ventures will ensure year-round supplies of uniform quality fruit, and per capita consump-tion of mangoes will increase (Johnson, 1995).
Acknowledgements
The authors acknowledge the contributions of the co-authors of Johnson et al. (1997), which this book chapter supercedes, and Leanne Taylor and Roberto Marques for assistance with references. The authors also thank the Depart-ment of Primary Industries and Fisheries for research programme support.
References
Abu-Goukh, A.B.A. and Mohamed, H.I. (2004) Effect of harvesting method on quality and shelf life of mango fruits. Tropical Science 44, 73–76.

G.I. Johnson and P.J. Hofman580
Adkins, M., Hofman, P.J. and Stubbings, B. (2002) Manipulating the Ripening of Tropical and Subtropical Fruits with 1-MCP (EthylblocR). Final Report for Project FR00029.Horticulture Australia Ltd, Sydney.
Adkins, M.F., Hofman, P.J., Stubbings, B.A. and Macnish, A.J. (2005) Manipulating avo-cado fruit ripening with 1-methylcyclopropene. Postharvest Biology and Technol-ogy 35, 33–42.
Agricultural and Processed Food Products Export Development Authority (APEDA) (2007) Ministry of Commerce and Industry, Government of India. Available at: http://www.apeda.com/apedawebsite/Announcements/GUIDELINES MANGOES TO USA.pdf (accessed 11 October 2008).
Akem, C.N. (2006) Mango anthracnose disease: present status and future research pri-orities. Plant Pathology 5, 266–273.
Alves, R.E., Filgueiras, H.A.C., Almeida, A.S., Pereira, M.E.C., Cocozza, F.M. and Jorge, J.T. (2004) Postharvest ripening of ‘Tommy Atkins’ mangoes on two maturation stages treated with 1-MCP. Acta Horticulturae 645, 627–632.
Amarante, C. and Banks, N.H. (2001) Postharvest physiology and quality of coated fruits and vegetables. Horticultural Reviews 26, 161–238.
Amesbury, K., Kernot, I. and Rudge, T. (2002) Tableland Marketing Mango Grading Guide. Available at: http://www.mangoes.net.au/images/pdf/Mango_Grade_Chart.pdf (accessed 10 March 2008).
Anesiadis, T., Karaoglanidis, G.S. and Tzavella-Klonari, K. (2003) Protective, curative and eradicant activity of the strobilurin fungicide azoxystrobin against Cercospora beticola and Erysiphe betae. Journal of Phytopathology 15, 647–651.
Anonymous (1983) Ethylene dibromide: intent to cancel registrations of pesticide prod-ucts containing ethylene dibromide; determination concluding the rebuttable pre-sumption against registration; availability of position documents. Federal Register48, 46234–46248.
Anonymous (1985) Treatments for fresh fruit and vegetables. In: Plant Quarantine Treat-ment Schedule. Australian Quarantine and Inspection Service, Canberra, pp. 21–23.
Anonymous (1986) Irradiation in the production, processing, and handling of food. Food and Drug Administration. Federal Register 51, 13376–13399.
Anonymous (1988) Conditions for importation of fruit and vegetables. In: Plant Quaran-tine Manual, section E. Australian Quarantine Inspection Service, Canberra, pp. 1–59.
Anonymous (1989) Guide to Food Transport. Fruit and Vegetables. Mercantila, Copen-hagen, pp. 162–163.
Anonymous (1992) New Zealand Ministry of Agriculture and Fisheries Treatment Re-quirements, NASS: 152.02, Border Protection Specifi cations: Clearance of Fresh Produce, Section 2.0, TB. Wellington, North Island, New Zealand.
Anonymous (1993) International Standardisation of Fruit and Vegetables: Mangoes. Or-ganisation for Economic Cooperation and Development. Available at http://ingen-taconnect.com/content/oecd/16080149/1993/00001993/00000001/5193033e
Anonymous (1994a) Plant Pest and Quarantine Treatment Manual. United States De-partment of Agriculture (USDA), Animal and Plant Health Inspection Service, Hyattsville, Maryland, USA.
Anonymous (1994b) Mango Farmer/Packhouse/Exporter Guidelines. South African Mango Growers’ Association, Tzaneen, South Africa.
Arafat, L.A.E.T. (2005) Chilling injury in mangoes. PhD thesis, Wageningen University, Wageningen, the Netherlands.
Arpaia, M.L. (1994) Preharvest factors infl uencing postharvest quality of tropical and subtropical fruit. HortScience 29, 982–985.
Askar, A. and Treptow, H. (1993) Quality Assurance in Tropical Fruit Processing. Springer-Verlag, Berlin.

Postharvest Technology 581
ASEAN_GAP (2007) Association of South-east Asian Nations (ASEAN) Good Agricul-tural Practice. Standards and Manuals. Available at: http://www.aphnet.org/gap/ASEANgap.html (accessed 31 October 2007).
Astridge, D. and Baron, Z. (2007a) Fruit spotting bug in mangoes. In: Delivering Mango Research. Proceedings of the Sixth Australian Mango Conference. Gold Coast, Queensland, Australia, pp. 15–16.
Astridge, D. and Baron, Z. (2007b) IPM in mangoes. In: Delivering Mango Research. Proceedings of the Sixth Australian Mango Conference. Gold Coast, Queensland, Australia, pp. 14–15.
Astridge, D. and Baron, Z. (2007c) Mango pulp weevil in mangoes of the Philippines. (Abstract). In: Delivering Mango Research. Proceedings of the Sixth Australian Mango Conference. Gold Coast, Queensland, Australia, p. 17.
Australian Quarantine and Inspection Service (AQIS) (2008a) AQIS Import Conditions Database. Search ‘mango’. Available at: http://www.aqis.gov.au/icon32/asp/ex_querycontent.asp (accessed 3 April 2008).
Australian Quarantine and Inspection Service (AQIS) (2008b) Phyto search. Available at: http://www.aqis.gov.au/phyto/asp/ex_search.asp (accessed 7 April 2008).
Bagshaw, B. and Brown, B. (1989) Disorders. In: Ridgeway, R. (ed.) Mango Pests and Disorders. Queensland Department of Primary Industries, Brisbane, Queensland, Australia, pp. 22–28.
Baker, I.W. (1986) Mango maturity investigations. In: Proceedings of the First Australian Mango Research Workshop. Commonwealth Scientifi c and Industrial Research Or-ganization (CSIRO), Melbourne, pp. 271–273.
Baldwin, E.A., Burns, J.K., Kazokas, W., Brecht, J.K., Hagenmaier, R.D., Bender, R.J. and Pesis, E. (1999) Effect of two edible coatings with different permeability character-istics on mango (Mangifera indica L.) ripening during storage. Postharvest Biology and Technology 17, 215–226.
Bally, I.S.E. (2007) The effect of preharvest nutrition and crop load on fruit quality and postharvest disease in mango (Mangifera indica L.). PhD thesis, University of Queensland, Brisbane, Queensland, Australia.
Bally, I.S.E., O’Hare, T.J. and Holmes, R.J. (1997) Detrimental effects of detergent in the development of mango skin browning. Acta Horticulturae 455, 612–621.
Bandyopadhyay, C., Gholap, A.S. and Mamdapur, V.R. (1985) Characterisation of alke-nyl resorcinol in mango (Mangifera indica L.) latex. Journal of Agricultural and Food Chemistry 33, 377–379.
Biosecurity New Zealand (BANZ) (2008) Import Health Standards. Available at: http://www.biosecurity.govt.nz/commercial-imports/import-health-standards/search (ac-cessed 7 April 2008).
Barba, R.C. (1974) Induction of fl owering of the mango by chemical spray. In: Proceed-ings of the Fifth Crop Science Society of the Philippines Meeting. pp. 154–160.
Bartley, J.P. and Schwede, A. (1987) Volatile fl avour components in the headspace of the Australian ‘Bowen’ mango. Journal of Food Science 52, 353–360.
Bender, R.J., Brecht, J.K., Sargent, S.A. and Huber, D.J. (2000) Mango tolerance to re-duced oxygen levels in controlled atmosphere storage. Journal of the American Society for Horticultural Science 125, 707–713.
Bezuidenhout, J.L.J., Robbertse, P.J. and Kaiser, C. (2005) Anatomical investigation of lenticel development and subsequent discolouration of ‘Tommy Atkins’ and ‘Keitt’ mango (Mangifera indica L.) fruit. Journal of Horticultural Science and Biotechnology 80, 18–22.
Blankenship, S.M. and Dole, J.M. (2003) 1-Methylcyclopropene: a review. Postharvest Biology and Technology 28, 1–25.
Boelema, T. (1987) Long-distance transport of avocados. South African Avocado Grow-ers’ Association Yearbook 10, 153–156.

G.I. Johnson and P.J. Hofman582
Bondad, N.D. and Linsagan, E. (1979) Flowering in mango induced with potassium ni-trate. HortScience 14, 527–528.
Bower, J.P., Dennison, M.T. and Fowler, K. (2003) Avocado and mango cold storage damage as related to water loss control. Acta Horticulturae 628, 401–406.
Brady, C. (1994) Biochemical and molecular approaches to fruit ripening and senes-cence. In: Champ, B.R., Highley, E. and Johnson, G.I. (eds) Postharvest Handling of Tropical Fruits. Australian Centre for International Agricultural Research (ACIAR) Proceedings 50. ACIAR, Canberra, pp. 205–217.
Brecht, J.K., Chau, K.V., Fonseca, S.C., Oliveira, F.A.R., Silva, F.M., Nunes, M.C.N. and Bender, R.J. (2003) Maintaining optimal atmosphere conditions for fruits and veg-etables throughout the postharvest handling chain. Postharvest Biology and Tech-nology 27, 87–101.
Brown, B.I., Wells, I.A. and Murray, C.F. (1986) Factors affecting the incidence and se-verity of mango sapburn and its control. ASEAN Food Joumal 2, 127–132.
Buchanan, A. (1994) Tropical fruits: the social political and economic issues. In: Champ, B.R., Highley, E. and Johnson, G.I. (eds) Postharvest Handling of Tropical Fruits. Australian Centre for International Agricultural Research (ACIAR) Proceedings 50. ACIAR, Canberra, pp. 18–26.
Bunt, C. and Piccone, M. (1994) Quality assurance: a total approach. In: Champ, B.R., Highley, E. and Johnson, G.I. (eds) Postharvest Handling of Tropical Fruits. Austra-lian Centre for International Agricultural Research (ACIAR) Proceedings 50. ACIAR, Canberra, pp. 56–64.
Burdon, J.N. and Moore, K.G. (1991) The soft-nose disorder of mango fruit. Planter 67, 477–481.
Bustos, M.E., Enkerlin, H.W., Toledo, A.J., Reyes, F.J. and Casimiro, G.A. (1992) Irradia-tion of mangoes as a quarantine treatment. In: Use of Irradiation as a Quarantine Treatment of Food and Agricultural Commodities. Proceedings of the Final Research Coordination Meeting, International Atomic Energy Agency (IAEA), Vienna, Austria.
Bustos, M.E., Enkerlin, W., Reyes, J. and Toledo, J. (2004) Irradiation of mangoes as a postharvest quarantine treatment for fruit fl ies (Diptera: Tephritidae). Journal of Eco-nomic Entomology 97, 286–292.
Castro, J.V., Pfaffenbach, L.B., Carvalho, C.R.L. and Rossetto, C.J. (2005) Effects of fi lm packaging and cold storage on postharvest quality of ‘Tommy Atkins’ mangoes. Acta Horticulturae 682, 1683–1687.
Catindig, J.L.A. and Kong, L.H. (2007) Asia Pacifi c Alien Species Database. Mango Pulp Weevil. Available at: http://apasd-niaes.dc.affrc.go.jp/ (accessed 31 October 2007).
Chalutz, E., Droby, S., Wilson, C.L. and Wisniewski, M.E. (1992) DV-induced resistance to postharvest diseases of citrus fruit. Journal of Phytochemistry and Photobiology 15, 367–374.
Chaplin, G.R., Nuevo, P.A., Graham, D. and Cole, S.P. (1986a) Chilling responses of ‘Kensington’ mango fruit stored under variable low temperature regimes. ASEANFood Journal 2, 133–138.
Chaplin, G.R., Graham, D. and Cole, S.P. (1986b) Reduction of chilling injury in mango fruit by storage in polyethylene bags. ASEAN Food Journal 2, 139–142.
Chaplin, G.R., Cole, S.P., Landragan, M., Nuevo, P.A., Lam, P.F. and Graham, D. (1991a) Chilling injury and storage of mango (Mangifera indica L.) fruit under low tempera-tures. Acta Horticulturae 291, 461–471.
Chaplin, G.R., McBride, R.L., Abdullah, A. and Nuevo, P.A. (1991b) Sensory and physi-cochemical quality of ‘Kensington’ mangoes after storage at low temperature. ASE-AN Food Journal 6, 109–113.

Postharvest Technology 583
Chin, D., Brown, H. and Thistleton, B. (2007) Mango pulp weevil in mangoes of the Philippines. In: Delivering Mango Research. Proceedings of the Sixth Australian Mango Conference. Gold Coast, Queensland, Australia, pp. 18–23.
Chitarra, A.B., Chitarra, M.I.F. and Evangelista, R.M. (2001) Biochemical changes in mango fruits, ‘Tommy Atkins’ treated with calcium chloride preharvest and stored under refrigeration. Acta Horticulturae 553, 79–81.
Cipollini, M.L. and Stiles, E.W. (1992) Relative risks of microbial rot for fl eshy fruits: signifi cance with respect to dispersal and selection for secondary defence. Advanc-es in Ecological Research 23, 35–91.
Coates, L.M. and Johnson, G.I. (1993) Effective disease control in heat-disinfested fruit. Postharvest News and Information 4, 35N–40N.
Coates, L.M., Johnson, G.I. and Cooke, A.W. (1993) Postharvest disease control in man-goes using high humidity hot air and fungicide treatments. Annals of Applied Biol-ogy 123, 441–448.
Coates, L.M., Davis, R.D. and Cooke, A.W. (1995) Postharvest disease control in mango – potential options for reducing chemical use. In: Proceedings of Mango 2000 Pro-duction Workshop, Townsville.
CODEX (2008a) CODEX Alimentarius Commission. Available at: http://www.codexali-mentarius.net/web/index_en.jsp (accessed 21 March 2008).
CODEX (2008b) Codex Standards List, 2008. CODEX Alimentarius – Current Offi cial Standards. Available at: http://www.codexalimentarius.net/web/standard_list.do?lang=en (accessed 21 March 2008).
CODEX-MRL (2008) Limits for Prochloraz – Assorted Tropical and Sub-tropical Fruits – Inedible Peel. Database Listings of Maximum Residue Limits; Extraneous Maximum Residue Limits. Offi cial Standards – Pesticide Residues in Food Searchable Data-base. World Health Organization (WHO)/Food and Agriculture Organization (FAO) Food Standards CODEX Alimentarius. Available at: http://www.codexalimentarius.net/mrls/pestdes/jsp/pest_q-e.jsp (accessed 4 April 2008).
Coetzer, L.A., Robbertse, P.J. and De Wet, E. (1991) Die invloed van boortoedienings op vrugproduksie en verkoelde opberging van mangos. South African Mango Growers’ Association Yearbook 11, 29–31.
Cojocaru, M., Droby, S., Glotter, E., Goldman, H., Gottlieb, H., Jacoby, B. and Prusky, D. (1985) 5-(12-heptadecerlyl)-resorcinol, the major component of the antifungal agent from the peel of mango fruit. Phytochemistry 25, 1093–1095.
Condé, B.D. and Pitkethly, R.N. (2007) Mango scab in Australia. In: Delivering Mango Research. Proceedings of the Sixth Australian Mango Conference. Gold Coast, Queensland, Australia, pp. 8–10.
Cooke, A.W. and Johnson, G.I. (1994) Mango postharvest disease control: effect of rain at harvest, fungicide treatments and fruit brushing on fruit appearance. In: Champ, B.R., Highley, E. and Johnson, G.I. (eds) Postharvest Handling of Tropical Fruits. Australian Centre for International Agricultural Research (ACIAR) Proceedings 50. ACIAR, Canberra, pp. 448–449.
Crous, P.W., Slippers, B., Wingfi eld, M.J., Rheeder, J., Marasas, W.F.O., Philips, A.J.L., Alves, A., Burgess, T., Barber, P. and Groenewald, J.Z. (2006) Phylogenetic lineages in the Botryosphaeriaceae. Studies in Mycology 55, 235–253.
Cunningham, I.C. (1986) Mango insect pests. In: Proceedings of the First Australian Mango Research Workshop. Commonwealth Scientifi c and Industrial Research Organization (CSIRO), Melbourne, pp. 221–224.
Cunningham, I.C. (1991a) Management of mango insect pests. Acta Horticulturae 291,379–388.
Cunningham, I.C. (1991b) Common mango scales in Queensland. Acta Horticulturae 291, 409–412.

G.I. Johnson and P.J. Hofman584
Cunningham, I.C. (1991c) Mango seed weevil in Queensland. Acta Horticulturae 291,413–417.
Dann, E., Zainuri, Hassan, K., Irving, D.E., Cooke, A.W. and Coates, L. (2005) Self-defence for mangoes. Mango Matters Autumn/Winter issue, 35–36.
Dann, E., Coates, L., Cooke, A.W., Smith, L. and Dean, J. (2007) Novel methods for integrated control of mango postharvest diseases. In: Delivering Mango Research. Proceedings of the Sixth Australian Mango Conference. Gold Coast, Queensland, Australia, pp. 5–7.
Davenport, T.L. and Núñez-Elisea, R. (1997) Reproductive physiology. In: Litz, R.E. (ed.) The Mango: Botany, Production and Uses. CAB International, Wallingford, UK, pp. 69–146.
DeEll, J.R. and Toivonen, P.M.A. (eds) (2003a) Practical Applications of Chlorophyll Fluo-rescence in Plant Biology. Kluwer Academic Publishers, Dordrecht, the Nether-lands, 280 pp.
DeEll, J.R. and Toivonen, P.M.A. (2003b) Use of chlorophyll fl uorescence in postharvest quality assessments of fruits and vegetables. In: DeEll, J.R. and Toivonen, P.M.A. (eds) Practical Applications of Chlorophyll Fluorescence in Plant Biology. Kluwer Academic Publishers, Dordrecht, the Netherlands, pp. 203–242.
de Jesus, L.R.A., Calumpang, S.M.F., Medina, J.R. and Ohsawa, K. (2007) Floral volatiles of Mangifera indica L. (cv. Carabao) attractive to Sternochetus frigidus (Fabr.) (Coleoptera: Curculionidae). The Philippine Agricultural Scientist 86, 282–289.
DeLong, J.M., Prange, R.K. and Harrison, P.A. (2007) Chlorophyll fl uorescence-based low-O2 CA storage of organic ‘Cortland’ and ‘Delicious’ apples. Acta Horti-culturae 737, 31–37.
Diedhiou, P.M., Mbaye, N., Drame, A. and Samb, P.I. (2007) Alteration of postharvest diseases of mango (Mangifera indica) through production practices and climatic factors. African Journal of Biotechnology 6, 1087–1094.
Dodd, J.C., Estrada, A.B., Matcham, J., Jeffries, P. and Jeger, M.J. (1991a) The effect of climatic factors on Colletotrichum gloeosporioides, the causal agent of mango an-thracnose, in the Philippines. Plant Pathology 40, 568–575.
Dodd, J.C., Bugante, R., Koomen, I., Jefries, P. and Jeger, M.J. (1991b) Pre- and post-harvest control of mango anthracnose in the Philippines. Plant Pathology 40, 576–583.
Dodd, J.C., Estrada, A. and Jeger, M.J. (1992) Epidemiology of Colletotrichum gloeospo-rioides in the tropics. In: Bailey, J.A. and Jeger, M.J. (eds) Colletotrichum: Biology, Pathology and Control. CAB International, Wallingford, UK, pp. 308–325.
Dodd, J.C., Prusky, D. and Jeffries, P. (1997) Fruit diseases. In: Litz, R.E. (ed.) The Mango: Botany, Production and Uses. CAB International, Wallingford, UK, pp. 257–280.
Droby, S., Prusky, D., Jacoby, B. and Goldman, A. (1986) The presence of an antifungal compound and its relation to the latency of Alternaria alternata in unripe peels of mango fruits. Physiological Plant Pathology 29, 173–183.
Droby, S., Prusky, D., Jacoby, B. and Goldman, A. (1987) Induction of antifungal resor-cinols in fl esh of unripe mango fruits and its relation to latent infection by Alter-naria alternata. Physiological and Molecular Plant Pathology 30, 285–292.
Ducamp Collin, M.-N., Arnaud, C., Kagy, V. and Didier, C. (2007) Fruit fl ies: disinfesta-tion, techniques used, possible application to mango. Fruits 62, 223–236.
du Plooy, G.W., van der Merwe, C. and Korsten, L. (2004) Differences in the surface structures of three mango cultivars and the effect of kaolin on these structures. South African Mango Growers’ Association Yearbook 24, 29–36.
du Plooy, G.W., van der Merwe, C.F. and Korsten, L. (2006) Lenticel discolouration in mango (Mangifera indica L.) fruit – a cytological study of mesophyll cells from af-fected tissue. Journal of Horticultural Science and Biotechnology 81, 869–873.
Eckert, J.W. (1983) Control of postharvest diseases with antimicrobial agents. In: Diseases of Fruits and Vegetables. North Atlantic Treaty Organization (NATO) Advisory

Postharvest Technology 585
Study Institute Series, Series A – Life Sciences. Available at http://www.fao.org/ agris/search/display.do?f=./1984/v1002/US8268297.xml; US8268297, New York, pp. 265–285.
Eksteen, G.J. (1990) Temperature changes during sea shipment of mangoes in ducted containers. South African Mango Growers’ Association Yearbook 10, 26–27.
El Ghaouth, A. and Wilson, C.L. (1995) Biologically based technologies for the control of postharvest diseases. Postharvest News Information 6, 5N–11N.
El Ghaouth, A., Arul, J., Asselin, A. and Benhamou, N. (1992a) Antifungal activity of chitosan on postharvest pathogens: induction of morphological and cytological alterations in Rhizopus stolonifer. Mycological Research 96, 769–779.
El Ghaouth, A.E., Ponnampalam, R., Castaigne, F. and Arul, J. (1992b) Chitosan coating to extend the storage life of tomatoes. HortScience 27, 1016–1018.
Environmental Protection Agency (EPA) (2002) Irradiation Phytosanitary Treatment of Imported Fruits and Vegetables Federal Register (Volume 67, Number 205). Rules and Regulations: 65016–65029. Available at: http://www.wais.access.gpo.gov (accessed 23 October 2008).
Environmental Protection Agency (EPA) (2008) The Phaseout of Methyl Bromide. Avail-able at: http://www.epa.gov/ozone/mbr/ (accessed 3 April 2008).
Erwin, D.C. (1973) Systemic fungicides: disease control, translocation and mode of action. Annual Review of Phytopathology 11, 389–400.
Esguera, E.B. and Lizada, M.C.C. (1990) The postharvest behaviour and quality of ‘Cara-bao’ mangoes subjected to vapour heat treatment. ASEAN Food Journal 5, 6–12.
Esguerra, E.B., Brena, S.R., Reyes, M.U. and Lizada, M.C.C. (1990) Physiological break-down in vapor heat-treated ‘Carabao’ mango. Acta Horticulturae 269, 425–434.
Estrada, A.B., Dodd, J.C. and Jeffries, P. (1993) Effect of environment on the in vitro growth and development of Colletotrichum gloeosporioides isolates from the Phil-ippines. Acta Horticulturae 341, 360–370.
EurepGAP (2007) What is EurepGAP? Available at: http://www.eurepgap.org/Languages/English/about.html (accessed 31 October 2007).
Eustice, R.F. (2007) Recent Developments in Food Irradiation: a Global Review. Min-nesota Beef Council. Available at: http://www.mnbeef.org/New%20Site/HTML/irradiationSub/2RecentDevelopments.htm (accessed 31 October 2007).
Everett, K.R., Owen, S.G. and Cutting, J.G.M. (2005) Testing effi cacy of fungicides against postharvest pathogens of avocado (Persea Americana cv. Hass). New Zea-land Journal of Plant Protection 58, 89–95.
Fallik, E. and Grinberg, S. (1992) Hinokitiol: a natural substance that controls postharvest dis-eases in eggplant and pepper fruits. Postharvest Biology and Technology 2, 137–144.
FAOSTAT (2008) FAOSTAT On-line. Rome. United Nations Food and Agriculture Orga-nization. Available at: http://faostat.fao.org/default.aspx?lang=en (accessed 21 March 2008).
Feygenberg, O., Hershkovitz, V., Ben-Arie, R., Jacob, S., Pesis, E. and Nikitenko, T. (2005) Postharvest use of organic coating for maintaining bio-organic avocado and mango quality. Acta Horticulturae 682, 1057–1061.
FFTC-GAP (2007) Good Agricultural Practice in Asia and Oceana. Available at http://agnet.org/library/ubk/54/
Fitzell, R.D. and Peak, C.M. (1984) The epidemiology of anthracnose disease in mango: inoculum sources, spore production and dispersal. Annals of Applied Biology 104,53–59.
Fitzell, R.D., Peak, C.M. and Darnell, R.E. (1984) A model for estimating infection levels of anthracnose disease of mango. Annals of Applied Biology 104, 451–458.
Follett, P.A. (2004) Irradiation quarantine treatments for mango seed weevil and Cryp-tophlebia spp. In: Irradiation as a Phytosanitary Treatment of Food and Agricultural

G.I. Johnson and P.J. Hofman586
Commodities. IAEA-TECDOC-1427. International Atomic Energy Agency (IAEA), Vienna, Austria, pp. 9–18.
Follett, P.A. and Gabbard, Z. (2000) Effect of mango weevil (Coleoptera: Curculionidae) damage on mango seed viability in Hawaii. Journal of Economic Entomology 93, 1237–1240.
Follett, P.A. and Lower, R.A. (2000) Irradiation to ensure quarantine security for Cryp-tophlebia spp. (Lepidoptera: Tortricidae) in Sapindacious fruits from Hawaii. Jour-nal of Economic Entomology 93, 1848–1854.
Follett, P.A. and Neven, L.G. (2005) Current trends in quarantine entomology. AnnualReview of Entomology 51, 359–385.
Follett, P.A., Yang, M., Lu, K. and Chen, T. (2007) Irradiation for postharvest control of quarantine pests. Formosan Entomology 27, 1–15.
Fonseca, M.J.O., Cecon, P.R., Salomão, L.C.C. and Pushmann, R. (2004a) Pulp and skin pigments in mango ‘Haden’ treated with fungicides and wax. Acta Horticulturae645, 633–638.
Fonseca, M.J.O., Salomão, L.C.C., Cecon, P.R. and Puschmann, R. (2004b) Fungicides and wax in postharvest preservation of mango ‘Haden’. Acta Horticulturae 645, 557–563.
Food and Fertilizer Technology Centre (FFTC) (2007) Good Agricultural Practice in Asia and Oceana. Available at: http://www.agnet.org/library/vbk/54/ (accessed 7 Octo-ber 2007).
Gaffney, J.J. and Armstrong, J.W. (1990) High-temperature forced-air research facility for heating fruits for insect quarantine treatments. Journal of Economic Entomology 83,1959–1964.
Good Agricultural Practice (GAP) Small Producers in Export Horticulture: a Guide to Best Practice. Natural Resources Institute. Available at htto://nri.org/NRET/
GLOBALGAP (2007) Global GAP. Available at: htttp://www.globalgap.org/cms/front_content.php?idart=3&idcat=9&lang=1 (accessed 31 October 2007).
Glogoski, A. (1995) Group marketing – redressing the balance of power. In: Smarter Marketing, Better Profi ts. Proceedings of the Second Annual Queensland Fruit and Vegetable Growers’ Conference, Brisbane, Queensland, pp. 1–6.
Goguey, T. (1993) Study of the effects of three fl ower-inducing substances on ‘Ken’ and ‘Zill’ mango. Acta Horticulturae 341, 216–224.
González-Aguilar, G.A., Fortiz, J., Cruz, R., Baez, R. and Wang, C.Y. (2000) Methyl jas-monate reduces chilling injury and maintains postharvest quality of mango fruit. Journal of Agricultural and Food Chemistry 48, 515–519.
Good Agricultural Practice (GAP) (2003) Small Producers in Export Horticulture. A Guide to Best Practice. Agricultural and Environmental Practices: Good Agricul-tural Practice. Natural Resources Institute. Available at: http://www.nri.org/NRET/SPCDR/Chapter4/agriculture-4-4.htm (accessed 7 October 2007).
Gould, W.P. and Sharp, J.L. (1990) Caribbean fruit fl y (Diptera: Tephritidae) mortality induced by shrink-wrapping infested mangoes. Journal of Economic Entomology 83, 2324–2326.
Griffi n, D. (1995) Grower networking to achieve international competitiveness. In: Smarter Marketing, Better Profi ts. Proceedings of the Second Annual Queensland Fruit and Vegetable Growers Conference, Brisbane, Queensland, pp. 7–16.
Grové, T., Steyn, W.P. and de Beer, M.S. (1997) Determining suitable non-damaging treatments for post-harvest disinfestation of fruit fl y in mango. South African Mango Growers’ Association Yearbook 17, 119–125.
GPOAccess (2008) Code of Federal Regulations. Available at: http://gpoaccess.gov/cfr/index.html (accessed 10 March 2008).

Postharvest Technology 587
Gullino, M.L. and Kuijpers, L.A. (1994) Social and political implications of managing plant diseases with restricted fungicides in Europe. Annual Review of Phytopathol-ogy 32, 559–579.
Hall, A., Bockett, G., Taylor, S., Sivamohan, M.K.V. and Clark, N. (2001) Why research partnerships really matter: innovation theory, institutional arrangements and im-plications for developing new technology for the poor. World Development 29, 783–797.
Han, J., Tian, S., Meng, X. and Ding, Z. (2006) Response of physiologic metabolism and cell structures in mango fruit to exogenous methyl salicylate under low-temperature stress. Physiologia Plantarum 128, 125–133.
Hansen, J.D. (1991) Mango weevil biology, fi eld control and postharvest technology: a review. International Journal of Tropical Agriculture 9, 234–244.
Hansen, J.D. (1993) Dynamics and control of the mango seed weevil. Acta Horticultu-rae 341, 415–420.
Hardy, S., Fallow, K. and Barkley, P. (2004) Using Copper Sprays to Control Diseases in Citrus. Citrus Fact Sheet, New South Wales Department of Primary Industries. Avail-able at: http://www.dpi.nsw.gov.au/__data/assets/pdf_fi le/0003/138171/copper-spray-citrus.pdf (accessed 3 January 2008).
HarvestWatch (2008) HarvestWatch™ – Monitoring Produce Health. Availablet at: http://www.fl intbox.com/technology.asp?page=2582 (accessed 28 February 2008).
Harvey, E.C. (1987) Maturity indices for quality control and harvest maturity. In: Prins-ley, R.T. and Tucker, G. (eds) Mangoes – a Review. The Commonwealth Secretariat, London, pp. 39–55.
Hassan, M.K. (2007) Constitutive alk(en)ylresorcinols and resistance to postharvest dis-ease in mango (Mangifera indica L.). PhD thesis, University of Queensland, Bris-bane, Queensland.
Hassan, M.K., Dann, E.K., Irving, D.E. and Coates, L.M. (2007) Concentrations of constitu-tive alk(en)ylresorcinols in peel of commercial mango varieties and resistance to post-harvest anthracnose. Physiological and Molecular Plant Pathology 17, 158–165.
Heather, N.W. (1994) Quarantine disinfestation of tropical fruits: non-chemical options. In: Champ, B.R., Highley, E. and Johnson, G.I. (eds) Postharvest Handling of Tropi-cal Fruits. Australian Centre for International Agricultural Research (ACIAR) Pro-ceedings 50. ACIAR, Canberra, pp. 272–279.
Heather, N. (2004) Generalized quarantine disinfestation research protocol. In: Irradiation as a Phytosanitary Treatment of Food and Agricultural Commodities. IAEA-TECDOC-1427. International Atomic Energy Agency (IAEA), Vienna, Austria, pp. 171–178.
Heather, N.W., Corcoran, R.J. and Banos, C. (1991) Disinfestation of mangoes with gamma irradiation against two Australian fruit fl ies (Diptera: Tephritidae). Journal of Economic Entomology 84, 1304–1307.
Hegnauer, R. (1994) Phytochemistry and chemotaxonomy of the Anacardiaceae with special emphasis on Mangifera. In: Kostermans, A.J.G.H. and Bompard, J.M. (1993) The Mangoes, their Botany, Nomenclature, Horticulture and Utilisation. AcademicPress, London, pp. 11–16.
Hewett, E. (2006) An overview of preharvest factors infl uencing postharvest quality of horticultural products. International Journal of Postharvest Technology and Innovation1, 4–15.
Higginbottom, D. (1995) Group marketing – the future. In: Smarter Marketing, Better Profi ts. Proceedings of the Second Annual Queensland Fruit and Vegetable Grow-ers Conference, Brisbane, Queensland, pp. 17–22.
Hilton, D.J. (1994) Impact and vibration damage to fruit during handling and transporta-tion. In: Champ, B.R., Highley, E. and Johnson, G.I. (eds) Postharvest Handling of

G.I. Johnson and P.J. Hofman588
Tropical Fruits. Australian Centre for International Agricultural Research (ACIAR) Proceedings 50. ACIAR, Canberra, pp. 116–126.
Hoa, T.T. and Ducamp, M.-N. (2008) Effects of different coatings on biochemical changes of ‘Cat Hoa Loc’ mangoes in storage. Postharvest Biology and Technology 48, 150–152.
Hofman, P.J. (1997) What causes green, ripe mangoes. Mango Care 20, 13–15.Hofman, P.J. (1998) Production factors infl uence fruit quality and response to postharvest
treatments. In: Coates, L.M., Hofman, P.J. and Johnson, G.I. (eds) Disease Control and Storage Life Extension in Fruit. Australian Centre for International Agricultural Research (ACIAR), Canberra, pp. 6–18.
Hofman, P.J. and Ledger, S.N. (2006) Using a supply chain approach to guide R&D. Acta Horticulturae 699, 219–226.
Hofman, P.J. and Mullen, C. (2005) The Role of Rootstocks and Nutrition in the Quality of Hass Avocado. Final Report of Project AV00013. Horticulture Australia Ltd, Sydney.
Hofman, P.J. and Smith, L.G. (1994) Preharvest effects on postharvest quality of sub-tropical and tropical fruit. In: Champ, B.R., Highley, E. and Johnson, G.I. (eds) Postharvest Handling of Tropical Fruits. Australian Centre for International Agricul-tural Research (ACIAR) Proceedings 50. ACIAR, Canberra, pp. 261–268.
Hofman, P.J., Smith, L.G. and Meiburg, G.F. (1995) Effect of position on the tree and time of fl owering on fruit quality in mango. In: Australasian Postharvest Horticul-ture Conference: Science and Technology for the Fresh Food Revolution, Mel-bourne p. 38.
Hofman, P.J., Smith, L.G., Joyce, D.C., Johnson, G.I. and Meiburg, G.F. (1997a) Bagging of mango (Mangifera indica cv. Keitt) fruit infl uences fruit quality and mineral com-position. Postharvest Biology and Technology 12, 83–91.
Hofman, P.J., Smith, L.G., Meiburg, G.F. and Giles, J.E. (1997b) Production locality affects mango fruit quality. Australian Journal of Experimental Agriculture 37, 801–808.
Hofman, P.J., Smith, L.G., Meiburg, G.F., Holmes, R. and Barker, J.A. (1998) Mango fruit quality is affected by production conditions. In: Coates, L.M., Hofman, P.J. and Johnson, G.I. (eds) Disease Control and Storage Life Extension in Fruit. Australian Centre for International Agricultural Research (ACIAR) Proceedings 81. ACIAR, Canberra, pp. 31–39.
Hofman, P.J., Jobin-Décor, M., Meiburg, G.F., Macnish, A.J. and Joyce, D.C. (2001) Rip-ening and quality responses of avocado, custard apple, mango and papaya fruit to 1-methylcyclopropene. Australian Journal of Experimental Agriculture 41, 567–572.
Holmes, R. (2003) Checklist for sapburn. Mango Care 39, 10–11.Holmes, R. and Bally, I. (1994) Harvest aid update. Mango Care 11, 10–11.Holmes, R.J., Ledger, S.N. and Macleod, W.N.B. (1993) Handling systems to reduce
sapburn. Acta Horticulturae 341, 528–532. Horn, F. and Horn, P. (2004) Fresh fruit fumigation with phosphine as alternative for
methyl bromide. Paper 58. In: Proceedings 2004 of the Annual International Re-search Conference on Methyl Bromide Alternatives and Emissions Reductions.Available at: http://mbao.org/2004/Proceedings04/mbrpro04.html (accessed 1 May 2008).
International Federation for Produce Standards (IFPS) (2008) Available at: http://www.plucodes.com/ (accessed 7 April 2008).
International Plant Protection Convention (IPPC) (2008) International Standards for Phy-tosanitary Measures (ISPMs). International Phytosanitary Portal. Available at: https://www.ippc.int/IPP/En/default.jsp (accessed 2 April 2008).
International Standardisation Organisation (ISO) (2008) Available at: http://www.iso.org/iso/home.htm (accessed 2 April 2008).

Postharvest Technology 589
International Standards for Phytosanitary Measures (ISPM) (2003) Guidelines for the use of irradiation as a phytosanitary measure. In: International Standards for Phytosani-tary Measures Secretariat of the International Plant Protection Convention, 2006.ISPM No. 18. Secretariat of the International Plant Protection Convention, Food and Agriculture Organization of the United Nations, Rome, Italy, pp. 209–220.
International Standards for Phytosanitary Measures (ISPM) (2007a) International Stan-dards for Phytosanitary Measures 1 to 29. Available at: https://www.ippc.int (accessed 31 October 2007).
International Standards for Phytosanitary Measures (ISPM) (2007b) Phytosanitary Treat-ments for Regulated Pests. ISPM No. 28. Available at: https://www.ippc.int (accessed 31 October 2007).
International Standards for Phytosanitary Measures (ISPM) (2008) Irradiation Treatment for Anastrepha ludens; A. serpentina and Others. Annex to ISPM No. 28. Available at: https://www.ippc.int (accessed 3 April 2008).
Jacobi, K.K. and Giles, J.E. (1997) Quality of ‘Kensington’ mango (Mangifera indica Linn.) fruit following combined vapour heat disinfestation and hot water disease control treatments. Postharvest Biology and Technology 12, 285–292.
Jacobi, K.K. and Gowanlock, D. (1995) Ultrastructural studies of ‘Kensington’ mango (Mangifera indica Linn.) heat injuries. HortScience 30, 102–103.
Jacobi, K.K. and Wong, I.S. (1992) Quality of ‘Kensington’ mango (Mangifera indica Linn.) following hot water and vapour heat treatments. Postharvest Biology and Technology 1, 349–359.
Jacobi, K.K., Coates, L.M. and Wong, L.S. (1994) Heat disinfestation of mangoes: effect on fruit quality and disease control. In: Champ, B.R., Highley, E. and Johnson, G.I. (eds) Postharvest Handling of Tropical Fruits. Australian Centre for International Agricultural Research (ACIAR) Proceedings 50. ACIAR, Canberra, pp. 280–287.
Jacobi, K.K., Wong, L.S. and Giles, J.E. (1995) Effect of fruit maturity on quality and physiology of high-humidity hot air-treated ‘Kensington’ mango (Mangifera indica Linn.). Postharvest Biology and Technology 5, 149–159.
Jacobi, K.K., MacRae, E.A. and Hetherington, S.E. (2001a) Loss of heat tolerance in ‘Kensington’ mango fruit following heat treatments. Postharvest Biology and Tech-nology 21, 321–330.
Jacobi, K.K., MacRae, E.A. and Hetherington, S.E. (2001b) Postharvest heat disinfesta-tion treatments of mango fruit. Scientia Horticulturae 89, 171–193.
Jeffries, P. and Koomen, I. (1992) Strategies and prospects for biological control of dis-eases caused by Colletotrichum. In: Bailey, J.A. and Jeger, M.J. (eds) Colletotri-chum: Biology, Pathology and Control. CAB International, WalIingford, UK, pp. 337–357.
Jessup, A.J., Dominiak, B., Woods, B., de Lima, C.P.F., Tomkins, A. and Smallridge, C.J. (2007) Area-wide management of fruit fl ies in Australia. In: Vreysen, M.J.B., Robin-son, A.S. and Hendrichs, A. (eds) Area-wide Control of Insect Pests – from Research to Field Implementation. Springer, Dordrecht, the Netherlands, pp. 685–697.
Jiang, Y. and Joyce, D.C. (2000) Effects of 1-methylcyclopropene alone and in combina-tion with polyethylene bags on the postharvest life of mango fruit. Annals of Applied Biology 137, 321–327.
Joel, D.M. (1978) The secretory ducts of mango fruits: a defence system effective against the Mediterranean fruit fl y. Israel Journal of Botany 1, 44–45.
Joel, D.M. (1980) Resin ducts in the mango fruit: a defence system. Journal of Experi-mental Botany 31, 1707–1718.
John, K.S., Bhat, S.G. and Rao, U.J.S.P. (2002) Involvement of peroxidase and polyphe-nol oxidase in mango sap-injury. Journal of Food Biochemistry 26, 403–414.

G.I. Johnson and P.J. Hofman590
Johnson, G.I. (1994) Mango, stem end rot, Part III. In: Ploetz, R.C., Zentmyer, G.A., Nishijima, W.T., Rohrbach, K.G. and Ohr, H.D. (eds) Compendium of Tropical Fruit Diseases. American Phytopathological Society (APS) Press, St Paul, Minnesota, pp. 39–41.
Johnson, G.I. (1995) Global outlook. In: Mango 2000 Marketing Seminar Handbook. Department of Primary Industry Brisbane, Queensland, pp. 21–30.
Johnson, G.I. (1997) Mango disease losses: balancing the economy and ecology. Acta Horticulturae 455, 575–586.
Johnson, G.I. and Coates, L.M. (1991) Quality assurance in tropical fruit – future disease control prospects. In: Sustainable Management of Pests, Diseases and Weeds, Aus-tralian Horticulture Clean and Green in the 90s, Proceedings of the First National Conference, Australian Society of Horticultural Science. Macquarie University, Syd-ney, pp. 71–76.
Johnson, G.I. and Coates, L.M. (1993) Postharvest diseases of mango. Postharvest News and Information 4, 27N–34N.
Johnson, G.I. and Heather, N.W. (1995) Disease and pest control in tropical fruit. In: Champ, B.R. and Highley, E. (eds) Postharvest Technology for Agricultural Products in Vietnam. Australian Centre for International Agricultural Research (ACIAR) Pro-ceedings 60. ACIAR, Canberra, pp. 100–126.
Johnson, G.I. and Hofman, P.J. (eds) (2004) Agri-product Supply-Chain Management in Developing Countries. Australian Centre for International Agricultural Research (ACIAR) Proceedings 119. ACIAR, Canberra.
Johnson, G.I. and Sangchote, S. (1994) Control of postharvest diseases of tropical fruits: challenges for the 21st century. In: Champ, B.R., Highley, E. and Johnson, G.I. (eds) Postharvest Handling of Tropical Fruits. Australian Centre for International Agricul-tural Research (ACIAR) Proceedings 50. ACIAR, Canberra, pp. 140–161.
Johnson, G.I., Muirhead, I.F., Mayers, P. and Cooke, T. (1989a) Diseases. In: Ridgeway, R. (ed.) Mango Pests and Disorders. Department of Primary Industries, Brisbane, Queensland, pp. 1–9.
Johnson, G.I., Muirhead, I.F. and Rappel, L.M. (1989b) Mango post-harvest disease con-trol: a review of research in Australia, Malaysia and Thailand. ASEAN Food Journal 4, 139–141.
Johnson, G.I., Boag, T.S., Cooke, A.W., Izard, M., Panitz, M. and Sangchote, S. (1990a) Interaction of postharvest disease control treatments and gamma radiation on man-goes. Annals of Applied Biology 116, 245–257.
Johnson, G.I., Sangchote, S. and Cooke, A.W. (1990b) Control of stem-end rot (Dothi-orella dominicana) and other postharvest diseases of mangoes (cv. Kensington Pride) during short- and long-term storage. Tropical Agriculture 67, 183–187.
Johnson, G.I., Mead, A.J, Cooke, A.W. and Dean, J.R. (1991) Mango stem end rot patho-gens – infection levels between fl owering and harvest. Annals of Applied Biology 119, 465–473.
Johnson, G.I., Mead, A.J., Cooke, A.W. and Dean, J.R. (1992) Mango stem end rot patho-gens – fruit infection by endophytic colonisation of the infl orescence and pedicel. Annals of Applied Biology 120, 225–234.
Johnson, G.I., Cooke, A.W. and Mead, A.J. (1993) Infection and quiescence of mango stem end rot pathogens. Acta Horticulturae 341, 329–336.
Johnson, G.I., Cooke, A.W., Mayers, P.E., Bagshaw, J.S., Ledger, S.N., Yuen, C.M.C., Tan, S.C. and Joyce, D.C. (1996) Mangoes. In: Coates, L., Cooke, T., Persley, D., Beattie, B. and Wade, N. (eds) Postharvest Diseases of Horticultural Produce. Vol. 2: Tropi-cal Fruit. Queensland Department of Primary Industries, Brisbane, Queensland, pp. 46–62.

Postharvest Technology 591
Johnson, G.I., Sharp, J.L., Milne, D.L. and Oosthuyse, S.A. (1997) Postharvest technol-ogy and quarantine treatments. In: Litz, R.E. (ed.) The Mango: Botany, Production and Uses, CAB International, Wallingford, UK, pp. 447–508.
Johnson, G.I., Highley, E. and Joyce, D.C. (eds) (1998) Disease Resistance in Fruit. Aus-tralian Centre for International Agricultural Research (ACIAR) Proceedings 80. ACIAR, Canberra.
Johnson, G.I., Weinberger, K. and Wu, M.H. (2008) The Vegetable Industry in Tropical Asia: an Overview of Production and Trade, with a Focus on Thailand, Indonesia, the Philippines, Vietnam and India [CD-ROM]. Shanhua, Taiwan: AVRDC - The World Vegetable Center. 56 pp. (Exploration series; no. 1.)
Joshi, G.D. and Roy, S.K. (1985) Spongy tissue in mango a physiological disorder. IndianHorticulture 29, 21–22.
Joyce, D.C. and Patterson, B. (1994) Postharvest water relations in horticultural crops: principles and problems. In: Champ, B.R., Highley, E. and Johnson, G.I. (eds) Post-harvest Handling of Tropical Fruits. Australian Centre for International Agricultural Research (ACIAR) Proceedings 50. ACIAR, Canberra, pp. 228–238.
Joyce, D.C., Hockings, P.D., Mazucco, R.A., Shorter, A.J. and Brereton, I.M. (1993) Heat treatment injury of mango fruit revealed by non-destructive magnetic resonance imaging. Postharvest Biology and Technology 3, 305–311.
Joyce, D.C., Macnish, A.J., Hofman, P.J., Simons, D.H. and Reid, M.S. (1999) Use of 1-methylcyclopropene to modulate banana ripening. In: Kanellis, A.K., Chang, C., Klee, H., Bleecker, A.B., Pech, J.C. and Grierson, D. (eds) Biology and Biotechnol-ogy of the Plant Hormone Ethylene II. Kluwer Academic Publishers, Dordrecht, the Netherlands, pp. 189–190.
Joyce, D.C., Shorter, A.J. and Hockings, P.D. (2001) Mango fruit calcium levels and the effect of postharvest calcium infi ltration at different maturities. Scientia Horticultu-rae 91, 81–99.
Kane, O. and Marcellin, P. (1978) Incidence of ripening and chilling the mitrochondria from mango fruits. Plant Physiology 61, 634–638.
Karunanayake, K.O.L.C. (2007) Natural defence mechanisms in mango fruit and their potential in management of postharvest diseases. PhD thesis, University of Peride-nya, Peridenya, Sri Lanka.
Katrodia, J.S. (1989) Spongy tissue in mango – causes and control measures. Acta Hor-ticulturae 231, 814–826.
Katrodia, J.S. and Sheth, I.K. (1989) Spongy tissue development in mango fruit of cultivar ‘Al-phonso’ in relation to temperature and its control. Acta Horticulturae 231, 827–834.
Ke, D. and Kader, A.A. (1992) Potential of controlled atmospheres for postharvest insect disinfestation of fruits and vegetables. Postharvest News and Information 3, 31N–37N.
Keil, H., Wasserman, D. and Dawson, E.R. (1946) Mango dermatitis and its relationship to poison-ivy hypersensitivity. Annals of Allergy 4, 268–281.
Kernot, I.I., Meurant, N., Holmes, R., MacLeod, N., Fullelove, G. and Bally, I. (eds) (1999) Mango Information Kit. Queensland Department of Primary Industries, Bris-bane, Queensland.
Kitagawa, H., Matsui, T., Kawada, K. and Agravante, J.U. (1992) Bagging of fruit on the tree to control disease. Acta Horticulturae 321, 871–875.
Kitinoja, L. and Kader, A.A. (2003) Small-scale Postharvest Handling Practices – a Man-ual for Horticultural Crops. University of California, Davis, California.
Kobiler, I., Shalom, Y., Roth, I., Akerman, M., Vinokur, Y., Fuchs, Y. and Prusky, D. (2001) Effect of 2,4-dichlorophenoxyacetic acid on the incidence of side and stem end rots in mango fruits. Postharvest Biology and Technology 23, 23–32.

G.I. Johnson and P.J. Hofman592
Kondo, S., Kittikorn, M. and Kanlayanarat, S. (2005) Preharvest antioxidant activities of tropical fruit and the effect of low temperature storage on antioxidants and jas-monates. Postharvest Biology and Technology 36, 309–318.
Koomen, I., Dodd, J.C., Jeger, M.J. and Jeffries, P. (1990) Postharvest biocontrol of an-thracnose disease of mangoes. Journal of the Science of Food and Agriculture 50,137–138.
Korsten, L. (2006) Advances in control of postharvest diseases in tropical fresh produce. International Journal of Postharvest Technology and Innovation 1, 48–61.
Korsten, L., Van Harmelen, M.W.S., Heitmann, A., De Villiers, E.E. and De Jager, E.S. (1991) Biological control of postharvest mango fruit diseases. South African Mango Growers’ Association Yearbook 11, 65–67.
Korsten, L., Lonsdale, J.H., De Villiers, E.E. and De Jager, E.S. (1992) Preharvest biologi-cal control of mango diseases. South African Mango Growers’ Association Yearbook 12, 72–78.
Korsten, L., De Villiers, E.E. and Lonsdale, J.H. (1993) Biological control of mango post-harvest disease in the packhouse. South African Mango Growers’ Association Year-book 13, 117–121.
Korsten, L., De Villiers, E.E., Wehner, F.C. and Kotze, J.M. (1994) A review of biological control of postharvest diseases of subtropical fruits. In: Champ, B.R., Highley, E. and Johnson, G.I. (eds) Postharvest Handling of Tropical Fruits. Australian Centre for International Agricultural Research (ACIAR) Proceedings 50. ACIAR, Canberra, pp. 172–185.
Kostermans, A.J.G.H. and Bompard, J.M. (1993) The Mangoes, their Botany, Nomencla-ture, Horticulture and Utilisation. Academic Press, London.
Kramer, A. (1973) Fruits and vegetables. In: Kramer, A. and Twigg, B.A. (eds) Quality Control for the Food Industry. Vol. 2. AVI, Westport, Connecticut, USA, pp. 157–228.
Kuo, L.S., Su, C.Y., Hseu, C.Y., Chao, Y.F., Chen, H.Y., Liao, J.Y. and Chu, C.F. (1987) Vapour Heat Treatment for Elimination of Dacus dorsalis and Dacus cucurbitae In-festing Mango Fruits. Taiwan Bureau of Commodity Inspection and Quarantine, Ministry of Economic Affairs, Taipei, Taiwan.
Lahav, E. and Whiley, A.W. (2002) Irrigation and mineral nutrition. In: Whiley, A.W., Schaffer, B. and Wolstenholme, B.N. (eds) The Avocado: Botany, Production and Uses. CAB International, Wallingford, UK, pp. 259–297.
Lalel, H.J.D. and Singh, Z. (2006) Controlled atmosphere storage of ‘Delta R2E2’ mango fruit affects production of aroma volatile compounds. Journal of Horticultural Sci-ence and Biotechnology 81, 449–457.
Lalel, H.J.D., Singh, Z. and Tan, S. (2003a) Glycosidically-bound aroma volatile com-pounds in the skin and pulp of ‘Kensington Pride’ mango fruit at different stages of maturity. Postharvest Biology and Technology 29, 205–218.
Lalel, H.J.D., Singh, Z. and Tan, S.C. (2003b) Maturity stage at harvest affects fruit ripen-ing, quality and biosynthesis of aroma volatile compounds in ‘Kensington Pride’ mango. Journal of Horticultural Science and Biotechnology 78, 225–233.
Lalel, H.J.D., Singh, Z. and Tan, S.C. (2004) Ripening temperatures infl uence biosynthe-sis of aroma volatile compounds in ‘Kensington Pride’ mango fruit. Journal of Hor-ticultural Science and Biotechnology 79, 146–157.
Ledger, S.N. (1986) Mango packaging. In: Proceedings of the First Australian Mango Research Workshop. Commonwealth Scientifi c and Industrial Research Organiza-tion (CSIRO), Melbourne, pp. 314–319.
Ledger, S.N. (1991a) Impact and pressure resistance of mangoes – postharvest handling of mangoes. Acta Horticulturae 291, 479–488.

Postharvest Technology 593
Ledger, S.N. (1991b) Post-harvest handling of mangoes. Acta Horticulturae 291, 479–488.
Ledger, S.N. (2003a) Don’t gas hot fruit. Mango Care 39, 2–3.Ledger, S.N. (2003b) Two handling systems. Mango Care 36, 13–14.Ledger, S.N. (2004) Control of Postharvest Diseases in Mango. Department of Primary
Industries and Fisheries (DPI&F) note. Department of Primary Industries and Fisher-ies, Brisbane, Australia.
Ledger, S.N. (2007) Managing mango ripening and storage rooms. In: Delivering Mango Research. The Sixth Amistar Australian Mango Conference. Gold Coast, Queensland, pp. 26–29. Available at: http://www.mangoes.net.au/downloads/File/PDF/industry_conference/mango_conference_07_proceedings.pdf (accessed 4 April 2008).
Ledger, S.N. and Bagshaw, J. (1994) Stimulating change in horticulture quality manage-ment. Australasian Postharvest Conference Proceedings. University of Queensland Gatton College, Lawes, Queensland, pp. 35–40.
Ledger, S.N. and Premier, R. (2006) Interpretive Guide for Association of South-east Asian Nations (ASEAN) Good Agricultural Practice (GAP) Produce Quality Module – Good Agricultural Practices for Production of Fresh Fruit and Vegetables in ASEAN Countries, Quality Assurance Systems for ASEAN Fruit and Vegetables Project. ASE-AN-Australia Development Cooperation Project. Available at: http://www.aphnet.org/gap/ASEANgap.html (accessed 1 May 2008).
Ledger, S.N., Campbell, T.P., Holmes, R.J. and Kernot, I.I. (2002a) The saleable life index. Mango Care 36, 2–3.
Ledger, S.N., Holmes, R. and Kernot, I. (2002b) Better mangoes – special edition. MangoCare 36, 1–14.
León, D.M., Ortega, D.A., Cabrera, H., de la Cruz, J., Parkin, K.L. and Garcia, H.S. (2000) Postharvest disinfestation of mango (Mangifera indica cv. Manila) with con-trolled atmospheres. Journal of Applied Horticulture 2, 71–75.
Lim, T.K. and Kuppelweiser, W. (1993) Mango sapburn amelioration in the Northern Territory. Acta Horticulturae 341, 518–527.
Lin, T.H., Tseng, F.C., Chang, C.R. and Wang, I.Y. (1976) Multiple treatment for disinfest-ing oriental fruit fl y in mangoes. Plant Protection Bulletin 18, 231–241.
Lizada, M.C.C. (1994) Fruit handling systems in developing countries. In: Champ, B.R., Highley, E. and Johnson, G.I. (eds) Postharvest Handling of Tropical Fruits. Austra-lian Centre for International Agricultural Research (ACIAR) Proceedings 50. ACIAR, Canberra, pp. 109–115.
Lonsdale, J.H. (1992) In search of an effective postharvest treatment for the control of postharvest diseases of mangoes. South African Mango Growers’ Association Year-book 12, 32–36.
Lonsdale, J.H. (1993) Strategies for the control of postharvest diseases of mango. SouthAfrican Mango Growers’ Association Yearbook 13, 109–116.
Loveys, B.R., Robinson, S.P., Brophy, J.J. and Chacko, E.K. (1992) Mango sapburn: com-ponents of fruit sap and their role in causing skin damage. Australian Journal of Plant Physiology 19, 449–457.
Lunn, D. (2004) Prochloraz. Food and Agriculture Organization (FAO) Pesticide Man-agement – Pesticide Specifi cations and Quality Control Standards. Available at: http://www.fao.org/ag/AGP/AGPP/Pesticid/JMPR/Download/2004_eva/Prochloraz.pdf (accessed 28 October 2007).
MacLeod, A.J. and Snyder, C.H. (1985) Volatile components of two cultivars of mango from Florida. Journal of Agriculture and Food Chemistry 33, 380–384.
MacLeod, A.J., MacLeod, G. and Snyder, C.H. (1988) Volatile aroma constituents of mango (cv. Kensington). Phytochemistry 27, 2189–2193.

G.I. Johnson and P.J. Hofman594
Mahayothee, B., Mühlbauer, W., Neidhart, S., Leitenberger, M. and Carle, R. (2004) Non-destructive determination of maturity of Thai mangoes by near-infrared spec-troscopy. Acta Horticulturae 645, 581–588.
Mahendra, M.S., Suryadi, M. and Padmono, N. (2002) Current status of the rural econ-omy and measures for revitalising it and increasing farmers’ income in Indonesia: development of substantial Gedong Gincu mango plantations in West Java. In: The Ninth Japanese International Research Centre for Agricultural Sciences Inter-national Symposium 2002 – Value-Addition to Agricultural Products. Tsukuba, Ja-pan, pp. 49–52.
Mangan, R.L. and Ingle, S.J. (1992) Forced hot-air quarantine treatment for mangoes infested with West Indian fruit fl y (Diptera: Tephritidae). Journal of Economic Ento-mology 85, 1859–1864.
Maqbool, M., Malik, A.U. and Jabbar, A. (2007) Sap dynamics and its management in commercial mango cultivars of Pakistan. Pakistan Journal of Botany 39, 1565–1574.
Marigheto, N., Venturi, L. and Hills, B. (2008) Two-dimensional NMR relaxation studies of apple quality. Postharvest Biology and Technology 48, 331–340.
Maxtend (2008) Maxim freshness – global service. Available at: www.maxtend.com.au (accessed 7 April 2008).
McGregor, B.M. (1987) Tropical Products Transport Handbook. United States Department of Agriculture (USDA), Agricultural Handbook No. 668. USDA, Washington, DC.
McKenzie, C.B. (1994) The background skin colour of exported mango fruit in relation to tree nitrogen status. South African Mango Growers’ Association Yearbook 14, 26–28.
McLauchlan, R.L. and Barker, L.R. (1994) Controlled atmospheres for ‘Kensington’ man-go storage: classical atmospheres. In: Johnson, G.I. and Highley, E. (eds) Develop-ment of Postharvest Handling Technology for Tropical Tree Fruits. Australian Centre for International Agricultural Research (ACIAR) Proceedings 58. ACIAR, Canberra, pp. 25–29.
McLauchlan, R.L. and Wells, L.A. (1994) Storage and ripening temperatures for ‘Kensington’ mangoes. In: Johnson, G.I. and Highley, E. (eds) Development of Postharvest Handling Technology for Tropical Tree Fruits. Australian Centre for International Agricultural Research (ACIAR) Proceedings 58. ACIAR, Canberra, pp. 25–29.
McMaugh, T. (2005) Guidelines for the surveillance for plant pests in Asia and the Pa-cifi c. Australian Centre for International Agricultural Research (ACIAR) Mongraph MN119 [English], MN119a [Indonesian]. ACIAR, Canberra. Available at: http://www.aciar.gov.au/publication/MN119+%5BEnglish%5D%2C+MN119a+%5BIndonesian%5D (accessed 1 May 2008.).
Mead, A.J. and Winston, E.C. (1991) Description of the disorder ‘stem-end cavity’, pos-sible causes and suggestions for reducing incidence in packing sheds. Acta Horti-culturae 291, 265–271.
Medlicott, A.P. (1985) Mango fruit ripening and the effects on maturity, temperature and gases. PhD thesis, Wolverhampton Polytechnic, Wolverhampton, UK.
Medlicott, A.P., Reynolds, S.B., New, S.W. and Thompson, A.K. (1987) Harvest maturity effects on mango fruit ripening. Tropical Agriculture 65, 153–157.
Medlicott, A.P., N’Diaye, M. and Sigrist, J.M.M. (1990a) Harvest maturity and concen-tration and exposure time to acetylene infl uence initiation of ripening in mangos. Journal of the American Society of Horticultural Science 115, 426–430.
Medlicott, A.P., Sigrist, J.M.M. and Sy, O. (1990b) Ripening of mangoes following low temperature storage. Journal of the American Society of Horticultural Science 115,430–434.

Postharvest Technology 595
Meiburg, G.F., Hofman, P.J., Smith, L.G., Cooke, A.W. and Barker, J.A. (1998) Quality of ‘Kensington’ mangoes after short duration exposure to high carbon dioxide concen-trations. In: Coates, L.M., Hofman, P.J. and Johnson, G.I. (eds) Disease Control in Storage Life Extension in Fruit. Australian Centre for International Agricultural Re-search (ACIAR) Proceedings No. 81. ACIAR, Canberra, pp. 55–60.
Milne, D.L. (1994) Postharvest handling of avocado, mango and lychee for export from South Africa. In: Champ, B.R., Highley, E. and ]ohnson, G.I. (eds) Postharvest Han-dling of Tropical Fruits. Australian Centre for International Agricultural Research (ACIAR) Proceedings 50. ACIAR, Canberra, pp. 73–89.
Minnis, D. (1993) Horticultural marketing – what’s really involved. In: Australasian Post-harvest Conference Proceedings, Department of Primary Industries, Gatton, Queen-sland, Australia, pp. 203–206.
Minnis, D.E. (1994) Prospects for marketing tropical fruits in Asia. In: Champ, B.R., Highley, E. and Johnson, G.I. (eds) Postharvest Handling of Tropical Fruits. Austra-lian Centre for International Agricultural Research (ACIAR) Proceedings 50. ACIAR, Canberra, pp. 56–64.
Mitcham, E.J. (2007) Heat with controlled atmospheres. In: Tang, J., Mitcham, E., Wang, S. and Lurie, S. (eds) Heat Treatments for Postharvest Pest Control: Theory and Practice. CAB International, Wallingford, UK, pp. 251–268.
Mitchell, G.E., McLauchlan, L., Isaacs, A.R., Williams, D.J. and Nottingham, S.M. (1992) Effect of low dose irradiation on composition of tropical fruits and vegetables. Jour-nal of Food Composition and Analysis 5, 291–311.
Mizrach, A. (2008) Ultrasonic technology for quality evaluation of fresh fruit and vege-tables in pre- and postharvest processes. Postharvest Biology and Technology 48, 315–330.
Moreno, M., Castell-Perez, M.E., Gomes, C., Da Silva, P.F. and Moreira, R.G. (2006) Effects of electron beam irradiation on physical, textural, and microstructural properties of ‘Tommy Atkins’ mangoes (Mangifera indica L.). Journal of Food Science 71, E80–E86.
Moreno, M., Castell-Perez, M.E., Gomes, C., Da Silva, P.F., Kim, J. and Moreira, R.G. (2007) Irradiation for postharvest quarantine insects. Formosan Entomology 27, 1–15.
Muirhead, I.F. (1976) Postharvest control of mango anthracnose with benomyl and hot water. Australian Journal of Experimental Agriculture and Animal Husbandry 16,600–603.
Muirhead, I.F. and Grattidge, R. (1986) Postharvest diseases of mango – the Queensland experience. In: Proceedings of the First Australian Mango Workshop. Common-wealth Scientifi c and Industrial Research Organization (CSIRO), Melbourne, pp. 248–252.
Muller, A.T. and Burt, J.R. (1989) Postharvest storage control of mango stem end rot with fungicidal dips. Australian Journal of Experimental Agriculture 29, 125–127.
Murray, E. and George, E. (1994) Fruit packing house operations to improve returns. In: Champ, B.C., Highley, E. and Johnson, G.I. (eds) Postharvest Handling of Tropical Fruits. Australian Centre for International Agricultural Research (ACIAR) Proceed-ings 50. ACIAR, Canberra, pp. 105–108.
Nair, S.S., Singh, Z. and Tan, S. (2003) Chilling injury development in ‘Kensington Pride’ mango fruit in relation to free polyamines. In: Australian Postharvest Horticulture Conference. Brisbane, Queensland, pp. 278–279.
Nascimento, A.S., Malavasi, A., Morgante, J.S. and Duarte, A.L.A. (1992) Hot-water im-mersion treatment for mangoes infested with Anastrepha fraterculus, A. obliqua,and Ceratitis capitata (Diptera: Tephritidae) in Brazil. Journal of Economic Entomol-ogy 85, 456–460.

G.I. Johnson and P.J. Hofman596
Natural Resources Institute (NRI) (2008a) Small Producers in Export Horticulture: a Guide to Best Practice. Elements for Successful Business. Natural Resources InstituteUK: the University of Greenwich. Available at: http://www.nri.org/projects/NRET/SPCDR/Chapter1/elements-1-5-1.htm (accessed 28 March 2008).
Natural Resources Institute (NRI) (2008b) Smallholders in Export Horticulture: a Guide to Best Practice. Natural Resources Institute UK: the University of Greenwich. Avail-able at: http://www.nri.org/projects/NRET/SPCDR/index.htm (accessed 1 April 2008).
Neven, L.G. (2008) Organic quarantine treatments for tree fruits. HortScience 43, 22–26.Nguyen, H.X. (2003) Pre-harvest and postharvest factors affecting skin colour and other
quality attributes of ‘Kensington Pride’ mango (Mangifera indica Linn.). PhD thesis, University of Queensland, Brisbane, Queensland.
Nguyen, H., Hofman, P.J., Smith, L.G., Stubbings, B., Adkins, M. and McConchie, R. (2002) Effect of ethylene and ripening temperatures on the skin colour and fl esh characteristics of ripe ‘Kensington Pride’ mango fruit. Acta Horticulturae 575, 635–642.
Nguyen, H., Hofman, P., Holmes, R., Bally, I., Stubbings, B. and McConchie, R. (2004) Effects of nitrogen on the skin colour and other quality attributes of ripe ‘Kensington Pride’ mango Mangifera indica L. fruit. Journal of Horticultural Science and Bio-technology 79, 204–210.
Nguyen, M.C., Wei, W., Vo, T.T., Rankin, M. and Russell, I. (2004) Getting farmers to work together: the experiences of mango growers in the Mekong Delta region of Vietnam. In: Johnson, G.I. and Hofman, P.J. (eds) Agriproduct Supply-Chain Man-agement in Developing Countries. Australian Centre for International Agricultural Research (ACIAR) Proceedings 119. ACIAR, Canberra, pp. 107–111.
O’Hare, T.J. (1994) The susceptibility of Thai and Australian mango cultivars to sap injury and possible means of control. In: Johnson, G.I. and Highley, E. (eds) Devel-opment of Postharvest Handling Technology for Tropical Tree Fruits. Australian Centre for International Agricultural Research (ACIAR) Proceedings 58. ACIAR, Canberra, pp. 21–24.
O’Hare, T.J. and Prasad, A. (1993) The alleviation of sap-induced skin injury by calcium hydroxide. Acta Horticulturae 321, 372–381.
O’Hare, T.J., McLaughlan, R.L. and Turner, C.D. (1994) Effect of co-shipment of exotic tropical fruit on existing fruit transported in bulk shipment. Proceedings of the Aus-tralasian Postharvest Conference. University of Queensland, Gatton, Queensland, pp. 245–250.
O’Hare, T.J., Bally, I.S.E., Dahler, J.M., Saks, Y. and Underhill, S.J.R. (1999) Characterisa-tion and induction of ‘etch’ browning in the skin of mango fruit. Postharvest Biol-ogy and Technology 16, 269–277.
Oka, K., Saito, F., Yasuhara, T. and Sugimoto, A. (2004) A study of cross-reactions be-tween mango contact allergens and urushiol. Contact Dermatitis 51, 292–296.
Oosthuyse, S.A. (1990) Effect of lowering the temperature of the cold storage regime of export mangoes after four weeks of storage. South African Mango Growers’ Asso-ciation Yearbook 10, 38–56.
Oosthuyse, S.A. (1991) Correlating pre-storage ripeness of mangoes with the proportion of good quality fruit present after four weeks of cold-storage. South African Mango Growers’ Association Yearbook 11, 49–53.
Oosthuyse, S.A. (1992) Long-term refrigerated storage of mangoes: what are the issues? South African Mango Growers’ Association Yearbook 12, 61–65.
Oosthuyse, S.A. (1993) Disorders of fi breless mangos growth in South Africa for export. South African Mango Growers’ Association Yearbook 13, 80–88.

Postharvest Technology 597
Oosthuyse, S.A. (1994) Quality of mature ‘Zill’ mangoes after long-term refrigerated storage as determined by pre-storage ripeness and cold-storage regime. South Afri-can Mango Growers’ Association Yearbook 14, 37–42.
Oosthuyse, S.A. (1995) The South African industry experience. South African Mango Growers’ Association Yearbook 15, 1–11.
Oosthuyse, S.A. (1998) Effect of environmental conditions at harvest on the incidence of lenticel damage in mango. South African Mango Growers’ Association Yearbook18, 15–17.
Oosthuyse, S.A. (1999) Effects of each of the stages in the pack-line on the incidence of lenticel damage in ‘Keitt’ and ‘Tommy Atkins’ mango. South African Mango Grow-ers’ Association Yearbook 19, 37–39.
Opara, L.U. and Nguyen, H.X. (1999) Postharvest handling and environmental factors infl uencing mango fruit quality – a review. Journal of South Pacifi c Agriculture 6, 1–19.
Pacifi c Data Systems (PDS) (2008) Ethylene/Ripening Gas Controlling System. Pacifi c Data Systems. Available at: http://www.pacdatasys.com.au/dataloggers/Systems_Overview.htm (accessed 7 April 2008).
Peacock, B.E. (1986) Postharvest handling of mangoes. In: Proceedings of the First Australian Mango Research Workshop. Commonwealth Scientifi c and Industrial Research Organization (CSIRO), Melbourne, pp. 295–313.
Peak, E.M., Fitzell, R.D., Hannah, R.S. and Batten, D.J. (1986) Development of a micro-processor-based data recording system for predicting plant disease based on studies on mango anthracnose. Computers and Electronics in Agriculture 1, 251–262.
Peña, J. (2004) Integrated pest management and monitoring techniques for mango pests. Acta Horticulturae 645, 151–161.
Penchaiya, P., Jansasithorn, R. and Kanlavanarat, S. (2006) Effect of 1-MCP on physio-logical changes in mango ‘Nam Dok Mai’. Acta Horticulturae 712, 717–721.
Pesis, E., Ampunpong, C., Shusiri, B. and Hewett, E.W. (1994) Enhancement of ethylene and CO2 production in apple fruit following short-term exposure to high CO2. Post-harvest Biology and Technology 4, 309–317.
Pesis, E., Aharoni, D., Aharon, Z., Ben-Arie, R., Aharoni, N. and Fuchs, Y. (2000) Modi-fi ed atmosphere and modifi ed humidity packaging alleviates chilling injury symp-toms in mango fruit. Postharvest Biology and Technology 19, 93–101.
Phakawatmongkol, W., Ketsa, S. and van Doorn, W.G. (2004) Variation in fruit chilling injury among mango cultivars. Postharvest Biology and Technology 32, 115–118.
Piñeiro, M., Berania, L. and Ríos, D. (2004) Improving the Quality and Safety of Fresh Fruits and Vegetables: a Practical Approach. Manual for Trainers. Food and Agricul-ture Organization (FAO), Rome. Available at: http://www.fao.org/docrep/007/y5488e/y5488e00.htm (accessed 1 May 2008).
Plan, M.R.R., Joyce, D.C., Ogle, H.J. and Johnson, G.I. (2002) Mango stem-end rot (Botryosphaeria dothidea) disease control by partial-pressure infi ltration of fungi-cides. Australian Journal of Experimental Agriculture 42, 625–629.
Ploetz, R.C. (ed.) (2003) Diseases of Tropical Fruit Crops. CAB International, Wallingford, UK.
Ploetz, R.C. (2004) The major diseases of mango: strategies and potential for sustainable management. Acta Horticulturae 645, 137–150.
Ploetz, R.C. (2007) Diseases of tropical perennial crops: challenging problems in di-verse environments. Plant Disease 91, 644–663.
Ploetz, R.C. and Prakash, O. (1997) Foliar, fl oral and soilborne diseases. In: Litz, R.E. (ed.) The Mango: Botany, Production and Uses. CAB International, Wallingford, UK, pp. 281–325.

G.I. Johnson and P.J. Hofman598
Poffl ey, M., Owens, G., Kulkani, V., Smith, E.S.C. and Conde, B. (1999) Mango Management – Flowering to Market. Agnote, 301 No. D9, December 1999. Agdex No: 234/11.
Plant Protection and Quarantine (PPQ) (2007) 5. Treatment Manual – Treatment Schedules T100 – Schedules for Fruit, Nuts, and Vegetables. United States Department of Ag-riculture (USDA) Animal and Plant Health Inspection Service (APHIS). Available at: http://www.aphis.usda.gov/import_export/plants/manuals/ports/downloads/treatment_pdf/05_02_t100schedules.pdf (accessed 31 October 2007).
Proctor, F.J. and Cropley, J.P. (1994) Trends and changes in the European market for tropical fruits and their impact on technology requirements. In: Champ, B.R., High-ley, E. and Johnson, G.I. (eds) Postharvest Handling of Tropical Fruits. Australian Centre for International Agricultural Research (ACIAR) Proceedings 50. ACIAR, Canberra, pp. 65–72.
Prusky, D. and Keen, N.T. (1993) Involvement of preformed antifungal compounds in the resistance of subtropical fruits to fungal decay. Plant Disease 77, 114–119.
Prusky, D. and Plumbley, R.A. (1992) Quiescent infections of Colletotrichum in tropical and subtropical fruit. In: Bailey, J.A. and Jeger, M.J. (eds) Colletotrichum: Biology, Pathology and Control. CAB International, Wallingford, UK, pp. 289–307.
Prusky, D., Fuchs, Y. and Zauberman, G. (1980) A method for preharvest assessment of latent infections in fruit. Annals of Applied Biology 98, 79–85.
Prusky, D., Keen, N.T., Sims, J.J. and Midland, S.L. (1982) Possible involvement of an antifungal compound in latency of Colletotrichum gloeosporioides in unripe avo-cado fruits. Phyiopathology 72, 1578–1582.
Prusky, D., Plumbley, R.A. and Kobiler, I. (1992) Modulation of natural resistance of avocado fruit by CO2 treatments. Physiological and Molecular Plant Pathology 39,325–334.
Prusky, D., Gat, Z. and Burd, P. (1993a) Effect of relative humidity during growth on the incidence of quiescent infections of Altemaria alternata. Plant Disease 77, 249–252.
Prusky, D., Kobiler, I., Zauberman, G. and Fuchs, Y. (1993b) Preharvest conditions and postharvest treatments affecting incidence of decay in mango fruits during storage. Acta Horticulturae 341, 307–320.
Prusky, D., Plumbley, R.A., Kobiler, I., Zauberman, G. and Fuchs, Y. (1993c) The effect of elevated CO2 levels on the symptom expression of Colletotrichum gloeospori-oides on avocado fruits. Plant Pathology 42, 900–904.
Prusky, D., Fuchs, Y., Kobiler, I., Roth, I., Weksler, A., Shalom, Y., Fallik, E., Zauberman, G., Pesis, E., Akerman, M., Ykutiely, O., Weissblum, A., Regev, R. and Artes, L. (1999) Effect of hot water brushing, prochloraz treatment and waxing on the inci-dence of black spot decay caused by Alternaria alternata in mango fruits. Posthar-vest Biology and Technology 15, 165–174.
Prusky, D., Kobiler, I., Akerman, M. and Miyara, I. (2006) Effect of acidic solutions and acidic prochloraz on the control of postharvest decay caused by Alternaria alternata in mango and persimmon fruit. Postharvest Biology and Technology 42, 134–141.
Regional Standards for Phytosanitary Measures (RSPM) (2004) Guidelines for the Estab-lishment, Maintenance and Verifi cation of Fruit Fly Pest Free Areas in North America. RSPM No. 17. North American Plant Protection Organisation (NAPPO). Available at: http://www.nappo.org/Standards/REVIEW/RSPMNo17-e.pdf (accessed 1 May 2008).
Reid, M.S. (2002) Ethylene in postharvest technology. In: Kader, A.A. (ed.) Postharvest Technology of Horticultural Crops, 3rd edn. University of California, Oakland, California, pp. 149–162.
Reuveni, M. (2000) Effi cacy of trifl oxystrobin (Flint), a new strobilurin fungicide, in con-trolling powdery mildews on apple, mango and nectarine, and rust on prune trees. Crop Protection 19, 335–341.

Postharvest Technology 599
Reuveni, M., Harpaz, M. and Reuveni, R. (1998) Integrated control of powdery mildew in fi eld-grown mango trees by foliar sprays of mono-potassium phosphate fertiliser, sterol inhibitor fungicides and the stroilurin Kresoxym-methyl. European Journal of Plant Pathology 104, 853–860.
Reyes, L.F. and Cisneros-Zevallos, L. (2007) Electron-beam ionizing radiation stress effects on mango fruit (Mangifera indica L.) antioxidant constituents before and during post-harvest storage. Journal of Agricultural and Food Chemistry 55, 6132–6139.
Reyes, M.U., Paull, R.E., Gautz, L.D., Armstrong, J.W. and Follett, P.A. (2000) Non-destructive inspection of mango fruit (Mangifera indica L.) with soft X-ray imaging. Acta Horticulturae 509, 787–792.
Robinson, S.P., Loveys, B.R. and Chacko, E.K. (1993) Polyphenol oxidase enzymes in the sap and skin of mango fruit. Australian Journal of Plant Physiology 20, 99–107.
Rolle, R.S. (2006) Improving postharvest management and marketing in the Asia-Pacifi c region: issues and challenges. In: Postharvest Management of Fruit and Vegetables in the Asia-Pacifi c Region – Reports of the Asian Productivity Organisation (APO) Seminar on Reduction of Postharvest Losses of Fruit and Vegetables held in India; and Marketing and Food Safety: Challenges in Postharvest Management of Ag-ricultural/Horticultural Products in Islamic Republic of Iran. Asian Productivity Organisation, pp. 23–31. Available at: http://www.apo-tokyo.org/00e-books/AG-18_PostHarvest.htm (accessed 1 May 2008).
Rosa, B.A., Aharoni, D., Feygenberg, O., Aharoni, N., Keynan, A. and Pesis, E. (2001) Effect of modifi ed atmosphere packaging on mango ripening. Acta Horticulturae553, 607–609.
Sadasivam, R., Muthuswamy, S., Sundaraj, J.S. and Vasudevan, V. (1971) Note on chill-ing injury in mango (Mangifera indica L.) fruits in refrigerated storage. Indian Jour-nal of Agricultural Science 41, 715–716.
Saranwong, S., Sornsrivichai, J. and Kawano, S. (2004) Prediction of ripe-stage eating quality of mango fruit from its harvest quality measured non-destructively by near infrared spectroscopy. Postharvest Biology and Technology 31, 137–145.
Schoorl, D. and Holt, J.E. (1982) Planning for change in horticulture – a case study in packing shed design. Agricultural Systems 8, 77–86.
Schoorl, D. and Holt, J.E. (1985) Design of a packing shed for the ripening, packing and handling of tomatoes. Queensland Agricultural Journal 111, 185–190.
Segarra-Carmona, A.E., Franqui, R.A., Ramirez-Ramos, L.V., Santiago, L.R. and Torres-Rivera, C.N. (1990) Hot-water dip treatments to destroy Anastrepha obliqua larvae (Diptera: Tephritidae) in mangoes from Puerto Rico. Journal of Agriculture of the University of Puerto Rico 74, 441–447.
Self, G., de Assis, J.S. and Caron, V.C. (2006) Effects of postharvest handling on lenticel spotting of ‘Tommy Atkins’ mangoes from Northeast Brazil. Acta Horticulturae 712, 543–550.
Seo, S.T., Chambers, D.L., Akamine, E.K., Komura, M. and Lee, C.Y.L. (1972) Hot water-ethylene dibromide fumigation-refrigeration treatment for mangoes infested by orien-tal and Mediterranean fruit fl ies. Journal of Economic Entomology 65, 1372–1374.
Seo, S.T., Kobayashi, R.M., Chambers, D.L., Steiner, L.F., Lee, C.Y.L. and Komura, M. (1974) Mango weevil: cobalt 60 gamma-irradiation of packaged mangoes. Journalof Economic Entomology 67, 504–505.
Sepiah, M. (1986) Effectiveness of hot water, hot benomyl and cooling on postharvest diseases of mango. ASEAN Food Journal 2, 117–120.
Seymour, G.B., N’diaye, M., Wainwright, H. and Tucker, G.A. (1990) Effects of cultivar and harvest maturity on ripening of mangoes during storage. Journal of Horticul-tural Science 65, 479–483.

G.I. Johnson and P.J. Hofman600
Sharp, J.L. (1992) Hot-air quarantine treatment for mango infested with Caribbean fruit fl y (Diptera: Tephritidae). Journal of Economic Entomology 85, 2302–2304.
Sharp, J.L. and Heather, N.W. (2002) Quarantine treatments for pests of tropical fruits. In: Peña, J.E., Sharp, J.L. and Mysoki, M. (eds) Tropical Fruit Pests and Pollinators – Biology, Economic Importance, Natural Enemies and Control. CAB International, Wallingford, UK, pp. 391–406.
Sharp, J.L. and Picho-Martinez, H. (1990) Hot-water quarantine treatment to control fruit fl ies in mangoes imported into the United States from Peru. Journal of Eco-nomic Entomology 83, 1940–1943.
Sharp, J.L. and Spalding, D.H. (1984) Hot water as a quarantine treatment for Florida mangos infested with Caribbean fruit fl y. Proceedings of the Florida State Horticul-tural Society 97, 355–357.
Sharp, J.L., Ouye, M.T., Thalman, R., Hart, W., Ingle, S. and Chew, V. (1988) Submersion of ‘Francis’ mango in hot water as a quarantine treatment for the West Indian fruit fl y and the Caribbean fruit fl y (Diptera: Tephritidae). Journal of Economic Entomology 81, 1431–1436.
Sharp, J.L., Ouye, M.T., Ingle, S.J. and Hart, W.G. (1989a) Hot-water quarantine treat-ment for mangoes from Mexico infested with Mexican fruit fl y and West Indian fruit fl y (Diptera: Tephritidae). Journal of Economic Entomology 82, 1657–1662.
Sharp, J.L., Ouye, M.T., Ingle, S.J., Hart, W.G., Enkerlin, W.R.H., Hilario Celedonio, H., Jorge Toledo, A., Stevens, L., Quintero, E., Reyes, J.F. and Schwarz, A. (1989b) Hot-water quarantine treatment for mangoes from the state of Chiapas, Mexico, infested with Mediterranean fruit fl y and Anastrepha serpentina (Wiedemann) (Diptera: Tephritidae). Journal of Economic Entomology 82, 1663–1666.
Shorter, A. and Joyce, D.C. (1994) Effect of surface coatings, on sap burn of ‘Kensington Pride’ mango fruit. Tropical Agriculture 71, 243–246.
Shukla, R.P. and Tandon, P.L. (1985) Bio-ecology and management of the mango weevil, Sternocbetus mangiferae (Fabricius) (Coleoptera: Curculionidae). InternationalJournal of Tropical Agriculture 3, 293–303.
Simmons, S.L. (1998) The effects of preharvest factors on postharvest quality of ‘Kensing-ton Pride’ mango (Mangifera indica L.). PhD thesis, University of Queensland, Bris-bane, Queensland.
Simmons, S.L., Hofman, P.J. and Hetherington, S.E. (1995) The effects of water stress on mango fruit quality. Proceedings of the Mango 2000 Production workshop, 191–197.
Simmons, S.L., Hofman, P.J., Whiley, A.W. and Hetherington, S.E. (1998) Effects of leaf:fruit ratios on fruit growth, mineral concentration and quality of mango (Mangifera indica L. cv. Kensington Pride). Journal of Horticultural Science and Biotechnology 73, 367–374.
Singh, R.N., Singh, G., Mishra, J.S. and Rao, O. (1987) Studies on the effect of pre- and post-harvest treatment of calcium nitrate and calcium chloride on the storage life of ‘Amrapali’ mango. Progressive Horticulture 19, 1–9.
Singh, Z. and Janes, J. (2001) Effects of postharvest application of ethephon on fruit rip-ening, quality and shelf life of mango under modifi ed atmosphere packaging. Acta Horticulturae 553, 599–602.
Singh, Z., Janes, J. and Tan, S.C. (2000) Effects of different surfactants on calcium uptake and its effects on fruit ripening, quality and postharvest storage of mango under modifi ed atmosphere packaging. Acta Horticulturae 509, 413–417.
Singh, Z., Janes, J. and Suresh, N. (2001) Packaging materials affect physiological weight loss, fruit colour and quality of mango during storage. Acta Horticulturae 553, 603–604.

Postharvest Technology 601
Singh, Z., Lalel, H.J.D. and Suresh, N. (2004) A review of mango fruit aroma volatile compounds – state of the art research. Acta Horticulturae 645, 519–527.
Siriphanich, J. (1994) Minimal processing of tropical fruits. In: Champ, B.R., Highley, E. and Johnson, G.I. (eds) Postharvest Handling of Tropical Fruits. Australian Centre for International Agricultural Research (ACIAR) Proceedings 50. ACIAR, Canberra, pp. 127–137.
Sivapalasingam, S., Barrett, E., Kimura, A., Van Duyne, S., De Witt, W., Ying, M., Frisch, A., Phan, Q., Gould, E., Shillam, P., Reddy, V., Cooper, T., Hoekstra, M., Higgins, C., Sanders, J.P., Tauxe, R.V. and Slutsker, L. (2003) A multistate outbreak of serotype newport infection linked to mango consumption: impact of water-dip disinfestation technology. Clinical Infectious Diseases 37, 1585–1590.
Slippers, B., Johnson, G.I., Crous, P.W., Coutinho, T.A., Wingfi eld, B.D. and Wingfi eld, M.J. (2005) Phylogenetic and morphological re-evaluation of the Botryosphaeriaspecies causing diseases of Mangifera indica. Mycologia 97, 102–113.
Smillie, R.M., Hetherington, S.E., Nott, R., Chaplin, G.R. and Wade, N.L. (1987) Appli-cations of chlorophyll fl uorescence to the postharvest physiology and storage of mango and banana fruit and the chilling tolerance of mango cultivars. ASEAN Food Journal 3, 55–59.
Smith, E.S.C. (1992) Fruit fl y disinfestation in mangoes by hot water dipping. In: Ooi, P.A.C., Lim, G.S. and Teng, P.S. (eds) Proceedings of the Third International Confer-ence on Plant Protection in the Tropics. Malaysian Plant Protection Society No. 6.Malaysian Plant Protection Society, Kuala Lumpur, pp. 188–195.
Smith, T.E., Hofman, P.J., Stephenson, R.A., Asher, C.J. and Hetherington, S.E. (1997) Improving boron nutrition improves ‘Hass’ avocado fruit size and quality. In: Cutting, J. (ed.) Conference ’97: Searching for Quality. Joint Conference of the Australian Avo-cado Growers’ Federation and the New Zealand Avocado Growers’ Association.Rotorua, New Zealand, pp. 131–137.
Snowdon, A. (1990) A Colour Atlas of Post-harvest Disease and Disorders of Fruits and Vegetables, Vol. 1. Fruits and General Introduction. Wolfe Scientifi c, London.
Snowdon, A. (1994) Diagnosing the causes of outturn problems in imported tropical fruits. In: Champ, B.R., Highley, E. and Johnson, G.I. (eds) Postharvest Handling of Tropical Fruits. Australian Centre for International Agricultural Research (ACIAR) Proceedings 50. ACIAR, Canberra, pp. 94–101.
Sonneveld, C. (2006) Measures to assure better food safety, marketing, and consumer satisfaction in fruit and vegetables. In: Postharvest Management of Fruit and Vege-tables in the Asia-Pacifi c Region – Reports of the Asian Productivity Organisation (APO) Seminar on Reduction of Postharvest Losses of Fruit and Vegetables held in India; and Marketing and Food Safety: Challenges in Postharvest Management of Agricultural/Horticultural Products in Islamic Republic of Iran. Asian Productivity Organisation, pp. 84–99. Available at: http://www.apo-tokyo.org/00e-books/AG-18_PostHarvest.htm (accessed 1 May 2008).
Sornsrivichai, J., Anusadorn, P., Oogaki, C. and Gemma, H. (1989) Storage life and qual-ity of mango (Mangifera indica L. cv. Keaw Sawoey) fruits stored in seal-packaging by plastic fi lms and under low pressure at different temperatures. Japanese Journal of Tropical Agriculture 33, 6–17.
Spalding, D.H., Benschoter, C.A., von Windeguth, D.L., King, J.R., Reeder, W.F. and Burditt, A.K., Jr (1977) Methyl bromide and phosphine fumigation injury to avo-cados and mangos. Proceedings of the Florida State Horticultural Society 90, 268–270.
Stephens, B.E. and Tanner, D.J. (2005) The harvest watch system – measuring fruit’s healthy glow. Acta Horticulturae 687, 363–364.

G.I. Johnson and P.J. Hofman602
Stovolt, G.E. and Dirou, G.E. (2004) Blight Disease in Mangoes. Agnote, DPI-489 First Edition 2004. Agdex 217/10. New South Wales Department of Primary Industries, New South Wales.
Subedi, P.P., Walsh, K.B. and Owens, G. (2007) Prediction of mango eating quality at harvest using short-wave near infrared spectrometry. Postharvest Biology and Tech-nology 43, 326–334.
Subramanayam, H., Krishnamurthy, S., Subhadra, N.V. and Dalal, V. (1980) Studies on internal breakdown, a physiological ripening disorder in Alphonso mangoes (Mangifera indica L.). Tropical Science 13, 203–209.
Suhaila, M. and Halim, M.T. (1994) Anti-fruit-fl y activity of extracts of black pepper and other edible plants. In: Champ, B.R., Highley, E. and Johnson, G.I. (eds) Postharvest Handling of Tropical Fruits. Australian Centre for International Agricultural Re-search (ACIAR) Proceedings 50. ACIAR, Canberra, pp. 371–372.
Sunagawa, K., Kume, K. and Lwaizumi, R. (1987) The effectiveness of vapour heat treat-ment against the melon fl y, Dacus cucurbitae Coquillett, in mango and fruit toler-ance to the treatment. Research Bulletin of Plant Protection Japan 23, 13–20.
Sunarharum, W.B. (2007) Identifi cation of key volatiles responsible for the unique aromas and fl avours of different mango varieties. MSc thesis, University of Queensland, Bris-bane, Queensland.
Sundravadana, S., Alice, D., Kuttalam, S. and Samiyappan, R. (2006) Control of mango anthracnose by azoxystrobin. Tunisian Journal of Plant Protection 1, 109–114.
Sundravadana, S., Alice, D., Kuttalam, S. and Samiyappan, R. (2007) Effi cacy of azox-ystrobin on Colletotrichum gloeosporiodes Penz. growth and on controlling mango anthracnose. Journal of Agricultural and Biological Science 2, 10–15.
Suresh, N., Singh, Z. and Tan, S.C. (2004) Chilling injury in relation to ethylene biosyn-thesis in ‘Kensington Pride’ mango fruit. Journal of Horticultural Science and Bio-technology 79, 82–90.
Suslow, T. (2000) Postharvest Handling for Organic Crops. University of California Pub-lication 7254. University of California, Davis, California.
Swaine, G., Melksham, K.J. and Corcoran, R.J. (1984) Dimethoate dipping of ‘Kensing-ton’ mango against Queensland fruit fl y. Australian Journal of Experimental Agricul-ture and Animal Husbandry 14, 620–623.
Taylor, R.W.D., Devereau, A.D. and Golob, P. (2002) Remedial treatments in pest man-agement. In: Golob, P., Farrell, G. and Orchard, J.E. (eds) Crop Post-harvest: Sci-ence and Technology, Vol. 1: Principles and Practice. Blackwell Science, Oxford, UK, pp. 321–359.
Terry, L.A. and Joyce, D.C. (2004) Elicitors of induced disease resistance in postharvest horticultural crops: a brief review. Postharvest Biology and Technology 32, 1–13.
Thomas, A.C. (1975) The Application of Ionizing Radiation to the Shelf Life Extension of Mangoes in South Africa. Republic of South Africa Atomic Energy Board, Pelinada-ba, Pretoria, PEL-244, South Africa.
Thomas, P. (1986) Radiation preservation of foods of plant origin. III. Tropical fruits: banana, mangoes, and papayas. Critical Reviews in Food Science and Nutrition 23, 147–206.
Thomas, P. and Joshi, M.R. (1988) Reduction in chilling injury in ripe ‘Alphonso’ mango fruit in cold storage by temperature conditioning. International Journal of Food Sci-ence and Technology 23, 447–455.
Thomas, P. and Oke, M.S. (1983) Improvement in quality and storage of ‘Alphonso’ mangoes by cold adaptation. Scientia Horticulturae 19, 257–262.
Thomas, P., Kannan, A., Degwekar, V.H. and Ramamurthy, M.S. (1995) Non-destructive detection of seed weevil-infested mango fruits by X-ray imaging. Postharvest Biol-ogy and Technology 5, 161–165.

Postharvest Technology 603
Thompson, A.K. (1998) Controlled Atmosphere Storage of Fruit and Vegetables. CAB International, Wallingford, UK.
Thompson, J.F. (2002) Tranportation. In: Kader, A.A. (ed.) Postharvest Technology of Horticultural Crops, 3rd edn. University of California, Oakland, California, pp. 259–269.
Thompson, J.F., Mitchell, F.G. and Kasmire, R.F. (2002) Cooling horticultural commodi-ties. In: Kader, A.A. (ed.) Postharvest Technology of Horticultural Crops, 3rd edn. University of California, Oakland, California, pp. 97–112.
Timmer, L.W. and Zitko, S.E. (1996) Evaluation of copper fungicides and rates of metallic copper for control of Melanose on grapefruit in Florida. Plant Disease 80, 166–169.
Torres, J.D., Talens, P., Carot, J.M., Chiralt, A. and Escrichea, I. (2007) Volatile profi le of mango (Mangifera indica L.) as affected by osmotic dehydration. Food Chemistry101, 219–228.
Truter, A.B. and Eksteen, G.J. (1987) Controlled and modifi ed atmospheres to extend storage life of avocados. South African Avocado Growers’ Association Yearbook 10, 151–153.
Underhill, S. and Dahler, J. (1995) Classifying skin browning. Mango Care 14, 6–7.Van der Linde, H.J. and Thord-Gray, R.S. (1986) Irradiation as a post harvest treatment
for mangoes. South African Mango Growers’ Association Yearbook 6, 24–26.Van Straten, B. and Oosthuyse, S.A. (1994) Die effek van koelopberging by 4 of 8°C op
die tempo van kwalititeit verslegting van ryp ‘Sensation’ mango vrugte. South Afri-can Mango Growers’ Association Yearbook 14, 34–36.
Varith, J., Sirikajornjaru, W. and Kiatsiriroat, T. (2007) Microwave-vapor heat disinfesta-tion on oriental fruit fl y eggs in mangoes. Journal of Food Processing and Preserva-tion 31, 253–269.
Velasco, L.R.I. and Medina, C. (2004) Soft X-ray imaging for non-destructive detection of mango pulp weevil (Sternochetus frigidus Fabr.) infestation in fresh mature green ‘Carabao’ mango (Mangifera indica L.) fruits. The Philippine Agricultural Scientist87, 160–164.
Vijaysegaran, S. (1994) Preharvest fruit fl y control: strategies for the tropics. In: Champ, B.R., Highley, E. and Johnson, G.I. (eds) Postharvest Handling of Tropical Fruits.Australian Centre for International Agricultural Research (ACIAR) Proceedings 50. ACIAR, Canberra, pp. 288–303.
Vinning, G.S. and Young, J. (2006) Linking production and marketing of fruit and vegeta-bles for better farm incomes in the Asia-Pacifi c region. In: Postharvest Management of Fruit and Vegetables in the Asia-Pacifi c Region – Reports of the Asian Productivity Organisation (APO) Seminar on Reduction of Postharvest Losses of Fruit and Veg-etables held in India; and Marketing and Food Safety: Challenges in Postharvest Management of Agricultural/Horticultural Products in Islamic Republic of Iran. Asian Productivity Organisation, pp. 53–69. Available at: http://www.apo-tokyo.org/00e-books/AG-18_PostHarvest.htm (accessed 1 May 2008).
von Windeguth, D.L. (1986) Gamma irradiation as a quarantine treatment for Caribbean fruit fl y infested mangos. Proceedings of the Florida State Horticultural Society 99,131–134.
Wagner, L.J. and Flores, H.E. (1994) Effect of taxol and related compounds on growth of plant pathogenic fungi. Phytopathology 84, 1173–1178.
Wainwright, H. and Burbage, M.B. (1989) Physiological disorders in mango (Mangiferaindica L.) fruit. Journal of Horticultural Science 64, 125–135.
Waite, G. (2002) Pests and pollinators of mango. In: Peña, J.E., Sharp, J.L. and Mysoki, M. (eds) Fruit Pests and Pollinators. Biology, Economic Importance, Natural Enemies and Control. CAB International, Wallingford, UK, pp. 103–130.

G.I. Johnson and P.J. Hofman604
Walker, K. (2007a) Mango Pulp Weevil (Sternochetus frigidus) Pest and Diseases Image Library. Available at: http://www.padil.gov.au (accessed 2 April 2008).
Walker, K. (2007b) Red Banded Mango Caterpillar (Deanolis sublimbalis). Pest and Diseases Image Library. Available at: http://www.padil.gov.au (accessed 31 October 2007).
Wall, M. (2008) Quality of postharvest horticultural crops after irradiation treatment. Stewart Postharvest Review 2008, 2, Article 1.
Wang, B., Wang, J., Liang, H., Yi, J., Zhang, J., Lin, L., Wu, Y., Feng, X., Cao, J. and Jiang, W. (2008) Reduced chilling injury in mango fruit by 2,4-dichlorophenoxyacetic acid and the antioxidant response. Postharvest Biology and Technology 48, 172–181.
Watada, A.E., Herner, R.C., Kader, A.A. and Romani, R.J. (1984) Terminology for the de-scription of developmental stages of horticultural crops. HortScience 19, 20–21.
Wells, I. and Littlemore, J. (1989) Dipping mangoes, watch the Benlate concentration. Queensland Fruit and Vegetable News 60, 18.
Whiley, A.W. (1999) Investigation into the effect of calcium on the fruit disorder ‘stem end cavity’ in the mango cultivar ‘Kensington Pride’. In: Whiley, A.W. (ed.) Flower-ing Behaviour and Subsequent Productivity in Mango. Final Report for Project no. 9012. Australian Centre for International Agricultural Research (ACIAR), Canberra.
Whiley, A.W. and Hofman, P.J. (2007) Development of Best Practice Pre- and Post-harvest Protocols for Production of Calypso™ mango. Final Report for Project FR02049. Horticulture Australia Ltd, Sydney.
Wikipedia (2007) Good Agricultural Practice. Available at: http://en.wikipedia.org/wiki/Good_Agricultural_Practices (accessed 31 October 2007).
Willingham, S.L., Coates, L.M., Cooke, A.W., Dean, J.R. and Beasley, D.R. (1999) Field management of mango postharvest pathogens. In: Asia-Pacifi c Plant Pathology for the New Millenium. 12th Biennial Australasian Plant Pathology Conference. Can-berra, p. 138.
Wills, R.B.H., Yuen, M.C.C., Sabari, Laksmi, L.D.S. and Suyanti (1988) Effect of calcium infi ltration on delayed ripening of three mango cultivars in Indonesia. ASEAN Food Journal 4, 67–68.
Wilson, C.L. and Pusey, P.L. (1985) Potential for biological control of postharvest plant diseases. Plant Disease 69, 375–378.
Wilson, C.L., El Ghaouth, A., Chalutz, E., Droby, S., Stevens, C., Lu, J.Y., Khan, V. and Arul, J. (1994) Potential of induced resistance to control postharvest diseases of fruits and vegetables. Plant Disease 78, 837–844.
Winston, E. (1986) Observations of internal mango fl esh breakdown: need for standard-ization of terminology. In: Proceedings of the First Australian Mango Research Workshop. Commonwealth Scientifi c and Industrial Research Organization (CSIRO), Melbourne, pp. 77–82.
Wittenberg, L. (2007) Mapping mango seed weevil. In: Delivering Mango Research. The Sixth Amistar Australian Mango Conference. Gold Coast, Queensland, pp. 31–33.
Wolstenholme, B.N. (2004) Nitrogen – the manipulator element: managing inputs and outputs in different enviroments. South African Avocado Growers’ Association Yearbook 27, 62–78.
World Trade Organisation (WTO) (2008) World Trade Organisation Technical Barriers to Trade. Available at: http://www.wto.org/english/tratop_e/tbt_e/tbt_e.htm (accessed 4 April 2008).
Wright, M.A. and Stringer, A. (1973) The toxicity of thiabendazole, benomyl, methyl benzimidazol-2-yl carbamate and thiophanate-methyl to the earthworm, Lumbri-cus terrestris. Pesticide Science 4, 431–432.
Yahia, E.M. (1994) The potential use of insecticidal atmospheres for mango, avocado, and papaya fruits. In: Champ, B.R., Highley, E. and Johnson, G.I. (eds) Postharvest

Postharvest Technology 605
Handling of Tropical Fruits. Australian Centre for International Agricultural Re-search (ACIAR) Proceedings No. 50. ACIAR, Canberra, pp. 373–374.
Yahia, E.M. (2006) Modifi ed and controlled atmospheres for tropical fruits. Stewart Post-harvest Review 2006(5), article 6.
Yahia, E.M. and Tiznando-Hernandez, M. (1993) Tolerance and responses of harvested mango to insecticidal low oxygen atmospheres. HortScience 28, 1031–1033.
Yahia, E.M. and Vazquez-Moreno, L. (1993) Responses of mango to insecticidal oxygen and carbon dioxide atmospheres. Food Science and Technology 26, 42–48.
Yarrow, R. and Chandler, L. (2007) Informal testing of the RBMC pheromone over early mango fruiting season at Lockerbie Station via Bamaga, Cape York Peninsula. In: Delivering Mango Research. The Sixth Amistar Australian Mango Conference. Gold Coast, Queensland, pp. 24–25.
Yashoda, H.M., Prabha, T.N. and Tharanathan, R.N. (2006) Mango ripening: changes in cell wall constituents in relation to textural softening. Journal of the Science of Food and Agriculture 86, 713–721.
Yogaratnam, N. and Johnson, D.S. (1982) The application of foliar sprays containing ni-trogen, magnesium, zinc and boron to apple trees. Journal of Horticultural Science57, 159–164.
Young, T.W. (1957) ‘Soft-nose’, a physiological disorder in mango fruits. Proceedings of the Florida State Horticulture Society 70, 280–283.
Young, T.W. and Miner, J.T (1961) Relationship of nitrogen and calcium to soft-nose disorder in mango fruit. Proceedings of the American Society of Horticultural Sci-ence 58, 201–208.
Yuen, C.M.C., Tan, S.C., Joyce, D. and Chettri, P. (1993) Effect of postharvest calcium and polymeric fi lms on ripening and peel injury in ‘Kensington Pride’ mango. ASE-AN Food Journal 8, 110–113.
Zainuri (2006) Defence mechanisms and induced resistance in ‘Kensington Pride’ man-go. PhD thesis, University of Queensland, Brisbane, Queensland.
Zainuri, Wearing, A.H., Coates, L. and Terry, L. (2001) Effects of phosphonate and salicylic acid treatments on anthracnose disease development and ripening of ‘Kensington Pride’ mango fruit. Australian Journal of Experimental Agriculture 41, 805–813.
Zainuri, Irving, D.E., Dann, E.K., Coates, L.M. and Wearing, A.H. (2003) Activating man-go fruit defence to anthracnose disease. In: Australian Postharvest Horticulture Conference. Brisbane, Queensland, pp. 149–150.
Zeng, K. and Waibo, J. (2005) Induced resistance to anthracnose (Colletotrichum gloeo-sporioides) in mango by postharvest treatment with salicylic acid. Journal of China Agricultural University 10, 36–40.
Zeng, K.F., Cao, J.K. and Jiang, W. (2006) Enhancing disease resistance in harvested mango (Mangifera indica L.) fruit by salicylic acid. Journal of the Science of Food and Agriculture 85, 694–698.

© CAB International 2009. The Mango, 2nd Edition: Botany, Production and Uses606 (ed. R.E. Litz)
16 World Mango Trade and the Economics of Mango Production
E.A. Evans and O.J. MendozaUniversity of Florida, Florida, USA
16.1 Introduction 60616.2 Recent Trends in World and USA Mango Production, Trade and Consumption 607
World situation 607USA mango production, imports and consumption 611
16.3 Sample Costs and Returns Associated with the Establishment and Production of Mango Orchards 613
General approach to estimating cost of production of orchard crops 614Main assumptions 614Discussion of establishment phase budget 617Discussion of production phase budget 621Profi tability analysis 624
16.4 Conclusions 626
16.1 Introduction
Worldwide mango production occurs in over 90 countries. Although only a relatively small proportion of total mango production enters international trade (<4%), the volume traded has increased substantially since the late 1990s. Among the factors responsible for the increased mango production, trade and consumption are lower prices, year-round availability, fewer horticultural trade barriers, changes in food consumption preferences, longer shelf life for perishables and consumer interest in healthier foods. Although not a major mango producer, the USA has developed most of the popular cultivars traded on the international market, and is the largest single-country mango importer. The costs and returns and general practices of establishing and maintaining orchards vary considerably from coun-try to country and within each country (different regions and production systems).

World Mango Trade and Economics 607
16.2 Recent Trends in World and USA Mango Production, Trade and Consumption
World situation
Asia accounts for approximately 77% of global mango production, and the Americas and Africa account for approximately 13% and 9%, respectively (FAOSTAT, 2007). In 2005, world production of mango was estimated to have reached 28.51 million t, an increase from the 27.82 million t recorded in the previous year. Between 1996 and 2005, production grew at an average annual rate of 2.6%. Table 16.1 shows the world’s top ten mango-producing coun-tries, which account for about 85% of the world’s production.
India is the largest producer, accounting for 38.58% of global production from 2003 to 2005. During that period, the Indian mango crop averaged 10.79 million t, followed by China and Thailand at 3.61 million t (12.90%) and 1.73 million t (6.20%), respectively. Other leading mango-producing coun-tries and their respective shares of world production during the 2003–2005 period include Mexico (5.50%), Indonesia (5.29%), Pakistan (4.48%), Brazil (4.30%), the Philippines (3.53%), Nigeria (2.61%) and Egypt (1.28%).
Although currently only 3.3% of the world production of mango is traded globally, this represents a noticeable increase over the quantities traded since the late 1980s. In terms of distribution, Mexico, Brazil, Peru, Ecuador and Haiti supply the majority of North America’s imports. India and Pakistan are the predominant suppliers to the West Asian market. South-east Asian coun-tries get most of their supplies from the Philippines and Thailand. European Union (EU) buyers source mango mainly from South America and Asia.
In 2005, global exports of mango reached 912,853 t, a slight decrease of 0.73% compared with the previous year, and were valued at US$543,100,000 (FAOSTAT, 2007). Table 16.2 shows the top ten major mango-exporting coun-tries. India is the largest producer but only recently has overtaken Mexico as the number one exporter of the fruit. For the 2003–2005 period, Mexico and India dominated the export trade with shares of 22.64% and 20.25%, respec-tively, followed by Brazil (13.18%) and Pakistan (6.94%). Other major export-ers include the Netherlands (major re-exporter), Peru, Ecuador, the Philippines, Thailand and China.
World imports of mango increased from 397,623 t in 1996 to 826,584 t in 2005. The USA is the number one importer of mango. During the 2003–2005 period, the USA imported 271,848 t, or approximately a third of the total mango imports (Table 16.3).
The Netherlands imported 88,300 t of mangoes (10.62%), although most of it is redistributed throughout the EU. Other prominent importing countries that are also major redistributors are the United Arab Emirates (6.82%) and Saudi Arabia (5.32%). Most of these imports are redistributed to other countries within the Middle East. Although China (4.91%) appears as a major importer, the quan-tities imported have been declining. For example, China imported 57,000 t in 2004 and only 19,000 t in 2005. This could be due to increases in domestic pro-duction in response to an increase in domestic demand driven by rising per
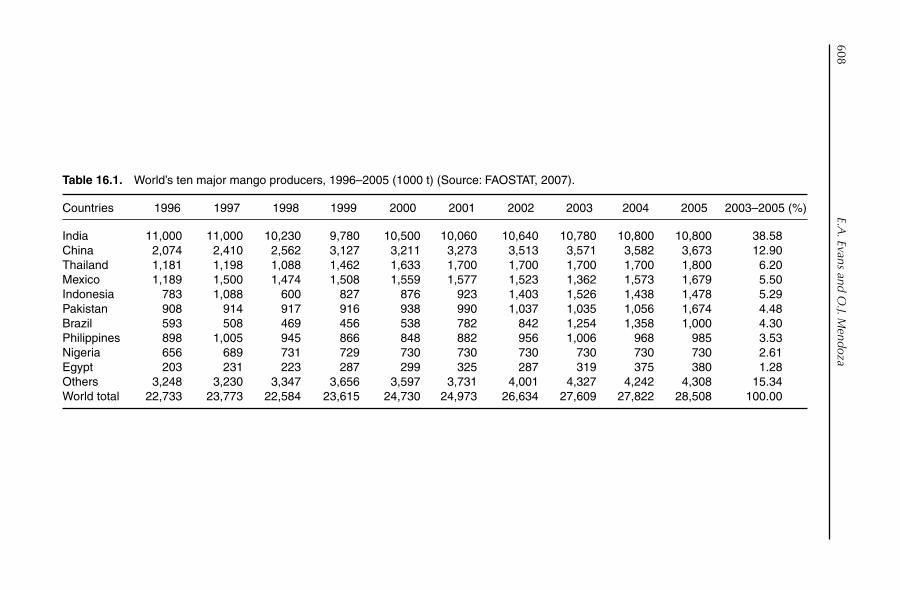
E.A. Evans and O
.J. Mendoza
608
Table 16.1. World’s ten major mango producers, 1996–2005 (1000 t) (Source: FAOSTAT, 2007).
Countries 1996 1997 1998 1999 2000 2001 2002 2003 2004 2005 2003–2005 (%)
India 11,000 11,000 10,230 9,780 10,500 10,060 10,640 10,780 10,800 10,800 38.58China 2,074 2,410 2,562 3,127 3,211 3,273 3,513 3,571 3,582 3,673 12.90Thailand 1,181 1,198 1,088 1,462 1,633 1,700 1,700 1,700 1,700 1,800 6.20Mexico 1,189 1,500 1,474 1,508 1,559 1,577 1,523 1,362 1,573 1,679 5.50Indonesia 783 1,088 600 827 876 923 1,403 1,526 1,438 1,478 5.29Pakistan 908 914 917 916 938 990 1,037 1,035 1,056 1,674 4.48Brazil 593 508 469 456 538 782 842 1,254 1,358 1,000 4.30Philippines 898 1,005 945 866 848 882 956 1,006 968 985 3.53Nigeria 656 689 731 729 730 730 730 730 730 730 2.61Egypt 203 231 223 287 299 325 287 319 375 380 1.28Others 3,248 3,230 3,347 3,656 3,597 3,731 4,001 4,327 4,242 4,308 15.34World total 22,733 23,773 22,584 23,615 24,730 24,973 26,634 27,609 27,822 28,508 100.00

World M
ango Trade and Economics
609
Table 16.2. World’s ten major mango-exporting countries, 1996–2005 (1000 t) (Source: FAOSTAT, 2007).
Countries 1996 1997 1998 1999 2000 2001 2002 2003 2004 2005 2003–2005 (%)
Mexico 148 187 209 204 207 195 195 216 213 195 22.64India 27 45 47 38 39 46 42 179 156 223 20.25Brazil 24 23 39 54 67 94 104 138 111 114 13.18Pakistan 18 25 39 41 48 52 48 60 82 49 6.94Netherlands 21 25 17 37 34 43 33 58 51 69 6.42Peru 11 6 11 20 21 27 35 40 60 58 5.71Ecuador 0 2 7 0 26 34 30 38 41 40 4.31Philippines 40 45 53 35 40 39 36 38 36 25 3.61Thailand 8 9 10 10 9 11 9 8 33 2 1.55China 12 7 9 10 5 5 15 22 10 4 1.31Others 80 104 87 103 132 121 127 126 127 135 14.08World total 391 478 529 552 628 666 673 923 920 913 100.00
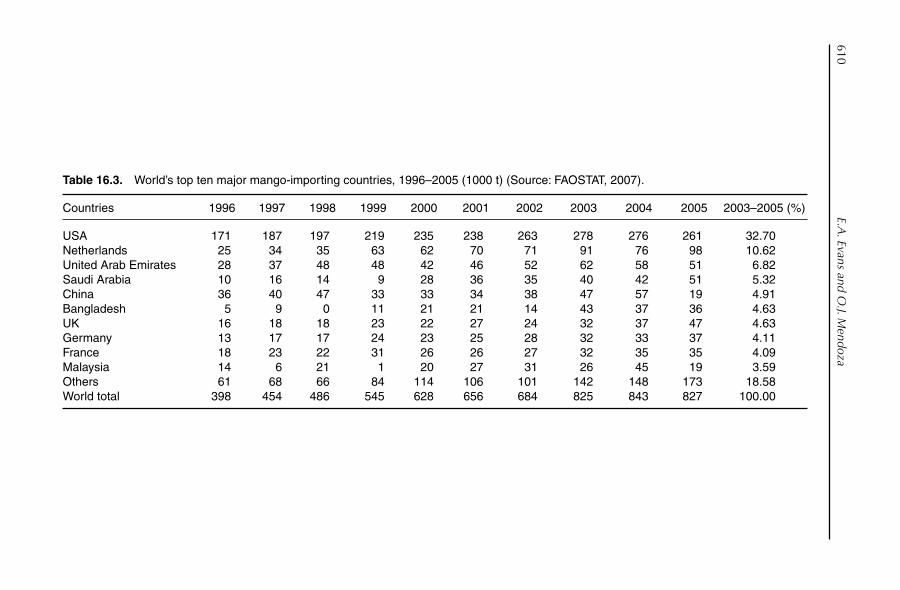
E.A. Evans and O
.J. Mendoza
610
Table 16.3. World’s top ten major mango-importing countries, 1996–2005 (1000 t) (Source: FAOSTAT, 2007).
Countries 1996 1997 1998 1999 2000 2001 2002 2003 2004 2005 2003–2005 (%)
USA 171 187 197 219 235 238 263 278 276 261 32.70Netherlands 25 34 35 63 62 70 71 91 76 98 10.62United Arab Emirates 28 37 48 48 42 46 52 62 58 51 6.82Saudi Arabia 10 16 14 9 28 36 35 40 42 51 5.32China 36 40 47 33 33 34 38 47 57 19 4.91Bangladesh 5 9 0 11 21 21 14 43 37 36 4.63UK 16 18 18 23 22 27 24 32 37 47 4.63Germany 13 17 17 24 23 25 28 32 33 37 4.11France 18 23 22 31 26 26 27 32 35 35 4.09Malaysia 14 6 21 1 20 27 31 26 45 19 3.59Others 61 68 66 84 114 106 101 142 148 173 18.58World total 398 454 486 545 628 656 684 825 843 827 100.00

World Mango Trade and Economics 611
capita income. Other noticeable importers include Bangladesh and the UK (4.63% each), Germany (4.11%), France (4.09%) and Malaysia (3.59%).
The most popular export mango cultivars continue to be ‘Kent’, ‘Tommy Atkins’, ‘Haden’ and ‘Keitt’. These cultivars have fruit with a red blush and are less fi brous, fi rmer and more suited for long-distance transportation. The green cultivars are only now being widely accepted in the international market and include cultivars such as ‘Ataulfo’ and ‘Amelie’. Other cultivars which are gain-ing signifi cance in international trade include: ‘Alphonso’, ‘Dudhpeda’, ‘Kesar’, ‘Sindhu’, ‘Pairi’, ‘Desi’, ‘Chausa’, ‘Langra’ and ‘Katchamita’. Most of these newer cultivars on the international scene are coming from India and Pakistan.
Since the late 1990s, per capita consumption has increased noticeably in the USA, Japan and China, mainly due to higher income levels, improved advertising and lower mango prices. On the international scene, prices for most mango varieties have declined considerably over the decade, dropping about 50%. For example, the average price for mango in the European mar-ket was US$12/kg in 1996, compared to just over US$6/kg today. The reduc-tion in price is partly due to the increased availability of tropical fruits. There is consensus, however, that prices have stabilized but could increase with proper promotional efforts.
Although the quantity of processed mango fruit that is traded internation-ally is small compared with the fresh fruit trade (<7%) there is evidence to suggest that it is increasing. Such products include mango juice, pickled man-goes, mango chutney, mango pulp and paste, mango purée, dried fruit, mango slices in brine and mango fl our. India is the main exporter of processed mango followed by Pakistan, Brazil and Zimbabwe. Major importers include the United Arab Emirates, Saudi Arabia, Kuwait, USA, UK and Canada.
USA mango production, imports and consumption
The USA is not a major mango producer even though most of the commer-cially traded varieties have been developed in Florida. USA production, mainly in Florida, remains fairly stable at a little under 3000 t/year.
The USA is currently the world’s leading importer of fresh mangoes, accounting for 32.70% of the total imports during the 2003–2005 period (FAOSTAT, 2007). Figure 16.1 shows the trend of mango imports into the USA between 1997 and 2006. Overall, the graph indicates a steady increase in the volume of mango imports. Between 1997 and 2006, imports increased from 187,193 t to 298,088 t, an average annual growth rate of 5.46%. Mango imports were valued at about US$233,100,000 in 2006 (USDA, Foreign Agri-cultural Service, 2007).
The main sources of USA imports of mango are Mexico, Peru, Ecuador and Brazil. Mexico is the main supplier of mango to the USA (60.78% share in 2006) (Fig. 16.2). In recent years, Brazil, Peru and Ecuador have become signifi cant exporters to the USA, competing with Mexico at the beginning and the end of the season. The USA exports very few of its mango imports, mainly to Canada and the UK.
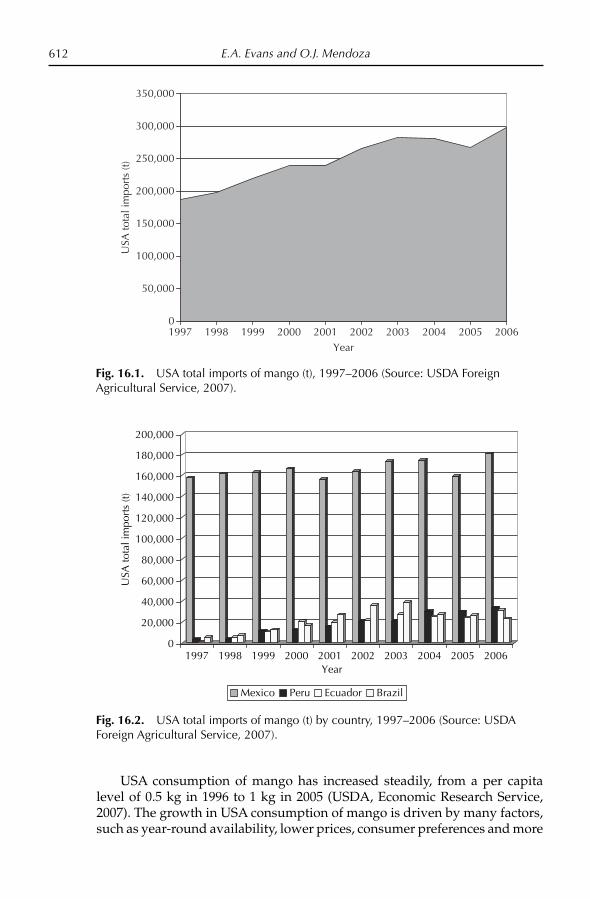
E.A. Evans and O.J. Mendoza612
USA consumption of mango has increased steadily, from a per capita level of 0.5 kg in 1996 to 1 kg in 2005 (USDA, Economic Research Service, 2007). The growth in USA consumption of mango is driven by many factors, such as year-round availability, lower prices, consumer preferences and more
0
50,000
100,000
150,000
200,000
250,000
300,000
350,000
1997 1998 1999 2000 2001 2002 2003 2004 2005 2006Year
USA
tota
l im
port
s (t)
Fig. 16.1. USA total imports of mango (t), 1997–2006 (Source: USDA Foreign Agricultural Service, 2007).
0
20,000
40,000
60,000
80,000
100,000
120,000
140,000
160,000
180,000
200,000
USA
tota
l im
port
s (t)
1997 1998 1999 2000 2001 2002 2003 2004 2005 2006Year
Mexico Peru Ecuador Brazil
Fig. 16.2. USA total imports of mango (t) by country, 1997–2006 (Source: USDA Foreign Agricultural Service, 2007).
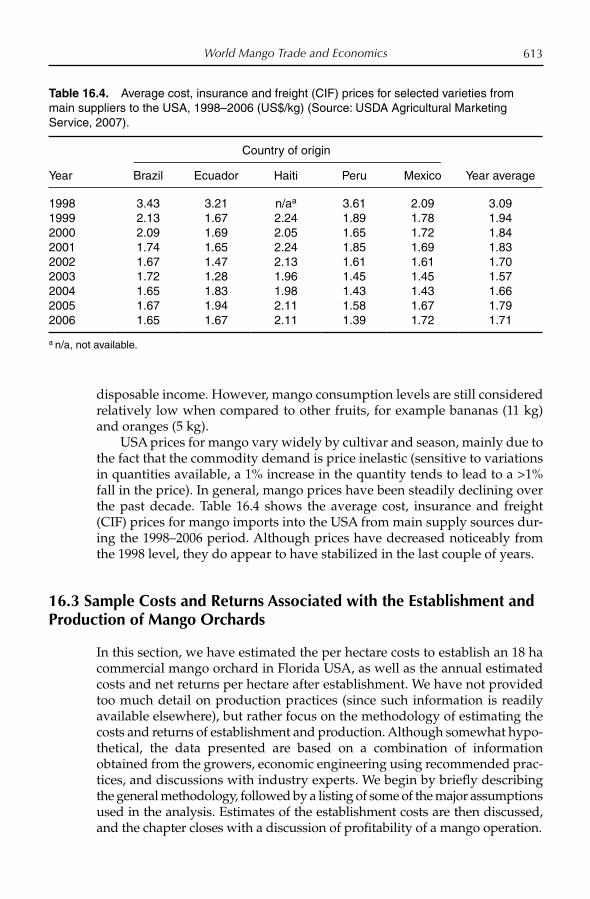
World Mango Trade and Economics 613
disposable income. However, mango consumption levels are still considered relatively low when compared to other fruits, for example bananas (11 kg) and oranges (5 kg).
USA prices for mango vary widely by cultivar and season, mainly due to the fact that the commodity demand is price inelastic (sensitive to variations in quantities available, a 1% increase in the quantity tends to lead to a >1% fall in the price). In general, mango prices have been steadily declining over the past decade. Table 16.4 shows the average cost, insurance and freight (CIF) prices for mango imports into the USA from main supply sources dur-ing the 1998–2006 period. Although prices have decreased noticeably from the 1998 level, they do appear to have stabilized in the last couple of years.
16.3 Sample Costs and Returns Associated with the Establishment and Production of Mango Orchards
In this section, we have estimated the per hectare costs to establish an 18 ha commercial mango orchard in Florida USA, as well as the annual estimated costs and net returns per hectare after establishment. We have not provided too much detail on production practices (since such information is readily available elsewhere), but rather focus on the methodology of estimating the costs and returns of establishment and production. Although somewhat hypo-thetical, the data presented are based on a combination of information obtained from the growers, economic engineering using recommended prac-tices, and discussions with industry experts. We begin by briefl y describing the general methodology, followed by a listing of some of the major assumptions used in the analysis. Estimates of the establishment costs are then discussed, and the chapter closes with a discussion of profi tability of a mango operation.
Table 16.4. Average cost, insurance and freight (CIF) prices for selected varieties from main suppliers to the USA, 1998–2006 (US$/kg) (Source: USDA Agricultural Marketing Service, 2007).
Year
Country of origin
Year averageBrazil Ecuador Haiti Peru Mexico
1998 3.43 3.21 n/aa 3.61 2.09 3.091999 2.13 1.67 2.24 1.89 1.78 1.942000 2.09 1.69 2.05 1.65 1.72 1.842001 1.74 1.65 2.24 1.85 1.69 1.832002 1.67 1.47 2.13 1.61 1.61 1.702003 1.72 1.28 1.96 1.45 1.45 1.572004 1.65 1.83 1.98 1.43 1.43 1.662005 1.67 1.94 2.11 1.58 1.67 1.792006 1.65 1.67 2.11 1.39 1.72 1.71
a n/a, not available.

E.A. Evans and O.J. Mendoza614
General approach to estimating cost of production of orchard crops
The general approach for estimating the cost of production of perennial crops (orchards and vineyards), which usually take more than a year to begin pro-duction, is to develop two separate budgets: one for the establishment phase and the other for the production phase. The establishment phase bud-get refl ects the sum total of all expenses (expressed on a yearly and per unit basis) that are incurred over the years to bring an orchard into meaningful (mature trees) production. Once this amount is determined, it is treated as if it were an expense incurred in the purchase of a ‘capital item’, for example machinery to be used in the production phase of the orchard. As in the case of a capital item, an equal annual amount (amortized amount) is charged to the production operation as an expense spread over the estimated future life of the orchard. In other words, the amortized amount is included as a non-cash overhead expenditure in the production phase budget when determin-ing net returns from the enterprise. The production phase budget estimates the costs and returns on an annual per unit basis associated with the mainte-nance of the orchard after it has been established. Although the procedure seems daunting, it is made much easier by using spreadsheet software with built-in formulas for calculating amortization.
Main assumptions
The following assumptions were used to estimate production costs and returns for 18 ha operations.
LandThe hypothetical farm consists of 20 ha. A mango orchard is being estab-lished on 18 ha, with roads, irrigation system and farmstead occupying 2 ha. The 18 ha orchard is considered large enough to use machinery and equip-ment effi ciently. The orchard is farmed by the owner with the help of hired part-time labour.
Site preparationIt is assumed that the land is relatively clear, with no costs being included for major land preparation such as timber clearing, rock removal or land level-ling. However, if these operations are required, they should be included. The main operations considered here are associated with fencing and road con-struction for travelling and harvesting. These operations are usually done 1 year prior to planting, but costs are shown in the fi rst year.
Planting, training and pruningTrees are planted on 7.5 m × 7.5 m spacing, or 175 trees/ha. The life of the orchard in this study is projected to be 25 years. The variety considered in this study is ‘Tommy Atkins’. Pruning and training begin in the third year, and labour time required for pruning increases in the successive years.

World Mango Trade and Economics 615
Hedging and topping operations are carried out immediately after fruit har-vesting.
FertilizationDuring the fi rst 3 years, an N-P-K fertilizer (6% nitrogen) is spread by hand six times each year. After year 3, the frequency of application decreases to four times each year. Table 16.5 shows the annual fertilizer rates assumed. In addition, the trees are given annual nutritional sprays of copper, zinc, man-ganese and boron. Iron is applied in chelated form as a soil drench two to three times each year.
IrrigationTotal irrigation costs include the cost of pumping water and irrigation labour. Water for our irrigation system is supplied from a well; therefore, the cost of the water is zero. Irrigation costs for individual orchards vary, depending on the amount of water pumped, pumping system, energy source and irrigation district.
Weed control managementWeeds in the tree rows are controlled with applied pre- and post-emergent (residual) contact herbicides such as Roundup®. With fi ve applications each year, spot sprays are applied at the rate of approximately 5 l/ha. However, as the trees grow larger and shade reduces weed growth, the number of appli-cations is reduced to approximately three per annum.
HarvestMango fruits are best harvested using clippers and hand-carried harvesting bags (about US$0.11/kg). With large trees, fruits are harvested using picking poles, with or without attached clippers, which are equipped with bags into which the fruit falls. After harvesting, the fruits are usually piled under a tree on the ground. The fruit is then loaded onto trucks in the fi eld and hauled to packing houses for US$0.11/kg.
Yields and returnsA major assumption is that the orchard requires approximately 7 years to reach maturity; hence the fi rst 6 years are considered the establishment phase. Even though trees require about 7 years before reaching maturity, they will start having saleable fruits from about the third year. Table 16.6 shows the typical yield assumptions used in the analysis. Annual gross fi eld yields at maturity are assumed to be 22,000 kg/ha.
Table 16.5. Annual fertilizer rates @ 6% N.
Year 1 2 3 4 5 6 7+
Rate (kg/ha) 236 472 707 786 1100 1257 1415

E.A. Evans and O.J. Mendoza616
LabourHourly wages for workers are US$10.00 for skilled workers and US$7.00 for fi eld workers. Adding 34% for the employer’s share of federal and state pay-roll taxes, insurance and other possible benefi ts, yields a labour rate of US$13/h for skilled labour and US$9/h for fi eld labour.
Cash overheadCash overhead consists of numerous cash expenses paid during each year that are assigned to the entire farm, not to a particular operation. These costs include property taxes, offi ce expenses, capital interest, liability and property insurance, equipment repairs, sanitation services and crop insurance.
PROPERTY TAXES. In the USA, most counties charge a base property tax rate of approximately 1% on the assessed value of the property.
INTEREST ON OPERATING CAPITAL. Interest on operating capital is based on cash operating costs and is calculated at a nominal rate of 5%/year. A nominal interest rate is the going market cost of borrowed funds.
INSURANCE. Insurance for farm investment differs depending on assets included and amount of coverage. Property insurance provides coverage for property loss and the standard practice in the USA according to the American Society of Agricultural Engineers (ASAE) is to charge about 0.67% of the average value of the assets over their useful life. Liability insurance covers accidents on the farm (US$580/year).
OFFICE EXPENSES. Offi ce and business operating expenses are estimated at US$355/ha, and include offi ce supplies, telephone, bookkeeping, account-ing, legal fees, road maintenance and miscellaneous administrative charges.
SANITATION SERVICES. Sanitation services offer portable toilets for orchards and cost the farm US$1900/year (US$106/ha). This cost includes delivery and servicing of a single toilet and washing unit.
CROP INSURANCE. Multi-peril crop insurance is purchased at a cost of US$445/ha.
Non-cash overheadNon-cash overhead is calculated as the capital recovery cost for equipment and other farm investments. Capital recovery cost is the annual depreciation
Table 16.6. Yield assumptions.
Year 1 2 3 4 5 6 7+
Yield (kg/ha) 0 0 1,700 4,500 9,000 15,500 22,000

World Mango Trade and Economics 617
and interest costs for a capital investment. It is the amount of money required each year to be set aside so that a capital item can be fully replaced at the end of its useful life. It can be viewed as the value of the portion of the capital item that was utilized in the production process during that year. The for-mula for the calculation of the annual capital recovery costs is the PMT Excel formula:
Capital Recovery = –PMT (I, N, (PP − SV)) + (I)*(SV)
Where:
PMT = the formula that calculates the payment for a loan based on con- ●
stant payments and a constant interest rate.I = the interest rate used to represent the cost of borrowed capital (in this ●
case, 5%).N = the number of years. ●
PP = the purchase price for the capital item. ●
SV = an estimate of the remaining value of an investment at the end of its ●
useful life. In this case, the percentage of remaining value is calculated from equations developed by the ASAE based on equipment type and years of life.
Farm equipment on a mango orchard in the region is purchased either new or used. The study shows the current purchase price for new equip-ment. The new purchase price is adjusted to 60% to indicate a mix of new and used equipment.
The orchard is irrigated using a sprinkler irrigation system. The life of the irrigation system is estimated at 20 years. The irrigation system is installed before the orchard is planted and includes costs of a pumping system, instal-lation labour and design and materials.
Although it is assumed that the grower owns the land, the going rental rate in South Florida of approximately US$2500/ha is used to refl ect the opportunity (economic) cost of land.
Discussion of establishment phase budget
Table 16.7 shows the layout of the establishment phase budget. The budget refl ects the annual per hectare costs incurred in establishing a mango orchard. Here it is assumed that a mango orchard reaches maturity in the seventh year. As such, the budget refl ects all associated cash and non-cash expenses incurred over 6 years.
The information required for each year is similar except for the fi rst 2 or 3 years. With reference to Table 16.7, year 1 includes pre-planting costs (site preparation and orchard layout), which in this case amounts to US$5000/ha. Most of the pre-planting costs most likely would occur in the year before, but the practice is to include these costs in year 1. Planting costs include new trees, digging holes for trees, setting trees, and wrapping and staking trees. Both pre-planting and planting costs appear only in year 1. Total per hectare
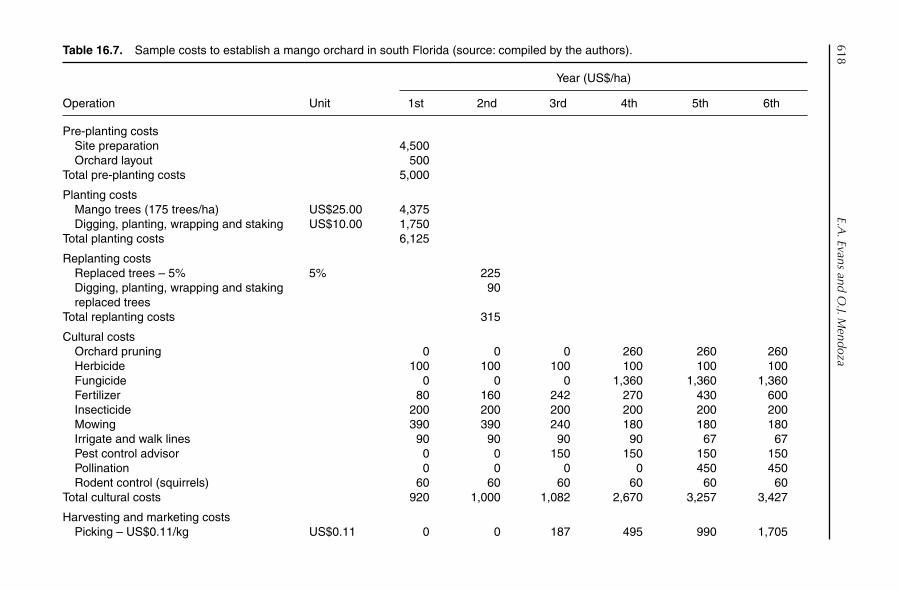
E.A. Evans and O
.J. Mendoza
618Table 16.7. Sample costs to establish a mango orchard in south Florida (source: compiled by the authors).
Operation Unit
Year (US$/ha)
1st 2nd 3rd 4th 5th 6th
Pre-planting costs Site preparation 4,500 Orchard layout 500Total pre-planting costs 5,000
Planting costs Mango trees (175 trees/ha) US$25.00 4,375 Digging, planting, wrapping and staking US$10.00 1,750Total planting costs 6,125
Replanting costs Replaced trees – 5% 5% 225 Digging, planting, wrapping and staking replaced trees
90
Total replanting costs 315
Cultural costs Orchard pruning 0 0 0 260 260 260 Herbicide 100 100 100 100 100 100 Fungicide 0 0 0 1,360 1,360 1,360 Fertilizer 80 160 242 270 430 600 Insecticide 200 200 200 200 200 200 Mowing 390 390 240 180 180 180 Irrigate and walk lines 90 90 90 90 67 67 Pest control advisor 0 0 150 150 150 150 Pollination 0 0 0 0 450 450 Rodent control (squirrels) 60 60 60 60 60 60Total cultural costs 920 1,000 1,082 2,670 3,257 3,427
Harvesting and marketing costs Picking – US$0.11/kg US$0.11 0 0 187 495 990 1,705
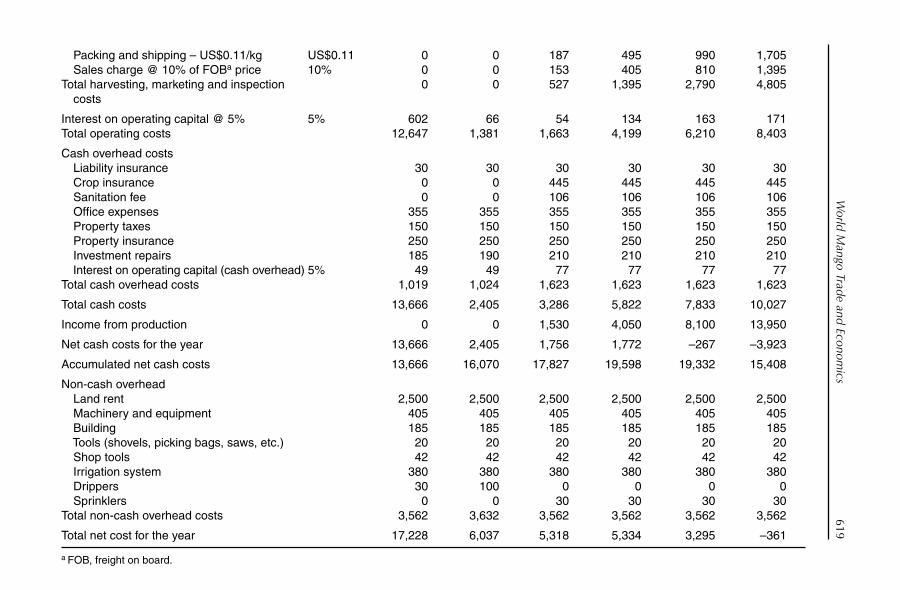
World M
ango Trade and Economics
619 Packing and shipping – US$0.11/kg US$0.11 0 0 187 495 990 1,705 Sales charge @ 10% of FOBa price 10% 0 0 153 405 810 1,395Total harvesting, marketing and inspection costs
0 0 527 1,395 2,790 4,805
Interest on operating capital @ 5% 5% 602 66 54 134 163 171Total operating costs 12,647 1,381 1,663 4,199 6,210 8,403
Cash overhead costs Liability insurance 30 30 30 30 30 30 Crop insurance 0 0 445 445 445 445 Sanitation fee 0 0 106 106 106 106 Offi ce expenses 355 355 355 355 355 355 Property taxes 150 150 150 150 150 150 Property insurance 250 250 250 250 250 250 Investment repairs 185 190 210 210 210 210 Interest on operating capital (cash overhead) 5% 49 49 77 77 77 77Total cash overhead costs 1,019 1,024 1,623 1,623 1,623 1,623
Total cash costs 13,666 2,405 3,286 5,822 7,833 10,027
Income from production 0 0 1,530 4,050 8,100 13,950
Net cash costs for the year 13,666 2,405 1,756 1,772 –267 –3,923
Accumulated net cash costs 13,666 16,070 17,827 19,598 19,332 15,408
Non-cash overhead Land rent 2,500 2,500 2,500 2,500 2,500 2,500 Machinery and equipment 405 405 405 405 405 405 Building 185 185 185 185 185 185 Tools (shovels, picking bags, saws, etc.) 20 20 20 20 20 20 Shop tools 42 42 42 42 42 42 Irrigation system 380 380 380 380 380 380 Drippers 30 100 0 0 0 0 Sprinklers 0 0 30 30 30 30Total non-cash overhead costs 3,562 3,632 3,562 3,562 3,562 3,562
Total net cost for the year 17,228 6,037 5,318 5,334 3,295 –361
a FOB, freight on board.

E.A. Evans and O.J. Mendoza620
pre-planting and planting costs are estimated at US$11,125 (US$5000 + US$6125). In year 1, it is also assumed that there are no replanting costs asso-ciated with replacement trees. However, provisions are made to accommo-date this cost in subsequent years. Here we assume that 5% of the trees planted in year 1 will be replaced in year 2. Cultural costs are then estimated, includ-ing standard operations such as pruning, fertilizing and mowing. In our example, the total cultural costs in year 1 are estimated at US$920/ha. Next are harvesting and marketing costs. This subheading includes the costs of such operations as picking, packing and shipping. Other charges include sales charges (if these operations were commissioned) and inspection and assessment fees. In both years 1 and 2, it is not expected that trees will have ‘saleable’ crops and, in fact, it is recommended that efforts should be made to defl ower such trees to prevent fruiting and to foster better tree growth. Con-sequently, there are no marketing costs in those years. To the sum for pre-planting, planting, replanting, cultural, and harvesting and marketing costs is added an interest charge of 5%. This refl ects the economic (opportunity cost) cost of money that was either borrowed or owner-fi nanced, to cover these expenses. Thus the total operating (variable) costs in year 1 is US$12,647/ha.
Overhead costs are fi xed costs (in the sense that they must be paid irre-spective of the level of production) that relate to the entire farm. For the pur-pose of budgeting they are allocated on a per hectare basis. There are two subcategories of overhead costs: cash overhead and non-cash overhead. Cash overhead costs must be paid with cash during the year. They include liability and crop insurance, taxes and offi ce expenses. Interest is charged on cash overhead costs. Total cash overhead costs plus total operating costs equals total cash costs. Total cash costs represent the total cash expended in any given year for producing and marketing a crop. In this example total cash costs for year 1 is US$13,666 (US$12,647 + US$1019)/ha. Non-cash over-head costs refl ect capital recovery associated with asset ownership is added later and discussed below.
The next major category is income from production. Mango trees will start supporting economic crops from about the third year, even though the trees are not yet fully mature. The income from the sales of these crops is deducted (credited) from the total cash costs for that year. In years 1 and 2, there is no income. However, in year 3 there is income. Based on our analysis, the income (US$1530) is then deducted from total cash costs for that year (US$3286) to give the net cash costs for the year (US$1756). The negative net cash costs recorded for year 5 (US$267) implies that, for that year, revenues from the sales of fruit exceeded total cash expenditures for that year (US$7833 − US$8100).
The accumulated net cash costs category is self-explanatory; it keeps a running tally of cash costs incurred during the establishment years. Thus, based on our analysis, the accumulated net cash costs in year 6 amounts to US$15,408/ha, refl ecting the sum of the net cash costs for years 1 through to 6, i.e. US$13,666 + US$2405 + US$1756 + US$1772 + (−US$267) + (−US$3923). This amount does not include capital recovery costs for equipment and machinery that a grower owns and that is used in establishing an orchard.

World Mango Trade and Economics 621
To complete the establishment phase budget, it is necessary to include the non-cash overhead charges. As previously defi ned, these are not cash payments. They include an estimate of the amount of money that should be set aside to help with replacing an investment item at the end of its useful life. They usually refl ect depreciation and interest charges associated with a capital item (i.e. the contribution of a capital item to the production process for a given year). If some of these services were commissioned they would be included in cash charges. The same is true for land rent. Here it is included among the non-cash overhead charges because we assumed that the grower already owns the land and therefore is not paying cash for the land. Since the land could have been sold and the proceeds placed in a bank to earn interest, the rent charged against production represents, in a sense, lost interest from not selling the land and placing the proceeds in a money-bearing bank account. In other words, it is included as payment to the owner for using his land in this manner. The sum of non-cash overhead and net cash cost equals the total net cost for that year. Table 16.7 shows that, in year 1, the total net cost is US$17,228/ha, out of which US$13,666 and US$3562 are the net cash costs and the non-cash overhead costs, respectively.
The fi nal category is total accumulated net costs, which keeps a tally of the total cost incurred since the start of the establishment phase. Based on the information provided in Table 16.7, the data reveal that, at the end of the 6 years, the investor would have invested US$36,850/ha to establish the orchard. Of this amount, the cash costs amount to US$15,408 and non-cash costs due to ownership of fi xed assets were estimated at US$21,442. Thus, the total cost to establish the 18 ha mango orchard is US$663,300 (18 × US$36,850) of which the cash amount (ignoring fi xed costs) would be US$277,344 (18 × US$15,408).
The full establishment costs of US$663,300 (US$36,850/ha) is then amortized at 5% real rate of interest for the estimated life of the orchard (25 years) to give an annual cost for capital recovery (interest and depreciation) of US$2615/ha. This amount will be charged as part of the non-cash over-head charges (Table 16.8) in computing the cost to produce a hectare of mango during the production phase (after the orchard has been established).
Discussion of production phase budget
Table 16.8 provides the estimates of the annual growing costs (after establish-ment) for mango in south Florida. The budget is similar to the one used in the establishment phase except that there are no provisions for pre-planting, planting and replanting expenses, and the estimates are for a typical year. Also of importance, and shown in italics in Table 16.8, is the amortized estab-lishment cost (US$2615/ha) obtained from Table 16.7. This refl ects the annual amount that must be recovered for the investment made in establishing an orchard.
Based on Table 16.8, it can be seen that the total annual cash cost to pro-duce a hectare of mango, assuming a yield of 22,000 kg/ha (Table 16.6) is
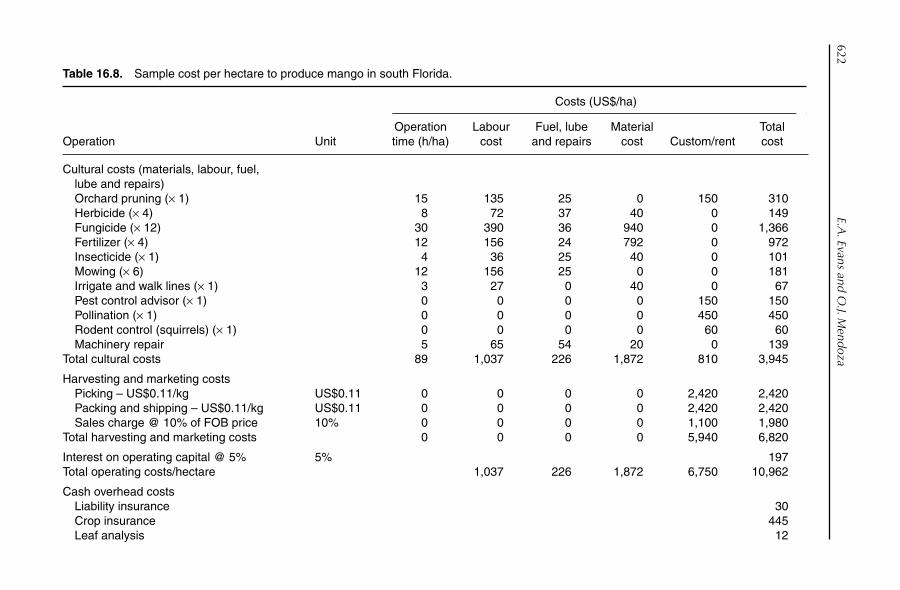
E.A. Evans and O
.J. Mendoza
622Table 16.8. Sample cost per hectare to produce mango in south Florida.
Operation Unit
Costs (US$/ha)
Operation time (h/ha)
Labourcost
Fuel, lube and repairs
Material cost Custom/rent
Total cost
Cultural costs (materials, labour, fuel, lube and repairs) Orchard pruning (× 1) 15 135 25 0 150 310 Herbicide (× 4) 8 72 37 40 0 149
Fungicide (× 12) 30 390 36 940 0 1,366 Fertilizer (× 4) 12 156 24 792 0 972 Insecticide (× 1) 4 36 25 40 0 101 Mowing (× 6) 12 156 25 0 0 181 Irrigate and walk lines (× 1) 3 27 0 40 0 67 Pest control advisor (× 1) 0 0 0 0 150 150 Pollination (× 1) 0 0 0 0 450 450 Rodent control (squirrels) (× 1) 0 0 0 0 60 60 Machinery repair 5 65 54 20 0 139Total cultural costs 89 1,037 226 1,872 810 3,945
Harvesting and marketing costs Picking – US$0.11/kg US$0.11 0 0 0 0 2,420 2,420 Packing and shipping – US$0.11/kg US$0.11 0 0 0 0 2,420 2,420 Sales charge @ 10% of FOB price 10% 0 0 0 0 1,100 1,980Total harvesting and marketing costs 0 0 0 0 5,940 6,820
Interest on operating capital @ 5% 5% 197Total operating costs/hectare 1,037 226 1,872 6,750 10,962
Cash overhead costs Liability insurance 30 Crop insurance 445 Leaf analysis 12
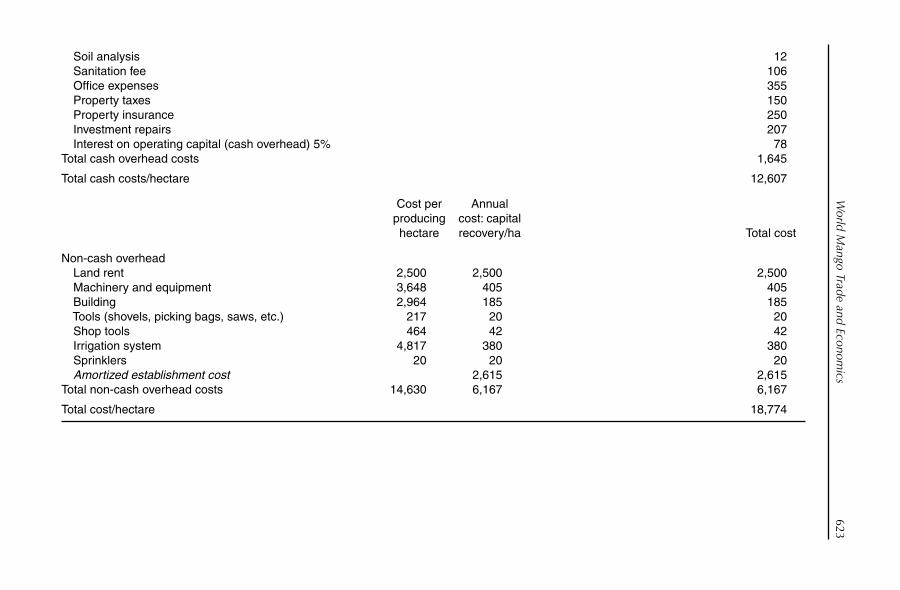
World M
ango Trade and Economics
623 Soil analysis 12 Sanitation fee 106 Offi ce expenses 355 Property taxes 150 Property insurance 250 Investment repairs 207 Interest on operating capital (cash overhead) 5% 78Total cash overhead costs 1,645
Total cash costs/hectare 12,607
Cost per producinghectare
Annual cost: capital recovery/ha Total cost
Non-cash overhead Land rent 2,500 2,500 2,500 Machinery and equipment 3,648 405 405 Building 2,964 185 185 Tools (shovels, picking bags, saws, etc.) 217 20 20 Shop tools 464 42 42 Irrigation system 4,817 380 380 Sprinklers 20 20 20
Amortized establishment cost 2,615 2,615Total non-cash overhead costs 14,630 6,167 6,167
Total cost/hectare 18,774

E.A. Evans and O.J. Mendoza624
US$12,607 or about US$0.57/kg. Of this total, cultural costs accounts for US$3945 (31%), harvesting and marketing costs amount to US$6820 (54%) and cash overhead costs US$1645 (13%). When non-cash overhead costs are added in, the economic cost to produce a hectare of mango in South Florida is estimated at US$18,774, or about US$0.85/kg.
Profi tability analysis
Table 16.9 utilizes the information presented in Tables 16.6 and 16.8 along with information obtained elsewhere to analyse the profi tability of mango production in South Florida based on 2006 data. Based on an average freight on board (FOB) price of US$0.90/kg and marketable yield of 22,000 kg/ha (actual yield would be slightly more), the gross returns are calculated as US$19,800/ha. Subtracting total operating costs of US$10,962 (cultural, and harvesting and marketing costs) from this amount gives a gross margin of US$8838/ha. In other words, based on these assumptions, the enterprise is profi table in the short run since gross margin is positive. This implies that the returns from crop sales are more than suffi cient to cover the variable costs incurred in the production and marketing of the crop and there are still funds left over to go towards paying overhead costs.
To be viable in the long run, the amount remaining should be able to cover overhead (cash and non-cash) and provide a reasonable return to the operator. Subtracting cash overhead costs (US$1645) and non-cash overhead costs (US$6167) from gross margin (US$8838) gives net returns of US$1026/ha. Viewed slightly differently, from total revenue of US$19,800 obtained from selling a hectare of the crop, after paying all costs (variable and fi xed) the amount of US$1026/ha remain as payment to management. This results in a return of approximately 7% on cash and non-cash investment.1
Given that prices and yield can vary, Table 16.10 shows a sensitivity anal-ysis for changes in price and yield variables. The sensitivity analysis is cal-culated on net returns but could be done on gross margin. As seen in Table 16.10, a 10% price increase combined with a 10% yield increase would result in over a 335% net return per hectare increase (from US$1026 to US$4460/ha). A 10% price increase would have a greater impact on net return than would a 10% yield increase. In the case of the former, the net returns would increase from US$1026/ha to US$2984/ha (191%). In the case of the latter, net returns would increase to US$2324 (127%). This underscores the importance of doing whatever is necessary to improve crop prices such as improving the quality of the product and engaging in direct marketing where possible.
Assuming marketable yields of 22,000 kg/ha, the break-even price, or the price at which net returns (economic profi ts) would be zero, is computed as US$0.85/kg. In other words, at this price the grower is just able to cover both variable and fi xed costs. Any FOB price above this would result in pos-itive net return and vice versa. Likewise, if prices were to remain at US$0.90/kg, growers would have to ensure that they obtain yields in excess of about 20,000kg/ha (break-even volume) to generate positive returns.
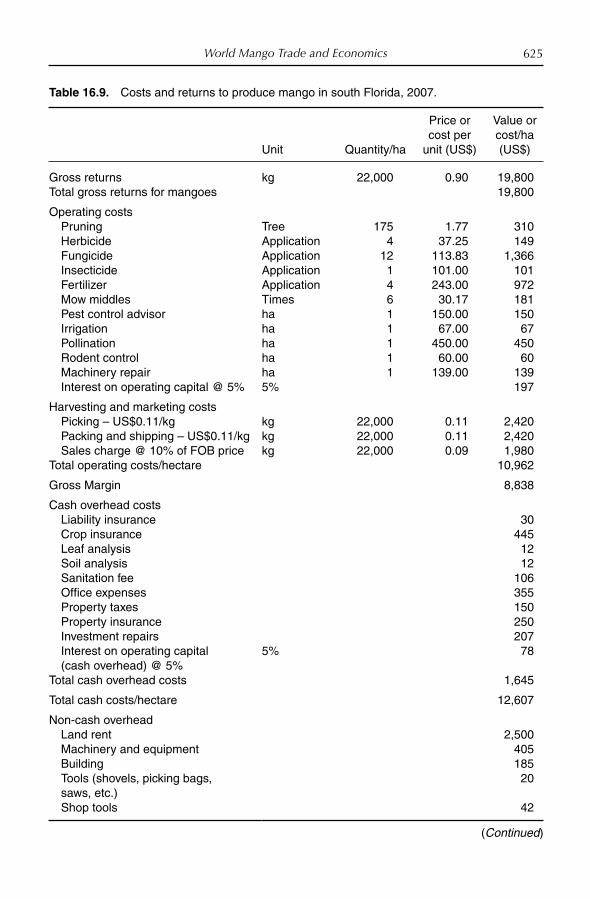
World Mango Trade and Economics 625
Table 16.9. Costs and returns to produce mango in south Florida, 2007.
Unit Quantity/ha
Price or cost per
unit (US$)
Value or cost/ha(US$)
Gross returns kg 22,000 0.90 19,800Total gross returns for mangoes 19,800
Operating costsPruning Tree 175 1.77 310Herbicide Application 4 37.25 149Fungicide Application 12 113.83 1,366Insecticide Application 1 101.00 101Fertilizer Application 4 243.00 972Mow middles Times 6 30.17 181Pest control advisor ha 1 150.00 150Irrigation ha 1 67.00 67Pollination ha 1 450.00 450Rodent control ha 1 60.00 60Machinery repair ha 1 139.00 139Interest on operating capital @ 5% 5% 197
Harvesting and marketing costsPicking – US$0.11/kg kg 22,000 0.11 2,420Packing and shipping – US$0.11/kg kg 22,000 0.11 2,420Sales charge @ 10% of FOB price kg 22,000 0.09 1,980
Total operating costs/hectare 10,962
Gross Margin 8,838
Cash overhead costsLiability insurance 30Crop insurance 445Leaf analysis 12Soil analysis 12Sanitation fee 106Offi ce expenses 355Property taxes 150Property insurance 250Investment repairs 207Interest on operating capital (cash overhead) @ 5%
5% 78
Total cash overhead costs 1,645
Total cash costs/hectare 12,607
Non-cash overheadLand rent 2,500Machinery and equipment 405Building 185Tools (shovels, picking bags, saws, etc.)
20
Shop tools 42
(Continued)
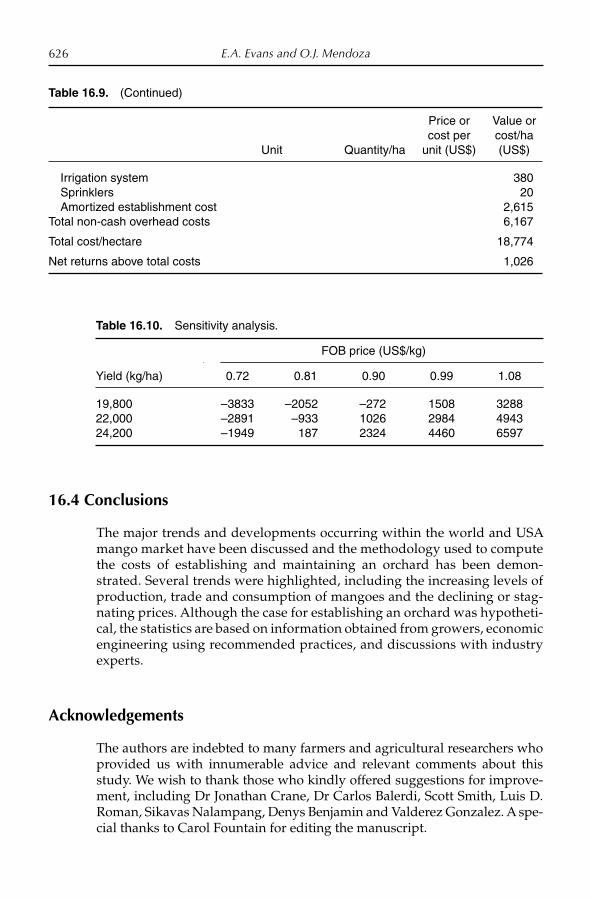
E.A. Evans and O.J. Mendoza626
16.4 Conclusions
The major trends and developments occurring within the world and USA mango market have been discussed and the methodology used to compute the costs of establishing and maintaining an orchard has been demon-strated. Several trends were highlighted, including the increasing levels of production, trade and consumption of mangoes and the declining or stag-nating prices. Although the case for establishing an orchard was hypotheti-cal, the statistics are based on information obtained from growers, economic engineering using recommended practices, and discussions with industry experts.
Acknowledgements
The authors are indebted to many farmers and agricultural researchers who provided us with innumerable advice and relevant comments about this study. We wish to thank those who kindly offered suggestions for improve-ment, including Dr Jonathan Crane, Dr Carlos Balerdi, Scott Smith, Luis D. Roman, Sikavas Nalampang, Denys Benjamin and Valderez Gonzalez. A spe-cial thanks to Carol Fountain for editing the manuscript.
Table 16.9. (Continued)
Unit Quantity/ha
Price or cost per
unit (US$)
Value or cost/ha(US$)
Irrigation system 380Sprinklers 20Amortized establishment cost 2,615
Total non-cash overhead costs 6,167
Total cost/hectare 18,774
Net returns above total costs 1,026
Table 16.10. Sensitivity analysis.
Yield (kg/ha)
FOB price (US$/kg)
0.72 0.81 0.90 0.99 1.08
19,800 –3833 –2052 –272 1508 328822,000 –2891 –933 1026 2984 494324,200 –1949 187 2324 4460 6597

World Mango Trade and Economics 627
Note
1This is obtained by adding the interests charged on operating and cash overhead costs to the net returns to give an adjusted net income. Next, subtract these interests from the total costs per hectare to give an adjusted total cost per hectare. Finally, express the adjusted net income as a percentage of the adjusted total cost per hectare.
References
FAOSTAT (2007) Food and Agriculture Organization of the United Nations database. Available at: http://faostat.fao.org/ (accessed 30 November 2007).
United States Department of Agriculture (USDA) Agricultural Marketing Service (2007) USDA, Agricultural Marketing Service: Fruit and Vegetable Market News Website. Available at: http://www.marketnews.usda.gov/portal/fv (accessed 31 March 2008).
United States Department of Agriculture (USDA) Economic Research Service (2007) Fruit and Tree Nut Yearbook, 2005. Available at: http://www.ers.usda.gov/Data/FoodConsumption/FoodAvailSpreadsheets.htm (accessed 31 March 2008).
United States Department of Agriculture (USDA) Foreign Agricultural Service (2007) USDA Foreign Agricultural Service: Market and Trade Data. Available at: http://www.fas.usda.gov/markettradedata.asp (accessed 3 November 2007).

© CAB International 2009. The Mango, 2nd Edition: Botany, Production and Uses628 (ed. R.E. Litz)
17 Fruit Processing
L.C. Raymundo, M.T. Ombico and T.M. de VillaUniversity of the Philippines Los Baños, Laguna, the Philippines
17.1 Introduction 62817.2 Dehydrated Mango Products 630
Dried mango 630Mango fruit bar 631Mango fruit roll 632Vacuum-puffed dried mango 632
17.3 Spray-dried Mango Powders 634Spray-dried mango fruit powder and instant mango juice 634Spray-dried green mango powder 635Spray-dried instant green mango shake 636
17.4 Capital Investment Costs 63717.5 Raw Material Requirements of the Mango Processing Plant 63817.6 Conclusion 639
17.1 Introduction
Processing is a value-adding step that fi lls the gap between farm production and marketing. The objective of processing is to convert highly perishable but prime quality farm commodities to more stable forms. When the process is accomplished with the least alteration in the nutritional value and aes-thetic properties of the food, high consumer acceptance is assured. Com-pletely new product lines can likewise be created out of fresh raw materials through processing. In other instances, the raw material may undergo such extensive physical alteration during processing that the product is used dif-ferently by consumers. Culinary experts devise new uses for these products to fi t the changing lifestyles of present-day consumers. The availability in the market, for example, of pre-mixed condiments and various meat powders used for fl avouring foods such as instant noodles and similar convenience

Fruit Processing 629
foods, has simplifi ed the life of working women who do not have the time or the skill to prepare food the way their mothers and grandmothers did at home. The products are affordable and easy to prepare; hence, consumer acceptance is high. In addition, these foods are found everywhere, from neighbourhood stores in the suburbs and countryside to supermarkets in urban centres. The product and process research and development work of food scientists and food technologists, the business acumen of entrepreneurs and the marketing expertise of product distributors ensure the availability of the food products.
The global competitiveness of agricultural produce of a country can be con-siderably enhanced by utilizing appropriate technologies to produce high-quality processed foods. Research focusing particularly along the areas of processing equipment, product design and packaging has made the latest techniques available for the manufacture of new generations of food prod-ucts. Tropical fruits have had an excellent record of breaking into the world market because of their exotic fl avours (Plate 83). When processed into purée, juice, nectar or simply dried fruit slices, they can be shipped long distances with only minimum changes in quality.
The popularity of fruits may be attributed to consumer perception of their health benefi ts. In the case of fruit juices, many consumers are now looking for healthy alternatives to the traditional carbonated beverages. Fruits are rich in dietary fi bre as well as phytonutrients, especially antioxidants, and have no cholesterol. The availability in the market of natural fruit juices derived from fresh fruits is a welcome alternative to synthetic juices that are being passed off as fruit juices but are, in reality, sugar-based, fruit-fl avoured beverages prepared from artifi cial ingredients.
Mango fruits are usually eaten fresh as dessert or as relish depending on their stage of maturity when picked. In the Philippines and elsewhere, fresh mangoes are available throughout the year because the fl ower-induction technology fi rst described by Barba (1974) allows growers to produce fruit out-of-season. The off-season fruits, however, are still expensive. Most mango growers have not put the technology into practice due to the high cost of additional farm inputs necessary for its successful implementation during the rainy season.
At the peak of the harvest season, on the other hand, oversupply of fresh mangoes depresses the market price to the detriment of the growers. Moreover, high temperatures combined with high relative humidity (RH) and intense sunlight during the harvest season accelerate the metabolic processes associated with ripening in fresh mangoes, rendering them sus-ceptible to microbial attack, particularly by Colletotrichum gloeosporioides Penz., the cause of anthracnose. The physico-chemical changes that occur during ripening also lead to fruit deterioration and loss of quality. Thus, fresh mangoes are processed to facilitate better distribution and to stabilize prices.
Mango processing was previously reviewed by Nanjundaswamy (1997), who described the status of processing technologies and products in India. This review discusses current trends in mango processing.

L.C. Raymundo et al.630
17.2 Dehydrated Mango Products
Dehydration works on the principle that by lowering the water (H2O) con-tent of foods below a certain threshold level, growth of many spoilage micro-organisms is prevented, thus preserving the food. As more and more H2O is removed from the material, its solids content becomes concentrated, further making the food less suitable for microbial growth.
The choice of the most appropriate drying system is determined largely by the cost-effectiveness of the process. Sun drying is the most inexpensive method for drying foods. The products, however, are rather susceptible to contamination by dirt, insects, rodents, faecal matter and microorganisms. The process also requires several days to dry each load or batch of raw mate-rials since it relies on the availability of sunlight. Temperature control is vir-tually impossible. Other systems have been designed that are more hygienic and equally cost-effective compared to sun drying. The use of solar dryers practically eliminates most of the inherent defects of sun drying. However, the reliance of solar dryers on sunlight as the source of energy for evaporat-ing H2O from the material being dried makes the system commercially unre-liable. Solar panel collector-equipped dryers with provision to store energy from sunlight can operate continuously and effi ciently.
A mechanical dryer such as the convection oven provides the processor with control over the system that is essential for a successful drying operation. It elim-inates most of the problems mentioned above. Although the initial investment cost is high due to the acquisition of the dryer, in the long run it is more eco-nomical than sun drying because the drying time per load is much shorter.
Oil-, steam-, liquid propane gas- or electric-powered air heaters are the alternative sources of energy for the dryers. Dryers have also utilized a wood-fi red furnace that heats the air entering the drying chamber. Its main advan-tage is that trimmings from raw materials, packing materials and other trash can be burned in the furnace, keeping the compound clean.
Dried mango
Dehydrated or dried mango slices are among the fi rst products manufac-tured commercially from ripe mango fruits that caught the attention of con-sumers in the international market for processed tropical fruits (Plates 83 and 84). The product was developed in Cebu, the Philippines, from where it spread to the neighbouring islands of Panay and Negros. It is now produced in many regions of the Philippine archipelago where mango is abundant. In addition, it is a popular product in Thailand and elsewhere in South and South-east Asia.
In the Philippines, the ‘Carabao’ mango is the preferred variety for dehy-dration or drying. The fruit should be at the fi rm-ripe stage. When over-ripe fruit is used as raw material, a dark-coloured product will invariably result. Although the dried pieces from over-ripe mangoes have a more distinct ripe mango fl avour that attracts customers, the shelf life is considerably shorter.

Fruit Processing 631
The fruit is washed thoroughly. The cheeks are sliced with a sharp stainless-steel knife. Each slice is then cut into two or three pieces length-wise. The fl esh is scooped from the skin with a stainless-steel scoop or ladle. These operations are done manually; however, a simple and novel peeling and slicing machine for ripe mangoes has been developed and patented in Australia (as cited by Nanjundaswamy, 1997).
Sugar syrup is prepared by adding 175 kg sugar, 1.7 kg citric acid and 0.85 kg sodium metabisulfi te in 175 l H2O to make a 45 Brix sugar concen-tration and heating to 90°C to dissolve the metabisulfi te. The prepared mango slices (1 t) are added to the syrup. The preparation is heated to 90°C and then allowed to stand for at least 6 h. Subsequently, the sugar concen-tration of the syrup is adjusted by dissolving more sugar until the concen-tration reaches 50 Brix using a hand refractometer. After 6 h, the mango pieces are again removed from the syrup and the sugar concentration is adjusted to 60 Brix using a hand refractometer for the ‘plumping’ stage. The syrup is reheated to 90°C; the mango slices are added to the syrup and soaked for a further 6 h.
The fi nal step in the process involves the removal of mango pieces from the syrup. They are lightly rinsed with H2O to remove the excess sugar that may crystallize on the surface of the mango during drying. The slices are then loaded in drying trays and dehydrated at 40–50°C in a convection dryer. Drying time usually requires 18 h. The dried mango pieces are unloaded from the dryer and reconditioned at room temperature overnight. Each piece is coated with confectioner’s sugar. The product is then packed in aluminium-foil-laminated pouches and sealed. Recent improvements in dryer design can reduce drying time to 8 h. As a result, up to three batches of prepared mango slices can be processed daily instead of only two batches every 36 h. The process for the production of dried mangoes has been described by Raymundo et al. (1999).
Mango fruit bar
The product is also commonly referred to as mango ‘leather’ (Plate 83). The processing of mango fruit bar has also been described previously by Amor-iggi (1992) and Raymundo et al. (1999, unpublished work, 2006). Purée pro-cessed from ripe ‘Carabao’ mango is the preferred raw material for the manufacture of mango fruit bar in the Philippines, although ‘Pico’ is also suitable because of the orange colour of the purée.
The total solids content of 1 t of mango purée is adjusted to 25 Brix with the use of a hand refractometer by adding sugar to the purée. The amount of sugar required depends on the initial total solids content of the mango purée. Then 2 kg each of citric acid and sodium metabisulfi te are blended into the purée. Juice of calamansi, also known as the Philippine lemon (× Citrofor-tunella microcarpa Wij. (Bunge); Citrus mitis Blanco) may be used instead of citric acid at the rate of c.20 kg per batch, although the resulting cost of the product will be slightly higher. There is no real difference in the appearance

L.C. Raymundo et al.632
and fl avour of the fi nished product. Citrus juice is generally utilized if the client prefers an all-natural product.
The prepared purée is heated for 2 min at 80°C with constant stirring to avoid scorching. The hot mixture is carefully transferred to stainless-steel drying trays. The trays are loaded into the dryer and dried for 14 h at 55°C. At the end of the drying operation, the dried sheets of mango should be pli-able and should not stick to the fi ngers when touched.
The sheets of mango are removed from the trays. Six layers of the dried mango leather are stacked on top of each other. The edges are trimmed and bars measuring 2 × 4 cm are cut from the stack. Each bar is coated with con-fectioner’s sugar, wrapped in cellophane then packed in labelled plastic bags.
Mango fruit roll
The processing of mango fruit roll has been described previously (UPLB, 1996; Raymundo et al., 1999, unpublished work, 2006). The product and pro-cess are similar to mango fruit bar, and they only differ in the presentation. The total solids content of 1 t of mango purée is adjusted to 25 Brix using a hand refractometer by adding sugar to the purée. The amount of sugar needed depends on the initial total solids content of the mango purée. Then 2 kg each of citric acid and sodium metabisulfi te are blended into the purée. As with mango fruit bar, citric acid may be replaced by calamansi juice at the rate of c.20 kg per batch without affecting the overall sensory quality of the fruit roll, if the client specifi es an all-natural product.
The prepared purée is then heated for 2 min at 80°C with constant stir-ring to avoid scorching. The hot mixture is carefully transferred to stainless-steel drying trays. The trays are loaded into a convection dryer and dried for 12–16 h at 55°C.
The dried sheets of mango are removed from the trays. Confectioner’s sugar is sprinkled over a stainless-steel-topped table to avoid sticking of the sheets on subsequent rolling. Each sheet is rolled manually into 1 cm diameter pieces. The ends are trimmed; and each roll is sliced into 5 cm long pieces. The pieces are coated with confectioner’s sugar and wrapped with cellophane. The rolls are then packed in appropriate containers.
Vacuum-puffed dried mango
The use of vacuum for puffi ng and drying mango and similar food materials is not as widespread as explosion puffi ng (Eskew et al., 1963), high tempera-ture short time (HTST) pneumatic drying and centrifugal fl uidized bed (CFB) drying (Brown et al., 1972), because the vacuum-puff dryer is still expensive. With the increasing consumer demand for high-quality processed foods and their willingness to pay a higher price for such products, vacuum-puff dehy-dration could become an economically viable investment opportunity for entrepreneurs, particularly in the mango processing industry.

Fruit Processing 633
Vacuum-puffed mango pieces will readily rehydrate due to the porosity created during the puffi ng and drying stages of the process. The signifi cantly shorter drying time (Candelaria and Raymundo, 1994b) also makes possible the drying of four loads of prepared mango slices in 24 h.
The ripe mangoes are washed thoroughly in chlorinated H2O, then sliced either mechanically or with a stainless-steel knife. The fl esh is scooped from the skin with a sharp stainless-steel ladle (Candelaria, 1993; Candelaria and Raymundo, 1994a). The fruit pieces are then heated to 90°C in 30 Brix syrup containing 1% sodium metabisulfi te, and steeped in the syrup for 4–6 h. The mango slices are removed from the syrup, rinsed briefl y in H2O, arranged in stainless-steel trays and loaded into the vacuum oven.
The mango pieces thus prepared are initially heated at a positive pres-sure of 40–50 kPa until the maximum tissue temperature of 100°C is reached, usually within 8 min. The pressure is released and the hot mango pieces are dried at −70 to −80 kPa vacuum at a temperature of 45–50°C. Total dehydration time under vacuum is 6 h. The above pressure-temperature combinations pro-vide the most desirable puff and rehydration characteristics (Candelaria and Raymundo, 1994b).
With the present technology, vacuum-puffed dried mango from a 1 t batch of prepared mango slices is more expensive to produce than convec-tion oven-dried mango (Table 17.1). The production cost needs to be reduced for the product to be market competitive. Research that focuses on devising a system to assure a continuous supply of mango fruits is required. The plant must operate on a year-round basis in order to optimize the use of its equip-ment and facilities.
The facilities can be used for the production of other vacuum-puffed fruits (i.e. bananas and muskmelon) as well as vegetables (i.e. white potato and maize kernels) (Candelaria, 1993; Candelaria and Raymundo, 1994b) among others, which can be used as raw materials in the manufacture of instant foods.
Table 17.1. A comparison of the profi tability of different mango product lines.
Product Volume (kg)Cost of goods
sold (US$)Gross profi t
(US$)Net profi t before
tax (US$)
Mango powdera 124,800 251,460 966,540 439,983Instant mango juicea 202,890 467,355 1,560,645 965,088Green mango powdera 141,221 245,226 1,166,984 600,216Instant green mango shakea 124,800 209,468 1,308,532 473,975Vacuum-puffed dried mangob 124,800 1,141,807 730,193 246,588Dried mangoc 67,392 398,492 140,644 84,386Mango fruit barc 115,200 921,600 461,816 277,090Mango fruit rollc 115,200 921,600 461,816 277,090
a US$10/kg powder.b US$15/kg.c US$8/kg.

L.C. Raymundo et al.634
17.3 Spray-dried Mango Powders
Spray-drying is a process in which a liquid feed is fi nely dispersed or atomized to form droplets, which are eventually sprayed into a heated air chamber. The process facilitates the rapid evaporation of H2O from the feed droplets, thereby forming the powder particles. The product obtained using the technology is a free-fl owing powder that may or may not be instantly soluble in H2O, depend-ing on the formulation of the liquid feed that has been used. By modifying the formulation and adjusting the process parameters, a plain powder is obtained that can be dry-mixed with sugar, modifi ed starches or similar components. The powder is used for fl avouring confectioneries and pharmaceutical prepa-rations as well as in the manufacture of baby foods and tropical fruit drinks fortifi ed with nutrients to replace those portions lost during processing.
Tropical fruit juice powders that are rapidly soluble in H2O are produced directly by spray-drying fruit juices and purées. By dry-mixing spray-dried plain fruit powder with sweeteners, a ready-to-drink juice is made. The latter method has the added advantage in that it allows formulation of exclusive blends of fruit drinks. As a result, consumers have a wider range of products from which to choose that will suit their individual preferences.
By converting fruit pulps into powder or by instantizing them, their shelf life is prolonged. Consequently, this value-adding step simplifi es exportation since many of the restrictions normally imposed on fresh produce by import-ing countries are offset by the process. Transportation cost is also reduced by at least 85%, which refl ects the amount of H2O removed from the fresh juice. Instant juices are more convenient for consumers since they can be reconsti-tuted easily. In the pharmaceutical and cosmetics industries, there is a large demand for natural tropical fruit powders as fl avouring and colouring agents to add to their usual line of fruit-fl avoured products. Powders are also more convenient to handle during the manufacturing process.
Spray-drying is by far the most cost-effective method for transforming fruit pulps into powder. Fruit pulp is very heat-sensitive and requires special treat-ments to produce competitively priced powders of superior quality. Further-more, because of the rapid drying cycle and simplicity of operation, continuous production is achieved which contributes to the low operational cost. The short holding time of the powder inside the drying chamber reduces the risks of pow-der burn. Human contact with the liquid feed and powder is also minimized as a result of the short holding time. The processes are, therefore, very hygienic and the product is ready for packaging as it leaves the dryer. Unlike other drying sys-tems, including convection oven-drying and drum-drying, there is no need to purchase a hammer mill for grinding the dry fl akes into a powder. The main drawback of spray-drying, however, is the relatively high initial investment cost.
Spray-dried mango fruit powder and instant mango juice
Khalid and Sial (1974), Anonymous (1998) and Diaz (2000) have discussed the recovery of fruit powder and production of instant mango juice powder

Fruit Processing 635
using the technology. Both products use mango purée as the raw material. They differ only in the composition of the liquid feed. The liquid feed is mango purée with the total solids adjusted to the right consistency, thereby allowing the purée to be discharged through a high-speed nozzle in the form of fi ne droplets into the drying chamber that quickly dries to a yellow free-fl owing powder. The patent application for the manufacture of spray-dried mango powders is still pending at the Philippine Patent Offi ce. The process parameters used, therefore, cannot be discussed in detail in this chapter. The parameters used are the same as reported previously, namely, the inlet tem-perature, the feed rate, air speed and the outlet temperature (Welti and Lafl u-ente, 1983; Liang and King, 1991; Anonymous, 1998). The powder of instant mango juice is comparatively dense so that it gets wet easily, enabling it to sink immediately during reconstitution. The plain mango fruit powder, on the other hand, does not disperse as readily; it has a tendency to clump on the H2O surface. However, it will dissolve quickly in hot H2O or with the use of an ordinary household blender.
A modest capacity spray-drying plant equipped with a NIRO SD 12.5R model spray-dryer made in Denmark, valued at approximately US$207,860, with an evaporative capacity of 25 kg H2O/h will require an initial invest-ment of about US$313,860 to make it operational. In addition, if fresh man-goes are be used as raw materials, a purée processing facility will cost around US$98,000. The fi gure represents expenses for buying and installation of equipment, and does not include the costs of the building, land and ancillary expenses such as environmental pollution control system and spray-dryer accessories.
At the evaporative capacity rate of 25 kg H2O/h, the facility can produce 124.8 t/year of plain mango powder and 202.8 t/year of instant mango juice at a total cost of US$251,460 and US$467,355, respectively (Table 17.1). The net profi t for plain mango powder is US$439,983 and US$965,088 for instant mango juice before taxes.
Spray-dried green mango powder
The purée of green mangoes can be converted to a powder (UPLB, 2005) just like the purée of ripe mangoes by spray-drying (Plate 85a). The spray-dried powder can be mixed with other condiments and used as a souring agent for exotic or native dishes, or as the raw material in the manufacture of instant green mango shake (see below). The powder may be dry-mixed with sugar, powdered honey, caramel powder or powdered sugar syrup to instantize it. During this process, the mixture can be fortifi ed with vitamins (e.g. ascorbic acid) and other nutrients.
The estimated cost of goods sold using a commercial spray-dryer with an evaporative capacity of 25 kg H2O/h is US$245,226/year (Table 17.1). At the rated capacity of the spray-drying plant of 141,221 kg/year, the net profi t before taxes for green mango fruit powder is US$600,216 at a selling price of US$10/kg of powder.

L.C. Raymundo et al.636
Spray-dried instant green mango shake
Green mango shake on cracked ice is a very popular thirst quencher in South-east Asia and elsewhere (Plate 85b). Its origin can be traced back to the coun-tryside where fi nely diced fresh green mango pieces are mixed with H2O, sugar and ice to make a cheap, wholesome summer drink during the mango season. The practice has since been modifi ed and upgraded to cater to upscale domestic markets and abroad.
The beverage is an excellent source of vitamin C. When the green mango purée is spray-dried, a free-fl owing white to greenish-white powder is pro-duced that will dissolve instantly even in cold H2O. The powdering process offers unprecedented convenience to the consumer, especially when the pow-der is packed in sachets. Since no artifi cial or synthetic colours and fl avour-ing agents are included in the liquid-feed formulation, the natural taste and aroma of green mango is retained in the powder.
Using a commercial spray-dryer with an evaporative capacity of 25 kg H2O/h, the estimated cost of goods sold is US$209,468/year. At the rated capacity of spray-drying plant of 124,800 kg, the net profi t before taxes for instant green mango shake is US$473,975/year at a selling price of US$10/kg of powder (Table 17.1).
The technology for the production of instant green mango shake and green mango powder was developed at UPLB (2005). If the proper feed for-mulation and process parameters are applied, spray-drying is an effi cient and hygienic method for producing cheap but high-quality mango fruit powder and instant mango juice. Table 17.2 demonstrates that the spray-dried mango powders have much lower microbial load, i.e. 2.8–10.4 × 102 colony forming units (cfu)/g, compared to the industry standard of 1 × 104 to 5 × 105 cfu/g total plate count. Except for mango powder, the mould and yeast counts are within the limit of 1 × 102 cfu/g. Locally refi ned sugar and imported modifi ed starch, on the other hand, have much higher mould and yeast counts than the Heinz standard for powdered products (Shapton
Table 17.2. Microbial load of raw materials and spray-dried mango fruit powder and instant mango juice (cfu/g).
Material Total plate count Mould count Yeast count
Mango powder 2.8 × 102 2 × 102 <10Instant mango juice 3.6 × 102 0.4 × 102 <10Green mango powder 10.4 × 102 0.95 × 102 <10Instant green mango shake 9.8 × 102 0.85 × 102 1 × 102
Mango purée 0.5 × 102 0.2 × 102 <10Green mango purée 0.8 × 102 0.1 × 102 <10Sugar, refi ned 11 × 102 43 × 102 20 × 102
Modifi ed starch 63 × 102 7.5 × 102 5 × 102
H.J. Heinz standard (Shapton and Shapton, 1991)
1 × 104 to 5 × 105 1 × 102 1 × 102

Fruit Processing 637
and Shapton, 1991) at 11 × 102 and 63 × 102 cfu/g total plate count, respec-tively. These materials are essential ingredients of the liquid-feed formula-tion used for the production of the spray-dried powders.
The formulation and processing of numerous mango products popu-lar in South Asian countries were reported previously (Nanjundaswamy, 1997).
17.4 Capital Investment Costs
An initial investment of approximately US$100,000 is needed for the estab-lishment of a mango dehydration plant. The facility can also be used for dry-ing other farm commodities such as vegetables, spices and other high-value crops. The cost of acquiring the land, building construction and ancillary costs such as environmental pollution abatement as well as quality assurance laboratory facilities are not included in the estimate.
A commercial dryer such as that manufactured by Tsung Hsing Food Machinery Co. Ltd of Taiwan with a loading capacity of 500–600 kg prepared mango slices costs c.US$11,000. At the rated capacity of the plant of 1 t per batch, 67,392 kg of dried mango pieces can be produced annually. The cost of goods sold is US$398,492 with a net profi t before taxes of US$84,386/year. If the facility is used for the manufacture of mango fruit bar and mango fruit roll, the annual production is 115,200 kg of either fruit bar or fruit roll with a net profi t before taxes of US$277,090/year (Table 17.1).
The total cost of equipment for the vacuum-puffi ng plant with a loading capacity of 1 t is approximately US$122,000, broken down as US$100,000 for fabrication of the dryer and US$22,000 for other plant equipment. The total capitalization, which includes the cost of raw materials, salaries and equip-ment per year, is US$648,240 excluding interest on capital.
Spray-dryers come in various capacities ranging from NIRO SD Micro Spray-dryer ideal for powdering small quantities of raw materials for research and development to units that produce powders at evaporative rates in excess of 500 kg H2O/h. The investment cost for equipment increases with the capac-ity of the dryer (Table 17.3). For a small-scale operation producing powders or fl ours from commercial purée, the initial investment is US$182,000 for a 5 kg H2O evaporative capacity dryer while a plant equipped with a 12 kg evapora-tive capacity dryer requires US$262,000 for equipment. On the other hand, an investment of US$313,860 is needed to equip a medium-scale drying plant with a 25 kg H2O/h evaporative capacity such as NIRO SD 12.5R.
The estimates do not include land, building and construction costs as well as the cost of pollution statement facilities. The cost of land is dependent on where the plant will be located while building and construction costs are determined by the capacity of the spray-dryer unit to be acquired.
Total capitalization is US$317,200, US$535,947 and US$806,265 for the 5 kg, 12 kg and 25 kg H2O/h evaporative capacity units, respectively. The estimate includes the cost of raw materials, salaries and equipment per annum. Interest on capital is not included.
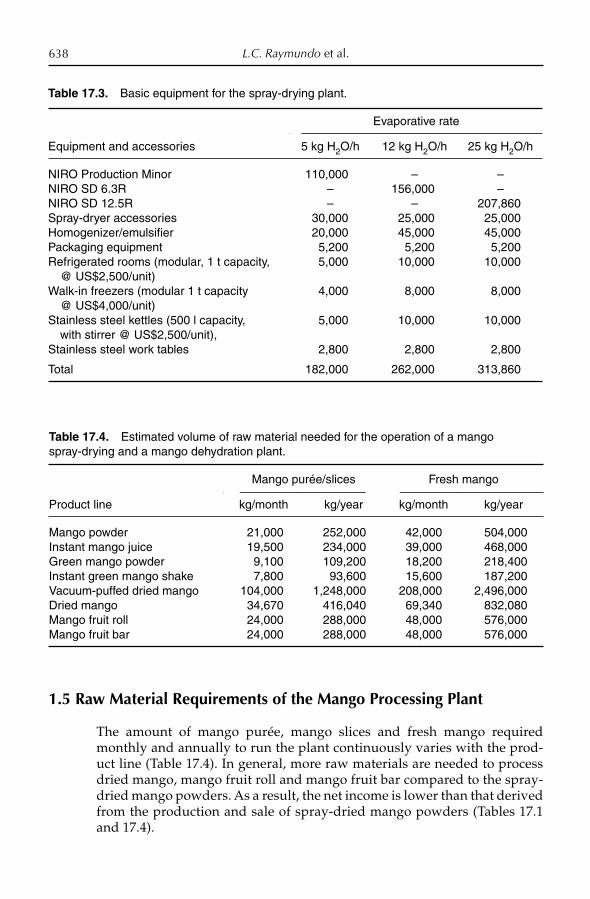
L.C. Raymundo et al.638
1.5 Raw Material Requirements of the Mango Processing Plant
The amount of mango purée, mango slices and fresh mango required monthly and annually to run the plant continuously varies with the prod-uct line (Table 17.4). In general, more raw materials are needed to process dried mango, mango fruit roll and mango fruit bar compared to the spray-dried mango powders. As a result, the net income is lower than that derived from the production and sale of spray-dried mango powders (Tables 17.1 and 17.4).
Table 17.3. Basic equipment for the spray-drying plant.
Equipment and accessories
Evaporative rate
5 kg H2O/h 12 kg H2O/h 25 kg H2O/h
NIRO Production Minor 110,000 – –NIRO SD 6.3R – 156,000 –NIRO SD 12.5R – – 207,860Spray-dryer accessories 30,000 25,000 25,000Homogenizer/emulsifi er 20,000 45,000 45,000Packaging equipment 5,200 5,200 5,200Refrigerated rooms (modular, 1 t capacity, @ US$2,500/unit)
5,000 10,000 10,000
Walk-in freezers (modular 1 t capacity @ US$4,000/unit)
4,000 8,000 8,000
Stainless steel kettles (500 l capacity, with stirrer @ US$2,500/unit),
5,000 10,000 10,000
Stainless steel work tables 2,800 2,800 2,800
Total 182,000 262,000 313,860
Table 17.4. Estimated volume of raw material needed for the operation of a mango spray-drying and a mango dehydration plant.
Product line
Mango purée/slices Fresh mango
kg/month kg/year kg/month kg/year
Mango powder 21,000 252,000 42,000 504,000Instant mango juice 19,500 234,000 39,000 468,000Green mango powder 9,100 109,200 18,200 218,400Instant green mango shake 7,800 93,600 15,600 187,200Vacuum-puffed dried mango 104,000 1,248,000 208,000 2,496,000Dried mango 34,670 416,040 69,340 832,080Mango fruit roll 24,000 288,000 48,000 576,000Mango fruit bar 24,000 288,000 48,000 576,000

Fruit Processing 639
The spray-drying facility requires 187.2–218.4 t/year of green mangoes and 468–504 t/year of ripe mango fruits at the evaporative capacity of 25 kg H2O/h to produce the volume of powders in Table 17.1. On the other hand, 576 t/year of ripe fruits are required to produce mango fruit roll or fruit bar. For dried mango, 832 t/year of fresh fruits are needed to produce 67.4 t/year of the product while to produce 124.8 t/year of vacuum-puffed dried mango, 2496 t/year of fresh fruits are necessary.
In terms of area, c.38–48 ha can supply the green mango requirement of the spray-drying facility. Approximately 100 ha are needed to produce 124.8 t of ripe mango powder or 202.9 t of instant mango juice from fresh ripe mango fruits. The fresh fruit requirement for the manufacture of fruit roll and fruit bar can be supplied by 115 ha of mangoes. The 832 t and 2496 t of fresh fruit are equivalent to the annual harvest from 166 ha and 500 ha of mangoes to produce 67.4 t and 124.8 t of dried mango and vacuum-puffed dried mango, respectively. The estimates are based on a yield of 5 t from 100 trees/ha as well as the biennial fruiting habit of mango.
17.6 Conclusion
Processing of mango is a profi table business venture once economies of scale are attained, i.e. when the processor has the proper proportions of raw materials, labour and machinery to meet a given market demand. Fresh mangoes are now available year-round. But the supply is still insuffi cient to satisfy the demand by the fresh fruit market and the mango-processing sector. It is, therefore, essential that a farming system for mango be designed in order to minimize the cost of production of off-season fruits and ensure the sustainable operation of mango processing facilities. Since raw material sourcing is the primary cause of the dif-fi culties encountered by processors of dried mangoes, mango in syrup and similar products, growers need more assistance for them to adopt the tech-nology for off-season mango fl ower induction. The problem is not as serious for product lines utilizing mango purée as starting material since the purée can be processed during the peak of the harvest season and held in cold stor-age for later use. Once the system is in place, the processed mango industry is expected to develop further and become a major revenue generator.
In the near future, with the advances in the fi eld of genetic engineering, it may be possible to eliminate the biennial fruiting habit of many current mango cultivars or, at the very least, minimize its infl uence on the perfor-mance of the crop. Species of Mangifera and some non-cultivated, wild types of mango can be the source of desirable traits that may be incorporated in the next generation of mango cultivars, such as their innate ability to bear fruits during the rainy season (see Bompard, Chapter 2, this volume).
The technologies for processing mangoes are readily available. Others are being developed in research laboratories to cope with the changing needs of consumers. The main problem is to ensure a continuous supply of high-quality fresh mango fruit in order to produce the prime quality commodities that consumers expect from the industry.

L.C. Raymundo et al.640
References
Amoriggi, G. (1992) The marvelous mango bar. Ceres 24, 25–28.Anonymous (1998) Instruction Manual for Spray Drying Plant. NIRO A/S Soeborg, Denmark.Barba, R.C. (1974) Induction of fl owering of the mango by chemical spray. Proceedings
of the Crop Science Society of the Philippines 5, 154–160.Brown, G.E., Farkas, D.F. and De Marchena, E.S. (1972) Centrifugal fl uidized bed.
Blanches, dryers and puff piece-form foods. Food Technology 26, 23–30.Candelaria, N.M. (1993) Dehydration of vacuum-puffed fruits and vegetables. PhD dis-
sertation, University of the Philippines Los Baños, Laguna, the Philippines.Candelaria, N.M. and Raymundo, L.C. (1994a) Comparative drying and reconstitution,
characteristics of some fruits and vegetables. Philippine Agriculturist 77, 321–326.Candelaria, N.M. and Raymundo, L.C. (1994b) Vacuum puffi ng and dehydration of fruits
and vegetables. Philippine Agriculturist 77, 251–260.Diaz, J.V. (2000) Development of spray-dried instant mango (Mangifera indica L.) juice pow-
der. MSc thesis, University of the Philippines Los Baños, Laguna, the Philippines.Eskew, R.K., Cording, J., Jr and Sullivan, J.F. (1963) A gun for explosive, puffi ng of dehy-
drated fruits and vegetables. Food Engineering 35, 91–96.Khalid, M.A. and Sial, M.B. (1974) Spray-drying of mango juice powder. Mesopotamia
Journal of Agriculture 9, 47–56.Liang, B. and King, L.J. (1991) Factors infl uencing fl ow patterns, temperature and conse-
quence drying rates in spray-drying. Drying Technology 9, 1–27.Nanjundaswamy, A.M. (1997) Processing. In: Litz, R.E. (ed.) The Mango: Botany, Pro-
duction and Uses. CABI International, Wallingford, UK, pp. 507–542.Raymundo, L.C., Ombico, M.T. and De Villa, T.M. (1999) Establishment of integrated
mango processing plant. In: Namuco, L.O. and Andam, C.J. (eds) Mango Production Manual. Philippine Council for Agriculture, Forestry and Natural Resources Research and Development (PCARRD), Los Baños, Laguna, the Philippines, pp. 97–119.
Raymundo, L.C., Ombico, M.T., De Villa, T.M. and Jaen, R.M. (2006) Processes of Fruits, Nuts, Vegetables and Other Tropical Foods. Fruit and Vegetable Laboratory, Food Science Cluster, University of the Philippines Los Baños, Laguna, the Philippines. (Unpublished)
Shapton, D.A. and Shapton, N.F. (1991) Principles and Practices for the Safe Processing of Foods. Butterworth-Heinemann Ltd, Oxford, UK.
University of the Philippines Los Baños (UPLB) (1996) Standardization of Processing Method and Evaluation of Different Packaging Material and Storage Conditions for Mango Roll. Annual Report, Institute of Food Science and Technology, College of Agriculture, University of the Philippines Los Baños, Laguna, the Philippines.
University of the Philippines Los Baños (UPLB) (2005) Spray-dried Instant Juice and Powder from Green Mango. Annual Report, Food Science Cluster, College of Agri-culture, University of the Philippines Los Baños, Laguna, the Philippines.
Welti, J.S. and Lafl uente, B. (1983) Spray-drying of comminuted orange products. I. In-fl uence of air and temperature and feed rate on product quality. Revista de Agro-nomica y Technologia de Alementos 23, 97–106.

© CAB International 2009. The Mango, 2nd Edition: Botany, Production and Uses(ed. R.E. Litz) 641
18 Biotechnology
R.E. Litz,1 M.A. Gómez-Lim2 and U. Lavi31University of Florida, Florida, USA
2CINVESTAV, Irapuato, Mexico3Agricultural Research Organization, Bet Dagan, Israel
18.1 Introduction 64118.2 Cell and tissue culture 643
Organogenesis 643Somatic embryogenesis 643Protoplast isolation and culture 650Potential for other regeneration pathways 651
18.3 Molecular Breeding and Genetics 651Marker assisted selection (MAS) 651Gene cloning 651Genomics 655
18.4 Genetic Engineering 656In vitro induced mutations 656Genetic transformation 659
18.5 In vitro conservation 660Medium-term storage 660Long-term storage 661
18.6 Conclusions 662
18.1 Introduction
Improvement of monoembryonic mangoes by selection of superior seedling trees resulting from open pollinations dates from approximately 500 years ago (the late 1500s), when the Mogul emperor Akbar established the Lakh Bagh, a garden of seedling mango trees near Dabangha in Bihar (see Mukher-jee and Litz, Chapter 1, this volume). Only a few years before this time, the Portuguese had introduced grafting techniques into India, and the superior selection mango trees within the Lakh Bagh were vegetatively propagated and named. Among these early mango selections were ‘Alphonso’, ‘Dashe-hari’, ‘Langra’, etc. All subsequently named mango selections have been

R.E. Litz et al.642
derived by identifying seedling trees within populations either from within uncontrolled open pollinations or, less commonly, among controlled polli-nated progenies.
Mango cultivar improvement programmes currently exist in several countries (see Iyer and Schnell, Chapter 4, this volume), and they address signifi cant production problems that have a genetic basis. Classical breeding of mango has obvious limitations, which include, the long juvenile period of mango trees (7 or more years), the low frequency of fruit set following con-trolled pollination, the period required for seedling trees to be evaluated for fruit production, tree architecture, and the cost of maintaining large popu-lations of seedling trees in order to observe segregation of important hor-ticultural traits. There is no single ideotype for mango; however, the most important attributes for scion cultivars must include: compact size, resistance to anthracnose and other limiting diseases, fruit production (which would include annual bearing and factors that affect fruit quality, i.e. shape, colour, fl avour and size). In many traditional mango-producing countries, consumer preference for mangoes has been quite conservative, and has been resistant to the introduction of new cultivars. Although there is a demand for improved clonal rootstocks, there are few breeding programmes that address this need. For mango rootstocks, the most important traits include tolerance of abiotic stress, polyembryony and the ability to limit vegetative growth of the scion (i.e. dwarfi ng).
Biotechnology refers to the application of molecular biology and somatic cell genetics to the improvement of plants. Biotechnology can resolve some of the most serious production problems of important mango cultivars and improve breeding methodologies. Genomic studies will ultimately associate genes with specifi c functions, and this will impact genetic engineering and molecular breeding of mango. Marker assisted selection (MAS) would facili-tate the screening of seedling populations for important horticultural traits. Genetic engineering would permit the targeting and alteration of specifi c horticultural traits in existing cultivars, without altering the integrity of clones. Mango improvement by modern genetics will be freed from the constraints of the lengthy juvenile period of the species and the additional years required for tree evaluation. Moreover, the effi cient management of mango plant genetic resources should be greatly facilitated within the next decade by advances that have been made in molecular biology, cell culture and cryo-preservation.
Mango biotechnology has been variously reviewed since 2002 (Litz and Gomez-Lim, 2002, 2005; Litz, 2003, 2004, 2008; Gomez-Lim and Litz, 2007; Krishna and Singh, 2007). In this chapter, we have addressed the current sta-tus of mango genomics, gene cloning and cell culture. Within the discussion of cell culture, the following areas of research are addressed: (i) the utiliza-tion and potential of cell culture for improving existing mango cultivars by invitro-induced mutation followed by selection and genetic transformation; and (ii) advances in medium- and long-term conservation of genetic resources. The model species Arabidopsis thaliana has provided a useful tool for studying the regulation of gene expression of important plant processes, for example

Biotechnology 643
embryo and organ development, including shoots, leaves, roots, fl owers and fruit. Genomic analysis and the identifi cation of horticulturally important genes that control these and other processes will ultimately enable us to under-stand and control many aspects of mango fruit production (i.e. disease and pest control, fruit quality, tree architecture, etc.). The development of molecu-lar approaches will also facilitate mango breeding through the identifi cation of DNA markers for important traits.
This chapter is a discussion of the current state of mango cell culture, gene cloning and the manipulation of cell cultures to address plant breeding objectives by genetic engineering.
18.2 Cell and Tissue Culture
Organogenesis
The earliest report of morphogenesis from mango cell cultures involved the differentiation of adventitious roots from callus that was initiated from mango cotyledons on Murashige and Skoog (1962) semi-solid induction medium (MS) that was supplemented with kinetin and naphthaleneacetic acid (NAA) (Rao et al., 1982). Of greater signifi cance was the later report that callus initiated from leaves from mature mango trees also had morphogenic potential. Raghuvanshi and Srivastava (1995) induced caulogenic cultures from fully expanded, young leaves of monoembryonic ‘Amrapali’. Disin-fested leaf pieces were initially explanted into liquid MS medium supple-mented with the antioxidant 0.05% polyvinylpyrolidone with 2 h subcultures for up to 24 h on a rotary shaker at 75 rpm. Leaf pieces were then transferred onto MS semi-solid medium supplemented with 13.0 M kinetin and 1.1 M indole acetic acid (IAA), the optimum growth regulator formulation for mul-tiple shoot development from leaf callus. Subculture of individual shoots onto semi-solid MS medium supplemented with 9.8 M indole butyric acid (IBA) stimulated root induction and development. Cultures were main-tained at 25°C with a 16 h photoperiod provided by cool white fl uorescent lights (50–70 mol/m2/s).
The major advantage of the organogenic pathway for regenerating mango is that cultures can be induced whenever there is a leaf fl ush; however, genetic manipulation studies require much higher rates of regeneration than has been hitherto reported.
Somatic embryogenesis
Genetic engineering of mango has been based upon the effi cient recovery of somatic embryos from embryogenic cultures. Effi cient regeneration of woody plants from cell cultures derived from mature phase materials is often diffi cult to achieve, and the optimum procedure generally must be determined empir-ically. Mangifera indica consists of two ecogeographic races: a monoembryonic

R.E. Litz et al.644
group that probably originated in north-eastern India and northern Myan-mar and a polyembryonic group that originated in South-east Asia. The pres-ence of nucellar embryos in seeds of polyembryonic mangoes demonstrates that cells of the nucellus of the species have morphogenic potential (see Mukherjee and Litz, Chapter 1, this volume). Sturrock (1968) reported that polyembryony in mango appeared to be inherited as a recessive trait; how-ever, Aron et al. (1998) have demonstrated that polyembryony is under the control of a single dominant gene. Somatic embryogenesis, which is the equiv-alent morphogenic response in vitro, is a dominant trait in several other spe-cies, for example lucerne (Reisch and Bingham, 1980) and orchardgrass (Gavin et al., 1989), although in red clover it apparently is conferred by a reces-sive gene (Broda, 1984). The presence of morphogenically competent cells in the nucellus is critical for the induction of embryogenic mango cultures of both monoembryonic and polyembryonic mango cultivars.
Induction of embryogenic mango cultures from the excised nucellus of immature mango seeds of polyembryonic and monoembryonic cultivars was fi rst described by Litz et al. (1982) and Litz (1984), respectively. The current standard protocol for induction and maintenance of embryogenic mango cultures and for development of somatic embryos is derived from DeWald et al. (1989a, b) and Litz et al. (1993). Table 18.1 provides citations for all invitro studies that have involved somatic embryogenesis of mango. The efforts in this fi eld have expanded considerably since the late 1990s.
It is impossible to predict the embryogenic response of the nucellus of different mango cultivars, irrespective of the seed type (i.e. polyembryonic or monoembryonic). Early reports suggested that the nucellus of polyembry-onic mangoes responded in vitro more readily than the nucellus of monoem-bryonic mangoes (Litz, 1986); however, this assumption no longer is considered to be valid. Litz et al. (1997) reported that the induction of embryogenic com-petence in the explanted nucellus of monoembryonic ‘Tommy Atkins’ can be inhibited by the ethylene antagonist aminoethoxyvinylglycine (AVG) and by dicyclohexylammonium sulfate (DCHA), an inhibitor of spermidine synthe-sis. In contrast, induction of embryogenic competence of the explanted nucel-lus of polyembryonic ‘Tuehau’ is unaffected by either AVG or DCHA. Litz and Schaffer (1987) demonstrated that somatic embryogenesis in mango is partially mediated by spermidine. The biosynthesis of ethylene and/or the sensitivity of nucellar tissue to ethylene may be important factors for the induction of embryogenic cultures from this tissue.
InductionThe induction of embryogenic competence is associated with the develop-mental stage of the nucellus at the time of explanting, and also can be af-fected by the physiological status of the tree (Litz, 1987). Fruits that are approximately 30–40 days after pollination contain seeds in which the nucel-lus is at the ideal stage for explanting: relatively thick and intact and easily removed. The embryo (mass) in fruit and developing seed of this age is usu-ally no more than half the length of the immature seed. Mango fruit of the appro-priate stage of development are surface-disinfested with 20–30% (v/v) domestic

Biotechnology 645
bleach containing Tween 20 for 30 min. The sterilant is rinsed from the fruit with three changes of sterile deionized or distilled water, and each fruit is bisected along its longitudinal axis without damaging the immature seed under axenic conditions. The immature seed is removed from the bisected fruit, which is also bisected carefully along its longitudinal axis. Manza-nilla Ramirez et al. (2000) obtained optimum induction with polyembryonic ‘Ataulfo’, monoembryonic ‘Tommy Atkins’ and monoembryonic ‘Haden’ nucellar explants when the embryo (mass) to immature seed ratio was 1:3. The zygotic embryo (of a monoembryonic cultivar) or polyembryonic mass (of a polyembryonic cultivar) is excised and discarded. The nucellus can be carefully peeled from the interior of the seedcoat using a sterile, fl at spatula. Following the transfer of the nucellus onto induction medium in sterile Petri dishes, the cultures are incubated in darkness at 25°C. Thereafter, it is essen-tial to subculture the nucellar explants onto fresh induction medium at daily intervals until the oxidation of the explant ceases; oxidation is associated with darkening of the medium around the explanted nucellus.
A summary of those mango cultivars that have been successfully estab-lished as embryogenic cultures is provided in Table 18.2. The effi ciency of
Table 18.1. Summary of published reports on somatic embryogenesis of mango from nucellar explants.
Country Reference
Colombia Flórez-Ramos et al. (2007)India Ara et al. (1998, 2000a, b)
Chaturvedi et al. (2004)Deore et al. (2000)Jana et al. (1994)Krishna and Singh (2007)Laxmi et al. (1999)Singh et al. (2001, 2002)Sulekha and Rajmohan (2003)Thomas (1999)
Mexico Manzanilla Ramirez et al. (2000)Rivera Domínguez et al. (2004)
Philippines Patena et al. (2002)USA DeWald et al. (1989a, b)
Lad et al. (1997)Litz (1984)Litz and Gomez-Lim (2005)Litz and Lavi (1997)Litz and Schaffer (1987)Litz and Vijayakumar (1988)Litz et al. (1982, 1983, 1984, 1995, 1997, 1998)Mathews et al. (1992)Monsalud et al. (1995)Pliego Alfaro et al. (1996b)
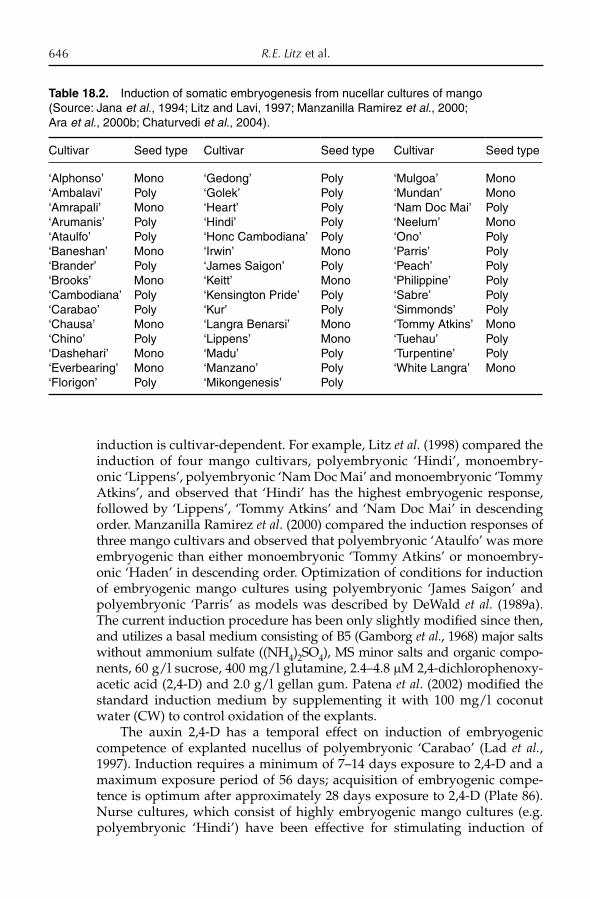
R.E. Litz et al.646
induction is cultivar-dependent. For example, Litz et al. (1998) compared the induction of four mango cultivars, polyembryonic ‘Hindi’, monoembry-onic ‘Lippens’, polyembryonic ‘Nam Doc Mai’ and monoembryonic ‘Tommy Atkins’, and observed that ‘Hindi’ has the highest embryogenic response, followed by ‘Lippens’, ‘Tommy Atkins’ and ‘Nam Doc Mai’ in descending order. Manzanilla Ramirez et al. (2000) compared the induction responses of three mango cultivars and observed that polyembryonic ‘Ataulfo’ was more embryogenic than either monoembryonic ‘Tommy Atkins’ or monoembry-onic ‘Haden’ in descending order. Optimization of conditions for induction of embryogenic mango cultures using polyembryonic ‘James Saigon’ and polyembryonic ‘Parris’ as models was described by DeWald et al. (1989a). The current induction procedure has been only slightly modifi ed since then, and utilizes a basal medium consisting of B5 (Gamborg et al., 1968) major salts without ammonium sulfate ((NH4)2SO4), MS minor salts and organic compo-nents, 60 g/l sucrose, 400 mg/l glutamine, 2.4–4.8 M 2,4-dichlorophenoxy-acetic acid (2,4-D) and 2.0 g/l gellan gum. Patena et al. (2002) modifi ed the standard induction medium by supplementing it with 100 mg/l coconut water (CW) to control oxidation of the explants.
The auxin 2,4-D has a temporal effect on induction of embryogenic competence of explanted nucellus of polyembryonic ‘Carabao’ (Lad et al., 1997). Induction requires a minimum of 7–14 days exposure to 2,4-D and a maximum exposure period of 56 days; acquisition of embryogenic compe-tence is optimum after approximately 28 days exposure to 2,4-D (Plate 86). Nurse cultures, which consist of highly embryogenic mango cultures (e.g. polyembryonic ‘Hindi’) have been effective for stimulating induction of
Table 18.2. Induction of somatic embryogenesis from nucellar cultures of mango (Source: Jana et al., 1994; Litz and Lavi, 1997; Manzanilla Ramirez et al., 2000; Ara et al., 2000b; Chaturvedi et al., 2004).
Cultivar Seed type Cultivar Seed type Cultivar Seed type
‘Alphonso’ Mono ‘Gedong’ Poly ‘Mulgoa’ Mono‘Ambalavi’ Poly ‘Golek’ Poly ‘Mundan’ Mono‘Amrapali’ Mono ‘Heart’ Poly ‘Nam Doc Mai’ Poly‘Arumanis’ Poly ‘Hindi’ Poly ‘Neelum’ Mono‘Ataulfo’ Poly ‘Honc Cambodiana’ Poly ‘Ono’ Poly‘Baneshan’ Mono ‘Irwin’ Mono ‘Parris’ Poly‘Brander’ Poly ‘James Saigon’ Poly ‘Peach’ Poly‘Brooks’ Mono ‘Keitt’ Mono ‘Philippine’ Poly‘Cambodiana’ Poly ‘Kensington Pride’ Poly ‘Sabre’ Poly‘Carabao’ Poly ‘Kur’ Poly ‘Simmonds’ Poly‘Chausa’ Mono ‘Langra Benarsi’ Mono ‘Tommy Atkins’ Mono‘Chino’ Poly ‘Lippens’ Mono ‘Tuehau’ Poly‘Dashehari’ Mono ‘Madu’ Poly ‘Turpentine’ Poly‘Everbearing’ Mono ‘Manzano’ Poly ‘White Langra’ Mono‘Florigon’ Poly ‘Mikongenesis’ Poly

Biotechnology 647
embryogenic competence from the nucellus of cultivars that are normally diffi cult to induce, for example polyembryonic ‘Nam Doc Mai’ (Litz et al., 1998). The nurse culture procedure involves explanting the nucellus onto sterile fi lter paper which has been moistened with induction medium and which overlays the highly embryogenic mango culture (polyembryonic ‘Hindi’) on semi-sterile induction medium. It is not clear if a nucellar callus is initiated from the explant prior to acquisition of embryogenic competence; however, somatic embryos can develop directly from the nucellus without an intermediate callus (Litz, 1987). Embryogenic nucellar cultures are recog-nizable ap-proximately 30 days after explanting; they are completely orga-nized, and consist of proembryonal somatic embryos, embryogenic cells, cell aggregates and proembryonic masses (PEMs) (Litz et al., 1993, 1995; Litz and Lavi, 1997).
MaintenanceEmbryogenic mango cultures are friable and cream to light brown in colour, although the cultures rapidly darken on semi-solid medium, and must therefore be subcultured at 3–4 week intervals. PEMs develop from globular somatic embryos in the presence of the primary induction agent, 2,4-D. The PEMs originate as globular somatic embryos, but their devel-opment as individual somatic embryos is arrested in the presence of 2,4-D. The PEMs increase in diameter with cells of the protoderm dividing rap-idly; secondary globular somatic embryos develop from these proliferating embryogenic cells. This highly repetitive pattern of somatic embryogen-esis in the presence of 2,4-D is the basis for maintenance of embryogenic cultures.
Embryogenic mango cultures can be maintained as proliferating PEMs either on semi-solid or in liquid induction medium. Maintenance of embryo-genic cultures of many cultivars is optimal in liquid induction medium sup-plemented with 4.8 M 2,4-D (Litz et al., 1984; DeWald et al., 1989a) (Plate 87); however, rapid proliferation of embryogenic suspension cultures is cultivar-dependent (Litz et al., 1993). Embryogenic suspension cultures are initiated by inoculating approximately 400 mg of PEMs into sterile 80 ml maintenance medium in 250 ml Erlenmeyer fl asks (or 200 mg of PEMs into 40 ml mainte-nance medium in 125 ml Erlenmeyer fl asks). The fl asks are maintained on a rotary shaker at 100–125 rpm in semi-darkness at 25°C with regular transfers of PEMs into fresh medium at 10–14 day intervals. Regular subculturing is essential to prevent loss of morphogenic potential and darkening of the tis-sue. The typical embryogenic suspension culture consists of PEMs, embryo-genic cells and multicellular complexes.
MaturationDevelopment of somatic embryos from embryogenic cultures maintained on semi-solid maintenance medium occurs sporadically and without synchroni-zation, due to the lack of direct contact of parts of a culture with medium containing 2,4-D. Exposure to 2,4-D is necessary for embryogenic culture proliferation, while at the same time, somatic embryo development is

R.E. Litz et al.648
inhibited. In order to stimulate somatic embryo development, embryogenic cultures must be transferred from maintenance medium to medium without 2,4-D in order to initiate effi cient somatic embryo development. For embryo-genic suspension cultures, the PEMs are decanted through sterile fi ltration fabric with a 1000 m opening size. The larger fraction (>1000 m diameter) is re-inoculated into liquid maintenance medium for continued proliferation and the smaller fraction is transferred either into liquid medium or onto semi-solid medium without 2,4-D in order to arrest repetitive somatic embryogenesis and to initiate somatic embryo development (Plate 88).
The plant growth media and conditions for stimulating somatic embryo development and maturation are based upon the protocol described by DeWald et al. (1989b) with minor alterations. Initially, a medium consisting of B5 major salts, MS minor salts and organic components, 60 g/l sucrose and 400 mg/l glutamine with or without 2.0 g/l gellan gum is utilized. Different mango cultivars require different periods for cotyledon differentiation following subculture to maturation medium (Litz et al., 1993). Either 4.65 M kinetin or 4.44 M benzyladenine (BA) can stimulate the development of cotyledons and the apical meristem and reduce the maturation period. The cultures are incubated in darkness at 25°C.
When growth of embryogenic cultures of highly responsive cultivars is optimized as suspension cultures, the early cotyledonary somatic embryos that develop in liquid medium are hyperhydric. Mathews et al. (1992) and Monsalud et al. (1995) demonstrated that hyperhydric somatic embryos can-not develop to maturity, and become necrotic. Hyperhydricity of mango somatic embryos can be reversed either by partially desiccating heart stage embryos (2–3 mm length) under high relative humidity (RH) for 24 h or by plating them on maturation medium solidifi ed with 6.0 g/l gellan gum (Monsalud et al., 1995). Reversion of hyperhydricity can result in precocious germination of mango somatic embryos, although this can be prevented if 500 M abscisic acid (ABA) is included in the modifi ed maturation medium.
Embryogenic cultures of different mango cultivars vary in their response to subculture from liquid maintenance medium onto maturation medium, and this must be determined empirically for each cultivar. For example, poly-embryonic ‘Hindi’ somatic embryo development (cotyledon differentiation) from embryogenic suspension cultures follows a step-wise procedure. The <1000 m diameter fraction from liquid maintenance is subcultured into liquid maturation medium until the early heart stage of somatic embryo development is apparent. Subsequently, the heart stage somatic embryos are subcultured onto semi-solid maturation medium. In contrast, the <1000 m fraction of monoembryonic ‘Keitt’ and polyembryonic ‘Carabao’ embryogenic suspension cultures can be transferred directly from liquid maintenance medium onto semi-solid maturation medium.
Mango zygotic embryos require approximately 4–5 months in order to develop to maturity in vivo, and mature embryos can be >6–8 cm long (Plate 89). Consequently, the plant growth media that have been formulated to stimulate growth and development of mango somatic embryos from the heart stage to maturity refl ect the differing requirements of these enlarging

Biotechnology 649
somatic embryos. The medium that has been utilized for development of mango somatic embryos to maturity consists of B5 major salts, MS minor salts and organic components, 400 mg/l glutamine, 20% (v/v) fi lter-sterilized CW, 40 g/l sucrose and 2.0 g/l gellan gum (DeWald et al., 1989b). As the somatic embryos enlarge, the sucrose concentration of maturation medium is gradually reduced to 10 g/l.
During somatic embryo development certain developmental anomalies become apparent, of which the most frequently observed are polycotyly, fas-ciation, absence of bipolarity, secondary somatic embryogenesis from the hypocotyl and precocious germination. Polycotyly and fasciation do not affect subsequent plant development; however, failure to address problems associated with absence of bipolarity, secondary embryogenesis and preco-cious germination can seriously impact the recovery of plants. Control of precocious germination of developing embryos is occasionally necessary, since immature somatic embryos that germinate before they are physiologi-cally mature cannot survive. Some of these developmental anomalies can be eliminated by maintaining relatively high sucrose concentrations and/or by incorporating 100 M ABA in the maturation medium (Monsalud et al., 1995; Pliego-Alfaro et al., 1996b). The cultures are maintained in darkness at 25°C during somatic embryo maturation.
GerminationWhen somatic embryos begin to germinate, they are fi nally transferred to light conditions. The radicle elongates, followed by growth of the taproot. The shoot apical meristem remains quiescent for approximately 2 weeks after germination, at which time the shoot elongates. Although many mango somatic embryos germinate under these conditions, their survival or conver-sion ex vitro is low, primarily due to apical shoot necrosis, a physiological disorder that is associated with calcium ion (Ca++) defi ciency. Different strat-egies have been attempted to improve the conversion rate (i.e. survival of somatic embryo-derived plants):
The period for embryogenic cultures in/on maintenance medium should 1. be minimal (Litz and Lavi, 1997).
Ara 2. et al. (1998) rescued in vitro microshoots obtained from germinated somatic embryos by pulsing them for 24 h with 24.6 M IBA in liquid medi-um followed by transfer to auxin-free medium in darkness for root develop-ment.
Somatic embryo shoots have also been rescued by micrografting the 3. shoots on decapitated in vitro-germinated seedling rootstocks (Plate 90).
Enhanced recovery of mango plantlets can occur following the induction 4. of photoautotropism by transfer of small plantlets onto minimal plant growth medium, containing <5% sucrose and 1% (w/v) activated charcoal (Litz et al., 1993). A fi lter-sterilized air mixture consisting of 20,000 ppm carbon dioxide (CO2) in a nitrogen gas carrier is introduced into the growing containers, and the plantlets are exposed to a 16 h photoperiod at 180 mol/s/m2 provided by cool white fl uorescent tubes.

R.E. Litz et al.650
Ara 5. et al. (1999) reported improved conversion following the encapsulation of early heart stage somatic embryos in calcium alginate-containing modifi ed standard mango medium with half-strength major salts and supplemented with 2.9 M gibberellic acid (GA3). The germination of encapsulated somatic embryos was almost 75% greater than non-encapsulated somatic embryos.
Protoplast isolation and culture
The isolation, culture and regeneration of plantlets from protoplasts isolated from embryogenic suspension cultures of monoembryonic ‘Amrapali’ has been described (Ara et al., 2000a). One gram of embryogenic culture was transferred from a 3–4-week-old suspension into 10 ml fi lter-sterilized growth medium consisting of B5 major salts, MS minor salts and organic components, supple-mented with 0.3 M sucrose, 0.4 M mannitol, 0.1 M sorbitol, 2.74 mM glutamine, 1.0% cellulase, 1.0% hemicellulase and 0.5% pectinase (Sigma) with gentle shak-ing in darkness at 25°C for 24 h. The digestion mixture was then passed through a sieve (50 m) in order to remove debris, and then centrifuged for 5 min at 100 g. The supernatant was discarded, and cell debris was resuspended and precipitated by centrifugation three times at 3 min for each centrifugation cycle. The pellet was fi nally resuspended in 1 ml medium, and layered on 3 ml sucrose solution (25% w/v) and centrifuged at 100 g for 7 min. Protoplasts were removed and cultured in maintenance medium, but modifi ed to contain 0.18 M sucrose, 2.74 mM glutamine and 4.5 M 2,4-D. Somatic embryos developed from PEMs following subculture onto somatic embryo development medium (without 2,4-D), and plantlets were recovered using standard procedures (see Somatic embryogenesis section in 18.2 Cell and tissue culture, this chapter).
Protoplast technology has not been utilized at this time for mango improvement, and the likelihood of its exploitation is diffi cult to predict. The sexual compatibilities of Mangifera spp. with the common mango are unknown. Many newly described Mangifera species tolerate stressful environmental conditions and have pest and disease resistance (see Bompard, Chapter 2, this volume); however, it is uncertain if they are isolated genetically from mango. Somatic hybridization might enable the genetic recombination of the common mango with some of the Mangifera species as a means for develop-ing new rootstocks. This approach, however, would almost certainly have little utility for scion development, since the common mango is a tetraploid, and the ploidy level of many of the Mangifera spp. is unknown. Somatic hybridization would result in hexaploids or octoploids, and introgression of useful traits into the common mango would be very diffi cult.
Potential for other regeneration pathways
The recovery of embryogenic haploid cultures from microspores has been described for several perennial fruit tree species, including Annona squamosa (Nair et al., 1983), Citrus aurantifolia (Chaturvedi and Sharma, 1985), Citrus

Biotechnology 651
microcarpa (Chen et al., 1980), Dimocarpus longan (Yang and Wei, 1984) and Litchi chinensis (Fu and Tang, 1983). Mangifera indica is thought to be an auto-tetraploid (2n = 4x = 40), and recovery of diploid (n = 2x = 20) plants would reveal the nature of the ancestral species and simplify genomic analysis. Chromosome doubling would restore autotetraploidy, and somatic hybrid-ization of diploid mangoes with wild Mangifera species would allow interest-ing genetic recombination.
18.3 Molecular Breeding and Genetics
The identifi cation and use of molecular markers for distinguishing mango from other Mangifera spp. and for identifying different mango cultivars has been discussed by Bompard, Chapter 2, this volume and by Iyer and Schnell, Chapter 4, this volume.
Marker assisted selection (MAS)
MAS offers great potential for improvement of quantitative traits in crop plants. There are clear advantages for the use of molecular markers in plant breeding, such as a decreased number of breeding generations, the availabil-ity of a uniform method for scoring, no need to use phenotypic scoring until the end and, fi nally, the possibility for obtaining information on the percent-age of genome contributed by each parent in the offspring. Although molecu-lar markers have been used for taxonomic purposes with mango (Schnell et al., 1995; López-Valenzuela et al., 1997; Eiadthong et al., 2000; Ravishankar et al., 2000; etc.) mango has not been the subject of MAS. It is notable that although mango has 40 chromosomes, it has a comparatively small haploid genome size (0.91 pg), which is only three times as large as the recently sequenced genome of A. thaliana (the plant with the smallest genome size known), about half that of tomato and comparable to that of rice (Arumuganathan and Earle, 1991). Clearly, the comparatively small mango genome would facilitate the identifi -cation of molecular markers and the creation of a genetic map.
Gene cloning
Genetic transformation of mango cultures involves the transfer of genes to manipulate specifi c processes. Genetic manipulation of any crop requires that relevant genes must be available. Mango has been the subject of molecu-lar research for several years, and several genes have been identifi ed.
Early studies showed that changes in mRNA and protein content occur during fruit ripening (López-Gómez and Gómez-Lim, 1992; Chaimanee et al., 1999). The level of a number of mRNAs (as assayed by in vitro translation) changes throughout the ripening process. This method for detecting changes in mRNAs has low sensitivity and, therefore, only the most abundant proteins

R.E. Litz et al.652
can be detected. For that reason, the molecular analysis of fruit ripening requires construction of a gene library. In mango, cDNA libraries have been constructed, mainly from ripe fruit, and screened using several approaches. The mRNA for virtually all the ripening-related genes isolated so far have been shown to be absent or at a low level in immature fruit, increase during ripening and decline as ripening progresses (Giovannoni, 2001). Unlike other fruits, none of the identifi ed genes in mango code for enzymes involved in the ripening process itself (see below).
Mango fruit are highly perishable commodities due to over-ripening, which is mainly caused by the sharp increase in ethylene production, which occurs simultaneously with the climacteric peak (Tucker and Grierson, 1987; see Brecht and Yahia, Chapter 14, this volume). Mangoes have poor storage quality and storage in controlled or modifi ed atmospheres has been associ-ated with physiological disorders (Chaplin, 1989). Genetic manipulation of mango fruit ripening represents an attractive alternative to extend storage life and, therefore, the isolation of mango genes coding for enzymes involved in ethylene biosynthesis has been a target for research.
The two key enzymes in the ethylene biosynthetic pathway are those catalysing the conversion of S-adenosylmethionine (SAM) to 1-aminocyclo-propane-1-carboxylic acid (ACC) and ACC to ethylene, i.e. ACC synthase and ACC oxidase or ethylene forming enzyme (EFE), respectively and cDNA clones coding for mango ACC synthase and ACC oxidase have been identi-fi ed (Gómez-Lim, 1993). Their expression during ripening was studied in pulp and peel in northern blot analysis-type experiments. The ACC synthase message is undetectable in unripe fruit and starts to appear in turning fruit, reaching a maximum in ripe fruit (Gómez-Lim, 1993). This pattern of expres-sion is similar in the peel and in the pulp; however, the message appears in the pulp before the peel. The ACC oxidase message shows similar kinetics in both types of tissue, but the message is clearly detectable before any ACC synthase message becomes detectable (Gómez-Lim, 1993). These results sug-gest that ACC oxidase is expressed before ACC synthase and that ripening starts on the inside of mango fruit and proceeds outwards. Ethylene-treated mango fruits show a different pattern of expression, with ACC oxidase and ACC synthase appearing initially in the peel.
If ethylene is being actively produced, the gas must be clearly perceived by plant cells and therefore ethylene sensitivity is important for the ripening process. Major advances in understanding ethylene signal transduction have come from a molecular genetic approach using ethylene responsive mutants of A. thaliana and tomato. Several genes coding for ethylene receptor homo-logues have been isolated from various plants (Johnson and Ecker, 1998). Genetic manipulation of the ethylene receptor represents an interesting alter-native to control ethylene production and delay ripening, particularly in those fruits where other alternatives, such as storage in controlled atmo-spheres, have not been effective (Wilkinson et al., 1997). A cDNA coding for a mango ethylene receptor has been isolated (Gutiérrez-Martinez et al., 2001). The message seems to be present at low levels in unripe fruit and to increase

Biotechnology 653
as the fruit ripens. Mechanical wounding appears to up-regulate the expres-sion of the receptor.
Several studies have convincingly shown that the profi le of released volatiles changes as ripening proceeds (Olle et al., 1998; Sakho et al., 1998; Ansari et al., 1999; Saby John et al., 1999; Andrade et al., 2000; Bender et al., 2000). It is likely that many of these compounds, most of which are terpe-noids, are determinants of fl avour and aroma and many of them originated from the metabolism of fatty acids via the -oxidation pathway. It is known that several fatty acids, particularly linoleic and oleic acids, decrease in con-centration during ripening. In this sense, a cDNA for thiolase, an enzyme from the -oxidation pathway, was identifi ed as having a ripening-specifi c pattern and to be up-regulated during ripening (Bojórquez and Gómez-Lim, 1995). A cDNA for acyl CoA oxidase, the key enzyme in the -oxidation path-way, has isolated from mango fruit and shown to behave similarly to thiolase (A. Nila-Mendez and M. A. Gómez-Lim, Irapuato, unpublished data). These enzymes might be involved in metabolism of fatty acids to produce volatile compounds. The contents of sugars and organic acids can infl uence the fl a-vour properties of mango (Malundo et al., 2001).
The manipulation of fruit aroma and fl avour is a long established research goal and, accordingly, the isolation of genes coding for enzymes involved in biosynthesis of these compounds has been targeted. The gene coding for alcohol acyl transferase, an enzyme presumably involved in the synthesis of compounds implicated in fruit fl avour, has been identifi ed in mango (GB: AX025510; patent WO 0032789-A 36).
Alternate oxidase is involved in the cyanide-resistant respiratory path-way. It has been studied mainly in thermogenic species, and its activity is correlated with heat production, necessary to volatilize foul-smelling com-pounds to attract insect pollinators. There is a signifi cant participation of this pathway in the climacteric of many fruit. A cDNA coding for mango alter-nate oxidase has been isolated and the message was detected by Northern blot analysis in unripe fruit and shown to increase substantially in ripe fruit (Cruz-Hernandez and Gómez-Lim, 1995). These results were correlated with similar increases in enzyme activity and protein accumulation. The tempera-ture in ripe monoembryonic ‘Alfonso’ fruit is up to 10°C higher than in unripe fruit and this has been attributed to the activity of alternate oxidase (Kumar et al., 1990). This extra heat might also serve to volatilize aroma-giving compounds. The results with alternate oxidase were confi rmed by Considine et al. (2001), who isolated several members of the multigene family of mango alternate oxidase and showed that they were expressed differen-tially during ripening. They also identifi ed a gene coding for an uncoupling protein whose mRNA peaked at the turning stage whereas the protein peaked at the ripe stage (Considine et al., 2001). They suggested a role for alternate oxidase and the uncoupling protein in post-climacteric senescence. Because both mRNA and protein for alternate oxidase and the uncoupling protein increased in a similar pattern, they hypothesized that their expression is con-trolled simultaneously.

R.E. Litz et al.654
Ripening of fruit involves a number of metabolic reactions, including synthesis and turnover of the plant cell wall. In mango, fruit ripening is char-acterized by a gradual softening, which is caused by progressive depolymer-ization of pectic and hemicellulosic polysaccharides with signifi cant loss of galactose, arabinose and mannose residues at the ripe stage (Yashoda et al., 2005). This depolymerization is caused by the activity of different hydrolytic enzymes that are secreted as ripening proceeds (i.e. polygalacturonase and
-galactosidase), which are present in different isoforms in mango pulp (Prasanna et al., 2005, 2006). Cell wall-metabolizing enzymes from mango, i.e. exopolygalacturonase (Chaimanee et al., 2000), endo-polygalacturonase (Sun-tornwat et al., 2000), -galactosidase (S. Parra-Arenas and M. A. Gómez-Lim, Irapuato, unpublished data) and pectate lyase (Chourasia et al., 2006), have been cloned and correlate closely with fruit ripening. Furthermore, an ethyl-ene-responsive, ripening-related expansin has also been identifi ed as being closely correlated with softening (Sane et al., 2005).
A cDNA clone coding for a homologue of the YPT/Rab class of small Guanosine-5’-triphosphate (GTP)-binding proteins has been identifi ed from mango and is induced during ripening (Zainal et al., 1996). These proteins appear to control the secretion of other proteins as well as the fusion of mem-branes in animal cells (Fischer von Mollard et al., 1994). Some of these small GTP-binding proteins have homologues in plants, albeit their role has not been well defi ned (Staehelin and Moore, 1995). It is tempting to speculate that, based on the pattern of expression, these small GTP-binding proteins facilitate secretion of various hydrolytic enzymes during fruit ripening.
Saiprasad et al. (2004) isolated fi ve ripening-related cDNAs by reverse transcription-polymerase chain reaction (RT-PCR). They showed similarity to PRl-1 protein, a transcription initiation factor, a CCR-4 protein and to an 18S ribosomal RNA gene and a 23S ribosomal RNA gene. Likewise, Lycett et al. (1997) identifi ed by differential screening six clones up-regulated dur-ing ripening. Unfortunately, none of these clones code for proteins directly associated with ripening. Only two of them showed homology to a plastid chromatin and Ypt/Rab11 class of small GTPase, respectively. I A tomato rab11-like cDNA was used for tomato transformation in antisense (Lu et al., 2001). The fruits changed colour as expected but failed to soften normally. This was accompanied by reduced levels of two cell wall hydrolases, pectin-esterase and polygalacturonase. There were other phenotypic effects in the plants, including determinate growth, reduced apical dominance, branched infl orescences, abnormal fl oral structure and ectopic shoots on the leaves. In some plants, ethylene production was reduced. These data suggest an additional role for the gene in other ripening processes.
Vasanthaiah et al. (2006) reported the characterization of 27 differentially expressed genes during mango internal breakdown and found catalase, superoxide dismutase, ubiquitin, alcohol dehydrogenase, coproporphyrino-gen oxidase and keratin-associated protein to be up-regulated; whereas, several ribosomal genes, cysthathionine gamma synthase and fructose bispho-sphate aldolase were down-regulated. The increased expression of catalase, coproporhyrinogen III oxidase and keratin genes during internal breakdown

Biotechnology 655
was interpreted as a sign of oxidative stress, which they hypothesized as one of the probable causes for this disorder.
In addition to these genes, several mango sequences have been reported in the GenBank. These include a genomic sequence for the large subunit of ribulose 1,5-bisphosphate carboxylase (U39269), cDNAs for two unidentifi ed clones (AF370123 and AF061639), a partial cDNA for LFY (AY189684), a cDNA for xyloglucan endo-transglycosylase (GenBank accession AY600965) and a putative Pto-like serine/threonine kinase gene (AY693369). The function of these sequences in mango development remains to be determined.
There have been several reports on therapeutic properties of extracts from mango leaves or seeds, including anti-infl ammatory effects (Beltrán et al., 2004; Garrido et al., 2004; Leiro et al., 2004), anthelminthic and antial-lergic properties (García et al., 2003a), anti-diarrhoeal properties (Sairam et al., 2003) and even an enhancement of the humoral immune response in mouse (Garcia et al., 2003b). It is likely that the constituents responsible for these therapeutic effects will be the subject of intense scrutiny and, if the right genetic elements are identifi ed, the genetic manipulation of the biosyn-thetic route may become a reality.
Genomics
The human genome project has been the catalyst for the development of sev-eral high-throughput technologies that have made it possible to map and sequence complex genomes. Many bacterial genomes and the genomes of Sacchromyces cerevisiae, Caenorhabditis elegans, Drosophila melanogaster, A. thali-ana, Oriza sativa and Populus trichocarpa have been fully sequenced. In addition, the National Center for Biotechnology Information Entrez Genome Projects web site reports that sequencing of several more plant genomes is in prog-ress. Nevertheless, the completion of the entire genomic sequence of a par-ticular organism represents the end of the structural genomics segment of the project. It is clear, therefore, that the identifi cation of every gene within the genomes of model organisms is only the initial step to understand what these genes do and how they interact to make up a living organism. Understand-ing the functions of the 20,000–50,000 genes comprising plant (and animal) genomes and the variations within a population and roles in normal devel-opment will represent a possibly greater task than the mapping and sequenc-ing efforts currently underway.
Understanding the function of genes and other parts of the genome is known as functional genomics, and research has involved model organisms such as A. thaliana and rice. Model organisms offer a cost-effective way to follow the inheritance of genes through many generations in a relatively short time. Functional genomics is characterized by high throughput or large-scale experimental methodologies combined with statistical and com-putational analysis of the results. The fundamental strategy in a functional genomics approach is to expand the scope of biological investigation from studying single genes or proteins to studying all genes or proteins at once in

R.E. Litz et al.656
a systematic fashion. Computational biology will perform a critical and expanding role in this area. Whereas structural genomics has been character-ized by data management, functional genomics will be characterized by min-ing the data sets for particularly valuable information. Functional genomics promises to rapidly narrow the gap between sequence and function and to yield new insights into the behaviour of biological systems. The essential requirement for the implementation of functional genomics is the availability of identifi ed sequences, which can be subsequently used in studies such as high-density arrays. Currently, there are very few sequenced genes from mango, and these are mainly fruit-specifi c, and an effort to increase this num-ber is needed. Considering the cDNA libraries prepared from ripe fruit in various laboratories around the world, an effort to determine expressed sequence tags (ESTs) would be worthwhile. At the same time, more cDNA libraries from tissues other than fruit are needed. At present, the technology for successful application of functional genomics to mango is well devel-oped, but the ‘raw material’ (i.e. identifi ed genes) is lacking. Nevertheless, over 1000 ESTs from ripe mango pulp have been sequenced and are in the process of being analysed by microarrays using RNA probes from different ripening stages and from different mango cultivars (G. Ramirez Zavala and M.A. Gómez-Lim, Irapuato, unpublished results).
On the other hand, proteomics, the large-scale study of protein structure and function, is still to be applied to mango. Proteomics is often considered the next step after genomics but in fact it is more challenging, because a pro-teome differs from cell to cell and constantly changes through its biochemical interactions with the genome and the environment. The Human Genome Project has revealed that there are far fewer protein-coding genes in the human genome than proteins in the human proteome (20,000–25,000 genes versus >500,000 proteins). The protein diversity is thought to be due to alter-native splicing and post-translational modifi cation of proteins and protein degradation (Reddy, 2007). Clearly, this discrepancy implies that protein diversity cannot be fully characterized by gene expression analysis alone. Thus proteomics may be more useful for characterizing cells and tissues.
18.4 Genetic Engineering
In vitro induced mutations
There is signifi cant consumer resistance to replacement of local mango cultivars by newer selections in many traditional mango-producing countries of South and South-east Asia. Serious production and postharvest problems that have a genetic cause (e.g. alternate and irregular bearing, susceptibility to anthra-cnose and mango malformation, tree shape and size, etc.) cannot be addressed in the long term using applied physiology. For example, paclobutrazol has been applied to alternate-bearing mango cultivars to promote fl owering in the off-years; however, this can result in severe decline in trees that have been treated for successive years. As a result, conventional mango breeding in

Biotechnology 657
India has focused upon the development of mango cultivars that are largely indistinguishable from traditional selections with respect to fruit size, appear-ance, taste, fl avour and overall quality.
Despite its impact on crop breeding (Maluszynski, 2001) and particularly on banana improvement (Novak et al., 1990), mutation breeding has not been successfully exploited for mango cultivar improvement by production of use-ful off-types of existing selections. There is some anecdotal evidence that somatic mutations can occur naturally in mango on the basis of variation that occurs within seed-propagated polyembryonic cultivars. There are reported to be marked phenotypic differences within polyembryonic ‘Kensington Pride’ trees in Australia, and among polyembryonic cultivars of South-east Asia (e.g. ‘Arumanis’, ‘Golek’, etc.). Different phenotypes of polyembryonic ‘Aru-manis’, for example, have been characterized as ‘Arumanis-1’, ‘Arumanis-2’, etc. Gan et al. (1981) and Litz et al. (1993) described isozyme variation within populations of South-east Asian polyembryonic mangoes.
The most important production and postharvest problem of mango in the humid tropics and subtropics is anthracnose, caused by Colletotrichumgloeosporiodes (Penz.) Penz. and Sacc. In Penz. The current strategies for con-trol of this disease involve the use of moderately resistant cultivars (i.e. monoembryonic ‘Calypso’, monoembryonic ‘Keitt’ and monoembryonic ‘Tommy Atkins’, etc.) and at least weekly applications of fungicides (i.e. benomyl or maneb or mancozeb) from the time of fl owering until harvesting (Dodd et al., 1997). This can result in as many as 25 spray applications in a season, and is considered an increasingly unsustainable agricultural practice.
The effect of γ-irradiation on embryogenic cultures of polyembryonic ‘Hindi’, monoembryonic ‘Keitt’ and monoembryonic ‘Tommy Atkins’ was reported by Litz (2001) as part of the Food and Agriculture Organization (FAO)/International Atomic Energy Agency (IAEA) Co-ordinated Research Project entitled Improvement of Tropical and Subtropical Fruit Trees through Induced Mutations and Biotechnology. Embryogenic mango cultures on semi-solid maintenance medium were exposed to 0–200 Gy irradiation provided by 60Co. The lethal dose for 50% mortality (LD50) for each cultivar was deter-mined: it was approximately 125 Gy for ‘Keitt’ and approximately 100 Gy for ‘Tommy Atkins’. The LD50 of embryogenic ‘Hindi’ cultures could not be established within this dosage range, possibly due to its high rate of prolif-eration relative to ‘Keitt’ and ‘Tommy Atkins’. This study was confi rmed by Manzanilla Ramirez et al. (2000). The main objective of these studies has been to develop appropriate technologies for utilizing induced mutations in vitro for recovery of useful off-types of existing cultivars.
Culture fi ltrates produced by pathogenic fungi and bacteria can be uti-lized not only to select for resistance to the pathogen in vitro (Hammerschlag, 1992), but also to induce the host resistance response. In order for in vitro selection to be used effectively, essential prerequisites include: (i) a highly morphogenic suspension culture; and (ii) evidence that the culture fi ltrate or phytotoxin can produce symptoms of the disease on plant organs as well as with cells in suspension cultures. Litz et al. (1991) fi rst reported that the C.

R.E. Litz et al.658
gloeosporiodes culture fi ltrate was effective as a selective agent in mango sus-pension cultures growing in maintenance medium formulation. Somatic embryos developed from embryogenic cells and PEMs that had survived exposure to C. gloeosporiodes culture fi ltrate, and regenerants appeared to show resistance to inoculation with the pathogen. Jayasankar et al. (1999) characterized the in vitro effects of the purifi ed C. gloeosporiodes phytotoxin, colletotrichin (Gohbara et al., 1977, 1978), and crude C. gloeosporiodes culture fi ltrate on the mortality and growth of polyembryonic ‘Hindi’ and polyem-bryonic ‘Carabao’ embryogenic cultures. In this study, the LD50 values for the effects of C. gloeosporiodes culture fi ltrate and colletotrichin on embryogenic cultures and the growth curves of challenged cultures were established. Later, embryogenic cultures of the same cultivars were either exposed con-tinuously for four cycles of challenge/selection/regrowth or were challenged for one, two, three and four complete cycles with colletotrichin and the par-tially purifi ed culture fi ltrate of C. gloeosporiodes (Jayasankar and Litz, 1998). At the end of each cycle, surviving PEMs were physically removed from the selection medium, cloned and then either rechallenged or subcultured onto somatic embryo maturation medium.
In order to induce the expression of anti-fungal genes in vitro, at least three successive challenges with either crude fi ltrate or colletotrichin were necessary. This was determined by co-culturing the challenged material with a virulent strain of C. gloeosporiodes. Maintenance medium was inoculated with challenged and selected embryogenic cultures at opposite sides of Petri dishes. After 3 weeks, a virulent strain of the pathogen was inoculated in the centre of each Petri dish. Co-culture of the pathogen with resistant cultures resulted in the suppression of mycelium growth; the anti-fungal nature of the PEMs increased with each cycle of challenge and selection, and was maxi-mum after three cycles. There was enhanced production of extracellular anti-fungal proteins chitinase and -1,3-glucanase in selected cultures. An additional chitinase isozyme at 45 kDa was observed with anti-fungal ‘Hindi’ cultures and at 25 kDa with anti-fungal ‘Carabao’ cultures, with respect to the controls. The anti-fungal nature of selected, resistant lines in suspension cultures and in somatic embryos was persistent for more than 2 years follow-ing selection. Various random amplifi cation of polymorphic DNA (RAPD) markers were associated with selected cultures that were strongly anti-fun-gal (Jayasankar et al., 1998). RAPD markers of the unchallenged controls and of leaves from the parent trees were identical, indicating that exposure to either colletotrichin or culture fi ltrate is essential for expression of anti-fungal genes. The RAPD results also demonstrated that embryogenic cultures are quite stable genetically, and the expression of anti-fungal genes is not the result of somaclonal variation. Furthermore, it seems highly probable that the phytotoxins themselves are highly mutagenic.
In vitro-induced mutation followed by selection could be a highly effi cient method for addressing specifi c breeding problems of mango assum-ing that effective selection agents are available. Unfortunately, at this time, there are relatively few such selection agents that can be utilized in this manner.

Biotechnology 659
Genetic transformation
Genetic transformation is currently the only practical solution for improving existing elite selections of perennial species for specifi c horticultural traits and for investigating gene function by interference RNA. Transformation of mango has been reviewed most recently by Litz and Gomez-Lim (2002, 2005) and Gomez-Lim and Litz (2007).
General protocolsMathews et al. (1992, 1993) fi rst reported the genetic transformation of mango using embryogenic cultures of polyembryonic ‘Hindi’ and of a monoembry-onic ‘Keitt’ zygotic embryo-derived embryogenic line, respectively. These two studies utilized two different disarmed, engineered strains of Agrobacterium tumefaciens: (i) strain C58C1 containing the plasmid pGV 3850::1103 with the selectable marker gene for neophosphate transferase (NPTII) which confers resistance to the antibiotic kanamycin, both of which were driven by the CaMV constitutive 35S promoter (Mathews et al., 1993); and (ii) strain A208 containing the plasmid pTiT37-SE::pMON9749, a co-integrate vector, with genes for NPTII and the scorable marker -glucuronidase (gus or uidA) with the 35S promoter (Mathews et al., 1992). A report by Cruz Hernandez et al. (1997) utilized A. tumefaciens strain LBA4404 containing NPTII, -glucuronidase (GUS) and genes that mediate a horticulturally useful trait in binary plasmid pBI121 with the CaMV 35S promoter. Mathews and Litz (1990) earlier had demonstrated that 12.5 g/ml kanamycin sulfate is toxic to embryogenic suspension cultures; whereas, much higher levels (200 g/ml kanamycin) are toxic to embryogenic cultures that are grown on semi-solid medium.
These genetic transformation reports have followed a similar two-step selection (Mathews et al., 1992; Cruz Hernandez et al., 1997). Embryogenic suspension cultures in their logarithmic phase of growth are separated by passing them through sterile fi ltration fabric (1000 m pore size), and the large fraction (>1000 m) is abraded with a sterile brush on sterile fi lter paper. The abraded PEMs are then incubated with acetosyringone-activated A. tumefaciens for 3 days in liquid maintenance medium, with subculture into fresh medium at 24 h intervals. The PEMs are then transferred onto semi-solid maintenance medium supplemented with 200 mg/l kanamycin sulfate and 500 mg/l cefotaxime. After 10 months on this selection medium, the PEMs are transferred to semi-solid maintenance medium containing 400 mg/l kanamycin sulfate. Proliferating cultures are subcultured in liq-uid maintenance medium containing 100 mg/l kanamycin sulfate, and somatic embryo development is initiated by subculture onto semi-solid maturation medium. Mathews et al. (1993) regenerated transgenic mango plants derived from a ‘Keitt’ zygotic embryo embryogenic culture and which had been transformed with pGV 3850::1103 containing the selectable marker gene nptII. Genetic transformation was confi rmed by: (i) growth in selection medium containing inhibitory levels of kanamycin sulfate; (ii) positive histochemical reaction for GUS with X-GLUC (Jefferson, 1987); and (iii) Southern hybridization.

R.E. Litz et al.660
Transient gene expression in embryogenic polyembryonic ‘Kensington Pride’ and polyembryonic ‘Carabao’ cultures has been described using a biolistic approach using two vectors: (i) pBI426 with GUS-NPTII under the control of a double CaMV 35S promoter; and (ii) pBINgfp-Ser, which con-tains NPTII and the green fl uorescent protein gene (gfp) (Cruz Hernandez et al., 2000).
Transformation with genes that mediate horticulturally signifi cant traitsLoss of mango fruit due to spoilage in storage and en route to markets accounts for a signifi cant proportion of total production in many developing countries that have poorly developed infrastructure (i.e. cold storage facili-ties, poor roads, unreliable transportation, etc.) (see Brecht and Yahia, Chap-ter 14, this volume) Mango has become an important export commodity for several developing countries. Extended shelf life and absence of physiologi-cal disorders that cause internal breakdown of fruit (e.g. ‘soft nose’ and ‘jelly seed’) of the most important export cultivars (e.g. monoembryonic ‘Tommy Atkins’) are potentially very important, therefore, for the valuable export trade and for domestic markets.
The mango is a climacteric fruit, and ethylene therefore is a critical regu-lator of the biochemical processes that occur during ripening. Certain rate-limiting genes that mediate ethylene production in mango have been cloned. Cruz Hernandez et al. (1997) described the genetic transformation of embryo-genic polyembryonic ‘Hindi’ mango cultures with mango ACC oxidase, ACC synthase and ACC alternative oxidase cloned in the antisense orientation and under the control of the CaMV 35S constitutive promoter in the pBI121 binary vector in A. tumefaciens strain LBA4404. Embryogenic cultures were transformed by the two-step procedure described above. Although the phe-notype of the transformed lines was not reported, the genetic transforma-tions were confi rmed in each case by the XGLUC reaction for GUS, growth in the presence of inhibitory levels of kanamycin sulfate, Southern blot hybrid-ization and NPTII amplifi cation by PCR. Successful regeneration of plants and inhibition of ethylene production by mature mango fruit could possibly resolve the production problem of premature ripening (jelly seed) and post-harvest loss due to spoilage.
18.5 In vitro Conservation
Medium-term storage
Mango embryos cannot tolerate desiccation during maturation, and devel-opment is not arrested; this is typical of recalcitrant seeds or embryos. With-out a period of developmental arrest, mango embryos develop to maturity and germinate in a continuous sequence. Mango seeds (and embryos) cannot survive for more than 3 or 4 weeks under minimal in vitro growth conditions (Parisot, 1988), and would perish after several days in a conventional seed bank.

Biotechnology 661
Monsalud et al. (1995) demonstrated that 4–5 mm long somatic embryos representing the late heart stage can be partially desiccated, and stored dry in Petri dishes for more than 30 days without any loss of viability. On the other hand, larger somatic embryos cannot survive partial desiccation. This study has interesting implications for future studies that could focus on the concept of an ‘artifi cial seed’ (i.e. somatic embryo) genebank for vegetatively propa-gated tropical fruit trees.
Mango somatic embryos have been manipulated in order to induce developmental arrest. Abscisic acid (ABA) is associated with the initiation of developmental arrest and acquisition of desiccation tolerance of orthodox type seeds and embryos. Abscisic acid causes developmental arrest in vitro at relatively low concentrations for somatic embryos of the orthodox type (Bew-ley and Black, 1985). Pliego Alfaro et al. (1996a, b) were able to arrest the development of late heart stage polyembryonic ‘Hindi’ somatic and nucellar embryos with ABA at 100 M and higher concentrations in the maturation medium. This strategy arrests growth and development for several months so long as mango embryos are on ABA-containing maturation medium. ABA also has a persistent residual effect, and mango somatic embryo develop-ment was inhibited for approximately 1 month after their transfer onto matu-ration medium without ABA. Increased osmolarity of the maturation medium also inhibited somatic embryo development; however, there was no residual effect following transfer of somatic embryos onto maturation medium with-out osmoticum.
Long-term storage
Embryogenic mango cultures cannot be stored indefi nitely, and lose their regeneration potential over time. The initiation of embryogenic cultures is dependent on fl owering and fruit set, which is strictly related to environmen-tal stimuli, and normally occurs one time each year or on alternative years with alternate-bearing selections. Long-term storage of embryogenic mango cul-tures is essential for genetic manipulation studies and will be increasingly important for the management of genetic resources. Embryogenic mango cul-tures have been cryopreserved by different procedures (Wu et al., 2003; Rajani Nadgauda and Pamela Moon, Homestead, Florida, USA, personal communi-cation). Wu et al. (2003) compared three cryopreservation protocols for embryo-genic cultures derived from monoembryonic ‘Zihua’ zygotic embryos: encapsulation-dehydration, pregrowth-dehydration and vitrifi cation. Encap-sulation-dehydration was unsuccessful, and only limited survival (8.3%) was obtained following desiccation of PEMs for 1 h to 58.5% moisture content prior to freezing in liquid nitrogen. Vitrifi cation, involving treatment of PEMs with plant vitrifi cation solution 2 (Sakai et al., 1991) for 20 min prior to freezing in liquid nitrogen, was successful (94.3%). Embryogenic ‘Hindi’ cultures have also been introduced into cryogenic storage (Rajani Nadgauda and Pamela Moon, Homestead, Florida, USA, personal communication) and somatic embryos have been recovered from these cultures. Two procedures were

R.E. Litz et al.662
successful: (i) stepwise cooling in which cryoprotected (5% DMSO and 5% glycerol) embryogenic cultures were cooled in ‘Mr Frosty’ containers at the rate of –1°C/min from room temperature (25°C) to –75°C followed by rapid cooling to –196°C; (ii) rapid cooling (vitrifi cation). After cryovials were removed from liquid nitrogen and rapidly warmed, cultures were thoroughly washed with maintenance medium and plated on semi-solid maintenance medium. Somatic embryo development was initiated by subculturing the PEMs on somatic embryo maturation medium.
18.6 Conclusions
Biotechnology tools have great potential for mango, including advanced micropropagation procedures, conservation and cultivar improvement. There are also several strategies to genetically manipulate the crop for biotechno-logical purposes. Genes in the antisense or sense orientation or RNAi (Small, 2007) can be utilized to inhibit specifi c genes. The increasing availability of identifi ed genes should facilitate a better understanding and genetic manip-ulation of specifi c developmental processes.
Considering the large number of mango microsatellite sequences identi-fi ed and reported in the GenBank, parameters such as heterozygosity, aver-age gene diversity, frequency of outcrossing, cultivar relationships and a mango genetic map should be determined in the near future. Correlations between morphological and DNA markers together with a linkage map should eventually enable MAS for mango improvement.
Most of the research quoted here has been carried out in a few centres; however, it is now widely acknowledged that biotechnology is mainstream research. It is hoped that more research centres will be involved.
References
Andrade, E.H.A., Maia, J.G.S. and Zoghbi, M. das G.B. (2000) Aroma volatile constitu-ents of Brazilian varieties of mango fruit. Journal of Food Composition and Analysis13, 27–33.
Ansari, S.H., Ali, M., Velasco-Negueruela, A. and Pérez-Alonso, M.J. (1999) Volatile constituents of the fruits of three mango cultivars, Mangifera indica L. Journal ofEssential Oil Research 11, 65–68.
Ara, H., Jaiswal, U. and Jaiswal, V.S. (1998) Rooting of microshoots of Mangifera indicaL. cv. Amrapali. Current Science 74, 240–242.
Ara, H., Jaiswal, U. and Jaiswal, V.S. (1999) Germination and plantlets regeneration from encapsulated somatic embryos of mango (Mangifera indica L.). Plant Cell Reports19, 166–170.
Ara, H., Jaiswal, U. and Jaiswal, V.S. (2000a) Plant regeneration from protoplasts of mango (Mangifera indica L.) through somatic embryogenesis. Plant Cell Reports 19, 622–627.
Ara, H., Jaiswal, U. and Jaiswal, V.S. (2000b) Somatic embryogenesis and plantlet regen-eration in Amrapali and Chausa cultivars of mango (Mangifera indica L.). CurrentScience 78, 164–169.

Biotechnology 663
Aron, Y., Czosnek, H., Gazit, S. and Degani, C. (1998) Polyembryony in mango (Mangifera indica L.) is controlled by a single dominant gene. HortScience 33, 1241–1242.
Arumuganathan, K. and Earle, E.D. (1991) Nuclear DNA content of some important plant species. Plant Molecular Biology Reporter 9, 208–218.
Beltrán, A.E., Alvarez, Y., Xavier, F.E., Hernanz, R., Rodriguez, J., Nunez, A.J., Alonso, M.J. and Salaices, M. (2004) Vascular effects of the Mangifera indica L. extract (Vimang). European Journal of Pharmacology 499, 297–305.
Bender, R.J., Brecht, J.K., Baldwin, E.A. and Malundo, T.M.M. (2000) Aroma volatiles of mature-green and tree-ripe ‘Tommy Atkins’ mangoes after controlled atmosphere vs. air storage. HortScience 35, 684–686.
Bewley, J.D. and Black, M. (1985) Seeds: Physiology of Development and Germination.Plenum, New York.
Bojórquez, G. and Gómez-Lim, M.A. (1995) Peroxisomal thiolase mRNA is induced during mango fruit ripening. Plant Molecular Biology 28, 811–820.
Broda, Z. (1984) Wegatatvymna popagacja koniczyny czerwonej (Trifolium pratense L.) proprzez kultury tkankowe ze szczegolnym uwzglednieniem genetycznego uwar-unkowania zdolnosci do regeneracji z kalusa. Rocsniki Akademii Rolniczej w Poz-naniu Rozprawy naukowe 140, 5–41.
Chaimanee, P., Suntornwat, O., Lerrtwikool, N. and Bungaruang, L. (1999) Changes in gene expression during mango fruit ripening, Journal of Biochemistry Molecular Biology and Biophysics 3, 75–79.
Chaimanee, P., Lertwikoon, N., Bungaruang, L. and Suntornwat, O. (2000) Exo-polygal- actanase from ripe mango (Mangifera indica Linn. Cv Nam doc Mai). Acta Horticul-turae 509, 171–176.
Chaplin, G.R. (1989) Advances in postharvest physiology of mango. Acta Horticulturae231, 639–648.
Chaturvedi, H.C. and Sharma, A.K. (1985) Production of androgenic plants of Citrusaurantifolia. Journal of Plant Physiology 119, 473–477.
Chaturvedi, H.C., Agnihotri, S., Sharma, M., Sharma, A.K., Jain, M., Gupta, P., Chour-asia, A. and Kidwai, N.R. (2004) Induced nucellar embryogenesis in vitro for clonal multiplication of Mangifera indica L. var. Ambalavi: a dwarfi ng rootstock. IndianJournal of Biotechnology 3, 22–228.
Chen, Z., Wang, M. and Liao, H. (1980) The induction of Citrus pollen plants in artifi cial media. Acta Genetica Sinica 7, 189–191 (in Chinese).
Chourasia, A., Sane, V.A. and Nath, P. (2006) Differential expression of pectate lyase during ethylene-induced postharvest softening of mango (Mangifera indica var. Dashehari). Physiologia Plantarum 128, 546–555.
Considine, M.J., Daley, D.O. and Whelan, J. (2001) The expression of alternative oxi-dase and uncoupling protein during fruit ripening in mango. Plant Physiology 126, 1619–1629.
Cruz-Hernandez, A. and Gómez-Lim, M.A. (1995) Alternative oxidase from mango (Mangifera indica, L.) is differentially regulated during fruit ripening. Planta 197, 569–576.
Cruz-Hernandez, A., Gomez-Lim, M.A. and Litz, R.E. (1997) Transformation of mango somatic embryos. Acta Horticulturae 455, 292–298.
Cruz-Hernandez, A., Town, L., Cavallaro, A. and Botella, J.lR. (2000) Transient and sta-ble transformation in mango by particle bombardment. Acta Horticulturae 509, 237–242.
Deore, A.C., Chavan, S.S., Deshpande, R.S. and Dhonushke, B.L. (2000) Factors affecting in vitro somatic embryogenesis from nucellar tissue of ‘Alphonso’ mango. Annals of Plant Physiology, 14, 182–186.

R.E. Litz et al.664
DeWald, S.G., Litz, R.E. and Moore, G.A. (1989a) Optimizing somatic embryo pro-duction in mango. Journal of the American Society for Horticultural Science 114, 712–716.
DeWald, S.G., Litz, R.E. and Moore, G.A. (1989b) Maturation and germination of man-go somatic embryos. Journal of the American Society for Horticultural Science 114, 837–841.
Dodd, J.C., Prusky, D. and Jeffries, P. (1997) Fruit diseases. In: Litz, R.E. (ed.) The Mango: Botany, Production and Uses. CAB International, Wallingford, UK, pp. 257–280.
Eiadthong, W., Yonemori, K., Kanzaki, S., Sugiura, A., Utsunomiya, N. and Subhadrab-andhu, S. (2000) Amplifi ed fragment length polymorphism analysis for studying genetic relationships among Mangifera species in Thailand. Journal of the American Society for Horticultural Science 125, 160–164.
Fischer von Mollard, G., Stahl, B., Li, C., Südhof, T.C. and John, R. (1994) Rab proteins in regulated exocytosis. Trends in Biochemical Sciences 19, 164–168.
Flórez-Ramos, C.P., Buitrago, M.E., Lentini, Z. and Cock, J. (2007) Somatic embryogen-esis and plantlet regeneration of mango (Mangifera indica L.). Acta Horticulturae738, 443–445.
Fu, L. and Tang, D. (1983) Induction of pollen plants of litchi tree (Litchi chinensis Sonn.) Acta Genetica Sinica 10, 369–374. (in Chinese with English abstract)
Gamborg, O.L., Miller, R.A. and Ojima, K. (1968) Plant cell cultures: I. Nutrient require-ments of suspension cultures of soybean root cells. Experimental Cell Research 50, 151–158.
Gan, Y.Y., Zaini, S. and Idris, A. (1981) Genetic variation in the grafted vegetatively propagated mango (Mangifera indica). Pertanika 4, 53–62.
García, D., Escalante, M., Delgado, R., Ubeira, F.M. and Leiro, J. (2003a) Anthelminthic and antiallergic activities of Mangifera indica L. stem bark components vimang and mangiferin. Phytotherapy Research 17, 1203–1208.
García, D., Leiro, J., Delgado, R., Sanmartin, M.L. and Ubeira, F.M. (2003b) Mangiferaindica L. extract (vimang) and mangiferin modulate mouse humoral immune re-sponses. Phytotherapy Research 17, 1182–1187.
Garrido, G., Gonzalez, D., Lemus, Y., Garcia, D., Lodeiro, L., Quintero, G., Delporte, C., Nuñez-Selles, A.J. and Delgado, R. (2004) In vivo and in vitro anti-infl ammatory activity of Mangifera indica L. extract (VIMANG). Pharmacology Research 50, 143–149.
Gavin, A.L., Conger, B.V. and Trigiano, R.N. (1989) Sexual transmission of somatic embryogenesis in Dactylis glomerata. Plant Breeding 103, 251–254.
Giovannoni, J. (2001) Molecular biology of fruit maturation and ripening. AnnualReview of Plant Physiology and Plant Molecular Biology 52, 725–749.
Gohbara, M., Kosuge, Y., Suzuki, A. and Tamura, S. (1977) Colletotrichin monohydrate methanol solvate. Acta Crystalogia 33, 1276–1278.
Gohbara, M., Kosuge, Y., Yamasaki, Y., Suzuki, A. and Tamura, S. (1978) Isolation, struc-tures and biological activities of colletotrichins, phytotoxic substances from Col-letotrichium nicotianae. Agricultural and Biological Chemistry 42, 1037–1043.
Gómez-Lim, M.A. (1993) Mango fruit ripening: physiology and molecular biology. Acta Horticulturae 341, 484–499.
Gómez-Lim, M.A. and Litz, R.E. (2007) Mango. In: Pua, E.C. and Davey, M.R. (eds) Biotechnology in Agriculture and Forestry. Vol. 60. Springer, Heidelberg, pp. 51–71.
Gutiérrez-Martinez, P., López-Gómez, R. and Gómez-Lim, M.A. (2001) Identifi cation of an etr1-homologue from mango expressing during fruit ripening and wounding. Journal of Plant Physiology 158, 101–108.

Biotechnology 665
Hammerschlag, F.A. (1992) Somaclonal variation. In: Hammerschlag, F.A. and Litz, R.E. (eds) Biotechnology of Perennial Fruit Crops. CAB International, Wallingford, UK, pp. 35–55.
Jana, M.M., Nadgauda, R.S., Rajmohan, K. and Mascarenhas, A.F. (1994) Rapid somatic embryogenesis from the nucelli of monoembryonic mango varieties. In Vitro Cel-lular and Developmental Biology 30, 55–57.
Jayasankar, S. and Litz, R.E. (1998) Characterization of embryogenic mango cultures selected for resistance to Colletotrichum gloeosporioides culture fi ltrate and phyto-toxin. Theoretical and Applied Genetics 96, 823–831.
Jayasankar, S., Litz, R.E., Schnell, R.J. and Cruz-Hernandez, A. (1998) Embryogenic mango cultures selected for resistance to Colletotrichum gloeosporioides culture fi ltrate show variation in random amplifi ed polymorphic DNA (RAPD) markers. InVitro Cellular and Developmental Biology 34, 112–116.
Jayasankar, S., Litz, R.E., Gray, D.J. and Moon, P.A. (1999) Responses of mango cultures and seedling bioassays to a partially purifi ed phytotoxin produced by a mango leaf isolate of Colletotrichum gloeosporioides Penz. In Vitro Cellular and Developmen-tal Biology 35, 475–479.
Jefferson, R.A. (1987) Assaying chimeric genes in plants: the GUS gene fusion system. Plant Molecular Biology Reporter 5, 387–405.
Johnson, P.R. and Ecker, J.R. (1998) The ethylene gas signal transduction pathway: a molecular perspective. Annual Review of Genetics 32, 227–254.
Krishna, H. and Singh, S.K. (2007) Biotechnological advances in mango (Mangifera in-dica L.) and their future implication in crop improvement–review. BiotechnologyAdvances 25, 223–243.
Kumar, S., Patil, B.C. and Sinha, K. (1990) Cyanide resistant respiration is involved in temperature rise in ripening mangoes. Biochemistry and Biophysics Research Com-mmunications 168, 818–822.
Lad, B.L., Jayasankar, S., Pliego-Alfaro, F.A., Moon, P.A. and Litz, R.E. (1997) Temporal effect of 2,4-D on induction of embryogenic nucellar cultures, culture maintenance and maturation of ‘Carabao’ mango somatic embryos. In Vitro Cellular and Devel-opmental Biology 33, 253–257.
Laxmi, D.V., Sharma, H.C., Kirti, P.B. and Mohan, M.L. (1999) Somatic embryogenesis in mango (Mangifera indica L.). Current Science 77, 1355–1358.
Leiro, J., Garcia, D., Arranz, J.A., Delgado, R., Sanmartin, M.L. and Orallo, F. (2004) An Anacardiaceae preparation reduces the expression of infl ammation-related genes in murine macrophages. International Immunopharmacology 4, 991–1003.
Litz, R.E. (1984) In vitro somatic embryogenesis from nucellar callus of monoembryonic mango. HortScience 19, 715–717.
Litz, R.E. (1986) The mango. In: Evans, D.A., Sharp, W.R. and Ammirato, P.V. (eds) Hand-book of Plant Cell Culture. Vol. IV Techniques and Applications. MacMillan Pub-lishing Co., New York, pp. 612–625.
Litz, R.E. (1987) Application of tissue culture to tropical fruit trees. In: Green, C.E., Som-ers, D.A., Hackett, W.P. and Biesboer, D.D. (eds) Plant Tissue and Cell Culture. Alan R. Liss, New York, pp. 407–418.
Litz, R.E. (2001) Recovery of mango plants with anthracnose resistance following mutation induction and selection in vitro with the phytotoxin(s) produced by Colletotrichum gloeosporioides Penz. In: Improvement of Tropical and Subtrop-ical Fruit Trees through Induced Mutations and Biotechnology. Report of the First Co-ordinated Research Project. Joint Food and Agriculture Organization (FAO)/International Atomic Energy Agency (IAEA) Division, Vienna, Austria, pp. 73–78.

R.E. Litz et al.666
Litz, R.E. (2003) In vitro regeneration and transformation of mango. In: Jaiwal, P.K. and Singh, R.P. (eds) Plant Genetic Engineering Vol. 6: Improvement of Fruit Crops. Sci Tech Publishing, Houston, pp. 23–40.
Litz, R.E. (2004) Biotechnology and Mango Improvment. Acta Horticulturae 645, 85–92.
Litz, R.E. (2008) Mango. In: Kole, C. and Hall, T.C. (eds) Compendium of Transgenic Crop Plants: Transgenic Tropical and Subtropical Fruits and Nuts. Blackwell Pub-lishing, Oxford, UK, pp. 163–174.
Litz, R.E. and Gomez-Lim, M.A. (2002) Genetic transformation of mango. In: Khacha-tourians, G., McHughern, A., Scorza, R., Nip, W.K. and Hui, Y.H. (eds) Transgenic Plants and Crops. Marcel Dekker, New York, pp. 421–436.
Litz, R.E. and Gomez-Lim, M.A. (2005) Mangifera indica Mango. In: Litz, R.E. (ed) Biotechnology of Fruit and Nut Crops. CAB International, Wallingford, UK, pp. 40–61.
Litz, R.E. and Lavi, U. (1997) Biotechnology. In: Litz, R.E. (ed.) The Mango: Botany, Pro-duction and Uses. CAB International, Wallingford, UK, pp. 401–423.
Litz, R.E. and Schaffer, B. (1987) Polyamines in adventitious and somatic embryogenesis in mango (Mangifera indica L.). Journal of Plant Physiology 128, 51–258.
Litz, R.E. and Vijayakumar, N. (1988) In vitro somatic embryogenesis from the nucellus of Mangifera indica L. Acta Horticulturae 231, 473–475.
Litz, R.E., Knight, R.L. and Gazit, S. (1983) Somatic embryos from cultured ovules of polyembryonic Mangifera indica L. Plant Cell Reports 1, 264–266.
Litz, R.E., Knight, R.J., Jr and Gazit, S. (1984) In vitro somatic embryogenesis from Mangifera indica L. callus. Scientia Horticulturae 22, 233–240.
Litz, R.E., Mathews, V.H., Hendrix, R.C. and Yurgalevitch, C. (1991) Mango somatic cell genetics. Acta Horticulturae 291, 133–140.
Litz, R.E., Mathews, V.H., Moon, P.A., Pleigo-Alfaro, F., Yurgalevitch, C. and DeWald, S.G. (1993) Somatic embryogenesis of mango (Mangifera indica L.). In: Reden-baugh, K. (ed.) SynSeeds: Application of Synthetic Seeds to Crop Improvement.CRC Press, Boca Raton, Florida, pp. 409–425.
Litz, R.E., Moon, P.A., Monsalud, M.J., Jayasankar, S. and Mathews, H. (1995) Somatic embryogenesis in Mangifera indica L. (mango). In: Jain, S.M., Gupta, P.K. and New-ton, R.J. (eds) Somatic Embryogenesis in Woody Plants Vol. 2 Angiosperms. Kluwer Academic Publishers, Dordrecht, the Netherlands, pp. 341–356.
Litz, R.E., Yurgalevitch, C. and Hendrix, R.C. (1997) Effects of 1-aminocyclopropane-1-carboxylic acid, aminoethoxyvinylglycine, methylglyoxal-bis-(guanylhydrazone) and dicyclohexylammonium sulfate on induction of embryogenic competence of mango nucellar explants. Plant Cell Tissue and Organ Culture 51, 171–176.
Litz, R.E., Hendrix, R.C., Moon, P.A. and Chavez, V.M. (1998) Induction of embryogenic mango cultures as affected by genotype, explanting, 2,4-D and embryogenic nurse culture. Plant Cell Tissue and Organ Culture 53, 13–18.
López-Gómez, R. and Gómez-Lim, M.A. (1992) Changes in mRNA and protein synthe-sis during ripening in mango fruit. Journal of Plant Physiology 141, 82–87.
López-Valenzuela, J.A., Martínez, O. and Paredes-López, O. (1997) Geographic differen-tiation and embryo type identifi cation in Mangifera indica cultivars using random amplifi ed polymorphic DNA markers. HortScience 32, 105–108.
Lu, C., Zainal, Z., Tucker, G.A. and Lycett, G.W. (2001) Developmental abnormalities and reduced fruit softening in tomato plants expressing an antisense Rab11 GTPase gene. The Plant Cell 13, 1819–1833.
Lycett, G.W., Zainal, Z.B., Los, M., Findlay C.L. and Tucker, G.A. (1997) Novel ripening-specifi c cDNA clones from mango fruit. Acta Horticulturae 455, 277–286.

Biotechnology 667
Malundo, T.M.M., Shewfelt, R.L., Ware, G.O. and Baldwin, E.A. (2001) Sugars and acids infl uence fl avor properties of mango (Mangifera indica L.). Journal of the American Society for Horticultural Science 126, 115–121.
Maluszynski, M. (2001) Offi cially released mutant varieties – the FAO/IAEA database. Plant Cell Tissue and Organ Culture 65, 175–177.
Manzanilla Ramirez, M.A., Robles Gonzalez, M.M. and Medina Urrutia, V.M. (2000) Avances sobre embriogenesis somatica en mango Ataulfo y Haden. In: Proceedings Simposio Mango Control de Floracion y Mejoramiento Genetico. Instituto National de Investigaciones Forestales y Agropecuarias, Apatzingan, Mexico, pp. 47–56.
Mathews, H., Litz, R.E., Wilde, D.H., Merkel, S. and Wetzstein, H.Y. (1992) Stable inte-gration and expression of -glucuronidase and NPT II genes in mango somatic embryos. In Vitro Cellular and Developmental Biology 28, 172–178.
Mathews, H., Litz, R.E., Wilde, H.D. and Wetzstein, H.Y. (1993) Genetic transformation of mango. Acta Horticulturae 341, 93–97.
Mathews, V.H. and Litz, R.E. (1990) Kanamycin sensitivity of mango somatic embryos. HortScience 25, 965–966.
Monsalud, M.J., Mathews, H., Litz, R.E. and Gray, D.J. (1995) Control of hyperhydricity of mango somatic embryos. Plant Cell Tissue and Organ Culture 42, 195–206.
Murashige, T. and Skoog, F. (1962) A revised medium for growth and bioassays with tobacco tissue cultures. Physiologia Plantarum 15, 473–497.
Nair, S., Gupta, P.K. and Mascarenhas, A.F. (1983) Haploid plants from in vitro anther culture of Annona squamosa L. Plant Cell Reports 2, 198–200.
Novak, F.J., Afza, R., van Duren, M. and Omar, M.S. (1990) Mutation induction by gamma irradiation of in vitro cultured shoot-tips of banana and plantain (Musacvs.). Tropical Agriculture 67, 21–28.
Olle, D., Baumes, R.L., Bayonove, C.L., Lozano, Y.F., Sznaper, C. and Brillouet, J.M. (1998) Comparison of free and glycosidically linked volatile components from polyembryonic and monoembryonic mango (Mangifera indica L.) cultivars. Journalof Agricultural and Food Chemistry 46, 1094–1100.
Parisot, E. (1988) Etude de la croissance rhythmique chez de jeunes manguiers (Mangiferaindica L.). Premiere partie: description, germination et conservation de graines polyembryonnees de manguier. Fruits 43, 97–105.
Patena, L.F., Carlos-Refuerzo, L.R. and Barba, R.C. (2002) Somatic embryogenesis and plantlet regeneration in mango (Mangifera indica L.). Vitro Cellular and Develop-mental Biology 38, 173–177.
Pliego-Alfaro, F., Monsalud, M.J., Litz, R.E., Gray, D.J. and Moon, P.A. (1996a) Effect of abscisic acid, osmolarity and partial desiccation on the development of recalcitrant mango (Mangifera indica L.) somatic embryos. Plant Cell Tissue and Organ Culture44, 63–70.
Pliego-Alfaro, F., Litz, R.E., Moon, P.A. and Gray, D.J. (1996b) Effect of abscisic acid, os-molarity and temperature on in vitro development of recalcitrant mango (Mangifera indica L.) nucellar embryos. Plant Cell Tissue and Organ Culture 44, 53–61.
Prasanna, V., Prabha, T.N. and Tharanathan, R.N. (2005) Multiple forms of beta-galactosidase from mango (Mangifera indica L. ‘Alphonso’) fruit pulp. Journal of the Science of Food and Agriculture 85, 797–803.
Prasanna, V., Prabha, T.N. and Tharanathan, R.N. (2006) Multiple forms of polygalactu-ronase from mango (Mangifera indica L. cv Alphonso) fruit. Food Chemistry 95, 30–36.
Raghuvanshi, S.S. and Srivastava, A. (1995) Plant regeneration of Mangifera indica using liquid shaker to reduce phenolic oxidation. Plant Cell Tissue and Organ Culture 41,83–85.

R.E. Litz et al.668
Rao, A.N., Sin, Y.M., Kathagoda, N. and Hutchinson, J.F. (1982) Cotyledon tissue culture of some tropical fruits. In: Rao, A.N. (ed.) Tissue Culture of Economically Important Plants. COSTED, Singapore, pp. 124–137.
Ravishankar, K.V., Anand, L. and Dinesh, M.R. (2000) Assessment of genetic relatedness among mango cultivars of India using RAPD markers. The Journal of Horticultural Science and Biotechnology 75, 198–201.
Reddy, A.S. (2007) Alternative splicing of pre-messenger RNAs in plants in the genomic era. Annual Review of Plant Biology 58, 267–294.
Reisch, B. and Bingham, E.T. (1980) The genetic control of bud formation from callus cultures of diploid alfalfa. Plant Science Letters 20, 71–77.
Rivera Domínguez, M., Manzanilla Ramirez, M.A., Robles González, M. and Gomez-Lim, M.A. (2004) Induction of somatic embryogenesis and plant regeneration of ‘Ataulfo’ mango (Mangifera indica L.). Plant Cell Tissue and Organ Culture 79, 101–104.
Saby John, K., Mohan Rao, L.J., Bhat, S.G. and Prasada Rao, U.J.S. (1999) Characteriza-tion of aroma components of sap from different Indian mango varieties. Phytochem-istry 52, 891–894.
Saiprasad, G.V.S., Anand, L., Ravishankar, K.V., Mythili, J.V., Nagesh, M. and Joshi, R. (2004) Isolation and characterization of mRNAs differentially expressed during rip-ening of mango fruits. Indian Journal of Biotechnology 3, 533–537.
Sairam, K., Hemalatha, S., Kumar, A., Srinivasan, T., Ganesh, J., Shankar, M. and Venka-taraman, S. (2003) Evaluation of anti-diarrhoeal activity in seed extracts of Mangiferaindica. Journal of Ethnopharmacology 84, 11–15.
Sakai, A., Kobayashi, S. and Oyama, I. (1991) Cryopreservation of nucellar cells of naval range (Citrus sinensis Osb. var brasiliensis Tanaka) by vitrifi cation. Plant Cell Reports 9, 30–33.
Sakho, M., Chassagne, D., Jaus, A., Chiarazzo, E. and Crouzet, J. (1998) Enzymatic mac-eration: effects on volatile components of mango pulp. Journal of Food Science 63, 975–978.
Sane, V.A., Chourasia, A. and Nath, P. (2005) Softening in mango (Mangifera indica cv. Dashehari) is correlated with the expression of an early ethylene responsive, ripening related expansin gene, MiExpA1. Postharvest Biology and Technology 38, 223–230.
Schnell, R.A., Ronning, C.M. and Knight, RJ., Jr (1995) Identifi cation of cultivars and validation of genetic relationships in Mangifera indica L. using RAPD markers. The-oretical and Applied Genetics 90, 269–274.
Singh, S.K., Sharma, H.C., Goswami, A.M. and Singh, S.P. (2001) Abscisic acid aided normal somatic embryogenesis from nucellar tissue in mango cv. Amprapali. In: Rai, M., Kumar, S., Singh, R. and Prasad, V.S.R.K. (eds) Recent Trends in Horticul-tural Research, pp. 161–164.
Singh, S.K., Sharma, H.C. and Singh, S.P. (2002) Abscisic acid aided normal somatic embryogenesis from nucellar tissue in mango cv. Amprapali. In: Kapoor, A.C. (ed.) Sustainability of Hill Agriculture: Emerging Trends and Possible Solutions. Indian Agricultural Research Institute, Delhi, pp. 295–299.
Small, I. (2007) RNAi for revealing and engineering plant gene functions. Current Opin-ion in Biotechnology 18, 148–153.
Staehelin, L.A. and Moore, I. (1995) The plant Golgi apparatus: structure, functional organization, and traffi cking mechanisms. Annual Review of Plant Physiology andPlant Molecular Biology 46, 261–288.
Sturrock, D. (1968) Genetics of mango polyembryony. Proceedings of the Florida State Horticultural Society 81, 311–314.

Biotechnology 669
Sulekha, G.R. and Rajmohan, K. (2003) Relative response of varieties and explants in the induction of somatic embryogenesis in mango (Mangifera indica L.). South Indian Horticulture 52, 5–12.
Suntornwat, N., Lertwikoon, L., Bungaruang, P., Chaimanell, J., Speirs, J. and Subhadra-bandhu, S. (2000) Cloning and characterization of putative, endopolygalacturonase cDNA from ripening mango (Mangifera indica Linn cv. Nam Dok Mai). Acta Horti-culturae 509, 153–158.
Thomas, P. (1999) Somatic embryogenesis and plantlet regeneration from nucellar tissue of monoembryonic mango. Journal of Horticultural Science and Biotechnology 74,135–139.
Tucker, G.A. and Grierson, D. (1987) Fruit ripening. In: Stumpf, P.K. and Conn, E.E. (eds) The Biochemistry of Plants Vol. 12. Academic Press, New York, pp. 265–313.
Vasanthaiah, H.K.N., Ravishankar, K.V., Shivashankara, K.S., Anand, L., Narayanas-wamy, P., Mukunda, G. and Prasad, T.G. (2006) Cloning and characterization of differentially expressed genes of internal breakdown in mango fruit (Mangifera in-dica). Journal of Plant Physiology 163, 671–679.
Wilkinson, J.Q., Lanahan, M.B., Yen, H.C., Giovannoni, J.J. and Klee, H.J. (1997) An ethylene-inducible component of signal transduction encoded by never-ripe. Science270, 1807–1809.
Wu, Y.-J., Huang, X.-L., Xiao, J.-N., Li, X.-J., Zhou, M.-D. and Engelmann, F. (2003) Cryopreservation of mango (Mangifera indica L.) embryogenic cultures. Cryo-Letters24, 303–314.
Yang, Y.Q. and Wei, W.X. (1984) Induction of anther-derived plantlet in longan. Acta Genetica Sinica 11, 288–293. (in Chinese with English abstract)
Yashoda, H.M., Prabha, T.N. and Tharanathan, R.N. (2005) Mango ripening-chemical and structural characterization of pectic and hemicellulosic polysaccharides. Car-bohydrate Research 340, 1335–1342.
Zainal, Z., Tucker, G.A. and Lycett, G.W. (1996) A rab11-like gene is developmentally regulated in ripening mango (Mangifera indica L.) fruit. Biochemica et Biophysica Acta 1314, 187–190.

This page intentionally left blank

671
Index
1-aminocyclopropane-1-carboxylic acid (ACC) 494, 652
Abscisic acid 143Alfa 45Algal leaf spot 232–233Alphonso 10, 45–46, 506Alternaria
leafspot 233–235rot 219–221
Alternate bearing 132–133, 145Alternative oxidase (Aox) 492–493, 653–654Amblypelta spp. 322–333Amelie 46Anastrepha spp. 319–320Anthesis 71–72, 136Anthracnose 235–236, 657–658
aetiology 237–239epidemiology 213–214, 239–240host-pathogen interactions 214–216management 216–219, 240post harvest 211–219resistance 11, 64, 77
Apicaldevelopment 100–104, 105, 128–129necrosis 240–241
Aroma 504–507, 544Arumanis 46Ataulfo 46–47Auxin
alternate bearing 132–133development 129–131
fl owering 117–118, 129–131fruit set 141–142somatic embryogenesis 646–649
B74 (Calypso) 47Bacterial canker 74–75, 77, 241–245Bactrocera spp. 78, 320–321
Ceratis spp. 321Banganpalli 47Beak 74Beta 47Biological control 330, 333, 338, 341, 342–343,
346microbial 326, 332parasitoids 325–326, 332, 334, 337post harvest 217predators 326–327, 334, 339
Black-banded disease 245Black mildew 246–247Black root rot 281Black spot see AlternariaBlack tip disorder 310–311Blackfl y 343Blossom blight 248–249Bombay Green 47–48, 77Boron 141, 422–424, 456, 457Botryodiplodia theobromae see Stem-end rotsBotryosphaeria dothidea 221–224, 253Brazil 436–439, 441, 449–451, 461, 463–464,
468, 470–471

Index672
Brazil continuedcultivars 86–87, 445–447fertilizer 452–456industry 434–435
Breeding 68, 89conventional 85–87early 9–11, 641–642gene cloning 651–655genetic transformation 515–516, 659–660genomics 642–643, 655–656inheritance 73–75marker assisted selection 81–83, 89, 651mutations 87–88, 656–658novel germplasm 36–37, 88–89objectives 75
disease resistance 77–78dwarfness 76fruit colour 76–77regular bearing 76open-pollination 78–79
problems 83–84selection 80–81
Brushing 554Budding 383–385Buds
apical development 100–104, 105, 128–129
hormonal control 129–130
C:N ratios 123CO2 181–182, 197–198
storage 506–507, 508–510, 573–575Calcium 419–422, 498, 541Cambodiana 48Canopy 2
cultivar description 44–63development 99–100management 133–134, 449–452, 463–468
Carabao 10, 48Carbohydrate
fl owering regulation 124–127, 132photosynthesis 184–186
Carotenoids 488–491, 498–502Cell culture 643–651Cephaleuros virescens see Algal leaf spotCeratis spp. 321Ceratocystis spp. 274–279Chausa 48–49Chromosome Number 72–73Classifi cation 5
Climate 436–440Coating 513–514, 566–567Cogshall 49Colletotrichum gloeosporioides see AnthracnoseConsumer education 578–579Consumption 607–613Controlled Atmospheres see Modifi ed
AtmospheresCoração de Boi 49Crop
establishment 613–621harvest 470–472production 198–200, 621–626
Cultivars 12–14, 43–44, 445–449description 44–63early 9–11evaluation 63–64 485–491internal fruit breakdown 307–308see also individually named cultivars
Cultivation 614–615history 5–11, 69–70rootstock 35–36, 83, 376–378, 385–386,
443–445see also Propagation
Cuttings 389–391Cycocel 121Cytokinins 118–119, 129–131, 142Cytology 72–73
Daminozide 121Damping off 284–285Dashehari 10, 49, 391–392Deanolis sublimbalis 331–332Decline disorders 249–256Development
costs 613–621crop 432–433fl ower 101–102, 105fruit 139infl orescence 105–108
models 124–133phenology 99–100root 129shoot 100–104tree 190–198
Disease 232, 542–543algal leaf spot 232–233alternaria
leaf spot 233–235rot 219–221

Index 673
anthracnose 11, 64, 77, 211–219, 235–241, 657–658
bacterial black spot 241–245bacterial canker 74–75, 77, 241–245black-banded disease 245black mildew 246–247black root rot 281black spot see Alternariablossom blight 248–249damping off 284–285decline disorders 249–256epidemiology 232–233, 244–245, 247,
249, 259, 273alternaria 219–220, 235anthracnose 213–214, 239–240malformation 265–268stem-end rots 222–223sudden decline 277–278
galls 256–259gray leafspot 259–260host-pathogen interaction 214–216,
220infection process 211–213, 219, 221–222,
240leaf blight 260–261malformation 77, 261–269management 233, 241, 245, 247, 255–256,
259, 273–274, 289alternaria 220–221, 235anthracnose 216–219, 240malformation 268–269post harvest 550–554stem-end rots 223–224sudden decline 278–279
nematode 254, 281–282parasitic plants 270Phoma spp.
blight 270leafspot 270–271
phytophthora diseases 282–284pink disease 271powdery mildew 77, 271–274resistance 12, 75, 241, 273, 279
anthracnose 11, 64, 77, 214–216bacterial canker 74–75, 77, 245mango malformation 77, 269powdery mildew 77
root rot 284–285scab 274scaly bark 256–259sclerotium rot 285
stem-end rots 221–224stigmina leafspot 279–281sudden decline 274–279verticillium wilt 286white root rot 286–288
Disorders see Physiological DisordersDistribution 10–11
species 20, 28–30DNA markers 5, 14, 369–374Domestication 9–10Dothiorella mangiferae see Stem-end rotsDouble working 387Dried products 630–639Drought 468–470
irrigation 193–194tolerance 188–189, 193
Dwarfness 73, 76
Ecology 20–22Economics 535
crop establishment 613–621production 621–626
Eldon 11Elsinoë mangiferae 274Embryo 4–5, 8–9
abortion 139–140somatic embryogenesis 643–650transformation 659–662
Environmentfl owering 111–116, 129, 134–135growth 190–193, 197–200internal fruit breakdown 306–307photosynthesis 173–174, 179–187stress management 468–470
Erythricium salmonicolor 271Espada 49–50, 327–328Ethylene 652–653
fl owering 116–117fruit set 143–144ripening 492–494, 515–516, 569–571
Ewais 50Excellent Succari 50Export 530–535Extrema 50
Fajri 50–51Fernandin 51Fertilizer 452–460, 615Flavour 4, 75, 79, 504–507, 544

Index674
Floodingmanagement 468–470photosynthesis 182–184tree growth 194
Florigon 11Flower 2–3
cultivar description 44–63development 101–102, 105pests 334–338pollination 71–72taxonomy 24–28
Flowering 98–99C:N ratio 123environment 111–116, 129–130, 134–135hormonal infl uence
auxin 117–118, 129–131cytokinins 118–119, 129–131ethylene 116–117fl origenic promoter (FP) 102, 108–110,
123, 131–132gibberellins 119–121, 135growth retardants 121–123
horticultural manipulation 123–124management 133–134, 463–468, 538mechanisms 105–108models 124–133sex ratio 134–136shoot initiation 105–106timing 74, 99–100
Florigenic Promoter (FP) 102, 108–110, 123, 131–132
Fruit 3–4, 516aroma 504–507, 544brushing 554colour 74, 76–77, 79, 191, 498–502,
538–540cracking 310cultivar 44–64development 139disease 210–225fl avour 4, 75, 79, 504–507, 544grading 554–557internal fruit breakdown 308–309, 456,
515–516novel 37nutrients 144–145, 410–411, 412, 414,
420–422, 423–424nutritional value 4, 485–491pests
fruit fl ies 318–328, 559–566fruitspotting bugs 332–333
mango seed borer 331–332seed weevil 78, 328–331thrips 333
photosynthesis 187post-harvest 470–472, 484–485
coating 513–514, 566–567damage 507–508packaging 511–513, 567–568sapburn 545–548, 550storage 506–507, 508–511transpiration 507
processing 628–629, 637–639dehydrated 630–633dried 634–637
ripening 484–485, 536–537, 569–571, 654–655
ethylene 492–494, 515–516, 569–571organic acids 494–495pattern 491–493phenolic compounds 502–504polysaccharides 496–498soluble sugars 495–496
set 140–145taxonomy 24–28weight 78see also Physiological Disorders
Fusarium spp. 281decemcellulare 256–259mangiferae 261–265
epidemiology 265–268management 77, 268–269
sterilihyphosum 264–266 subglutinans 77, 263
Galls 256–259Gene cloning 651–655Genetic
analysis 642–643, 655–656microsatellites 11, 69–70, 81, 82–83,
369–374taxonomy 30–31
erosion 11–12transformation 515–516, 659–660
Genovea 51Germination 377–378, 649–650Germplasm conservation 11–12Gibberellin 328
alternate bearing 132–133fl owering 110, 117–118, 122–123, 135fruit set 142–143

Index 675
propagation 377–378Glenn 11, 51Glomerella cingulate see AnthracnoseGolek 51–52Good Agricultural Practice 531–535, 555–556Grading 554–557Grafting 378–383Gray leafspot 259–260Growth
crop 432–433fl ower 101–102, 105fruit 139infl orescence 105–108
models 124–133phenology 99–100root 129shoot 100–104tree 190–198, 463–468
Growth retardants 121–123
Haden 11, 52, 78Harvest 470–472, 535–538, 544–548Hemicriconemoides mangiferae 254Himsagar 52Hindi Besennara 52Hindi Khassa 52–53History
cultivation 9–10trade 34–35
Honey dew 246–247Hormone
auxin 117–118, 129–133, 141–142, 646–649
balance 129–130, 131–132cytokinins 118–119, 129–131, 142ethylene 116–117, 143–144, 492–494,
515–516, 569–571, 652–653fl origenic promoter (FP) 102, 108–110,
123, 131–132fl owering models
comprehensive conceptual 128–133tri-facter hypothesis 127–128
fruit development 139gibberellin 110, 117–123, 132–133, 135,
142–143, 328, 377–378growth retardants 121–123rooting 387–391vegetative promoter (VP) 110–111,
131–132Human nutrition 4, 485–491
Humidity 182Hybridization 31, 36Hypocryphalus mangiferae 277–278
Idiocerus spp. 78Incompatibility 72India
cultivarbreeding 85–86, 87–88, 641–642origins 8–10, 32–33, 69
production 11, 607Induction 106–108, 123, 131–132Infl orescence
development 101–102, 105photosynthesis 186–187sex ratio 134–136
Inheritance 73–75Insect
honey dew 246–247pollination 138–139resistance 78, 327–328, 330, 341traps 322–324, 329–330, 332, 334,
335–336, 338, 341, 342, 343–344, 347vectors 267, 277–278see also Pests
Internal fruit breakdown 304–306, 515–516cause 306–307management 307–309
Iron 418–419, 459, 460Irrigation 193–194, 460–463Irwin 53
Julie 53
Keitt 53–54, 448, 498–500Kensington Pride 54, 75, 86, 377Kent 54Khanefy 54Kyo Savoy 55
Langra 10, 55, 391Lasiodiplodia spp. 250–251Layering 387–389Leaf 2
age 184cultivar description 44–63development 100–104

Index676
Leaf continuedtaxonomy 25–26temperature 179–181
Leaf blight 260–261Lenticel
damage 538–540spot 311
Lepidoptera 336–337Light
crop production 199–200photosynthesis 177–179tree growth 190–192
Lumpy Tissue 309–310
Mabrouka 55Macrophoma mangiferae 260–261Madame Francis 55–56Magnesium 414–415Malformation 261–265
epidemiology 265–268management 77, 268–269
Mallika 56, 327Management
crop 432–433cultivar 445–449harvest 470–472, 544–548preharvest 535–544
disease 216–221, 223–224, 233, 235, 240–241, 245, 247, 250–256, 259, 268–269, 273–274, 278–279, 289
environmental stress 468–470fertilizer 452–460fl owering 133–134, 463–468growth 463–468irrigation 460–463nutrient 411orchard 199–200, 449–452soil 440–443
Manganese 417–418, 459Mangifera 5, 19–20
altissima 5–8, 27blommesteinii 24caesia 5, 24–25, 31caloneura 26casturi 28, 30cochinchinensis 25–26conservation 11–12decandra 24distribution 28–30ecology 20–22
foetida 5, 24–25, 30–31gedebe 25, 36griffi thii 26, 36indica 5–9, 31, 32, 35–36, 68–69kemanga 5–8, 24–25laurina 8, 26–27, 31, 89magnifi ca 27–28microphylla 26minor 27odorata 8, 24–25, 31origin 31–32pajang 8, 24–25, 30pentandra 8, 26quadrifi da 28, 30, 36superba 24torquenda 27, 30
Mango bud mite 267Mango hopper 78, 334–336Mango seed borer 331–332Manila 10, 56Manzanillo 56Marker Assisted Selection 81–83, 89Marketing 530, 531, 578–579Maturity 535–538, 544–545, 572Mealybugs 343–344Meliola mangiferae 246–247Mesk 56–57Mexico 435, 439, 441–442, 451, 461–462,
464–465, 468–469, 471, 611cultivars 447fertilizer 456–457
Micropropagation 391Microsatellites 11, 69–70, 81, 82–83, 369–374Midge 334, 339–340Mites 340–341Models
carbohydrate-regulated 124–127hormone-regulated 127–133
Modifi ed Atmospheres 508–510, 573–575injury 510–511insecticidal 514–515packaging 511–513
Molecularmarker 5, 14, 69–70, 369–374, 658
assisted breeding 81–83, 89taxonomy 30–31
Monoembryonic 4–5, 8–9, 32–33, 369, 376–377
Mulgoa 10–11, 57Mulgoba see MulgoaMutations 87–88, 656–658

Index 677
Nabeel 57Nam Doc Mai 57–58Neelum 58Nematode 254, 281–282Neofusicoccum spp. 248–249, 251–253Nitrogen 407–411, 452–460, 538–539Nutrients 440–443
analysis 405–407boron 141, 422–424, 456, 457C:N ratios 123calcium 419–422, 498, 541carbohydrate 124–127, 132, 184–186defi ciency 409–410, 412, 413, 414–417,
418, 419, 420, 423, 456fertilizer 452–460fl owering 114–115, 123, 124–127, 130,
132, 133, 418fruit 144–145, 410–411, 412, 414,
420–422, 423–424internal fruit breakdown 308–309, 456iron 418–419, 459, 460magnesium 414–415management 411manganese 417–418, 459nitrogen 407–411, 452–460, 538–539phosphorus 411–412, 452–460potassium 412–414, 452–460, 541sulfur 415–416toxicity 409–410, 412, 413, 414–417, 418,
419, 420, 423uptake 409, 411–412, 413, 414, 415, 416,
417–418, 419–420, 422–423yield 410–411, 412, 414, 417, 418,
420–421, 423–424zinc 416–417, 456, 460
Nutrition 4, 485–491Nuwun Chan 58
Oidium mangiferae 77Okrung 58Orchard 199–200, 449–452 613–621Organic acids 494–495Osteen 11, 59
Packaging 511–513, 514–515, 567–568, 575Packhouse 548–550
brushing 554disease control 550–554disinfestation 557–566
grading 554–557market preparation 566–571
Paclobutrazol (PBZ) 110–111, 121–123, 133, 135, 143, 463, 467
Pairi 59, 77Palmer 59Panicle 104–105Parasitic plants 270Pectin 497–498Personnel 534–535Pestalotiopsis mangiferae 259–260
see also Stem-end rotsPests 317–318, 345–346, 542–543
blackfl y 343bud mite 267control 322, 346–347, 557–566
biological 325–327, 330, 332, 333, 334, 335, 337, 338, 339, 341, 342–343
chemical 324–325, 330, 332, 333, 335, 337, 338, 339, 341, 345
cultural 327, 330, 344, 560–564host resistance 78, 327–328, 330, 341monitoring 322–324, 329–330, 332, 334,
335–336, 338, 341, 342, 343–344fruit fl ies 318–319
Anastrepha spp. 319–320Bactrocera spp. 78, 320–321Ceratis spp. 321control 322–328, 559–566
fruitspotting bugs 332–333lepidoptera 336–337mango hoppers 78, 334–336mango seed borer 331–332mealybugs 343–344midge 334, 339–340mites 340–341scale 342–343, 344–345seed weevil 78, 328–331termites 345thrips 333, 337–338, 339nematodes 254, 281–282whitefl y 343yield loss 321, 330, 331–332, 343see also Insects
Pesticides 542–543foliar diseases 235, 240, 261, 268–269,
273–274paclobutrazol (PBZ) 110–111, 121–123,
133, 135, 143, 463, 467pests 324–325, 330, 332, 333, 335, 337,
338, 339, 341, 346, 514–515

Index678
Pesticides continuedpost harvest 216–219, 220–221, 224,
550–554residues 533
Phenolic compounds 502–504Phenology 99–100Philippines 8–10, 33Phoma spp. 270–271Phomopsis mangiferae see Stem-end rotsPhosphorus 411–412, 452–460Photoperiod 113–114Photosynthesis 172–177
atmospheric CO2 181–182carbohydrate accumulation 184–186fl ooding 182–184fruit 187humidity 182infl orescence 186–187leaf 179–181, 184light 177–179
stomatal conductance 173–174Physiological Disorders 303–304, 311–312,
507, 510–511, 540–542black tip disorder 310–311chilling injury 507–508, 571–573fruit cracking 310heat injury 508internal fruit breakdown 304–306,
515–516cause 306–307management 307–309
lenticel spot 311lumpy tissue 309–310ricey tissue 310
Phytophthora diseases 282–284Pigments 488–491, 498–502Pink disease 271Pollen 136–137Pollination 71–72, 137–139, 136
controlled 80open 78–79
Polyembryonic 4–5, 32–33, 70–71, 84, 369–374, 377, 385–386
Polyphenol oxidase (PPO) 504Polyploidy 73Polysaccharides 496–498Postharvest 484–485
coatings 513–514, 566–567damage 507–508disease 210–211, 224–225
alternaria rot 219–221
anthracnose 211–219stem-end rots 221–224
management 470–472packhouse management 548–566quality assurance 531–535sapburn 545–548storage 506–507, 508–510, 514–515transpiration 507
Potassium 412–414, 452–460, 541Powdery mildew 77, 271–274Precocity 73, 135Processing 628–629, 637–639
dehydrated 630–633dried 634–637
Productioncrop 198–200
yield 434–436market 530–531region 434–436, 606–613
climate 436–440volume 14–15
Propagation 367–369, 391–392, 443–445seed 369–374vegetative 374–375
budding 383–385cuttings 389–391double working 387grafting 378–383layering 387–389micropropagation 391rootstock 376–378, 385–386, 443–445top working 386–387
Protoplast 650–651Pseudomonas syringae pv, syringae 240–241Pythium vexans 284–285
Quality assurance 531–535, 555–557Quarantine 328, 532–533, 557–566
Reproductionanthesis 71–72, 136cytology 72–73fl oral biology 134–139incompatibility 72pollen 136–137pollination 71–72, 136, 137–139polyembryonic 4–5, 70–71, 369–374
origin 32–33Rhinocladium corticola 245

Index 679
Rhizoctonia solani 284–285Ricey tissue 310Rigidoporus lignosus 286–288Ripening 484–485, 536–537, 569–571,
654–655ethylene 492–494, 515–516, 569–571organic acids 494–495pattern 491–493phenolic compounds 502–504polysaccharides 496–498soluble sugars 495–496
Root 129, 380Rooting 387–391Rootstock 35–36, 83, 376–378, 385–386,
443–445Rosa 59–60
Salinity 196–197Sandersha 11, 327Sapburn 545–548, 550Scab 274Scale 342–343, 344–345Scaly bark 256–259Sclerotium rot 285Seed 4–5, 369–374
cultivar description 44–63germination 377–378hormones 130, 132–133mango seed borer 331–332weevil 78, 328–331
Seedbed 377–378Sensation 60Sex ratio
fl owering 134–136fruit set 140–141
Shootchimeric 107–108development 100–104formation 128–129initiation 105–106, 110–111, 129–131reproductive 104–105
Smartfresh 571Smith 11, 328Soil
analysis 405–407crop production 440–443fl ooding 182–184, 194–195temperature 192–193
Soluble sugars 495–496Somatic embryogenesis 643–650
Sooty blotch 246–247Sooty mould 246–247Source-sink balance 184–186Stem-end rots 221–224Stenochetus mangiferae 78Stenospermocarpy 139–140Sternochetus spp. 78, 328–331Stigmina leafspot 279–281Stomatal conductance 173–174Storage 506–507, 508–510, 514–515, 540–542,
569, 571–575Stress 37, 468–470Sudden decline 274–279Sulfur 415–416Suvarnarekha 60
Tahar 60Taimour 61Taiwan 435, 439, 442, 447–448, 457, 465–466,
469, 471Taxonomy 19–20, 22–30Temperature
crop production 198–199fl owering 111–113, 129, 134–135growth 192–193internal fruit breakdown 306photosynthesis 179–181
Termites 345Thailand 88Thiabendazole (TBZ) 551–553Thrips 333, 337–338, 339Tissue culture 643–651Tommy Atkins 11, 61, 323, 328, 448, 498–500Top working 386–387Totapuri 61, 327, 506Trade 34–35, 530–535, 606–611Transport 533–534, 548, 575–578Tree growth 190–198, 463–468Triazoles see Paclobutrazol (PBZ)Turpentine 11, 62, 78, 377
USA 462–463, 466–468, 469–470, 471–472, 611–613
costs 613–626cultivars 78, 448–449cultivation 10–11, 70fertilizer 457–460production 435–436, 439–440, 442–443,
451–452

Index680
Vallenato 62Van Dyke 62Vegetative Promoter (VP) 110–111, 131–132Verticillium wilt 286Volatiles 504–507
Water relations 188–190fl owering 113–114, 130irrigation 460–463
Whitefl y 343White root rot 286–288White Succari 62Wind 195–196
Xanthomonas campestris pv. mangiferaeindicae 241–242, 244–245
aetiology 242–244resistance 74–75, 77, 245
Yield 170–172, 615, 624crop production 198–200, 434–436nutrient 410–411, 412, 414, 417, 418,
420–421, 423–424photosynthesis 172–177regular bearing 73, 76tree growth 190–198
Zebda 63Zinc 416–417, 456, 460






























