Embed Size (px)
Citation preview

MARIO MAYES
THE MEIOTIC ARREST OF BOVINE OOCYTES
Thèse présentée
à la Faculté des études supérieures de l'Université Laval
pour l’obtention du grade de Philosophiae Doctor (Ph. D.)
Département des sciences animales
FACULTE DES SCIENCES DE L`AGRICULTURE ET DE L`ALIMENTATION UNIVERSITÉ LAVAL
QUÉBEC
JUILLET 2002 © Mario Mayes, 2002

RÉSUMÉ
Les ovocytes bovins incubés in vitro reprennent la méiose alors que ceux incubés en présence de
mono-couches de cellules de la thèque demeurent au stade de vésicule germinale (GV). Les
présentes expériences ont été menées sur des ovocytes bovins afin de mieux comprendre les
facteurs impliqués dans le contrôle de la maturation des ovocytes. Les objectives étaient 1) de
déterminer si la morphologie du complexe ovocyte-cumulus (COCs) est corrélée avec les
cinétiques de la reprise méiotique suivant une période d’arrêt méiotique induite. 2) Étudier la
fonction des inhibiteurs de phosphodiesterase (PDE) spécifiques seuls ou en présence de une
monocouche des cellules de la thèque sur l’arrêt méiotique des ovocytes bovins. 3) Étudier le
rôle de la voie de signalisation de la protéine kinase A et de la protéine kinase C dans la
modulation de la reprise méiotique des ovocytes bovins. 4) Faire une caractérisation plus
poussée du facteur d’inhibition sécrété par les cellules de la thèque. Ces études démontrent que
les COCs bovins reprennent la méiose plus rapidement après une courte période d’arrêt méiotique
que les COCs non traités. Les ovocytes atrésiques ou dénudés sont donc moins nombreux au
stage de GV au début de la maturation que les COCs sains ou légèrement atrésiques. Les
inhibiteurs de phosphodiesterase (PDE) de type 3 ont maintenu les ovocytes bovins en arrêt
méiotique alors que les inhibiteurs de type 4 n’ont eu aucun effet. De plus, la combinaison des
inhibiteurs PDE et des cellules de la thèque montre un effet additif. Les COCs incubés avec des
cellules de la thèque sont restés en arrêt méiotique alors que ceux incubés en présence de cellules
de granulosa ont repris la méiose. Les cellules de la granulosa non traitées ont annulé l’effet
inhibiteur des cellules de la thèque sur la maturation des COCs bovins. Par ailleurs, les COCs
sont restés en arrêt méiotique lorsque des analogues d’AMPc ont été utilisés en conjonction avec
des cellules de granulosa et de la thèque. Finalement, le facteur inhibiteur sécrété par les cellules
de la thèque dans le milieu conditionné est (a) insensible à la chaleur, aux enzymes protéolytiques
et à un traitement au charbon, (b) sensible à un traitement au chloroforme, (c) concentré dans la
fraction plus petite que 1000 daltons du milieu conditionné et (d) les patrons de pics
chromatographiques sont différents entre le milieu conditionné et le milieu frais. D’autres études
seront nécessaires pour l’isolation complète du facteur(s) inhibiteur(s) sécrété(s) par les cellules
de la thèque responsable du maintien de l’arrêt méiotique des ovocytes bovins.
ii

ABSTRACT Bovine oocytes incubated in vitro resume meiosis, whereas most of those incubated with theca
cell monolayers remain at the germinal vesicle (GV) stage. Experiments were carried out using
bovine oocytes to understand the factors involved in the control of oocyte maturation. The
objectives were 1) to determine whether or not the morphology of cumulus oocyte complexes
(COC) is correlated with the kinetics of meiotic resumption following a period of induced
meiotic arrest. 2) To investigate the role of type-specific phosphodiesterase (PDE) inhibitors
alone or in conjunction with theca cell monolayers on meiotic arrest of bovine oocytes. 3) To
study the role of the protein kinase A and protein kinase C signaling pathways in the control of
meiotic resumption of bovine oocytes incubated with granulosa cells, theca cell monolayers or
both cell types. 4) To further characterize the inhibitory factor secreted by the theca cell
monolayers. These studies show that bovine COCs resume meiosis faster following a short
period of induced meiotic arrest than untreated COCs. Atretic or denuded oocytes have fewer
oocytes at the GV stage at the onset of maturation compared to healthy or slightly atretic COCs.
Type 3 phosphodiesterase (PDE) inhibitors maintained bovine oocytes in meiotic arrest, whereas
type 4 PDE inhibitors did not have an effect. Furthermore, PDE inhibitors and the theca cell
monolayers show an additive effect. COCs incubated with the theca cell monolayer remained in
meiotic arrest, whereas those incubated with granulosa cells resumed meiosis. Untreated
granulosa cell abrogate the inhibitory effect of the theca cell monolayers on the maturation of
COCs. However, COCs remain in meiotic arrest when cAMP analogs are used in conjunction
with both granulosa and theca cells. The inhibitory factor(s) secreted by the theca cell
monolayers is (a) not sensitive to heat, proteolytic enzymes and charcoal treatment, (b) sensitive
to chloroform treatment, (c) has a Mr less than 1000 daltons and (d) the patterns of
chromatographic peaks are different in conditioned medium and fresh medium. Further work is
needed to fully isolate and identify the factor produced by the theca cells responsible for
maintaining bovine oocytes in meiotic arrest.
iii

AVANT-PROPOS J’aimerais exprimer ma gratitude envers mon directeur de recherche M. Marc-André Sirard, pour
m’avoir donné l’opportunité de poursuivre mes études au niveau du Doctorat. Je crois que ce fût
une expérience enrichissante pour chacun de nous.
J’aimerais également souligner le travail fait par les membres du comité examinateur: Mme
Janice Bailey Ph.D., Mme Trudee Fair Ph.D., M. Michel Fortier Ph.D. ainsi que M. François
Richard Ph.D.
J’aimerais aussi exprimer ma reconnaissance envers la Fondation de l’Université Laval pour
m’avoir octroyé une bourse d’étude.
De sincères remerciements à Karine Coenen, Susan Novak, Patrick Bélanger et André Roy pour
toute l’assistance technique qu’ils m’ont apporté. Leur contribution m’ayant rendu la vie plus
facile au cours des années. J’aimerais également remercier les membres du Centre de Recherche
en Biologie de la Reproduction (C.R.B.R.)
Je remercie, du fond du cœur, les personnes qui ont été là pour moi dans les bons et moins bons
moments et tout spécialement les personnes suivantes: Anne Rieth, François Richard, Gisèle
LaPointe, Shannon Scott, Janice Bailey, Denis Garneau, Céline Campagna, Charlotte Dubé,
Denise Lelièvre, Marilène Gosselin, Melinda Boice, Jocelyn Desrochers et les membres du Club
de course à pied “La Foulée”. Je me considère comme une personne très chanceuse d’avoir pu
vous connaître et d’avoir partager d’inoubliables moments avec tous et chacun d’entre vous.
Finalement, j’aimerais remercier mes parents et ma famille pour tous les sacrifices qu’ils ont faits
afin de me permettre de poursuivre mes études supérieures.
iv

ACKNOWLEDGEMENTS I would like to express my gratitude to my research Director, Marc-André Sirard for giving me
the opportunity to pursue a Doctoral degree. I believe it has been a learning experience for both
of us.
I would also like to recognize the work done by the members of my examining committee: Janice
Bailey Ph.D., Trudee Fair Ph.D., Michel Fortier, Ph.D, and François Richard Ph.D.
I am very thankful to La Foundation de l`Université Laval for awarding me a scholarship.
I would like to acknowledge the technical support provided by Karine Coenen, Susan Novak,
André Roy and Patrick Bélanger. Your technical expertise and knowledge made my life easier
throughout these years. I would also like to thank the members of Le Centre de Recherche en
Biologie de la Reproduction (CRBR).
I give thanks with all my heart the people who have been there for me in the good and not so
good times. I specially would like to recognize the following people: Anne Rieth, François
Richard, Gisèle LaPointe, Shannon Scott, Janice Bailey, Denis Garneau, Céline Campagna,
Charlotte Dubé, Denise Lelièvre, Marilène Gosselin, Melinda Boice, Jocelyn Desrochers and the
members of the Running Club La Foulée. I consider myself a very lucky person for having met
and shared some unforgettable time with all of you.
Finally, I would like to recognize my parents and family for all the sacrifices that they have made
to allow me to pursue an advance degree.
v

TABLE OF CONTENTS RÉSUMÉ......................................................................................................................................... ii ABSTRACT................................................................................................................................... iii AVANT-PROPOS ..........................................................................................................................iv ACKNOWLEDGEMENTS .............................................................................................................v List of abbreviations...................................................................................................................... xii General Introduction ........................................................................................................................1 1 CHAPTER I Oogenesis and Meiotic Arrest. .....................................................................................................3
1.1 The Ovary.........................................................................................................................4 1.2 The Follicle ......................................................................................................................5 1.3 Folliculogenesis................................................................................................................6
1.3.1 Primordial germ cells (PGC)....................................................................................6 1.3.2 Oogonia ....................................................................................................................7 1.3.3 Primordial follicles...................................................................................................8 1.3.4 Primary Follicles ....................................................................................................11 1.3.5 Secondary Follicles ................................................................................................11 1.3.6 Tertiary or Antral Follicles.....................................................................................11
1.4 Developmental Competence ..........................................................................................14 1.5 The Estrous Cycle ..........................................................................................................16 1.6 Follicular Waves in Cattle..............................................................................................16 1.7 Apoptosis........................................................................................................................18 1.8 Meiosis ...........................................................................................................................19 1.9 Oocyte Maturation..........................................................................................................20
1.9.1 Oocyte maturation: In vivo vs. in vitro ..................................................................21 1.9.2 Nuclear maturation.................................................................................................23 1.9.3 Cytoplasmic maturation .........................................................................................27
1.10 The role of M-Phase Factor in oocyte maturation..........................................................29 1.11 Meiotic Arrest ................................................................................................................31
1.11.1 Transcription inhibitors ..........................................................................................31 1.11.2 Protein synthesis inhibitors ....................................................................................33 1.11.3 Phosphatase inhibitors............................................................................................35 1.11.4 Protein kinase inhibitors.........................................................................................36 1.11.5 Protein kinase A .....................................................................................................38 1.11.6 Phosphodiesterase inhibitors ..................................................................................40 1.11.7 Protein kinase C .....................................................................................................41 1.11.8 Purines....................................................................................................................42 1.11.9 Physiological methods of maintaining meiotic arrest in vitro................................42
1.12 Hypothesis......................................................................................................................49 1.13 References ......................................................................................................................50
vi

2 CHAPTER II The Influence of Cumulus-Oocyte Complex Morphology and Meiotic Inhibitors on the Kinetics of Nuclear Maturation in Cattle. .......................................................................67
2.1 RÉSUMÉ........................................................................................................................68 2.2 ABSTRACT...................................................................................................................69 2.3 INTRODUCTION..........................................................................................................69 2.4 INTRODUCTION..........................................................................................................70 2.5 MATERIALS AND METHODS ...................................................................................72
2.5.1 Preparation of Theca Cell Monolayers...................................................................72 2.5.2 Oocyte Collection...................................................................................................73 2.5.3 Oocyte Classification .............................................................................................74 2.5.4 Experimental Design ..............................................................................................74 2.5.5 Oocyte Fixation ......................................................................................................75
2.6 Statistical Analysis .........................................................................................................75 2.7 RESULTS.......................................................................................................................75 2.8 DISCUSSION ................................................................................................................76 2.9 ACKNOWLEGMENTS.................................................................................................78 2.10 REFERENCES...............................................................................................................79
3 CHAPTER III Effect of Type 3 and Type 4 Phosphodiesterase Inhibitors on the Maintenance of Bovine Oocytes in Meiotic Arrest. .......................................................................................84
3.1 RÉSUMÉ........................................................................................................................85 3.2 ABSTRACT...................................................................................................................86 3.3 INTRODUCTION..........................................................................................................86 3.4 MATERIALS AND METHODS ...................................................................................89
3.4.1 Preparation of Theca Cell Monolayers...................................................................89 3.4.2 Collection of COCs ................................................................................................90 3.4.3 Chemicals ...............................................................................................................90 3.4.4 Denuded Oocytes ...................................................................................................90 3.4.5 Fixation of Oocytes ................................................................................................91
3.5 EXPERIMENTAL DESIGN..........................................................................................91 3.5.1 Exp 1: PDE inhibitors and bovine DO...................................................................91 3.5.2 Exp 2: PDE inhibitors and bovine COC.................................................................91 3.5.3 Exp 3: PDE inhibitors and bovine COCs incubated with theca cell monolayers ..91
3.6 STATISTICAL ANALYSIS..........................................................................................92 3.7 RESULTS.......................................................................................................................92 3.8 DISCUSSION ................................................................................................................93 3.9 ACKNOWLEDGEMENTS ...........................................................................................96 3.10 REFERENCES...............................................................................................................96
4 CHAPTER IV Granulosa Cells Reverse the Inhibitory Effect of Theca Cell Monolayers
vii

on the Maturation of Bovine Cumulus-Oocyte Complexes. ....................................................103 4.1 RÉSUMÉ......................................................................................................................105 4.2 ABSTRACT.................................................................................................................106 4.3 INTRODUCTION........................................................................................................106 4.4 MATERIALS AND METHODS .................................................................................109
4.4.1 Preparation of Theca Cell Monolayers.................................................................109 4.4.2 Preparation of Granulosa Cells ............................................................................110 4.4.3 Collection of COCs ..............................................................................................110 4.4.4 Chemicals .............................................................................................................111 4.4.5 Evaluation of Nuclear Maturation........................................................................111
4.5 EXPERIMENTAL DESIGN........................................................................................111 4.6 STATISTICAL ANALYSIS........................................................................................111 4.7 RESULTS.....................................................................................................................112
4.7.1 Incubation of COCs with granulosa cells.............................................................112 4.7.2 Incubation of COCs with theca cell monolayers..................................................112 4.7.3 Incubation of COCs with granulosa and theca cell monolayers. .........................112
4.8 DISCUSSION ..............................................................................................................113 4.9 ACKNOWLEDGMENTS............................................................................................114 4.10 REFERENCES.............................................................................................................115
5 CHAPTER V Partial Characterization of the Meiosis Inhibiting Factor Secreted by Bovine Theca Cell Monolayers. ...........................................................................................................120
5.1 RÉSUMÉ......................................................................................................................121 5.2 ABSTRACT.................................................................................................................122 5.3 INTRODUCTION........................................................................................................123 5.4 MATERIALS AND METHODS .................................................................................124
5.4.1 Preparation of Theca Cell Monolayers.................................................................124 5.4.2 Collection of COCs ..............................................................................................125 5.4.3 Denuded Oocytes .................................................................................................126 5.4.4 Fixation of Oocytes ..............................................................................................126 5.4.5 Culture media .......................................................................................................126 5.4.6 Chromatography...................................................................................................130
5.5 EXPERIMENTAL DESIGN........................................................................................130 5.5.1 Exp 1: Effect of medium replacement on the nuclear maturation of COCs incubated with theca cell monolayers. .....................................................131 5.5.2 Exp 2: Effect of heat-treated CM on the nuclear maturation of oocytes incubated with theca cell monolayers. ......................................................131 5.5.3 Exp 3: Effect of dextran:charcoal-treated CM .....................................................131 5.5.4 Exp 4: Effect of medium supplemented with cut off fractions of CM.................131 5.5.5 Exp 5 Effect of enzymatic treatment of CM-cFCS and FM-cFCS ......................131 5.5.6 Exp 6 Effect of dialysis treatment of CM-cFCS and FM-cFCS ..........................132 5.5.7 Exp 7 Effect of chloroform treatment of CM and CM-cFCS ..............................132 5.5.8 Exp 8 Effect of the organic phase of cCM-cFCS.................................................132 5.5.9 Exp 9 Effect of different chromatographic fractions of CM-cFCS......................132
viii

5.6 STATISTICAL ANALYSIS........................................................................................132 5.7 RESULTS.....................................................................................................................133 5.8 DISCUSSION ..............................................................................................................136 5.9 ACKNOWLEDGMENTS............................................................................................140 5.10 REFERENCES.............................................................................................................140
6 GENERAL CONCLUSIONS ..............................................................................................153
ix

List of figures Figure 1. 1 Diagram of a mammalian ovary.................................................................4
Figure 1. 2 Life history of ovarian follicles ................................................................10
Figure 1. 3 Schematic representation of the growth, capacitation and
maturation of the bovine oocyte throughout folliculogenesis. .................13
Figure 1. 4 Schematic illustration of two-way functional coupling between FSH and the follicles in the bovine. .........................................................18
Figure 1. 5 Bovine cumulus-oocyte complex (COC) with compact layers of cumulus cells at the beginning of in vitro maturation. .............................21
Figure 1. 6 Bovine cumulus-oocyte complex (COC) with expanded layers of cumulus cells after 24h of in vitro maturation. ........................................25
Figure 1. 7 The stages of nuclear maturation of bovine oocytes. ..............................26
Figure 2. 1 Nuclear status of bovine oocytes at 0 h after an incubation of 4 h...........81 Figure 2. 2 Nuclear status of bovine COCs of Class 1 after 0 or 4 h
incubation in Cycloheximide (CX) and theca cell-free medium or 4 h of inhibited GVBD by theca cell monolayers or CX followed by another 4 h period of incubation in CX and theca cell-free medium.....................................................................................................82
Figure 2. 3 Nuclear status of bovine oocytes of Classes 1 to 2 at 0 h, 4 h of incubation or after 24 h co-incubation with theca cell monolayers. ........83
Figure 3. 1 Effect of specific PDE inhibitors on the maturation of denuded bovine oocytes........................................................................................100
Figure 3. 2 Effect of specific PDE3 and PDE4 inhibitors on the maturation of bovine COCs. .........................................................................................101
Figure 3. 3 Effect of specific PDE3 and PDE4 inhibitors on the maturation of bovine COCs incubated with theca cell monolayers..............................102
x

Figure 4. 1 Modulation of the meiotic arrest of bovine COCs incubated for 24 h in the presence of granulosa cells, theca cell monolayers or both cell types.........................................................................................118
Figure 5. 1 Effect of fresh culture medium on the nuclear maturation of
oocytes cocultured with theca cell monolayers......................................144
Figure 5. 2 Effect of heat-treated conditioned medium on nuclear maturation of oocytes cocultured with theca cell monolayers..................................145
Figure 5. 3 Effect of charcoal-treated conditioned medium on nuclear maturation of oocytes cocultured with theca cell monolayers. ..............146
Figure 5. 4 Effect of different cut-off fractions of medium on nuclear maturation of oocytes cocultured with theca cell monolayers. ..............147
Figure 5. 5 Effect of chloroform-treated conditioned medium on nuclear maturation of oocytes cocultured with theca cell monolayers. ..............148
Figure 5. 6 Effect of the organic phase of chloroform-treated conditioned medium on nuclear maturation of oocytes cocultured with theca cell monolayers. .....................................................................................149
Figure 5. 7 Effect of incubating bovine COCs with theca cell monolayers and culture medium supplemented with the chromatographic fractions obtained from the supernatant of chloroform treated conditioned medium mix............................................................................................150
Figure 5. 8 Chromatographic comparison of CM and FM samples .........................152
xi

List of abbreviations AC adenylate cyclase AI anaphase I ANOVA analysis of variance BAPTA/AM 1, 2-bis(o-aminophenoxy)ethane-N,N,N',N'- tetraacetic acid-acetoxymethyl BFF bovine follicular fluid BL-I butyrolactone-I BSA bovine serum albumin Ca2+, Mg2+ calcium, magnesium Cdk1 cyclin dependent kinase 1 Cil cilostamide cFM-cFCS chloroform treated-FM supplemented with choroform-treated FCS cCM-cFCS chloroform treated-CM supplemented with chloroform-treated FCS CM conditioned medium CM-cFCS conditioned medium supplemented with chloroform-treated FCS CM-cFCS Pooled samples of CM-cFCS CO2 carbon dioxide COC cumulus-oocyte complex COCG COC connected to the membrana granulosa cAMP cyclic adenosine monophosphate cGMP cyclic guanosine monophosphate CSF cytostatic factor DAG diacylglycerol dbcAMP dibutyril cyclic AMP DC dextran:charcoal DNA deoxyribonucleic acid DNase dexosyribonuclease DO (OD) Denuded oocyte DRB 5,6-dichloro-1-ß-ribofuranosylbenzimidazole EDTA ethylenediaminetetraacetic acid ET erreur type FCS fetal calf serum FK forskolin FM fresh unconditioned medium FM-cFCS Fresh unconditioned medium supplemented with chloroform-treated FCS FM-cFCS mix Pooled samples of FM-cFCS FSH follicle stimulating hormone GV germinal vesicle GVBD germinal vesicle Breakdown h hour(s) Hepes 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid hnRNA heterogeneous nuclear RNA IBMX 3-isobutyl-1-methylxanthine IP3 inositol triphosphate
xii

IU international unit IVM/ IVF/IVC in vitro maturation / in vitro fertilization / in vitro culture kDa kilo daltons LH luteinizing hormone M stage of cell division MI, MII metaphase I, metaphase II Mil milrinone MAPK mitogen activated protein kinase min minute(s) MIS müllerian inhibiting substance ml, µl, l milliliter, microliter, liter mM, µM, M millimolar, micromolar, Molar mg, µg, ng, g milligram, microgram, nanogram, gram mm, µm, nm, m millimeter, micrometer, nanometer, meter MPF M-phase promoting factor / maturation promoting factor mRNA messenger RNA Mr relative molecular weight n number of observations NaF sodium fluoride NMWL nominal molecular weight limit OA okadaic acid OMI oocyte maturation inhibitor P probability p34cdc2 catalytic subunit of MPF of 34 kDa PBS phosphate buffered saline PCM preconditioned medium PDE phosphodiesterase PDE3A / PDE4 type 3A phosphodiesterase /type 4 phosphodiesterase PGC primordial germ cell pH hydrogen potential PIP2 phosphoinositol bi-phosphate PKA/PKC protein kinase A/ protein kinase C RNA ribonucleic acid rRNA ribosomal RNA Rol rolipram RT room temperature (21°C) SE standard error sec seconds TCM-199 tissue culture medium-199 TLH hepes buffered tyrode`s medium TI telophase I v:v volume:volume w/v weight/volume 5' –AMP 5'–adenosine monophosphate 6-DMAP 6-dimethylaminopurine
xiii

8-Br-cAMP 8-bromo-3',5'-cAMP °C degree celsius % percentage
xiv

General Introduction Although the germ cells are not important to the survival of the individual, they are important for
the survival of the species and the continuation of the life cycle from generation to generation.
The union of the oocyte and the spermatozoa at fertilization gives rise to a new individual. The
mammalian ovary is the organ responsible for the production of mature oocytes and the
production of hormones that permit the development of secondary sexual characteristics and the
successful completion of pregnancy. The ovarian follicle contains a single oocyte surrounded by
multiple layers of distinct somatic cell types. The follicle provides the nutrients and regulatory
signals required for oocyte growth and maturation. Follicles start to grow early during postnatal
life and continue to grow until menopause. Oogenesis occurs early during fetal life and the
number of oocytes present in the ovary is not renewable. The oocyte, which starts with a 4n
complement of DNA, must undergo meiosis, a process of cell division characteristic of germ
cells, to become a haploid egg (1n DNA). Meiosis is initiated at approximately day 75-80 post-
conception. The oocyte progresses through several transitory stages of prophase and arrests at
diplotene stage [Erikson, 1966 #945]. The oocyte is in meiotic arrest at the germinal vesicle
(GV) stage. Oocytes remain in meiotic arrest for many months or even years until they either
ovulate or become degenerated. The maturation of mammalian oocytes in vivo requires the pre-
ovulatory peak of LH; whereas in vitro oocyte maturation occurs spontaneously upon removal of
the oocyte from the follicular environment (Pincus and Enzmann, 1935). Oocyte maturation
involves changes at the nuclear and cytoplasmic level that render the oocyte capable of
undergoing fertilization and embryo development. The mature oocyte undergoes fertilization and
the life cycle begins anew.
Surgical aspiration (Brackett et al., 1982), laparascopy (Sirard and Lambert, 1986) and
ultrasound-guided aspiration (Bousquet et al., 1995) have been used to collect oocytes from fetal,
prepubertal, pregnant and nonpregnant animals. However, most embryos produced in vitro
originate from oocytes collected from ovaries obtained at the slaughterhouse. These ovaries
collected post-mortem provide an inexpensive source of a large number of immature oocytes that
allow large-scale production of embryos in vitro and the testing of new technologies for research
and agriculture applications.

2
Oocytes matured in vitro or in vivo have similar rates of nuclear maturation, fertilization and
cleavage, but differ in their developmental potential (Blondin et al., 1996a; Sirard and Blondin,
1996). In vivo maturation of oocytes yields greater percentages of blastocysts when compared to
in vitro maturation (IVM) (Greve et al., 1987; Leibfried-Rutledge et al., 1987). Oocytes obtained
from postmortem ovaries are extremely heterogeneous in terms of quality, meiotic and
developmental competence (Gordon and Lu, 1990). Information on the meiotic process is
necessary to realize the full potential of in vitro systems for producing embryos since a possible
cause of developmental failure is the incompetence of oocytes from smaller follicles to complete
maturation. If these oocytes are to be used in technologies such as in vitro fertilization, cloning
or transgenesis, then it becomes essential to learn how to control their maturation. A better
understanding of the meiotic control mechanism and of the signaling system from the follicles
will be required to improve the in vitro developmental competence in bovine immature oocytes.
The control of oocyte maturation may appear to be simple, but in fact it is quite complex.
The present thesis will present current knowledge on the control of oocyte maturation and using
the cow oocyte as a model to discuss our results and understand this very important aspect of
oocyte development. The working hypothesis is that inhibition of meiosis of bovine oocytes by
the theca cells involves the interactions of multiple signaling pathways. Specifically, the thesis
explores the effects of morphology on the kinetics of nuclear maturation, the effect of PDE
inhibitors on oocyte maturation, the effect of modulating the PKA and PKC signaling pathways
and the partial characterization of the inhibitory factor secreted by the theca cell monolayers.

CHAPTER I Oogenesis and Meiotic Arrest.

4
The Ovary In mammals, the ovary is the female gonad responsible for the differentiation and release of a
mature oocyte for fertilization and successful propagation of the species (Figure 1.1). Equally
important, the ovary is an endocrine organ that produces steroids to allow the development of
female secondary sexual characteristics and support pregnancy. The bovine ovary has an ovoid
shape and an approximate size of 4 x 2 x 1.5 cm (Dyce et al., 1987). The outermost layer
covering the ovary consists of germinal epithelium (Figure 1.1). Directly underneath the
germinal epithelium there is a layer of dense connective tissue known as the tunica albuginea.
The ovarian follicles, in conjunction with surrounding fibroblasts, collagen and elastic fibers,
form the ovarian cortex located under the tunica albuginea. The ovarian medulla contains the
blood vessels, lymphatic vessels and the nervus terminals. The formation of a functional ovary
depends on three major events taking place during early stages of gonadogenesis: the initiation
of meiosis, the formation of follicles and the differentiation of steroid producing cells.
Figure 1. 1 Diagram of a mammalian ovary.
(Senger, 1997).

5
The Follicle An ovarian follicle is a highly complex unit consisting of distinct cell types. The ovarian follicle
is comprised of several layers of somatic cells surrounding a fluid-filled cavity “Antrum” in
which the oocyte surrounded by somatic cells is bathed. The fluid found in the antrum is known
as follicular fluid. The follicle provides a microenvironment for oocyte growth and it is
responsible for the production of hormones (Gordon, 1994). The oocyte and its companion
somatic cells comprising the follicular unit maintain close association throughout development
from primordial to preovulatory stages. The walls of mature preovulatory follicles consist of
membrana granulosa, theca interna and theca externa. Granulosa cells are cells of epithelial origin
essential for the growth and survival of the oocyte. Granulosa cells are not a homogeneous tissue
but rather specialized subpopulations consisting of the corona radiata, cumulus cells, mural and
antral granulosa cells. The cumulus cells are the subpopulation of ovarian granulosa cells that
surrounds the oocyte. The cumulus cells in close contact with the oocyte are known as corona
radiata. They are in close contact with the oocyte through cytoplasmic extensions across the zona
pellucida (De Loos et al., 1991). The corona radiata communicates the oocyte with other
cumulus cells through the gap junctions thus forming an electrophysiological syncytium (De
Loos et al., 1991). The cumulus cells nourish the oocyte (Buccione et al., 1990) and participate
in the formation of the zona pellucida. In addition, cumulus cells have also been implicated in the
modulation or generation of oocyte maturation inhibitors (Tsafriri et al., 1982; Eppig and Downs,
1984).
The granulosa cells closest to the basement membrane are known as mural granulosa cells, while
those closest to the follicular antrum are known as antral granulosa cells. The follicular antrum
contains the follicular fluid that is made of blood exudate modified by local secretions and
metabolism (Gordon and Lu, 1990). The subpopulations of granulosa cells differ in their
distribution of receptors and steroidogenic characteristics (Rouillier et al., 1998; Rouillier et al.,
1996; Rouillier et al., 1994). The basement membrane or lamina propia is an acellular layer
present between the theca and granulosa cells. The basement membrane contains several types of

6
collagen (collagen IV alpha 1 and alpha 2, reduced amounts of alpha 3-alpha 5), fibronectin,
laminin and proteoglycans (Rodgers et al., 1999). The theca interna and theca externa are
stromal or fibroblastic cells that constitute the outermost coat of the preovulatory follicle. An
extensive capillary network irrigates theca cells. The theca interna are the major source of
androgens during the final stage of development of the Graafian follicle (Moor, 1977). Follicles
can be classified as primordial follicles, preantral follicles (primary and secondary follicles),
antral and preovulatory follicles.
Folliculogenesis Folliculogenesis is the process responsible for the development of ovulatory follicles and the
release of one or more mature oocytes at a fixed interval throughout the reproductive life of a
female. Folliculogenesis is resumed after a long quiescent phase and involves sequential
subcellular and molecular transformations by various components of the follicle. During
postnatal life, ovarian follicles continue to grow, mature and either ovulate or regress. Follicles
are recruited continuously until the original store is exhausted. The follicle population in the cow
is divided more or less evenly between each of the two ovaries. In monotocous species, the
follicle selected to ovulate is the fastest growing functional unit in the body of the adult female
mammal. The bovine follicle grows 300 to 400 fold in diameter from the primary (50 µm) to the
preovulatory (15-20 mm) stage (Rajakoski, 1960b). The entire process of follicular growth from
the primordial stage (50-60 µm) to the ovulatory stage (10-15 mm) takes approximately 180 days
in cattle (Lussier et al., 1987). This extended period allows the building of the zona pellucida and
the accumulation of a number of known and unknown products required for the fertilization and
early embryonic development events. Follicles either ovulate a mature oocyte or undergo atresia.
Primordial germ cells (PGC) Oocytes present in the adult ovary develop from a definite number of primordial germ cells
(PGC). The mechanisms by which the PCG migrate from the extragonadal sites to the gonadal
ridge are not well understood (Bryskov and Hoyer, 1994). They form the cortical proliferations
and germ cell cords to form the primitive ovary and give rise to oogonia (Eddy et al., 1981;
Hashimoto and Eguchi, 1955; Witschi, 1948). The migration of PGC is due to a combination of

7
passive transfer and self-propulsion (Kuwana and Fujimoto, 1983). Primordial germ cells
proliferate during migration and have undergone six or more divisions by the time they colonize
the future gonad. The c-kit protein and its ligand (KL) appear to play an important role in the
proliferation, survival and migration of PGC (Besmer et al., 1993). Once established in the
developing ovary, the proliferating PGC begin to differentiate into oogonia.
Oogonia The oogonia are the stem cells that give rise to all the oocytes in the ovary (Rüsse, 1983). The
PGC loose the ability to undergo amoeboid movement, stain less intensively for alkaline
phosphatase activity and they become more spherical with fewer cytoplasmic organelles
(Donovan et al., 1986). The population of oogonia goes through a predetermined, species-
specific, number of mitotic cycles until the cells enter the prophase of meiosis and become
oocytes. The prophase of meiosis is traditionally separated into five sequential stages: leptotene,
zygotene, pachytene, diplotene and diakinesis. The leptotene stage is resumed by the end of an
active period of pre-meiotic DNA synthesis. At the leptotene stage, each chromosome condense
from its interphase conformation to produce a fine discrete thread. Each chromosome has
replicated and consists of two sister chromatids. Zygotene starts as soon as the synapsis or
intimate pairing of homologous chromosomes is initiated. Each gene is brought in close
juxtaposition with its homologous gene on the opposite chromosome. Each chromosome pair is
usually called a bivalent, but each homologous chromosome consists of two sister chromatids or
tetrad. The cells are said to have entered the pachytene stage of prophase as soon as synapsis is
completed. The synaptonemal complexes are structures that hold the two homologous
chromatids together. The maternal and paternal chromatids undergo homologous recombination
to allow the exchanges or crossovers between two nonsister chromatids. When the oocyte
reaches the pachytene stage, it becomes enclosed in a follicle. The diplotene stage starts with the
unpairing of homologous chromosomes in each bivalent after the crossovers are completed. The
bivalent remains joined by one or more chiasmata (crossover-sites). The chromosomes take a
diffused aspect that permits a period of active RNA synthesis. The nucleus of diplotene oocytes
is known as germinal vesicle (GV). The oocyte at the GV stage is a diploid cell (2n), which has
twice the normal amount of DNA since the first meiotic prophase is stopped at diplotene since

8
the early post-natal period. The oocytes are considered to be in meiotic arrest. Oocytes remain in
meiotic arrest for many months or even years. The mammalian ovary has only a finite supply of
oocytes. It is estimated that bovine ovaries contain an estimated 420,000 at birth that is reduced
to less than 3000 by 20 years of age (Erikson, 1966; Gosden, 1995).
Primordial follicles The oogonia enlarge and initiate meiosis independently of any endocrine stimulation to form
primary oocytes. The primary oocyte becomes arrested at the pachytene or dictyate stage of the
first meiotic prophase (Byskov and Hoyer, 1994). In cattle, meiosis begins on the 82nd day of
gestation (Rüsse, 1983). Primordial follicles consist of a dictyate-oocyte surrounded by a single
layer of flattened pregranulosa cells (Byskov, 1978). The oocyte and the follicular cells are
interdependent. The oocyte requires the presence of granulosa cells to grow and survive (Byskov
and Lintern-Moore, 1973; Picton, 2001). Pregranulosa cells rest on a delicate basement
membrane opposite the stromal cells that give origin to the theca cells (Gougeon, 1996). The
precursors of theca cells may already be present at the very outset of follicular growth
(Hirshfield, 1991b). It has been proposed that theca cells are already present in primordial and
primary rat follicles (Hirshfield, 1991b). This would suggest that the interaction of the theca and
granulosa cells might play a role in the regulation of the growth and differentiation of the follicle
throughout all the stages of folliculogenesis.
Primordial follicles are the fundamental developmental unit of the mammalian ovary. The store
of primordial follicles is not renewable and serves the entire reproductive life span of the adult
(Figure 1.2). Primordial follicles in the bovine are first detected on the 90th day of gestation
(Erikson, 1966; Szollosi, 1991). In cattle, the primordial follicles have a diameter equal to or
smaller than 40 µm (Braw-Tal and Yossefi, 1997; Fair, 1995; Van Den Hurk et al., 1997; van
Wezel and Rodgers, 1996). Primordial follicles each contain one oocyte measuring
approximately 30 µm in diameter (Picton, 2001). Primordial follicles are located in the
peripheral cortex of the ovary (Zamboni, 1974). As the follicles and oocytes start to grow, they
move deeper into the cortex of the ovary. The follicle grows out through the cortex as the antrum
develops and becomes visible on the surface of the ovary. The mechanism of activation of
primordial follicles and oocytes remains unknown. As soon as the primordial follicle store is

9
established at approximately day 140 day of gestation in the cow, follicle recruitment begins and
continues without halting for the rest of life or until the ovary is depleted.
10
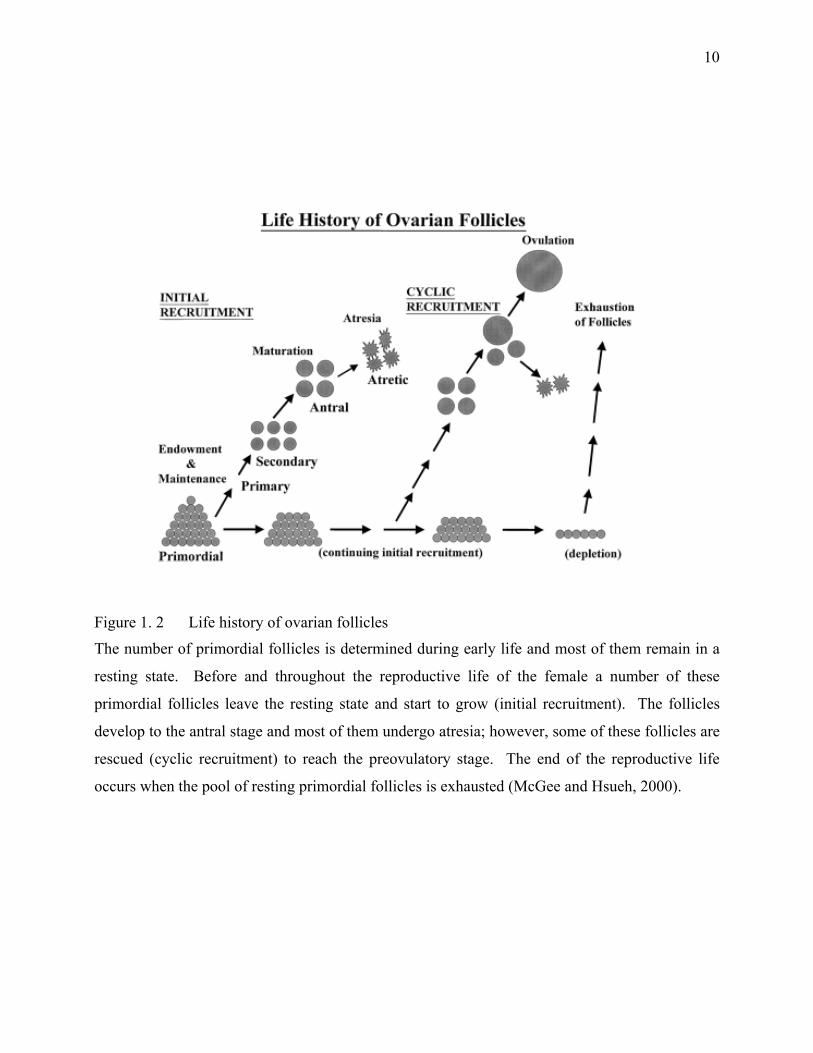
Figure 1. 2 Life history of ovarian follicles
The number of primordial follicles is determined during early life and most of them remain in a
resting state. Before and throughout the reproductive life of the female a number of these
primordial follicles leave the resting state and start to grow (initial recruitment). The follicles
develop to the antral stage and most of them undergo atresia; however, some of these follicles are
rescued (cyclic recruitment) to reach the preovulatory stage. The end of the reproductive life
occurs when the pool of resting primordial follicles is exhausted (McGee and Hsueh, 2000).

11
Primary Follicles Bovine primordial follicles are activated to become primary follicles on the 140th day of gestation
(Rüsse, 1983). A primary follicle contains an oocyte with a diameter of approximately 30 µm
that is surrounded by cuboidal granulosa cells (Hyttel et al., 1997; Rüsse, 1983; van Wezel and
Rodgers, 1996). The bovine oocyte starts to grow when there are approximately 40 granulosa
cells present (Braw-Tal and Yossefi, 1997). Thus oocytes from primordial and primary follicles
are not significantly different in size. However, important changes take place during the primary
follicle stage. The corona radiata develops gap junctions with the oocyte and the zona pellucida
begins to form between the two cell types (Fair, 1995; Picton, 2001; Rüsse, 1983). The zona
pellucida will not completely surround the oocyte until the follicle reaches the late preantral stage
(Braw-Tal and Yossefi, 1997).
Secondary Follicles Secondary follicles appear on day 210 of gestation in cattle when the follicular cells of the
primary follicles undergo intensive mitotic division. A secondary follicle contains at least two
layers of granulosa cells and the oocyte measures between 50 and 60 µm in diameter (Hyttel et
al., 1997; Rüsse, 1983). Furthermore, the theca cells are identifiable outside the basement
membrane and the follicle contains a fine capillary network.
Tertiary or Antral Follicles Tertiary or antral follicles are characterized by the presence of a cavity known as antrum. The
antrum is a cavity filled with follicular fluid. The antrum is first detected in bovine follicles
measuring between 0.12 and 0.28 mm in diameter (Lussier et al., 1987; Monniaux et al., 1997).
The first antral follicles appear around the 230th day of gestation in the bovine (Rüsse, 1983).
Tertiary follicles have an extensive network of gap junctions that permits the transfer of nutrients
and regulatory signals between the oocyte and the granulosa cells (Espey, 1994). Theca cells
appear to originate from the stromal mesenchyme (Harrison and Weir, 1977; Weakly, 1966).
Granulosa cells first acquire the LH receptor when the follicle measures between 8-9 mm in

12
diameter (Ireland and Roche, 1983a). The perivitelline space is formed between the oocyte and
the zona pellucida just before ovulation. The blood supply of the theca layer changes
dramatically during the final stages of follicle maturation and develops into an inner plexus in the
theca interna and an outer plexus derived from the stromal capillaries (Yamada et al., 1995).
The intensity of RNA synthesis in oocytes correlates well with the stage of their growth and their
corresponding changes in morphology of nucleoli [Crozet, 1989 #747]. The RNA synthetic
activity decreases gradually as the oocytes reach their full size. Bovine oocytes from follicles
between 0.5 and 3 mm actively synthesize RNA (Crozet, 1989). RNA synthesis decreases after
the follicle size has reached 3 mm (Crozet, 1989). However, low levels of transcription continue
until the oocyte has reached a diameter of 110 µm (Fair et al., 1995). The continued nucleolar
transcription may play an important role in the acquisition of the developmental competence of
the oocyte. Oocytes with a diameter larger than 110 µm have an electron-dense fibrillar nucleoli
and lacked transcriptional activity (Fair et al., 1996; Hyttel et al., 1997).
The germinal vesicle or nucleus of the bovine oocyte contains one or two nucleoli (Crozet, 1989;
Crozet et al., 1986). Nucleoli are the sites of rRNA synthesis. Primordial follicles contain
transcriptionally inactive nucleoli that are characterized by the presence of exclusively granular
components interspaced with multiple vacuoles, whereas transcriptionally active nucleoli have a
fibrillo-granular appearance (Hyttel et al., 1997). When an oocyte begins to grow in the
primordial follicle, the uncondensed loops of chromatin in the dictyate state ensure transcription
of the required elements. The mRNA product is either translated immediately or stored under
special conditions. The oocyte has the ability to accumulate mRNA in a stable form. After the
beginning of maturation, transcription stops within hours. It is postulated that oocytes originating
from competent or differentiated follicles contain the right mRNA in sufficient amount. These
mRNA accumulated in the oocyte are important during the progression of the embryo through the
maternal to zygotic transition.
Bovine antral follicles require approximately 42 days to reach the preovulatory size. Thus the
development of the follicle through the antral stage requires a period equivalent to 2 estrous
cycles (Lussier et al., 1987).
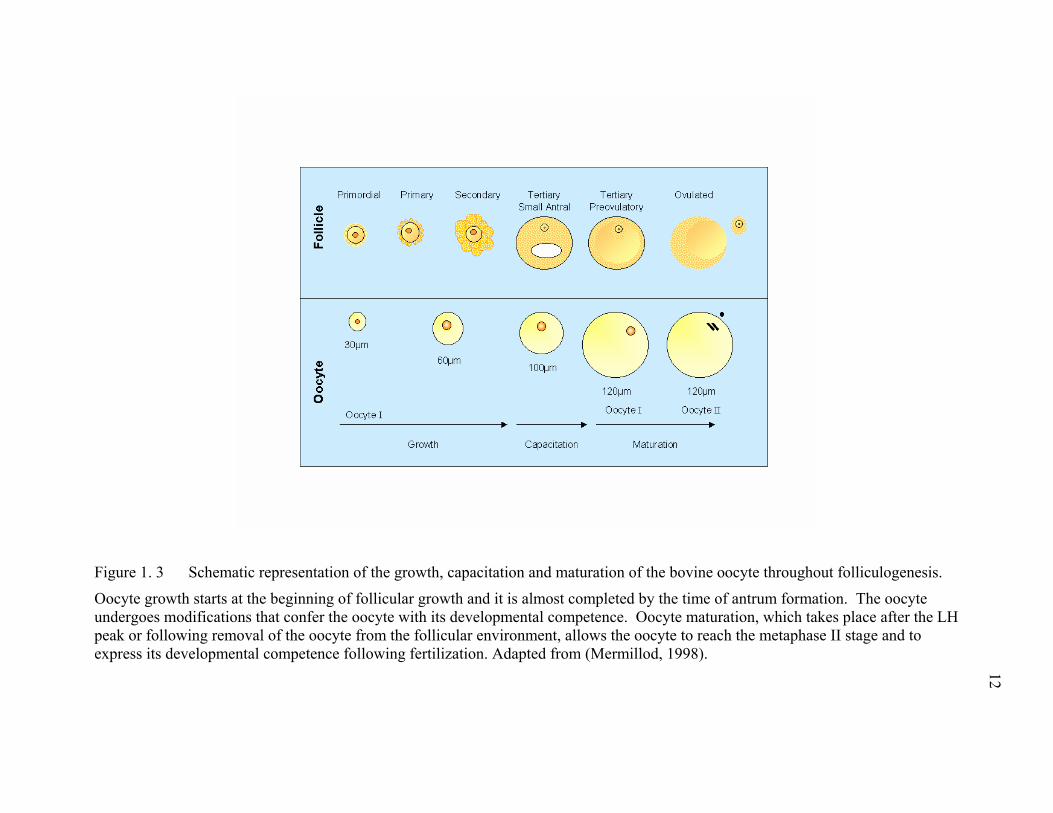
Figure 1. 3 Schematic representation of the growth, capacitation and maturation of the bovine oocyte throughout folliculogenesis.
Oocyte growth starts at the beginning of follicular growth and it is almost completed by the time of antrum formation. The oocyte undergoes modifications that confer the oocyte with its developmental competence. Oocyte maturation, which takes place after the LH peak or following removal of the oocyte from the follicular environment, allows the oocyte to reach the metaphase II stage and to express its developmental competence following fertilization. Adapted from (Mermillod, 1998). 12

14
Developmental Competence Developmental competence is the ability of the oocyte to produce normal, viable and fertile
offspring after fertilization. The final steps of oocyte maturation are crucial to the acquisition of
functional properties necessary for further development (Hyttel et al., 1997). The developmental
competence of the oocyte is acquired within the ovary during the stages that precede ovulation or
in case of in vitro maturation, precede the isolation of the oocyte from its follicle.
The acquisition of developmental competence is a gradual process during follicular development.
Gradual increase refers to the percentage of competent oocytes and not to a higher competence of
a single oocyte (Gandolfi, 1998). Developmental competence is a difficult parameter to assess
since embryonic development may fail due to reasons independent of oocyte quality.
Developmental competence is usually expressed as the percentage of oocytes that can develop to
the blastocyst stage (Gandolfi, 1998). However, development to the blastocyst stage does not
guarantee that the embryo will develop to term. Other aspects used to evaluate developmental
competence include morphological evaluations such as number of blastomeres or the ratio
between inner cell mass and trophoectoderm cell numbers. Functional evaluations such as
metabolic rates and the ability to resume development after freezing should also be considered to
provide a more complete idea of the developmental potential of the oocyte.
Multiple studies have been carried out to study the factors affecting the developmental
competence of the oocyte. The size (Pavlok et al., 1992; Tan and Lu, 1990) and the quality of the
follicle of origin (Blondin et al., 1995; Blondin and Sirard, 1994; Hazeleger et al., 1995)
influence the developmental capacity of bovine oocytes. Studies that followed the fate of
individual oocytes according to the specific follicle of origin have corroborated that
developmental competence of the oocyte increases with follicular size (Blondin and Sirard,
1995). Oocytes from bovine follicles greater than 6 mm in diameter produce blastocysts in vitro
at substantially greater rates than those from 2 to 6 mm follicles (Lonergan et al., 1994b) and
follicles smaller than 2 mm yield oocytes capable of fertilization, but lack the ability to cleave
beyond the 8-cell stage (Pavlok et al., 1992). The follicle must reach a diameter of at least 2-3
mm before the oocyte reaches a satisfactory developmental competence.

15
Some oocytes have acquired an intrinsic capacity to develop into an embryo after IVM-IVF-IVC
at the follicular stage of 3 mm, but and the proportion of competent oocytes does not increase
during development up to 7 mm (Hendriksen et al., 2000). It appears that the oocyte requires an
additional "prematuration" to express this competence (Hendriksen et al., 2000). In vivo, this
pre-maturation occurs during preovulatory growth before the LH surge. Other factors besides
follicular size may be critical for the oocyte to acquire developmental competence. Some large
follicles contain developmentally incompetent oocytes; while, some medium follicles contain
competent oocytes (Blondin and Sirard, 1995). Follicular atresia may promote the acquisition of
developmental competence (Blondin and Sirard, 1995; Hendriksen et al., 2000). However, the
health of the follicle in terms of atresia cannot be used to predict the developmental competence
of the oocyte. Oocytes obtained from slightly atretic, intermediate or nonatretic follicles posses
similar developmental competence (Blondin et al., 1996b; Blondin and Sirard, 1995).
The ovarian morphology is another parameter used to estimate the developmental competence of
the oocyte. The number and size of the follicles present in the ovary at the time of aspiration may
be used to select oocytes with higher developmental competence. Oocytes retrieved from ovaries
that have at least one follicle larger than 10 mm in diameter or with more than 10 follicles of 2 to
5 mm have a high developmental potential. In contrast, oocytes retrieved from ovaries with
fewer than 10 follicles of 2 to 5 mm or no follicle larger than 10 mm reached the blastocyst stage
at a lower rate and the blastocysts had low cell numbers (Gandolfi et al., 1997). The composition
of the follicular fluid may also play a role in developmental competence (Arlotto et al., 1996;
Hazeleger et al., 1995; Madison et al., 1992; Pavlok et al., 1992). The developmental
competence of the oocyte may also be lost during in vitro maturation. Extended incubation
during oocyte maturation leads to decreased developmental competence that could be the result
of oocyte aging. The time at which the oocytes are aspirated from the post-mortem ovaries
affects the development rates. A delay of 4 h between the slaughter of the cow and oocyte
aspiration has been reported to yield the highest rates of development after 5 days of in vitro
development (Blondin et al., 1995). Cumulus cells are sometimes lost during the oocyte
aspiration. The number and quality of cumulus cells surrounding the oocyte are important in
developmental competence (Blondin and Sirard, 1995; Gandolfi et al., 1997). The developmental

16
competence of bovine oocytes with corona cells only (corona-enclosed oocytes) is not
comparable with that of COC (Leibfried Rutledge et al., 1989).
The Estrous Cycle Heifers have their first estrous cycle between 8 and 12 months of age. On average, the estrous
cycle in the cow lasts approximately 21 days. The period of receptivity to be mounted or estrus
lasts approximately 12 to 15 h and ovulation takes place 10 to 12 h after this period. The estrous
cycle is divided into a luteal and a follicular phase. The luteal phase starts after ovulation when
the corpus luteum (CL) is formed from the wall of the collapsed follicle and continues until day
16-18. Progesterone is the main hormone secreted during the luteal phase. The follicular phase
starts after the regression of the corpus luteum, which occurs approximately on day 18 of the
estrous cycle. The preovulatory follicle produces high levels of estradiol leading to the standing-
heat behavior and stimulating the release of LH to induce ovulation. If the female does not
become pregnant, then the CL regresses and the level of FSH, which is released by the pituitary
gland increases and a new cycle begins. If the female is pregnant, then the CL persists and
continues to secrete progesterone.
Follicular Waves in Cattle Cows have either 2 or 3 follicular waves per estrous cycle (Ginther et al., 1989c; Knopf et al.,
1989; Sirois and Fortune, 1988) and each follicular wave lasts between 7-9 days (Evans et al.,
1994). The number of waves per cycle is independent on the age or breed of the animal. A
follicular wave consists of a cohort of follicles that undergoes synchronous development during
the final phase of growth (Ginther et al., 1989a; Ginther et al., 1989b; Knopf et al., 1989). The
wave-like pattern of follicular growth has been observed in heifers as young as two weeks of age.
The follicles continue to grow and regress throughout the prepubertal period, however, ovulation
does not occur until the animal attains puberty. The follicular waves continue even during
pregnancy (Bergfelt and Ginther, 1996; Ginther et al., 1996). The emergence of successive
follicular waves during the estrous cycle has been associated with increases in circulating
concentrations of FSH (Adams et al., 1992; Hamilton et al., 1992; Suntherland et al., 1994),
which precede the wave emergence by 1 day (Adams et al., 1992)(Figure 1.4). The FSH receptor

17
is expressed in granulosa cells of all growing follicles, starting in some follicles with only one
layer of granulosa cells (Xu et al., 1995).
Follicular growth involves three separate processes: recruitment, selection and dominance
(Ginther et al., 2001a; Ginther et al., 2001b). Follicle recruitment is a process whereby a cohort
of antral follicles begins to grow beyond 4 mm in diameter in the presence of sufficient
gonadotropin to permit their progress toward ovulation. The maturational step associated with
follicle recruitment is the appearance of aromatase activity within the granulosa layer. The
aromatase activity is first detected in bovine follicles with a diameter of 3 to 4 mm. (Ginther et
al., 1996). These follicles are now susceptible to undergo recruitment. A follicle is selected out
of a cohort of follicles to undergo preferential growth and to become the dominant follicle, while
the other follicles become subordinate. The dominant follicle avoids atresia, inhibits the
recruitment of a new cohort of follicles and acquires competence to achieve ovulation (Adams et
al., 1992; Ginther et al., 1989a; Ginther et al., 1989c; Spicer and Echternkamp, 1986). The
declining concentrations of FSH after wave emergence have been implicated in the mechanism of
selection of a dominant follicle (Adams et al., 1993). If this dominant follicle develops during
the follicular phase, it ovulates. However, dominant follicles that develop during the luteal phase
of the estrous cycle, regress rather than ovulate, due to the absence of the high preovulatory LH
levels (Webb et al., 1992).
18
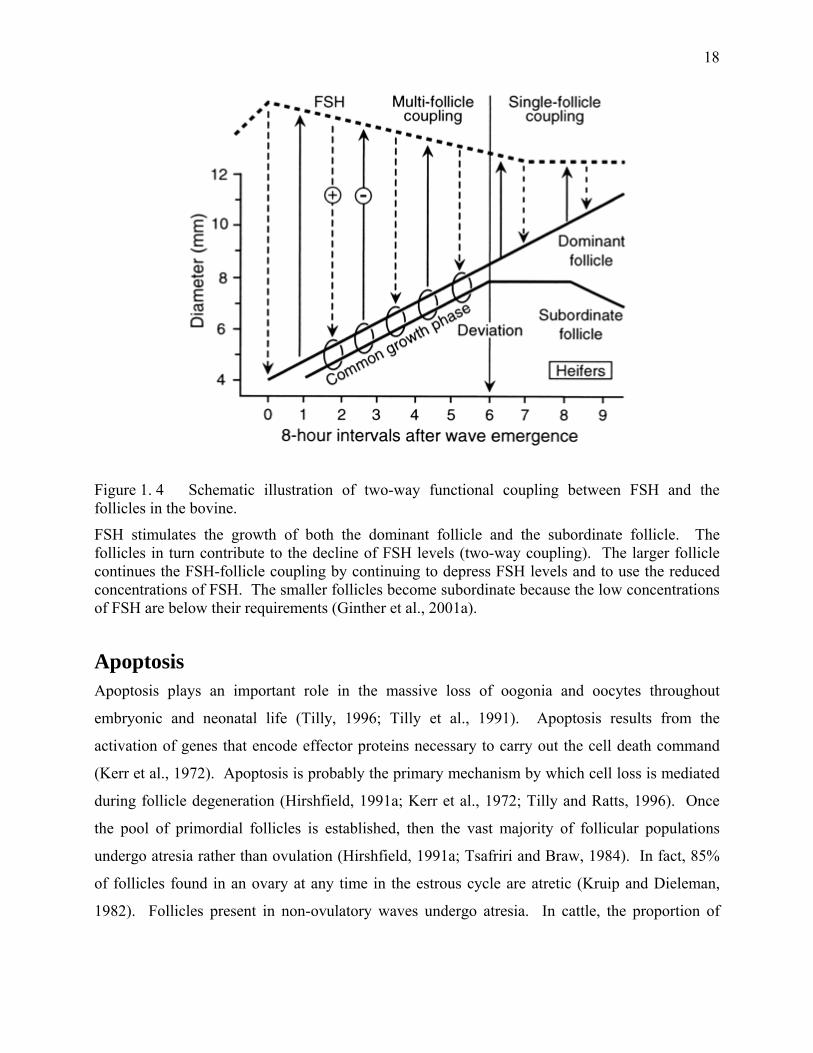
Figure 1. 4 Schematic illustration of two-way functional coupling between FSH and the follicles in the bovine.
FSH stimulates the growth of both the dominant follicle and the subordinate follicle. The follicles in turn contribute to the decline of FSH levels (two-way coupling). The larger follicle continues the FSH-follicle coupling by continuing to depress FSH levels and to use the reduced concentrations of FSH. The smaller follicles become subordinate because the low concentrations of FSH are below their requirements (Ginther et al., 2001a).
Apoptosis Apoptosis plays an important role in the massive loss of oogonia and oocytes throughout
embryonic and neonatal life (Tilly, 1996; Tilly et al., 1991). Apoptosis results from the
activation of genes that encode effector proteins necessary to carry out the cell death command
(Kerr et al., 1972). Apoptosis is probably the primary mechanism by which cell loss is mediated
during follicle degeneration (Hirshfield, 1991a; Kerr et al., 1972; Tilly and Ratts, 1996). Once
the pool of primordial follicles is established, then the vast majority of follicular populations
undergo atresia rather than ovulation (Hirshfield, 1991a; Tsafriri and Braw, 1984). In fact, 85%
of follicles found in an ovary at any time in the estrous cycle are atretic (Kruip and Dieleman,
1982). Follicles present in non-ovulatory waves undergo atresia. In cattle, the proportion of

19
atretic follicles increases with follicle size (Rajakoski, 1960a). A higher proportion of bovine
oocytes with degenerated cumuli are recovered from follicles over 3 mm in diameter than from
follicles 1 to 3 mm in diameter (Leibfried and First, 1979b). The first signs of atresia are
manifested by the degeneration of the granulosa cells that loose their aromatase activity and
undergo apoptosis (Gordon and Lu, 1990). Later on, the theca cells undergo hypertrophy and
their androsterone production decreases (Driancourt, 1991; Greenwalt and Terranova, 1988).
The oocyte is affected only at the very last stages of follicular atresia (Kruip and Dieleman, 1982;
Leibfried and First, 1979b).
Under normal conditions, the oocytes acquire their developmental competence late in the
follicular phase of the estrous cycle (Blondin et al., 1995). The late onset of atresia may allow
the oocytes to retain their developmental competence for a period of time even if follicular atresia
has already started (Hazeleger et al., 1995). It is also possible that the positive effects of atresia
on in vitro developmental potential might be the result of a longer growth period during which
the oocyte is preparing itself for maturation, fertilization and development (de Wit, 2001). On
the other hand, the developmental competence of the oocyte in vitro will be lower if the degree of
follicular atresia is too advanced (Blondin and Sirard, 1995).
Cumulus-oocyte complexes with signs of atresia undergo organelle rearrangement and nuclear
changes that show some similarities with the processes associated with final maturation in the
dominant oocytes. This phenomenon has been described as "pseudomaturation" (Assey et al.,
1994). Pseudomaturation might occur because follicular atresia mimics some of the post-LH
changes occurring in the follicle (Wise and Maurer, 1994; Wise et al., 1994). The rapid decrease
in estradiol, the rise in androgen, progesterone and PGE2, the presence of inflammatory
conditions (Ireland and Roche, 1982; Ireland and Roche, 1983b) perhaps induce a series of
changes in the oocyte that are extremely important in the acquisition of developmental
competence (Blondin et al., 1996b).
Meiosis Meiosis from the Greek Meiōsis, meaning reduction, consists of two successive cell divisions
following one round of DNA replication. Meiosis gives rise to four haploid cells from a single

20
diploid cell. This type of cell division is characteristic of germ cells. Meiosis up to the diplotene
stage occurs in the fetal ovary. During the first meiotic division, maternal and paternal genes are
exchanged before the pairs of chromosomes are divided into two daughter cells, each containing
1n chromosome and 2c DNA. The second meiotic division occurs without being preceded by
DNA synthesis and nuclear reformation. Haploid germ cells are formed with a 1n set of
chromosomes and 1c DNA. The two meiotic divisions of the oocyte are asymmetrical, resulting
in expulsion of polar bodies. Meiosis in each female germ cell results in a single egg and two
polar bodies.
Oocyte Maturation Oocyte maturation is a complex phenomenon during which the oocyte progresses from the
diplotene to the metaphase II stage (nuclear maturation) (Figure 1.7). The transition from the
diplotene stage to metaphase is called diakinesis. The oocyte resumes meiosis in response to the
ovulatory LH surge (Callesen et al., 1986; Channing et al., 1978; Dieleman et al., 1983; Ireland
and Roche, 1982; Masui and Clarke, 1979; Peng et al., 1991; Pincus and Enzmann, 1935) or
removal from the follicle (Pincus and Enzmann, 1935). During diakinesis, the nuclear membrane
starts to fold, the nuclear pores disappear and then the nuclear membrane fragments before
rapidly disappearing to leave only small sacs with double walls (Kubelka et al., 1988; Szollosi et
al., 1972). These events are known as germinal vesicle breakdown (GVBD), which is the first
visible sign of meiotic resumption. The nucleolus disappears rapidly after coming in contact with
the cytoplasm. In cattle, GVBD occurs within hours after removal from the follicle or the
ovulatory LH signal. In cattle, by 6.6 h of culture, 50% of the oocytes had undergone GVBD (De
Loos et al., 1994; Sirard et al., 1989). Then, the chromosomes condense further. The
kinetochores appear and the microtubules pull the chromosomes and they form the metaphasic
plate of MI. The separation of the homologous chromosomes and the migration of the
chromosomes to their respective poles take place during anaphase I. During telophase I, the
chromosomes found at each pole are surrounded by a nuclear membrane. The second meiotic
division without chromosome replication takes place immediately and the oocyte reaches the
metaphase II. The oocyte remains arrested at the metaphase II stage until fertilization takes place
and the oocyte completes meiosis and forms the pronucleus.
21
Oocyte maturation also involves transformations at the cytoplasmic level that prepare the cell to
support fertilization and early embryonic development (cytoplasmic maturation). The completion
of nuclear maturation alone does not guarantee subsequent embryo development (Sirard et al.,
1989; Yang et al., 1998).
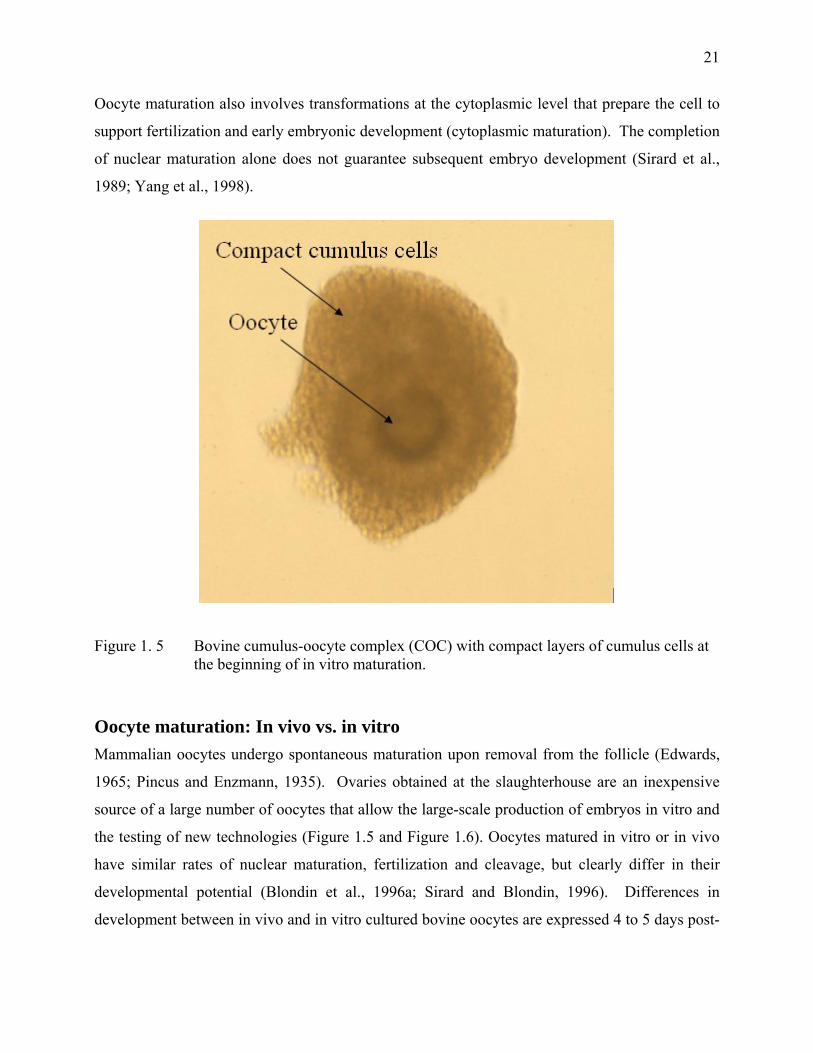
Figure 1. 5 Bovine cumulus-oocyte complex (COC) with compact layers of cumulus cells at the beginning of in vitro maturation.
Oocyte maturation: In vivo vs. in vitro Mammalian oocytes undergo spontaneous maturation upon removal from the follicle (Edwards,
1965; Pincus and Enzmann, 1935). Ovaries obtained at the slaughterhouse are an inexpensive
source of a large number of oocytes that allow the large-scale production of embryos in vitro and
the testing of new technologies (Figure 1.5 and Figure 1.6). Oocytes matured in vitro or in vivo
have similar rates of nuclear maturation, fertilization and cleavage, but clearly differ in their
developmental potential (Blondin et al., 1996a; Sirard and Blondin, 1996). Differences in
development between in vivo and in vitro cultured bovine oocytes are expressed 4 to 5 days post-

22
fertilization at the morula-blastocyst stage (Blondin et al., 1996a; Hyttel et al., 1997). In vivo
maturation of oocytes yields greater percentages of blastocysts when compared to in vitro
maturation (IVM) (Greve et al., 1987; Leibfried-Rutledge et al., 1987). The fact that
approximately 85% of ovulated oocytes in inseminated cows develop into an embryo capable of
establishing pregnancy is indicative of the high developmental competence of fully capacitated
oocytes that have matured and ovulated in vivo. On the contrary, only one third of in vitro
matured oocytes develop to the morula-blastocyst stage regardless of whether they are fertilized
in vivo (Gordon and Lu, 1990) or in vitro (Brackett and Zuelke, 1993; Dominko and First, 1997;
Gordon, 1994).
Important factors either in the form of proteins or in the form of stable mRNAs are stored during
oocyte growth and final follicular maturation after the growth has been completed (Blondin and
Sirard, 1995). Oocytes matured in vitro bypass oocyte capacitation (Hyttel et al., 1997) and other
substantial changes that take place in vivo under the influence of LH and the follicular
environment (Motlik and Fulka, 1981). The term “oocyte capacitation” describes the
ultrastructural modifications that take place in the oocyte of dominant follicles before the LH
peak (Figure 1.3). These modifications permit the oocyte to attain full developmental
competence (Hyttel et al., 1997). The formation of the male pronuclei is significantly reduced in
oocytes matured in vitro (69%) when compared to oocytes matured in vivo (88%) (Leibfried-
Rutledge et al., 1987). However, superstimulation of cows with 6 doses of FSH and a dose of LH
after a 48 h coasting period results in 80% ± 9% (mean ± SEM) blastocyst yield when the ovum
pick up is carried out 6 h after the LH injection (Blondin et al., 2002). The developmental
potential of the COC is greatly affected by the coasting period between hormonal stimulation and
ovary collection (Blondin and Sirard, 1997) and the interval between ovaries collection and
oocyte aspiration (Blondin et al., 1995).
The oocyte ovulated during the normal estrous cycle of the cow originates from the dominant
follicle. The dominant follicle grows from 2 to 15 mm in approximately 5 days (Driancourt,
1991). Most oocytes collected for in vitro maturation originate from subordinate or growing
follicles that are at least 4 to 10 days away from any possible ovulation. Even though most of

23
these oocytes complete their nuclear maturation, few develop to the blastocyst stage (van de
Leemput et al., 1999).
Even stringent selection procedures do not always yield oocytes capable of developing to the
blastocyst stage after standard in vitro maturation/in vitro fertilization (IVM/IVF) procedures. It
is hypothesized that these oocytes that fail to undergo normal fertilization and development is a
result of an incomple cytoplasmic maturation.
Oocyte maturation in vitro is independent of the estrous cycle (Arlotto et al., 1996; Fukui and
Sakuma, 1980). Germinal vesicle breakdown occurs earlier and meiotic maturation proceeds
more rapidly during in vitro than in vivo maturation (Hyttel et al., 1997). Different patterns of
protein synthesis have been reported for oocytes matured in vivo versus those matured in vitro
(Kastrop et al., 1991a). Differences can also be found at the ultrastructural levels such as the
localization of cortical granules (Szollosi, 1967). Finally, cumulus cell expansion is more
extensive in vivo than in vitro (Hendriksen et al., 2000).
Nuclear maturation Nuclear maturation refers to the progression of the oocyte nucleus from the germinal vesicle to
the metaphase II stage. Nuclear maturation involves GVBD, condensation of chromosomes,
metaphase I spindle formation, separation of the homologous chromosomes with extrusion of the
first polar body and arrest at metaphase II (Kubelka et al., 1988). The nuclear membrane starts to
fold, the nuclear pores disappear and then the nuclear membrane undergoes fragmentation and
rapidly disappears (Szollosi et al., 1972). A period of 24 h is necessary for a bovine oocyte to
complete nuclear maturation (Sirard et al., 1989). It appears that nuclear maturation follows the
same pattern in vivo and in vitro (Hyttel et al., 1986b). Nuclear maturation involves changes in
protein synthesis patterns (Hunter and Moor, 1987). Bovine oocytes undergo marked changes in
the patterns of protein synthesis after GVBD in vitro and in vivo, whereas oocytes that remain at
GV stage have consistent protein synthesis patterns (Kastrop et al., 1990b; Kastrop et al., 1991a).
The ability of the oocyte to complete meiosis is known as meiotic competence. Meiotic
competence is acquired gradually during follicular growth. Oocytes first acquire the capacity to

24
undergo GVBD and chromosome condensation, then further follicular development is required to
acquire the ability to progress to the metaphase I (Tsafriri and Channing, 1975) and finally they
acquire the ability to reach metaphase II (Sorensen and Wassarman, 1976). The ability to
complete the MI to MII transition coincides with the achievement of full size and with the
process of nucleolar compaction (Motlik et al., 1984).
Growing oocytes can be categorized as incompetent or competent to resume meiosis (Arlotto et
al., 1996; Szybek, 1972). Incompetent bovine oocytes remain at the GV stage because they do
not have enough cyclin B to progress beyond prophase I in sufficient quantities (Levesque and
Sirard, 1996). Meiotic competence is associated with an increased concentration of p34, which is
being accumulated at the end of oocyte growth (Chesnel and Eppig, 1995; de Vantery et al.,
1996; de Vantery et al., 1997), and also on specific dephosphorylations of inactive
phosphorylated p34 on threonine and tyrosine residues by cdc25 phosphatase (Dunphy and
Kumagai, 1991). The acquisition of full meiotic competence coincides with the reduction of the
nucleolar transcriptional activity in bovine oocytes (Hyttel et al., 1997; Motlik et al., 1984).
Meiotic competence is closely correlated with oocyte size, which in turn is correlated with
follicle size (Armstrong, 2001). The size of the antral follicle at which the oocyte acquires
meiotic competence is species-specific (Wickramasinghe and Albertini, 1993). Bovine oocytes
acquire the ability to complete GBVD and meiosis by the time the antral follicle reaches 2-3 mm
in diameter (Fair et al., 1995; Lonergan et al., 1994b; Motlik and Fulka, 1986). Meiotic
competence is also related to oocyte diameter, since bovine oocytes must have a diameter of 110
µm to complete nuclear maturation to the MII stage (Fair et al., 1995; Otoi et al., 1997).
Bovine oocytes with an inside-zona diameter smaller than 95 µm are unable to resume meiosis in
vitro. A high proportion of bovine oocytes are able to resume meiosis to the MI stage once the
oocyte diameter is at least 100 µm (Fair et al., 1995; Otoi et al., 1997). However, the oocyte must
measure 110 µm or more to reach the MII stage (Fair et al., 1995). The ability to develop to the
blastocyst stage in vitro increases with oocyte growth (Arlotto et al., 1996; Fair et al., 1995;
Harada et al., 1997). Cleavage and blastocyst rates increased in parallel with meiotic competence
and significantly higher developmental rates have been obtained when the diameter of fertilized
oocytes is greater than 120 µm (Hazeleger et al., 1995). The developmental potential is

25
apparently similar in oocytes originating from non-atretic and early atretic follicles (Blondin et
al., 1996a; Blondin and Sirard, 1995; Fair et al., 1995). Once the oocyte becomes meiotically
competent, inhibitory factors are necessary to maintain the oocyte in meiotic arrest.
Figure 1. 6 Bovine cumulus-oocyte complex (COC) with expanded layers of cumulus cells after 24h of in vitro maturation.
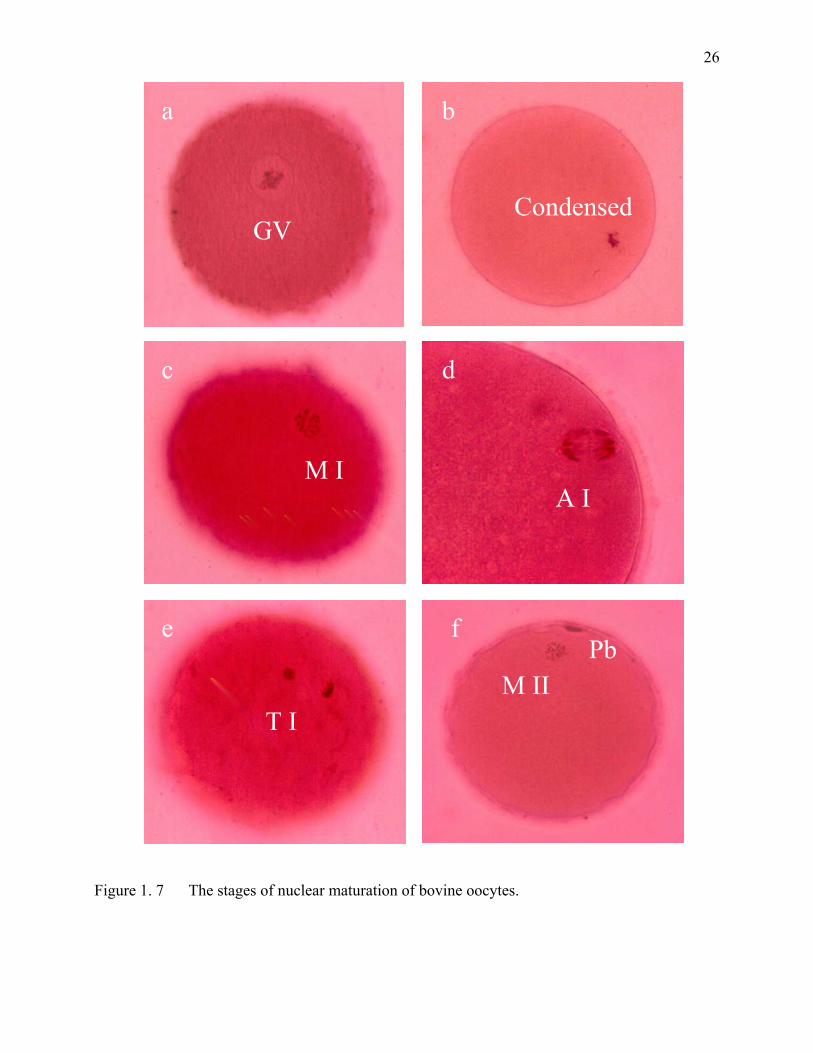
26
Figure 1. 7 The stages of nuclear maturation of bovine oocytes.
Condensed
b
GV
a
M I
c
A I
d
T I
e Pb
M II
f

27
The average time that the oocytes spend at each stage is represented in the table below. Time
zero represents the beginning of in vitro maturation. Adapted from Sirard et al, 1989.
Photo Nuclear stages Time (h) a Germinal vesicle (GV) 0-6.6 Germinal vesicle breakdown (GVBD) 6.6-8.0 b Condensed 8.0-10.3 c Metaphase I (M I) 10.3-15.4 d Anaphase I (A I) 15.4-16.6 e Telophase I (T I) 16.6-18.0 f Metaphase II (MII) and Polar body (Pb) 18.0-24.0
Cytoplasmic maturation Cytoplasmic maturation describes both the ultrastructural changes that take place in the oocyte
from the GV to the MII stage and the acquisition of developmental competence of the oocyte
(Calarco, 1995; Ducibella et al., 1994; Duranthon and Renard, 2001; Hyttel et al., 1986a; Hyttel
et al., 1986b; Shamsuddin et al., 1993). Cytoplasmic maturation is indirectly and retroactively
assessed as the ability of the mature oocyte to undergo normal fertilization, cleavage and
blastocyst development. Other indirect morphological parameters taken into account to evaluate
cytoplasmic maturation include cumulus cell expansion, expulsion of the polar body and an
increased perivitelline space (Kruip et al., 1983).
Morphological and ultrastructural changes The ultrastructural changes that take place during oocyte maturation are important to achieve
cytoplasmic maturation and ensure further development. The germinal vesicle is eccentrically
located in bovine oocytes with a diameter smaller than 110 µm, whereas the germinal vesicle of
oocytes with a diameter greater or equal to 110 µm is located close to the zona pellucida (Fair,
1995). The nucleus of bovine oocytes arrested at the GV stage contains one or two nucleoli. The
nucleoli found in the oocyte from primordial follicles are inactive, whereas the nucleoli found in
oocytes from preantral follicles with a diameter of 0.5 to 1.6 mm show active synthesis of RNA
(Crozet et al., 1986). The nucleoli of oocytes from follicles with a diameter of 3-4 mm condense

28
and transcription decreases. Nucleolus inactivation occurs during the growth of the bovine
oocyte from about 110 to 120 µm (Fair et al., 1996).
The location and morphology of mitochondria change throughout the development of the oocyte.
Mitochondria are centrally located in oocytes with a diameter smaller than 100 µm (Fair, 1995).
Mitochondria are located in the periphery of oocytes larger than 110 µm (Fair, 1995; Fair et al.,
1997). The movement of mitochondria is dependent on the microtubules (Kim et al., 1996).
Mitochondria morphology is different according to oocyte diameter. Mitochondria are round
when the oocyte has a diameter smaller than 100 µm; oval when the oocyte diameter is between
100 and 110 µm and hooded when the oocyte diameter is 110 µm (Fair, 1995; Zamboni, 1974).
The Golgi apparatus is responsible for producing the cortical granules and the zona pellucida.
The number of Golgi apparatus present in the oocyte increases as the diameter of the follicle
increases (Fair, 1995). The change in location of cortical granules constitutes the most obvious
ultrastructural sign of cytoplasmic maturation. The cortical granules that originate from the
Golgi apparatus are originally located in the center of the oocyte. However, as the oocyte
progresses to the metaphase I stage, the cortical granules translocate to the periphery of the
oocyte and become attached to the plasma membrane (Cran, 1989). The pattern and location of
cortical granules seem especially crucial for normal fertilization (Hyttel et al., 1986a; Hyttel et
al., 1986b; Hyttel and Madsen, 1987). The release of the contents of the cortical granules
changes the physical and chemical properties of the zona pellucida, increases the size of the
perivitelline space and prevents polyspermy (Hyttel et al., 1986a; Kruip et al., 1983).
The cumulus cells nourish the oocyte and provide key products for the acquisition of
developmental competence (Figure 1.6). The level of glutathione is low in the immature oocyte,
it increases during oocyte maturation to reach a maximum level at metaphase II and then drop
during the early stages of embryonic development. Glutathione is involved in the reduction of
the disulfide bonds during sperm chromatin decondensation and in the replacement of protamines
by histones (Perreault et al., 1988). The absence or small number of cumulus cells has a negative
effect on embryo production (Blondin and Sirard, 1995; Sirard et al., 1988). Denuded oocytes in
culture are capable of reaching the MII stage, but they are incapable of undergoing normal
fertilization and development (Zhang et al., 1995).

29
The removal of cumulus-oocyte gap junctions from the oolemma is temporarilly correlated with
GVBD in bovine oocytes (Hyttel, 1987; Hyttel et al., 1986a; Sutovsky et al., 1993) both in vivo
(Hyttel et al., 1986a) and in vitro (Hyttel, 1987). However, metabolic assays have shown that
GVBD in cattle oocytes takes place before any detectable decrease in the transport of small
radiolabeled molecules between the cumulus and the oocytes (De Loos et al., 1991; Laurincik et
al., 1992).
The role of M-Phase Factor in oocyte maturation Maturation promotion factor (MPF) was originally reported as a factor found in frog oocytes
(Rana pipiens) that induced oocytes to resume meiosis (Masui and Markert, 1971). Maturation
promotion factor was later renamed M-phase promoting factor because it is present in dividing
cells from almost all eucaryotes and is equally effective at promoting entry into the M phase
irrespective of the species of recipient or donor cells (Nurse, 1990). M-phase factor regulates the
G2/M transition in both meiotic and somatic cells. Thus far MPF has been shown to be active
during oocyte maturation of all studied species.
MPF is a protein kinase whose activity triggers a series of reactions ultimately leading to nuclear
membrane breakdown, chromosome condensation and spindle formation and therefore entry into
meiosis and second meiotic arrest. MPF is a protein complex composed of two subunits: p34cdc2
and cyclin B (Gautier et al., 1988). P34cdc2 is a 32 kDa protein homologous of the cell division
cycle gene of yeast cdc2 (Erikson and Maller, 1989). Cyclin B is a protein of 45 kDa whose
synthesis coincides with the activation of p34 (Levesque and Sirard, 1995; Wu et al., 1997a).
Thus, cyclin B is the regulatory component and p34cdc2 is the catalytic component of MPF. In
bovine oocytes, cyclin B is the limiting factor for meiotic resumption (Levesque and Sirard,
1996). The injection of cyclin B into cycloheximide-treated bovine oocytes results in meiotic
resumption (Levesque and Sirard, 1996).
The activation of MPF requires first the formation of the p34cdc2-cyclin B complex and second,
phosphorylation and dephosphorylation of p34cdc2 itself (Christmann et al., 1994; Jacobs, 1992;
Murray, 1993; Norbury et al., 1991; Nurse, 1990). Cyclin B is synthesized and accumulated
during interphase, while p34cdc2 is always present in the cytoplasm (Solomon et al., 1990). The

30
cyclic nature of MPF is explained by the phases of cyclin B synthesis and degradation. The
degradation of cyclin B initiates the metaphase-anaphase transition and the decline in p34cdc2
activity (Hashimoto and Kishimoto, 1988). Thus, the levels of MPF in the GV oocyte are low,
but rise at GVBD and are at a maximum as MI is reached; there is then a rapid loss in activity at
anaphase-telophase followed by a further rise as the oocyte enters MII (Fulka et al., 1992; Naito
and Toyoda, 1991; Wu et al., 1997b). This second level is maintained until completion of MII at
fertilization or parthenogenetic activation elicits the second rapid fall (Fulka et al., 1992;
Hashimoto and Kishimoto, 1988). The peak levels of MPF activity in bovine oocytes correspond
with the two metaphases: 9-12 h and after 18 h of in vitro culture. The temporary decline in MPF
activity occurs in bovine between the two metaphases (Wehrend and Meinecke, 2001).
Meiotic resumption and the maturation process are controlled by protein kinases and
phosphatases, which modulate cellular processes by phosphorylation and dephosphorylation
events. In the bovine species, meiotic resumtpion
MPF seems to be the universal regulator of the M-phase in mitotic and meiotic cell cycles
(Fissore et al., 1996; Wu et al., 1997a). M-phase factor phosphorylates a number of proteins and
it is believed to be responsible for nuclear membrane disintegration, chromatin condensation and
microtubular reorganization during meiosis (Kastrop et al., 1990b; Peter et al., 1990; Verde et al.,
1992). Initially, the oocyte acquires the ability to fully condense its chromatin. Chromatin
condensation requires active MPF, which is predetermined by a sufficient amount of cyclin B1
and p34cdc2 in an activated form (Levesque and Sirard, 1996). The oocyte then acquires the
capacity to form a MI plate and a functional spindle. The mechanism of chromosome separation
is not simply related to spindle formation, because phosphatase I inhibitors such as okadaic acid
allow their separation but not their alignment on the spindle (Levesque and Sirard, 1995). The
next step is progression to the second metaphase plate and extrusion of the polar body. The last
meiotic event is arrest at the MII step, which requires functional cytostatic factor (CSF) (Masui,
1990; Masui and Markert, 1971). The arrest at MII is maintained by a cytostatic factor (CSF).
CSF promotes meiotic arrest by either directly or indirectly stabilizing MPF activity during MII
and hence prevents oocytes from progressing beyond metaphase (Murray et al., 1989; Sagata et
al., 1989). The metaphase II arrest is maintained until meiosis resumes due to sperm-mediated or

31
parthenogenic activation. These events release the metaphase II arrest through inactivation of
MPF by degradation of the cyclin component (Murray, 1992).
Meiotic Arrest The maturation of bovine oocytes is a protracted process characterized by multiple stop-go
controls. The first meiotic arrest occurs during prenatal life when the oocyte proceeds through
the first stages of meiosis and stops at the diplotene stage of the first meiotic division. The
oocyte remains in meiotic arrest for many months or even years until the meiotic process is
resumed by the preovulatory surge of LH (Hyttel et al., 1997), atresia (Assey et al., 1994) or
removal from the follicle (Pincus and Enzmann, 1935). Bovine oocytes are competent to
complete meiosis when the oocyte diameter measures more than 110 µm (Fair et al., 1995; Hyttel
et al., 1997). It is generally accepted that the follicular cells secrete factors needed to maintain
meiotic competent oocytes in meiotic arrest. The second meiotic arrest takes place when the
oocyte reaches the metaphase stage of the second meiotic division. The oocyte completes
meiosis in response to fertilization or parthenogenic activation.
Studies on oocyte maturation have permitted the development of effective treatments to maintain
bovine oocytes in meiotic arrest in vitro. Meiotic arrest in vitro has been studied using either
chemicals or follicular fractions to block processes essential for oocyte maturation.
Transcription inhibitors Transcription inhibitors prevent bovine oocytes from resuming meiosis (Hunter and Moor, 1987;
Martus and Farin, 1994; Memili and First, 1998; Tatemoto and Terada, 1995). Bovine oocytes
actively synthesize heterogeneous nuclear RNA (hrRNA) and ribosomal RNA (rRNA) until the
oocyte has reached a diameter of 110 µm (Fair et al., 1996). The use of transcription inhibitors
has shown that RNA synthesis by the cumulus cells is required for the resumption of meiosis in
bovine oocytes (Tatemoto and Terada, 1995). Nuclear maturation occurs independently of
transcription within the oocyte (Galli and Moor, 1991).

32
Alpha-amanitin Alpha amanitin, a transcription inhibitor, is isolated from the mushroom Amanita phalloides.
Alpha amanitin (α-amanitin) inhibits mRNA synthesis by preferentially binding to the RNA
polymerase II. Ovine (Osborn and Moor, 1983), bovine (Hunter and Moor, 1987) and porcine
(Meinecke and Meinecke-Tillmann, 1993) oocytes remain in meiotic arrest when treated with α-
amanitin. The synthesis of RNA during the first hours of culture is required for meiotic
resumption in bovine COCs (Tatemoto and Terada, 1995). It has also been reported that α-
amanitin is unable to inhibit GVBD in the absence of FSH (de Wit, 2001). However, it is
unknown as to why FSH is required for the action of α-amanitin.
Bovine COCs treated with α-amanitin remain in meiotic arrest (Hunter and Moor, 1987), whereas
denuded oocytes (DO) treated with α-amanitin resume meiosis (Hunter and Moor, 1987; Kastrop
et al., 1991b; Osborn and Moor, 1983). Consequently, inhibition of RNA synthesis in the
cumulus cells of bovine COCs must occur within the first hour after the onset of culture to
prevent meiotic resumption (Tatemoto et al., 1994). The sensitivity of COCs to α-amanitin
decreases as the level of atresia increases (de Wit, 2001). The meiotic arrest induced by α-
amanitin is only partially reversible following removal of the inhibitor from the culture medium
(Hunter and Moor, 1987; Meinecke and Meinecke-Tillmann, 1993; Motlik et al., 1989). Alpha
amanitin treatment results in higher rates of oocyte degeneration (Hunter and Moor, 1987)
DRB 5,6-dichloro-1-B-D-ribofuranosylbenzymidazole (DRB) is a reversible inhibitor of the synthesis
of hnRNA and it interferes with the function of RNA polymerase II at the level of transcription
initiation (Egyhazi, 1975; Zandomeni et al., 1983). Bovine COCs treated with DRB remain in
meiotic arrest (Farin and Yang, 1994; Martus and Farin, 1994), while denuded oocytes treated
with DRB resume meiosis (Farin and Yang, 1994). In addition, DRB requires the presence of
supplemental gonadotropins (LH/FSH) to maintain the oocytes in meiotic arrest (Farin and Yang,
1994). However, DRB effectively inhibits RNA transcription regardless of whether
gonadotropins are present or not. Gonadotropins may facilitate the action of DRB by delaying
the initial transcriptional events that occurs rapidly after removal of the oocyte from the follicle,
thus rendering a higher number of COCs sensitive to DRB. Alternatively, gonadotropins may

33
facilitate the uptake of DRB into the COC (Farin and Yang, 1994). However, the exact role of
supplemental gonadotropins in the action of DRB remains unknown. DRB appears to be
involved in suppressing the stimulatory action of gonadotropin-induced maturation (Farin and
Yang, 1994).
DRB treatment blocks cumulus cell expansion; but unlike α-amanitin, DRB is not associated
with oocyte degeneration (Farin and Yang, 1994). To effectively suppress meiotic resumption,
DRB must be supplied at least every 8 h during the culture period (Martus and Farin, 1994). The
action of DRB is fully reversible when exposure to the inhibitor is limited to a maximum of 10 h.
Even though DRB is an analog of adenosine, its inhibitory action on meiotic resumption is
exerted through its role as an RNA synthesis inhibitor (Farin and Yang, 1994) because bovine
COCs treated with adenosine analogs resume meiosis (Sirard, 1990; Sirard and First, 1988).
Protein synthesis inhibitors Translation or protein synthesis inhibitors are the most effective inhibitors of the spontaneous
maturation of bovine oocytes. Mouse oocytes do not require new protein synthesis to resume
meiosis in vitro (Fulka et al., 1986). In contrast, oocytes require new protein synthesis to resume
meiosis in cattle (Hunter and Moor, 1987; Kastrop et al., 1990a; Kastrop et al., 1990b; Simon et
al., 1989; Sirard et al., 1989; Tatemoto and Horiuchi, 1995), pigs (Fulka et al., 1986; Kubelka et
al., 1988; Motlik et al., 1991), goat (Le Gal et al., 1992) and sheep (Moor and Crosby, 1986).
Oocytes from these species can be reversibly maintained at the GV stage by culturing them in the
presence of protein synthesis inhibitors (Hunter and Moor, 1987; Sirard and First, 1988). In vitro
maturation does not affect the ability of bovine oocyte to synthesize proteins (Sirard and Coenen,
1994). In fact, protein synthesis in bovine oocytes is maximal during the initial 12 h of IVM
(Shamsuddin et al., 1993; Wu et al., 1996). Bovine oocytes require the synthesis of new protein
during the first 8 h of culture to resume meiosis (Kastrop et al., 1991b; Tatemoto et al., 1994).
Protein synthesis inhibitors prevent meiotic resumption by inhibiting the synthesis of cyclin B
(Levesque and Sirard, 1996; Wu et al., 1997a). Meiotically incompetent bovine oocytes lack
cyclin B1, which is required for the autoamplification of MPF and meiotic resumption (Levesque
and Sirard, 1996). The activity of MPF depends on active protein synthesis (Hunter and Moor,

34
1987) and its state of phosphorylation. Thus, bovine oocytes treated with inhibitors of protein
synthesis or phosphorylation inhibitors remain in meiotic arrest.
Bovine oocytes are sensitive to protein synthesis inhibitors only during the first half-period of the
time required for GVBD (Fulka et al., 1986; Motlik et al., 1991). Newly synthesized proteins are
also necessary for the transition from metaphase I to metaphase II, as well as for the extrusion of
the first polar body. Otherwise, chromatin decondensation occurs and pronuclei-like structures
are observed (Sirard et al., 1989).
Cycloheximide Cycloheximide is a protein synthesis inhibitor of eukaryotic cells. This tetracycline-analog stops
peptide elongation by interacting with peptidyl transferase; it thus interfers with protein synthesis
by retaining the peptide on the 80S ribosome (Barbacid et al., 1975; Vasquez, 1974). The
addition of cycloheximide to the incubation medium within 6 h from the onset of in vitro
maturation prevents bovine oocytes from resuming meiosis (Tatemoto and Terada, 1995).
Cycloheximide prevents meiotic resumption in either COC or DO by blocking the synthesis of
proteins associated with the nuclear envelope and other stage-specific events (Hunter and Moor,
1987; Levesque and Sirard, 1995; Levesque and Sirard, 1996; Simon et al., 1989; Sirard et al.,
1989; Tatemoto et al., 1994). The inhibitory effect of cycloheximide on meiotic resumption is
reversible. Removal of the oocytes from the inhibitory conditions allows approximately 80% of
the oocytes to reach metaphase II and 20% of those develop to the blastocyst stage after
fertilization (Fulka et al., 1986; Hunter and Moor, 1987; Lonergan et al., 1997; Simon et al.,
1989; Sirard and First, 1988; Tatemoto et al., 1994); whereas control oocytes incubated in TCM
supplemented with 10% FCS was fully reversible and 45% of the oocytes developed to the
blastocyst stage after fertilization. Blastocysts obtained from cycloheximide-treated oocytes have
the same number of cells as blastocysts obtained from non-treated oocytes (Lonergan et al.,
1997). However, oocytes treated with cycloheximide for more than 12 h have decreased rates of
development to the blastocyst stage (Lonergan et al., 1997). The maturation and fertilization of
oocytes previously treated with cycloheximide produced live calves thus corroborating the
reversibility of this protein synthesis inhibitor (Motlik et al., 1990; Saeki et al., 1997; Saeki et al.,
1998). The nuclear maturation of cycloheximide-treated oocytes is accelerated and completed 4

35
h faster than control oocytes, but the same interval of maturation is necessary to complete meiosis
I for subsequent development to blastocyst (Kubelka et al., 1988; Saeki et al., 1998; Tatemoto et
al., 1994). Furthermore, cycloheximide prevents GVBD but treated oocytes show some degree
of chromosome condensation. This suggests that some of the steps involved in GVBD are
independent of protein synthesis (Mayes and Sirard, 2001; Simon et al., 1989).
Puromycin Puromycin is a structural analog of tRNA that attaches itself to the growing end of the
polypeptide chains during protein synthesis. As a result, the growing chain is prematurely
released from the ribosome and protein synthesis is not completed (Vasquez, 1974). Bovine
oocytes treated with concentrations of 75 to 100 µg/mL of puromycin remain in meiotic arrest,
while oocytes treated with lower doses of puromycin resumed meiosis (Motlik et al., 1990). The
inhibitory effect of puromycin is reversible. Puromycin accelerates the nuclear maturation of
bovine oocytes since treated oocytes resumed meiosis 2-3 h faster than control oocytes (Motlik et
al., 1990).
Phosphatase inhibitors Phosphatase inhibitors play a role in oocyte maturation by controlling the phosphorylation status
of the p34cdc2 component of MPF (Levesque and Sirard, 1996).
Okadaic acid Okadaic acid (OA) is a potent inhibitor of phosphatase type 2A and, to a lesser extent,
phosphatase type 1 (Alexandre et al., 1991). Okadaic acid treatment leads to an increase in
overall protein phosphorylation and allows the dissolution of the nuclear membrane. Okadaic
acid accelerates GVBD in bovine oocytes by 2 h when compared to untreated oocytes (Kalous et
al., 1993a). Bovine oocytes treated with OA do not progress beyond the late diakinesis–
metaphase I stage because it affects the alignment of the chromosomes on the metaphase plates
(Kalous et al., 1993a; Levesque and Sirard, 1995; Levesque and Sirard, 1996). Continuous
exposure of bovine oocytes to okadaic acid results in an abortive M-phase without spindle
formation (Kalous et al., 1993; Levesque and Sirard, 1996). Okadaic acid fully reverses the

36
inhibitory effects of 6-DMAP and cycloheximide on the maturation of bovine oocytes (Kalous et
al., 1993a; Tatemoto and Terada, 1998). This suggest that okadaic acid may positively regulates
MPF activation, probably by influencing the phosphorylation / dephosphorylation stage of p34cdc2
(Kalous et al., 1993a).
Vanadate Vanadate is an inhibitor of protein tyrosine phosphatase and maintains bovine oocytes in meiotic
arrest (Lonergan et al., 1997). Vanadate prevents the dephosphorylation of p34cdc2; thus
preventing MPF activation in vitro (Choi et al., 1991; Solomon et al., 1990). Treatment of
bovine oocytes with vanadate is reported to be reversible only in 56% of oocytes, with only 6%
reaching metaphase II (Lonergan et al., 1997), whereas the treatment of bovine oocytes with 6-
DMAP is fully reversible after a period of incubation without the inhibitor (100% GVBD and
83% of the treated oocytes reached the MII stage) (Lonergan et al., 1997). The irreversible effect
of vanadate on oocyte maturation may be the result of microtubule damage (Aquino et al., 1995).
However, vanadate reversibly inhibits the meiotic resumption of rat oocytes (Goren and Dekel,
1994). Further suggesting differences in the control of meiois across different species.
Protein kinase inhibitors The pattern of protein phosphorylation in bovine oocytes undergoes major changes just before
the resumption of meiosis (Kastrop et al., 1990b). Extensive phosphorylation takes place during
the first 3 h of in vitro maturation of bovine oocytes (Kastrop et al., 1990b). The change in
protein phosphorylation is correlated with the activation of two major M-phase kinases: p34cdc2
protein kinase and mitogen-activated protein kinase (MAPK). Both kinases are activated around
the time of GVBD in bovine oocytes (Motlik et al., 1998). Phosphorylation inhibitors prevent
meiotic resumption by acting on regulatory proteins that have kinase activity (Lonergan et al.,
1997). These inhibitors are used to control the phosphorylation status of specific residues of
p34cdc2; therefore; protein kinase inhibitors block MPF activity to maintain bovine oocytes in
meiotic arrest. Butyrolactone, 6-DMAP and roscovitine inhibit cyclin-dependent kinase 1 (cdk1)
activity by competing with ATP for binding to the catalytic site. Studies were also carried out
with the protein tyrosine kinase inhibitors genistein and tyrphostin. Genistein does not affect the

37
meiotic resumption of bovine COCs (Levesque and Sirard, 1995). Whereas, bovine COCs
treated with tyrphostin remain in meiotic arrest (Sirard et al., 1998).
6-DMAP The puromycin analog 6-dimethylaminopurine (6-DMAP) is a potent, broad-spectrum
serine/threonine kinase inhibitor that prevents the burst in phosphorylation that occurs shortly
before GVBD in bovine oocytes (Avery et al., 1998; Dode and Adona, 2001; Faerge et al., 2001;
Fulka et al., 1991; Levesque and Sirard, 1995; Lonergan et al., 1997). 6-DMAP is active
regardless of the presence or absence of cumulus cells (Fulka et al., 1991). However, bovine
oocytes remain in meiotic arrest only if they are treated with 6-DMAP within the first 4 h of
culture (Fulka et al., 1991). The meiotic arrest of bovine oocytes treated with 6-DMAP is
reversible, but between 60 and 80% of treated oocytes have abnormal metaphase plates compared
to untreated oocytes (Avery et al., 1998; Fulka et al., 1991). Abnormal metaphase plates might
result from impaired spindle microtubules (Simili et al., 1997). Cortical granules in 6-DMAP
treated oocytes are normally distributed, but they are larger and less electron dense than controls
(Avery et al., 1998). Protein synthesis proceeds in the presence of 6-DMAP (Fulka et al., 1991;
Kastrop et al., 1990b; Kastrop et al., 1991a), but it changes the phosphorylation and migration
patterns of certain proteins (Levesque and Sirard, 1995). Two proteins with molecular weights of
46 kDa and 29 kDa were not synthesized in oocytes treated with 6-DMAP (1 mM) and that
proteins in the 50 kDa range presented an isoelectric point more basic than in than the meiosis–
conductive treatemnt. (Levesque and Sirard, 1996). Development to the blastocyst stage is
impaired in bovine oocytes that have previouly been treated with 6-DMAP (Dode and Adona,
2001; Kubish et al., 1995; Lonergan et al., 1997). Some embryos escape the negative side effects
of 6-DMAP-treatment and the resulting blastocysts are not different from control blastocysts in
terms of morphology and kinetics (Avery et al., 1998).
Butyrolactone Butyrolactone I (BL-I), a specific inhibitor of cdk, inhibits the resumption of meiosis in bovine
oocytes (Faerge et al., 2001; Kitawa et al., 1993; Kubelka et al., 2000; Lonergan et al., 2000;
Motlik et al., 1998). Butyrolactone prevents the phosphorylation of p34cdc2 kinase, but it has little

38
effect on MAPK (Kubelka et al., 2000). The effect of BL-I is dose-dependent and reversible
under conditions permissive to normal oocyte maturation. Once the inhibitor is removed,
butyrolactone-treated oocytes have similar developmental potential to the blastocyst stage as
untreated oocytes following maturation, fertilization, and culture in vitro. (Kubelka et al., 2000;
Lonergan et al., 2000). These results demonstrate the feasibility of maintaining bovine oocytes in
artificial meiotic arrest without compromising their subsequent developmental competence
(Lonergan et al., 2000). However, it is important to point out that fewer butyrolactone-treated
oocytes than untreated oocytes reach the blastocyst stage following vitrification (Lonergan et al.,
2000).
Roscovitine Roscovitine is a potent inhibitor of MPF kinase activity (Meijer et al., 1997). Roscovitine
maintains bovine oocytes in meiotic arrest for at least 24 h (Mermillod et al., 2000). The
cleavage rate and the development to the blastocyst stage are not different between treated and
untreated oocytes indicating that the inhibitory effect of roscovitine is fully reversible (Mermillod
et al., 2000). Simultaneous treatment of bovine oocytes with butyrolactone and roscovitine
permit the use of smaller doses of each inhibitor that maintain a reversible state of meiotic arrest
without comprising subsequent development to the blastocyst stage (Ponderato et al., 2001).
Protein kinase A Cyclic AMP exerts its effects in animal cells mainly by activating an enzyme called cyclic AMP-
dependent protein kinase (PKA), which catalyzes the transfer of the terminal phosphate group
from ATP to specific serine or threonine residues of selected proteins. In its inactive state, PKA
is a complex of two regulatory subunits and two catalytic subunits. The binding of 4 cAMP
molecules to the regulatory subunits alters their conformation causing them to dissociate from the
PKA complex. The released catalytic subunits are thereby activated to phosphorylate the serine
or threonine residues of substrate proteins. The biochemical properties of the regulatory subunits
distinguish the different isoenzymes of the PKA type I and type II (Downs and Hunzicker-Dunn,
1995). Bovine oocytes are not as sensitive as oocytes from other species to protein kinase
stimulation via cAMP accumulation (Sirard and First, 1988).

39
Cyclic AMP is synthesized from ATP by the plasma-membrane-bound enzyme adenylate cyclase
(AC), and it is rapidly and continuously degraded by one or more cAMP-phosphodiesterase
(PDE), which hydrolyze cAMP to adenosine 5’-monophosphate (5’-AMP). It has also been
reported the existence of a soluble form of adenylate cyclase (sAC) in cytosolic extracts from
mammalian testis (Braun and Dods, 1975; Buck et al., 1999). The existence of a sAC in the
oocyte would imply that cAMP could be generated at a distance from the membrane and closer to
the site of action. The existence of soluble and membrane bound forms of AC in the oocyte
would increase the complexity of the regulation of the signals mediated by cAMP.
In laboratory animals, modification of the amount of cAMP in the oocyte, using additives such as
dibutyryl cAMP or PDE inhibitors, is used to control GVBD (Dekel, 1988; Downs et al., 1989).
In cattle, a rise in cAMP will delay GVBD in bovine COC or DO (Sirard et al., 1989). Treatment
of bovine DO with cAMP analogs results in a transitory inhibition of meiotic resumption.
Bovine oocytes treated with dibutyryl cyclic AMP (dbcAMP) or 8-bromo-3',5'-cAMP (8-Br-
cAMP) transiently remain in meiotic arrest (Homa, 1988; Sirard and First, 1988). Dibutyryl
cAMP is less effective than 8-Br-cAMP in maintaining meiotic arrest (Homa, 1988). The
differences in effectiveness to maintain the meiotic arrest of bovine oocytes may be explained by
the fact that dbcAMP is degradable by phosphodiesterases, 8-Br-cAMP is only partially
degradable (Beebe et al., 1988). However, the inhibitory effect of these cAMP-analogs is only
temporary. There is no significant effect of these inhibitors on the meiotic arrest of treated or
untreated oocytes after 24 h of incubation (Homa, 1988). Treatment with 8-bromo-cAMP
induced a significant increase in the proportion of oocytes remaining at the MI compared with
control cells in which the majority had reached anaphase I or beyond (Homa, 1988).
Bovine COCs contain adenylate cyclase in both the cumulus cells and the oocyte (Kuyt et al.,
1988). Adenylate cyclase was localized specifically between the junctional complexes of
cumulus cells and on the cumulus cell processes contacting the oocyte. Adenylate cyclase is also
present in the plasma membrane of the bovine oocyte (Kuyt et al., 1988). Thus, bovine oocytes
are able to synthesize cAMP to affect their in vitro maturation (Bilodeau et al., 1993).
Stimulation of adenylate cyclase results in increased levels of cAMP in the oocyte. Forskolin, a
direct stimulator of the catalytic subunit of adenylate cyclase, increases the synthesis of cAMP in

40
both bovine COC and DO (Bilodeau et al., 1993; Seamon et al., 1981). Forskolin treatment
significantly increases the number of bovine oocytes in meiotic arrest (Homa, 1988; Kuyt et al.,
1988; Richard et al., 1997; Rose-Hellekant and Bavister, 1996). The inhibitory effect of
forskolin on oocyte maturation is increased when used in combination with 3-isobutyl-1-
methylxanthine (IBMX) (Bilodeau et al., 1993; Homa, 1988). Large doses of sodium fluoride
(NaF), a stimulator of adenylate cyclase (Sirard, 1990), block COC or DO at the GV stage.
Sodium fluoride can alter the function of the adenylate cyclase by irreversibly coupling the alpha
subunit of the Gs to the catalytic component (Ross and Gilman, 1980).
Cholera toxin and pertussis toxin modify G protein function by using NAD to ADP-ribosylate Gs
and Gi respectively; these modifications lead to the irreversible stimulation of AC by Gs or the
inability of Gi to turn off the enzyme (Birmbaumer et al., 1987; Gilman, 1987). Bordetella
pertussis toxin and invasive AC therefore reversibly maintains bovine oocytes and COCs in
meiotic arrest (Aktas et al., 1995a; Aktas et al., 1995b).
Phosphodiesterase inhibitors Phosphodiesterases (PDEs) degrade cyclic nucleotides by hydrolizing the 3’ phosphodiester bond
of the 3’ 5’ purine ribose cyclic monophosphate cAMP and cGMP. The resulting biologically
inactive monophosphates do not activate their respective protein kinases. At least 11 PDE
families have been identified based on their kinetics and substrate characteristics, inhibitor
profiles, allosteric activators and inhibitors and amino acid sequences (Beavo, 1995; Conti, 2000;
Conti and Jin, 1999; Conti et al., 1991; Fisher et al., 1998). The use of non-selective PDE
inhibitors such as IBMX has shown that PDEs are involved in the maturation of bovine oocytes
(Sirard and First, 1988). It is important to clarify that PDE8 and PDE9 are not inhibited by
IBMX (Soderling and Beavo, 2000). Approximately 45% of bovine COCs treated with 0.2 mM
IBMX are temporarily prevented from resuming meiosis (Sirard and First, 1988). The effect of
IBMX on bovine oocytes is similar to the effect of cAMP analogues and results in a delayed
GVBD followed by normal maturation or a metaphase I block depending on the product and the
time of addition after oocyte aspiration (Ball et al., 1984; Sirard, 1989; Sirard and First, 1988).
The role of PDE isoenzymes in oocyte maturation is now studied using isoenzyme-specific PDE
inhibitors. These inhibitors act mainly as active site competitors (Atienza et al., 1999).

41
Isoenzyme specific PDE inhibitors have permitted the differentiation between different PDE
families. The PDE3 family or cGMP-inhibited PDE (cGI-PDE) shows high affinity to cAMP as
a substrate but it is inhibited by micromolar concentrations of cGMP (Loten et al., 1978; Meacci
et al., 1992; Taira et al., 1993). Specific PDE3A inhibitors such as cilostamide and milrinone
prevent the maturation of rat oocytes both in vivo and in vitro (Conti, 2000; Harrison et al., 1986;
Tsafriri et al., 1996). Thus the presence of an active PDE3A has been demonstrated in rat
oocytes (Richard et al., 2001). The PDE4 family or cAMP-specific PDE hydrolyzes cAMP with
high affinity. However, the PDE4 family, unlike the PDE3 family, is not inhibited by cGMP
(Colicelli et al., 1989; Swinnen et al., 1989). Rolipram, a specific PDE4 inhibitor, has permitted
the localization of PDE4 to the granulosa cells of rat COCs (Tsafriri et al., 1996).
Protein kinase C The protein kinase C (PKC) family phosphorylates proteins rich in serine-threonine residues.
This family of multiple isoforms mediates the signal transduction of extracellular signals into
intracellular signals (Jaken, 1996; Jones, 1998). The physiological activators of PKC are Ca2+ and
diacylglycerol (DAG) (Bell, 1986). These activators are generated by phospholipase C
hydrolysis of phosphoinositol bi-phosphate (PIP2) molecules to generate DAG and inositol
triphosphate (InsP3). Inositol triphosphate induces the release of Ca2+ from the intracellular
compartments. There is a homology between DAG and the tumor-promoting phorbol esters,
which irreversibly activate PKC by binding to its regulatory subunits (Castagna et al., 1982;
Jones, 1998; Niedel et al., 1983). The stimulation of PKC in denuded bovine oocytes did not
have any effect on oocyte maturation (Homa, 1991; Rose-Hellekant and Bavister, 1996).
Neomycin inhibits the action of phospholipase C by preventing phosphoinositide hydrolysis and
the subsequent generation of second messengers. Neomycin is a polycationic aminoglycoside
antibiotic that binds with high affinity to phosphatidylinositol 4,5-biphosphate (Gabev et al.,
1989). Neomycin prevents denuded bovine oocytes from resuming meiosis probably by
decreasing the mobilization of intracellular calcium (Homa, 1991). Treatment of oocytes with
neomycin must occur within 3 h following removal from the follicle to prevent meiotic
resumption (Homa, 1991).

42
Calcium, however, plays a role in the maturation of mammalian oocytes (Collas et al., 1993a;
Collas et al., 1993b; Homa, 1995; Homa et al., 1993). Luteinizing hormone induces calcium
transients in the oocyte (Homa et al., 1993). The initial calcium transient results from the
mobilization of intracellular calcium and then subsequent transients result from an influx of
extracellular calcium (Davis et al., 1987). Calcium transients also occur at fertilization and
appear to be required for the release of the second metaphase arrest in the bovine oocyte (Collas
et al., 1993a; Collas et al., 1993b). The absence of external calcium by incubating the oocytes in
Ca2+ -free or Ca2+ -depleted medium has no effect on GVBD (Homa, 1995; Leibfried and First,
1979a; Maruska et al., 1984). In addition, treatment of bovine oocytes with 0.1 to 5 mM EDTA,
an extracellular calcium chelator, has no effect on meiotic arrest (Sirard et al., 1998). Chelation
of the intracellular calcium prevents meiotic resumption (Homa, 1995). Oocytes loaded with the
intracellular calcium chelator BAPTA/AM exhibit a dose-dependent inhibition of GVBD, even in
the presence of extracellular calcium (Homa, 1991). However, the culture of oocytes in Ca2+-
deficient medium suppresses polar body formation (Jagiello et al., 1982). Furthermore, bovine
COCs incubated in Ca2+ and Mg2+ free medium do not complete meiosis I (Jagiello et al., 1982).
Purines The purines, adenosine and hypoxanthine, maintain mouse oocytes in meiotic arrest (Downs,
1993b; Downs, 1999; Downs and Eppig, 1987; Shim et al., 1992), whereas in bovine oocytes,
their inhibitory effect is only transient (Sirard and First, 1988). Porcine follicular fluid and
bovine follicular fluid contain both of these purines in sufficient amounts to maintain mouse
oocytes in meiotic arrest (Downs et al., 1986; Downs et al., 1985; Kadam and Koide, 1990). On
the other hand, bovine oocytes treated with adenosine resume meiosis (Sirard, 1990; Sirard and
First, 1988). It is doubtful that purines have a similar role in these two species. The inhibitory
action of purines is possibly explained by their capacity to decrease phosphodiesterase activity
(Downs et al., 1986; Downs et al., 1989).
Physiological methods of maintaining meiotic arrest in vitro Physiological methods of maintaining bovine oocytes in meiotic arrest include incubation of
COCs with follicular fluid (Sirard and First, 1988), co-culture with granulosa or theca cell

43
monolayers (Richard and Sirard, 1996a; Richard and Sirard, 1996b; Richard and Sirard, 1998),
follicular hemisections (Richard and Sirard, 1996a; Sirard and Coenen, 1993) or by culturing
intact antral follicles (Nashta et al., 1998).
Follicular fluid In addition to providing nourishment to the oocyte, follicular fluid is also involved in the
regulation of oocyte maturation. Follicular fluid has been reported to have no effect, stimulate or
inhibit oocyte maturation (Downs et al., 1985; Leibfried and First, 1980a; Lonergan et al., 1994a;
Naito et al., 1988; Romero-Arredondo and Seidel, 1994; Sirard et al., 1995; Tsafriri et al., 1977).
The effect of follicular fluid on oocyte maturation depends on the size of the follicle (Lonergan et
al., 1994b; Sirard et al., 1995; Sun et al., 1994; Vatzias and Hagen, 1994) the stage of the estrous
cycle (Rabahi et al., 1993; Romero-Arredondo and Seidel, 1994; Sirard et al., 1995) and the
fraction of follicular fluid used (Daen et al., 1994). The origin of follicular fluid is important but
the effect of its addition is not uniform over oocytes originated from follicles of different sizes
(Carolan et al., 1996). The fate of a given pool of oocytes is determined by the status of
individual oocytes rather than the status of the follicular fluid supplement (Carolan et al., 1996).
The inhibitory effect of follicular fluid on oocyte maturation has been reported in mice, sheep,
cattle, pigs, hamsters and humans (Chang, 1955; Downs and Eppig, 1984; Hillensjo et al., 1978;
Racowsky and Baldwin, 1989; Sirard and First, 1988; Sun et al., 1994; Tsafriri et al., 1976).
Bovine follicular fluid
Bovine follicular fluid (BFF) completely and reversibly inhibits the spontaneous meiotic
resumption of mouse (Kadam and Koide, 1990; Sato and Koide, 1984) and hamster oocytes
(Gwatkin and Andersen, 1976). On the other hand, the maturation of bovine oocytes is only
transiently delayed by bovine follicular fluid (Leibfried and First, 1980a; Sirard and Bilodeau,
1990b; Sirard and First, 1988). The presence of bovine follicular fluid in the incubation medium
greatly reduces the number of granulosa cells required to maintain bovine COC in meiotic arrest
(Sirard and Bilodeau, 1990b). Bovine follicular fluid collected from both small and medium
follicles at estrus had the greatest ability to prevent meiotic resumption, while the follicular fluid
collected from large follicles at estrus had the smallest ability to maintain meiotic arrest (Ayoub

44
and Hunter, 1993). The quantity of bovine follicular fluid present in the culture medium has an
inverse linear relationship with the nuclear maturation of bovine oocytes (Sirard et al., 1992;
Sirard and First, 1988). Bovine oocytes isolated from the follicular environment rapidly become
committed to undergo meiotic resumption (Sirard and First, 1988; Sirard et al., 1989). Indeed,
oocytes become committed to undergo GVBD even when short washing procedures are carried
out in the absence of follicular fluid (Sirard and First, 1988).
Potential inhibitory factors have been isolated from bovine follicular fluid. A peptide factor with
an estimated Mr of 8000 isolated from bovine follicular fluid blocks the progesterone-induced
maturation of Xenopus oocytes, while the same factor does not block the meiotic resumption of
mouse oocytes (Kadam and Koide, 1991). The presence of a soluble protein kinase has also been
reported in bovine and human follicular fluid (Yang et al., 1993). The soluble PKA is probably a
product of the granulosa cells of the follicles and suggests that a highly phosphorylated state
would sustain the meiotic arrest. In addition, the presence of a protein with a Mr of
approximately 6000 found in bovine follicular fluid is suggested to play a role in the control of
meiosis (Dostal and Pavlok, 1996). This protein does not prevent the meiotic resumption of
denuded oocytes. Supplementation of the incubation medium with 2.0 mg/ml of the isolated
protein induces meiotic arrest in all treated oocytes (Dostal and Pavlok, 1996).
Linoleic acid may play a role in maintaining the meiotic arrest of bovine oocytes in vivo.
Linoleic acid (18:2) is the most abundant unsaturated fatty acid found in bovine follicular fluid.
The concentration of linoleic acid decreases significantly during follicular development (Homa
and Brown, 1992). Incubation of bovine COC with linoleic acid significantly inhibited GVBD in
a dose-dependant manner, while all the other fatty acids tested did not have an effect on oocyte
maturation (Homa and Brown, 1992). As mentioned before, purines have a weak effect on the
inhibition of meiotic resumption of bovine oocytes (Sirard and First, 1988). Hypoxanthine is
present in bovine follicular fluid (Kadam and Koide, 1990), but the quantity of purines found in
bovine follicular fluid are not comparable to those found in mouse follicular fluid (Downs et al.,
1989; Eppig et al., 1985).
Porcine follicular fluid

45
The inhibitory activity present in porcine follicular fluid has been attributed to the presence of
Oocyte Maturation Inhibitor (OMI) (Sato and Ishibashi, 1977; Tsafriri and Channing, 1975b;
Tsafriri et al., 1982; Tsafriri et al., 1976). Oocyte maturation inhibitor was isolated from a low
molecular-weight fraction of porcine follicular fluid that exerts an inhibitory action on nuclear
maturation of porcine (Jagiello et al., 1977; Tsafriri and Channing, 1975b), mouse (Tsafriri,
1979) and rat oocytes (Tsafriri et al., 1977). OMI is produced by porcine granulosa cells and
released into the follicular fluid (Stone et al., 1978; Tsafriri et al., 1976). The approximate
molecular weight of OMI is 2000 daltons (Hillensjo et al., 1979; Leibfried and First, 1980a).
OMI is active in COC but not in DO, thus cumulus cells are required to maintain the oocyte in
meiotic arrest. The inhibitory effect of OMI is reversed by LH or by removal from the culture
medium (Stone et al., 1978). OMI remains active even after freezing/thawing, charcoal
extraction and heating to 60 ºC (Tsafriri et al., 1976). However, OMI is inactivated by trypsin
treatment. It is hypothesized that OMI might be follistatin or at least part of this follicular
protein. However, other laboratories have reported that porcine (Leibfried and First, 1980a;
McGaughey, 1978; Racowsky and McGaughey, 1982) or cow oocytes (Sreenan, 1970) resume
meiosis in the presence of porcine follicular fluid.
Hypoxanthine is another inhibitory product found in the low molecular weight fraction of porcine
follicular fluid (Downs et al., 1985; Downs and Eppig, 1984). The levels of hypoxanthine found
in porcine follicular fluid are sufficient to prevent meiotic resumption in mouse oocytes (Downs
et al., 1985).
Mouse follicular fluid
The inhibitory activity of mouse follicular fluid has been attributed to the presence of
hypoxanthine and adenosine (Downs et al., 1985; Downs et al., 1989). Both of these purines are
found in high quantities in mouse follicular fluid (Downs et al., 1989; Eppig et al., 1985). The
simultaneous addition of hypoxanthine and adenosine to the incubation medium inhibits the in
vitro maturation of mouse oocytes (Downs et al., 1986; Eppig et al., 1985). The action and
uptake of both of these compounds does not require the presence of cumulus cells (Eppig and
Downs, 1987; Shim et al., 1992). However, mouse COCs are more sensitive to hypoxanthine

46
than denuded oocytes (Eppig et al., 1985). Hypoxanthine acts as a non-specific inhibitor of PDE
activity in the oocyte (Downs et al., 1989; Racowsky, 1991).
Rat follicular fluid
The ability of rat follicular fluid to prevent rat oocytes from resuming meiosis is due to the
presence of the Müllerian Inhibiting Substance (MIS) (Takahashi et al., 1986). The rat granulosa
cells produce MIS. Müllerian inhibiting substance prevents the meiotic resumption of rat COC
or DO. The inhibitory effect of MIS is dose-dependent, reversible and independent of cAMP.
The action of MIS is not affected by the presence of FSH, LH, progesterone, estradiol or
testosterone. Müllerian inhibiting substance does not appear to act through the adenylate cyclase
system and it was hypothesized that it might act by inhibiting a tyrosine kinase (Takahashi et al.,
1986).
Whole follicles Isolated oocytes spontaneously resume meiosis in vitro, whereas follicle-enclosed oocytes remain
in meiotic arrest unless they have been exposed to gonadotropins in vivo or in vitro (Dekel, 1995;
Pincus and Enzmann, 1935; Tsafriri et al., 1996; Whitaker, 1996).
Follicle hemisections Complete follicles or follicle hemi-sections maintain oocytes in meiotic arrest in vitro in the
sheep (Moor and Trounson, 1977), pig (Saito and Ishibashi, 1977) and cow (Sirard et al., 1992).
Bovine COCs incubated with follicle hemisections remain in meiotic arrest even without direct
contact between the oocyte and the follicular wall (Carbonneau and Sirard, 1994; De Loos et al.,
1994; Leibfried and First, 1980a; Leibfried and First, 1980b; Richard and Sirard, 1996a; Sirard
and Coenen, 1993). Follicle stimulating hormone and LH do not reverse the inhibitory action of
the follicular wall on the meiotic resumption of bovine COCs incubated within their own
follicular wall hemisection (Carbonneau and Sirard, 1994). The follicular wall does not maintain
bovine denuded oocytes in meiotic arrest (Carbonneau and Sirard, 1994). The cumulus cells are
important in transmitting or processing the inhibitory signal from the follicular wall to the oocyte
as with most inhibitors. It appears that the number of COCs in meiotic arrest is proportional to

47
the quantity of tissue present in the culture medium (Sirard and Coenen, 1993). However, the
inhibitory effect of the follicle wall is irreversible when a large number of hemisections are added
to the culture medium (Sirard and Coenen, 1993).
Granulosa cells Bovine granulosa cells have a moderate ability to prevent bovine oocytes from resuming meiosis
in vitro (Kotsuji et al., 1994; Sirard and Bilodeau, 1990a; Sirard and Bilodeau, 1990b; Sirard et
al., 1992). High concentrations of bovine granulosa cells (10 to 100 million cells /ml) maintain
bovine oocytes in meiotic arrest, whereas low concentrations of granulosa cells (1 to 5 million
cells /ml) stimulate the oocyte to resume meiosis in vitro (Sirard and Bilodeau, 1990b), especially
if the culture medium includes gonadotropins (Sirard and Bilodeau, 1990a) or estrus serum (Lu et
al., 1987). The inhibitory effect of granulosa cells is amplified by the presence of BFF and direct
contact between the granulosa cells and the cumulus cells (Sirard et al., 1992). Direct contact
between the granulosa cells and COC appears necessary to maintain bovine oocytes in meiotic
arrest. Embedding of COCs in agar prior to culture with granulosa cells removes the inhibitory
effect of granulosa cells (Sirard, 1991; Sirard and Bilodeau, 1990b). Bovine granulosa cells in
suspension are more effective in preventing meiotic resumption of bovine COCs than granulosa
cell monolayers (Sirard and Bilodeau, 1990b). Monolayers of bovine granulosa cells with their
conditioned medium provide inhibitory conditions to 33% of those oocytes adhering to the
monolayer (Sirard and Bilodeau, 1990a). However, neither suspension of fresh bovine granulosa
cells (Leibfried and First, 1980a; Sirard and Bilodeau, 1990b) nor monolayers of bovine
granulosa cells fully prevents GVBD in bovine COCs (Richard and Sirard, 1996b; Sirard and
Bilodeau, 1990a).
At least 50% of bovine COC connected to the membrana granulosa (COCG) remain in meiotic
arrest for at least 22 h (van Tol et al., 1996). The effect of the membrana granulosa is reversed by
the addition of FSH to the culture medium (van Tol et al., 1996). Incubation of COCGs in the
presence of theca cells enhances the proportion of oocytes in meiotic arrest even in the presence
of added FSH (van Tol and Bevers, 1998). Thus, theca cells secrete a factor that prevents the
GVBD induced by FSH on COCGs. Furthermore, follicle hemisections, which contain both
granulosa and theca cells, inhibit the meiotic resumption of COCs even in the presence of

48
gonadotropin hormones (Carbonneau and Sirard, 1994). In conclusion, bovine granulosa cells
can participate in maintaining meiotic arrest, but the main inhibitory signal comes from theca
cells (Richard and Sirard, 1996a).
Heterologous cultures have shown that the inhibitory factors produced by porcine membrana
granulosa cells reversibly maintain the meiotic arrest of bovine COCs (Kalous et al., 1993b; Petr
et al., 1989). In these experiments, the cumulus cells were closely associated with granulosa
cells, but no gap junctions were formed among heterologous granulosa cells and the expansion of
the bovine cumulus cells did not occur (Kalous et al., 1993b).
Theca cells Theca cells maintain bovine oocytes in meiotic arrest during in vitro maturation (Richard, 1997;
Richard and Sirard, 1996b). Theca cells are obtained by enzymatically digesting layers of theca
externa and theca interna of follicles ranging from 2 to 5 mm in diameter. Theca interna or theca
externa are equally effective in maintaining bovine COCs in meiotic arrest (Richard and Sirard,
1996a). On the other hand, bovine COCs cocultured with granulosa cell monolayers resume
meiosis (Richard and Sirard, 1996b). Theca cell monolayers capable of maintaining meiotic
arrest secrete two proteins of approximate Mr of 214 000 and 190 000 daltons (Richard and
Sirard, 1998). The protein pattern of theca cell monolayers is different from the pattern of
proteins secreted by granulosa cell monolayers. In addition, the secretion of the 214 000 dalton
protein decreases when the theca cell monolayers are grown on collagen-coated wells.
Consequently, the 214 000 dalton protein may be involved in the process of maintaining bovine
oocytes in meiotic arrest (Richard and Sirard, 1998).
Theca cell monolayers maintain COCs but not DOs in meiotic arrest (Richard and Sirard, 1996b;
Richard and Sirard, 1998). Consequently, the inhibitory factor acts on the oocyte through or in
association with the cumulus cells (Richard and Sirard, 1996b). However, DOs cocultured with
COCs significantly increased the percentage of COCs maintained in meiotic arrest. This suggests
that denuded oocytes produce a factor that modulates either the theca cells or the cumulus cells to
increase the efficiency of the inhibitory pathway (Richard and Sirard, 1998).

49
Replacing fetal calf serum with bovine serum albumin (BSA) or growing theca cell monolayers
on collagen-coated wells results in loss of the inhibitory activity (Richard and Sirard, 1998).
Theca cell monolayers require the presence of 10% FCS to produce the inhibitory effect on
oocyte maturation (Richard and Sirard, 1998). Direct contact between the theca cells and the
COCs is not required to maintain bovine oocytes in meiotic arrest because the theca cell
monolayers release the inhibitory factor into the culture medium. Furthermore, the inhibitory
effect of theca cells is fully reversible for up to 24 h (Richard and Sirard, 1996a).
Hypothesis Oocyte maturation is a complex process resulting from the multiple interactions between the
oocyte and the surrounding follicular cells. The maturation of bovine oocytes occurs
spontaneously in vitro. It is possible to modulate the meiotic resumption of bovine oocytes in
vitro with either follicular or chemical products. The present work was aimed to better
understand the meiotic resumption of bovine oocytes in vitro. Experiments were carried out
using bovine oocytes collected from cow ovaries obtained at the slaughterhouse. The working
hypothesis is that inhibition of meiosis of bovine oocytes by the theca cells involves the
interaction between multiple signaling pathways. Therefore, the following hypothesis were
tested:
1) The morphology of the bovine cumulus-oocyte complex has an impact on the resumption of
meiosis and the effectiveness of meiotic inhibitors.
2) The inhibition of specific subtypes of phosphodiesterases in the oocyte and the cumulus cells
of bovine cumulus oocyte complexes may be used to control meiotic arrest.
3) The interactions between the granulosa and theca cells are important to modulate the
resumption of meiosis of bovine COCs in vitro. Moreover the modulation of the PKA signaling
pathway is involved in controlling oocyte maturation.
4) Theca cells produce a factor that is responsible for the maintenance of bovine oocytes in
meiotic arrest. Partial characterization of this inhibitory factor may be carried out by simple
biochemical tests.

50
References Adams GP, Kot K, Ginther OJ. Selection of a dominant follicle and suppression of follicular
growth in heifers. Anim Reprod Sci 1993; 30: 259-71. Adams GP, Matteri RL, Kastelic JP, Ko JC, Ginther OJ. Association between surges of follicle-
stimulating hormone and the emergence of follicular waves in heifers. J Reprod Fertil 1992; 94: 177-88.
Aktas H, Wheeler MB, First NL, Leibfried-Rutledge ML. Maintenance of meiotic arrest by increasing [cAMP]i may have physiological relevance in bovine oocytes. J Reprod Fertil 1995a; 105: 237-45.
Aktas H, Wheeler MB, Rosenkrans CF, Jr., First NL, Leibfried-Rutledge ML. Maintenance of bovine oocytes in prophase of meiosis I by high [cAMP]i. J Reprod Fertil 1995b; 105: 227-35.
Alexandre H, Van Cauwenberge A, Tsukitani Y, Mulnard J. Pleiotropic effect of okadaic acid on maturing mouse oocytes. Development 1991; 112: 971-80.
Aquino FP, Naito K, Cruz LC, Sato E, Toyoda Y. Effects of vanadate on meiotic maturation of porcine oocytes in vitro. J Reprod Dev 1995; 41: 271-276.
Arlotto T, Schwartz JL, First NL, Leibfried Rutledge ML. Aspects of follicle and oocyte stage that affect in vitro maturation and development of bovine oocytes. Theriogenology 1996; 45: 943-956.
Armstrong DT. Effects of maternal age on oocyte developmental competence. Theriogenology 2001; 55: 1303-22.
Assey RJ, Hyttel P, Greve T, Purwantara B. Oocyte morphology in dominant and subordinate follicles. Mol Reprod Dev 1994; 37: 335-344.
Atienza JM, Susanto D, Huang C, McCarty AS, Colicelli J. Identification of inhibitor specificity determinants in a mammalian phosphodiesterase. J Biol Chem 1999; 274: 4839-47.
Avery B, Hay-Schmidt A, Hyttel P, Greve T. Embryo development, oocyte morphology, and kinetics of meiotic maturation in bovine oocytes exposed to 6-dimethylaminopurine prior to in vitro maturation. Mol Reprod Dev 1998; 50: 334-44.
Ayoub MA, Hunter AG. Inhibitory effect of bovine follicular fluid on in vitro maturation of bovine oocytes. J Dairy Sci 1993; 76: 95-100.
Ball GD, Leibfried ML, Ax RL, First NL. Maturation and fertilization of bovine oocytes in vitro. J Dairy Sci 1984; 67: 2775-85.
Barbacid M, Fresno M, Vazquez D. Inhibitors of polypeptide elongation on yeast polysomes. J Antibiot (Tokyo) 1975; 28: 453-62.
Beavo JA. Cyclic nucleotide phosphodiesterases: functional implications of multiple isoforms. Physiol Rev 1995; 75: 725-48.
Beebe SJ, Beasley-Leach A, Corbin JD. cAMP analogs used to study low-Km, hormone-sensitive phosphodiesterase. Methods Enzymol 1988; 159: 531-40.
Bell RM. Protein kinase C activation by diacylglycerol second messengers. Cell 1986; 45: 631-2. Bergfelt DR, Ginther OJ. Ovarian, uterine and embryo dynamics in horses versus ponies. Journal
of Equine Veterinary Science 1996; 16: 66-72. Besmer P, Manova K, Duttlinger R, Huang E, Packer A, Gyssler C, et al. The kit-ligand (steel
factor) and its receptor c-kit/W:pleiotropic roles in gametogenesis and melanogenesis. Development Suppl 1993: 125-137.

51
Bilodeau S, Fortier MA, Sirard MA. The effect of adenylate cyclase stimulation on meiotic resumption and cyclic AMP content of zona-free and cumulus-enclosed bovine oocytes in vitro. J Reprod Fertil 1993; 97: 5-11.
Birmbaumer L, Codina J, Mattera R, Atani A, Sherer N, Goro MJ, et al. Signal transduction by G proteins. 1987; Kidney International.
Blondin P, Bousquet D, Herménégilde T, Barnes F, Sirard MA. Manipulation of follicular development to produce developmentally competent bovine oocytes. Biol Reprod 2002; 66: 38-43.
Blondin P, Coenen K, Guilbault LA, Sirard MA. Superovulation can reduce the developmental competence of bovine embryos. Theriogenology 1996a; 46: 1191-1203.
Blondin P, Dufour M, Sirard MA. Analysis of atresia in bovine follicles using different methods: flow cytometry, enzyme-linked immunosorbent assay, and classic histology. Biol Reprod 1996b; 54: 631-7.
Blondin P, Guilbault LA, Sirard MA. In vitro production of bovine embryos: Developmental competence is acquired before maturation. Theriogenology 1995; 43: 168.
Blondin P, Sirard MA. The influence of oocyte and follicular morphology on developmental competence in superovulated heifers. Theriogenology 1994; 41: 164.
Blondin P, Sirard MA. Oocyte and follicular morphology as determining characteristics for developmental competence in bovine oocytes. Mol Reprod Dev 1995; 41: 54-62.
Blondin P, Sirard MA. The time interval between FSH-P administration and slaughter can influence the developmental competence of beef cattle oocytes. Theriogenology 1997; 47: 184.
Bousquet D, Milovanov C, Bell JC, Durocher J, Smith LC. Nuclear and cytoplasmic maturation of oocytes aspirated from large follicles in superovulated heifers. Theriogenology 1995; 43: 172.
Brackett BG, Zuelke KA. Analysis of factors involved in the in vitro production of bovine embryos. Theriogenology 1993; 39: 43-64.
Brackett RB, Bousquet D, Boice ML, Donawick WJ, Evans JF, Dressel MA. Normal development following in vitro fertilization in the cow. Biol Reprod 1982; 27: 147-58.
Braun T, Dods RF. Development of a Mn-2+-sensitive, "soluble" adenylate cyclase in rat testis. Proc Natl Acad Sci U S A 1975; 72: 1097-101.
Braw-Tal R, Yossefi S. Studies in vivo and in vitro on the initiation of follicle growth in the bovine ovary. J Reprod Fertil 1997; 109: 165-171.
Bryskov AG, Hoyer PE. Embryology of mammalian gonads and ducts. In: Knobil E and Neill J, editors. The physiology of reproduction. Vol 1. New York: Raven Press, 1994: 265-302.
Buccione R, Schroeder AC, Eppig JJ. Interactions between somatic cells and germ cells throughtout mammalian oogenesis. Biol Reprod 1990; 43: 543-47.
Buck J, Sinclair ML, Schapal L, Cann MJ, Levin LR. Cytosolic adenylyl cyclase defines a unique signaling molecule in mammals. Proc Natl Acad Sci U S A 1999; 96: 79-84.
Byskov AG. The anatomy and ultrastructure of the rete system in the fetal mouse ovary. Biol Reprod 1978; 19: 720-35.
Byskov AG, Hoyer PE. Embryology of mammalian gonads and ducts. In: Knobil E and Neill JD, editors. The Physiology of Reproduction. New York: Raven Press Ltd, 1994: 487-540.
Byskov AG, Lintern-Moore S. Follicle formation in the immature mouse ovary:the role of the rete ovarii. Journal of Anatomy 1973; 116: 207-17.

52
Calarco PG. Polarization of mitochondria in the unfertilized mouse oocyte. Dev Genet 1995; 16: 36-43.
Callesen H, Greve T, Hyttel P. Preovulatory endocrinology and oocyte maturation in superovulated cattle. Theriogenology 1986; 43: 1115-28.
Carbonneau G, Sirard MA. Influence of follicular wall on meiotic resumption of bovine oocytes when cultured inside or outside hemisections. J Reprod Dev 1994; 40: 125-132.
Carolan C, Lonergan P, Monget P, Monniaux D, Mermillod P. Effect of follicle size and quality on the ability of follicular fluid to support cytoplasmic maturation of bovine oocytes. Mol Reprod Dev 1996; 43: 477-83.
Castagna M, Takai Y, Kaibuchi K, Sano K, Kikkawa U, Nishizuka Y. Direct activation of calcium-activated, phospholipid-dependent protein kinase by tumor-promoting phorbol esters. J Biol Chem 1982; 257: 7847-51.
Chang M. The maturation of rabbit oocytes in culture and their maturation, activation, fertilization, and subsequent development in the fallopian tubes. J Exp Zool 1955; 128: 378-405.
Channing CP, Hillensjo T, Schaerf FW. Hormonal control of oocyte meiosis, ovulation and luteinization in mammals. Clin Endocrinol Metab 1978; 7: 601-24.
Chesnel F, Eppig JJ. Synthesis and accumulation of p34cdc2 and cyclin B in mouse oocytes during acquisition of competence to resume meiosis. Mol Reprod Dev 1995; 40: 503-8.
Choi T, Aoki F, Mori M, Yamashita M, Nagahama Y, Kohmoto K. Activation of p34cdc2 protein kinase activity in meiotic and mitotic cell cycles in mouse oocytes and embryos. Development 1991; 113: 789-95.
Christmann L, Jung T, Moor RM. MPF components and meiotic competence in growing pig oocytes. Mol Reprod Dev 1994; 38: 85-90.
Colicelli J, Birchmeier C, Michaeli T, O'Neill K, Riggs M, Wigler M. Isolation and characterization of a mammalian gene encoding a high- affinity cAMP phosphodiesterase. Proc Natl Acad Sci U S A 1989; 86: 3599-603.
Collas P, Fissore R, Robl JM, Sullivan EJ, Barnes FL. Electrically induced calcium elevation, activation, and parthenogenetic development of bovine oocytes. Mol Reprod Dev 1993a; 34: 212-23.
Collas P, Sullivan EJ, Barnes FL. Histone H1 kinase activity in bovine oocytes following calcium stimulation. Mol Reprod Dev 1993b; 34: 224-31.
Conti M. Phosphodiesterases and cyclic nucleotides signaling in endocrine cells. Mol Endocrinol 2000; 14: 1317-1327.
Conti M, Jin SL. The molecular biology of cyclic nucleotide phosphodiesterases. Prog Nucleic Acid Res Mol Biol 1999; 63: 1-38.
Conti M, Jin SL, Monaco L, Repaske DR, Swinnen JV. Hormonal regulation of cyclic nucleotide phosphodiesterases. Endocr Rev 1991; 12: 218-34.
Cran DG. Cortical granules during oocyte maturation and fertilization. J Reprod Fertil Suppl 1989; 38: 49-62.
Crozet N. Nucleolar structure and RNA synthesis in mammalian oocytes. J Reprod Fertil Suppl 1989; 38: 9-16.
Crozet N, Kanka J, Motlik J, Fulka J. Nucleolar fine structure and RNA synthesis in bovine oocytes from antral follicles. Gam Res 1986; 14: 65-73.

53
Daen FP, Sato E, Naito K, Toyoda Y. The effect of pig follicular fluid fractions on cumulus expansion and male pronucleus formationin porcine oocytes matured and fertilized in vitro. J Reprod Fertil 1994; 101: 667-73.
Davis JS, Weakland LL, Farese RV, West LA. Luteinizing hormone increases inositol trisphosphate and cytosolic free Ca2+ in isolated bovine luteal cells. J Biol Chem 1987; 262: 8515-21.
De Loos F, Kastrop P, Van Maurik P, Van Beneden TH, Kruip TAM. Heterologous cell contacts and metabolic coupling in bovine cumulus oocyte complexes. Mol Reprod Dev 1991; 28: 255-9.
De Loos FA, Zeinstra E, Bevers MM. Follicular wall maintains meiotic arrest in bovine oocytes cultured in vitro. Mol Reprod Dev 1994; 39: 162-5.
de Vantery C, Gavin AC, Vassalli JD, Schorderet-Slatkine S. An accumulation of p34cdc2 at the end of mouse oocyte growth correlates with the acquisition of meiotic competence. Dev Biol 1996; 174: 335-44.
de Vantery C, Stutz A, Vassalli JD, Schorderet-Slatkine S. Acquisition of meiotic competence in growing mouse oocytes is controlled at both translational and posttranslational levels. Dev Biol 1997; 187: 43-54.
de Wit AAC. Bovine cumulus-oocyte-complex quality is reflected in sensitivity for α-amanitin, oocyte diameter and developmental capacity. Anim Reprod Sci 2001; 65: 51-65.
Dekel N. Regulation of oocyte maturation. The role of cAMP. Ann N Y Acad Sci 1988; 541: 211-6.
Dekel N. Molecular control of meiosis. Trends in Endocrinology and Metabolism 1995; 6: 165-169.
Dieleman SJ, Kruip TAM, Fontijne P, de Jong WHR, van der Weyden GC. Changes in oestradiol, progesterone and testosterone concentration in follicular fluid and in micromorphologyof preovulatory bovine follicles relative to the peak of luteinizing hormone. J Endocrinol 1983; 97: 31-42.
Dode MA, Adona PR. Developmental capacity of Bos indicus oocytes after inhibition of meiotic resuption by 6-dimethylaminopurine. Anim Reprod Sci 2001; 65: 171-80.
Dominko T, First NL. Timing of meiotic progression in bovine oocytes and its effect on early embryo development. Mol Reprod Dev 1997; 47: 456-67.
Donovan PJ, Stott D, Cairns LA, Heasman J, Wylie CC. Migratory and postmigratory mouse primordial germ cells behave differently in culture. Cell 1986; 44: 831-8.
Dostal J, Pavlok A. Isolation and characterization of maturation inhibiting compound in bovine follicular fluid. Reprod Nutr Dev 1996; 36: 681-90.
Downs SM. Purine control of mouse oocyte maturation - evidence that nonmetabolized hypoxanthine maintains meiotic arrest. Mol Reprod Dev 1993; 35: 82-94.
Downs SM. Uptake and metabolism of adenosine mediate a meiosis-arresting action on mouse oocytes. Mol Reprod Dev 1999; 53: 208-21.
Downs SM, Coleman DL, Eppig JJ. Maintenance of murine oocyte meiotic arrest: uptake and metabolism of hypoxanthine and adenosine by cumulus cell-enclosed and denuded oocytes. Dev Biol 1986; 117: 174-83.
Downs SM, Coleman DL, Ward-Bailey PF, Eppig JJ. Hypoxanthine is the principal inhibitor of murine oocyte maturation in a low molecular weight fraction of porcine follicular fluid. Proc Natl Acad Sci U S A 1985; 82: 454-8.

54
Downs SM, Daniel SA, Bornslaeger EA, Hoppe PC, Eppig JJ. Maintenance of meiotic arrest in mouse oocytes by purines: modulation of cAMP levels and cAMP phosphodiesterase activity. Gamete Res 1989; 23: 323-34.
Downs SM, Eppig JJ. Cyclic adenosine monophosphate and ovarian follicular fluid act synergistically to inhibit mouse oocyte maturation. Endocrinology 1984; 114: 418-27.
Downs SM, Eppig JJ. Induction of mouse oocyte maturation in vivo by perturbants of purine metabolism. Biol Reprod 1987; 36: 431-7.
Downs SM, Hunzicker-Dunn M. Differential regulation of oocyte maturation and cumulus expansion in the mouse oocyte-cumulus cell complex by site-selective analogs of cyclic adenosine monophosphate. Dev Biol 1995; 172: 72-85.
Driancourt MA. Follicular dynamics in sheep and cattle. Theriogenology 1991; 35: 55-68. Ducibella T, Duffy P, Buetow J. Quantification and localization of cortical granules during
oogenesis in the mouse. Biol Reprod 1994; 50: 467-73. Dunphy WG, Kumagai A. The cdc25 protein contains an intrinsic phosphatase activity. Cell
1991; 67: 189-96. Duranthon V, Renard JP. The developmental competence of mammalian oocytes: a convenient
but biologically fuzzy concept. Theriogenology 2001; 55: 1277-89. Dyce KM, Sack WO, Wensing CJG. The pelvic and reproductive organs of the female ruminants.
Philadelphia: W B Saunders Company, 1987. Eddy EM, Clark JM, Gong D, Fenderson BA. Origin and migration of primordial germ cells in
mammals. Gam Res 1981; 4: 333-62. Edwards R. Maturation in vitro of mouse, sheep, cow, pig, rhesus monkey and human ovaries.
Nature 1965; 208: 349-351. Egyhazi E. Inhibition of balbiani ring RNA synthesis at the initiation level. Proc Natl Acad Sci U
S A 1975; 72: 947-50. Eppig JJ, Downs SM. Chemical signals that regulate mammalian oocyte maturation. Biol Reprod
1984; 30: 1-11. Eppig JJ, Downs SM. The effect of hypoxanthine on mouse oocyte growth and development in
vitro: maintenance of meiotic arrest and gonadotropin-induced oocyte maturation. Dev Biol 1987; 119: 313-21.
Eppig JJ, Ward-Bailey PF, Coleman DL. Hypoxanthine and adenosine in murine ovarian follicular fluid: concentrations and activity in maintaining oocyte meiotic arrest. Biol Reprod 1985; 33: 1041-9.
Erikson BH. Development and senescence of the post natal bovine ovary. Journal of Animal Science 1966; 25: 800-05.
Erikson E, Maller JL. Biochemical characterization of the p34cdc2 protein kinase component of purified maturation-promoting factor from Xenopus eggs. J Biol Chem 1989; 264: 19577-82.
Espey LL. Current status of the hypothesis that mammalian ovulation is comparable to an inflammatory reaction. Biol Reprod 1994; 50: 233-8.
Evans AC, Adams GP, Rawlings NC. Follicular and hormonal development in prepubertal heifers from 2 to 36 weeks of age. J Reprod Dev 1994; 102: 462-70.
Faerge I, Mayes M, Hyttel P, Sirard MA. Nuclear ultrastructure in bovine oocytes after inhibition of meiosis by chemical and biological inhibitors. Mol Reprod Dev 2001; 59: 459-67.

55
Fair T. Oocyte growth in cattle: Ultrastructure, transcription and developmental competence. Department of Animal Science and Production. Dublin: University College Dublin, 1995: Thesis
Fair T, Hulshof SCJ, Hyttel P, Greve T, Boland M. Oocyte ultrastructure in bovine primordial to tertiary follicles. Anat Embryol 1997; 195: 327-336.
Fair T, Hyttel P, Greve T. Bovine oocyte diameter in relation to maturational competence and transcriptional activity. Mol Reprod Dev 1995; 42: 437-42.
Fair T, Hyttel P, Greve T, Boland M. Nucleus structure and transcriptional activity in relation to oocyte diameter in cattle. Mol Reprod Dev 1996; 43: 503-12.
Farin CE, Yang L. Inhibition of germinal vesicle breakdown in bovine oocytes by 5,6- dichloro-1-beta-D-ribofuranosylbenzimidazole (DRB). Mol Reprod Dev 1994; 37: 284-92.
Fisher DA, Smith JF, Pillar JS, St Denis SH, Cheng JB. Isolation and characterization of PDE9A, a novel human cGMP-specific phosphodiesterase. J Biol Chem 1998; 273: 15559-64.
Fissore RA, He CL, Vande Woude GF. Potential role of mitogen-activated protein kinase during meiosis resumption in bovine oocytes. Biol Reprod 1996; 55: 1261-70.
Fukui Y, Sakuma Y. Maturation of bovine oocytes cultured in vitro: relation to ovarian activity, follicular size and the presence or absence of cumulus cells. Biol Reprod 1980; 22: 669-73.
Fulka J, Jr, Motlik J, Fulka J, Jilek F. Effect of cycloheximide on nuclear maturation of pig and mouse oocytes. J Reprod Fertil 1986; 77: 281-285.
Fulka J, Jr., Jung T, Moor RM. The fall of biological maturation promoting factor (MPF) and histone H1 kinase activity during anaphase and telophase in mouse oocytes. Mol Reprod Dev 1992; 32: 378-82.
Fulka J, Jr., Leibfried-Rutledge ML, First NL. Effect of 6-dimethylaminopurine on germinal vesicle breakdown of bovine oocytes. Mol Reprod Dev 1991; 29: 379-84.
Gabev E, Kasianowicz J, Abbot T, McLaughlin S. Binding of neomycin to phosphatidylinositol 4,5-biphosphate (PIP2). Biochem Biophys Acta 1989; 979: 105-12.
Galli C, Moor RM. Somatic cells and the G2 to M-phase transition in sheep oocytes. Reprod Nutr Dev 1991; 31: 127-34.
Gandolfi F. Embryonic development as a function of oocyte competence. In: Lauria A, Gandolfi F, Enne G and Gianaroli L, editors. Gametes development and function. Rome: Serono Symposia, 1998: 337-53.
Gandolfi F, Luciano AM, Modina S, Ponzini A, Pocar P, Armstrong DT, et al. The in vitro developmental competence of bovine oocytes can be related to the morphology of the ovary. Theriogenology 1997; 48: 1153-1160.
Gautier J, Norbury C, Lohka M, Nurse P, Maller J. Purified maturation promoting factor contains the product of a Xenopus homolog of the fission yeast cell cycle control gene cdc2. Cell 1988; 54: 433-39.
Gilman AG. G-proteins: transducers of receptor-generated signals. Annual Review of Biochemistry 1987; 56: 615-30.
Ginther OJ, Beg MA, Bergfelt DR, Donadeu FX, Kot K. Follicle selection in monovular species. Biol Reprod 2001a; 65: 638-47.
Ginther OJ, Bergfelt DR, Beg MA, Kot K. Follicle selection in cattle: relationships among growth rate, diameter ranking, and capacity for dominance. Biol Reprod 2001b; 65: 345-50.

56
Ginther OJ, Kastelic JP, Knopf L. Composition and characteristics of follicular waves during the bovine estrous cycle. Anim Reprod Sci 1989a; 20: 187-200.
Ginther OJ, Kastelic JP, Knopf L. Intraovarian relationshiop among dominant and subordinate follicles and corpus luteun in heifers. Theriogenology 1989b; 32: 787-95.
Ginther OJ, Knopf L, Kastelic JP. Temporal associations among ovarian events in cattle during oestrous cycles with two and three follicular waves. J Reprod Fertil 1989c; 87: 223-30.
Ginther OJ, Wiltbank MC, Fricke PM, Gibbons JR, Kot K. Selection of the dominant follicle in cattle. Biol Reprod 1996; 55: 1187-94.
Gordon I. Laboratory production of cattle embryos. Wallingford, UK: CAB International, 1994. Gordon I, Lu KH. Production of embryos in vitro and its impact on livestock production.
Theriogenology 1990; 33: 77-87. Goren S, Dekel N. Maintenance of meiotic arrest by a phosphorylated p34cdc2 is independent of
cyclic adenosine 3',5'-monophosphate [published erratum appears in Biol Reprod 1994; Dec;51(6):1330]. Biol Reprod 1994; 51: 956-62. Gosden RG. Ovulation 1: Oocyte develoment throughout life. In: Grudzinskas JG and Yovich JL,
editors. Gametes - The oocyte. Vol III. Cambridge: Cambridge University Press, 1995: 119-49.
Gougeon A. Regulation of ovarian follicular development in primates: facts and hypothesis. Endocrine Rev 1996; 17: 121-155.
Greenwalt GS, Terranova PF. Follicular selection and its control. In: Knobil E and Neill J, editors. Physiology of Reproduction. New York: Raven Press, 1988: 387-445.
Greve T, Xu KP, Callesen H, Hyttel P. The effects of exogenous gonadotropins on oocyte and embryo quality in cattle. J of In Vitro Fertilization and Embryo Transfer 1987; 4: 281-5.
Gwatkin RB, Andersen OF. Hamster oocyte maturation in vitro: inhibition by follicular components. Life Sci 1976; 19: 527-36.
Hamilton SA, Xu ZZ, Kieborz KR, Youngquist RS, Garverick HA. Relationship between ovarian follicular dynamics and follicle stimulating hormone levels during the bovine estrous cycle. J Anim Sci 1992; 70 (Suppl 1): 261.
Harada M, Miyano T, Matsumura K, Osaki S, Miyake M, Kato S. Bovine oocytes from early antral follicles grow to meiotic competence in vitro: Effect of FSH and hypoxanthine. Theriogenology 1997; 48: 743-755.
Harrison RJ, Weir BJ. Structure of the mammalian ovary. In: Zuckerman L and Weir BJ, editors. The ovary I: General Aspects. New York: Academic Press, 1977.
Harrison SA, Reifsnyder DH, Gallis B, Cadd GG, Beavo JA. Isolation and characterization of bovine cardiac muscle cGMP-inhibited phosphodiesterase: a receptor for new cardiotonic drugs. Mol Pharmacol 1986; 29: 506-14.
Hashimoto N, Kishimoto T. Regulation of meiotic metaphase by a cytoplasmic maturation-promoting factor during mouse oocyte maturation. Dev Biol 1988; 126: 242-52.
Hashimoto Y, Eguchi Y. Histological observations on the gonads in the cattle and horse fetus. Japanese Journal of Zoo Technology 1955; 26: 259-266.
Hazeleger NL, Hill DJ, Stubbings RB, Walton JS. Relationship of morphology and follicular fluid environment of bovine oocytes to their developmental potential in vitro. Theriogenology 1995; 43: 509-522.

57
Hendriksen PJ, Vos PL, Steenweg WN, Bevers MM, Dieleman SJ. Bovine follicular development and its effect on the in vitro competence of oocytes. Theriogenology 2000; 53: 11-20.
Hillensjo T, Batta SK, Schwartz-Kripner A, Wentz AC, Sulewski J, Channing CP. Inhibitory effect of human follicular fluid upon the maturation of porcine oocytes in culture. J Clin Endocrinol Metab 1978; 47: 1332-5.
Hillensjo T, Channing CP, Pomerantz SH, Schwartz-Kripner A. Intrafollicular control of oocyte maturation in the pig. In Vitro 1979; 15: 32-9.
Hirshfield AN. Development of follicles in the mammalian ovary. International Review of Cytology 1991a; 124: 46-101.
Hirshfield AN. Theca cells may be present at the outset of follicular growth. Biol Reprod 1991b; 44: 1157-62.
Homa ST. Effects of cyclic AMP on the spontaneous meiotic maturation of cumulus-free bovine oocytes cultured in chemically defined medium. J Exp Zool 1988; 248: 222-31.
Homa ST. Neomycin, an inhibitor of phosphoinositide hydrolysis, inhibits the resumption of bovine oocyte spontaneous meiotic maturation. J Exp Zool 1991; 258: 95-103.
Homa ST. Calcium and meiotic maturation of the mammalian oocyte. Mol Reprod Dev 1995; 40: 122-134.
Homa ST, Brown CA. Changes in linoleic acid during follicular development and inhibition of spontaneous breakdown of germinal vesicles in cumulus-free bovine oocytes. J Reprod Fertil 1992; 94: 153-60.
Homa ST, Carroll J, Swann K. The role of calcium in mammalian oocyte maturation and egg activation. Human Reproduction 1993; 8: 1274-1281.
Hunter AG, Moor RM. Stage dependent effects of inhibiting RNA and protein synthesis on meiotic maturation of bovine oocytes in vitro. J Dairy Sci 1987; 70: 1646-1651.
Hyttel P. Bovine cumulus-oocyte disconnection in vitro. Anat Embryol (Berl) 1987; 176: 41-4. Hyttel P, Callesen H, Greve T. Ultrastructural features of preovulatory oocyte maturation in
superovulated cattle. J Reprod Fertil 1986a; 76: 645-56. Hyttel P, Fair T, Callesen H, Greve T. Oocyte growth, capacitation and final maturation in cattle.
Theriogenology 1997; 47: 23-32. Hyttel P, Madsen I. Rapid method to prepare mammalian oocytes and embryos for transmission
electron microscopy. Acta Anat 1987; 129: 12-4. Hyttel P, Xu KP, Smith S, Greve T. Ultrastructure of in-vitro oocyte maturation in cattle. J
Reprod Fertil 1986b; 78: 615-25. Ireland JJ, Roche JF. Development of antral follicles in cattle after prostaglandin-induced
luteolysis: Changes in serum hormone, steroids in follicular fluid and gonadotropin receptors. Endocrinol 1982; 111: 2077-2086.
Ireland JJ, Roche JF. Development of non ovulatory antral follicles in heifers: changes in steroid in follicular fluid and receptors for gonadotrophins. Endocrinology 1983a; 112: 150-56.
Ireland JJ, Roche JF. Growth and differentiation of large antral follicles after spontaneous luteolysis in heifers: changes in concentration of hormones in follicular fluid and specific binding of gonadotropins to follicles. J Anim Sci 1983b; 57: 157-67.
Jacobs T. Control of the cell cycle. Dev Biol 1992; 153: 1-15. Jagiello G, Ducayen MB, Downey R, Jonassen A. Alterations of mammalian oocyte meiosis I
with divalent cations and calmodulin. Cell Calcium 1982; 3: 153-62.

58
Jagiello G, Graffo J, Ducayen M, Prosser R. Further studies of inhibitors of in vitro mammalian oocyte maturation. Fertil Steril 1977; 28: 476-81.
Jaken S. Protein kinase C isozymes and substrates. Curr Opin Cell Biol 1996; 8: 168-73. Jones KT. Protein kinase C action at fertilization: overstated or undervalued? Rev Reprod 1998;
3: 7-12. Kadam AL, Koide SS. Identification of hypoxanthine in bovine follicular fluid. J Pharm Sci
1990; 79: 1077-82. Kadam AL, Koide SS. A follicular fluid factor inhibiting Xenopus oocyte maturation. Endocr
Res 1991; 17: 343-55. Kalous J, Kubelka M, Rimkevicova Z, Guerrier P, Motlik J. Okadaic acid accelerates germinal
vesicle breakdown and overcomes cycloheximide- and 6-dimethylaminopurine block in cattle and pig oocytes. Dev Biol 1993a; 157: 448-54.
Kalous J, Sutovsky P, Rimkevicova Z, Shioya Y, Lie BL, Motlik J. Pig membrana-granulosa cells prevent resumption of meiosis in cattle oocytes. Mol Reprod Dev 1993b; 34: 58-64.
Kastrop PM, Bevers MM, Destree OH, Kruip TAM. Analysis of protein synthesis in morphologically classified bovine follicular oocytes before and after maturation in vitro. Mol Reprod Dev 1990a; 26: 222-6.
Kastrop PM, Bevers MM, Destree OH, Kruip TAM. Changes in protein synthesis and phosphorylation patterns during bovine oocyte maturation in vitro. J Reprod Fertil 1990b; 90: 305-10.
Kastrop PM, Bevers MM, Destree OH, Kruip TAM. Protein synthesis and phosphorylation patterns of bovine oocytes maturing in vivo. Mol Reprod Dev 1991a; 29: 271-5.
Kastrop PMM, Hulshof SCJ, Bevers MM, Destree OHJ, Kruip TAM. The effects of α-amanitin and cycloheximide on nuclear progression, protein synthesis, and phosphorylation during bovine oocyte maturation in vitro. Mol Reprod Dev 1991b; 28: 249-254.
Kerr JF, Wyllie AH, Currie AR. Apoptosis: a basic biological phenomenon with wide-ranging implications in tissue kinetics. Br J Cancer 1972; 26: 239-57.
Kim NH, Day BN, Lee HT, Chung KS. Microfilament assembly and cortical granule distribution during maturation, parthenogenetic activation and fertilisation in the porcine oocyte. Zygote 1996; 4: 145-9.
Kitawa M, Okabe T, Ogino H, Matsumoto H, Suzuki-Takahashi I, Kokubo T, et al. Butyrolactone I, a selective inhibitor of cdk2 and cdc2 kinase. Oncogene 1993; 8: 2425-32.
Knopf L, Kastelic JP, Schallenberger E, Ginther OJ. Ovarian follicular dynamics in heifers Test of a two-wave hypothesis by ultrasonically monitoring individual follicles. Domest Anim Endocrinol 1989; 6: 111-9.
Kotsuji F, Kubo M, Tominaga T. Effect of interactions between granulosa and thecal cells on meiotic arrest in bovine oocytes. Journal of Reproduction and Fertility 1994; 100: 151-156.
Kruip T, Cran D, Van Beneden T, Dieleman S. Structural changes in bovine oocytes during final maturation in vitro. Gam.Res 1983; 8: 29-47.
Kruip TA, Dieleman SJ. Macroscopic classification of bovine follicles and its validation by micromorphological and steroid biochemical procedures. Reprod Nutr Dev 1982; 22: 465-73.

59
Kubelka M, Motlik J, Fulka JJ, Prochazka R, Rimkevikova Z, Fulka J. Time sequence of germinal vesicle breakdown in pig oocytes after cycloheximide and p-aminobenzamidine block. Gam. Res. 1988; 19: 423-431.
Kubelka M, Motlik J, Schultz RM, Pavlok A. Butyrolactone I reversibly inhibits meiotic maturation of bovine oocytes, Without influencing chromosome condensation activity. Biol Reprod 2000; 62: 292-302.
Kubish HM, Larson MA, Barnes F, Sikes JD, Roberts RM. Transgene expression in IVM/IVF-derived bovine embryos after delay on maturation with 6-dimethylaminopurine. Theriogenology 1995; 43: 255.
Kuwana T, Fujimoto T. Active locomotion of human primordial germ cells in vitro. Anat Rec 1983; 204: 21-5.
Kuyt JR, Kruip TA, de Jong-Brink M. Cytochemical localization of adenylate cyclase in bovine cumulus-oocyte complexes. Exp Cell Res 1988; 174: 139-45.
Laurincik J, Kroslak P, Hyttel P, Pivko J, Sirotkin AV. Bovine cumulus expansion and corona-oocyte disconnection during culture in vitro. Reprod Nutr Dev 1992; 32: 151-61.
Le Gal F, Gall L, De Smedt V. Changes in protein synthesis pattern during in vitro maturation of goat oocytes. Mol Reprod Dev 1992; 32: 1-8.
Leibfried L, First NL. Effects of divalent cations on in vitro maturation of bovine oocytes. J Exp Zool 1979a; 210: 575-80.
Leibfried L, First NL. Effect of bovine and porcine follicular fluid and granulosa cells on maturation of oocytes in vitro. Biol Reprod 1980a; 23: 699-704.
Leibfried L, First NL. Follicular control of meiosis in the porcine oocyte. Biol Reprod 1980b; 23: 705-9.
Leibfried ML, First NL. Characterization of bovine follicular oocytes and their ability to mature in vitro. J Anim Sci 1979b; 48: 76-86.
Leibfried Rutledge ML, Critser ES, Parrish JJ, First NL. In vitro maturation and fertilization of bovine oocytes. Theriogenology 1989; 31: 61-74.
Leibfried-Rutledge ML, Critser ES, Eyestone WH, Northey DL, First NL. Development potential of bovine oocytes matured in vitro or in vivo. Biol Reprod 1987; 36: 376-83.
Levesque JT, Sirard MA. Effects of different kinases and phosphatases on nuclear and cytoplasmic maturation of bovine oocytes. Mol Reprod Dev 1995; 42: 114-21.
Levesque JT, Sirard MA. Resumption of meiosis is initiated by the accumulation of cyclin B in bovine oocytes. Biol Reprod 1996; 55: 1427-1436.
Lonergan P, Carolan C, Mermillod P. Development of bovine embryos in vitro following oocyte maturation under defined conditions. Reprod Nutr Dev 1994a; 34: 329-39.
Lonergan P, Dinnyes A, Fair T, Yang X, Boland M. Bovine oocyte and embryo development following meiotic inhibition with butyrolactone I. Mol Reprod Dev 2000; 57: 204-209.
Lonergan P, Khatir H, Carolan C, Mermillod P. Bovine blastocyst production in vitro after inhibition of oocyte meiotic resumption for 24 h. J Reprod Fertil 1997; 109: 355-65.
Lonergan P, Monaghan P, Rizos D, Boland MP, Gordon I. Effect of follicle size on bovine oocyte quality and developmental competence following maturation, fertilization, and culture in vitro. Mol Reprod Dev 1994b; 37: 48-53.
Loten EG, Assimacopoulos-Jeannet FD, Exton JH, Park CR. Stimulation of a low Km phosphodiesterase from liver by insulin and glucagon. J Biol Chem 1978; 253: 746-57.

60
Lu KH, Gordon I, Gallagher M, McGovern H. Pregnancy established in cattle by transfer of embryos derived from in vitro fertilisation of oocytes matured in vitro. Vet Rec 1987; 121: 259-60.
Lussier JG, Matton P, Dufour JJ. Growth rates of follicles in the ovary of the cow. J Reprod Fertil 1987; 81: 301-07.
Madison V, Avery B, Greve T. Selection of immature bovine oocytes for developmental potential in vitro. Anim Reprod Sci 1992; 27: 1-11.
Martus NS, Farin CE. Effectiveness of DRB for inhibiting germinal vesicle breakdown in bovine oocytes. Theriogenology 1994; 42: 1295-1302.
Maruska DV, Leibfried ML, First NL. Role of calcium and the calcium-calmodulin complex in resumption of meiosis, cumulus expansion, viability and hyaluronidase sensitivity of bovine cumulus-oocyte complexes. Biol Reprod 1984; 31: 1-6.
Masui Y. The cytostatic factor (CSF) that causes metaphase arrest in amphibian eggs. Vol H-45, 1990.
Masui Y, Clarke HJ. Oocyte maturation. Int Rev Cytol 1979; 57: 185-282. Masui Y, Markert CL. Cytoplasmic control of nuclear behavior during maturation of frog
oocytes. J Exp Zool 1971; 177: 129-46. Mayes MA, Sirard MA. The influence of cumulus-oocyte complex morphology and meiotic
inhibitors on the kinetics of nuclear maturation in cattle. Theriogenology 2001; 55: 911-22.
McGaughey RW. In vitro oocyte maturation. In: Daniel Jr JC, editor. Methods in mammalian reproduction. New York: Academic Press, 1978: 1-20.
McGee EA, Hsueh AJ. Initial and cyclic recruitment of ovarian follicles. Endocr Rev 2000; 21: 200-14.
Meacci E, Taira M, Moos M, Jr., Smith CJ, Movsesian MA, Degerman E, et al. Molecular cloning and expression of human myocardial cGMP-inhibited cAMP phosphodiesterase. Proc Natl Acad Sci U S A 1992; 89: 3721-5.
Meijer L, Borgne A, Mulner O, Chong JP, Blow JJ, Inagaki N, et al. Biochemical and cellular effects of roscovitine, a potent and selective inhibitor of the cyclin-dependent kinases cdc2, cdk2 and cdk5. Eur J Biochem 1997; 243: 527-36.
Meinecke B, Meinecke-Tillmann S. Effects of alpha-amanitin on nuclear maturation of porcine oocytes in vitro. J Reprod Fertil 1993; 98: 195-201.
Memili E, First NL. Developmental changes in RNA polymerase II in bovine oocytes, early embryos, and effect of alpha-amanitin on embryo development. Mol Reprod Dev 1998; 51: 381-9.
Mermillod P. La production d'embryons bovins in vitro. Tours: Université François Rabelais, 1998: 33.
Mermillod P, Tomanek M, Marchal R, Meijer L. High developmental competence of cattle oocytes maintained at the germinal vesicle stage for 24 hours in culture by specific inhibition of MPF kinase activity. Mol Reprod Dev 2000; 55: 89-95.
Monniaux D, Monget P, Besnard N, Huet C, Pisselet C. Growth factors and antral follicular development in domestic ruminants. Theriogenology 1997; 47: 3-12.
Moor RM. Sites of steroid production in ovine graafian follicles in culture. J Endocrinol 1977; 73: 143-50.

61
Moor RM, Crosby IM. Protein requirements for germinal vesicle breakdown in ovine oocytes. J Embryol Exp Morphol 1986; 94: 207-220.
Moor RM, Trounson AO. Hormonal and follicular factors affecting maturation of sheep oocytes in vitro and their subsequent developmental capacity. J Reprod Fertil 1977; 49: 101-9.
Motlik J, Crozet N, Fulka J. Meiotic competence in vitro of pig oocytes isolated from early antral follicles. J Reprod Fertil 1984; 72: 323-8.
Motlik J, Fulka J. In vitro maturation of mammalian oocytes. Ontogenez 1981; 12: 435-42. Motlik J, Fulka J. Factors affecting meiotic competence in pig oocytes. Theriogenology 1986; 25:
87-96. Motlik J, Fulka J, Jr., Prochazka R, Rimkevicova Z, Kubelka M, Fulka J. RNA and protein
synthesis requirements for the resumption of meiosis in rabbit oocytes: the role of cumulus cells. Reprod Nutr Dev 1989; 29: 601-9.
Motlik J, Lie B, Shioya Y. Two sensitivity levels of cattle oocytes to puromycin. Biol Reprod 1990; 43: 994-8.
Motlik J, Nagai T, Kikuchi K. Resumption of meiosis in pig oocytes cultured with cumulus and parietal granulosa cells - The effect of protein synthesis Inhibition. J Exp Zool 1991; 259: 386-391.
Motlik J, Pavlok A, Kubelka M, Kalous J, Kalab P. Interplay between CDC2 kinase and MAP kinase pathway during maturation of mammalian oocytes. Theriogenology 1998; 49: 461-469.
Murray AW. Creative blocks: cell-cycle checkpoints and feedback controls. Nature 1992; 359: 599-604.
Murray AW. Turning on mitosis. Curr Biol 1993; 3: 291-3. Murray AW, Solomon MJ, Kirschner MW. The role of cyclins synthesis and degradation in the
control of maturation promoting factor activtiy. Nature 1989; 339: 280-6. Naito K, Fukuda Y, Toyoda Y. Effects of porcine follicular fluid on male pronucleus formation
in porcine oocytes matured in vitro. Gamete Res 1988; 21: 289-95. Naito K, Toyoda Y. Fluctuation of histone H1 kinase activity during meiotic maturation in
porcine oocytes. J Reprod Fertil 1991; 93: 467-73. Nashta AA, Waddington D, Campbell BK. Maintenance of bovine oocytes in meiotic arrest and
subsequent development in vitro: a comparative evaluation of antral follicle culture with other methods. Biol Reprod 1998; 59: 255-62.
Niedel JE, Kuhn LJ, Vandenbark GR. Phorbol diester receptor copurifies with protein kinase C. Proc Natl Acad Sci U S A 1983; 80: 36-40.
Norbury C, Blow J, Nurse P. Regulatory phosphorylation of the p34cdc2 protein kinase in vertebrates. Embo J 1991; 10: 3321-9.
Nurse P. Universal control mechanism regulating the onset of M-phase. Nature 1990; 344: 503-8. Osborn JC, Moor RM. Time-dependent effects of a a-amanitin on nuclear maturation and protein
synthesis in mammalian oocytes. J Embryol Exp Morphol 1983; 73: 317-38. Otoi T, Yamamoto K, Koyama N, Tachikawa S, Suzuki T. Bovine oocyte diameter in relation to
developmental competence. Theriogenology 1997; 48: 769-774. Pavlok A, Lucas-Hahn A, Niemann H. Fertilization and developmental competence of bovine
oocytes derived from different categories of antral follicles. Mol Reprod Dev 1992; 31: 63-7.

62
Peng XR, Hsueh AJ, LaPolt PS, Bjersing L, Ny T. Localization of luteinizing hormone receptor messenger ribonucleic acid expression in ovarian cell types during follicle development and ovulation. Endocrinology 1991; 129: 3200-7.
Perreault SD, Barbee RR, Slott VL. Importance of glutathione in the acquisition and maintenance of sperm nuclear decondensing activity in maturing hamster oocytes. Dev Biol 1988; 125: 181-6.
Peter M, Nakagawa J, Doree M, Labbe JC, Nigg EA. Identification of major nucleolar proteins as candidate mitotic substances of cdc2 kinase. Cell 1990; 60: 791-801.
Petr J, Zetova L, Fulka J, Jr., Jilek F. Quantitative inhibitory influence of porcine cumulus cells upon the maturation of pig and cattle oocytes in vitro. Reprod Nutr Dev 1989; 29: 541-9.
Picton HM. Activation of follicle development: the primordial follicle. Theriogenology 2001; 55: 1193-210.
Pincus G, Enzmann EV. The comparative behavior of mammalian eggs in vitro and in vivo. J. Exp. Med. 1935; 62: 665-675.
Ponderato N, Lagutina I, Crotti G, Turini P, Galli C, Lazzari G. Bovine oocytes treated prior to in vitro maturation with a combination of butyrolactone I and roscovitine at low doses maintain a normal developmental capacity. Mol Reprod Dev 2001; 60: 579-85.
Rabahi F, Monniaux D, Pisselet C, Durand P. Control of in vitro maturation of bovine cumulus-oocyte complex by preovulatory granulosa cells. Mol Reprod Dev 1993; 34: 431-442.
Racowsky C. Follicular control of meiotic maturation in mammalian oocytes. In: Bavister BD, editor. Preimplantation embryo development. New York: Springer-Verlag, 1991: 22-37.
Racowsky C, Baldwin KV. In vitro and in vivo studies reveal that hamster oocyte meiotic arrest is maintained only transiently by follicular fluid, but persistently by membrana/cumulus granulosa cell contact. Dev Biol 1989; 134: 297-306.
Racowsky C, McGaughey RW. Further studies of the effects of follicular fluid and membrana granulosa cells on the spontaneous maturation of pig oocytes. J Reprod Fertil 1982; 66: 505-12.
Rajakoski E. The follicular system in sexually mature heifers with special reference to seasonal, cyclical and left-right variations. Acta Endocrinol Suppl 1960a; 52: 1-68.
Rajakoski E. The ovarian follicular system in sexually mature heifers with special reference to seasonal, cyclical and left-right variations. Acta Endocrinol 1960b; 52: 6-68.
Richard FJ. Effect de différentes composantes cellulaires du follicule ovarien sur le contrôle de la maturation nucléaire chez les ovocytes de bovin. Sciences Animales. Ste. Foy, Québec, Canada: Université Laval, 1997: Thesis.
Richard FJ, Fortier MA, Sirard MA. Role of the cyclic adenosine monophosphate-dependent protein kinase in the control of meiotic resumption in bovine oocytes cultured with thecal cell monolayers. Biol Reprod 1997; 56: 1363-9.
Richard FJ, Sirard MA. Effects of follicular cells on oocyte maturation. I: Effects of follicular hemisections on bovine oocyte maturation in vitro. Biol Reprod 1996a; 54: 16-21.
Richard FJ, Sirard MA. Effects of follicular cells on oocyte maturation. II: Theca cell inhibition of bovine oocyte maturation in vitro. Biol Reprod 1996b; 54: 22-8.
Richard FJ, Sirard MA. Theca cell monolayers that inhibit maturation of bovine oocytes show differences in their protein secretion pattern. Mol Reprod Dev 1998; 50: 200-6.
Richard FJ, Tsafriri A, Conti M. Role of phosphodiesterase type 3A in rat oocyte maturation. Biol Reprod 2001; 65: 1444-51.

63
Rodgers RJ, van Wezel IL, Irving-Rodgers HF, Lavranos TC, Irvine CM, Krupa M. Roles of extracellular matrix in follicular development. J Reprod Fertil Suppl 1999; 54: 343-52.
Romero-Arredondo A, Seidel GE, Jr. Effects of bovine follicular fluid on maturation of bovine oocytes. Theriogenology 1994; 41: 383-394.
Rose-Hellekant TA, Bavister BD. Roles of protein kinase A and C in spontaneous maturation and in forskolin or 3-isobutyl-1-methylxanthine maintained meiotic arrest of bovine oocytes. Mol Reprod Dev 1996; 44: 241-9.
Ross EM, Gilman AG. Biochemical properties of hormone sensitive adenylate cylase. Annu Rev Biochem 1980; 49: 533-64.
Rouillier P, Matton P, Dufour M, Sirard MA, Guilbault LA. Steroid production, cell proliferation, and apoptosis in cultured bovine antral and mural granulosa cells: development of an in vitro model to study estradiol production. Mol Reprod Dev 1998; 50: 170-7.
Rouillier P, Matton P, Sirard MA, Guilbault LA. Follicle-stimulating hormone-induced estradiol and progesterone production by bovine antral and mural granulosa cells cultured in vitro in a completely defined medium. J Anim Sci 1996; 74: 3012-9.
Rouillier P, Saumande J, Sirard MA, Matton P, Guilbault LA. Comparison of the FSH-induced estradiol-17-beta production by bovine antral and mural granulosa cells cultured in vitro in a completely defined medium. Theriogenology 1994; 41: 286.
Rüsse I. Oogenesis in cattle and sheep. Bibl Anat 1983; 24: 77-92. Saeki K, Nagao Y, Kishi M, Nagai M. Developmental capacity of bovine oocytes following
inhibition of meiotic resumption by cycloheximide or 6-dimethylaminopurine. Theriogenology 1997; 48: 1161-1172.
Saeki K, Nagao Y, Kishi M, Nagai M, Iritani A. Timing of completion of the first meiotic division in bovine oocytes after maintenance of meiotic arrest with cycloheximide and their subsequent development. J Vet Med Sci 1998; 60: 523-526.
Sagata N, Watanabe N, Vande Woude GF, Ikawa Y. The c-mos proto-oncogene product is a cytostatic factor responsible for meiotic arrest in vertebrate eggs. Nature 1989; 342: 512-8.
Saito E, Ishibashi T. Meiotic arresting action of the substance obtained from cell surface of porcine ovarian granulosa cells. Jap J Zootech Sci 1977; 48: 22-6.
Sato E, Ishibashi T. Meiotic arresting action of the substance obtained from cell surface of porcine ovarian granulosa cells. Jpn J Zootech Sci 1977; 48: 22-6.
Sato E, Koide SS. A factor from bovine granulosa cells preventing oocyte maturation. Differentiation 1984; 26: 59-62.
Seamon KB, Padgett W, Daly JW. Forskolin: unique diterpene activator of adenylate cyclase in membrane and intact cells. Proc Natl Acad Sci U S A 1981; 78: 3363-7.
Senger PL. Pathways to pregnancy and parturition. Pullman, WA: Current Conceptions, Inc, 1997.
Shamsuddin M, Larsson B, Rodriguez-Martinez H. Maturation-related changes in bovine oocytes under different culture conditions. Anim Reprod Sci 1993; 31: 49-60.
Shim C, Lee DK, Lee CC, Cho WK, Kim K. Inhibitory effect of purines in meiotic maturation of denuded mouse oocytes. Mol Reprod Dev 1992; 31: 280-6.
Simili M, Pellerano P, Pigullo S, Tavosanis G, Ottaggio L, Saint-Georges L, et al. 6-DMAP inhibition of early cell cycle events and induction of mitotic abnormalities. Mutagenesis 1997; 12: 313-9.

64
Simon M, Jilek F, Fulka J, Jr. Effect of cycloheximide upon maturation of bovine oocytes. Reprod Nutr Dev 1989; 29: 533-40.
Sirard MA. Temporary inhibition of in vitro meiotic resumption by adenylate cyclase stimulation in immature bovine oocytes. Theriogenology 1989; 31: 257.
Sirard MA. Temporary inhibition of meiosis resumption in vitro by adenylate cyclase stimulation in immature bovine oocytes. Theriogenology 1990; 33: 757-67.
Sirard MA. Developmental capacity acquisition requires exposure of uncondensed chromosomes before maturation of bovine oocytes. Bull Assoc Anat (Nancy) 1991; 75: 93-98.
Sirard MA, Bilodeau S. Effects of granulosa cell co-culture on in-vitro meiotic resumption of bovine oocytes. J Reprod Fertil 1990a; 89: 459-65.
Sirard MA, Bilodeau S. Granulosa cells inhibit the resumption of meiosis in bovine oocytes in vitro. Biol Reprod 1990b; 43: 777-83.
Sirard MA, Blondin P. Oocyte maturation and IVF in cattle. Animal Reproduction Science 1996; 42: 417-426.
Sirard MA, Coenen K. The co-culture of cumulus-enclosed bovine oocytes and hemisections of follicles: Effects on meiotic resumption. Theriogenology 1993; 40: 933-942.
Sirard MA, Coenen K. The effect of hormones during in vitro meiotic inhibition with cycloheximide on subsequent development of bovine oocytes. Biol Reprod 1994; 50: 361.
Sirard MA, Coenen K, Bilodeau S. Effect of fresh or cultured follicular fractions on meiotic resumption in bovine oocytes. Theriogenology 1992; 37: 39-58.
Sirard MA, First NL. In vitro inhibition of oocyte nuclear maturation in the bovine. Biol Reprod 1988; 39: 229-34.
Sirard MA, Florman HM, Leibfried-Rutledge ML, Barnes FL, Sims ML, First NL. Timing of nuclear progression and protein synthesis necessary for meiotic maturation of bovine oocytes. Biol Reprod 1989; 40: 1257-1263.
Sirard MA, Lambert RD. Birth of calves after in vitro fertilization using laparoscopy and rabbit oviduct incubation of zygotes. Vet Rec 1986; 119: 167-9.
Sirard MA, Parrish JJ, Ware CB, Leibfried-Rutledge ML, First NL. The culture of bovine oocytes to obtain developmentally competent embryos. Biol Reprod 1988; 39: 546-52.
Sirard MA, Richard F, Mayes M. Controlling meiotic resumption in bovine oocytes: A review. Theriogenology 1998; 49: 483-497.
Sirard MA, Roy F, Mermillod P, Guilbault LA. The origin of follicular fluid added to the media during bovine IVM influences embryonic development. Theriogenology 1995; 44: 85-94.
Sirois J, Fortune JE. Ovarian follicular dynamics during the estrous cycle in heifers monitored by real-time ultrasonography. Biol Reprod 1988; 39: 308-17.
Soderling SH, Beavo JA. Regulation of cAMP and cGMP signaling: new phosphodiesterases and new functions. Curr Opin Cell Biol 2000; 12: 174-9.
Solomon MJ, Glotzer M, Lee TH, Philippe M, Kirschner MW. Cyclin activation of p34cdc2. Cell 1990; 63: 1013-1024.
Sorensen RA, Wassarman PM. Relationship between growth and meiotic maturation of the mouse oocyte. Dev Biol 1976; 50: 531-6.
Spicer LJ, Echternkamp SE. Ovarian follicular growth, function and turnover in cattle: a review. J Anim Sci 1986; 62: 428-51.
Sreenan J. In vitro maturation and attempted fertilization of cattle follicular oocytes. J Agric Sci Camb 1970; 75: 393-96.

65
Stone SL, Pomerantz SH, Schwartz-Kripner A, Channing CP. Inhibitor of oocyte maturation from porcine follicular fluid: further purification and evidence for reversible action. Biol Reprod 1978; 19: 585-92.
Sun FJ, Holm P, Irvine B, Seamark RF. Effect of sheep and human follicular fluid on the maturation of sheep oocytes in vitro. Theriogenology 1994; 41: 981-88.
Suntherland SJ, Crowe MA, Boland M, Roche JF. Selection, dominance and atresia of follicles during the oestrous cycle of heifers. J Reprod Dev 1994; 101: 547-55.
Sutovsky P, Flechon JE, Flechon B, Motlik J, Peynot N, Chesne P, et al. Dynamic changes of gap junctions and cytoskeleton during in vitro culture of cattle oocyte cumulus complexes. Biol Reprod 1993; 49: 1277-87.
Swinnen JV, Joseph DR, Conti M. The mRNA encoding a high-affinity cAMP phosphodiesterase is regulated by hormones and cAMP. Proc Natl Acad Sci U S A 1989; 86: 8197-201.
Szollosi D. Development of cortical granules and the cortical reaction in rat and hamster eggs. Anat Rec 1967; 159: 431-46.
Szollosi D. Maturation de l`ovocyte. Paris: Ministère de la Recherche et de la Technologie (DIST), 1991.
Szollosi D, Calarco P, Donahue R. The nuclear envelope: its breakdown and fate in mammalian oogonia and oocytes. Anat Rec 1972; 174.
Szybek K. In vitro maturation of oocyte from sexually inmmature mice. J Endocrinol 1972; 54: 527-28.
Taira M, Hockman SC, Calvo JC, Belfrage P, Manganiello VC. Molecular cloning of the rat adipocyte hormone-sensitive cyclic GMP- inhibited cyclic nucleotide phosphodiesterase. J Biol Chem 1993; 268: 18573-9.
Takahashi M, Koide SS, Donahoe PK. Mullerian inhibiting substance as oocyte meiosis inhibitor. Mol Cell Endocrinol 1986; 47: 225-34.
Tan SJ, Lu KH. Effects of different oestrous stages of ovaries and sizes of follicles on generation of bovine embryos in vitro. Theriogenology 1990; 33: 335.
Tatemoto H, Horiuchi T. Requirement for protein synthesis during the onset of meiosis in bovine oocytes and its involvement in the autocatalytic amplification of maturation-promoting factor. Mol Reprod Dev 1995; 41: 47-53.
Tatemoto H, Horiuchi T, Terada T. Effects of cycloheximide on chromatin condensations and germinal vesicle breakdown (GVBD) of cumulus-enclosed and denuded oocytes in cattle. Theriogenology 1994; 42: 1141-1148.
Tatemoto H, Terada T. Time-dependent effects of cycloheximide and alpha-amanitin on meiotic resumption and progression in bovine follicular oocytes. Theriogenology 1995; 43: 1107-1113.
Tatemoto H, Terada T. Involvement of cumulus cells stimulated by FSH in chromatin condensation and the activation of maturation-promoting factor in bovine oocytes. Theriogenology 1998; 49: 1007-1020.
Tilly JL. Apoptosis and ovarian function. Reviews of Reproduction 1996; 1: 162-72. Tilly JL, Kowalski KI, Johnson AL, Hsueh AJW. Involvement of apoptosis in ovarian follicular
atresia and postovulatory regression. Endocrinology 1991; 129: 2799-2801. Tilly JL, Ratts VS. Biological and clinical importance of ovarian cell death. Contemporary
Obstetrics and Gynecology 1996; 41: 59-86.

66
Tsafriri A. Oocyte maturation in mammals. In: Jones E, editor. The vertebrate ovary. New York: Plenum Press, 1979: 409-42.
Tsafriri A, Braw RH. Experimental approaches to atresia in mammals. Oxford Reviews of Reproductive Biology 1984; 6: 226-65.
Tsafriri A, Channing CP. Influence of follicular maturation and culture conditions on the meiosis of pig oocytes in vitro. J Reprod Fertil 1975a; 43: 149-52.
Tsafriri A, Channing CP. An inhibitory influence of granulosa cells and follicular fluid upon porcine oocyte meiosis in vitro. Endocrinology 1975b; 96: 922-7.
Tsafriri A, Channing CP, Pomerantz SH, Lindner HR. Inhibition of maturation of isolated rat oocytes by porcine follicular fluid. J Endocrinol 1977; 75: 285-91.
Tsafriri A, Chun SY, Zhang R, Hsueh AJ, Conti M. Oocyte maturation involves compartmentalization and opposing changes of cAMP levels in follicular somatic and germ cells: studies using selective phosphodiesterase inhibitors. Dev Biol 1996; 178: 393-402.
Tsafriri A, Dekel N, Bar-Ami S. The role of oocyte maturation inhibition in follicular regulation of oocyte maturation. Journal of Reproduction and Fertility 1982; 64: 541-551.
Tsafriri A, Pomerantz SH, Channing CP. Inhibition of oocyte maturation by porcine follicular fluid: Partial characterization of the inhibitor. Biol Reprod 1976; 14: 511-516.
van de Leemput EE, Vos PL, Zeinstra EC, Bevers MM, van der Weijden GC, Dieleman SJ. Improved in vitro embryo development using in vivo matured oocytes from heifers superovulated with a controlled preovulatory LH surge. Theriogenology 1999; 52: 335-49.
Van Den Hurk R, Bevers MM, Beckers JF. In vivo and in vitro development of preantral follicles. Theriogenology 1997; 47: 73-82.
van Tol HT, Bevers MM. Theca cells and theca-cell conditioned medium inhibit the progression of FSH-induced meiosis of bovine oocytes surrounded by cumulus cells connected to membrana granulosa. Mol Reprod Dev 1998; 51: 315-21.
van Tol HTA, Vaneijk MJT, Mummery CL, Vandenhurk R, Bevers MM. Influence of FSH and hCG on the resumption of meiosis of bovine oocytes surrounded by cumulus cells connected to membrana granulosa. Mol Reprod Dev 1996; 45: 218-224.
van Wezel IL, Rodgers RJ. Morphological characterization of bovine follicles and their environment in vivo. Biol Reprod 1996; 55: 1003-11.
Vasquez D. Inhibitors of protein synthesis. FEBS Lett 1974; 40: S63-S64. Vatzias G, Hagen DR. Effects of follicular fluid aspirated from three follicular sizes on in vitro
maturation of porcine oocytes. Biol Reprod 1994; 50: 168. Verde F, Dogterom M, Stelzer E, Karsenti E, Leibler S. Control of microtubule and chromatin
behavior follow MAP kinase activity but not MPF activity during meiosis in mouse oocytes. Development 1992; 120: 1017-25.
Weakly BS. Electron microscopy of the oocyte and granulosa cells in the developing ovarian follicles of the golden hamster. J. Anat 1966; 100: 503-34.
Webb R, Gong JG, Law AS, Rusbridge SM. Control of ovarian function in cattle. J Reprod Fertil (Suppl) 1992; 45: 141-56.
Wehrend A, Meinecke B. Kinetics of meiotic progression, M-phase promoting factor (MPF) and mitogen-activated protein kinase (MAP kinase) activities during in vitro maturation of

67
porcine and bovine oocytes: species specific differences in the length of the meiotic stages. Anim Reprod Sci 2001; 66: 175-84.
Whitaker M. Control of meiotic arrest. Rev Reprod 1996; 1: 127-35. Wickramasinghe D, Albertini DF. Cell cycle control during mammalian oogenesis. Curr Top Dev
Biol 1993; 28: 125-53. Wise T, Maurer RR. Follicular development, oocyte viability and recovery in relation to
follicular steroids, prolactin and glycosaminoglycans throughout the estrous period in superovulated heifers with a normal LH surge, no detectable LH surge, and progestin inhibition of LH surge. Domest Anim Endocrinol 1994; 11: 35-58.
Wise T, Suss U, Stranzinger G, Wuthrich K, Maurer RR. Cumulus and oocyte maturation and in vitro and in vivo fertilization of oocytes in relation to follicular steroids, prolactin, and glycosaminoglycans throughout the estrous period in superovulated heifers with a normal LH surge, no detectable LH surge, and progestin inhibition of LH surge. Domest Anim Endocrinol 1994; 11: 59-86.
Witschi E. Migration of the germ cells of human embryos from the yolk sac to the primitive gonadal fold. Contrib Embryol 1948; 32: 67-80.
Wu B, Ignotz G, Currie WB, Yang X. Dynamics of maturation-promoting factor and its constituent proteins during in vitro maturation of bovine oocytes. Biol Reprod 1997a; 56: 253-9.
Wu B, Ignotz G, Currie WB, Yang X. Expression of Mos proto-oncoprotein in bovine oocytes during maturation in vitro. Biol Reprod 1997b; 56: 260-5.
Wu B, Ignotz GG, Currie WB, Yang X. Temporal distinctions in the synthesis and accumulation of proteins by oocytes and cumulus cells during maturation in vitro of bovine oocytes. Mol Reprod Dev 1996; 45: 560-5.
Xu Z, Garverick HA, Smith GW, Smith MF, Hamilton SA, Youngquist RS. Expression of follicle-stimulating hormone and luteinizing hormone receptor messenger ribonucleic acids in bovine follicles during the first follicular wave. Biol Reprod 1995; 53: 951-7.
Yamada O, Abe M, Takehana K, Hiraga T, Iwasa K, Hiratsuka T. Microvascular changes during the development of follicles in bovine ovaries: a study of corrosion casts by scanning electron microscopy. Arch Histol Cytol 1995; 58: 567-74.
Yang LS, Kadam AL, Koide SS. Identification of a cAMP-dependent protein kinase in bovine and human follicular fluids. Biochem Mol Biol Int 1993; 31: 521-5.
Yang X, Kubota C, Suzuki H, Taneja M, Bols PE, Presicce GA. Control of oocyte maturation in cows--biological factors. Theriogenology 1998; 49: 471-82.
Zamboni L. Fine morphology of the follicle wall and follicle cell-oocyte association. Biol Reprod 1974; 10: 125-49.
Zandomeni R, Bunick D, Ackerman S, Mittleman B, Weinmann R. Mechanism of action of DRB. III Effect on specific in vitro initiation of transcription. J Mol Biol 1983; 167: 561-74.
Zhang L, Jiang S, Wozniak PJ, Yang X, Godke RA. Cumulus cell function during bovine oocyte maturation, fertilization, and embryo development in vitro. Mol Reprod Dev 1995; 40: 338-44.

CHAPTER II The Influence of Cumulus-Oocyte Complex Morphology and Meiotic Inhibitors on the Kinetics of Nuclear Maturation in Cattle.
Cet article a été publié dans la revue “Theriogenology ” sous la référence suivante: M. A. Mayes
and M. A. Sirard. The influence of cumulus-oocyte complex morphology and meiotic inhibitors
on the kinetics of nuclear maturation in cattle. Theriogenology 2001; 55:911-22.

68
RÉSUMÉ Cette étude évaluait si les classes morphologiques préétablies de complexes ovocyte-cumulus
(COCs) bovins différaient dans leur cinétique de la reprise de la méiose après 4 h d’incubation et
si le facteur ‘’temps’’ de la reprise de la méiose des COCs différait après une période de maintien
de l’arrêt méiotique. Les COCs bovins ont été aspirés de follicules de 2- à 5- mm et classifiés
selon l’état de leur cytoplasme et des cellules de cumulus (classes 1 à 3 ). Les COCs ont été fixés
par groupe de 15 à 20 au temps 0h ou après avoir été incubés pour une période de 4 h. De plus,
les COCs évalués comme classe 1 ont été préalablement incubés pour 4 h sur une monocouche de
cellules de thèque ou en présence de 2 µg/mL de cycloheximide, rincés puis incubés en présence
de cycloheximide dans un milieu sans cellule de thèque pour encore 4 h. Les ovocytes ont été
fixés et évalués pour l’état de la maturation nucléaire. Les résultats démontrent qu’à 0h, les
COCs de la classe 3 avaient moins d’ovocytes au stade GV que les COCs des classes 1 et 2
(respectivement 69,3 ± 3,2 vs 88,8 ± 3.4% et 86,9% GV ± 4,3% ET; P < 0,05). Après 4 h
d’incubation, toutes les classes de COCs ont démontré une diminution significative quand ou
nombre de COCs au stade de GV. Les COCs maintenus en arrêt méiotique puis incubés pour 4 h
ont repris la méiose plus rapidement que les COCs incubés en présence de cycloheximide dans
un milieu sans cellules de thèque (19,4 ± 2,5 ; 33,3 ± 7,3 et 59,9 ± 6,5% GV ± ET,
respectivement). Les COCs de la classe 3 ont moins d’ovocytes au stade de GV au début de
l’incubation que les ovocytes des autres classes. Le nombre de COCs au stade de GV après 4 h
d’incubation en présence de cycloheximide dans un milieu sans cellules de thèque n’est pas
significativement différent du nombre de COCs incubés en présence de la monocouche de
cellules de thèque pour 24 h (58,8 ± 6,5 vs 56,4 ± 6,4% GV ± ET; respectivement). Nos résultats
indiquent que l’habileté des cellules de thèque à maintenir les ovocytes au stade de GV peut être
limité aux ovocytes qui n’étaient pas engagés ou stimulés in vivo à reprendre la méiose, tel
qu’indiqué par leur cinétique de maturation plus rapide.

69
ABSTRACT This study evaluated whether pre-established morphological classes of bovine cumulus oocyte
complex (COCs) differ in their kinetics of meiosis resumption after 4 h of incubation and
whether the timing of COCs resumption of meiosis differed after a period of maintained meiotic
arrest. Bovine COCs were aspirated from 2- to 5- mm follicles and classified according to the
state of their cumulus cells and cytoplasm (Classes 1 to 3). Groups of 15 to 20 COCs were fixed
at 0 h or after an incubation period of 4 h. In addition, COCs from Class 1 were first incubated
for 4 h on a theca cell monolayer or in the presence of 2 µg/ml of cycloheximide, rinsed and then
incubated in cycloheximide and theca cell-free medium for another 4 h. Oocytes then were fixed
and evaluated for state of nuclear maturation. Results show that at 0 h, COCs from Class 3 have
fewer oocytes at the GV stage than COCs from Class 1 and Class 2 (respectively 69.3 ± 3.2 vs
88.8 ± 3.4% and 86.9% GV ± 4.3% SEM; P < 0.05). After 4 h of incubation, all COCs classes
show a significant decrease in the number of COCs at the GV stage. The COCs maintained in
meiotic arrest and then incubated for 4 h resume meiosis faster than COCs incubated in
cycloheximide and theca cell-free medium (19.4 ± 2.5, 33.3 ± 7.3 and 59.9 ± 6.5% GV SEM,
respectively). The COCs of Class 3 have fewer oocytes at the GV stage at the beginning of
incubation than all other classes. The number of COCs at the GV stage after 4 h of incubation in
cycloheximide and theca cell-free medium is not significantly different than those COCs
incubated in the presence of theca cell monolayers for 24 h (58.8 ± 6.5 vs. 56.4 ± 6.4% SEM,
respectively). Our results indicate that the ability of theca cells to maintain oocytes at the GV
stage could be limited to those oocytes that were not committed or primed in vivo to resume
maturation as indicated by their faster maturation kinetics.
INTRODUCTION This study evaluated whether pre-established morphological classes of bovine cumulus oocyte
complex (COCs) differ in their kinetics of meiosis resumption after 4 h of incubation and
whether the timing of COCs resumption of meiosis differed after a period of maintained meiotic
arrest. Bovine COCs were aspirated from 2- to 5- mm follicles and classified according to the
state of their cumulus cells and cytoplasm (Classes 1 to 3). Groups of 15 to 20 COCs were fixed

at 0 h or after an incubation period of 4 h. In addition, COCs from Class 1 were first incubated
for 4 h on a theca cell monolayer or in the presence of 2 µg/ml of cycloheximide, rinsed and then
incubated in cycloheximide and theca cell-free medium for another 4 h. Oocytes then were fixed
and evaluated for state of nuclear maturation. Results show that at 0 h, COCs from Class 3 have
fewer oocytes at the GV stage than COCs from Class 1 and Class 2 (respectively 69.3 ± 3.2 vs
88.8 ± 3.4% and 86.9% GV ± 4.3% SEM; P < 0.05). After 4 h of incubation, all COCs classes
show a significant decrease in the number of COCs at the GV stage. The COCs maintained in
meiotic arrest and then incubated for 4 h resume meiosis faster than COCs incubated in
cycloheximide and theca cell-free medium (19.4 ± 2.5, 33.3 ± 7.3 and 59.9 ± 6.5% GV SEM,
respectively). The COCs of Class 3 have fewer oocytes at the GV stage at the beginning of
incubation than all other classes. The number of COCs at the GV stage after 4 h of incubation in
cycloheximide and theca cell-free medium is not significantly different than those COCs
incubated in the presence of theca cell monolayers for 24 h (58.8 ± 6.5 vs. 56.4 ± 6.4% SEM,
respectively). Our results indicate that the ability of theca cells to maintain oocytes at the GV
stage could be limited to those oocytes that were not committed or primed in vivo to resume
maturation as indicated by their faster maturation kinetics.
INTRODUCTION In vivo, bovine oocytes resume meiosis after the pre-ovulatory LH peak, while resumption of
meiosis in vitro occurs spontaneously when the cumulus-oocyte complex (COCs) is removed
from the follicle and incubated under suitable conditions [1]. During resumption of meiosis, the
oocyte nucleus (known as germinal vesicle (GV)) undergoes germinal vesicle breakdown
(GVBD), which involves a gradual chromatin condensation, disappearance of the nucleolus and
disintegration of the nuclear membrane [2]. The oocyte then progresses through the Metaphase I,
Anaphase I and Telophase I stages. After the extrusion of the first polar body, the oocyte
becomes arrested at Metaphase II until reactivated at fertilization.
De novo protein synthesis is required before GVBD takes place in species such as the bovine [3],
porcine [4], caprine [5], ovine [6] and Xenopus [7]. Oocytes from these species can be reversibly
maintained in a state of meiotic arrest by culturing them in the presence of a protein synthesis

inhibitor such as cycloheximide [8, 9]. Cycloheximide is an inhibitor of peptidyl transferase,
which prevents resumption of meiosis by blocking the synthesis of stage-specific proteins [10].
The effects of cycloheximide on oocyte maturation are reversible, as shown by the birth of live
calves after the maturation and fertilization of oocytes previously treated with this protein
synthesis inhibitor [11, 12]. Addition of cycloheximide to the incubation medium within 6 h
from the onset of IVM prevents bovine oocytes from resuming meiosis [13].
Bovine oocytes co-incubated with confluent theca cell monolayers are prevented from reaching
the GVBD. This inhibitory effect of theca cells on resumption of meiosis is reversible upon
incubation of oocytes in theca cell-free medium [14-16].
Techniques used for the in vitro production of bovine embryos take advantage of the large
number of oocytes harvested from ovaries collected at the slaughterhouse. However, the in vitro
production of blastocysts is limited to approximately 35 to 40% of all the oocytes harvested [17].
Numerous evaluation methods were applied to select those oocytes that yield the highest
percentages of blastocysts after fertilization. These methods allocated COCs into specific
categories and evaluated the percentages of blastocyst after fertilization to explain how the
morphology of the COCs correlates with the developmental competence of the oocyte [18-20].
Traditionally, COCs with a homogeneous cytoplasm and multiple compact layers of cumulus
cells were considered to be developmentally competent, while COCs showing signs of atresia or
granulations of the cytoplasm were normally discarded. Recent work in our laboratory correlated
the visual appearance of bovine COCs and their follicle of origin with the oocyte`s
developmental competence [21]. In that study, COCs were allocated to six different classes
based on the aspect of the cytoplasm and the surrounding cumulus cells. Classes 1 and 2 include
COCs with multiple layers of compact cumulus cells and a homogeneous or slightly granulated
cytoplasm, while Class 3 includes those oocytes with a homogeneous cytoplasm and slightly
expanded outer layers of cumulus cells. Classes 4 to 6 include respectively COCs with fully
expanded cumulus cells, only corona radiata or denuded oocytes [21]. Classes 1 and 2 originate
from "healthy" follicles, while Classes 3 to 6 originate from follicles with obvious signs of
atresia. The Class 3 COCs, which originate from slightly atretic follicles, showed the highest
percentage of development to the morula stage.

Recent experiments in our laboratory showed that adequate pretreatment of the cow before
oocyte aspiration and culture in vitro significantly enhanced the developmental capacity of the
oocytes [22]. To recreate this follicle effect in vivo with oocytes coming from nontreated cows
we would need a culture system, that could maintain meiotic arrest and gene transcription while
the right follicular signal (once found) could be applied.
Immature bovine oocytes used in laboratory studies are usually obtained from ovaries collected
post-mortem. These oocytes are extremely heterogeneous in terms of quality, meiotic and
developmental competence. Therefore, it is important to define the parameters used to select
oocytes for in vitro maturation and the effects of manipulating oocyte maturation in vitro when
studies on meiotic arrest of bovine oocytes. The kinetics of meiotic resumption could indicate
either the beginning of pseudo-atresia or the ability of the oocyte to develop further. Therefore
this article is based on the hypothesis that the morphology of the bovine cumulus-oocyte complex
has an impact on the resumption of meiosis and the effectiveness of meiotic inhibitors. In this
study, we evaluated how pre-established classes of bovine COCs differ in their ability to resume
meiosis after 4 h of incubation and whether the timing of COC meiotic resumption differed after
a 4 h period of meiotic arrest maintained by cycloheximide or co-incubation with theca cell
monolayers.
MATERIALS AND METHODS
Preparation of Theca Cell Monolayers Bovine theca cell monolayers were prepared as previously described [14]. Bovine follicles (2 to
5 mm) were chosen randomly from ovaries that were kept on ice in bags containing gauze soaked
with saline solution. The follicles were isolated from ovaries and dissected free of stromal tissue
with scissors. Theca cells were harvested exclusively from transparent follicles containing at
least 75% of the granulosa layer intact and unexpanded COCs [14, 23, 24]. Selected follicles
(n=12) were placed in a calcium- and magnesium-free Hanks’ balanced salt solution and cut into
hemisections with a scalpel. Granulosa cells and COCs were removed by scraping the
hemisections with a fine glass loop made from a Pasteur pipette. This technique is used to
effectively isolate theca cell layers. The theca layers were enzymatically digested in 15 mL of

modified Hanks’ solution supplemented with 2125 U of collagenase Type II, 1600 U trypsin
from porcine pancreas, 1 mg DNAse Type I and 2 mg EDTA (all from Sigma Company, St.
Louis, MO). Enzymatic digestion took place for 1 h at 38.5ºC in an atmosphere of 95% air and
5% CO2. Theca layers were passed through a 10-mL pipette every 20 min. After incubation,
larger pieces of remaining follicular hemisections were withdrawn from the solution. The
reaction was stopped by the addition of 4 mL of washing medium, consisting of HEPES-buffered
Tyrode’s medium (TLH, pH 7.4) [25] supplemented with 10% fetal calf serum (heat-treated FCS
from MEDICORP, Montreal, PQ, Canada), 0.2 mM pyruvic acid and 50 µg/mL gentamicin
sulfate. Cells then were centrifuged (10 min at 2000 g), resuspended and washed a second time.
The first and second pellets were resuspended in 10 mL and 1 mL, respectively, of washing
medium. Theca cells were counted with a hemocytometer and seeded in a 24-well plate (Falcon
Becton Dickinson, Rutherford, NJ) at a concentration of 1.5 to 2.0 x 105 cells/mL. The culture
medium, consisting of TCM 199 with Earle’s salts (Gibco Laboratories, Grand Island, NY) and
bicarbonate (Sigma), 10% FCS, 0.2 mM pyruvic acid and 50 µg/mL gentamicin sulfate, was pre-
exposed to culture conditions (38.5ºC, 5% CO2:95% air atmosphere with 100% humidity) for at
least 2 h. The culture medium was replenished every 48 h, and follicular cells were cultured for 5
to 7 days. When the monolayers were confluent, the medium was replenished and conditioned
for 48 h.
Oocyte Collection Bovine ovaries at various stages of their reproductive cycle were collected at the slaughterhouse
and transported to the laboratory in saline solution kept between 30 and 35ºC. The saline
solution consisted of 0.9% NaCl (w:v supplemented with 100,000 IU penicillin, 100 mg
streptomycin and 250 µg amphotericin B per liter (Sigma)). The contents of 1- to 5-mm follicles
were aspirated using a 10 mL syringe and an 18-gauge needle. The follicular contents were
pooled in 50 mL conical tubes. After sedimentation, COCs were recovered with the use of a
stereomicroscope.

Oocyte Classification Recovered COCs were allocated to different groups depending on the status of the cumulus cells
and the homogeneity of the cytoplasm. The classification of bovine COCs according to the status
of the cytoplasm and number of cumulus layers was reported previously [21]. For these
experiments, the first group of COCs had at least five layers of compact cumulus cells and a
cytoplasm that was either homogeneous or showed a dark zone around the periphery (Class 1).
The second group of COCs showed a slight expansion of the outer layers of cumulus cells and
slight granulation of the cytoplasm (Class 2). The third group consisted of COCs with atretic or
incomplete cumulus (Class 3). Once classified, COCs then were rinsed twice in the supernatant
of follicular fluid (centrifuged for 10 min at 325 g at 25ºC) and allocated to their respective
treatments.
Experimental Design A total of 990 oocytes (6 repetitions) were selected and then allocated in groups of 15 to 20 to
specific treatments. All treatments were represented in each replicate. The COCs from Classes
1, 2 and 3 were denuded and fixed immediately to evaluate the percentage of oocytes arrested at
the GV stage at time zero. Alternatively, COCs from each group were allowed to mature for 4 h
by incubating them in 1 mL of TCM 199 medium supplemented with 10% FCS, 0.2 mM pyruvic
acid and 50 µg/mL gentamicin sulfate (cycloheximide (CX) and theca cell-free medium). The
medium was allowed to equilibrate to incubation conditions for at least 2 h before beginning the
treatments. At the end of the treatment, the COCs were denuded and fixed for evaluation of
nuclear status. In addition, COCs from Class 1 were incubated for 4 h under conditions that
reversibly prevent GVBD (culture medium supplemented with 2 µg/mL of cycloheximide or
incubation with a theca cell monolayer) and then rinsed twice for 5 min with CX and theca cell-
free medium. The COCs were then incubated for a second period of 4 h in CX and theca cell-
free medium. At the end of treatment, COCs were denuded and fixed. An additional treatment
consisted of COCs from Class 1 incubated with theca cell monolayers for 24 h.

Oocyte Fixation At the end of each treatment, COCs were transferred into small centrifuge tubes containing 500
µL of TLH and were vortex-agitated for 7 min to remove the cumulus cells. Completely denuded
oocytes were recovered under a stereomicroscope and were transferred onto a glass slide in a
small drop of medium. The coverslip was kept in contact with the oocyte with a mix of Vaseline
and paraffin wax. The coverslip was fixed in place with epoxy glue and the slides were
immersed in fixative (ethanol: acetic acid, 3:1) for a minimum of 24 h. Oocytes then were
stained with 1% aceto-orcein and were examined for nuclear morphology with a phase contrast
microscope at x100 and x400 magnification [26].
Statistical Analysis Oocytes with a nuclear membrane present were classified as GV stage, while those without a
nuclear membrane were classified as having undergone germinal vesicle breakdown (GVBD) and
resumption of meiosis. Results are expressed as means ± standard error (SEM). Data were
analyzed by a two-way ANOVA. When ANOVA indicated significant treatment effect (P <
0.05), treatment differences were compared by Duncan’s multiple range test.
RESULTS Evaluation of the status of nuclear maturation of COCs shows that Classes 1 and 2 have similar
proportions of COCs at the GV stage at time zero (88.8 ± 3.4% and 86.9 ± 4.3% SEM,
respectively), while COCs Class 3 already show a significantly lower proportion of COCs at the
GV stage (69.3 ± 3.2%). After 4 h of incubation, COCs from Classes 1 and 2 still have higher
numbers of COCs at the GV stage (59.8 ± 6.5% and 79.9 ± 6.0%, respectively) than COCs of
Classes 3 (42.4 ± 3.2%). Regardless of classification, all COCs classes show decreased
percentages of COCs at the GV stage after 4 h of incubation (Figure 2.1).
The COCs of Class 1 also were evaluated to determine if a period of meiotic arrest maintained
during 4 h with 2 µg/mL cycloheximide or incubation with theca cell monolayers had an effect
on the rate of resumption of meiosis. After a 4 h incubation in CX and theca cell-free medium,
more COCs of Class 1 (59.9% GV ± 6.5% SEM) remained in meiotic arrest than those COCs

treated with cycloheximide or incubated with theca cells (19.4% GV ± 2.5% SEM and 33.3% GV
± 7.3% SEM, respectively) (Figure 2.2).
The number of COCs at the GV stage after an incubation of 4 h in CX and theca cell-free
medium was not significantly different from the number of COCs incubation with theca cells for
24 h (58.8% ± 6.5 vs. 56.4 ± 6.4% SEM, respectively) (Figure 2.3).
DISCUSSION Previous work by our group suggested that the morphological evidence in the COCs of the early
process of follicular atresia might enhance the oocyte`s developmental competence [21]. This
study shows that the number of COCs at the GV stage at 0 h is also correlated with the COC
morphology. There is no significant difference in the number of oocytes at the GV stage found in
"healthy" COCs (Class 1) or slightly atretic COCs (Class 2) at 0 h. However, the proportion of
COCs showing advanced stages of atresia with very few or no surrounding cumulus cells (Class
3) at the GV stage at the beginning of the incubation is significantly lower. This trend is
maintained even after an incubation of 4 h, when a larger percentage of COCs from Class 1 are
still at the GV stage when compared to Class 3. In fact, COCs class 3 at 0 h have comparable
percentages of COCs at the GV stage than classes 1 and 2 after 4 h of incubation. This strongly
suggests that the most atretic oocytes have already started the process of resumption of meiosis
before the onset of incubation. These data provide further support that the control of meiotic
arrest is severely impaired in follicles undergoing atresia maybe due to decreased membrane
integrity.
Similar observations were reported for equine oocytes [27, 28]. In vitro maturation of horse
oocytes showed that a higher proportion of oocytes with expanded cumuli mature in vitro
compared to oocytes with compact cumuli. The expansion of the cumuli was negatively
correlated with the health of the follicle. Oocytes with expanded cumuli are considered to
originate from atretic follicles, while oocytes with compact cumuli come from "healthy" follicles.
Horse oocytes with expanded cumuli appear to have greater meiotic competence than oocytes
from "healthy" follicles. This might result from a loss of suppressive activity by the degenerating
follicle on the oocyte’s nuclear maturation [27]. Changes in the condensation of chromatin at the

GV stage has been correlated with meiotic competence of horse oocytes. These changes were
reported in the pre-ovulatory follicle, but may also occur during follicular atresia [28].
In humans, oocytes from atretic follicles also resume meiosis faster than oocytes from healthy
follicles [29]. In addition, "healthy follicles" contain oocytes arrested at the GV stage, while at
least 10% of atretic follicles show GVBD [30]. Similar trends are also observed in nonhuman
primates. Cynomolgus monkey oocytes recovered from atretic follicles resumed meiosis faster
than those oocytes collected from "healthy" follicles [31].
We also compared cycloheximide treatment with theca cell monolayer incubation with respect to
the frequency of resumption of meiosis of COCs of Class 1 after a second incubation period of 4
h. Overall, COCs exposed to cycloheximide or co-incubated with theca cell monolayers do not
show significant differences in the number of oocytes at the GV stage after the second 4 h
incubation. The effectiveness of cycloheximide as an inhibitor of resumption of meiosis has been
studied extensively [3, 11, 12]. Cycloheximide added at the beginning of the incubation period is
capable of blocking bovine COCs at the GV stage for at least 24 h. The percentages of COCs
maintained in meiotic arrest after 24 h is similar to the percentage of GV found at time zero [12].
Since it has been shown that bovine oocytes incubated in the presence of cycloheximide do not
resume meiosis, then the percentage of COCs found at the GV stage after an exposure to 4 h of
cycloheximide should not significantly differ from those found at the beginning of incubation.
Therefore, it is technically possible to compare COCs incubated in CX and theca cell-free
medium for 4 h with COCs incubated for a total incubation period of 8 h (4 h in repressed
resumption of meiosis and 4 h in CX and theca cell-free medium). Based on these assumptions
we can infer that treated COCs resume meiosis faster than untreated COCs incubated for 4 h.
Furthermore, there was no significant difference in the frequency of COCs at the GV stage
whether COCs were treated with cycloheximide or coincubated with theca cell monolayers after
an incubation of 4 h in CX and theca cell-free medium. Even though these treatments were not
significantly different, there is a trend suggesting that, after the second 4 h incubation period,
cycloheximide treated oocytes undergo GVBD faster than COCs incubated with theca cell
monolayers (Figure 2.2). The meiotic resumption of COCs following a short period of
maintained in meiotic arrest in vitro is accelerated. This suggests that a single treatment might

not be enough to prevent all the processes involved in meiotic resumption and further suggests
that multiple mechanisms are involved in the maintenance of meiotic arrest. It is probable that
during the period of artificial meiotic arrest, the oocyte still undergoes changes that allow a faster
meiotic resumption upon removal of the inhibitory factor.
Furthermore, the percentage of COCs at the GV stage after 4 h of incubation is not significantly
different than the percentage of COCs at the GV stage after 24 h incubation with theca cell
monolayers. This suggests that within these classes there is a subpopulation of oocytes that is
responsive to the inhibitory action of the theca cells, thus remaining at the GV stage. This further
suggests that either the inhibitory action of theca cell monolayers is not identical to the in vivo
conditions or that oocytes show a different response threshold to the inhibitory factors. A 4 h
interval from the time the ovaries are collected at the slaughterhouse to the time the COCs are
aspirated has a beneficial effect on the maturation of the COCs and the zygote’s developmental
competence [22]. This interval may induce a state similar to the early stages of atresia [21]. The
4 h interval and its positive effect on development might indicate that atresia begins during
transport of the ovaries, thus resembling the early events of the maturation process. In particular,
bovine oocytes need to synthesize cyclin B to resume meiosis [32]. The interval might also
induce the accumulation of a small amount of cyclin B, which may render a certain proportion of
the COCs population more susceptible to resume meiosis [32].
In conclusion, our study shows that more atretic or denuded COCs went through the early phase
of nuclear maturation after 4 h of incubation than "healthy" or slightly atretic COCs. We also
found that the inhibitory action of theca cells monolayers might be effective only on a specific
COC subpopulation. This new information is important to understand the heterogeneity observed
in oocyte quality from follicles of different health status.
ACKNOWLEGMENTS This work was supported by the Natural Sciences and Engineering Research Council of Canada
(NSERC) and Semex Canada. The authors gratefully acknowledge the assistance of Dr. Gisèle
LaPointe for her comments on the manuscript.

79
REFERENCES 1. Pincus, G. and E.V. Enzmann, The comparative behavior of mammalian eggs in vitro and
in vivo. J. Exp. Med., 1935. 62: p. 665-675. 2. Kubelka, M., et al., Time sequence of germinal vesicle breakdown in pig oocytes after
cycloheximide and p-aminobenzamidine block. Gam. Res., 1988. 19: p. 423-431. 3. Sirard, M.A., et al., Timing of nuclear progression and protein synthesis necessary for
meiotic maturation of bovine oocytes. Biol Reprod, 1989. 40: p. 1257-1263. 4. Fulka, J., Jr, et al., Effect of cycloheximide on nuclear maturation of pig and mouse
oocytes. J Reprod Fertil, 1986. 77: p. 281-285. 5. Le Gal, F., L. Gall, and V. De Smedt, Changes in protein synthesis pattern during in vitro
maturation of goat oocytes. Mol Reprod Dev, 1992. 32(1): p. 1-8. 6. Moor, R.M. and I.M. Crosby, Protein requirements for germinal vesicle breakdown in
ovine oocytes. J Embryol Exp Morphol, 1986. 94: p. 207-220. 7. Wasserman, W.J. and Y. Masui, Effects of cycloheximide on a cytoplasmic factor
initiating meiotic naturation in Xenopus oocytes. Exp Cell Res, 1975. 91(2): p. 381-8. 8. Hunter, A.G. and R.M. Moor, Stage dependent effects of inhibiting RNA and protein
synthesis on meiotic maturation of bovine oocytes in vitro. J Dairy Sci, 1987. 70: p. 1646-1651.
9. Sirard, M.A. and N.L. First, In vitro inhibition of oocyte nuclear maturation in the bovine. Biol Reprod, 1988. 39(2): p. 229-34.
10. Vasquez, D., Inhibitors of protein synthesis. FEBS Lett, 1974. 40: p. S63-S64. 11. Saeki, K., et al., Developmental capacity of bovine oocytes following inhibition of
meiotic resumption by cycloheximide or 6-dimethylaminopurine. Theriogenology, 1997. 48: p. 1161-1172.
12. Saeki, K., et al., Timing of completion of the first meiotic division in bovine oocytes after maintenance of meiotic arrest with cycloheximide and their subsequent development. J Vet Med Sci, 1998. 60(4): p. 523-526.
13. Tatemoto, H. and T. Terada, Time-dependent effects of cycloheximide and alpha-amanitin on meiotic resumption and progression in bovine follicular oocytes. Theriogenology, 1995. 43: p. 1107-1113.
14. Richard, F.J. and M.A. Sirard, Effects of follicular cells on oocyte maturation. II: Theca cell inhibition of bovine oocyte maturation in vitro. Biol Reprod, 1996. 54(1): p. 22-8.
15. Richard, F.J. and M.A. Sirard, Theca cells inhibit the spontaneous meiotic resumption in bovine oocytes. Biol Reprod, 1994. 50(Suppl. 1): p. 305.
16. Richard, F.J. and M.A. Sirard, Effects of follicular cells on oocyte maturation. I: Effects of follicular hemisections on bovine oocyte maturation in vitro. Biol Reprod, 1996. 54(1): p. 16-21.
17. Brackett, B.G. and K.A. Zuelke, Analysis of factors involved in the in vitro production of bovine embryos. Theriogenology, 1993. 39(1): p. 43-64.
18. De Loos, F.A., E. Zeinstra, and M.M. Bevers, Follicular wall maintains meiotic arrest in bovine oocytes cultured in vitro. Mol Reprod Dev, 1994. 39(2): p. 162-5.
19. Hawk, H.W. and R.J. Wall, Improved yields of bovine blastocyst from in vitro-produced oocytes. I. Selection of oocytes and zygotes. Theriogenology, 1994. 41: p. 1571-1583.

20. Hazeleger, N.L. and R.B. Stubbings, Developmental potential of selected bovine oocyte cumulus complexes. Theriogenology, 1992. 37: p. 219.
21. Blondin, P. and M.A. Sirard, Oocyte and follicular morphology as determining characteristics for developmental competence in bovine oocytes. Mol Reprod Dev, 1995. 41(1): p. 54-62.
22. Blondin, P., L.A. Guilbault, and M.A. Sirard, In vitro production of bovine embryos: Developmental competence is acquired before maturation. Theriogenology, 1995. 43(1): p. 168.
23. Carbonneau, G. and M.A. Sirard, Influence of follicular wall on meiotic resumption of bovine oocytes when cultured inside or outside hemisections. J Reprod Dev, 1994. 40: p. 125-132.
24. McNatty, K.P., et al., Some aspects of thecal and granulosa cell function during follicular development in the bovine ovary. J Reprod Fertil, 1984. 72(1): p. 39-53.
25. Bavister, B.D., M.L. Leibfried, and G. Lieberman, Development of preimplantation embryos of the golden hamster in a defined culture medium. Biol Reprod, 1983. 28(1): p. 235-47.
26. Motlik, J., H.H. Koefoed-Johnsen, and J. Fulka, Breakdown of the germinal vesicle in bovine oocytes cultivated in vitro. J Exp Zool, 1978. 205(3): p. 377-83.
27. Hinrichs, K. and K.A. Williams, Relationships among oocyte-cumulus morphology, follicular atresia, initial chromatin configuration, and oocyte meiotic competence in the horse. Biol Reprod, 1997. 57(2): p. 377-84.
28. Hinrichs, K., Cumulus expansion, chromatin configuration and meiotic competence in horse oocytes: a new hypothesis. Equine Vet J Suppl, 1997(25): p. 43-6.
29. Lefevre, B., A. Gougeon, and J. Testart, In-vitro oocyte maturation: some questions concerning the initiation and prevention of this process in humans. Human Reprod, 1987. 2: p. 495-497.
30. Gougeon, A. and J. Testart, Germinal vesicle breakdown in oocytes of human atretic follicles during the menstrual cycle. J Reprod Fertil, 1986. 78: p. 389-401.
31. Lefevre, B., et al., In vivo changes in oocyte germinal vesicle related to follicular quality and size at mid-follicular phase during stimulated cycles in the cynomolgus monkey. Reprod Nutr Dev, 1989. 29(5): p. 523-31.
32. Levesque, J.T. and M.A. Sirard, Resumption of meiosis is initiated by the accumulation of cyclin B in bovine oocytes. Biol Reprod, 1996. 55(6): p. 1427-1436.

81
Figure 2. 1 Nuclear status of bovine oocytes at 0 h after an incubation of 4 h.
Bovine COCs were either fixed at 0 hours or incubated for 4 hours and then fixed. Different letters indicate significant differences (P < 0.05) according to Duncan's multiple range test. Data shown were derived from 6 replicates, 990 oocytes.

82
Figure 2. 2 Nuclear status of bovine COCs of Class 1 after 0 or 4 h incubation in Cycloheximide (CX) and theca cell-free medium or 4 h of inhibited GVBD by theca cell monolayers or CX followed by another 4 h period of incubation in CX and theca cell-free medium.
The COCs were incubated with theca cells (PT) or 2 µg/ml cycloheximide (PC) for 4 h, then rinsed and incubated for another 4 h in CX and theca cell-free medium. The COCs were then fixed. As controls, COCs were fixed at 0 h or after 4 h of incubation. Different letters indicate significant differences (P < 0.05) according to Duncan`s multiple range test.
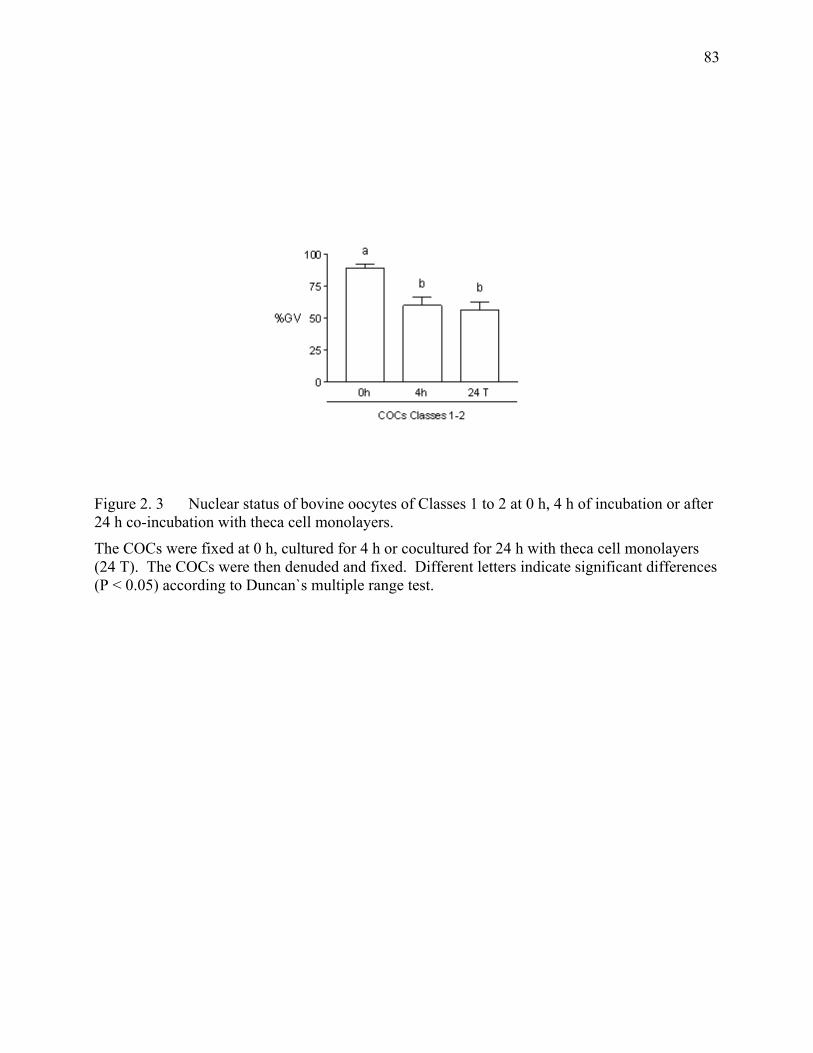
83
Figure 2. 3 Nuclear status of bovine oocytes of Classes 1 to 2 at 0 h, 4 h of incubation or after 24 h co-incubation with theca cell monolayers.
The COCs were fixed at 0 h, cultured for 4 h or cocultured for 24 h with theca cell monolayers (24 T). The COCs were then denuded and fixed. Different letters indicate significant differences (P < 0.05) according to Duncan`s multiple range test.

CHAPTER III Effect of Type 3 and Type 4 Phosphodiesterase Inhibitors on the Maintenance of Bovine Oocytes in Meiotic Arrest.
Cet article a été publié dans la revue “Biology of Reproduction ” sous la référence suivante:
M. A. Mayes and M. A. Sirard. Effect of Type 3 and Type 4 Phosphodiesterase Inhibitors on
the Maintenance of Bovine Oocytes in Meiotic Arrest. Biol Reprod 2002; 66:180-4.

85
RÉSUMÉ L’utilisation d’inhibiteurs à large spectre a d’abord suggéré que les phosphodiesterases
(PDEs) sont impliquées dans la maturation des ovocytes bovins. La modulation de chaque
famille de PDE est maintenant possible grâce à l’utilisation d’inhibiteurs spécifiques pour
chaque type de PDE nouvellement développés. Cette étude a évalué l`effet des inhibiteurs de
PDE spécifiques aux types 3 et 4 sur l’arrêt méiotique des complexes ovocyte-cumulus bovins
(COCs) et des ovocytes dénudés (ODs). L`influence des inhibiteurs en présence ou absence
d’une monocouche de cellules de thèque a aussi été évalué. Les COCs ont été aspirés
d’ovaires provenant de l’abattoir. Les ODs et les COCs ont été incubés 12 h dans un milieu
de culture en présence ou absence d’inhibiteurs de PDE de type 3 (10 et 20 µM cilostamide
ou 10 et 50 µM milirinone) ou de type 4 (10 et 50 µM rolipram). Les ovocytes ont alors été
fixés et classifiés selon le statut de leur maturation nucléaire. Les COCs ont par la suite été
coincubés avec une monocouche de cellules de thèque non traitées ou traitées avec des
différents inhibiteurs spécifiques de PDE. Les ODs et les COCs bovins incubés dans le
milieu de culture ont repris la méiose, mais l’ajout d’inhibiteurs de la PDE3 (cilostamide ou
milrinone) au milieu de culture a résulté en un arrêt méiotique. Par contre, l’ajout de rolipram
au milieu de culture n’a pas prévenu la maturation ovocytaire. De plus, les inhibiteurs de
PDE3, mais non ceux de PDE4, ont eu un effet additif sur l’action inhibitrice des
monocouches de cellules de thèque sur la maturation ovocytaire. Ces résultats supportent que
l’inhibition des PDE3 prévient la reprise de la méiose des ovocytes bovins, alors que
l’inhibition des PDE4 ne bloque pas la maturation ovocytaire même sous des conditions
inhibitrices normales. L’effet additif des inhibiteurs des PDE3 sur l’habileté des cellules de
thèque à maintenir les ovocytes bovins en arrêt méiotique suggère que les PDEs de type 3
aient un rôle important à jouer dans la reprise de la méiose chez les ovocytes bovins.

ABSTRACT The use of broad-spectrum inhibitors first suggested that phosphodiesterases (PDEs) are
involved on the maturation of bovine oocytes. Modulation of individual PDE families is now
possible with the use of newly developed type-specific PDE inhibitors. This study evaluated
the role of type 3- and 4-specific PDE inhibitors on the meiotic arrest of bovine cumulus-
oocyte complexes (COCs) and denuded oocytes (DOs). It also evaluated the role of these
specific inhibitors on meiotic arrest when COCs are incubated in the presence or absence of
theca cell monolayers. Bovine COCs were aspirated from ovaries collected at the abattoir.
Denuded oocytes and COCs were incubated for 12 h in culture medium alone or culture
medium containing the type 3 PDE inhibitors: cilostamide (10 and 20 µM), milrinone (10 and
50 µM) or the type 4 PDE inhibitor rolipram (10 and 50 µM). Oocytes were then fixed and
classified according to the status of nuclear maturation. Cumulus-oocyte complexes were
coincubated with untreated theca cell monolayers or theca cell monolayers treated with the
different specific PDE inhibitors. Bovine COCs or DOs incubated in culture medium
resumed meiosis, but supplementation of the culture medium with PDE3 inhibitors
cilostamide or milrinone resulted in meiotic arrest. On the other hand, supplementation of the
culture medium with rolipram did not prevent oocyte maturation. Furthermore, PDE3
inhibitors, but not PDE4 inhibitors, had an additive effect on the inhibitory action of theca cell
monolayers on oocyte maturation. These data support that inhibition of PDE3 prevents the
meiotic resumption of bovine oocytes, whereas inhibition of PDE4 does not block oocyte
maturation even under normally inhibitory conditions. The additive effect of PDE3 inhibitors
on the ability of theca cells to maintain bovine oocytes in meiotic arrest suggests that type 3
PDE has an important role in meiotic resumption of bovine oocytes.
INTRODUCTION Bovine oocytes enter meiosis early during fetal development, but they soon become arrested
at the diplotene stage of the first meiotic division and remain arrested until undergoing
ovulation, atresia or artificial removal from the follicle. However, bovine oocytes must
measure at least 110 µM in diameter to be considered competent to resume meiosis and to
complete their nuclear maturation [1, 2]. The resumption of meiosis occurs in vivo following
the endogenous preovulatory surge of LH [3], whereas in vitro, oocytes spontaneously resume
meiosis following their removal from the follicular microenvironment [4].

The somatic components of the follicle have been implicated in the maintenance of meiotic
arrest [5-7]. Original studies proposed that the inhibitory factor(s) originated from granulosa
cells [8, 9]. However, further experiments showed that isolated theca cell layers or theca cells
cultured in monolayers secrete an inhibitory factor capable of maintaining cumulus-enclosed
oocytes (Cumulus-oocyte complexes [COCs]) in meiotic arrest [7, 10]. Denuded oocytes
(DOs) do not respond to the inhibitory action of theca cell monolayers [7, 10].
The second messenger cAMP has been shown to play a key role in the maintenance of bovine
oocytes in meiotic arrest. High levels of cAMP in the oocyte induced by the addition of
membrane permeable cAMP derivatives such as dbcAMP or 8-bromo-3’ 5’ cAMP (8-br-
cAMP) have an inhibitory effect on meiotic resumption [11, 12]. However, high levels of
cAMP present in cumulus cells result in meiotic resumption [13, 14]. Therefore, the levels of
cAMP present in the somatic or germ cell compartment could be used to modulate the meiotic
resumption of bovine oocytes.
The total levels of cAMP found in the oocyte depend primarily on the rate of synthesis by
adenylate cyclase (AC) and the rate of degradation by phosphodiesterases (PDE). In the
bovine, AC activity has been localized to the cumulus cells and to the plasma membrane of
the oocyte [13, 15]. Early studies showed that purine and nucleosides influence the activity of
adenylate cyclase [16]. However, it is uncertain whether the oocyte is able to produce enough
cAMP to prevent meiosis resumption [17].
Phosphodiesterases hydrolyze the 3’ phosphoester bond of the 3’ 5’ purine ribose cyclic
monophosphate cAMP and cGMP, thereby transforming them to their corresponding
biologically inactive monophosphates. At least 11 different PDE families have been
identified since the original purification and characterization of PDE activity [18-22].
Phosphodiesterase families can be identified based on their kinetics and substrate
characteristics, inhibitor profiles, allosteric activators and inhibitors and amino acid sequences
[20]. Phosphodiesterases are composed of a catalytic domain, which is connected to the
amino and carboxy-terminal domains by hinge regions [21]. It is believed that the catalytic
domain is structurally homologous between PDEs of the same family, while the other
domains act as modulators of the catalytic center [19].

Original experiments used non-selective or first-generation inhibitors, such as IBMX (3-
isobutyl-1-methylxanthine), papaverine, theophylline and pentoxifylline, to study PDE
activity [23]. These inhibitors exert no directed action against any one type of PDE, but in
general inhibit many isozymes. These studies suggested that PDEs are involved in oocyte
maturation [24]. Their lack of specificity, inhibition of adenosine action and induction of
toxic side effects of these inhibitors limit their usefulness. Approximately 45% of bovine
COCs incubated in the presence of 0.2 mM IBMX are temporarily prevented from resuming
meiosis [11]. More recently, second-generation or isozyme-specific PDE inhibitors have
permitted evaluation the physiological role of specific PDE isoenzymes in the overall cyclic
nucleotide degradation pathway of the intact cell. These inhibitors act mainly as active site
competitors and are used to develop pharmacological treatments with less undesirable side
effects [25]. In addition, pharmacological profiles of individual PDE isoenzymes can be
created based on their response to specific PDE inhibitors. The concentration at which these
specific inhibitors are used plays a role in their specificity [26-28]. Recent studies used
specific PDE inhibitors to demonstrate the presence of PDE3A in rat and mouse oocytes.
These inhibitors prevent oocyte maturation both in vivo and in vitro [18, 29].
The PDE3 family shows high affinity to cAMP as a substrate and is inhibited by the presence
of cGMP, which binds tightly to the enzyme and it is poorly hydrolyzed. Thus, micromolar
concentrations of cGMP greatly inhibit cAMP hydrolysis [30-32]. This PDE family, which is
also known as cGMP inhibited PDE (cGI-PDE), has at least two different gene products:
PDE3A and PDE3B. Cilostamide and milrinone are specific PDE3 inhibitors [33].
The PDE4 family or cAMP-specific PDE hydrolyzes cAMP with high affinity. However, the
PDE4 family, unlike the PDE3 family, is not inhibited by cGMP [34, 35]. This family has at
least 4 different gene products with 2 or more alternative splice variants differently expressed
in different tissues [20]. Rolipram is routinely used as a specific PDE4 inhibitor [25, 29].
This study was undertaken to evaluate the specific role of PDE inhibitors on meiotic
resumption of bovine oocytes. Since high concentrations of cAMP in the cumulus cells
compared to high concentrations of cAMP in the oocyte result in opposite effects on meiotic
resumption [13], it is quite possible that these cell types have different phosphodiesterases.
This article is based on the hypothesis that the inhibition of specific subtypes of
phosphodiesterases in the oocyte and the cumulus cells of bovine cumulus-oocyte complexes

may be used to control meiotic arrest. The focus of this study was placed on the role of type 3
and type 4 phosphodiesterases. The effect of type-specific inhibitors was also evaluated in
the presence of absence of theca cell monolayers.
MATERIALS AND METHODS
Preparation of Theca Cell Monolayers Bovine theca cell monolayers were prepared as previously described [10]. Bovine follicles
(2-5mm) were randomly chosen from ovaries that were kept on ice in bags containing gauze
soaked with saline solution. Follicles were dissected free of stromal tissue with scissors.
Theca cells were harvested exclusively from transparent follicles containing at least 75% of
the granulosa layer intact and unexpanded COC. Selected follicles (n=12) were placed in a
calcium- and magnesium-free Hanks Balanced Salt Solution and cut into hemisections with a
scalpel. Granulosa cells and COCs were removed by scraping the hemisections with a fine
glass loop made from a Pasteur pipette. This technique is used to effectively isolate theca cell
layers. The theca layers were enzymatically digested in 15 ml of modified Hanks solution
supplemented with 2125 U of collagenase type II, 1600 U trypsin from porcine pancreas, 1
mg DNAse type I and 2 mg EDTA (all from Sigma). Enzymatic digestion took place for 1 h
at 38.5°C in an atmosphere of 95% air and 5% CO . Theca layers were passed through a 10-
ml pipette every 20 min. Following incubation, larger pieces of remaining follicular
hemisections were withdrawn from the solution. The reaction was stopped by the addition of
6 ml of washing medium, consisting of HEPES-buffered Tyrodes medium (TLH, pH 7.4) [36]
supplemented with 10% fetal calf serum
2
(heat-treated FCS from MEDICORP, Montreal, PQ,
Canada), 0.2 mM pyruvic acid and 50 µg/ml gentamicin sulfate. Cells were then centrifuged
(10 min at 2000 x g), resuspended, and washed a second time. The first and second pellets
were resuspended in 10 ml and 1 ml, respectively, of washing medium. Theca cells were
counted with a hemocytometer and seeded in a 24-well plate (Falcon Becton Dickinson,
Rutherford, NJ) at a concentration of 1.5-2.0 x 10 cells/ml. The culture medium, consisting
of TCM-199 with Earle’s salts (Gibco Laboratories, Grand Island, NY) and bicarbonate
(Sigma Company, St. Louis MO), 10% FCS, 0.2 mM pyruvic acid and 50 µg/ml gentamicin
sulfate, was pre-exposed to culture conditions (38.5°C, 5% CO :95% air atmosphere with
100% humidity) for at least 2 h. The culture medium was replenished every 48 h, and
5
2

follicular cells were incubated for 5-7 days. When the monolayers were confluent, the
medium was replenished and conditioned for 48 h.
Collection of COCs
Bovine ovaries at various stages of their reproductive cycle were collected at the
slaughterhouse and transported to the laboratory in saline solution kept between 30-35ºC.
The saline solution consisted of 0.9% NaCl (w:v supplemented with 100 000 IU penicillin,
100 mg streptomycin and 250 µg amphotericin B per liter (All from Sigma Chemical
Company Co, St. Louis, MO)). The contents of 1-5 mm follicles were aspirated using a 10-ml
syringe and an 18-gauge needle. The follicular contents were pooled in 50-ml conical tubes.
After sedimentation, the cumulus-enclosed oocytes (COCs) were recovered with the use of a
stereomicroscope. The COCs used in these experiments had at least 5 layers of cumulus cells.
The COCs were rinsed twice in the supernatant of follicular fluid (centrifuged for 10 min at
350 x g) and then transferred to the respective treatments.
Chemicals
The doses and PDE inhibitors tested in this study were: cilostamide (10 and 20 µM),
milrinone (10 and 50 µM) or rolipram (10 and 50 µM). The chemicals were added to the
culture medium or the theca cell monolayers at approximately 4 h before the oocytes were
added. Millimolar stock concentrations of PDE inhibitors were dissolved in dimethyl
sulfoxide (DMSO), stored at –20 ºC, and diluted into culture medium. Final concentrations of
DMSO never exceeded 0.1%, and equal amounts of carrier were added to control groups of
cells.
Denuded Oocytes In experiment 1, cumulus cells were removed from the COCs before culturing them in the
presence of PDE inhibitors. Selected COCs were vortexed for 10 min in sterile centrifuge
tubes containing follicular fluid supernatant (prepared as indicated above). After vortexing,
oocytes were rinsed in follicular fluid supernatant. Denuded oocytes with a homogeneous
cytoplasm were then allocated to their respective treatments.

Fixation of OocytesAt the end of each treatment, COCs were transferred into 2-ml centrifuge tubes containing
500 µl of TLH and vortex-agitated for 7 min to remove the cumulus cells. Denuded oocytes
were recovered under a stereomicroscope and transferred onto a glass slide in a small drop of
medium. A Vaseline:paraffin wax mixture was used to maintain the coverslip in contact with
the oocytes. The coverslip was fixed in place with epoxy glue, and the slides were immersed
in a fixative solution (ethanol:acetic acid, 3:1) for a minimum of 24 h. Oocytes were then
stained with 1% aceto-orcein and examined for nuclear morphology with a phase contrast
microscope at 100 and 400x magnification [37].
EXPERIMENTAL DESIGN
Exp 1: PDE inhibitors and bovine DO In experiment 1, COCs were denuded and then allocated in groups of 15-20 oocytes per
treatment. Denuded oocytes were either fixed at time zero or incubated for 12 h in culture
medium or culture medium supplemented with the PDE inhibitors: cilostamide (10 and 20
µM), milrinone (10 and 50 µM) or rolipram (10 and 50 µM). In addition, a group of oocytes
were treated with 0.1% DMSO to evaluate if the carrier itself used at the highest dose had an
effect on meiotic resumption.
Exp 2: PDE inhibitors and bovine COC In experiment 2, COCs were incubated in culture medium or culture medium supplemented
with the same inhibitors PDE inhibitors used in experiment 1. COCs were denuded after 12 h
of treatment and fixed as previously described.
Exp 3: PDE inhibitors and bovine COCs incubated with theca cell monolayers In experiment 3, groups of 15-20 COCs were incubated for 12 h with untreated theca cell
monolayers or theca cell monolayers supplemented with the same PDE inhibitors used in
experiment 2. The effect of DMSO on oocyte maturation was also tested by culturing COCs
in the presence of theca cell monolayers supplemented with 0.1% DMSO.

STATISTICAL ANALYSIS The status of nuclear maturation of 1201 oocytes was evaluated with a contrast microscope
immediately after staining with aceto-orcein. Oocytes with a nuclear membrane present were
classified as germinal vesicle (GV) stage, while those without a nuclear membrane were
classified as having undergone germinal vesicle breakdown (GVBD) and thus meiotic
resumption. Results are expressed as means ± SEM. Data were analyzed by a two-way
ANOVA. When ANOVA indicated a significant effect of treatment (P<0.05), treatment
differences were compared by the Duncan multiple range test.
RESULTS In experiment 1, COCs were denuded of their cumulus cells (i.e., DOs) and then incubated in
the presence of specific PDE inhibitors for 12 h. Denuded oocytes incubated in the culture
medium alone or in the presence of rolipram at either 10 or 20 µM did not remain at the GV
stage (3.3 ± 1.7%; 3.0 ± 3.3% and 0 ± 0% GV, respectively). However, DOs incubated for 12
h in the presence of 10 and 20 µM cilostamide or 50 µM milrinone remained in meiotic arrest
(61.2 ± 9.5%, 68.1 ± 7.7% and 49.2 ± 14%, respectively). However, milrinone did not
prevent meiotic resumption when used at the 10 µM dose. Denuded oocytes incubated with
PD3 inhibitors remain in meiotic arrest, whereas DOs incubated with PDE4 inhibitors resume
meiosis (Figure 3.1).
In experiment 2, COCs were incubated in the presence or absence of specific PDE inhibitors
for 12 h. Cumulus-oocyte complexes incubated in culture medium resumed meiosis, as did
those COCs incubated with the type 4 PDE inhibitor rolipram. In contrast, COCs incubated
in the presence of PDE3 inhibitors did not resume meiosis. Cilostamide used at 10 and 20
µM maintained COCs at the GV stage (30.4 ± 15.2% and 46.3 ± 5.3% GV, respectively).
Incubation of COCs in the presence of 50 µM milrinone maintained 22.5 ± 0.3% of treated
COCs at the GV stage. However, milrinone used at the 10 µM dose maintained only 3.9 ±
2% of those COCs treated in meiotic arrest, which was not significantly different than the
percentages of meiotic arrest obtained when COCs are left untreated. Cumulus-oocyte
complexes treated with PDE3 inhibitors remain in meiotic arrest, whereas those treated with
PDE4 inhibitors resumed meiosis (Figure 3.2).

In experiment 3, COCs were coincubated with theca cell monolayers in the presence or
absence of type-specific PDE inhibitors. Theca cell monolayers maintained 36.5% ± 3.4 of
the oocytes in meiotic arrest. The addition of 0.1% DMSO to the coincubated COCs and
theca cell did not have a significant effect. The addition of rolipram, a type 4 PDE inhibitor,
at either 10 or 50 µM, had no effect on the meiotic arrest maintained by theca cells.
The addition of PDE3 inhibitors to the coincubation significantly increased the number of
oocytes arrested at the GV stage (P< 0.05). The percentage COCs at the GV stage rose to
more than 80% when COCs were incubated with theca cells and treated with cilostamide
(Figure 3.3). Milrinone at both 10 and 50 µM also significantly increased the rate of meiotic
arrest (61% ± 4.9% and 83.2% ± 7.6% GV, respectively). Under these experimental
conditions, cilostamide and milrinone were both equally effective in enhancing the ability of
theca cells to maintain the COCs in meiotic arrest.
DISCUSSION The second messenger cAMP plays a crucial role in the maturation of mammalian oocytes.
However, the exact role of cAMP in the maintenance of bovine COCs in meiotic arrest is not
entirely clear. It would seem that high levels of cAMP in the cumulus cells that surround the
oocyte induce meiotic resumption, while high levels of cAMP inside the oocyte result in
meiotic arrest [13]. The extensive network of gap junctions between the oocyte and the
cumulus cells could facilitate the transfer of cAMP between these two compartments [38].
Nonetheless, the direct transfer of cAMP from the cumulus cells to the oocyte has not been
fully demonstrated [39], but when cAMP is high in cumulus cells, then it is also high in the
oocyte [14, 40]. High levels of cAMP in the cumulus cells may induce the release of a
signal(s) that triggers meiotic resumption despite the presence of high levels of cAMP in the
oocyte [13].
Raising the levels of cAMP in bovine oocytes by treating them with an activator of AC
(forskolin), or cAMP analogs (dbcAMP and 8-br-cAMP) transiently delays meiotic
resumption, thus supporting the idea that high levels of cAMP in the oocyte affect oocyte
maturation [14]. Electron microscopy studies have shown that the increased presence of AC
in the cumulus cell-projections contacting the oocyte occurs only following forskolin
stimulation. In the absence of this stimulation, the oocyte might not have enough AC

enzymatic activity to affect its nuclear maturation [15]. It has been hypothesized that the
cumulus cells might transfer cAMP to the cumulus oocyte and are thus involved in the
regulation of meiotic arrest [41-43].
Preventing the degradation of cAMP by means of broad-spectrum PDE inhibitors such as
IBMX can also transiently delay meiotic resumption [11]. Interestingly, IBMX does not
inhibit PDE8 and PDE9 [44]. A transient effect of IBMX on meiotic arrest was observed
after incubating bovine COCs with IBMX for 8 h; however, this effect was not noticeable
after 24h [13]. In addition, IBMX is more effective in preventing the resumption of meiosis
in zona-free oocytes than in COCs. Treatment of COCs with IBMX increases the levels of
cAMP in both the cumulus cells and the oocyte. Cumulus-oocyte complexes treated with
IBMX contain approximately twice the level of cAMP found in untreated COCs [13].
In this study, PDE3 and PDE4 inhibitors were used to determine whether or not type 3 and
type 4 phosphodiesterases play a role in modulating the resumption of meiosis of bovine
oocytes. The PDE3 inhibitors: cilostamide and milrinone, maintained bovine oocytes in
meiotic arrest. On the other hand, the PDE4 inhibitor rolipram used under the same
experimental conditions did not block oocyte maturation. These results suggest a role for
PDE3 in the meiotic arrest of bovine COCs and more effectively in denuded bovine oocytes.
The ability of cilostamide and milrinone to inhibit meiotic resumption suggests that inhibition
of PDE3 prevents meiotic resumption in bovine oocytes [45]. Similar results have been
observed in rat oocytes both in vitro [29] and in vivo [46]. Cilostamide is more effective than
milrinone in maintaining meiotic arrest in bovine oocytes. A higher dose of milrinone is
required to maintain rat oocytes in meiotic arrest [29].
Theca cell monolayers have been shown to be effective in maintaining bovine COCs, but not
DOs, in meiotic arrest [10, 47]. Our laboratory has demonstrated that the inhibitory factor
produced by the theca cells requires that cumulus cells be in direct contact with the oocyte.
The cumulus cells play a key role in either transporting or processing the inhibitory factor
produced by the theca cell monolayers [10]. In addition, modulation of cAMP is important in
controlling meiotic arrest of bovine COCs incubated with theca cell monolayers [47]. The
addition of PDE3 inhibitors to the coincubation of COCs and theca cell monolayers had an
additive effect on meiotic arrest. Incubation of COCs in the presence of theca cell

monolayers and type 3 PDE inhibitors resulted in higher rates of meiotic arrest than those
obtained by incubating COCs in the presence of untreated theca cell monolayers. However,
the type 4 PDE inhibitor rolipram did not have an additive effect. The theca cell factor(s)
may indirectly act by decreasing the activity of type 3 phosphodiesterases. We suggest that
these two inhibitory mechanisms are complementary or maybe alternative methods to
maintain meiotic arrest. Phosphodiesterase inhibitors will act by regulating cAMP, whereas
theca cells might use one or more alternative mechanisms. Unfortunately, the mechanism(s)
used by theca cell monolayers to maintain meiotic arrest has not yet been elucidated.
Another way to explain these results is based on the COCs ability to respond to the inhibitory
signals. There might be differences in the oocytes’ ability to respond to the inhibitory signals
despite the care taken to select, as much as possible, a morphologically homogenous
population of COCs. This hypothesis is supported by the fact that neither system is capable
of maintaining all of the treated oocytes in meiotic arrest. When used in conjunction, both
systems might be able to target more oocytes than either one alone. The remaining
unresponsive oocytes are probably already committed to meiotic resumption.
Incubation of oocytes with rolipram had no significant effect under any of the other
experimental conditions tested. This finding suggests that PDE4 is not involved in the
maintenance of meiotic arrest of bovine oocytes. Due to the considerable amount of research
carried out in vitro [25, 29, 48] and in vivo with rolipram [46] the authors believe that
product’s inability to maintain meiotic arrest is not a result of a lack of permeability across the
plasma membrane.
Differential regulation of cAMP levels by PDEs might explain the role of cAMP in meiotic
resumption. The localization of active PDE subtypes to different cell type compartments such
as somatic or germ cell appears to play an important role in the regulation of meiosis [29, 38].
It was recently reported that treatment of mural granulosa cells with the PDE4 inhibitor
rolipram resulted in a 2.5-fold increase in the level of cAMP over controls, whereas treatment
with the PDE3 inhibitor milrinone did not rise cAMP levels in those cells [49]. Therefore,
preferential activation of certain PDE subtypes in the cumulus cells or the oocyte might
determine whether the oocyte remains in meiotic arrest or proceeds to meiotic resumption.

In conclusion, the results of this study show that inhibition of PDE3 prevents meiotic
resumption of bovine oocytes, whereas inhibition of PDE4 does not block oocyte maturation.
Furthermore, the additive effect of PDE3 inhibitors on the meiotic arrest maintained by theca
cell monolayers suggests that PDE3 plays a role in meiotic maturation of bovine oocytes.
ACKNOWLEDGEMENTS This work was supported by the Natural Sciences and Engineering Research Council of
Canada (NSERC) and Semex Canada. The authors would like to express their gratitude to
François Richard, Ph.D. for his critical review of the manuscript.
REFERENCES 1. Fair T, Hyttel P, Greve T. Bovine oocyte diameter in relation to maturational
competence and transcriptional activity. Mol Reprod Dev 1995; 42: 437-442. 2. Hyttel P, Fair T, Callesen H, Greve T. Oocyte growth, capacitation and final
maturation in cattle. Theriogenology 1997; 47: 23-32. 3. Hyttel P, Callesen H, Greve T. Ultrastructural features of preovulatory oocyte
maturation in superovulated cattle. J Reprod Fertil 1986; 76: 645-656. 4. Pincus G, Enzmann EV. The comparative behavior of mammalian eggs in vitro and in
vivo. J. Exp. Med. 1935; 62: 665-675. 5. De Loos FA, Zeinstra E, Bevers MM. Follicular wall maintains meiotic arrest in
bovine oocytes cultured in vitro. Mol Reprod Dev 1994; 39: 162-165. 6. Carbonneau G, Sirard MA. Influence of follicular wall on meiotic resumption of
bovine oocytes when cultured inside or outside hemisections. J Reprod Dev 1994; 40: 125-132.
7. Richard FJ, Sirard MA. Effects of follicular cells on oocyte maturation. I: Effects of follicular hemisections on bovine oocyte maturation in vitro. Biol Reprod 1996; 54: 16-21.
8. Sirard MA, Bilodeau S. Granulosa cells inhibit the resumption of meiosis in bovine oocytes in vitro. Biol Reprod 1990; 43: 777-783.
9. Sirard MA, Coenen K, Bilodeau S. Effect of fresh or cultured follicular fractions on meiotic resumption in bovine oocytes. Theriogenology 1992; 37: 39-58.
10. Richard FJ, Sirard MA. Effects of follicular cells on oocyte maturation. II: Theca cell inhibition of bovine oocyte maturation in vitro. Biol Reprod 1996; 54: 22-28.
11. Sirard MA, First NL. In vitro inhibition of oocyte nuclear maturation in the bovine. Biol Reprod 1988; 39: 229-234.
12. Downs SM, Hunzicker-Dunn M. Differential regulation of oocyte maturation and cumulus expansion in the mouse oocyte-cumulus cell complex by site-selective analogs of cyclic adenosine monophosphate. Dev Biol 1995; 172: 72-85.
13. Bilodeau S, Fortier MA, Sirard MA. The effect of adenylate cyclase stimulation on meiotic resumption and cyclic AMP content of zona-free and cumulus-enclosed bovine oocytes in vitro. J Reprod Fertil 1993; 97: 5-11.

14. Aktas H, Wheeler MB, Rosenkrans CF, Jr., First NL, Leibfried-Rutledge ML. Maintenance of bovine oocytes in prophase of meiosis I by high [cAMP]i. J Reprod Fertil 1995; 105: 227-235.
15. Kuyt JR, Kruip TA, de Jong-Brink M. Cytochemical localization of adenylate cyclase in bovine cumulus-oocyte complexes. Exp Cell Res 1988; 174: 139-145.
16. Polan ML, DeCherney AH, Haseltine FP, Mezer HC, Behrman HR. Adenosine amplifies follicle-stimulating hormone action in granulosa cells and luteinizing hormone action in luteal cells of rat and human ovaries. J Clin Endocrinol Metab 1983; 56: 288-294.
17. Tsafriri A, Dekel N. Molecular mechanisms in ovulation. In: Findlay JK (ed.) Molecular Biology of the Female Reproductive System. San Diego: Academic Press; 1994: 207-258.
18. Conti M. Phosphodiesterases and cyclic nucleotides signaling in endocrine cells. Mol Endocrinol 2000; 14: 1317-1327.
19. Conti M, Jin SL. The molecular biology of cyclic nucleotide phosphodiesterases. Prog Nucleic Acid Res Mol Biol 1999; 63: 1-38.
20. Beavo JA. Cyclic nucleotide phosphodiesterases: functional implications of multiple isoforms. Physiol Rev 1995; 75: 725-748.
21. Conti M, Jin SL, Monaco L, Repaske DR, Swinnen JV. Hormonal regulation of cyclic nucleotide phosphodiesterases. Endocr Rev 1991; 12: 218-234.
22. Fisher DA, Smith JF, Pillar JS, St Denis SH, Cheng JB. Isolation and characterization of PDE9A, a novel human cGMP-specific phosphodiesterase. J Biol Chem 1998; 273: 15559-15564.
23. Downs SM, Daniel SA, Bornslaeger EA, Hoppe PC, Eppig JJ. Maintenance of meiotic arrest in mouse oocytes by purines: modulation of cAMP levels and cAMP phosphodiesterase activity. Gamete Res 1989; 23: 323-334.
24. Sadler SE, Maller JL. A similar pool of cyclic AMP phosphodiesterase in Xenopus oocytes is stimulated by insulin, insulin-like growth factor 1, and [Val12,Thr59]Ha-ras protein. J Biol Chem 1989; 264: 856-861.
25. Atienza JM, Susanto D, Huang C, McCarty AS, Colicelli J. Identification of inhibitor specificity determinants in a mammalian phosphodiesterase. J Biol Chem 1999; 274: 4839-4847.
26. Reeves ML, Leight BK, England PJ. The identification of a new cyclic nucleotide phosphodiesterase activity in human and guinea-pig cardiac ventricle. Biochem J. 1987; 241: 535-541.
27. Weishaar RE, Burrows SD, Kobylarz DC, Quade MM, Evans DB. Multiple molecular forms of cyclic nucleotide phosphodiesterase in cardiac and smooth muscle and in platelets. Isolation, characterization, and effects of various reference phosphodiesterase inhibitors and cardiotonic agents. Biochem Pharmacol 1986; 35: 787-800.
28. Hidaka H, Hayashi H, Kohri H, Kimura Y, Hosokawa T, Igawa T, Saitoh Y. Selective inhibitor of platelet cyclic adenosine monophosphate phosphodiesterase, cilostamide, inhibits platelet aggregation. J Pharmacol Exp Ther 1979; 211: 26-30.
29. Tsafriri A, Chun SY, Zhang R, Hsueh AJ, Conti M. Oocyte maturation involves compartmentalization and opposing changes of cAMP levels in follicular somatic and germ cells: studies using selective phosphodiesterase inhibitors. Dev Biol 1996; 178: 393-402.

30. Loten EG, Assimacopoulos-Jeannet FD, Exton JH, Park CR. Stimulation of a low Km phosphodiesterase from liver by insulin and glucagon. J Biol Chem 1978; 253: 746-757.
31. Meacci E, Taira M, Moos M, Jr., Smith CJ, Movsesian MA, Degerman E, Belfrage P, Manganiello V. Molecular cloning and expression of human myocardial cGMP-inhibited cAMP phosphodiesterase. Proc Natl Acad Sci U S A 1992; 89: 3721-3725.
32. Taira M, Hockman SC, Calvo JC, Belfrage P, Manganiello VC. Molecular cloning of the rat adipocyte hormone-sensitive cyclic GMP- inhibited cyclic nucleotide phosphodiesterase. J Biol Chem 1993; 268: 18573-18579.
33. Harrison SA, Reifsnyder DH, Gallis B, Cadd GG, Beavo JA. Isolation and characterization of bovine cardiac muscle cGMP-inhibited phosphodiesterase: a receptor for new cardiotonic drugs. Mol Pharmacol 1986; 29: 506-514.
34. Swinnen JV, Joseph DR, Conti M. The mRNA encoding a high-affinity cAMP phosphodiesterase is regulated by hormones and cAMP. Proc Natl Acad Sci U S A 1989; 86: 8197-8201.
35. Colicelli J, Birchmeier C, Michaeli T, O'Neill K, Riggs M, Wigler M. Isolation and characterization of a mammalian gene encoding a high- affinity cAMP phosphodiesterase. Proc Natl Acad Sci U S A 1989; 86: 3599-3603.
36. Bavister BD, Leibfried ML, Lieberman G. Development of preimplantation embryos of the golden hamster in a defined culture medium. Biol Reprod 1983; 28: 235-247.
37. Motlik J, Koefoed-Johnsen HH, Fulka J. Breakdown of the germinal vesicle in bovine oocytes cultivated in vitro. J Exp Zool 1978; 205: 377-383.
38. Eppig JJ, Downs SM. Chemical signals that regulate mammalian oocyte maturation. Biol Reprod 1984; 30: 1-11.
39. Schultz RM, Montgomery RR, Belanoff JR. Regulation of mouse oocyte meiotic maturation: implication of a decrease in oocyte cAMP and protein dephosphorylation in commitment to resume meiosis. Dev Biol 1983; 97: 264-273.
40. Aktas H, Wheeler MB, First NL, Leibfried-Rutledge ML. Maintenance of meiotic arrest by increasing [cAMP]i may have physiological relevance in bovine oocytes. J Reprod Fertil 1995; 105: 237-245.
41. Dekel N. Spatial relationship of follicular cells in the control of meiosis. Prog Clin Biol Res 1988; 267: 87-101.
42. Dekel N. Regulation of oocyte maturation. The role of cAMP. Ann N Y Acad Sci 1988; 541: 211-216.
43. Dekel N, Aberdam E, Sherizly I. Spontaneous maturation in vitro of cumulus-enclosed rat oocytes is inhibited by forskolin. Biol Reprod 1984; 31: 244-250.
44. Soderling SH, Beavo JA. Regulation of cAMP and cGMP signaling: new phosphodiesterases and new functions. Curr Opin Cell Biol 2000; 12: 174-179.
45. Mayes MA. Effect of type 3 and type 4 phosphodiesterase inhibitors on the maintenance of bovine cumulus-enclosed oocytes in meiotic arrest. Theriogenology 2000; 53: 459.
46. Wiersma A, Hirsch B, Tsafriri A, Hanssen RG, Van de Kant M, Kloosterboer HJ, Conti M, Hsueh AJ. Phosphodiesterase 3 inhibitors suppress oocyte maturation and consequent pregnancy without affecting ovulation and cyclicity in rodents. J Clin Invest 1998; 102: 532-537.
47. Richard FJ, Fortier MA, Sirard MA. Role of the cyclic adenosine monophosphate-dependent protein kinase in the control of meiotic resumption in bovine oocytes cultured with thecal cell monolayers. Biol Reprod 1997; 56: 1363-1369.

48. DiSanto ME, Heaslip RJ. Rolipram inhibition of phosphodiesterase-4 activation. Eur J Pharmacol 1995; 290: 169-172.
49. Thomas RE, Armstrong DT, Gilchrist RB. Differential effects of specific phosphodiesterase isoenzyme inhibitors on bovine oocyte meiotic maturation. Dev Biol 2002; 244: 215-225.
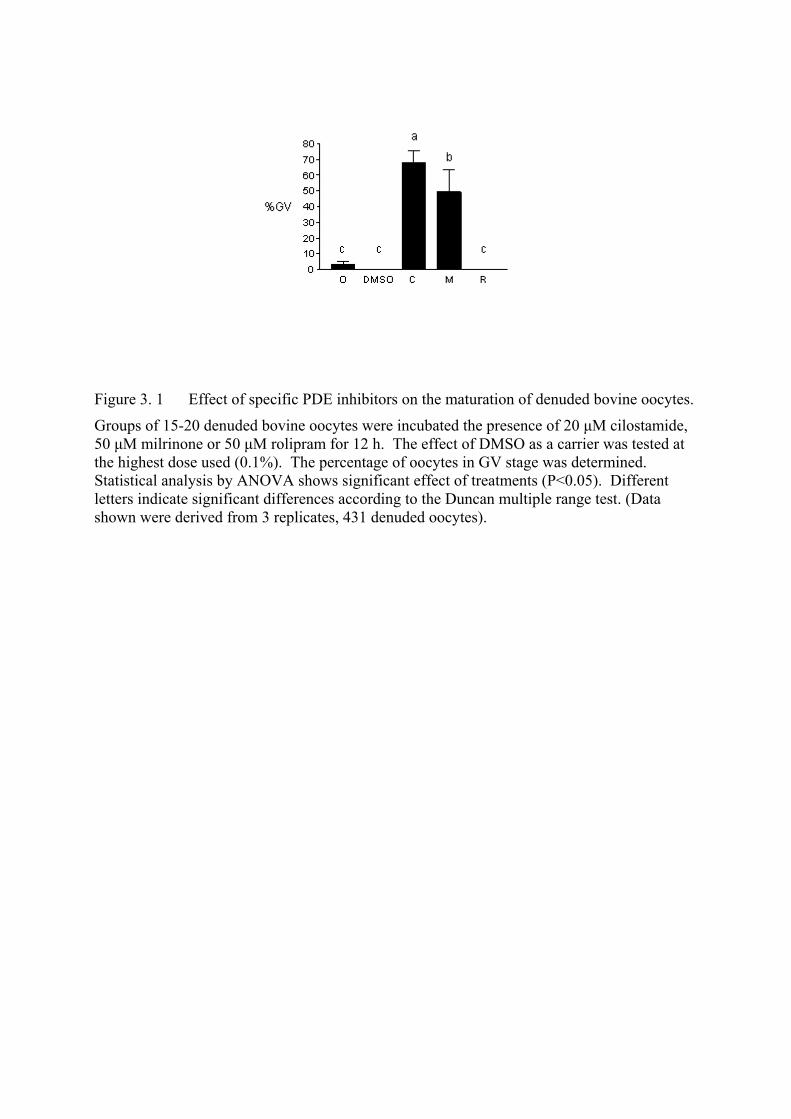
Figure 3. 1 Effect of specific PDE inhibitors on the maturation of denuded bovine oocytes.
Groups of 15-20 denuded bovine oocytes were incubated the presence of 20 µM cilostamide, 50 µM milrinone or 50 µM rolipram for 12 h. The effect of DMSO as a carrier was tested at the highest dose used (0.1%). The percentage of oocytes in GV stage was determined. Statistical analysis by ANOVA shows significant effect of treatments (P<0.05). Different letters indicate significant differences according to the Duncan multiple range test. (Data shown were derived from 3 replicates, 431 denuded oocytes).

Figure 3. 2 Effect of specific PDE3 and PDE4 inhibitors on the maturation of bovine COCs.
Bovine COCs were incubated in the presence of 20 µM cilostamide, 50 µM milrinone or 50 µM rolipram for 12 h. The percentage of oocytes in GV stage was determined. Statistical analysis by ANOVA shows significant effect of treatments (P<0.05). Different letters indicate significant differences according to the Duncan multiple range test. (Data shown were derived from 3 replicates, 360 COCs).

Figure 3. 3 Effect of specific PDE3 and PDE4 inhibitors on the maturation of bovine COCs incubated with theca cell monolayers.
Bovine COCs were incubated for 12 h in contact with theca cell monolayers in presence of 20 µM cilostamide, 50 µM milrinone, 50 µM rolipram. The percentage of oocytes in GV stage was determined. Statistical analysis by ANOVA shows significant effect of treatments (P< 0.05). Different letters indicate significant differences according to Duncan multiple range test. (Data shown were derived from 3 replicates, 410 COCs).

CHAPTER IV Granulosa Cells Reverse the Inhibitory Effect of Theca Cell Monolayers on the Maturation of Bovine Cumulus-Oocyte Complexes.

Cet article a été soumis pour publication dans la revue Biology of Reproduction sous la
référence suivante : M. A. Mayes and M. A. Sirard. Granulosa cells reverse the inhibitory
effect of theca cell monolayers on the maturation of bovine cumulus-oocyte complexes.

RÉSUMÉ Les monocouches de cellules de thèque maintiennent les COCs bovins dans un état d`arrêt
méiotique réversible. L`objectif de cet étude était d`évaluer si les cellules de granulosa
provenant de follicules de grandes tailles (8-15 mm de diamètre) sont aptes à renverser l’effet
inhibiteur des monocouches de cellules de thèque sur l`arrêt méiotique des COCs bovins. Afin
de comprendre l’effet des cellules de granulosa, les COCs ont été incubés avec des
monocouches de cellules de thèque, des cellules de granulosa ou les deux types de cellules.
Chacun des types de combinaisons de cellules-COCs ont été traitées ou non avec 10 ng/ml de
LH, 2 mM de dbcAMP ou 10 µM de PMA. Des COCs on été évalues pour l`état de maturation
nucléaire après 24 h de traitement.. Indépendamment du traitement, les COCs incubés avec les
cellules de granulosa reprennent la méiose, alors que les COCs incubés avec les monocouches
de cellules de thèque, qui ont été traitées ou non, sont demeurées en arrêt méiotique. Les
cellules de granulosa renversent l`effet inhibiteur des monocouches de cellules de thèque sur la
maturation des COCs bovins à moins qu’elles ne soient traitées avec 2 mM dbcAMP.
L`activation de la voie de signalisation de la PKA en presence des cellules de granulosa
empêche celles-ci de renverser l`effet inhibiteur des monocouches de cellules de thèque.

106
ABSTRACT Theca cell monolayers maintain bovine cumulus-oocyte complexes (COCs) in a state of
reversible meiotic arrest. The objective of this study was to evaluate if granulosa cells from large
follicles (8-15 mm diameter) are able to counteract the inhibitory effect of theca cell monolayers
on the meiotic arrest of bovine COCs. In an attempt to understand the effect of the granulosa
cells, COCs were incubated with theca cell monolayers, granulosa cells or both cell types. Each
COCs-cell type combination was either left untreated or treated with 10 ng/mL LH, 2 mM
dbcAMP or 10 µM PMA. After 24h of treatment, COCs were evaluated for nuclear maturation.
Regardless of the treatment, COCs incubated with granulosa cells resumed meiosis; whereas
COCs incubated with treated or untreated theca cell monolayers remained in meiotic arrest.
Granulosa cells reversed the inhibitory effect of the theca cell monolayers on meiotic resumption
of bovine COCs unless they were treated with 2 mM dbcAMP. The activation of the PKA
signaling pathway in presence of granulosa cells prevents them from reversing the inhibitory
effect of theca cell monolayers.
INTRODUCTION Fully-grown mammalian oocytes must undergo meiotic maturation to become fertilizable haploid
eggs. Meiosis starts early during fetal development but it soon stops at the diplotene stage of
prophase I. The nucleus of arrested oocytes is known as germinal vesicle (GV). Oocytes remain
in this state of meiotic arrest for many months or even years until stimulated to resume meiosis at
the time of ovulation. The resumption of meiosis is characterized by changes in oocyte protein
phosphorylation [1], dissolution of the nuclear membrane and condensation of the chromosomes.
This process is known as germinal vesicle breakdown (GVBD). The oocyte pro ceeds through
several stages to become arrested at the metaphase II until fertilization triggers the expulsion of
the second polar body and meiosis is completed.
The follicle plays a role in modulating the growth, development and meiotic status of the oocyte.
Follicular wall sections [2], follicular hemisections [3] or theca cell monolayers maintain
cumulus-oocyte complexes (COCs) in a state of reversible meiotic arrest [4]. The theca cells

107
incubated as explants or monolayers are capable of maintaining cumulus-oocyte complexes
(COCs) but not denuded oocytes (DO) in a state of reversible meiotic arrest for at least 24 h. The
cumulus cells play an important role in transmitting and/or modulating the inhibitory effect of
theca cells on oocyte maturation [3, 4]. The theca cell monolayers secrete the inhibitory factor(s)
into the incubation medium. The factor is active even without direct contact between the oocyte
and the theca cell monolayer [3-5]. Theca cells or hemisections from small follicles (2-5 mm in
diameter) [6, 7] or sections of membrana granulosa attached to the COCs [3, 8] have also been
reported to exert an inhibitory effect on the meiotic process.
Meiotic resumption in vitro occurs spontaneously after the oocyte is removed from the follicle
and incubated in simple medium [9]. In vivo, luteinizing hormone (LH) triggers the resumption
of meiosis in bovine COCs. The signaling pathway by which the preovulatory LH surge induces
meiotic resumption is not fully resolved, however, in rats, treatment of follicles with cyclic
adenosine monophosphate (cAMP) can induce the same effects as LH [10]. Furthermore, it is
known that the binding of LH to its receptor on the plasma membrane of ovarian cells can
potentially activate the signaling pathways of protein kinase A (PKA) and protein kinase C
(PKC) [11, 12].
Luteinizing hormone activates adenylate cyclase, inducing a rapid increase in cAMP that results
in the activation of PKA and protein phosphorylation [1, 13-15]. Protein kinase A consists of 2
regulatory subunits and 2 catalytic subunits. The binding of 4 molecules of cAMP to the
regulatory subunits activates the catalytic subunits, which phosphorylate proteins at serine or
threonine residues. It has been proposed that high levels of cAMP maintain oocytes in meiotic
arrest [16], while a transient increase may trigger meiotic resumption [17]. The absolute level of
cAMP inside the oocyte might not be as important in triggering oocyte maturation as the
magnitude of the change in cAMP [18]. Thus, cAMP also plays a dual role in oocyte maturation.
Meiotic inhibition of bovine oocytes can be achieved at least temporarily by raising the
intracellular levels of cAMP with cAMP analogues [19-21], stimulators of adenylate cyclase [22,
23] or by specifically inhibiting type 3 phosphodiesterase expressed in the ovary [24, 25].
Luteinizing hormone is also reported to activate the protein kinase C (PKC) signaling pathway.
Luteinizing hormone activates phospholipase C, which increases phosphoinositide turnover and

108
the production of diacylglycerol, which activates PKC [26-29]. The physiological activators of
PKC are calcium (Ca2+) and diacylglycerol (DAG). Protein kinase C phosphorylates proteins
rich in threonine-serine residues [30]. Tumor promoting phorbol esters such as phorbol 12-
myristate-13-acetate (PMA) replace DAG and bind the regulatory subunit of PKC [30-32]. The
activation of PKC by phorbol esters is irreversible. The stimulation of PKC in denuded bovine
oocytes does not have any effect on oocyte maturation [33], although activation of the PKC
signaling pathway with PMA induces ovulation in perfused rabbit ovaries [12].
The expression of the LH receptor (LHr) mRNA depends on cell type and follicle size. In theca
cells, the LHr mRNA is detected in theca interna cells from follicles as small as 4 mm in diameter
[34]. Functional LHr are present in mature granulosa cells originating from follicles measuring at
least 8 mm in diameter [8, 35]. The induction of LHr in granulosa cells is mediated by concerted
actions of both FSH and estradiol [26, 27]. The responses of mature granulosa cells to LH
stimulation include: steroidogenesis, luteinization and ovulation [27]. The cumulus cells and the
oocyte do not have LHr [8, 36, 37]. This suggests that the effect of LH on meiotic resumption is
mediated through the action of LH on follicular cells.
Bovine follicles incubated in vitro maintain their respective oocytes in meiotic arrest [38].
Granulosa cells are involved in the maintenance of meiotic arrest of bovine oocytes in intact
follicles [3, 39]. However, the reconstruction of a bovine follicle in vitro induces the granulosa
cells to stimulate oocyte maturation. Research was conducted to test the hypothesis that the
interactions between the granulosa and theca cells are important to modulate the meiotic
resumption of bovine COCs in vitro. Moreover the modulation of the PKA signaling pathway is
involved in controlling oocyte maturation. This study evaluated the meiotic status of bovine
COCs incubated in the presence of granulosa cells, theca cell monolayers or both granulosa and
theca cell monolayers. In addition, COCs incubated under these conditions were left untreated or
treated with LH, dbcAMP (PKA activator) or PMA (PKC activator). Our objective was to
determine if granulosa cells could, by themselves or in the presence of LH, PKA or cAMP
analogs, reverse the inhibitory effect of theca cell monolayers.

109
MATERIALS AND METHODS
Preparation of Theca Cell Monolayers Bovine theca cell monolayers were prepared as previously described [4]. Bovine follicles (2-5
mm) were randomly chosen from ovaries that were kept on ice in bags containing gauze soaked
with saline solution. Follicles were dissected free of stromal tissue with scissors. Theca cells
were harvested exclusively from transparent follicles containing at least 75% of the granulosa
layer intact and unexpanded COC. Selected follicles (n=12) were placed in a calcium- and
magnesium-free Hanks Balanced Salt Solution and cut into hemisections with a scalpel.
Granulosa cells and COCs were removed by scraping the hemisections with a fine glass loop
made from a Pasteur pipette. This technique is used to effectively isolate theca cell layers. The
theca layers were enzymatically digested in 15 ml of modified Hanks solution supplemented with
2125 U of collagenase type II, 1600 U trypsin from porcine pancreas, 1 mg DNAse type I and 2
mg EDTA (all from Sigma). Enzymatic digestion took place for 1 h at 38.5°C in an atmosphere
of 95% air and 5% CO2. Theca layers were passed through a 10-ml pipette every 20 min.
Following incubation, larger pieces of remaining follicular hemisections were withdrawn from
the solution. The reaction was stopped by the addition of 6 ml of washing medium, consisting of
HEPES-buffered Tyrodes medium (TLH, pH 7.4) [40] supplemented with 10% fetal calf serum
(heat-treated FCS from MEDICORP, Montreal, PQ, Canada), 0.2 mM pyruvic acid and 50 µg/ml
gentamicin sulfate. Cells were then centrifuged (10 min at 2000 x g), resuspended, and washed a
second time. The first and second pellets were resuspended in 10 ml and 1 ml, respectively, of
washing medium. Theca cells were counted with a hemocytometer and seeded in a 24-well plate
(Falcon Becton Dickinson, Rutherford, NJ) at a concentration of 1.5-2.0 x 105 cells/ml. The
culture medium, consisting of TCM-199 with Earle’s salts (Gibco Laboratories, Grand Island,
NY) and bicarbonate (Sigma Company, St. Louis MO), 10% FCS, 0.2 mM pyruvic acid and 50
µg/ml gentamicin sulfate, was pre-exposed to culture conditions (38.5°C, 5% CO2: 95% air
atmosphere with 100% humidity) for at least 2 h. The culture medium was replenished every 48
h, and follicular cells were incubated for 5-7 days. When the monolayers were confluent, the
medium was replenished and conditioned for 48 h.

110
Preparation of Granulosa Cells Granulosa cells were prepared as described by Rouiller and collaborators [41]. Bovine ovaries at
various stages of their reproductive cycle were collected at the slaughterhouse and transported to
the laboratory in saline solution kept between 30-35ºC. The saline solution consisted of 0.9%
NaCl (w:v supplemented with 100 000 IU penicillin, 100 mg streptomycin and 250 µg
amphotericin B per liter). Antral granulosa cells were collected from 8-15 mm follicles.
Follicles were punctured using a 5-ml syringe and an 18-gauge needle and rinsed 10 times with
0.5 ml Menezo B2 medium supplemented with 100 IU/ml of heparin (Organon Teknika, Toronto,
Ontario). Cells were centrifuged for 10 min at 300 g at room temperature (RT) and the
supernatant discarded. Cells were resuspended in 0.5 ml of Menezo B2 medium and centrifuged
for 5 min at 50 g at RT to remove most of the red blood cells. Cells were then incubated for 15
min at 38.5°C in an atmosphere of 95% air and 5% CO2. Cells were incubated in Menezo B2
medium supplemented with 2.47 mg/ml EGTA to improve cell dissociation [42]. After a 7 min
centrifugation at 300 g at RT, cells were resuspended in Menezo B2 medium and cell viability
determined by trypan blue exclusion (0.13% v/v, final concentration) using a hemocytometer. In
an attempt to recreate a bovine follicle in vitro, granulosa cells were incubated in Millicell inserts
(approximately 150,000 cells per insert) (Millipore Co., Bedford, MA). The inserts were placed
in 24-well plates 2 h before the addition of the COCs. Granulosa cells were either incubated alone
or in the presence of theca cell monolayers.
Collection of COCs
Bovine ovaries were collected as described above. The contents of 1-5 mm follicles were
aspirated using a 10-ml syringe and an 18-gauge needle. The follicular contents were pooled in
50-ml conical tubes. After sedimentation, cumulus-oocytes complexes (COCs) were recovered
with the use of a stereomicroscope. Cumulus-oocytes complexes used in these experiments had at
least 5 layers of cumulus cells. COCs were rinsed twice in the supernatant of follicular fluid
(centrifuged for 10 min at 350 g) and then transferred to the respective treatments.

111
Chemicals
All chemicals were purchased from Sigma Chemical (St. Louis, MO). Stock concentrations of
LH and dbcAMP were dissolved in TCM-199, whereas the stock concentration of PMA was
dissolved in dimethyl sulfoxide (DMSO). Final concentrations of DMSO never exceeded 0.1%
and equal amounts of carrier were added to control groups of cells.
Evaluation of Nuclear Maturation
At the end of each treatment, COCs were transferred into 2 ml centrifuge tubes containing 500 µl
of TLH and vortex-agitated for 7 min to remove the cumulus cells. Denuded oocytes were
recovered under a stereomicroscope and transferred onto a glass slide in a small drop of medium.
A Vaseline:paraffin wax mixture was used to maintain the coverslip in contact with the oocytes.
The coverslip was fixed in place with epoxy glue and the slides were immersed in a fixative
solution (ethanol:acetic acid, 3:1) for a minimum of 24 h. Oocytes were then stained with 1%
aceto-orcein and examined for nuclear morphology with a phase contrast microscope at 100 and
400x magnification [43].
EXPERIMENTAL DESIGN Cumulus-oocyte complexes were incubated in the presence of theca cell monolayers, granulosa
cells or both for 24 h. Groups of 15-20 COCs were then left untreated or treated with 10 ng/ml
LH, 2 mM dbcAMP or 10 µM PMA. Each chemical was added at a maximum volume of 10 µl.
The effect of DMSO on oocyte maturation was tested by culturing COCs in the presence of theca
cell monolayers supplemented with 0.1% DMSO. The final concentration of DMSO never
exceeded 0.1%. After treatment, COCs were denuded, fixed and stained to evaluate their stage of
nuclear maturation.
STATISTICAL ANALYSIS Oocytes were classified as GV, representing meiotic arrest, or GVBD (including early
condensation, metaphase I to metaphase II), representing meiotic resumption. Values are
expressed as means of percentages ± SEM. Data were analyzed as a 4 x 3 factorial in a complete

112
block design with 4 replicates. Cell type and treatment were the two components of the model.
There was no replicate effect.(P<0.05) Data were analyzed using the Non-Parametric One-Way
ANOVA procedure (NPAR1WAY) of SAS (Statistical Analysis System (SAS) Institute Inc,
Cary, NC, U.S.A., Release 6.12). Significant treatment differences (P<0.05) were compared with
the least square means using the probability of differences.
RESULTS
Incubation of COCs with granulosa cells. Our results show that granulosa cells (150,000 cell/insert) did not maintain COCs at the GV stage
(0% GV ± 0). Regardless of the treatment, COCs incubated in the presence of granulosa cells
resumed meiosis. The percentages of oocytes at the GV stage after treatment of granulosa cells
with 10 ng/ml LH, 10 µM PMA or 2mM dbcAMP were respectively 1.9 ± 1.9% GV, 0 ± 0%
and 6.3 ± 6.3% (Figure 4.1).
Incubation of COCs with theca cell monolayers. On the other hand, incubation of COCs with theca cell monolayers resulted in 43.5 ± 6.9% GV.
The treatments of the theca cell monolayer with LH, PMA or dbcAMP did not have a significant
effect on the percentage of ooycte in meiotic arrest when compared to controls (34.6 ± 11.9%;
34.2 ± 6.5%, 28.1 ± 6.3% and 43.5 ± 6.9% GV; respectively). Furthermore, the treatment of the
theca cell monolayers with 0.1% DMSO (data not shown) did not have any significant effect on
the ability of theca cells to maintain meiotic arrest.
Incubation of COCs with granulosa and theca cell monolayers. Bovine oocytes incubated in the presence of granulosa cells resumed meiosis, whereas those
incubated in the presence of theca cells remained in meiotic arrest. However, when COCs were
incubated in presence of both cell types: untreated granulosa cells abrogated the inhibitory effect
of theca cells on meiotic arrest. The treatment of both cell types with LH or stimulation of the
PKC signaling pathway with PMA did not have a significant effect on the meiotic arrest of COCs
when compared to COCs incubated with untreated granulosa and theca cells (16.1 ± 5.6%, 4.8 ±

113
4.7% and 8.8 ± 6.6% GV, respectively). The inhibitory effect of theca cell monolayers was not
abrogated when both cell types where treated with 2 mM dbcAMP. Furthermore, the percentage
of oocytes at the GV stage when both cell types were present and PKA was stimulated was not
significantly different than COCs incubated only with theca cells (53.4 ± 10.2 vs. 43.5 ± 6.9%
GV, respectively).
DISCUSSION This article focuses on the control of the meiotic resumption of bovine oocytes by the different
follicular components in vitro. Bovine COCs incubated with untreated theca cell monolayers
remain in meiotic arrest. The inhibitory effect of theca cell monolayers on COCs was not
reversed by cAMP analogs (dbcAMP) that activate PKA or tumor promoting phorbol esters
(PMA) acting on the PKC or by the addition of the natural agonist LH. Doses of up to 100 ng/ml
of LH failed to raise the percentage of COCs in meiotic arrest above the level obtained with
untreated theca cells (results not shown). Since both of these pathways are involved in the LH
signaling cascade and none of them reversed the inhibitory effect of theca cells, then it seems that
LH itself will not reverse the theca cell inhibitory effect. Furthermore, the presence of LH
receptors on small to medium size follicles has been localized exclusively to the theca cells [8].
Furthermore, the cumulus cells of COCs originating from small-medium follicles do not respond
to the LH surge because they lack specific mRNA associated with the LHr. Incubation of COCs
in the presence of theca cell monolayers and type 3 phosphodiesterase (PDE3) inhibitors
(cilostamide or milrinone) results in higher rates of meiotic arrest than those obtained by
incubating COCs in the presence of untreated theca cell monolayers [24]. This suggests that
increased cAMP level in theca cells in the presence of LH is not enough to maximize its effect on
meiotic arrest. The PKA inhibitor H-89 (N-{2-(p-bromocinnamylamino)ethyl}–5-isoquinole-
sulfonamide) maintains bovine COCs but not DO in meiotic arrest [44]. Also, theca cell
monolayers treated with H-89 do not maintain the COCs in meiotic arrest. This suggests that the
activity of the inhibitory factor produced by the theca cell monolayers is diminished by PKA.
This is in agreement with the present results since stimulation of the PKA pathway with dbcAMP
did not reverse the inhibitory effect of theca cells.

114
Bovine COCs incubated with high concentrations of fresh granulosa cells (above 10 million
cells/ml) collected from small follicles remain in meiotic arrest [6, 7]. However, low
concentrations of fresh granulosa cells (2x106 cells /ml) do not maintain bovine COCs in meiotic
arrest [6]. Cumulus-oocyte complexes incubated with fresh granulosa cells collected from large
follicles (8-15 mm) resumed meiosis. Treatment of granulosa cells with LH, dbcAMP or PMA
does not prevent meiotic resumption of bovine COCs (Figure 4.1).
In contrast to granulosa cells, COCs incubated with theca cell monolayers remain in meiotic
arrest. Untreated granulosa cells are able to abrogate the inhibitory effect of theca cell
monolayers on COC maturation. Neither, LH or PMA interfered with the ability of granulosa
cells to reverse the theca cell meiotic arrest of bovine COCs. This is in agreement with the idea
that oocyte maturation takes place in vitro independently of LH stimulation. In contrast,
treatment of granulosa cells and theca cell monolayers with dbcAMP resulted in percentages of
meiotic arrest that are not significantly different than those induced by theca cells alone. This is
interesting since we know that LH activates PKA by increasing the levels of cAMP. Increasing
the levels of cAMP by cAMP-analogs such as dbcAMP under these conditions effectively
bypasses the LH receptor and suggests that activation of PKA in granulosa cells is responsible for
maintaining meiotic arrest in conjunction with the theca cell monolayers. However, activation of
PKA in granulosa cells alone does not maintain the meiotic arrest of bovine oocytes.
Overall, this study suggest that fresh granulosa cells collected from large follicles are capable of
reversing the inhibitory activity of theca cell monolayers on oocyte maturation. In addition, the
activation of the PKA signaling pathway in granulosa cells prevents them from reversing the
inhibitory activity of the theca cells. Further experiments should test the same experimental
design but using granulosa cells collected from small follicles to gain a better understanding of
the interaction between granulosa cells and theca cells in vitro.
ACKNOWLEDGMENTS This work was supported by the Natural Sciences and Engineering Research Council of Canada
(NSERC) and Semex Canada. The authors gratefully acknowledge Dr. Susan Novak for her

115
assistance with the statistical analysis and Dr. François Richard for the critical review of the
manuscript.
REFERENCES 1. Schultz RM, Montgomery RR, Belanoff JR. Regulation of mouse oocyte meiotic
maturation: implication of a decrease in oocyte cAMP and protein dephosphorylation in commitment to resume meiosis. Dev Biol 1983; 97: 264-273.
2. De Loos FA, Zeinstra E, Bevers MM. Follicular wall maintains meiotic arrest in bovine oocytes cultured in vitro. Mol Reprod Dev 1994; 39: 162-165.
3. Richard FJ, Sirard MA. Effects of follicular cells on oocyte maturation. I: Effects of follicular hemisections on bovine oocyte maturation in vitro. Biol Reprod 1996; 54: 16-21.
4. Richard FJ, Sirard MA. Effects of follicular cells on oocyte maturation. II: Theca cell inhibition of bovine oocyte maturation in vitro. Biol Reprod 1996; 54: 22-28.
5. van Tol HT, Bevers MM. Theca cells and theca-cell conditioned medium inhibit the progression of FSH-induced meiosis of bovine oocytes surrounded by cumulus cells connected to membrana granulosa. Mol Reprod Dev 1998; 51: 315-321.
6. Sirard MA, Bilodeau S. Effects of granulosa cell co-culture on in-vitro meiotic resumption of bovine oocytes. J Reprod Fertil 1990; 89: 459-465.
7. Sirard MA, Bilodeau S. Granulosa cells inhibit the resumption of meiosis in bovine oocytes in vitro. Biol Reprod 1990; 43: 777-783.
8. van Tol HTA, Vaneijk MJT, Mummery CL, Vandenhurk R, Bevers MM. Influence of FSH and hCG on the resumption of meiosis of bovine oocytes surrounded by cumulus cells connected to membrana granulosa. Mol Reprod Dev 1996; 45: 218-224.
9. Pincus G, Enzmann EV. The comparative behavior of mammalian eggs in vitro and in vivo. J. Exp. Med. 1935; 62: 665-675.
10. Dekel N, Galiani D, Sherizly I. Dissociation between the inhibitory and the stimulatory action of cAMP on maturation of rat oocytes. Mol Cell Endocrinol 1988; 56: 115-121.
11. Mattioli M, Barboni B, Gioia L. Activation of protein kinase A and protein kinase C mediates the depolarising effect of LH in ovine cumulus-corona cells. J Endocrinol 1996; 150: 445-456.
12. Kaufman G, Dharmarajan AM, Takehara Y, Cropp CS, Wallach EE. The role of protein kinase-C in gonadotropin-induced ovulation in the in vitro perfused rabbit ovary. Endocrinology 1992; 131: 1804-1809.
13. Adashi EY, Resnick CE. 3',5'-cyclic adenosine monophosphate as an intracellular second messenger of luteinizing hormone: application of the forskolin criteria. J Cell Biochem 1986; 31: 217-228.
14. Petersson F, Andersson RG, A AsB, Hammar M. Early effects of hCG on human testicular cyclic AMP content, protein kinase activity, in-vitro progesterone conversion and the serum concentrations of testosterone and oestradiol. Int J Androl 1988; 11: 179-186.
15. Davis JS, Weakland LL, West LA, Farese RV. Luteinizing hormone stimulates the formation of inositol trisphosphate and cyclic AMP in rat granulosa cells. Evidence for

116
phospholipase C generated second messengers in the action of luteinizing hormone. Biochem J 1986; 238: 597-604.
16. Cho WK, Stern S, Biggers JD. Inhibitory effect of dibutyryl cAMP on mouse oocyte maturation in vitro. J Exp Zool 1974; 187: 383-386.
17. Dekel N. Regulation of oocyte maturation. The role of cAMP. Ann N Y Acad Sci 1988; 541: 211-216.
18. Downs S, M, D, S, A, J, E, J, J. Induction of maturation in cumulus enclosed mouse oocytes by FSH and EGF: evidence for a positive stimulus of somatic cell origin. J Exp Zool 1988; 245: 86-96.
19. Sirard MA. Developmental capacity acquisition requires exposure of uncondensed chromosomes before maturation of bovine oocytes. Bull Assoc Anat (Nancy) 1991; 75: 93-98.
20. Bilodeau S, Fortier MA, Sirard MA. The effect of adenylate cyclase stimulation on meiotic resumption and cyclic AMP content of zona-free and cumulus-enclosed bovine oocytes in vitro. J Reprod Fertil 1993; 97: 5-11.
21. Homa ST. Effects of cyclic AMP on the spontaneous meiotic maturation of cumulus-free bovine oocytes cultured in chemically defined medium. J Exp Zool 1988; 248: 222-231.
22. Aktas H, Wheeler MB, First NL, Leibfried-Rutledge ML. Maintenance of meiotic arrest by increasing [cAMP]i may have physiological relevance in bovine oocytes. J Reprod Fertil 1995; 105: 237-245.
23. Aktas H, Wheeler MB, Rosenkrans CF, Jr., First NL, Leibfried-Rutledge ML. Maintenance of bovine oocytes in prophase of meiosis I by high [cAMP]i. J Reprod Fertil 1995; 105: 227-235.
24. Mayes MA, Sirard MA. Effect of type 3 and type 4 phosphodiesterase inhibitors on the maintenance of bovine cumulus-enclosed oocytes in meiotic arrest. Biol Reprod 2002; 66: 180-184.
25. Thomas RE, Armstrong DT, Gilchrist RB. Differential effects of specific phosphodiesterase isoenzyme inhibitors on bovine oocyte meiotic maturation. Dev Biol 2002; 244: 215-225.
26. Richards JS. Hormonal control of gene expression in the ovary. Endocr Rev 1994; 15: 725-751.
27. Shi H, Segaloff DL. A role for increased lutropin/choriogonadotropin receptor (LHR) gene transcription in the follitropin-stimulated induction of the LHR in granulosa cells. Mol Endocrinol 1995; 9: 734-744.
28. Gudermann T, Birnbaumer M, Birnbaumer L. Evidence for dual coupling of the murine luteinizing hormone receptor to adenylyl cyclase and phosphoinositide breakdown and Ca2+ mobilization. Studies with the cloned murine luteinizing hormone receptor expressed in L cells. J Biol Chem 1992; 267: 4479-4488.
29. Dimino MJ, Snitzer J, Brown KM. Inositol phosphates accumulation in ovarian granulosa after stimulation by luteinizing hormone. Biol Reprod 1987; 37: 1129-1134.
30. Jones KT. Protein kinase C action at fertilization: overstated or undervalued? Rev Reprod 1998; 3: 7-12.
31. Castagna M, Takai Y, Kaibuchi K, Sano K, Kikkawa U, Nishizuka Y. Direct activation of calcium-activated, phospholipid-dependent protein kinase by tumor-promoting phorbol esters. J Biol Chem 1982; 257: 7847-7851.

117
32. Niedel JE, Kuhn LJ, Vandenbark GR. Phorbol diester receptor copurifies with protein kinase C. Proc Natl Acad Sci U S A 1983; 80: 36-40.
33. Rose-Hellekant TA, Bavister BD. Roles of protein kinase A and C in spontaneous maturation and in forskolin or 3-isobutyl-1-methylxanthine maintained meiotic arrest of bovine oocytes. Mol Reprod Dev 1996; 44: 241-249.
34. Xu Z, Garverick HA, Smith GW, Smith MF, Hamilton SA, Youngquist RS. Expression of follicle-stimulating hormone and luteinizing hormone receptor messenger ribonucleic acids in bovine follicles during the first follicular wave. Biol Reprod 1995; 53: 951-957.
35. Kawate N, Inaba T, Mori J. A quantitative comparison in the bovine of steroid and gonadotropin receptors in normally developing follicles and in follicular and luteinized cysts. Anim Reprod Sci 1990; 23: 273-281.
36. Eppig JJ, Wigglesworth K, Pendola F, Hirao Y. Murine oocytes suppress expression of luteinizing hormone receptor messenger ribonucleic acid by granulosa cells. Biol Reprod 1997; 56: 976-984.
37. Eppig JJ, Pendola FL, Wigglesworth K. Mouse oocytes suppress cAMP-induced expression of LH receptor mRNA by granulosa cells in vitro. Mol Reprod Dev 1998; 49: 327-332.
38. Fouladi Nashta AA, Waddington D, Campbell KH. Maintenance of bovine oocytes in meiotic arrest and subsequent development In vitro: A comparative evaluation of antral follicle culture with other methods. Biol Reprod 1998; 59: 255-262.
39. Kotsuji F, Kubo M, Tominaga T. Effect of interactions between granulosa and thecal cells on meiotic arrest in bovine oocytes. Journal of Reproduction and Fertility 1994; 100: 151-156.
40. Bavister BD, Leibfried ML, Lieberman G. Development of preimplantation embryos of the golden hamster in a defined culture medium. Biol Reprod 1983; 28: 235-247.
41. Rouillier P, Matton P, Sirard MA, Guilbault LA. Follicle-stimulating hormone-induced estradiol and progesterone production by bovine antral and mural granulosa cells cultured in vitro in a completely defined medium. J Anim Sci 1996; 74: 3012-3019.
42. Saumande J. Culture of bovine granulosa cells in a chemically defined serum-free medium: the effect of insulin and fibronectin on the response to FSH. J Steroid Biochem Mol Biol 1991; 38: 189-196.
43. Motlik J, Koefoed-Johnsen HH, Fulka J. Breakdown of the germinal vesicle in bovine oocytes cultivated in vitro. J Exp Zool 1978; 205: 377-383.
44. Richard FJ, Fortier MA, Sirard MA. Role of the cyclic adenosine monophosphate-dependent protein kinase in the control of meiotic resumption in bovine oocytes cultured with thecal cell monolayers. Biol Reprod 1997; 56: 1363-1369.
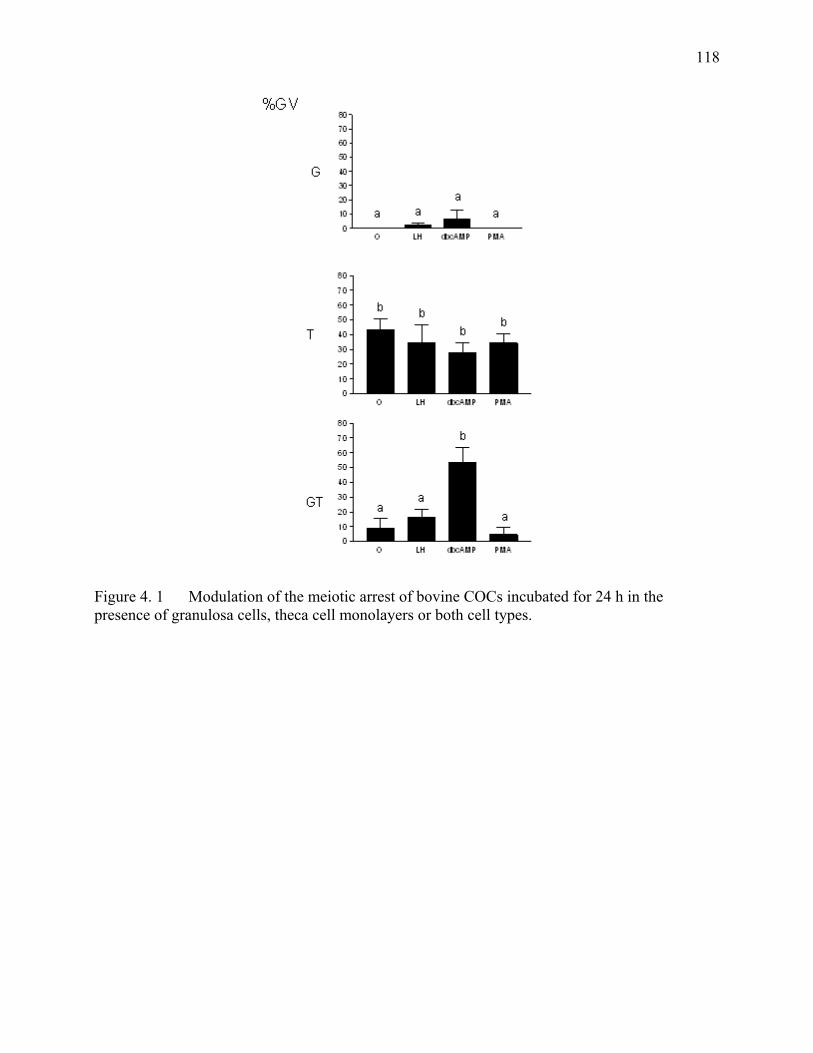
118
Figure 4. 1 Modulation of the meiotic arrest of bovine COCs incubated for 24 h in the presence of granulosa cells, theca cell monolayers or both cell types.

119
Modulation of the meiotic arrest of bovine COCs incubated for 24 h in the presence of granulosa cells, theca cell monolayers or both cell types.
Groups of 15-20 bovine COCs were incubated for 24h in the presence of granulosa cells (G), theca cell monolayers (T) or both granulosa and theca cell monolayers (GT). The x-axis represents the treatments: Control (0), 10 ng/ml LH (LH) or 10 µM PMA (PMA) or 2 mM dbcAMP (dbcAMP). The y-axis represents the percentage of oocytes arrested at the GV stage. Values are expressed as the percentage means of GV ± SEM. Data were analyzed as a 4 x 3 factorial in a complete block design with 4 replicates. Cell type and treatment were the two components of the model. Data were analyzed using the NPAR1WAY procedure of SAS. Significant treatment differences (P<0.05) were compared with the least square means using the probability of differences. Data shown were derived from 3 replicates and 681 COCs. Bars with different superscripts a and b are significantly different across the three panels at P< 0.05

CHAPTER V Partial Characterization of the Meiosis Inhibiting Factor Secreted by Bovine Theca Cell Monolayers.

121
RÉSUMÉ La culture in vitro des ovocytes bovins mène à la reprise de la méiose. Les monocouches des
cellules de la thèque retiennent les ovocytes bovins au stade de vésicule germinale. Les cellules
de la thèque obtenues par la digestion de la paroi folliculaire ont été incubées avec le milieu de
culture TCM-199 supplémenté avec 10% de sérum de veau fœtal. Une fois que les cellules de la
thèque ont formé une monocouche, le milieu a été conditionné par celles-ci pendant 48 heures.
Les ovocytes ont été sélectionnés et incubés avec les monocouches de cellules de la thèque en
groupes de 15 à 20 ovocytes par traitement. Le milieu conditionné (MC) a été traité soit à la
chaleur, aux enzymes protéolytiques, au charbon, au chloroforme ou passé sur différents filtres.
Lorsque le MC est remplacé par un MC traité à la chaleur ou au charbon, le pourcentage des
ovocytes maintenus au stade de GV demeure élevé; cependant, lorsque le MC est remplacé par
un MC traité au chloroforme, le pourcentage diminue significativement (p< 0.05). Le filtrat du
MC passé sur un filtre de 1 000 daltons maintient un pourcentage d’ovocytes au stade GV
significativement (p < 0.05) plus élevé que toutes les autres fractions. Les fractions de MC qui
ont été obtenues par la chromatographie de milieu conditionné avec une colonne C18 a produit
une fraction active. Les fractions obtenues par la chromatographie ont été analysées avec un
spectromètre de masse et les résultats ont indiqué qu’il y a des différences dans le patron des pics
chromatographiques du MC et MF. En conclusion, le facteur inhibiteur sécrété par les cellules de
la thèque dans le milieu conditionné est (1) insensible à la chaleur, aux enzymes protéolytiques et
à un traitement au charbon, (2) sensible à un traitement au chloroforme, (3) concentré dans la
fraction plus petite que 1 000 daltons du milieu conditionné et (4) les patrons de pics
chromatographiques sont differentes entre le MC et le MF. D’autres études seront nécessaires
pour l’isolation complète du facteur(s) inhibiteur(s) sécrété(s) par les cellules de la thèque.

122
ABSTRACT In vitro culture of bovine oocytes leads to meiotic resumption. Theca cell monolayers maintain
bovine oocytes at the GV stage. Theca cells obtained by digesting the follicular wall with
collagenase, were cultured in TCM-199 supplemented with 10% fetal calf serum. Once theca
cells reached confluency, medium was conditioned with the monolayers for 48 h. Groups of 15
to 20 oocytes were selected and cocultured with monolayers for 18 h. Conditioned medium
(CM) was treated with heat, proteolytic enzymes, charcoal, chloroform or cut-off filtered. When
medium was replaced either with heat-treated, proteolytic enzyme-treated or with charcoal-
treated conditioned medium, the percentage of oocytes at the GV stage remained high; however,
when medium was replaced with chloroform-treated conditioned medium, the percentage (p <
0.05) decreased. The filtrate fraction of conditioned medium lower than 1000 daltons maintained
a significantly (p < 0.05) higher percentage of oocytes at the GV stage than other fractions of
conditioned medium. Testing of the fractions of CM obtained by chromatography of CM with a
C18 column yielded an active fraction. Analysis of the chromatographic fractions with a mass
spectrometer indicated that there are differences in the pattern of chromatographic peaks of CM
and fresh medium (FM). In conclusion, the theca cell factor(s) secreted in conditioned medium
to maintain oocytes in meiotic arrest is (1) not sensitive to heat, proteolytic enzymes and charcoal
treatment, (2) sensitive to chloroform treatment, (3) lower than 1 000 daltons and (4) the patterns
of chromatographic peaks are different in CM and FM. Further work is needed to fully isolate
the factor produced by the theca cells responsible for maintaining bovine oocytes in meiotic
arrest.

123
INTRODUCTION Bovine oocytes enter meiosis early during fetal development, but they soon become arrested at
the diplotene stage of the first meiotic division and remain arrested for many months or years
until stimulated to resume meiosis at the time of ovulation, atresia or removal from the follicle.
In vivo, bovine oocytes resume meiosis following the endogenous preovulatory surge of LH [1],
whereas in vitro, bovine oocytes spontaneously resume meiosis upon removal from the follicle
[2-4]. However, the bovine oocytes must have an inside diameter or at least 110 µm to be
meiotically competent [10, 11].
During resumption of meiosis, the oocyte undergoes germinal vesicle breakdown (GVBD), which
involves a gradual chromatin condensation, disappearance of the nucleolus and disintegration of
the nuclear membrane [5]. The oocyte then progresses through the Metaphase I, Anaphase I and
Telophase I stages. After the extrusion of the first polar body, the oocyte becomes arrested at
Metaphase II until reactivated at fertilization. Bovine oocytes complete their nuclear maturation
after 24 h of in vitro maturation [6]. Bovine oocytes apparently follow the same pattern of
nuclear maturation in vivo and in vitro [7]. Bovine oocytes become committed to undergo
GVBD within a short period after their removal from the follicle [8]. After 6.5 h of culture, the
nuclear membrane of most competent bovine oocytes undergoes breakdown [6], and in vivo,
GVBD is observed to occur 4-8 h after the preovulatory LH peak [9].
Bovine oocytes require new protein synthesis during the first 8 h of culture to resume meiosis [6,
12, 13] and the first 4-5 h of culture is the critical period for the synthesis of proteins needed for
GVBD [14, 15]. Meiotically incompetent bovine oocytes lack cyclin B1, which is required for
the autoamplification of MPF and meiotic resumption [16]. Thus, bovine oocytes can be
prevented from resuming meiosis by inhibiting the synthesis of cyclin B [16, 17]. The activity of
MPF depends on active protein synthesis [18] and its state of phosphorylation. Thus, bovine
oocytes treated with inhibitors of protein synthesis or phosphorylation remain in meiotic arrest.

124
The somatic components of the follicle have been implicated in the maintenance of meiotic arrest
[19-21]. The follicle plays a role in modulating the growth, development and meiotic status of the
oocyte. Bovine oocytes within intact follicles or incubated with follicular hemisections or pieces
of the follicular wall remain in meiotic arrest [19, 20, 22]. Granulosa cells from small follicles
(2-5 mm in diameter) or sections of membrana granulosa attached to the COCs have also been
reported to exert an inhibitory effect on the meiotic process [21, 23]. Bovine granulosa cells are
important in the transmission of inhibitory signals and close contact between granulosa cells and
cumulus cells enhances the percentage of oocytes maintained at the GV stage [19, 24, 25]. The
maturation of bovine oocytes is only transiently delayed by bovine follicular fluid [8, 25, 26].
The quantity of bovine follicular fluid present in the culture medium has an inverse linear
relationship with the nuclear maturation of bovine oocytes [8, 27]. The presence of bovine
follicular fluid in the incubation medium greatly reduces the number of granulosa cells required
to maintain bovine COC in meiotic arrest [25]. Theca cells incubated as explants or monolayers
are capable of maintaining cumulus-oocyte complexes (COCs) but not denuded oocytes (DO) in
a state of reversible meiotic arrest for at least 24 h [21, 28]. The theca cell monolayers secrete the
inhibitory factor(s) into the incubation medium. The factor is active even without direct contact
between the oocyte and the theca cell monolayer [21, 28, 29]. Our hypothesis is that the factor
secreted by the theca cell monolayers, which is responsible for the maintenance of bovine
cumulus-oocyte complexes in meiotic arrest may be partially characterized using simple
biochemical tests. This study focuses in the characterization and isolation of the factor produced
by the theca cells that is responsible for maintaining bovine oocytes in meiotic arrest.
MATERIALS AND METHODS
Preparation of Theca Cell Monolayers Bovine theca cell monolayers were prepared as previously described [28]. Bovine follicles (2-
5mm) were randomly chosen from ovaries that were kept on ice in bags containing gauze soaked
with saline solution. Follicles were dissected free of stromal tissue with scissors. Theca cells
were harvested exclusively from transparent follicles containing at least 75% of the granulosa
layer intact and unexpanded COC. Selected follicles (n=12) were placed in a calcium- and

125
magnesium-free Hanks Balanced Salt Solution and cut into hemisections with a scalpel.
Granulosa cells and COCs were removed by scraping the hemisections with a fine glass loop
made from a Pasteur pipette. This technique is used to effectively isolate theca cell layers [21].
The theca layers were enzymatically digested in 15 ml of modified Hanks solution supplemented
with 2125 U of collagenase type II, 1600 U trypsin from porcine pancreas, 1 mg DNase type I
and 2 mg EDTA (all from Sigma). Enzymatic digestion took place for 1 h at 38.5°C in an
atmosphere of 95% air and 5% CO2. Theca layers were passed through a 10-ml pipette every 20
min. Following incubation, larger pieces of remaining follicular hemisections were withdrawn
from the solution. The reaction was stopped by the addition of 6 ml of washing medium,
consisting of HEPES-buffered Tyrodes medium (TLH, pH 7.4) [30] supplemented with 10% fetal
calf serum (heat-treated FCS from MEDICORP, Montreal, PQ, Canada), 0.2 mM pyruvic acid
and 50 µg/ml gentamicin sulfate. Cells were then centrifuged (10 min at 2000xg), resuspended,
and washed a second time. The first and second pellets were resuspended in 10 ml and 1 ml,
respectively, of washing medium. Theca cells were counted with a hemocytometer and seeded in
a 24-well plate (Falcon Becton Dickinson, Rutherford, NJ) at a concentration of 1.5-2.0 x 105
cells/ml. Theca cells were grown in basic culture medium (described below). The culture
medium was replenished every 48 h, and the theca cells were incubated for 5-7 days until they
formed a monolayer.
Collection of COCs Bovine ovaries at various stages of their reproductive cycle were collected at the slaughterhouse
and transported to the laboratory in saline solution kept between 30-35ºC. The saline solution
consisted of 0.9% NaCl (w:v supplemented with 100 000 IU penicillin, 100 mg streptomycin and
250 µg amphotericin B per liter (All from Sigma Chemical Company Co, St. Louis, MO)). The
contents of 1-5 mm follicles were aspirated using a 10-ml syringe and an 18-gauge needle. The
follicular contents were pooled in 50-ml conical tubes. After sedimentation, the cumulus-
enclosed oocytes (COCs) were recovered with the use of a stereomicroscope. The COCs used in
these experiments had at least 5 layers of cumulus cells (Class 3) [Mayes, 2001 #723]. The
COCs were rinsed twice in the supernatant of follicular fluid (centrifuged for 10 min at 350x g)
and then transferred to the respective treatments.

126
Denuded Oocytes Denuded oocytes (DOs) were obtained by removing the surrounding cumulus cells from COCs.
Briefly, selected COCs were vortexed for 10 min in sterile centrifuge tubes containing follicular
fluid supernatant (prepared as indicated above). Oocytes were then rinsed in follicular fluid
supernatant and those with a homogeneous cytoplasm were allocated to their respective
treatments.
Fixation of Oocytes At the end of each treatment, COCs were transferred into 2-ml centrifuge tubes containing 500 µl
of TLH and vortex-agitated for 7 min to remove the cumulus cells. Denuded oocytes were
recovered under a stereomicroscope and transferred onto a glass slide in a small drop of medium.
A Vaseline:paraffin wax mixture was used to maintain the coverslip in contact with the oocytes.
The coverslip was fixed in place with epoxy glue, and the slides were immersed in a fixative
solution (ethanol:acetic acid, 3:1) for a minimum of 24 h. Oocytes were then stained with 1%
aceto-orcein and examined for nuclear morphology with a phase contrast microscope at 100 and
400x magnification [31].
Culture media The basic culture medium used to carry out these experiments consisted of TCM-199 with
Earle’s salts (Gibco Laboratories, Grand Island, NY) and bicarbonate (Sigma Company, St. Louis
MO), 10% FCS, 0.2 mM pyruvic acid and 50 µg/ml gentamicin sulfate and was pre-exposed to
culture conditions (38.5°C, 5% CO2:95% air atmosphere with 100% humidity) for at least 2 h.
Fresh unconditioned medium (FM)
Fresh unconditioned medium (FM) is the basic culture medium prepared at the same time than
conditioned medium but never exposed to the theca cell monolayer. It was kept in the incubator
at 38.5°C, 5% CO2:95% air atmosphere with 100% humidity for 48 h. The fresh medium was
stored under the same conditions as conditioned medium

127
Theca cell conditioned medium (CM) Theca cell conditioned medium was obtained by incubating the basic culture medium with
confluent theca cell monolayers for 48 h (CM). The conditioned medium was pooled from
individual wells and centrifuged at 300xg at RT for 10 min. The supernatant was collected and
stored at -20˚C until treated. After the treatment, CM was kept at -80˚C until used. Three hours
before the coculture of COC with theca cell monolayers, culture medium was replaced with the
treated-CM. The CM was always tested to verify its efficacy in preventing bovine COCs from
resuming meiosis. The CM was considered acceptable to do further experiments when the
percentage of GV was at least 50%.
Treatment of FCS with chloroform Fetal calf serum (FCS) was treated with chloroform (10:20, v/v, serum:chloroform), vortex-
agitated (30 sec) and followed by centrifugation (300xg at room temperature (RT) for 5 min).
Chloroform treatment was repeated 4 times. Chloroform-treated serum (cFCS) was exposed to
nitrogen for at least 30 min to remove the chloroform solvent. Chloroform-treated FCS was
frozen at -20˚C until used.
Fresh unconditioned medium with chlorofom-treated FCS (FM-cFCS) FM-cFCS is the basic culture medium in which 10% FCS was substituted with 10% chloroform-
treated FCS. (cFCS) FM-cFCS was processed and stored as FM.
Conditioned medium with chloroform-treated FCS (CM-cFCS) CM-cFCSis the basic culture medium in which 10% FCS was substituted with 10% cFCS and
then conditioned by the theca cell monolayer for 48 h.
Treatment of Conditioned Medium
Enzymatic Digestion The CM-cFCS and FM-cFCS were each treated with trypsin, protease, α-chymotrypsin, DNase (2
Units/µl), RNAse (10 Units/µl) or catalase. After each treatment, the media were heated to
100˚C for 5 min to inactivate the enzymes, then cooled and stored.

128
Dialysis The CM-cFCS and FM-cFCS were dialyzed for 24 h at 4°C using a cellulose ester dialysis
membrane with a pore size of either 14 000 or 3 500 daltons. The dialyzed and non-dialyzed
fractions were collected and roto-evaporated. The samples were reconstituted to the original
osmolarity and stored at – 80°C until needed.
Treatment of CM-cFCS with solvents The CM-cFCS was treated 4 times with chloroform (10:20, v:v, CM:chloroform), vortex-agitated
(30 sec) and followed by centrifugation (300xg at room temperature for 5 min). Chloroform-
treated CM-cFCS was exposed to nitrogen for at least 30 min to remove chloroform solvent. In
order to evaluate the effect of the organic phase on nuclear maturation, the resuspension medium
was supplemented with the organic phase of Chloroform-treated CM-cFCS. Chloroform was
collected from the extraction of CM-cFCS and evaporated by heating the test tube in a boiling
water bath. Different concentrations of the remaining chloroform (organic phase) were added to
the resuspension medium, TCM-199 supplemented with 0.3% BSA (fraction V, Sigma), 0.2 mM
pyruvic acid, and 50 µg/ml gentamicin sulfate. Resuspension medium was exposed to nitrogen
until the organic phase was evaporated. The culture medium was also similarly treated with
acetone, 95% ethanol, methanol or benzene. Furthermore, the CM-cFCS was extracted with
chloroform under acidic, neutral or basic conditions.
Heat treatment of CM The CM and CM-cFCS were heat-treated in a boiling water bath either at 100˚C for 5 min or at
56˚C for 1 h in a water bath to evaluate whether the theca cell factor(s) was heat-stable. The CM
and CM-cFCS were cooled and then stored at – 80°C until needed.
NMWL Fractioning Both CM-cFCS and FM-cFCS were filtered through Ultrafree Millipore filters to obtain the
respective cut-off fractions. Filters had a Nominal Molecular Weight Limit (NMWL) of 100, 30,
10 and 1 kDa. The cut-off filtered medium was diluted 1.5 times with a solution of TCM-199
with 0.2 mM pyruvic acid, and 50 µg/ml gentamicin sulfate. The fractioned medium was stored
at – 80°C until needed.

129
Dextran : Charcoal The CM was treated twice with dextran:charcoal (0.12 g/l: 12 g/l, Sigma) at 4˚C [32]. The first
treatment was overnight and the second for 3 h. Charcoal was removed by centrifugation (20
000xg at 4˚C for 15 min) followed by filtration through a series of 0.45-µm filter and finally
through a 0.22-µm filter (Millipore, Bedford, MA). These treatments were either followed or
preceded by heating the CM at 100˚C for 5 min in a boiling water bath.
Sep Pak Plus minicolumns Reversed-phase chromatography of CM-cFCS was carried out using Sep-Pak Plus minicolumns.
The sorbents used were C18, aminopropyl (NH2), Diol and cyanopropyl (CN). The minicolumns
were rinsed sequentially with solutions of 100% acetonitrille (ACN), 80% ACN and 0.1%
trifluoroacetic acid (TFA) and finally with 0.1% TFA solution. The CM-cFCS sample (200 ml)
was lyophyllized and reconstituted with water (5 ml) to a concentration of 40x. The concentrated
sample was loaded and the unretained fraction was collected. The minicolumn was rinsed with
0.1% TFA in water and the rinse fraction was collected. The sample was eluted with 80% ACN
+ 20% water with 0.1% TFA. The retained and eluted fractions were collected, lyophylized,
reconstituted and tested. Normal phase chromatography was used for the Sep Pak Plus
minicolumns with the following sorbents: Florisil, Alumina A, B and N, NH2, CN and Diol. The
minicolumn was rinsed with hexane, loaded with the CM-cFCS sample, rinsed with hexane and
eluted with water. The eluted fractions were lyophylized, reconstituted with TCM-199 and
adjusted to an osmolarity of approximately 290 mMol per kg.
Preparation of CM and FM Mixes Conditioned medium with chloroform-treated FCS (CM-cFCS) was collected, centrifuged and
frozen at –80°C on a weekly basis. Aliquots of pooled samples of CM-cFCS from a given week
were tested to evaluate the ability of the CM-cFCS to maintain bovine COCs in meiotic arrest.
Samples with the ability to maintain at least 50% of the oocytes in meiotic arrest were pooled
(CM-cFCS mix) and fractioned using Centricon Plus-80 centrifugal filter units with a 5000
NMWL. The CM was centrifuged at 3000xg at 4°C until the initial sample volume of 80 mL was
reduced to approximately 300 µl. The fraction smaller than 5000 NMWL was lyophylized and

130
concentrated. The sample was then centrifuged. The precipitate and supernatant were separated,
reconstituted and tested for the presence of the inhibitory activity. The inhibitory activity was
found in the precipitate fraction. Fresh unconditioned medium with chlorofom-treated FCS (FM-
cFCS) was prepared by pooling together the samples of FM-cFCS collected weekly. The
samples were processed as the conditioned medium.
Chromatography Both CM-cFCS mix and FM-cFCS mix were tested to evaluate their ability to maintain meiotic
arrest. The CM-cFCS mix (6 mL) was lyophylized and reconstituted to 200 µl water and 50 µl
ethanol, (6000 µl / 250 µl yields a mix concentrated 24 times). The reconstituted mix was
centrifuged and the supernatant and precipitate separated. Liquid chromatography of the
supernatant was carried out in a Waters 2690 LC system using a Zorbax C18 column (7.5 x 4.6
mm) and eluted with a linear gradient of a mixture of ethanol, 10% ACN and 1 mM trifluoracetic
acid.
Chromatography A: Fractions were collected every 3 minutes and stored at – 80°C until needed.
The chromatograms were represented in terms of mass spectrometer. The same procedure was
carried out with FM-cFCS mix. The fractions of CM-cFCS mix were reconstituted and adjusted
to an osmolarity of approximately 290 mMol per kg. Chromatography B: the sample was
acidified with 0.1% formic acid and the fractions were collected every minute from 0.5 to 7.5 min
after injecting the sample. The fractions of CM-cFCS mix were reconstituted and adjusted to an
osmolarity of approximately 290 mMol per kg.
EXPERIMENTAL DESIGN In all of the following experiments, the treated medium was transferred to the theca cell
monolayer approximately 3 h before transferring the COCs to the theca cell monolayer. COCs
were incubated for 18 h (unless otherwise indicated) and then fixed as previously described.

131
Exp 1: Effect of medium replacement on the nuclear maturation of COCs incubated with theca cell monolayers. In experiment 1, the theca cell conditioned medium (CM) or conditioned medium supplemented
with choroform-treated FCS (CM-cFCS) were replaced with either FM or conditioned medium
preconditioned by a different theca cell monolayer (PCM).
Exp 2: Effect of heat-treated CM on the nuclear maturation of oocytes incubated with theca cell monolayers. Experiment 2 evaluated if the inhibitory factor produced by the theca cell monolayers is heat
stable. The conditioned media (CM and CM-cFCS) were heat-treated in a boiling water bath
either at 100°C for 5 min or at 56°C for 1 h.
Exp 3: Effect of dextran:charcoal-treated CM Experiment 3 evaluated whether dextran:charcoal (DC) treatment of the CM removes the
inhibitory factor produced by the theca cell monolayers. Conditioned medium was either left
untreated or treated once (1X) or twice (2X) with dextran charcoal. The DC treatment of CM
was followed or preceded by heating the CM at 100°C for 5 min.
Exp 4: Effect of medium supplemented with cut off fractions of CM In experiment 4, the culture medium was supplemented with different cut-off fractions of CM
and FM (100, 30, 10 and 1 kDa).
Exp 5 Effect of enzymatic treatment of CM-cFCS and FM-cFCS In experiment 5a, CM-cFCS and FM-cFCS were each treated with trypsin and then protease or
vice versa. The media were enzymatically treated for 1h at 37°C. After each treatment, the
enzymes were inactivated by heating the medium in a water bath to 95°C for 10 min.
Additionally, the same protocol was used to test the effect of treating the CM-cFCS and FM-
cFCS with α-chymotrypsin and proteinase K.
In experiment 5b, CM-cFCS and FM-cFCS were first treated for 1 h at 37°C with DNase I or
RNase, then treated with protease (1 h at 37°C) and finally heat inactivated as in experiment 5a.

132
Exp 6 Effect of dialysis treatment of CM-cFCS and FM-cFCS In experiment 6, the dialyzed and non-dialyzed fraction of CM-cFCS and FM-cFCS were
lyophyllized and reconstituted to the original osmolarity. The oocytes were fixed after 12 h of
treatment.
Exp 7 Effect of chloroform treatment of CM and CM-cFCS Experiment 7 studied whether the theca cell factor(s) were sensitive to chloroform treatment.
The CM and CM-cFCS were treated with chloroform to test the ability of the conditioned
medium to maintain COCs in meiotic arrest. The chloroform-treated conditioned medium
containing chloroform-treated FCS will be abbreviated (cCM-cFCS).
Exp 8 Effect of the organic phase of cCM-cFCS In experiment 8, the organic fraction of cCM-cFCS was tested at different concentrations on
TCM-199 supplemented with 0.3% BSA (fraction V, Sigma), 0.2 mM pyruvic acid and 50 µl
gentamicin.
Exp 9 Effect of different chromatographic fractions of CM-cFCS. The eluted and unretained fractions of CM-cFCS were tested for the C18, aminopropyl (NH2),
Diol and cyanopropyl (CN) Sep Pak Plus minicolumns. The eluted fractions of CM-cFCS were
tested for the Florisil, Alumina A, B and N, NH2, CN and Diol. Sep Pak Plus minicolumns. The
individual fractions were tested in the presence of theca cell monolayers and COCs to evaluate
their ability to maintain the COCs in meiotic arrest.
STATISTICAL ANALYSIS Oocytes were classified as germinal vesicle stage (GV stage) representing those in meiotic arrest,
as intermediate stage (including germinal vesicle breakdown [GVBD], early condensation, and
metaphase-I [M-I]) and as mature stage (anaphase-I, telophase-I and metaphase-II [M-II]).
Stained oocytes presenting abnormal chromatin configuration or absence of chromatin were
considered degenerated. Values are expressed as means of percentages ± SE. In all experiments,
results were analyzed by a two-way ANOVA. Treatment and replicate were the 2 components of

133
the model. When ANOVA indicated a significant effect of treatment (P < 0.05), treatment
differences were compared by Duncan's multiple range test.
RESULTS Each treatment was assessed by evaluating the percentage of oocytes maintained in meiotic arrest
after coculturing COC with theca cell monolayers. This thesis corroborates the experiments
(1,2,3,4,7 and 8), which were originally carried out by Francois Richard during his Ph.D.
program. This chapter will be submitted for publication as a collaborative effort of both authors.
In experiment 1, bovine COCs remained in meiotic arrest when incubated with a theca cell
monolayer and conditioned medium (CM). On the other hand, the number of bovine COCs in
meiotic arrest significantly decreases (P < 0.05) when the CM is replaced by fresh medium (FM)
(78.9 ± 1.8 vs. 12.8 ± 6.7). However, bovine COCs remain in meiotic arrest even after replacing
the CM with medium conditioned by a different theca cell monolayer (PCM) (Figure 5.1a).
Similar results were observed when the conditioned medium contained chloroform-treated FCS
(CM-cFCS) (Figure 5.1b). These results indicate that the incubation medium must be
conditioned for 48 h by the theca cell monolayers to maintain bovine oocytes in meiotic arrest.
The CM can be replaced from one monolayer to another without affecting its ability to maintain
bovine COCs in meiotic arrest. Thus the theca cell inhibitory factor(s) is secreted into the
conditioned medium. Furthermore, CM-cFCS also maintains bovine COCs in meiotic arrest.
In experiment 2, heat-treatment of CM at 100ºC for 5 min or at 56ºC for 60 min did not
compromise the ability of the CM to maintain bovine COCs in meiotic arrest. Heat-treatment of
CM resulted in less than 4% of degenerated oocytes (data not shown). Furthermore, preliminary
experiments showed that keeping the CM at 38ºC for more than 7 days does not affect its
inhibitory ability (data not shown). The inhibitory activity of both the CM (Figure 5.2a) and CM-
cFCS (Figure 5.2b) were not affected by the heat treatment.
In experiment 3, the extraction of CM with dextran-charcoal did not decrease the percentage of
oocytes at the GV stage compared to control (64.6 ± 2.1 vs 63.9 ± 2.8, respectively) (Figure 5.3).
In fact, a double extraction of the CM with dextran-charcoal did not decrease the percentage of

134
oocytes at the GV stage when compared to controls 65.0 ± 5.0 vs. 64,6 ± 2,1, respectively). Heat
treatment (100ºC for 5 min) before or after the double dextran-charcoal extraction of the CM did
not have a significant effect on the inhibitory activity. Less than 2% of the oocytes incubated in
the dextran-charcoal and/or heat treated CM degenerated (data not shown).
Experiment 4 showed that the inhibitory activity of CM was recovered in the filtrate of the 1000
NMWL cut-off filter (Figure5.4a). In fact, the 1000 NMWL cut-off fraction and the non-
fractioned (whole) CM were equally effective in maintaining bovine oocytes in meiotic arrest
(61.0 ± 1.5 vs. 66.5 ± 5.8, respectively). Whole fresh unconditioned medium (FM) or the
individual cut-off fractions of FM failed to maintain bovine oocytes in meiotic arrest (Figure
5.4b). Consistently, the percentage of oocytes arrested at the GV stage was significantly lower
(p < 0.05) for each FM fraction when compared to the respective fraction of CM. Less than 4%
of oocytes were degenerated (data not shown).
In experiment 5a, treatment of the CM-cFCS with trypsin and protease (60% GV) did not affect
the ability of CM (57.14% GV) to maintain bovine COCs at the GV stage. The same lack of
effect on the inhibitory activity was observed after treatment with α-chymotrypsin and proteinase
K (57.89% GV) compared to controls (57.89% GV). Bovine COCs incubated with FM-cFCS or
FM-cFCS treated with the same enzymes resumed meiosis.
In experiment 5b, CM-cFCS treated with DNase or RNase and then digested with protease
retained the inhibitory effect on meiotic resumption (47.06%, GV 55.56% and GV 61.11% GV;
Dnase, Rnase and controls respectively). The same treatment applied to the FM-cFCS did not
prevent the oocytes from resuming meiosis (16.67% GV,15% GV and 16.67% GV; DNase,
RNase and controls, respectively).
In experiment 6, the fractions of CM-cFCS and FM-cFCS collected after dialysis (retentate and
dialyzed fractions) failed to maintain bovine COCs in meiotic arrest (6.25% GV, 0% GV and
52,63% GV; retentate, dialyzed and control respectively).
In experiment 7, the percentage of COCs in meiotic arrest significantly decreased compared (p <
0.05) when the COCs were incubated in Chloroform-treated CM (Chloro-CM) when compared to

135
control (4.8 ± 3.1 vs. 61.2 ± 9.9% GV ± SEM, respectively) (Figure 5.5a). Similar results were
obtained when the cCM-cFCS was tested (Figure 5.5b). Treatment of CM with chloroform
results in the formation of three different layers. The uppermost layer is the chloroform-treated
CM, the intermediate layer has a gel-like consistency and the bottom layer is the chloroform.
The COCs resumed meiosis after being incubated with any of the three different layers. The
inhibitory activity of CM was removed or destroyed by the chloroform-treatment. Solubilization
of the intermediate layer with solvents such as acetone, ethanol or DMSO failed to liberate the
inhibitory factor and to maintain the bovine COCs at the GV stage. Less than 2% of oocytes
were degenerated (data not shown).
In experiment 8, Bovine COCs incubated in medium containing the organic fraction of
chloroform-treated conditioned medium containing chloroform-treated FCS (cCM-cFCS)
resumed meiosis (Figure 5.6). Less than 3% of oocytes were degenerated (data not shown).
Increasing the concentration of the extracted organic fraction in the incubation medium did not
significantly increase the percentage of oocytes at the GV stage to control levels (27.6 ± 4.3 vs.
68.5 ± 1.8% GV ± SEM, respectively).
In experiment 9, the eluted and unretained fractions of CM-cFCS, obtained by reverse-phase
chromatography, were tested for the presence of the inhibitory activity. The eluted fractions of
the C18, aminopropyl (NH2), Diol and cyanopropyl (CN) minicolumns failed to extract the active
factor from the CM. The active factor was present in the unretained fraction of the NH2
minicolumn. Furthermore, the eluted fractions of the Florisil, Alumina A, B and N, NH2
minicolumns obtained by normal phase chromatography did not maintain bovine COCs at the GV
stage.
The CM-cFCS mix and the eluted fractions of CM-cFCS mix were tested for the presence of the
inhibitory factor produced by the theca cell monolayers. The COCs incubated with the first
fraction (0-3 min elution time) remained in meiotic arrest, whereas those COCs incubated with
the other fractions resumed meiosis (Figure 5.7a). The chromatographic conditions were then
modified: the sample was acidified and the fractions were taken every minute for 6.5 min. These
modifications were carried out to obtain a better separation of the active factor. Testing of the

136
fractions revealed that the Fraction 1 (0.5-1.5 min) contained the active factor (Figure 5.7b and
Figure 5.8).
DISCUSSION Bovine cumulus oocyte complexes (COCs) incubated in vitro resume meiosis [2]. Theca cell
monolayers maintain bovine COCs in meiotic arrest, thus preventing the spontaneous resumption
of meiosis [28, 33]. Theca cells play an important role in the structural integrity of the ovarian
follicle and the control and maintenance of ovarian function. The role of theca cells in
maintaining meiotic arrest may be different according to species. Theca cells play an important
role in maintaining bovine oocytes in meiotic arrest [21, 28, 33]. However, mouse oocytes found
in growth differentiation factor 9 deficient females grow and remain in meiotic arrest even
though the follicles lack the theca cells [34]. This suggests that mouse theca cells are not
exclusively implicated in the maintenance of meiotic arrest.
Previous studies have shown that theca cell monolayers require specific conditions to produce the
factors responsible for maintaining bovine COCs in meiotic arrest [33]. Theca cells monolayers
must condition the culture medium for at least 48 h and require culture media containing 10%
fetal calf serum (FCS). The supplementation of incubation medium with serum facilitates the
maintenance of cells in culture, but it also interferes with the androgen production and hormone
responsiveness of theca cells [35]. However, replacing the fetal calf serum with 10% estrous cow
serum, 1% fetal calf serum or 0.3% BSA leads to a significant decrease in the percentage of
oocytes that remain at the GV stage [33]. Clearly, fetal calf serum contains certain factors
required by the theca cell monolayer to maintain bovine COCs in meiotic arrest. The factors are
extracted with charcoal but unaffected by chloroform. The fraction of FCS with the molecular
weight greater than 100 kDA contains the required factor(s). It has been proposed that inhibitors
found in follicular fluid may be derived from the larger serum molecules within an intermediate
cell type responsible for producing the inhibitory factor(s) [36].
Multiple studies have reported on the interactions that take place in vitro between the theca cell
secretions and granulosa cells [35, 37-41]. Bovine granulosa cell monolayers cultured with
conditioned medium transiently inhibit nuclear maturation of oocytes [42]. However, this effect

137
is significantly lower using conditioned medium without the monolayers and none of oocytes is
maintained in meiotic arrest using fresh medium [42].
Numerous studies have suggested that close contact between granulosa cells and COC is
necessary to promote meiotic arrest [19, 23, 27, 43]. The ability of hamster oocytes to remain at
the GV stage depends on the extent of contact between the grafted oocyte and the membrane
granulosa within the complete follicle [44]. Cumulus cells must be present to process and/or
transfer the theca cell inhibitory factor to the oocyte since the theca cell inhibitory factor does not
work on denuded bovine oocytes. The inhibitory factor is active even in the absence of direct
contact between the theca cell and COCs suggesting that the inhibitory factor is secreted into the
culture medium [28]. . Theca cell monolayers must be present or the conditioned medium
replaced every 4 h to maintain bovine COCs in meiotic arrest [33]. The presence of theca cell
monolayers is required to promote the inhibitory effect of CM. Coculturing COCs in fresh
unconditioned medium (FM) even in the presence of theca cell monolayers significantly (p <
0.05) decreases the percentage of oocytes at the GV stage (Figure 5.1). Furthermore, the
inhibitory factors must be present at the onset of in vitro maturation because it is known that
bovine oocytes become committed to resuming meiosis shortly after their removal from the
follicle [8]. This might explain why the incubation medium must be conditioned to effectively
maintain bovine oocytes at the GV stage.
Previous reports have suggested that the inhibitory factor(s) secreted into the culture medium are
unstable [42] or highly labile [21, 24, 45]. The present study confirms that the factor secreted by
the theca cell factor is not degraded by heat treatment (Richard, 1997) (Figure 5.2). In addition,
The theca cell factor is still biologically active after freezing and thawing and after prolonged
periods of incubation at 38.5 ºC (data not shown).
Dextran:charcoal does not remove the inhibitory factors present in the theca cell monolayer
conditioned medium (Figure 5.3). Charcoal treatment is used to extract greater than 98% of
estradiol-17β, testosterone and progesterone from bovine follicular fluid [46] and small
hydrophobic molecules [47]. Thus the factor(s) produced by the theca cell monolayer is probably
not a steroid, small hydrophobic molecule or other charcoal-extractable substances. These

138
observations would suggest that the product effect is labile in presence of COCs but not on its
own.
The oocyte maturation inhibitor (OMI) found in porcine follicular fluid inhibits the meiotic
resumption of rodent [48-50] and porcine oocytes [51, 52]. The OMI of porcine follicular fluid
which requires the presence of cumulus cells is not extracted by charcoal, is heat stable [50] and
corresponds to a molecular weight of less than 2 000 [51]. This is in agreement with our results
in the bovine since the inhibitory factor produced by the theca cell monolayer is found in the cut-
off filtered fraction of CM smaller than 1000 daltons (Figure 5.4). Other studies also support that
the maturation inhibitory factor is a low molecular weight product. The molecular weight of the
bovine follicular fluid factor(s) is between 1 000 and 10 000 [49], less than 10 000 [48], which
respectively inhibits hamster, rat oocytes. A factor from bovine granulosa cells that inhibits
mouse oocyte maturation is estimated to be less than 6 000 daltons [53]. In addition, the meiotic
arresting substance from porcine granulosa cells was found to have a molecular weight range
between 1 450 and 3 000 [54, 55]. Linoleic acid (18:2) a polyunsaturated fatty acid present in
high quantities in bovine follicular fluid (bFF) has been reported to inhibit the nuclear maturation
of porcine and bovine denuded oocytes [60]. However, other studies have shown that the meiotic
arresting substance is insoluble in acetone and ether [55], or ether extraction of the bovine
follicular fluid inhibitor is without any effect [49]. This suggests that the factor produced by the
theca cells is not linoleic acid or a closely related lipid.
Hypoxanthine inhibits the maturation of mouse oocytes and it is the predominant low molecular
weight component of porcine follicular fluid [56]. Hypoxanthine is also found in bovine
follicular fluid [57]. However, the inhibitory factor secreted by the theca cell monolayer is not
hypoxanthine. Hypoxanthine has a transient effect in preventing meiotic resumption of bovine
oocytes [8].
The inhibitory action of OMI is inactivated by trypsin [51]. On the other hand, treatment of the
theca cell inhibitory factor with trypsin and protease or α-chymotrypsin and proteinase K does
not remove the inhibitory activity of CM. This suggests that the inhibitory factor produced by the
theca cell monolayer is not a protein or a large peptide. The inhibitory activity could potentially
be a small peptide that is not degraded by the action of the proteolytic enzymes tested so far.

139
Furthermore, DNase or RNase treated CM continues to inhibit the meiotic resumption of bovine
oocytes, thus suggesting that the inhibitory factor is probably not a large fragment of DNA or
RNA. Due to the lack of effect of DNase and RNase on the inhibitory activity and the fact that
cyclic nucleotides play an important role in oocyte maturation, it is possible that the inhibitory
factor might be a nucleotide. The theca cell factor(s) described here seems similar to OMI but
different than hypoxanthine.
Chloroform is routinely used to extract lipids. Extraction of lipids with chloroform is preferred
over extraction with ether since chloroform is considered to be less damaging to lipids.
Chloroform may also extracts non-lipid materials such as urea, amino acids and even sugars [58].
Treatment of CM with chloroform yields three layers: the top layer is the chloroform-treated CM,
then an intermediate gel-like layer and then a final layer of chloroform. All of these layers have
been tested, but the inhibitory action is not completely restored in any of the 3 fractions and the
COCs resume meiosis. The factor produced by the theca cell monolayer is either sensitive to
chloroform extraction or modified to an inactive form (Figure 5.6). Even after solubilization of
the gel-like layer, the inhibitory activity is no longer present (Figure 5.6). A different study has
reported that the theca cell factor produced by theca cell hemisections is not degraded by
chloroform treatment [59].
Normal or reverse phase chromatography with the different Sep Pak minicolumns did not yield
an eluted fraction containing the inhibitory activity. The goal was to be able to remove unwanted
components from the CM and to obtain a fraction with higher concentrations of the inhibitory
factor. Probably the inhibitory activity rested bound to the sorbent and thus unavailable. Dialysis
also proved to be ineffective to purify the sample.
Chromatography of the conditioned medium with a C18 column yielded several fractions of CM.
Testing of these fractions indicated that the inhibitory factor is found eluted from the column in
the first fraction (Figure 5.7a). Analysis with a mass spectrometer indicated that there are
differences in the pattern of chromatographic peaks of CM and FM (Figure 5.8).
In conclusion, the inhibitory factor produced by the theca cell factor(s) is (1) not sensitive to heat,
proteolytic enzymes and charcoal treatment, (2) sensitive to chloroform treatment, and, (3)

140
smaller than 1 000 dalton and (4) the conditioned medium has a chromatographic pattern
different than the pattern of FM. Other investigations are necessary to determine the nature of the
inhibitory factor.
ACKNOWLEDGMENTS The authors gratefully acknowledge the assistance of Mr. Patrick Bélanger for performing the
chromatographic analysis and Dr. François Richard for the critical review of the manuscript.
REFERENCES 1. Hyttel P, Callesen H, Greve T. Ultrastructural features of preovulatory oocyte maturation
in superovulated cattle. J Reprod Fertil 1986; 76: 645-656. 2. Pincus G, Enzmann E. The comparative behavior of mammalian eggs in vitro and in vivo.
J. Exp. Med. 1935; 62: 665-675. 3. Edwards R. Maturation in vitro of mouse, sheep, cow, pig, rhesus monkey and human
ovaries. Nature 1965; 208: 349-351. 4. Chang M. The maturation of rabbit oocytes in culture and their maturation, activation,
fertilization, and subsequent development in the fallopian tubes. J Exp Zool 1955; 128: 378-405.
5. Kubelka M, Motlik J, Fulka JJ, Prochazka R, Rimkevikova Z, Fulka J. Time sequence of germinal vesicle breakdown in pig oocytes after cycloheximide and p-aminobenzamidine block. Gam. Res. 1988; 19: 423-431.
6. Sirard MA, Florman HM, Leibfried-Rutledge ML, Barnes FL, Sims ML, First NL. Timing of nuclear progression and protein synthesis necessary for meiotic maturation of bovine oocytes. Biol Reprod 1989; 40: 1257-1263.
7. Hyttel P, Xu KP, Smith S, Greve T. Ultrastructure of in-vitro oocyte maturation in cattle. J Reprod Fertil 1986; 78: 615-625.
8. Sirard MA, First NL. In vitro inhibition of oocyte nuclear maturation in the bovine. Biol Reprod 1988; 39: 229-234.
9. Kruip T, Cran D, Van Beneden T, Dieleman S. Structural changes in bovine oocytes during final maturation in vitro. Gam.Res 1983; 8: 29-47.
10. Fair T, Hyttel P, Greve T. Bovine oocyte diameter in relation to maturational competence and transcriptional activity. Mol Reprod Dev 1995; 42: 437-442.
11. Hyttel P, Fair T, Callesen H, Greve T. Oocyte growth, capacitation and final maturation in cattle. Theriogenology 1997; 47: 23-32.
12. Tatemoto H, Horiuchi T, Terada T. Effects of cycloheximide on chromatin condensations and germinal vesicle breakdown (GVBD) of cumulus-enclosed and denuded oocytes in cattle. Theriogenology 1994; 42: 1141-1148.
13. Kastrop PMM, Hulshof SCJ, Bevers MM, Destree OHJ, Kruip TAM. The effects of a-amanitin and cycloheximide on nuclear progression, protein synthesis, and

141
phosphorylation during bovine oocyte maturation in vitro. Mol Reprod Dev 1991; 28: 249-254.
14. Simon M, Jilek F, Fulka J, Jr. Effect of cycloheximide upon maturation of bovine oocytes. Reprod Nutr Dev 1989; 29: 533-540.
15. Motlik J, Lie B, Shioya Y. Two sensitivity levels of cattle oocytes to puromycin. Biol Reprod 1990; 43: 994-998.
16. Levesque JT, Sirard MA. Resumption of meiosis is initiated by the accumulation of cyclin B in bovine oocytes. Biol Reprod 1996; 55: 1427-1436.
17. Wu B, Ignotz G, Currie WB, Yang X. Dynamics of maturation-promoting factor and its constituent proteins during in vitro maturation of bovine oocytes. Biol Reprod 1997; 56: 253-259.
18. Hunter AG, Moor RM. Stage dependent effects of inhibiting RNA and protein synthesis on meiotic maturation of bovine oocytes in vitro. J Dairy Sci 1987; 70: 1646-1651.
19. De Loos FA, Zeinstra E, Bevers MM. Follicular wall maintains meiotic arrest in bovine oocytes cultured in vitro. Mol Reprod Dev 1994; 39: 162-165.
20. Carbonneau G, Sirard MA. Influence of follicular wall on meiotic resumption of bovine oocytes when cultured inside or outside hemisections. J Reprod Dev 1994; 40: 125-132.
21. Richard FJ, Sirard MA. Effects of follicular cells on oocyte maturation. I: Effects of follicular hemisections on bovine oocyte maturation in vitro. Biol Reprod 1996; 54: 16-21.
22. Sirard MA, Coenen K. The co-culture of cumulus-enclosed bovine oocytes and hemisections of follicles: Effects on meiotic resumption. Theriogenology 1993; 40: 933-942.
23. van Tol HTA, Vaneijk MJT, Mummery CL, Vandenhurk R, Bevers MM. Influence of FSH and hCG on the resumption of meiosis of bovine oocytes surrounded by cumulus cells connected to membrana granulosa. Mol Reprod Dev 1996; 45: 218-224.
24. Sirard MA, Bilodeau S. Effects of granulosa cell co-culture on in-vitro meiotic resumption of bovine oocytes. J Reprod Fertil 1990; 89: 459-465.
25. Sirard MA, Bilodeau S. Granulosa cells inhibit the resumption of meiosis in bovine oocytes in vitro. Biol Reprod 1990; 43: 777-783.
26. Leibfried L, First NL. Effect of bovine and porcine follicular fluid and granulosa cells on maturation of oocytes in vitro. Biol Reprod 1980; 23: 699-704.
27. Sirard MA, Coenen K, Bilodeau S. Effect of fresh or cultured follicular fractions on meiotic resumption in bovine oocytes. Theriogenology 1992; 37: 39-58.
28. Richard FJ, Sirard MA. Effects of follicular cells on oocyte maturation. II: Theca cell inhibition of bovine oocyte maturation in vitro. Biol Reprod 1996; 54: 22-28.
29. van Tol HT, Bevers MM. Theca cells and theca-cell conditioned medium inhibit the progression of FSH-induced meiosis of bovine oocytes surrounded by cumulus cells connected to membrana granulosa. Mol Reprod Dev 1998; 51: 315-321.
30. Bavister BD, Leibfried ML, Lieberman G. Development of preimplantation embryos of the golden hamster in a defined culture medium. Biol Reprod 1983; 28: 235-247.
31. Motlik J, Koefoed-Johnsen HH, Fulka J. Breakdown of the germinal vesicle in bovine oocytes cultivated in vitro. J Exp Zool 1978; 205: 377-383.

142
32. Lussier JG, Carruthers TD, Murphy BD. Effects of bovine follicular fluid and partially purified bovine inhibin on FSH and LH release by bovine pituitary cells in culture. Reprod Nutr Dev 1993; 33: 103-119.
33. Richard FJ. Effect de différentes composantes cellulaires du follicule ovarien sur le contrôle de la maturation nucléaire chez les ovocytes de bovin. Ste. Foy, Québec, Canada: Université Laval; 1997 Thesis.
34. Dong JW, Albertini DF, Nishimori K, Kumar TR, Lu NF, Matzuk MM. Growth differentiation factor-9 is required during early ovarian folliculogenesis. Nature 1996; 383: 531-535.
35. Roberts AJ, Skinner MK. Hormonal regulation of thecal cell function during antral follicle development in bovine ovaries. Endocrinology 1990; 127: 2907-2917.
36. Jagiello G, Graffo J, Ducayen M, Prosser R. Further studies of inhibitors of in vitro mammalian oocyte maturation. Fertil Steril 1977; 28: 476-481.
37. Roberts AJ, Skinner MK. Transforming growth factor a nd b differentially regulate growth and steroidogenesis of bovine thecal cells during antral follicle development. Endocrinology 1991; 129: 2041-2048.
38. Skinner MK, Keski-Oja J, Osteen KG, Moses HL. Ovarian thecal cells produce transforming growth factor-b which can regulate granulosacell growth. Endocrinology 1987; 121: 786-792.
39. Skinner MK. Transforming growth factor production and action in the ovarian follicle: Theca cell-granulosa cell interactions. In: Hirshfield AN (ed.) Growth factors in the ovary. New York: Plenum Press; 1989: 141-150.
40. Skinner MK, Coffey RJ. Regulation of ovarian cell growth through the local production of transforming growth factor-a by theca cells. Endocrinology 1988; 123: 2632-2638.
41. Bendell JJ, Lobb DK, Chuma A, Gysler M, Dorrington JH. Bovine theca cells secrete factor(s) that promote granulosa cell proliferation. Biol Reprod 1988; 38: 790-797.
42. Kotsuji F, Kubo M, Tominaga T. Effect of interactions between granulosa and thecal cells on meiotic arrest in bovine oocytes. Journal of Reproduction and Fertility 1994; 100: 151-156.
43. Foote WD, Thibault C. Recherches experimentales sur la maturation in vitro des ovocytes de truie et de veau. Ann Biol Anim Biochim Biophys 1969; 9: 329-349.
44. Racowsky C, Baldwin KV. In vitro and in vivo studies reveal that hamster oocyte meiotic arrest is maintained only transiently by follicular fluid, but persistently by membrana/cumulus granulosa cell contact. Dev Biol 1989; 134: 297-306.
45. Sirard MA. Temporary inhibition of in vitro meiotic resumption by adenylate cyclase stimulation in immature bovine oocytes. Theriogenology 1989; 31: 257.
46. Lussier JG, Carruthers TD. Endocrine and superovulatory response in heifers pre-treated with FSH or bovine follicular fluid. Theriogenology 1989; 31: 779-794.
47. Downs SM, Eppig JJ. Cyclic adenosine monophosphate and ovarian follicular fluid act synergistically to inhibit mouse oocyte maturation. Endocrinology 1984; 114: 418-427.
48. Tsafriri A, Channing CP, Pomerantz SH, Lindner HR. Inhibition of maturation of isolated rat oocytes by porcine follicular fluid. J Endocrinol 1977; 75: 285-291.
49. Gwatkin RB, Andersen OF. Hamster oocyte maturation in vitro: inhibition by follicular components. Life Sci 1976; 19: 527-536.

143
50. Tsafriri A, Dekel N, Bar-Ami S. The role of oocyte maturation inhibition in follicular regulation of oocyte maturation. Journal of Reproduction and Fertility 1982; 64: 541-551.
51. Tsafriri A, Pomerantz SH, Channing CP. Inhibition of oocyte maturation by porcine follicular fluid: Partial characterization of the inhibitor. Biol Reprod 1976; 14: 511-516.
52. Hillensjo T, Channing CP, Pomerantz SH, Schwartz-Kripner A. Intrafollicular control of oocyte maturation in the pig. In Vitro 1979; 15: 32-39.
53. Sato E, Koide SS. A factor from bovine granulosa cells preventing oocyte maturation. Differentiation 1984; 26: 59-62.
54. Sato E, Ishibashi T. Meiotic arresting action of the substance obtained from cell surface of porcine ovarian granulosa cells. Jpn J Zootech Sci 1977; 48: 22-26.
55. Sato E, Ishibashi T, Iritani A. Meiotic arresting substance separated from porcine ovarian granulosa cells and hypothetical arresting mechanism of meiosis. In: Channing CP, Segal SJ (eds.), Intraovarian control mechanisms. New York: Plenun Publishing Corp; 1982: 161-173.
56. Downs SM, Coleman DL, Ward-Bailey PF, Eppig JJ. Hypoxanthine is the principal inhibitor of murine oocyte maturation in a low molecular weight fraction of porcine follicular fluid. Proc Natl Acad Sci U S A 1985; 82: 454-458.
57. Kadam AL, Koide SS. Identification of hypoxanthine in bovine follicular fluid. J Pharm Sci 1990; 79: 1077-1082.
58. Norbert TW. Fundamentals of clinical chemistry. Toronto: W.B. Saunders Company; 1970.
59. van Tol HT, Bevers MM. Partial characterization of the factor in theca-cell conditioned medium that inhibits the progression of FSH-induced meiosis of bovine oocytes surrounded by cumulus cells connected to the membrana granulosa. Mol Reprod Dev 2001; 60: 418-424.
60. Homa ST, Brown CA. Changes in linoleic acid during follicular development and inhibition of spontaneous breakdown of germinal vesicles in cumulus-free bovine oocytes. J Reprod Fertil 1992; 94: 153-160.
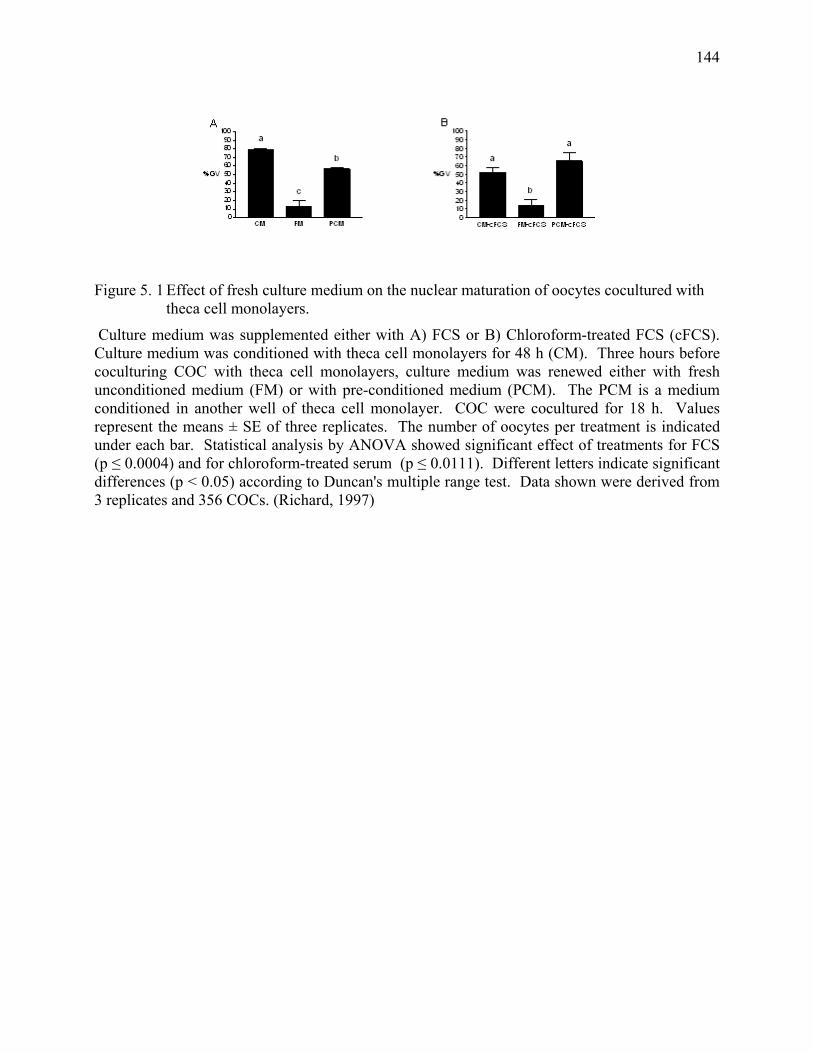
144
Figure 5. 1 Effect of fresh culture medium on the nuclear maturation of oocytes cocultured with theca cell monolayers.
Culture medium was supplemented either with A) FCS or B) Chloroform-treated FCS (cFCS). Culture medium was conditioned with theca cell monolayers for 48 h (CM). Three hours before coculturing COC with theca cell monolayers, culture medium was renewed either with fresh unconditioned medium (FM) or with pre-conditioned medium (PCM). The PCM is a medium conditioned in another well of theca cell monolayer. COC were cocultured for 18 h. Values represent the means ± SE of three replicates. The number of oocytes per treatment is indicated under each bar. Statistical analysis by ANOVA showed significant effect of treatments for FCS (p ≤ 0.0004) and for chloroform-treated serum (p ≤ 0.0111). Different letters indicate significant differences (p < 0.05) according to Duncan's multiple range test. Data shown were derived from 3 replicates and 356 COCs. (Richard, 1997)
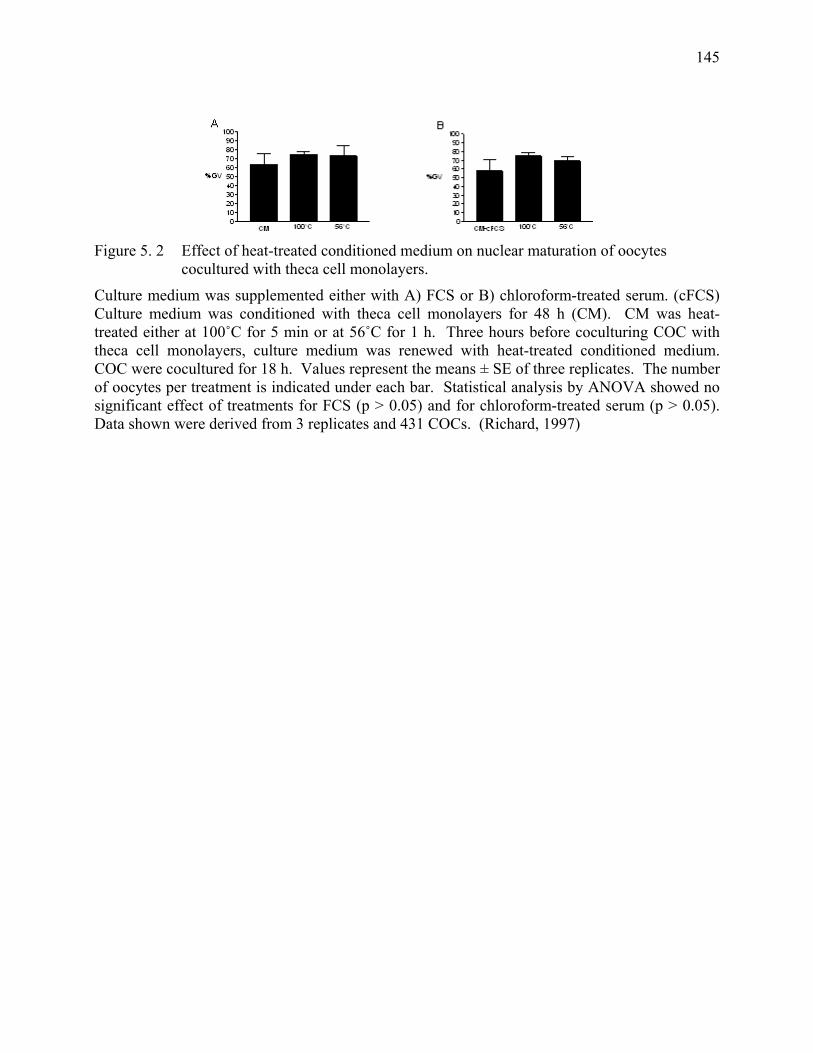
145
Figure 5. 2 Effect of heat-treated conditioned medium on nuclear maturation of oocytes
cocultured with theca cell monolayers.
Culture medium was supplemented either with A) FCS or B) chloroform-treated serum. (cFCS) Culture medium was conditioned with theca cell monolayers for 48 h (CM). CM was heat-treated either at 100˚C for 5 min or at 56˚C for 1 h. Three hours before coculturing COC with theca cell monolayers, culture medium was renewed with heat-treated conditioned medium. COC were cocultured for 18 h. Values represent the means ± SE of three replicates. The number of oocytes per treatment is indicated under each bar. Statistical analysis by ANOVA showed no significant effect of treatments for FCS (p > 0.05) and for chloroform-treated serum (p > 0.05). Data shown were derived from 3 replicates and 431 COCs. (Richard, 1997)
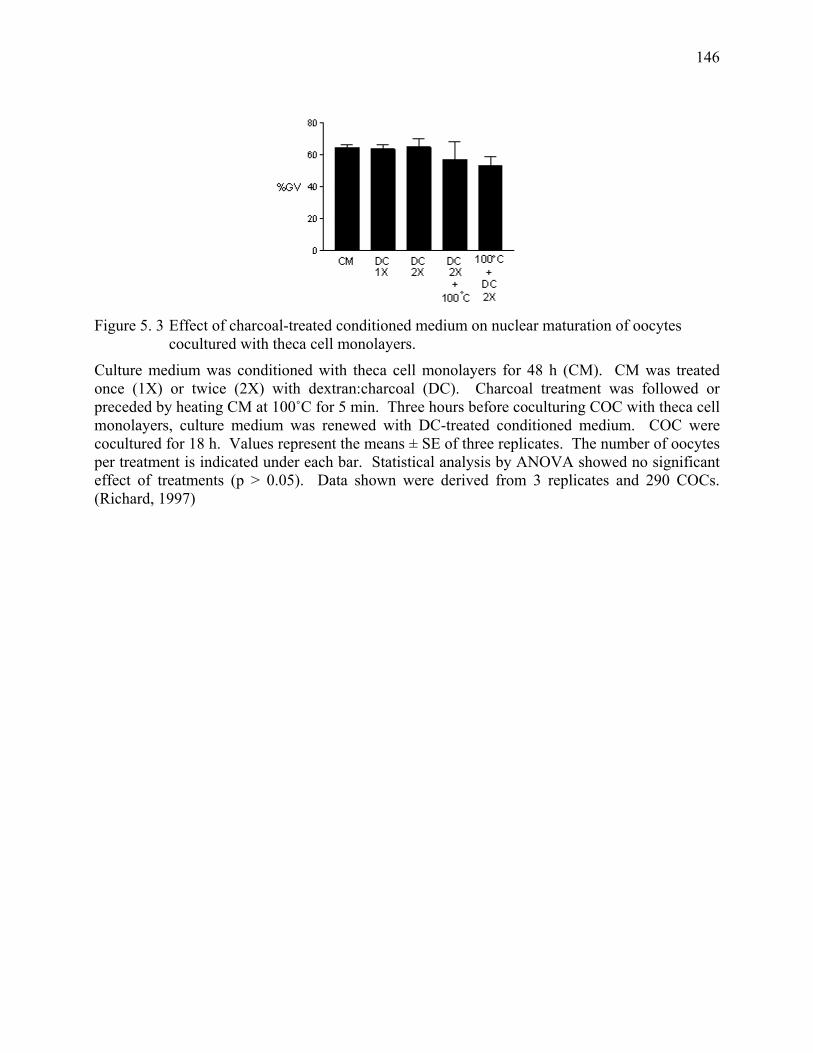
146
Figure 5. 3 Effect of charcoal-treated conditioned medium on nuclear maturation of oocytes
cocultured with theca cell monolayers.
Culture medium was conditioned with theca cell monolayers for 48 h (CM). CM was treated once (1X) or twice (2X) with dextran:charcoal (DC). Charcoal treatment was followed or preceded by heating CM at 100˚C for 5 min. Three hours before coculturing COC with theca cell monolayers, culture medium was renewed with DC-treated conditioned medium. COC were cocultured for 18 h. Values represent the means ± SE of three replicates. The number of oocytes per treatment is indicated under each bar. Statistical analysis by ANOVA showed no significant effect of treatments (p > 0.05). Data shown were derived from 3 replicates and 290 COCs. (Richard, 1997)

147
Figure 5. 4 Effect of different cut-off fractions of medium on nuclear maturation of oocytes cocultured with theca cell monolayers.
Conditioned medium (CM) shown in panel A and Fresh unconditioned medium (FM) shown in panel B were fractionated with different cut-off filters (300, 100, 30, 10, 1 kDa). The non-fractionated medium is identified «Total». Three hours before coculturing COC with theca cell monolayers, culture medium was renewed with cut-off filtered fractions of either FM or CM. COC were cocultured for 18 h. Values represent the means ± SE of three replicates. The number of oocytes per treatment is indicated under each bar. Statistical analysis by ANOVA showed significant effect of treatments (p < 0.0001). Different letters indicate significant differences (p < 0.05) according to Duncan's multiple range test. Data shown were derived from 5 replicates and 367 COCs. (Richard, 1997)
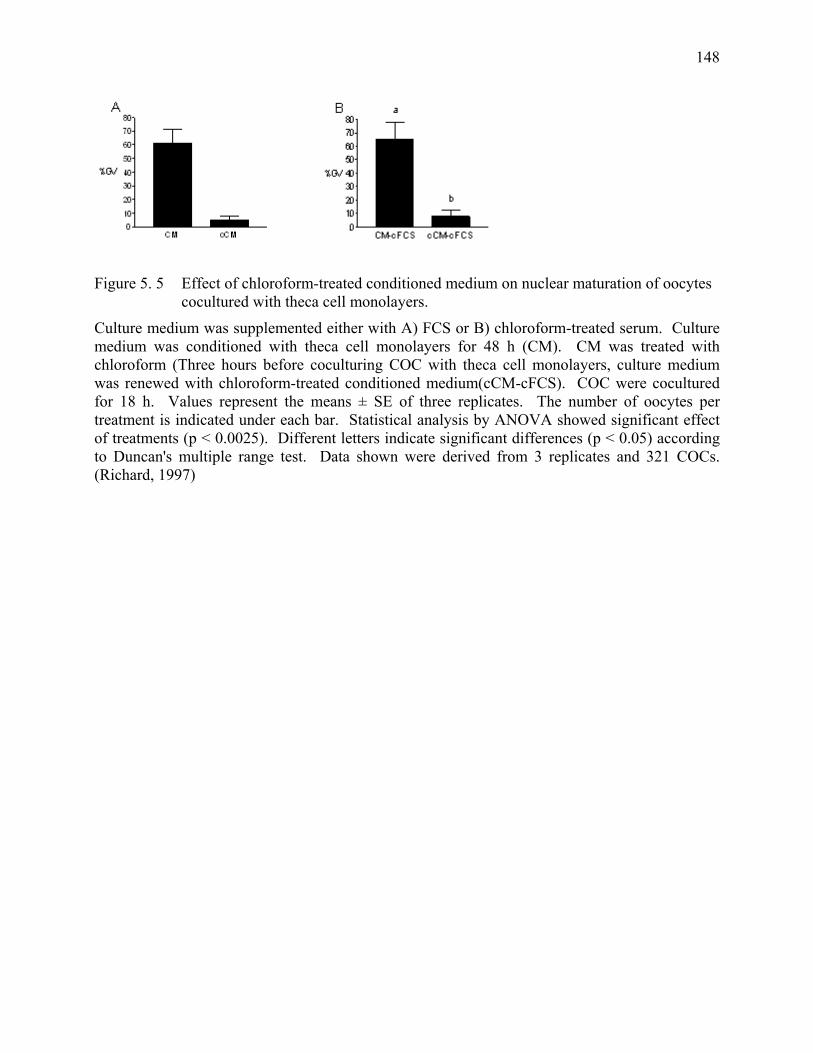
148
Figure 5. 5 Effect of chloroform-treated conditioned medium on nuclear maturation of oocytes cocultured with theca cell monolayers.
Culture medium was supplemented either with A) FCS or B) chloroform-treated serum. Culture medium was conditioned with theca cell monolayers for 48 h (CM). CM was treated with chloroform (Three hours before coculturing COC with theca cell monolayers, culture medium was renewed with chloroform-treated conditioned medium(cCM-cFCS). COC were cocultured for 18 h. Values represent the means ± SE of three replicates. The number of oocytes per treatment is indicated under each bar. Statistical analysis by ANOVA showed significant effect of treatments (p < 0.0025). Different letters indicate significant differences (p < 0.05) according to Duncan's multiple range test. Data shown were derived from 3 replicates and 321 COCs. (Richard, 1997)
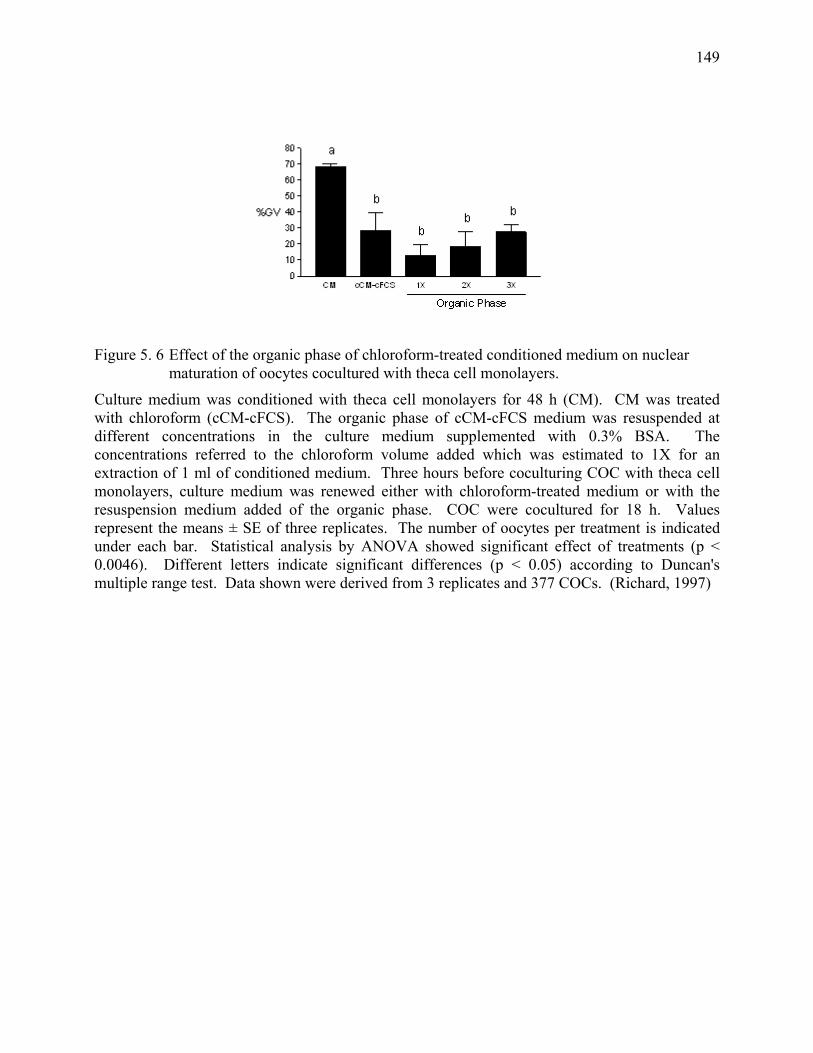
149
Figure 5. 6 Effect of the organic phase of chloroform-treated conditioned medium on nuclear maturation of oocytes cocultured with theca cell monolayers.
Culture medium was conditioned with theca cell monolayers for 48 h (CM). CM was treated with chloroform (cCM-cFCS). The organic phase of cCM-cFCS medium was resuspended at different concentrations in the culture medium supplemented with 0.3% BSA. The concentrations referred to the chloroform volume added which was estimated to 1X for an extraction of 1 ml of conditioned medium. Three hours before coculturing COC with theca cell monolayers, culture medium was renewed either with chloroform-treated medium or with the resuspension medium added of the organic phase. COC were cocultured for 18 h. Values represent the means ± SE of three replicates. The number of oocytes per treatment is indicated under each bar. Statistical analysis by ANOVA showed significant effect of treatments (p < 0.0046). Different letters indicate significant differences (p < 0.05) according to Duncan's multiple range test. Data shown were derived from 3 replicates and 377 COCs. (Richard, 1997)
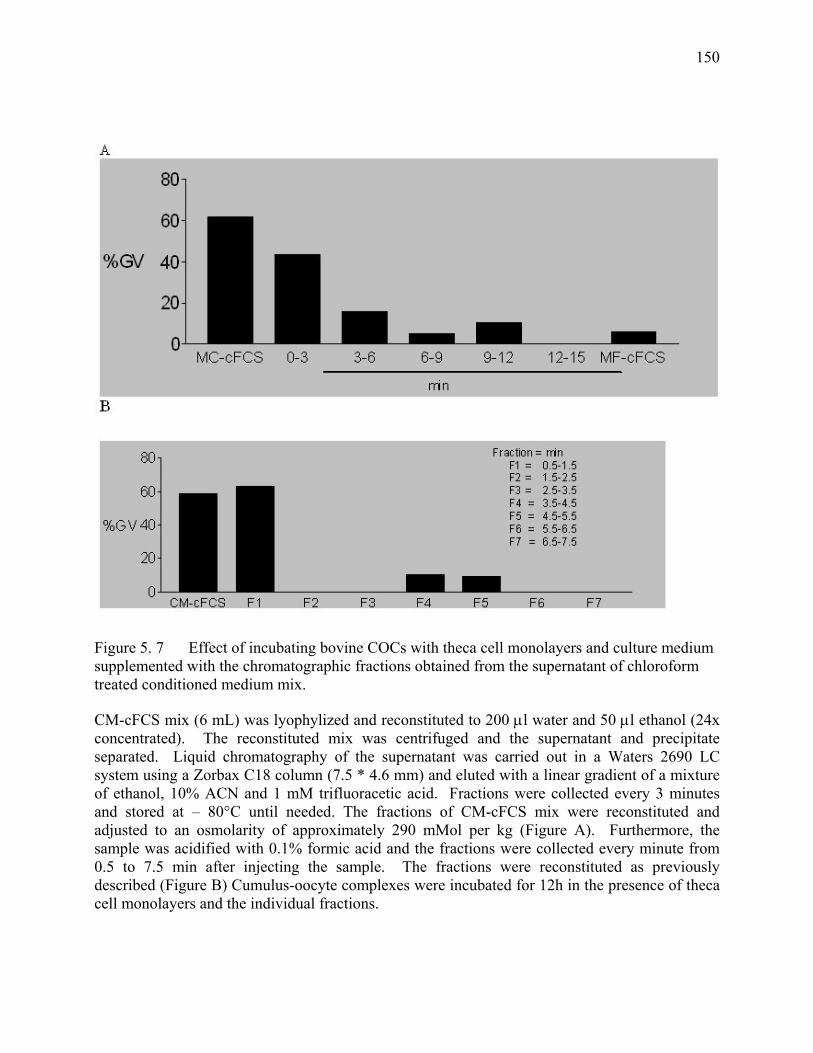
150
Figure 5. 7 Effect of incubating bovine COCs with theca cell monolayers and culture medium supplemented with the chromatographic fractions obtained from the supernatant of chloroform treated conditioned medium mix.
CM-cFCS mix (6 mL) was lyophylized and reconstituted to 200 µl water and 50 µl ethanol (24x concentrated). The reconstituted mix was centrifuged and the supernatant and precipitate separated. Liquid chromatography of the supernatant was carried out in a Waters 2690 LC system using a Zorbax C18 column (7.5 * 4.6 mm) and eluted with a linear gradient of a mixture of ethanol, 10% ACN and 1 mM trifluoracetic acid. Fractions were collected every 3 minutes and stored at – 80°C until needed. The fractions of CM-cFCS mix were reconstituted and adjusted to an osmolarity of approximately 290 mMol per kg (Figure A). Furthermore, the sample was acidified with 0.1% formic acid and the fractions were collected every minute from 0.5 to 7.5 min after injecting the sample. The fractions were reconstituted as previously described (Figure B) Cumulus-oocyte complexes were incubated for 12h in the presence of theca cell monolayers and the individual fractions.
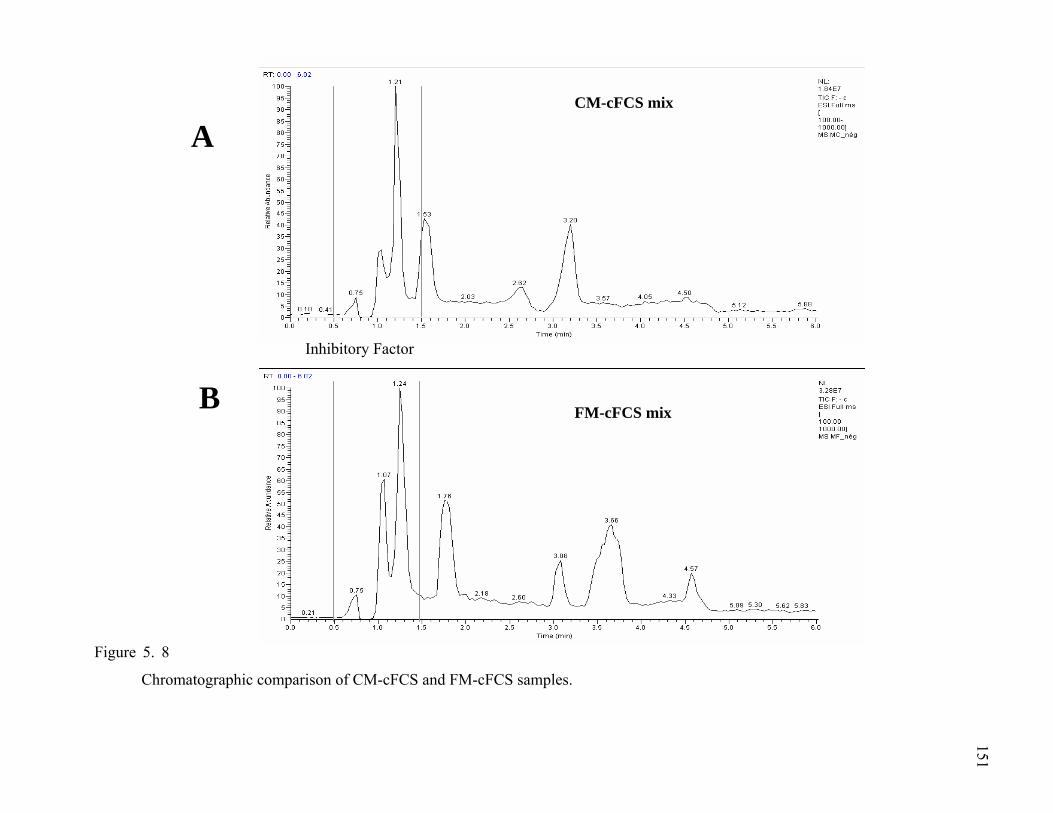
Figure 5. 8
Chromatographic comparison of CM-cFCS and FM-cFCS samples.
FM-cFCS mix
CM-cFCS mix
B
FactorInhibitory
A
151

152
Chromatographic comparison of CM-cFCS and FM-cFCS samples
CM-cFCS (Figure 5. 8a) and FM-cFCS (Figure 5. 8b) mixes were filtrated to obtain the fractions smaller than 5000 daltons. The samples were lyophilized lyophylized and reconstituted to 200 µl water and 50 µl ethanol (24x concentrated). The reconstituted mix was centrifuged and the supernatant and precipitate separated. The samples were then acidified with 0.1% formic acid. Liquid chromatography of the supernatant was carried out in a Waters 2690 LC system using a Zorbax C18 column (7.5 * 4.6 mm) and eluted with a linear gradient of a mixture of ethanol, 10% ACN and 1 mM trifluoracetic acid. The fractions were collected every minute from 0.5 to 6.5 min after injecting the sample. The fractions of CM-cFCS mix were reconstituted and adjusted to an osmolarity of approximately 290 mMol per kg. Cumulus-oocyte complexes were incubated for 12h in the presence of theca cell monolayers and the individual fractions.

153
GENERAL CONCLUSIONS Oocyte maturation is the first step towards a successful production of embryos in vitro.
Ovaries collected postmortem are the source of a large number of inexpensive and easily
obtainable immature cumulus-oocyte complexes (COCs). Bovine cumulus oocyte complexes
(COCs) incubated in vitro resume meiosis, whereas most of those incubated with theca cell
monolayers remain at the germinal vesicle (GV) stage. The oocytes proceed from the
germinal vesicle (GV) to the metaphase II stage. Important ultrastructural changes, that
prepare the oocyte to undergo fertilization and to sustain embryo development, take place
during oocyte maturation. It is hypothesized that incubation of oocytes under conditions that
maintain meiotic arrest at the GV stage might provide them with the opportunity to enhance
their developmental competence. Theca cells secrete an inhibitory factor that maintains
oocytes in a state of reversible meiotic arrest. Theca cells are a physiological alternative to
the pharmacological products used to maintain oocytes in meiotic arrest. Experiments were
carried out using bovine oocytes as a model to further study meiotic arrest and oocyte
maturation.
Bovine COCs harvested from postmortem ovaries at the slaughterhouse are an extremely
heterogeneous population. These COCs are usually selected and grouped in terms of their
morphology. In chapter II, our study showed that COC morphology is correlated with the
kinetics of meiotic resumption following a period of induced meiotic arrest. Differences
between the COC categories are detectable at the onset of in vitro maturation. In fact, the
number of COCs at the GV stage at time zero significantly decreases COCs with advanced
levels of atresia. This difference is maintained even after 4 h of in vitro culture. This strongly
supports the idea that the ability of the follicle to maintain bovine oocytes in meiotic arrest
becomes impaired as the follicle undergoes atresia. The degenerating follicle appears to lose
its suppressive activity on oocyte maturation. The kinetics of meiotic resumption after a
period of maintained meiotic arrest suggests that treated oocytes resume meiosis faster than
those untreated. Some COCs do not respond to the inhibitory factor produced by the theca
cell monolayers. Thus suggesting that the factor produced by the theca cells might only be
effective on a specific COC subpopulation. Furthermore, this study shows that the response
of immature oocytes to inhibitors of meiosis is affected by the post-mortem manipulations.

154
The specific role of cAMP in controlling the meiotic arrest of bovine oocytes is not well
understood. It has been proposed that high levels of cAMP in the cumulus cells stimulates
meiotic resumption, while high levels of cAMP in the oocyte promote meiotic arrest. Broad-
spectrum phosphodiesterase inhibitors such as IBMX have a transient effect on the meiotic
arrest of bovine oocytes. These non-specific inhibitors increase the levels of cAMP in both
the cumulus and the oocyte. Therefore, chapter III evaluated the ability of type-specific
phosphodiesterases (PDE) inhibitors to modulate the meiotic arrest of bovine oocytes.
Bovine oocytes treated with type 3 phosphodiesterase inhibitors remained in meiotic arrest,
whereas those treated with type-4 phosphodiesterase inhibitors resumed meiosis. These
results suggest that type 3 phosphodiesterase (PDE3) plays a role in meiotic arrest, whereas
type 4 phosphodiesterase (PDE4) does not have an inhibitory effect. The theca cell
monolayers and the type specific PDE inhibitors have an additive effect when used in
combination. The inhibitory factor produced by the theca cells appears to act upstream from
the site of action of PDE3 inhibition. The factor secreted by the theca cells may act by
decreasing the activity of PDE3. The pathways used by the theca cell monolayers or PDE
inhibitors may be complementary or alternative. On the other hand, the fact that COCs
constitute a heterogeneous population suggests that the COCs unresponsive to both treatments
may already be committed to resume meiosis. The response of the COCs to cAMP might be
explained by the preferential activation of PDE subtypes in the cumulus or the oocyte.
Further studies on the effect of type specific PDE inhibitors on the actual levels of cAMP in
the COC would provide a more complete idea of the role of PDE on oocyte maturation. In
addition it is also important to evaluate the reversibility of type-specific inhibitors.
The preovulatory peak of LH induces bovine oocytes to resume meiosis in vivo. It is known
that LH acts via the PKA signaling pathway, but other reports indicate that LH also acts
through the PKC signaling pathway. In chapter IV, the role of the PKA and PKC signaling
pathways in modulating the meiotic resumption of bovine oocytes was evaluated to better
understand and control oocyte maturation. Treatment of the theca cell monolayers with either
dbcAMP, PMA or LH did not reverse the inhibitory effect of the theca cells on meiotic
resumption. These results, in conjunction with the results of theca cells and PDE inhibitors,
suggests that high levels of cAMP in the theca cell monolayers is not enough to maximize
their effect on meiotic arrest. Cumulus-oocyte complexes incubated with granulosa cells
from large follicles resumed meiosis. Supplementation of the medium with dbcAMP, PMA

155
or LH did not have an effect on meiotic arrest. Untreated granulosa cell abrogate the
inhibitory effect of the theca cell monolayers on the maturation of bovine COCs. This is in
agreement with the fact that in vitro maturation of bovine oocytes is independent of LH
action. Interestingly, increasing the levels of cAMP with cAMP-analogs suggested that
activation of PKA in granulosa cells maintains the meiotic arrest in conjunction with the theca
cells. These results should be followed up with studies on the role of protein kinase C
inhibitors on oocyte maturation to better understand oocyte maturation. An interesting follow
up study would measure the levels of cAMP following treatment of bovine COCs with
phosphodiesterase inhibitors in the presence of granulosa cells. This study would help to
clarify the role of PDE , granulosa cells and cAMP in the maturation of bovine oocytes.
Chapter V describes experiments designed to further characterize the inhibitory factor
secreted by the theca cell monolayers. The inhibitory factor secreted by the theca cells is not
affected by either heat or multiple cycles of freezing and thawing. The conditioned medium
retains its inhibitory activity after treatment with charcoal or proteolytic enzymes. However,
the factor is either no longer present or active following chloroform-extraction of the
conditioned medium. The secreted factor is found in the cut-off filtered fraction of CM
smaller than 1000 daltons. The inhibitory factor has much in common with a previously
reported oocyte maturation inhibitor (OMI). However, the theca cell factor is not affected by
trypsin treatment, whereas OMI is degraded. The inhibitory factor is also resistant to the
other proteolytic enzymes tested. This suggests that inhibitory factor secreted by the theca
cells is not a protein or a large peptide. However, the inhibitory activity could potentially be a
small peptide that is not degraded by the action of the proteolytic enzymes tested so far.
Hypoxanthine, while an effective inhibitor of meiotic resumption in rodent oocytes, has only
a transient effect in preventing bovine oocytes from resuming meiosis. Supplementation of
incubation medium with hypoxanthine does not maintain bovine oocytes in meiotic arrest.
Due to the lack of effect of DNase and RNase on the inhibitory activity and the fact that
cyclic nucleotides play an important role in oocyte maturation, it is possible that the inhibitory
factor might be a nucleotide other than hypoxanthine. Chromatographic separation of the
conditioned medium yielded a fraction with the active but yet unknown component.
Comparison of the conditioned and fresh medium fractions indicated differences in the pattern
of chromatographic peaks. It is important to pursue the characterization of the inhibitory

156
factor(s) produced by the theca cell monolayers to better understand oocyte development and
to improve the developmental competence of immature oocytes in vitro.





























