Embed Size (px)
Citation preview

lable at ScienceDirect
Placenta 29 (2008) 892–897
Contents lists avai
Placenta
journal homepage: www.elsevier .com/locate/placenta
Three-dimensional Arrangement of the Capillary Bed and Its Relationshipto Microrheology in the Terminal Villi of Normal Term Placenta
M. Jirkovska a,*, J. Janacek b, J. Kalab a,b, L. Kubınova b
a Institute of Histology and Embryology, First Faculty of Medicine, Charles University, Albertov 4, CZ-12801 Prague 2, Czech Republicb Department of Biomathematics, Institute of Physiology, v.v.i., Academy of Sciences of the Czech Republic, Prague, Czech Republic
a r t i c l e i n f o
Article history:Accepted 4 July 2008
Keywords:3D reconstructionAngiogenesisConfocal microscopyImage analysisMurray’s lawPlasma skimmingSpatial arrangement
* Corresponding author. Fax: þ4202 24919899.E-mail address: [email protected] (M. Jirkovska).
0143-4004/$ – see front matter � 2008 Elsevier Ltd.doi:10.1016/j.placenta.2008.07.004
a b s t r a c t
Spatial arrangement of the capillary bed, manifestations of its growth and symmetry of capillarybranching were studied in peripheral villi of normal human placenta at term using confocal microscopyand image analysis. Unlike the model that has been accepted so far, it was shown that the arrangement ofthe capillary bed in terminal villi varied from simple, U-like loops to a richly branched network. Threedifferent categories of terminal villi (TV) were recognised:
(1) TV developing and protruding from an existing terminal villus, whose capillaries supply developingvilli;
(2) TV protruding from the mature intermediate villus in pairs or more numerous groups in which thecapillary bed is formed by capillaries connecting both the mature intermediate villus vascular bedand the capillary bed of the neighbouring terminal villus;
(3) separate TV with a capillary bed leading directly off and joining the vessels of the mature interme-diate villus.
Signs of capillary elongation and sprouting were observed in the villous capillary bed. Based on theassessment of the mean cross-sectional areas of capillaries constituting simple, Y-like capillary bifur-cation in terminal villi, the capillary branching was found to be asymmetric. Therefore, we conclude thatthe conditions for the ‘‘plasma skimming’’ effect are met in human placenta.
� 2008 Elsevier Ltd. All rights reserved.
1. Introduction
Knowledge of the spatial organisation of the microvascular bedis of great importance in understanding its function in differentorgans. Due to its role in transport between the mother and fetus,the placental microcirculation is a decisive factor in the appropriateintrauterine development of each individual. It is therefore inten-sively studied in both normal and pathological placenta.
As commonly accepted, the vast majority of the microvascularbed of the human term placenta is located in the mature inter-mediate and terminal villi. The mature intermediate villi encom-pass arterioles, thin capillaries and venules, from which capillariesof terminal villi arise. They run in a tight relationship with thetrophoblast, together forming a placental barrier that separatesmaternal and fetal blood and, at the same time, transports nutrients
All rights reserved.
and metabolites. In order to meet fetal requirements, the area of thetrophoblast enlarges due to the mitotic division of cytotrophoblastcells and their subsequent fusion with syncytiotrophoblast,whereas the capillary wall area enlarges by angiogenesis. Signs ofthose processes are observed in conventional microscopic sections,but no information is available on the spatial organisation of thevillous capillary bed and its changes.
Few studies on the three-dimensional (3D) arrangement of thevillous capillary bed have been carried out in the last five decades.Due to different methodical approaches (injection of contrastmedium and conventional light microscopy, corrosion casts, scan-ning electron microscopy, physical wax reconstruction) and alsobecause the different stages of placental samples were examined,the models that were designed were barely comparable. Crawford[1] interpreted the villous capillary bed as being a long, non-branched, wavy capillary running garland-like throughout thegrape-like arranged terminal villi of term placenta. On the otherhand, in terminal villi of immature placenta, Boe [2] demonstratedcapillary beds consisting of capillaries parallel with the villous axis,

M. Jirkovska et al. / Placenta 29 (2008) 892–897 893
mutually connected by dense, short segments mainly in the villoustips. According to Arts’s scheme [3], longitudinally oriented capil-laries of terminal villi are mutually connected by short capillarysegments, arise directly from arteries and run into veins. Scanningelectron microscopy of corrosion casts [4,5] showed more preciselysome variability in the capillary beds in terminal villi and theirrelationships to vessels in mature intermediate villi.
The authors of the latest concept used two methodical approaches,namely SEM analysis of corrosion preparations, and physical 3Dreconstruction based on photomicrographs of serial histologicalsections [6–8]. In their scheme, arterioles and paravascular capillariesof intermediate villi continue as coiled and sinusoidally dilatedcapillaries of the terminal villi. They are 3000–5000 mm long and runas predominantly hairpin-like capillaries throughout several succes-sive terminal villi, in some places forming protrusions or new villi bybulging against the trophoblast. Capillary sprouts with blind endswere described and interpreted for the first time in that paper as beingthe initial stage of newly formed capillaries [6,9].
Unlike the findings described above, our analysis, based on 3Dimages of capillaries acquired by confocal microscopy, has shownthat only 44% of the terminal villi contain a simple, hairpin- or U-like capillary loop in normal placenta. In 32% of villi, the capillarybed takes the form of a simple bifurcation (Y); other villi havemulti-branched capillaries [10]. A high proportion of villi withbranched capillaries leads us to the question: how could such anarrangement be useful for placental function?
As observed many years ago and repeatedly studied in detaillater on, at the asymmetric branching of microvessels withdiameter less than about 35 mm, asymmetric red cell distributionand separation of blood plasma take place. It results in theheterogeneity of the haematocrit in the daughter branches. Thisphenomenon, called ‘‘plasma skimming’’, influences local circu-lation [11–13]. The studies dealing with plasma skimming andother microrheological effects were carried out in vivo predomi-nantly on easily accessible animal organs, e.g. frog skin [11],animal mesentery [13] or muscle [14], and quite exceptionally inhumans, on the nerve root [15]. As for placenta, to our knowledge,the only reference to plasma skimming in the fetoplacentalmicrovascular bed is represented by a short speculative paragraphbased on Boe’s model of villous circulation [16]. The question ofwhether this effect occurs in human placenta has not yet beenanswered, most likely due to the absence of appropriate methodsof examination of the human placental microvascular bed.
Thanks to the recent technical progress it is possible to analyseplacental microvasculature using sophisticated methods, e.g. bymicrocomputed tomography; however, these techniques haveserious limits: it is very difficult, if even possible, to achievecompletely homogenous filling of all vessels by the contrastmedium, this procedure can cause collapse of the villous capillaries[17], and its resolution is still too low to resolve vessels with diam-eters smaller than 30 mm, which completely excludes analysis of thecapillary bed [18]. Therefore, microscopic techniques are stillnecessary for the examination of placental capillaries. Here, confocalmicroscopy in particular is a useful tool, enabling 3D images to beacquired with sufficient resolution as well as good detection of celland tissue compartments stained by relevant methods [19].
In this paper our aim was to demonstrate the spatial arrange-ment of the villous placental capillaries and to assess the symmetryof their bifurcation in order to judge if conditions required for the‘‘plasma skimming’’ effect are met.
2. Materials and methods
2.1. Specimen preparation and image acquisition by confocal microscopy
Human placentas were obtained with informed consent from three healthymothers (one secundipara and two tertiparae) undergoing spontaneous delivery in
the 39th or 40th week of gestation. Repeated examinations carried out duringpregnancy showed no signs of hypertension, preeclampsia or diabetes. The neonateswere healthy, and without congenital defects. Samples taken following systematicuniform random sampling no later than 10 min after delivery were fixed in 4%formaldehyde containing 0.5% eosin for at least 24 h, and embedded in paraffin wax.Thick sections (120 mm) were cut by a microtome HM 304 E (MICROM Laborgerate,Walldorf, Germany). Well-developed terminal villi lying completely inside the thicksections were sampled in an unbiased manner as described in Jirkovska et al. [10].Stacks of optical sections (Fig. 1A), 1 mm apart, encompassing the segments of thevillous tree containing the selected villi, were collected by a Bio-Rad MRC 600 laserscanning confocal microscope (double excitation at 488 and 567 nm, emission filtertransmitting>560 nm) using the planapochromat oil immersion objective with�40magnification and N.A.¼1.3. The collected series were used for the demonstration ofdifferent arrangements of villous capillaries, and those encompassing the Y-likecapillary bed were analysed for symmetry of capillary bifurcation.
2.2. 3D reconstructions of the villous capillary bed
Surface models of villi and capillaries were created and rendered using theimage analysis program Ellipse (ViDiTo, Kosice, Slovakia), supplied with plug-inmodules for the creation and visualisation of 3D surface models [20]. The coordi-nates of the contours of a selected class produced by the Ellipse Colon module wereconverted into a 3D binary image. To remove irregularities due to the hand drawingof contours, the image was resampled as a grey image with lower resolution andtreated by 3D Gaussian filtration. The triangulated isosurface of the grey value wasdetected. Partial models of the villous surface and capillaries were combined and theimages of the resulting 3D scenes were rendered [21].
2.3. Examination of symmetry of capillary bifurcationin terminal villi
Seventeen series of optical sections of terminal villi encompassing the Y-likecapillary bed were used for the assessment of the symmetry of capillary bifurcation.The mean cross-sectional area of each capillary branch was measured as follows:first, in subsequent optical sections, the contours of the villous surface and capillarybranches, except areas of bifurcation, were digitised using the Ellipse software(Fig. 1A). The volume of each capillary branch was measured by voxel countingapplied to the voxel representation of this branch delimited by the digitisedcontours (Fig. 1B). The length of the capillary branch was estimated by our Tracerplug-in module running in the Ellipse software environment, based on the lengthmeasurement of manually traced lines going through the capillary axis (Fig. 1C).Finally, the mean cross-sectional area of the capillary branch was estimated by theratio of the branch volume to its length, and the capillary diameter was calculated. Inorder to determine whether our data correspond with the theoretical assumption ofMurray’s law [22], the cubed diameters of the parent branches were compared withsums of the cubed diameters of the daughter branches using a paired t-test.Diameters d0> d1> d2 of each branching were used for the calculation of thebifurcation exponent D, such that
dD0 ¼ dD
1 þ dD2
3. Results
3.1. 3D reconstructions of the villous capillary bed
In order to demonstrate various forms of terminal villi and theircapillary bed, and stages of their growth and development, three-dimensional surface reconstructions of segments of peripheralplacental villi were arranged in a gallery in Fig. 2. Three differentcategories of terminal villi can be observed according to the rela-tionship of their capillary bed to the vascular bed of the matureintermediate villus. The first category is represented by terminalvilli developing and protruding from an existing terminal villus, andcontaining various forms of the capillary bed, e.g. a U-like loop orsimple bifurcation. Capillaries of those terminal villi originate in thecapillary bed of the pre-existing terminal villus, and the wholecluster is supplied from vessels of a mature intermediate villus(Fig. 2A, B, L). Another type of terminal villus protrudes from themature intermediate villus, usually in pairs (Fig. 2C). One or twocapillary segments shared by two neighbouring villi run just underthe trophoblast following the villous outer surface (Fig. 2D–G), othercapillaries connect them with the vessels of the mature interme-diate villus. And last but not least, separate villi with a capillary bedleading directly off and joining the vessels of the mature
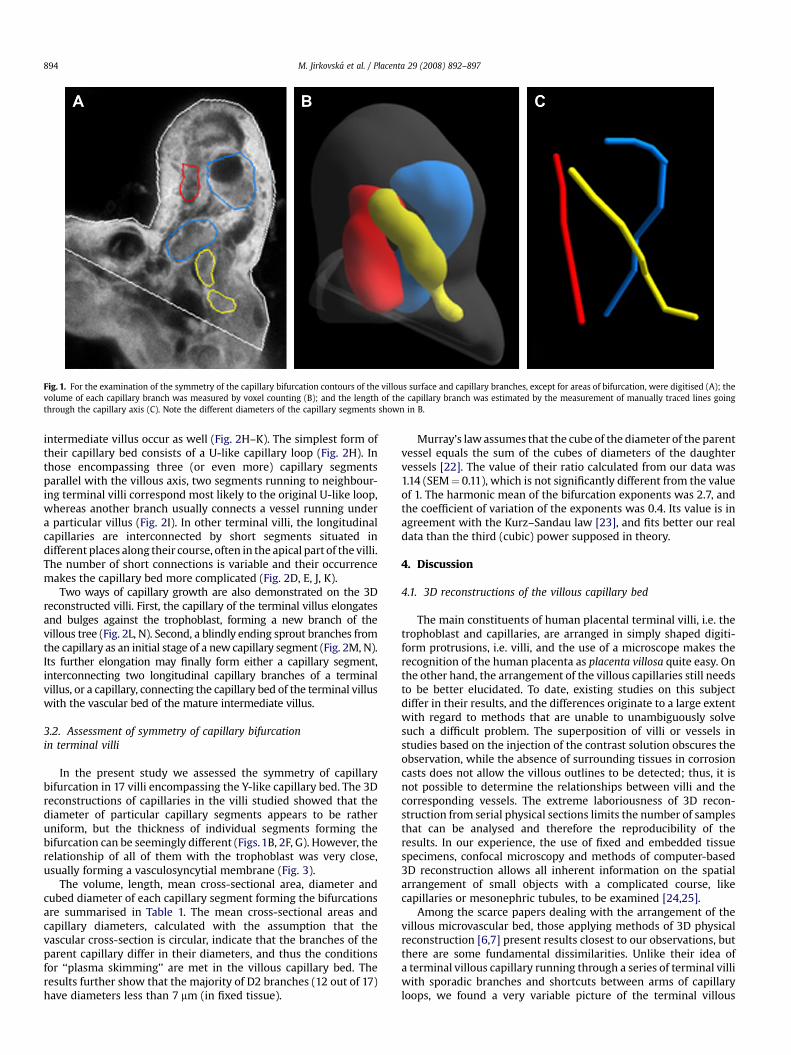
Fig. 1. For the examination of the symmetry of the capillary bifurcation contours of the villous surface and capillary branches, except for areas of bifurcation, were digitised (A); thevolume of each capillary branch was measured by voxel counting (B); and the length of the capillary branch was estimated by the measurement of manually traced lines goingthrough the capillary axis (C). Note the different diameters of the capillary segments shown in B.
M. Jirkovska et al. / Placenta 29 (2008) 892–897894
intermediate villus occur as well (Fig. 2H–K). The simplest form oftheir capillary bed consists of a U-like capillary loop (Fig. 2H). Inthose encompassing three (or even more) capillary segmentsparallel with the villous axis, two segments running to neighbour-ing terminal villi correspond most likely to the original U-like loop,whereas another branch usually connects a vessel running undera particular villus (Fig. 2I). In other terminal villi, the longitudinalcapillaries are interconnected by short segments situated indifferent places along their course, often in the apical part of the villi.The number of short connections is variable and their occurrencemakes the capillary bed more complicated (Fig. 2D, E, J, K).
Two ways of capillary growth are also demonstrated on the 3Dreconstructed villi. First, the capillary of the terminal villus elongatesand bulges against the trophoblast, forming a new branch of thevillous tree (Fig. 2L, N). Second, a blindly ending sprout branches fromthe capillary as an initial stage of a new capillary segment (Fig. 2M, N).Its further elongation may finally form either a capillary segment,interconnecting two longitudinal capillary branches of a terminalvillus, or a capillary, connecting the capillary bed of the terminal villuswith the vascular bed of the mature intermediate villus.
3.2. Assessment of symmetry of capillary bifurcationin terminal villi
In the present study we assessed the symmetry of capillarybifurcation in 17 villi encompassing the Y-like capillary bed. The 3Dreconstructions of capillaries in the villi studied showed that thediameter of particular capillary segments appears to be ratheruniform, but the thickness of individual segments forming thebifurcation can be seemingly different (Figs. 1B, 2F, G). However, therelationship of all of them with the trophoblast was very close,usually forming a vasculosyncytial membrane (Fig. 3).
The volume, length, mean cross-sectional area, diameter andcubed diameter of each capillary segment forming the bifurcationsare summarised in Table 1. The mean cross-sectional areas andcapillary diameters, calculated with the assumption that thevascular cross-section is circular, indicate that the branches of theparent capillary differ in their diameters, and thus the conditionsfor ‘‘plasma skimming’’ are met in the villous capillary bed. Theresults further show that the majority of D2 branches (12 out of 17)have diameters less than 7 mm (in fixed tissue).
Murray’s law assumes that the cube of the diameter of the parentvessel equals the sum of the cubes of diameters of the daughtervessels [22]. The value of their ratio calculated from our data was1.14 (SEM¼ 0.11), which is not significantly different from the valueof 1. The harmonic mean of the bifurcation exponents was 2.7, andthe coefficient of variation of the exponents was 0.4. Its value is inagreement with the Kurz–Sandau law [23], and fits better our realdata than the third (cubic) power supposed in theory.
4. Discussion
4.1. 3D reconstructions of the villous capillary bed
The main constituents of human placental terminal villi, i.e. thetrophoblast and capillaries, are arranged in simply shaped digiti-form protrusions, i.e. villi, and the use of a microscope makes therecognition of the human placenta as placenta villosa quite easy. Onthe other hand, the arrangement of the villous capillaries still needsto be better elucidated. To date, existing studies on this subjectdiffer in their results, and the differences originate to a large extentwith regard to methods that are unable to unambiguously solvesuch a difficult problem. The superposition of villi or vessels instudies based on the injection of the contrast solution obscures theobservation, while the absence of surrounding tissues in corrosioncasts does not allow the villous outlines to be detected; thus, it isnot possible to determine the relationships between villi and thecorresponding vessels. The extreme laboriousness of 3D recon-struction from serial physical sections limits the number of samplesthat can be analysed and therefore the reproducibility of theresults. In our experience, the use of fixed and embedded tissuespecimens, confocal microscopy and methods of computer-based3D reconstruction allows all inherent information on the spatialarrangement of small objects with a complicated course, likecapillaries or mesonephric tubules, to be examined [24,25].
Among the scarce papers dealing with the arrangement of thevillous microvascular bed, those applying methods of 3D physicalreconstruction [6,7] present results closest to our observations, butthere are some fundamental dissimilarities. Unlike their idea ofa terminal villous capillary running through a series of terminal villiwith sporadic branches and shortcuts between arms of capillaryloops, we found a very variable picture of the terminal villous
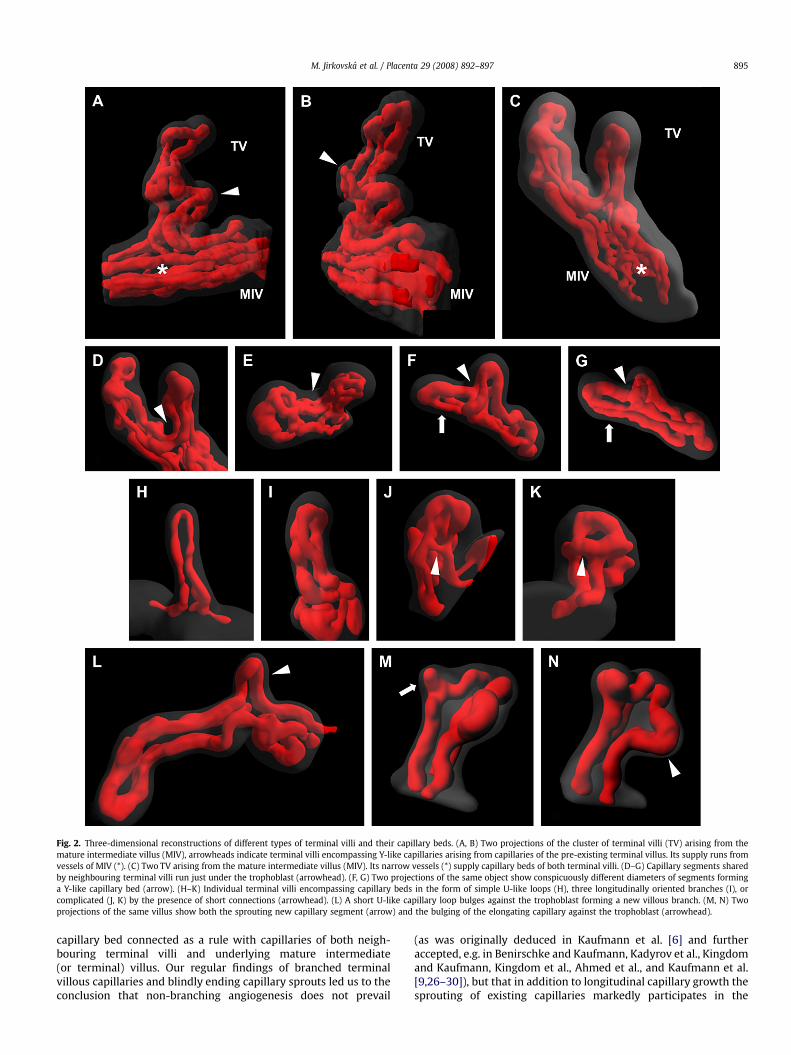
Fig. 2. Three-dimensional reconstructions of different types of terminal villi and their capillary beds. (A, B) Two projections of the cluster of terminal villi (TV) arising from themature intermediate villus (MIV), arrowheads indicate terminal villi encompassing Y-like capillaries arising from capillaries of the pre-existing terminal villus. Its supply runs fromvessels of MIV (*). (C) Two TV arising from the mature intermediate villus (MIV). Its narrow vessels (*) supply capillary beds of both terminal villi. (D–G) Capillary segments sharedby neighbouring terminal villi run just under the trophoblast (arrowhead). (F, G) Two projections of the same object show conspicuously different diameters of segments forminga Y-like capillary bed (arrow). (H–K) Individual terminal villi encompassing capillary beds in the form of simple U-like loops (H), three longitudinally oriented branches (I), orcomplicated (J, K) by the presence of short connections (arrowhead). (L) A short U-like capillary loop bulges against the trophoblast forming a new villous branch. (M, N) Twoprojections of the same villus show both the sprouting new capillary segment (arrow) and the bulging of the elongating capillary against the trophoblast (arrowhead).
M. Jirkovska et al. / Placenta 29 (2008) 892–897 895
capillary bed connected as a rule with capillaries of both neigh-bouring terminal villi and underlying mature intermediate(or terminal) villus. Our regular findings of branched terminalvillous capillaries and blindly ending capillary sprouts led us to theconclusion that non-branching angiogenesis does not prevail
(as was originally deduced in Kaufmann et al. [6] and furtheraccepted, e.g. in Benirschke and Kaufmann, Kadyrov et al., Kingdomand Kaufmann, Kingdom et al., Ahmed et al., and Kaufmann et al.[9,26–30]), but that in addition to longitudinal capillary growth thesprouting of existing capillaries markedly participates in the
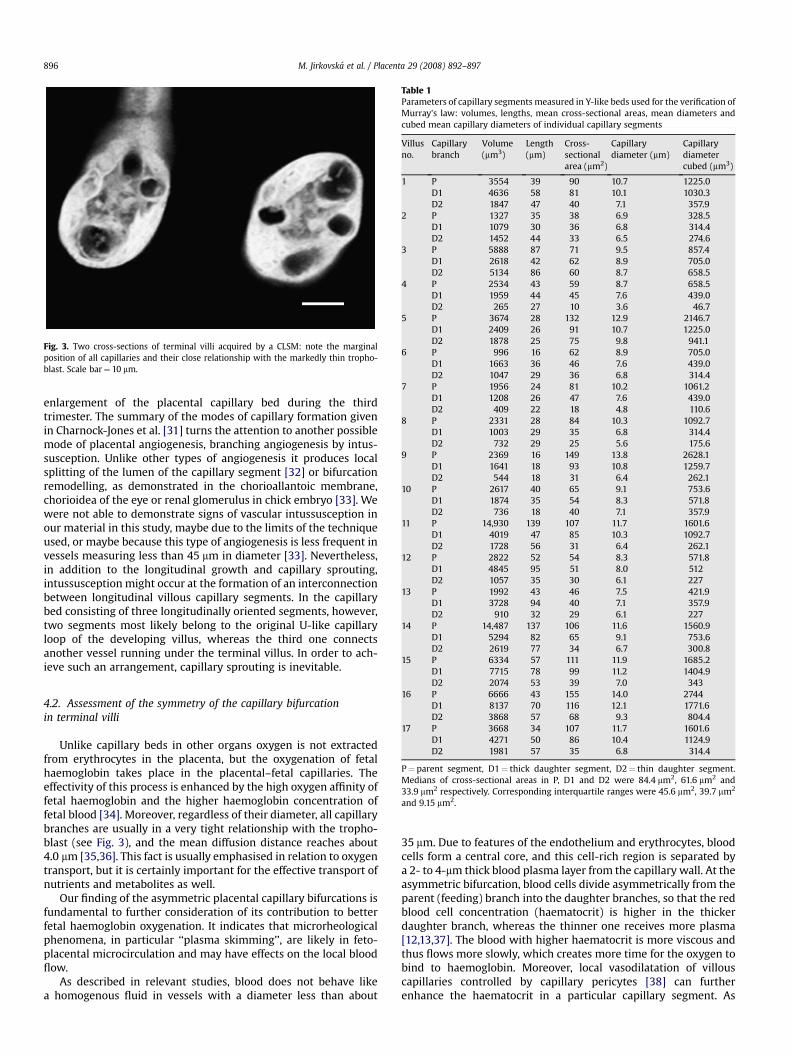
Fig. 3. Two cross-sections of terminal villi acquired by a CLSM: note the marginalposition of all capillaries and their close relationship with the markedly thin tropho-blast. Scale bar¼ 10 mm.
Table 1Parameters of capillary segments measured in Y-like beds used for the verification ofMurray’s law: volumes, lengths, mean cross-sectional areas, mean diameters andcubed mean capillary diameters of individual capillary segments
Villusno.
Capillarybranch
Volume(mm3)
Length(mm)
Cross-sectionalarea (mm2)
Capillarydiameter (mm)
Capillarydiametercubed (mm3)
1 P 3554 39 90 10.7 1225.0D1 4636 58 81 10.1 1030.3D2 1847 47 40 7.1 357.9
2 P 1327 35 38 6.9 328.5D1 1079 30 36 6.8 314.4D2 1452 44 33 6.5 274.6
3 P 5888 87 71 9.5 857.4D1 2618 42 62 8.9 705.0D2 5134 86 60 8.7 658.5
4 P 2534 43 59 8.7 658.5D1 1959 44 45 7.6 439.0D2 265 27 10 3.6 46.7
5 P 3674 28 132 12.9 2146.7D1 2409 26 91 10.7 1225.0D2 1878 25 75 9.8 941.1
6 P 996 16 62 8.9 705.0D1 1663 36 46 7.6 439.0D2 1047 29 36 6.8 314.4
7 P 1956 24 81 10.2 1061.2D1 1208 26 47 7.6 439.0D2 409 22 18 4.8 110.6
8 P 2331 28 84 10.3 1092.7D1 1003 29 35 6.8 314.4D2 732 29 25 5.6 175.6
9 P 2369 16 149 13.8 2628.1D1 1641 18 93 10.8 1259.7D2 544 18 31 6.4 262.1
10 P 2617 40 65 9.1 753.6D1 1874 35 54 8.3 571.8D2 736 18 40 7.1 357.9
11 P 14,930 139 107 11.7 1601.6D1 4019 47 85 10.3 1092.7D2 1728 56 31 6.4 262.1
12 P 2822 52 54 8.3 571.8D1 4845 95 51 8.0 512D2 1057 35 30 6.1 227
13 P 1992 43 46 7.5 421.9D1 3728 94 40 7.1 357.9D2 910 32 29 6.1 227
14 P 14,487 137 106 11.6 1560.9D1 5294 82 65 9.1 753.6D2 2619 77 34 6.7 300.8
15 P 6334 57 111 11.9 1685.2D1 7715 78 99 11.2 1404.9
M. Jirkovska et al. / Placenta 29 (2008) 892–897896
enlargement of the placental capillary bed during the thirdtrimester. The summary of the modes of capillary formation givenin Charnock-Jones et al. [31] turns the attention to another possiblemode of placental angiogenesis, branching angiogenesis by intus-susception. Unlike other types of angiogenesis it produces localsplitting of the lumen of the capillary segment [32] or bifurcationremodelling, as demonstrated in the chorioallantoic membrane,chorioidea of the eye or renal glomerulus in chick embryo [33]. Wewere not able to demonstrate signs of vascular intussusception inour material in this study, maybe due to the limits of the techniqueused, or maybe because this type of angiogenesis is less frequent invessels measuring less than 45 mm in diameter [33]. Nevertheless,in addition to the longitudinal growth and capillary sprouting,intussusception might occur at the formation of an interconnectionbetween longitudinal villous capillary segments. In the capillarybed consisting of three longitudinally oriented segments, however,two segments most likely belong to the original U-like capillaryloop of the developing villus, whereas the third one connectsanother vessel running under the terminal villus. In order to ach-ieve such an arrangement, capillary sprouting is inevitable.
D2 2074 53 39 7.0 34316 P 6666 43 155 14.0 2744
D1 8137 70 116 12.1 1771.6D2 3868 57 68 9.3 804.4
17 P 3668 34 107 11.7 1601.6D1 4271 50 86 10.4 1124.9D2 1981 57 35 6.8 314.4
P¼ parent segment, D1¼ thick daughter segment, D2¼ thin daughter segment.Medians of cross-sectional areas in P, D1 and D2 were 84.4 mm2, 61.6 mm2 and33.9 mm2 respectively. Corresponding interquartile ranges were 45.6 mm2, 39.7 mm2
and 9.15 mm2.
4.2. Assessment of the symmetry of the capillary bifurcationin terminal villi
Unlike capillary beds in other organs oxygen is not extractedfrom erythrocytes in the placenta, but the oxygenation of fetalhaemoglobin takes place in the placental–fetal capillaries. Theeffectivity of this process is enhanced by the high oxygen affinity offetal haemoglobin and the higher haemoglobin concentration offetal blood [34]. Moreover, regardless of their diameter, all capillarybranches are usually in a very tight relationship with the tropho-blast (see Fig. 3), and the mean diffusion distance reaches about4.0 mm [35,36]. This fact is usually emphasised in relation to oxygentransport, but it is certainly important for the effective transport ofnutrients and metabolites as well.
Our finding of the asymmetric placental capillary bifurcations isfundamental to further consideration of its contribution to betterfetal haemoglobin oxygenation. It indicates that microrheologicalphenomena, in particular ‘‘plasma skimming’’, are likely in feto-placental microcirculation and may have effects on the local bloodflow.
As described in relevant studies, blood does not behave likea homogenous fluid in vessels with a diameter less than about
35 mm. Due to features of the endothelium and erythrocytes, bloodcells form a central core, and this cell-rich region is separated bya 2- to 4-mm thick blood plasma layer from the capillary wall. At theasymmetric bifurcation, blood cells divide asymmetrically from theparent (feeding) branch into the daughter branches, so that the redblood cell concentration (haematocrit) is higher in the thickerdaughter branch, whereas the thinner one receives more plasma[12,13,37]. The blood with higher haematocrit is more viscous andthus flows more slowly, which creates more time for the oxygen tobind to haemoglobin. Moreover, local vasodilatation of villouscapillaries controlled by capillary pericytes [38] can furtherenhance the haematocrit in a particular capillary segment. As

M. Jirkovska et al. / Placenta 29 (2008) 892–897 897
shown on 3D reconstructions, the interconnected capillary bed ofneighbouring terminal villi or villous clusters actually representssuccessive bifurcations. It is likely that such an arrangementprovides working conditions for a very dynamic system where theasymmetric distribution of erythrocytes at upstream bifurcationinfluences the distribution of erythrocytes at downstream bifur-cation (as suggested in studies using computational and physicallaboratory models [39,40]). Considering possible and probablerepeated redistribution of erythrocytes at consecutive capillarybifurcations before they enter the venous system, we judge that thebranching of capillaries can contribute to the higher efficiency ofthe haemoglobin oxygenation in placenta.
The theory of Murray’s law is based on the premise that the flowin the physiological vascular system with the optimal arrangementis achieved with the least possible energy consumption. It issupposed that in such ramified networks, the relationship betweenthe diameters of the parent and daughter vessels is substantial, andis represented by the so-called bifurcation exponent D (see alsoSection 2). Provided that the fluid flows with the minimum energyconsumption and constant shear stress, its value is 3 [22]. Analysisof experimental data has corrected its value to about 2.7 for arte-rioles [33]. The bifurcation exponent calculated from our data isidentical; thus, the first evidence that bifurcation of the fetopla-cental capillaries meets the assumptions of Murray’s law is pre-sented in our study.
Acknowledgements
We thank an anonymous reviewer for a helpful discussion. Thisstudy was supported by the Grant Agency of the Czech Republic,project no. 304/05/0153, by the Ministry of Education of the CzechRepublic, project no. LC06063, and by the Academy of Sciences ofthe Czech Republic (Grant AV0Z50110509).
Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.placenta.2008.07.004.
References
[1] Crawford JM. The foetal placental circulation. IV. The anatomy of thevillus and its capillary structure. J Obstet Gynaecol Br Emp 1956;63:548–52.
[2] Boe F. Studies on the human placenta. III. Vascularization of the young foetalplacenta. A. Vascularization of the chorionic villus. Acta Obstet Gynecol Scand1969;48:159–66.
[3] Arts NFT. Investigations on the vascular system of the placenta. I. Generalintroduction and the fetal vascular system. Am J Obstet Gynecol 1961;82:147–58.
[4] Akiba K, Kuwabara Y, Mizuno M, Fukuda S. Semiquantitative study of thevascularisation of human term placenta using scanning electron microscopy.J Clin Electron Microsc 1987;20:5–6.
[5] Burton GJ. The fine structure of the human placental villus as revealed byscanning electron microscopy. Scanning Microsc 1987;1:1811–28.
[6] Kaufmann P, Bruns U, Leiser R, Luckhardt M, Winterhager E. The fetal vascu-larisation of term human placental villi. II. Intermediate and terminal villi.Anat Embryol 1985;173:203–14.
[7] Leiser R, Luckhardt M, Kaufmann P, Winterhager E, Bruns U. The fetal vascu-larisation of term human placental villi. I. Peripheral stem villi. Anat Embryol1985;173:71–80.
[8] Leiser R, Krebs C, Ebert B, Dantzer V. Placental vascular corrosion cast studies:a comparison between ruminants and humans. Microsc Res Tech 1997;38:76–87.
[9] Benirschke K, Kaufmann P. Pathology of the human placenta. New York:Springer; 1995. pp. 136.
[10] Jirkovska M, Kubınova L, Janacek J, Moravcova M, Krejcı V, Karen P. Topologicalproperties and spatial organization of villous capillaries in normal and dia-betic placentas. J Vasc Res 2002;39:268–78.
[11] Krogh A. Studies on the physiology of capillaries. II. The reactions to localstimuli of the blood vessels in the skin and web of the frog. J Physiol1921;55:412–22.
[12] Gaehtgens P. Distribution of flow and red cell flux in the microcirculation.Scand J Clin Lab Invest Suppl 1981;156:83–7.
[13] Pries AR, Ley K, Claasen M, Gaehtgens P. Red cell distribution at microvascularbifurcations. Microvasc Res 1989;38:81–101.
[14] Kim S, Popel AS, Intaglietta M, Johnson PC. Effect of erythrocyte aggregation atnormal human levels on functional capillary density in rat spinotrapeziusmuscle. Am J Physiol Heart Circ Physiol 2006;290:H941–7.
[15] Dezawa A, Unno K, Yamane T, Miki H. Changes in the microhemodynamics ofnerve root retraction in patients with lumbar spinal canal stenosis. Spine2002;27:2844–9.
[16] Jønsson V, Bock JE, Nielsen JB. Significance of plasma skimming and plasmavolume expansion. J Appl Physiol 1992;72:2047–51.
[17] Langheinrich AC, Wienhard J, Vormann S, Hau B, Bohle RM, Zygmunt M.Analysis of the fetal placental vascular tree by X-ray micro-computedtomography. Placenta 2004;25:95–100.
[18] Rennie MY, Whiteley KJ, Kulandavelu S, Adamson SL, Sled JG. 3D visualisationand quantification by microcomputed tomography of late gestational changesin the arterial and venous feto-placental vasculature of the mouse. Placenta2007;28:833–40.
[19] Ockleford CD, Mongan LC, Hubbard ARD. Techniques of advanced lightmicroscopy and their applications to morphological analysis of human extra-embryonic membranes. Microsc Res Tech 1997;38:153–64.
[20] Janacek J, Kubınova L. 3D reconstruction of surfaces captured by confocalmicroscopy. Acta Stereol 1998;17:259–64.
[21] Jirkovska M, Kubınova L, Janacek J, Kalab J. 3-D study of vessels in peripheralplacental villi. Image Anal Stereol 2007;26:165–8.
[22] Sherman TF. On connecting large vessels to small. The meaning of Murray’slaw. J Gen Physiol 1981;78:431–53.
[23] Kurz H, Sandau K. Modeling of blood vessel development – bifurcation patternand hemodynamics, optimality and allometry. Comments Theor Biol1997;4:261–91.
[24] Jirkovska M, Kubınova L, Krekule I, Hach P. Spatial arrangement of fetalplacental capillaries in terminal villi: a study using confocal microscopy. AnatEmbryol 1998;197:263–72.
[25] Jirkovska M, Naprstkova I, Janacek J, Kucera T, Macasek J, Karen P, et al. Three-dimensional reconstructions from non-deparaffinized tissue sections. AnatEmbryol 2005;210:163–73.
[26] Kadyrov M, Kosanke G, Kingdom J, Kaufmann P. Increased fetoplacentalangiogenesis during first trimester in anaemic women. Lancet1998;352:1747–9.
[27] Kingdom J, Kaufmann P. Oxygen and placental vascular development. In:Roach RC, Wagner PD, Hackett PH, editors. Hypoxia: into the next millennium.New York: Kluwer Academic/Plenum Publishing; 1999.
[28] Kingdom J, Huppertz B, Seaward G, Kaufmann P. Development of the placentalvillous tree and its consequences for fetal growth. Eur J Obstet Gynecol ReprodBiol 2000;92:35–43.
[29] Ahmed A, Dunk C, Ahmad S, Khaliq A. Regulation of placental vascularendothelial growth factor (VEGF) and placenta growth factor (PlGF) andsoluble Flt-1 by oxygen – a review. Troph Res, 14. Placenta 2000;21(Suppl.A):S16–24.
[30] Kaufmann P, Mayhew TM, Charnock-Jones DS. Aspects of human fetoplacentalvasculogenesis and angiogenesis. II. Changes during normal pregnancy.Placenta 2004;25:114–26.
[31] Charnock-Jones DS, Kaufmann P, Mayhew TM. Aspects of human fetoplacentalvasculogenesis and angiogenesis. I. Molecular regulation. Placenta2004;25:103–13.
[32] Davies JA. Mechanisms of morphogenesis. Elsevier Academic Press; 2005.277–280.
[33] Djonov VG, Kurz H, Burri PH. Optimality in the developing vascular system:branching remodeling by means of intussusception as an efficient adaptationmechanism. Dev Dyn 2002;224:391–402.
[34] Petersen OH, editor. Human physiology. 5th ed. Blackwell Publishing; 2007.p. 473–4.
[35] Mayhew TM, Joy CF, Haas JD. Structure-function correlation in the humanplacenta: the morphometric diffusing capacity for oxygen at full term. J Anat1984;139:691–708.
[36] Burton GJ, Ingram SC, Palmer ME. The influence of mode of fixation onmorphometrical data derived from terminal villi in the human placenta atterm: a comparison of immersion and perfusion fixation. Placenta1987;8:37–51.
[37] Enden G, Popel AS. A numerical study of plasma skimming in small vascularbifurcations. J Biomech Eng 1994;116:79–88.
[38] Oishi K, Kamiyashiki T, Ito Y. Isometric contraction of microvascular pericytesfrom mouse brain parenchyma. Microvasc Res 2007;73:20–8.
[39] Carr RT, Wickham LL. Plasma skimming in serial microvascular bifurcations.Microvasc Res 1990;40:179–90.
[40] Carr RT, Xiao J. Plasma skimming in vascular trees: numerical estimates ofsymmetry recovery lengths. Microcirculation 1995;2:345–53.





























