Embed Size (px)
Citation preview

Unilateral carrageenan injection into muscle or joint induceschronic bilateral hyperalgesia in rats
Rajan Radhakrishnana,b, Steven A. Moorec,d, and Kathleen A. Slukaa,b,c,*aGraduate Program in Physical Therapy and Rehabilitation Science, University of Iowa, 100 MedicalEducation Building #1-252, Iowa, IA 52242-1190, USAbPain Research Program, University of Iowa, Iowa, IA 52242-1190, USAcNeuroscience Graduate Program, 1178ML, College of Medicine, Iowa, IA 52242-1190, USAdDepartment of Pathology, College of Medicine, 200 Hawkins Drive, 5239B Roy Carver Pavilion,Iowa, IA 52242-1190, USA
AbstractChronic musculoskeletal pain is a major clinical problem and there is a general lack of animal modelsto study this condition. Carrageenan is commonly used to produce short-lasting acute inflammationand hyperalgesia in animal models. However, the potential of carrageenan to produce chronic, long-lasting hyperalgesia has not been evaluated. In the present study, we investigated the long-term effectsof carrageenan injected into joint or muscle in rats. Rats were injected with 0.3, 1 or 3% carrageenanin one knee joint or gastrocnemius muscle and hyperalgesia to mechanical (measured as decreasedwithdrawal threshold) and heat (measured as decreased withdrawal latency) stimuli of both pawsassessed before and at varying times after injection, through 8 weeks. Histological changes wereexamined only after injection of 3% carrageenan. Three percent carrageenan injected in the muscleor knee produced hyperalgesia to mechanical and heat stimuli ipsilaterally, which lasted 7–8 weeksand spread to the contralateral side 1–2 weeks after injection. One percent carrageenan injected tothe knee joint or gastrocnemius muscle, produced hyperalgesia that was shorter-lasting and remainedipsilateral; 0.3% carrageenan injected into the knee joint or gastrocnemius muscle had no effect.Three percent carrageenan injected into the skin surrounding the knee joint did not producehyperalgesia. A similar pattern of inflammatory changes was observed histologically for both thejoint and muscle tissues. Acute inflammation was observed for the first 24 h with edema andneutrophilic infiltration evident as early as 4 h. At 1 week, the inflammation converted to primarilya macrophage response with scattered mast cells. The data suggest that animals injected with 1 or3% carrageenan in the knee joint or gastrocnemius muscle could be used as models of acuteinflammation through 24 h and chronic inflammation after 1 week. Furthermore, 3% carrageenaninjected into deep tissues produces hyperalgesia that spreads to the contralateral side, at the sametime period as the inflammation transforms from acute to chronic.
KeywordsCarrageenan; Hyperalgesia; Muscle; Knee; Chronic; Bilateral; Inflammation
© 2003 International Association for the Study of Pain. Published by Elsevier Science B.V. All rights reserved.*Corresponding author. Tel.: +1-319-335-9791; fax: +1-319-335-9707. E-mail address: [email protected] (K.A. Sluka)..
NIH Public AccessAuthor ManuscriptPain. Author manuscript; available in PMC 2009 August 26.
Published in final edited form as:Pain. 2003 August ; 104(3): 567–577.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

1. IntroductionChronic pain due to arthritis or neuropathy is a major clinical problem globally. To study theunderlying pathological mechanisms of chronic pain, a number of good experimental animalmodels have been developed over the years (De Castro Costa et al., 1981; Bendele et al.,1999; Kehl et al., 2000; Sluka et al., 2001). However, there is a general lack of experimentalmodels for chronic musculoskeletal pain. Most of the animal models available to studymusculoskeletal pain are of acute nature and the chronic studies carried out to date do notextend for more than 3 or 4 weeks. Acute, short-lasting hyperalgesia models may not share thesame mechanisms of chronic pain and may not be suitable to study the chronic painmechanisms.
Complete Freund’s adjuvant (CFA), carrageenan (alone or in combination with kaolin),zymosan or urate crystals are used to induce experimental inflammation/arthritis and chronicpain in animal models (Pearson and Wood, 1959; Winter et al., 1962; Di Rosa et al., 1971;Keystone et al., 1977; Coderre and Wall, 1987; Decaris et al., 1999). CFA normally consistsof heat-killed bacteria, Mycobacterium butyricum or Mycobacterium tuberculosis, as anemulsion in sterile mineral oil (Pearson and Wood, 1958; Swingle and Grant, 1977). CFAcauses systemic effects like febrile response and inflammation at distant locations due to theirsystemic spread (Philippe et al., 1997; Deacris et al., 1999). Therefore, their use may not beappropriate to study-specific mechanisms, especially the neuronal mechanisms, of paininduction and/or maintenance. CFA (Haak et al., 1996) and zymosan, a glycan derived fromyeast cell wall (Keystone et al., 1977), produce an immune-mediated chronic inflammationwhereas agents like carrageenan, probably, produce a non-immune-mediated inflammation(Di Rosa et al., 1971; Moreno, 1993; Guthrie et al., 1996). Also, the severity of CFA-inducedarthritis is strain specific (Swingle et al., 1969; Muir and Dumonde, 1982; Crowe et al.,1985). The potential of carrageenan in inducing chronic hyperalgesia has not been evaluatedso far. Lambda carrageenan (type IV) is a water-extractable polysaccharide derived frommarine plants, Gigartina aciculaire and Gigartina pistillata (Sigma Chemical Co. Catalogue).It causes inflammation and hyperalgesia when injected into tissues but not on topicalapplication or ingestion in reasonable quantities (Nicklin and Miller, 1984). The mechanismsof carrageenan-induced acute inflammation and the subsequent hyperalgesia have been studiedextensively (for review, see Schaible and Grubb, 1993; Mense, 1993). The acute inflammatoryprocess is characterized by the accumulation of neutrophils in the perivascular space (Diehl etal., 1988), which is accompanied by the local release of noxious chemicals such as glutamate,prostaglandins, histamine and serotonin (Neil et al., 1987; Guilbaud et al., 1989; Nantel et al.,1999; Lawand et al., 2000; Hong et al., 2002). These noxious chemicals sensitize primaryafferents resulting in primary hyperalgesia (Berberich et al., 1988; Diehl et al., 1988; Schaibleand Schmidt, 1985, 1988; Hargreaves et al., 1988; Kehl et al., 2000).
Injection of carrageenan into deep tissues activates dorsal horn neurons causing centralsensitization either spinally or supraspinally (Schaible et al., 1987; Neugebauer and Schaible,1990; Dougherty and Willis, 1992; Hoheisel et al., 1993, 1995; Urban et al., 1999). The centralsensitization, together with the increased sensitivity of the peripheral nociceptors, is manifestedas secondary hyperalgesia (Sluka and Westlund, 1993). Secondary hyperalgesia is usuallyobserved in the areas adjacent to the injury and sometimes in distal locations, which could bedue to neuronal changes occurring spinally or supraspinally (Coderre and Melzack, 1985;Urban et al., 1999; Chen et al., 2000, Sluka et al., 2001; Sluka, 2002).
The mechanisms of pain and hyperalgesia induced by injecting inflammatory agents intodifferent tissues like muscle, joint and skin are thought to be different (Sluka, 2002; reviewedby Mense, 1993 and Schaible and Grubb, 1993). The afferent innervation of the spinal cordfrom skin and deep tissues like muscles and joints are also different (Mense and Craig, 1988;
Radhakrishnan et al. Page 2
Pain. Author manuscript; available in PMC 2009 August 26.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Craig et al., 1988; Willis and Coggeshall, 1991). It has been reported that stimulation of C-fibers from a muscle nerve causes a long-lasting enhancement of the ventral root reflexcompared to C-fibers from a cutaneous nerve (Wall and Woolf, 1984). Stimulation of deeptissues like muscle or knee joint with capsaicin produces a longer-lasting hyperalgesiacompared to the stimulation of cutaneous tissue (Sluka, 2002).
With the above observations in mind, the potential of lambda carrageenan to induce chronichyperalgesia, after injection to different tissues viz. muscle, joint and skin, was investigated inthe present study using behavioral testing. Since histological changes are known to occur duringchronic inflammation, a histopathological study of the tissues, parallel to the behavioral studieswere also carried out.
2. Materials and methods2.1. Animals
Sprague—Dawley rats (Harlan, USA), weighing 225–300 g, kept at 12 h dark—light cyclewith free access to standard rat chow and water, were used for the experiments. Animals werebrought to the behavioral testing room the day before, to acclimatize them to the testingenvironment. Behavioral tests were usually done between 9 a.m. and 2 p.m. except for the 8 htesting after inflammation, which was done before 5 p.m. All experiments were approved byUniversity of Iowa Animal Care and Use Committee and were carried out according to theguidelines of the International Association for the Study of Pain and National Institute of Health(Zimmermann, 1983).
2.2. Heat testingAnimals were kept in plexi-glass restrainers on an elevated platform with a clear glass top forabout 30 min for acclimatization. A high-intensity radiant heat source was used as the stimulus.The heat source was positioned on the plantar skin of the hind limb and the beam was switchedon, simultaneously starting a built-in timer. When the animal withdrew the paw abruptly toheat stimulus, the heat source and timer were stopped. The duration in seconds from the startof heat application to paw withdrawal was taken as the paw withdrawal latency (PWL). PWLswere determined five times bilaterally, with an interval of 5 min between each test and themean of five readings was taken as the PWL for a particular time point. The intensity of theheat source was kept constant in all experiments with the aid of a constant voltage-powersupply. This method of testing results in an increase in the paw temperature of the animal untilit withdraws the paw and the voltage was set to produce an intensity required to obtain a baselineresponse time between 12 and 16 s. Cut-off time was set to 30 s to minimize heat damage tothe skin. The validity and reliability of this testing method was previously established(Hargreaves et al., 1988; Sluka et al., 1999). A decrease in withdrawal latency is interpretedas heat hyperalgesia for the purpose of this study.
2.3. Mechanical testingAnimals were kept in a plexi-glass restrainer on an elevated platform with a mesh wire top.Threshold to mechanical stimuli was tested using von Frey filaments with increasing bendingforce as described elsewhere (Sluka et al., 2001). Briefly, the filament with the lowest thresholdwas applied to the plantar surface of the hind limb two times and observed for a withdrawal.If there was no response, the next higher force filament was tested. The value of the lowestforce filament causing a withdrawal of the paw was taken as the mechanical threshold. Onehundred and sixty-two millinewton was set as the cut-off. The following bending forces wereassessed: 8, 12, 16, 32, 44, 56, 75, 104, 162 mN. The reliability of this testing method waspreviously established (Gopalkrishnan and Sluka, 2000). A decrease in withdrawal thresholdis interpreted as mechanical hyperalgesia in this study.
Radhakrishnan et al. Page 3
Pain. Author manuscript; available in PMC 2009 August 26.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

2.4. ProtocolSeven groups of animals were used for the behavioral testing. Three groups were given a singleinjection of 0.3 (n = 6), 1 (n = 6) or 3% (n = 12) lambda carrageenan (Type IV, 100 μl, dissolvedin sterile saline, Sigma Chemical Company, St. Louis, USA) in the left knee joint anteriorly.The other three groups were given a single injection of 0.3 (n = 6), 1 (n = 6) or 3% (n = 12)carrageenan (100 μl) in the left gastrocnemius muscle belly. Another group (n = 6) was givena single injection of 3% carrageenan in the skin surrounding the anterior aspect of the knee.All carrageenan injections were given under halothane anesthesia (2–4% v/v in oxygen). PWLsto heat and mechanical withdrawal thresholds were recorded in all groups, except the skin-injected group, before and 4, 8, 24 h, 1, 2, 3, 4, 5, 6, 7 and 8 weeks after injection of carrageenan.In the skin group, the PWLs and mechanical withdrawal threshold were measured before and2 and 4 h after injection.
2.5. Histological studiesSeven groups of two animals each were injected with 3% carrageenan into the knee joint underlight halothane anesthesia. Groups 1–7 were sacrificed at 4, 8, 24 h, 1, 2, 4 and 8 weeks,respectively, after the injection. Both ipsi- and contralateral knee joints were dissected andfixed in 10% formalin. In another seven groups of two animals each, 3% carrageenan wasinjected in the gastrocnemius muscle and the muscle was dissected and fixed according to thesame protocol and time schedule as the knee joint. Knee joints were subjected to decalcificationprior to embedment in paraffin. Paraffin sections of all tissues were stained with hematoxylinand eosin (H and E) and examined by light microscopy. Analysis of histological findings wasdescriptive and performed in a blinded fashion by a pathologist.
2.6. Statistical analysisStatistical analyses were performed using SPSS 10.1 statistical software. The level ofsignificance was set at P < 0.05. Differences in PWL in heat testing were determined acrosstime (baseline to 8 weeks) and among different carrageenan doses (0.3, 1 and 3%) usingrepeated measures of analysis of variance (ANOVA) followed by Tukey’s post hoc test fordifferences between groups and paired t-test for differences from baseline. For mechanicaltesting, non-parametric tests were used. Friedman’s ANOVA assessed differences across time,followed by sign test to determine differences from baseline. Kruskal—Wallis one-wayANOVA was used to test differences between groups followed by sign test to determinedifferences between individual groups. The area under the curve (AUC) for time—responsecurves for PWL and mechanical thresholds were calculated for individual animals usingSigmaplot® software which takes into account time on X-axis and response on the Y-axis.Student’s t-test was used to compare differences between group means. All data are presentedas mean ± SEM.
3. Results3.1. Knee-PWL to heat
Intraarticular injection of carrageenan produced an ipsilateral decrease in the PWL to radiantheat that was long-lasting depending on the dose. Baseline withdrawal latencies were 17.7 ±0.57 s and decreased to 12.0 ± 0.38 s within 4 h after injection of 3% carrageenan into the kneejoint. Injection of 3% carrageenan produced a decrease in PWL ipsilaterally by 4 h that lastedthrough 6 weeks (F1,11 = 3.7, P = 0.02) (Fig. 1A). Contralaterally, there was a significantdecrease in PWL to heat (F1,11 = 12.9, P = 0.001) after injection of 3% carrageenan into theknee (Fig. 1B). Lower doses of carrageenan produced only ipsilateral effects that were shorter-lasting, 24 h for 0.3% and up to 3 weeks for 1% carrageenan (Fig. 1B). Analysis of the AUC
Radhakrishnan et al. Page 4
Pain. Author manuscript; available in PMC 2009 August 26.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

for the PWL to heat was greater ipsilaterally for the group that received 3% carrageenan whencompared to the 0.3% and contralaterally compared to 0.3 or 1% dose (Fig. 2A).
3.2. Muscle-PWL to heatIntramuscular injection of 3% carrageenan decreased PWL to heat ipsilaterally that was long-lasting, i.e. decreased through 8 weeks (F1,11 = 16.7, P = 0.006) (Fig. 1D). Baseline withdrawallatencies were 17.2 ± 0.94 s and decreased to 11.8 ± 0.47 s within 4 h after injection of 3%carrageenan into the gastrocnemius muscle. There was a contralateral decrease in PWL to heatwith 3% i.m. carrageenan by 1 week that also lasted through 8 weeks (F1,11 = 21.4, P = 0.04)(Fig. 1C). There were no significant differences in PWL to heat for the group injected witheither 0.3 or 1% carrageenan into the gastrocnemius muscle, ipsi- or contralaterally. Analysisof the AUC for the PWL to heat from the group that received 3% carrageenan was significantlygreater both for ipsi- and contralateral sides than the group that received 1% carrageenan (Fig.2C).
3.3. Knee-mechanical thresholdThe mechanical withdrawal threshold decreased ipsilaterally following intraarticular injectionof carrageenan (χ2 = 54.2, P = 0.0001). This decrease was dependent on dose and differencesbetween doses occurred 4 (P = 0.03), 8 (P = 0.001), 24 h (P = 0.05), 3 (P = 0.02), 5 (P = 0.004),6 (P = 0.02), 7 (P = 0.001) and 8 weeks (P = 0.02) after carrageenan. The group that received3% carrageenan showed significantly decreased withdrawal thresholds compared to 0.3%group at 4, 8, 24 h, 3, 4, 5 and 7 weeks and compared to 1% group at 24 h, 3, 4, 5, 6 and 7weeks. Ipsilaterally, 0.3% carrageenan did not produce a significant decrease in mechanicalwithdrawal threshold at any time after injection. Injection of 1% carrageenan into the kneejoint showed significant decreases compared to baseline 4, 8, 24 h and 1 week, ipsilaterally.Injection of 3% carrageenan produced significant decreases in withdrawal threshold tomechanical stimuli 4, 8, 24, 1, 3 and 5 weeks compared to baseline values (Fig. 3A). Analysisof the AUC of time—response curves from the group that received 3% carrageenan wassignificantly greater ipsilaterally, than the group that received 0.3 and 1% carrageenan.
A contralateral decrease in mechanical threshold also occurred with 3% carrageenan by 3weeks that remained through 6 weeks. Significant differences between doses occurred 3 (P =0.02), 4 (P = 0.005), 5 (P = 0.005), 6 (P = 0.03) and 7 weeks (P = 0.03) after intraarticularinjection of carrageenan with the group that received 3% carrageenan showing greaterreductions in withdrawal thresholds compared to 0.3 (3, 4 and 5 weeks) and 1% carrageenan(3, 4, 5, 6 and 7 weeks) (Fig. 3B). Analysis of the AUC for time—response curves from thegroup with 3% carrageenan was significantly greater than the group with 1% carrageenan (Fig.2B).
3.4. Muscle-mechanical thresholdThe mechanical withdrawal threshold decreased ipsilaterally following intramuscular injectionof carrageenan (χ2 = 35.8, P = 0.0001). This decrease was dependent on dose and differencesbetween doses occurred 4 (P = 0.001), 8 (P = 0.001), 24 h (P = 0.004), 1 (P = 0.003), 2 (P =0.006), 3 (P = 0.04), 4 (P = 0.01), 5 (P = 0.005), 6 (P = 0.01) and 8 weeks (P = 0.05) aftercarrageenan injection. The group received 3% carrageenan showed significantly decreasedwithdrawal thresholds ipsilaterally, compared to 0.3% group at 4, 8, 24 h, 3, 4, 5, 6 and 8 weeksand compared to 1% group at 4, 8 h, 1, 2, 3, 4, 5 and 6 weeks. Ipsilaterally, 0.3% carrageenandid not produce a significant decrease in mechanical withdrawal threshold at any time afterinjection. Injection of 1% carrageenan into the gastrocnemius muscle showed significantdecreases in mechanical withdrawal threshold compared to baseline, 4 and 8 h ipsilaterally.Injection of 3% carrageenan produced significant decreases in withdrawal threshold tomechanical stimuli 4, 8, 24 h, 1, 2, 4 and 5 weeks compared to baseline values (Fig. 3C).
Radhakrishnan et al. Page 5
Pain. Author manuscript; available in PMC 2009 August 26.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Analysis of the AUC of time—response curves from the group that received 3% carrageenanwas significantly greater ipsilaterally than the group that received 0.3 and 1% carrageenan (Fig.2D).
A contralateral decrease in mechanical threshold also occurred following intramuscularinjection of 3% carrageenan by 3 weeks that remained through 6 weeks (Fig. 3D). Significantdifferences between doses occurred 1 (P = 0.008), 2 (P = 0.03), 3 (P = 0.03), 4 (P = 0.004), 5(P = 0.002), 6 (P = 0.007), 7 (P = 0.02) and 8 weeks (P = 0.03) after intramuscular injectionof carrageenan with the group that received 3% carrageenan showing greater reductions inwithdrawal thresholds compared to 0.3 (1–8 weeks) and 1% carrageenan (1–8 weeks). Analysisof the AUC for the time—response curves from the contralateral paw showed that 3%carrageenan was significantly greater than the group with 1% (Fig. 2D).
3.5. SkinInjection of 3% carrageenan into the skin surrounding the anterior aspect of the knee did notproduce any significant changes in withdrawal latency to heat or withdrawal threshold tomechanical stimuli (data not shown). The withdrawal latency to heat ipsilaterally was 17.6 ±1.1 s before injection and 15.7 ± 0.75 s and 16.9 ± 0.87 s 2 and 4 h after injection of 3%carrageenan into the skin. Mechanical withdrawal threshold remained at a median value of 162mN before and after injection of 3% carrageenan into the skin.
3.6. Spontaneous pain behaviorAnimals showed spontaneous pain behaviors such as guarding the injected paw and weight-bearing on the contralateral paw during the first 24–48 h. After 48 h, there was no sign ofspontaneous pain except that there was curling of the paw ipsliaterally for 1–2 weeks.
3.7. Histology studiesBoth knee and muscle developed similar patterns of acute inflammation during the first weekfollowing carrageenan injection (Fig. 4). Mild hemorrhage, edema and minimal inflammatorycell infiltrates (mostly neutrophils) were seen 4 h after injection in both tissues. Moreneutrophils were found at 8 h. At 24 h, acute inflammation was severe and accompanied bymyonecrosis and fibrinous exudate into the joint space. By 1 week, the inflammation hadconverted to primarily a macrophage response with a few scattered mast cells. Muscleinflammation was mostly epimysial and perimysial. Changes at week 2 were very similar tothat at week 1 for muscle and joint tissue. By the fourth week, macrophages were still present,but less in number and by the eighth week, only mild chronic inflammation (primarilymacrophages) was seen. There was no evidence of inflammatory cell infiltrates in thecontralateral knee joint or muscle tissues at any time points (Fig. 4).
4. DiscussionThe present study examined dose effects, time course and laterality of carrageenan-inducedhyperalgesia, when injected unilaterally into knee joint or gastrocnemius muscle in rats. Threepercent carrageenan injected unilaterally in either muscle or knee joint produces acuteunilateral hyperalgesia to heat and mechanical stimuli that spreads contralaterally within 1–2weeks and is long-lasting (weeks). Lower doses of carrageenan produce acute unilateralhyperalgesia that is transient. Injection of carrageenan into the skin in a comparable dermatometo the muscle and joint did not produce hyperalgesia to mechanical or heat stimuli.Histologically, there is an acute inflammatory response with neutrophilic infiltration for thefirst week. This converts to a macrophage-dominated chronic inflammation by 1 week thatlasts through 8 weeks. Thus, carrageenan can be used as a model of acute inflammation of
Radhakrishnan et al. Page 6
Pain. Author manuscript; available in PMC 2009 August 26.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

muscle or joint for the first 24 h and as a model of chronic inflammatory hyperalgesia after 1–2 weeks.
4.1. Differences between cutaneous and deep somatic painPain originating from muscle or joint is uniquely different from that of pain originating fromskin. Muscle pain is diffuse, longer lasting and more unpleasant (Torebjork et al., 1984;Marchettini et al., 1996; Svensson et al., 1997; Witting et al., 2000). This may relate todifferences in central anatomical pathways or different biochemical mediators. Dorsal rootganglion (DRG) cells that innervate muscle and joint have more calcitonin gene-related peptideand substance P and less isolectin B4 and somatostatin when compare to DRG cells innervatingskin (O’Brien et al., 1989; Plenderleith and Snow, 1993). The central projections from primaryafferents innervating muscle and joint are predominately to laminae I and deeper dorsal horn,while those from cutaneous tissue also project to laminae II (Mense and Craig, 1988; Craig etal., 1988; Willis and Coggeshall, 1991; Mense, 1993; Schaible and Grubb, 1993). Thus, painfrom muscle or joint would be expected to produce a different patterned response than painfrom skin.
4.2. Chronic hyperalgesiaIt has been previously shown that most of the injected carrageenan is removed within 48 hfrom the knee joint (Santer et al., 1983) and possibly faster from other tissues. However,secondary hyperalgesia produced by carrageenan persists even after 48 h and transforms intochronic hyperalgesia that is long-lasting (weeks). The induction, maintenance and spread ofchronic hyperalgesia could result from a series of peripheral and central changes occurring atthe site of insult and at spinal or supraspinal sites.
Peripherally, carrageenan injected into the muscle causes the symptoms of myositis viz.hyperemia, edema and infiltration by neutrophils (Berberich et al., 1988). When injected intothe knee joint, carrageenan leads to synthesis and release of inflammatory mediators, whichcause edema and rapid infiltration of neutrophils within the first few hours (Schaible and Grubb,1993). Injection of carrageenan/kaolin mixture into the knee joint causes an immediate increasein glutamate and nitric oxide metabolites in the knee joint, which persists for hours (Lawandet al., 2000). The increase is prevented by intraarticular administration of lidocaine suggestingthat the glutamate is released from neuronal endings in the joint (Lawand et al., 2000). In thepresent study, when carrageenan was injected into the knee joint or muscle, an acute phase ofinflammation followed by a chronic phase, lasting up to 8 weeks, was observed. The localchanges occurring after carrageenan insult are likely responsible for the sensitization of theperipheral nociceptors and primary afferents, which contribute to the development ofsecondary hyperalgesia, along with central changes (Mense, 1993; Schaible et al., 2002).
Injection of carrageenan into the paw of rats induces cyclooxygenase-2 (COX-2) and producesprostaglandin E2 locally (Di Rosa et al., 1971; Nantel et al., 1999), induces COX-2 mRNA inthe lumbar spinal cord (Hay and de Belleroche, 1997) and releases glutamate, aspartate,substance P, nitric oxide and prostaglandin E2 in the dorsal horn (Schaible et al., 1990; Slukaand Westlund, 1992; Sorkin et al., 1992; Yang et al., 1996; Rivot et al., 2002). Carrageenanhas also been found to cause an acute increase in immunoreactive calcitonin gene-relatedpeptide and substance P in the spinal cord that lasts through at least 1 week (Garry andHargreaves, 1992; Sluka and Westlund, 1993). Some or all of these spinal changes could beresponsible for the central sensitization occurring spinally, following carrageenan-inducedinflammation of a joint or muscle (Schaible et al., 1987; Dougherty and Willis, 1992; Hoheiselet al., 1995). Supraspinal involvement in secondary hyperalgesia produced by peripheral tissueinjury, including knee joint carrageenan, has been reported (Pertovaara, 1998; Urban et al.,
Radhakrishnan et al. Page 7
Pain. Author manuscript; available in PMC 2009 August 26.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

1996, 1999). The chronic phase of hyperalgesia probably is maintained by the centralsensitization.
4.3. Dose dependence of hyperalgesia and inflammationOur data also indicate that the duration and contralateral spread of chronic hyperalgesiadepends on the severity of initial inflammation produced by carrageenan and the site ofinjection, i.e. deep tissues. In a different study from our laboratory (Sluka, 2002), capsaicininjected into cutaneous tissue produces only short-lasting secondary mechanical hyperalgesiawhereas injection of capsaicin into deep tissues, muscle or joint produces a long-lastingbilateral mechanical hyperalgesia in rats. Zymosan injected around the sciatic nerve at a lowdose produces ipsilateral mechanical hyperalgesia whereas higher doses produce a bilateralhyperalgesia that is not a result of systemic spread of Zymosan (Chacur et al., 2001). A dose-dependent increase in hyperalgesia induced by carrageenan paw inflammation also occurs butremains ipsilateral and is short-lasting (Hargreaves et al., 1988). Further, CFA or calcitoningene-related peptide injected ipsilaterally into the joint dose-dependently producescontralateral inflammation and hyperalgesia that depends on central neural mechanisms(Donaldson et al., 1993, 1995; Rees et al., 1996; Bileviciute et al., 1993, 1998). It has to be re-emphasized in this context that, in the current study, there was a contralateral spread ofmechanical and heat hyperalgesia with 3% carrageenan, both in muscle and joint-injectedanimals, whereas, there was no contralateral spread with the lower concentrations (0.3 and 1%)and with skin injections. It may, therefore, be hypothesized that, greater the initial tissue insultand deeper the injected tissue, hyperalgesia will be more chronic and bilateral.
4.4. Contralateral spread of hyperalgesiaBilateral effects of unilateral injury have been reported and reviewed by many investigators(Fitzgerald, 1982; Woolf, 1983; Coderre and Melzack, 1985; Kayer and Guilbaud, 1987;Bileviciute et al., 1993; Donaldson et al., 1993; Mapp et al., 1993; Kissin et al., 1998; Chen etal., 1999, Decaris et al., 1999; Donaldson, 1999, Koltzenburg et al., 1999; Lowrie, 1999;Chacur et al., 2001; Sluka et al., 2001; Sluka, 2002). C-fiber stimulation of the nerve to thegastrocnemius—soleus muscle or application of mustard oil intraarticularly produces bilateralincreases in the flexion reflex (Woolf, 1983; Woolf and Wall, 1986). The contralateral increasein the flexion reflex is unaffected by blockade of afferent input from the site of injury (Woolf,1983). Similarly, Sluka et al. (2001) showed that unilateral injection of acidic saline producesa long-lasting bilateral hyperalgesia in rats, which is not abolished by lidocaine injection intothe gastrocnemius muscle or unilateral dorsal rhizotomy. Also, capsaicin injected into themuscle or knee joint produces a long-lasting bilateral mechanical, but not heat hyperalgesia(Sluka, 2002). This mechanical hyperalgesia is dependent on the early activation of the cAMPpathway spinally during the first 24 h after capsaicin injection (Sluka, 2002). Chen et al.(2000) showed that a unilateral subcutaneous injection of bee venom into the plantar surfaceof the hindpaw produces bilateral heat and mechanical hyperalgesia in rats, after 4 h. Thecontralateral hyperalgesia is not abolished after ipsilateral sciatic nerve axotomy, but thedevelopment of contralateral effect is prevented by prior administration of N-methyl-D-aspartate (NMDA) or non-NMDA receptor antagonists intrathecally, indicating a centralsensitization mediated by spinal excitatory amino acids (Chen et al., 2000). Thus, contralateralspread of hyperalgesia likely depends on plastic changes in the central nervous system.
Following carrageenan-induced hindpaw inflammation in rats, phosphorylated-cAMP-responsive element binding protein (P-CREB) increases bilaterally in the spinal cord(Messersmith et al., 1998). Although an increase in P-CREB is not a sufficient parameter, it isa necessary factor for nociceptive-specific increases in c-fos expression (Messersmith et al.,1998), which could increase neuronal excitability and hence, hyperalgesia. Bilateral increasesin substance P and calcitonin gene-related peptide were also observed in the spinal dorsal horn
Radhakrishnan et al. Page 8
Pain. Author manuscript; available in PMC 2009 August 26.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

during the acute phase of experimentally induced monoarthritis in rat knee (Mapp et al.,1993) and for up to 1 week following carrageenan knee joint inflammation (Sluka andWestlund, 1993). Thus, bilateral changes in transcription factors and neurotransmitters maycontribute to the long-lasting, bilateral hyperalgesia associated with carrageenan-inducedmuscle or joint inflammation.
Although most of the above mentioned mechanisms and other reviews (Koltzenburg et al.,1999) on the bilateral effects of unilateral injury points towards a spinal role, a supraspinalmechanism cannot be ruled out, since supraspinal centers are involved in the development andmaintenance of secondary hyperalgesia (Herrero and Cervero, 1996; Pertovaara, 1998; Urbanet al., 1996, 1999). For instance, A- and C-fiber-mediated wind-up of flexor motoneuronsfollowing knee joint injection of carrageenan is prevented by spinal transection (Herrero andCervero, 1996). Further, inactivation of the rostral ventral medulla (RVM) by lidocainereverses and lesion of RVM by pre-treatment with ibotenic acid completely blocks secondaryheat hyperalgesia produced by knee joint injection of carrageenan (Urban et al., 1999). Thesemanipulations in the RVM do not affect the primary hyperalgesia produced by carrageenaninjected into the plantar paw. Thus, supraspinal centers play a major role in the production andmaintenance of secondary hyperalgesia. It is quite possible that the contralateral hyperalgesiaobserved in the current study could be mediated by RVM or other supraspinal centers throughdescending facilitatory pathways, although we do not have data to support these suggestions.
Interestingly, the contralateral hyperalgesia did not occur until 1–2 weeks after induction ofinflammation. The reasons for this are unclear, but may relate to conversion of the inflammatoryresponse from acute to chronic, since the contralateral spread follows the same time course.Alternatively, this could result from initiation of gene transcription mediating plastic changesin the central nervous system as a consequence of the tissue injury.
4.5. Histological changesHistopathological examination of the tissues in the current study shows inflammatory changesthat parallel the long-lasting hyperalgesia observed. The chronic hyperalgesia, therefore, couldbe maintained by chronic inflammation observed in the tissues. However, there are nocontralateral signs of inflammation observed in our studies, supporting a neuronal role, eitherspinal or supraspinal, for the contralateral spread of hyperalgesia. Several previous studiesshow a bilateral development of articular inflammation and degeneration after a unilateralinjection of CFA (Donaldson et al., 1995; Decaris et al., 1999). The absence of the contralateralhistological changes in our studies could be due to the difference in the inflammatory agentused, i.e. carrageenan vs. CFA. The histopathological changes observed in the present studycorrelate well with the hyperalgesic time periods observed with 3% carrageenan injection. Bothmuscle and joint tissues on the ipsilateral side show the earliest signs of acute inflammationaround 4–8 h, changing to chronic inflammation by 1–2 weeks that persists for up to 8 weeks.Both muscle and joint groups show hyperalgesia starting by 4 h, with the 3% group showingcontinued hyperalgesia through 7–8 weeks.
5. ConclusionFindings from this study clearly show a dose-dependent development of chronic hyperalgesiafollowing injection of carrageenan into muscle or joint in the rat and contralateral spread ofhyperalgesia at the higher dose. The data also support the earlier findings that injury to deepertissues result in a robust and long-lasting contralateral hyperalgesia compared to cutaneousinsult. The study provides animal models for acute as well as chronic hyperalgesia, one inducedby muscle inflammation and the other by joint inflammation with carrageenan, both of whichare possibly maintained by spinal or supraspinal neuronal mechanisms.
Radhakrishnan et al. Page 9
Pain. Author manuscript; available in PMC 2009 August 26.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

AcknowledgementsThe authors wish to thank Tammy Lisi, Chris Bromley and Jan Rogers for excellent technical assistance and Ms CarolLeigh for assistance with manuscript preparation. This work was supported by National Institutes of Health grantsR01 NS39734 and K02 AR02201 (KAS).
ReferencesBendele A, McComb J, Gould T, McAbee T, Sennello G, Chlipala E, Guy M. Animal models of arthritis:
relevance to human disease. Toxicol Pathol 1999;27:134–42. [PubMed: 10367688]Berberich P, Hoheisel U, Mense S. Effects of a carrageenan-induced myositis on the discharge properties
of group III and IV muscle receptors in the cat. J Neurophysiol 1988;59:1395–409. [PubMed: 3385466]Bileviciute I, Lundeberg T, Ekblom A, Theodorsson E. Bilateral changes of substance P-, neurokinin A-,
calcitonin gene-related peptide- and neuropeptide Y-like immunoreactivity in rat knee joint synovialfluid during acute monoarthritis. Neurosci Lett 1993;153:37–40. [PubMed: 8510822]
Bileviciute I, Stenfors C, Thodorsson E, Lundeberg T. Unilateral injection of calcitonin gene-relatedpeptide (CGRP) induces bilateral oedema formation and release of CGRP-like immunoreactivity inthe rat hindpaw. Br J Pharmacol 1998;125:1304–12. [PubMed: 9863661]
Chacur M, Milligan ED, Gazda LS, Armstrong C, Wang H, Tracey KJ, Maier SF, Watkins LR. A newmodel of sciatic inflammatory neuritis (SIN): induction of unilateral and bilateral mechanical allodyniafollowing acute unilateral peri-sciatic immune activation in rats. Pain 2001;94:231–44. [PubMed:11731060]
Chen H-S, Chen J, Sun Y-Y. Contralateral heat hyperalgesia induced by unilateral bee venom injectionis produced by central changes: a behavioral study in the conscious rat. Neurosci Lett 2000;284:45–8. [PubMed: 10771158]
Chen J, Luo C, Li HL, Chen HS. Primary hyperalgesia to mechanical and heat stimuli followingsubcutaneous bee venom injection into the plantar surface of hindpaw in the conscious rat: acomparative study with the formalin test. Pain 1999;83:67–76. [PubMed: 10506673]
Coderre TJ, Melzack R. Increased pain sensitivity following heat injury involves a central mechanism.Behav Brain Res 1985;15:259–62. [PubMed: 4005034]
Coderre TJ, Wall PD. Ankle joint urate arthritis (AJUA) in rats: an alternative animal model of arthritisto that produced by Freund’s adjuvant. Pain 1987;28:379–93. [PubMed: 3574965]
Craig AD, Hepplemann B, Schaible H-G. The projection of the medial and posterior articular nerves ofthe cat’s knee to the spinal cord. J Comp Neurol 1988;276:279–88. [PubMed: 2464629]
Crowe WE, Battisto JR, Smith RN. The autologous mixed lymphocyte reaction is decreased in Freund’sadjuvant-injected rats of arthritis-susceptible and -insusceptible strains. Arthritis Rheum1985;28:537–41. [PubMed: 3159391]
De Castro Costa M, De Sutter P, Gybels J, Van Hees J. Adjuvant-induced arthritis in rats: a possibleanimal model of chronic pain. Pain 1981;10:173–85. [PubMed: 7267134]
Decaris E, Guingamp C, Chat M, Philippe L, Grillasca JP, Abid A, Minn A, Gillet P, Netter P, TerlainB. Evidence for neurogenic transmission inducing degenerative cartilage damage distant from localinflammation. Arthritis Rheum 1999;42:1951–60. [PubMed: 10513812]
Di Rosa M, Giroud JP, Willoughby DA. Studies of the mediators of the acute inflammatory response. JPathol 1971;104:15–29. [PubMed: 4398139]
Diehl B, Hoheisel U, Mense S. Histological and neurophysiological changes induced by carrageenan inskeletal muscle of cat and rat. Agents Actions 1988;25:210–3. [PubMed: 3218593]
Donaldson LF, Seckl JR, McQueen DS. A discrete adjuvant-induced monoarthritis in the rat: effects ofadjuvant dose. J Neurosci Methods 1993;49:5–10. [PubMed: 8271831]
Donaldson LF, McQueen DS, Seckl JR. Neuropeptide gene expression and capsaicin-sensitive primaryafferents: maintenance and spread of adjuvant arthritis in the rat. J Physiol 1995;486:473–82.[PubMed: 7473211]
Donaldson LF. Unilateral arthritis: contralateral effects. Trends Neurosci 1999;22:495–6. [PubMed:10530993]
Radhakrishnan et al. Page 10
Pain. Author manuscript; available in PMC 2009 August 26.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Dougherty PM, Willis WD. Enhanced responses of spinothalamic tract neurons to excitatory amino acidsaccompany capsaicin-induced sensitization in the monkey. J Neurosci 1992;12:883–94. [PubMed:1545244]
Fitzgerald M. Alterations in the ipsi- and contralateral afferent inputs of dorsal horn cells produced bycapsaicin treatment of one sciatic nerve in the rat. Brain Res 1982;248:97–107. [PubMed: 6289998]
Garry MG, Hargreaves KM. Enhanced release of immunoreactive CGRP and substance P from spinaldorsal horn slices occurs during carrageenan inflammation. Brain Res 1992;582:139–42. [PubMed:1379871]
Gopalkrishnan P, Sluka KA. Effect of varying frequency, intensity and pulse duration of TENS on primaryhyperalgesia in inflamed rats. Arch Phys Med Rehabil 2000;81:984–90. [PubMed: 10896017]
Guilbaud G, Benoist JM, Eschalier A, Gautron M, Kayser V. Evidence for peripheral serotonergicmechanisms in the early sensitization after carrageenin-induced inflammation: electrophysiologicalstudies in the ventrobasal complex of the rat thalamus using a potent specific antagonist of peripheral5-HT receptors. Brain Res 1989;502:187–97. [PubMed: 2819456]
Guthrie AJ, Short CR, Swan GE, Mulders MS, Killeen VM, Nurton JP. Characterization of a sterile soft-tissue inflammation model in thoroughbred horses. J Vet Pharmacol Ther 1996;19:44–9. [PubMed:8992025]
Haak T, Delverdier M, Amardeilh MF, Oswald IP, Toutain PL. Pathologic study of an experimentalcanine arthritis induced with complete Freund’s adjuvant. Clin Exp Rheumatol 1996;14:633–41.[PubMed: 8978958]
Hargreaves K, Dubner R, Brown F, Flores C, Jones J. A new and sensitive method for measuring thermalnociception in cutaneous hyperalgesia. Pain 1988;32:77–88. [PubMed: 3340425]
Hay C, de Belleroche J. Carrageenan-induced hyperalgesia is associated with increased cyclo-oxygenase-2 expression in spinal cord. NeuroReport 1997;8:1249–51. [PubMed: 9175123]
Herrero JF, Cervero F. Supraspinal influences on the facilitation of rat nociceptive reflexes induced bycarrageenan monoarthritis. Neurosci Lett 1996;209:21–4. [PubMed: 8734900]
Hoheisel U, Mense S, Simons DG, Yu X-M. Appearance of new receptive fields in rat dorsal horn neuronsfollowing noxious stimulation of skeletal muscle: a model for referral of muscle pain. Neurosci Lett1993;153:9–12. [PubMed: 8510831]
Hoheisel U, Sander B, Mense S. Blockade of nitric oxide synthase differentially influences backgroundactivity and electrical excitability in rat dorsal horn neurones. Neurosci Lett 1995;188:143–6.[PubMed: 7540740]
Hong SK, Han JS, Min SS, Hwang JM, Kim YI, Na HS, Yoon YW, Han HC. Local neurokinin-1 receptorin the knee joint contributes to the induction, but not maintenance, of arthritic pain in the rat. NeurosciLett 2002;322:21–4. [PubMed: 11958834]
Kayer V, Guilbaud G. Local and remote modifications of nociceptive sensitivity during carrageenan-induced inflammation in the rat. Pain 1987;28:99–107. [PubMed: 3822499]
Kehl LJ, Trempe TM, Hargreaves KM. A new animal model for assessing mechanisms and managementof muscle hyperalgesia. Pain 2000;85:333–43. [PubMed: 10781907]
Keystone EC, Schorlemmer HU, Pope C, Allison AC. Zymosan-induced arthritis: a model of chronicproliferative arthritis following activation of the alternative pathway of complement. Arthritis Rheum1977;20:1396–401. [PubMed: 911357]
Kissin I, Lee SS, Bardley EL. Effect of prolonged nerve block on inflammatory hyperalgesia in rats:prevention of late hyperalgesia. Anesthesiology 1998;88:224–32. [PubMed: 9447876]
Koltzenburg M, Wall PD, McMahon SB. Does the right side know what the left is doing? Trends Neurosci1999;22:122–7. [PubMed: 10199637]
Lawand NB, McNearney T, Westlund KN. Amino acid release into the knee joint: key role in nociceptionand inflammation. Pain 2000;86:69–74. [PubMed: 10779662]
Lowrie MB. Contralateral effects of peripheral nerve injury. Trends Neurosci 1999;22:496–7. [PubMed:10529816]
Mapp PI, Terenghi G, Walsh DA, Chen ST, Cruwys SC, Garrett N, Kidd BL, Polak JM, Blake DR.Monoarthritis in the rat knee induces bilateral and time-dependent changes in substance P andcalcitonin gene-related peptide immunoreactivity in the spinal cord. Neuroscience 1993;57:1091–6.[PubMed: 7508583]
Radhakrishnan et al. Page 11
Pain. Author manuscript; available in PMC 2009 August 26.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Marchettini P, Simone DA, Caputi G, Ochoa JL. Pain from excitation of identified muscle nociceptorsin humans. Brain Res 1996;75:109–16. [PubMed: 8973804]
Mense S, Craig AD. Spinal and supraspinal terminations of primary afferent fibers from thegastrocnemius—soleus muscle in the cat. Neuroscience 1988;26:1023–35. [PubMed: 3200424]
Mense S. Nociception from skeletal muscle in relation to clinical muscle pain. Pain 1993;54:241–89.[PubMed: 8233542]
Messersmith DJ, Kim DJ, Iadarola MJ. Transcription factor regulation of prodynorphin gene expressionfollowing rat hind paw inflammation. Brain Res Mol Brain Res 1998;53:259–69.
Moreno JJ. Effect of aristolochic acid on arachidonic acid cascade and in vivo models of inflammation.Immunopharmacology 1993;26:1–9. [PubMed: 8407280]
Muir VY, Dumonde DC. Different strains of rats develop different clinical forms of adjuvant disease.Ann Rheum Dis 1982;41:538–43. [PubMed: 6812510]
Nantel F, Denis D, Gordon R, Northey A, Cirino M, Metters KM, Chan CC. Distribution and regulationof cyclooxygenase-2 in carrageenan-induced inflammation. Br J Pharmacol 1999;128:853–9.[PubMed: 10556918]
Neil A, Benoist JM, Kayser V, Guilbaud G. Initial nociceptive sensitization in carrageenin-induced ratpaw inflammation is dependent on amine autacoid mechanisms: electrophysiological and behaviouralevidence obtained with a quaternary antihistamine, thiazinamium. Exp Brain Res 1987;65:343–51.[PubMed: 2881800]
Neugebauer V, Schaible HG. Evidence for a central component in the sensitization of spinal neuronswith joint input during development of acute arthritis in cat’s knee. J Neurophysiol 1990;64:299–311. [PubMed: 2388073]
Nicklin S, Miller K. Effect of orally administered food-grade carrageenans on antibody-mediated andcell-mediated immunity in the inbred rat. Food Chem Toxicol 1984;22:615–21. [PubMed: 6381264]
O’Brien C, Woolf CJ, Fitzgerald M, Lindsay RM, Molander C. Differences in the chemical expressionof rat primary afferent neurons which innervate skin, muscle or joint. Neuroscience 1989;32:493–502. [PubMed: 2555742]
Pearson CM, Wood FD. Studies on polyarthritis and other lesions induced in rats by injection ofmycobacterial adjuvant. I. General clinical and pathologic characteristics and some modifyingfactors. Arthritis Rhuem 1959;2:440–59.
Pertovaara A. A neuronal correlate of secondary hyperalgesia in the rat spinal dorsal horn is submodalityselective and facilitated by supraspinal influence. Exp Neurol 1998;149:193–202. [PubMed:9454628]
Philippe L, Gegout-Pottie P, Guingamp C, Bordji K, Terlain B, Netter P, Gillet P. Relations betweenfunctional, inflammatory, and degenerative parameters during adjuvant arthritis in rats. Am J Physiol1997;273:R1550–6. [PubMed: 9362323]
Plenderleith MB, Snow PJ. The plant lectin Bandeiraea simplicifolia I-B4 identifies a subpopulation ofsmall diameter primary sensory neurones which innervate the skin in the rat. Neurosci Lett1993;159:17–20. [PubMed: 8264961]
Rees H, Sluka KA, Lu Y, Westlund KN, Willis WD. Dorsal root reflexes in articular afferents occurbilaterally in a chronic model of arthritis in rats. J Neurophysiol 1996;76:4190–3. [PubMed: 8985913]
Rivot JP, Montagne-Clavel J, Besson JM. Subcutaneous formalin and intraplantar carrageenan increasenitric oxide release as measured by in vivo voltammetry in the spinal cord. Eur J Pain 2002;6:25–34.[PubMed: 11888225]
Santer V, Sriratana A, Lowther DA. Carrageenan induced arthritis: V. A morphologic study of thedevelopment of inflammation in acute arthritis. Semin Arthritis Rheum 1983;13:160–8. [PubMed:6673111]
Schaible H-G, Ebersberger A, Von Banchet GS. Mechanisms of pain in arthritis. Ann N Y Acad Sci2002;966:343–54. [PubMed: 12114291]
Schaible H-G, Grubb BD. Afferent and spinal mechanisms of joint pain. Pain 1993;55:5–54. [PubMed:8278210]
Schaible H-G, Jarrott B, Hope PJ, Duggan AW. Release of immunoreactive substance P in the spinalcord during development of acute arthritis in the knee joint of the cat: a study with antibodymicroprobes. Brain Res 1990;529:214–23. [PubMed: 1704282]
Radhakrishnan et al. Page 12
Pain. Author manuscript; available in PMC 2009 August 26.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Schaible H-G, Schmidt RF, Willis WD. Enhancement of the responses of ascending tract cells in the catspinal cord by acute inflammation of the knee joint. Exp Brain Res 1987;66:489–99. [PubMed:3475211]
Schaible H-G, Schmidt RF. Effects of an experimental arthritis on the sensory properties of fine articularafferent units. J Neurophysiol 1985;54:1109–22. [PubMed: 4078610]
Schaible H-G, Schmidt RF. Time course of mechanosensitivity changes in articular afferents during adeveloping experimental arthritis. J Neurophysiol 1988;60:2180–94. [PubMed: 3236065]
Sluka KA. Stimulation of deep somatic tissue with capsaicin produces long-lasting mechanical allodyniaand heat hypoalgesia that depends on early activation of the cAMP pathway. J Neurosci2002;22:5687–93. [PubMed: 12097520]
Sluka KA, Christy MR, Peterson WL, Rudd SL, Troy SM. Reduction of pain related behaviors with eithercold or hot treatment in an animal model of acute arthritis. Arch Phys Med Rehabil 1999;80:313–7.[PubMed: 10084440]
Sluka KA, Kalra A, Moore SA. Unilateral intramuscular injections of acidic saline produce a bilateral,long-lasting hyperalgesia. Muscle Nerve 2001;24:37–46. [PubMed: 11150964]
Sluka KA, Westlund KN. An experimental arthritis in rats: dorsal horn aspartate and glutamate increases.Neurosci Lett 1992;145:141–4. [PubMed: 1361220]
Sluka KA, Westlund KN. Behavioral and immunohistochemical changes in an experimental arthritismodel in rats. Pain 1993;55:367–77. [PubMed: 7510059]
Sorkin LS, Westlund KN, Sluka KA, Dougherty PM, Willis WD. Neural changes in acute arthritis inmonkeys. IV. Time-course of amino-acid release into the lumbar dorsal horn. Brain Res Rev1992;17:39–50. [PubMed: 1638274]
Svensson P, Beydoun A, Morrow TJ, Casey KL. Human intramuscular and cutaneous pain:psychophysical comparisons. Exp Brain Res 1997;114:390–2. [PubMed: 9166929]
Swingle KF, Grant TJ. Evaluation of some standard arthritogenic adjuvants (‘modified perrigens’) inmale rats. Agents Actions 1977;7:459–63. [PubMed: 412400]
Swingle KF, Jaques LW, Kvam DC. Differences in the severity of adjuvant arthritis in four strains ofrats. Proc Soc Exp Biol Med 1969;132:608–12. [PubMed: 5355113]
Torebjork HE, Ochoa JL, Schady W. Referred pain from intraneural stimulation of muscle fascicles inthe median nerve. Pain 1984;18:145–56. [PubMed: 6709382]
Urban MO, Jiang MC, Gebhart GF. Participation of central descending nociceptive facilitatory systemsin secondary hyperalgesia produced by mustard oil. Brain Res 1996;737:83–91. [PubMed: 8930354]
Urban MO, Zahn PK, Gebhrat GF. Descending facilitatory influences from the rostral medial medullamediate secondary, but not primary hyperalgesia in the rat. Neuroscience 1999;90:349–52. [PubMed:10215139]
Wall PD, Woolf CJ. Muscle but not cutaneous C-afferent input produces prolonged increases in theexcitability of the flexion reflex in the rat. J Physiol 1984;356:443–58. [PubMed: 6520794]
Willis, WD.; Coggeshall, RE. Sensory mechanisms of the spinal cord. Plenum Press; New York, NY:1991.
Winter CA, Risley EA, Nuss GW. Carrageenin-induced edema in hind paw of rat as an assay forantiinflammatory drugs. Proc Soc Exp Biol Med 1962;111:544–7. [PubMed: 14001233]
Witting N, Svensson P, Gottrup H, Arendt-Nielsen L, Jensen TS. Intramuscular and intradermal injectionof capsaicin: a comparison of local and referred pain. Pain 2000;84:407–12. [PubMed: 10666547]
Woolf CJ. Evidence for a central component of post-injury pain hypersensitivity. Nature 1983;306:686–8. [PubMed: 6656869]
Woolf CJ, Wall PD. Relative effectiveness of C primary afferent fibers of different origins in evoking aprolonged facilitation of the flexor reflex in the rat. J Neurosci 1986;6:1433–42. [PubMed: 3711988]
Yang LC, Marsala M, Yaksh TL. Characterization of time course of spinal amino acids, citrulline andPGE2 release after carrageenan/kaolin-induced knee joint inflammation: a chronic microdialysisstudy. Pain 1996;67:345–54. [PubMed: 8951928]
Zimmermann M. Ethical guidelines for investigations of experimental pain in conscious animals. Pain1983;16:109–10. [PubMed: 6877845]
Radhakrishnan et al. Page 13
Pain. Author manuscript; available in PMC 2009 August 26.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
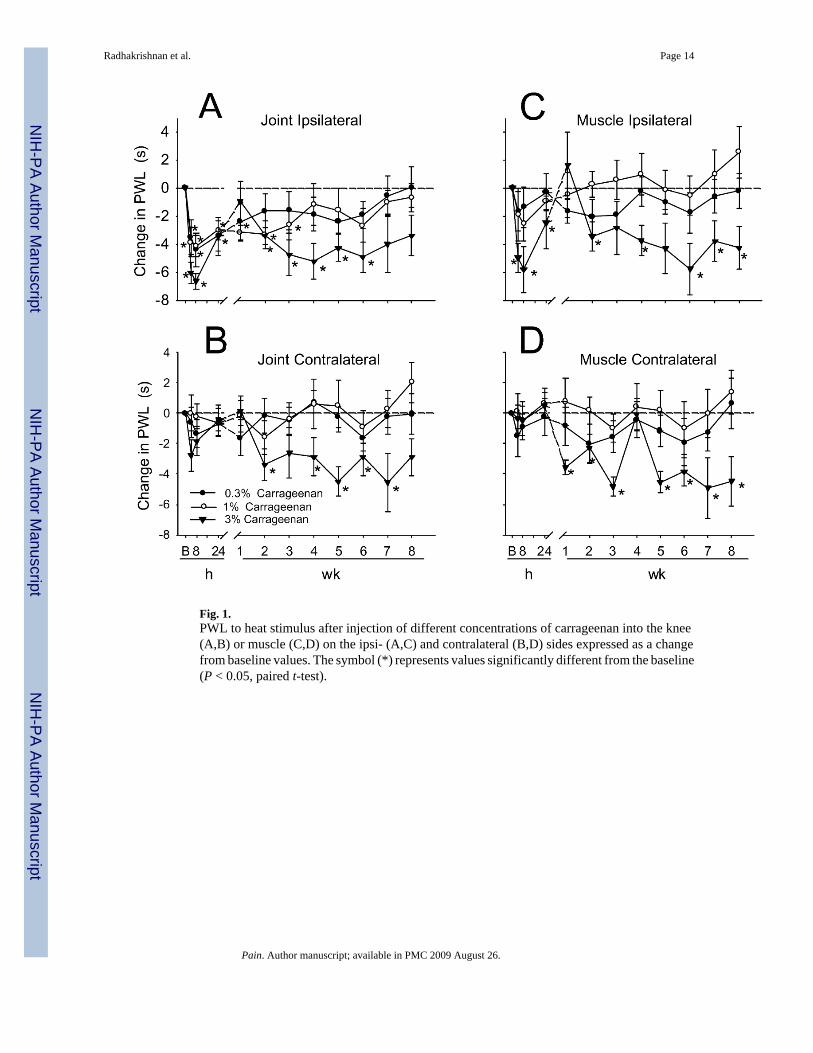
Fig. 1.PWL to heat stimulus after injection of different concentrations of carrageenan into the knee(A,B) or muscle (C,D) on the ipsi- (A,C) and contralateral (B,D) sides expressed as a changefrom baseline values. The symbol (*) represents values significantly different from the baseline(P < 0.05, paired t-test).
Radhakrishnan et al. Page 14
Pain. Author manuscript; available in PMC 2009 August 26.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
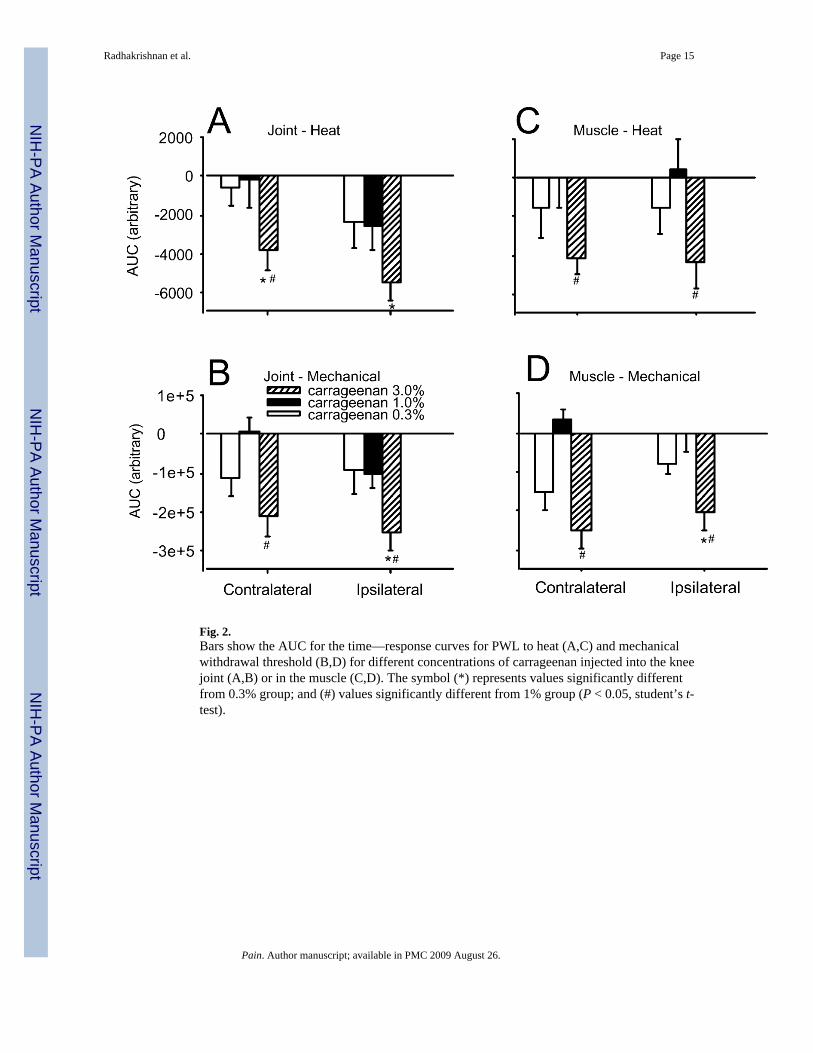
Fig. 2.Bars show the AUC for the time—response curves for PWL to heat (A,C) and mechanicalwithdrawal threshold (B,D) for different concentrations of carrageenan injected into the kneejoint (A,B) or in the muscle (C,D). The symbol (*) represents values significantly differentfrom 0.3% group; and (#) values significantly different from 1% group (P < 0.05, student’s t-test).
Radhakrishnan et al. Page 15
Pain. Author manuscript; available in PMC 2009 August 26.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
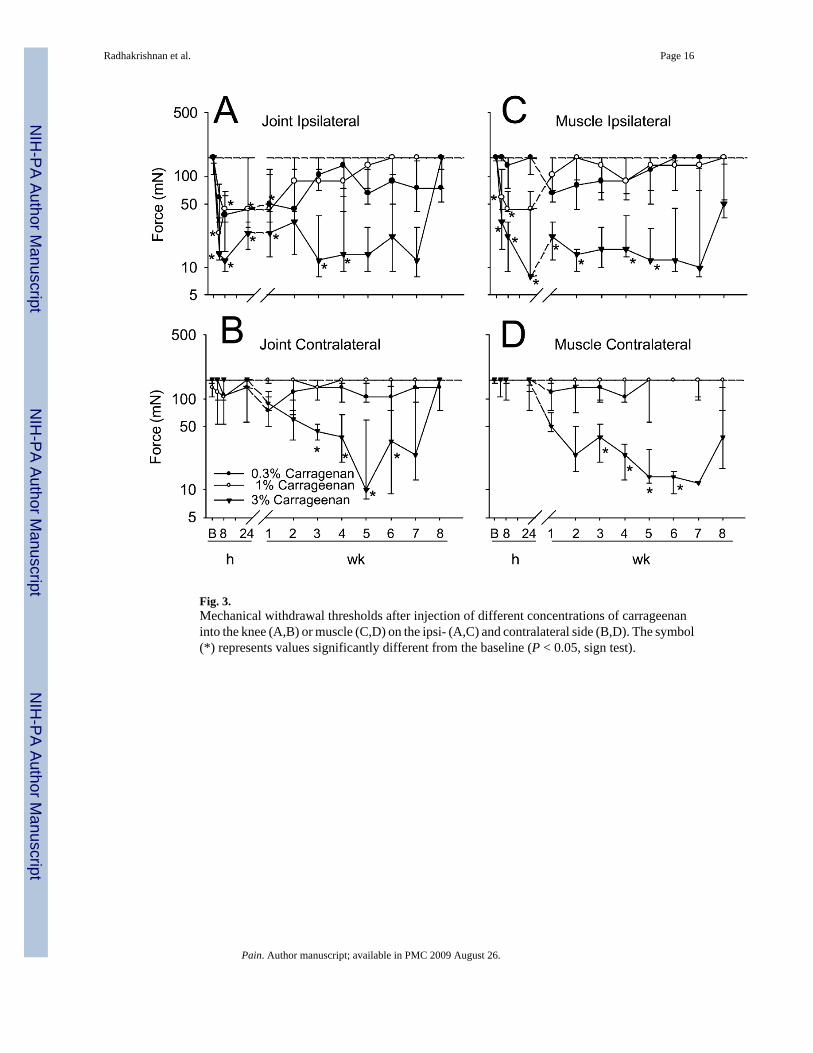
Fig. 3.Mechanical withdrawal thresholds after injection of different concentrations of carrageenaninto the knee (A,B) or muscle (C,D) on the ipsi- (A,C) and contralateral side (B,D). The symbol(*) represents values significantly different from the baseline (P < 0.05, sign test).
Radhakrishnan et al. Page 16
Pain. Author manuscript; available in PMC 2009 August 26.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
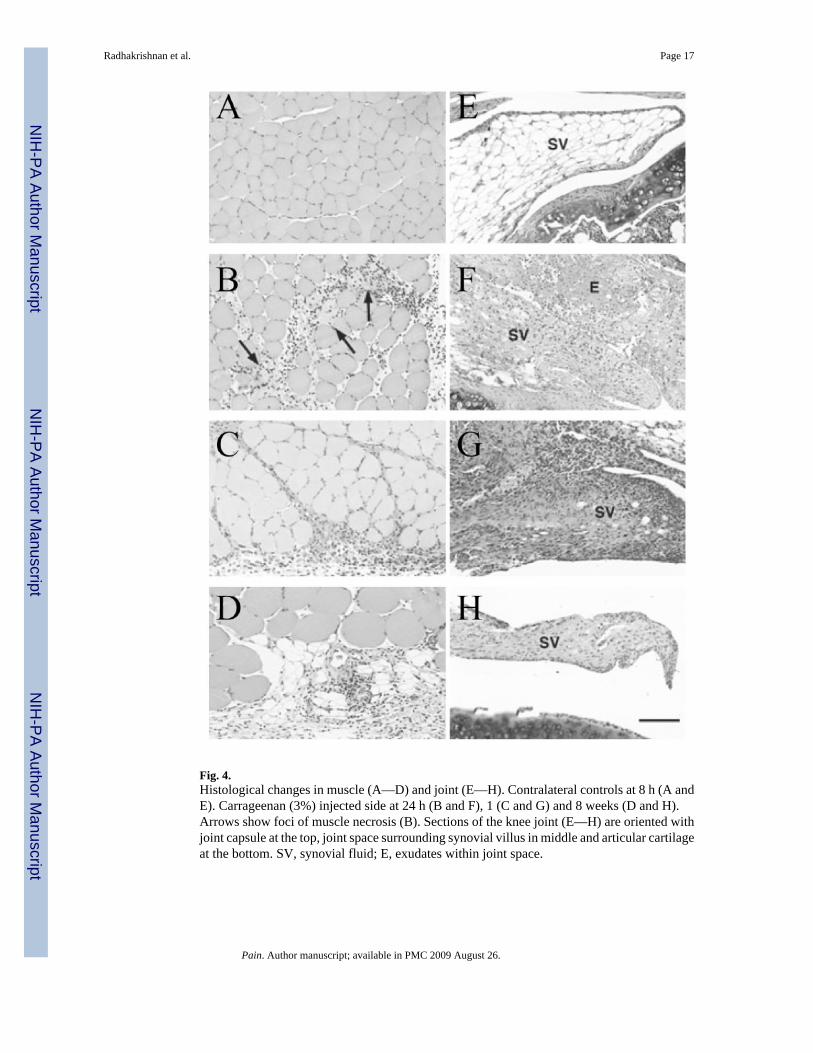
Fig. 4.Histological changes in muscle (A—D) and joint (E—H). Contralateral controls at 8 h (A andE). Carrageenan (3%) injected side at 24 h (B and F), 1 (C and G) and 8 weeks (D and H).Arrows show foci of muscle necrosis (B). Sections of the knee joint (E—H) are oriented withjoint capsule at the top, joint space surrounding synovial villus in middle and articular cartilageat the bottom. SV, synovial fluid; E, exudates within joint space.
Radhakrishnan et al. Page 17
Pain. Author manuscript; available in PMC 2009 August 26.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript





























