Embed Size (px)
Citation preview

(Aus dem Physiologischen Laboratorium der Biologischen Station an der Murmankiiste.)
Untersuehungen iiber den respiratorischen Gasweehsel bei B alanus crenatus bei verschiedenem Salzgehalt
des Aullenmilieus. I. Mitteilung.
(~ber den Sauerstoffverbrauch im Wassermflieu bei verschiedenem Salzgehalt.
Von
Eugen Kreps. Mit 4 T e x t a b b i l d u n g e n .
('Eingegangen am 3. Dezember 1928.)
I~ .R. H6ber 1 bezeichnet die Seewirbellosen als ,,poikilosmotische" Tiere
wegen ihrer schwaeh entwickelten osmoregulatorischen Funktion. Da die Seewirbellosen unf~hig sind, die osmotische Konzentration ihres inneren Milieus (milieu interieur im Sinne yon Claude Bernard) konstant zu erhalten, sind sie in der l~egel sehr empfindlich gegen alle scharfen Schwankungen des Salzgehalts in dem umgebenden Milieu. Dafiir sprechen sowohl die Verteilung der Formen in der Natur, als auch speziell ange- stellte Versuche. Die sch~dliehe Wirkung der Ver~nderung des Salz- gehalts, und speziell der VersfiBung, wurde an verschiedenen Seetieren yon vielen Forschern, angefangen mit Paul .Bert (Plateau, J. Loeb u. a.), studiert. ~Es ist daher begreiflich, dab gerade die ,,homoiosmotischen" Tiere sich als solche erweisen, die leicht aus Seewasser in SiiBwasser und umgekohrt fibergehen; hierzu gehSren haupts~chlich Fische, z. B. Wan- derfische, die zur Laiehzeit aus der See in Flfisse herfiberziehen. Die See- wirbellosen haben bei weitem in den meisten F~llen keine Gelegenheit , einigermaBen starken osmotischen Schwankungen im AuBenmilieu zu begegnen.
Einzelne Formen aus der groBen Menge yon den Evertebraten, die in der Regel flit die Ver~nderung der Meerwassersaliniti~t sehr emp- findlich sind, erregen jedoch unsere Aufmerksamkeit dureh eine unge-
1 R. Hbr Physikalische Chemie der Zelle und der Gewebe. 5. Aufl. 1922.

216 E. Kreps: Untersuchungen fiber den respira$orischen Gaswechsel
meine Stabilit/~t hhlsiehtlich der Salinit/~tsschwankungen der Umgebung. Einer besonders scharf ausgepr/~gten Eurichalinit/s begegnen wir bei einigen festsitzenden litoralen Formen, die infolge ihrer Existenzbedin- gungen best/indig sehr scharfe Schwankungen der Salzkonzentration des sie umspiilenden Wassers zu erleiden haben.
Eine spezielle Forschung, die yon uns an 2 Arten yon Ranken- fiiBern (Cirripedia), Balanus balanoides und Balanus erenatus 1, durch- gefiihrt wurde, zeigte, da$ diese Formen f/~hig sind, Salinit/~tssehwan- kungen yon vSllig siiBem Wasser bis 700/o0 zu ertragen, wobei besonders bemerkenswert ist, dab sie sogar scharfe und schnelle Uberg/~nge nieht ffirehten. Weiterhin wurde uns klar, dab eine gewisse minimale Schwel- lenkonzentration bestehe, deren Uberschreiten die normale Existenz der Balanl mit den ffir dieselben charakteristischen rhythmisehen Be- wegungen und der intensiven Atmungst/~tigkeit unmSglich maeht. Diese ,kritische Konzentration" ist ffir diese Arten sehr best/~ndig und 4 bis 60/00 gleich.
Es ergab sieh, dab bei einer weiteren Verminderung des Salzgehaltes des Auflenmilieus die Balani nicht umkommen, sondern in einen beson. deren Zustand i~bergehen, der dutch vollstdindige Unbeweglichkeit charak. terisiert wird. In einem solchen Zustand yon Scheintod oder ,,Salzsehlaf" kSnnen die Balani lange Zeit (nieht weniger als 3 Woehen) in vollkommen sfiI~em Wasser verweilen, indem sie bei der Riiekkehr in normale Be- dlngungen wieder aufleben. Dank dleser Eigentiimliehkeit k6nnen diese Formen ungiinstige Perioden starker VersiiBung vertragen und n~hern sich, was ihr Verh/iltnis zu schroffen Ver/inderungen der Salzkonzentra- tion betrifft, den Wanderfischen und anderen homoiosmotisehen Tieren. Jedoch die Unabhs yore Aul]enmilieu bei diesen und jenen ist auf ganz verschiedenen Meehanismen begrfindet.
Bei den homoiosmotischen Formen besteht dank der speziellen osmoregulatorisehen Vorrichtungen eine mehr oder weniger konstante osmotische Konzentration des Blutes und der entsprechenden K6rper- fliissigkeiten (Bottazzi, Loeb u.a.). Die Zellen dieser Tiere bleiben be- s$/~ndig in den Bedingungen eines praktisch unver/inderten gesamten osmotischen Druckes, sowie eines partiellen Druckes einzelner Salze.
Ganz anders steht die Frage bei den Balani. Eine Reihe von Ver- suchen, welehe das Austreten der Salze aus dem K6rper beim ~bergehen in ein weniger salzhaltiges Meerwasser behandeln, sowie die unmittel- bare krioskopische Untersuchung der Kfrperfliissigkeit zeigen, dal~ die osmotisehe Konzentration derselben eine Funktion der Salzkonzentra- tion im Aul3enmilieu ist ~. Die Ver/~nderung der Salzkonzentration im
1 E. Kreps, Arb. d. Ges. Natuff. zu Leningrad 55, 11 (1925) (russ.). 2 Die Beschreibung der diesbeziiglichen Versuche bildet den Gegens$~nd
einer besonderen Abhandlung.

bei Balanus crenatus bei verschiedenem Salzgehalt des Aul]enmilieus. I. 217
inneren Milieu, d. h. das, was bei dem wcit fiberragenden Teit der Tiere mit dem Tode und demnach mit Reaktionen irreversibler Art im Zellen- plasma verbunden ist, ffihrt bei dem Balanus und den fibrigen ibm ~hn- lichen Tieren blol] zu einer temporKren Funktionshemmung, zu irgend- welchen reversiblen l~rozessen, die nicht mit ernsten St6rungen der phy- sisehen nnd ehemisehen Zellenstruktur verbunden sind. Der yon uns beim Batanus beobachtete Zustand des Scheintodes in stark versfiBtem Milieu erinnert/iuBerlich an andere Zust~nde herabgesetzter Lebenst~tig- keit, wie an den Zustand des ,,Winterschlafes", an den Zustand vorfiber- gehenden Eintrockncns odor an anaerobe Lebensphasen einiger Orga- nismen. ])as weitere Studium der Frage fiber die Anpassung der Meer- eicheln an die Bedingungen des sehwankenden Salzgehaltes wurde nach verschiedenen Richtungen hin geffihrt und befaBte sich mit der Wirkung einzelner Ionen des Meerwassers (E. Krepsl), der Durchl~ssigkeit dcr KSrperoberfl~ehe des Balanus, den Schwankungen des osmotischen Druckes in der KSrperflfissigkeit und endlich mit der physiologischen Untersuchung des Zustandes des Seheintodes, der als ,,Salzschlaf" be- zeichnet werden kann.
II. Ein genaueres physiologisches Studium des ,,Salzschlafes" der Balani
wurde mit dcr Un~ersuehung des Atmens dieser Tiere begonnen, da das Atmen den zentralen Lebensproze2 vorstellt, der dem ganzen kompli- zierten 1Vfechanismus der Lebenserscheinungen zugrunde liegt. Vorlie- gende Mitteilung ist das Ergebnis einer Anzahl yon Versuchen fiber den Gaswechsel der Balani bei verschiedcner gesamter Salzkonzentration im Augenmilieu.
Wir wahlten fiir unsere Versuche 2 Gruppen yon Balanus crena$us; die 1. Gruppe aus 22 und die 2. Gruppe aus 24 an Dimensionen mehr oder weniger gleiehen auf 4 Mytilusmuseheln sitzenden Individuen. Die Oberfl/~ehe des lKalk- h/~uschens der Tiere und der leeren Muscheln warden sehr sorgf/~ltig von Algen und Bryozoa gereinigt, mit denen die Balanuskolonien gew6hnlich bewaehsen sind. ]3eide Gruppen waren am 3. VII. 1926 vom Ufer genommen worden und wurden bis zum 30. VIII. zum Versuch verwandt. Keines der Tiere kam urn, und zum SchluB der ganzen Versuehsserie batten alle Exemplare ein vollkommen gesundes und normales Aussehen, und ihr Verhalten in gew6hnHchem Meerwasser unterschied sich nieht yon dem Verhalten der eben yon der Litoralzone gebraehten Balani ~. Die 1. Gruppe diehte uns zu Versuchen mit mehr versiiBten, die 2. mit mehr salzhaltigen Milieus.
Unsere Versuche zeigten, dab beim Transport der Tiere aus ihrer natfirliehen Umgebung in aquariale Verh~ltnisse, die dazu dutch spezi-
1 E. Kreps, Arb. d. biolog. Station an der Murmankfiste 2, 25 (1926) (russ.). Bei einiger Effahrung und einiger Kenntnis der Balani und ihres Ver-
haltens, sowie der Art und des Rhythmus ihrer motorischen Ti~tigkeit kann der Beobaehter leicht versehiedene Abweichungen yore normalen Verhalten der Tiere merken (E. Kreps 1926).

218 E. Kreps: Untersuohungen tiber den respiratorischen Gaswechsel
fische Bedingungen des Respirationsversuchs erschwert waren, die Inten- sit~t der Lebensprozesse langsam, aber best~ndig geschw~cht wurde. Periodisch angestellte und bei mSglichst gleichen Standardbedingungen ausgefiihrte Respirationsversuche zeigten ein allm~hliches Fallen des Sauerstoffverbrauchs, das nach einer bestimmten Kurve verlief. Dieses mul~te bei dem endgiiltigen Berechnen der Resultate unserer Experi. mente fiber den Einflul] des Salzgeha]tes auf die Energie der Atmungs- prozesse in Betracht genommen und ein en~sprechendes Korrektiv ge- funden werden.
Unter den F~ktoren, die den Gaswechsel beeinflussen, ist bekanntlieh die Muskelarbeit yon besonders grol~er Bedeutung. Es ist daher natfir- ]ich, dab der Sauerstoffverbrauch w~hrend des Salzsehlafes bei voll- kommenem Ruhestand des Muskelsystems um viele Male geringer sein muB als bei st~rkeren Salinit~ten in den Verh~ltnissen einer kontinuier- lichen rhythmischen Bewegung der Rankenfiil~chen. Im weiteren ist es klar, dab es schwer ist, einzelne Versuche miteinander zu vergleichen, die bei verschiedenen, aber genfigend hohen Salinit~ten stattfinden, da die ungleichen Werte fiir den Sauerstoffverbrauch nieht als Resultat des unmittelbaren Einflusses der Salzkonzentration auf den Grundgas- wechsel erseheinen kSnnten, sondern als Folge einer ungleichen Muskel- arbeit, die sich in den Versuchsbedingungen quantitativ sehr sehwer bestimmen l~l]t. Es lag der Gedanke nahe, alle Experimente unter ~Tar- kose, z. B. Urethannarkose durchzufiihren. Krogh 1 hat die vollkommene Anwendbarkeit des Urethans bei Gasweehselexperimenten mit Wasser- tieren best~tigt, aber speziell die Balani erwiesen sich als sehr wenig emp- findlich gegen Ethyl-Urethan. Es mul~ten ungemein grol~e Dosen (mehr als .1,5% im Laufe yon 6 Stunden) angewendet werden, um eine vSllige Hemmung der motorischen T~tigkeit zu bewirkcn. Da aber unser Ar- beitsplan das Ausffihren einer ]angen Reihe yon Versuehen mit denselben Individuen in sich schlol], so schien es uns nicht zweckm~ltig, yon einer solchen chronischen Vergiftung der Tiere Gebraueh zu maehen. Ubrigens gab uns die Muskelarbeit nicht viel Bedenken, und zwar aus folgendem Grunde. Wie vorhergehende Beobaehtungen gezeigt haben ~, beginnen die Balani beim Transport aus einem Wasser mit voller ozeanischer Salinit~t (gegen 33--340/00) in versfiBtes Wasser mit einer Salinit~t unter" 10--11~ nicht vor dem 5.--6. Tage zu ,,atmen", d. h. mit den Rankenfiil~chen (cirri) rhythmische Bewegungen zu machen, die den Zweck haben, das Wasser in der MantelhShle zu erneuern. Bei S z 13--140/00 bcginnen die Atmungsbewegungen gew5hnlich am 3. Tage. Wenn die S unter 5--6~ ist - - kritische Konzentration - - , so setzen die Atmungsbewegungen nicht ein, solange die Tiere sich in
1 Krogh, Internat. Rev. d. Itydrobiel. 6, 42 (1914). Kreps, 1925 und 1926, I.e.

bei Balanus crenatus bei verschiedenem Salzgehalt des Aul]enmilieus. I. 219
dieser Salzkonzcntration befinden. Da unsere Hauptaufgabe in dem Stu- dium des Gaswechsels in stark versfiBten Milieus, diesseits und ]enseits der kritisehen Konzentration, d .h . in den Verh~ltnissen vollstandiger Bewegungslosigkeit bestand, so erschien die Frage yon dem Einflul] der Muskelarbeit als nicht besonders wesentlich.
Die Hauptfrage unserer Arbeit war folgende: Ob die Verdnderungen des Salzgehalts (folglieh auch des osmotischen Druckes) einen Ein/lu[3 au] den Gaswechsel eines voll]commen ruhenden Tieres ausi~ben, dessen funk- tiona]e Aktivitat auf das Minimum herabgesetzt ist ? Ob die GrS[3e des Standardmetabolismus, im Sinne yon Krogh, als eine Fun]ction der osmo- tischen Konzentration des Au]3enmilieus erscheine ? In der uns zug~ng- lichen Literatur fanden wir nun einzelne Daten fiber den EinfluB yon S auf die Intensit~t der Assimilation (Legendre 1, Fromageot ~) und des Atmens bei den Seealgen (Inman3). Diese oder jene Antwort auf die gestellten Fragen kormten jedoch ~ieles in dem Mechanismus der Ent- wicklung und im Wesen des Salzschlafes erkl~ren sowie iiberhaupt im Problem der Anpassung der Seewirbellosen an eine Existenz unter den Bedingungen schwankenden Salzgehaltes.
Der Frage fiber den EinfluB des Salzgehalts auf das Atmen der Wasser- tiere ist die interessante Arbeit von Schlieper 4 gewidmet. Dieser Autor weist auf eine Erschwerung der Atmung im SiiBwasser im Vergleich zum Salzwasser hin. Den Grund dafiir sieht Schlieper in einer erleichterten C02-Abgabe beim Atmen im Salzwasser, was auf einem reicherem Gehalt an Monocarbonaten, die die ausgesehiedene Kohlensaure chemiseh bin- den, beruht.
III.
Methodisehes. Die Methode der Respirationsversuche war sehr einfach. Ihr Prinzip ist alt
und stamm~ noch aus der Zcit Humboldts und Proven~als 5. Das Versuchsobjekt wird in ein bestimmtes, abgeschlosscncs Wasservolumcn gebracht, dessen Gas- bestand vor und nach dem Versuch analysiert wird. Was die Genauigkei~ der Resultate betrifft, so steht diese Methode den komplizierten Respirationsapparaten fiir Wassertiere nicht nach, die yon Jolyet und Regnard 6, Gr~hant ~, Vernon s, Zuntz 9,
1 Legendre, C. r. Soc. Biol. 85, 222 (1921). 2 F~omageot, C. r. Acad. Sci. 177, 779 (1923). - - Bull. Soc. de Chim. biol.
6, 169 (1924). 3 Inman, J. gcn. Physiol. 3, 663 und 4, 1 (1921).
Schlieper, Naturwiss. 1928, Nr 14. 5 Humboldt et Provencal, l~em. de Phys. ct de Chim. de la Soc. 4'Arcueil
1809. 6 Jolyet et t~egnard, Arch. de Physiol. 4, 44 (1877). 7 Gr~hant, C. r. Acad. Sci. 38, 421 (1886). s Vernon, J. of Physiol. 19, 18 (1895/96), o Zuntz, Arch. Anat. u. Physiol. 1901, 543.

2 2 0 E. Kreps: Untersuchungen fiber den respiratorischen Gaswechsel
Bounhiol 1 u. a. vorgeschlagen worcIen sind. A . Krogh 2, eine Autorit~t in den Fragen fiber den Gaswechsel, sagt von dieser Methode, dab sie ebenso einfach als vorzfiglieh sei (,,is as excellent as i t is simple").
Als Respirationsgef~Be dienten uns 2 Glaszylinder, die mit einem gut an- gesehliffenen Deckel lest verschlossen werdcn konnten. Das Volumen des Wassers be[ eingebrachten Tieren war fiir den I. Zyiinder = ~10 ecru, ffir den 2. = 750 com.
Fiir jeden Versuch wurden in einem Glasbehglter einige Liter Wasser yon einem bestimmten Salzgehalt bereitet, indem wir Meerwasser, das aus einer Tiefe yon 16 m (S = 33 Prom.) genommen wurde, mit verschiedenen Volumina sfil~en oder destilHerten Wassers verdfinnten. Die endgfiltige Salinit~t der erhaltenen Misehung wurde geprfift durch Titration mit A g N Q and nach K n u d s e n s Tabellen berechnet.
Das Wasser wurde filtriert, die Temperatur bis auf 6 ~ gebracht und das Wasser bei dieser Temperatur mittels Durchblasen von Aul]enluft durch einen porSsen Stein im Laufe yon 2 Stunden mit Sauerstoff bis auf den atmospharischen Sauerstoffdruck gess 1/z Stunde vor Begirm des Exper~mentcs wurde mi~ dem DurchbIasen aufgeh5rt. Auf diese Weise gingen wit hei allen Experimenten vom Zustand der Sauerstoffs~ttigung des Wassers aus.
Die Experimente yon Henze a u. a. habcn gezeigt, da/~ bei Wassertieren mi t guter Zirkulation und intensiver Kiemenatmung, sowie bei den halb durehsichtigen hyalinen pelagischen Formen mit einem sehr unbedeutenden Gehalt yon Trocken- substanz der O~-Verbrauch vom 02-Druck in weitem Mal~e unabhimgig ist. Bei Formen mit einem grol~en Gehalt yon Trockensubstanz und einem unvollkommenen Atmungs- und Zirkulati0nsapparat dagegen h~ngt der Os-Verbrauch deutlich yon dem Oz-Druck im Wasser ab. Nach den Angaben yon MacClenclon 4, sowie denjenigen yon Amberson, Mayerson und Scott a ist sogar bei vielen hSberen Wirbel- losen der Oz-Verbrauch dirckt proportional dcm 0~-Druck im Mcerwasser.
Die Versuchstiere wurden in das Respirationsgefa/3 gebracht und gleichzeitig das Gefal3 und mehrere (6--7) Winklersehe Flaschen yon 160--180 ccm Inhal t mittels mehrerer parallel geleiteter SiphonrShren mit dem bereiteten Wasser an- gefiillt. Darauf wurde das Gef~l~ sofort mit dem angeschliffenen Deckel luftdicht verschlossen, so dab unter dem Deckel nicht das geringste Luftbli~sehen fibrig blieb, und darauf das ganze Gef~l~ in einen grol3en Kfibel mit Wasser versenkt, dessen Temperatur = 6 ~ erhalten wurde. Die Temperaturschwankungcn waren im Laufe des Versuches nieht grO~er als • A~e Prozeduren, sowie d~s Ex- periment selbst, verliefen in einem kiihlen, sehwaeh erleuehteten Aquariumraum.
Von den gleichzeitig geffillten Winklerschen Flaschen wurden 3- -4 als Kon- trollglhser fiir die ganze Versuchsdauer in demselben Kfibel mit Wasser gestellt, die fibrigen aber wurden sofort zur Analyse des Wassers auf Oz- und CO~-Gehalt und zur p~-Bestimmung angewandt. Der Sauerstoff wurde naeh der Winklerschen l~Iethode, CO~ nach dem Verfahren yon Tornoe 6 bestimmt. Diese Methodc gibt die Menge ,,der gesamten Kohlensaure" im Meerwasser, d .h . die Summa der freien und der gebundenen COe an.
CO s-Bes t immung.
Nine unmittelbare Bestimmmlg der Kohlens~ure im Wasser wurde nicht in allen Versuchen ausgeffihrt. In den meisten Experimenten wurde der Gehal~
1 Bounhiol , Bull. seient. France et Belg. 39, 227 (1905). 2 Krogh, The Respiratory Exchange of Animal and 1VIam 1916, 49. 3 Henze, Biochem. Z. 26, 255 (1910). 4 MacClendon, J. of biol. Chem. 28, 135 (I916). 5 Amberson, Mayerson and Scott, J. gen. Physiol. 7, Nr 1 (1924). 6 Tornoe, Den norske Nordhavsexpedition 1876/78 I (1880).

bei Balanus crenatus bei verschiedenem Salzgehalt des Aul]enmilieus. I. 221
an gesamter Kohlens/~ure auf Grund der Alkalinitat und der Wasserstoffionen- konzentration mit l-Iilfe yon spezicllen Tabellen berechnet, welche die Abhiingig- keit zwischen der Alkalinit~t, der gesamten CO S, der freien COs und dem p~ im Mcerwasser feststellen. Solche Tabellcn sind auf Grund experimentcller Arbciten yon MacClendon 1, Kur t Buch ~ und E. Kreps ~ fiir Seewasser verschiedenen Salz- gehaltes ausgearbeitet worden.
Durch das Benutzen dieser Tabellen wird die Arbeit ungemein vereinfacht und das langwierige und umsts Bestimmen der gesamten CO S erschein~ dabei ats iiberfliissig. Das Bestimmen der Alkalinit/~t in unseren Vcrsuchen wurde gew6hnlich im Ausgangswasscr nach der Mcthode von Mac Clendon 1 mit dem Indicator Brom-Kresolpurpur gemacht.
Das PH des ~eerwassers wurde colorimetrisch nach der Methode yon Sorensen mit den Indicatoren yon Clark und /J~b~ bemesscn. Bei allen /~-Berechnungen wurde natiirlich der Salzfehler des Indicators beriicksichtigt und die entsprechende Korrektur durchgefiihrt. Als Indicator benutzten wir hauptsi~chlich Thymolblau und Kresolrot, welche praktisch denselben Salzfehler geben. Be] der colorimetri- schen Bestimmung der Reaktion des IVIeerwassers mit den Indicatoren yon Clark kann das p~ ohne Schwierigkeiten bis auf 0,02 fcstgestellt werden.
Leeren des Ge/Sfles.
Nach Schlul~ des Vcrsuches wurde das Glas geSffnet und das darin enthaltene Wasser schnell und gleichzeitig mittels raehrerer SiphonrShren in 3--4 Winklersche l~laschen hiniibergeleitct. 2 dersclben dienten der 0~-Bestimmung, die iibrigen der Bestimmung yon CO S und p~. Das Volumcn der Flaschen wurde in der Be- rechnung gew/thlt, dab bei ihrem Anfiillen das im Respirationszylinder cnthaltene Wasser ohne Rest verbraucht wurde. Letzterer UmsSand war yon groBer Wichtig- keit. Die Tiere, welche sich im Zylinder befinden, vcr~ndern den Gasbestand des sic umgebenden Wassers, wobei die ihnen n/~chstgelegenen benachbarten Schichten am meisten vers erscheinen. Das Ausgleichen des Gasdruckes in der Fliissigkeit vollzieht sich durch Diffusion schr langsam. Etwaige Konvektions- str6me, die zur Vermischung beitragen, sind infolge vollst~ndiger Homothermie nicht vorhanden. Es ist daher klar, dab man ungleiche Resu]tate erhalten wiirde, wenn roan die Prnben zur Analyse verschiedenen Wasserschichten entnehmcn wollte. Notwendigerweise mul~te man entweder das Wasser griindlich vermischen, ohne die Gcsamtmenge der gelSstcn Gase zu veriindern, oder aber das ganze im Glase enthaltene Wasser mul~te der Analyse unterworfcn werden.
Da uns keine speziellen komplizierten und kostspieligen Respirationsapparate mit Vorrichtungen zum bestandigen Zh'kulieren und Vermischen des Wasscrs zur Vcrfiigung standen, umgingcn wir diese Schwierigkeit, indem wit" zu gleicher Zeit nnd untcr ganz glcichen Bedingungen das ganze zu untersuchende Wasser in 3--4 Winklersche Gl/~ser verteilten. Die Analysen dieser Wasserportionen gaben ~uflerst iibereinstimmende Resultate, die als vollkommen getreue Wiedergabe des Gasbestandes der ganzen, im Respirationsgefa~e enthaltenen Wasscrmenge gelten konnten.
Versuchsdauer. Die Dauer der Versuche war yon der Salinit~t des angewandten Wassers
abh~ngig. Bei hSheren Salinit/~ten, wo die Balani ihre lebhaften rhythmischen Respirationsbewcgungen nicht unterbrachen und wo dcr O~-Verbrauch und die
1 Mac Clendon, Gault and Mulholland, Papers from Dep. Marine Biol. Carnegie Inst. II, 21 (1917).
JK. Buch, Soc. Scient. Fennica. Finl. hydrograph, biol. Unters. 191~, Nr 14. a Kreps, Intcrnat. Rev. d. ttydrobiol. 15, 240 (1926).

222 E. Kreps: Untersuchungen tiber den respiratorischen Gasweehsel
CO~-Abgabe bedeutende Werte zeigten, dauerte der Versuch 5 S~unden. Bei Versuehen mit stark versfil3tem Wasser, bei denen der O~-Verbraueh bedeutend langsamer vor sigh ging, dauerte der Versueh 8 Stunden.
DiG Versuehsdauer war so gew~thl~ worden, dab der 02-Verbraueh 15--20 % des Ausgangsvorrates an 02 nieht iibersehritt. Naeh dem Experiment blieben noch nieht weniger als 5--6 cem 02 pro 1 1 Wasser. Diejenigen geringen Steige- rungen der CO~-Tension, die der Zehrung yon 1/6--1/5 des vorhandenen 02 ent- spraehen, konnten an und fiir sieh auf die Intensit~t des Gaswechsels keine Wirkung haben. Auf Grund eines sehr reiehlichen experimentellen Ma.terials, sowohl seines eigenen als aueh desjenigen anderer Forscher, kommt Krogh 1 zu dem SchluB, dub dig Steigerung der CO~-Tension, die dem Verbrauch eines Drittels des vor- handenen O~ entspricht, gar keine Wirkung auf die Respirationstatigkeit der Seetiere ausfibt und der Gaswechsel vollkommen normal bleibt. Der Grund dafiir liegt 1. in dem sehr hohen Absorptionskoeffizienten der Kohlens~ure e und 2. darin, dal] der grSBte Teil der produzierten CO 2 dureh die Carbonate des Meer- wassers (Alkalinita~) gebunden wird.
l~iltrieren des Wassers.
Da das Meerwasser kein s~eriles Nilieu vorstellt, sondern immer eine be- deutende Quantit~t yon Phyto- und Zooplankton und Bakterien enthglL so finden im Wasser selbst wi~hrend des Stehens Atmungs- und Assimilationsprozesse start, die das Ergebnis des Versuehs beeinflussen k6nnten (Winterslein3). Das Fil~rieren des Wassers verringerte in bedeutendem Mal3e die darin stattfindenden chemisehen Prozesse, ohne sit jedoch endgfiltig zu beseitigen. Deswegen muBte das Wasser des Respirationsg]ases nicht mit dem Ausgangswasser, sondern mit dem Wasser aus den Kontrollgl~sern verglichen werden, dig wi~hrend des ganzen Versuehes ~ich in VerMltnissen befanden, welche mi~ denjenigen des Respirationsglases vollkommen iibereinstimmten. Das Niehtzusammenfallen des O~-Gehaltes und des TH in dem Ausgangs- und in dem Kontrollwasser war sehr gering und liel~ sieh etwa mit 1--2 Hundertstel eines Kubikzentimeters pro 1 1 bemessen.
IV. Die erste Per iode der Arbe i t m i t den beiden Ba lan ig ruppen war vor-
l~ufigen Orient ierungsversuchen geweiht. Die Balan i l eb ten in Aquar i en , in denen das Wasse r tggl ich dureh frisches erse tz t und a u g e r d e m 2 - - 3 maI a m Tage aer ie r t wurde. W~hrend dieser vorl~ufigen Per iode, die vom 3. V I I . - - 7 . V I I I . w/ihrte, wurde mi t beiden Gruppen eine Reihe yon Versuchen vorgenommen, mi t der 1. Gruppe meis t bei S = 330/00, m i t der 2. Gruppe bei S = 12~ 0. Die 2. Gruppe bl ieb im Laufe yon 20 Tagen unausgese tz t bei 12~ was der mi t t l e r en Sal ini t / i t des Oberf lgehenwassers im Meere zur gegebenen Jahresze i t en tsprach . Schwache Respi ra t ions- bewegungen zeigten sieh schon a m 3. 4. Tage nach Beginn des Aufent - ha l t s bei dieser Saliniti~t; vom 5. Tage an und wei ter gingen sie sehon in vo l lkommen normale r Weise vor sich. Angefangen mi t dem 10. Tage
1 Krogh, Respiratory Exchange of Animal and Man. 1916. 2 Nach Krogh ist fiir Meerwasser bei S = 350/00 der Absorptionskoeffizient
yon CO~=1,41 bei 0~ 1,17 bei 5~ 0,99 bei 10~ 0,85 bei 15 ~ a Winterstein, B!oehem." Z. 19, 427 (1909).

bei Balanus crenatus bei verschJedenem S~lzgehalt des Au$enmilieus. I. 223
und bis zum 20. warden mit dieser Gruppe 7 Respirationsversuehe ge- maeht bei S = 12~ (siehe welter Tab. 5).
Naeh den ersten Orientierungsversuchen wurde mit beiden Gruppen eine Reihe yon systematischen Experimenten nach einem bestimmten Plan durchgefiihr~. Diese Periode umfaBte die Zeit vom 7.--30. VII I . Die Versuche wurden t/~glieh gemacht. Bei der 1. Gruppe waren SS -----yon 0--14,5~ bei der 2. Gruppe yon 8--33o/00 .
Versuchsardnung. Ln der zu erforsehenden Salinit~t braehte die Gruppe 3 Tage der Reihe nach
zu, wobei jeden Morgen ein Respirationsversuch bei derselben Salinit~t gemaeht wurde, lqach 3 solehen Experimen~n blieben die Tiere bei derselben S noeh bis zum Morgen des n~chsten Tages, we sie unmittelbar in das Respirationsgefal~ gebraeht warden, welches sehon mit normalem Meerwasser bei 8 ~ 33~ ft an- gefiillt war. Nach diesem Versuch blieben die Tiere bei S~33~ Am n~ehsten Tage wurde wieder ein Respirationsversuch mit derselben S=33~ durch- geftihrt. Auf diese Weise wurdc das 1. Experiment bei S-~ 330/00 mit'Tieren vorgenommen, die eben aus einem versiil]ten Milieu genommen worden waren, mit dem sie sehon ins Gleichgewicht gekommen waren. Zur Zeit des 2. Versuches in normalem hleerwasser kamen die Balani mit dem neuen, mehr salzhaltigen Milieu ins Gleichgewicht. Sofort naeh diesem 2. Versuch bei S = 330/00 warden die Balani schon in das versfil~te Milieu gebrg~ht, dem die n~chsten 3 Versuehs- rage gewidmet wurden.
Aus dem Gesagten erhellt, dab nach je 3 Tagen Aufenthalt und Be- obachtung in einer etwaig verminderten Salinit~t 2 Tage folgten, we die Ver~uchsobjekte sich in normalem Meerwasser aufhielten. Diese Er- holungszeit in einem Milieu mit yeller Salinit~t hatte den Zweck, die Tiere wieder herzustellen und sie zu Anfang jeder neuen Triade in gleiche Verh/~ltnisse zu bringen.
Weiterhin ist zu ersehen, da$ vor Beginn eines jeden ersten Ver- suehs im versiii~ten Milieu, die Tiere sich schon 18--20 S~unden in diesem Milieu befunden hasten. Jeder erste Versuch bei S ~ 33~ dagegen wurde ohne jegliche Vorbereitung der Tiere angestellt, und erst zum zwei- ten Versuch bei dieser Salini~t konnte angenommen werden, dal~ die Balani mit dem neuen Milieu gewissermaSen ins Gleiehgewicht gekommen waren.
Es mul~ bemerkt werden, dal~ die Tiere/~ul~erlich sich sehr bald or- helen, naehdem man sie in die normale Salinit~t hiniiberbringt. Naeh einem etwa 4t~gigen A~ffenthalt in einem sehr versiiBten Milieu, w~hrend dessert die Tiere sich im Zustande des Seheintodes befanden, 5ffnen sie, nachdem sie in gewShnliches Meerwasser kommen, schon nach 5---8 Mi-
1 Ausnahmen wurden nur in wenigen einzelnen F/~llen mit einem speziellen Zweek gemacht, z.B. Experiment 27a mit der 2. Gruppe, we die Balani gleieh naeh dem 3. Versueh mit S=21Prom. in S = 33Prom, umgesetz$ Wurden, in der sie bis zum naehsten Tage blieben, an dem der Respirationsversuch mit 8 ~- 33Prom. gemaeht wurde.
224 E. Kreps: Untersuehungen fiber den respiratorisehen Gaswechsel
nuten die Klappen der Schalen (scutum und tergum), strecken ihre Ran- kenfiil~chen heraus und fangen an, mit denselben zu arbeiten, erst ]ang- sam, nach 20--30 Minuten aber schon beinahe normal. Eine Abweiehung vonder Norm wurde nieht yon seiten des Charakters der rhythmischen Bewegungen, sondern yon seiten des 02-Verbrauches beobachtet, welcher bei diesem Versueh in der Regel immer gesteigert war, worauf wir sparer noeh zurfickkommen werden. Als ,,Norm", nach der wir fiber den all- gemeinen Zustand der Tiere w~hrend der gegebenen Periode urteilen k6nnen, erscheinen die Resultate des 2. Experiments bei S----33~ Diese Standardversuche fielen naeh je 4 Tagen (3 Tage mit versii~tem Wasser und der 1. Tag mit S ~ 33~ auf den 5. Tag.
Wir beginnen mit den Standardversuehen. Die GrSBe des Gasweehsels w~hrend eines solehen Experiments kann bei Gleiehheit der fibrigen Be- dingungen als Kriterium des allgemeinen Zustandes der Tiere dienen.
Zu den Standardversuehen kSnnen fo]gende Experimente gerechnet werden (Tab. 1 und 2):
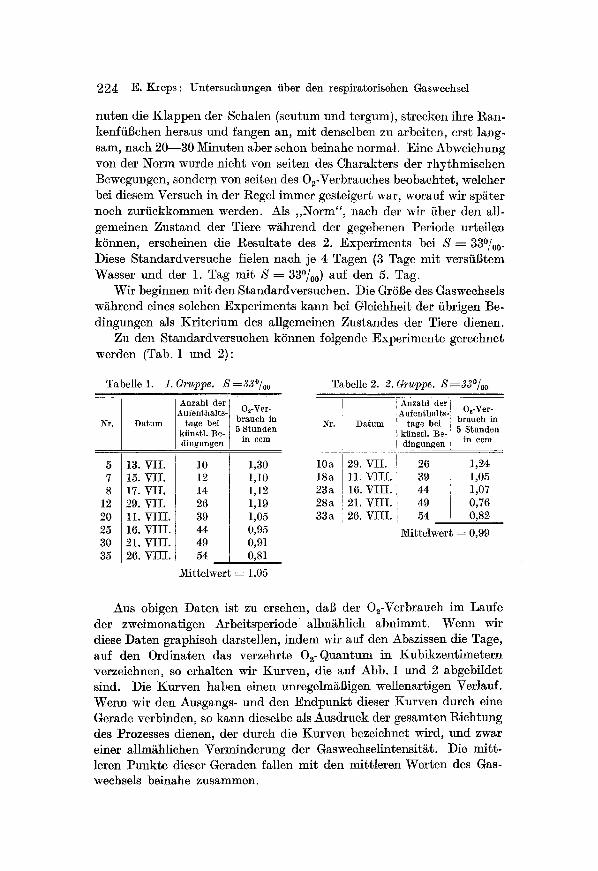
Tabelle 1. 1. Gruppe. S =33~
Anzahl der ~ufenthalts- O~-Ver-
Nr. Datum rage bei brauch in kfinstl. Be- 5 Stunden dingungen in ccm
5 7 8
12 20 25 30 35
13. VII. 15. VII. 17. VII. 29. VII. 11. VIII. 16. VIII. 21. VIII. 26. VIII.
10 1,30 12 1,10 14 1,12 26 1,19 39 1,05 44 0,95 49 0,91 54 0,81
Mittelwert : 1,05
Tabelle 2. 2. Gruppe. S~33~
O2-Ver- Nr. Datum brauch in
5 Stunden in ccm
10a 18a 23a 28a 33a
29. VII. 11. VIII. 16. VIII. 21. VIII. 26. VIII.
26 1,24 39 1,05 44 1,07 49 0,76 54 0,82
~4ittelwert = 0,99
Aus obigen Daten ist zu ersehen, daI~ der 02-Verbrauch im Laufe der zweimonatigen Arbeitsperiode ' allmiihlich abnimmt. Wenn wir diese Daten graphisch darstellen, indem wir auf den Abszissen die Tage, auf den Ordinaten das verzehrte O~-Quantum in Kubikzentimetern verzeichnen, so erhalten wir Kurven, die auf Abb. 1 und 2 abgebildet sind. Die Kurven haben einen unregelm~l~igen wellenartigen Verlauf. Wenn wir den Ausgangs- und den Endpunkt dieser Knrven dureh eine Gerade verbinden, so kann dieselbe als Ausdruck der gesamten Richtung des Prozesses dienen, der durch die Kurven bezeichnet wird, und zwar einer allm~hlichen Verminderung der Gaswechselintensit~t. Die mitt- ]eren Punkte dieser Geraden fallen mit den mittleren Werten des Gas- weehse]s beinahe zusammen.
bei Balanus crenatus bei verschiedenem Salzgehalt des Aul~enmilieus. I. 225
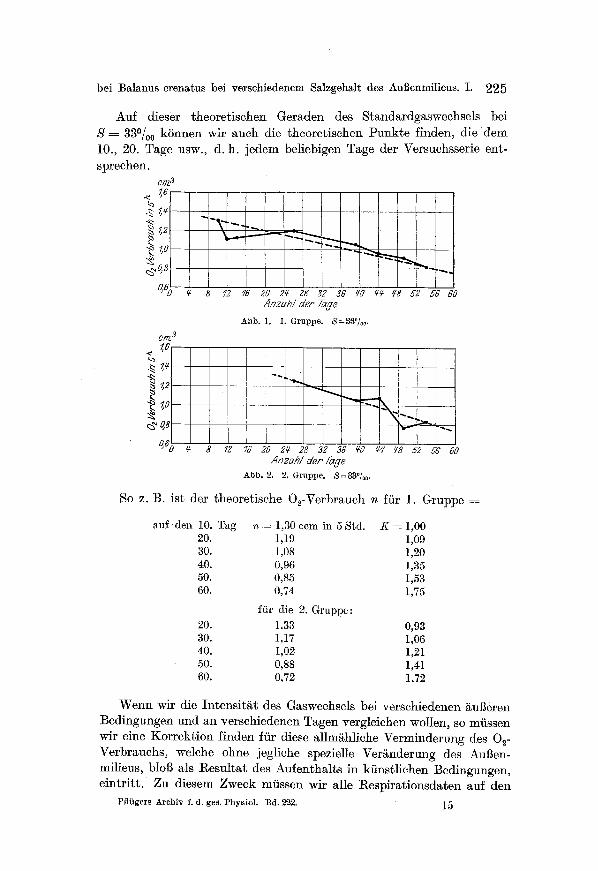
Auf dieser theoretisehen Geraden des Standardgaswechsels bei S 330/oo kSnnen wit aueh die theoretischen Punkte linden, die dem 10., 20. Tage usw., d. h jedem beliebigen Tage der Versuchsserie ent- sprechen.
C]7/3
"~2 , ; . . . , ~ ~
I 0,@ ~_ 8 12 16 2o 2r 23 32 3G go ~ ~3 52 5G 60
Anzahl der Ta#e
Abb. 1. i . Gruppe. ~=33~
crn 3 .e ~61
.~ 1,4
8 ~Z 1~ 2a 2g 28 32 36 fo ~ ~8 52 5G 50 A n z ~ h / t i e r Z~#e
Abb. 2. 2. Gruppe. S=33~
So z. B. ist der theoretische O~-Verbraueh n ffir 1. Gruppe =
auf .den 10. Tag n ~ 1,30 ccm in 5 Std. 20. 1,19 30. 1,08 40. 0,96 50. 0,85 60. 0,74
~tir die 2. Gruppe: 20. 1,33 30. 1,17 40. 1,02 50. 0,88 60. 0,72
K ~ 1,00 1,09 ],20 1,35 1,53 1,75
0,93 1,06 1,21 1,41 ] .72
Wenn wir die Intensit~t des Gaswechsels bei verschiedenen ~ul~eren Bedingungen und an versehiedenen Tagen vergleiehen wollen, so mfissen wir eine Korrektion linden ffir diese allm~hliche Verminderung des 0 2- Verbrauehs, welehe ohne jegliehe spezielle Ver~ndermlg des Au•en- milieus, bloS a]s Resultat des Aufenthalts in kiinstliehen Bedingungen, eintritt. Zu diesem Zweck mfissen wir alle Respirationsdaten auf den
Pfliigers k reh iv f. d. ges. Physiol . Bd. 222. 15
226 E. Kreps: Untersuchungen tiber den respiratorisohen Gaswechsel
Zustand der Tiere zurfiekffihren, in dem sie sich an irgendeinem beliebig gew/ihlten Tage befanden, z. B. am Tage des ersten Standard-Respira- tionversuches (Mr wghlen den ersten Tag, da die LebenstiLtigkeit der Tiere damals der Norm ngher stand als in den n/~chsten Tagen). Ffir die 1. Gruppe ist es der Tag, we der 02-Verbraueh = 1,3 ecru wi~hrend 5 Stunden, ffir die 2. Gruppe 1,24 eem w/ihrend 5 Stunden war, Um die Bereehnung ffir diesen Tag zu maehen, mfissen wir bloB das Resultat eiues jeden Experiments mit dem entsprechenden Quotienten K multi-
plizieren, der ffir die 1. Gruppe -- 1,3 , ffir die 2. Gruppe = 1,24 ist. n q$
Mit ttilfe dieses Quotienten kann man bis zu einem gewissen Grade den Einflul~ des Aufenthalts in ungfinstigen kiinstlichen Bedingungen, der
~5
,
o,2
/ /
J / !
r 0 $ ~/ 6 8 10 52 5g Y6 58
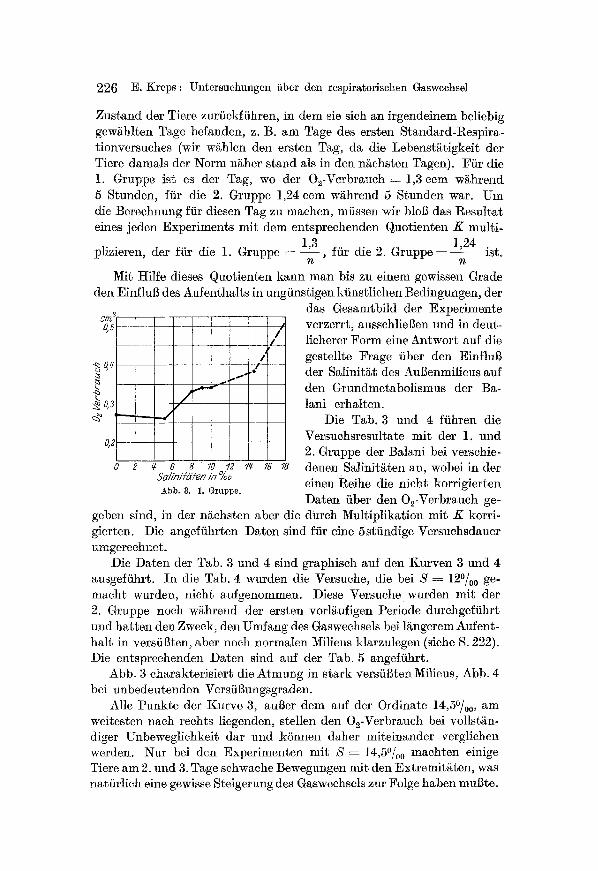
&d/n//h'/en 1)7 %o Abb. 8. 1. Gruppe.
das Gesamtbild der Experimente verzerrt, aussehliegen tmd in deut- lieherer Form eine Antwort auf die gestellte Frage fiber den Einflug der Salinitgt des Augenmilieus auf den Grundmetabolismus der Ba- lani erhalten.
Die Tab. 3 und 4 ffihren die Versuehsresultate mit der 1. und 2. Gruppe der Balani bei versehie- denen Salinit/Lten an, wobei in der einen Reihe die nieht korrigierten Daten fiber den 02-Verbraueh ge-
geben sind, in der n/iehsten aber die dutch Multiplikation mit K korri- gierten. Die angeffihrten Daten sind fiir eine 5stiindige Versuehsdauer umgereehnet.
Die Daten der Tab. 3 und 4 sind graphiseh auf den Kurven 3 und 4 ausgeffihrt. In die Tab. 4 wurden die Versuche, die bei S = 12~ o ge- maeht wurden, nieht aufgenommen. Diese Versuche wurden mit der 2. Gruppe noch w~hrend der ersten vorl/~ufigen Periode durehgeffihrt und hatten den Zweek, den Umfang des Gaswechsels bei l~ngerem Aufent- halt in versfigten, aber noch normalen Milieus klarzulegen (siehe S. 222). Die entsprechenden Daten sind auf der Tab. 5 angefiihrt.
Abb. 3 charakterisiert die Atmung in stark versiigten Milieus, Abb. 4 bei unbedeutenden Versfigungsgraden.
A]le Punkte der Kurve 3, auger dem auf der Ordinate 14,5~ am weitesten nach reehts liegenden, stellen den 02-Verbrauch bei vollst/in- diger Unbeweglichkeit dar und kSnnen daher miteinander vergliehen werden. Nut bei den Experimenten mit S = 14,5~ machten einige Tiere am 2. und 3. Tage sehwache Bewegungen mit den Extremiti~ten, was natfirlich eine gewisse Steigerung des Gasweehsels zur Folge haben mugte.
bei BalanUs crenatus bei verschiedenem Salzgehalt des AuBenmilieus. I. 227
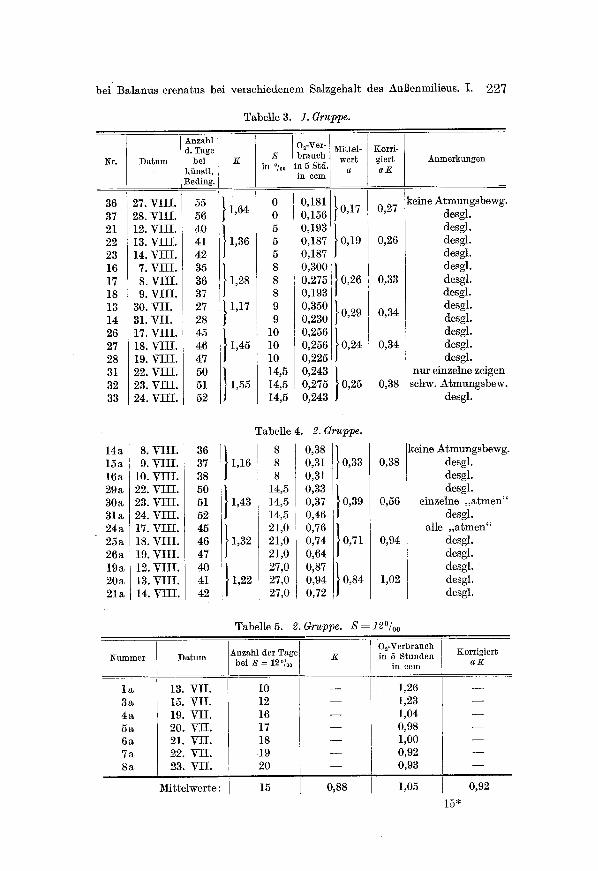
Tabelle 3. 1. Grus
Nr.
36 37 21 22 23 16 17 18 13 14 26 27 28 31 32 33
14a 15a 16a 29a 30a 31a 24a 25a 26a 19a 20a 21a
Anzahl d. Tage
D a t u m bei ki inst l . Beding.
27. VIII. 55 28. VIII. 56 12. VIII. 40 13. VIII. 41 14. VIII. 42 7. VIII. 35 8. VIII. 36 9. VIII. 37
30. VII. 27 31. VII. 28 17. VIII. 45 18. VIII. 46 19. VIII. 47 22. VIII. 50 23. VIII. 51 24. VIII. 52
8. VIII. 36 9. VIII. 37
10. VIII. 38 22. VIII. 50 23. VIII. 51 24. VIII. 52 17. VIII. 45 18. VIII. 46 19. VIII. 47 12. VIII. 40 13. VIII. 41 14. VIII. 42
K
1,28
1,17
1,45
1,55
S in ~
0 0 5 5 5 8 8 8 9 9
10 10 I0 14,5 14,5 14,5
Oz-Ver- brauch
in 5 Std. in ccm
0,181 0,156 0,193 0,187 0,187 0,300 0,275 0,193 0,350 0,230 0,256 0,256 0,225 0,243 0,275 0,243
~Iittel- wert
a
0,17
0,19
0,26
0,29
0,24
0,25
Korr i - g ie r t a K
0,27
0,26
Tabelle 4. 2. Gruppe.
8 1,16 8
8 14,5
1,43 14,5 14,5 21,0
1,32 21,0 21,0 27,0
1,22 27,0 27,0
9,38 } 9,31 0,33 0,31 0,33 } 0,37 0,39 0,46 0,76 } 0,74 0,71 0,64 0,87 } 0,94 0,84 0,72
0,33
0,34
0,34
0,38
A n m e r k u n g e n
keine Atmungsbewg. desgl. desgl. desgl. desgl. desgl. desgl. desgl. desgl. desgl. desgl. desgl. desgl.
nur einzelne zeigen schw. Atmungsbew.
desgl.
keine Atmungsbewg. 0,38 desgl.
desgl.
0,56
0,94
1,02
Tabelle 5. 2. Gruppe. S = 12~
desgl. einzelne ,,atmen"
desgl. alle ,,atmen"
desgl. desgl. desgl. desgl. desgl.
N u m m e r
3a 4a 5a 6a 7a 8a
D a t u m
13. VII. 15. VII. 19. VII. 20. VII. 21. VII. 22. VII. 23. VII.
Mittelwerte:
Anzahl der Tage bei S = 12"/oo
10 12 16 17 18 19 2O
15
K
0,88
O~-Verbrauch in 5 S tunden
in ccm
1,26 1,23 1,04 0,98 1,00 0,92 0,93
1,05
Korr ig ie r t a K
0,92
15"

228 E. Kreps: Untersuchungen fiber den respiratorisehen Gaswechsel
Bei S ---- O~ d. h. in siiBem Wasser und bis 5o/00 erwiesen sieh die Werte der 02-Zehrung minimal und naeh Korrektion praktisch gleieh groB. Bei den S S - 8~ 90/00 und 10~ ist beinahe kein Unterschied zwischen den Werten des O~-Verbrauchs zu merken, wobei diese Werte diejenigen bei den S~linit~ten 5o/00 und 0~ urn 25% /ibersteigen.
Bei S ~ 14,5~ o, wo sehon eine sehwache motorisehe T~tigkeit ein- setzt, und der dureh dieselbe bedingte Metabolismus sich zu dem ur- spriinglichen Standardmetabolismus addiert, beobaehten wir noeh eine Steigerung des 02-Verbr~uch (ungef~hr urn 10~
Der Absehnitt der Kurve, der auf das Gebiet vSttiger Unbeweglieh- keit f~llt, erh~]~ auf diese Weise einen Wendepunkt zwischen 50/oo und 80/00 . Diese Salinit~ten entspreehen gerade den kritisehen Konzentr~- tionen, die ffir die litoralen Ba]ani yon uns festgestellt wurden (5~ f/h" B. erenatus und 6--70/00 ffir B. balanoides). Das sind diejenigen Schwel- lenkonzentrationen des AuBenmilieus, deren Herabsetzung die Balani in einen Zustand yon Anabiose bringt, indem sie so ]ange verbleiben, bis die Salzkonzentration des AuBenmilieus in genfigendem Mal]e gestiegen ist.
Es erweist sich also, dab in dlesem Gebiet geringen Salzgehaltes (0--10~ der Stand~rdmetabolismus selbst, der mit keiner siehtbaren /iuBeren T~tigkeit verbunden ist, yon der Salzkonzentration des um- gebenden Milieus abhiingig ist. Diese Abh/~ngigkeit erscheint nicht als eine ununterbrochene regelm/iBige Funktion, und die Kurve, welche diese Abh~tngigkeit darstellt, maeht art einer bestimmten Stelle einen ziemlich scharfen Knick.
In der Abb. 4 entspreehen 4 experimentell festgestellte Punkte ( S S ~- 12~ 21~ 270/00 und 330/0o) den Versuchen, in denen die Tiere vollkommen normal ihre Atembewegungen machten 2. Bei den Versuchen mit S = 14,5~ waren die Bewegungen schwaeh; die Tiere bat ten nur 3 Tage bei dieser Salinit~t zugebraeht und sich an d~s versfiBte Milieu nicht gew6hnt. In den Versuehen mit S = 80/o0 waren iiberhaupt keine Bewegungen zu merken.
Die Kurve zeigt, dab dort, wo die respiratorischen Bewegungen frei vor sich gehen, der G~swechsel viel h6here Werte gibt als beim Zustand der Unbewegliehkeit der Tiere. Wir k6nnen in der Kurve aueh bei ffeien Atmungsbewegungen einen bestimmben Zusammenh~ng zwischen der Salinit~t des AuBenmflieus und dem verbrauehten Sauerstoff bemerken: je h6her die S, desto starker die O2-Zehrung. Die Bedeutung des Salz-
1 Bei der 2. Gruppe in der Kurve 4 wurde bei derselben S = 14,5%o eine intensivere Bewegung der FfiBchen und infolgedessen ein grSBerer Oz-Verbraueh beobachtet.
2 Die lebhafte motorisehe T/~tigkeit bei der geringen S : i2~ 1/~l]t sieh, wie schon erw/~hnt wurde, dadureh ertd~ren, dal] die Tiere schon viele Tage (mehr als 10) sieh in diesem S~lzgehalt befanden und Zeit gehab~ hatten, sieh an den- selben zu ,,gew6hnen".
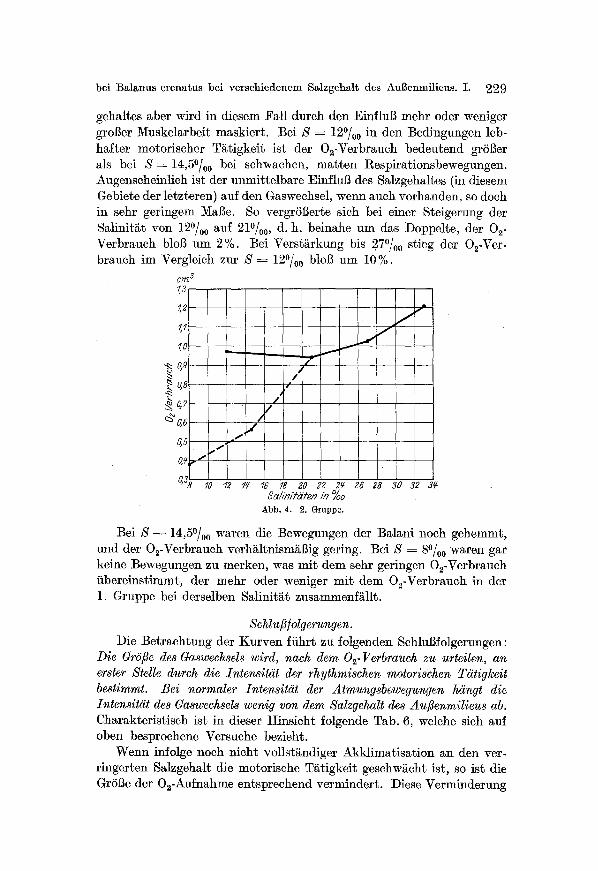
bei Bal~nus crenatus bei verschiedenem Salzgehalt d~s AuBenmilieus. I. 229
gehaltes aber wird in diesem Fall durch den Einflu]3 mehr oder weniger groSer Muskelarbeit maskiert. Bei S = 12~ in den Bedingungen leb- hafter motorischer T~tigkeit ist der 02-Verbrauch bedeutend grSl3er als bei S ~ 14,5~ bei schwachen, marten Respirationsbewegungen. Augenscheinlich ist der unmittelbare EinfluB des Salzgehaltes (in diesem Gebiete der letzteren) auf den Gaswechsel, wenn auch vorhanden, so doch in sehr geringem MaBe. So vergrSBerte sich bei einer Steigerung der Salinit~t yon 12~ 0 auf 21O/oo, d. h. beinahe um das Doppelte, der 02- Verbrauch bloB um 2%. Bei Verst~rkung bis 270/oo stieg der 02-Vcr- brauch im Vergleich zur S = 12~ bloB um 10%.
CT/Z 3 7,3
J 7j f i
~ i I
~ o,6 , / ' 0,5 ! - - . . f ' 0,r ~ f
0~3~ 7o ~z I~ r 78 30 z2 2~ 2~ 28 30 32 3~ Salinitd~en /n %o
Abb. 4. 2. Grupl~e.
Bei S - 14,5O/oo waren die Bewegungen der Balani noch gehemmt, und der O2-Verbrauch verh~ltnism~$ig gering. Bei S = 80/00 waren gar keine Bewegungen zu merken, was mit dem sehr geringen 02-Verbrauch iibereinstimmt, der mehr oder weniger mit dem O2-Verbrauch in der 1. Gruppe bei derselben Salinit~t zusammenf~llt.
Schlufi /olgerungen.
Die Betrachtung der Kurven fiihrt zu folgenden Schlul3folgerungen: Die GrS/3e des Gaswechsels wird, nach dem 0~-Verbrauch zu urteilen, an erster Stelle dutch die Intensit~it der rhythmischen motorischen Ti~tigkeit beztimmt. Bei normaler Intensitii t der Atmungsbewegungen Mingt die Intensitii t des Gaswechsels wenig yon dem Salzgehalt des Auf lenmil ieus ab. Charakteristisch ist in dieser Hinsicht folgende Tab. 6, welche sich auf oben besprochene Versuche bezieht.
Wenn infolge noch nicht vollst~ndiger Akklimatisation an den ver- ringerten Salzgehalt die motorische T~tigkeit geschw~cht ist, so ist die GrSI~e der O2-Aufnahme entsprechend vermindert. Diese Verminderung

230 E. Kreps: Untersuchungen fiber den respiratorischen Gaswechsel
Tabelle 6. 2. Grulgpe.
]gummer Datum
24a 17. VIII. 25a 18. VIII. 26a 19. VIII. 27a 20. VIII. 28a 21. VIII.
Anzahl der Tage bei
kfinstl. Be- dingungen
45. 46. 47. 48. 49.
So in 100
21 21 21 33 33
O~-Ver- brauch in
5 Std.(ohne Korrektur)
0,76 0,74 0,64 0,70 0,76
Anmerkungen
LebhafteAtmungsbeweg. desgl. desgl. desgl. desgl.
kann einerseits als das Resultat herabgesetzten 02-Verbrauchs infolge der gesehw/~ehten funktionellen Aktivitiit des Muskelapparats ange- sehen werden, andererseits abet als Resultat verminderter Zufuhr yon 0 3 infolge der Schw/~chung und Verlangsamung der Atembewegungen.
Wenn auch eine Abh/ingigkeit zwischen der Salzkonzentration im um- gebenden Milieu und dem Standardmetabolismus im Sinne yon Krogh vorhanden ist, so wird bei einer solchen Versuchsanordnung in diesem Salinit/~tsgebiet diese Abh/~ngigkeit dureh alas Hinzukommen der Muskel- arbeit vollkommen verdunkelt.
Die Beziehungen kl/~ren sich in den Bedingungen vSlliger Abwesenheit yon Muskelarbeit bei stark verminderten Salinit~ten. Erstens ist in diesem Fall der Gesamtverbrauch yon Oz bedeutend geringer, was vor allem wahrscheinlich als direktes Resultat der Abwesenheit yon Muskelarbeit anzusehen ist. I m Durchschnitt gerechnet ist hier der O~-Verbraueh im Vergleieh zur normalen aktiven Existenz der Balani 3- -4rea l geringer. Zweitens hs der O~-Verbrauch in stark versiiBten Milieus bei voll- kommener Unbeweglichkeit der Tiere yon dem Salzgehalte des A u/3enmilieus ab. Diese Abh/~ngigkeit ist der Art, dab der Gasweehsel, wenn der Salz- gehalt 6%0 nieht fibersteigt, sieh auf ein bestimmtes Minimalniveau stellt, das bei weiterer Verminderung der Salinit/it nicht mehr, oder bei- nahe nicht mehr, herabgesetzt wird. Dieses Niveau des Gaswechsels ist sowohl ffir Sfigwasser als ffir S = 50/00 ein und dasselbe. In diesem Sali- nit/~tsgebiet finder kein , ,Auf leben", kein Ubergehen zum aktiven Leben bei den Balani start ; dieselben verfallen in den Zustand des,, Salzsehlafes".
Wenn der Salzgehalt hSher ist als diese ,,kritische Konzentration" - - gleich 8--10~ o - - , so i~bersteigt der Gaswechsel um 25 30% das Min i - mum. Er besteht a]s soleher, solange die Tiere gar keine Bewegungen machen. Wir wissen aber, dab dieses gerade diejenigen Salinitgten sind, bei denen die Balani mit der Zeit mi t ihren rhythmischen Bewegungen beginnen. Gleichzeitig mit der Rfickkehr der motorischen T/~tigkeit wird aueh der Umfang des Gaswechsels zunehmen, bis er sein Maximum erreicht, wenn die Bewegungen sieh zu ihrer gr6gten Intensit/~t ent- wickeln.

bei Balanus crenatus bei verschiedenem Salzgehalt des Aul~enmilieus. I. 231
Diese Daten geben der ,,kritisehen Konzentrat ion" eine neue Deft- nition, auBer der schon auf Seite 216 erwghnten. Die l~ritische Konzen- tration ist diejenige Konzentration des hypotonischen Aufienmilieu, bei der die Kurve des respiratorischen Gaswechsels bei volllcommen ruhenden Tieren einen Umschwung nach unten zeigt, bei der der Ubergang yon einem Gaswechselniveau zu einem andern, minimalen, stattfindet, der dem Zustande des Salzsehlafes entsprieht.
V .
Es erhebt sich nun die Frage, ob bei regungslosen Tieren bei sehr herabgesetztem Gasweehsel in stark versfiBten Milieus ein v61liges Uber- einstimmen zwisehen dem Verbrauch yon 03 und dem Bediirfnis nach ibm sich einstellt ? Ob die einzelnen Phasen des Stoffwechsels mifeinander ins Gleichgewicht kommen ? Zur Beanfwortung dieser Frage verfiigen wir fiber folgende D~ten:
Wenn wit die Balani nach 3---4tiigigem Au/enthalt bei vermindertem Salzgehalt und im Zustande der Regungslosiglceit direlct in das Respirations- glas bringen und einen Versuch mit S = 33 % anordnen, so erhalten wit einen gegen die Norm bedeutend vergr6[3erten Wert /i~r den 03- Verbrauch. Am folgenden Tage, nach einem 24stfindigen Aufenthalt der Tiere in normaler Salinitgt, f/~llt der O2-Verbrauch bis zur normalen GrSBe. Dieses Faktum offenbarte sieh unabanderlich, regelm~Big nach jeder Versuchs- triade mit verdiinntem Wasser. Zur Illustration fiihren wir Tab. 7 an.
Tabelle 7.
5!ach 3 t ig igem Aufen~halt bei
S = ~
Durchschnit ts- verbrauch -zon O~
in dem angegebenen Salzgehalt (mit
Korrektur) in ecru
I O~-Verbrauch O:-Verbrauch bei I
S=33.~/o0 so,of t nach[ bei S = 33~ dem Ubersetzen ausl am folgenden d. versiiBten Milieu / Tage (ohne (ohne Korrektur) . Korrektur)
in ecru I in ecru I
Steigerung des 02-Verbrauchs am 1. Tage im Vergleich zum
2. Tage in %
5,0 8,0
10,0 14,5
0,26 0,33 0,34 0,38
1. Gruppe: 1,62 1,34 1,13 1,16
0,95 1,05 0,91 0,81
70 28 24 43
2. Gruppe: s,o ! 0,38 1,85 ] 1,05 I 76
14,5 f 0,56 1,17 i 0,82 I ~3
Beim Hiniiberbringen in S = 33~ aus einer Umgebung, wo die Atmungst~tigkeit normal vonstatten ging, wird eine solehe Steigerung des Oz-Verbrauchs nicht beobaehtet (Tab. 8).

232 E. Kreps: Untersuchungen tiber den respiratorischen Gaswechsel
Tabelle 8. 2. Gruppe.
Durchschnitts- 02-Verbrauch bei 3 ~0- f . ~ -I O~-Verbrauch bei Differenz verbraueh r O l l O~ ~'~ ~ /~ SO or~ acn ~ Naoh 3 Tagen bei der gegebenen S dem Ubersetzen 8=~]0~ am z. Tage zwischen dem
bei S=~/~ (mit Korrek~ur) (ohne Korrektur) (ohne Korrektur) 1. und 2. Tage in ecm in eem m ecru in %
27 21
1,02 0,94
0,95 ] 1,07 0,70 I 0,76
- - I1 - - 8
Die ~ngofiihrten FakCa sprechon ~ugenscheinlioh daffir, dab wahrend des Aufenthalts in oinem entsalzten Milieu, in Verhal~nissen eines herab- gesot, zten O~-Vorbrauohs, im Organismus der B~lani eine Anhs gowisser Stoffweohselprodukte stattfindet, die einer Oxydation untor- worfen werden muBton, ~ber nicht oxydiert geblieben sind. Diese ,,Sauer- stoffschuld" verursaoht oinen gesteigerten O~-Verbraueh, sobald d~s Tier in Verh/~l~nisse zurfiokkehrt, in denen es soinen Geweben das Zukommen eines genfigenden 0 3- Quan~ums gestat ten kann.
Analoge T~tsaehen wurden bei einer Reiho yon Tieron beobaohtet, besonders bei Wirbellosen, die befghigt sind, mehr oder minder lange Zoib in anaeroben Verh~ltnissen zu existieren. So fand Lesser 1, dab beim Regenwurm (Lumbrieus terrestris), der lange ohne Sauerstoff leben kann, sioh im Laufe einer solohen Periode versohiedene Fetts~uren bilden, die bei normaler, oxydativer Spaltung der Nghrstoffe sich nicht offenbaren. Wenn wir naeh einer anoxybiotischen Poriode dem Wurme S~uerstoff geben, so beobachton wit eine abnorm grebe Zehrung desselben mit einem sehr niedrigen respiratorisehen Quotienton. Dabei warden die Produk~e des unvollsbandigen Zorfalls schnoU oxydiert.
Es liegt nahe, auch einigo Analogien mit dem Wintorsehlaf der Sguge- tiere durchzuf/ihren. So meint Naga i 2, dab wghrend des Wintersehlafs im Organismus der S~ugetiere abnorme, metabolische Prozesse start- linden und, speziell, eine unvollst/~ndige Oxydation. W~hrond des Sohla- fes werden sehr niodrigo respiratorisohe Quotiente beobaohtet; im H a m erschoinb viol Milchsgure, der Harnstoffgeh~l~ wird geringer, derjonige der Aminos~uren grSBor.
Der ProzeB des Erw~ehens bUS dam Wintersehlaf wird yon einem enormen Steigen dos Gasweehsels begleibet (Dubois s, Pembrey 4, Wein- land und Riehl 5, Henriques 6 u. a.). Die Ursaehe dieser Gaswoehselstei- gerung ist noah nicht ganz klar. Zum Teil kann sie dureh erh6hte
1 Lesser, Z. Biol. 52, 282 (1909); 53, 532 (1910) and 54, 1 (1910). Nagai, Z. f. allg. Physiol. 9, 243 (1909).
3 Dubois, J. de Physiol. 1899, 1020. 4 Pembrey, J. of Physiol. 27, 66 (1901) und 29, 195 (1903). 5 Weinland und Riehl, Z. Biol. 49, 37 (1907) und ~0, 75 (1908). 6 Henriques, Skand. Arch. Physiol. 25, 15 (1911).

bei Balanus crenatus bei verschiedenem Salzgehalt des Aul3enmilieus. I. 233
Muskelt~tigkeit, durch s~arkes Sichschiitteln und andere Bewegungen erkl~rt werden.
Das in unseren Experimenten beobachtete Steigen des O~-Verbrauchs fand immer nach 3---4 Tagen des ,,Salzschlafes" start. Es ist uns nicht bekannt, ob dieser Zustand yon ,,Sauerstoffschuld" bei l~ngerem Aufent- half in sehr geringen S~]init~ten weiterbesfeht, wir nehmen uns ,,/or, bei weiterer Arbeit diese Frage zu erlautern.
Zusammenfassung. 1. Der respiratorische Gaswechsel und im speziellen der O~-Verbrauch
bei Balanus crenatus wird in erster Linie dutch die Intensitat der rhyth- mischen Atmungst~tigkeit bestimmt. Bei normaler Intensitaf der letz- teren, die bei Salinitaten yon 12--35O/oo stattfindet, ist die GrSl3e des Gas- wechsels praktisch yon dem SMzgehalf des Aul3enmilieus unabh~ngig.
2. In stark verminderten Salinit~ten unter 10~ in den Bedingungen einer vSlligen Abwesenheit yon Muskelarbeit, ist der gesamte Sauer- stoffverbrauch bedeutend niedriger, was wahrscheinlich als direkte Folge der Muskelruhe zu erklaren ist. Im Durchschnitt isf in diesem Fat1 der O~-Verbrauch im Vergleich zur normalen aktiven Existenz der Balani 3--4 real geringer.
3. In stark versfil~tem Milieu (0--10~ h~ngt der O~-Verbrauch bei vSlliger Bewegungslosigkeit yon dem Salzgeha]t des Aul]enmilieus ab. Wenn die Salinit~t unter 6~ ist, so stellt sich der Gaswechsel auf dem niedrigsten, dem minimalen Niveau ein, das sowohl fiir Siil~wasser, als fiir S = 6O/0o gleich ist. Ubersteigt die Salinitgt diese ,,kritische Kon- zentration" yon 6o/0o und kommt sie 7--8--90/00 gleich, so~ist der Gas- wechsel 25--30% hSher als der minimale.
4. Solange die Balani in einem Milieu sind, dessen Salinit~t die kri- tische Konzen~ration = 6~ nicht iibersteigt, befinden sie sich im Zu- stande des Salzschlafes, aus dem sie beim Hinfiberkommen in hShere Salinitgten erwachen.
5. Beim Hiniiberkommen aus einem stark versiiBten Milieu in ein salzhaltigeres ist der 02-Verbrauch sofort nach dem Ubergang vom Salz- schlaf zum aktiven Zustand bedeutend h6her als der normale. Am zwei- ten Tage f~llt der 02-Verbrauch bis zur Norm.


![Untersuchungen zur herzfrequenzvariabilität bei herzkranken kindern und jugendlichen [electronic resource]](https://img.pdfslide.net/doc/110x75/63572da6dffe30f4b50c78e7/untersuchungen-zur-herzfrequenzvariabilitaet-bei-herzkranken-kindern-und-jugendlichen.jpg)













![Pelagius in Irland [microform] : texte und untersuchungen zur](https://img.pdfslide.net/doc/110x75/631feb7c9353b08ff5016b51/pelagius-in-irland-microform-texte-und-untersuchungen-zur-.jpg)












