Embed Size (px)
Citation preview

1
Lehninger Principles of BiochemistryFourth Edition
Chapter 19:Oxidative Phosphorylationand Photophosphorylation
Copyright © 2004 by W. H. Freeman & Company
David L. Nelson and Michael M. Cox

2
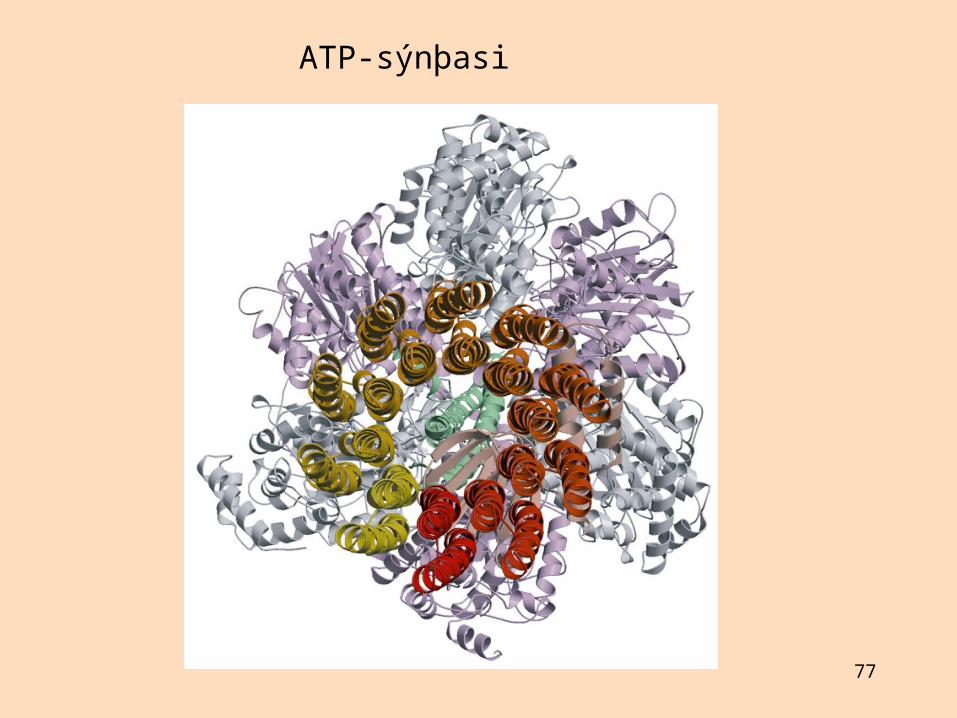
F1-eining ATP-sýnþasa ADP: gult, ATP: rautt

3

4
Hvað er ildisháð fosfórýlering (oxidative phosphorylation)?
Rafeindir eru fluttar frá afoxuðum kóensímum, NADH og FADH2, eftir röð rafeindaberaSúrefni (“ildi”) er notað til oxunarinnar, og það afoxast í vatn, en kóensímin enduroxast
Um leið verður ATP-myndun frá ADP og Pi (fosfórýlering)Fosfórýleringin gerist ekki nema súrefni sé tils staðar, er ildisháð

5
Hvað er ildisháð fosfórýlering (oxidative phosphorylation)?
Fulloxun fæðuefna gefur margfalt meira ATP eða orku en loftfirrt efnaskipti (anaerobic)Fita gefur 9 kcal/g, kolhýdröt 4 kcal/g, prótein 4 kcal/g, alkóhól 7 kcal/g
Oxun NADH ~ 2,5 ATPOxun FADH2 ~ 1,5 ATP

6
Hvar gerist ildisháð fosfórýlering?
Ildisháð fosfórýlering gerist í innri himnu mítókondríaÍ einni lifrarfrumu eru allt að 2000 mítókondríur, algengt að um 100 mítókondríur séu í frumuUm helmingur massa hjartavöðva, sem reiðir sig á loftháð efnaskipti, er mítókondríurMítókondríur eru einnig kallaðar “hvatberar”, “kyndikorn” eða “orkukorn”Sítrónusýruhringur og oxun fitusýra, sem leggja til afoxuð kóensím, gerast í mergholi (matrix) mítókondría
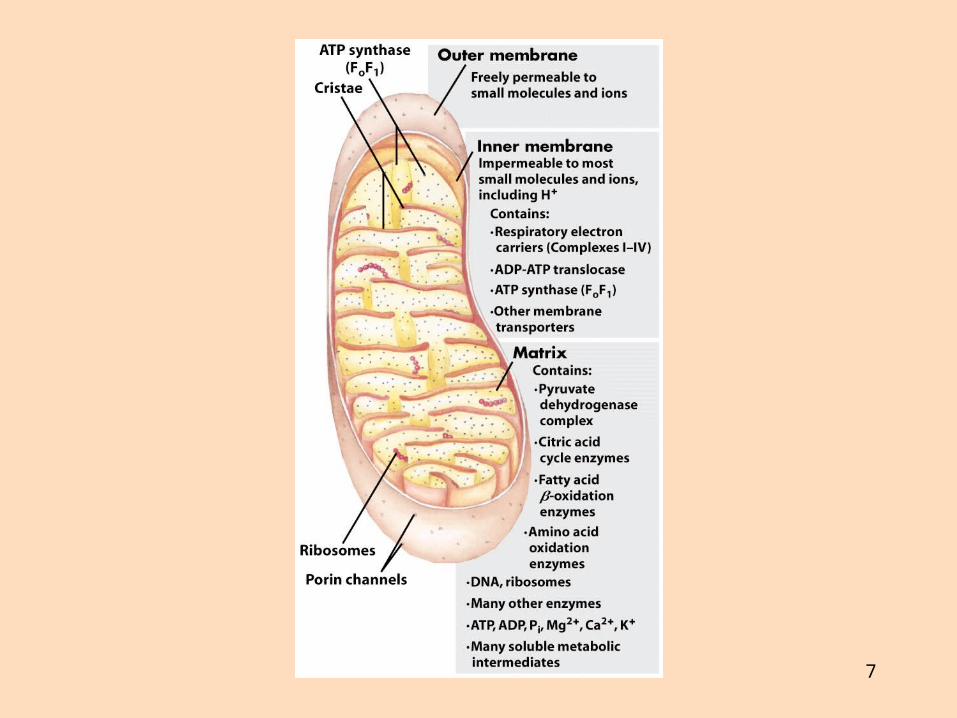
7

8
Rafeindaberar og ildisháð fosfórýlering
Rafeindaberar (að mestu sýtókróm) mynda keðju sem er í innri himnu mítókondríaÞessi keðja er kölluð rafeindaberakeðja eða öndunarkeðja
Við rafeindaflutning (oxun kóensíma á kostnað súrefnis) losnar frjáls orka sem er notuð til að dæla prótónum út úr mergholi mítókondría gegn styrkleikastigli

9
Rafeindaberar og ildisháð fosfórýlering
Við það myndast prótónustigull og himnuspennaATP myndast frá ADP og Pi (fosfórýlering) og það er losað frá ensíminu ATP-sýnþasa þegar prótónur flæða aftur inn í merghol undan stigli
Oxun og fosfórýlering eru tengd með prótónustigli

10
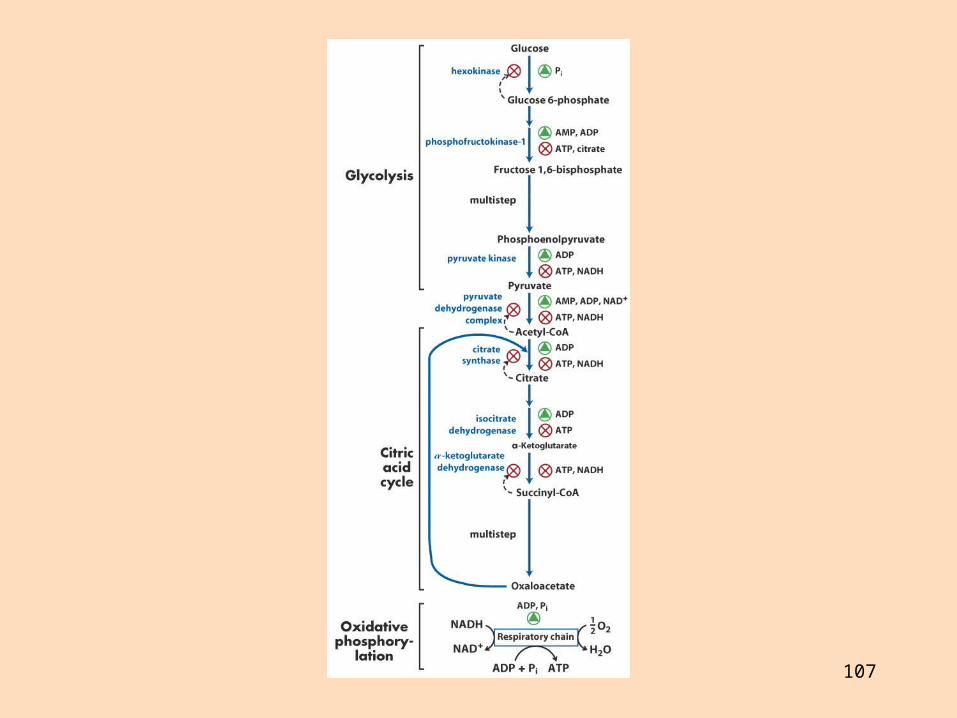
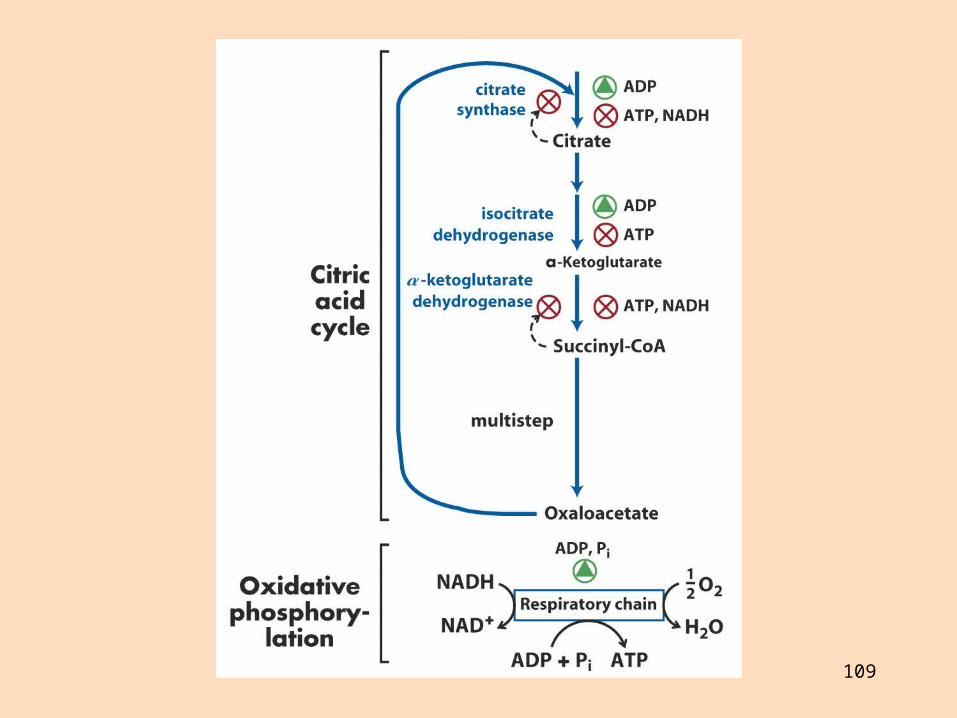
Rafeindaberar og ildisháð fosfórýlering Gerð rafeindaberakeðjunnar
Rafeindaberakeðjan er gerð úr fjórum rafeindaberandi ensímakomplexum (prótónudælum) sem eru tengdir saman með tveimur rafeindaberum sem fljóta í innri himnunni
Ensímakomplexarnir eru gerðir úr mörgum próteinum Sum próteinin innihalda járn og brennistein sem taka þátt í rafeindaflutningnum
Þau breyta um lögun í oxunar- og afoxunarhvörfum
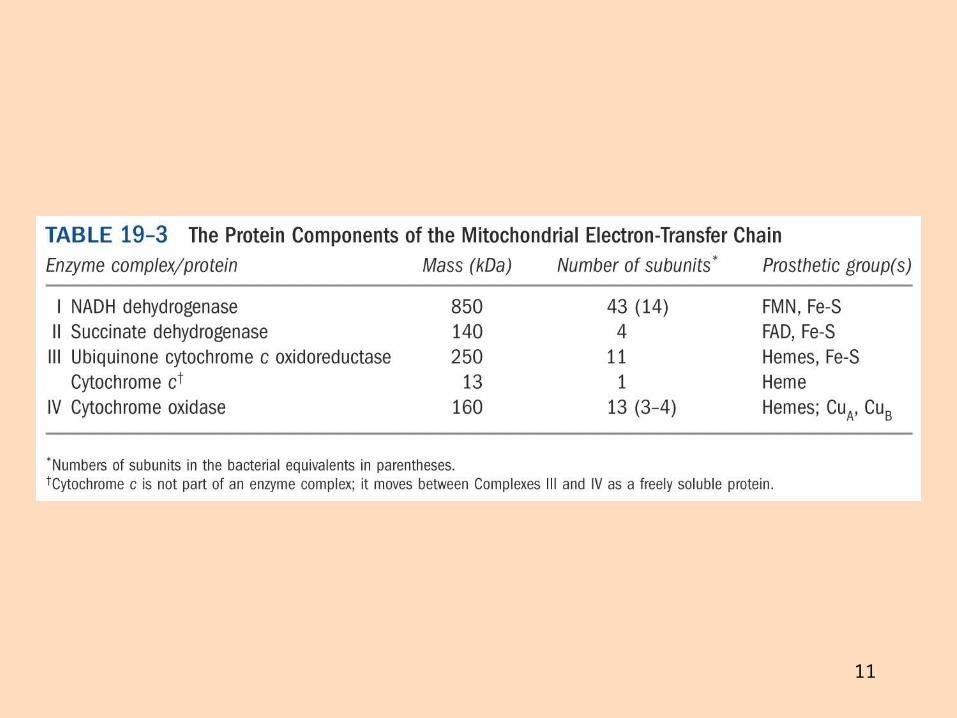
11
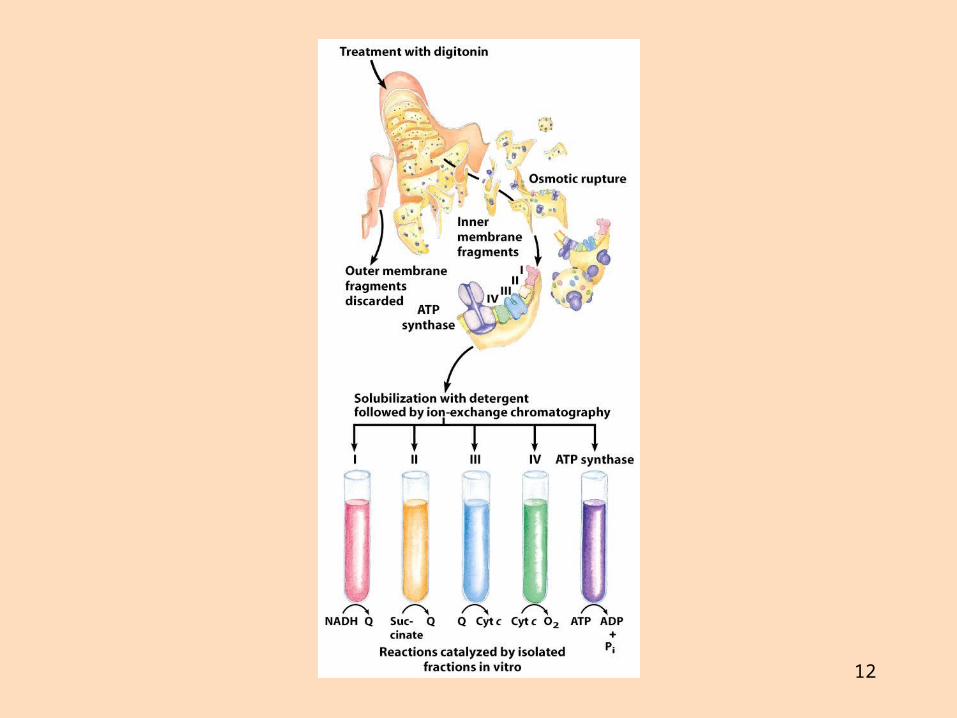
12
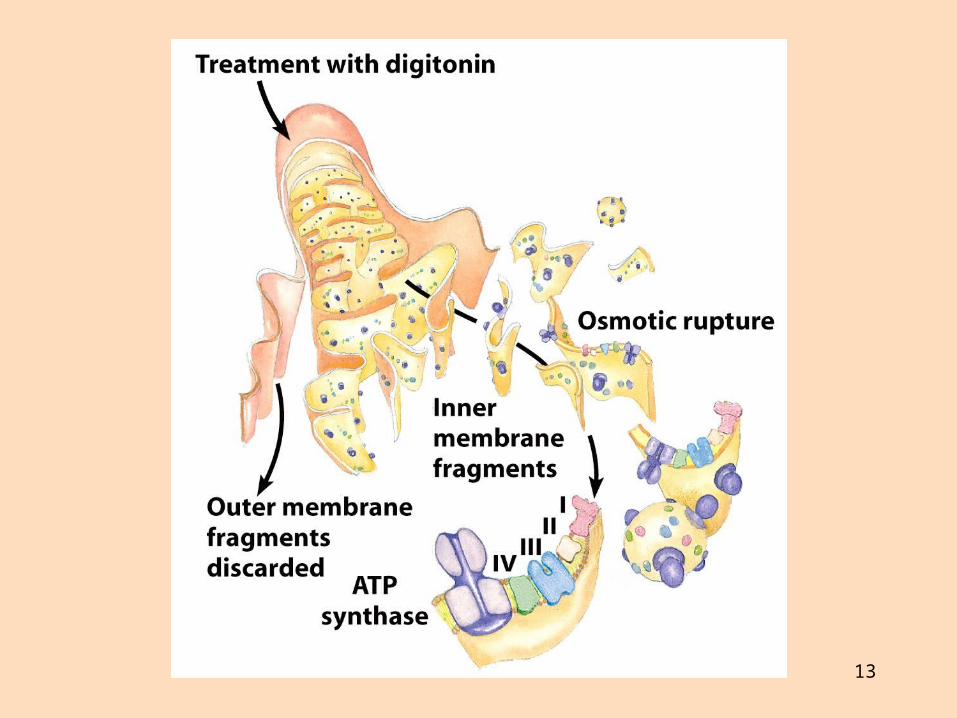
13
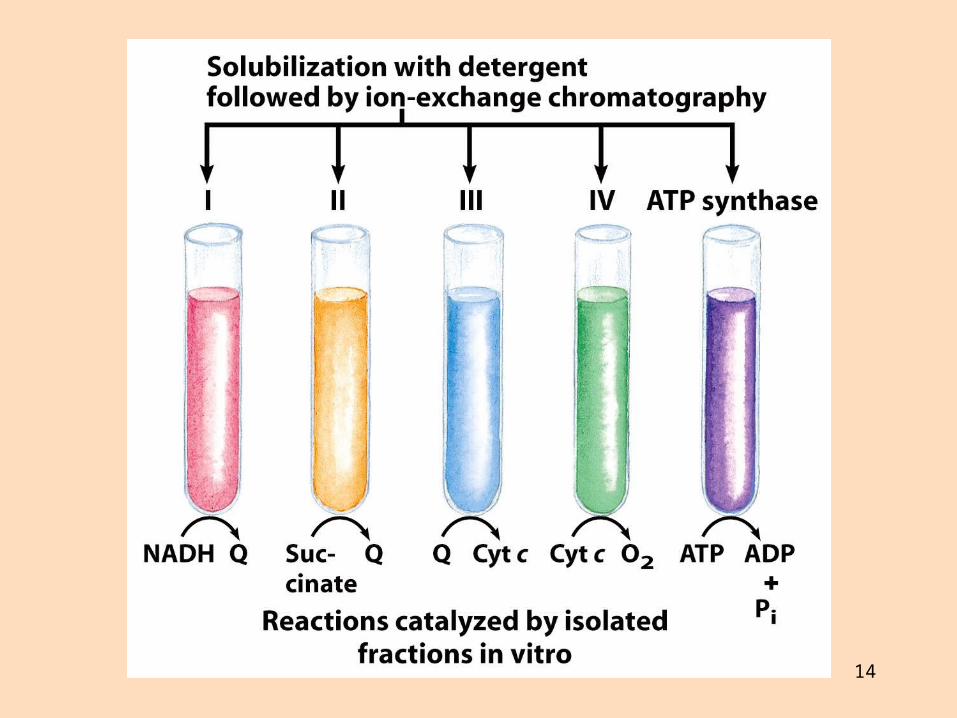
14

15
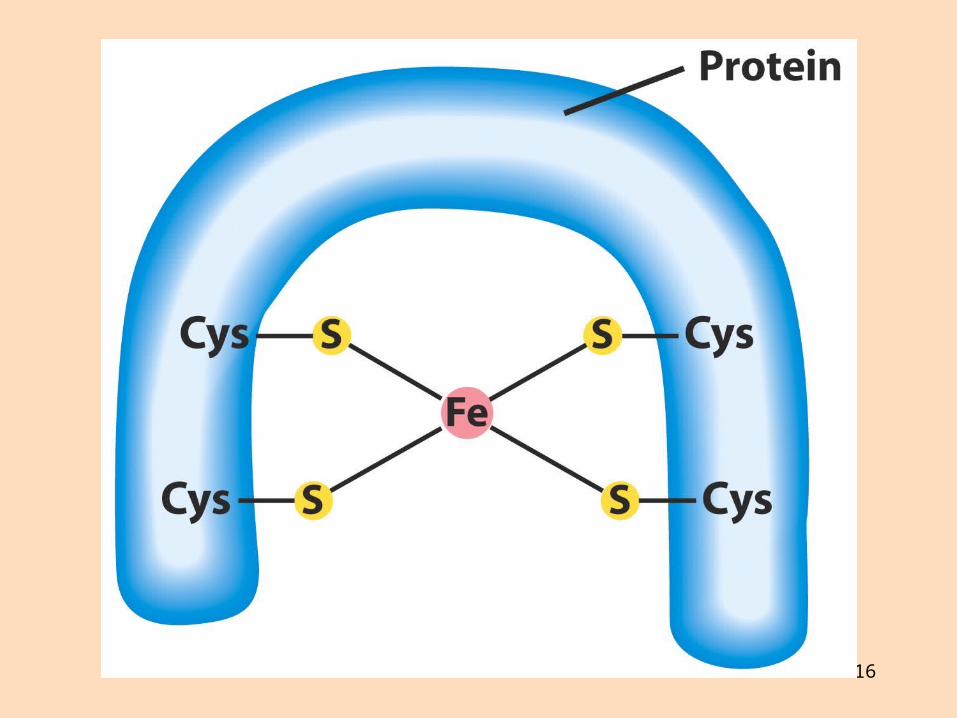
16

17
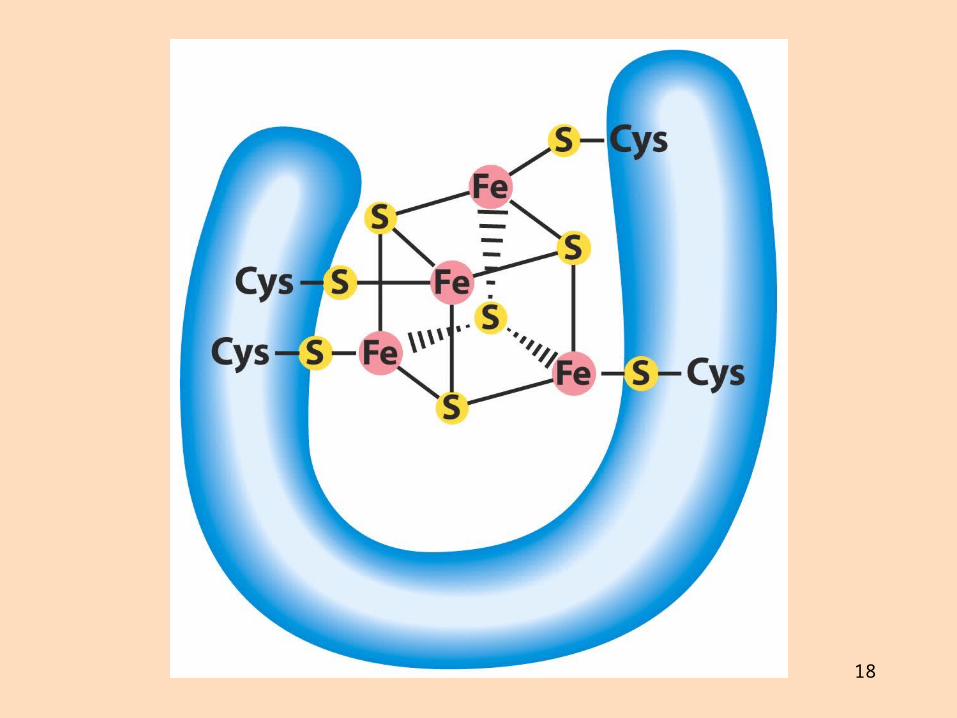
18

19
Ferredoxín úr bakteríu

20
Rafeindaberar og ildisháð fosfórýlering Gerð rafeindaberakeðjunnar
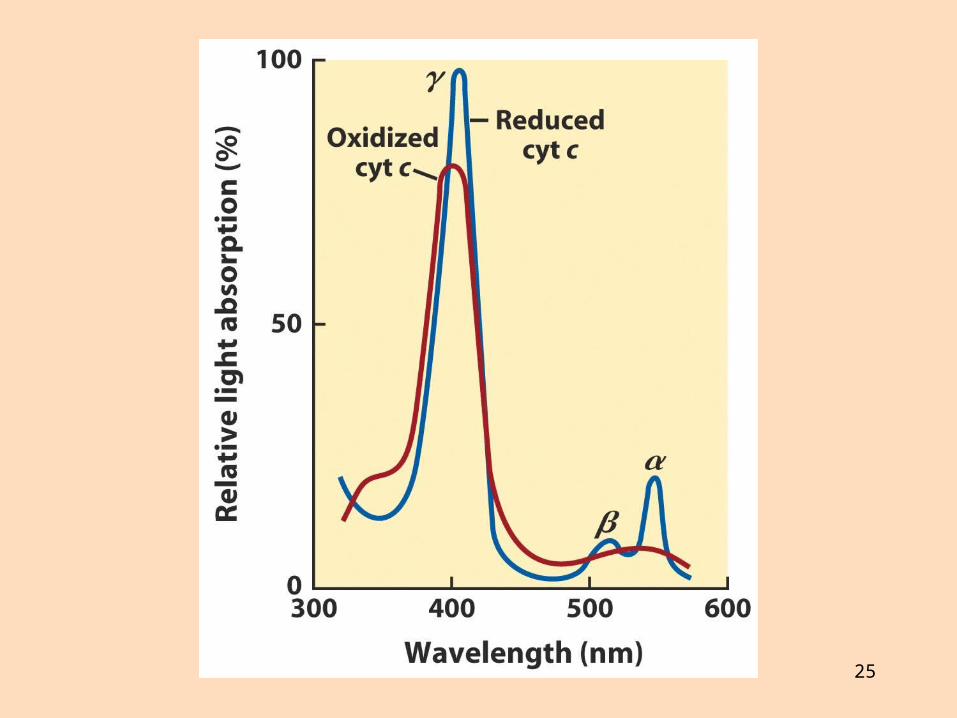
Önnur prótein eru sýtókróm með járnbundinn hemhóp Þau breyta um litrófseiginleika við oxun og afoxun
Ekki er talið að einingar rafeindaberakeðjunnar raðist skipulega upp í innri himnu mítókondría
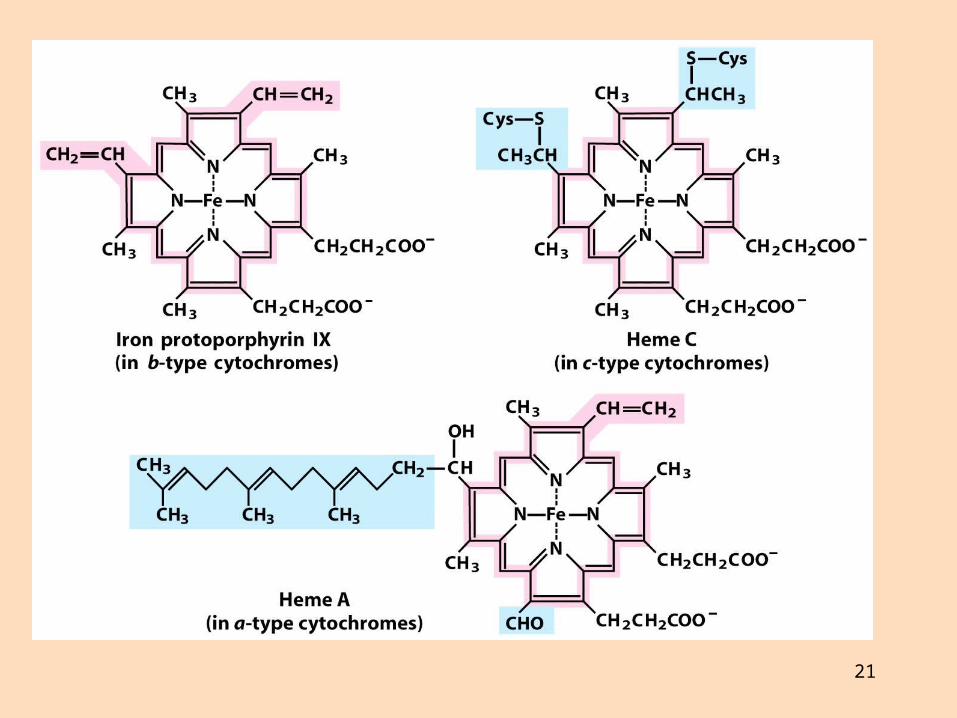
21

22

23

24
25

26
Rafeindaberar og ildisháð fosfórýlering Gerð rafeindaberakeðjunnar
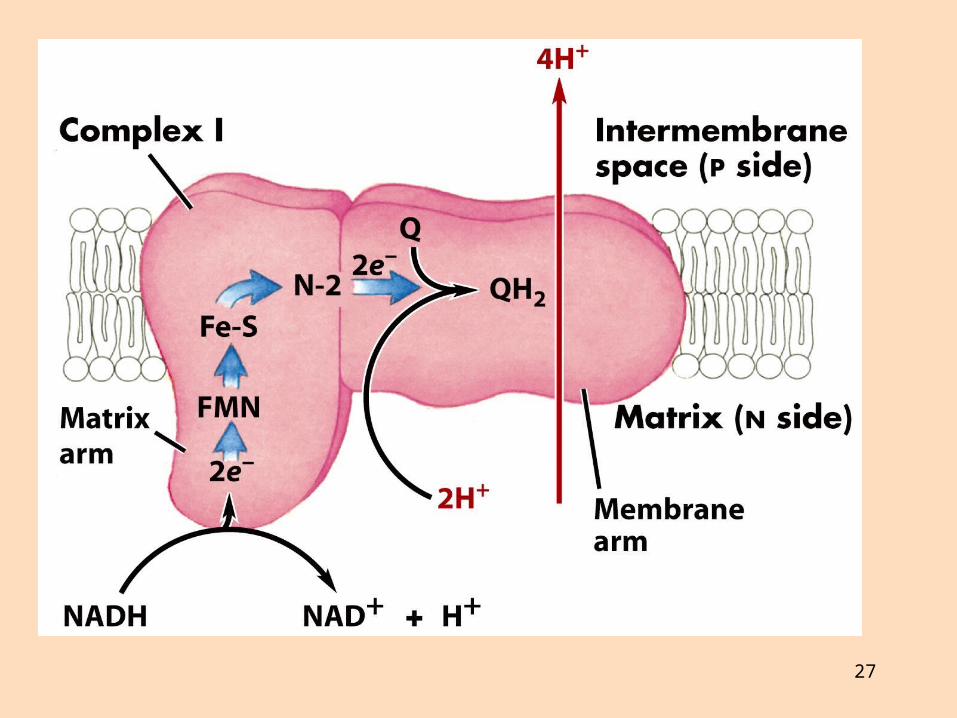
Komplex I (NADH-dehýdrógenasi) tekur við rafeindum frá NADH og flytur á úbikvinón (UQ)
Úbikvinón (rafeindaberi) tekur við FADH2 og tengir NADH-redúktasa við röð sýtókróma
27

28

29

30

31

32
Rafeindaberar og ildisháð fosfórýlering Gerð rafeindaberakeðjunnar
Komplex II er súkksínatdehýdrógenasi auk Fe-S-próteina og er ekki prótónudæla

33

34
Komplex II

35
Rafeindaberar og ildisháð fosfórýlering Gerð rafeindaberakeðjunnar
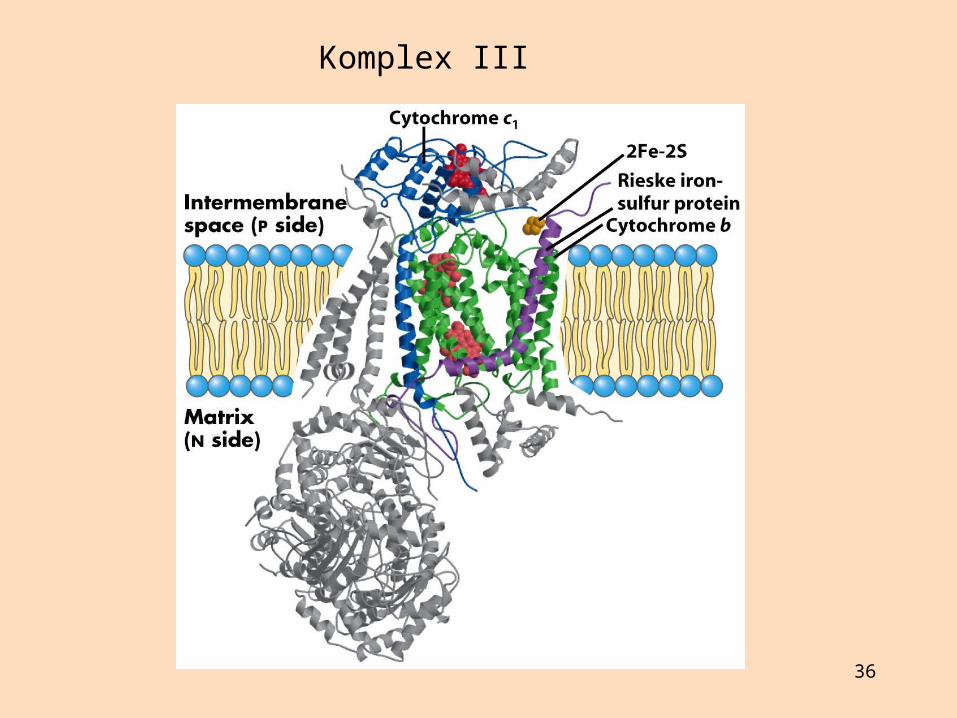
Komplex III: Sýtókróm-bc1-komplex (sýtókrómredúktasi), röð sýtókróma og Fe-S-próteina flytur rafeindir frá úbikvínón að sýtókrómi c (leysanlegur rafeindaberi)
Sýtókróm c er lítið prótein sem hefur verið raðgreint í fjölmörgum lífverum
36
Komplex III

37

38
39

40
Rafeindaberar og ildisháð fosfórýlering Gerð rafeindaberakeðjunnar
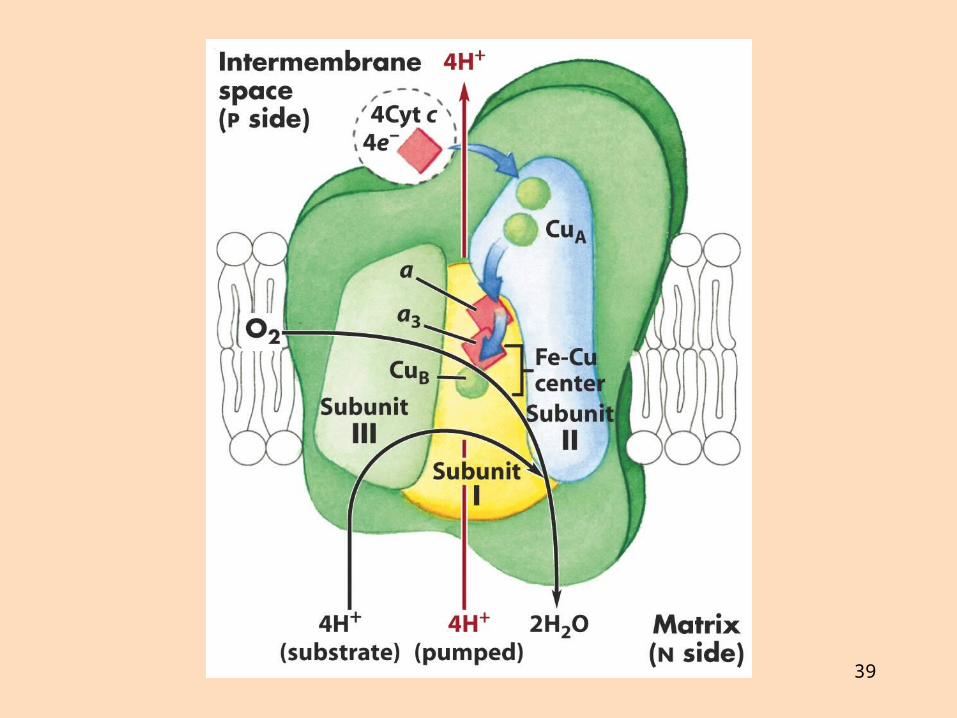
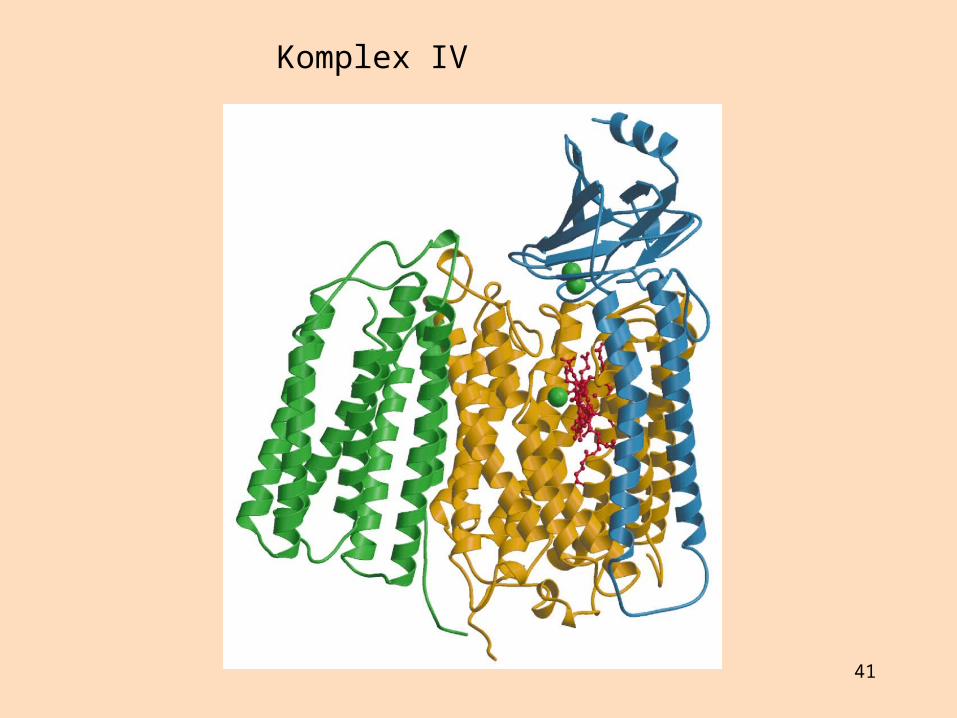
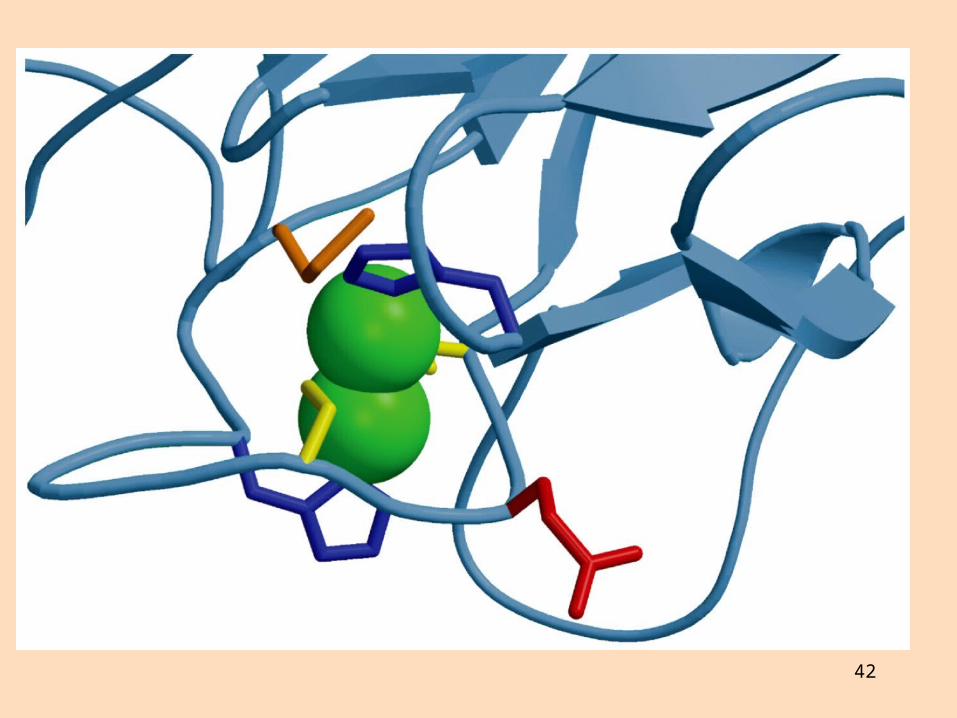
Sýtókrómoxídasi (komplex IV) flytur rafeindir frá sýtókrómi c til súrefnis sem afoxast í vatn
41
Komplex IV
42

43
Hvernig gerist ildisháð fosfórýlering?
Eldri tilgátur:“Chemical coupling hypothesis”, Slater, 1953Tenging við orkuríkt millistigsefni knýr áfram ATP-myndun
“Conformational coupling hypothesis”, Boyer, 1965Prótein sem breytir um lögun knýr áfram ATP-myndun (er að hluta til rétt)
Viðtekin tilgáta:“Chemiosmotic hypothesis”, Peter Mitchell, 1961

44

45

46
Hver eru helstu atriði kemíosmótísku tilgátunnar?
1. Við rafeindaflutning (oxun kóensíma á kostnað súrefnis) í rafeindaberakeðju er prótónum dælt úr mergholi
mítókondría gegn styrkleikastigli2. Við það myndast prótónustigull og himnuspenna, þ. e. prótónukraftur
ATP myndast frá ADP og fosfati (fosfórýlering) og það er losað frá ensíminu ATP-sýnþasa við að prótónur
flæða aftur inn í merghol undan stigliÞetta er í eðli sínu gagnhverf breyting
3. Innri himna mítókondría er ógegndræp fyrir prótónur og aðrar jónir4. Í innri himnu mítókondría eru burðarprótein sem stuðla að
sérhæfðum flutningi prótóna, jóna og nauðsynlegra millistigsefna í efnaskiptum

47

48
Hve mörgum prótónum er dælt þegar rafeindir flytjast eftir rafeindaberakeðju?
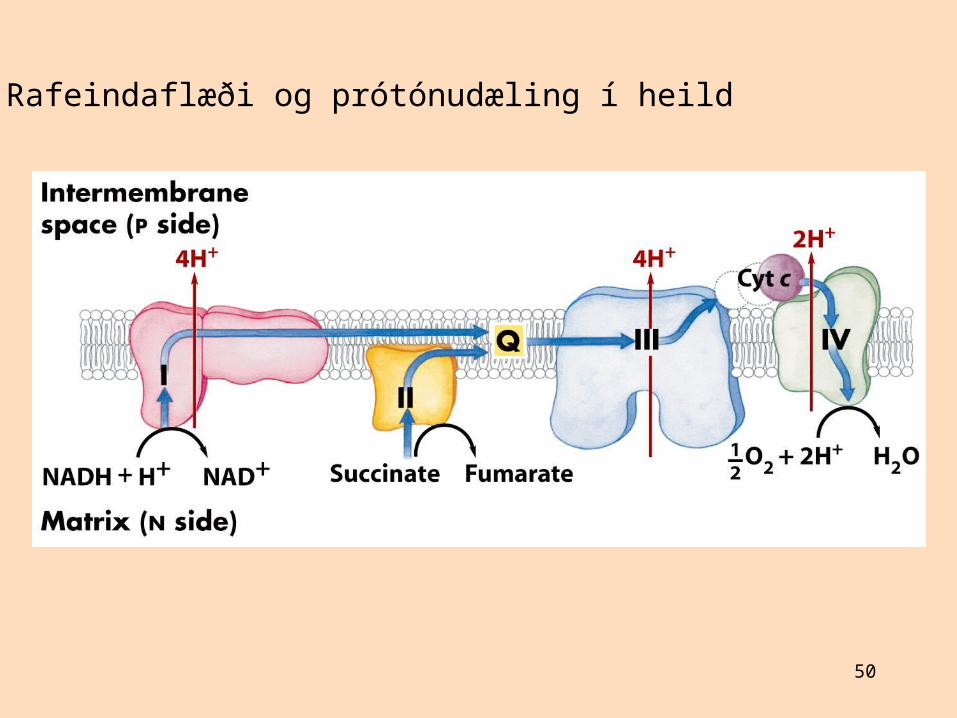
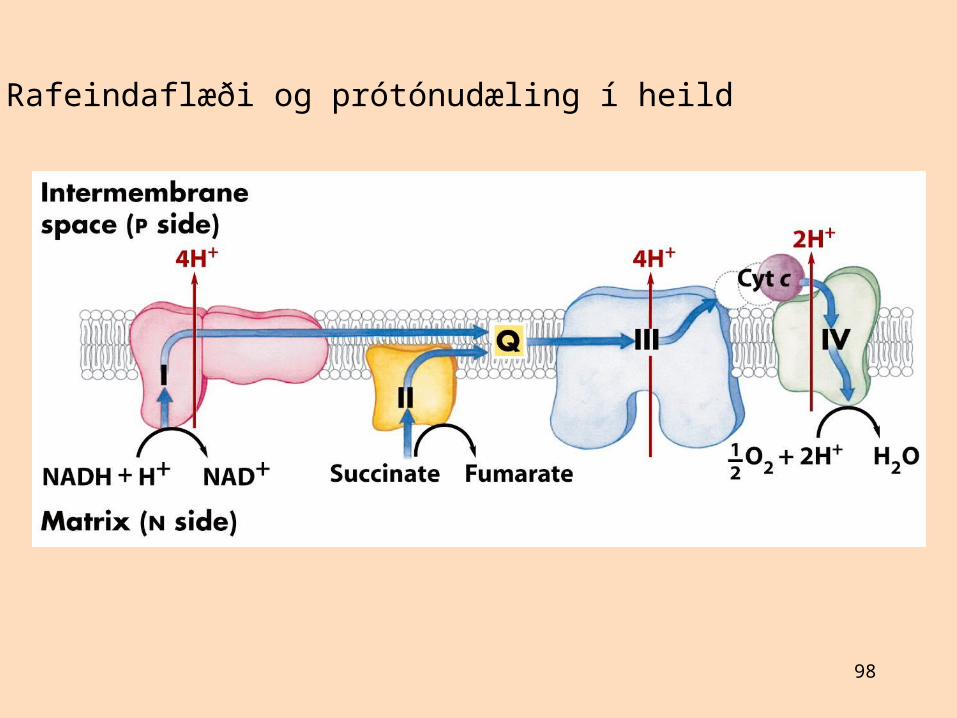
Ekki er vitað nákvæmlega hve mörgum prótónum er dælt út úr mergholi við rafeindaflutningTalið er að NADH-dehýdrógenasi (komplex I) dæli 4 prótónum út úr mergholiTalið er að sýtókróm-bc1-komplex (komplex III) dæli 4 prótónum út úr mergholiTalið er að sýtókrómoxídasi (komplex IV) dæli 2 prótónum út úr mergholiKomplex II, ubíkvinón og sýtókróm c eru ekki prótónudælur

49
Hve mörgum prótónum er dælt þegar rafeindir flytjast eftir rafeindaberakeðju?
Ekki er vitað nákvæmlega hvernig prótónum er dælt út í mergholRafeindaberarnir geta breytt um lögun við rafeindaflutning og dælt prótónum um leiðEinnig myndast spennumunur sem er leggur til mikinn hluta prótónukrafts
50
Rafeindaflæði og prótónudæling í heild

51
Hvernig er sýnt fram á tengsl prótónudælingar við myndun ATP?
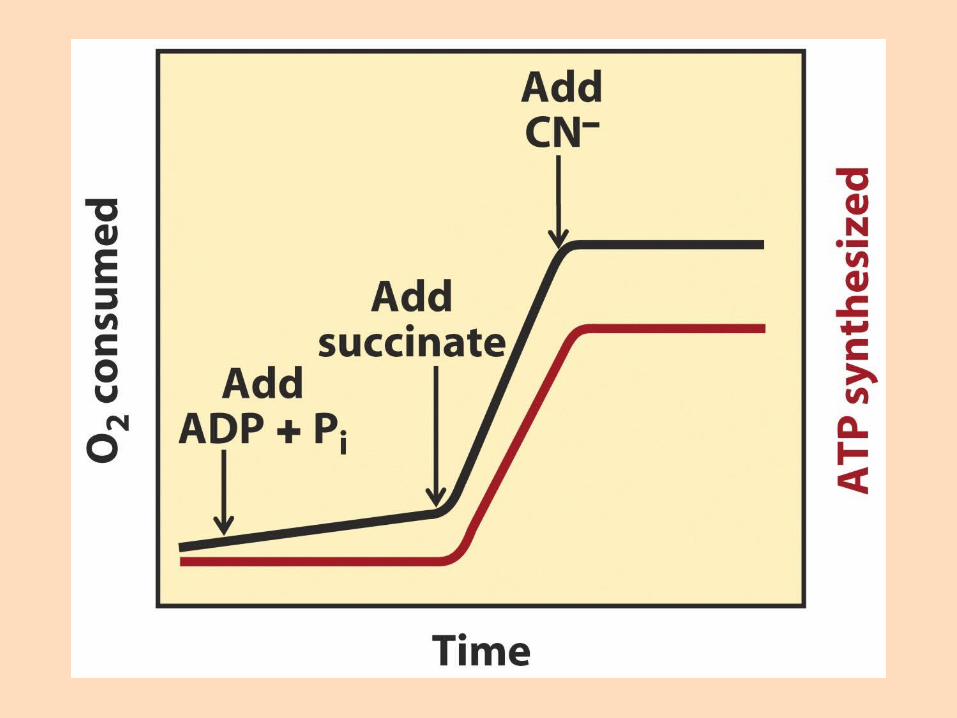
Súrefniselektróða er einfalt tæki, tengt við sírita, sem mælir súrefnisstyrk í lokuðu hvarfahólfiSúrefnisnotkun er hæg þó að oxanlegum efnum sé bætt í sviflausn með mítókondríum
Sé mældum skammti af ADP bætt í lausnina eykst öndunarhraðiHann er stöðugur þar til ADP er upp urið
52
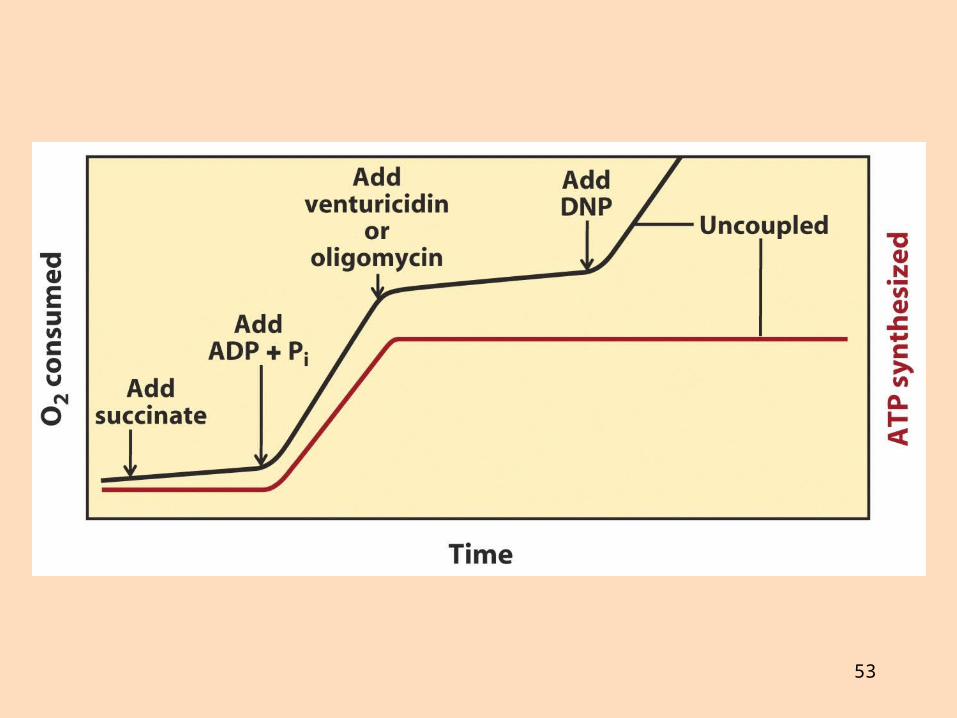
53

54

55
Hvernig er sýnt fram á tengsl prótónudælingar við myndun ATP?
1. ATP afrakstur ýmissa hvarfefna -ketóglútarat: NADH ~ 2,5 ATP súkksínat: FADH2 ~ 1,5 ATP askorbat: ~ 0,5 ATP
2. Varmafræðilegar forsendur G˚´ við rafeindaflutning milli komplexa er borið saman við orku sem þarf til ATP-myndunar

56
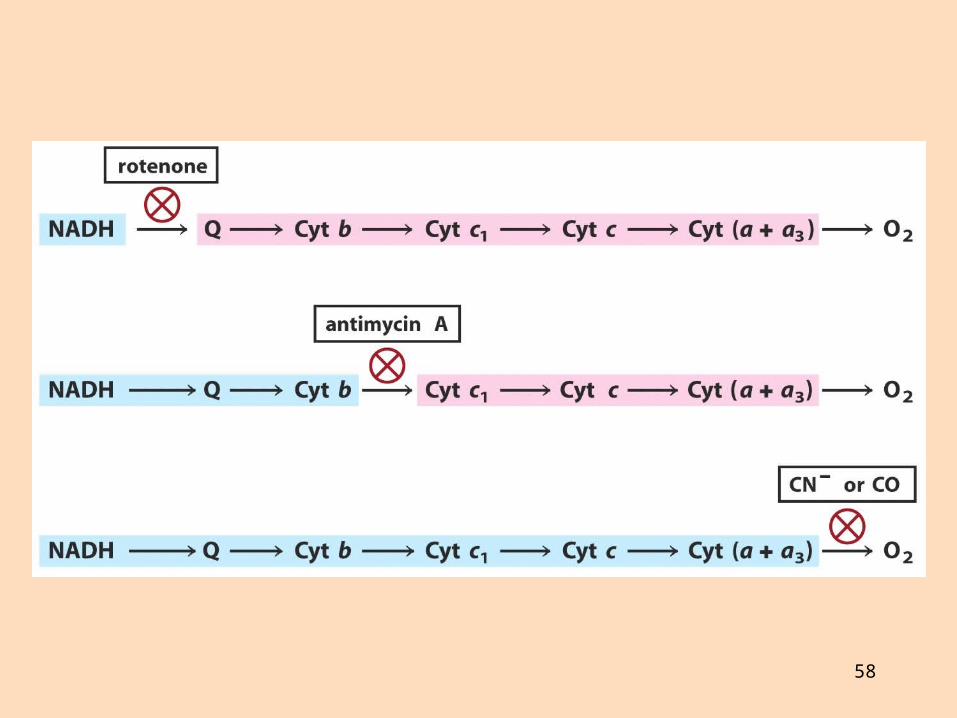
Hvernig er sýnt fram á tengsl prótónudælingar við myndun ATP?
3. Sérhæfð hindrun rafeindaflæðis:
rótenón hindrar komplex I (NADH-dehýdrógenasa/úbikvinón)
antimýsín A hindrar komplex III (sýtókróm-bc1-komplex)
sýaníð, azíð og kolmónoxíð hindra komplex IV (sýtókrómoxídasa)
4. Sérhæfð hindrun ATP-sýnþasa með ólígómýsíni

57
58

59
Hvernig er sýnt fram á tengsl prótónudælingar við myndun ATP?
5. Notkun frátengimiðla
6. Notkun sérhæfðra hindra ATP-ADP translókasa (atraktýlósíð og bongkrekínsýra)
7. Notkun endursamsettra himnudælukerfa

60
Hvaða atriði styðja kemíosmótísku tilgátuna?
1. Prótónustigull myndast við rafeindaflutning
2. Unnt er að knýja áfram ATP-myndun með því að breyta pH
3. Innri himna mítókondría verður að vera heil og óskemmd
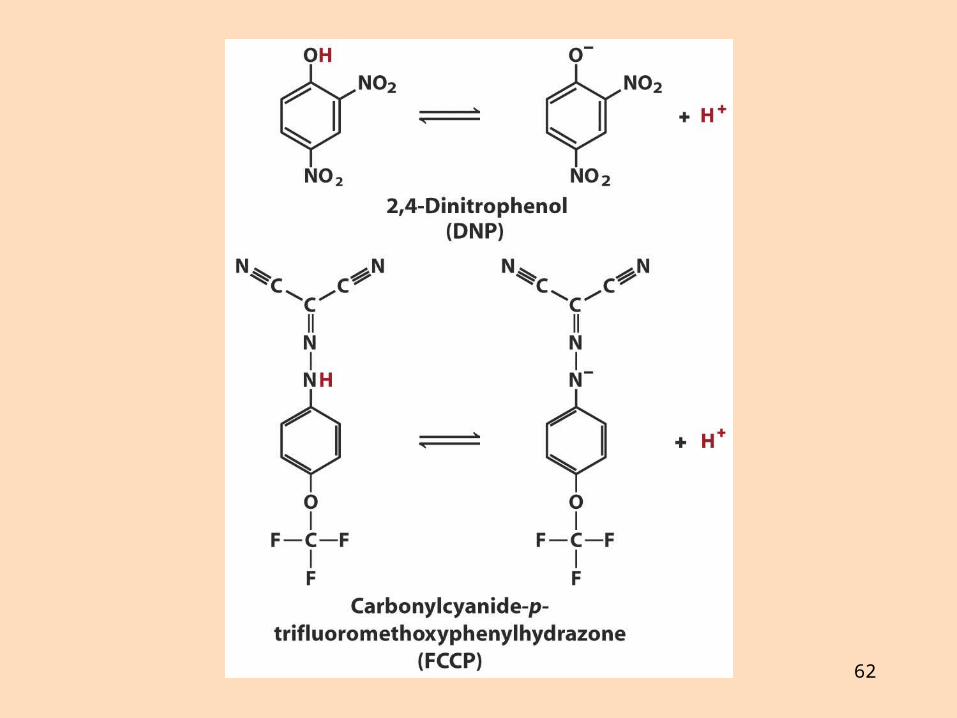
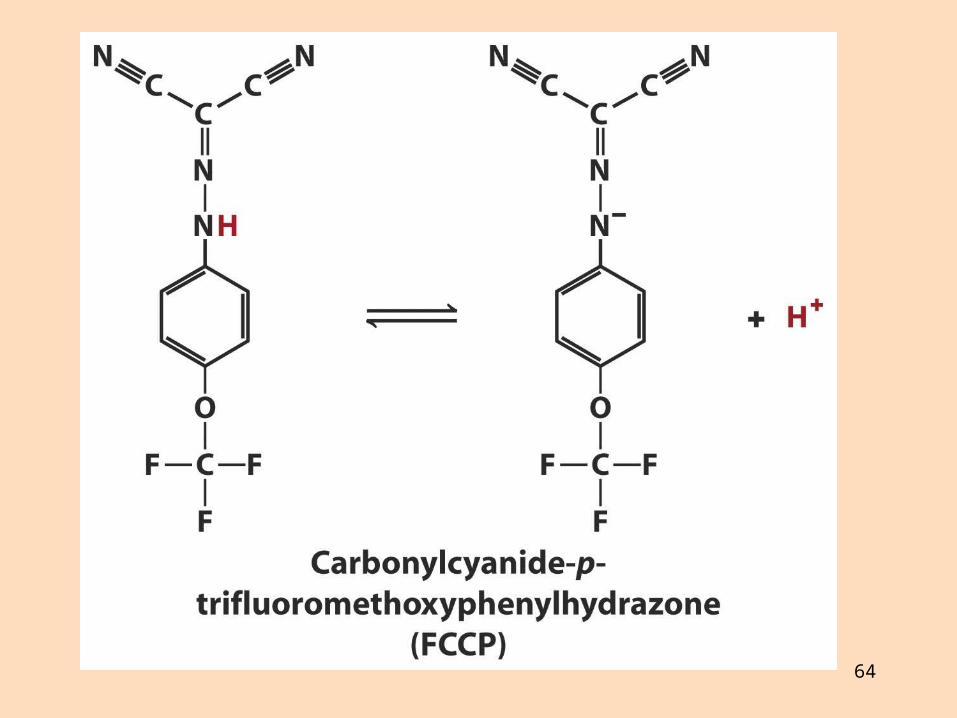
4. Efni sem ferja prótónur yfir innri himnuna eyðileggja prótónustigulinn og frátengja oxun og fosfórýleringu,
þau eru frátengimiðlar, t. d. 2,4-dínítrófenól
5. Svipuð kerfi er að finna í ljóstillífun og fornbakteríum
6. Endursamsett himnudælukerfi geta myndað ATP

61
62

63
64

65
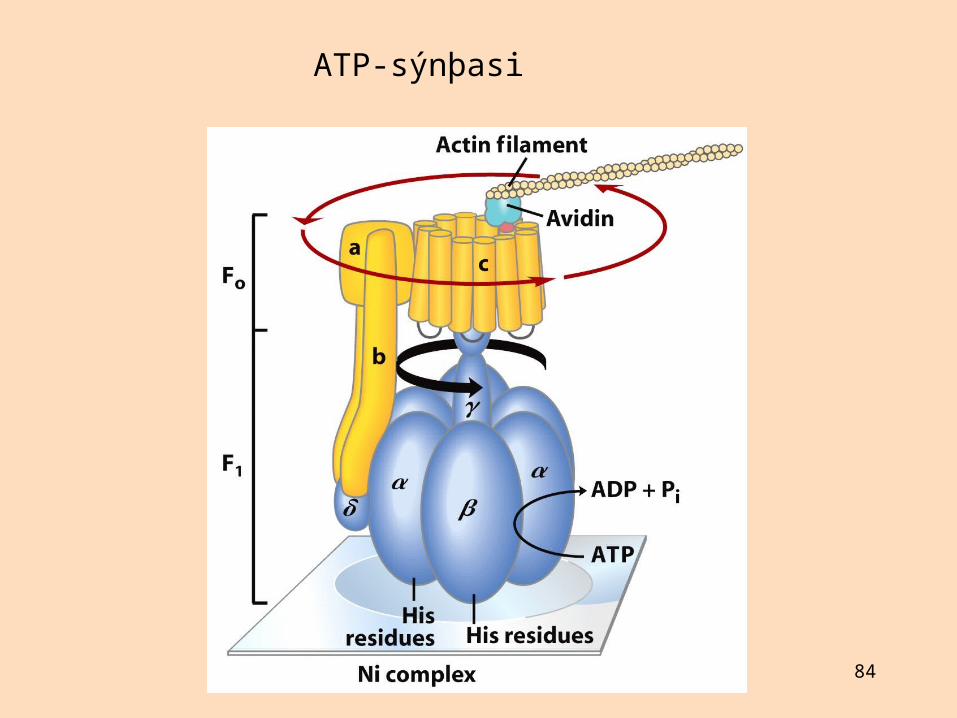
Endursamsett himnudælukerfi
Lípósóm eru gagnleg módel til að kanna flutninga yfir frumuhimnur
Þau geta einnig verið hentug burðarefni fyrir lyf
Með lípósómatækni er unnt er er að endursamsetja starfandi frumuhimnur úr hreinsuðum og einangruðum byggingarhlutum þeirra (úthverfar himnublöðrur)

66
Endursamsett himnudælukerfi
Unnt er að gera blöðrur úr innri himnu mítókondría þar sem F1-einingin snýr út
Slíkt kerfi getur bæði myndað ATP og hvatt vatnsrof þess
Svipað endursamsett dælukerfi gert úr ljósknúinni prótónudælu úr fornbakteríum myndar ATP

67

68
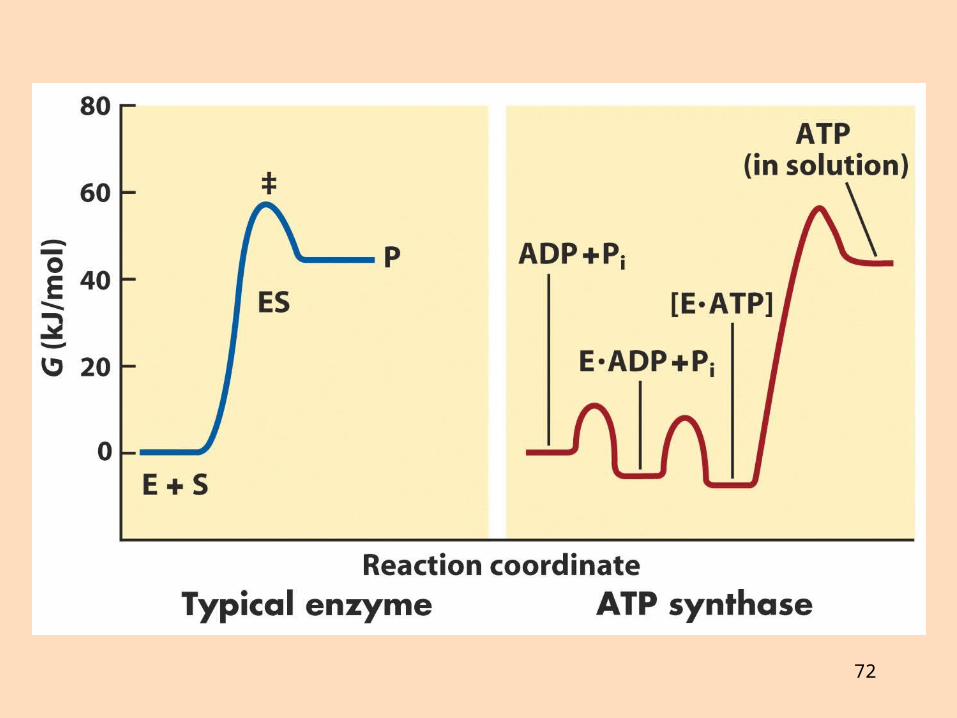
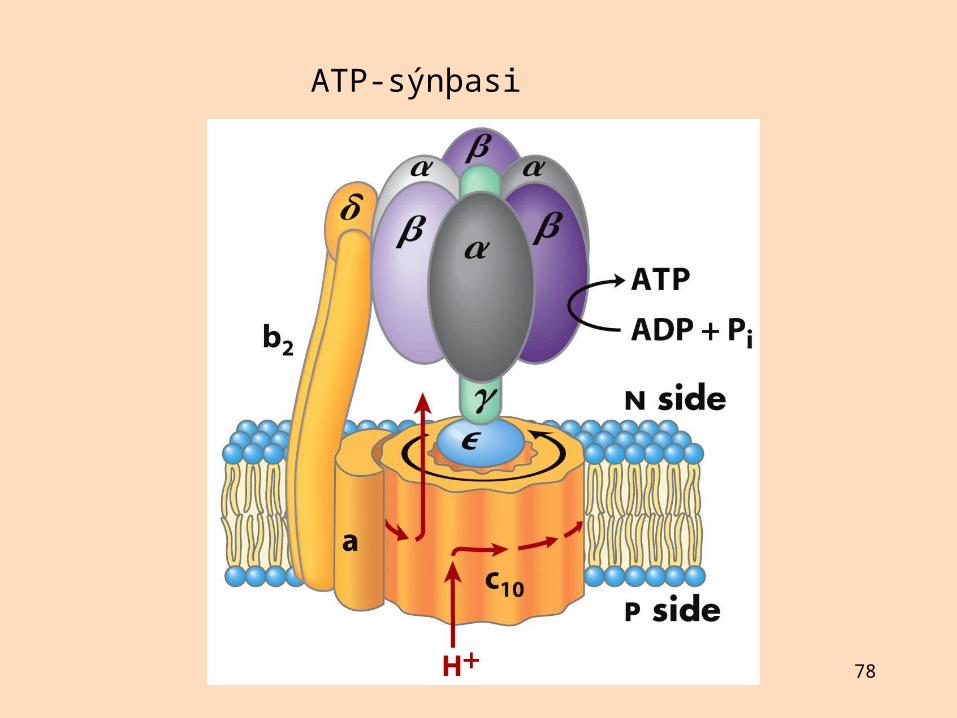
Hvernig myndast ATP?
ATP myndast í ensímakomplex, sem kallast ATP-sýnþasi eða FoF1-ATPasi eða komplex V
Fo-eining er vatnsfælin: göng sem spanna innri himnu og hleypa prótónum í gegnÓlígómýsín hindrar Fo-eininguna í ATP-sýnþasa (oligomycin-sensitive, Fo)F1-eining er vatnsleysin, situr innan á innri himnu, tengd við Fo-einingu: hvetur ATP-myndun

69
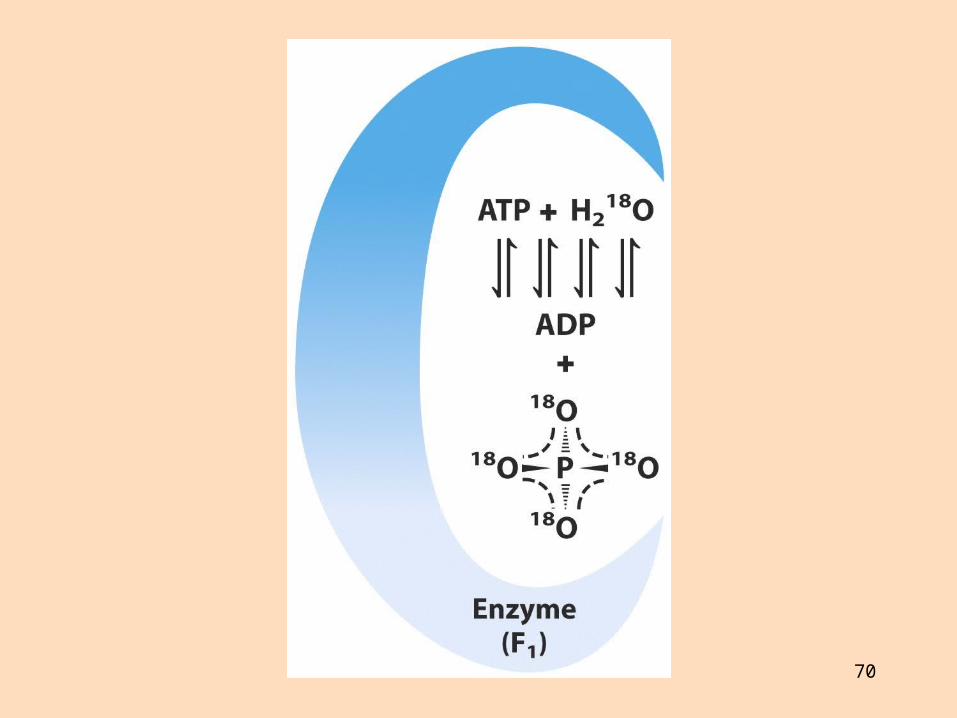
Hvernig myndast ATP?
Á rafsjármyndum sést Fo-eining tengd við stilk sem tengir hana við hnúðlaga F1-einingu
Prótónur sem flæða um Fo-einingu losa emsímbundið ATP af F1-einingu Talið er að þrjár prótónur þurfi til að losa eina ATP-sameind Einn hluti ATP-sýnþasa hreyfist við prótónuflutning og losun ATP
70

71
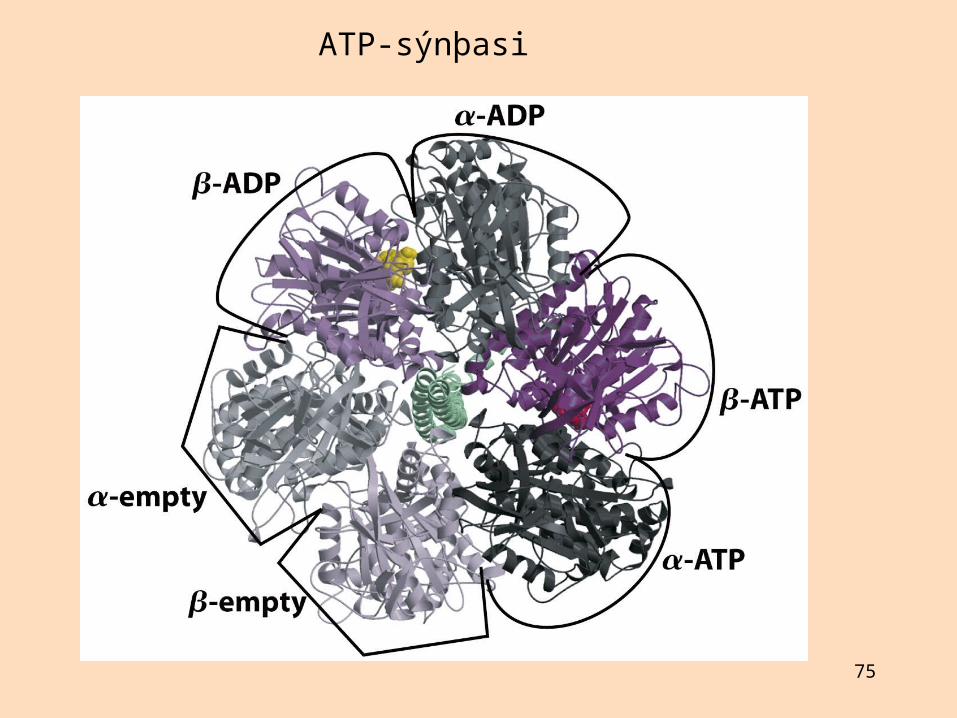
F1- eining í ATP-sýnþasa
72

73
ATP-sýnþasi

74
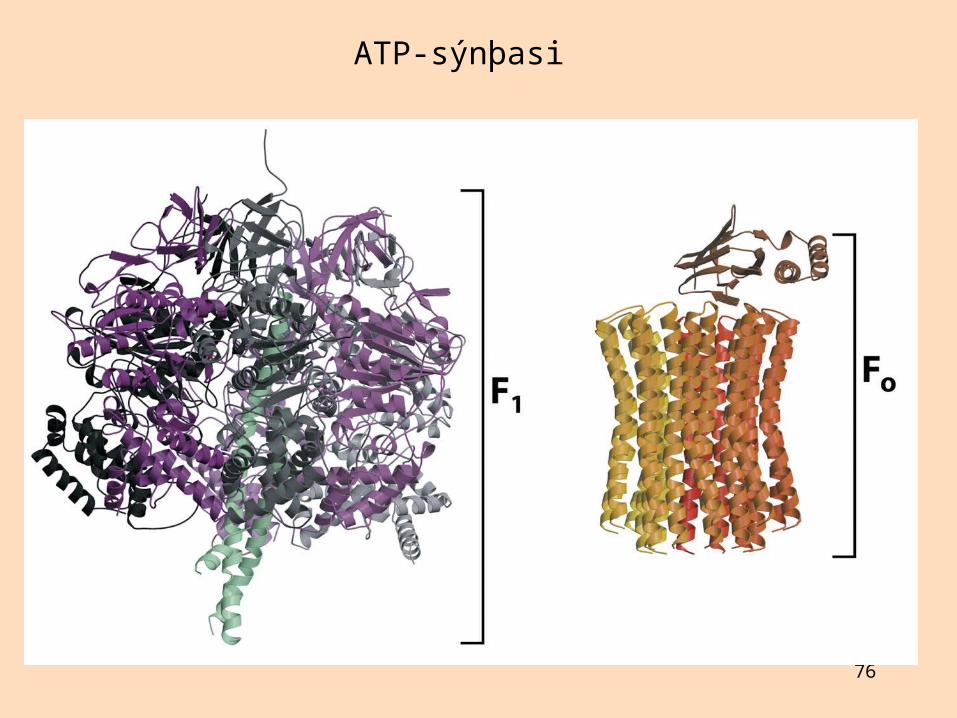
ATP-sýnþasi
75
ATP-sýnþasi
76
ATP-sýnþasi
77
ATP-sýnþasi
78
ATP-sýnþasi

79
80

81

82
ATP-sýnþasi

83
ATP-sýnþasi
84
ATP-sýnþasi

85

86
Flutningskerfi í mítókondríuhimnu
Innri himna mítókondría er ógegndræp fyrir margar litlar lífsameindir:
Ógegndræp fyrir: Gegndræp fyrir (flutt sem):NADH Malat, glýseról fosfatFitu-asýl-CoA Fitu-asýl karnitínAsetýl-CoA SítratOxalóasetat Pýrúvat, aspartat, malatFlestar amínósýrur Glútamat, aspartat, pýrúvat

87
Flutningskerfi í mítókondríuhimnu
Þörf er fyrir þessi efni bæði í frymi og mítókondríumFruman skiptist í hólf (compartments), þ. e. mítókondríur og frymi með tilliti til margra efnaFlutningskerfi eða ferjur (carriers) sjá um flutning efna milli frymis og mítókondríaFlest þessi kerfi eru antiportkerfi, en sum eru symportkerfiKerfin eru himnuspannandi prótein sem hafa svipaða byggingu
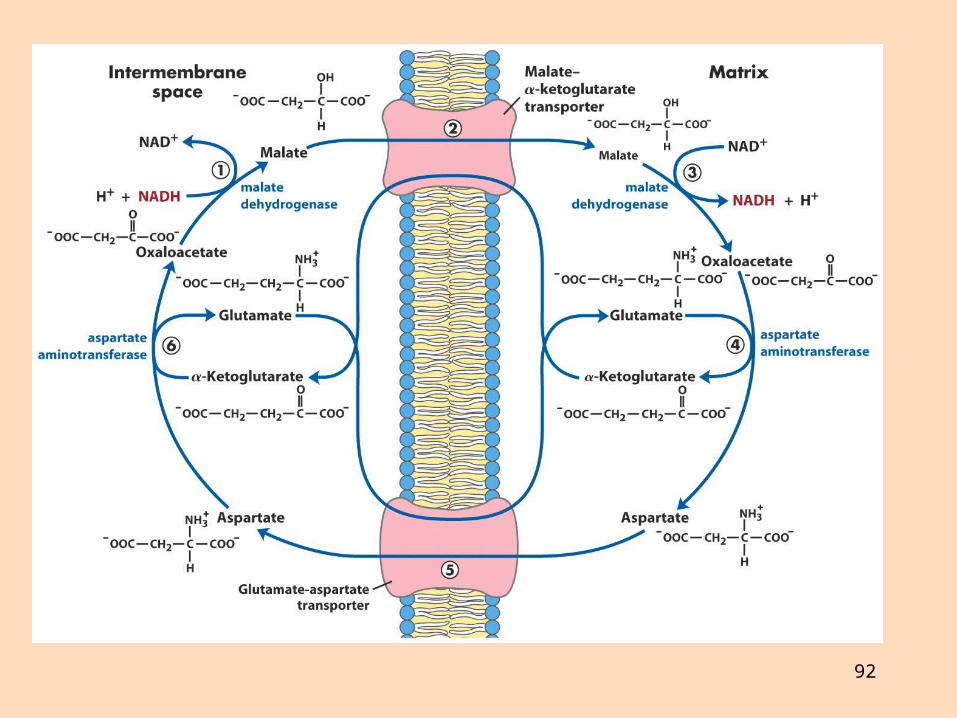
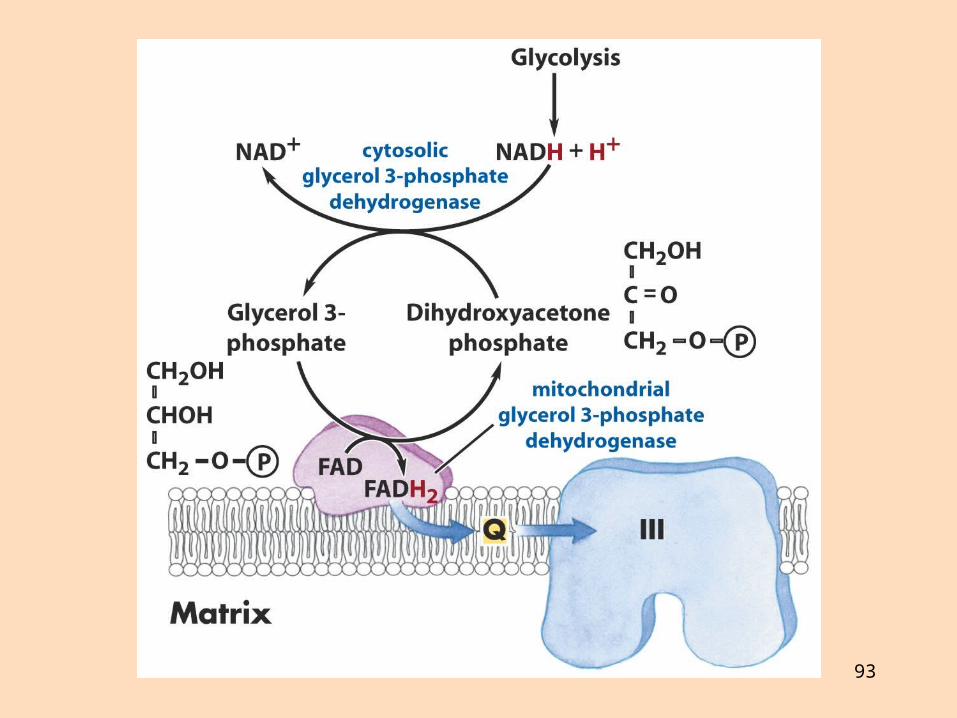
Hvernig flyst NADH úr frymi í mítókondríur?Malatbrúin er gagnhverf, en glýserólfosfatbrúin ógagnhverfGlýserólfosfatbrúin gefur FADH2 innan mítókondría, en malatbrúin NADH

88
Stýring ATP-smíðar og flutningur þess út úr mítókondríum
ADP-styrkur stýrir hraða ildisháðrar fosfórýleringarÞetta kallast öndunarstýring (respiratory control)

89
Stýring ATP-smíðar og flutningur þess út úr mítókondríum
Hlutföll NAD+/NADH og ATP/ADP hafa áhrif á virkni margra ensímaKomplex IV er stýrt með fosfórýleringu og defosfórýleringu
Skjaldkirtilshormón hafa margvísleg áhrifÞau örva smíð frátengipróteina
Þau bindast Komplex IV sem dælir þá færri prótónumÞau örva einnig smíð Na+-K+-ATPasa

90
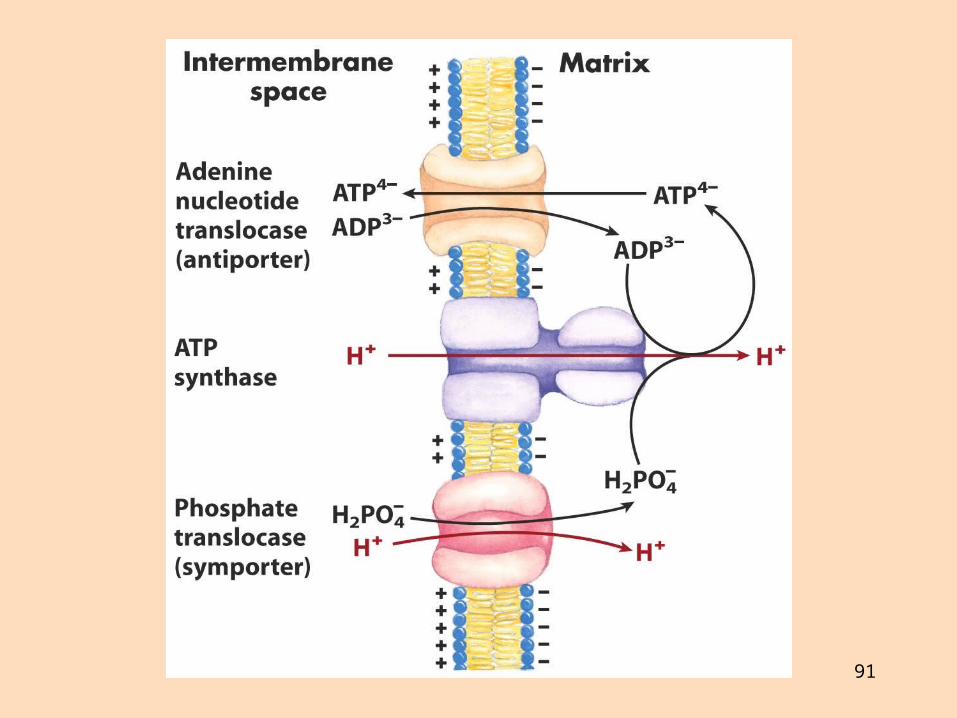
Stýring ATP-smíðar og flutningur þess út úr mítókondríum
Flutningur ADP inn í mítókondríur er tengdur flutningi ATP út úr þeim með ATP-ADP-translókasa (antiport-kerfi)
Atraktylosíð og bongkrekinsýra hindra translókasann
91
92
93

94
Hve mikið ATP myndast?
Lengi var talið að P/O hlutföllin væru 3:1 fyrir hvarfefni sem mynda NADH (-ketóglútarat), 2:1 fyrir hvarfefni sem mynda FADH2 (súkksínat), 1:1 fyrir hvarfefni sem oxast í komplex IV (t. d. askorbat)

95
Hve mikið ATP myndast?
Nú er almennt talið að P:O hlutföllin séu ~ 2,5 fyrir -ketóglútarat, ~ 1,5 fyrir súkksínat, ~ 0,5 fyrir askorbat
Þessar niðurstöður fást úr útreikningum á nettó prótónu og rafeindaflutningum
Við íhugun kemíosmótísku tilgátunnar sést að ekki er nauðsynlegt að heilar tölur fáist fyrir P/O hlutföll

96
Hve mikið ATP myndast?
Einfaldir útreikningar á ATP-myndun frá ýmsum hvarfefnum eru á þessa leið:Það þarf 3 prótónur til að losa eina ATP-sameind frá ATP-sýnþasaFlutningur einnar ADP-sameindar inn í mítókondríur og flutningur einnar ATP-sameindar út í frymi krefst flutnings einnar prótónu inn í mítókondrur
Þannig þarf að dæla 4 prótónum út úr mergholi til þess að knýja smíð einnar ATP-sameindar sem er komin í frymi

97
Hve mikið ATP myndast?
Fyrir hvert rafeindapar sem flyst frá NADH til súrefnis er 10 prótónum dælt út úr mergholi, 10/4 = 2,5 ATP myndastVið að oxa FADH2 er dælt 6 prótónum og 1,5 ATP myndast, við oxun askorbats er dælt 2 prótónum og 0,5 ATP myndast
Ekki er fullvíst hve nákvæmar þessar tölur eru
98
Rafeindaflæði og prótónudæling í heild

99
Hve mikið ATP myndast?
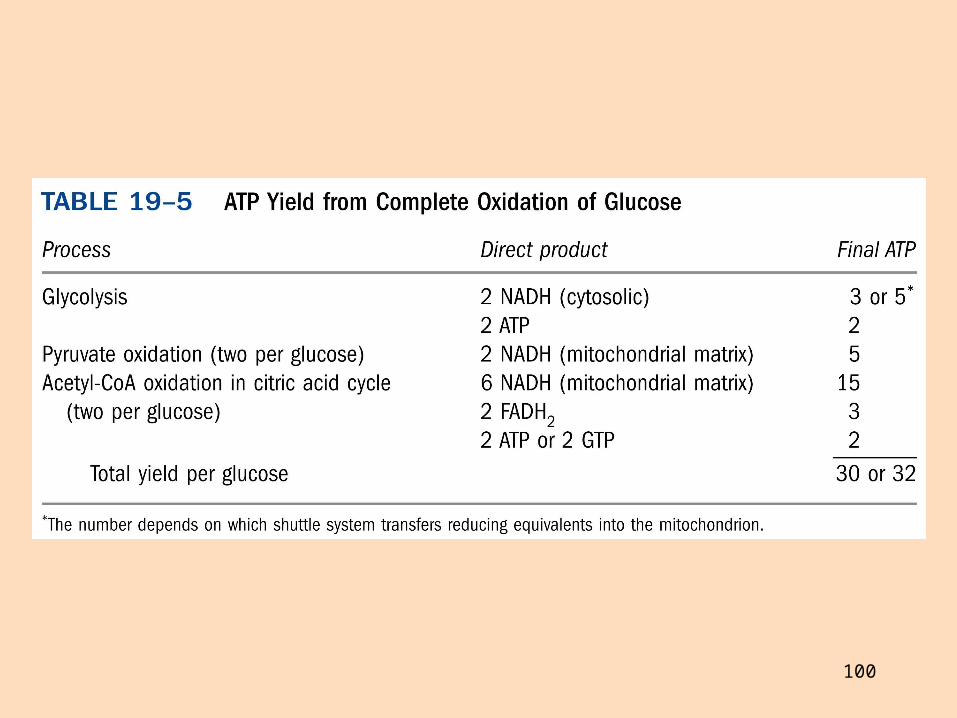
Af þessu leiðir að minna ATP myndast við fullbruna fæðuefna en áður hefur verið talið
ATP-myndun við oxun palmítats nú talin vera 106 (ekki 129)
Við fulloxun glúkósa myndast 30 eða 31 ATP í stað 36 eða 38
Fulloxun glúkósa gefur mismunandi mikið ATP eftir því hvernig NADH úr frymi er flutt (glýserólfosfatferja notar FADH2, en malatferja NADH)
100

101
Frátengimiðlar og brúnn fituvefur
Frátengimiðlar (uncouplers) eru veikar sýrur, leysanlegar í frumuhimnuÞeir flytja prótónur inn yfir innri himnu mítókondría - ATP-myndun verður ekkiÞeim má ekki rugla saman við hindra (inhibitora)Áhrifum frátengimiðils má líkja við rafrás þar sem skammhlaup verður

102
Frátengimiðlar og brúnn fituvefur
Í brúnum fituvef er prótein (uncoupling protein) sem stuðlar að flutningi prótóna inn í mítókondríur
Þetta er mikilvægt til varmamyndunar í ungviði og í dýrum sem leggjast í dvalaBrúnn fituvefur er ekki talinn skipta miklu máli varðandi offitu

103

104

105
Prótónukraftur (proticity)
Prótónustigull og spennumunur myndast í elektrónuflutningskeðju og í ljóstillífunPrótónukraftur er notaður til að knýja mörg ferli í lífríkinu: virkur efnaflutningur, varmi, ATP-smíð, hreyfingar baktería
Til þess að prótónustigull geti orðið þarf að aðskilja tvo vatnsfasa með þunnri lípíðahimnu

106
107

108
109

110
Mítókondríugen
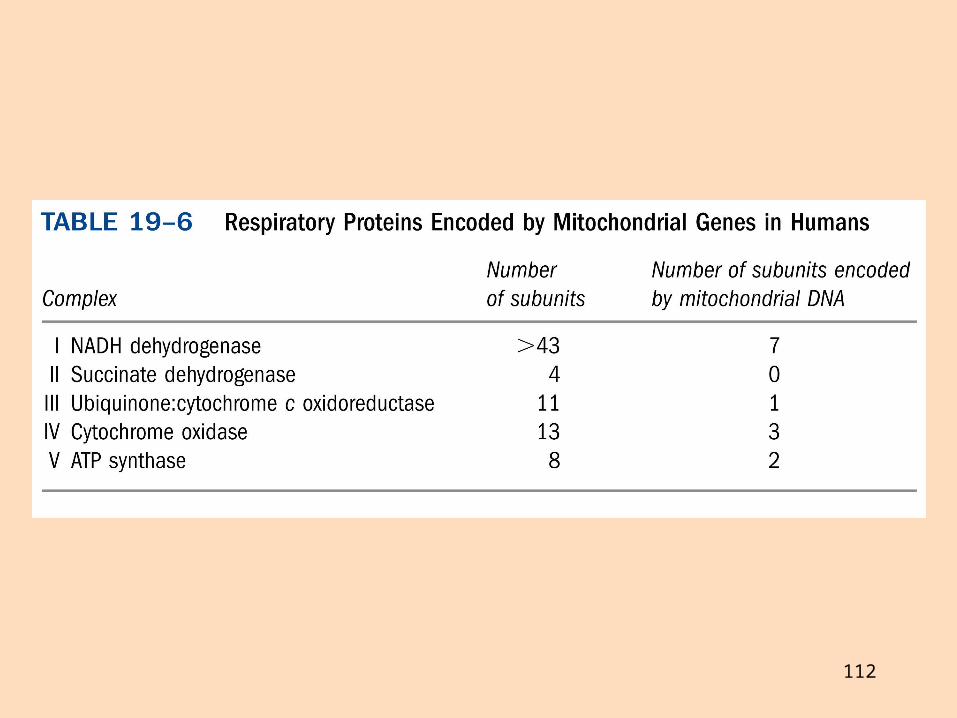
Mítókondríur dýra hafa eigið erfðaefni, hringlaga, tvíþátta DNA (mt-DNA) sem skráir 37 gen (13 prótein, 2 rRNA og 22 tRNA)Önnur mítókondríugen eru skráð í kjarna og próteinin flutt inn í mítókondríurMítókondríugen fósturs koma öll frá móður og stökkbreytast mun hraðar en önnur genÍ nokkrum tilvikum skrá táknar aðrar amínósýrur í mítókondríu-DNA en í erfðalyklinumMítókondríur þróuðust sennilega frá samlífisbakteríumTil eru sjaldgæfir erfðakvillar þar sem stökkbreytingar verða á mítókondríugenum

111
Mítókondríugen
Hafa mítókondríur þróast frá samlífisbakteríum?Vísbendingar:Genin eru hringlagaNokkrir táknar eru öðruvísi, og eru þeir sömu og í sumum bakteríumÞað er næstum ekkert kólesteról í innri himnu mítókondría og ekkert í frumuhimnu baktería
112

113
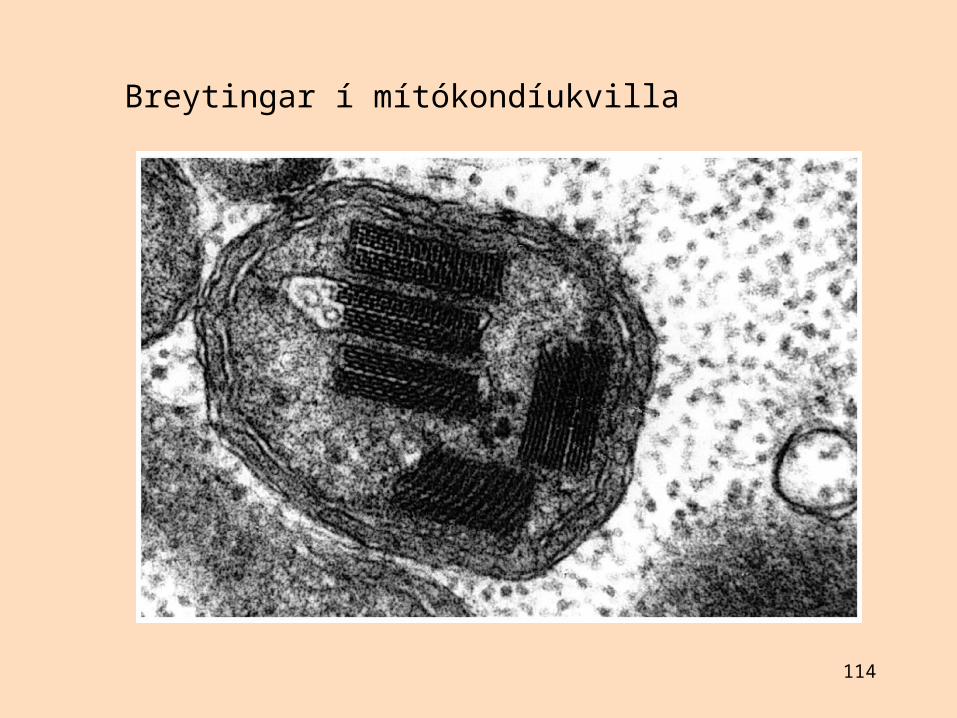
114
Breytingar í mítókondíukvilla

![[PPT]Chapter 18 Oxidative phosphorylation and … · Web viewTitle Chapter 18 Oxidative phosphorylation and photophosphorylation Last modified by h Document presentation format 全屏显示(4:3)](https://img.pdfslide.net/doc/110x75/5aee63477f8b9ac62b8c00c8/pptchapter-18-oxidative-phosphorylation-and-viewtitle-chapter-18-oxidative.jpg)










![SMJK PEREMPUAN CHINA PULAU PINANG SMJK ... - · PDF fileThe ATP is produced by oxidative phosphorylation. ... Distinguish between cyclic and non-cyclic photophosphorylation. [2]](https://img.pdfslide.net/doc/110x75/5abe113d7f8b9aa3088c6ff7/smjk-perempuan-china-pulau-pinang-smjk-atp-is-produced-by-oxidative-phosphorylation.jpg)
















