Embed Size (px)
Citation preview

‘H Editing of 2-Deuterated Exogenous and Natural Endogenous D-Glucose in Biological Samples Monique Biesemans, Franqois Kayser, Marc Van Cauteren, Nancy J. Rehrer, F’eter Neirinck, Kenny de Meirleir, Willy J. Malaise, Rudolph Willem
A spin-echo based ’H homonuclear pulse sequence, enabling the selective editing of homonuclear first-order &multiplet types, is presented and analyzed. Its effectiveness in quanti- fication applications is assessed. Its potential usefulness in the quantitative distinction between 2-deuterated and natural o-,glucose in biological samples is briefly discussed, with Bm- plhasis on studies of hexose metabolism conducted in vitro. Key words: ’H NMR editing; &modulation; glucose quantifi- cation; plasma glucose.
INTRODUCTION
The question of distinguishing and quantifying exoge- nous administered from endogenous natural D-glucose in cellular extracts and other biological samples is of major biomedical importance in studies of the metabolism of carbohydrates in living organisms. This is more particu- larly the case in the measurement of D-glucose turnover, a:; performed in either the whole organism or in isolated cells incubated in vitro.
Alternatives to the use of radiotracer techniques, to NMR-editing methods using exogenous 13C-labeled sub- sirates (1, 2) or to “biochemical” editing in ‘H spectra, as proposed recently (3), may prove useful in the quantifi- ccition of exogenous and endogenous glucose. Using deu- terated substrates can he appropriate to this purpose.
Jue presented homonuclear ‘H editing sequences ex- ploiting both JHH or JCH modulations, combining selec- tive and nonselective pulses, with the goal of specifically elditing a mixture of [3-I3C]- and [3- ”C]-L-alanine (I), as well as [3-I3C]-alanine alone in the latter mixture (4). Jue et al. proposed also a so-called “anti-editing” spin echo sequence (5). enabling the specific editing of ‘H-singlets and central multiplet components of odd multiplets (trip-
MRM 30:120-123 (1993) From the VUB, High Resolution NMR Centre (M.B., F.K., R.W.), VUB, Bio- medical MR Centre of the Academic Hospital (M.V.C.), VUB, Human Phys- iology and Sports Medicine Department N.J.R., P.N., K.D.M.), and ULB, Laboratory of Experimental Medicine, Erasmus Medical School (W.J.M.), Free University of Brussels, Belgium. Address correspondence to: Rudolph Willem, Ph.D., Vrije Universiteit Brus- sel, High Resolution NMR Centre, HNMR & AOSC, Room 8G508, Pleinlaan 2, 8-1050 Brussel, Belgium. Received December 2, 1992; revised February 11, 1993; accepted February 12, 1993. This work was supported by the Grant No. 9.0050.90 of the Belgian “Natio- nale Loterii” and by a grant of the Research Council of the Free University of Brussels (“Vrije Universiteit Brussel”) (M. B., R. W,). Grants from the “Min- istere des Affaires Culturelles du Luxembourg” (Grant No. BFR90/036) and from the “Ministere de Education Nationale du Luxembourg” are acknowl- edged (F. K.).
Copyright 0 1993 by Williams & Wilkins All rights of reproduction in any form reserved.
0740-31 94/93 $3.00
lets, quintuplets, etc. . .), under suppression of any other homonuclear first-order multiplets. The present paper proposes likewise a homonuclear editing sequence, that allows the use of ’H- rather than 13C-enriched substrates, and exploits solely the homonuclear JHH modulation in an appropriate phase cycling scheme. Its effectiveness in distinguishing, editing and quantifying exogenous [Z- ’HI- and endogenous natural [2 - lH]-~-glucose is illus- trated with edited ‘H spectra acquired from human plasma samples to which known amounts of deuterated o-glucose were added for methodology set up purposes.
MATERIALS AND METHODS
All lH spectra discussed below were obtained at 500.13 MHz on a Bruker AMX5OO spectrometer, using water signal presaturation for 4 s prior to the application of the phase-cycled sequence presented in the Principles sec- tion. The spectra were obtained either from model solu- tions containing variable concentrations of [ 2-’H]- and [2-’H]-~-glucose in 99.8% ’H,O in the range 0.1-3 mg/ ml, or from samples of lyophilized blood plasma from a healthy volunteer, which were dissolved in 99.8% ’H’O and to which appropriate amounts (0.1-3 mg/ml) of [ Z - ’H]-o-glucose were added. The repetition delay was held constant throughout all acquisitions at 9 s. The number of scans was 32, a multiple of 4, as required by the pulse sequence (see Principles section).
Calibration straight lines for quantification purposes were obtained from glucose solutions and plasma sam- ples with reference to an external chloroform standard. The signal of the latter was acquired simultaneously to those of the actual sample from a sealed capillary of ca. 85 mm length and 1.5 mm cross-section, introduced into the bottom of the 5-mm NMR tube containing the sample of interest. The natural endogenous o-glucose concentra- tion in the plasma sample reconstituted in ’H,O was determined by an adduct method. It consisted of adding known aliquots of D-glucose to the plasma sample with unknown concentration, to determine the slope of the straight line reflecting the signal integral increase as a function of the added D-glucose, and to multiply the in- verse of the latter slope by the area of the D-glucose signal obtained from the sample prior to the glucose aliquot additions (see Results and Discussion section).
PRINCIPLES
Our approach is based on the spin echo sequence (6), applied four times with incremented delays in a FID- coaddition scheme, as in the original “anti-editing” pro- posal of Jue et al. (5) (Fig. 1).
120

'H-Etfiting of D - G ~ U C O S ~ in Biological Samples 121
90°, 1 80°,
ACQUISITION
FIG. 1. Spin echo sequence (3, 4) applied in an add-subtract scherne with the delays T = N/4J ( N = 1, 2, 3, 4)
Three important new features, however, characterize our implementation of the sequence.
FiIst, we use shorter delays, r = Nl4J, with N = 1, 2 , 3 and 4, compared with 7' = 27 = (2n+1)/2J, with n = 0, 1, 2, and 3 as in the sequence of Jue et al. (5). This enables reduction of T2 attenuation in the detected spin echo, providing a better signal-to-noise ratio. In addition, this reduces residual artifacts of suppression of undesired signals during echo coaddition, resulting from increasing extent of transverse relaxation as the applied echo delay IS increased.
Secondly, we implement receiver phase cycling into the spin echo acquisitions. This enables selection at will of several types of multiplet pattern, instead of singlets solelv (5).
Last but not least, the phase cycling schemes intro- duced allow simplification of the multiplet patterns, in that i hey usually select only part, sometimes even only one single line of the multiplet. This leads to an appre- ciablls simplification of the JHH multiplet patterns and suppresses redundant spectral information.
Because only single quantum coherences are generated during the application of the sequence (first-order ap- proximation), a simple Bloch equation based formalism suffices to describe the procedure of multiplet compo- nent selection and to understand the above mentioned features. Thus, the quadrature signal obtained for a typ- ical doublet D after acquisition obeys the equation:
Table 1 Amplitude and Phase of Low-Field and High-Field Components of Doublets as Well as of Singlets, as Obtained from Eqs. [3]-[5], Neglecting Relaxation during the Echo Period 27
1 2 3 4 Signal sum:
1 2 3 4 Sicgnal sum: 1 2 3 4 Signal sum:
90 180 270
0
270 180 90 0
where ? refer to whether the low-field (+) resp. up-field (-1 components of the doublet D are considered (assum- ing J > o), T represents the delay as defined in Fig. 1, t represents the acquisition time, vD represents the reso- nance frequency of the doublet, and MOD represents its equilibrium magnetization. Taking trivially J = 0 reduces the above expression to a quadrature singlet signal S obeying the equation:
t
[21
where the symbols have the same meaning as in Eq. [ I ] , except for the use of S for singlet instead of D for doublet. Introducing simultaneously into Eqs. [I] and (21 both the delay parameter T, expressed as N/4J, and the influence of phase cycling the receiver, using the parameter +,-,,, de- fined as the shift in the phase of the receiver with respect to the phase of the 180" pulse in the experiment with T = N/4 J leads to
- 12Trvsl 2 T
S,(f) = iM,,,e-Z e -<
[31
where T, relaxation is assumed to be negligible during the delay 27 (i.e., 2 d T , << 1).
Table 1 gives the expressions for the amplitude and phase of the signals SD+(t,N,+N), SD-(t,N,+N), and Ss(t, N,+N)r obtained from Eqs. [3] to [5] respectively, for the four acquisitions with T = Nl4J, N = 1, 2 , 3, 4 and their sum, applied to three different receiver phase schemes. It reveals that with the phase cycle schemes (+N = 90, 180, 270, O"), (+N = 270, 180, 90,O") and (& = 0,0, 0, 0") the high field doublet component, SD-(f ,N,+J, the low field doublet component, SD+(t,N,+N), and the singlet S,(t,N,+,) are respectively edited after coadding of the four FIDs. In practice we achieved this coadding in the time domain. The latter phase cycle scheme is the special case used by Jue (5) for the editing of singlets.
Similar arguments can be used to edit other multiplet components according to the phase cycling scheme used. However, singlets, central components (m = 0 compo- nent) and m = ?2, 2 4 , 2 6 . . .-components of odd mul- tiplets are nonseparable because they have identical evo- lutions during the echo delay. The same mutual relationship exists form = 21, 1-3, 55. . .-components of odd multiplets, and for m = +1/2, -312, +5/2 , -712, . . .-components on the one hand, for -112, +3/2, -512, +7/2, . . .-components on the other hand of even multip- lets. Such sets of multiplet components, which are mu- tually indistinguishable in the present editing technique, correspond to aliasing multiplet components as far as their evolution during the echo delay is concerned; that is, they obey the same equation of time evolution prior to acquisition. Table 2 gives an overview of the phase cy- cling scheme to be applied to edit specifically either type of multiplet component.
1;!2 Biesemans et al.
Table 2 Multiplet-Component Types, Evolution Functions, Taking T = M4J, and Phase Cycling Schemes to be Applied for Their Specific Editing -
Evolution Receiver function phase cycling Drior to scheme for Multiplet-component type
acquisition specific editing
m = 0, +2, *4, . . . 1 rn=&l,+3,+5, . . . eiN?r 180, 0, 180, 0 rn = +1/2, -312, +5/2, . . . e-(iN?r)/z rn = -1123 +312, -512, . . . e+(iNn)/2
-
0, 0, 0, 0
270, 180, 90, 0 90, 180, 270, 0
RESULTS AND DISCUSSION
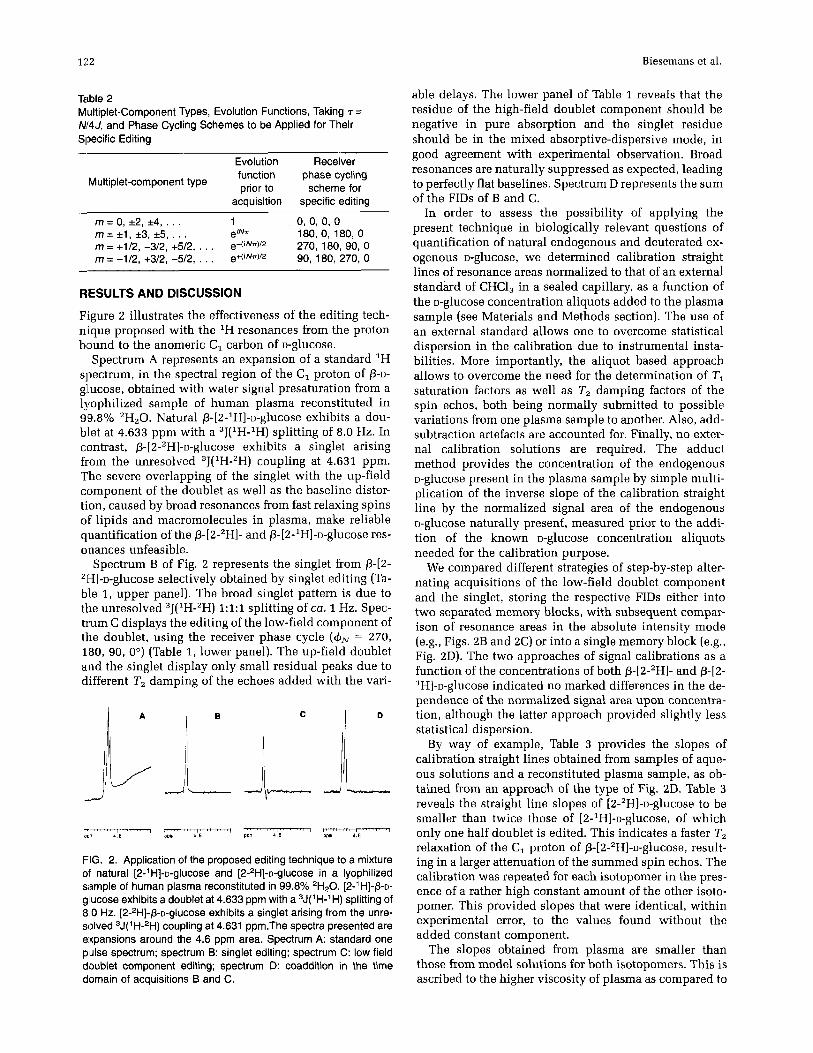
Figure 2 illustrates the effectiveness of the editing tech- nique proposed with the 'H resonances from the proton bound to the anomeric C, carbon of D-glucose.
Spectrum A represents an expansion of a standard 'H spectrum, in the spectral region of the C, proton of p-D- gllucose, obtained with water signal presaturation from a lgophilized sample of human plasma reconstituted in 99.8% *H,O. Natural p-[2-1H]-~-glucose exhibits a dou- blet at 4.633 ppm with a 3J(1H-1H) splitting of 8.0 Hz. In contrast, ~-[2-2H]-u-glucose exhibits a singlet arising from the unresolved ,J('H-'H) coupling at 4.631 ppm. The severe overlapping of the singlet with the up-field component of the doublet as well as the baseline distor- tion, caused by broad resonances from fast relaxing spins of lipids and macromolecules in plasma, make reliable quantification of the P-[2-'H]- and p-[2-1H]-o-glucose res- onances unfeasible.
Spectrum B of Fig. 2 represents the singlet from P-[2- 21-I]-~-glucose selectively obtained by singlet editing (Ta- ble 1, upper panel). The broad singlet pattern is due to the unresolved "(IH-"H) 1:1:1 splitting of ca. 1 Hz. Spec- trum C displays the editing of the low-field component of the doublet, using the receiver phase cycle (& = 270, 180, 90, 0") (Table 1, lower panel). The up-field doublet and the singlet display only small residual peaks due to different T, damping of the echoes added with the vari-
A B
FIG. 2. Application of the proposed editing technique to a mixture of natural [2-l H]-~-glucOSe and [2-zH]-~-glucose in a lyophilized sample of human plasma reconstituted in 99.8% 'H20. [2-' HI-P-D- glucose exhibits a doublet at 4.633 ppm with a ,J('H-'H) splitting of 8 0 Hz. [~-'H]-P-D-~IUCOS~ exhibits a singlet arising from the unre- solved ,J(I H-'H) coupling at 4.631 ppm.The spectra presented are expansions around the 4.6 ppm area. Spectrum A: standard one pulse spectrum; spectrum B: singlet editing; spectrum C: low field doublet component editing; spectrum 0: coaddition in the time domain of acquisitions B and C.
able delays. The lower panel of Table 1 reveals that the residue of the high-field doublet component should be negative in pure absorption and the singlet residue should be in the mixed absorptive-dispersive mode, in good agreement with experimental observation. Broad resonances are naturally suppressed as expected, leading to perfectly flat baselines. Spectrum D represents the sum of the FIDs of B and C.
In order to assess the possibility of applying the present technique in biologically relevant questions of quantification of natural endogenous and deuterated ex- ogenous D-glucose, we determined calibration straight lines of resonance areas normalized to that of an external standard of CHC1, in a sealed capillary, as a function of the D-glucose concentration aliquots added to the plasma sample (see Materials and Methods section). The use of an external standard allows one to overcome statistical dispersion in the calibration due to instrumental insta- bilities. More importantly, the aliquot based approach allows to overcome the need for the determination of TI saturation factors as well as T, damping factors of the spin echos, both being normally submitted to possible variations from one plasma sample to another. Also, add- subtraction artefacts are accounted for. Finally, no exter- nal calibration solutions are required. The adduct method provides the concentration of the endogenous o-glucose present in the plasma sample by simple multi- plication of the inverse slope of the calibration straight line by the normalized signal area of the endogenous D-glucose naturally present, measured prior to the addi- tion of the known u-glucose concentration aliquots needed for the calibration purpose.
We compared different strategies of step-by-step alter- nating acquisitions of the low-field doublet component and the singlet, storing the respective FIDs either into two separated memory blocks, with subsequent compar- ison of resonance areas in the absolute intensity mode (e.g., Figs. 2B and 2C) or into a single memory block (e.g., Fig. 2D). The two approaches of signal calibrations as a function of the concentrations of both p-[2-'H]- and p42- lH]-~-glucose indicated no marked differences in the de- pendence of the normalized signal area upon concentra- tion, although the latter approach provided slightly less statistical dispersion.
By way of example, Table 3 provides the slopes of calibration straight lines obtained from samples of aque- ous solutions and a reconstituted plasma sample, as ob- tained from an approach of the type of Fig. 2D. Table 3 reveals the straight line slopes of [ 2-ZH]-~-glucose to be smaller than twice those of [2-'H]-~-glucose, of which only one half doublet is edited. This indicates a faster T, relaxation of the C, proton of P-[2-*H]-~-glucose, result- ing in a larger attenuation of the summed spin echos. The calibration was repeated for each isotopomer in the pres- ence of a rather high constant amount of the other isoto- pomer. This provided slopes that were identical, within experimental error, to the values found without the added constant component.
The slopes obtained from plasma are smaller than those from model solutions for both isotopomers. This is ascribed to the higher viscosity of plasma as compared to

‘H-Editing of o-Glucose in Biological Samples 123
Table 3 Slopes of Calibration Straight Lines in ml . mg-l with Standard Error and Correlation Coefficient for the C, Proton Resonances of p- [2-2H]-o-Glucose and p- [2-’H]-o-Glucose in 99.8% 2H20) and in Plasma”
Substrate in 99.8% 2H20 in plasmab ~ ~ ~ ~ ~~ ~
[2-2H]-~-Glucose 0.896 k 0.040; 0.996c 0.676 0.026; 0.997
[2-1H]-o-Glucose 0.599 k 0.026; 0.993 0.508 k 0.016; 0.998e 0.855 r 0.01 9; 0.999’
0.585 & 0.021 ; 0.995d
a The integrated areas normalized to that of the external chloroform standard of the edited C, proton singlet of p-[2-2H]-o-glucose and doublet low field component of p-[2-’H]-o-gluCOSe are measured as a function of added D-
glucN3se concentration aliquots; the calibration was achieved with seven concentrations ranging from 0.1 to 1.5 mg/ml.
Lyophilized plasma, reconstituted in 99.8% 2H20, containing 0.61 mg/ml of endogenous natural D-glucose.
Calibration in the presence of 1.5 mg/ml of added [2-’H]-~-glucose. Calibration in the presence of 1.5 mg/ml of added [2-ZH]-~-glUCOSe. Calibration straight line having nonzero intercept at the abscissa because
of the presence of 0.61 mglml of endogenous natural o-glucose. The latter concmtration was determined from the product of the inverse slope of the straight line with the normalized signal area of the endogenous [2-’H]-D- gluclxe naturally present, measured prior to the addition of known D-glucose aliqLiots for calibration purposes.
model solutions resulting in higher T2 damping. This underlines the need for an adduct based calibration with known concentration aliquots of both [2-2H]-~-glucose ancl [2-1H]-o-gl~icose after measurement of their reso- nance area in the sample of interest itself, because the potential variation in plasma viscosity is expected to re- suli in significant T, damping variations. This makes transfers of calibration data from one plasma sample to another inappropriate.
As an example, the good linear correlation obtained in experiment e of Table 3 illustrates the effectiveness of this method.
COINCLUSION
The ‘H editing technique clearly enables quantitative de- terminations of exogenous [2-’H]-o-glucose in the pres- ence of endogenous natural [2-1H]-~-glucose. With appro- priate stock solutions at hand, the adduct calibration of the plasma sample is not too time consuming (ca. 2 h for preparing and acquiring spectra for a 7-aliquot calibra- tion).
The oral ingestion of about 20 g of P-[2-2H]-o-glucose by a human individual would be required for a concen- tration of about 0.8 mg/ml of the deuterated substrate to be observable with reasonable accuracy in the presence of natural endogenous P-D-glucose. This makes serial ex- periments in humans, for instance in applications of sports medicine or diabetology, economically prohibi-
tive. However, the approach is of potential interest in several biological investigations conducted in vitro. In this respect, the following two applications are illustra- tive. First, in the study of D-glucose cycling in the reac- tions catalyzed by hexokinase isoenzymes and glucose- 6-phosphatase, e.g., in hepatocytes, the generation of [2- ‘H]-~-glucose from natural D-glucose could be monitored in cells incubated in the presence of ’H,O. This approach offers the advantage over current tritium based radioiso- topic techniques (7, 8) to avoid the sophisticated proce- dures required to identify and quantify [2-3H]-~-glucose in samples obtained after incubation of cells in the pres- ence of unlabelled D-glucose and 3H20. Moreover, in this application, the underestimation of futile cycling attrib- utable to isotopic discrimination in the reaction by phos- phoglucoisomerase will be less marked with deuterated than with tritiated water (9). Second, by incubating eryth- rocytes in a small volume of medium containing [2-’H]- D-glucose, it would be possible to establish the time course for the exchange between influent [2-2H]-~-glu- cose and effluent natural D-glucose, which is present in- side the erythrocytes at the onset of incubation. Our ap- proach would be especially relevant to the erythrocytes of adult nonprimate animals, in which the isotopic equil- ibration of o-glucose across the plasma membrane repre- sents a delayed phenomenon and, hence, coincides with the time-related generation of large amounts of its metab- olites (10).
ACKNOWLEDGMENTS
We are indebted to Dr. D. Halliday, CRC Nutrition Research Unit, Harrow, England, for providing samples of [2- ’H]-~-glucose.
REFERENCES 1. 2.
3.
4. 5.
6. 7.
8.
9.
10.
T. Jue, J . Magn. Reson. 73, 524 (1987). R. Gruetter, E. J. Novotny, S. D. Boulware, D. L. Rothman, G. I. Shulman, G. F. Mason, R. G. Shulman, W. V. Tamborlane, Proc. Nafl . Acad. Sci. U S A 89, 1109 (1992). R. Gruetter, D. L. Rothman, E. J. Novotny, G. I. Shulman, J. W. Prichard, R. G. Shulman, Magn. Reson. Med. 27, 183 (1992). T. Jue, J. Magn. Reson. 76, 321 (1988). T. Jue, Y. Chung, R. G. Shulman, J. Magn. Reson. 76, 178 (1988). R. Freeman, H. D. W. Hill, J. Chem. Phys. 54, 301 (1971). A. Khan, V. Chadramouli, C.-G. Ostenson, B. Ahren, W. C. Schuman, H. Low, B. R. Landau, S. Efendic, I. B i d . Chem. 264, 9732 (1989). V. Liemans, A. Sener, W. J. Malaise, Biochem. Int. 24, 391 (1991). W. J. Malaise , F. Malaise-Lagae, V. Liemans, R. Ottinger, R. Willem, Mol. Cell. Biochem. 93, 153 (1990). J. Elbrink, I. Bihler, Science 188, 1177 (1975).





























