Embed Size (px)
Citation preview

news and views
Ubiquitin biology: an old dog learns an old trick
Cecile M. Pickart
Regulated protein degradation in eukaryotes occurs principally through covalent tagging of substrates with ubiquitin, thereby targeting them for destruction by 26S proteasomes. Classical allostery has now been added to the repertoire of mechanisms that can modulate ubiquitin tagging, allowing feed-forward regulation to be achieved through targeted protein destruction.
ukaryotic organisms use diverse mech-anisms to regulate metabolic path-ways. One common mechanism is the
allosteric activation or inhibition of rate-limiting enzymes1, and another involves thedegradation of enzymes2 or dedicated regu-latory proteins3 by the ubiquitin–proteas-ome system. In a recent issue of Nature,Turner et al.4 have shown that these regula-tory strategies can be functionally linked.
The majority of short-lived proteins aretargeted for destruction through conjuga-tion with ubiquitin, which provides a recog-nition signal for 26S proteasomes5. Thisproteolytic system is known for its ability todegrade diverse substrates with high specifi-city, properties that are brought about by anextensive array of enzymes known as ubiqui-tin–protein (E3) ligases. E3 ligases recognizetheir substrate(s) through specific sequencemotifs, or degrons; because each degron hasits own E3, the destruction of many differentsubstrates can be independently regulated.Several mechanisms for regulating degrada-tion are now known, all of which affect theubiquitin-tagging step2,4,5. Recognition caninvolve altering the properties of the degronitself; for example, this motif may requirephosphorylation to be recognized by the E3,or it may be masked as a result of interactionwith another protein. Alternatively, covalentmodification of the E3 may regulate its abil-ity to recognize the degron. In still othercases, regulation can involve changes inexpression of the E3. Until now, however,there have been no examples of regulationbeing effected through the binding of a smallmolecule to an E3.
The paradigm for specificity in the ubiq-uitin system was developed on the basis ofthe properties of an E3 known as Ubr1 (orE3α), as revealed through the work of theVarshavsky and Hershko laboratories. Ubr1is a large protein (relative molecular mass225,000) with several distinct functionaldomains4. It has two sites that bind to sub-strates through their amino termini — atype-1 site that recognises basic residuessuch as arginine, and a type-2 site that bindsto bulky hydrophobic residues such as leu-cine (Fig. 1). Although the positions of these
sites within the Ubr1 polypeptide are yet tobe discovered, both sites have been exten-sively characterized through competitionstudies6,7. Ubr1 is the E3 for an arm of theubiquitin system known as the N-end rulepathway, the name reflecting the fact thatUbr1 binds to model substrates throughtheir N-terminal residues6–8. In this work,much of it published over a decade ago,dipeptides played an important role by vir-tue of their ability to competitively inhibit
Ubr1, provided that the peptide and thesubstrate shared the same type of N-termi-nal residue6,7 (Fig. 1). Now, dipeptides havebeen shown to allosterically activate Ubr1-dependent degradation of a substrate that isrecognized in a distinct manner4.
The binding of type-1 substrates isinhibited by dipeptides with basic N termini(such as Arg-Ala, but not Ala-Arg), whereasthe binding of type-2 substrates is blockedby dipeptides with hydrophobic N termini
E
Figure 1 Modulation of Ubr1 activity by dipeptides. The binding to Ubr1 of a substrate with a type-1 N-terminal residue is blocked by a type-1 dipeptide (a), whereas the binding of a type-2 substrate is blocked by a type-2 dipeptide (b). Cup9 binds to Ubr1 at a distinct site and is constitutively ubiquitinated (c); this reaction is stimulated by the binding of type-1 or type-2 dipeptides to their respective sites (d). Large ovals represent substrate molecules; yellow circles represent ubiquitins. e, Mechanism of feed-forward regulation by dipeptides.
Cup9 2 1
a
b
c
d
e
Ubr1
Ubr1 Cup9
Cup9 2 1
Cup9 2 1
Cup9 2 1
Cup9 2 1
Ptr2 Dipeptide import
Type 1: Arg, Lys, His
Type 2: Leu, Ile, Phe, Tyr, Trp
© 2000 Macmillan Magazines LtdNATURE CELL BIOLOGY | VOL 2 | AUGUST 2000 | www.nature.com/ncb E139
news and views
(such as Leu-Ala, but not Ala-Leu). Ubr1also contains a binding site for a conjugat-ing enzyme partner, and a RING-fingermotif that is required for the chemistry ofthe conjugation reaction, but is dispensiblefor the binding of type-1 and type-2 sub-strates. The properties of Ubr1, as eluci-dated using model substrates, are strikinglyconserved in evolution7. However, formuch of its scientific life, Ubr1 was anenzyme in search of a biological substrate.
A breakthrough came with the findingthat mutations in Ubr1 inhibit dipeptideuptake in the yeast Saccharomycescerevisiae9. A yeast transcription factor
called Cup9 proved to be the relevantsubstrate3. Regulatory molecules such astranscription factors are frequently sub-strates of the ubiquitin system, for reasonsthat are easily understood — if a proteinturns over rapidly, its steady-state level willbe extremely sensitive to changes in therates of its synthesis and degradation. Cup9represses the import of peptides by bindingto sequences upstream of the PTR2 gene,which encodes a di/tripeptide transporter.Robust peptide import requires that Cup9is maintained at a low level through Ubr1-dependent ubiquitination, which leads tothe turnover of Cup9 by proteasomes (Fig.
1e). However, the recognition of Cup9 byUbr1 involves a noncanonical internaldegron3, rather than the Cup9 N terminus.
Turner and colleagues have now linkedthese apparently disparate substrate-bind-ing properties of Ubr1 in an unexpectedand elegant way. They have shown that theturnover of Cup9 in yeast cells is stimulatedby dipeptides with type-1 or type-2 N ter-mini, but not by control dipeptides withstabilizing N termini. This N-terminal spe-cificity mimics what would be expected ifdipeptides bind to the type-1 and type-2‘substrate’ sites of Ubr1 and allostericallyactivate the ubiquitination of Cup9 that is
Growth and polarity: the case for Scribble
The epithelial membranes of any organism consist of cells that have a regular columnar shape and defined apical–basolateral polarity. During carcinogenesis, epithelial cells lose these characteristics, as well as control of cell proliferation, and form disorganized cellular structures that have the potential to develop into metastases. Drosophila genetics has always been used to study epithelial morphogenesis and cell proliferation. However, very few genes have been identified that seem ultimately to control both.
Recently, Bilder and Perrimon (Nature 403, 676–680, 2000) reported the isolation of Scribble (Scrib), a member of the LAP (Leucine-rich repeat and PDZ domain) family of proteins (Nature Cell Biol. 2, E114, 2000). Scrib is required to maintain apical–basal polarity, as are the two further LAP proteins recently identified by Borg et al. (Nature Cell Biol. 2, 407–414, 2000) and Legouis et al. (Nature Cell Biol. 2, 415–422, 2000), although their precise role in this process is unknown.
Bilder et al. (Science 289, 113–116, 2000) went on to investigate the mechanism of Scrib’s activity. In epithelial cells Scrib co-localizes to the septate junction (the Drosophila homologue of the vertebrate tight junction) with Discs-large (Dlg). Dlg is a PDZ-domain protein that functions in cell polarity and as a tumour suppressor. The expression of both Scrib and Dlg overlaps with that of lethal giants larvae (Lgl), another known tumour suppressor. Bilder and colleagues conducted a screen to identify other genes that, like scrib, are required to maintain the polarity of follicle cells in the Drosophila ovary, and identified a new allele of lgl, which showed a similar phenotype in follicle cells to that of Scrib mutants.
The authors then returned to embryonic epithelia and noticed that in embyros with mutations in lgl or dlg, apical–basolateral polarity was altered, in the same way as in scrib mutants. Furthermore, they examined another Drosophila epithelium, the wing disc, and found that Scrib-mutant cells overproliferated and
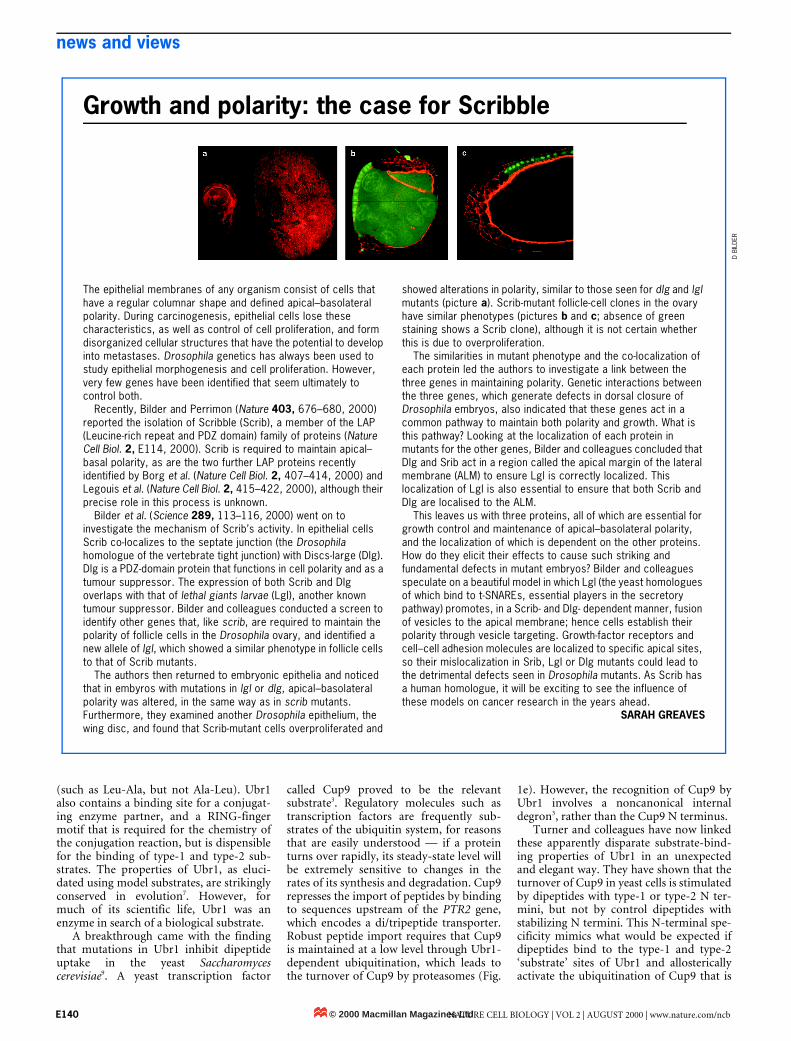
showed alterations in polarity, similar to those seen for dlg and lgl mutants (picture a). Scrib-mutant follicle-cell clones in the ovary have similar phenotypes (pictures b and c; absence of green staining shows a Scrib clone), although it is not certain whether this is due to overproliferation.
The similarities in mutant phenotype and the co-localization of each protein led the authors to investigate a link between the three genes in maintaining polarity. Genetic interactions between the three genes, which generate defects in dorsal closure of Drosophila embryos, also indicated that these genes act in a common pathway to maintain both polarity and growth. What is this pathway? Looking at the localization of each protein in mutants for the other genes, Bilder and colleagues concluded that Dlg and Srib act in a region called the apical margin of the lateral membrane (ALM) to ensure Lgl is correctly localized. This localization of Lgl is also essential to ensure that both Scrib and Dlg are localised to the ALM.
This leaves us with three proteins, all of which are essential for growth control and maintenance of apical–basolateral polarity, and the localization of which is dependent on the other proteins. How do they elicit their effects to cause such striking and fundamental defects in mutant embryos? Bilder and colleagues speculate on a beautiful model in which Lgl (the yeast homologues of which bind to t-SNAREs, essential players in the secretory pathway) promotes, in a Scrib- and Dlg- dependent manner, fusion of vesicles to the apical membrane; hence cells establish their polarity through vesicle targeting. Growth-factor receptors and cell–cell adhesion molecules are localized to specific apical sites, so their mislocalization in Srib, Lgl or Dlg mutants could lead to the detrimental defects seen in Drosophila mutants. As Scrib has a human homologue, it will be exciting to see the influence of these models on cancer research in the years ahead.
SARAH GREAVES
D BI
LDER
© 2000 Macmillan Magazines LtdE140 NATURE CELL BIOLOGY | VOL 2 | AUGUST 2000 | www.nature.com/ncb

news and views
bound at a distinct site. Because it is difficultto measure ubiquitination in vivo, theauthors confirmed this model by showingthat dipeptides with type-1 or type-2 N ter-mini, but not control peptides, stimulatethe Ubr1-dependent ubiquitination ofCup9 in an in vitro reaction involving exclu-sively purified recombinant proteins. Thefaithful recapitulation of the in vivo specifi-city in this in vitro experiment indicates thatthe intact dipeptide, and not a secondarymetabolite, is the actual regulatory entity.
Is the ability of dipeptides to promotethe ubiquitination and turnover of Cup9biologically relevant? The answer is emphat-ically yes. The dipeptides that stimulateUbr1 also induce the expression of PTR2messenger RNA, indicating that the acceler-ation of Cup9 turnover may be sufficient toincrease the number of transporters in themembrane4. Strikingly, although neither alow concentration of Leu-Ala nor a highconcentration of Ala-Leu could individuallysupport the growth of a leucine auxotroph,combining the two facilitated growth, indi-cating that Leu-Ala allosterically stimulatesthe uptake of Ala-Leu. Together, these find-ings show that allosteric activation of Ubr1is an efficient mechanism for feed-forwardregulation of peptide import. The conservedproperties of Ubr1 (ref. 7) and the presencein mammals of Ptr2-related transporters9
indicate that a similar allosteric mechanismcould potentially regulate dipeptide uptakein metazoans.
Important questions remain. First andforemost, exactly how do dipeptides stimu-
late Ubr1? Modulation of substrate bindingis a time-honoured allosteric mechanism1,but the Ubr1 reaction offers further possi-bilities. Like all E3s, Ubr1 must undergomany catalytic cycles before it generates auseful product; this is because proteasomesrecognize most efficiently those substratesthat have been tagged with many ubiquitinmolecules in the form of a polymeric chain.As an alternative to stimulating substratebinding, dipeptides could enhance the rateor processivity of chain elongation onCup9. In addition, the assumption thatdipeptides and substrates that share similarN termini bind to the same site on Ubr1should be tested more rigorously, now thatit is clear that peptides can act allosterically.If this assumption is correct, then it is pos-sible that different types of Ubr1 substratescould allosterically regulate each other’sdegradation. Finally, what is the biologicalrationale for restricting the feed-forwardregulation to dipeptides with type-1 ortype-2 N termini? One possibility, that suchpeptides are prioritized for import, seems tobe excluded by the rather broad specificityof the Ptr2 transporter9.
Ubr1 is first enzyme in the ubiquitin sys-tem that is known to be regulated in thismanner, but it is unlikely to be the last. Oneattractive idea is that feedback inhibitionoccurs through allosterically induced deg-radation of a rate-limiting metabolicenzyme. Such an effect could involve theactivation of an E3 (as in this case). Otherpossibilities include modulating the prop-erties of the substrate’s degron, altering
subunit interactions in a multisubunit E3,and modulating the interaction of an E3with upstream conjugating enzymes.Hampton et al.2 have shown that the ubiq-uitin-mediated turnover of yeast hydroxyl-methylglutaryl-CoA reductase is stimulatedby a downstream metabolite of the meval-onate pathway, perhaps through an allos-teric effect on substrate (reductase)properties. Allosteric modulation of theubiquitin–proteasome pathway could alsohave a function in the glucose-inducedturnover of fructose bisphosphatase10.Allostery should prove to be a powerfuladdition to the regulatory arsenal of theubiquitin system, and in the future maypresent attractive targets for drug design.hCecile M. Pickart is in the Department of Biochemistry and Molecular Biology, Johns Hopkins University School of Public Health, 615 North Wolfe Street, Baltimore, Maryland 21205, USA.e-mail: [email protected]
1. Perutz, M. F. Q. Rev. Biophys. 22, 139–237 (1989).
2. Gardner, R. G. & Hampton, R. Y. J. Biol. Chem. 274, 31671–31678 (1999).
3. Byrd, C., Turner, G. C. & Varshavsky, A. EMBO J. 17, 269–277
(1998).4. Turner, G. C., Du, F. & Varshavsky, A. Nature 405, 579–583
(2000).
5. Hershko, A. & Ciechanover, A. Annu. Rev. Biochem. 67, 425–480 (1998).
6. Reiss, Y., Kaim, D. & Hershko, A. J. Biol. Chem. 263, 2693–2698
(1988).7. Gonda, D. K. et al. J. Biol. Chem. 264, 16700–16712 (1989).
8. Bachmair, A., Finley, D. & Varshavsky, A. Science 234, 179–186
(1986).9. Alagramam, K., Naider, F. & Becker, J. M. Mol. Microbiol. 15,
225–234 (1995).
10. Schüle, T., Rose, M., Entian, K-D., Thumm, M. & Wolf, D. H.
EMBO J. 19, 2161–2167 (2000).
LAP proteins: what’s up with epithelia?
Peter J. Bryant and Adrien Huwe
How cells maintain their overall shape and size, and the related question of how proteins and other molecules reach and stay at their specific subcellular locations, are among the most difficult and exciting problems in cell biology. Three recent studies have made a significant contribution to this area by identifying new proteins, called LAP proteins, that have critical functions in maintaining the shape and apical-basal polarity of epithelial cells.
AP (leucine-rich repeats and PDZdomains) proteins contain either 1 or 4copies of the PDZ domain (named for
its occurrence in the PSD-95, Discs-Largeand ZO-1 proteins) at their carboxy terminiand 16 leucine-rich repeats (LRRs) at theiramino termini (Fig. 1). Both types of domainprobably function mainly in binding to otherproteins. Recently, Borg et al.1 and Legouis et
al.2, both in last month’s issue of Nature CellBiology, together with Bilder and Perrimon3,have reported the discovery of new LAP pro-teins that control cell size and shape as well assubcellular protein localization.
The first member of the LAP family,Densin-180, was identified, not in epithelialcells, but in the postsynaptic densities of therat brain, where it may be involved in the reg-
L
© 2000 Macmillan Magazines LtdNATURE CELL BIOLOGY | VOL 2 | AUGUST 2000 | www.nature.com/ncb
ulation of calcium/calmodulin-dependentprotein kinase II4. The newly discovered LAPproteins have been found in the epithelialcells of flies, worms, and humans, where theyseem to be critical for cell structure as well asfor the organization and regulation of the sig-nal-transduction pathways that controlgrowth and differentiation.
Epithelial cells are distinctly polarized —they have apical and basolateral domains,each of which have different membrane,extracellular-matrix and sub-membranecytoskeletal components (Fig. 2). Thesedomains are separated by specialized junc-tional complexes, which are also responsiblefor the lateral adhesion within the sheet andfor the sealing of the barrier that restrictsdiffusion between fluids that are apical andbasal to the layer. In arthropods the junc-tional complex includes adherens junctions,which are situated in the most apical posi-tion on the lateral cell membrane, and sep-tate junctions, which are immediately basalto them. In vertebrates the septate junctionsare replaced by tight junctions, which areapical rather than basal to the adherensjunctions. Some of the protein constituentsof these junctions are well known5 (Fig. 1).
E141


![Integrating the Healthcare Enterprise€¦ · Document Source Document ConsumerOn Entry [ITI Document Registry Document Repository Provide&Register Document Set – b [ITI-41] →](https://img.pdfslide.net/doc/110x75/5f08a1eb7e708231d422f7c5/integrating-the-healthcare-enterprise-document-source-document-consumeron-entry.jpg)


























