Embed Size (px)
Citation preview

NATURE BIOTECHNOLOGY VOL 19 FEBRUARY 2001 http://biotech.nature.com 153
To fully benefit from the remarkably comprehensive new informa-tion about the Drosophila and C. elegans genomes, it would be atremendous advantage to be able to isolate large quantities ofmutant embryos and embryo populations at defined developmentalstages. The combination of nearly a century of mutant collectionand analysis, in concert with new technologies such as DNAmicroarrays, provide an opportunity to understand genetic net-works, reveal regulatory pathways, and detect responses to environ-mental perturbations. Of the estimated 13,601 Drosophila genes,about 3,600–4,000 are expected to be essential for survival.Thousands of existing lethal mutations are kept as stocks of het-erozygous flies. One quarter of the embryos produced by such apopulation will be homozygous for the mutation of interest1. Thethreefold excess of embryos carrying the wild-type gene, often on abalancer chromosome, creates problems for many types of biochem-ical and molecular experiments (Fig. 1A).
To overcome this problem, we have designed, built, and tested aninstrument that sorts embryos of a particular genotype according totheir levels of fluorescence. The machine was designed for use withDrosophila embryos, but the same principles can be applied to sortingembryos from other species, such as C. elegans, that have been engi-neered to contain fluorescent cells. We initially developed the machinefor Drosophila because of the available genetic tools and the recent nearcompletion of the whole-genome sequencing project.
To ensure that lethal mutations of interest are inherited by thenext generation, Drosophila geneticists use balancer chromosomes.These chromosomes are multiply inverted to minimize recombina-tion events, and often carry lethal mutations so that homozygousbalancer embryos die. The homozygous mutant embryos also die, sothe only embryos that survive into adulthood are the heterozygousmutant/balancer embryos (Fig. 1A). Adults containing a balancerchromosome produce 25% homozygous embryos, the embryos thatmost people would like to study. A number of fly strains have a geneencoding green fluorescent protein (GFP) inserted into the balancerchromosome2. It is therefore possible to distinguish, in living
embryos, homozygous mutant embryos that lack GFP expressionfrom heterozygous and homozygous-balancer embryos that haveGFP (Fig. 1A). This paper describes a machine that can sortDrosophila embryos based on the presence or absence of GFP. Themachine can separate the 25% homozygous mutant embryos fromthe 75% that contain the GFP-balancer.
Cell sorting is similar in basic principles to embryo sorting.Fluorescence-activated cell sorting (FACS) is widely used to isolatesubpopulations of cells based on antigen display, nucleic acid content,and gene expression3,4. During FACS analysis, cells are observed inlaminar flow, which is then dispersed into droplets of solution suchthat only one cell is contained within a droplet5. The cells are sorted byelectrostatic deflection of the droplet, displacing it away from the lam-inar flow of solution. Whole embryos are too large to be sorted usingconventional FACS technology. Drosophila embryos are 450–500 µmlong and 150–200 µm wide6 and C. elegans first-instar larvae (L1) are250 µm long7; a typical mammalian cell has a diameter of 10–20 µm.
ResultsMechanics of the embryo sorter. An overview of the instrument(Fig. 1B) outlines how it operates. Collected embryos are main-tained suspended in buffer in a chamber using a magnetic stirrer. Aperistaltic pump continuously moves solution through the chamberinto an optical cuvette (Fig. 2A). Solution enters and exits the cham-ber at a fixed rate (∼ 6 ml/min). Some of the suspended embryos ran-domly enter the flow of liquid that is exiting the chamber. This flowcarries the embryos through the optical cuvette, which consists of a0.4 mm square glass tube embedded in a holder containing immer-sion oil. The diameter of the tube restricts the embryos to two possi-ble orientations (Fig. 2B).
Embryos passing through the cuvette are illuminated by an argonlaser (488 nm). Light emitted from an embryo is detected perpen-dicular to the laser excitation light (Fig. 2C). The emitted light pass-es through an aspheric condenser lens that collects and collimatesthe light and then through a plano-convex focusing lens followed by
Automated sorting of live transgenic embryos
Eileen E.M. Furlong, David Profitt, and Matthew P. Scott*
Departments of Developmental Biology and Genetics, Howard Hughes Medical Institute, Beckman Center B300, 279 Campus Drive, Stanford University School of Medicine, Stanford, CA 94305-5329. *Corresponding author ([email protected]).
Received 7 August 2000; accepted 10 November 2000
The vast selection of Drosophila mutants is an extraordinary resource for exploring molecular eventsunderlying development and disease. We have designed and constructed an instrument that automati-cally separates Drosophila embryos of one genotype from a larger population of embryos, based on a flu-orescent protein marker. This instrument can also sort embryos from other species, such asCaenorhabditis elegans. The machine sorts 15 living Drosophila embryos per second with more than 99%accuracy. Sorting living embryos will solve longstanding problems, including (1) the need for large quan-tities of RNA from homozygous mutant embryos to use in DNA microarray or gene-chip experiments,(2) the need for large amounts of protein extract from homozygous mutant embryos for biochemicalstudies, for example to determine whether a multiprotein complex forms or localizes correctly in vivowhen one component is missing, and (3) the need for rapid genetic screening for gene expressionchanges in living embryos using a fluorescent protein reporter.
Keywords: Drosophila, embryo, development, green fluorescent protein
RESEARCH ARTICLES©
2001
Nat
ure
Pu
blis
hin
g G
rou
p
htt
p:/
/bio
tech
.nat
ure
.co
m© 2001 Nature Publishing Group http://biotech.nature.com

two dichroic mirrors (optical cutoff at 498 nm and 520 nm) (Fig. 2D). Light is detected at three points along the light path: (1) Adiode, measuring light that has passed through the lenses, detects allinterruptions of the light. The diode signal, marking the position ofeach passing embryo, is analyzed for amplitude peaks by a dedicatedmicrocontroller. (2) A photomultiplier tube (PMT-1) detects thelight between the 498 nm and 520 nm dichroic mirrors. This lightpasses through a narrow filter (510 ± 5 nm) before reaching thePMT-1 and therefore detects mainly the GFP signal. (3) PMT-2detects light that passes through the 520 nm dichroic mirror. Thislight passes through a 515–60 nm barrier filter before reaching thePMT-2, which detects the background fluorescence or autofluores-cence emanating from the yolk of the embryo. When the diodedetects an embryo, the microcontroller measures the voltage detect-ed for the GFP (VGFP), autofluorescence (Vauto), and threshold signal(Vth) from the PMTs. The threshold signal is chosen by the user andis the level of GFP above or below which the machine should save anembryo. The computer draws peaks for each embryo, one for theGFP level and a second for autofluorescence. To obtain GFP-containing embryos, the microcontroller evaluates the conditionVGFP – Vauto > Vth. If the condition is true, the signals “New embryodetected” and “Save” are generated. If the condition is false, the sig-nal “New” is the only output. Conversely, to obtain embryos that donot contain any GFP, a condition VGFP – Vauto > Vth “false” would gen-erate a “New embryo detected” and a “Save” output. The “New” and“Save” outputs are sent to electromagnets that control the switchthat directs embryos into one tube or another.
The switch is composed of a waste tube and a save tube, separat-ed by a very thin central wall (Fig. 2G, H). The embryo and solutionflow is always aligned to the waste tube, which is the default position.The two tubes are attached to a rare-earth neodymium super-magnet suspended between two electromagnets. Electric currentapplied in one direction to the electromagnets exerts force on thesuspended magnet in one direction. Reversing the current producesa force in the opposite direction. The magnet moves the switch to theleft or right, that is, to the “Save” or “Waste” position.
The embryo–buffer suspension passes through the optical cuvetteat a rate of ∼ 6 ml/min. Buffer containing the embryos drips from theend of the optical cuvette into the switching mechanism. To facilitaterapid and accurate switching, the drop containing the embryo entersa high-speed fluid stream from a second pump (Fig. 2F). Themechanical switch is aligned underneath the high-speed stream ofliquid to collect the embryos (below the blue arrow in Fig. 2C).
The timing of the embryo-sorting switch is controlled by a sec-ond dedicated microcontroller that receives the Save and New out-puts from the optics microcontroller. For any fluid flow rate there isa predictable time from embryo detection to the arrival of theembryo at the sorting switch. For example, with a fluid flow of ∼ 6 ml/min there is an 11–12 ms delay from the time of embryodetection to the time when the embryo reaches the switch. If a Wasteembryo is ahead of a Save embryo, sufficient time must be allowedfor the Waste embryo to pass through the switch before the switch
154 NATURE BIOTECHNOLOGY VOL 19 FEBRUARY 2001 http://biotech.nature.com
RESEARCH ARTICLES
Figure 1. Sorting embryos based on their expression of GFP. (A) Drosophila lethal mutations are maintained over a balancerchromosome. Balancer chromosomes are multiply invertedchromosomes, to minimize recombination events, that often carrylethal mutations. They allow the stable maintenance of recessive lethalmutations, because the only embryos that can survive to adulthood arethe heterozygous embryos (mutation/balancer). However, for anyembryo collection from these parents, only 25% of the embryos containthe homozygous mutation of interest. Using a GFP marker gene on thebalancer chromosome, it is possible to distinguish, in living embryos,between this minority population of mutant embryos and theirbalancer-containing siblings. (B) Schematic diagram of the embryosorter outlines how the machine operates. Suspended embryosrandomly enter the fluid flow through the optical cuvette, where theyare illuminated by an argon laser (488 nm). The emitted light is detectedat three points along the light path: (1) a diode, (2) photomultiplier tube1 (PMT-1), which detects the light between the 510 ± 5 nm (GFP), and (3) PMT-2, which detects all the light that passes through the 520 nmdichroic mirror (autofluorescence). If the GFP peak is above or below adefined threshold, a signal is sent to the mechanical switch to direct theembryo flow to either the save or waste tube.
Figure 2. Optical system and the mechanical switch used in theembryo sorter. (A) Embryos pass through an optical cuvettecomposed of a 0.4 mm diameter square tube, enclosed in a squarechamber. (B) Schematic diagram of the 0.4 mm square tube. Thediameter of the tube restricts the number of possible orientations ofthe embryos. (C) The optical cuvette is aligned with the opticschamber, ensuring appropriate alignment of the viewing windows soemitted light from the embryo passes through the focusing lens to thePMTs. (D) Side view of the inside of the optics chamber. The emittedlight (red arrow) passes through this chamber and is detected by thediode and the PMTs. (E) End-on view of the optics cuvette shown in(A). The embryos exit the central hole in a droplet of solution beforereaching the switch. (F) The end of the cuvette shown in (E) attachesto a metal fluid ring. The fluid ring has an inflow of solution at highspeed; the drop of solution containing the embryo enters a sheath ofliquid and exits through the central hole in a stream of liquid. (G) Viewof the embryo switch from above. This switch is aligned underneaththe cuvette-fluid ring, underneath the blue arrow in (C). (H) Side viewof the embryo switch. The default position is for the switch to bealigned to the waste tube; the switch moves to the right to collect aSave embryo. (Yellow arrows indicate direction of fluid and embryoflow. Red arrows indicate direction of the light path.)
A
B
©20
01 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://b
iote
ch.n
atu
re.c
om
© 2001 Nature Publishing Group http://biotech.nature.com
NATURE BIOTECHNOLOGY VOL 19 FEBRUARY 2001 http://biotech.nature.com 155
moves to collect the Save embryo. Conversely, if a Save embryo isahead of a Waste embryo, the switch must return to the Waste posi-tion before the second embryo arrives. Timing is accomplished by asoftware algorithm that effectively maps the position of the embryoin real time as it travels from the detector to the sorting switch.
A detailed description of how to build the sorter is available assupplementary information (in the Web Extras page of NatureBiotechnology Online).
Viability of sorted embryos. We determined what effect sortinghas on the viability of the embryos. Three independent embryo col-lections from three different stages in development were analyzed.The fly strain was wild type (Canton S). Each embryo collection waspassed through the embryo sorter for 1 h. The embryos were count-ed and allowed to develop to adulthood at 25°C. After 14 days, >93%of the embryos had reached adulthood (Fig. 3). No significant dif-ference was observed between sorted embryos and those that wereheld in sorting buffer for 1 h or just dechorionated. Any deleteriouseffects of sorting are evidently due to incubating the embryos inbuffer or to dechorionation, rather than to the machine itself.
Sorting speed for transgenic GFP-bearing embryos. The optimalspeed of embryo sorting is determined both by the machine’s theo-retical sorting speed and by the density of the embryos that will besorted. The mechanical switch in the embryo sorter can open andclose in 10 ms, giving a theoretical sorting speed of approximately100 embryos/s. However, operating the machine at this speed wouldgreatly reduce the efficiency of sorting. An adequate time intervalmust be preserved between Waste and Save embryos in order toensure that a Waste embryo has passed through the switch beforeflipping the switch to collect a Save embryo. The instrument is set sothat if a Waste embryo is too close to a Save embryo (that is, within10 ms), both embryos are sent to the waste tube. A lower embryodensity results in greater spacing between embryos, therefore reduc-ing the probability of two embryos being within 10 ms of each other.Similar logic applies to FACS sorting.
Empirically, the optimal sorting rate is 15 embryos/s. To maintain aconstant embryo density, the embryo sorter uses a two-chamber system: a high-density embryo chamber, and a low-densityembryo chamber. Freshly collected embryos are added to the high-density chamber. The computer monitors the total number of detectedembryos, the number of saved embryos, and the rate of embryo flow
per second. Normally embryos flow from the low-density chamber to the cuvette. If the rate of embryo passage fallsbelow a defined threshold (i.e., 15 embryos/s) the computer sends asignal to a fluid valve. The valve redirects the fluid flow to pass throughthe high-density chamber into the low-density chamber and then tothe optical cuvette. This moves embryos from the high-density cham-ber to the low-density chamber. Maintaining the embryo density inthis way permits the embryos to be efficiently sorted at a constant rate.
Using the optimal sorting speed, we have routinely separated13,000 homozygous embryos from a population of ∼ 39,000embryos in 1 h.
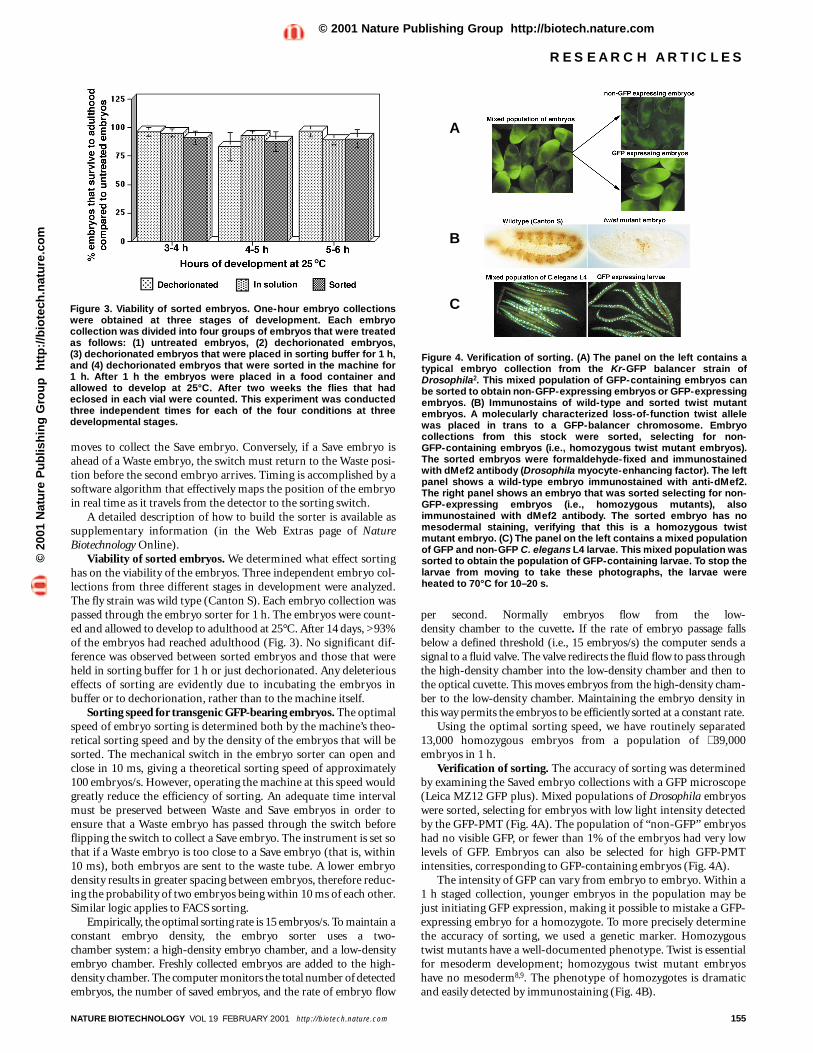
Verification of sorting. The accuracy of sorting was determinedby examining the Saved embryo collections with a GFP microscope(Leica MZ12 GFP plus). Mixed populations of Drosophila embryoswere sorted, selecting for embryos with low light intensity detectedby the GFP-PMT (Fig. 4A). The population of “non-GFP” embryoshad no visible GFP, or fewer than 1% of the embryos had very lowlevels of GFP. Embryos can also be selected for high GFP-PMTintensities, corresponding to GFP-containing embryos (Fig. 4A).
The intensity of GFP can vary from embryo to embryo. Within a1 h staged collection, younger embryos in the population may bejust initiating GFP expression, making it possible to mistake a GFP-expressing embryo for a homozygote. To more precisely determinethe accuracy of sorting, we used a genetic marker. Homozygoustwist mutants have a well-documented phenotype. Twist is essentialfor mesoderm development; homozygous twist mutant embryoshave no mesoderm8,9. The phenotype of homozygotes is dramaticand easily detected by immunostaining (Fig. 4B).
RESEARCH ARTICLES
Figure 3. Viability of sorted embryos. One-hour embryo collectionswere obtained at three stages of development. Each embryocollection was divided into four groups of embryos that were treatedas follows: (1) untreated embryos, (2) dechorionated embryos, (3) dechorionated embryos that were placed in sorting buffer for 1 h,and (4) dechorionated embryos that were sorted in the machine for 1 h. After 1 h the embryos were placed in a food container andallowed to develop at 25°C. After two weeks the flies that hadeclosed in each vial were counted. This experiment was conductedthree independent times for each of the four conditions at threedevelopmental stages.
Figure 4. Verification of sorting. (A) The panel on the left contains atypical embryo collection from the Kr-GFP balancer strain ofDrosophila2. This mixed population of GFP-containing embryos canbe sorted to obtain non-GFP-expressing embryos or GFP-expressingembryos. (B) Immunostains of wild-type and sorted twist mutantembryos. A molecularly characterized loss-of-function twist allelewas placed in trans to a GFP-balancer chromosome. Embryocollections from this stock were sorted, selecting for non-GFP-containing embryos (i.e., homozygous twist mutant embryos).The sorted embryos were formaldehyde-fixed and immunostainedwith dMef2 antibody (Drosophila myocyte-enhancing factor). The leftpanel shows a wild-type embryo immunostained with anti-dMef2.The right panel shows an embryo that was sorted selecting for non-GFP-expressing embryos (i.e., homozygous mutants), alsoimmunostained with dMef2 antibody. The sorted embryo has nomesodermal staining, verifying that this is a homozygous twistmutant embryo. (C) The panel on the left contains a mixed populationof GFP and non-GFP C. elegans L4 larvae. This mixed population wassorted to obtain the population of GFP-containing larvae. To stop thelarvae from moving to take these photographs, the larvae wereheated to 70°C for 10–20 s.
A
B
C
©20
01 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://b
iote
ch.n
atu
re.c
om
© 2001 Nature Publishing Group http://biotech.nature.com

Embryos were collected and sorted from a fly stock carrying aloss-of-function twist allele in trans to a GFP-balancer chromosome.Non-GFP containing embryos (i.e., homozygous twist mutantembryos) were formaldehyde-fixed10 and immunostained with anantibody that recognizes the mesoderm (Fig. 4B). Sorted immuno-stained embryos were examined under a microscope to countembryos that looked normal (i.e., were not homozygous twistmutants) and therefore were sorting errors. Only 15 out of 1,700sorted embryos were wild type, indicating an error rate of 0.88%.Therefore the machine operates at >99% accuracy. This accuracywas not reduced when the embryos were sorted at a rate higher than15 embryos/s, although as noted above the efficiency went down.
Mixed populations of C. elegans L4 larvae were also successfullysorted, selecting larvae that contain GFP (Fig. 4C). The larvae werehighly motile after sorting, indicating that the sorting process hadlittle or no effect on viability. The machine functions between 80 and99% accuracy when sorting C. elegans. The variability in the sortingaccuracy is due to the size differences between C. elegans L4 larvaeand Drosophila embryos. Our machine was scaled for use withDrosophila embryos. The optical cuvette was manufactured so thatonly one Drosophila embryo can pass through the optical cuvette at atime. Caenorhabditis elegans L4 larvae are narrower than Drosophilaembryos (Drosophila embryos being 150–200 µm wide and C. ele-gans L4 larvae 50–80 µm wide), and they can sometimes pass eachother while traversing the optical cuvette between the point of detec-tion and the point of sorting. The accuracy for C. elegans sorting canreadily be improved by manufacturing smaller optical cuvettes andidentifying the optimal size.
DiscussionApplications of the embryo sorter. An important use of the embryosorter is to separate homozygous Drosophila mutant embryos fromtheir GFP-balancer siblings. At present the molecular and biochemi-cal analysis of intact homozygous mutant embryos requires manualsorting of thousands of embryos in order to separate mutants fromthe 75% of the embryos that contain a balancer chromosome. As aresult of the time-consuming and laborious task of hand sortingmutant embryos, biochemical and molecular biology analyses ofDrosophila mutant embryos have been severely restricted.Embryonic development occurs very rapidly (22 h in Drosophila and15 h in C. elegans), making it impossible to hand sort large quantitiesof embryos within a narrow window of time. Automatic sorting of15 living embryos per second with more than 99% accuracy makespossible many previously difficult experiments.
Large quantities of mutant embryos will facilitate whole-genomegene expression analyses of mutants using DNA microarrays. For atypical Drosophila complementary DNA (cDNA) microarray exper-iment, we currently make a fluorescent cDNA probe from 2.5–4 µgof polyA+ RNA. This amount of RNA is purified from ∼ 3,000–4,000embryos, a number easily obtained with the sorter. These studieswill yield insights into molecular ramifications of losing a gene func-tion. For example, embryos that are homozygous mutant for a tran-scription factor can be isolated and the messenger RNA (mRNA)used in microarray experiments to identify direct and indirect tran-scriptional targets. Although many transcription factors are known,few of the genes they regulate have been identified. A much morecomplete analysis of gene regulatory relationships will yield exten-sive insights into cell fate determination, organogenesis, oncogene-sis, and cell proliferation mechanisms. Homozygous mutantembryos can also be used for biochemical studies to analyze the abil-ity of proteins to form multisubunit complexes or to have a normalsubcellular localization when one member of a complex is absent.
The sorter will be useful for selecting GFP-containing embryos.More than 93% of the embryos that pass through the embryo sortersurvive to adulthood, so the sorter can be used to carry out genetic
screens using changes in the production of a fluorescent protein toreveal a mutation. For example, genetic screens could be conductedto identify new oncogenes by isolating embryos with more GFP as aresult of an increased number of GFP-expressing cells. The sortedembryos develop into adults, allowing the maintenance of a newstrain of flies of the desired genotype.
Usefulness of multiple fluorescent protein tags. At present, weare using GFP-labeled embryos. Other fluorescent proteins, includ-ing red fluorescent protein (RFP), cyan fluorescent protein (CFP),and blue fluorescent protein (BFP)11, can be used to do two-colorembryo sorting using appropriate filter sets. For example, to selectembryos that contain CFP but not GFP, the machine can separatenon-GFP embryos from GFP-expressing embryos. After changingfilters, the selected non-GFP embryos can be sorted according toCFP content. In the future, additional filter sets and a third PMT canbe added to detect both fluorescent channels simultaneously.
As more enhancers become characterized, it may be possible tocreate a set of Drosophila and C. elegans strains that each express afluorescent protein at a specific stage of development. This wouldallow stage-specific embryo sorting. Any variability in microarraygene expression profiles that result from slight staging differenceswould be greatly reduced.
Experimental protocolDrosophila embryos have an outer membrane, the chorion, which is oftenremoved, without harming the embryos, to significantly enhance the GFPintensity. The chorion is removed by placing the embryos in 50% bleach for 2 min and then washing the embryos through a sieve into a solution ofPBS/2% Tween-20. All of the sorting is conducted in this buffer. The Tweendoes not affect viability (Fig. 4) and greatly reduces clumping of the embryos.The embryos are placed into the high-density embryo chamber. Once all theair bubbles have been flushed out of the system, some embryos are added tothe low-density embryo chamber and sorting begins. Caenorhabditis eleganslarvae were washed off agar plates in PBS and collected by centrifugation.Bacteria were removed by washing four times in PBS. The larvae were sortedin PBS/0.1% Tween-20. A detailed description of how to build the sorter isavailable as supplementary information.
Note: Supplementary information can be found on the NatureBiotechnology website in Web Extras (http://biotech.nature.com/web_extras).
AcknowledgmentsWe are grateful to Dr. Stephen Smith for his suggestions and discussions on theoptics used in the machine. We thank Dr. Roel Nusse for his careful reading of themanuscript, and Dr. Allan Spradling for advice. The C. elegans larvae werekindly provided by Drs. Peter J. Roy and Stuart Kim. E.F. was supported by aEuropean Molecular Biology Organization fellowship and a Stanford BerryFellowship. The research was supported by the Howard Hughes Medical Instituteand DARPA grant number N00014-98-1-0689.
1. Mendel, G. Versuche über Pflanzen-Hybriden. Vorgelegt in den Sitzungen 8 (1865).2. Casso, D., Ramirez-Weber, F.A. & Kornberg, T.B. GFP-tagged balancer chromo-
somes for Drosophila melanogaster. Mech. Dev. 88, 229–232 (1999).3. Melamed, M.R, Lindmo, T.L. & Mendelsohn, M.L. Flow cytometry and sorting.
(Wiley-Liss, New York, NY; 1990).4. Shapiro, H.M. Practical flow cytometry. (Wiley-Liss, New York, NY; 1995).5. Crosland-Taylor, P.J. A device for counting small particles suspended in a fluid
through a tube. Nature 171, 37–38 (1953).6. Ashburner, M. Drosophila, a laboratory handbook. (Cold Spring Harbor
Laboratory Press, Cold Spring Harbor, NY; 1989).7. Wood, W.B. The nematode Caenorhabditis elegans. (Cold Spring Harbor
Laboratory Press, Cold Spring Harbor, NY; 1988).8. Thisse, B., el Messal, M. & Perrin-Schmitt, F. The twist gene: isolation of a
Drosophila zygotic gene necessary for the establishment of dorsoventral pattern.Nucleic Acids Res. 15, 3439–3453 (1987).
9. Reuter, R. & Leptin, M. Interacting functions of snail, twist and huckebein duringthe early development of germ layers in Drosophila. Development 120,1137–1150 (1994).
10. Goldstein, L.S. & Fyrberg, E.A. Practical uses in cell and molecular biology.(Academic Press, San Diego, CA; 1994).
11. Ellenberg, J., Lippincott-Schwartz, J. & Presley, J.F. Dual-colour imaging withGFP variants. Trends Cell Biol. 9, 52–56 (1999).
156 NATURE BIOTECHNOLOGY VOL 19 FEBRUARY 2001 http://biotech.nature.com
RESEARCH ARTICLES©
2001
Nat
ure
Pu
blis
hin
g G
rou
p
htt
p:/
/bio
tech
.nat
ure
.co
m© 2001 Nature Publishing Group http://biotech.nature.com

![Integrating the Healthcare Enterprise€¦ · Document Source Document ConsumerOn Entry [ITI Document Registry Document Repository Provide&Register Document Set – b [ITI-41] →](https://img.pdfslide.net/doc/110x75/5f08a1eb7e708231d422f7c5/integrating-the-healthcare-enterprise-document-source-document-consumeron-entry.jpg)



























