Embed Size (px)
Citation preview

Signaling and Regulation
A Coactivator Role of CARM1 in the Dysregulation ofb-Catenin Activity in Colorectal Cancer CellGrowth and Gene Expression
Chen-Yin Ou1, Melissa J. LaBonte2, Philipp C. Manegold3, Alex Yick-Lun So4,5, Irina Ianculescu1,Daniel S. Gerke1, Keith R. Yamamoto4,5, Robert D. Ladner2, Michael Kahn1,3,Jeong Hoon Kim6, and Michael R. Stallcup1
AbstractAberrant activation of Wnt/b-catenin signaling, resulting in the expression of Wnt-regulated oncogenes, is
recognized as a critical factor in the etiology of colorectal cancer. Occupancy of b-catenin at promoters of Wnttarget genes drives transcription, but the mechanism of b-catenin action remains poorly understood. Here, weshow that CARM1 (coactivator-associated arginine methyltransferase 1) interacts with b-catenin and positivelymodulates b-catenin–mediated gene expression. In colorectal cancer cells with constitutively high Wnt/b-cateninactivity, depletion of CARM1 inhibits expression of endogenousWnt/b-catenin target genes and suppresses clonalsurvival and anchorage-independent growth.We also identified a colorectal cancer cell line (RKO) with a low basallevel of b-catenin, which is dramatically elevated by treatment withWnt3a. Wnt3a also increased the expression ofa subset of endogenous Wnt target genes, and CARM1 was required for the Wnt-induced expression of thesetarget genes and the accompanying dimethylation of arginine 17 of histone H3. Depletion of b-catenin from RKOcells diminished the Wnt-induced occupancy of CARM1 on a Wnt target gene, indicating that CARM1 isrecruited to Wnt target genes through its interaction with b-catenin and contributes to transcriptional activationby mediating events (including histone H3 methylation) that are downstream from the actions of b-catenin.Therefore, CARM1 is an important positive modulator of Wnt/b-catenin transcription and neoplastic trans-formation, and may thereby represent a novel target for therapeutic intervention in cancers involving aberrantlyactivated Wnt/b-catenin signaling. Mol Cancer Res; 9(5); 660–70. �2011 AACR.
Introduction
Wnt/b-catenin signaling is indispensible for the devel-opment of the gastrointestinal system, and aberrant activa-tion of this pathway is implicated in disease, notably incolorectal cancer (1–3). A hallmark of Wnt pathway activa-tion is the elevation of b-catenin protein levels. In theabsence of Wnt signaling, b-catenin protein is targeted to a
multisubunit degradation complex that is composed of thescaffolding proteins adenomatous polyposis coli (APC) andAxin, and the protein kinases casein kinase 1a (CK1a) andGSK3b. Phosphorylation of b-catenin within this complextargets it for polyubiquitination by bTrCP and subse-quently for proteasome-mediated proteolysis (3). BindingofWnt ligands to transmembrane-frizzled receptors disruptsthe destruction complex, and thereby stabilizes and causesaccumulation of b-catenin protein. b-catenin then translo-cates into the nucleus, where it binds to various transcrip-tion factors, including members of the lymphoid enhancerfactor (LEF)/T cell factor (TCF) family, and functions as atranscription coactivator to activate the expression of Wnttarget genes (3).Various mutations in components of the Wnt signaling
pathway, which are frequently observed in colorectal cancer,result in an abnormally elevated b-catenin protein level; thisleads to constitutive formation of LEF/TCF-b-catenincomplexes and alters expression of b-catenin–regulatedgenes, including well-known proto-oncogenes. Proteinsencoded by b-catenin–controlled genes likely cooperatein neoplastic transformation. In support of this notion,analysis of Wnt pathway component mutations in mice andidentification of mutations in human clinical specimens has
Authors' Affiliations: Departments of 1Biochemistry and Molecular Biol-ogy and 2Pathology, and 3Eli and Edythe Broad Center for RegenerativeMedicine and Stem Cell Research, Keck School of Medicine, University ofSouthern California, Los Angeles; 4Department of Cellular and MolecularPharmacology, 5Chemistry and Chemical Biology Graduate Program,University of California, San Francisco, California; 6Samsung MedicalCenter and Center for Clinical Research, Samsung Biomedical ResearchInstitute, Sungkyunkwan University School of Medicine, Seoul, Korea
Note: Supplementary data for this article are available at Molecular CancerResearch Online (http://mcr.aacrjournals.org/).
Corresponding Author:Michael R. Stallcup, Department of Biochemistryand Molecular Biology, Norris Comprehensive Cancer Center, Universityof Southern California (USC), 1441 Eastlake Avenue, NOR 6316, Universityof Southern California, Los Angeles, CA 90089-9176. Phone: 323-865-3852; Fax: 323-865-3866. E-mail: [email protected]
doi: 10.1158/1541-7786.MCR-10-0223
�2011 American Association for Cancer Research.
MolecularCancer
Research
Mol Cancer Res; 9(5) May 2011660
on June 21, 2018. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst April 8, 2011; DOI: 10.1158/1541-7786.MCR-10-0223

shown that abnormal stabilization of b-catenin leads tocolon tumorigenesis (4). Wnt/b-catenin target genesinclude known proproliferative genes (e.g., c-Myc; ref.5), antiapoptosis genes (e.g., survivin; refs. 6, 7), and genesinvolved in metastasis (e.g., S100A4; ref. 8). GPR49, anewly identified Wnt target gene, encodes an orphan Gprotein–coupled receptor and was proposed as a potentialmarker of tumor-initiating colorectal cancer stem cells (9).In addition, GPR49 was found overexpressed in colontumors compared with normal colon tissues; and knock-down of GPR49 in colon cancer cells induced apoptosis(10). Thereby, GPR49, in concert with other Wnt/b-cate-nin targets, promotes growth and/or survival of cancers withdefects in b-catenin regulation.When bound to target genes through its interaction with
LEF/TCF transcription factors, b-catenin functions as aprimary coactivator by recruiting additional coactivators,each of which makes specific contributions to the remodel-ing of chromatin conformation and/or the assembly of anactive transcription complex. Therefore, it is crucial toidentify and characterize the additional coactivators thatmodulate or mediate the transcriptional activity ofb-catenin. Coactivator CARM1 (coactivator-associatedarginine methyltransferase 1) belongs to the protein argi-nine methyltransferase (PRMT) family that methylatesarginine residues in proteins. CARM1 methylates a subsetof proteins that play crucial roles in transcription. Forinstance, CARM1 methylates arginine 17 of histone H3(H3R17) at promoters, and this event is associatedwith transcriptional activation (11, 12). CARM1 functionsas a transcriptional coactivator for many different DNA-binding transcriptional activator proteins, including estro-gen receptor, androgen receptor, c-Fos, peroxisome prolif-erator-activated receptor g , and NFkB (11). It waspreviously shown that CARM1 binds to b-catenin in vivoand functions in synergy with b-catenin as a coactivator forLEF1-mediated expression of artificial and transiently trans-fected reporter plasmids, suggesting the role of CARM1 inb-catenin signaling (13).Here, we report that CARM1 is overexpressed in human
colon cancer cell lines. Importantly, we report that CARM1plays a critical role in clonal survival and anchorage-inde-pendent growth of colorectal cancers by mediating Wnt/b-catenin signaling. Wnt3a signaling induced CARM1occupancy at the promoter of an endogenous Wnt targetgene in its native chromosomal location; CARM1 occu-pancy correlated with arginine dimethylation of histone H3(H3R17me2). Indeed, we showed that endogenousCARM1 is required for both Wnt3a-dependent gene acti-vation and H3 arginine dimethylation at a target genepromoter in colon cancer cells. Moreover, we examinedthe mechanism by which CARM1 directs Wnt/b-cateninsignaling. We showed that the methyltransferase domain ofCARM1 specifically interacts with b-catenin; b-cateninrecruits CARM1 to the promoter; and regulated transcrip-tion is dependent on CARM1. Interestingly, CARM1synergized with the coactivator p300 to further enhanceWnt/b-catenin activity. Collectively, our studies suggest
that CARM1 plays an essential role in oncogenic growthof colon cancers through the positive regulation of Wnt/b-catenin oncogenes, and is thereby a potential therapeutictarget in cancers involving abnormally activated Wnt/b-catenin signaling.
Materials and Methods
For detailed description of Materials and Methods, referto Supplementary data. Briefly, plasmids, cell culture,luciferase reporter gene assay, glutathione S transferase(GST) pull down, chromatin immunoprecipitation (ChIP),quantitative reverse-transcriptase PCR (qRT-PCR), cellproliferation assay, and colony formation assay in soft agarwere done as described previously (14). CARM1 andb-catenin were stably depleted by short-hairpin RNA(shRNA)–expressing lentiviruses. Colony formation assaywas performed as described previously (15).
Results
CARM1 interacts with b-catenin, but not LEF1Previous studies in our laboratory have shown the in vivo
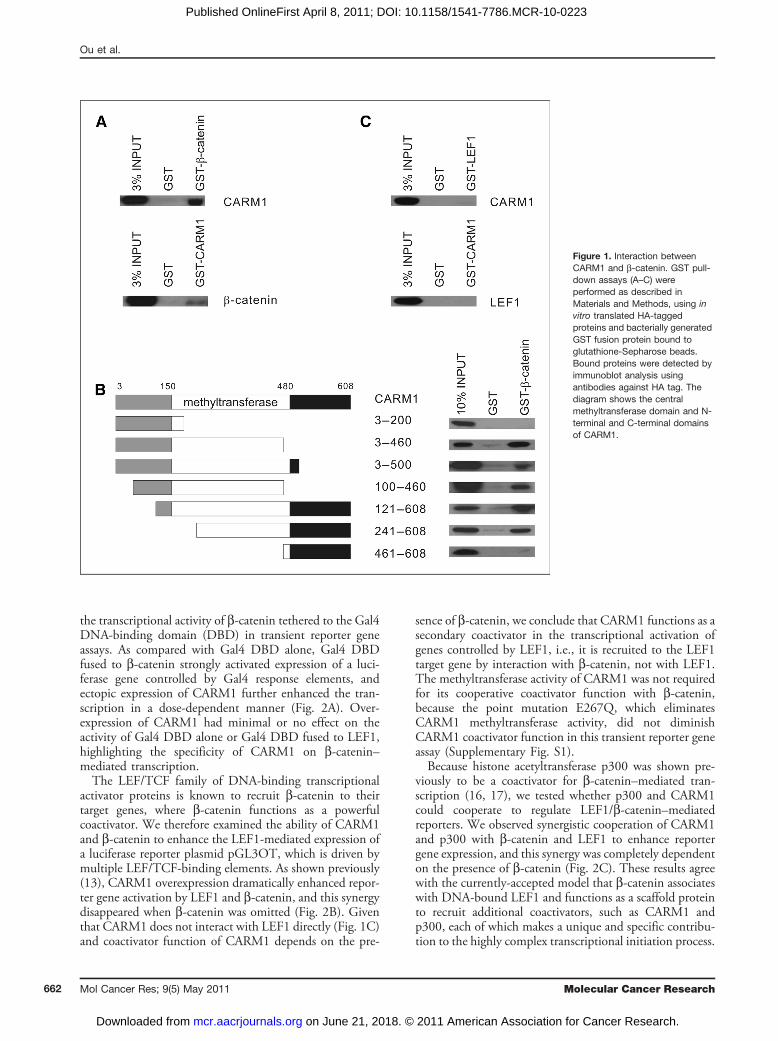
interaction between CARM1 and b-catenin (13). To definethe precise domain(s) by which CARM1 interacts withb-catenin, HA-tagged CARM1 and selected fragments weresynthesized in vitro and incubated with GST-b-cateninfusion protein bound to glutathione-Sepharose beads.The associated bound proteins were purified and analyzedby immunoblot. Full-length HA-CARM1 was bound byGST-b-catenin, but not by GST (Fig. 1A, upper); andconversely, b-catenin synthesized in vitro also bound spe-cifically to full-length GST-CARM1 (lower). Variousdomains of CARM1 were synthesized in vitro and testedfor binding to GST-b-catenin (Fig. 1B). The N-terminaland C-terminal domains of CARM1 (amino acids 3–200and 461–608, respectively) did not interact with b-catenin,whereas 5 overlapping fragments of CARM1 (amino acids3–460, 3–500, 100–460, 121–608, and 241–608) boundspecifically to GST-b-catenin, indicating that the methyl-transferase domain (which resides within amino acids 150–480) is responsible for b-catenin binding. Because themethyltransferase domain is conserved among membersof the PRMT family (11), b-catenin may also interact withother PRMTs besides CARM1. Because b-catenin func-tions as a coactivator for the LEF/TCF family of DNA-binding transcriptional activator proteins, we also testedwhether there is an interaction between CARM1 and LEF1;however, no interaction was detected by GST pull-downassay (Fig. 1C), indicating that CARM1 interaction withb-catenin is specific and may play a functional role in Wnt-dependent gene induction.
CARM1 and b-catenin cooperate as transcriptionalcoactivators for LEF1To address whether the binding of CARM1 to b-catenin
could modulate b-catenin–mediated transcription, wemonitored the influence of CARM1 overexpression on
CARM1 and b-Catenin in Aberrant Growth of Colon Cancer Cells
www.aacrjournals.org Mol Cancer Res; 9(5) May 2011 661
on June 21, 2018. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst April 8, 2011; DOI: 10.1158/1541-7786.MCR-10-0223
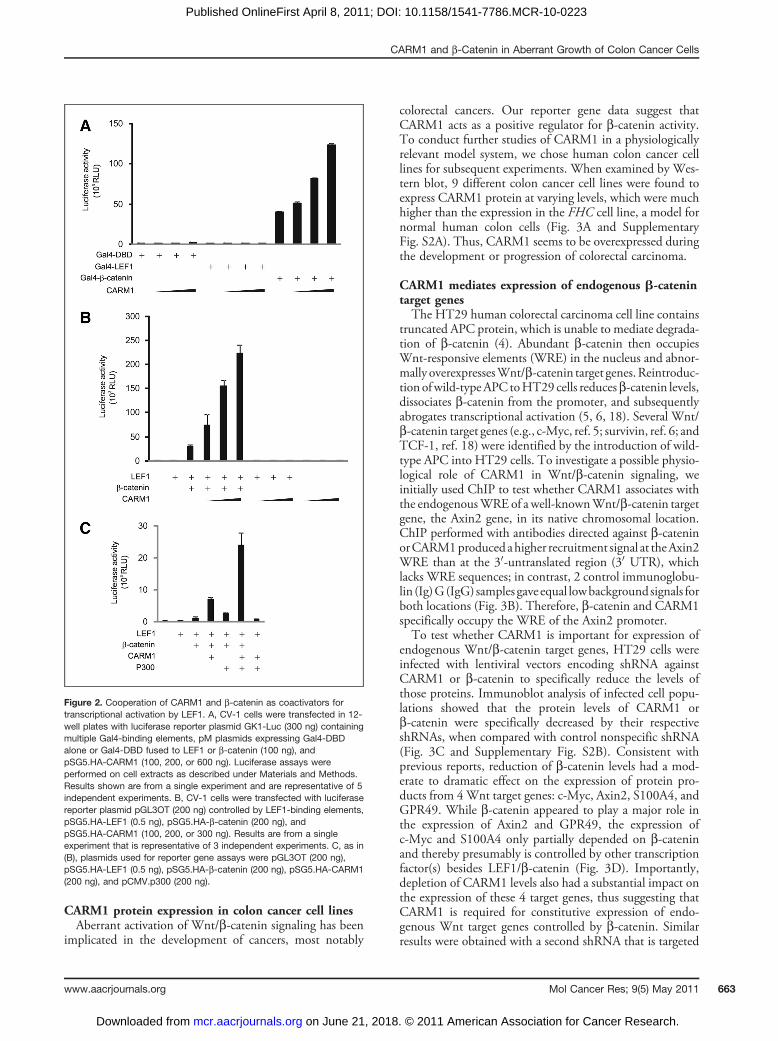
the transcriptional activity of b-catenin tethered to the Gal4DNA-binding domain (DBD) in transient reporter geneassays. As compared with Gal4 DBD alone, Gal4 DBDfused to b-catenin strongly activated expression of a luci-ferase gene controlled by Gal4 response elements, andectopic expression of CARM1 further enhanced the tran-scription in a dose-dependent manner (Fig. 2A). Over-expression of CARM1 had minimal or no effect on theactivity of Gal4 DBD alone or Gal4 DBD fused to LEF1,highlighting the specificity of CARM1 on b-catenin–mediated transcription.The LEF/TCF family of DNA-binding transcriptional
activator proteins is known to recruit b-catenin to theirtarget genes, where b-catenin functions as a powerfulcoactivator. We therefore examined the ability of CARM1and b-catenin to enhance the LEF1-mediated expression ofa luciferase reporter plasmid pGL3OT, which is driven bymultiple LEF/TCF-binding elements. As shown previously(13), CARM1 overexpression dramatically enhanced repor-ter gene activation by LEF1 and b-catenin, and this synergydisappeared when b-catenin was omitted (Fig. 2B). Giventhat CARM1 does not interact with LEF1 directly (Fig. 1C)and coactivator function of CARM1 depends on the pre-
sence of b-catenin, we conclude that CARM1 functions as asecondary coactivator in the transcriptional activation ofgenes controlled by LEF1, i.e., it is recruited to the LEF1target gene by interaction with b-catenin, not with LEF1.The methyltransferase activity of CARM1 was not requiredfor its cooperative coactivator function with b-catenin,because the point mutation E267Q, which eliminatesCARM1 methyltransferase activity, did not diminishCARM1 coactivator function in this transient reporter geneassay (Supplementary Fig. S1).Because histone acetyltransferase p300 was shown pre-
viously to be a coactivator for b-catenin–mediated tran-scription (16, 17), we tested whether p300 and CARM1could cooperate to regulate LEF1/b-catenin–mediatedreporters. We observed synergistic cooperation of CARM1and p300 with b-catenin and LEF1 to enhance reportergene expression, and this synergy was completely dependenton the presence of b-catenin (Fig. 2C). These results agreewith the currently-accepted model that b-catenin associateswith DNA-bound LEF1 and functions as a scaffold proteinto recruit additional coactivators, such as CARM1 andp300, each of which makes a unique and specific contribu-tion to the highly complex transcriptional initiation process.
Figure 1. Interaction betweenCARM1 and b-catenin. GST pull-down assays (A–C) wereperformed as described inMaterials and Methods, using invitro translated HA-taggedproteins and bacterially generatedGST fusion protein bound toglutathione-Sepharose beads.Bound proteins were detected byimmunoblot analysis usingantibodies against HA tag. Thediagram shows the centralmethyltransferase domain and N-terminal and C-terminal domainsof CARM1.
Ou et al.
Mol Cancer Res; 9(5) May 2011 Molecular Cancer Research662
on June 21, 2018. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst April 8, 2011; DOI: 10.1158/1541-7786.MCR-10-0223
CARM1 protein expression in colon cancer cell linesAberrant activation of Wnt/b-catenin signaling has been
implicated in the development of cancers, most notably
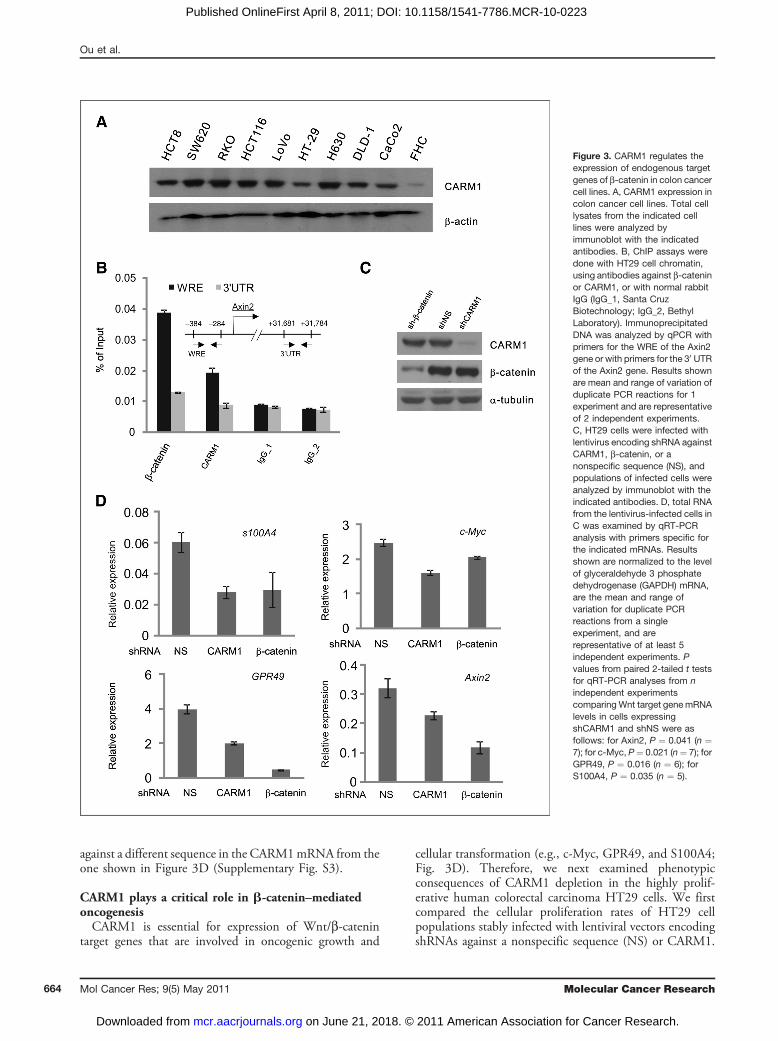
colorectal cancers. Our reporter gene data suggest thatCARM1 acts as a positive regulator for b-catenin activity.To conduct further studies of CARM1 in a physiologicallyrelevant model system, we chose human colon cancer celllines for subsequent experiments. When examined by Wes-tern blot, 9 different colon cancer cell lines were found toexpress CARM1 protein at varying levels, which were muchhigher than the expression in the FHC cell line, a model fornormal human colon cells (Fig. 3A and SupplementaryFig. S2A). Thus, CARM1 seems to be overexpressed duringthe development or progression of colorectal carcinoma.
CARM1 mediates expression of endogenous b-catenintarget genesThe HT29 human colorectal carcinoma cell line contains
truncated APC protein, which is unable to mediate degrada-tion of b-catenin (4). Abundant b-catenin then occupiesWnt-responsive elements (WRE) in the nucleus and abnor-mally overexpressesWnt/b-catenin target genes.Reintroduc-tion ofwild-typeAPC toHT29 cells reducesb-catenin levels,dissociates b-catenin from the promoter, and subsequentlyabrogates transcriptional activation (5, 6, 18). Several Wnt/b-catenin target genes (e.g., c-Myc, ref. 5; survivin, ref. 6; andTCF-1, ref. 18) were identified by the introduction of wild-type APC into HT29 cells. To investigate a possible physio-logical role of CARM1 in Wnt/b-catenin signaling, weinitially used ChIP to test whether CARM1 associates withthe endogenousWREof a well-knownWnt/b-catenin targetgene, the Axin2 gene, in its native chromosomal location.ChIP performed with antibodies directed against b-cateninorCARM1produced ahigher recruitment signal at theAxin2WRE than at the 30-untranslated region (30 UTR), whichlacks WRE sequences; in contrast, 2 control immunoglobu-lin (Ig)G(IgG) samples gave equal lowbackground signals forboth locations (Fig. 3B). Therefore, b-catenin and CARM1specifically occupy the WRE of the Axin2 promoter.To test whether CARM1 is important for expression of
endogenous Wnt/b-catenin target genes, HT29 cells wereinfected with lentiviral vectors encoding shRNA againstCARM1 or b-catenin to specifically reduce the levels ofthose proteins. Immunoblot analysis of infected cell popu-lations showed that the protein levels of CARM1 orb-catenin were specifically decreased by their respectiveshRNAs, when compared with control nonspecific shRNA(Fig. 3C and Supplementary Fig. S2B). Consistent withprevious reports, reduction of b-catenin levels had a mod-erate to dramatic effect on the expression of protein pro-ducts from 4Wnt target genes: c-Myc, Axin2, S100A4, andGPR49. While b-catenin appeared to play a major role inthe expression of Axin2 and GPR49, the expression ofc-Myc and S100A4 only partially depended on b-cateninand thereby presumably is controlled by other transcriptionfactor(s) besides LEF1/b-catenin (Fig. 3D). Importantly,depletion of CARM1 levels also had a substantial impact onthe expression of these 4 target genes, thus suggesting thatCARM1 is required for constitutive expression of endo-genous Wnt target genes controlled by b-catenin. Similarresults were obtained with a second shRNA that is targeted
Figure 2. Cooperation of CARM1 and b-catenin as coactivators fortranscriptional activation by LEF1. A, CV-1 cells were transfected in 12-well plates with luciferase reporter plasmid GK1-Luc (300 ng) containingmultiple Gal4-binding elements, pM plasmids expressing Gal4-DBDalone or Gal4-DBD fused to LEF1 or b-catenin (100 ng), andpSG5.HA-CARM1 (100, 200, or 600 ng). Luciferase assays wereperformed on cell extracts as described under Materials and Methods.Results shown are from a single experiment and are representative of 5independent experiments. B, CV-1 cells were transfected with luciferasereporter plasmid pGL3OT (200 ng) controlled by LEF1-binding elements,pSG5.HA-LEF1 (0.5 ng), pSG5.HA-b-catenin (200 ng), andpSG5.HA-CARM1 (100, 200, or 300 ng). Results are from a singleexperiment that is representative of 3 independent experiments. C, as in(B), plasmids used for reporter gene assays were pGL3OT (200 ng),pSG5.HA-LEF1 (0.5 ng), pSG5.HA-b-catenin (200 ng), pSG5.HA-CARM1(200 ng), and pCMV.p300 (200 ng).
CARM1 and b-Catenin in Aberrant Growth of Colon Cancer Cells
www.aacrjournals.org Mol Cancer Res; 9(5) May 2011 663
on June 21, 2018. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst April 8, 2011; DOI: 10.1158/1541-7786.MCR-10-0223
against a different sequence in the CARM1mRNA from theone shown in Figure 3D (Supplementary Fig. S3).
CARM1 plays a critical role in b-catenin–mediatedoncogenesisCARM1 is essential for expression of Wnt/b-catenin
target genes that are involved in oncogenic growth and
cellular transformation (e.g., c-Myc, GPR49, and S100A4;Fig. 3D). Therefore, we next examined phenotypicconsequences of CARM1 depletion in the highly prolif-erative human colorectal carcinoma HT29 cells. We firstcompared the cellular proliferation rates of HT29 cellpopulations stably infected with lentiviral vectors encodingshRNAs against a nonspecific sequence (NS) or CARM1.
Figure 3. CARM1 regulates theexpression of endogenous targetgenes of b-catenin in colon cancercell lines. A, CARM1 expression incolon cancer cell lines. Total celllysates from the indicated celllines were analyzed byimmunoblot with the indicatedantibodies. B, ChIP assays weredone with HT29 cell chromatin,using antibodies against b-cateninor CARM1, or with normal rabbitIgG (IgG_1, Santa CruzBiotechnology; IgG_2, BethylLaboratory). ImmunoprecipitatedDNA was analyzed by qPCR withprimers for the WRE of the Axin2gene or with primers for the 30 UTRof the Axin2 gene. Results shownare mean and range of variation ofduplicate PCR reactions for 1experiment and are representativeof 2 independent experiments.C, HT29 cells were infected withlentivirus encoding shRNA againstCARM1, b-catenin, or anonspecific sequence (NS), andpopulations of infected cells wereanalyzed by immunoblot with theindicated antibodies. D, total RNAfrom the lentivirus-infected cells inC was examined by qRT-PCRanalysis with primers specific forthe indicated mRNAs. Resultsshown are normalized to the levelof glyceraldehyde 3 phosphatedehydrogenase (GAPDH) mRNA,are the mean and range ofvariation for duplicate PCRreactions from a singleexperiment, and arerepresentative of at least 5independent experiments. Pvalues from paired 2-tailed t testsfor qRT-PCR analyses from nindependent experimentscomparingWnt target genemRNAlevels in cells expressingshCARM1 and shNS were asfollows: for Axin2, P ¼ 0.041 (n ¼7); for c-Myc, P¼ 0.021 (n¼ 7); forGPR49, P ¼ 0.016 (n ¼ 6); forS100A4, P ¼ 0.035 (n ¼ 5).
Ou et al.
Mol Cancer Res; 9(5) May 2011 Molecular Cancer Research664
on June 21, 2018. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst April 8, 2011; DOI: 10.1158/1541-7786.MCR-10-0223
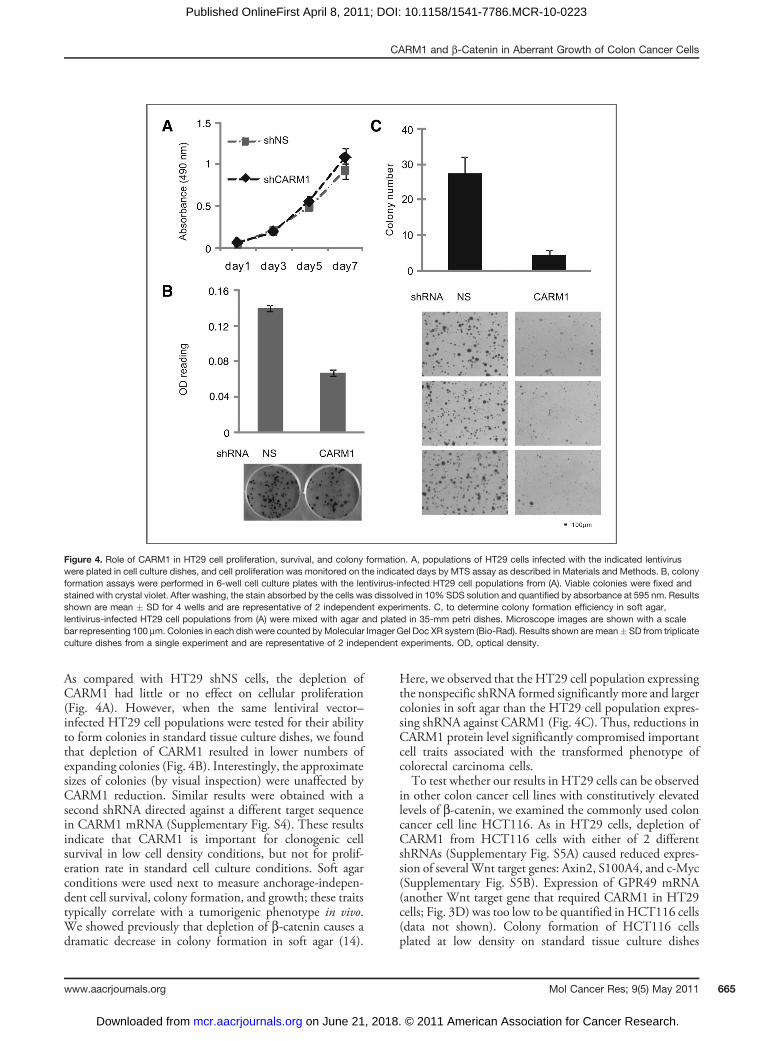
As compared with HT29 shNS cells, the depletion ofCARM1 had little or no effect on cellular proliferation(Fig. 4A). However, when the same lentiviral vector–infected HT29 cell populations were tested for their abilityto form colonies in standard tissue culture dishes, we foundthat depletion of CARM1 resulted in lower numbers ofexpanding colonies (Fig. 4B). Interestingly, the approximatesizes of colonies (by visual inspection) were unaffected byCARM1 reduction. Similar results were obtained with asecond shRNA directed against a different target sequencein CARM1 mRNA (Supplementary Fig. S4). These resultsindicate that CARM1 is important for clonogenic cellsurvival in low cell density conditions, but not for prolif-eration rate in standard cell culture conditions. Soft agarconditions were used next to measure anchorage-indepen-dent cell survival, colony formation, and growth; these traitstypically correlate with a tumorigenic phenotype in vivo.We showed previously that depletion of b-catenin causes adramatic decrease in colony formation in soft agar (14).
Here, we observed that the HT29 cell population expressingthe nonspecific shRNA formed significantly more and largercolonies in soft agar than the HT29 cell population expres-sing shRNA against CARM1 (Fig. 4C). Thus, reductions inCARM1 protein level significantly compromised importantcell traits associated with the transformed phenotype ofcolorectal carcinoma cells.To test whether our results in HT29 cells can be observed
in other colon cancer cell lines with constitutively elevatedlevels of b-catenin, we examined the commonly used coloncancer cell line HCT116. As in HT29 cells, depletion ofCARM1 from HCT116 cells with either of 2 differentshRNAs (Supplementary Fig. S5A) caused reduced expres-sion of several Wnt target genes: Axin2, S100A4, and c-Myc(Supplementary Fig. S5B). Expression of GPR49 mRNA(another Wnt target gene that required CARM1 in HT29cells; Fig. 3D) was too low to be quantified in HCT116 cells(data not shown). Colony formation of HCT116 cellsplated at low density on standard tissue culture dishes
Figure 4. Role of CARM1 in HT29 cell proliferation, survival, and colony formation. A, populations of HT29 cells infected with the indicated lentiviruswere plated in cell culture dishes, and cell proliferation was monitored on the indicated days by MTS assay as described in Materials and Methods. B, colonyformation assays were performed in 6-well cell culture plates with the lentivirus-infected HT29 cell populations from (A). Viable colonies were fixed andstained with crystal violet. After washing, the stain absorbed by the cells was dissolved in 10% SDS solution and quantified by absorbance at 595 nm. Resultsshown are mean � SD for 4 wells and are representative of 2 independent experiments. C, to determine colony formation efficiency in soft agar,lentivirus-infected HT29 cell populations from (A) were mixed with agar and plated in 35-mm petri dishes. Microscope images are shown with a scalebar representing 100 mm. Colonies in each dish were counted byMolecular Imager Gel Doc XR system (Bio-Rad). Results shown are mean� SD from triplicateculture dishes from a single experiment and are representative of 2 independent experiments. OD, optical density.
CARM1 and b-Catenin in Aberrant Growth of Colon Cancer Cells
www.aacrjournals.org Mol Cancer Res; 9(5) May 2011 665
on June 21, 2018. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst April 8, 2011; DOI: 10.1158/1541-7786.MCR-10-0223
was also substantially reduced by depletion of CARM1 byeither of the 2 shRNAs against CARM1 (SupplementaryFig. S5C). Thus, the requirement of CARM1 for expressionof Wnt/b-catenin target genes and for colony formation ischaracteristic in multiple colon cancer cell lines with con-stitutively high b-catenin levels.
Wnt3a ligand induces recruitment of b-catenin andCARM1 to WREs associated with endogenous Wnttarget genesCARM1 depletion reduced the expression of Wnt/
b-catenin target genes in HT29 and HCT116 cells, indi-cating that CARM1 is involved in the transcriptionalactivation process directed by b-catenin. However, crosstalk between the Wnt/b-catenin cascade and other signalingcascade(s) that converge on the regulation of the same geneshas been reported, and thus, it is possible that the action ofCARM1 onWnt/b-catenin target gene expression is not theresult of direct interaction of CARM1 with b-catenin atLEF/TCF binding sites, but through interaction ofCARM1 with other transcription factors associated withthe same promoters. Therefore, we identified a cell systemthat allowed us to test the role of CARM1 inWnt-inducibleexpression of endogenous target genes.RKO human colon carcinoma cells have low cellular
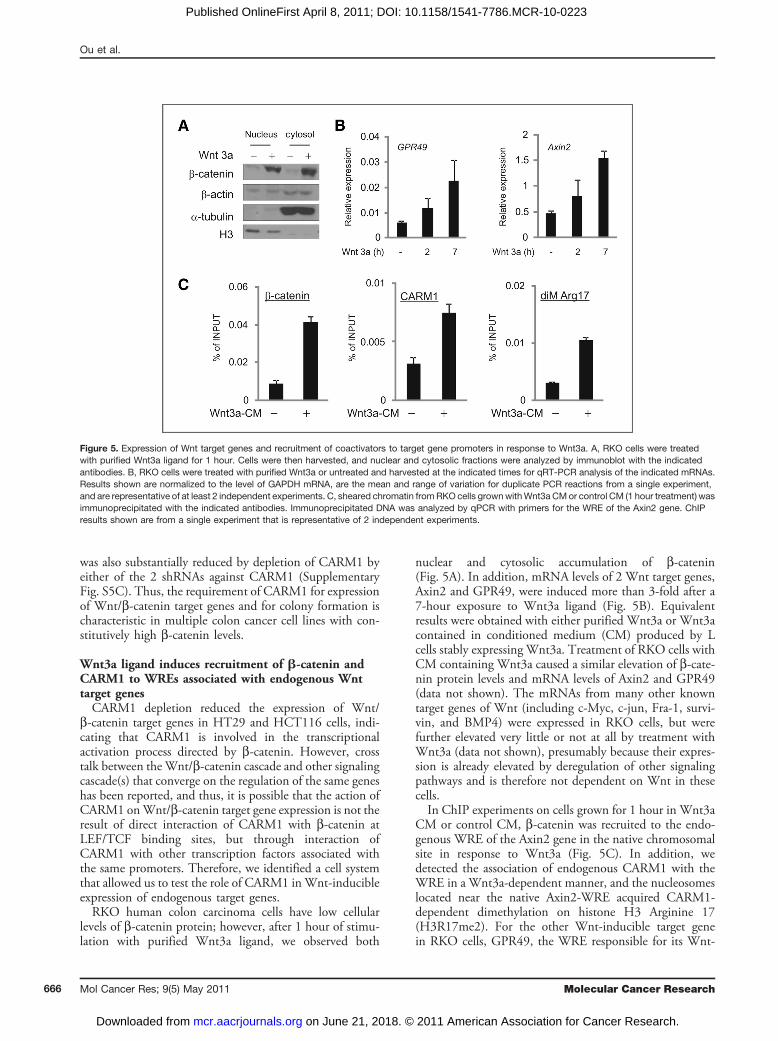
levels of b-catenin protein; however, after 1 hour of stimu-lation with purified Wnt3a ligand, we observed both
nuclear and cytosolic accumulation of b-catenin(Fig. 5A). In addition, mRNA levels of 2 Wnt target genes,Axin2 and GPR49, were induced more than 3-fold after a7-hour exposure to Wnt3a ligand (Fig. 5B). Equivalentresults were obtained with either purified Wnt3a or Wnt3acontained in conditioned medium (CM) produced by Lcells stably expressing Wnt3a. Treatment of RKO cells withCM containing Wnt3a caused a similar elevation of b-cate-nin protein levels and mRNA levels of Axin2 and GPR49(data not shown). The mRNAs from many other knowntarget genes of Wnt (including c-Myc, c-jun, Fra-1, survi-vin, and BMP4) were expressed in RKO cells, but werefurther elevated very little or not at all by treatment withWnt3a (data not shown), presumably because their expres-sion is already elevated by deregulation of other signalingpathways and is therefore not dependent on Wnt in thesecells.In ChIP experiments on cells grown for 1 hour in Wnt3a
CM or control CM, b-catenin was recruited to the endo-genous WRE of the Axin2 gene in the native chromosomalsite in response to Wnt3a (Fig. 5C). In addition, wedetected the association of endogenous CARM1 with theWRE in a Wnt3a-dependent manner, and the nucleosomeslocated near the native Axin2-WRE acquired CARM1-dependent dimethylation on histone H3 Arginine 17(H3R17me2). For the other Wnt-inducible target genein RKO cells, GPR49, the WRE responsible for its Wnt-
Figure 5. Expression of Wnt target genes and recruitment of coactivators to target gene promoters in response to Wnt3a. A, RKO cells were treatedwith purified Wnt3a ligand for 1 hour. Cells were then harvested, and nuclear and cytosolic fractions were analyzed by immunoblot with the indicatedantibodies. B, RKO cells were treated with purified Wnt3a or untreated and harvested at the indicated times for qRT-PCR analysis of the indicated mRNAs.Results shown are normalized to the level of GAPDH mRNA, are the mean and range of variation for duplicate PCR reactions from a single experiment,and are representative of at least 2 independent experiments. C, sheared chromatin fromRKO cells grownwithWnt3a CMor control CM (1 hour treatment) wasimmunoprecipitated with the indicated antibodies. Immunoprecipitated DNA was analyzed by qPCR with primers for the WRE of the Axin2 gene. ChIPresults shown are from a single experiment that is representative of 2 independent experiments.
Ou et al.
Mol Cancer Res; 9(5) May 2011 Molecular Cancer Research666
on June 21, 2018. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst April 8, 2011; DOI: 10.1158/1541-7786.MCR-10-0223

inducible expression has not been identified. However,although the expression of other classical Wnt target genesis not upregulated by Wnt in these cells, it is still possiblethat b-catenin is recruited to the WREs associated withthese genes in response to Wnt. Therefore, we examinedoccupancy of the WRE associated with the survivin gene, aclassical Wnt target gene that is expressed in RKO cells butis not further induced by Wnt3a (data not shown). Weobserved that, as with the Axin2 WRE, Wnt3a treatmentincreased occupancy of the survivinWRE by both b-cateninand CARM1 (Supplementary Fig. S6). Thus, Wnt-indu-cible occupancy by b-catenin and CARM1 is observed evenwhen Wnt does not cause the increased expression of aclassical Wnt target gene. Our results show that RKO cellsrepresent a novel and convenient system for studying themechanism of Wnt-regulated gene expression.
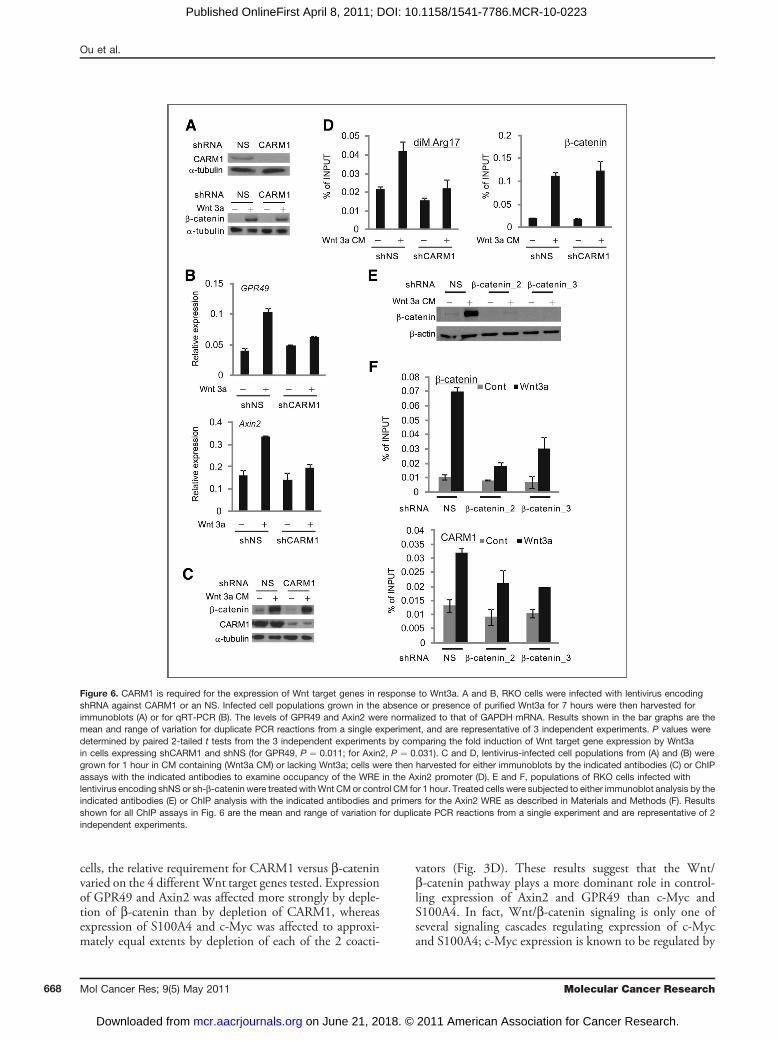
CARM1 is required for Wnt3a-dependenttranscriptional activationTo assess the requirement for CARM1 in Wnt3a-depen-
dent activation of the Axin2 and GPR49 genes, CARM1expression was efficiently and specifically silenced in RKOcells by infection with lentiviral vectors encoding shRNAagainst CARM1 (Fig. 6A). Upon CARM1 silencing, theWnt3a-induced increase in Axin2 and GPR49 was almosteliminated, whereas normal induction of expression of thesegenes was observed in cells expressing a nonspecific shRNA(Fig. 6B). However, depletion of CARM1 did not interferewith the Wnt3a-induced accumulation of b-catenin(Fig. 6A). Thus, the influence of CARM1 in Wnt3a/b-catenin signaling is not due to an effect on b-cateninprotein accumulation but rather events downstream.
Wnt3a-dependent recruitment of CARM1 to Axin2WRE requires b-cateninTo examine the mechanism of CARM1 recruitment to
WREs and the contributions of CARM1 to Wnt3a-depen-dent target gene activation, ChIP assays were performed forthe Wnt response elements in the Axin2 promoter in RKOcells expressing shRNA against CARM1, b-catenin, or anNS. Immunoblots confirmed the efficient and specificdepletion of CARM1 and the expected increase in b-cateninprotein levels in response to CM containing Wnt3a(Fig. 6C). ChIP assays showed that depletion of CARM1essentially eliminated the Wnt3a-induced dimethylation ofarginine 17 of histone H3 in nucleosomes associated withthe Axin2 WRE; but CARM1 depletion had no effect onthe Wnt3a-regulated recruitment of b-catenin to the Axin2WRE (Fig. 6D). In contrast, reduction of cellular levels ofb-catenin by either of 2 different shRNAs (Fig. 6E) dimin-ished the recruitment of CARM1 to the Axin2 WRE(Fig. 6E). Thus, b-catenin recruitment does not dependon the recruitment of CARM1; instead b-catenin recruit-ment is required for the recruitment of CARM1. Thisshows that CARM1 is required for events downstreamfrom the recruitment of b-catenin and CARM1, includingthe dimethylation of arginine 17 of histone H3. Theseresults are consistent with the results of transient reporter
gene assays, which showed that CARM1 action as a coacti-vator for LEF1 is dependent on the presence of b-catenin(Fig. 2).
Discussion
The Wnt/b-catenin signaling cascade is highly conservedduring evolution and plays a crucial role in the embryonicdevelopment of all animal species. Wnt signals regulatecellular processes such as cellular proliferation, survival, anddifferentiation; misregulation of this signaling pathway isheavily implicated in the development of cancers, mostnotably colorectal cancers (3, 4). Uncontrolled Wnt signal-ing leads to constitutive activation of many oncogenes thatare tightly regulated and only activated transiently duringnormal development. The pivotal mediator that responds toupstream Wnt signals and executes the downstream geneactivation program is b-catenin. Stimulatory Wnt ligandsinduce elevated levels of b-catenin that binds to transcrip-tion factors associated with the promoters of Wnt targetgenes; b-catenin recruits other transcriptional coactivatorsthat activate transcription of the Wnt target genes (3, 19).Identification and functional investigation of proteins thatbind b-catenin and cooperate in regulation of Wnt targetgenes will thus provide novel and exciting insights into thehighly complicated mechanism of Wnt target gene activa-tion. b-catenin–associated coactivators may also representnovel targets for therapeutic intervention against cancerswith dysregulated Wnt signaling.CARM1 is a member of the PRMT family, and methy-
lates a large number of cellular substrates including histonesand nonhistone proteins involved in transcription, RNAprocessing, and other cellular functions. Deregulation ofCARM1 expression is implicated in the pathogenesis ofcancers such that CARM1 expression has been correlatedwith tumor staging (20). CARM1 functions as a transcrip-tional coactivator for several different types of DNA-bind-ing transcriptional activator proteins, and thus alteredCARM1 expression likely affects many transcriptional pro-grams that target genes to control proliferation rate or otheroncogenic properties. In fact, studies from the MCF7 breastcancer cell line showed that CARM1 is a positive regulatorof estrogen receptor-responsive genes and is essential forestrogen-stimulated growth and proliferation of breast car-cinomas (21). Therefore, to further explore the mechanisticknowledge of how deregulated CARM1 might transformcells, we tested whether CARM1 is transcriptionallyinvolved in oncogenic activation of the Wnt/b-cateninsignaling cascade in human colorectal carcinomas.We showed that CARM1 interacts with b-catenin
(Fig. 1) and cooperates with b-catenin at the transcriptionallevel in transient reporter gene assays (Fig. 2). In a morephysiologically relevant model, CARM1 was essential forexpression of several Wnt target genes (including knownoncogenes) in the HT29 andHCT116 colorectal cancer celllines (Fig. 3D and Supplementary Fig. S5) that haveaberrant activation of Wnt/b-catenin signaling and thusconstitutive b-catenin–regulated transcription. In HT29
CARM1 and b-Catenin in Aberrant Growth of Colon Cancer Cells
www.aacrjournals.org Mol Cancer Res; 9(5) May 2011 667
on June 21, 2018. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst April 8, 2011; DOI: 10.1158/1541-7786.MCR-10-0223
cells, the relative requirement for CARM1 versus b-cateninvaried on the 4 differentWnt target genes tested. Expressionof GPR49 and Axin2 was affected more strongly by deple-tion of b-catenin than by depletion of CARM1, whereasexpression of S100A4 and c-Myc was affected to approxi-mately equal extents by depletion of each of the 2 coacti-
vators (Fig. 3D). These results suggest that the Wnt/b-catenin pathway plays a more dominant role in control-ling expression of Axin2 and GPR49 than c-Myc andS100A4. In fact, Wnt/b-catenin signaling is only one ofseveral signaling cascades regulating expression of c-Mycand S100A4; c-Myc expression is known to be regulated by
Figure 6. CARM1 is required for the expression of Wnt target genes in response to Wnt3a. A and B, RKO cells were infected with lentivirus encodingshRNA against CARM1 or an NS. Infected cell populations grown in the absence or presence of purified Wnt3a for 7 hours were then harvested forimmunoblots (A) or for qRT-PCR (B). The levels of GPR49 and Axin2 were normalized to that of GAPDH mRNA. Results shown in the bar graphs are themean and range of variation for duplicate PCR reactions from a single experiment, and are representative of 3 independent experiments. P values weredetermined by paired 2-tailed t tests from the 3 independent experiments by comparing the fold induction of Wnt target gene expression by Wnt3ain cells expressing shCARM1 and shNS (for GPR49, P ¼ 0.011; for Axin2, P ¼ 0.031). C and D, lentivirus-infected cell populations from (A) and (B) weregrown for 1 hour in CM containing (Wnt3a CM) or lacking Wnt3a; cells were then harvested for either immunoblots by the indicated antibodies (C) or ChIPassays with the indicated antibodies to examine occupancy of the WRE in the Axin2 promoter (D). E and F, populations of RKO cells infected withlentivirus encoding shNS or sh-b-catenin were treated withWnt CM or control CM for 1 hour. Treated cells were subjected to either immunoblot analysis by theindicated antibodies (E) or ChIP analysis with the indicated antibodies and primers for the Axin2 WRE as described in Materials and Methods (F). Resultsshown for all ChIP assays in Fig. 6 are the mean and range of variation for duplicate PCR reactions from a single experiment and are representative of 2independent experiments.
Ou et al.
Mol Cancer Res; 9(5) May 2011 Molecular Cancer Research668
on June 21, 2018. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst April 8, 2011; DOI: 10.1158/1541-7786.MCR-10-0223

many transcription factors, including nuclear receptors andAP1 (22, 23), and the S100A4 gene is reported to be underthe control of transcription factor NFAT5 (nuclear factor ofactivated T cells 5; ref. 24).Since CARM1 works as a coactivator for multiple tran-
scription factors and Wnt target genes are controlled byWnt/b-catenin signaling and transcription factors regulatedby other signaling cascades (13, 25), it is difficult to assesswhether the requirement of CARM1 for constitutively highexpression of Wnt target genes in cells with deregulatedWnt/b-catenin signaling is due to CARM1 actions throughLEF1/b-catenin or other transcription factors that bind tothe same promoter. To address this problem, we identified acell line in which we could observe Wnt-inducible geneexpression. After screening a number of colon cancer celllines, we identified RKO as a colon cancer cell line that hasnormal Wnt/b-catenin signaling. Importantly, in RKOcells, the induction of Axin2 and GPR49 transcriptionby Wnt3a ligands was blocked by CARM1 depletion(Fig. 6B), andWnt3a ligand caused recruitment of CARM1to the WRE of the Axin2 promoter, resulting in enhancedmethylation of Arg 17 of histone H3 in nucleosomesassociated with the WRE (Fig. 5C). Together, these resultsindicate that CARM1 acts directly at the Axin2 promoter tomediate the actions of LEF1/b-catenin in response to Wntsignaling. Therefore, CARM1 is a bona fide coactivator forWnt/b-catenin signaling.Our results have also begun to dissect the mechanism of
CARM1 action. Depletion of b-catenin from RKO cellsdiminished Wnt-induced occupancy of the Axin2 WRE byCARM1, thus showing that CARM1 recruitment inresponse toWnt3a depends upon the presence of b-catenin.In contrast, depletion of CARM1 levels did not diminishWnt-induced accumulation of b-catenin in the cells or therecruitment of b-catenin to the Wnt target gene; instead,CARM1 depletion reduced the methylation of Arg 17 onhistone H3 at the WRE (Fig. 6D). These findings indicatethat mechanistically, CARM1 acts downstream fromb-catenin and facilitates transcription complex formationby facilitating steps that are downstream from the recruit-ment of b-catenin and CARM1 to the promoter. In theLEF1/b-catenin reporter gene system, methyltransferaseactivity of CARM1 was not required for its transcriptionalcoactivator activity (Supplementary Fig. S1). The C-term-inal activation domain of CARM1 may be responsible forthe coactivator function of CARM1 in this case (26). In
addition, the importance of CARM1 enzymatic activity inthe expression of endogenous Wnt target genes should betested, given that: (i) plasmid-based reporter gene studiesmay omit the contribution of core histone and its tail to theregulation of transcriptional activation, and (ii) previousreports have indicated that arginine-specific histone methy-lation by CARM1 is a significant part of the transcriptionalactivation process for some genes.Abnormal expression of CARM1 has been linked to
human prostate (20, 27), breast (28), and colorectal cancers(29). In this report, we showed that CARM1 protein levelsare higher in a panel of colorectal cancer cell lines than in acell line that more closely resembles normal colon epithelialcells (Fig. 3A). Moreover, CARM1 silencing adverselyaffected clonal survival and anchorage-independent growthof colorectal cancer cell lines with constitutively high levelsof b-catenin (Fig. 4 and Supplementary Fig. S5). TheseCARM1-dependent phenotypes are presumably due to thefact that CARM1 depletion causes reduced expression ofgenes involved in Wnt-driven tumorigenesis (e.g., c-Myc;ref. 5), metastasis (e.g., S100A4; ref. 8), and prevention ofapoptosis (e.g., GPR49; ref. 10). Thus, CARM1 plays a keyrole in Wnt signaling through its role as a transcriptionalcoactivator that mediates the actions of b-catenin on Wnttarget genes; some of the Wnt target genes that requireCARM1 are critical for maintaining important aspects ofthe transformed phenotype of colon cancer cell lines. There-fore, selective inhibition of CARM1 methyltransferaseactivity or CARM1 binding to b-catenin may be a potentialstrategy for therapeutic treatment of abnormally activatedWnt/b-catenin signaling in colorectal cancers.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
We thank Ms. K. Chang for expert technical assistance.
Grant Support
This work was supported by grants DK055274 to M.R. Stallcup and CA20535 toK.R. Yamamoto from the National Institutes of Health.
Received May 21, 2010; revised February 25, 2011; accepted March 21, 2011;published OnlineFirst April 8, 2011.
References1. Polakis P. Wnt signaling and cancer. Genes Dev 2000;14:1837–51.2. Reya T, Clevers H. Wnt signalling in stem cells and cancer. Nature
2005;434:843–50.3. MacDonald BT, Tamai K, He X. Wnt/beta-catenin signaling: compo-
nents, mechanisms, and diseases. Dev Cell 2009;17:9–26.4. Klaus A, Birchmeier W. Wnt signalling and its impact on development
and cancer. Nat Rev Cancer 2008;8:387–98.5. He TC, Sparks AB, Rago C, Hermeking H, Zawel L, da Costa LT, et al.
Identification of c-MYC as a target of the APC pathway. Science1998;281:1509–12.
6. Zhang T, Otevrel T, Gao Z, Ehrlich SM, Fields JZ, Boman BM.Evidence that APC regulates survivin expression: a possible mechan-ism contributing to the stem cell origin of colon cancer. Cancer Res2001;61:8664–7.
7. Kim PJ, Plescia J, Clevers H, Fearon ER, Altieri DC. Survivin andmolecular pathogenesis of colorectal cancer. Lancet 2003;362:205–9.
8. Stein U, Arlt F, Walther W, Smith J, Waldman T, Harris ED, et al. Themetastasis-associated gene S100A4 is a novel target of beta-catenin/T-cell factor signaling in colon cancer. Gastroenterology 2006;131:1486–500.
CARM1 and b-Catenin in Aberrant Growth of Colon Cancer Cells
www.aacrjournals.org Mol Cancer Res; 9(5) May 2011 669
on June 21, 2018. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst April 8, 2011; DOI: 10.1158/1541-7786.MCR-10-0223

9. Barker N, van Es JH, Kuipers J, Kujala P, van den Born M, CozijnsenM, et al. Identification of stem cells in small intestine and colon bymarker gene Lgr5. Nature 2007;449:1003–7.
10. McClanahan T, Koseoglu S, Smith K, Grein J, Gustafson E, Black S,et al. Identification of overexpression of orphan G protein-coupledreceptor GPR49 in human colon and ovarian primary tumors. CancerBiol Ther 2006;5:419–26.
11. Bedford MT, Clarke SG. Protein arginine methylation in mammals:who, what, and why. Mol Cell 2009;33:1–13.
12. Lee YH, Stallcup MR. Minireview: protein arginine methylation ofnonhistone proteins in transcriptional regulation. Mol Endocrinol2009;23:425–33.
13. Koh SS, Li H, Lee YH, Widelitz RB, Chuong CM, Stallcup MR.Synergistic coactivator function by coactivator-associated argininemethyltransferase (CARM) 1 and beta-catenin with two differentclasses of DNA-binding transcriptional activators. J Biol Chem2002;277:26031–5.
14. Ou CY, Kim JH, Yang CK, Stallcup MR. Requirement of cell cycle andapoptosis regulator 1 for target gene activation by Wnt and beta-catenin and for anchorage-independent growth of human coloncarcinoma cells. J Biol Chem 2009;284:20629–37.
15. LaBonte MJ, Manegold PC, Wilson PM, Fazzone W, Louie SG, LenzHJ, et al. The dual EGFR/HER-2 tyrosine kinase inhibitor lapatinibsensitizes colon and gastric cancer cells to the irinotecan activemetabolite SN-38. Int J Cancer 2009;125:2957–69.
16. Hecht A, Vleminckx K, Stemmler MP, van Roy F, Kemler R. Thep300/CBP acetyltransferases function as transcriptional coacti-vators of beta-catenin in vertebrates. EMBO J 2000;19:1839–50.
17. Sun Y, Kolligs FT, Hottiger MO, Mosavin R, Fearon ER, Nabel GJ.Regulation of beta -catenin transformation by the p300 transcrip-tional coactivator. Proc Natl Acad Sci U S A 2000;97:12613–8.
18. Roose J, Huls G, van Beest M, Moerer P, van der Horn K,Goldschmeding R, et al. Synergy between tumor suppressorAPC and the beta-catenin-Tcf4 target Tcf1. Science 1999;285:1923–6.
19. Mosimann C, Hausmann G, Basler K. Beta-catenin hits chromatin:regulation of Wnt target gene activation. Nat Rev Mol Cell Biol2009;10:276–86.
20. Hong H, Kao C, Jeng MH, Eble JN, Koch MO, Gardner TA, et al.Aberrant expression of CARM1, a transcriptional coactivator of andro-gen receptor, in the development of prostate carcinoma and andro-gen-independent status. Cancer 2004;101:83–9.
21. Frietze S, Lupien M, Silver PA, Brown M. CARM1 regulates estrogen-stimulated breast cancer growth through up-regulation of E2F1.Cancer Res 2008;68:301–6.
22. Iavarone C, Catania A, Marinissen MJ, Visconti R, Acunzo M, Tar-antino C, et al. The platelet-derived growth factor controls c-mycexpression through a JNK- and AP-1-dependent signaling pathway. JBiol Chem 2003;278:50024–30.
23. Dubik D, Shiu RP. Transcriptional regulation of c-myc oncogeneexpression by estrogen in hormone-responsive human breast cancercells. J Biol Chem 1988;263:12705–8.
24. Chen M, Sinha M, Luxon BA, Bresnick AR, O’Connor KL. Integrinalpha6beta4 controls the expression of genes associated with cellmotility, invasion, and metastasis, including S100A4/metastasin. JBiol Chem 2009;284:1484–94.
25. Chesnutt C, Burrus LW, Brown AM, Niswander L. Coordinate regula-tion of neural tube patterning and proliferation by TGFbeta and WNTactivity. Dev Biol 2004;274:334–47.
26. Teyssier C, Chen D, StallcupMR. Requirement for multiple domains ofthe protein arginine methyltransferase CARM1 in its transcriptionalcoactivator function. J Biol Chem 2002;277:46066–72.
27. Majumder S, Liu Y, Ford OH III, Mohler JL, Whang YE. Involvement ofarginine methyltransferase CARM1 in androgen receptor function andprostate cancer cell viability. Prostate 2006;66:1292–301.
28. El Messaoudi S, Fabbrizio E, Rodriguez C, Chuchana P, Fauquier L,Cheng D, et al. Coactivator-associated arginine methyltransferase 1(CARM1) is a positive regulator of the Cyclin E1 gene. Proc Natl AcadSci U S A 2006;103:13351–6.
29. Kim YR, Lee BK, Park RY, Nguyen NT, Bae JA, Kwon DD, et al.Differential CARM1 expression in prostate and colorectal cancers.BMC Cancer; 10:197.
Ou et al.
Mol Cancer Res; 9(5) May 2011 Molecular Cancer Research670
on June 21, 2018. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst April 8, 2011; DOI: 10.1158/1541-7786.MCR-10-0223

2011;9:660-670. Published OnlineFirst April 8, 2011.Mol Cancer Res Chen-Yin Ou, Melissa J. LaBonte, Philipp C. Manegold, et al. Expression-Catenin Activity in Colorectal Cancer Cell Growth and Gene
βA Coactivator Role of CARM1 in the Dysregulation of
Updated version
10.1158/1541-7786.MCR-10-0223doi:
Access the most recent version of this article at:
Material
Supplementary
http://mcr.aacrjournals.org/content/suppl/2011/05/17/1541-7786.MCR-10-0223.DC1
Access the most recent supplemental material at:
Cited articles
http://mcr.aacrjournals.org/content/9/5/660.full#ref-list-1
This article cites 28 articles, 14 of which you can access for free at:
Citing articles
http://mcr.aacrjournals.org/content/9/5/660.full#related-urls
This article has been cited by 8 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
SubscriptionsReprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. (CCC)Click on "Request Permissions" which will take you to the Copyright Clearance Center's
.http://mcr.aacrjournals.org/content/9/5/660To request permission to re-use all or part of this article, use this link
on June 21, 2018. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst April 8, 2011; DOI: 10.1158/1541-7786.MCR-10-0223





























