Embed Size (px)
Citation preview

Models and Technologies
A Novel Fully Humanized 3D Skin Equivalent toModel Early Melanoma InvasionDavid S. Hill1, Neil D.P. Robinson2, Matthew P. Caley3, Mei Chen4, Edel A. O'Toole3,Jane L. Armstrong1,5, Stefan Przyborski2, and Penny E. Lovat1
Abstract
Metastatic melanoma remains incurable, emphasizing theacute need for improved research models to investigate theunderlying biologic mechanisms mediating tumor invasion andmetastasis, and to develop more effective targeted therapies toimprove clinical outcome. Available animalmodels ofmelanomadonot accurately reflect humandisease and current in vitrohumanskin equivalent models incorporating melanoma cells are notfully representative of the human skin microenvironment. Wehave developed a robust and reproducible, fully humanizedthree-dimensional (3D) skin equivalent comprising a stratified,
terminally differentiated epidermis and a dermal compartmentconsisting of fibroblast-generated extracellular matrix. Melanomacells incorporated into the epidermis were able to invade throughthe basement membrane and into the dermis, mirroring earlytumor invasion in vivo. Comparison of our novel 3D melanomaskin equivalent with melanoma in situ and metastatic melano-ma indicates that this model accurately recreates features ofdisease pathology, making it a physiologically representativemodel of early radial and vertical growth-phase melanomainvasion. Mol Cancer Ther; 14(11); 2665–73. �2015 AACR.
IntroductionCutaneous metastatic melanoma remains one of the most
deadly forms of cancer, with a rapidly increasing incidence,mortality, and public health burden. Although early-stage mela-noma is largely curable through surgical resection, continued5-year survival rates of only 5% to 19% for advanced disease(1) reflect the lack of consistently beneficial treatments for met-astatic melanoma. Improved research models are thereforeurgently needed to investigate the underlying biologic mechan-ismsmediating tumor invasion and subsequentmetastasis, and tofacilitate the development of more effective targeted therapies toimprove clinical outcome.
Human skin comprises an upper epidermal layer containingmainly keratinocytes in close association withmelanocytes, and alower dermal layer containing multiple cell types, includingfibroblasts that synthesize extracellular matrix (ECM) compo-nents to support cellular growth (2). Keratinocytes form a pro-
liferative basal layer and differentiate as they move toward thesurface of the skin, whereas melanocytes, the precursor cells ofmelanoma, proliferate less frequently and remain at the epider-mal–dermal junction where they interact with basal layer kerati-nocytes to regulate tanning of the skin in response toUV radiation(3). A basement membrane, composed of matrix molecules,including laminin isoforms and type IV, VII, and XVII collagens,separatemelanocytes and keratinocytes from the papillary dermis(4). However, when melanocytes become transformed, hyper-proliferative and migratory melanoma cells invade through thebasement membrane into the dermis. Therefore, models that aimto investigate early melanoma development must recreate themicroenvironment of this distinct cellular niche (5).
Although mouse xenograft models of melanoma in immu-nocompromised mice are commonly used to investigate tumordevelopment, progression, and therapeutic response, they donot accurately recreate the microenvironment of human mel-anoma at either the primary or distant site. As such, thesemodels cannot recapitulate the initial events leading to earlyinvasion through the basement membrane or dissemination ofmelanoma cells throughout the skin and to subsequent met-astatic sites. Furthermore, although spontaneous mouse mel-anoma models (6–8) are useful for investigating the earlystages of mouse melanoma development, significant differ-ences between the architecture of human and rodent skin (9),as well as differences observed in the histopathologic featuresof human and murine melanoma subtypes (10) make itdifficult to extrapolate results from these studies into a clin-ically relevant context.
To more accurately investigate early-stage human melanoma,full-thickness in vitro skin equivalent models incorporating mel-anoma cells have been developed, which allow investigation ofmelanoma migration and invasion from the epidermis into thedermis (11–14). However, such equivalents comprise a dermalcomponent created from fibroblasts embedded in bovine orrat-tail collagen, which as well as contracting over time leadingto distortion and disruption of the equivalent, are not
1Dermatological Sciences, Institute of Cellular Medicine, NewcastleUniversity, Newcastle-upon-Tyne,UnitedKingdom. 2School of Biolog-ical and Biomedical Sciences, Durham University, Durham, UnitedKingdom. 3Centre for Cutaneous Research, Barts and the LondonSMD, Queen Mary University of London, Blizard Institute, London,United Kingdom. 4Norris Comprehensive Cancer Centre, Universityof Southern California, Los Angeles, California. 5Faculty of AppliedSciences, University of Sunderland, Sunderland, United Kingdom.
Note: Supplementary data for this article are available at Molecular CancerTherapeutics Online (http://mct.aacrjournals.org/).
P.E. Lovat and S. Przyborski are joint senior authors.
Corresponding Authors: Penny Lovat, Newcastle University, DermatologicalSciences, Institute of Cellular Medicine, The Medical School, Framlington Place,Newcastle-upon-Tyne NE2 4HH, United Kingdom. Phone: 44-191-2227170;E-mail: [email protected]; Stefan Przyborski, School of Biological andBiomedical Sciences, Durham University, South Road, Durham DH1 3LE, UnitedKingdom. Phone: 44 191 3343988; E-mail: [email protected]
doi: 10.1158/1535-7163.MCT-15-0394
�2015 American Association for Cancer Research.
MolecularCancerTherapeutics
www.aacrjournals.org 2665
on June 24, 2020. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst September 1, 2015; DOI: 10.1158/1535-7163.MCT-15-0394

representative of the normal human skin microenvironment asthey include nonhuman ECM components. Alternatively,although decellularized human skin models offer a human skinmicroenvironment, variability between donors results in incon-sistent melanoma migration, which affects the reproducibility ofthese assays (15).
This study describes a novel in vitromodel for the investigationof early melanoma invasion, such as that which occurs in radialand vertical growth phase melanoma, within a fully humanizedcutaneous microenvironment. We have developed a unique full-thickness three-dimensional (3D) skin equivalent (organotypicskin culture) through the incorporation of an inert porous scaf-fold (16) with appropriate pore sizes to support the 3D growthand cell–cell contact of primary human dermal fibroblasts. Fibro-blasts are stimulated to produce their own ECM constituents(17, 18), forming a stable dermal component that is physiolog-ically representative of normal human skin. Following addition ofprimary human keratinocytes, cross-talk between fibroblasts andkeratinocytes facilitates the development of a permissive micro-environment conducive to long-term culture (19). This is consis-tent with previous studies showing the stratum corneum of skinequivalents formed on fibroblast-derived matrix contains a con-siderably higher concentration of natural moisturizing factorcompared with animal collagen-based skin equivalents, thusallowing cultures to be maintained for up to 20 weeks (20).
Seedingmelanoma cells onto thedermal equivalent prior to theincorporation of primary human keratinocytes, rather thanimplanting melanoma spheroids directly into the dermis (21),or suspending melanoma cells in hydrogel (22), places themelanoma cells in their original microenvironmental niche with-in the skin, resulting in subsequent proliferation and nest forma-tion at the epidermal–dermal junction prior to invasion throughthe basement membrane. We demonstrate that active invasion ofmelanoma cells results in breakdown of basement membranecomponents type IV and VII collagens, accurately recapitulatingthe pattern of early melanoma invasion observed in humancutaneous tumors in vivo, thus providing a valuable tool toinvestigatemechanismsmediatingmelanoma initiation and earlystages of disease progression.
Materials and MethodsCell culture
Primary human neonatal foreskin fibroblasts (CellnTec) werecultured in Media A (Table 1) for up to seven passages. Immor-talized mouse embryonic 3T3 fibroblasts (ATCC-CCL-92) werecultured in Media D. Following informed consent, primaryhuman keratinocytes derived from surplus skin obtained frompatients (ages between 20 and 55 years) undergoing routinesurgery (for which full ethical approval was obtained; NationalResearch Ethics reference, Newcastle and North Tyneside 108/H0906195 for all studies with human tissue) were isolatedby incubating the skin in dispase (Scientific Laboratory Supplies)for 12 to 18 hours at 4�C to separate the epidermis from thedermis before dissociating the epidermis with trypsin/EDTA(Scientific Laboratory Supplies; ref. 23) for 5 minutes at 37�Cand subsequently cultured in Media E for up to two passages.Keratinocytes were then further cocultured with mitomycin C(Sigma-Aldrich) treated 3T3 feeder cells (24) at 1:1 ratio inMediaB (based on ref. 25) for up to three passages, changing the mediaevery day. Following detachment with trypsin/EDTA, keratino-
cytes were subsequently incubated with an equal volume ofsoybean trypsin inhibitor (Sigma-Aldrich) and centrifuged at300 � g for 5-minutes prior to resuspension in fresh culturemedia and subsequent culture. The humanmetastatic melanomacell line SK-mel-28 (LGC Standards; ATCC-HTB-72) and theprimary human melanoma cell line WM35 (Coriell Cell Repos-itories) were obtained in 2011 and are tested every 6 months forMelan-A expression by immunofluorescence, with BRAF muta-tional status confirmed by real-time PCR (26), and cultured inMedia A as previously described (27). All cells were cultured at37�C in a humidified atmosphere with 5% CO2 in air.
Human skin equivalent preparationTwelve-well formatAlvetex scaffolds (Reinnervate Ltd.,Reprocell
Group) were pretreated with 70% ethanol in a 6-well plate accord-ing to the manufacturer's instructions. A total of 2.0� 106 primaryhuman neonatal foreskin fibroblasts were seeded onto Alvetex in100mLMediaAand incubatedat37�C, inahumidifiedatmosphereof5%CO2 inair for1.5hours.NinemillilitersofMediaAþ100mg/mL ascorbic acid (Sigma-Aldrich)were subsequently applied to thebottomofeachwell to gentlyflood the insert prior to incubation fora further 18 days, changing media every 3.5 days, to allow theformation of a dermal equivalent. Dermal equivalents were sub-sequently washed with 10 mL phosphate-buffered saline (PBS;Sigma-Aldrich) prior to the addition of 4 mL Media B to the outerside of the insert such that the bottom of each dermal equivalentwas in contact with the media. To establish a melanoma 3Dequivalent, 2.0 � 104 melanoma cells were applied to the dermalequivalent in100mLMediaB and incubationat 37�Ccontinued fora further 3 hours. In the meantime, primary human keratinocyteswere harvested by differential trypsinization, discarding the 3T3feeder cells, and 2.0 � 106 keratinocytes seeded onto dermalequivalents (with or without melanoma cells) in 100 mL MediaB and incubation continued for a further 3 hours. Fivemilliliters ofMedia B was then applied to the outer side of each well to gentlyflood the inside of the insert prior to further incubation at 37�C for3 days, changing the media every day. On day 21, the insert wasremoved from the 6-well plate and placed into a well insert holderin a deep petri dish (Reinnervate Ltd., Reprocell Group) on themiddle rung of the stand. Thirty milliliters of Media C was thenadded to the dish, such that the bottom of the equivalent was incontact with the media but the upper surface remained exposed tothe air and incubation continued at 37�C in 5% CO2 for 14 days,changing themedia every 3.5 days, to allow the formation of a full-thickness skin equivalent.
Scanning electron microscopySkin equivalent or primary tissue samples of normal human
skin were fixed in a 1:1 mix of DMEMmedia and double strengthfixation buffer [16% PFA, Sigma-Aldrich; 25% glutaraldehyde,Agar Scientific; 0.2mol/L sodium cacodylate, Agar Scientific) for 5to 10 minutes at room temperature. Samples were then trans-ferred to a new tube and incubated in single-strength fixationbuffer (8% PFA, 12.5% glutaraldehyde, 0.1 mol/L sodium caco-dylate) at 4�C for 1 hour prior to washing in PBS 3 time for 5minutes each. Samples were subsequently cut into 2- to 3-mm2
squares and immersed in postfixation buffer [1% osmium tetrox-ide (Agar Scientific)) in 0.1 mol/L sodium cacodylate] at 4�C for60minutes beforewashing in 0.1mol/L sodium cacodylate buffertwice for 10minutes each. Following dehydration through a seriesof ethanol washes (30%, 50%, 70%, 80%, 90%, 95%, and 100%)
Hill et al.
Mol Cancer Ther; 14(11) November 2015 Molecular Cancer Therapeutics2666
on June 24, 2020. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst September 1, 2015; DOI: 10.1158/1535-7163.MCT-15-0394

each for 15minutes, samples were then dried using a critical pointdryer (Baltec CPD030; Pf€affikon ZH), coated in 5 nmof platinumusing a Cressington Coating System 328 (Cressington ScientificInstruments) and visualized using a Leica S5200 scanning elec-tron microscope (Leica Microsystems).
Immunofluorescent analysis of skin biomarkersFormalin-fixed, paraffin-embedded primary human tissue
samples derived from an in situ melanoma or an AJCC stage IVmetastatic melanoma were used as a comparative to 3D humanmelanoma skin equivalents. All samples were processed forhematoxylin and eosin staining or immunohistochemistry, aspreviously described (28, 29). Sections (5 mm) were incubatedwith 1:1,000mouse antihuman type III collagen (kindly suppliedby Dr. Rachel Watson, Manchester University, Manchester, UK;Abcam; ab23445), 1:1,000 mouse anti-type IV collagen (Abcam;ab6586), 1:400 rabbit anti-type VII collagen (kindly supplied byDr.MeiChen,Norris ComprehensiveCancerCentre,University ofSouthernCalifornia, Los Angeles, CA; ref. 30), 1:1,000 rabbit anti-cytokeratin 1 (Abcam; ab93652), 1:1,000mouse anti-cytokeratin
14 (Abcam; ab7800), 1:1,000 mouse anti-involucrin (Abcam;ab68), or 1:250 mouse anti-Melan-A (Abcam; ab731) primaryantibodies diluted in PBS þ 5% BSA overnight at 4�C. Primaryantibody binding was detected with secondary Alexa Fluor 488goat anti-mouse (Life Technologies) or Alexa Fluor 488 goat anti-rabbit antibodies (Life Technologies) and cell nuclei counterstained with DAPI (1 mg/mL; Life Technologies) diluted in PBSþ 5% BSA for 1 hour at room temperature. Sections were finallymounted under glass coverslips in Vectorshield mounting media(Vector Laboratories) and visualized using either a LeicaDMI3000B (Leica Microsystems) or an Axioimager Z2 (Carl ZeissLtd.).
ResultsGeneration of a full-thickness human skin equivalent
Alvetex porous polymer scaffolds were used to create a full-thickness human skin equivalent in the absence of any animalmatrix components (Fig. 1). Pretreatment of Alvetex by immer-sion in 70% ethanol rendered it hydrophilic allowing media and
Table 1. Media components for the production of full-thickness melanoma skin equivalents
Media Component (stock conc.) Volume or weight Final concentration
A Dulbecco's Modified Eagle Medium^ 500 mL —
Fetal calf serum& 50 mL �10%Penicillin/streptomycin/amphotericin^ 5 mL �1%
B Dulbecco's Modified Eagle Medium^ 375 mL —
Ham's F12 nutrient mixture& 125 mL —
Chelex-treated� fetal calf serum& 25 mL �5%Cholera toxin (0.85 mg/mL)& 5 mL 8.5 ng/mLHydrocortisone (0.5 mg/mL)& 400 mL 0.4 mg/mLInsulin (10 mg/mL)& 250 mL 5 mg/mLAdenine (6 mg/mL)& 2 mL 24 mg/mLRecombinant human epidermal growth factor (0.2 mg/mL)& 50 mL 10 ng/mLPenicillin/streptomycin/amphotericin^ 5 mL �1%
C Dulbecco's Modified Eagle Medium^ 375 mL —
Ham's F12 nutrient mixture& 125 mL —
Fetal calf serum& 50 mL �10%Cholera toxin (0.85 mg/mL)& 5 mL 8.5 ng/mLHydrocortisone (0.5 mg/mL)& 400 mL 0.4 mg/mLRecombinant human epidermal growth factor (0.2 mg/mL)& 50 mL 10 ng/mLTransferin (10 mg/mL)& 250 mL 5 mg/mLPenicillin/streptomycin/amphotericin^ 5 mL �1%L-ascorbic acid (10 mg/mL) added fresh& 100 mL per 10 mL media 100 mg/mL
D Dulbecco's Modified Eagles Medium^ 500 mL —
Iron-fortified newborn calf serum& 25 mL �5%Penicillin/streptomycin/amphotericin^ 5 mL �1%
E MCDB153 with L-glutamine, 28 mmol/L HEPES& 500 mL —
Histidine& 18.65 mg 240 mmol/LIsoleucine& 49.2 mg 750 mmol/LMethioninen& 6.7 mg 90 mmol/LPhenylalanine& 7.45 mg 90 mmol/LTryptophan& 4.6 mg 45 mmol/LTyrosine& 9.775 mg 100 mmol/LEthanolamine (98% w/v)& 3.05 mL 100 mmol/LPhosphorylethanolamine& 7.05 mg 100 mmol/LCalcium chloride (1 mol/L)& 20 mL 40 mmol/LSodium bicarbonate& 588 mg 15 mmol/LHuman keratinocyte growth supplement� 5 mL �1%Penicillin/streptomycin/amphotericin^ 5 mL �1%Adjust to pH 7.2
NOTE: Volumeorweight andfinal concentration of each component inMedia A–E required for producing full-thicknessmelanoma skin equivalents (seeMaterials andMethods for use of each media). The suppliers of each component are indicated by superscripts:^, Scientific Laboratory Supplies Ltd.;&, Sigma-Aldrich CompanyLtd.;�, Life Technologies Ltd.; � , Bio-Rad Laboratories Ltd.
Cutaneous Melanoma Invasion in Human 3D Skin Equivalents
www.aacrjournals.org Mol Cancer Ther; 14(11) November 2015 2667
on June 24, 2020. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst September 1, 2015; DOI: 10.1158/1535-7163.MCT-15-0394

cells to enter the 3D matrix. Alvetex scaffolds were subsequentlywashedwith culturemedia to remove the ethanol and seededwithprimary human neonatal foreskin fibroblasts, prior to culture for18 days in Media A (Table 1) supplemented with ascorbic acid topromote synthesis of collagen polypeptides through the proces-sing of procollagens to collagen a-chains (17). Primary humankeratinocytes isolated from the epidermis of normal human skinwere then seeded onto the upper surface and cultured for 3 days inMedia B. The upper surface was subsequently exposed at the air–liquid interface for 14 days to induce keratinocyte differentiation,whereas the lower surface remained in contact with Media C,resulting in the formation of a full-thickness human skinequivalent.
Cell numbers, media components, and time intervals for eachstep of the protocol were optimized to allow full scaffold colo-nization by dermal fibroblasts (Fig. 2A) and the establishment ofan intact, fully stratified epidermis with keymorphologic featuresof a stratum basale, stratum spinosum, and stratum corneum(Fig. 2B, 20� magnification; Fig. 2C, 10� magnification). Elec-
tronmicrographs indicate the structure andporosity of theAlvetexscaffold membrane (Fig. 2D), supporting fibroblast growth inthree dimensions and facilitating the establishment of a full-thickness human skin equivalent (Fig. 2E)with clearmorphologicsimilarities to normal human skin (Fig. 2F).
Primary human keratinocytes and fibroblasts in organotypicculture form a humanized skin microenvironment
Normal human skin comprises a dermal layer and a multilay-ered epidermis, each layer of which displays a distinct proteinexpression profile (Fig. 3A). The dermis contains extracellularmatrix components, including type I and III collagens, whereasthe epidermis is characterized by the expression of various cyto-keratins that are differentially regulated within different layers ofthe epidermis, reflecting the progressive stages of normal humankeratinocyte differentiation.Histologic analysis of our established3D skin equivalent (Fig. 3B) demonstrated morphologic similar-ities to that of normal human skin (Fig. 3C), and a comparativecommercially available model (Mattek EpidermFT; Fig. 3D); inparticular, the presence of a fully developed stratum corneumwas evident, indicating keratinocyte differentiation and barrierformation.
Immunofluorescent staining also revealed the expression ofhuman type III, IV, and VII collagens, cytokeratin 1 and 14 as wellas involucrin (Fig. 3), to varying degrees in the 3D skin equivalent,normal human skin, and the Mattek EpidermFT. Dermal fibro-blasts contained within the 3D skin equivalent for 35 days clearlyexpressed type III collagen (Fig. 3T),which, albeit not as abundantas expression observed in normal human skin (Fig. 3U), never-theless indicated the production of human extrcellular matrix,critical to the long-term maintenance of the skin equivalent. Incontrast, however, less human type III collagen expression wasobserved in the Mattek EpidermFT (Fig. 3V), likely due to theirconstruction mainly being based on the use of bovine type Icollagen thatmay suppress further ECMproduction by the dermalfibroblasts. The 3D skin equivalent model also demonstratedproduction of human type I collagen (data not shown).
The basement membrane components type IV and VII col-lagenswere clearly expressed at the epidermal–dermal junction ofboth the 3D skin equivalent (Fig. 3N and Q) and normal humanskin (Fig. 3O and J), indicating an interaction between fibroblastsand keratinocytes and synthesis of a de novo basementmembrane.However, although expression of type IV collagen was partiallyobserved between the epidermal and dermal layers within theMattek EpidermFTmodel, therewas noevidence for theorganizedexpression of type VII collagen (Fig. 3P and S). It is possible,however, that the Mattek EpidermFT model may not have beencultured for sufficient time to enable type VII collagen organiza-tion and the formation of a basement membrane comparable tonormal skin (31).
Expression of cytokeratin 14 by keratinocytes within the 3Dskin equivalent also indicated the formation of a stratum basale(Fig. 3K), resembling that of normal human skin (Fig. 3L).Keratinocytes within the 3D skin equivalent appeared to undergonormal differentiation as demonstrated by the expression ofcytokeratin 1 and involucrin in suprabasal and terminal layerkeratinocytes, indicative of stratum spinosum and stratum gran-ulosum formation, respectively (Fig. 3H and E), and again indic-ative of the pattern of epidermal differentiation observed innormal human skin (Fig. 3I and F). Furthermore, althoughexpression of cytokeratin 14 (Fig. 3M) was observed in Mattek
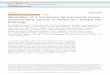
Pretreat Alvetex
Seed human dermal fibroblasts
Seed human melanoma cells
Seed human epidermalkeratinocytes
Media A
Media B
Media B
Media C
Media A
18 days
3 days
14 days
3 hours18 days
Raise to air/liquid interface
Formation of full-thickness3D (melanoma) skin equivalent
Figure 1.Schematic protocol for the formation of full-thickness human skinequivalents. Pretreat inert Alvetex polymer scaffold in 70% ethanol beforethoroughly washing in Media A, then seed with 5.0 � 105 human dermalfibroblasts. Culture fibroblast-seeded Alvetex in Media A for 18 days to createa dermal equivalent. If establishing amelanoma full-thickness skin equivalent,seed 2.0 � 104 metastatic melanoma cells onto the dermal equivalent andculture in Media B for 3 hours prior to the addition of primary humankeratinocytes. Alternatively, to create a full-thickness skin equivalent, add2.0 � 106 keratinocytes directly onto the dermal equivalent. Culture the(melanoma) equivalent fully submerged in Media B for 3 days beforeexposing the upper surface of the equivalent to the air–liquid interphase andcontinuing culture for 14 days with the lower surface in contact with MediaC (see Materials and Methods for full protocol).
Hill et al.
Mol Cancer Ther; 14(11) November 2015 Molecular Cancer Therapeutics2668
on June 24, 2020. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst September 1, 2015; DOI: 10.1158/1535-7163.MCT-15-0394

EpidermFT, cytokeratin I (Fig. 3J), and involucrin (Fig. 3G)expression was less well defined, indicating formation of a stra-tum basale but ineffective keratinocyte differentiation in thismodel. The establishment of an organotypic skin equivalent onAlvetex scaffolds therefore accurately recreates the microenviron-ment of normal human skin. This was subsequently used toinvestigate melanoma cell behavior in vitro.
Melanoma cell invasion through the basement membrane offully humanized 3D skin equivalents recreates the progressivehistopathologic features of melanoma invasion in human skin
The potential for human melanoma cell lines derived fromeither primary ormetastatic tumors to invade the pore structure ofAlvetex scaffolds was verified in the absence of primary fibroblasts(data not shown). To model melanoma invasion, metastaticmelanoma cells were applied to preestablished fibroblast-con-taining Alvetex dermal equivalents prior to the incorporation ofkeratinocytes at a slightly lower ratio (100:1) to the physiologicratio of keratinocytes-to-melanocytes in normal human skin(36:1; ref. 32), in order to prevent tumor cell over growth withinthe epidermis prior to the observation of dermal invasion. His-tologic staining of a 3D skin equivalent 2 weeks after incorpo-ration with metastatic SK-mel-28 melanoma cells demonstratedthe development of melanoma nests at the epidermal–dermaljunction (Fig. 4A), verified by the expression of the melanocytelineage-specific marker Melan-A (Fig. 4E). Immunofluorescentstaining for the human basement membrane components type IVcollagen (Fig. 4I) and type VII collagen (Fig. 5A) 2 weeks after theincorporation ofmelanoma cells into the skin equivalent revealedintact expression of both markers and clear localization of mel-
anoma cells above both type IV (Fig. 4M and Q) and type VIIcollagens (Fig. 5E and I). However, culture for a further 2 weeksresulted in the invasion of Melan-A–positive melanoma cells intothe dermal component (Fig. 4B and F), accompanied by disrup-tion of type IV collagen (Fig. 4J) and loss of type VII collagen(Fig. 5B). Furthermore, costaining for Melan-A and either type IVcollagen (Fig. 4N and R) or type VII collagen (Fig. 5F and J)demonstrated disruption of these basement membrane compo-nents coincided with melanoma invasion, indicating SK-mel-28melanoma cells actively invade from the epidermis into thedermis of the skin equivalent through the basement membrane.Similar results were also obtained with skin equivalents incorpo-rating Melan-A–positive primary WM35 melanoma cells, whereagain tumor invasion through the basement membrane, albeitless than metastatic SK-mel-28, was observed with a concurrentdisruption of type IV collagen at 4weeks (Supplementary Fig. S1).
To validate whether invasion of melanoma cells within a 3Dskin equivalent accurately reflects the progressive stages of clinicaldisease, the effect of melanoma cells on type IV and VII collagenswas investigated in a formalin-fixed paraffin-embedded in situmelanoma or in a primary tumor derived from a patient withmetastatic disease. Histologic staining and Melan-A immunos-taining of the melanoma in situ (Fig. 4C and G) confirmed aminimally invasive tumor accompanied by continuous and intactexpression of both type IV (Fig. 4K) and type VII collagens(Fig. 5C) at the epidermal–dermal junction. Co-immunostainingdemonstrated that in preinvasive melanomas, cells are locatedabove type IV (Fig. 4O and S) and type VII collagens (Fig. 5G andK), indicating an intact basement membrane that reflects thehistopathologic features observed in 3D skin equivalents 2 weeks
Figure 2.Validation of dermal and epidermalstructure in full-thickness human skinequivalents. A, representativephotomicrographs of hematoxylinand eosin (H&E) stained Alvetexseededwith human dermal fibroblastsafter culture in Media A for 18 days.B and C, representativephotomicrographs showing H&E-stained 35-day full-thickness humanskin equivalents at 20� and 10�magnification respectively. D,representative electronmicrographsof a noncellularized Alvetex scaffold;E, 35-day full-thickness human skinequivalent; or F, normal humanskin. A–C, scale bars, 100 mm; D–F,scale bars, 75 mm; Epi, epidermis;Der, dermis.
Cutaneous Melanoma Invasion in Human 3D Skin Equivalents
www.aacrjournals.org Mol Cancer Ther; 14(11) November 2015 2669
on June 24, 2020. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst September 1, 2015; DOI: 10.1158/1535-7163.MCT-15-0394

after postincorporation with melanoma cells. Conversely, histol-ogy and immunostaining for the expression of Melan-A in themetastatic melanoma (Fig. 4D and H) revealed highly invasive
tumor cells with disrupted type IV (Fig. 4L) and VII collagens(Fig. 5D) expression. Active invasion of this advanced metastaticmelanoma, resulting in loss or disruption of type IV (Fig. 4P and
Figure 3.Expression of epidermal, dermal, or basement membranemarkers in full-thickness human skin equivalents comparedwith human skin and Mattek EpiDermFT. A, schematicillustrating dermal and epidermal protein marker expression.B–D, representative photomicrographs showing ahematoxylin and eosin (H&E) stained full-thickness humanskin equivalent (B), normal human skin (C), or a MattekEpiDermFT (D). Representative fluorescentphotomicrographs for the expression of involucrin (E–G),cytokeratin I (CK 1; H–J), cytokeratinXIV (CK 14; K–M), type IVcollagen (N–P), type VII collagen (Q–S), or type III collagen(T–V) in full-thickness human skin equivalents, normalhuman skin, or Mattek EpiDermFT. B–D and T–V, scale bars,75 mm; E–S, scale bars, 25 mm.
Hill et al.
Mol Cancer Ther; 14(11) November 2015 Molecular Cancer Therapeutics2670
on June 24, 2020. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst September 1, 2015; DOI: 10.1158/1535-7163.MCT-15-0394

T) and type VII collagens (Fig. 5H and I), similarly reflected thehistopathologic features observed in 3D skin equivalents 4 weeksafter incorporation of SK-mel-28 metastatic melanoma cells.Collectively, these data indicate that our novel 3D skin equivalentaccurately recreates the progressive histopathologic features ofmelanoma invasion in human skin and the applicability of thisnovel organotypic skin model as a valuable tool for the investi-gation of early melanoma invasion.
DiscussionThis studydemonstrates the generationof a novel full-thickness
human skin equivalent bearing morphologic and structural sim-ilarity to normal human skin within 35 days. We have optimizedand validated a protocol for the construction of an organotypicskin model from primary human fibroblasts and keratinocytesthat accurately recreates themicroenvironment of normal human
skin, as demonstrated by the production of human extracellularmatrix component type III collagen, as well as the distinct expres-sion profile of basement membrane proteins type IV and VIIcollagens, and epidermal differentiation markers cytokeratin 14,and involucrin. Incorporation of melanoma cells into their orig-inal environmental niche at the epidermal–dermal junctiondemonstrates that tumor cells retain their proliferative and inva-sive potential, forming melanoma clusters before invadingthrough the basement membrane into the dermis.
Comparative histopathologic features observed in primarymelanomas, derived from differing American Joint Committeeon Cancer (AJCC) disease stages (33), confirm that the 3D skinequivalent model is physiologically representative of clinicaldisease. Conversely, although Mattek EpidermFT expressed typeIV collagen, the lack of human type III and VII collagen expressionsuggests that the reduced longevity of this model will limit its usefor the investigation of less-invasive melanoma cells.
Figure 4.Early cutaneousmelanoma invasion infull-thickness human skin equivalentsresults in disruption of basementmembrane component type IVcollagen. Representativephotomicrographs showinghematoxylin and eosin (H&E) stainedfull-thickness melanoma skinequivalents (MSE) at 2 weeks (A) or4 weeks (B) after inoculation withmelanoma cells, highlightingclusters/nests of melanoma cells atthe dermal/epidermal junction atweek 2, which subsequently invadethrough the basement membrane atweek 4 (black arrowheads); andH&E-stained sections of a melanomain situ (C) or a primary superficialspreading malignant melanoma (D;invasive melanoma; black dotted linesillustrate the tumor boundary).Representative fluorescentphotomicrographs for the expressionof Melan-A (red; E–H) or type IVcollagen (green; I–L) in 2-week (E andI) or 4-week (F and J) MSEs,melanoma in situ (G and K), or aninvasive melanoma (H and L; redarrows illustrate intact type IVcollagen whereas white arrowsillustrate where type IV collagen islost). M–P, overlay fluorescentphotomicrographs showing relativeexpression of Melan-A and type IVcollagen in 2-week (M) and 4-week(N) MSEs, melanoma in situ (O), andan invasive melanoma (P; notemelanoma cells have invaded fromright to left) with white boxeshighlighting area magnified in Q–T(blue, DAPI). Q–T, 63� magnificationof Melan-A and type IV collagen in 2-week (Q) and 4-week (R) MSEs,melanoma in situ (S), and an invasivemelanoma (T). A–P, scale bars, 100mm; Q–T, scale bars, 25 mm.
Cutaneous Melanoma Invasion in Human 3D Skin Equivalents
www.aacrjournals.org Mol Cancer Ther; 14(11) November 2015 2671
on June 24, 2020. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst September 1, 2015; DOI: 10.1158/1535-7163.MCT-15-0394

Interestingly, our data demonstrate that, although invasionof both SK-mel-28 and WM35 melanoma cells through thebasement membrane of the 3D skin equivalent resulted in thebreakdown and disruption of type IV collagen, there appearedto be an increase in type IV collagen surrounding invadingtumor cells, consistent with previous observations showingthat increased type IV collagen expression parallels melanomaprogression (34, 35) and which is directly required for mel-anoma metastasis (36). However, increased type IV collagen inthis context is likely independent of its function as a basementmembrane component as it does not form a continuousmembrane structure. Collectively, these data support thevalidity of the 3D skin equivalent as a representative modelof early melanoma invasion in vivo. Furthermore, becausechemokines and growth factors, including IGF-1 (37), areknown to drive melanoma invasion, the present model mayalso offer a means through which to study the effect ofmodulating such factors within melanoma cells on earlytumor invasion.
In addition to confirming the presence of distinct skin layerswithin the skin equivalent, our data demonstrate the presence ofregular compacted areas within the epidermis (Fig. 2B), whichmay represent important microenvironmental niche areas of theskin where skin stem cells may reside (38–41). Importantly, thesedata confer the additional potential utility of our 3D skin equiv-alent model for the investigation of dermal stem cell and hairfollicle biology.
Furthermore, although the model presented is an allogeneicskin equivalent specifically developed for the investigation ofmelanoma invasion, itmay be readily adapted into an autologoussetting for the investigation of immunologic pathologies, oradapted through the addition of endothelial cells to the lowersurface for studies of angiogenesis within the skin or developmentof tumor neovasculature. Grafting the 3D skin equivalent ontoimmunocompromised mice, in line with studies in alternativeskin equivalent models (42), may also represent a useful means
through which to investigate tumor cell dissemination from theskin to secondary sites.
In summary, the 3D skin equivalent model presented repre-sents a robust and reproducible assay that is widely applicable todermatological research, mimicking the morphology and micro-environment of normal human skin more accurately than previ-ous assays. The demonstration of the applicability of this modelfor the investigation of the early stages of human melanomainvasion therefore renders it a valuable tool for defining andevaluating urgently required novel drug targets and personalizedtherapies.
Disclosure of Potential Conflicts of InterestStefan Przyborski is a consultant/advisory board member for and is former
Director of Reinnervate. No potential conflicts of interest were disclosed by theother authors.
Authors' ContributionsConception and design: J.L. Armstrong, S. Przyborski, P.E. LovatDevelopment of methodology: D.S. Hill, N.D.P. Robinson, E.A. O'Toole,J.L. Armstrong, S. Przyborski, P.E. LovatAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.):D.S. Hill, N.D.P. Robinson, M.P. Caley, S. Przyborski,P.E. LovatAnalysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis):D.S. Hill, N.D.P. Robinson,M.P. Caley, E.A. O'Toole,S. Przyborski, P.E. LovatWriting, review, and/or revision of the manuscript: D.S. Hill, N.D.P.Robinson, E.A. O'Toole, J.L. Armstrong, S. Przyborski, P.E. LovatAdministrative, technical, or material support (i.e., reporting or organizingdata, constructing databases): D.S. Hill, M. Chen, S. Przyborski, P.E. LovatStudy supervision: S. Przyborski, P.E. Lovat
Grant SupportThis work was predominantly supported in the United Kingdom by grants
from The JGW Patterson Foundation (D.S. Hill, J.L. Armstrong, P.E. Lovat) andTheNewcastleHealthcare Charity (P.E. Lovat) with additional support from theNational Council for Reduction, Refinement and Replacement of Animals inResearch (NC3Rs; D.S. Hill, P.E. Lovat), the Biotechnology and Biological
Figure 5.Early cutaneousmelanoma invasion infull-thickness human skin equivalentresults in disruption of basementmembrane component type VIIcollagen. A–D, representativefluorescence photomicrographs oftypeVII collagen (green) expression in2-week (A) and 4-week (B)melanomaskin equivalents (MSE), melanomain situ (C), and a primary superficialspreading malignant melanoma (D;invasive melanoma). E–H, overlayfluorescence photomicrographsshowing relative expression ofMelan-A (red) and type VII collagen in2-week (E) and 4-week (F) MSEs,melanoma in situ (G) and an invasivemelanoma (H) with white boxeshighlighting area magnified in I–L(blue, DAPI). I–L, 63�magnification ofMelan-A and type VII collagen in2-week (I) and 4-week (J) MSEs,melanoma in situ (K), and an invasivemelanoma (L).A–H, scalebars, 100mm;I–L, scale bars, 25 mm.
Mol Cancer Ther; 14(11) November 2015 Molecular Cancer Therapeutics2672
Hill et al.
on June 24, 2020. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst September 1, 2015; DOI: 10.1158/1535-7163.MCT-15-0394

Sciences Research Council (BBSRC; N.D.P. Robinson and S. Przyborski), andDEBRAUK (M.P. Caley, E.A.O'Toole).Work in theUnited States of Americawassupported by NIH (M. Chen, grants RO1 AR47981 and RO1 AR33625).
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby marked
advertisement in accordance with 18 U.S.C. Section 1734 solely to indicatethis fact.
Received May 14, 2015; revised July 29, 2015; accepted August 14, 2015;published OnlineFirst September 1, 2015.
References1. Sandru A, Voinea S, Panaitescu E, Blidaru A. Survival rates of patients with
metastatic malignant melanoma. J Med Life 2014;7:572–6.2. SimpsonCL, PatelDM,GreenKJ.Deconstructing the skin: cytoarchitectural
determinants of epidermal morphogenesis. Nat Rev Mol Cell Biol2011;12:565–80.
3. Bandarchi B, Jabbari CA, Vedadi A, Navab R. Molecular biology of normalmelanocytes and melanoma cells. J Clin Pathol 2013;66:644–8.
4. Fleischmajer R, Utani A, MacDonald ED, Perlish JS, Pan TC, Chu ML, et al.Initiation of skin basement membrane formation at the epidermo-dermalinterface involves assembly of laminins through binding to cell membranereceptors. J Cell Sci 1998;111 (Pt 14):1929–40.
5. Boyce ST. Design principles for composition and performance of culturedskin substitutes. Burns 2001;27:523–33.
6. DhomenN, Reis-Filho JS, daRochaDias S,HaywardR, Savage K,Delmas V,et al. Oncogenic Braf induces melanocyte senescence and melanoma inmice. Cancer Cell 2009;15:294–303.
7. KumasakaMY, Yajima I,HossainK, IidaM,Tsuzuki T,OhnoT, et al. Anovelmouse model for de novo Melanoma. Cancer Res 2010;70:24–9.
8. Dankort D, Curley DP, Cartlidge RA, Nelson B, Karnezis AN, DamskyWEJ,et al. Braf(V600E) cooperates with Pten loss to induce metastatic melano-ma. Nat Genet 2009;41:544–52.
9. Khavari PA. Modelling cancer in human skin tissue. Nat Rev Cancer2006;6:270–80.
10. Walker GJ, Soyer HP, Terzian T, Box NF. Modelling melanoma in mice.Pigment Cell Melanoma Res 2011;24:1158–76.
11. Li L, Fukunaga-Kalabis M, Herlyn M. The three-dimensional human skinreconstructmodel: a tool to study normal skin andmelanoma progression.J Vis Exp 2011;54:pii 2937.
12. Monteiro-Riviere NA, Inman AO, Snider TH, Blank JA, Hobson DW.Comparison of an in vitro skin model to normal human skin for derma-tological research. Microsc Res Tech 1997;37:172–9.
13. Berking C, Herlyn M. Human skin reconstruct models: a new applicationfor studies of melanocyte and melanoma biology. Histol Histopathol2001;16:669–74.
14. Meier F, Nesbit M, Hsu MY, Martin B, Van Belle P, Elder DE, et al. Humanmelanoma progression in skin reconstructs: biological significance ofbFGF. Am J Pathol 2000;156:193–200.
15. Eves P, Layton C, Hedley S, Dawson RA, Wagner M, Morandini R, et al.Characterization of an in vitromodel of humanmelanoma invasion basedon reconstructed human skin. Br J Dermatol 2000;142:210–22.
16. Bokhari M, Carnachan RJ, Cameron NR, Przyborski SA. Novel cell culturedevice enabling three-dimensional cell growth and improved cell function.Biochem Biophys Res Commun 2007;354:1095–100.
17. Peterkofsky B. The effect of ascorbic acid on collagen polypeptide synthesisand proline hydroxylation during the growth of cultured fibroblasts. ArchBiochem Biophys 1972;152:318–28.
18. Nusgens BV,Humbert P, Rougier A,ColigeAC,HaftekM, Lambert CA, et al.Topically applied vitaminC enhances themRNA level of collagens I and III,their processing enzymes and tissue inhibitor of matrix metalloproteinase1 in the human dermis. J Invest Dermatol 2001;116:853–9.
19. Boehnke K,MiranceaN, Pavesio A, FusenigNE, Boukamp P, Stark HJ. Effectsof fibroblasts and microenvironment on epidermal regeneration and tissuefunction in long-term skin equivalents. Eur J Cell Biol 2007;86:731–46.
20. ElGhalbzouri A, Commandeur S, Rietveld MH, Mulder AA, Willemze R.Replacement of animal-derived collagen matrix by human fibroblast-derived dermal matrix for human skin equivalent products. Biomaterials2009;30:71–8.
21. Vorsmann H, Groeber F, Walles H, Busch S, Beissert S, Walczak H, et al.Development of a human three-dimensional organotypic skin-melanomaspheroid model for in vitro drug testing. Cell Death Dis 2013;4:e719.
22. Leight JL, Tokuda EY, Jones CE, Lin AJ, Anseth KS. Multifunctionalbioscaffolds for 3D culture of melanoma cells reveal increased MMP
activity and migration with BRAF kinase inhibition. Proc Natl Acad SciU S A 2015;112:5366–71.
23. Sharpe GR, Gillespie JI, Greenwell JR. An increase in intracellular freecalcium is an early event during differentiation of cultured human kera-tinocytes. FEBS Lett 1989;254:25–8.
24. Vollmers A, Wallace L, Fullard N, Hoher T, Alexander MD, Reichelt J. Two-and three-dimensional culture of keratinocyte stem and precursor cellsderived from primary murine epidermal cultures. Stem Cell Rev 2012;8:402–13.
25. Stark HJ, Baur M, Breitkreutz D, Mirancea N, Fusenig NE. Organotypickeratinocyte cocultures in defined medium with regular epidermal mor-phogenesis and differentiation. J Invest Dermatol 1999;112:681–91.
26. Hiscutt EL, Hill DS, Martin S, Kerr R, Harbottle A, Birch-Machin M, et al.Targeting X-linked inhibitor of apoptosis protein to increase the efficacy ofendoplasmic reticulum stress-induced apoptosis for melanoma therapy.J Invest Dermatol 2010;130:2250–8.
27. Armstrong JL, Hill DS, McKee CS, Hernandez-Tiedra S, Lorente M, Lopez-Valero I, et al. Exploiting cannabinoid-induced cytotoxic autophagy todrive melanoma cell death. J Invest Dermatol 2015;135:1629–37.
28. Knight E, Murray B, Carnachan R, Przyborski S. Alvetex(R): polystyrenescaffold technology for routine three dimensional cell culture. MethodsMol Biol (Clifton, NJ) 2011;695:323–40.
29. Ellis RA, Horswell S, Ness T, Lumsdon J, Tooze SA, Kirkham N, et al.Prognostic impact of p62 expression in cutaneous malignant melanoma.J Invest Dermatol 2014;134:1476–8.
30. ChenM, PetersenMJ, LiHL, Cai XY,O'Toole EA,WoodleyDT.Ultraviolet Airradiation upregulates type VII collagen expression in human dermalfibroblasts. J Invest Dermatol 1997;108:125–8.
31. Betz P, Nerlich A, Wilske J, Tubel J, Wiest I, Penning R, et al. The time-dependent rearrangement of the epithelial basementmembrane in humanskin wounds–immunohistochemical localization of collagen IV and VII.Int J Legal Med 1992;105:93–7.
32. Hoath SB, Leahy DG. The organization of human epidermis: functionalepidermal units and phi proportionality. J Invest Dermatol 2003;121:1440–6.
33. Balch CM, Gershenwald JE, Soong SJ, Thompson JF, Atkins MB, Byrd DR,et al. Final version of 2009 AJCC melanoma staging and classification.J Clin Oncol 2009;27:6199–206.
34. Rotte A, Martinka M, Li G. MMP2 expression is a prognostic marker forprimary melanoma patients. Cell Oncol (Dordrecht) 2012;35:207–16.
35. Hofmann UB, Westphal JR, Waas ET, Zendman AJ, Cornelissen IM, RuiterDJ, et al. Matrix metalloproteinases in human melanoma cell lines andxenografts: increased expression of activated matrix metalloproteinase-2(MMP-2) correlates with melanoma progression. Br J Cancer 1999;81:774–82.
36. Shaverdashvili K, Wong P, Ma J, Zhang K, Osman I, Bedogni B. MT1-MMPmodulates melanoma cell dissemination and metastasis through activa-tion of MMP2 and RAC1. Pigment Cell Melanoma Res 2014;27:287–96.
37. Friedl P, Wolf K. Tumour-cell invasion andmigration: diversity and escapemechanisms. Nat Rev Cancer 2003;3:362–74.
38. Ghazizadeh S, Taichman LB. Organization of stem cells and their progenyin human epidermis. J Invest Dermatol 2005;124:367–72.
39. Wong VW, Levi B, Rajadas J, LongakerMT, Gurtner GC. Stem cell niches forskin regeneration. Int J Biomater 2012;2012:926059.
40. Bickenbach JR. Identification and behavior of label-retaining cells in oralmucosa and skin. J Dent Res 1981;60 Spec No C:1611–20.
41. Taylor G, Lehrer MS, Jensen PJ, Sun TT, Lavker RM. Involvement offollicular stem cells in forming not only the follicle but also the epidermis.Cell 2000;102:451–61.
42. Kiowski G, Biedermann T, Widmer DS, Civenni G, Burger C, Dummer R,et al. Engineering melanoma progression in a humanized environment invivo. J Invest Dermatol 2012;132:144–53.
www.aacrjournals.org Mol Cancer Ther; 14(11) November 2015 2673
Cutaneous Melanoma Invasion in Human 3D Skin Equivalents
on June 24, 2020. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst September 1, 2015; DOI: 10.1158/1535-7163.MCT-15-0394

2015;14:2665-2673. Published OnlineFirst September 1, 2015.Mol Cancer Ther David S. Hill, Neil D.P. Robinson, Matthew P. Caley, et al. Melanoma InvasionA Novel Fully Humanized 3D Skin Equivalent to Model Early
Updated version
10.1158/1535-7163.MCT-15-0394doi:
Access the most recent version of this article at:
Material
Supplementary
http://mct.aacrjournals.org/content/suppl/2015/09/01/1535-7163.MCT-15-0394.DC1
Access the most recent supplemental material at:
Cited articles
http://mct.aacrjournals.org/content/14/11/2665.full#ref-list-1
This article cites 41 articles, 5 of which you can access for free at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://mct.aacrjournals.org/content/14/11/2665To request permission to re-use all or part of this article, use this link
on June 24, 2020. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst September 1, 2015; DOI: 10.1158/1535-7163.MCT-15-0394