Embed Size (px)
Citation preview

Proc. Natd. Acad. Sci. USAVol. 91, pp. 6614-6618, July 1994Genetics
A yeast system for expression of human cystathionine f3-synthase:Structural and functional conservation of the human andyeast genes
(homcystlnuria/inherlted metabolic dlsorder/Sacchromyces cerevisiae/CYS4)
WARREN D. KRUGER* AND DAVID R. Cox*Department of Psychiatry, University of California at San Francisco, San Francisco, CA 94143
Communicated by Ira Herskowitz, March 14, 1994 (received for review May 17, 1993)
ABSTRACT Human cystathionine P-synthase (CBS; EC4.2.1.22) deficiency results In a recessive genetic disorderwhose clinical and biochemical manifestations vary greatlyamong affected individuals. In an effort to identify and analyzemutations in the human CBS gene, we have developed a yeastexpression system for human CBS. We have cloned andsequenced a human cDNA that codes for CBS and haveexpressed the human CBS protein in yeast cells lacking endog-enous CBS. The human enzyme produced in yeast is functionalboth in Wiro and in vivo. We have also cloned and sequenced theyeast gene, CYS4, that codes for CBS. The predicted humanand yeast CBS proteins are 38% identical and 72% similar toeach other, as well as sharin sgnificant similarity withbacterial cysteine synthase. These results demonstrate theevolutionary conservation of CBS and establish the utility of ayeast expression system for studying human CBS.
Cystathionine P-synthase (CBS) deficiency in humans is arare autosomal recessive disease, which is the most commoncause of homocystinuria. Individuals with this disorder havea variety of clinical phenotypes including ocular, skeletal,neurological, and cardiovascular defects, which are believedto result primarily from elevated plasma homocysteine lev-els. Age of onset, symptoms, and severity of the disease canvary greatly among patients (1). Some, but not all, individualswith homocystinuria respond to increased dietary levels ofvitamin B6 with an increase in residual enzyme activityassociated with a lowering of plasma homocysteine levels.Within sibships, almost complete concordance is observedfor responsiveness and nonresponsiveness to vitamin B6 (2).Concordance within sibships is also observed in the residualCBS activity found in cultured fibroblast cells derived fromaffected individuals (3). Residual enzyme activity in fibro-blast extracts ranges from undetectable to 10%6 of normalcontrols. Generally, vitamin B6 responders have higher levelsof residual CBS activity than nonresponders (3-6). Theseobservations suggest that different mutations within the CBSgene alter the enzyme activity in discrete ways; thus, differ-ent CBS mutations may be related to the clinically observedheterogeneity found in patients with CBS deficiency.A life-threatening complication of CBS deficiency is
thromboembolism (1). Untreated vitamin B6 responsive andnonresponsive individuals have a 4% risk per year of athrombolytic event (2). Thrombosis can occur at any age andcan involve both large and small arteries. Studies also suggestthat individuals heterozygous for mutations in CBS are atincreased risk for premature peripheral and cerebral arterialdisease (7-10). These studies show a correlation betweenreduced levels of CBS activity and premature vascular dis-ease. Since ='1 in 200 people in the general population is a
heterozygote (1), it has been suggested that these individualsmay account for a large fraction of those at risk for stroke.However, testing this hypothesis is difficult due to the lack ofan accurate assay that identifies heterozygotes (11).To identify and study mutations that cause homocysti-
nuria, an effective assay for the human CBS gene product isrequired. Such an assay system must be able to distinguishDNA polymorphisms with no phenotypic effect from muta-tions that affect enzymatic function. In addition, the assaymust be able to isolate individual alleles for study.We report here the development of such an experimental
system for the study of human CBS using Saccharomycescerevisiae. We describe the cloning and sequencing of ahuman CBS-encoding cDNAt and show that expression ofthis cDNA is able to functionally complement a yeast strainthat lacks endogenous CBS activity. Additionally, we havecloned and sequenced the yeast gene encoding CBS (CYS4).The predicted yeast and human CBS proteins are 72% similarto each other and have significant similarity to the predictedrat CBS protein and bacterial cysteine synthase. Theseresults show the strong evolutionary conservation of CBSand suggest the utility of this yeast system for detection andanalysis of mutations in the human CBS coding region.
MATERIALS AND METHODSYeast Strains and Genetic Methxds. Yeast strains WC6-6a
(MATa ura3-52 cys4-1 cys2-1) and WC44a (MATa ura3-52)were constructed by crossing JW1-2c (MATa cys2-1 cys4-1)(12) and CY70 (MATa trpl ura3-S2 his3 leu2) (13). Standardyeast genetic methods were performed essentially as de-scribed (14).
Cloning of Human CBS cDNA. A human cDNA libraryderived from HepG2 cells constructed in vectorpAB23-BXN(15) was used as a template for PCR amplification using theCBS-specific primers 5'-TCCATTTATATGAAATGTCC(forward) and 5'-TGGGTGGCTCAGCATCCTCC (reverse)(16). PCR was carried out in 67 mM Tris (pH 8.8), 6.7 mMMgCl2, 16.6mM NH4SO4, 10mM 2-mercaptoethanol, and 6.7,uM EDTA. The cycling parameters were 30 sec at 94CC, 30sec at 500C, and 30 sec at 720C for 40 cycles. Bacterial cellscontaining the human cDNA inserts were diluted in liquidmedium and used to inoculate cultures at -50,000 cells pertube. Miniprep DNA was prepared and analyzed for thepresence of a CBS-encoding cDNA by PCR, and the productwas visualized by agarose gel electrophoresis. One of thepositive pools was diluted and used to inoculate 10 new poolsat 8000 cells each. One ofthese pools tested positive by PCR,
Abbreviations: CBS, cystathionine (-synthase; ORF, open readingframe.*Present address: Department of Genetics, School of Medicine,Stanford University, Stanford, CA 94305.tThe sequences reported in this paper have been deposited in theGenBank data base (accession nos. L14577 and L14578).
6614
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.

Proc. Natl. Acad. Sci. USA 91 (1994) 6615
and this pool was plated out for individual colonies, whichwere subsequently screened by hybridization with the PCR-generated human CBS fragment as probe. Probe was pre-pared by random priming (17), and hybridizations werecarried out using Biotrans membranes (ICN) under condi-tions recommended by the manufacturer. The final clone wasdesignated pHCBSi.
Cloning of CYS4 and Construction of a CYS4 Nuil Allele.Previous studies have shown that yeast containing cys4-1 donot exhibit CBS activity (12). Therefore, strain WC6-6a wastransformed with a yeast genomic library constructed inYCp5O (18) and plated out on SC-URA (14) plates supple-mented with 20 A. of glutathione (30 mg/ml) as a source ofcysteine. Approximately 2000 transformants were then rep-lica plated to unsupplemented SC-URA plates. Positive col-onies were retested and cured of the plasmid by passage on5-fluoroorotic acid plates (19). DNA was prepared from theyeast and used to transform Escherichia coli. The recoveredplasmid was named pYCBS. Restriction fragments weresubcloned into YCp5O as indicated in Fig. 1.A null allele of CYS4 was constructed as follows: Initially,
the 3.2-kb BamHI-Sph I fragment containing CYS4 wassubcloned into pUC18. The resulting construct was then cutwith Bgl II, and a 2.2-kb LEU2 fragment with BamHI endswas inserted. The LEU2 fiagment was derived from pCD7-B,a derivative of pRB758. The resulting clone, pUC:YCBS::LEU2 (Fig. 1C), was digested with Sph I and used totransform CY70. Nineteen out of 20 Leu+ colonies examinedhad a Cys- phenotype. Proper integration was confirmed byhybridization.
Sequencing. A 2-kb EcoRI fragment, a 1.2-kb EcoRI-SphI fragment, and a 3.2-kb BamHI-Sph I fragment from theyeast clone pYCBS (containing the CYS4 gene) were sub-cloned into pVZ1+ for sequencing. The entire EcoRI frag-ment containing the human CBS cDNA from clone pHCBSiwas also cloned into pVZ1+. pVZ1+ is pBS+ with anexpanded polylinker (20). All sequencing was done usingCsCl-purified double-stranded DNA as template and Seque-nase. The following primers, in addition to the vector T7 andT3 primers, were used in the CYS4 sequencing: 5'-GACCAATATAACAATATGAT, 5'-CTGGCATTGCGG-TACTGGTCCAG, 5'-TTCTAACGAAGGTGTCTTTG, 5'-AATGGCTAAATCTGAGCAGC, and 5'-GTTGGGCTA-AAATTGAACCG. The following primers were used in thehuman CBS sequencing: 5'-CTGGCATTGCGGTACTG-GTCCAG, 5'-CGCCCACCAATGCCAGGTTC, 5'-GCTC-CGACTTCACTTCTGG, 5'-CTTCCTTATCCTCTGG-GAAC, 5'-CGTTGCTCTTGAACCACTTG, 5'-GTGGAG-GATGCTGAGCCA,5'-AGTGGGCACGGGCGGCACCA,5'-CACCACGGGCGCCTGGTCGA, 5'-TGGTGGTGCAC-GAGCAGATC, 5'-AGGAGGGCCAGCGCTGGCTG, and5'-TGCGGTCCTTCACGCTCCCG.
Construction of pHCBS and pAHCBS. The plasmidpAB23BXBN is a 2-,um shuttle vector containing a multiplecloning site placed in between glyceraldahyde-3-phosphatedehydrogenase promoter and terminator sequences (15). Theoriginal cDNA clone, pHCBSi, had the human CBS cDNAinsert in the antisense orientation relative to the promoter inthe vector pAB23BXN. Therefore, pHCBSi DNA was di-gested with Bgl II, which released the insert, and subse-quently treated with T4 ligase. One of the resulting clonescontained the insert in the correct orientation and was calledpHCBS. To delete the 5' untranslated region of the CBScDNA, we made use of an Sph I site containing the predictedstart ATG for the human CBS gene. The plasmidpVZ1+:HCBS was partially digested with Sph I, and aresulting 2.2-kb fragment containing the human CBS cDNAand 3' polylinker sequences (containing a Sal I site) wasisolated and inserted into pUC18. The insert was removedfrom this plasmid by digestion with Sal I and was subse-
quently inserted into pBX-X at theXho I site. Plasmid pBX-Xis identical to pAB23BXN except for replacement of themultiple cloning site with a single Xho I site; it was createdby digestion of pHCBSi with Bgl II, isolation of the vectorbackbone, and ligation in the presence ofXho I linker DNA.CBS Enzyme Assays. Yeast extracts were prepared from
100-ml cultures grown to midlogarithmic phase in SC-URAmedium. Cells were harvested at 40C, washed once withice-cold lysis buffer (20 mM Na2PO4, pH 7.5), suspended in500 jul of lysis buffer, and transferred to tubes containing 500y4 of acid-washed glass beads (0.2 Am). Cells were lysedusing a Biospec Products (Bartlesville, OK) minibeadbeaterfor 2 min (4 30-sec pulses). Each lysed cell suspension wasthen centrifuged in a Fisher microcentrifuge for 10 min toremove insoluble material. Protein concentrations were de-termined with the Bio-Rad protein assay reagent, usingbovine serum albumin as an internal standard. CBS enzymeassays were conducted as described (21), except assay vol-umes were 200 A., and the reaction time was 4 hr. Cystathio-nine was measured as described (21) by mixing 70 ,ul of theassay reaction with 1 ml of Ninhydrin reagent and boiling for5 min. OD45o was measured using a Beckman model DU-50spectrophotometer. Levels of cystathionine produced werequantitatively determined by comparing results against astandard of known cystathionine concentration. The assaywas linear over the time tested.
RESULTSCloning of a Human CBS cDNA. A human CBS cDNA
clone was isolated using two primers derived from the 3'untranslated region of the human CBS gene and used to mapthe gene on chromosome 21 (16). These primers were used toscreen by PCR a human HepG2 cDNA library (15) for thepresence of a 323-bp product. The library was subsequentlydivided into subpools and screened again. One subpool wasidentified that contained the CBS cDNA. This pool wasplated out, and the colonies were screened by hybridizationusing the radiolabeled 323-bp CBS PCR product as a probe.Three positive clones were obtained, all of which had anidentical 2.5-kb cDNA insert. A map of this clone and thesequencing strategy are shown in Fig. 1A. The nucleotidesequence has been deposited in GenBank (accession no.L14577). The cDNA contains one very long open readingframe (ORF) of 1656 bp and two short ORFs upstream. TheDNA sequence of the long open reading frame has 80%oidentity with the rat CBS cDNA type I, which is one of thefour splice variants of the rat CBS cDNA previously de-scribed (22). At the amino acid level, the similarity is evenmore striking. Greater than 90%o of the amino acids are similarbetween the predicted rat and human proteins (see Fig. 2).
Cloning of the Yeast Gene Ending CBS. An S. cerevisiaegenomic clone encoding CBS was isolated by complementa-tion of a yeast strain containing the cys4-1 mutation (WC6-6a). Previous work had shown that yeast containing thismutation are unable to form colonies in the absence ofexogenous cysteine and that extracts from this strain lackdetectable CBS activity in vitro (ref. 12; Table 1). This yeaststrain was transformed with a yeast genomic library, andseven colonies were identified that grew in the absence ofexogenous cysteine (see Materials and Methods). This phe-notype was found to be plasmid dependent. Additionally,extracts made from cells carrying the plasmid had CBSenzyme activity in vitro (Table 1). The plasmids were recov-ered, and restriction analysis indicated that they all containedidentical inserts. Subcloning of the insert followed by trans-formation into WC6-6a revealed that an Sph I-BamHI sub-fiagment was sufficient for complementation and that anEcoRI site must lie in or very near the gene (see Fig. 1B).Sequencing in both directions from the EcoRI site revealed
Genetics: Kruger and Cox

Proc. Natl. Acad. Sci. USA 91 (1994)
A - 500 bpR S R
pHCBS I-,--- 1-
Cl N
R SpAHCBS I
B
pYCBS
complementation
+
-~1 kb
R S RH XR SI I~I II
I
HX B
pLR R R
R RpSR l _
S R BpSB I I I
C
pUC:YCBS::LEU2 LEU2
S 7 Bl ..I
+
+
C , N
FIG. 1. Structure and sequencing strategy of the human CBS geneand yeast CYS4 gene. (A) The structure and sequencing strategy ofthe human cDNA is shown. R, EcoRI; S, Sph I; H, HindHI; X, XbaI; B, BamHI. The column on the right ofeach clone indicates whetherthat piece could complement yeast strain WC6-6a (Cys-) for growth.The open reading frame and its directionality are indicated by theopen rectangle. (B) The structure of the original yeast genomic insertcontaining the yeast CBS gene (CYS4) is shown. Also shown arethree subfiagments derived from the insert. The sequencing strategyof the open reading frame that encodes yeast CBS is shown. (C)Structure of the disruption allele for CYS4.
that the site was contained within a large ORF encoding aprotein of 506 amino acids in length (GenBank accession no.L14578). The DNA sequence was then used to scan theGenBank data base for sequence homology. The DNA se-quence shares 50%6 identity with a rat cDNA for CBS. Thisstrong similarity was also seen at the protein level (Fig. 2).This similarity, along with the ability of the plasmid topromote CBS enzyme activity in yeast extracts, indicatesthat we have cloned the structural gene of the yeast CBSenzyme.
We constructed a null allele of CYS4 by insertion of theLEU2 gene near the beginning of the coding region (Fig. 1C).This insertion disrupts the gene at amino acid position 72.Congenic yeast carrying this mutation exhibit a Cys- phe-notype and are indistinguishable from cells carrying thecys4-1 allele (data not shown).
Expression of Human CBS in Yeast Lacking EndogenousCBS. The human CBS cDNA was tested for its ability topromote growth of cys4 yeast in the absence of exogenouscysteine. Initially, the entire human CBS cDNA was inserteddownstream from the glyceraldehyde-3-phosphate dehydrog-enase promoter present in pAB23BXN (15). The resultingplasmid, pHCBS, was used to transform cys4-1 strain WC6-6a. No growth was detected in the absence of exogenous
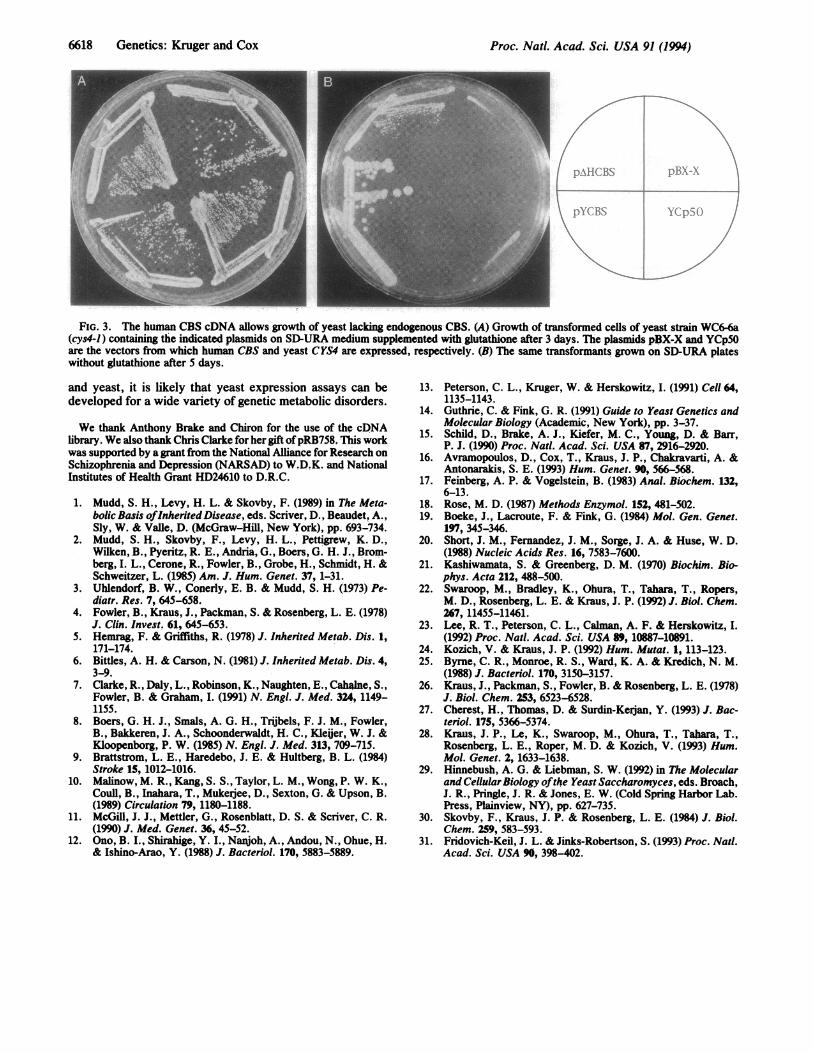
cysteine (data not shown). However, when the construct wasreengineered such that the 5' untranslated region of thehuman CBS cDNA was deleted (see Materials andMethods),growth was observed. As shown in Fig. 3, the plasmidexpressing the truncated human CBS cDNA, pAHCBS,allows strain WC6-6a to form colonies in the absence ofexogenous cysteine, while the same strain containing theexpression vector alone, pBX-X, does not. The figure alsoshows that cells containing pAHCBS do not grow as rapidlyas cells containing aplasmid expressing CYS4, pYCBS, in theabsence of exogenous cysteine. Identical results were ob-served using a yeast strain carrying a null allele ofCYS4 (Fig.1C; data not shown).CBS activity was also assayed from whole-cell extracts
made from these cells in vitro. CBS activity is detected invitro in extracts made from yeast expressing the human CBScDNA, but not from yeast with the expression vector alone(Table 1). This activity, however, is only about 20%o of theactivity seen in wild-type yeast extracts or in extracts madefrom the mutant strain carrying the yeast CBS gene on aplasmid. Thus, the reduced levels of activity observed in vitrocorrelate with the slow growth of the strain in vivo.
DISCUSSIONThis report describes the cloning and sequencing of a humancDNA clone and a yeast genomic clone encoding CBS. Wedemonstrate that the human gene is able to substitute func-tionally for the yeast gene in both in vivo and in vitro assays.CBS catalyzes the formation of cystathionine from homo-
cysteine and serine. This reaction involves the creation of asulfide linkage between the homocysteine and serine and theproduction of water as a leaving group. Both the human andyeast CBS proteins are able to carry out this reaction (12, 24).It is therefore not surprising that these two enzymes share alarge degree of sequence similarity. The yeast protein isslightly shorter than the human protein (506 vs 552 aminoacids). The alignment (Fig. 2) indicates that most of thisdifference occurs in the N terminus. No splice acceptorsequences were detected upstream of the predicted yeaststart codon, suggesting that this difference is genuine. Thefunction ofthe N terminus of the mammalian CBS protein hasnot been determined.The predicted yeast protein shares 38% identity and 72%
similarity over its entire length with the human protein.Interestingly, the human and yeast proteins also have signif-icant similarity with bacterial cysteine synthase A, which isencoded by the cysK gene (25). The human and yeast CBSproteins are, respectively, 37% and 34% identical to the cysKgene product over its entire length. Cysteine synthase cata-lyzes the addition of inorganic sulfide to O-acetylserine toform cysteine with acetic acid as a leaving group. Thus bothCBS and cysteine synthase condense serine with sulfur-containing compounds to form cysteine. Additionally, bothmammalian CBS and bacterial cysteine synthase use pyri-doxal phosphate as a cofactor (21, 25). There is one excep-tionally large block of identity between amino acids 144 and155 of human CBS in which 10 of 11 amino acids are identicalin all four CBS-related proteins. We speculate that this regioncould be involved in the binding of the common elements:serine, pyridoxal phosphate, and sulfur. Interestingly, mam-malian CBS can perform the bacterial reaction in vitro (26).The similarity of these two enzymatic reactions suggests thatthe condensation of serine with sulfur compounds to producecysteine must have occurred early in evolution.
Recently, after this work had been submitted, both thehuman and yeast CBS DNA sequences were published. Theyeast CBS DNA sequence published by Cherest et al. (27)and the one published here differ at three positions: 205,408,and 1239. The coding region for the human CBS cDNA
6616 Genetics: Kruger and Cox

Proc. Natl. Acad. Sci. USA 91(1994) 6617
61 13X -T p ED 1y1aAnfP1TI4 I
CfP1II R S G7P 11 S APMS[ T 1KS PT [T5K F A4MWPfF W lRT3A[P5 1R C 17WA[QReSFIsAIITA[A[r('l42;1I616_h )l-A'[JU J1Q)1) 11 V Q 1. K S ( K-. P R V WIGS1C A 6 '. P(IT YVPl1KW'C IBS j K 1
h(CB 7 -14r(BS 7(x140V(CBS I TcvcK 1-16
hCBlS 144-215rCB S 141-212vCBS 7X1-149cvsK 6 7 1I5
hC 9 21 6-2117rUS 2113 2I77vWBS 1511-219
ysK 156 202
hCBS 211 .5lrI S 2711 348vCIS 221- 290cyK 2(11-274
hCli S 52-40(rCBS 349-404v 'lI2H 1 1362cvsK 279-:118
hClBS 411 4111r IB'cS 41')5 476CiBS .10,4133
h(H15 4 12 539rUHBS 477-541vCBS 434-501
S II, P1DI! K i MVKI 'NK I ,K6Klp(1 1 11AKC K- T.NAiiBVRIJKi 1 I1RmT KKD&R UD I 1. Krt DU I1. P 1) I R K I G N TP MV R I N R I KNAGIKCELILAKC FFNAGS K :. S I R MI JRAG KpG- TI I
Q A S IrI I V NTP I 1 ALqK 1. P A 1. 71 K [A ELYNPGGS K D IAI S MW- A S RII BP 1S11MSE3~JI1> I_ I I
NT.PLVW"1RI-I
-NTVGATAGI-hA. AAV--V RCI1Cl I VMFP .K S KKVDrL:RALGAT IYR 7TTNARFDS PF S IT V GVAVWRI K N EI P£PTS GNT.[;GLA1AAAV K-G.R C II V M EKMS. M K V DVL R.ALAE I vRTPTPTN.,RRFDS P FS IIV GiVAWR 1. K N E. I PsrT5 iNTst1LAl mGA! KGYR II MP£.KM N KK. VL.KALGA.E I RTPTTfA AWDS P E SII I GVAMK I., . I PF T!.LGNTOIALAV V MS I R KLKALG SLV.RTA-
IN SII L1D YRt'AS NPI.ARYDT ADEFI. QQCI)- ---CiKVDMIVASAV2TGGTITGI ARKI[AEKcpcGCRI I r]Ns-SI LI)°C Y R S A S NIP 1. Alff Y D1 A EN A 1. QQ CM 1) C;-- K V 1) M 1, Y A S MA .T G Y.X ;(lI I(GAVILA)QYN IN FRF GREIQR 1. N 1, FI. NLAVVArAGTG1TI S KYI1. KLQN KI Q I V GEK YLLOsAPANE GKDTUPE-- LT -.[QnVVTIJAG TGiO ? T I T KCI K 'T ) L.JT-9
DPECGS- I Tr. A P F-QNTQTVQT1E VT -G Y D F IFPT V L.."RTVV1DWFKS NDF F. A F T F A I MLFAI LC G G S A)PI(;s- II-IIAP I1N UTIAYIVIGI Y L:FIF YLDRAVVDRWFKS N DD D F FARMIA ILCGGSSSU}P>S- I AQF I N II) TISY VEGSYDPVPt.LARK 1, I DMWYKT DD F K YAR I S G GSS G
[E_jjBTUP I A p I P CAXPF I F.A 1;!D K 1.V D K1V I G I I T RT]R E ALEG 1TIAS[Wi 1 jVWVTA V-;IA G- Q FI. Fl C Q R N- Y V I1T-F D -V R N Y%MT K F ETS D R WME Q K G F E K EFI I E K K P WIA MA V A V KA A Q IlK 6 V R V V I I P US V R YN MS K F L S D K WML QK 6 MK E- E 1I]V K R PAT A Y VV 111D 11PFV|lATD K NINPDI RY 1. TKVR F S S KK1.LT S T T K Y n*[ AAA 1-X1.-1)13 R 5TVKNIN 1R ;F TY 1. Q Q
[WWI 1. R Y Q a: L. 9 1. S A P T. T V 1, P T T T fGll T ETI T- R-'KCGF DiOXAP V V D EAp VI ,G MVTI1. 0 N \4 , S S LIAC.,\G K VQ-- n GWWILR V E1.S S A P 1 T V I. P T V T ICE T I AI I. R E K IF 1) Q A P V V N F SHA I L 6 MNiV 1. G N M1.. S S I A (iK Y|R|P511~~~~~~~~~~~~~~~~~ IYV K ,KDM
I I K Q FlR S ItII.F"ISIT! IT M rA YVI II Q I Q -- Y II S T[K Q R QJM[F G V V T A I nIK V YKQ-KIPI1I11 11HL I SHI 1 1 MDHI Al 'I VII1 Q I QjSR7QAWSI '.616Pl DR N1 NIIVISIK Q JMVF GC; V\ T A D)
JIWIKF1~V1~]UNN N VLS YFN S( K K K K XFT N 5I1. SR iFS K S N;TRI-MI.
hWBM 54111.2 F'iQ<.A RDQ[1) WQrU4BS9-561 1.1: V IARLUQIRJvC(BS SO 50 ! I 3
FIG. 2. Sequence alignment of CBS from human (hCBS), rat (rCBS) (22), S. cerevisiae (yCBS), and cysteine synthase A (cycK) from E.coli (25). Boxed residues show similarity using the human protein as the standard. Conservative groupings are (L, V, I, M), (F, W, Y), (A, G),(S, T), (R, K), (D, E), (D, N), and (E, Q). Boldfaced and shaded residues show identities found in all four proteins.
described by Kraus et al. (28) and the one published herediffer at four positions: 341, 884, 1082, and 1132.
Despite a billion years of evolutionary divergence betweenhuman and yeast, it has proved possible to functionallycomplement yeast lacking endogenous CBS by expression ofthe human CBS protein. The human CBS protein expressedin yeast functions well enough to allow growth of yeast cellsthat lack endogenous CBS activity. We were able to achievecomplementation only after deletion of the 5' leader sequencefrom the human CBS cDNA. The failure of the entire cDNAto complement growth is probably due to the presence oftwosmall ORFs in the 5' leader sequence. It has been shown innumerous instances that the presence of an ORF in the 5'untranslated region of yeast genes can have a dramatic effecton translation (29). Even with the 5' leader sequences re-moved, the growth rate of the cells expressing the humanCBS cDNA was significantly slower than the same cellscontaining the native yeast gene. This difference correlateswith the measurement of CBS enzyme activity in vitro. Wefound that CBS enzyme activity in extracts made from cellsexpressing the human cDNA is about 20% the levels of theextracts made from cells containing the yeast gene. Becausethe yeast CYS4 gene and the human CBS gene are expressedfrom different promoters, we can draw no conclusions as towhether the lower enzyme activity is due to reduction ofexpression or enzyme activity.
Table 1. CBS activity in whole-cell extracts prepared from yeastGenotype Strain Insert CBS activityCYS4 WC4-4a None 29.8 ± 1.4cys4 WC6-6a yCBS 27.7 ± 7.5cys4 WC6-6a hCBS 6.2 ± 3.9cys4 WC6-6a None ND
CBS activity was measured as described in Materials and Meth-ods. The enzyme activity is expressed in nmol of cystathionine permin per mg ofprotein. Each value represents the mean ± SD of threeindependent assays. ND, not detected (<1.0) and was the back-ground level for the bovine serum albumin control. yCBS, yeastCBS; hCBS, human CBS.
The ability to complement yeast cells lacking yeast CBSactivity with the human cDNA provides an assay system toidentify and characterize the effects of various mutations inhuman CBS. Such an assay system is significant because itallows analysis of mutations at a functional level.Using the yeast assay for human CBS in combination with
reverse transcription-PCR, it should be possible to detectheterozygotes in the general population. These studies couldfirmly establish whether heterozygotes are actually at in-creased risk of stroke. Some potential limitations of thisapproach include misdiagnosis due to mutations introducedby PCR and inability to detect mutations that lie outside thecoding region. Additionally, it may be that certain mutationsharmful in humans do not give phenotypes in yeast.Kozich and Kraus (24) have successfully used a bacterial
expression system for the human CBS cDNA to identify andanalyze two mutations in the CBS gene. This system essen-tially uses the bacteria as a factory to produce the enzyme andthen analyzes the product in vitro. In contrast, the yeastsystem described here can monitor activity both in vitro aridin vivo. Thus the yeast system can examine human CBSfunction in the context of the entire cysteine metabolicpathway.The yeast expression system should be useful in analysis of
allelic interactions between different human CBS mutations.Because CBS is normally present as a homotetramer (30),certain heterotetramers may have altered function. By al-lowing analysis ofalleles singly and in combination with otheralleles, the yeast system should be particularly well suited forsuch subunit interaction studies.
Similar yeast systems have recently been developed tostudy other human diseases. A yeast expression system forthe study of human galactose-1-phosphate uridylyltrans-ferase has been successfully used to identify and characterizea mutation in a patient with galactosemia (31). Anotheryeast-based expression system has been described for galac-tokinase, which may be useful in analyzing mutations thatcause cataracts in newborns (23). Given the high degree ofconservation in basic metabolic pathways between human
Genetics: Kruger and Cox
Proc. Natl. Acad. Sci. USA 91 (1994)
/
/
l~ K
FiG. 3. The human CBS cDNA allows growth of yeast lacking endogenous CBS. (A) Growth of transformed cells of yeast strain WC6-6a(cys4-1) containing the indicated plasmids on SD-URA medium supplemented with glutathione after 3 days. The plasmids pBX-X and YCp5Oare the vectors from which human CBS and yeast CYS4 are expressed, respectively. (B) The same transformants grown on SD-URA plateswithout glutathione after 5 days.
and yeast, it is likely that yeast expression assays can bedeveloped for a wide variety of genetic metabolic disorders.
We thank Anthony Brake and Chiron for the use of the cDNAlibrary. We also thank Chris Clarke for her gift ofpRB758. This workwas supported by a grant from the National Alliance for Research onSchizophrenia and Depression (NARSAD) to W.D.K. and NationalInstitutes of Health Grant HD24610 to D.R.C.
1. Mudd, S. H., Levy, H. L. & Skovby, F. (1989) in The Meta-bolic Basis ofInherited Disease, eds. Scriver, D., Beaudet, A.,Sly, W. & Valle, D. (McGraw-Hill, New York), pp. 693-734.
2. Mudd, S. H., Skovby, F., Levy, H. L., Pettigrew, K. D.,Wilken, B., Pyeritz, R. E., Andria, G., Boers, G. H. J., Brom-berg, I. L., Cerone, R., Fowler, B., Grobe, H., Schmidt, H. &Schweitzer, L. (1985) Am. J. Hum. Genet. 37, 1-31.
3. Uhlendorf, B. W., Conerly, E. B. & Mudd, S. H. (1973) Pe-diatr. Res. 7, 645-658.
4. Fowler, B., Kraus, J., Packman, S. & Rosenberg, L. E. (1978)J. Clin. Invest. 61, 645-653.
5. Hemrag, F. & Griffiths, R. (1978) J. Inherited Metab. Dis. 1,171-174.
6. Bittles, A. H. & Carson, N. (1981) J. Inherited Metab. Dis. 4,3-9.
7. Clarke, R., Daly, L., Robinson, K., Naughten, E., Cahalne, S.,Fowler, B. & Graham, I. (1991) N. Engl. J. Med. 324, 1149-1155.
8. Boers, G. H. J., Smals, A. G. H., Tribels, F. J. M., Fowler,B., Bakkeren, J. A., Schoonderwaldt, H. C., Kleier, W. J. &Kloopenborg, P. W. (1985) N. Engl. J. Med. 313, 709-715.
9. Brattstrom, L. E., Haredebo, J. E. & Hultberg, B. L. (1984)Stroke 15, 1012-1016.
10. Malinow, M. R., Kang, S. S., Taylor, L. M., Wong, P. W. K.,Coull, B., Inahara, T., Mukeijee, D., Sexton, G. & Upson, B.(1989) Circulation 79, 1180-1188.
11. McGill, J. J., Mettler, G., Rosenblatt, D. S. & Scriver, C. R.(1990) J. Med. Genet. 36, 45-52.
12. Ono, B. I., Shirahige, Y. I., Nanjoh, A., Andou, N., Ohue, H.& Ishino-Arao, Y. (1988) J. Bacteriol. 170, 5883-5889.
13. Peterson, C. L., Kruger, W. & Herskowitz, I. (1991) Cell 64,1135-1143.
14. Guthrie, C. & Fink, G. R. (1991) Guide to Yeast Genetics andMokcular Biology (Academic, New York), pp. 3-37.
15. Schild, D., Brake, A. J., Kiefer, M. C., Young, D. & Barr,P. J. (1990) Proc. Natl. Acad. Sci. USA 87, 2916-2920.
16. Avramopoulos, D., Cox, T., Kraus, J. P., Chakravarti, A. &Antonarakis, S. E. (1993) Hum. Genet. 90, 566-568.
17. Feinberg, A. P. & Vogelstein, B. (1983) Anal. Biochem. 132,6-13.
18. Rose, M. D. (1987) Methods En ymol. 152, 481-502.19. Boeke, J., Lacroute, F. & Fink, G. (1984) Mol. Gen. Genet.
197, 345-346.20. Short, J. M., Fernandez, J. M., Sorge, J. A. & Huse, W. D.
(1988) Nucleic Acids Res. 16, 7583-7600.21. Kashiwamata, S. & Greenberg, D. M. (1970) Biochim. Bio-
phys. Acta 212, 488-500.22. Swaroop, M., Bradley, K., Ohura, T., Tahara, T., Ropers,
M. D., Rosenberg, L. E. & Kraus, J. P. (1992) J. Biol. Chem.267, 11455-11461.
23. Lee, R. T., Peterson, C. L., Calman, A. F. & Herskowitz, I.(1992) Proc. Natl. Acad. Sci. USA 89, 10887-10891.
24. Kozich, V. & Kraus, J. P. (1992) Hum. Mutat. 1, 113-123.25. Byrne, C. R., Monroe, R. S., Ward, K. A. & Kredich, N. M.
(1988) J. Bacteriol. 170, 3150-3157.'26. Kraus, J., Packman, S., Fowler, B. & Rosenberg, L. E. (1978)
J. Biol. Chem. 253, 6523-6528.27. Cherest, H., Thomas, D. & Surdin-Kerjan, Y. (1993) J. Bac-
teriol. 175, 5366-5374.28. Kraus, J. P., Le, K., Swaroop, M., Ohura, T., Tahara, T.,
Rosenberg, L. E., Roper,'M. D. & Kozich, V. (1993) Hum.Mol. Genet. 2, 1633-1638.
29. Hinnebush, A. G. & Liebman, S. W. (1992) in The Molecularand CellularBiology ofthe Yeast Saccharomyces, eds. Broach,J. R., Pringle, J. R. & Jones, E. W. (Cold Spring Harbor Lab.Press, Plainview, NY), pp. 627-735.
30. Skovby, F., Kraus, J. P. & Rosenberg, L. E. (1984) J. Biol.Chem. 259, 583-593.
31. Fridovich-Keil, J. L. & Jinks-Robertson, S. (1993) Proc. Nati.Acad. Sci. USA 90, 398-402.
6618 Genetics: Kruger and Cox

![Three Main Causes of Homocystinuria: of Metabolism ... · most frequent causes are classical homocystinuria [deficiency of cystathionine beta-synthase (CBS)], methylmalonic aciduria](https://img.pdfslide.net/doc/110x75/5e951dcb19bd325819567b57/three-main-causes-of-homocystinuria-of-metabolism-most-frequent-causes-are.jpg)




![$YGD GH &iGL] - movilidadgranada.com · f3 f3 f3 f3 f3 f3 f3 f3 f3 f3 f3 f3f3 f3 f3 f3 f3 f3 f3 f3 f3 f3 f3 f3 f3 f3 f3 f3 f3f3 f3 f3 f3 f3 f3 f3 f3 f2 f2 f2 f2 f2 f2 f2 f2 f2 f2](https://img.pdfslide.net/doc/110x75/5bb5162f09d3f2b63a8c0773/ygd-gh-igl-f3-f3-f3-f3-f3-f3-f3-f3-f3-f3-f3-f3f3-f3-f3-f3-f3-f3-f3-f3-f3.jpg)





















