Embed Size (px)
Citation preview

R E S E A R C H A R T I C L E
ADH1expression inverselycorrelateswithCDR1andCDR2 inCandidaalbicans fromchronicoral candidosis inAPECED (APS-I) patientsEmilia Siikala1,2,3, Paul Bowyer4, Malcolm Richardson5, Harri Saxen6, Dominique Sanglard2 & RiinaRautemaa1,3,4
1Department of Bacteriology and Immunology, Haartman Institute, University of Helsinki, Helsinki, Finland; 2Institute of Microbiology, University of
Lausanne and University Hospital Center, Lausanne, Switzerland; 3Department of Oral and Maxillofacial Diseases, Helsinki University Central Hospital,
Helsinki, Finland; 4The University of Manchester, Manchester Academic Health Science Centre, Translational Research Facility, School of Translational
Medicine, and University Hospital of South Manchester, Wythenshawe Hospital, Manchester, UK; 5Mycology Reference Centre, University Hospital of
South Manchester, Wythenshawe Hospital and The University of Manchester, Manchester Academic Health Science Centre, Translational Research
Facility, School of Translational Medicine, Manchester, UK; and 6Hospital for Children and Adolescents, Helsinki University Central Hospital, University of
Helsinki, Helsinki, Finland
Correspondence: Emilia Siikala, Department
of Bacteriology and Immunology, Haartman
Institute, University of Helsinki,
Haartmaninkatu 3, PO Box 21, FI-00014
Helsinki, Finland. Tel.: 1358 919 126 377;
fax: 1358 919 126 382; e-mail:
Received 28 October 2010; revised 28 January
2011; accepted 9 May 2011.
Final version published online 16 June 2011.
DOI:10.1111/j.1567-1364.2011.00739.x
Editor: Richard Calderone
Keywords
resistance; ADH1; CDR1; CDR2; acetaldehyde.
Abstract
Expression of the alcohol dehydrogenase gene ADH1, which converts ethanol into
carcinogenic acetaldehyde, significantly inversely correlated with the expression of
CDR1 and CDR2, genes linked to azole resistance in Candida albicans isolated
from chronic oral candidosis in autoimmune polyendocrinopathy–candidosis–
ectodermal dystrophy (APECED, APS-I) patients. This is a novel link between
candidal two-carbon metabolism genes and azole resistance.
Introduction
Autoimmune polyendocrinopathy–candidosis–ectodermal
dystrophy (APECED), also called autoimmune polyendo-
crine syndrome type I (APS-I), is a rare autosomal recessive
disease causing T-cell-mediated dysfunction of the immune
system (Husebye et al., 2009). Most patients suffer from
chronic mucocutaneous candidosis of the oral and oesopha-
geal mucosa, which may result in the development of
squamous cell carcinoma (Rautemaa et al., 2007a). A high
oral and oesophageal carcinoma prevalence of 10.3% has
been reported among APECED patients over the age of 25
years (Rautemaa et al., 2007a).
Acetaldehyde is a highly toxic and mutagenic product of
alcohol fermentation and metabolism and it is especially
linked to cancers of the digestive tract (Secretan et al., 2009;
Uittamo et al., 2009). The latest consensus meeting of the
International Agency for Research on Cancer of the WHO
reclassified acetaldehyde as a class I carcinogen in association
with alcohol consumption (Secretan et al., 2009). According
to our previous results, Candida albicans isolates from
APECED patients can produce carcinogenic levels of acetalde-
hyde in vitro in both ethanol and glucose incubation (Uittamo
et al., 2009). Oxidation of ethanol into acetaldehyde is
catalysed by the alcohol dehydrogenase enzyme (Adh), which
is encoded by a single ADH gene. In C. albicans, only one Adh
isoenzyme has been identified, suggesting that one enzyme,
Adh1, operates in a bidirectional manner in both ethanol
fermentation and metabolism (Bertram et al., 1996).
Most APECED patients receive repeated courses of azole
antifungals and many have developed resistance, mainly to
fluconazole (Rautemaa et al., 2007b). Decreased fluconazole
susceptibility has been found to correlate with a high num-
ber of prophylactic and therapeutic courses of fluconazole,
ketoconazole and topical miconazole (Siikala et al., 2009).
The main molecular mechanism behind resistance in
FEMS Yeast Res 11 (2011) 494–498c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
YEA
ST R
ESEA
RC
H

C. albicans isolates from APECED patients is upregulation
of CDR1 and CDR2, the genes linked to azole resistance,
followed by gain-of-function mutations in the mutual
activator, the transcriptional activator TAC1 (Siikala et al.,
2010). Multidrug efflux transporters of the ATP-binding
cassette (ABC) superfamily and of the major facilitator class
play a key role in the low level of accumulation of azoles in
the yeast cell (Sanglard et al., 1995). CDR1 and CDR2 and
genes for ABC transporters have been shown to be upregu-
lated in resistant strains, leading to an enhanced efflux of the
drug (Sanglard et al., 1995). CDR2 expression is absent in
sensitive C. albicans strains, but its level of expression is
known to increase during the development of azole resis-
tance (Sanglard et al., 1997). Therefore, it can be seen as a
useful marker of CDR-mediated resistance.
In order to determine whether ADH1, the catalyst of
ethanol oxidation, is regulated in C. albicans isolated from
APECED patients with variable minimum inhibitory concen-
trations (MICs) to fluconazole, we analyzed the transcript
levels for ADH1. We also wanted to determine whether there
was a correlation between levels of expression of ADH1 and
fluconazole MICs as well as the expression of CDR1 and
CDR2, genes linked to azole resistance. As overexpression of
ADH1 is likely to lead to increased levels of carcinogenic
aldehyde; we also wanted to analyse whether CDR-mediated
fluconazole resistance correlated with ADH1 expression and
the possible risk for carcinogenicity of the infection.
Materials and methods
Candida albicans isolates
Candida albicans strains isolated during 1995–2007 from the
oral cavities of nine APECED patients suffering from CMC
were used in this study (one to three isolates/patient).
Strains that had been typed using multilocus sequencing
typing were selected (Siikala et al., 2010). Isolates with
identical diploid sequence type (DST) and MICs were
excluded to ensure that all isolates were independent.
However, isolates with similar DSTs and different MICs
were included to distinguish the role of different MICs in a
genetically closely related strain. A total of 19 strains
consisting of five resistant (R) strains (fluconazole
MICZ64 mg L�1), 11 susceptible dose-dependent (DD)
strains (fluconazole MIC 16 � MIC � 64 mg L�1) and three
susceptible (S) strains (fluconazole MIC � 8 mg L�1) were
included (National Committee for Clinical Laboratory
Standards, 2002). The small number of susceptible isolates
is due to decades of azole treatment leading to azole
resistance and lack of susceptible isolates in the APECED
patient population.
The C. albicans isolates had been identified from patient
samples using conventional culture and identification
methods at the Clinical Microbiology Laboratory of the
Helsinki University Central Hospital. The identification of
C. albicans was based on colony morphology on
CHROMagars Candida medium (CHROMagar, Paris,
France) and a negative Bichro-Dublis latex coagglutination
test result for Candida dubliniensis (Fumouze Diagnostics,
Levallois-Perret, France). The strains had been stored in
milk–glycerin at � 70 1C.
Growth media
The C. albicans strains were subcultured on YEPD agar [1%
Bacto peptone (Difco Laboratories, Basel, Switzerland),
0.5% yeast extract (Difco Laboratories) and 2% glucose
(Fluka, Buchs, Switzerland)].
Susceptibility testing
The susceptibility profiles of the isolates for fluconazole
were reanalysed by Etest (AB Biodisk, Sweden) according to
the manufacturer’s instructions and as described previously
(Arendrup et al., 2001). Quality control for susceptibility
testing was performed using C. albicans strains ATCC 90028
and ATCC 24433 and Candida glabrata strain ATCC 90030.
Northern blotting
Small-scale isolation of total RNA was performed from cells
grown to the logarithmic growth phase in YEPD medium at
30 1C with constant shaking. Northern blotting was per-
formed for CDR1, CDR2 and ADH1 as described previously
(Sanglard et al., 1995). RNA samples were separated by
agarose gel electrophoresis and transferred to a nitrocellu-
lose membrane using the Vacuum Blotting System (Hoefer
Scientific Instruments, San Fransisco, CA). Probes were
labelled with [a-32P]dATP with random priming using the
MegaPrime DNA Labelling System dNTP Kit (GE Health-
care, Waukesha, WI) according to the manufacturer’s in-
structions. CDR1, CDR2 and ACT1 probes were prepared
by PCR as described earlier (Sanglard et al., 1995). The
primers for the PCR-amplified ADH1-probe were forward,
50-GCAAGCTTATTCAGAATTTTCAGAGGTGC-30, and
reverse, 50-CAACTGGTGTCCAATACGTATCTACTCAAG-
30. Radioactive signals were revealed by exposure to a Kodak
BioMax MR film (GE Healthcare). Signals obtained in
blotted membranes were quantified by counting of radio-
activity (Typhoon Trio, GE Healthcare). As a control for the
evaluation of the expression levels of CDR1 and CDR2, the
membranes were hybridized with ACT1 and the amount of
RNA was normalized according to the expression of ACT1.
The strain with the lowest ADH1 expression was chosen as a
baseline control strain and the expression levels of the other
isolates were quantified relative to this strain.
FEMS Yeast Res 11 (2011) 494–498 c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
495ADH1 and CDR correlate in azole-resistant Candida
Statistical analysis
Data were analysed using GRAPHPAD PRISM version 5.00
(GraphPad Inc., San Diego, CA). Geometric means and
range were used for the analyses of MICs and the mean
and range for the analysis of gene expression levels. Spear-
man’s rho (rS) was used for the analyses of correlations.
Correlations are presented with a 95% confidence interval
(x to x) and P-value. P values of o 0.05 were considered
statistically significant.
Results
Susceptibility profiles of C. albicans isolates
The MICs for fluconazole of all the 19 C. albicans isolates
ranged from 1 to 128 mg L�1, with a geometric mean of
33 mg L�1 (1). The geometric mean of the three
susceptible isolates with fluconazole MICs � 8 mg L�1 was
3.63 mg L�1 (range 1–8). All susceptible isolates were
isolated from different patients (Patients 2, 4 and 5). The
geometric mean of the 11 susceptible DD isolates
with fluconazole MICs 16 � MIC � 64 mg L�1 was 37.9
(range 24–48). The geometric mean of the five resistant
with fluconazole MICsZ64 mg L�1 isolates was 91.6
(range 64–128).
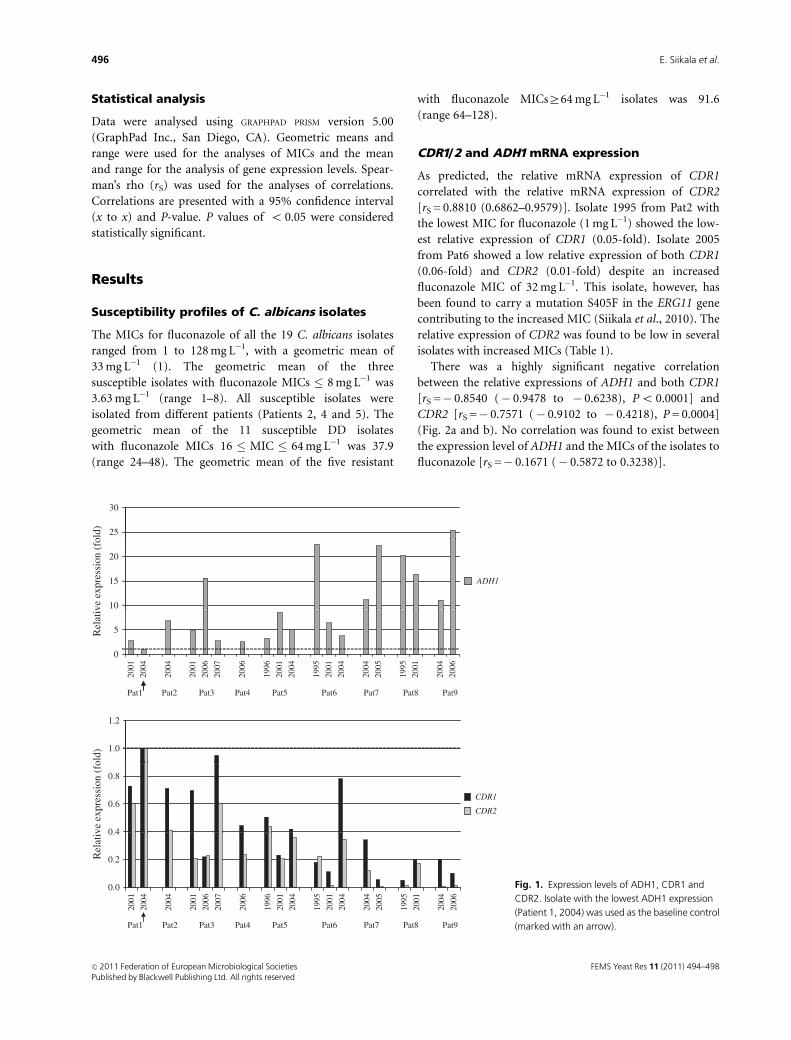
CDR1/2 and ADH1mRNA expression
As predicted, the relative mRNA expression of CDR1
correlated with the relative mRNA expression of CDR2
[rS = 0.8810 (0.6862–0.9579)]. Isolate 1995 from Pat2 with
the lowest MIC for fluconazole (1 mg L�1) showed the low-
est relative expression of CDR1 (0.05-fold). Isolate 2005
from Pat6 showed a low relative expression of both CDR1
(0.06-fold) and CDR2 (0.01-fold) despite an increased
fluconazole MIC of 32 mg L�1. This isolate, however, has
been found to carry a mutation S405F in the ERG11 gene
contributing to the increased MIC (Siikala et al., 2010). The
relative expression of CDR2 was found to be low in several
isolates with increased MICs (Table 1).
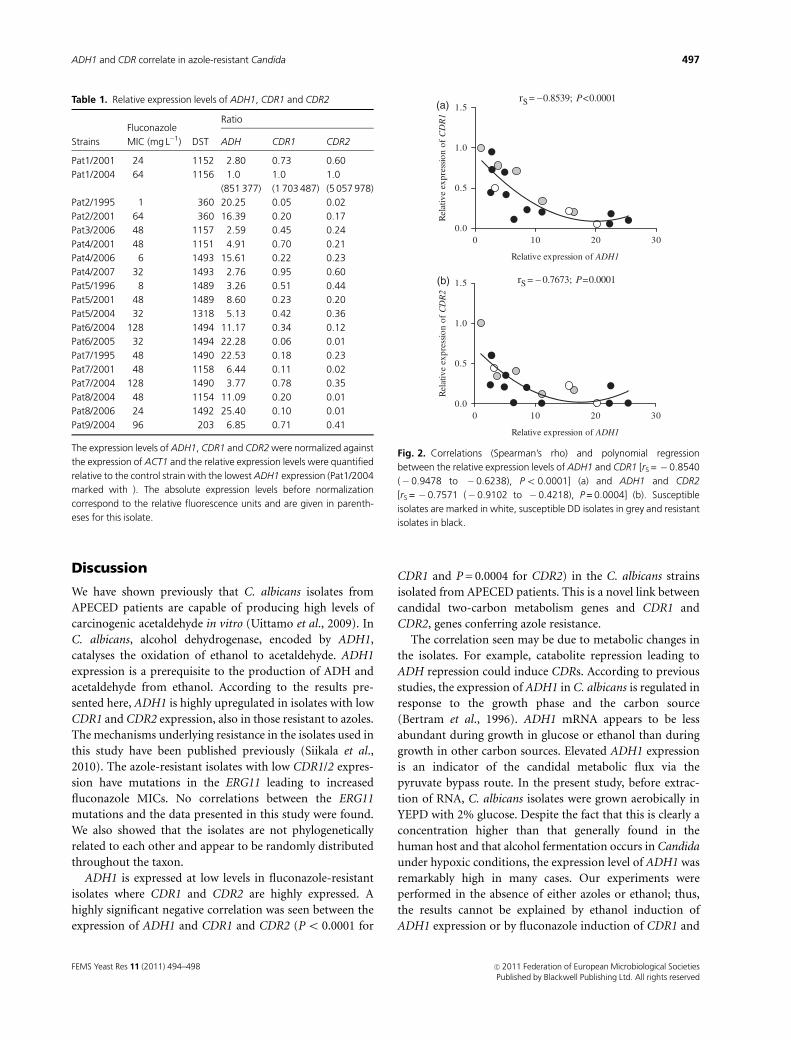
There was a highly significant negative correlation
between the relative expressions of ADH1 and both CDR1
[rS =� 0.8540 (� 0.9478 to � 0.6238), Po 0.0001] and
CDR2 [rS =� 0.7571 (� 0.9102 to � 0.4218), P = 0.0004]
(Fig. 2a and b). No correlation was found to exist between
the expression level of ADH1 and the MICs of the isolates to
fluconazole [rS =� 0.1671 (� 0.5872 to 0.3238)].
25
30
10
15
20
ADH1
0
5
2001
2004
2004
2001
2006
2007
2006
1996
2001
2004
1995
2001
2004
2004
2005
1995
2001
2004
2006
Rel
ativ
e ex
pres
sion
(fo
ld)
Rel
ativ
e ex
pres
sion
(fo
ld) 1.0
1.2
Pat1 Pat2 Pat3 Pat4 Pat5 Pat6 Pat7 Pat8 Pat9
0.6
0.4
0.8
CDR1
CDR2
0.0
0.2
001
004
004
0 01
006
007
0 06
996
001
004
9 95
001
004
004
005
995
001
0 04
006
2 0 20 20 20 20 20 20 19 20 20 19 20 20 20 2 0 19 20 20 20
Pat1 Pat2 Pat3 Pat4 Pat5 Pat6 Pat7 Pat8 Pat9
Fig. 1. Expression levels of ADH1, CDR1 and
CDR2. Isolate with the lowest ADH1 expression
(Patient 1, 2004) was used as the baseline control
(marked with an arrow).
FEMS Yeast Res 11 (2011) 494–498c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
496 E. Siikala et al.
Discussion
We have shown previously that C. albicans isolates from
APECED patients are capable of producing high levels of
carcinogenic acetaldehyde in vitro (Uittamo et al., 2009). In
C. albicans, alcohol dehydrogenase, encoded by ADH1,
catalyses the oxidation of ethanol to acetaldehyde. ADH1
expression is a prerequisite to the production of ADH and
acetaldehyde from ethanol. According to the results pre-
sented here, ADH1 is highly upregulated in isolates with low
CDR1 and CDR2 expression, also in those resistant to azoles.
The mechanisms underlying resistance in the isolates used in
this study have been published previously (Siikala et al.,
2010). The azole-resistant isolates with low CDR1/2 expres-
sion have mutations in the ERG11 leading to increased
fluconazole MICs. No correlations between the ERG11
mutations and the data presented in this study were found.
We also showed that the isolates are not phylogenetically
related to each other and appear to be randomly distributed
throughout the taxon.
ADH1 is expressed at low levels in fluconazole-resistant
isolates where CDR1 and CDR2 are highly expressed. A
highly significant negative correlation was seen between the
expression of ADH1 and CDR1 and CDR2 (Po 0.0001 for
CDR1 and P = 0.0004 for CDR2) in the C. albicans strains
isolated from APECED patients. This is a novel link between
candidal two-carbon metabolism genes and CDR1 and
CDR2, genes conferring azole resistance.
The correlation seen may be due to metabolic changes in
the isolates. For example, catabolite repression leading to
ADH repression could induce CDRs. According to previous
studies, the expression of ADH1 in C. albicans is regulated in
response to the growth phase and the carbon source
(Bertram et al., 1996). ADH1 mRNA appears to be less
abundant during growth in glucose or ethanol than during
growth in other carbon sources. Elevated ADH1 expression
is an indicator of the candidal metabolic flux via the
pyruvate bypass route. In the present study, before extrac-
tion of RNA, C. albicans isolates were grown aerobically in
YEPD with 2% glucose. Despite the fact that this is clearly a
concentration higher than that generally found in the
human host and that alcohol fermentation occurs in Candida
under hypoxic conditions, the expression level of ADH1 was
remarkably high in many cases. Our experiments were
performed in the absence of either azoles or ethanol; thus,
the results cannot be explained by ethanol induction of
ADH1 expression or by fluconazole induction of CDR1 and
Table 1. Relative expression levels of ADH1, CDR1 and CDR2
Strains
Fluconazole
MIC (mg L�1) DST
Ratio
ADH CDR1 CDR2
Pat1/2001 24 1152 2.80 0.73 0.60
Pat1/2004� 64 1156 1.0
(851 377)
1.0
(1 703 487)
1.0
(5 057 978)
Pat2/1995 1 360 20.25 0.05 0.02
Pat2/2001 64 360 16.39 0.20 0.17
Pat3/2006 48 1157 2.59 0.45 0.24
Pat4/2001 48 1151 4.91 0.70 0.21
Pat4/2006 6 1493 15.61 0.22 0.23
Pat4/2007 32 1493 2.76 0.95 0.60
Pat5/1996 8 1489 3.26 0.51 0.44
Pat5/2001 48 1489 8.60 0.23 0.20
Pat5/2004 32 1318 5.13 0.42 0.36
Pat6/2004 128 1494 11.17 0.34 0.12
Pat6/2005 32 1494 22.28 0.06 0.01
Pat7/1995 48 1490 22.53 0.18 0.23
Pat7/2001 48 1158 6.44 0.11 0.02
Pat7/2004 128 1490 3.77 0.78 0.35
Pat8/2004 48 1154 11.09 0.20 0.01
Pat8/2006 24 1492 25.40 0.10 0.01
Pat9/2004 96 203 6.85 0.71 0.41
The expression levels of ADH1, CDR1 and CDR2 were normalized against
the expression of ACT1 and the relative expression levels were quantified
relative to the control strain with the lowest ADH1 expression (Pat1/2004
marked with�). The absolute expression levels before normalization
correspond to the relative fluorescence units and are given in parenth-
eses for this isolate.
rS= −0.8539; P<0.0001
0 10 20 300.0
0.5
1.0
1.5
Relative expression of ADH1
Rel
ativ
eex
pres
sion
ofC
DR
1
rS = − 0.7673; P=0.0001
0 10 20 300.0
0.5
1.0
1.5
Relative expression of ADH1R
elat
ive
expr
essi
onof
CD
R2
(a)
(b)
Fig. 2. Correlations (Spearman’s rho) and polynomial regression
between the relative expression levels of ADH1 and CDR1 [rS = �0.8540
(� 0.9478 to �0.6238), Po 0.0001] (a) and ADH1 and CDR2
[rS = � 0.7571 (�0.9102 to � 0.4218), P = 0.0004] (b). Susceptible
isolates are marked in white, susceptible DD isolates in grey and resistant
isolates in black.
FEMS Yeast Res 11 (2011) 494–498 c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
497ADH1 and CDR correlate in azole-resistant Candida

CDR2 expression. Interestingly, progesterone exposure has
been shown to upregulate CDR1 and CDR2 expression and
to a lesser extent ADH1 expression in C. albicans (Banerjee
et al., 2007).
ADH1 has been shown to be downregulated in Candida
biofilms and it has been suggested to play a role in quorum
sensing during their formation (Chandra et al., 2001;
Mukherjee et al., 2006; De Monte et al., 2007). Biofilm
formation contributes to antifungal resistance in Candida
(Chandra et al., 2001). In our study, however, experiments
were performed with planktonic cells and low ADH1
expression by some isolates cannot be explained by biofilm
formation. It is possible that the expression of both ADH1
and CDR1 is affected by the same regulator. Regardless of
the mechanism, azole exposure resulting in CDR upregula-
tion could lead to ADH1 downregulation and decreased
exposure carcinogenic acetaldehyde. The mechanisms
behind this phenomenon will require further study.
Acknowledgements
We thank Francoise Ischer for her technical help and
expertise and Professor Seppo Sarna for statistical assistance.
The study was supported by grants from Helsinki University
Central Hospital, the Finnish Dental Association Apollonia,
the Yrjo Jahnsson Foundation, the Daisy and Yrjo Eskola
trust fund of the Paulo Foundation and the University of
Helsinki.
References
Arendrup M, Lundgren B, Jensen IM, Hansen BS & Frimodt-
Møller N (2001) Comparison of Etest and a tablet diffusion
test with the NCCLS broth microdilution method for
fluconazole and amphotericin B susceptibility testing of
Candida isolates. J Antimicrob Chemoth 47: 521–526.
Banerjee D, Martin N, Nandi S, Shukla S, Dominiquez A,
Mukhopadhyay G & Prasad R (2007) A genome-wide steroid
response study of the major human fungal pathogen Candida
albicans. Mycopathologia 164: 1–17.
Bertram G, Swoboda RK, Gooday GW, Gow NAR & Brown AJP
(1996) Structure and regulation of the Candida albicans ADH1
gene encoding an immunogenic alcohol dehydrogenase. Yeast
12: 115–127.
Chandra J, Kuhn DM, Mukherjee PK, Hoyer LL, McCormick T &
Ghannoum MA (2001) Biofilm formation by the fungal
pathogen Candida albicans – development, architecture and
drug resistance. J Bacteriol 183: 5385–5394.
De Monte S, d’Ovidio F, Danø S & Sørensen PG (2007)
Dynamical quorum sensing: population density encoded in
cellular dynamics. P Natl Acad Sci USA 104: 18377–18381.
Husebye ES, Perheentupa J, Rautemaa R & Kampe O (2009)
Clinical manifestations and management of patients with
autoimmune polyendocrine syndrome type I. J Intern Med
265: 514–529 (review).
Mukherjee PK, Mohamed S, Chandra J et al. (2006) Alcohol
dehydrogenase restricts the ability of the pathogen Candida
albicans to form biofilm on catheter surfaces through an
ethanol-based mechanism. Infect Immun 74: 3804–3816.
National Committee for Clinical Laboratory Standards (2002)
Reference Method for Broth Dilution Antifungal Susceptibility
Testing of Yeasts – Second Edition: Approved Standard M27-A2.
NCCLS, Wayne, PA.
Rautemaa R, Hietanen J, Niissalo S, Pirinen S & Perheentupa J
(2007a) Oral and oesophageal squamous cell carcinoma – a
complication or component of autoimmune
polyendocrinopathy–candidiasis–ectodermal dystrophy
(APECED, APS-I). Oral Oncol 43: 607–613.
Rautemaa R, Richardson M, Pfaller M, Koukila-Kahkola P,
Perheentupa J & Saxen H (2007b) Decreased susceptibility of
Candida albicans to azole antifungals: a complication of long-
term treatment in autoimmune polyendocrinopathy–
candidiasis–ectodermal dystrophy (APECED) patients.
J Antimicrob Chemoth 60: 889–892.
Sanglard D, Kuchler K, Ischer F, Pagani J-L, Monod M & Bille J
(1995) Mechanisms of resistance to azole antifungal
agents in Candida albicans isolates from AIDS patients
involve specific multidrug transporters. Antimicrob Agents
Ch 39: 2378–2386.
Sanglard D, Ischer F, Monod M & Bille J (1997) Cloning of
Candida albicans genes conferring resistance to azole
antifungal agents: characterization of CDR2, a new multidrug
ABC transporter gene. Microbiology 143: 405–416.
Secretan B, Straif K, Baan R et al. (2009) A review of human
carcinogens – Part E: tobacco, areca nut, alcohol, coal smoke,
and salted fish. Lancet Oncol 10: 1033–1034.
Siikala E, Richardson M, Pfaller MA, Diekema DJ, Messer SA,
Perheentupa J, Saxen H & Rautemaa R (2009) Candida
albicans isolates from APECED patients show decreased
susceptibility to miconazole. Int J Antimicrob Ag 34: 607–609.
Siikala E, Rautemaa R, Richardson M, Saxen H, Bowyer P &
Sanglard D (2010) Persistent Candida albicans colonization
and molecular mechanisms of azole resistance in APECED.
J Antimicrob Chemoth 65: 2505–2513.
Uittamo J, Siikala E, Kaihovaara P, Salaspuro M & Rautemaa R
(2009) Chronic candidosis and oral cancer in APECED-
patients: production of carcinogenic acetaldehyde from
glucose and ethanol by Candida albicans. Int J Cancer 124:
754–756.
FEMS Yeast Res 11 (2011) 494–498c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
498 E. Siikala et al.





























