Embed Size (px)
Citation preview

(2?)UNIVERSIDAD COMPLUTENSEDE MADRID
FACULTAD DEVETERINARIA
DEPARTAMENTODENUTRICION Y BROMATOLOGIA III
(HIGIENE Y ThCNOLOGIADE LOS ALIMENTOS)
Áeromonasspp.MOVILES:
FACTORESDE VIRULENCIA EN CEPASAISLADAS
DE ALIMENTOS Y DE HECESDIARREICAS HUMANAS
Memoriaqueparaopwal gradode DoctorenVeterinariapresentala LicenciadaM~ CarmenPm Arias.
Madrid.julio dc 1995.

TELE. 34 394 31 49
rAx: 34-9,1 -29437 43
UNIVERSIDAD COMPLUTENSE DE MADRID
FACULTAD DE VETERINARIA
OPTO. DE NUTRICION Y BROMATOLOGíA IIIHIGIENE Y TECNOLOGíA DE LOS ALIMENTOS
CIUDAD UNIVERSITARIA
26040 MADRID
JOSETORMO IGUACEL Y CARMEN CASAS VALENCIA, PROFESORESDE
NUTRICION Y BROMATOLOGIA DE LA FACULTAD DE VETERINARIA DE LA
UNiVERSIDAD COMPLUTENSEDEMADRID,
CERTIFICAN:
Que la tesis doctoraltitulada ‘¾4eromonasspp. móviles: Factores de
virulencia en cepasaisladasde alimentosy de hecesdiarreicashumanas’.dela queesautorala Licenciadaen VeterinariaM’ CarmenPm Arias,se harealizadoen el
DepartamentodeNutrición y BromatologíaIII (Higieney Tecnologíade los Alimentos)
bajo ladirecciónconjunta delos quesuscribeny cumple lascondicionesexigidas para optar
al título de DoctorenVeteriimrit
Madrid, 14 dejulio de 1995
CVa’ Sr tao
Fdo.: JoséTormoIguacel Fdo.:Carmen CasasValencia

A Tuco
A mispadres

Algunas vecesbe creídobasta
seiscosasimposiblesantesdel
desayuno
La Reina Blanca
“Alicia atravésdelespejo”

AGRADECIMIENTOS
Al ProfesorDr. José TormoIguacel.a quien estatesisselo debe todoy yo le debo
muchomás.
A la ProfesoraDra. Carmen Casas Valencia, por suinestimabley siempreacertada
asistenciay supervisión.
Además, quiero darleslas gracias por el tratoy el cariñoque he recibido en estos
cuatroaños.
Al ProfesorDr. BernabéSanzPérezporacogermeen elDepanamentoquedirigía en
el momentode mi incorporacióny por el interésdemostradoen mitrabajoy formación.
Al ProfesorDr. JuanAntonio OrdóñezPereda,actualDirector del Departamento,por
su accesibley desinteresadaayuda,prestadaen todo momento.
Al ProfesorDr. GonzaloGarcíade FemandoMinguillón, que con sugenerosidady
clarividenciahafacilitado,de formamuy importante,la realizaciónde estetrabajo.
A la Profesora Dra.PalomaMorales,porsuinestimableayuda en laejecucióndelos
ensayosde citotoxicidady, en lamisma línea,a los Profesores Dres.NP IsabelCambero,
Lorenzode la Hoz, Juan Miguel Rodríguez,Juan1. Azcona,W LuisaGarcíay Dolores
Selgasporecbarmeunamano, aclarar mis dudasy, sobretodo,porsuamistad.
También,quieroexpresarmi agradecimientoa todas las personascon las que he
convivido en este Departamentopor los consejos,el apoyo, la amistad, el humory los
buenosratos,en especial, a Gonzalo Anguita,SergioMano, IsabelGonzález, Manuela
Fernándezy Daniel López.
Al Dr. JorgeReina, médico del HospitalSon Duretade Palmade Mallorca, por
proporcionarmelas cepasdeAeromonasspp.aisladasen hecesdiarreicashumanas.
También, quiero agradecer la colaboraciónde los miembrosdel ServicioCentral de
MicroscopiaElectrónicade la Universidad Complutensey del Serviciode Microscopia
Electrónicade la Facultadde Veterinaria,especialmente,a Agustín Fernándezy a Ana
Vicente.

ME
ab
y
ME
u’
Vb
Al Ministerio de Educacióny Cienciapor la concesiónde una becadel Programa
Generalde Formaciónde Personal Investigador.
A Salva,aLourdesy al restode misamigos.
A mis padres,a mis hermanos,aManolo,aEsther,a JoséJuan,a Matilde,a Kiko y a
Mana porsu cariñoy confianza.
Por último a Tuco, por su colaboracióntantoen la parteexperimentalcomo en la
escrituradeestatesisy, sobretodo porque,sorprendentemente,mesiguesoportando.

INDICE

INDICE
Página
1.-INTRODUCCION 1
1.1. - Taxonomía 4
1.1.1. - Precedentes históricos 4
1.1.2. - Clasificación 7
1.1.2.1.-Gruposde hibridación: identificación bioquímica 9
1.2. - Aislamiento e identificación de Áeromonasspp. móviles 14
13. - Epidemiología u
1.3.1. - Heces humanas ¡y
1.3.2. - Alimentos: Factoresque influyen en la supervivencia
y en el crecimiento 19
1.3.3. Procesos clínicos 22
1.3.3.1. - Gastroenteritisasociadasa alimentos 22
1.3.3.2. - Infecciones extraintestinales 23
1.3.4. Agua y portadores animales 24
1.4. - Factores de virulencia 27
1.4.1. - Productos exocelulares 27
1.4.1.1. - Exotoxinas 27
1.4.1.1.1. - Enterotoxina citotónica 28
1.4.1.1.2. - Enterotoxinacitotóxica 29
1.4.1.1.3. - Hemolisinas 31
1.4.1.1.4. - Mecanismode acciónde la 13-hemolisina 32
1.4.1.1.5.- Factor que presentareaccionescruzadascon la toxina del
cólera (CTC) 33

9>
ir
u’
ev
U)
e
u’
a
t
INDIcE
1.4.1.2. - Proteasas 34
1.4.1.3. - Sideróforos 34
¡.4.2. - Característicasasociadasa la célula bacteriana 35
1.4.2.1. - Fimbrias o pili 35
¡.4.2.2.- Proteínasde la membrana externade la paredbacteriana
(OMPs) 38
1.4.2.3. - Lipopolisacáridosde la membrana externade la pared
bacteriana (LPSs) 39
¡.4.2.4. - Flagelo 39
1.4.2.5. - Capa extra, proteicay externaa la paredbacteriana(capa 5).... 40
1.4.2.6. - Invasividad 42
1.4.2.7. - Resistenciaal suero 42
1.4.2.8. - Plásmidos 43
II. - OBJETIVOS DEL TRABAJO 45
III. - MATERIAL Y METODOS 49
111.1. - Material y equipo 51
111.2. - Productos biológicos 52
111.2.1. - Microorganismos 52
111.2.2. - Alimentos 52
111.2.3. - Aguas 52
111.2.4. - Preparados hematológicos 52
111.2.5. - Lineas celulares estables 54
111.3. - Reactivos 54
111.4. - Métodos 54
fi

INDICE
111.4.1. - Preparaciónde los medios de cultivo 54
111.4.2. - Preparaciónde soluciones 54
111.4.3. - Recuentos microbianos 54
111.4.4. - Aislamiento de colonias 55
111.4.5. - Toma de muestras 55
111.4.6. - Mantenimiento de las cepas 55
111.4.7. - Revitalización de las cepas 55
111.4.8. - Manejo de líneas celulares estables 56
111.4.8.1. - Mantenimiento 56
111.4.8.2. - Recuentoscelulares 56
¡[1.4.8.3. - Cultivo de células Vero 56
111.4.8.4. - Cultivo de células de mieloma 57
111.4.9. - Técnicapara compararel comportamientode los
mediosselectivos(AAA, AGAP y mA) en el
aislamiento de Aeromonas spp.móviles 58
IH.4.10. - Técnicapara investigarel comportamientode A.
sobria CECT 837 en los medios selectivos(AAA,
AGAP y mA) modificados 61
111.4.10.1. - Antibiograma 62
111.4.11. - Técnicasde enriquecimiento,aislamiento e
identificación de Aeromonas spp. móviles en
aguas y alimentos 63
111.4.11.1. - Enriquecimientoy aislamiento 63
111.4.11.2.- Identificación bioquímica de géneroy especie 64
111.4.11.2.1. - Pruebas 64
111.4.11.2.1.1. - Oxidasa 64
111.4.11.2.1.2.- Fermentaciónde glucosa
(Agar con hierro de Kligler) 65
111.4.11.2.1.3. - Gram 65
111.4.11.2.1.4. - ONPG 65
u’

E.
u
*
e
e
e
a-
aL
e
INDICE
111.4.11.2.1.5. - ONasa . 66
11L4.1l.2.l.6. - Catalasa 66
111.4.11.2.1.7. - Crecimiento a 370C 66
111.4.11.2.1.8.- Crecimiento sin CINa 66
111.4.11.2.1.9.- Resistenciaal agentevibriostático 0/129 67
11L4.l1.2.l.lO. - Hidrólisis del almidón 67
11L4.11.2.1.11.- Fennentaciónde azúcares(D-manitol. i-inositoly
salicina) 67
111.4.11.2.1.12.- Descarboxilaciónde la oniitinay (o dihidrólisis)
de la arginina 68
111.4.11.2.1.13.- Producciónde H2S del tiosulfato sádico 68
111.4.11.2.1.14.- Producción de ureasa 68
111.4.11.2.1.15.- Producciónde gasa partir de glucosa 69
111.4.11.2.116.- Hidrólisis de la esculina 69
111.4.11.2.1.17.- Voges Proskauer 69
111.4.11.2.1.18.- Crecimientocon cianuropotásico 70
111.4.12. - Técnica para la estimación de la resistencia al cloro.... 70
1114.13. - Técnicas para la estimaciónde la resistenciaen
distintas condiciones de cultivo(Temperatura,cloruro
sódico, pH y nitrito sódico 70
¡11.4.14. - Técnicaspara la determinaciónde características
relacionadascon la virulencia en Aeromonasspp.
móviles 72
111.4.14.1. - Factores extracelulares 72
111.4.14.1.1. - Obtenciónde sobrenadantes 72
111.4.14.1.1.1.- A distintastemperaturasy tiempos 72
111.4.14.1.1.2.- Tratadospor calor 73
111.4.14.1.1.3.- Concentradosy fraccionadospor diafihración 73
lv

INDICE
111.4.14.1.1.4.- Correspondientesa las distintasfasesdel
crecimiento bacteriano 77
111.4.14.1.2. - Pruebashemolíticas 77
111.4.14.1.2.1.- En agar sangrede caballo o cordero 77
111.4.14.1.2.2.- En eritrocitosde conejoo corderoen suspensión... 77
111.4.14.1.2.3.- En eritrocitos de conejoincluidosen agar 78
111.4.141.3. - Pruebasoroteolíticas 78
111.4.14.1.3.1. - Caseína 78
111.4.14.1.3.2. - Elastina 79
111.4.14.1.4. - Pruebaslipolíticas 79
111.4.14.1.5.- Pruebasde toxicidad sobre cultivoscelulares 81
111.4.14.1.5.1. - Células Vero 81
111.4.14.1.5.2.- Células de mieloma 85
111.4.14.2. - Propiedades superficiales 86
111.4.14.2.1.- Hemaglutinación con eritrocitosde corderoy conejo 86
111.4.14.2.2.- Inhibición de la hemaglutinaciónpor0-manosa,
L-fucosa y D-galactosa 87
111.4.14.2.3. - Autoaglutinación 87
111.4.14.2.4.- Aglutinación con acriflavina 88
111.4.14.2.5.- Pruebaspara estimar lahidrofobicidadcelular 88
111.4.14.2.5.1.- Unión a filtros de nitrocelulosa(FNC) 90
111.4.14.2.5.2.- Adsorción ahidrocarburos(n-hexadecano,n-octano
y p-xileno) 90
111.4.14.2.6.- Inclusiónde las muestraspara la observación delas
estructurasbacterianasconel microscopioelectrónico
de transmisión 91
111.4.14.3. - Característicasbioquímicas 93
111.4.14.3.1. - Voges Proskauer 93
111.4.14.3.2.- Fermentaciónde la arabinosa 94
‘Vv

Mv
e,
ME
a
ME
u’
U)
a-
a-
a-
ir
e
a-,
u’
0
tú
INDICE
111.4.14.3.3. - Descarboxilaciónde la Usina 94
111.4.15. - Análisis estadístico 94
IV. RESULTADOS Y DISCUSION 95
IVA. - Medios de cultivo específicosnara el aislamientode
Aeromonas spp. móviles 97
lvii. - Recuperación de Acromonasspp. móviles e inhibición
de la flora natural de los alimentos en los medios mA,
AGAP y AAA 97
IVA.2. - Comportamiento de A. sobria CECT 837 sobre los
medios mA, AGAP y AAA 99
¡Vi. - Aislamiento e identificación de Aeromanasspp. móviles ... 102
IV.2.11. - Pruebasbioquímicasseleccionadaspara su
identificación y diferenciación 102
IV.2.2. Influencia del almacenamiento de los alimentos a baja
temperatura en su aislamiento 104
IV.2.3. - Incidencia en aguas y alimentos 104
IV.2.3.1. - Aguas 104
IV.2.3.2. - Carnes 105
FV.2.3.3. - Pescadosy mariscos los
IV.2.3.4. - Leche cruda y queso fresco 106
IV.2.3.5. - Ensaladaspreparadas 106
IV.2.4. - Distribución en especiesde las cepasaisladasen los
distintos alimentos 107
IV.3. - Resistencia al cloro de seis cepasdiarreicas y seis
alimentarias de Aeromonas spv. móviles 107

INDICE
IV.4. Influencia de diferentes parámetros exógenossobre elcrecimientode seis cepasdiarreicasy seis alimentariasde
4eromonas son. móviles
IV.4.1.
lV.4.2.
IV.4.3.
IV.4.4.
IV.4.5.
- Temperatura
- Cloruro sódico
-pH
- Nitrito sódico
- Efecto combinado de los parámetros ensayados
IVA. - Factores de virulencia de Aeromonas spp. móviles
IV.5.1. - Productos exocelularesde cepasdiarreicas y
alimentarias
IV.5.1.1. - Hemolisinas
108
108
lb
110
111
111
113
113
113
IV.5.1.1.1.-
IV.5.1.1.2.-
IV.5.l.I.3. -
IV.5.1.1.4.-
Hemolisisdesarrolladaporlas colonias bacterianas sobre
agar sangre
Hemolisisdesarrolladaporel sobrenadantecompletolibre
de célulassobre eritrocitosen suspensión
Hemolisis desarrollada por elsobrenadantelibre de
células,concentradoy fraccionadopordiafiltración,sobre
eritrocitos de conejo
Evoluciónde la producciónde hemolisinas,activas sobre
eritrocitosde conejo, enfuncióndelcrecimiento
bacteriano
IV.5.1.2. - Enzimas (proteasasy lipasas)
IV.5.l.2.l. - Proteasas
113
114
117
118
119
119
Vn

‘E
e,
e.
ir
ME
a.
ev
*1
*
a
U)
u,
ME
e-
e
ME,
ev
*7
ME
INDICE
IV.5.l.2.l.l. -
IV.5. 1.2.1.2.-
IV.5.l.2.1.3. -
IV.5. 1.2.2. -
IV.5.1.2.2.l.-
JV.5.1.2.2.2.-
IV.5.l.2.2.3. -
Caseinolisisy elastinolisis desarrolladaporel
sobrenadante completolibre de células
Caseinolisisdesarrolladaporel sobrenadantelibre
decélulas,concentradoy fraccionadopor
diafiltración
Evoluciónde la produccióndecaseinasaen función
del crecimiento bacteriano
Lioasas
Lipolisis desarrollada porelsobrenadantecompleto
libre de células
Lipolisis desarrollada por elsobrenadantelibre de
células,concentradoy fraccionadopordiafiltración..
Evoluciónde la producciónde lipasasen función
del crecimiento bacteriano
IV.5.1.3. - Citotoxinas
IV.5.1.3.1.
IV.5.1.3.2.
- Citotoxicidaddesarrolladapor elsobrenadantecompleto
libre decélulassobre líneascelularesestables
- Citotoxicidaddesarrolladaporel sobrenadantelibre de
células, concentradoy fraccionadopordiafiltración, sobre
células Vero
IV.5.2. - Propiedadessuperficialesde cepasdiarreicas y
alimentarias
IV.5.2.1.
IV.5.2.2. -
IV.5.2,3.
IV.5.2.4.
IV.5.2.5.
- Hemaglutinaci6n
Inhibición de la hemaglutinación porD-manosa,L-fucosa
y O-galactosa
• Autoaglutinación
- Aglutinación con acriflavina
- Hidrofohicidad superficial
119
119
120
121
121
121
122
122
123
125
126
126
127
129
130
130
VIII

INDICE
IV.5.2.5.l. - Unión a filtros de nitrocelulosa 131
IV.5.2.5.2. - Adsorción a hidrocarburoslfguidos 131
IV.5.2.5.2.1. - Al hexadecano 131
IV.5.2.5.2.2. - Al octano 132
IV,5.2.5.2.3. - Al xileno 132
IV.5.2.6. - Aspectosmorfológicosde la superficiebacteriana 133
IV.5.2.6.l. - Capa S 133
IV.5.2.6.2. - PiIi 134
IV.5.3. - Características bioquímicas de cepas diarreicas y
alimentarias 135
IV.5.3.1. - Voges Proskauer 13é
IV.5.3.2. - Descarboxilaciónde la lisina 136
IV.5.3.3. - Fermentaciónde la arabinosa 136
IV.t4. - Asociacionesdetectadasentre los factores indicadores
de virulencia 137
IVa. - ANEXO (Tablas y Figuras) 141
y. - CONCLUSIONES 207
VI. - BIBLIOGRAFIA 211

1. INTFIOOUCCIQN

ab
Mv
a-
ab
u,
u’
0
a
U)
e
ME.
es
e
e
e;
a
0
u,

IMrRODUCCION
En la décadade los 80 tienelugarunaexplosiónde interés científicode los miembros
del géneroAeromonascomopatógenosparael hombrey animalesdebido,principalmente.
a la asociaciónde estas bacterias con procesosgastroentéricoshumanos.Actualmente,
existenotrasáreasde interéscreciente, comola complejataxonomíay los factores asociados
a lavirulencia,potencialmenteresponsablesde infeccioneshumanasy animales.
En distintaspartes delmundo sehan descritoprocesosgastroentéricosasociados,
aunqueno necesariamentecausados porAeromonasspp. (Graceyy col., 1982;Janday
Duffey, 1988; Altweggy Geiss,1989);al parecerestaasociaciónes más frecuente enniños
menoresde 2 aflos, adultos mayoresde 50 añosy personasinmunodeficientes(Burke y
col., 1983a;Agger, 1986; SanJoaquiny Pickett, 1988; Kuijper y col., 1989a;Gluskin y
col., 1992)habiéndose detectadoun mayornúmerode casos enverano (Burkey col.,
1984b;Aggery col., 1985; Nishikaway Kishi, 1988;Wilcox y col., 1992).En general, las
cepasposeenfactores indicadoresde virulencia como enterotoxinas, citotoxinas,
hemolisinas,proteasasy o capacidadinvasiva,aunqueno está clara larepercusiónrelativa
de estaspropiedades en lapatogenicidad,de modoque no todaslas cepasque las poseen
parecenservirulentas,siendo necesaria la combinaciónde estaspropiedadesbacterianas
con factorespredisponentesen el hospedador paraque aparezca ladiarrea (Kirov,1993).
La hospitalización,terapia antimicrobiana, neutralización del ácido gástricoo inhibición dela secreciónácida,enfermedadeshepáticaso desórdenesintestinales,asícomolascirugías
gástricao entérica,cáncerde colon, hemorragiasgastrointestinalese inflamaciones
idiopáticasseconsideranfactorespredisponentesporpartedel hospedador(Goodwin y
col., 1983;Georgey col., 1985; Moyer, 1987).
No existeun modeloanimalquepermitareproducir ladiarreaasociada aAeromonas,
por lo quelos postuladosde Koch,necesariosparaconsideraraun microorganismocomo
agente causalde unaenfermedadinfecciosa,no se cumplen(Janda, 1991).Pazzagliay col.
(1994)hanintentandoestablecerun modeloanimal administrando108ufe/dla durante 4días
a ratonesalimentados con dietas hipoproteicas,sin detectarsíntomasde enfermedaden
ningún animal. Morgany col. (1985),observanque sólo dospersonasde 57 voluntarios
sanosdesarrollaron diarreatras ingerir dosis superiores a1010 bacterias procedentesde
hecesdiarreicasy concapacidadtoxigénica.Sin embargo,en este estudio no se tienen en
cuentalas propiedades adhesivasde las cepaso el nivel de inmunidadde estas personas.
La principal fuentede infecciónpareceser elagua.Así, se handetectado recuentos
elevadosde Aerornonasen, prácticamente,todos los tipos de aguas,inclusoen las
sometidasa cloración (Hazen y col., 1978; Burke y col., 1984b; Schubert,1991).
considerándoseel aguano tratadacomo un importantefactorde riesgo (Moycr, 1987>.
3

e,
Mv
Mv
e,
WC
Mv
9>
Mv
e
I,VrRODUCCION
El aislamientode estosmicroorganismosa partir dealimentosesun hechofrecuente
(Callistery Agger, 1987;Okrendy col., 1987;Abeytay Wekell, 1988;Nishikaway Kishi,
1988; Palumboy col., 1989) por lo que, potencialmente,puedenincluirse dentrode los
patógenoscausantesde toxiinfeccionesalimentarias(Buchanany Palumbo,1985;Morgany
Wood, 1988; Wadstil5my Ljungh. 1991). Además, Palumbo(1986)y Beuchat(1991)
entre otros autores, compruebanque ciertas cepas, inclusoenterotoxigénicas,son
psicrótrofaspor lo quela refrigeraciónde los alimentosno seríasuficientepara frenar su
desarrollo.
Numerososinvestigadoreshan detectadoenterotoxinas producidasporestegrupo
bacteriano(Burke y col., 1981b;Palumboy col., 1987; Houstony col., 1991; Condony
col., 1992) incluso enalimentos refrigerados(Majeedy MacRae,1991).Esto supondría
que la presenciade un número elevadodeAeromonasenalimentosno sólo significarfaun
riesgo de infección, sino tambiénde intoxicación. No obstante,los pocos casosde
gastroenteritisasociadosaAeromonasregistradoshastael momento,presentanun periodo
de incubacióndemasiado largo (> 6h.)como paraserreflejo de unaintoxicación (Tabla
1.1).
El controvertido papeldeAeromonasspp.móviles en las toxiinfeccionesalimentarias
es una realidaddesdeque losestudiosmicrobiológicos.epidemiológicos,clínicos e
inmunológicosindicanque,al menos algunas cepas,sonenteropatógenas.
1.1. - Taxonomía
1.1.1. - Precedenteshistóricos
El primerautorque describióun miembropertenecienteal actualgéneroAeromonas
fue, probablemente,Zimmennarrn(1890). Esteinvestigadoraisléunabacteriadel agua
potablede la ciudadde Chemnitzquesecaracterizabaporsucrecimientopuntiformeen agar
gelatina,por lo que la denominóBacillus puncratus.En añosposteriores,varios autores
describieroncepas similares,procedentesde distintos orígenesacuáticos(Franklandy
Franldand,1889; Burckhardt,1917) y su denominación pasóde Bacterium punctarum
(Lehmanny Neumann,1899)aAchronzobacíer punctatum(Bergeyy col., 1923), siendo,
posteriormente,transferidaal géneroPseudomonas,como Rs. punctata (Scbliperclaus,
1930; Hitchner, 1948). Finalmente,en la séptima edicióndel Manual Bergey(Snieszko,
1957)estemicroorganismoseregistracomoAeromonaspunetata.
Un año despuésde la publicaciónde Zimmermann(1890).Sanarelli(1891),aisló un
4

a’
oC
COC
‘osc
a’—
—O
CO
C—
a’
¿e~
——
.~.~
*cd
c~O
QN
cdc
c
eO
~—
>.‘n
~5
~.E
E
>-.
>‘~
cdcd
~<
.0.0
—,
<O
O
cd—
5
O—
—‘e
uci~
——
ci,,-~
O~
•-‘e
00
¡O
’eO
uci,os
os~‘
~‘.—
oo
o
0
uo
*OCQNu,
eo00
‘Ji
*.*
—~
OQN
.t.c
a~
oso..
ct
CC
OO
O>
~
.c.c
0000
——
o,
OO
ct,‘e
‘ecd
.—.0
28
E
bo
o~
cdC
dCJ
-4-4
OCt
ci,o
o?
en
Ito
3c-~
~-.N
i00
00
C4y
~A
sc
cdCu
cl
~r—
en‘n
Oen
OO
—Q
NIt
Aen
~.~
cl
O
O
‘Oe‘o
—.
rscd...s
O~c~j
ucd
Ccd
E~
99
9~
.~.~
~5
~-
e~
c.~o
,o
~5
55
.c~
x~
o—
cC
C.0
uu:
-~
u:
cdte5-e
ueeE
eu.-1>1
1<
41
FC.1-1u
eE-ie‘oE0<o,
tu..
euouFCu-o81~cdu
•5
.—a
eosaaeFCuE-3-uu-4
o’>5
‘jioc’>
‘Oo
.~z
u:
5

a-
a-
a-
a-
a-
a-
a-
a-
WC
a-
0
IWFRODUCCI’ON
bacilo de sangrey linfa de rana,quedesignécomoBacillus hydrophilusfuscas.El estudio
comparativode estemicroorganismocon la descripciónmicrobiológica¡ralizadaporErnst
(1890) dclbacilo Bacilasranicida (agentecausaldela enfermedadprimaveralde las ranas),
llevó aSanarellia la conclusióndequelas dosespecies debíande seríasmismas.Además,
este autorrechazóel términoranicida, yaque estebacilo tambiénpodíaseragenteinfeccioso
de pecesy animalesde sangrecaliente.En el Manual Bergey (Bergeyy col., 1923)a esta
especiese ledenominaProreus hydrophilus,manteniéndosecon estenombre(Reedy
Toner, 1942; Kulp y Borden, 1942; Guthrie y Hitchner,1943) hasta que setransfirió al
géneroPseudomonas(Hitchner, 1948). En la séptima edición delManual Bergey
(Snieszko,1957)sereclasificócomoAeromonashydrophila.
Hammer(1917) esel primer autorque describe la presencia deAeromonasen
alimentos,al aislarunabacteriaen leche alteradaque denominéBacilas ichthyosm¿us.
Entrelos primeros investigadores queobservaronAeromonasspp.asociadasa infecciones
humanaso animales secita a Aiken y col. (1936)y Miles y Halnan(1937).En 1954.Hill
y col, aíslanuna cepa, a partir de unprocesosepticémicodesarrollado enun hombrey.
posteriormente,Caselitz(1955)le asignael nombrede V¡briojamaicensis.
La especie denominada actualmenteA. caviae fue descritapor primeravez por
Scherago(1936)comoPseudomonascaviae, agentecausantede la septicemia epizoóticade
los cobayas.Más tarde, Liu (1962)y Schubert(1964)establecieronqueestebacilopodía
serunacepaanaerógenadel grupoAeronzonasy la llamaronA. puncratasubsp.caviae.
En 1936, Kluyver y van Niel proponenel nombrede Aeromonas;Stanier(1943)
apoyaestainiciativay, finalmente,la séptimaedicióndel ManualBergey( Snieszko,1957)
incluye estegénerodentrode la familia Pseudonwnadaceae.La octava(Schubert,1974)y
novena(Popoff, 1984)ediciónreclasificanestegénerodentrodela familia Vibrionaceae.
En 1969 Schubert,describedos especies:A. hydrophila (con tressubespecies:A.
hydrop/i¡la, A. anaerogenesy A. proreolytica)y A. punetata (con dossubespecies:A.
puncrwayA. caviae).
Los datos expuestos,antenormente,sehan recogidode los trabajospublicadospor
von Graevenitz(1987)y Reina(1992).
En 1976,Popoffy Veron estudiaronel comportamientode 68 Aeronwnasmesófilas
(anteriormentedenominadasA. hydrophilayA.puncunta)frentea203 pruebas bioquímicas
y fisiológicas,diferenciandoA. hydrophita (con las biovariedadesX1 y X2) y A. sobria.Posteriormente,Popoffy col. (1981) pudieroncomprobar,estudiando lahomologíadel
ADN. que lastresespeciespreviamentedescritas(A. bvdrophila.A. sobria yA. caviar,
6

I,VFRODUCCION
estaúltima sería la biovariedadX2 de A. hydrophila)erangenéticamentedistintasy que,al
menosdentrode cadauna de ellas, seintegrabandos o tres gruposde hibridación,lo que
hacía suponer laexistenciade másespeciesde lasqueen esemomentopodíandiferenciarse
bioquimicamente.
1.1.2. - Clasificación
El géneroAeromonasestáconstituidopor formasbacilaresrectaso curvas,de
extremosredondeados,de 0,3 a 1,0 gm de diámetroy de 1,0 a 3,5 jxm de longitud.
Aparecencomocélulasaisladas,en parejaso formandocadenascortas.Gram-negativas,
generalmentemóviles (hayespeciesinmóviles) con un único flagelo polar. Anaerobias
facultativas,conmetabolismorespiratorioy fermentativode la glucosa.Catalasay oxidasa
positivasy resistentesal agentevibriostítico 2,4,diamino-6,7,diisopropilpteridina(0/129).
El porcentajemolecularde G-¡-Cdel ADN es57-63(Bd, Tm) (Joshepy col., 1987).
Las principalescaracterísticasquepermitendiferenciar el grupo deAeromonasspp.
móviles de otros microorganismosfrecuentesen los mismosambientesradicanen su
carácteroxidasapositivo, quelas diferenciade la familiaEnterobacteriaceae;su capacidad
parafennentarloscarbohidratosquelas distinguedePseudomonasspp.;su crecimientoen
caldo nutritivo en ausenciade CíNa y resistenciaal agentevibriostético 0/129,en
contraposiciónal comportamientode Vibrios spp.halófilos; la producciónde DNasa, el
fracasoparametabolizarel inositol y. como seha dicho anteriormente,la resistenciaal
agentevibriostático0/129 que lasdistinguedePlesiomonasshigelloides.A diferenciade las
aeromonasmóviles y mesófilasAeromonas salmonicidaes inmóvil y no crece a370C
(Holmbergy Farmer,1984;Popoff, 1984;von Graevenitz,1985).
Actualmente,elgéneroAeromonasestáincluido junto con Vibrio, Photobacreriumy
Plesiomonasen la familia Vibrionaceae(Popoff, 1984; von Graevenitz,1985). Estos
autoresdescribendosgruposdeAeromonas,el primeroformadopor un grupo homogéneo
y reducidodecepaspsicrótrofas,inmóviles,incapacesdecrecer a370C,productorasde un
pigmentopardosolubleen agua,altamentepatógenasparapecesaunqueno para elhombre,
cuya únicaespecieesA. salmonicida.que incluye tres subespecies:salmonicida,
achromogenesy mausocida. El segundogrupo, amplioy heterogéneo, constituidopor
aeromonasmóviles mesófilas,a menudo denominadasgenéricamentecomoA. hydrophila,
.7

—
a-
a-
a-
a-.
a-
a-
u,
IArTRODUCCION
y dividido en tresespecies A. hydrophila.A. caviaey A. sobria.El interésen los últimos
añosse ha centrado en estesegundogrupo porsu implicaciónen enfermedadeshumanas.
Las diferencias bioquímicas entre estastresespeciesde aeromonasmóviles sebasan
en laincapacidaddeA. sobriade hidrolizarla esculina.creceren presenciadeCNK. utilizar
la L-arginina, L-histidinay L-arabinosay fermentar la salicina,mientrasque, al igual que
A. hydrophila. producegas de laglucosa,H2S de la cisteina,aunquemuestraun
comportamientovariableen el Voges Proskauer(producciónde acetoinaa partir de la
glucosa).A. caviae, porel contrario,no producegas,ni H2S, ni acetoina. siendo positiva
(al igual queA. hydrophila)en elrestode las pruebasen las quefracasaA. sobria.(Popoff,
1984).
Colwell y col. (1986) alestudiarlas fraccionesSS y 16Sdel ARN, hanobservado
que la evoluciónfilogenéticade las cepasincluidasen elgéneroAeromonasdifieren lo
suficientede la de los microorganismos pertenecientesa lasfamiliasEnrerobacteriaceaey
Vibrionaceae,como paraexcluiríasde estaúltima, y proponenla creaciónde la familia
Aeromonadaceae.Ruimy y col. (1994) coinciden en lanecesidadde crearestafamilia, al
analizarla secuenciade la subunidadpequeñadel ARNr.
Los estudiosrealizadospor Fanning y col. (1985), confirmaron los resultados
obtenidos previamenteporPopoffy col.(1981), demostrando la existencia decepasqueno
podían incluirseen ningunade las especiesconocidashasta elmomento,asícomodistintos
gruposde hibridación,en basea la cinéticadeasociaciónADN-ADN, dentrode cadauna de
lasespeciesbioquimicamentedefmidas.
Alíen y col. ya describieronen 1983 una nuevaespeciepertenecienteal género
Aeromonas,denominadaA. mediay caracterizadapor serinmóvil y producirun pigmento
pardo.
En 1987, Hickman-Brennery col., estudiangenéticamente11 cepasque,por su
semejanzacon y. cholerae yserornitina descarboxilasanegativas,hablansido previamente
designadascomo“Grupo ent&ico 77”. En estetrabajosecomprobóque podríanconstituir
unanuevaespecie(A. veronhi).cuyas principales características bioquímicasradicabanen
carecerde omitinadescarboxilasa yposeer argininadihidrolasa.En este mismo estudio se
comparóla cepatipo de A. veronii con otrasdel géneroAeromonas,diferenciándoseun
grupocon un elevadogradode hibridación genética quesc caracterizabapor ser ornitina

IATRODUCCION
descarboxilasanegativo y arginina dihidrolasa y lisina descarboxilasapositivo; la
confirmaci6nde estosdatos conllevaríala existenciade un biogrupodentro de esta especie
(Fanning y col.. 1985). Hickman-Brennery col. (1988), comprobaronque 8 cepas
similares aVibrio darnsela,aunquecrecíanen ausenciade CINa y hablansidoconsideradas
como ‘Grupo entérico 501”, en realidad, pertenecíanal géneroAeromonasy se
correspondíancon la especiedenominadaA. schuberrii. Un segundogrupo,actualmente
designadocomo“Grupo de Aerotnonas501”, essimilar aA. schubertiiaunquemuestra
ciertas diferenciasfenotípicas(indol positivoy lisina descarboxilasanegativo).
Schuberty Hegazy(1988)caracterizanaA. eucrenophilaque,bioquimicanhente,se
correspondecon cepasaemgénicasdeA. caviaey querepresentarlala especieanteriormente
denominadaA. punctatasubsp.punctata.
Carnahany col. (1991c)detectanun nuevogrupode hibridación,denominadoA.
jandaei,similaraA. sobriaperosacarosanegativo.Posteriormente,sedescribióotro grupo
dehibridación,caracterizado porsu sensibilidada la ampicilina,que se denominóA. troja
(Carnahany col., 1991b).
En 1992, Martínez-Murciay col., basándoseen la secuenciagenéticade la fracción
165 del ARN, describenunanuevaespecie,A. allosaccharophila,cuyacepatipo es CECT
4199. Estudiosde letalidaden pecesparecen indicarque esta nuevaespeciepuede que,
únicamente,sea patógenaoportunista,al contrarioqueA, jandaeique se ha manifestado
comopatógenoprimario(Estevey col., 1995).
Schuberty col. (1990a,b) propusieron dos especiesde Aeromonasmesófdas:A.
enteropelogenesy A. ichihiosmia,aunquela secuenciacióndela fracción165 del ARNr ha
demostradoque estos microorganismosson idénticos a A. troja y A. veronii,
respectivamente(Collins y col., 1993).
1.1.2.1. - Gruposde Hibridación: identificación bioquímica
Los gmpos de hibridación(GH) sehanconstituidoen función de la reasociación del
ADN de las cepas.Los criteriosempleadosparaestablecerlos 13 gruposde hibridación
(Tabla 1.2), descritoshastaahora,sebasanen lastasasrelativas deunióndel ADN. La
especie genéticaincluidaen cadagrupode hibridación,lo estáen funciónde la cepatipo que
se integre en él(Janda,1991).De estemodosehanconstituidoonce de los trecegruposde
hibridación, ya que el2 (GH2) y el II (GHl 1) no han podido serasimiladosen las
especies descritas(Janda,1991).
La especie fenotípicaA.hydrophilaenglobatresgruposde hibridación(GRI. GH2 y
9

1! uy a- u’ u’ a- a- u
——
—o
Ni
—..,
‘oo
—3
ON~.
A.~
.~
~-j
—
ca
~b
.—
..
&E’
~~
9.-t
O•-
.—
~.
~~
-~
-
4,
4,
‘t‘~
‘<t
‘1~
-
~:i.
~
~~
E’~
~%~
.~
.~
-e
-..
—
caca
caca
caca
ZZ
caca
caca
ca—
.—
—.
——
.—
.O
rs.
__
+$
±+
++
‘7%~
++
++
~-
~.+
+1
:1:+
‘-~
-~\.
~‘—
1‘—
¿1
o II rsrs
~+ +
5.1’
5E.
o‘e
9
o o o ‘e rs. -f a CL o o, ‘SI o E. 46
~ 5. o o o o, o o o ce E CL O o, o 5 o,
CjE
”
‘3 9> a—
eL
o ctE
Ca
roo
ea o a
—e a 9
E.o
4— *4~
• o
‘oc~ E.
zt.<
1<
~.
a— tfi
oC
a a’ 2 u! e a o 2 u, 1• a o a a e os o E cd -I a o E. o
lo

!ATRODUCC!ON
GH3); lacepatipo de A. hydrophi¡aATCC 7966estáincluidaen elGHl, porlo que este
grupo constituye la especie(genética)A. hydrnphila. Lacepacipo deA. saimonicidaATCC
33658se integraen el GH3, por lo que las diferenciasfenotípicas(inmóvil y pigmento
pardo)que,originalmente,distinguíanlas cepasde A. salmonicidade las Aeromonas
mesófilasahora,únicamente,definen unasubespeciedentro del GH3 (Janda,1991).
Existen controversiasen relacióncon el GH4, ya queA. caviae y A.punetatatienen la
mismacepatipo (ATCC 15468)(Farmery col., 1986).
El GH5 contienela cepatipo A. mediaATCC 33907.Hickman-Brennery col. (1988)
observandos gruposde hibridación,relacionadosgenéticamente,quedenominanGH5A yGH5B;A. mediasubsp.new namela incluyenen el GH5A y aA. mediasubsp.mediaenel GH5B.Debidoalas importantesdiferenciasfenotípicasdetectadasentrelascepasdeA.
media subsp.media,aisladasen aguasy en muestrasclínicas,es preciso apuntarlaposibilidaddequeexistandos biogrupos: el1, constituidoporcepasinmóvilese incapacesde usarla sacarosa comoúnica fuentede carbonoy el biogrupoII, que incluidacepasmóvilesque crecencon dichoazúcar(Allen y col., 1983).
Los gruposde hibridaciónGH6 (A. eucrenophila)y GH7 (A. sobria) comprendencepasquehastael momentono hansidoaisladasde muestrasclínicas.
La mayoríadelas cepasdeA. sobria,aisladasenla clínica,no seintegranenel GH7
(quecontienelacepatipo deA. sobriaCW 7433),sinoen el GH8 (queincluye la cepatipode A. veroniiATCC 9071), porlo que los aislamientosclínicos de A. sobriaenrealidad
corresponderíanaA. veronilbiotiposobria (Reina, 1992).Los gruposGH8 y GH1O seestablecieronindependientementey son distintos
bioquimicamente,para Hickman-Brennery col. (1987)el GH8 corresponderíaal fenotipode A. sobria y el GH1O al de A. veronii. Sin embargo, recientemente,Kuijper y col.(198%)han comprobado queestosdosgrupossonidénticosgenéticamentey, ya queelGH1O sedescribióantes,el GHS debeconsiderarseun biotipo deA. veronii. Altwegg y
col. (1990) hancomprobadoquela cepatipo A. sobria CII’ 7433 (GH7) muestraescasacorrelacióncon lascaracterísticasbioquímicasobservadasen los aislamientosclínicos,mientrasquela cepatipo A. sobria ATCC 9071 (incluida enel 0H8) seasociaen mayormedida.
Kuijper y col. (1989b), trabajandocon 142 cepas, compararonlos gruposestablecidospor hibridacióndel ADN conlos obtenidossegúnla clasificación fenotípicade
Popoff y col. <1981). llegando a observarseríasdiferencias;así, de las26 cepas
identificadasfenotípicamentecomoA. hydrophilasólo 15 pertenecíanal genotipoCHI y el
ti

u’
u
St
a-
St
St
a-
a-
St
u’
u’
IJVTROOUCC¡ON
restose repartíaentreel GH2, GI-13, GH5A y GH8.Janda(1991)opinaque lascepasdeAeromonasdebenconsiderarsegenéricamente
comoAeronwnasspp. o integrarseen el complejodeA. hydrophila.Es decir, sólo sedebeidentificarel género, yaque elpapelde las especiesen los procesosgastroentéricosno está
establecidode formaconcluyente, laidentificacióngenéticade lasespeciesresultacaray labioquímicaes excesivamentelaboriosay puededar lugar a errores,sin olvidar quenoexisten informes de comunidadesu hospitalessobre brotes de gastroenteritispor
Aeromonasquejustifiquenla identificaciónespecífica.Porel contrario,otros autoresconsideran importante diferenciarlas especies, desde
que seconoce quela mayoríade las cepas fecales humanas(> 85%) pertenecena Gui,GH4y GH8 (A. hydrophila,A. caviaeyA. veronil biotipo sobria)y el 15%restanteaA.
media (GH5) (Carnahany col., 1991a, 1992; Abbott y col., 1992).Al parecer,A. veronil
biotipo sobria (GH8) es elgrupo de mayor virulencia (Daily y col., 1981; Janda y col.,1984b;Kirov y col., 1986;Carelloy col.. 1988;Havelaary coL, 1992; Kirov y Hayward,
1993),A. caviae(GH4) esmás común endiarreasinfantiles (Altwegg, 1985; Namdariy
Botione, 1990a,b,1991>,A. veronhibiotipo veronii (OHiO), A. janáaei (GH9) y A. troja
(GHl4) se aíslande hecesdiarreicascon pocafrecuencia (Camahany col., 1991b)y A.
schubertiiseha asociadoa heridasinfectadasa travésdel agua,perono a gastroenteritis(Hickman-Brennery col., 1988;Caniahany col.. 1989b).En lo queserefiereal complejodeA. hydrophila, lascepaspertenecientesal GHl, frecuentemente se aíslande muestrasclínicasy producenfactoresde virulencia, mientras queel GH2 no elaboraestosfactoresy
el GH3 preferentementeseaíslade muestrasambientales(Kirov y col., 1994).Carnahany col. (1991a)proponenun sencilloesquemade sietepruebasbioquímicas,
denominado“Aerokey II” (Figura 1.1), paradiferenciarlas especiesfenotípicas.Por otraparte,Abbou y col. (1992),recomiendanentre19 y 24 pruebas bioqufluicaspara identificar
los gruposde hibridación,siendonecesarias9 pruebas(Usina descarboxilasa,ornitinadescarboxilasa,argininadihidrolasa.hidrólisis de la esculina,gas de glucosa,VogesProskauer,fermentaciónde la arabinosa,del ¡nanitol y de la sacarosa)parasepararel
complejoA. hydrophila(GHL, GH2 y GH3),A. caviae (GH4 y GHS), el dúoGHI2 yOH 13 y, de forma individualizada,el 0H6. G87,OHE, GH9, GH1O y 0H11. La
diferenciaciónde los tres gruposde hibridaciónqueconstituyenA. hydrophilase puede
realizaren función deotras sietepruebas(producciónde ácidode 0-ramnosa,D-sorbitol,
salicinay lactosa,oxidacióndel gluconato,elastasay producciónde ácido fenilpirúvico).Los genotipos quetienenel fenotipodeA. caviae (GH4, GH5y GH6) se diferencianen la
12

INTRODUCCION
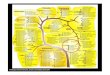
Figura Li. - Aerokey II (Carnahan etal., 1991)
A. trota
Producciónde indol
A. schubertii
VogesProskauer
A. jandaei
A. veronil biotipo sobria A. hydrophila
Agromonasspp.
Hidrólisis de la esculina
+
Gas deglucosa
+
A. caviae
Fermentacióndela arabinosa
A. hydrophila
Resistenciaa lacefalotina(30~.tg)
A. veronil biotipo veronil
++
Fermentacióndela sacarosa
+ +
13

a-
a-
!MTRODUCCION
utilizacióndel citrato,producciónde ácidodel glicerol y de la 0-manosa,H2S de la cisteina,
hemolisis,pirazinamidasay producciónde ácidofenilpirúvico. A. eucrenophila(GH6)podríadiferenciarsepor la producciónde gas,pero éstaesdébil en comparacióncon la
capacidad aerogénicade A. hydrophiloo A. veronii, porlo que incluirla en esteesquema
evitaríaun posibleerrorde identificación.Finalmente,la lipolisis sobre aceitede maíz, la
producciónde ácido a partirde la celobiosay de H2S de la cisteina,así como lasensibilidad
a la cefalotinay a la ampicilina sonlas cincopruebasadicionales quediferencianA. troja y
A. schubertii.
Recientemente,Hiinninen (1994) observaque en el complejoA. hydrophila,el 0111
puedediferenciarsedel GH2y GH3 en funciónde la temperatura máximade crecimiento,
mientrasque, en los patronesobtenidosen el análisiselectroforéticode las proteínas,el
GH3 posee una banda de pesomolecularvariableentre24, 25 o 26 kDa, queno apareceen
las cepas delOHí y GH2. Ash y col. (1993) hanpropuestoel usode la reacciónen cadena
de la polimerasay sondas génicasparala identificacióndeA. schuberñiy A. jandaei.
1.2. - Aislamiento e identificación de Aeromonasspp. móviles
En unprincipio seemplearonmedios diseñadosparael aislamientodeenterobacterias
(Pitarangsiy col., 1982; Champsaury col., 1982; Le Chevalliery col.. 1982) y vibrios
marinos(Graceyy col., 1982). El grupode Aeromonasspp. incluyemicroorganismos
lactosapositivosy lactosa negativos(Popoffy Veron, 1976),porlo que noesadecuado el
usode medios entéricos, como el agarMacConkeyo el agar azulde metileno eosina(von
Graevenitz,1985; von Gravenitzy Bucher, 1983). Además,Desmondy Janda (1986)
compruebanque los mediosutilizadosparael aislamientode patógenos entéricos pueden
inhibir el crecimientodeAeromonos.
Von Graevenitzy Bucher(1983) evaluan mediosadaptadoso desarrolladosparael
aislamiento específicode Aeromonasa partirde heceshumanas,recomendando el aguade
peptonaalcalinacomomedio deenriquecimientofrenteal caldotriptona sojacon ampicilina.
Porotro lado, sehandescritocomo medios sólidosselectivosel agarinositol verdebrillante
salesbiliares,el agardextrina fucsina sulfito (descrito en1967 por Schubertparael
aislamiento específicode Aeromonasen aguas),el agar xilosa desoxicolato sódico citrato
(Shready col., 1981)o el agar prilxilosaampicilina (Rogol y col.. 1979).Estos autores
observaronque elagarpeptonaextracto de carnede vaca glicógeno (McKoyy Pilcher,
1974)no diferenciaestosmicroorganismosde la flora normalde las hecesy el medio
14

IMTRODUcCION
Riniler Shotts (Shottsy Rimíer, 1973) no es lo suficientementeinhibidor para otros
microorganismosgram-negativos.El agar DNasaazul de toluidina ampicilina (von
Oraevenitzy Zinterbofer, 197<)), el medio mA (Rippey y Cabelli, 1979) y el agar sal
almidón xilosa lisinadesoxicolatosódico (Roland, 1977). tampocodieron buenos
resultados.
Otro medio con elque varios autores obtuvieronaislamientosfue el agar sangrede
corderocon 10 ó 15gg/l de ampicilina (Aggery col., 1985;Oraceyy col., 1982), siendo
ademásadecuado para realizar la pruebadela oxidasa,pues lafermentaciónde azúcaresen
otrosmediosalterael pHy dificulta surealización.
Palumboy col. (1985a) desarrollanun medioespecífico(agaralmidónampicilina),para lacuantificacióndirectadeAeromonasspp.en alimentos,quecontieneampicilina(10¡¿gil) comoagenteselectivoy almidóncomodiferencial,aunquetieneel inconvenientedeno
separarAeromonasde Vibrio spp..Altorfer y col. (1985) proponenel agar cefsulodínirgasán novobiocina(agarCIN),
descritopor Schiemannen 1979 parael aislamiento deYersinia enterocolitica,aunquerecomiendandisminuirla concentracióndecefsulodin(CIN II) parala detecciónsimultánea
de Yersínia spp. y Aeromonasspp. Fricker y Tompsett (1989) destacanel buencomportamientodel mediodeAeromonas(Difco) en el análisisde alimentos.
Para el análisis de aguasse han desarrolladodos medios, previa filtración yconcentracióna travésde membranas:el agarde Rimíer Shoíts(Shottsy Rimíer, 1973)quecontienecitrato, novobiocinay desoxicolato sádicocomo agentesselectivosy usina,ornitina y maltosacomo diferencialesy el medio mA (Rippey y Cabelli, 1979) conampicilina, etanol y desoxicolatosádico como agentesselectivosy trealosacomo
diferencial. Posteriormente,Havelaary col. (1987, 1988)proponenel agardextrina
ampicilina(ADA), aunqueno diferenciaAeromonasspp.de Vibrios marinos. Huguety
Ribas(1991) modificanel medio glutamatoalmidónampicilina (OSP),propuesto porKielwein en 1969parael aislamientosimultáneodeAeromonasy Pseudomonas,añadiendo
10 ¡xg/l de C-glucosay 20 de ampicilina(SGAP-lOC), conlo quesemejora larecuperación
deAeromonasa partir de muestrasacuáticasy seinhibeel crecimientode Pseudonwnas.
Holmes y Sartory (1993)empleanel medio de Aeronwnasde Ryan (comercializadopor
Oxoid) y resaltansubuencomportamiento.Warburtony col. (1994)diseftanun protocolo
paraaislarAeromonasa partir del agua,que incluye filtración a través de membranas
hidrofóbicas,revitalizaciónen agar tripticasasoja y siembraen dos medios selectivos:
mediodeAerornonasy agartrealosaAeromonas.
Aunquemuchosautoreshanempleadola siembra directaen placa.la recuperaciónde
15

e
St
a-
a
St
a.
a-
e
a-
a-
St
e
St
INTRODUCC(ON
Acromonas aumenta, considerablemente,cuandoserealizaun enriquecimientoprevio(von
Graevenitzy Bucher,1983).Okrend(1987)comparael efectode medios selectivosen larecuperacióndcAeromonas,utilizandocomomediosde enriquecimientoaguade peptona
alcalina(pH 8.6),caldo triptonasoja, caldotriptona sojaconampicilina.caldo triptona sojacon CINay caldotriptona.observandolosmejoresresultadosconel caldotriptona sojaconampicilina. Sin embargo,muchosinvestigadoresprefierenemplearel agua depeptonaalcalinapuesno impide la detecciónde cepassensiblesa la ampicilina(Varnam y Evans,
1991).
Un medio que podríaresultarútil en la identificación,seríael AH, desarrolladoporKaper y col. (1979),ya quepermite realizar, simultáneamente,cinco pruebas bioquímicas.
El géneroAeromonaspuedeserdiferenciadoa través del sistema API 20E, aunque se
requierenotras pruebascomplementarias,comola sensibilidadal agentevibriostático0/129
(Varnamy Evans,1991).El apartadoIV.2.1 recogemásdatos sobreestepunto.
Ademásde los sistemasclásicosde aislamiento eidentificación,cienosautores
proponenotrosmás actuales como el“ON microplate”, basadoen la oxidaciónde sustratos
carbonados,capaz dediferenciarA. hydrophila, A. sobriay A. caviae(Carnahany col.,
1989a).Lasensibilidadalacefalotinay la actividadpirazinamidasa se hanpropuestocomo
pruebasespecíficasparadiferenciarA. sobriadel restode lasespecies.(Janday Motyl,
1985; Carnaham y col., 1990).Sugita y col. (1994)señalanla posibilidad de identificar
especiesrealizando lahibridaciónenmicroplacas.
El serotipadoseha realizadoen cepaspatógenaspara los peces, pero hay pocos
estudiosefectuadossobrecepasde Aeronwnasspp. potencialmentepatógenaspara el
hombre. Fricker (1987)utilizando 16 antisuerosfrentea antígenoslipopolisacáridos.
desarrollaun esquemacapazde tipificar el 46%de las cepasaisladasen heces humanas,
Thomasy col. (1990)reconocen,provisionalmente,52 serogrupos.Janday col. (1994a>indican queel 60% de las cepasimplicadasen septicemias pertenecena los serogrupos0:11,0:16,0:18 yO:34.
Otros sistemasde tipificaciónsebasanen la comparaciónde los patronesproteicosobtenidosmediantemarcajeradioactivo (Stephesony col., 1987), tinción con plata
(Millership y Want. 1989, 1993), quimioluminiscencia(Carey y col., 1994) o
“inmunoblotting” (Mulla y Millership, 1993)de los componentescelularesseparadospor
SDS-PAGE. Frecuentemente,tambiénse realiza la tipificación a nivel molecular,
empleando elanálisisdel ADNcon endonucleasasderestriccióny la electroforesisen gel de
poliacrilamidade las cubiertas celulares(Kuijper y col.. 1989c).Huys y col. (1994,1995).
16

INTRODUCCION
al igual que Kampfery col. (1994)y Altwegg y Kersters (1994),proponeny empleanel
análisis, por cromatografíagaseosa,de los ácidos grasoscelularespara identificar las
especiesfenotípicasy los gruposde hibridaciónincluidosen estegénero.
1.3. - Epidemioloela
1.3.1. - Heces humanas
Las publicacionessobredatosepidemiológicosde Aeromonasspp. sonescasas,
debido,fundamentalmente,a quelas tasasde aislamientoen los mediosentéricosestándar
son bajas y no existe una clara evidencia de que estos microorganismos sean
enteropatógenos,comoparajustificar suinvestigaciónespecíficaen alimentossospechosos
o en heces.Sin embargo,la tasa deaislamientosaumentaconsiderablemente cuando,
previamente,seefectúaenriquecimientoy seutilizan mediosespecíficos(Burke y col.,
1983a).
EnThailandia,Pitarangsiy col. (1982)observanun mayornivel de aislamientosentre
la población indígena(9-30%)sin síntomas de diarreao con unarápidarespuestaal
tratamiento.Por el contrario,Echeverriay col. (1984)detectanque el aislamientode
Aeromonases,significativamentemásfrecuente,en personas condiarrea(31-48%) que en
individuossin síntomasaparentes (9-15%),cuandosetratade sujetosreciénllegados alas
zonasafectadas.Graceyy col. (1984)en Asia, Black (1986) en áreasde Latinoaméricay
Africa y Mattila y col. (1993) enFinlandia incluyenestosmicrorganismos dentrode los
agentescausantes de ladenominada‘diarreade los viajeros”.
En un estudio realizadoen Perth(Australia),Burke y col. (1983a)comprobaron que
la deteccióndeAeromonasfuesignificativamente másaltaen niñoscon diarrea (10,8%)que
encontrolesasintomáticos(0,7%) observando, además,una marcada estacionalidaden los
mesescálidosde veranoy otoño,comotambién señalanGraceyy col. (1982),Janday col.
(1983a),Aggery col. (1985)y Georgey col. (1985).Contrariamente,en Italia lastasasde
aislamientofueronescasas(1%) sin diferencias marcadasentrelos casossintomáticosy loscontroles(Figura y col.,1986).En Bangladesh(Kay y col., 1986,citado en Sacky col.,
1987)el uso de enriquecimientoprevio y de mediosselectivosha puestode manifiestouna
tasaelevadade aislamientosen pacientescon diarreaaguda(33%).
En Perú(Gilman, R.H., citado en Sacky col., 1987) secontrolaron 400familias
(2000personas)de bajonivel económicoa lo largode dosaños,analizandola presenciade
Aeromonasspp.en hecesdiarreicasy normales,previoenriquecimientoen aguade peptona
17

r
a-
St
a!
a-
a-
a
e
e,
IWTRODUCCION
alcalinay aislamientosobreagarsangreampicilina.obteniéndoseun númerosuperiorde
aislamientosen heces diarreicas.Las mayoresdiferenciassedetectaronen el grupodeedad
inferior a seis meses (16% en heces diarreicasy 9% en controles). Unaño después, al
analizarel grupo deedadinferior a3 años,seobservarontambiéndiferencias,peromenos
marcadas(9% en hecesdiarreicasy 4,3%en hecesnormales).A lo largode esteperiodo,
Aeromonasspp. se aisló, deformaconstante,en el aguade bebidade estacomunidad.En
este sentido.Graceyy col. (1982)observanque el 13% de los procesosgastroentéricos
asociadosaAeromonasafectana niños menoresde 6 meses,el 57,4%entre6 mesesy 2
años,el 22%a niñosde 2 a 5 añosy sólo el ‘7,6% a mayoresde 5 años.Estosestudios
sugierenque, al menosalgunascepas, sonenteropatógenasy que lasexposiciones
primariasdaríanlugaral desarrollode laenfermedad(niñospequeñosy visitantesazonas
endémicas).
EnEgipto se handetectadoAeromonasspp.en hecesdiarreicas deniñoscon mayor
frecuenciaque en no diarreicas,aunquela actividadenterotoxigénicano se asoció
preferentementea unasu otras cepas(Ghanemy col., 1993).
En Queensland (Australia)se haobservadola presenciadeAeromonasen las heces
diarreicasde pacientes inmunocompetentes,distribuidosen dos gruposdeedad:menoresde5 añosy mayoresde 25 (Ashdowny Koehler, 1993).
San Joaquiny Pickett (1988) y Kuijper y col. (1989a) observanque la diarreaasociadaaAeromonassedetecta,fundamentalmente,en niños de edadescomprendidasentre6 mesesy 5 añosy que las cepas citotoxigénicas parecensermás frecuentesen
personasmayoresde 50 años. En conformidadcon estosdatos,Aggery col. (1985) y
Millership y col. (1983) tambiénobservaronesta mayor incidenciaen niños y personasmayores.Namdari y Bottone (1990a) apuntan quecienosalimentos infantiles pueden
incrementarel riesgo deinfecciónal favorecerun aumentoconsiderable del pHintestinal
(superiora 7,5). De la Morenay col. (1993) aíslanespeciesmóviles deAeronionas,
principalmentedeA. caviae,en hecesdiarreicasdeniños,
Otros autoreshan aisladotasasmoderadasde estos microorganismos encasosdecolitis crónicas,aunqueno en las biopsiasde los tejidos lesionados (Robertsy col., 1987;
Domany coL, 1989;Farrayey col., 1989;Wiloughby y col., 1989).
Durantemucho tiempo,estegrupode microorganismosse haconsideradocomo
oportunistaen individuos inmunodeficientes(Stelma, 1989). En los últimos años,
numerososautoreshan manifestadoque podríanserademáspatógenosprimarios,puesto
que tambiénse han aisladoen procesos infecciososde personasnormales(Rhamony
Willoughby, 1980; Janday col.. l983a),destacandoel aislamientodeA. hydroph¡la de un
18

INTRODUCCION
proceso septicémicomortal desarrolladoen un jovenaparentementesanoy de un hombre
inmunocompetentede 37 añosconmeningitisy bacteriemia(Qadri y col., 1976).
Aeromonases un génerode presenciatransitoriaen el tracto gastrointestinaldel
hombre.Las tasasde aislamiento a partirde heceshumanasson másaltasen regiones
tropicales,como Asia,Sudaméricay Australiaqueen EstadosUnidosy Europa. Parece ser
queno son necesariasprecaucionesespecialescon respecto alos portadoresde Acromonas,
quepuedanmanipularalimentos(Varnamy Evans,1991).
1.3.2. - Alimentos: Factores que influyen en la supervivencia y en el
crecimiento
Este género seconsideraparte de la flora acrobia total presente, deformanatural, en
alimentos(Palumboy col., 1985a).Palumboy col. (1985a)observanla presenciade A.
hydrophilaen distintos pescadosy mariscos,carnes rojasy de avesy leche cruda.Myersy
col. (1982) y Simardy col. (1984) la aíslan decarnesenvasadasal vacío. Numerosos
autoreshan comunicadoel aislamientode Aeromonasspp. en estosproductosde origen
animal, inclusoenvasados en atmósferasmodificadas(Eddy y Kitchell, 1959;Gardner,
1965; Kielwein y col., 1969; Kleeberger,1975; Toule y Murphy, 1978; Enfordsy col.,
1979; Grau, 1981; Blickstandy Molin, 1983; Hudsony DeLacy,1991;Hanninen,1993;Hudsony Mott, 1993). Tambiénse han detectado con frecuenciaen vegetalescrudos
(Callister y Agger, 1987; Hudsony DeLacy, 1991). Gobaty Jemmi (1993)señalansupresenciaen alimentoscrudos,contasasde 106 ufc/g, y listos para el consumo, aniveles
de 10~ ufc/g. Otros productosinvestigadoscomo helados,huevos, ancasde rana ycaracolestambiénhanresultadopositivos(Huntery Burge, 1987;Varnamy Evans,1991).
La incidenciade las distintasespeciesdeAeromonasen alimentos varía,en funciónde
la zonageográficaconsiderada. Las cepas detectadasen aguasy alimentosaustralianos,que
mostraronpropiedadesasociadasa virulencia, se incluían,principalmente, dentrode los
gruposA. veronhlbiotipo sobriay A. hydrophlla (Greenaway,1988; Majeed y col.,
1989a,b;Kirov y col., 1990; Ibrahim y Mac Rae, 1991; Kirov y col., 1993c).A. veronii
biotipo sobria es la especie aisladacon másfrecuenciaen casosde gastroenteritisen
Australia (Kirov y col., 1986). También se hadetectadoen el 24% del totalde los
aislamientosobtenidosen alimentos japoneses (Nishikaway Kishi, 1988) y en el 42%de
los alimentosingleses (Frickery Tompsett,1989).Sin embargo,en otras partes del mundo
como en USA (Palumboy col., 1989),en Dinamarca (Knochely Jeppcscn.1990) y en
19

4
St
a-
a!
e
u.
e
a-
a-
a-
St
e
a!
e
e,
IMTRODUCCION
NuevaZelanda(Hudsony De Lacy, 1991)A. veronil biotipo sobriaprácticamenteno se
encontróen alimentos,predominandoA. hydrophilayA. caviae (Janday col., 1984b;Altwegg, 1985; Altwegg y Geiss. 1989; Moyer y col., 1991). Recientemente,se ha
comprobadoque las tres especiescomprendenmicroorganismospotencialmente
enterotoxigénicos,independientementede la fuente de aislamiento,ya que cepasque
inicialmenteno provocabanacúmulode fluidos, comenzaron aproducir enterotoxinas
despuésde uno atres pasesporasaintestinal ligadade conejo(Singh y Sanyal,1992a,b).
Hussleiny col. (1991, 1992)observanque la secuenciacorrespondientea la citotoxina
hemolítica(aerolisina)esun gen estructuralampliamentedistribuido en las cepasdeAeromonasde estastres especies.
Actualmente,losconsumidoresdemandanalimentosmásnaturales,pocoprocesadosy sin aditivos; en consecuencia,la refrigeración seha erigidocomounade lasprincipalesmedidasparacontrolar el desarrollode microorganismospatógenosy alterantesen
alimentos. Aunque las aeromonasmuestranuna temperaturaóptima y máxima decrecimientomesófila, sepuedendefinir comomicroorganismos psicrótrofossi, deacuerdo
con Eddy(1960>,se asumequeel término psicrótrofocorrespondea los microorganismoscapacesdecrecera temperaturas derefrigeración,independientementede sutemperatura
óptimade desarrollo.Muchosautores handescritoel crecimientodeAeromonas,inclusodecepas exotoxigénicas,a bajas temperaturas(Palumboy col., 1985a;Callister y Agger,1987;Berrangy col., 1989;Kirov y col., 1990;Beuchat,1991). La temperatura mínimadecrecimientodependede la cepaen cuestióny Rouf y Rigney (1971), Palumboy col.
(1985b),Walkery Stringer,1987y Kirov y col. (1993a,b)encuentranquealgunasdeellaselaboranfactoresasociadosa virulenciaa temperaturasde crecimientoentre4,4 y 60C.Kirov y col. (1993a) observaronen una cepaaisladaen leche de cabrauna temperaturamínimadecrecimientode -5,30C,óptimade 19%?y máximade430C,mientrasque elresto
de lascepasaisladasde alimentos,aunquecrecieronabajastemperaturas,precisaronmás
de seis díasparaque lapoblaciónaumentasesignificativamente.
Numerososautoreshan comunicadola capacidadde cepasde Aeromonaspara
producirexotoxinasa bajas temperaturasen mediosdecultivo (Knochel, 1989a;Majeedycol., 1990; Krovaceky col., 1991; Eley y col., 1993; Kirov y col., 1993a)y alimentos
(Majeedy MacRae,1991; Kirov y Brodribb, 1993)aunque,en estosúltimos. Todd y col.
(1989) detectanactividadesenterotóxicas,citotóxicas y hemolíticas inferioresa las
registradasen mediosbacteriológicosa las mismastemperaturas.Contrariamente,Kirov y
Brodribb (¡993)señalanque.exceptoen preparadosalimenticiosinfantilesy leche Uf-IT, la
20

IMrRODUCCION
producciónde exotoxinases taneficazen alimentoscomoen mediosde cultivo.
La concentraciónde sal, el pH, la atmósferay la flora competitivainfluyen en el
crecimientoy supervivenciade lasAeromonasen alimentosa bajas temperaturas(Palumbo.1988; Palumboy Buchanan,1988;Knochel, 1990; Beuchat,1991; Palumboy col., 1991,1992).Así, la toleranciaa la sal y al pH se reduceconsiderablementea temperaturasderefrigeración,siendoincapacesde crecer con3-3,5% de CINao a pHs inferioresa 6
(Palumbo,1988; Palumboy Buchanan,1988). Sin embargo,cuandola temperaturadeincubaciónespróximaala óptimade crecimiento puedenmultiplicarseinclusoen presenciade 6% deCíNa(Knochel,1990)y a pH4,5 (Palumboy col., 1985b).
No hay evidenciaclara dequelasAeromonasseanlo suficientementecompetitivas,
frenteal resto dela flora presenteen los alimentos,paraalcanzartasasimportantesabajastemperaturas(Palumboy Buchanan,1988; Inghamy Potter, 1988; Kirov y col., 1990;Erickson y Jenkins,1992). La disminucióndel oxígenoen la atmósferaquerodeaa losalimentosparecefavorecerel crecimiento deAeromonas,posiblemente, porque retarda el
crecimientode la flora aerobia competitiva(Buchanany Palumbo,1985).Palumboy col.(1992)observanqueotros parámetroscomoel pH, el CíNa y los nitritos puedensernecesariosparacontrolarel crecimientode algunasespeciesde Aeromonasa bajastemperaturas.
Aeromonasspp. forma partede la flotaalterantede alimentos refrigerados,como
carnesrojas (Jay,1967; Len, 1987),pollo (Toule y Murphy, 1978)pescado (Gorczycaycol., 1985; Gramy col., 1987), mariscos(Smith y col., 1984) y huevosenterospasteurizados(Mackenziey Skerman,1982). Especialmenteen alimentoscocinados,mantenidosen refrigeración,puedendar lugar a unarápidaalteración(Toule y Murphy,1978). Sinembargo,en leche UHT esposiblellegar a alcanzartasasmuy elevadassin
modificacionesorganolépticas(Kirov y col., 1993c).Goldeny col. (1989)han demostrado lainhibición específicadel dióxidode carbono
sobreel crecimientodeA. hydrophila, aunqueGilí y Reichel (1989) e Ingham(1990)
observarondesarrollo bacteriano a100C en alimentos envasadosen atmósferasdeCO2. Sin
embargo,los ambientesricos en nitrógenofavorecenel crecimientode célulaslesionadaseindemnesa 5
0C; este hecho se atribuyeal funcionamientodeficientedel sistemaprotectorfrente al oxígeno(enzima superóxido-dismutasa)en Aercmonasa 50C, por lo que, el
aumentoen la concentraciónde nitrógeno, con la consiguientedisminución enla deoxígeno,tendríaun efectoprotector(Goldeny col., 1989).No obstante.Manoy Garcíade
Femando(1995)observanque el tiempo de duplicaciónde este microorganismo sobre
21

w.
a-
a-
a!
a-
pv
e
mt
a-
INTROOIIICCIOW
carnede cerdoy pavo,esmásprolongadocuandocreceen 100%de nitrógeno que cuando
lo haceen aire.En “mince” y “surimi”, elaboradoscon pocasal, A. hydrophila crecebiena50C, pero
se inhibe cuandoel ‘surimi” contiene2,5-3% de NaCí (Inghamy Potter, 1988).En carne
de cerdoenvasadaal vacío,un 3% deCINa essuficienteparaimpedir la multiplicaciónde
Aerornonas(Palumbo,1988).
También seha investigadola influenciadel sorbatoy del humo líquido (Gram,1991),
poniéndosede manifiestoque el sorbato(1000 ¡.iglml) combinadocon la sal impide el
crecimiento atemperaturas elevadas(25-37%?).Sin embargo,el humolíquido sólo eseficaz
cuandola contaminacióninicial esbaja(< 10~ ufc/ml).Aeronwnasseelimina de los alimentos porlos tratamientostérmicosdiseñadospara
SamoneHa (Condón y col., 1992; Palumboy col., 1991). No obstante,las curvasde
supervivenciamuestranunarespuestadifásica,debidoa la presencia deunasubpoblaciónmoderadamentetermorresistente(Palumboy col., 1987).Tambiénesrelativamentesensible
a las radiaciones ionizantes,con valoresDentomoa 1,4-2,2¡cOy (Palumboy col., 1986).Nishikaway col. (1993)han observado queestosmicroorganismossonmássensiblesa lostratamientostérmicosy a la acidezque otros patógenospresentesen alimentoscomo
Escherichiacdi 0157:H7,Staphilococcusaureusy Solnionella ¡yphimurium.
1.3.3. - Procesos clínicos
1.3.3.1. - Gastroenteritisasociadasa alimentos
Los pocoscasos publicadosde gastroenteritisasociadasal consumode alimentos,en
los quese han implicado a las aeromonasmóviles, se recogenen la Tabla 1.1. Los
alimentos sospechososfueron, principalmente,ostrasy otrosmariscos.En todoslos casos,
el cocinado,previo al consumode los alimentos,presumíblemente,fue inadecuadoo
incluso inexistente. Altweggy col. (1991)encuentranpruebasconcluyentesde que losalimentospuedenser vehículosde transmisión,en las gastroenteritisasociadasaAeromonas,al hallarunafuerte asociaciónentrecepasaisladasen gambascontaminadasy
en lashecesdiarreicasde un pacienteque consumiódicho producto.Sin embargo,el
númerode casos detectadoses bajo y los alimentosimplicadosno son aquellosque, enteoría,consumenlos gruposde poblaciónmásafectados(niños pequeños),por lo que
Kirov (1993) indicaque, o bien la transmisiónpor alimentosno es frecuente,o bien los
adultospodadoresasintomáticos,contaminadosa partir de alimentos,sedanel vehículo
99

IWTRODVCCION
transmisorde la gastroenteritisen niños del entorno familiar. También, es preciso
considerarlos casosde gastroenteritis,asociadosaAeromonasporalimentos,queno sondetectados,y queforman partede eseelevadoporcentaje(40%)de infeccionestransmitidas
poralimentosqueno aparecen enlasestadísticasoficiales.La investigaciónsistemáticade
Aeromonasen infeccionesasociadasal consumode alimentos aclararla,en parte,estos
problemas.Los estudios inmunológicos, realizadosrecientemente,han proporcionado
información importante sobrela actividad enteropatógenade Aeromonasen el hombre.
Kuijper y col. (1990) observan,en pacientes congastroenteritisagudasasociadasaAeromonas,una respuesta inmunológicaespecíficafrentea variasestructurascelularesy ala citolisinadela cepahomóloga,mediantela técnicaELISA; sin embargo,los resultadosobtenidospor radioinmunoensayono fueron concluyentesporque el suero no sólo
reaccionófrente a cepashomólogasy heterólogasde Aeromonassino tambiénfrente aEscherichiacoli y Vibrio cholerae.Jiangy col. (1990) detectaron,en 12 de 13 pacientes
infectadosconA. hydrophilay A. sobria,respuestasinmunológicasespecíficasfrente acepashomólogas.En estosindividuosse registróun aumentosignificativo dela secreción
de inmunoglobulinaA frentealos componentes lipopolisacáridosde pesomolecularentre20 y 80 kDa de lascepasinfectantes.Si secontrastasenestosresultadossepodríahablardeun avanceimportanteen la confirmacióndeAeromonascomopatógenogastrointestinal.
Se handescritodos tiposde síndromesgastrointestinalesasociadosaAeromonas
(Graceyy col., 1982).El primeroy máscomúnesun procesosimilar al cólera,quecursa
conhecesacuosasy fiebre moderada,pudiendoaparecervómitosenniños menoresde dosañosy, en ocasiones,doloresabdominales.El segundotipo, quese ha observadoen,aproximadamente,el 25% de los casos detectados (Georgey col., 1985; Graceyy col.,
1982), sedesarrollade forma similar ala disentería, con diarreamucohemorrágica,raravezcon vómitosy, en ocasiones,con doloresabdominales.Estosprocesos,generalmente,tienenbuenpronóstico(Holmbergy Farmer,1984),aunquealgunos casoshan puestoenpeligrola vida de los pacientes (Rhamony Willoughby, 1980;Champsaury col., 1982).
1.3.3.2.- Infecciones extraintestinales
La vía de entradaen infeccionessistémicases, probablemente,a través del tracto
gastrointestinalcomoconsecuenciade la ingestiónde alimentoscontaminados(Harris ycol., 1985; Beebe,1986). Se han aisladoAeromonasspp.en procesossepticémicos(vonGraevenitzy Mensch,1968; Slotnick, 1970; Davis y col., 1978; Abbott et al., 1994),
23

r
mt
a-
mt
a-
mt
a
e
e
IKFRODUCCION
meningitis(Qadri y col., 1976; Ellison y Mostow, 1984; Panasy col., 1993) y heridas
infectadas(von Graevenitzy Mensch,1968;Wolff y col., 1980;Ellison y Mostow, 1984;Newton y Kennedy. 1993; GoId y Salit, 1993). En ninguno de estoscasosse detectó,
únicamente,Aeromonas.La mayoríade las septicemiasy meningitisatribuidas aAeronwnassehandesarrolladoen personasinmunodeficientes,principalmente, en casosde
leucemiay cirrosis (Davis y col., 1978; Wolff y col., 1980; Ellison y Mostow, 1984;
Harris y col.. 1985) aunquetambiénse ha detectadosepticemiaen personasanémicasy
sometidasa hemodiálisis (Wolffy col., 1980). Otras manifestacionesmásinusualesdeestosprocesosextraintestinalesincluyeninfeccionesoculares,osteomielitis,infeccionesdeltracto urinario y respiratorio,peritonitis y endocarditis(Slotnick, 1970; Tumbulíy col.,1984; Muñozy col., 1994;Pietuy col., 1993;Poiriery col., 1993).La tasade mortalidad
en infeccionesextraintestinalesasociadasa Aeromonasen personasinmunodeficientessuperael 60%(Abeytay Wekell, 1988).
Las infeccionesde heridasporAero.’nonasestándirectamenterelacionadascon la
exposición deltraumatismoal aguao tierra (Davis y col., 1978; Wolff y col., 1980); enindividuos sanos sedesarrollancomo heridaslocalizadas, mientrasque en sujetos
inmunodeficientespueden derivaren septicemiasy darlugar,en ocasiones,al fallecimiento(Wolff y col., 1980;Krovaceky col., 1993).
Recientemente,se ha descritoun nuevosíndromeasociadoaAeromonas,comoconsecuencia delempleo desanguijuelasconfinesterapéuticos(Abrutyn, 1988).Esteviejoprocedimientose utiliza actualmenteparatratar congestionesvenosas,subsiguientesa.cirugíasplásticaso microvasculares,y se han descritoal menosocho casosde heridasinfectadasporAeromonasderivadasde dichapráctica(Dickson y col., 1984;Mercery col.,
1987; Snowery col., 1989).Todoslos casos remitieroncondesbridaciónde la heriday oterapia antimicrobiana.La fuente de esta infección pareceserla sanguijuelaHirudo
medicinalis,que mantienesimbióticamenteaeromonasen su intestinopara facilitarla
digestiónenzimáticade la sangre queingiere. En otrassanguijuelas,utilizadas confinesmedicinales,tambiénsehan detectadoestosmicroorganismoscomo partede la flora
intestinal (Willcen y Appleton, 1993;Bkkel y col., 1994).De las recientesespeciesincorporadasal génerosóloA. schubertiiseha aisladode
procesosextraintestinales(sangrey heridas) (Hickman-Brennery col.. 1988).
1.3.4. - Agua y portadores animales
Acromonas esun género ubicuoque se ha aisladode muchosambientes,pero al
24

1AtTRODUCCION
parecerel hábitatmás frecuente esel agua(Hazeny col., 1978; Grabowy DuPreez,1979;Biamon y Hazen,1983: Hazen,1983).En ríos y lagosse handetectadotasasdesde<
ufc/ml hasta>to~ ufc/ml; miembros deestegénerose han aislado enbotellasde aguamineral,pantanos(Abeytay Wekell, 1988) ypozos, tambiénen aguadorada(LeChevalier,
1980. 1982; Burke y col., 1984c) y en distintos puntosde sistemasmunicipalesdedistribución de agua(Clark y col., 1982; LeChevalliery col., 1982: Notermansy col.,1986).En Suiza,el aguade un pozo mostró640 ufc/mldeA. hydrophila,considerándoseal génerocomoun marcador decalidadde aguamáseficaz quelos colifornieso E. cdi
(Krovaceky col., 1989).En Holanda.van derKooij (1988)proponeunatasamáximade20 ufc/ml de Aeromonasen aguade bebida y de 200 ufc/ml. en los sistemasdedistribución.Actualmente,en Canadála presenciade Aeromonasen agua embotelladaestáregulada,siendoel límite admisible deO ufc/100ml de agua(Warburtony col., 1994).
El aguade bebidase suele contaminara partir de efluentesdepuradoso no,
presentandoalgunosdeellos, comolos de los mataderos,un alto contenidoen nutrientesque favorecensumultiplicación (García-Lópezy col., 1993).Wadstrómy Ljungh (1991)adviertendel riesgo quesuponenlos fallosen el procesadodel aguade bebiday VamamyEvans(1991)resaltanquela actualtendenciaaemplear aguaembotelladaen la alimentación
de niñosy personasinmunodeficientesentrañaun peligropotencial, yaqueconstituyenlosgruposde poblaciónconmayorriesgode padecerinfeccionesporAeromonas.
El númerodeAeromonasen el aguadependede la polución,estado tróficodel aguaytemperaturaambiente.Su tasa esmayor en aguascorrientesque en lasestancadas,
encontrándoseconmásfrecuenciaen aguascon ciertaconcentracióndesales, conpH 5,2 a9,8 y temperaturasde 40C a 450C. No pareceestarpresenteen lagoshipersalinosni enaguas termales(>450C) y tampocoen las muy contaminadas;ademáspresentavariacionesestacionalescon mayorincidencia enépocascálidas(con temperaturas>200C) (Hazenycol.. 1978). Kapery col. (1981> apuntanque la detecciónde Aeromonasen aguasdisminuye cuandose trata de aguascon baja concentraciónde oxígenodisuelto.
Aeromonas,probablemente,senutredelfitoplactony pareceserqueexiste correlaciónentreel númerodeAeromonasy la concentraciónde clorofila y fósforo (van derKooij, 1988).Por estarazón, la cargade Aeromonases un buen indicadordel estado tróficodel agua
(Rippeyy Cabelli, 1980).Aeronwnasescapazde creceren aguas residualeshasta niveles delO~-lO~ ufe/mí.
Estemicroorganismoes frecuenteen sistemasdomésticos,como tuberíasde drenajede
agua sucia, sumideros, pilasde fregar y tuberíasde salida de electrodomésticos
(lavaplatos).Sin embargo,la polución que puedanrepresentarestasfuentesno permite
25

w
a-
e,
u
a-
pv
a!
e
u
a
u
u
u
u
IAPTRODUCCION
explicarel alto númerodeAeromonaspresentesen el ambiente (Vamaniy Evaus.1991).
Estemicroorganismotambiénse ha aislado en aguasalinay estuarios(Williams y
LaRoche, 1985; Abeyta y col., 1986), aunque en estosambientessu presenciaes
transitoria.
Los animalesde abastopuedenactuarcomo portadoressanos,yaquesehanaisladoAeromonasen susheces,en proporciones queoscilan desdeel 4% (Sterny col., 1987)hasta el 11,8% (Gray, 1984). Wohlegemuthy col. (1972) relacionaron estosmicroorganismoscon brotesde abortosbovinos y Dobrescu (1978) con diarreaenlechones.Morsey Hind (1984)detectaronAeromonasenun 5.7% de nóduloslinfáticosde
cerdoy en avesseha puestode manifiestoun alto nivel de portadores.Por otra parte,Pasqualey col. (1994) describenun broteinfecciosocausadoporA. hydrophilaentortugas
conun95% de mortalidade indican queestosanimalespuedenactuarcomo reservoriosnaturales.Tambiénestos microorganismosse hanvisto implicadosen diversasinfeccionesde animalespoiquilotermos(Heywood,1968; Austiny Austin, 1987).En estesentido,una
de lascausasa las queseatribuyeel declive de las poblacionesde anfibios, en distintaszonas delmundo,hacereferenciaalas infeccionesporA. hydrophila (Blausteiny Wake,1995).
26

IAifRODUCCION
1.4. - Factores de virulencia
Aunquela mayoríade las investigaciones,hoy día, secentranen la identificaciónde
factoreso mecanismosde virulencia,potencialmenteimplicadosen infeccioneshumanasoanimales,sólo la capa S de A. salmonicida (Kay y col.. 1981) se ha asociado,
inequívocamente,a la patogenicidadde cepasde estaespecieen peces.La mayoríadelosfactores producidospor Aeromonasspp. seconsideranindicadoresde virulenciaporinferencia, es decir, por presentar moléculas similaresa otras que determinanlapatogenicidadde otrosmicroorganismos,comoE. coli. La falta de un modeloanimal quereproduzcala gastroenteritisasociadaa Aeromonas incrementael interésen el
establecimientode un esquemabasado enpropiedadesestructuralesy enzimáticas.asociadasa la célula, que puedanestarimplicadasen las infeccionesporAeromonasdehombresy animales.
1.4.1. - Productosexocelulares
1.4.1.1. - Exotoxinas
Atendiendoasurelacióncon la bacteria,las toxinassehandividido en exotoxinasyendotoxinas.Lasexotoxinassonsustanciassolublesy difusiblesquela bacteriasintetizaenel cursode sudesarrolloy secretaal exterior, mientrasquelasendotoxinasestánligadasadeterminadas estructurasy seconsiderancompuestostóxicospreformados,que seliberanpor lisis de la bacteria. Sin embargo,el hecho de que las endotoxinasseencuentrenlocalizadasen la superficiebacterianay de quealgunasexotoxinasse liberen,en granparte,
por lisis de la bacteriao estén asociadasa componentesestructuralesha hechoque, aúnmanteniéndoseestaclasificación,seconsideremás apropiado dividirlas,por sunaturalezaquímica,en toxinasproteicasy lipopolisacáridas.
Segúnsu importanciaen la patogeniade las enfermedades,las exotoxinasseclasificanen toxinas mayores,cuandoporsí solassonresponsablesdel cuadro clínico,y
toxinasmenores,cuandosólo coadyuvana la acciónpatógenajunto con otros factores.Segúnel sitio deacciónsedividenen toxinas queactúansobre lasmembranasy toxinasquelo hacensobre compuestosintracelularesy éstas,a su vez, segúnsu especificidady
tropismos,se pueden clasificaren toxinasde acción general,neurotoxinasy enterotoxinas.Lasenterotoxinasmuestran unaacciónespecíficasobre las célulasintestinales,queda
lugara la presentaciónde diarrea.Se clasificanen enterotoxinascitotónicascuandosólo

pv
a-
e
e
e
pv
e
IWTRODUCCION
producenalteracionesfuncionalesen el metabolismodel enterocitoy enterotoxinas
citotóxicascuandoproducen alteracionesanatómicaso necróticas(Pumarola,1987a).
Sehan descritodos tiposde enterotoxinasenAeronwnas:
- la enterotoxinacitotónica,similar a la toxinacoléricay ala toxina termolábilde E.
coli, potencialmenteresponsablede la diarreaacuosa (Dubeyy col., 1980;Ljungh y col.,
1982;Chakrabortyy col., 1984).Estatoxina redondeay estimulala esteroidogénesisde las
célulasY1 (Turnbull y col., 1984).- la enterotoxina citotóxica(Cumberbachty col., 1979;Johnsony Lior, 1981;Asaoy
col., 1984; Stelmay col., 1986),queda lugar asíntomas similaresa ladisentería,al igual
quealgunasespeciesde Salmonellay Shigella,caracterizadapor diarreamucohemorrágica.
Estossíntomassehandetectadosóloen un 20% de las infeccionesasociadasaAeronionas
(Graceyy col., 1982). En estudiosrealizadosen diferenteslineascelulares (Vero,bit 407,CHO, HeLa, etc.), los efectoscitotóxicos de los sobrenadantesde los cultivos deAeromonasdan lugar alesioneso muerte celular(Boulangery col., 1977; Barery col.,
1986; Kindschuhy col., 1987).
Al parecer,la enterotoxinacitotónicasedetectacon menosfrecuenciaquela citotóxica
(Stelma,1988)y, sin embargo,la mayoríade las infecciones relacionadasconAeromonas
muestransíntomas asociadosa laenterotoxinacitotónica.Además,Morgany col. (1985)
administraroncepascitotóxicas (dosis de 1Wt a 1010) a 57 voluntarioshumanos,observandounadiarrea débil ensolodosde ellos,y Kindschuhy col. (1987) afirmanque
el gradodecitotoxicidadde las cepasno secorrelaciona con la gravedadde los síntomasdeenfermedad(diarreao sepsis).por lo quesepodría sugerirqueesta enterotoxinacitotóxica
no juegaun papel prioritario en la patogénesis.Por otro lado, Potomski y col. (1987b)indicanquesólo la enterotoxinacitotóxicaes factorde virulencia,yaque fue la única queasociaronala apariciónde diarrea.
1.4.1.1.1.- Enterotoxina citotónica
Ljungh y col. (1981, 1982),en funcióndel punto isoeléctricoy la filtración porgel,separarondoshemolisinas(a y ~)y unaenterotoxina.Las característicasdela enterotoxina,parcialmentepurificada,secorrespondíancon la de unaenterotoxinacitotónica:las célulasy i scredondearonsin producirsemuertecelular,se vio estimuladala síntesisde AMPc y lasecreciónde esteroidesy, en asaligada de intestinode conejo,sedetectóun incrementoenla acumulaciónde fluidos. La inoculaciónintradérmicaenconejo produjosólo induración
28

INTRODUCCJON
de la piel y no resultóletal para elratón.El pesomolecularseestimóen 15.000 daltons.Las
actividadesbiológicas permanecieronestablesa 560C durante 10 minutos, perodesaparecierondespuésdeun tratamiento t¿rmicode 600C-2()minutos.No sehan detectadoreaccionescruzadascon latoxina del cólerani conlas hemolisinas(a y ~),aunquesehaobservadosuneutralizaciónconun antisuerohomólogo.
Dubey y col. (1980) purificaron tambiénunaenterotoxinacitotónica, aunquenodeterminaronsu pesomolecular.Chakrabortyy col. (1984)donanel gende la enterotoxina
citotónicade A. hydrophilaenE. cali y las enterotoxinasqueobtienenson establesa560C-20 minutos,elonganlascélulasCHO, provocanacúmulode fluidosen asa ligadade
intestinodeconejoy difierendela toxinacoléricay dela toxinaTL deE. cotí
Goslingy col. (1993) aislarony purificaronunaenterotoxinacitotónicade 15 kDa y
detectaronotro pico con estaactividad,que parecíacorrespondera otraenterotoxinacitotí5nicadistinta.
Chopray col. (1994) indican que Aeromonasspp. producedos enterotoxinascitotónicas:unatermolábily la otra termoestable.
Muchosinvestigadoreshandetectadola enterotoxina citotónica enescasonúmerode
cepas.En estesentido,Johnsony Lior (1981) no encontraronactividadcitotónica enningunade las 73 cepasutilizadasy Seidier y col. (1980) sólo observaron actividadcitotónica en el 6% de las cepas.Stelma (1988)basándose enestosdatos, sugiereque la
enterotoxinacitotónica debeser menosfrecuente quela citotóxica. Sin embargo,elsíndrome diarreicomás frecuentementeasociadoa Aeromonascoincide con el
correspondienteaeste grupode enterotoxinas.
1.4.1.1.2. - Enterotoxinacitotóxica
Cumberbatchy col. (1979) relacionaronlas actividadescitotóxica. hemolíticayenterotóxica,evidenciandola existenciade unaenterotoxina citotóxica,al igual queJohnson
y Lior (1981) quetambién relacionaronla actividadcitotóxicacon la enterotóxica.Sin
embargo,es en 1984cuandoMao y col., purificaronuna»-hemolisinaapartirde unacepaaisladade un pacientecon diarrea,que fue citotóxicasobrecélulas Vero y enterotóxica,
tanto enasa ligadade intestinode conejocomoen la pruebadel ratón lactante.Por otraparte, era capazde provocarla muerteen ratonesun minuto despuésde la inoculación
intravenosa,por lo que presentabapropiedadesenterotóxicas,¡3-hemolíticasy citotóxicas.El pesomolecularestimadofue de 60.00<) daltonsy seinactivabaa 560C en 5 minutos.Stelmay col. (1986) observan queel sueroelaborado(reine a estahemolisinaneutraliza
2’)

—Q
9:
u,
e
u
a-
a-
a-
u
a-
pv
a
a-
INTRODUCCION
completamentela actividad de cuatrocepas13-hemolíticas,por lo queconcluyenque la
¡3-hemolisina,por sí sola, puededeterminarcambiosen la permeabilidadintestinal.
Hostackay col. (1982). Blochy Monteil (1989),Asao y col. (1984). Bunning y col.
(1986), Stelma y col. (1986)y Burke y col. (1987) describieronmoléculascon estas
características.
Burke y col. (198 la) trabajancon 100 cepasde A. hydrophila. examinandosusactividadeshemolítica,citotóxicay enterotóxica. Aunqueencontraron una relacióntotal
entrela producciónde hemolisinas, citotoxinasy enterotoxinas,no hallaronunaasociación
constanteentrelas diferentescepas,por lo queconsideraronquehemolisis,citotoxicidadyenterotoxicidad,en ocasiones,podríanno ser propiedadesde una misma molécula.Además,observaronquela actividadenterotóxicadetectadaen ratapersistíadespuésdecalentarel sobrenadantelibre de célulasy seneutralizabacon laantitoxinacolérica,mientras
que la enterotoxicidaden ratoneslactantes eraextremadamentetermolábil y no seneutralizabacon dichosuero,por lo queexistiríandos enterotoxinasdistintas.
Jiwa (1983), trabajandocon cepasde Aeromonasde diversasfuentes (heceshumanas,pescado,conejos, lombrices), encuentrarelación entre citotoxicidad,
enterotoxicidady alteraciónde la permeabilidaden piel de conejo,por lo queatribuyetodasestasactividadesala enterotoxina.
Chopray col. (1986)pusieronde manifiestodos enterotoxinasen los filtradosde A.
hydrophil.a:unapresentaba reaccionescruzadas con laantitoxinacoléricay eraterniolábil,y
otra de carácterheterólogoy termoestable(1000C- 20 minutos).Potomskiy col. (1987a)purificaronunaproteína¡3-hemolítica,citotóxicay enterotóxicaqueno reaccionaba conlaantitoxina coléricay era termolábil.Mientras que Rose y col. (1989a,b)aislaronunaenterotoxinacitolitica, queya ha sido secuenciada(Chopra y col., 1993) y quepuede
detectarse,inmunológicamente,conanticuerposelaboradosfrenteala toxinacolérica.En 68 cepasde origen clínicoy ambiental,Watsony col. (1985) observaronque el
41% de las aisladasen casosde diarreano eranenterotoxigénicas.Esto sugiereque la
gastroenteritispodríadeberseaotros factoreso quelos métodosempleados paradeterminarlaenterotoxicidad(asa ligadade intestinodeconejoy ratoneslactantes)no permitendetectarla actividadde estamolécula.Por otra parte,el aislamientode Aeromonasspp. en hecespodríaúnicamentereflejar su ingesta,mientrasque lapresenciade unacargaelevadasedeberíaa la alteraciónde la ecologíaintestinal.Por todo ello, parecenecesarioprecisaryaclararlas relacionesentre bacterias,enterotoxinasy diarrea.
3(j)

IWTRODUCCION
1.4.1.1.3.- Hemolisinas
Lashemolisinasson proteínasextracelulares citoliticasmuy frecuentementeasociadas
aA. hydrophilay A. sobria (Brendeny Janda,1987; Kozaki y col., 1988). Pertenecenalgrupode citolisinasbacterianasformadorasde porosy causantesde Vrdida del contenidocitoplásmico poralteraciónde la membranacelular(Bhakdiy Tranum-Jensen,1988).
Ljungh y col., (1981)handescritodoshemolisinasenAeromonas:
- la a-hemolisina,caracterizadapor producirunazonaopacade hemolisis enagarsangre,que seinactiva a 560C en 10 minutosy que producemodificacionesmorfológicasencultivos celulares.No esenterotóxicaen asaligadade intestino de conejo,pero porinoculaciónintradérmicacausainduración,roturade capilaresy dennonecrosis,provocandoademásmuerteen el ratón.Supesomolecularaproximadoesde 65.000daltonsy supuntoisoeléctricode 4,8.- la p-hemolisina,detectadaen grandescantidadescuando latemperaturade incubaciónesde 370C, seinactiva conel mismo tratamientotérmico y da lugar a unazonaclara dehemolisisen agarsangrey vacuolización irreversibledel citoplasmaen ensayoscelulares.No es enterotóxicaen asaligadade intestinode conejo,pero sí letal en el ratónpor
inoculaciónintraperitoneal,presentandolosmismosefectossobrepiel de conejo quelacz-hemolisina.Supesomolecularseestimóen tomoa 49.000-53.0000daltonsy supuntoisoeléctricoen 5.5. Se la conocetambiéncomo aerolisina’(Bernheimery Avigad, 1974) ypresentareaccióninniunológicacruzadaconla hemolisinaa.
Al menos se han descrito otras dos 13-hemolisinas, distintas y con actividad
enterotóxica,producidasporAeromonas(Mao y col., 1984;Asaoy col., 1986; Kozaki y
col., 1987). Ambasproteínastienen un pesomolecularaproximado de 50.000 daltons,
aunquesus puntosisoeléctricosdifieren notablemente (5.3 a54 frentea7.6).Los ensayosde inmunodifusión indican que estasdos jB-hemolisinas están inmunológicanienterelacionadas(Asaoy col., 1986).
La escasaactividad enterotóxicade la [3-hemolisinaparcialmentepurificadaporLjung
y col (1981)y la elevadaactividadde la hemolisinadetectadapor Maoy col. (1984)sobreasa ligadade intestinode conejo,probablemente,se deba a lasdistintas técnicasde
purificaciónempleadas(Stelma.1988).Chakrabortyy col. (1984) han señaladoquelos clonesde E. cotíqueportaban los
genesde la hemolisinay la citotoxinade A. hydrophilason inactivos sobre elasaligadade
intestinodeconejo,posiblemente,por la incapacidadde E. cotí paraliberarlashemolisinasal medio (Chakrabortyy col.. 1986).
31

r
u
a-.
u
a-
a
a-
a
e
0<
e
IWrRODUCCION
1.4.1.1.4. - Mecanismode acciónde la [3-hemolisina
Lashemolisinasde Aeromonasproducenalteracionesen la permeabilidadvascular,
edemasy necrosis(Chakrabortyy col., 1986). Scheffer y col. (1988) estudiaronlacapacidadde las bacterias productorasde hemolisinasparaliberarfactoresmediadoresde lainflamación. La a-hemolisinade E. cali y la ¡3-hemolisinadeAeromonasson potentes
estimuladoresde la liberaciónde leucotrienos(LTB4 y LTC4) a partir de granulocitos
humanos.El primero esquimiotácticoparaneutrófilosy eosinófiloshumanosy el segundo
es un mediadorde las reaccionesanafilácticas,implicado en la broncoconstricción,producciónde mucusy aumentode la permeabilidad vascular.La liberaciónde estosmediadoresde la inflamaciónpuedenser la causa del incrementode la permeabilidadvasculary de la formaciónde edemasporhemolisinas,mientrasquela interacciónde laaerolisinacon los polimorfonucleocitoshumanospuede dar lugar a la liberacióndemetabolitos,apartir deestascélulas,queestimulenla secreciónen la mucosa.Estos hechos
podríanexplicarla apariciónde la diarrea.Chakrabortyy col. (1987) constataronque unacepamutante, deficiente en la
[3-hemolisina,eramenos tóxicaparael ratón,no causólesionesnecróticasen el puntodeinyeccióny no sedetectóen cultivos realizados24 horas despuésde la inoculación,mientrasquela cepapaterna[3-hemolíticaeramástóxica,produjo necrosisy seaisló hasta14 díasdespuésdesu inoculación.
Garlandy Buckley (1988)comprobaronque la13-hemolisinaesun proteínahidrofílicay solublequesesecretacomounaprotoxinainactivay se reactivapor la eliminaciónde 25aminoácidosdel extremoC terminal. La rupturade los glóbulosrojos implicatres fases:unión a los receptores, agregaciónde toxinasy rupturade la membrana deleritrocito.Vander Goot y col. (1993) observanque tanto la proaerolisinacomola forma activasondímerosde similarestabilidady solublesen agua.La protoxinaseactivaría eliminandoelpéptidodel carbonoterminal, que no estaríaimplicado en la formación de porosen
membranas,pudiendoentonces oligomerizarsey darlugar a la forma activa(Van derGooty col., 1994a,b).
Bunningy col. (1986)utilizaroncélulasde melanomaparacomprobarla producción
de los efectos,mediadosporel AMPe, de la [3-hemolisinapurificada (inhibicióndelcrecimientocelular, incrementode la actividad tirosinasay acúmulo de melanina)
característicosde las enterotoxinascitotónicas. Esta 13-hemolisina purificada era
32

INTRODUCCION
extremadamentetóxica para las célulasde melanornay no se laconsiderócon efectos
citotónicos.
1.4.1.1.5.- Factor que presentareaccionescruzadascon la toxinadel cólera(CTC)
Jiwa(1983),Burke y col. (1984b)y Tumbulí y col. (1984)han observado reacción
cruzadaentrela toxina coléricay la enterotoxinadeAeronionasspp. El usode la antitoxinacoléricaen técnicas ELISAmostróla existenciade un factor enterotóxicoy hemolítico en
Aeromonasspp.,que reacciona conlos anticuerposde la antitoxinacolérica. Enningún
casoseha podidodetectar homologíaentreeste factory la toxinacoléricapor lo queno seha podidodenominartoxina“cólera-hice”,sino“factor quepresentareaccionescruzadasconla toxina colérica” o “CTC”. La similitud antigénicay el hechode que seobservaraincrementointestinalde fluidossugierequeesenterotóxicoy, por tanto, un posiblefactordevirulenciaenAeromonasspp. (Campbelly Houston,1985).
Schultzy McCardell(1988)utilizaronsondasgénicasespecíficas, sintetizadasapartir
deciertasregionesde lassubunidadesA y B de la toxinacolérica,y comprobaronqueestas
sondashibridabanconel ADN de algunascepasdeA. hydrophila;además,la enterotoxinaqueproducíanestascepasreaccionabaconel suerode antitoxinacoléricay sela podía
describir comounaenterotoxinacitotónica.Potomskiy col. (1987b)detectaronunatoxina,a partir de cepasfecalesdeAeromonasspp.,que reaccionabaconla antitoxinacolérica,no
era citotóxica ni hemolíticay tenía efectoscixotónicos. Hoy día, aunqueno estácompletamenteestablecido,algunosautoresopinan que laenterotoxina citotónicay este
factorsepuedenconsiderarmoléculasmuy similareso inclusoidénticas(Janda.1991). Noobstante,hay queteneren cuentaque laenterotoxina citotónica,descritapor Ljungh y col.(1981)y Chakrabortyy col. (1984),esmástermorresistentey no presentareacciones
cruzadasconla toxinadelcólera,porlo queen todocaso,este factorquese inactivaa560Cen 10 minutos(Caliill, 1990)constituidaunaenterotoxinacitotónica distinta.
Las reaccionesinmunológicascruzadasdetectadasentrelasenterotoxinascitotóxicasdescritaspor Asaoy col. (1984),Howard y Buckley (1986>,Chakrabortyy col. (1986) yBunningy col. (1986)sugierenqueson moléculassimilares,aunque puedenpresentar
diferenciasestructuralesy funcionales importantesy, en cualquier caso,no se relacionanantigénicanientecon la toxinacolérica(Cahilí, 1990). Por todo ello, parece quesólo laenterotoxinacitotónicay la enterotoxinacitolítica purificadapor Rosey col. (1989a.b) seasocianconla toxinadel cólera.
33

pv
pv.
u
u
a-
IWTRODUCC!ON
1.4.1.2. - Proteasas
Las proteasas producen una acción patógena de lesión directa de los tejidos,
facilitando así lainvasividad(Sakai, 1985>; además, pueden tenerun efecto indirectoal
activarel precursorde la [3-hemolisina (Leungy Stevenson,1988). Ciertasproteasas
exocelulares,producidasporotrasbacteriasgram-negativas,como laelastasa(33 kDa)de
Pseudonwnasaeruginosay algunas proteasasde Serradarnarcescens(50y 56 kDa)tienen
un papelimportanteenla patogenicidad(Wandersman,1989).Aeromonasspp.secretaal menoscuatroo cincoproteasas,en funcióndel pH óptimo
y la especificidaddel sustrato(Nieto y Ellis, 1986).Las dosproteasasprincipaleshansido
descritaspor Leungy Stevenson(1988);unade ellasesunametalproteasatermoestabley,
la otra, unaserinoproteasatermolábil.La mayoríade las cepasde Aeromonas,analizadas
inmunológicamente,secretanambasproteasas.Loewy y col. (1993) han purificado ycaracterizadounanueva zinc-proteinasade 19 kDa.Aunqueel papelde las proteasasno está
claro, la LD50 parael salmóny la truchade unacepamutante,deficiente enproteasas,fue
de 1000a 10000vecesmásaltaque ladela cepapaterna,productorade proteasas(Sakai,1985).
L4.1.3. a Sideróforos
Los sideróforosson compuestosde bajo pesomolecular conunaalta afinidad ycapacidadde unión a distintasformasorgánicase inorgánicasde hierro, particularmentebajo condicioneslimitantes para estemetal (Crosa,1989>.Estos compuestos puedendesarrollarimportantes papelesenelestablecimientode la infección, ya que el hierroes
fundamentalpara el crecimientode la mayoríade las especiesbacterianas.Los
microorganismospatógenosson capacesde obtenerel hierro a partir de formascombinadas,mediantela síntesisde sustanciasquelanteso secuestradorasde hierro(sideróforoso quelinas)(Pumarola,1987b),asociándoseciertossideróforosextracelulares
abacteriasgram-negativas,comoE. colí, productorasde enfermedadesinvasivas(Payne,
1988). Fn el caso de Aeromonasspp., Janday col. (1994a) observanque las cepasimplicadasen bacteriemias.elaboran sideróforos,son muy hemolíticasy proteolíticasyresistentesal complemento.
Casi todaslas cepasde A. salmonicida,A. hvdrophila.A. sobria y A. caviaeelaboran
34

II’ffRODUCCION
uno o mástipos de sideróforos(Barghouthiy col., 1981; Chany Trust, 1983). En A.
salmonicidaexisteun mecanismode adquisiciónde hierro,que precisaun sideróforodebajo pesomolecular(Chany Trust, 1983).En el casode las Aeromonasmesótilas,lamayoríade las cepasde A. hydrophilay A. caviaey, algunasde A. sobria, producenunsideróforodenominadoamonabactina,o bien,el llamado enterobactina(Barghouthiy col.,1981, 1989; Zywno y col., 1989). Massady col. (1991)observandosmecanismosde
adquisiciónde hierro en distintas cepasde este género,uno de ellos, mediadopor la
amonabactina,permite utilizarel hierro de la Fe-transferrina,mientrasel otro proceso,independientede lossideróforos,accede alhierro de distintoscompuestoshemo.Telfordy
col. (1994)describen4 variedadesde amonabactinas producidaspordistintascepasdeA.
hydrophila bajo condicionesrestrictivasde hierro. Massady col. (1994)señalanquelasíntesisdelácido 2-3 dihidroxibenzoico, presenteen sideróforos, estácodificadaen dosgruposde genesdistintos, unodeellosdetectadoen las cepasproductoras deenterobactinay el otro en las de amonabactina.
1.4.2.- Característicasasociadas ala célula bacteriana
Muchosfactoresasociadosa la patogenicidad,comolos determinantes deinvasividad
o deresistenciaal suero,lasproteínasde la membranaexternareguladorasde hierro y lacapacidadpara enfrentarseal sistema inmunemediante lavariación antigénica nohansidoobjeto de un estudioprofundoque, sin embargo,podría aportardatosmuy interesantes(Finlay y Falkow, 1989;Smith, 1990).
1.4.2.1. - Fimbrias o pili
La presenciade adhesinasen las fimbrias o en otrasestructurasde la superficie
bacteriana(proteínasde la membrana externa,lipopolisacáridosy flagelos) explica la
adhesiónde las bacteriasgram-negativas.En bacteriasgram-positivas,la adherenciase
encuentraasociadaa otros mecanismos(ácido lipoteitóico, glicocalix u otrasestructurassuperficiales).
La adherenciarequiereel contactode la bacteriacon la célulapero, comoambassuperficiessonelectronegativas,se creaunafuerzade repulsión,quesólo esneutralizada,
en parte, por débilesimpulsosde atracción(fuerzasde Van der Waals,enlacesiónicos,
enlacesde hidrógeno,interacciones hidrofóbicas)que danlugara una barreraelectrostáticaque dificulta la aproximación.La presenciaen la superficie de la bacteriade moléculas
3ji

O
a-
0.
IWTRODUCCION
irolóbicasy. enespecialde adhesinas,que por supequeñotamañono estánsujetasalas
fuerzasde repulsión,facilitan el contacto.Una infección es un proceso secuencialcuyaprimera etapasería la fijación y
colonizaciónde las bacteriaspatógenasen las célulasepitelialesde la puertade entrada.Laadherenciade las bacteriaspatógenasal epitelioevita sueliminaciónpor factoresmecánicosy facilita sudesarrolloy multiplicación.Todoello, no sólorepresenta una protecciónfrentea losagentesexternos,sinoque, además,les permitealcanzarel númerocrítico de bacteriaso la concentraciónadecuadade productosparapoderiniciar la infección. El estudiode la
relaciónentreadherenciae infectividad ha demostradoque, en la mayoríade los casos,existeun gradodecorrelaciónalto,tanto en las infecciones invasivascomopor toxinas.Porotra parte, seha sugeridoque los fenómenosde adherenciatambiénintervienenen la
difusiónde infecciones bacterianassecundarias.Sehaseñalado quelas alteracionesde lascélulasepiteliales,producidasen la infecciónprimaria, podríandarlugara la pérdida deciertassustanciasde la superficiede la célula(fibronectina),quefacilitadael acceso alos
receptores,o biena la formaciónde otros nuevos,comosesuponequeocurreen las células
infectadasporvirus.Sin embargo,la adhesiónno constituyeun factor indispensableparala infección,
puesanulandoo reduciendolos mecanismosde defensade la mucosa,las bacteriassincapacidadde adhesiónpuedenmultiplicarsee incluso producirunaacciónpatógena.Enconsecuencia, lacapacidadde adherenciarepresentauna ventajaecológicapara lacolonizacióneinfecciónperono constituyeun requisitoimprescindible.
Lasfimbrias o pili sonapéndicesfilamentososde naturalezaproteicaconcapacidadantigénicay, frecuentemente,portadoresde adhesinas,que presentan lapropiedadde
combinarsecon receptoresde naturalezapolisacárida, localizadosen la superficiede las
célulasepiteliales,medianteuniones deltipo azúcar-lectina.La zonacombinanteestáconstituidapor un pequeñosegmentode las subunidades proteicasde la fimbria (regiónamino-terminal).
Lasfimbrias sepuedendiferenciarporunaseriede caracterescomo eltamaño,peso
molecular,composición enaminoácidosy poderinmunógeno y.además,en relacióncon lapresenciade adhesinas, porla capacidadde fijaciónselectivaen los receptoresde diversascélulasy especiesde hematíes responsablesdel fenómenodehemaglutinación;también,porla naturalezaquímicade estosreceptoresy, sobretodo,porque laadiciónde los mismosode sustanciasanálogaspermiteinhibir específicamenteel fenómenode adherenciao dehemaglutinación.Las adhesinasde las fimbrias, atendiendo a su sensibilidadal
monosacáridomanosa,se dividen en dosgrandesgrupos: sensiblesy resistentesa la
36

¡A’TRODUCCION
manosa.
Las fimbrias manosa-sensibles,MS o fimbrias de tipo 1 son las másfrecuentesyestánpresentesen las bacteriasgram-negativas.Presentanla propiedadde combinarseconreceptoresquecontienenD-manosa,queseencuentranen muchasglicoproteinas delmoco,
así como enlos fagocitosy hematíesde diversasespeciesde animalesy del hombre, dando
lugaral fenómenode la hemaglutinación, reacciónque seinhibepor laadiciónde n-manosa
al medio. Estasfimbrias seconsideranpoco relacionadascon elpoder patógeno,pero
intervienenen el procesodecolonización.
Lasfimbrias manosa-resistenteso MR, por el contrario,sonmenos frecuentesy secaracterizanporqueproducenla aglutinación de los hematíes,aún en presenciadeD-¡nanosa.Recientemente,seha demostradoquelas fimbriasmanosa-resistentesno formanun grupohomogéneo,sinoqueagwpantodaslas fimbriasquesecombinan conreceptoresespecíficosqueno contienenmanosa.Lamayoríade las fimbriasdeE. coli asociadas conpatogenicidad en el hombrey animalessonmanosa-resistentes(Pumarola,1987c).
Existeunasubclasede fimbriasque intervienenen la conjugaciónbacteriana. Estasfimbrias sonmáslargas,flexuosasy poconumerosas(1 a4 por bacteria)y finalizanen una
especiede botón. Son necesariasparatransferirel materialgenético desde las bacteriasdonantesalasreceptoras durantela conjugaciónbacteriana(García-Rodríguez.1987).
Los estudiosrealizadosconmicroscopiaelectrónicahan puestodemanifiesto quealmenos existendos tipos morfológicosde fimbrias en cepasdeAcromonasclínicasy
ambientales(Carrello y col., 1988):lospili rígidos(numerososporcélula)con unalongitudde 0,6 a 2,0 ~tm.,formados porsubunidadesde pesomolecularde 17.000a 18.000,y lospili flexibles (1-60 porcélula),denominados “minipilin”, porel bajopesomolecularde lasubunidad quelos constituye (4.000)(Ho y col., 1990;Honmay Nakasone,1990).Satoycol. (1989) hanobservadoqueno existenreaccionescruzadasentrelospili rígidosy los
antígenosde los factoresde colonización1 y II de E. cali. Ho y col. (1990)secuencianlaunidadconstituyentede lospili flexible (“mini pilir”), observandoqueestamoléculade 46aminoácidospareceser totalmentenuevay no muestrahomologíaconotrassecuenciasde
pilí conocidas.La expresióndel “mini pilin” se puederegularambientalmente,siendomáximaa 220C, enmedioliquido y en presencia dehierro reducido,mientrasquelospili
rígidosparecenserconstitutivosy aparecenen todaslas condiciones.Estos hechossugierenque la síntesisde los pili flexibles podría ser un requisito para la colonización enhospedadoresmamíferosdependientede las condicionesdel medio.
37

r
w
a-
a-
o
o
a-
a-
e
e;
e
O
‘y
¡PURO»UCCION
1.4.2.2. - Proteínasde la membranaexternade la paredbacteriana(OMPs)
Aoki y Holland (1985) han separadoelectroforéticamentelas proteínasde la
membranaexternade ochocepasdeA. hydrophila y otras ochodeA. salmonicida.En el
casode A. hydrophila, las proteínasde la membranaexternason distintasde unacepaaotra, aunquela mayoríadeellasconteníanunaproteínade 36 kDa; porel contrario, enlas
cepasdeA. salmonicidaseobservaronmuchasproteínascomunesatodaslascepas, siendola más abundanteuna de 32 kDa. Bajo condicionesrestrictivasde hierro, tanto A.
hydrophila cornoA. salmonicida,sintetizaronnuevasproteínasde la membranaexternacon
unpesomolecular entre68 y 93 kDa. Kuijpery col. (1989c)estudian lasproteínasde la
membranaexternaen 46 cepasfecales,apareciendolas bandasmásprominentesentre25 y45 kDa.En las cepasdeA. caviae,los perfilesde las proteínas fueronsimilares,mientrasqueen las cepasdeA. hydrophila y A. veronhl biotipo sobria seobservóunamarcadaheterogeneidad.
Atkinsony col. (1987)señalaronque lahemaglutinacióny laproducciónde piEl son
independientes,al observarque lacapacidadhemaglutinanteseasociaaunaproteínade lamembranaexternaquepermitiríaa las células adherirsea receptores(antígenoH) de los
eritrocitoshumanos.Estos receptorestambién seencuentranen lascélulas intestinales,porlo queestaproteínade la membrana externafacilitaríala uniónalascélulasintestinales,en
las que la producciónde enterotoxinas adquieresu máxima relevancia.Por otra parte,Wilcox y col. (1994)observanla expresiónde nuevasproteínasde la membranaexterna,
que no aparecenen caldo de cultivo, cuandoel crecimientoocurreen contenidoilealhumano.Quinn y col. (1994) aíslan, mediante cromatografíade afinidad basadaen la
reactividadconcarbohidratos,dosproteínas formadorasde poro en bicapaslipídicasque
podrían actuar comoadhesinas.
Lasaeromonassoncapacesde adherirseaeritrocitos,levaduras,células delepitelio
bucal y a la mucosaintestinalde conejopor mecanismos queimplican la existenciadeproteínasno finibriadasdela superficiebacteriana(Atkinson y Trust,1980; Levetty Daniel,1981). Singh y col. (1993), indicanque la adhesión alepitelio intestinalpuedeestarmediadapor factoresde colonizaciónfimbriadoso no y hemaglutinanteso no. Nevesy col.
(1994), detectanadherencia agregativaa células HEp-2en A. hydrophilay A. sobria,
similara la enteroadherenciaagregativadetectadaenE. cdi.Grayy Kirov (1993) observanqueA. vcronhi biotipo sobria prescntaunacapacidaddeadherenciaacélulasHEp-2superioral resto de las especies.
38

INTRODUCCION
1.4.2.3.- Lipopolisacáridosde la membranaexternade la pared bacteriana(LPSs)
Forman parte de la membranaexterna de la pared celular de las bacterias
gram-negativas,asociándoseal antígenoO y a las endotoxinas.En el lipolisacáridode lamembranaexternade la paredbacterianase describentresfracciones(Pumarola,1987a):
- Fracciónexternao cadenaslateralesdel lipopolisacárido,constituidapor unidadesdeoligosacáridos,cuyo número, composicióny secuencia diferirían segúnla especie
bacterianay seríanresponsablesde laespecificidaddel antígenoO.- Fracción centralo núcleodel polisacárido(core), formadoporoligosacáridosy
KDO (ácidoketodesoxioctónico) queseríacomúnentrelas bacterias gram-negativas,yexplicaríalas reaccionescruzadasquese presentanentrelas distintasespecies.
- Fraccióninternao lípido A, queseidentificaríaconla endotoxina,responsablede latoxicidad y de ciertasmanifestaciones clínicassimilaresen infeccionescausadaspordistintasespecies,que se liberaríapor lisis celular. Su toxicidad es inferior a la de las
exotoxinas, diferenciándosede éstasenquesu acciónesinespecífica.sontermoestables,nosedetoxican poraccióndel formol porlo queno setransformanen toxoidesy tienenescasopoderinmunógeno.
Al analizarlospolisacáridosdela regiónespecífica(cadenas lateralesO) del LPSpor
SDS-PAGEsehandetectadoal menostres modelos.Uno deellos,de carácterhomogéneo,serelaciona conel serogrupo0:11 deAeromonasy, al parecer, este tipode LPS desempeñaun importantepapel en el anclaje de la capa 5. Además,a esteserogrupose le harelacionadoconla mayorvirulenciaenpecesy con laresistenciaala acciónbactericidadel
suero. (Dooley y col., 1985; Kokka y col., 1991b).Estevey col. (1994),describendosgruposde cepaspatógenas,uno de ellos constituidopor cepasPAB+, pertenecientesalserotipo0:19,quepresentancadenasde tamañohomogéneoen el LPSy unaproteínade lamembranaexternade 50 kDa.Lascepasdel otro grupofueronserológicamentevariables,PAB-, presentaronun LPSheterogéneoy OMPs de 33 a 45 kDa.Porotro lado, Franckiy
Chang(1994)indicanqueel LPS podríaactuarcomounaadhesinaenA. sobria.
1.4.2.4. - Flagelo
Los miembrosdeAeromonasspp. son generalmentemóvilescon un único flagelopolarde 1,7 jlm, aunqueen cultivosjóvenes,a vecesseobservanflagelos lateralescortos(< 1,7 ~tm)(Stelma, 1989). Existe muy poca informaciónsobre el flagelode Aeromonas
spp..apesarde quela movilidad serelacionacon la patogenicidadya que. únicamente,las
3<)

0<
0<
o
st
o
IWTRODUCCION
especiesmóvilesseconsideranpotencialmentepatógenasparael hombre.
1.4.2.5.- Capa extra,proteicay externaa la paredbacteriana(CapaS)
La presenciade una “monocapamacromolecularperiódica”, de estructura
paracristalina.sobrela paredbacteriana seobservó,por primeravez, hace42 años,enfotografías realizadascon un microscopio electrónico(Houwink, 1953,citado en Sleyrty
Messner,1983).Posteriormente, se hacomprobadoque estacapaesmásfrecuente deloqueen unprincipio sesospechaba,observándosesu presenciaen Acinetobacterspp.,
Campylobacterfetus,Aeromonassalmonicida,distintas especiesde Bacillus, deClostridium,de Laciobaciflusy en otrosmicroorganismos(Yaxnaday col., 1981;Ishiguroy col., 1981; Evenberg y col., 1982; Tsuboi y col., 1982; Sleyrt y Messner,1983). La
capa5 se puedeconsiderarcomo la membranabiológicamássencilla desarrolladaalo largode la evolución;generalmente,estáformadaporun sólo polipéptidohomogéneode pesomolecularalto y, ocasionalmente,por carbohidratoscomo componentesminoritariosenforma de glicoproteinas.Los pesos molecularesdeterminadospor SDS-PAGEvaríande 40a200kDa.
Estádirectamenteimplicada enlas interaccionesentre la célula y su entorno y,desaparececuandola bacteria secultivade forma prolongadaen el laboratorio, indicando
quesu presenciaesun elementode protección paralos microorganismosque la poseen(Sleytr, 1978).
Se precisan5 x ío~ monómerosde la capa 5 para darlugar a una cadenabidimensionaltotalmentecerradaalrededorde un bacilo. Enalgunosmicrorganismos,la
síntesis de laproteínade la capaS pareceestar completamente controlada, dada la escasa
cantidadque se detectaen el medio de crecimiento(Thorney col., 1976),mientras que,en
otros,la producciónes excesiva(Beveridgey Murray, 1975; Smity col., 1981; Ya¡nadaycol., 1981).La capaSno sólodesempeñaunaacciónprotectorafrenteaataquesexternoseinternos, sinoque tambiéncontribuyea mantenerla morfologíacelulary juegaun papel
importanteenla adhesión. La existenciade canales,de 2 a3 nm de diámetro,no impediríael pasodenutrientesy productosde degradación,pero si podríaprotegera las célulasde laacciónde enzimaslíticas y de otros agenteshostilesdel entorno(Sleyrt,1978; Beveridge.
1979; Stewarty Beveridge,1980; Stewarty Murray. 1982). Seha observadoque lacapaSpuede evitarla liberaciónde macromoléculas,aumentando laconcentracióncelularsobre
todo en la fase exponencialde crecimiento(Sleyrt y Messner.1983). Ademásde las
funciones dependientesdel tamañode los poros,la capaS puedeactuarcomo una barrera
40

LVTRODUCCION
físicafrentea la respuestainmunedel hospedador (Trusty col, 1980; Ishiguroy col., 1981;Evenbergy Lugtenberg,1982; Evenbergy col., 1982;Munn y col., 1982).En ocasiones
se opone a lainfecciónporbacteriófagos (Ishiguroy col., 1981), aunquetambiénpuede
presentarreceptoresespecíficosa cienosbacteriófagos(Sleyrty Messner,1983).Además,
la capaS seconsideraimplicadaen fenómenosde adhesión como autoaglutinaciónyadhesiónaepitelios (Sleyrty Messner,1983), enel intercambioiónico (Beveridge,1978;Beveridge,1979) y en el efectotampónfrentea ácidose hidrogenionesde lacélula y el
entorno(Sleyrt y Messner,1983).En lo que serefiere a Aeromonas,en 1981 Kay y col, identificaron, con el
microscopioelectrónicode transmisión, unacapaadicional(originalmentellamadacapaA)externaala paredcelularen cepasautoaglutinantesdeA. salmonicida(HG3) patógenasparapeces.Los estudiosbioquímicosy genéticosindican queestácompuestapor una simplecadenaproteicasuperficialde 49 a 51 kDa (Kay y col., 1984). La capa5 confiere unahidrofobicidadsuperficialelevadaa las cepasautoaglutinantesdeA. salmoniciday eselfactordevirulenciamásimportante(Trusty col., 1983;Bellandy Trust. 1985;Ishiguroycol., 1985). La capacidadde unión al rojo Congo,la resistenciaa la lisis mediadapor elcomplementoy la asociacióna monocitosfagocíticosparecen relacionarsecon lapresenciade la capa5 enA. salmonicida(Ishiguro y col., 1985; Munn y col., 1982; Trust y col.,1983).
Posteriormente,en varias investigacionessimultáneas,se detectaroncepasde A.
hydrophila(GHI) y deA.veroniibiotipo sobria(GHE), provistasdecapaS,implicadasen
infeccioneshumanasy animales(Dooley y Trust, 1988; Paulay col., 1988; Kokkay col.,1991a). Las cepasde origen humanose aislaron, principalmente,en infeccionesextraintestinales,comoperitonitisy bacteriemias(Janday col., 1987). Independientemente
de su procedencia (humano,animal o ambiental),estascepas presentaron idénticasestructurasy propiedadesgenéticas,asícomo, característicasfenotípicas similares(Paulay
col., 1988;KokJcay col., 1991a).El análisiselectroforéticode lasproteínassuperficialesdela capa 8,indicé queson moléculasácidasconun pesode 52 a 58 kDa (Dooley y col.,1988; Kolcka y col., 1990, 1991a). La composiciónen aminoácidosde estasproteínasresultó similara ladelas proteínasdeA. salinonicida,aunqueno seobservóhomologíaenla secuenciaaminoacídicaterminal, queesrepetitivaen lascepasde A. sainionicida(Kayy
col., 1984; Dooley y col., 1988). También sedetectaron diferenciasfuncionalesconrespectoa la capaS de A. salmonicida,ya que no seobservóque seincrementasela
hidrofobicidadde la superficiecelular, ni la capacidadde unión del rojo Congo (Paulay
col., 1988). Recientemente.Thomasy Trust (1995) handonadoy expresadoen E. cotí la
41

‘y
0<
e
0<
St
e
a-
St
a-
e
5,
st
IMPRODUCCION
subunidadproteicaqueconstituyeestacapa.Kokka y col. (199 Ib) afirmanque lapresenciade capa5 no estádirectamenteimplicadaen la patogenicidadparael ratóneindican quesupapelen lapatogénesispuedesertotalmentedistinto al de lacapaSdeA. salmonicida.En
cualquier caso,las cepasdeA. hydrophilacon capaS, pertenecientesal serotipo0:11,se
consideranaltamentevirulentasparahombresy peces (Dooleyy Trust. 1988;Paulay col.,
1988).Algunasinvestigacionesseroepidemiológicas ponende manifiestoqueeste tipode
cepas representanel 20-30%de todas las de A. hydrophilay A. veronhi biotipo sobria
aisladasde infeccioneshumanas(Kokka y col.. 1992). En este sentido,Merino y col.(1993) han desarrollado unapruebainmunoenzimáticaparadetectarestetipo de cepasen
alimentos.
1.4.2.6. - Invasividad
La capacidadde Aeromonaspara producir síntomasdisentéricospodríarelacionarseconla invasividadcelular,comoocurreen Salmonellay Shigella(Watsony col., 1985).Al
investigar cepasde origen fecal, seobservóque la invasividad eramás frecuenteen A.
sobriaqueen el resto delas especies,aunqueestacaracterísticano se relacionabaconla
enterotoxicidad.Sinembargo, enel casodeA. hydrophila,sísedetectóasociaciónentrelaapariciónde síntomasde disenteríay la invasividad delas cepas(Watsony col., 1985).En1985, Lawsony col. (1985)observaronque6 de 10 cepasdeA. hydrophilaerancapacesde invadir célulasHEp-2; la composiciónplasmídicade estascepasinvasivassugirió que,probablemente,los genesresponsablesselocalizabanenel cromosomay que,en cualquier
caso,no estabaclarala relaciónentre invasividadiii viti-o y patogenicidad¡ti vivo.Gray y
col. (1990) comprobaronque entreel 14 y el 36% de las cepasde Aerontonasinvaden
célulasHEp-2,siendotambiénA. sobriala másinvasiva.Nishikaway col. (1994)señalanla adhesióne invasiónen célulasdecarcinomade colonhumano(Caco-2).
1.4.2.7. - Resistenciaal suero
MuchasAeromonas,tantomesófilascomo psicrótrofas.son resistentes a la lisis
mediadapor el complementohumano,de pecesy conejos(Munn y col.. 1982;Janday col.,1984a). Lamayoríade las cepasdeA. hydrophila y A. sobria son másrefractariasa laacción bactericidadel sueroque A. caviae; las cepas sensiblesparecenactivar elcomplementodando lugar aunarápidalisis (Brendeny Janda,1986).Mittal y col. (1980)asociaronla resistenciaa la acciónbactericidadel suerocon cepas altamentevirulentaspara
42

INTRODUCCION
peces. Sinembargo,Montenegroy col. (1985) no han detectado,en 66 cepasdeAeromonas,el gentraT,quecodificaunaproteínade la membranaexterna,responsable dela resistenciaal suerode E. cotí. Merino y col. (1994)observan que la resistencia a la lisis
mediadapor el complemento, ademásdeserdependientede la temperaturade crecimientoy
de asociarsea distintasformasdellipopolisacárido,puededebersea la rápida degradación
de algunoselementosde estesistema, impidiendola formacióndel complejo lítico.Janday
col. (1994b)handetectadoresistenciaa la acción líticadelcomplemento,principalmente,encepas del serogrupo0:11y provistasde capaS.
1.4.2.8. - Plásmidos
Entreel 20 y el 100% de las cepasdeA. salmonicida,A. hydrophilay A. sobria
examinadasportabanplásmidosde pesomolecularentre2 y 100 MDa (Toranzoy col.,1983a;Changy Bolton, 1987; Belland y Trust, 1989). La mayoríade las cepasde A.
salmonicidapresentanperfilesplasmídicosbastantehomogéneos.En A. hydrophilay A.
sobria, la frecuenciade plásmidosesmásbaja (20 a 58%)y las variacionesen los perfiles
plamídicossonmayores.Muchosde los plásmidosdeAeromonassonfactoresR que llevanmarcadoresde resistenciafrente a la tetraciclina,ampicilina, tobramicinay kanamicina
(Toranzo y col., 1983a; Hedgesy col., 1985; Chang y Bolton, 1987). Toranzo y col.
(1983a) observaronen una cepade A. hydrophila,“curada” de plásmidos,numerosasdiferenciascon respectoa la cepapaterna,que afectabana la producciónde indol ygelatinasa,alaspropiedadessupeificialesy ala resistenciaala tetraciclina.Posteriormente,Hanesy Chandíer(1993) detectanun plásmido de 40 MDa queregulala adhesividadacélulasCaco-2, laactividadhemolíticay la resistenciaaantibióticos.
En losúltimos añosseha intentadoestablecerel papelquelasaeromonasjueganenlas enfermedadeshumanasy animales.Los mayoresavanceshacen referenciaal
conocimientode nuevasespecies,la identificaciónde estructuras (capaS, “mini pum”) y la
caracterizacióngenéticade factoresde virulencia(hemolisinasy proteasas).Sin embargo,quedan muchascuestionespor resolver, siendoprimordial el desarrollode un modeloanimal dondese reproduzcala diarreaasociadaa Aeroinonasy, por tanto, seaposibleestablecer,inequívocamente,el papel de Atromonas comopatógenosgastrointestinales.
43

a-
St
a-
0’
a-
St
a-
st
e
st
st
0<
IWTRODUCCJON
Tambiénson necesariaslas investigacionessobreproductos extracelularrsy asociadosa la
célula,encepas patógenasy no patógenas, paradeterminarlos factoresimplicados en la
virulencia.La frecuenciade los distintosgruposde hibridaciónrequiere tambiénun estudio
másprofundo con el fin de aclararla epidemiología deestetipo de infecciones(Janda,
1991).Por otro lado,son necesariosexámenesrutinariosde hecesdiarreicasy de alimentossospechosospara valorar la importancia sanitariade las gastroenteritisasociadasa
Aeromonasspp. transmitidaspor alimentos.Algunosestudiossugierenquesu presenciaen
alimentosrepresentaríaun riesgobajo,ya quetantoestosmicrorganismos comosustoxinas
sontermolábiles,la producciónde toxinases máselevada enmediosbacteriológicos queenlos alimentosy. aunque algunas cepaspuedencrecercompetitivanienteen alimentosa50C,
la detecciónde unacargaelevadaen los mismosno pareceserfrecuente(Kirov, 1993).
Hasta que las cuestionesconcernientesa la patogenicidadde Aeromonasy su
comportamientoen alimentosno esténtotalmenteresueltas,la presenciade un númeroelevadodeAeromonasen alimentosdeberíaserconsiderada,únicamente,como un riesgopotencialparala salud, principalmente,en personasinmunodeficientes.
44

II. OBJETIVOS DEL TRABAJO

t
st.
mt
0<
ab
0<
sfr -
0<
e
e
e
0<-

OBJbIT VOSDEL TRABAJO
El planteamientode este trabajosecentréen el estudiocomparativode cepasdeAeromonasspp.móvilesaisladasde alimentosy de heces diarreicashumanas,en cuanto a
susposibilidadesde llegara produciruna acciónpatógenaenel hombre.En consecuencia,unavezseleccionadoslos mediosde cultivo y técnicasde identificacióny aislamientode
másgarantíay despuésde compararla resistenciade ambostipos decepas,se abordéel
problemadel estudioy valoraciónde los factoresde virulenciaasociados alos productos
exocelulares,propiedadessuperficialesy característicasbioquímicasde las cepasde
Aeromonasspp. Paraconseguir estosobjetivosse estructuróel trabajodel siguientemodo:
- Evaluacióndel comportamientode tres mediosselectivospara el aislamientodeAeronzonasspp.móviles,así como selecciónde las pruebasbacteriológicasy bioquímicas
quegarantizanla correctaidentificación deeste grupobacterianoy sudiferenciaciónanivelde especie.
- Determinaciónde su incidenciaen alimentos españoles(carnes, pescados, mariscos,etc.)recogidosde supermercadosy establecimientosde venta alpúblico.
- Comprobaciónde la resistenciadeAeromonasspp.frenteadiferentesparámetrosfísicosy
químicos(temperatura,pH, clorurosódico,nitritos, cloro, antibióticos, etc.).
- Estudiode los factoresde virulenciadecepasaisladasen alimentosy en hecesdiarreicashumanas:
Actividad hemolitica (eritrocitosde corderoy conejoy sangrede caballoy cordero).
• Actividadenzimética(proteasasy lipasas).Citotoxicidad(lineascelularesestablesVero y mieloma).Hemaglutinacióne inhibiciónde lahemaglutinaciónporazúcares.Autoaglutinacióny aglutinaciónconacriflavina.
• Hidrofobicidadsuperficial(uniónafiltros de nitrocelulosay adsorciónahidrocarburos).Detección depilí y de la capa5 mediante microscopiaelectrónicade transmisión.
• Característicasbioquímicasquepudieranrelacionarseconla patogenicidad.Asociacionespresentesentrelos factoresde virulencia estudiados.
47

r
r
mt
ab
a-
ab.
mt
a-
u
a-
e
a-
SI
SI

III. MA TERIA L Y METODOS

r
ab
a-
a-
e
SI
e
a-
e
e
mt
a-
5,
mt
a-

MATERIAL Y MFYTODOS
111.1. - Material y equipo
- Materialde vidrio (pipetas,tubos,matraces,etc.) “Pyrex” o de calidadsimilar.
- Material de plásticoestéril (microplacas.placasde petri, botellas,tubos,etc.) “Nunc”,“Falcon” y “Sterilin”
- Micropipetasautomáticas P-20,P-200,P-1000y P-5000(Gilson),pipeteadorautomático
(Falcon).agitadores,etc.
- Destiladorde agua “Pobel”.
- Homogeneizador“Stomacher”mod.400 Colworth.
- Balanzas“A/D” mod.ER-120A. “A/D” mod.ER-600Ay mod. ER-3000A.- Ph-metroMetrohmmod.654.- Autoclaves“Selecta”mod.437-G.- Estufadeaire forzado“Heraeus” mod. KFTU-K.- Filtros “Millipore” de 0,22 y 0,45 jm de diámetrode poro. Lafiltración de grandes
volúmenesserealizóconun sistemaconectadoauna bombade vacíoEYELA A-3S.- Sistemade ultrafiltración tangencial“Minisette” TM de “FILTRON”, conectadoa unabombaperistáltica“Masterflex” y con 3 paquetescompactos,“cassettes”o cartuchosde
membranas(serie“Omega”, “FILTRON”) específicospararealizarcortes molecularesa50,30 y 8 kDa.- Filtros denitrocelulosa“Millipore” mod.SCWP.- Centrífugas“Sorvail” mods. RC-5B y GLC-l con rotores SS-34y OSA y
microcentrífuga“Heraeus” mod.Biofugeconrotor 1220.- Sistemade evaporación-centrifugaciónal vacío “Howe” mod. GV1 conectadoa una
bombade vacío“Titan” mod. Trap.
- Cámarade flujo laminar“Telstar” mod.CE-A.- Estufas“Heraeus” mods.K.B-500, B6200y BK-600 y “Selecta”,mod.Termotronic338.
Bañosde aguao propilenglicolprovistos contermostatos“Selecta”,mod.Tectron.Bañodeaguacon agitaciónorbital “GFL” mod. 1092 conectadoa una unidadde refrigeración“Braun” mod. Frigomix U.
- Microscopioóptico “Nikon”, mod. L-Ke, equipadocon un dispositivodecontrastedefases.- Ultraniicrotomo“LKiB” mod.UltratomeIII,- Microscopioelectrónicode transmisión“Zeiss’ mod. 902 conequipo fotográfico.- Espectrofotómetros“Kontron” mod. Uvikon 820 y “Hitachi” mod. U-2000, conimpresoratérmica“Kontron” mod. Uvikon LS Thermoprinter4B. Lectorde microplacas
SI

a-
a-
ab
mt
0;
0;
a
a
e
MATERIAL Y METOI)OS
“Cultelc” mod.Tietertek MultiskanPlus.
- Congeladores“Liebherr”, mod. GT6102 y “Sanyo” mod. Ultra low. Frigorífico
“Kelvinator” mod. AKR. Recipientede Nitrógenolíquido
- Cámaracon equipo de filtración-esterilizaciónde aire “Telstar” y acondicionadorde
temperatura“Interelisa”, provisto de: campanade flujo laminar “MDH” mod. 20134;
centrífuga“Sorvalí” mod. TC6 con un rotor H-400;incubadorde£02con camisade agua
“REVCO” mod.Ulúma; bañode aguacon temrnstato‘Bunsen” mod.BA-12; microscopio
invertido equipadoconcámarafotográfica“Olympus” mod.CK2 y frigorífico y congelador
“Liebherr”.
1111.2. - Productos biológicos
111.2.1. - Microorganismos
Seemplearon100cepasdeAeronionasspp.:4 procedentesde la ColecciónEspaflolade Cultivos Tipo (Burjasot, Valencia),15 aisladaspor el Dr. J.Reinaen hecesdiarreicashumanas(Hospital Son Dureta, Palmade Mallorca) y el resto obtenidasa partir de
alimentosyaguas.(Tabla111.1).
111.2.2. - Alimentos
Se tomaronmuestrasde los siguientesalimentos:carnes(vacuno,cerdo, polío ycordero),pescados(pescadilla,gallo, jurel, sardina,trucha),mariscos(chirlas, mejillones,gambas,cigalas),lechecruda,quesofrescoy ensaladaspreparadas.
111.2.3. - Aguas
Serecogieron muestrasde aguapotabledorada,aguapotableno doraday aguanopotable.
111.2.4. - Preparados hematológicos
La sangrede caballoy cordero,asícomo,las suspensionesdc eritrocitosdeconejoy
corderofueronsuministrados por“Biomerieux”.
52

Tabla 111.1 - Cepas de Aeromonasspp. móviles empleadas en este trabajo
Especie Origen Especie Origen
A. hydrophilaA. hydrophilaA. hydrophilaA. hydrophilaA. hydrophila
A. sobriaA. sobriaA. sobriaA. sobriaA. sobriaA. caviaeA. caviaeA. caviaeA. caviaeA. caviae
A. hydrophitaA. hydrophilaNoclasificada
A. sobriaA. hydrophilaA. hydrophilaNo clasificadaA. hydrophita
A. sobriaNo clasificadaNo clasificadaA. hydrophitaNo clasificada
A. caviaeA. sobria
NoclasificadaA. hydrophila
A. sobriaA. hydrophitaNoclasificadaNoclasificada
A. sobriaNo clasificadaA. hydrophitaA. hydropbilaA. bydrophilaA. hydrophilaNo clasificada
A. sobriaA. sobria
CECT (839)*CECT (398)*CECT (838)*CECT (837)*
HecesdiarreicashumanasHecesdiarreicashumanasHecesdiarreicashumanasHecesdiarreicashumanasHecesdiarreicashumanasHecesdiarreicashumanasHecesdiarreicashumanasHecesdiarreicashumanasHecesdiarreicashumanasHecesdiarreicashumanasHecesdiarreicashumanasHecesdiarreicashumanasHecesdiarreicasbumanasHecesdiarreicashumanasHecesdiarreicashumanas
Aguano potableAguano potableAguano potableAgua no potableAgua no potableAgua nopotableAgua nopotableAgua nopotableAgua nopotableCarnede pollo
CarnedecorderoCarnedevaca
Carnede corderoCarnede corderoCarnede corderoCarnede polloCarnede polloCarnede polloCarnedevacaQueso frescoQueso fresco
Carne de polloCarne de pollo
CarnedecorderoCarnede polloCarnede cerdoCarnede cerdo
Carnede corderoCarnede corderoCarnede cerdo
5051525354555657585960616263646566676869707172737475767778798081828384858687888990919293949596979899loo
No clasificadaNoclasificadaNoclasificada
A. sobriaA. caviae
A. hydrophi¡aA. hydrophi¡a
A. sobriaNo clasificada
A. sobriaNo clasificadaNo clasificadaNo clasificadaA. hydrophilaNo clasificadaA. hydrophilaNo clasificadaA. hydrophila
A. sobriaNo clasificadaNo clasificadaA. hydrophilaNo clasificadaA. hydrophilaA. hydrc’philaA. hydrophilaA. hydrophilaNo clasificadaA. hydrophila
A. sobriaA. hydrophilaA. hydrophitaNoclasificada
A. caviaeA. sobria
A. hydrophilaA. hydrophila
A. caviaeA. hydrophila
A. sobriaA. caviaeA. sobriaA. sobria
No clasificadaA. hydrophitaNo clasificadaA. hydrophila
A. caviaeA. hydrophila
A. sobriaA. hydrophila
Carne de polloCarnede polloCarnede polloCarnede cerdoCarnedecerdoCarnedecerdoCarnede cerdoCarnedecerdoCarne de polloCarnede polloCarne de cerdoCarne de cerdoQuesoftescoCarnede vraCarne de pollo
PescadillaPescadilla
JurelJurelJurel
PescadillaGalloGallo
SardinaJurel
PescadillaJurel
SardinaTruchaTrucha
JurelPescadillaPescadillaMej iliónChuñaChirla
GambasGambasMejillónMejillónMejillónChina
GambasCigalaChirlaCigala
MejillónMejillónMejillónChirla
Leche crnda
* - Nómerocori-espond~eflteenla Colección Espaflolade CultivosTi~ (CECT) (Budasol. Valencia)
1 A. hydrophila2 A. hydrophila3 A. caviae4 A. sobria
5678910111213141516171819
202122232425262728293031323334353637383940414243444546474849
53

r
a-
a-
*
a-
0;
e
e
O’
e
e
O’
*
MATERIAL YMP~ODOS
111.2,5. - Líneas celulares estables
Se utilizó la líneacelularVero (riñón demonoverdeafricano),cedidaporel CentrodeBiología Molecularde la Universidad Autónomade Madrid y célulasde mielomade ratón
(P3X63-Ag8.653),procedentesdel Instimto Llorentede Madrid.
111.3. - R&aflixn~
Los reactivosutilizadosfueronde la calidad“reactivo” suministradospor “Merck”,“Sigma”, “Fluka” y “Panreac”.
Los productosempleadosen las experienciasmicrobiológicasprocedieronde lasfirmas “Oxoid”, “Merck” y “Difco”.
Los cultivos celularesserealizaroncon los productossuministradospor “Gibeo”.
111.4. - Mkladns
111.4.1. - Preparación de los medios de cultivo
Sellevó a cabo,de acuerdocon lasinstruccionesde los suministradores.Seajustaron
al pHcorrespondientey seesterilizarona 1200C y una atmósferade presióndurante20minutos, exceptoaquellosmedioso suplementosen los queseespecificaotrométododeesterilización,
III.4,L Prepar-adón-cle---soluciones
Se realizaron en aguadestilada,exceptoaquellasenlasqueseindicabaotrodiluyenteparticular.
111.4.3. - Recuentos microbianos
Pararealizarlos recuentos se hicierondilucionesdecimalesdelasmuestrasen suerofisiológico (0,85%)o en aguade peptona(1%) estériles.Las dilucionesse sembraron(en
masao en superficie)en los mediossólidos.utilizando dosplacasparacadadilución.
54

MATERIAL Y METanOS
111,4.4. - Aislamiento de colonias
Se realizó de acuerdocon el método descrito por Ordóñez (1978).Paraello senumeraronlas coloniasde las placas,en las queaparecían entre100 y 200 colonias,y seeligió el 20%de ellas,de acuerdoconunatablade númerosestrictamentealeatorios.
111.4.5. - Toma demuestras
Lasmuestrasde un mismoalimentoseadquirieronen distintosdíasy comerciosde
Madrid. La lechecrudaprocedióde la granjaPriégolaLa Chirigota deMadrid. El agua
potabledorada serecogiódela redde distribución deMadrid,el agua potableno doradadeun manantialsituadoen el Parque delOestede Madrid y las muestrasde aguano potablede
distintostramosde losñosManzanaresy Guadarrania.
111.4.6. - Mantenimiento de las cepas
Medio empleado:
Caldo triptona sojacon extractode levadura <TSB-f.YE):
Caldo triptonasoja: 31g11
Digetidopancreáticode caseína 17 g/lDigeridopapafnicodeseinilladesoja 3 g/lClomro sódico 5 g/lFosfatopotÁsico dibésico 2,5 gIlGlucosa 2,5 g/í
Extracto de levadura 6g/l
Todaslas cepasse conservaron a-80~Cy a ~20oC,en caldotriptonasojaconextractodelevadura(0,6%)(TSB+YE) y glicerol (12 a 15%).
111.4.7. - Revitalización de las cepas
Previamentea la realizaciónde los ensayosse revitalizaron lascepas,partiendo
siemprede un cultivo purode 24h obtenidoenTSB+YE a280£.
55

r
a-:
rs
a-
St
a-
St
e
e
St
0;
O’
•5
St-
MATERIAL YMKTODOS
111.4.8. - Manejo de lineas celulares estables
111.4.8.1. - Mantenimiento
Seresuspendieron5x106 célulasporml en la mezclade congelación(90% de suero
fetal bovino y 10% de dimetilsulfóxido) y se dispensanen volumenesde 1 ml en vialesestériles.Se conservarona -850Cen un arcén congeladory a ~l50ÓCen nitrógenoliquido.
Tras la descongelación,las célulassemantuvieronen los mediosde cultivo a 370C,conun 5% de CO
2 y una humedadrelativapróximaal 100%,sustituyendoel medio de
cultivo por mediofresco reciénpreparadoy ajustandoel númerode células, siemprequeseconsideró necesario,con el fin de mantenerla poblacióncelularen fasede crecimientoexponencial.
111.4.8.2. - Recuentos celulares
El númeroaproximadode célulasseestiméenunacámarade Neubauercon laayudade un microscopioinvertido. Previamente, lascélulasde mielomase tiñeron(Id) conThpanazul al 2% en PBS.
111.4.8.3.- Cultivo de célulasVero
Medio deFaglemodificado por Dulbecco:
CaCI2.2H20Fe(No3).9H20KCIMgSO4.7H20
NaCJNaHC%NaH2PO4.2H20O-GlucosaFenolrojoPiruvatodesodioL-Arginina.HCIL-CistinaL-GlutaminaGlicinaL-Histidina IICI.1120L-lsoleucinaI~-Leucina
264
0,10
400200
64003700
141100015HO8448580
mg/l
mgflmg/lmg/lmg/1mg/lmg/lmg/1mg/lmg/lmg/1mg/lmg/]
30 mg/l42 mg/l
105 mg/l105 mg/I
L-Lisina HCIL-Metionina
L-FenilalaninaL-SermaL TreoninaL-TriptófanoL-TirosinaL-ValinaPantotenatocálcicoCloruro de colinaAcido fálico1-InositolNicotinamidaPifidoxal HORiboflavinaTiaminaHO
56
146
30
6642
9516
72944447,24
mg/l
mg/1mgIlmg/l
mg/]mg/lmg/lmg/lmg/1mg/lmglImg/lmg/l
4 mg/l0,4 rng/l
4 mg/1

MATERIAL YMETODOS
El medio decultivo completoutilizadoparalascélulasVero contenía:
- MediodeEaglemodificadoporDulbecco- Suerofetal bovino (previamenteinactivadoa 56oC~ 10minutos)- Soluciónde Penicilina-Estreptomicina (10000UI y 10 mg/mí, resp.)- Soluciónde L-Glutamina200mM
100ml
lOmí
lml
imí
Las condicionesdecrecimientofueron37”C, 5%de CO2y humedad relativaalta.
111.4.8.4- Cultivo de célulasde mieloma
Medio RPMI 1640sin glutamina
Ca(N03)2.4H20KCIMgSO4.7H20NaCíNaIICO3NaH2PO4(anlúd)O-GlucosaGlutationaFenolrojoL-ArgininaL-AsparraginaÁcido L-AspárticoL-CistinaÁcido L-OlutémicoGlicinaL-HistidinaL-Hidroxiprolina[-IsoleucinaL-Leucina
L-Lisina HCI
1004001006600200080020001520050205020101520505040
mg!)mg/lmg/lmg/lmg/l
mg!)mg!)mg/lmg/lmg/lmg!)mg!)mg!)mg/1mg!)mg!)mg!)mg!)mg!)mg!)
L-MetioninaL-FenilalaninaL-ProlinaL-SerinaL-TreoninaL-Triptéfano
L-TirosinaL-ValinaBiotinaPantotenatocálcicoCloruro decolinaÁcido félicoi-lnositolNicotinamidaÁcido Para-aminobenzoicoPiridoxinaMCIRiboflavinaTiaminaMCIVitaminaB12
15
15
2030205
20200.20.331351110,210,005
El mediode cultivo empleadoenestalíneacelularse componíade:
- MedioRPMI 1640sin glutamina- Suero fetalbovino (previamenteinactivadoa56
0C- 10minutos)- Soluciónde Penicilina-Estreptomicina (10000UI y 10 mg/mí, resp.)- Soluciónde L-Glutaniina200mM
Lascondicionesde crecimientofueron370C, 5% de£02 y humedad relativaalta.
mg/lmg/lmg/lmg/lmg/lmg/l
mg/lmg/lmg!)m~lmg/lmg!)mg!)mgIlmg/lmg/lmg!)mg/lmg/l
100 ml
iSmí
lmlimí
57

St
8W
w
a-
e
It
a-
St
e
e
MATERIAL YMFTODOS
111.4.9. - Técnica para comparar el comportamiento de los medios selectivos
(AAA, AGAP y mA) en el aislamiento de Aeromonas spp. móviles.
- Mediode Aeronwnos(mA) (Ryan, 1985.DatosproporcionadosporOxoid):
Proteosa-peptonaExtracto de levaduraClorhidrato de L-lisinaClorhidrato de L-argininaInositolLactosaSorbosaXilosa
5 gIl33,522,51,533,75
gilg/lg/lg/lg/l
gIlgil
Salesbiliares n03TiosulfatosédicoClorurosédicoCitratoamónicofénicoAzul de bromotimolAgarAzul de timolAmpicilina
pH 8±0,1
Las coloniasdeAeronionassonde colorverde etNaconel centroinésoscuroy de 0,5a 1,5 mm dediámetro.
- Agarglutainato penicilina(AGAP) (Kielwein, 1969):
L(+)glutamato sédicoAlmidón hidrosolubleDihidrógeno fosfatopotásicoSulfato demagnesioRojo de fenolAgarPimaricinaPenicilina
10 gil20 g/l20 gil0,5 gIl0.36 g/l
gIlgil
120,01100.000
pH 7,2 + 02
Aparecencolonias amarillasde2-3 mm dediámetrorodeadasdeunazona ainauiflenta.
- Agaralmidónaxnpicilina(AAA> (Palumboel al. 1985a):
Almidón solubleAgarconrojo de fenol:
Proteosa peptonanlExtractodecarneClorurosédicoRojo fenolAgar
10150,02515
Ampicilina
10 g/l31 g/l
g/lgilgilg/lg/l
0,01 g/lpH 7,0±0,1
58
310,6750.80,0412,50,040,005
g/lg/l
gIlgilg!)g/lgIlg/l

MATERIAL YMETODOS
Aeronionasspp.dsrn lugaracoioniasamarillas,de 3-5 mm de diámetro,quese caracterizanporque
despuésdeafiadir 5 mIdelugol <Soluciónyodo yoduradadeLugol: Yodo 1 g; YoduropoAsico2 g;
Agua destilada200 mí)en la superficie de la placase tornan de colormiel con un halo amarillo
- Agar triptona sojaconextractode levadura(TSA+YE>:
TSB+YE,suplementadoconagar (1,5%)
Losantibióticosseañadieronalosmediosdecultivo enformadesoluciónesterilizada
por filtración (poro = 22 I.tnx)
Anuasy alimentos: aguadorada,leche cruda,carnespicadasde vacuno,cerdo y poíío,
pescadilla,trucha,chirlasy gambas.
£~¡a~:A. hydrophilaCECT 839, A. hydrophilaCECT398, A. caviaeCECT 838 y A.sobria£ECT837.
~i~¡: Decadaalimentoseobtuvieron cincofracciones: alimentossólidos(10g triturados
y homogeneizadosen 90 ml de agua de peptonaal 1%); alimentoslíquidos (10 ml
homogeneizadosen 90 mide aguade peptonaal 1%).
Las cepasdeAeromonasse sembraron en TSB+YEy seincubarona280C durante
18h. La concentraciónaproximadade los cultivosde las cuatrocepassedeterminóporespectrofotometríaa600 nm,mediantela utilizaciónde rectaspatróncalculadaspreviamente
paracada cepa(Figuras111.1, 111.2,111.3 y 111.4),que relacionabanla absorbanciaa 600nm con el 1og
10 de las ufc/m] decadacultivo en fasedecrecimientoexponencial.
Cuatro de las muestrasobtenidasdecadaalimentose inocularoncon 1 ml de un
cultivo de cadaunade las cepasde Aeromonasspp., variando elvolumendel alimento
inoculadode modo que, laconcentraciónfmal decada cepafueseaproximadamentede ío~
ufe/mí de muestrade alimento. A continuación, se efectuaronlos recuentosde todaslas
muestrasen los tres mediosespecíficosdescritos.La quintamuestrade cadaalimentose
utilizó pararealizarlos recuentosde la flora naturalde los alimentos,en agartriptona sojay
en los tresmediosde cultivo selectivosparaAeromonas.
Lasplacasse incubarona280C-24h, exceptolas del medio AGAP quese incubaron
durante48h.Trasrealizarlos recuentos,la capacidadde inhibición de losmediosselectivos
59

e
e
a-
e’
a-
st
Ir
u-
e
u
e
u
e
e
MATERIALYMETODOS
1,2
0,8
0,4
0,0
y - 9,2265 + l,1323x R = 0.956(p<0,01)
7,5 8.0 8,5 9,0 9.5
¡ng ufe/mí
Figura 111.1 - Relaciónentrela absorbancia(600 orn) yel log ufc/ml deun cultivo deA. hydrophilaCECT 398
iw
o=‘a
aoa
.0
ou,
.0
1,2
0,8
0,4
0,0
y - 5.0045+ O,66547x R 0.998(p <0,0001)
7,5 8,0 8,5 9,0 9,5
log ufe/mí
Figura 111.2 - Relaciónentre la absorbancia(600 nnfl y el log ufc/ml deun cultivo deA. hydrophila CECT839
1,2
0,8
0,4
0,0
y - 5,3765+ 0.71186x R = 0,988<0,01)
‘7,5 8,0 8,5 9,0 9,5
Iog ufclmnl
Figura 111.3- Relación entre la absorbancia(600 nmn) y el Iog ufc/ml de un cultivo deA. sobña CIICT 837
ia
ee‘e
aoaa.01.o
.0
1,2
0,8
0,4
0,0
y - 6.6688 + O.8769x R = 0,997(p<0,O01)
7,5 8.0 8,5 9,0 9.5
IOg i¡fchul
Figura 111.4 . Relación entre laabsorbancía(600 nm) y elJog ufc/rnl de un cultivo deA. caWae CECT 838
60
ia
o=~0
aoua
.03-ou,
.0
ia
ee‘a
aoaa
.03-ou,.0

MATERIAL YMETODOS
frentea la flora naturalde los distintosalimentossedeterminócomparandolos recuentosde
coloniasen estos medioscon los obtenidosen TSA+YE. Finalmente,la capacidadde
recuperaciónde las cepasde Aeromonasspp. móviles se determinécomparandolos
recuentos en los medios específicoscon el nivel deinoculacióndeterminado por
espectrofotometría.
111.4.10. - Técnica para investigar el comportamiento de A. sobria CECT
837 en los medios selectivos (AAA, AGAP y mA) modificados
Laconcentraciónfmal de antibióticos- AAA sinampicilina- AAA con0,005gIl deampicilina
- mA sin ampicilina
- mA con0,01 gIl de ampicilina- AGAP sin penicilinani pimaricina
- AGAP sin penicilina- mA con 25.000IJIA de penicilina
- mA con50.000UT/l de penicilina
- mA con75.000131/1 de penicilina- mA con 100.000UI/l de penicilina- AGAP con 25.000UI/l de penicilina
- AGAP con50.000UI/l de penicilina
- AGAP con 75.000UN de penicilina
en losmediosfue la siguiente:
Para estudiar elposibleefectoinhibidor
- mAcon0,025gIl de rojo fenol- AAA modificado sin rojo fenol y con0,005g/l de ampicilina:
Agarnutritivo:
PeptonaExtractodecarne
Cloruro sódicoAlmidónAmpicilina
delrojo fenol seprepararon los medios:
31 g/l
5 gil3 gIl
5 gil10 g/l0,005 g/l
61

Ir
e
e
y.
r
a-:
Ir
Ir
Ir
e
e
e
Ir
MATERIALYMETODOS
- AAA modificadoconrojo fenol y 0,005g/l deanipiciina:
AAA modificadosin rojo fenol y con0,005 gIl de ampicilina
Suplementadocon 0,025gIl de rojo fenol
Para observar la influenciade la interacciónde la ampicilina con el pH. la xilosa,
lisina libre, argininalibrey el extractode levaduraseprepararonlos siguientesmedios:
-mApI-{7
-AAA
-AAA
-AAA
-AAA
-AAA
-AAA
(0,005gil
(0.005gIl
(0,005gIl(0,005gIl
(0,005gIl
(0,005gIl
de ampiciina)pH 8
de anipiciina)con 3,75g/l dexiosa
de ampicilina)con 3,5 g/l de L-lisinade ampidiina)con2 gIl deL-arginina
deampicilina) con 3,5 gIl deL-argininay 2 g/l deL-lisina
de ampicilina) con 3 gIl de extracto de levadura
Todos
suplementos
los medios se esterilizaronpor calentamientohastaebullición y los
de antibióticosy xilosaseesterilizaronpor filtración.
I~nia: Los recuentosdeA. sobriaCECT 837 en los mediosmodificadosserealizaron, a
partir de un cultivo en TSB+YE, incubadoa280C durante18 h.
Los recuentosobtenidosen TSA+YE se utilizaroncomo referenciaparaexpresar,enforma de porcentaje, la recuperación deA. sobriaCECT 837enlos mediosen estudio.
111.4.10.1.- Antibiograma
Se empleóel medio Mueller-Hintoncompuestopor:
InfusióndecarneÁcidosdecasaminoAlmidónAgar
g/lgígRgil
y discosimpregnadosen los siguientesantibióticos:
Mido nalidfxico (30 ~‘g)Tetmaciclina(30íg)Estreptomicina(1 Ogg>Oxaciina(1 I’g)Neomicina(30 ~íg)Cefoxitina (30 ~sg)Polirnixina 13(300(71)
Lincomicina(2 gg)A¡npicilina (10 i’g)Penicilina6(10 171)Cloranfenicol<30 pg)Sulfisoxazol(300gg)Gentamicina(10 ~tg)Nitrofunmtoína(300pg)
Kanajnicina(30 ig)Cefalotina(30 ~íg)Trimctroprim-Sulfamctoxazol(25 pg)Eritromicina(15 j.tg)Bacitracina(10 UI)Novobiocina (30¡ig)
30017,51,517
62

MATERIAL YMETODOS
Se prepararoncultivos en TSB+YE, que sediluyeron en suerofisiológico hasta
obtener unaconcentraciónaproximadade í0~ ufc/ml. Se añadieron5 ml de esteinóculoa la
superficiedel mediodistribuidoen placas,eliminando,despuésde 5 minutosde reposo, el
líquido sobrante.
Los discosimpregnadosde antibióticos secolocaronsobrela superficie delmedio
inoculadoy, tras 15 minutosde reposo,las placas sc incubarona 280C durante18 horas.
La sensibilidado resistenciade los microrganismosse determinó enfunción del
didmetrodel halode inhibición alrededorde los discos(Tablasproporcionadaspor la firma
suministradora).
111.4.11. - Técnicas de enriquecimiento, aislamiento e identificación deÁeromonasspp. móviles en aguas y alimentos
Seutilizaronlas siguientes muestras:2 de agua potabledorada,3 de agua potableno
doraday 7 de agua nopotable; 5 muestrasde carnede cerdo,5 decordero,5 de vacay 5 de
pollo; 5 muestrasde trucha,5 de pescadilla,5 dejurel, 5 de sardinay 5 de gallo; 5 muestras
dechirla, 5 de gamba,5 de cigalay 5 de mejillón; 5 muestrasde lechecruda,5 de queso
fresco y5 deensaladaspreparadas.
LapresenciadeÁeromonasseinvestigó,encadamuestra, el díade su adquisicióny
cuandoaparecieronlos primeros signos de alteración enesamismamuestramantenidaa
100C.
Ifl.4.1 1.1. - Enriquecimientoy aislamiento
Medio de enriquecimiento:Aguade peptonaalcalina:
Peptona 10 gIlCloruro sódico 10 g/l
pH 8,6El pH sc ajustó con NaOH lN
Medio de aislamiento: mediodeAeronionas(mA)
Lasmuestrassólidasse trituraronen condicionesasépticas.Se tomaron25 g de losalimentossólidosy 25 ml de las muestraslíquidas que sehomogeneizaron, enamboscasos,con 225 ml de aguade peptonaalcalina.El homogeneizadose mantuvo a 280C
63

‘4
Ir
Ir
r
MATERIAL VUfLIODOS
durante24h.
Las diluciones decimalesdel homogeneizadoenriquecidosc sembraronen lasuperficie delmediodeAeromonas(mA), previamentedistribuidoen placas.Despuésde la
incubación(300C-24h),lascoloniascon lascaracterísticasde Aeronwnasse sembraronen
tubos inclinadosde TSA+YE y se mantuvieron a40C, parasu posterioridentificación
bioquímica.Lascepasserevitalizaronantesde realizarlas pruebasbioquímicas.
111.4.11.2.- Identificación bioquímicade géneroy especie
En todas las cepasconsideradascomoAeromonasspp. móviles secomprobósucapacidadparaproducircitocromooxidasay fermentarla glucosaenel agarde Kligler. Enlas cepasoxidasa(+) y Kligler (+) se comprobóquecumplíanlas siguientescaracterísticas:gram(-), ONPG (+), DNasa(+), catalasa(+), crecimientoa370<2(+), crecimiento sinCINa
(+>. resistenciaal agentevibriostático0/129(+), hidrólisisdel almidón(+), fermentación
delD-manitol (+) e 1-inositol (-), ornitina descarboxilasa(-), argininadihidrolasa(+),
producciónde H2S del tiosulfato(-) y ureasa(-).
DeacuerdoconPopoff(1984),el criterio seguidopandiferenciarlas especiesfue:
Pruebasbioquímicas A. hydrophlla A. caviae A. sobria
Producciónde gasde la glucosa + - +
Hidrólisisde la esculina + + -
Fermentaciónde la salicina + + -
VogesProskauer + - +
CrecimientoconCNK + + -
111.4.11.2.1.- Pruebas
III.4.1l.2.l.l. - Oxidasa
Se emplearonbastonesimpregnadosen una solución de oxalatoN~N-dimetil-p-
fenilenediamina,zicido ascórbicoy a-naftol.
El bastónse poneen contactocon unacolonia obtenidaen agartriptona sojacon
extractode levadura(0.6%) de 18-24h; laapariciónde un colorazul púrpuraen elextremo
64

MATERIAL Y METODOS
del bastón indicaque lareacciónespositiva.
111.4.11.2.1.2.- Fermentaciónde glucosa(Agarcon hierrode Kligler)
Agar conhierrode Kligler:
“Lab-Lemco”en polvo 3 gIlExtractode levadura 3 gilPeptona 20 g/lClorurosédico 5 gIlLactosa 10 g/lGlucosa 1 gIlCitrato fénico 0,3 gIlTiosulfatode sodio 0,3 g/lRojo fenol 0,05 gIlAgar 12 gIl
Los tubosesterilizadosse dejaronsolidificar en posición inclinada,formando una
pendienteen lapartesuperiordel agar. A partir de un cultivo revitalizado se realizó la
siembraensuperficiey porpicadura hasta labasedel tubo. Seincubaron a280<2durante48h.
Unapendienteroja y un fondo amarillo, cono singas,indican lafermentaciónde la
glucosa perono de la lactosa delmedio.Cuandotantola pendientecomoel fondo presentan
una reacción ácida(amarillos),con o sin gas,seha producidola fermentaciónde lalactosa.
Cuandono se aprecian cambiosdecolor, los microorganismosno fermentanningunodelos
dosazúcares.
Ill.4.1l.2.l.3. - Gram
Seefectuócon cristal violeta comocoloranteprincipaly safranina como colorantede
contraste.
Tras realizar latinción, las bacteriasgram-negativasseobservabande color rojo-rosa,
mientrasque laafinidad tintorial de la paredde las gram-positivas dabalugara color
púrpura.
III.4.ll.2.1.4. - ONPG
La actividadIS-galactosidasadelas cepassedeterminóempleandodiscosimpregnados
en el compuesto(J-nitrofenoI-~-D-galactopiranósido(ONPG). Para ello,se suspendióuna
65

a-
a-
“4
y
vi
e’
e’
Ir
MATERIAL YMETODOS
colonia en un tubo deensayocon 0.2 ml de una soluciónestéril de CINa al 0,85%,
añadiendo,acontinuación,un discode ONPG.Los tubosse incuban a370C. La lecturase
realizaa los 20 minutosya las 4 horas.La apariciónde un color amarilloen el discoyen el
medio indica la hidrólisisdel O-nitrofenol43-O-galactopiranósido.
111.4.11.2.1.5.- DNasa
Agarpara lapruebade DNasa
Triptosa 20 gIlMido desoxinibonucléico 2 gIl
5 gIlAgar 15 gil
Paradeterminarla actividaddesoxirribonucleasa,se realizó unasiembraen estrías
sobreel agar DNasa. Estas placas se incubaron a280<2 durante24h. A continuación,las
placasseinundaroncon unasoluciónde ácidoclorhídrico0.IN. Los microorganismosquedegradanel ADN del mediodanlugar a laapariciónde halostransparentesalrededorde las
estrías.
111.4.ll.2.l.6. - Catalasa
Serealizóla siembraenTSA+YE. Las placasseincubaron a280<2durante24h. Se
vertió sobrela superficie del cultivo1 ml deperóxidode hidrógenode 10 vol. La pmebase
considerópositivacuando aparecíaunamarcadaefervescenciasobrelascolonias.
I1I.4.1l.2.l.7.- Crecimientoa 370<2
Sesembraron sobreTSA+YE. Despuésde incubarlas placasa 370C durante7 díasse
observólapresenciao ausenciade crecimiento.
llI.4.ll.2.l.8. - Crecimientosin CiNa
Los microorganismos sesembraronsobrecaldonutritivo:
Exrodc carne 3 gil
Peptona 5 g/l
Despuésde incubarlas tubosa 370C durante7 díasse observó latransparenciao
66

MATERIAL YMKrODOS
turbidezdel medio.
HI.4.ll.2.l.9. - Resistencia alagente vibriostático01129
Seemplearondiscosimpregnadoscon 150gg delagentevibriostárico 0/129 (fosfato
2-4-diamino-6-7-diisopropilpteridina).La siembrase realizó enestríassobreplacasde
TSA±YE,depositándoseun discosobre cadaestría.Las placasse incubaron a280<2durante
48h. Lascepas se consideraronresistentescuandoel crecimientoen la estríafueuniforme,
sin observarsehalosde inhibición alrededordel disco.
u1.4.1l.2.1.10.- Hidrólisis del almidón
Para estimar la hidrólisis delalmidónse empleóunamodificación delagaralmidón
ampicilina,descritoporPalumbo(1985),como medio específico para elaislamientode
Aeromonas,a partir de alimentos.Estemedio semodificó, excluyendode sucomposición
la ampicilina, quedandoconstituidopor:
Agarconrojo de fenol 31 g/l
Almidón lOg/l
El mediose sembró e incubó a280<2durante24 h; posteriormente,seañadieron5 ml
de la soluciónde Lugol a cadaplaca.Las cepasquehidrolizaron el almidón dieronlugara
halos amarillos alrededorde las colonias. Cuando las cepas eranamilasanegativas,la
superficiedelagar,incluidos los limites de lascolonias,tomócolor violetaoscuro.
111.4.11.2.1.11.- Fermentaciónde azúcares(D-manitol, i-inositol y salicina)
Se preparóun mediobaseconstituidopor:
Agar infusiónde ternera 25 gIl
Infusióndecarne¡nagrade ternera 500 g/lProteosapeponan03 10 g/lCloruro sádico 5 gIl
Púrpurade bromocresol 0,02 gil
Estemedio sesuplementécon 10 g/l de inositol, manitol o salicina.
(1’

r
‘4
e’
e’
a-
si
vi
a
e’
Ir
e’
y
MATERIAL YMLTODOS
Los tubos sesembrarone incubarona 280<2 durante48h. La fermentaciónse
evidencióporel cambiode colordel indicadorde pH desdepúrpuraa amarillo.
Ill.4.1l.2.l.12. - Descarboxilaciónde laornitina y (o dihidrólisis) de laarginina
Medio basede Moeller paradescarboxilasa:
Extxauode vacuno 5 gIlPeptona 5 gIlGlucosa 0,5 gilPúrpurade bxx>mocresol 0,01 gilRojo creso! 0,005 gilPiridoxal 0.005 gil
Estemediose suplementócon 10 gIl de ornitinao arginina,ajustandoa continuación
elpH a 6. Los caldos inoculados seincubarona 280<2, un máximode 4 días.Enestetiempo
seexaminóla apariciónde color púrpura
Lll.4.11.2.1.13. - Producciónde 1125 del tiosulfatosódico
La producciónde ácido sulfídrico a partirdel tiosulfatose pusode manifiesto por el
ennegrecimientodel agarhierrode Kligler (descritoen el apanado111.4.11.2.1.2)y del agar
lisina hierro(descritoen elapartado1114.14.3.3).
I11.4.1l.2.l.14.- Produciónde ureasa
Laproducciónde ureasa se detectéempleandola basede agarde ureasuplementada,
despuésde suesterilizacióny enfriamientohasta5O~C, con una soluciónde ureaestéril,
obteniendoel siguientemedio:
Peptona 1 gilGlucosa 1 gilClorurosédico 5 giíFosfaopotásico monobésico 2 g/l
20 g/lRojo fenol 0,012 g/lÁgar 15 g/l
El medio serepartió en tubos,quesolidificaron en posicióninclinada,en los quese
inocularon los microoganismos.Los tubos se incubarona 280C durante24h. Los
microorganismosqueno producenureasano modificanelcolor del medio;la detecciónde
68

MATERIAL YMFJTODOS
ureasa implicael cambiode colorde amarilloarojo púrpura.
III.4.ll.2.l.15. - Producciónde gasapartirdeglucosa
Se preparóel medio baseutilizado paradetectarla fermentaciónde azúcares
(III.4.l1.2.l.ll). Este medio se suplementó con10 g/l de glucosay serepartióen tubos,
con campanaDurham.
Los tubossesembrarony se incubaron a280<2durante 48h. Laproducciónde gas se
detectó por la aparición de unaburbujade gasen la campana.
1II.4.11.2.1.l6.- Hidrólisis de la esculina
Seempleó el agar con esculinay bilis.
Extractodecarne 3 gilPeptona 5 g/lBilis de buey 40 g/lEsculina 1 g/lCintoférrico 0.5 g/lAgar 15 g/l
Las placassesembrarony se incubaron a28~Cdurante24h. Laaparicióndecolor
negroalrededor de lascoloniasesconsecuenciade lahidrólisisdelaesculina.
I11.4.1l.2.1.17.- VogesProskauer
Seutilizó el medioRojo de metilo VogesProskauer (MRVP)distribuidoen tuboscon
5 mí:
Peptonadecarne 7 g/íGlucosa 5 gIlTampóndefosfatos 5 gIl
Lostubosseinoculany seincubana30~Cdurante24h.A cadatubo sele añade:- 0,5 ml de una solución dea-naftolal 6 % en etanol
- 0,5 ml de KOH al 40% enaguadestilada
Despuésde agitar los tubos y exponerlos al aire seobservacolor rojo (+) en la
superficiedel medio,como consecuencia de laproducciónde acetilmetilcarbinola partirde
la glucosadel medio.
69

‘4
‘4
a-
a-
a-
e’
e’
si.
a-
vi
e’
MATERIAL YMETODOS
1I1.4.1l.2.l.18.- Crecimientocon cianuro potásico
Se empleóel medio basede <2NK suplementado con unasolución de CNK,
esterilizadapor filtración, que se añadía despuésde la esterilizacióny enfriamiento del
mediohasta500C:
Proteosapeptonanl 3 gIlFosfatodisódico 5.64 gIlFosfatomonopotásico 0,225 g/1Clorurosádico 5 gIlCianuro potásico 0,075 g/l
Despuésde incubarlas tubos, previamente sembradosa 280<2 durante7 días,se
observó latransparenciao turbidezdel medio.
111.4.12. - Técnica para la estimación de la resistenciaal cloro
Se emplearoncomprimidosesterilizantes que contenían:Halazona 5 mg
Excipientee. s. p. 1 comprimido
Seprepararoncultivos en5 ml deTSB+YE, incubándolosa280<2durante24 horas,y
diluyéndolosen suerofisiológicohastala concentración adecuada,
Para cada cepaseprepararon4 matracescon 1 litro de suerofisiológico estéril,
inoculadoscon 1O~ ufeímí, aproximadamente. Tresdeestosmatracessedoraroncon 2.5, 5
y 10 mg de halazona,dandolugar a niveles de 1,2, 2,5 y 5 ppm de cloro activo,
respectivamente,el cuartomatrazseutilizó comocontrol no dorado.Tras30 minutosen
agitación,serealizaronrecuentosdetodoslos matracesenTSA+YE.
111.4.13. - Técnica para la estimación de la resistencia en distintas
condiciones de cultivo (temperatura, cloruro sódico, pH y nitrito sódico)
Seempleó TSB+YE,comomediobase,quesemodificabaporla adición de cloruro
sódico,nitrito sódicoo variacióndel pH, esterilizándoseporfiltración (poro= 0,22 gm). El
medio modificado, de acuerdocon la variable en estudio,se dispenséen microplacas
estériles(250 gí porpocillo), obteniéndosecuatropocilloscon las mismascondicionesde
cultivo; dosde ellossesembraroncon It) ¡fi del inéculoy los otrosdosseutilizaron como
controlesno inoculados.
70

MATERIAL YMETODOS
Lasmicroplacasseincubarondentrode cajasherméticascon aguaen el fondo para
mantenerunahumedad elevada.A lo largo de la incubacióny a intervalos adecuados se
midió la absorbancia a620 nm de cadapocillo en un lectorde microplacas.De acuerdo con
Ferreiray Lund (1987), el crecimientose considerópositivo cuandola mediaaritméticade
la absorbanciade los pocillos inoculadoseraal menos0,025 unidadessuperiora la de los
dos pocillosno inoculados.
Seobtuvo una curva decrecimientoparacadacepa,encadauna delascondicionesde
cultivo estudiadasy, apartirdeella, secalcularonlos siguientes parámetros:
D: Tiempo(horas)que tarda endetectarseel crecimiento
P: Tiempo(horas)necesarioparaalcanzarunatasabacterianade i<Pufcímlg: Tiempode generacióno duplicaciónde la poblaciónbacterianaM: Máxima absorbanciadetectadadurante la incubación(620nm).
La relaciónentreel númerode bacteriasviablespor ml y la densidad óptica se
determinóen un cultivo deA. sobria 13 en TSB+YE incubadoa 280<2, realizandolosrecuentossobreTSA+YE y midiendola absorbanciaa620 nmen el lectorde microplacas
cadados horas(Figura111.5).
y = 4.1525e-2+ 3,5120e-LOxR = 0,998
1,2
1o<.4‘o 0.80
auaSSo 0,4a
1E+9 2E+9 3E+9 4E+9
ufc/ml
Figura 111.5 - Relaciónentrelaabsorbancia(620 nm)y el númerode ufe/mídc un cultivo deA. sobria 13
71

‘e
e’
e’
e’
e’
e,
a
a-
u
e’
MA TERlA ¿ YMETODOS
Temperatura:El efectode la temperaturase determinóapH 7, 0,5%dc CINa y en ausencia
de nitritos; paraello las placasse incubarona4, 10, 28. 37 y 450<2.
Ckruxs~54i~a:Seregistróel crecimientode las cepas a 0.5,2,5, 4,5 y 6.5% deCINa.
manteniendoentodos los casosunatemperaturade 280<2,pH 7 y ausenciade nitritos.
ph: Los parámetrosconstantesdurantela incubaciónfueron la temperatura(28~C), el
clorurosódico (0,5%)y la ausenciade nitritos, comprobandoen estas condiciones elefecto
del pH (3,5; 4,5; 6; 7; 8,5)sobree] crecimiento.
Nixriin..s~dLn: Para estudiar la influencia del nitrito sódico semantuvo constantela
temperatura(28~<2).el clorurosádico(0,5%)y el pH (7) y se adicionaron almedio 0, 63,
125,250, 500 y 1000pg/ml de nitrito sódico.
Posteriormente,serealizarontodaslas combinacionesposiblesentrela temperatura(4, 10, 28 y 370C), laconcentraciónde clorurosádico(0.5 y 2,5%),el pH (6. 7 y 8,5) y la
concentraci6nde nitritos (0, 63, 125, 250, 500y 1000ng/ml). Unicanienteseemplearon
los valoresen los que,previamente,al estudiar elefectoindividual decadaparámetrosobre
el desarrollode lascepasde Aeromono.sse habladetectado crecimiento.
111.4.14. - Técnicas para la determinación de características relacionadas
con la virulencia de Áeromónas spp. móviles
111.4.14.1.- Factores extracelulares
111.4.14.1.1.- Obtenciónde sobrenadantes
III.4.14.1.1.l. - A distintas temperaturasy tiempos
Las cepasdeAeromonasspp.se sembraronen 10 ml de TSB+YE distribuidosen
matracesde 100 ml. La incubaciónserealizóen agitación(150rpm) adiferentestiemposytemperaturas:
- Parael estudiode la actividadhemolítica:70C~336 horas;200C-5()horas;28oC~24
horas;370<218horas
- Paradetectarlas actividadesproteolíticay lipolítica: 280C~48horas
72

.
MATERIAL YMKTODOS
- Paradetectar laactividadcitotóxica: 280C~24horas
El sobrenadante seobtuvo (Figura111.6) porcentrifugación(10.000rpm, 30’~, 4~C)
Seemplearonfiltros con un tamañode porode 0,22 ~tm.recogiendo el filtrado en viales
estériles.El sobrenadantese almacenóa 40<2 y se utilizó en las24 h siguientesa su
obtención.
111.4.14.1.1.2.- Tratadosporcalor
Volúmenesde 1 ml de los sobrenadantes,obtenidosapartir de loscultivosincubados
a 280<2-24h,semantuvieron enun bañotermostítico,a 560Cdurante10 minutos.
111.4.14.1.1.3.- Concentradosy fraccionadospor diafiltración
Cadacepase sembró en22 matraces(de500 mí) queconteníanvolumenesde 100ml
deTSR-YE.La incubación serealizóconagitación (150rpm) a 28~<2 durante24 horas.Los
cultivos se centrifugaron(10.000 rpm, 30m, 40<2) y se filtraron (poro = 0,45 gm),
obteniéndose2 litros de sobrenadantefiltrado.
La diafiltración, tal y como indica la Figura 111.7, se basa en realizar dosultrafiltraciones:en primer lugar, serealizóla ultrafiltración del sobrenadante(2 litros),
obteniéndose1,9 litros de filtrado y 100 ml de retenido,quequedaen la partesuperior delsistema.La segundaultraíiltración serealizó adicionando2 litros de agua destilada,que al
atravesarel cartucho purificanel retenidoy dan lugar a2 litros de filtrado, del quese
aprovechanlos primeros100ml. Finalmente,serecogenlos 100 ml de retenido.
Con cadasobrenadantcse realizarontresdiafiltracionessucesivas, atravésde tres
cartuchoso paquetesde membranaspoliméricasdepoliéstersulfona,para realizarcortesmolecularesa 50, 30 y 8 kDa(Figura111.8.).Con ello seobtuvieroncuatro fracciones,tres
de ellas concentradasunas20 veces:fraccióndePM> 50 kDa, fracciónde PM c 50 y > 30
kDa y fracciónde PM < 30 y > 8 kDa y unacuartasin concentrar:fracción de PM <8 kDaEsta última fracción se concentró. aproximadamente20 veces, por
evaporación-centrifugaciónal vacíoa temperaturainferiora300C.Las fracciones se esterilizaron porfiltración (poro = 0,22 ~tnm).dispensándoseen
alícuotasde 1 ml en vialesestérilesquese conservarona -200C hastasu utilización.
73

*
e’
0
e’
e,
a-
a-
e.
e’
e,
e.
MATERIALYMETODOS
Figura 111.6-Preparacióndel sobrenadante
Mantenimientode cepas:850<2
(15% glicerol)
Revitalización:TSB + 0,6% YE
28~C - 24h.
Condiciones de cultivo:TSB + 0,6% YE
150 rpmTemperatura/tiempovariable
Centrifugación:10000r.p.m.3Oxninutos
40<2
/<
Filtracióndel sobrenadante:
poro= 0.22 gm.
o
Almacenamientodel sobrenadantefiltrado:24 h - 4’>C máximo
o -200C
74
e
,, t/,,, t/
\ ‘. ‘

aO
u,—
‘Oau
O-~5
a
.~
a>‘oo>
auEa>~0CA,
ro
‘a1Eu.1uuEuurgEuaL
u,
‘eauu,O‘ea‘Ou‘e
a‘Oo
viNaaaOua‘Ouuu1~
75

e.
a-
-t
e’
e’
e,
e,
e,
e’
e
e.’
e’
MAYLA’L41. Y 411<1 OLIOS
Figura 111.8 - Esquemadel fraccionamientodelsobrenadante por diafiltración
Sobrenadantefiltrado (2 litros)
$Diafiltración a través del cartucho de 50kDa
Retenido(100 mí)=Fracciónde PM >50kDaConcentradaunas20 veces
Filtrado (2litros) PM <50 kDa
$Diaflítración a través del cartucho de 30 kDa
Retenido(100 mí)=FraccióndePM < SOy >30kDa
Concentradaunas20 veces
Filtrado (2 litros) PM <30kDa
yDiafiltración a travésdel cartucho de 8 kDa
Retenido(100 mí)=
Fracciónde PM .c 30 y > E kDaConcentradaunas20 vecesFiltrado (2 litros) = Fracciónde PM < 8 kDa
Sin concentrar
76

MATERIAL YMKTODOS
111.4.14.1.1.4.- Correspondientesa las distintasfasesde crecimientobacteriano
Cadacepasesembróen 12 matracesde 50 ml quecontenían5 ml de TSB+YE. Losmediosinoculadosse incubarona280<2,enagitación(150rpm) durante22 horas.Cadadoshorasseretiró un matrazpararealizarel recuentoen TSA+YE y obtenerel sobrenadantetaly como se ha indicadoanteriormente(Figura111.6).El sobrenadante sedividió en alícuotasde 1 ml quesemantuvierona -200Chastasuutilización.
111.4.14.1.2.- Pruebas hemolíticas
111.4.14.1.2.1.- En agarsangrede caballoo cordero (Burkey col., 198la)
El medio TSA+YE, despuésde esterilizado,se dejó enfriar hasta450C, paraadicionarleun 5% desangrede caballoo cordero desfibrinadaestéril. El medio serepartióen placasquesesembraroneincubarona200C-50h,280C-24hy 370C-18h.
Lahemolisisu sedetectópor la aparición dehalosopacos alrededor delascolonias,mientrasquela cepas13-hemolíticasdieronlugarahalostransparentes.
III.4.14.1.2.2.- En eritrocitosde conejoo corderoen suspensión(Burke y col., 1981a)
Tampónfosfato salino (PBS)pH 7,2:
CINa 8 gIl
KH2PO4 0,2 g/l
Na2HPO4.12H20 2,9 gIl
<21K 0,2 g/l
Los eritrocitosde corderoo conejo se lavaron tresvecesen PBS estéril pH7,2 antes
de preparar lasuspensiónde eritrocitos al1%en PBSestéril.
Se utilizaron microplacasde 96 pocillos de fondo cóncavo.En cadapocillo se
añadieron100 fl de sobrenadanteo de las correspondientes diluciones doblesdel mismo,
en PBSestéril,y el mismovolumen de la suspensiónde eritrocitos.Cadamicroplacaincluía
dos pocillos con TSB+YE y otros dos con PBS estéril como controles.Las placas se
incubaronlh a370<2 y. a continuación,el mismotiempoa 40<2.
77

e.
e.
e.?
a-
a-
e,
st
MAJERIAI, YMUOLh)S
El úulo hemolitico representabala inversade la dilución másalta queproductael
100% de hemolisis. Lascepasse consideraronhemolíticascuandomostrarontítulos
superioresa 2.
111.4.14.1.2.3.- Eneritrocitosdeconejoincluidos en agar(Ljungh y col., 1981)
Paradetectarel tipo de hemolisis se preparóun gel de agar purificado(10 gIl). Unavez esterilizadose dejó enfriarhasta450<2, añadiéndoleun 5% de eritrocitosde conejo,
previamentelavadosen PBSestéril.Sobre portaobjetosde 7,5 cm x 2,5 cm, situadossobreunamesaniveladora,se
depositaron5 ml del agarcon eritrocitosa 450<2. Se efectuaroncortescircularessobree]agarsolidificadoparaobtenertrespocillos, de 5 mm de diámetro,en cadaportaobjetos.Encada pocillo se depositaron,por duplicado, 50 pJ de las diferentes fraccionesdelsobrenadante.Dospocillos conteníanTSB+YEy otrosdosPBSestéril querepresentabanlos correspondientescontroles.
Los portaobjetossemantuvieronen unacubeta, herméticamentecerrada,en cuyofondo sehabíadepositadounapequeñacantidadde una soluciónde azidade sodioal 1%.Lacubetasemantuvoa370C 1 día,y a4~C 1 hora.Despuésdeestaincubaciónseobservóla presenciade halosde hemolisis (ao lO-
111.4.14.1.3.- Pruebasproteolíticas
111.4.14.1.3.1.- Caseína(Garcíade Femandoy Fox, 1991)
A 1 ml de sobrenadanteseañadió1 ml de azocaseinaal 0,8% en tampón Tris-ClH
(2M, pH 7) y se incubó a370<2durante2 horas,en un bailotermostático.
A continuación, seadicionaron2 ml de ácidotricloroacético(TCA) al 6% y sefiltró
todo elvolumen a travésde papel Whatmann0 42. La absorbanciadel filtrado se midió a
440 nm.
Paracadacepaserealizó un controlnegativomezclando,1 ml desobrenadantey 1 ml
de azocaseinaal 0,8%en tampónTris-CIH (2M, pH 7)y añadiendo,inmediatamente,2 ml
de TCA (6%). Los tubos seincubarona 370<2 durante2 horasy su contenido se pasóa
travésde papel Waíhmann0 42. El filtrado obtenido se empleó paraajustara O la
absorbanciaa 440 nm, leyendo,a continuación,la absorbanciadesarrolladapor las
proteasas presentesen el sobrenadantede la cepacorrespondiente.
78

)
414 FERIAL YMEYODOS
Se considerócomo unidad proteolítica. laactividad enzimática de 1 ml de
sobrenadanteque produceun incrementode 0.1 unidadesde la absorbanciaa 440 nm
(Mateosy col., 1993).
Absorbanciaa440 ninUnidadesproteolíticas= _______________
0,1
111.4.14.1.3.2.- Elastina (Bjorn y col., 1979)
A 1 ml de sobrenadante seadicionaron2 ml de tampónTris-maleato(0,lM, pH 7),
suplementadocon Cl2Ca (0,OOIM) y con 10 mg de Rojo Congo-elastina.Se incubóen un
bañotermostático a37~<2 durante2 horasen agitación(200rpm).
La reaccióncesapor la adiciónde 2 ml de tampónfosfato sódico(O,7M, pH 6). El
Rojo Congo-elastinano hidrolizado pennaneceinsoluble, siendoeliminado por filtración
(Wathniann042). La absorbanciadel filtrado semide a495 nm.
Paracada cepase preparóun blancocon 1 ml de sobrenadante,2 ml de tampón
Tris-maleato(0,IM, pH 7) suplementadocon CI2Ca (0,OO1M) y con 10 mg de Rojo
Congo-elastinay 2 ml de tampónfosfato sódico(0,7M, pH 6), que seincubaa 370<2 en
agitación(200rpm) durante2 horas.Despuésde filtrar a travésde papel Wathmann0 42, se
ajustaa O la absorbanciaa 440 mix midiendo,a continuación, la actividad elastinolítica del
sobrenadante correspondientey calculandolas unidades proteolíticasde acuerdocon lo
descritoen el apartado111.4.14.1.3.1.
111.4.14.1.4.- Pruebaslipolíticas (McKellar.1986
Reactivos:
TampónBES:
BES 10.64 gIl
Taurocolatosódico 3,2 gIl
Cloruro cálcico 0,11 gIl
Soluciónde timerosalal 5% 1 ml/]
El pH seajustaa 7
79

e.
a-
e.
a--
e.
e,
e.,
e
MAJÍRIAL YMU 0h05
Solucióndej3-naftilcaprilato:
Se disuelven0,27 g de 13-naftilcaprilato en 10 ml de
dimetilsulfóxido (DMSO).
Solución de ‘FastBlue’:Sedisuelven0,041g de “FastBlue” en 1 ml de DMSOEsta solución se preparainmediatamenteantes de su
utilización.
A cadatuboy, porduplicado,seañadieron2 ml detampónBES, 20 hl de la solución
de j3-naftil caprilatoy 50 ¡.tl de sobituadante.Seagité e incubó a 40~Cdurante30 minutos
en un bañotermostático.A continuación,seañadieron20 t~ de la soluciónde “Fast Blue’,
manteniendolos tubosa400<2durante5 minutosmás.
Para parar la reacción seañadieron200gl deTCA (0,72N¿). Laextracciónserealizó
con 5 ml deacetatode etilo, agitandofuertementehasta observarcolorblanquecinoen la
fase inferior. La separación defasessellevó a cabo centrifugando a 3000 rpm durante10
minutos. Finalmente,la absorbanciade la fasesuperiorsemidió a 540 nm.
El controlnegativo, utilizadoparaajustaraO el espectrofotómetrocon laabsorbancia
seleccionada,serealizó sustituyendolos 50 ~ilde sobrenadantepor SO ¡tI de aguadestilada.
La cantidad absoluta de13-naftolliberada,comoconsecuenciade la actividad lipolítica
de los sobrenadantes,se extrapolé de un recta patrón, previamenteelaborada, que
relacionabaabsorbancia a540 nm y j.tg de13-naftol presentes(Figura111.9).
Parahacerla rectapatrón se partióde unasoluciónde 13-naftol 3,7 mM. Los 50 gl de
los sobrenadantesse sustituyeronpor 50 ¡tI de estasolución y sus diluciones, que
representabanla adición de 26,5 p.g, 13.5 ¡tg, 5,3 ¡tg, 2,65 ¡tg y 0,53 ¡tg de 13-naftol en
cantidades absolutas.El procesofue idénticoa] realizadoconlos sobrenadantes.
y -7,3004e-3+ 2.9631e~2x It = 0.999(pCOXUOI)
E O,fl
o0,6 Figura 111.9 - Relaciónente los pg de ~-nafloI
Itliberados, comoconsecuenciade la actividad lipolítica.
.5 0,4 y la absorbanciaa 540¡un.
.. 0.2
•< 0,0•300 10 20
pg ¡3-naftol
80

MATERIAL YMIETODOS
111.4.14.1.5.- Pruebasde roxi.gjd~I~hrecultivos celulares
Todaslas incubacionesdelos cultivoscelularesserealizarona370<2con 5% deCO2 y
humedadrelativaalta. Despuésde utilizar los sobrenadanteen los ensayoscitotóxicosse
comprobó suesterilidadincubándolos1 semanaa 280C.
111.4.14.1.5.1.- Células Vero (Jenssen,1984)
Reactivos:
Solución salinade Hanksin Ca~ni Mg~ con HEPES(HBSS 1 HEPES):
- Solución salinade Hanksin Ca~ni Mg~ (HBSSI 100 ml
- Soluciónde Penicilina-Estreptomicina (10000UI y 10 mg/mi, resp.) 2 ml
- Tampón HEPES 1M 2m1
Soluciónsalina de Hank (HflSS~):
CaCI2.2H20 185 mg!! NaCI 8000 mg/l
XCI 400 mg/l NaHCO3 350 mg/1KH2PO4 60 mg/I Na2HPO4 48 mg/1MgCl2.611~O 100 mg!! D-Glucosa 1 mg/lMgSO4.7H20 100 mg/l Rojo fenol 0,01 mg/l
Soluciónsalinade Hanksin Ca~~ ni Mg~~ (14H55-):
KCl 400 mg/I NaHCO3 350 mg/1KH2PO4 60 mg!! O-Glucosa 1 mgIlNa2HPO4 48 mg/l Rojo fenol 0,01 mg/1NaCí 8000 mg!!
Tripsina-EDTAensoluciónsalina:
Tripsina 0,5 gIllIBIA 0,2 gIl
Siembrade célulasVero (Figura111.10)
Se inocularon5x105 célulasen frascosde 25 ml con 5 ml de medio completo,
incubándolosdurante24 horas,con el fin dequelas célulasseadhierana la pareddel frasco
y comiencea formarse la monocapa.Este hechopermite cambiar el medio con facilidad
mediante elsimplevaciadodelos frascos.

*
e.
a-
u-
u
e
st
e
e
e
e
e
e
MATERIAL Y AWTODUS
Figura III. 10 - Esquemadel ensayocitotóxico sobre células VeroJenssen (1984)
5x105 célulasVero 5 ml demedio fresco
24 b
1 ml sobrenadante+4 ml medio fresco
< medio usado
lSh
4 ml HHSS+
4 ml HBSS+
5 ml demedio fresco
24h
4 ml HiBSS- 1 HEPES
sobrenadante+ medio usado
- .- a- ‘- .-~ — 10 mmHiHSS- 1 HEPES
1 ml Tripsina/ FDTA
5 mm
5 ml HBSS+ —~
—~ Tripsina 1 EDTA
loo
5 mi de medio fresco
7 días Q r~ Tinción Giemsa —~
82
medio usado
o

MATERIAL YMETODOS
Tratamientode las células
El medio utilizado se eliminó de los frascos, sustituyéndolopor medio completo
fresco (4 ml 1 frasco) y 1 ml de cadasobrenadanteo de susdilucionesrealizadaspor
duplicadoen mediofresco.El control positivose obtuvoañadiendo1 ml de una soluciónde
etilmetanosulfonato(1,515 mg! mí). Seeligió estaconcentración (equivalentealapresencia
dc 303 ¡íg deetilmetanosulfonatopor ml de medio en contactocon las células) porque,
previamente,se había determinadoque dabalugara un 50% de supervivenciaal actuar
sobre células Vero (Figura111.11).Comocontrolnegativoseutilizó un frascoquecontenía
únicamente5 ml de medio de cultivo fresco.También se realizóun control conTSB±YE.
Los frascosse incubaron durante18 horasy se observó elefectode las distintasdiluciones
del sobrenadantesobrela morfología delas células de la monocapa. El medio con el
sobrenadante se eliminóde los frascos lavandolas célulasdosvecescon 4 ml de la solución
salinade Hank (HBSS+). Después seañadía,de nuevo,medio completo fresco(5 mí!
frasco) antes deincubarlos frascosotras24 horas.
‘4
sioo4>u
80u4>~ 601-~> 40osu,~, 20
~ omo ~oo 5oo
Figura 11.11 - Disminución de lasupervivenciade lascélulasVero enfuncióndel aumentoenlaconcentracióndeebhnetanosulfonato
Supervivenciade las células
A partir de los frascoscorrespondientesa las diluciones delsobrenadanteen los que
schabían observado célulasmuertas,lesionadase intactas seefectuaron recuentosde las
célulasviables.
300 400
gg/mnl de etilmetanosulfonato
120 y = 122,11 - O,23775x R= 0,982(pc 0,05)
813

a-
e
e
e
e
5<
e
e.
a
e
e
e
e
e
MA iItRIA 1. YMICIOI)OS
Parael lo, se climi rió cl mcdjo dc los ¡ascosy seañadieron,a continuación , a cada
ti no, 4 ml dc la sol tic ion sal¡mx dc ¡-lank, sin Ca+±ni Mg+±y con II EPES (HB SS- ¡
HEPES), ¡ncuh-andolos durante It) minulos. Este tampón se retiró de los fi-ascos y se
adicionó tripsina-EIYIA(3 mi/frasco), ineubándose duranteSminutos. A conUnuación,se
eliminó la tíipsinay se añadieron,5 ml de tampónsalinode Hank.Trasdespegarlas células
de los frascos se realizaronrecuentos, diluyendola suspensióncelularen la soluciónsalina
de Hank (HBSSfl. Las diluciones seleccionadassedepositaban,por triplicado,en el centro
de placas(6cm dediámetro),dispersando1(X) ~dde la dilución de célulasen 5 ml de medio
completo.Las placas seincubarondurante?días.
Tras eliminarel mediode las placasse realizó la tinción conGiemsade las células
viablesque sehabíandesarrollado hastaformar coloniasvisiblesa simple vista (Figura
111.12).Para ello,se cubrenlas placascon 5 ml de metanol (100%)durante20 minutos.A
continuación, se elimina el metanol y se añaden5 ml de coloranteGicmsaal 10%.
Transcurridos20 minutosselavan las placascon aguadestiladay secuentanlas colonias
celularesviables quehan tomadocolor violeta.
Figura 111.12 -
Coloniasde célulasVero,viables,tcñi(Iascon(iiiernsa
84

MATERIAL YMEZDDOS
Paraobtenerel porcentajede supervivenciacorrespondientea cadadilución del
sobrenadante,se compararon elnúmerode célulasviablesdetectadasen los frascos,
tratadoscon las dilucionesdel sobrenadante,y el númerode célulascontadasen el frasco
sin trataro control negativo,queequivalíaa un 100% de supervivencia.De estemodo,para
cadasobrenadante, seelaboréunamcta (Figura111.13)querelacionabala mediaaritmética
de los porcentajesde supervivenciacelular, obtenidosen seisensayosdistintos,con la
dilución delsobrenadante.A partirde estarectasedeterminéla dilución quedaba lugaral
50%de supervivencia(o 50%demuerte)celular.
y = ~4,6614+ 0,65034x R = 0,993
íoo- (p<0,O1)
‘4 75..u4.
t 50a.
4>~ 25-*
0-20 60 100 140
Inversa de la dilución del sobrenadante
Figura 111.13- Aumento de la supervivenciadelas células Veroen función de la dilución progresivadel sobrenadantede la cepa77
111.4.14.1.5.2.- Célulasde mieloma (Eley y col., 1993)
Reactivos:
Solución salinade HankconHEPES(HBSS~1 HEPES):
- SoluciónsalinadeHank (HBSS~) 100 ni]
- SolucióndePenicilina-Estreptomicina (10000UI y 10 mg/ml. resp.) 2 ml
-TampónHEPESIM 2m1
85

0:
e-
a
a-
5<
5<
ea-
e
MAYIERIAL Y A/Ph 4.>oS
Se emplearonmicroplacasestérilesde 96 pocillos con tapa.En cadapocillo se
añadieron,porduplicado,50 ¡tI del sobrenadantedela cepaproblemadeAeromonasspp. y
sus dilucionesdobles en solución salinade Hank con HEPES (HBSS~¡ HEPES).A
continuación,seadicionaron200 ~tlde medio frescocompleto que contenía unas5 000
células de mieloma ensuspensión.Las microplacasseincubaron18 horas.
El control positivo se realizó añadiendoa los pocillos unos 400 ¡tg de
etilmetanosulfonato porml de suspensióncelular, ya que, previamente,se había
comprobadoqueestaerala concentraciónmás bajade EMS que provocaba la muerte del
100% de las células.El control negativo conteníaúnicamentela suspensión celular.
También serealizaroncontrolesdelTSB+YE y del HBSS~1 HEPES.
Se consideróque existía muertecelularcuandocon el microscopio invertido se
observabancélulas pequeñasy arrugadas, considerándose como título citotóxicola dilución
másalta en la quetodaslas célulasdelpocillo teníanestas características.
El ensayoserepitió tres vecesy se determinó la mediaaritméticade los títulos
obtenidos.
111.414.2.- Propiedadessuperficiales
111.4.14.2.1.- Hemaglutinaciónconeritrocitosdecorderoy conejo(Burkey col., 1984a)
Los eritrocitosse lavarontresveces conPBSestéril,centrifugandoa 1200rpm 10”~ y40<2, para prepararuna suspensión de eritrocitosde corderoo de conejo al5% en PBS
estéril,
Las cepasseinocularonen 5 ml de TSB+YE y se incubarona 280C durante18 h.
Estos cultivos se vertieron sobre placasde TSA+YE, que se dejaron secar a370<2durante
30-45 minutos y seincubarona 280C durante24 horas. Las coloniasdesarrolladas se
suspendieronen 2 ml de PBS estéril. A continuación,se hizo el recuentode las
suspensiones en TSA+YE.Seconsiguieronconcentracionesaproximadasde 1012 ufelmí.
La aglutinaciónde eritrocitosseobservóen unaplacadecristal, situadasobreun foco
de luz, en la quesedepositaban20 ¡tI de la suspensión bacterianay el mismovolumende la
de eritrocitos.En todaslas placasseincluyó un control negativo,sustituyendolos 20 ¡tI de
la suspensiónbacterianapor 20 ¡tI de PBSestéril.
Las cepasfuertementehemaglutinantesdieron lugar a unaaglutinacióncompletae
inmediata.En las cepashemaglutinantes débilesse observóunareacción incompletao no
instantáneay en lascepasno hemaglutinantesno seobservaba agregaciónde eritrocitosen 5
86

MATERIALY METODOS
minutos.
111.4.14.2.2.- Inhibición de la hernaglutinaciónpor0-manosa,t-fucosay o-galactosa
Los azúcares(D-manosa, L-fucosay D-galactosa)se prepararon al 3% en PBSestéril.
Para cada cepahemaglutinantey sobreunamismaplacadecristalsedepositaron:
- 20 ¡tI de la suspensión bacteriana+ 20 ¡tí de lasoluciónde D-manosa,L-fucosao
D-galactosa+ 20 pl de la suspensiónde eritrocitos.
- 20 pl de la suspensión bacteriana+ 20 ¡tI de la suspensión de eritrocitos.
Además,seefectuaroncontrolesdel PBS y de las solucionesde azúcares(20 ¡ti de la
soluciónde azúcar+ 20 gí de ]a suspensióndeeritrocitos).
Se consideró que un azúcar inhibía la hemaglutinacióncuando las cepas
hemaglutinantesno mostraban estapropiedado la hemaglutinaciónfuerte setransformaba
en débil. Este ensayoserealizóporduplicado.
111.4.14.2.3.- Autoaglutinación(Janday col., 1987)
Caldoinfusióncerebrocowzón(BHm):
Infusióndecerebrodetemen 200 gIlInfusióndecorazónde vacuno 250 gIlProteosapeptona 10 gIlGlucosa 2 gIlClorurosédico 5 gIlFosfatosédicodibásico 2,5 gIlAcidop-aminobenzóico 0.05 g/l
Lascepasse sembraronen tuboscon 6 ml decaldoinfusión cerebrocorazón(BHLB)
y se incubarona 300C durante18 horas.
LascepasSP+ (self pelleting’) fueron aquellas en las que se detectóprecipitación
espontáneaduranteel crecimiento,observándoseun botón celularen el fondo del tuboy
escasaturbidezen la partesuperior.Las cepasSP-crecieron de formahomogéneaen todo
el tubo.
Paracuantificarla agregacióncelular duranteel crecimiento,semidió la absorbancia a
540 nm de la partesuperiordel cultivo (A). A continuación,seagitael tubo paravolver a
medir la absorbanciadel cultivo (B). Con estosdatos se calculó el porcentajede
precipitaciónespontánea:
87

r..
“y
3v
5<
e-
a,
MAThRJAL YMEIUDOS
A511>d¿B - A540 deA%SP= XlOO
A540de E
Después,cadacultivo seagitéde nuevoy sedividió en dosfraccionesde 3 ml. Una
de estas fracciones semantuvoa 1000C, en un bailode polietilenglicol,durante1 hora (C),
mientrasla otra permanecía elmismo tiempo,a temperatura ambiente(D). En las cepas
PAB+ (“precipitationaftaboiing”) scobservóprecipitacióncelular(grumos)en elfondo
del tubomantenidoa 1000C. Los tubos a 1000C delas cepasPAB- y los mantenidosa
temperaturaambientepresentaronun aspectohomogéneo.
Para determinar elporcentajedeprecipitaciónde los cultivos a 1000C-lh,semidió la
absorbancia a540nm, de la fase superiorde cadacultivo a 1000Cy a temperaturaambiente,
aplicandolasiguiente fórmula:
As4~~deD- A540deC
%SP= X100A5~deD
El ensayose realizó dosveces.Se consideraroncepasautoaglutinantes(AA+)
aquellas quemostraronun comportamientoSP+y/o PAB+ (Figura111.14).
111.4.14.2.4.- Aglutinacióncon acriflavina (Janday col., 1987)
Se prepararonsuspensiones bacterianasconcentradas,tal y como se describe en el
apanado111.4.14.2.1,y unasoluciónde acriflavinaal 0,4% en PBS estéril.Sobreunaplacade vidrio situada sobreun foco de luz, se mezcló una gota de la
suspensión bacterianay unagotade la soluciónde acriflavina.Seconsideróquelas cepas
eran positivaspara estecaráctercuandoaparecían agregadosbacterianosen los cinco
segundosposterioresa lamezcla.El ensayose realizópor duplicado.
111.4.14.2.5.- Pruebasparaestimarla hidrofobicidadcelulaj
Las cepas scsembraronen TSB+YE y se incubarona 280C durante16-18horas.Los
cultivos se centrifugaron (7000rpm, 10, 40C) y el sedimentoseresuspendióen PES (pH
7). La absorbanciaa 540 nm de la suspensiónbacteriana seajustéa 1 con este mismo
sg

-
MA TERIAL YMUnDOS
Figura 111.14 - Autoaglutinación(3anda
Cultivo BHIB (6 ml.)
u300C-18h.
Agitación y divisiónen dosalicuotasde 3 ml.
u1000C - lh.
y-)PAB+ PAB-
Tem. amb.•h.
cControl
Tem.amb.
+h.
uControl
1000C - lh.
o
o
¿PAB+ PAB-
SP+yPAB+Ii
SP-yPAB+ SP-yPAB-SP+yPAB-
AM- AA+
y col., 1987)
SPi~
nJ
&
sp-
o
u
AA+ AA
89

r
a-
a-
3v
e
“y
“y
.2
a-
a-2
0:
MA/LB ¡AL YMJ~7OVOS
tampón. La hidrofobicidadsuperficialse midió tresvecescon cadauno de los métodos que
sedescribenacontinuación.
11L4.14.2.5.1.- Unión afiltros de nitrocelulosa(FN(S) (Lachicay Zink, 1984)
Sepasaron3 ml de cadasuspensión bacterianaa travésde los filtros de nitrocelulosa.
A continuación,semidió la absorbanciaa 540 mxi delcontrol (suspensiónbacterianasin
filtrar) y delfiltrado, determinándoseel porcentajede unión al filtro:
A5~delcontrol - A5~ del filtrado% Unión al FNC = _______________ XlOO
A540del control
Las cepas se consideraron“fuertementehidrofóbicas”cuandoesteporcentajeera
superior al70%, “medianamentehidrofóbicas” si seencontrabaentreel 35 y el 70% y
“poco hidrofóbicas”si erainferior al 35% (clasificándolascomono hidrofóbicas cuando era
cercanoal 0%).
111.4.14.2.5.2.- Adsorción a hidrocarburos(n-hexadecano,n-octanoy p-xileno)
(Rosenbergy col., 1980;Sweety col., 1987)
La suspensiónbacterianase distribuyó envolunienesde3 ml queseañadían a tubos
con 1 ml de n-hexadecano,ti-octano o p-xyleno. Estaoperaciónse realizóporduplicado.
Como control seutilizó un tubo con3 ml de suspensiónbacteriana.
Los tubos seincubarona 30~Cdurante10 minutos.Se agitaronfuertementedurante
30 segundosy se incubaronde nuevo a 3Q0(S durante30 minutos. Acontinuación,se
recogió la faseinferior acuosa,borboteándolacon aire 1 minuto. Porúltimo, se determinó
la absorbancia a540 mxi de lafaseacuosay del control;estasmedidasseemplearonpara
determinarla hidrofobicidadsuperficialen función de la adsorcióna los hidrocarburos:
A540del control - A540 de la faseacuosa% Adsorciónal hidrocarburo= _______________________________ XlOO
A540del control
Seconsideraroncepas“fuertementehidrofóbicas’ las que mostraron unaadsorción
90

MAITRIAL Y MLI ODOS
superioral 70%. Cuando laadsorciónse encontraba entreel 35 y el 70% seregistraron
como“medianamentehidrofóbicas”.Unaadsorcióninferioral 35%indicó que lasuperficie
bacterianaera “poco hidrofóbica” y ‘no hidrofóbica” cuandolos valoreseran próximos al
0%.
111.4.14.2.6.- Inclusiónde lasmuestraspara la observaciónde las estructurasbacterianas
con elmicroscopioelectrónicode transmisión(Janday col., 1987)
Reactivos:
TampónMilloning pH 7,3 0,lM, parafijación y paralavado:
Solución1 -
Solución2 -
Solución3 -
Solución4 -
NaH2PO4.H20
NaOH
Glucosa
CI2Ca
Solución A- 830 ml de la solución1 + 170 ml de la solución2. Seajusta
el pH a7,3
TampónMifloning (parafijación):
900 ml de la soluciónA + 100ml de la solución3 + 5 ml de la solución4
TampónMifloning pH 7,3 0,IM (paralavado):
TampónMilloning (parafijación) + 68,4 gIl de sacarosa.
MezcladeMullenhavero Epon-Araldita:
Epon812
Araldita (502, 6005 epoxyresma)
Dibutiltalato(plastificador)
DMP-30 (acelerador)
15
55
2-4
1,15-3
ml
ml
ml
22,6
25,2
54
10
g/l
gIl
g/l
g/l
91

r
3v
a
e
a,
e
e
e
a
“y
0:
“y
a
0:
MATERIAL Y MUÚDOS
Soluciónde citratode plomo:
Pb(N03), 1,33
Na3(C6H507).2H-,O 1,76
Na3(C6H5045 l/2H20 2,14Aguadestiladadesmineralizada 30
Antesde utilizarla se le añaden8 ml de NaOH IN,
(poro= 0,22 p.im)
g
g
gml
se centrifugay sefiltra
Técnica(~I9:
1) Obtencióndelas célulasbacterianas:
Cadacepase sembróen 5 ml de TSB+YE y se incubó a280C durante24h. A
continuación,el cultiQo secentrifugó, lavandoel sedimento entampón
Miloning (para lavado). Porúltimo, secentrifugóde nuevo.
2) Fijaciónde las células:
El sedimentose suspendió en2 ml de glutaraldehídoal 3% en tampón
Milloning (para fijación)y semantuvo 1 h a 4~(2. Seguidamente, se centrifugó, se
lavó con tampónMilloning (paralavado)durante20 h a40(2y se volvió acentrifugar.
A continuación,el sedimento se resuspendióen 2 ml de osmioal 1% en tampón
Milloning (parafijación) y semantuvo 1 horaa temperaturaambiente.Porúltimo, se
centrifugó y selavó, tres veces, conagua destilada,10 minutos atemperatura
ambiente.
3) Tinción del sedimento:
Despuésde lavar el sedimentoen agua destilada sele añadieron2 ml de una
soluciónde acetatodeuranilo al 2% en aguadestilada(filtrada justo antes de su
utilización; poro= 0,22 ¡tm) y se mantuvo 1 hora a temperatura ambiente.A
continuación,se lavé tres veces en aguadestilada, 10 minutos atemperatura
ambiente.
* PodaslascentrlttIgaci(M1CSdeLa suspensiénbacterianase wali¡aroii a 20(X) rpm. 10’” y 4W.
92

MATERIAL YMLTODOS
4) Deshidratación del sedimento:
Se realizóa temperaturaambientecon la siguientesecuencia:
- 2 ml deetanol de300 durante10”‘
- 2 ml deetanolde500 durante10”’
- 2 ml de etanolde70~ durante10”’
- 2 ml deetanolde 800durante10”’
-2 ml deetanolde96~ durante10”’
- 2 mIdeetanolde 1000 duranteIOm
mide etanolde 1000 durante10”‘
5) Inclusiónen resma:
Al sedimento,una vez deshidratado, se le añadieron de 2 ml de óxido de
propileno que se mantuvo 10 minutos a temperaturaambiente,antesde su
eliminación.Estaoperaciónseefectuódosveces.Despuésse leañadieron2 ml dela
mezcla(1:1) de óxido de propilenoy epon-araldita, manteniéndolo 24h a40(S• El
sedimento,en pequeñasfracciones,se colocó en moldes que se rellenaron,
posteriormente,con epon-araldita.Una vezsolidificadala resma, a 600C-24h,se
obtuvieronlos cortesde 60-90nm de grosor.
6)Tuición delos eones:
Las rejillas, portandolos cortes,se sumergieronen unasolución deacetatode
uranio al 2% (filtrada justo antesde su utilización; poro= 0,22 ¡tm) durante3
minutosy, posteriormente,en aguadestiladadurante1 minuto. Despuésdesecarlas
con papel, se volvieron a sumergiren la solución decitrato de plomo, durante4
minutos, y en agua destilada,duranteun minuto. Por último, se secarony se
observaronal microscopioelectrónicode transmisióna 80 kilovoltios.
111.4.14.3.- Característicasbioquímicas
111.4.14.3.1.- Voges Proskauer
Descritoen el apartado111.4.11.2.1.17
93

r
e
a,
e
a
e
e
e
u
0:
MAIliRIAI, YMLI ‘IDOS
111.4.14.3.2.- Fermentaciónde la arabinosa
Se realizó como se detallaen el apanado111.4.11.2.1.11,añadiendo10 g/l dc
arabinosaal medio basedescrito.
111,4.14.3.3.- Descarboxilaciónde la usina
Seempleó el agarlisina hierro:
Peptonadecarne 5 g/lExtractode 1ev~lum 3 g/l]X+)-Glucosa 1 gIlL-Lis¡namonoclorhidrato 10 g/lTiosulfatosálico 0,04 g/lCitrato deamónicofénico 0,5 gIlPúrpuradebromocresol 0,02 gIlAgar 12,5 gIl
El medio esterilizado serepartióen tubosquesedejaronenfriaren posicióninclinada;
la siembraserealizó sobrela pendientey en picadura hasta labasede cadatubo. La
incubaciónse realizó a 280C-18h. Los microorganismosque descarboxilanla lisina
producenun viraje del indicadorde pH acolor violeta.
111.4.15. - Análisis estadístico
Serealizaronlas pruebasestadísticasde “t” de Studenty de “Ji” cuadrado, utilizando
el programa“Stat View”, en un ordenador Macintosh.
94

IV. RESULTADOS YDISCUSION

r
it
it
it
o.
e
a
e
*

RESULÍADOS Y DISCUSIQN
IV.]. - Medios de cultivo específicospara el aislamiento de Aero~gjzas
sp»~mxiln
¡Vii. Recuperación de Aeromonasspp. móviles e inhibición de la floranatural de los alimentos en los medios mA, AGAP y AAA
Unavez descartados,parael aislamientodeAeromonasspp. móviles, losmediosdecultivo disefiadosparamicroorganismosentéricosy vibrios marinos,sehanensayadouna
ampliavariedadde mediosespecíficosen todo tipo de muestras,con resultadosescasamente
concluyentes.
La faltade resultados significativosha hechonecesariala comparaciónde los medios
disponibles,con el fin de elegir el másapropiadopara el posterioraislamientode
Aeromonasspp.a partirde alimentos.
Se haestudiadoel comportamientode tresmediosselectivospropuestosparael
aislamientodeAeromonasspp.móviles:mediodeAeromonaso mA (Ryan,1985,datosno
publicados. Disponiblecomercialmenteen Oxoid), agarglutaznatoalmidónpenicilina o
AGAP (Kielwein, 1969)y agar almidónampidiinao ANAL (Palumboy col., 1985a).
La selectividadde estosmediosparaAeromonasspp.móvilesse estiméestudiando
su capacidadde inhibición frente aotrosmicroorganismospresentesde forma naturalen los
alimentosy su capacidadde recuperaciónpara las especiesmóviles deAeromonas,
previamenteinoculadasen estosalimentos.Otra técnicapropuesta para laevaluación
sistemáticade mediosde cultivo esla “técnicaecométrica”,desarrolladaporMossell y col.
(1983), que fue descartadaporsucaráctercualitativo,faltade uniformidaden las siembras
y complejidadde los resultados.
La capacidadde inhibición frentea la flora naturalde los alimentossedeterminé
comparandolos recuentos obtenidos,a partirde distintos alimentos (aguadorada,leche
cruda, carnesde vaca, polloy cerdo, pescadilla,trucha,chirlasy gambas)sobrelos medios
específicosen estudio,con los registrados sobre agartriptonasoja(TSA). Como se observa
en la TablaIVA (VéaseAnexo IVa, página143), la inhibiciónfue superioren mA, quesólo
permitió el crecimientodel 61,64%de la floranatural,mientrasqueen AAA y AGAP se
detectaronel 75,94%y el 75,54%,respectivamente,delnúmerototal de microorganismos
obtenidos sobre agar triptonasoja.Esteefectofue aúnmásintensoen el casoparticularde
la leche cruda, donde se observó queel mediode mA únicamentedetectéel 42,37%de la
flora total, frenteal 90,60%en AAA y AGAP. Posiblemente este hecho se debaa que el
mA incorporaen su formulaciónsalesbiliares,queafectanmuy directamenteal desarrollo
97

‘y
e
e
e
e
*
e
RLSULT4DQS1’ DiSCUSIÓN
de las bacterias lácticas.Los recuentosde microorganismosaerobios totales sobre agar
triptonasojamostraronquela cargamicrobianade los distintos alimentososcilabaentreiO~
y l0~ ufc/ml o g de alimento,exceptoen el aguadoradadondela cargaestimadafue de 1
ufe/mí. Las ampliasvariacionesen las desviacionesestándarposiblemente sc deban a la
heterogeneidadde lasmuestras dealimentos.
La capacidad derecuperaciónparalas especies móvilesdeAeromonas,sedeterminé
inoculandolos alimentoscitadosconA. hydrophila 839,A. hydrophila 398,A. caviae838
y A. sobria 837, procedentesde la ColecciónEspañolade Cultivos Tipo (CECT), y
comparandolos recuentosobtenidosen los distintos mediosespecíficoscon losnivelesde
inoculaciónde cadaalimento, determinadosporespectrofotometría.Los recuentosde los
diferentesalimentos contaminadosconcadaunade estascepas,osciléen tomoa io~ ufe/mí
o g de alimento.Considerandolas cuatrocepasconjuntamente(TablaIV.2. VéaseAnexo
IVa, página144)se observa queel mediomA alcanzaun porcentajede recuperación,para
las cepasde Aeromonasinoculadasen los alimentos,de 94,11%,superiora ACIAP y AAA
(80,14%y 80,01%,respectivamente).Contrariamente, Ribasy col., (1991)no encuentran
diferenciasen el número deAeromo¡w.sspp.aisladas,apartir de distintos tipos de aguas,
utilizandovariosmedios:Acromonasmediumde Rippeyy Cabelíl,(1979),agaralmidónampicilinade Palumboy col. (1985a), agardextrinaampicilinade Havelaary col., (1987)y
agaralmidón glutamato arnpicilinapenidiina-IOCde Huguety Ribas, (1991).Sin embargo,
Fricker y Tompseu,(1989), comparan los resultadosobtenidos al aislar estos
microorganismosen agar MacConkey, agar sangreampicilina y medio de Aeromonas
(Difco), registrandolos mejoresresultadoscon esteúltimo. Knockel (1989b) destacael
mejorcomportamientodel agaralmidónampicilina, frente al agar dextrinafucsina sulfito.
agarsangreampicilina (lOmgJl.) y agarsangreanipicilina (30m8i1.). Sterny col. (1987)
empleanagaralmidón anipicilina, agarMacConkey ampicilina,agarinfusión cerebro
corazón,agar infusión cerebrocorazón sangreampicilina y agar cefsulodín irgasán
novobiocina,consiguiendoun mayor número deaislamientoscon el agar almidón
ampicilina.Respectoal agaralmidónampicilina y al agar dextrina ampicilina, Varnamy
Evans (1991) y Stelma(1989)advienensobre el posiblecrecimientode Vibrios spp.
amilolíticos. principalmente,en muestrasde mariscoy pescado.Varnamy Evans (1991)
tambiénseñalanque el medio de Aeromonas(Difco) esuno de los másempleadosen la
industria alimentaria, aunquetiene la desventajade permitir el crecimientode otros
microorganismosquedan lugara coloniasde similar morfología alas de Aeromonasspp.
móviles. Un estudio reciente (Holmes y Sartory. 1993) ha confirmado el buen
comportamientodel mediodeAeromonasde Ryan (mA) paraaislarestosmicroorganismos
98

RESULTADOSY DISCUSÍUN
a partirde aguas,al observarque presentamayorsensibilidady especificidadqueel agar
xilosaampicilina y el mediode Aeromonas(Difeo). Contrariamente,Bemagozziy col.
(1994)han selialadoque el medio de Aerotnonasessuperiora los mediosde Ryan y de
Rimler-Shotts. Porotro lado, Gobaty Jemmi (1995),comparanel comportamientode 7
medios selectivosy 2 mediosde enriquecimientoparael aislamientoen alimentos,
obteniendolos mejoresresultadoscon aguade peptonaalcalina,agarsangre ampicilina(30
gg) y agarsales biliares irgasán verdebrillante, recomendandoel empleode más deun
medio sólido.
El comportamientosuperiordelmA parala recuperacióndeAeromonasspp.móviles
sedebe, probablemente, a la fuerteinhibición quepresentanlos otrosdosmediossobreA.
sobriaCECT838. Así,mientraslos porcentajesde recuperacióndeA. hydrophilaCECT
839, A. hydrophilaCECT 398 y A. caviaeCECT 838 fueron satisfactoriosy bastantes
similares paralos tres medios,superandoen todos los casos el 90% (Tablas IV.3,IV.4 y
IIV.5. VéaseAnexoIVa, páginas145 a 147),conA. sobria CECT837 (TablaIV.6. Véase
Anexo ¡Va,página148) fue mejorla recuperaciónsobremA (95,48%),que sobre los otros
dosmedios(31,61%en AAA y 33,02%enAGAP). Estosresultados,podríanjustificar, elescasonúmerodeaislamientosde A. sobria detectadospor Callister y Agger (1987)yHudsony De Lacy (1991) en alimentosy Figuray col. (1986)en procesos clínicos.Del
mismo modo,Stem y col., (1987), trabajando con muestrasde hecespreviamente
inoculadasconAeromonasobtienenunabajarecuperacióndeA. sobriay A. caviaeen los
mediosquecontienenantibióticos.
En definitiva,el mediodeAeronzonases el que presentamayorpoderde inhibición
frentea la flora naturalde los alimentosy máscapacidadde recuperaciónde lasespecies
móviles de Aeromonas,siendoporello el mediomásselectivode los estudiadosparael
aislamiento deAeromonasspp.móvilesa partirde alimentos.El mediodeAeromonasse
puedeconsiderarel más indicadopara ladetecciónde las especiesmóvilesdeAeromonas
(sobretodo de A. sobria),en el casode muestrasde alimentoscon unacargamicrobiana
elevada.En productoscon una floramoderaday cuandose pretendaaislarA. hydrophila
y/o A. caviaesepodríaemplear el agaralmidónampicilinao el agar glutamato almidón
penicilinaconresultadossatisfactorios.
IV.L2. - Comportamientode A. sobria CECT 837 sobre los medios mA,
AGAP y AAA.
La inhibición del crecimientodeA. sobriaCECT837 detectada sobre AAAy AGAP,
99

a.
RLSULJAD(’SY L)/S((15/QN
en comparacióna su comportamientosobre elmedio niA, hizo necesarioconsiderarla
composiciónde dichos medioscon elfin de investigarlas posibles causasdeestehecho.
El mA contieneampicilina(Smg./L),el AAA también,peroen unaconcentracióndos
veces superior (lOmg./l.) y el AGAP lleva penicilina G sádica(l00.OOOU.I./l.) y
pimaricina(lOmgJl.). Paracomprobarsi estos antibióticoseran la causade la inhibición se
realizóun antibiogrania(técnicade Mueller-Hinton) frentea lascuatro cepasdeAeromonas,
empleandoveinte antibióticos,entrelos que seincluyeronla ampicilinay la penicilina O
sódica. En la Tabla IV.7 (VéaseAnexo IVa, página149)seobserva quelas cuatro cepas
fueron sensiblesal ácidonalidixico, tetracidina,estreptomicina,cloranfenicol. sulfisoxazol,
gentamicina,nitrofurantoina,kanamicinay trimetroprim-sulfametoxazol.En este sentido,
Fainsteiny col. (1982)y Fassy Barnishan (1981)señalanunasusceptibilidadmarcada a la
tetraciclina,cloranfenicoly trimetroprim-sulfometoxazol, mientrasque Rahimy col. (1984)
describencepasresistentesa estosantibióticos. Por el contrario, las cuatro cepas fueron
resistentesfrente a la oxacilina, lincomicina, bacitracinay novobiocintEn todoslos casos
se observéun grado desensibilidadmedio frente a la polimixina B, mientras que el
comportamientode cada cepafue variablefrente a laneomicina,cefoxitina, ampicilina,
penicilina, cefalotinay eritromicina.A. sobria 837 fue sensiblea la ampidilinay a la
penicilina,antibióticosutilizadosfrecuentementeenmediosselectivosparaelaislamientode
Aeromonasspp. móviles,y que podrían limitar la detección de ciertas cepas sobre estos
medios. Estos resultados estaríande acuerdocon los de Seidiery col. (1980)y Rahim y
col. (1984) quetambiénhanencontradocepassusceptiblesa laampicilina.Contrariamente.
Villuendasy col., (1991) señalan quetodaslascepasclínicassonresistentesa la ampicilina
y Monta y col. (1994) han detectadobeta-lactamasasen las 182 cepas analizadas.A.
hydrophilaCECT839,A. /zydrophilaCECT398 y A. caviaeCECT838 eran resistentes a
ambosantibióticos.
Estosresultadosexplicaríanla inhibición detectadaen el crecimientode A. sobria
CECT837 sobre AAAy AGAP. La falta de inhibición del crecimientodeA. sobriaCECT837, en el mediode Ryan(mA), podríadebersea que laconcentraciónde ampicilinaen este
medio esinferior a la presenteenel agar almidón ampicilina. En la TablaIV .8 (Véase
Anexo IVa, página150)se observaqueel crecimientode esta cepa seinhibió con 10 mg./l.
de ampicilina, tanto en mA comoen AAA, sin embargo,cuandola concentraciónde
ampicilinaerade 5 mg.f1. en ambos medios,sólo sedetectéinhibición en el AAA. Esta
experienciatambién aclaréque la inhibiciónde A. sobria CECT 837, en agar glutamato
almidón penicilina,se debía a la presenciade l00.OOOU.I./l. de penicilina 6 sádica,
mientrasque la pimaricina,con un papel fundamentalmenteantifúngico,no inhibió el
loo

RESULTADOSY L)ISCUSIQN
crecimientodeA. sobriaCECT 837.
La actividadinhibidorade Smg./l.de ampicilinasobreel crecimientodeA. sobria
CECT 837, desapareceen el medio mA. En basea ello, se pensóque podríadetectarseun
efectosimilar empleandodistintasconcentracionesde penicilina O sódicasobreel citado
medio,sin embargo, se comprobóqueel crecimientodeA. sobriaCECT837 erainhibido,
prácticamenteconigual intensidad,en los mediosmA y AGAP,cuando laconcentraciónde
penicilina G oscilaba entre25.000y 100.OOOU.IIl.(TablaIV.9. Véase AnexoIVa, página
151).La inhibición del crecimientodeA. sobriaCECT 837 sobre elmedio AAA podría
explicarse por unainteracciónsinérgicade la ampicilina con el rojo fenol,utilizado como
indicador del pH en estemedio. El efectoinhibidor del rojo fenol, presenteen elmedio
AAA a unaconcentraciónde0,25mg.¡1.y ausenteen el mA, podríapotenciarlaacciónde la
ampicilina.La TablaIV.l0 (VéaseAnexo IVa, página152)muestra larecuperacióndeA.
sobria CECT837 en mA y AAA con y sin O,25mg./l.de rojo fenol,incluyendoenambos
casosSmgil. de ampicilina.De acuerdocon estosresultados, el rojo fenol nomodificala
actividad inhibidorade laampidiina(concentraciónde 5 mgJl.), sobre elcrecimientodeA.
sobriaCEO?837,en los medios AAAy mA.
Ladiferenciade pH (pH 8 en mA y 7 en AAA) también seconsiderócomoposible
causa deldistinto comportamientode la ampidiinaen estosdosmedios.En laTablaIV. 11
(Véase AnexoIVa, página153)seobservanlos resultadosobtenidosal modificarel pH, en
presenciadeSmg./l. de ampidiinaen ambosmedios.Seconcluyequela inhibiciónsobreel
crecimientodeA. sobriaCECT837,detectada únicamenteen el agaralmidónampicilina,es
independientedelpH, al menosconsideradoaisladamente.
También se controlaronotros componentes,presentesexclusivamenteen elmedio
mA, porsu posiblecapacidadpara protegero potenciar el crecimientodeA. sobriaCECT837, en presenciade Smg./l.deanipiciina.Paraello, comparamos larecuperacióndeeste
microorganismoen los mediosmA y AAA,con Smg. de ampicilina porlitro y adicionados
con xilosa (TablaIV.12. Véase Anexo IVa,página154), usinay arginina libres (Tabla
IV.13. VéaseAnexo IVa. página155) y extractode levadura (TablaIV.14. VéaseAnexo
IVa, página156), frentea los mediosoriginalesmA y al AAA, modificadosúnicamenteen
su contenidoen ampicilina (Smg.Il.). En ningún caso sepudo demostrarque lafalta de
inhibición de la ampicilina(Smg./l.) sobreA. sobriaCECT 837,al emplearmA sedebieraa
la presenciade las sustanciascontrastadas.
Posiblemente, algunoo algunosde los numerosos componentes presentesen el medio
mA, actuandode forma individualo asociada,acomplejeno inactivena la ampicilina.
101

“y
RL SU/JA J)< )S Y DIS CUS/QN
cuando su concentración esde Smg./l.,provocandola pérdidade su efectoinhibidor sobre
el crecimiento deA. sobriaCECT837. En cualquiercaso,unaconcentraciónde ampicilina
de lOmg./I. en dichomedio provocauna notoria inhibición sobreel crecimientode esta
cepa,equiparablea ladetectadaen los mediosAAA y AGAP, siendopor ello aconsejable
no modificar la concentraciónde arnpicilinaen el medio original de Ryan(mA), con elfin
dedetectaraquellascepasde Aeromonasspp. móvilesque puedansersensibles a este
antibiótico.
¡Vi. - Aislamiento e identificación de Aeromonasspp. móviles
La incidenciadeAeromonasspp. móviles en alimentos esrelativamentepoco
conocida, aunque sesabeque soncontaminanteshabitualesde los mismosy queparticipan
en laalteraciónde los alimentosrefrigerados(Buchanany Palumbo,1985;Palumboy col.,
1985a). La asociaciónentre las especiesmóviles de Aeromonase infecciones
gastrointestinalesen el hombre(Freij, 1987)justificael interésactualporconocerel papel
quepuedanjugaren los alimentos.
El aislamientode estosmicroorganismosserealizósobreun total de 92 muestras de
diferentesalimentosy aguas, previoenriquecimientoselectivoen aguade peptona alcalina
(pH 8,6), ya que dalugara unamayoreficaciaen la detecciónde especiesmóviles de
Aeromonasen muestras clínicasy medioambientales,sobretodo, cuandoen ellasno existe
un número elevadode microorganismos(Millership y Chattopadhyay,1984).
Deacuerdo con los resultadosobtenidoscon anterioridad,al comparar la eficaciade
tres mediosespecíficospara elaislamientode Aeromonasspp.móviles de alimentos, el
mediode Ryano mA (Oxoid) se seleccionéparainvestigarla presenciadeAeromonasspp.
en las muestras dealimentos.
IV.2.l. - Pruebas bioquímicas seleccionadaspara su identificación y
diferenciación
La identificaciónde Aeronionasspp. móvilessebaséen unaamplia gamade pruebas
bioquímicas(oxidasa,Kligler, gram, ONPO, DNasa,catalasa,crecimiento a 370C,
crecimientosin CINa, hidrólisis de almidón, fermentaciónde D-manitol e ¡-inositol,
descarboxilaciónde la ornitina.dihidrólisis dela arginina,producciónde H2S del tiosulfato
102

RESULJADOS Y DISCUSIÓN
y ureasa),de acuerdocon Popoff (1984>.Stelma (1989)y Varnam y Evans (1991). Las
cepasque no fermentaron el0-manitol,descarboxilaron la ornitinao no hidrolizaronla
argininano seexcluyeron,yaque Kampfery Altwegg (1992)señalan queA. schubertiino
fermentael D-manitol y Hickman-Brennery col. (1987) que A. veronhi es ornitina
descarboxilasapositivay arginina dihidrolasanegativa.Hudsony De Lacy (1991)y Córreay col. (1993)utilizan, también,un esquemade identificaciónsimilar.
Otrossistemas menoslaboriososy másrápidos hansidoutilizadosporotrosautores.Así, Overmany col. (1985), comparanlos sistemasAPI 20E, API RE y API NFT,obteniendolos mejoresresultadoscon el API 20E, al igual que Abeytay col. (1986)y
Stelmay col. (1986).En estesentido,Figuray Guglielmetti(1987)consideranadecuadostantoel sistemaAPI 20E como el API2ONE. Sin embargo, Toranzoy col. (1986)afirman
que la utilidad del sistemaAPI 20E esrelativa,siendo necesarias pruebasbioquímicas
complementarias comofermentaciónde laarabinosa,gelatinasa,Voges Proskauery
utilización del citrato. Coincidiendoconestosautores,Georgey col. (1986)empleanel
sistema API 20E complementadocon gramn, oxidasa,fermentaciónde la glucosayresistenciaal agente vibriostático 0/129, mientras que Kirovy col. (1990)agregana esta
pautade identificaciónla pruebade la movilidady la reacciónen el mediode Kaper(Kaper
y col., 1979). Krovacek y col. (1992a)afíadenestasmismaspruebascomplementarias,exceptuandola reacciónen el mediode Kaper.al sistema API2ONE.Recientemente,Ogdeny col. (1994),al compararla eficacia delos sistemasAPI 20E, API 20NEy Microbact24E,
consiguenlos mejoresresultadoscon los dos últimoscitados.Otro método rápidode
identificacióny clasificación enespecies,propuestoporKtihn y col. (1992),esel sistema
PhP(Pheneplate),basado enel seguimiento espectrofotométricode la cinética de 48pruebasbioquímicas realizadasen placasdepoliestirenode96 pocillos. Hastael momento,parece que estossistemasno son lo suficientementefiablesparautilizarlos, de forma
exclusiva,en la identificación delas especiesmóviles deAeromonas.En distintalínea,sehanpropuestootrossistemasde identificaciónrápidosbasadosenel análisisde lasecuencia
deARNr lÓS (Dorsch,y col.. 1994)yen la detección inmunoenzimáticadeA. hydrophita
serotipo0:11 (Merino y col., 1993).
Janda (1991) señala que la diferenciación bioquímicade los grupos dehibridaciónes
aún confusay requiereun número elevadode pruebas bioquímicasqueconllevenun claro
riesgode identificaciones erróneas, proponiendo comoalternativaa los métodosgenéticos,
la diferenciaciónfenotípicaclásicadeA. hydrophita,A. sobriay A. caviae.
Enfocandoel trabajo desdeun punto de vistahigiosanitarioy no taxonómico,la
103

‘y
t
u
u
RASLIIAI)QS Y DISULIS/ON
clasificaciónde lasespeciesmóvilesde Aeromonasserealizóde acuerdocon el criterio de
Popoff (1984),diferenciandoA. hydrophila,A. caviaeyA.sobriaen basealaproducción
de gasde la glucosa,hidrólisisde la esculina,fermentaciónde la salicina, VogesProskaucr
y crecimientocon KCN. Lascepasquebioquimicamenteno encajabanen ninguna dcesas
tresespecies,se incluyeronen el grupode Aerornonasspp. móvilesno clasificadas.El
esquemadeclasificaciónelegidoessimilar al utilizadoporStemy col. (1987),Kirov y col.(19%),Hudsony DeLacy (1991)y Pofféy OpdeBeeck(1991). Porotro lado,VarnamyEvans(1991) señalanque lossistemasrápidosAPI 2ONE, ATB 32Ey API 20E no sonútiles, por sí solos,para la diferenciaciónde estastresespecies.Figura y Guglielmetti
(1987)proponenel factor ‘CAMP-like” pataestefin. Okrendy col. (1987) sebasan
exclusivamente en laproducciónde gasde glucosay la hidrólisisde laesculina.Namdariy
Bottone (1989),tambiénutilizan un protocolosimplificadopara definirestastresespecies,
añadiendoel denominado“fenómenosuicida” a lasdospruebas citadas.
IV.2.2. Influencia del almacenamientode los alimentosa baja temperatura
en su aislamiento
En la Tabla IV.l5 (VéaseAnexo IVa, página157) se observala detecciónde
Aeromonasspp.en el 40,21%de las muestrasde alimentosanalizadas el día ceroo día de
su adquisiciónen el mercado,cifra queaumentóhastael 53,26%(Tabla IV.16. Véase
Anexo IVa, página 158) tras el almacenamientode las muestrasa 100C, cuandose
apreciabansignoslevesde alteraciónen el alimento.El aumentodel númerodemuestras
positivas, trasla conservaciónde los alimentosa bajastemperaturas, coincidecon losresultadosobtenidosporPalumboy col. (1985b),Callistery Agger (1987)y Hudsony
Avery (1994)y podríadeberse,no sólo al efectoselectivode lasbajastemperaturas,sinotambiénal aumentogeneralde lacargamicrobianadelos alimentosen funcióndel tiempode
almacenamiento.
Es importantedestacarque,con ciertafrecuencia,se aislémásde unacepao especie
distintaa partir de unamismamuestrade alimento.
IV.2.3. - Incidenciaen aguas y alimentos
IV.2.3.1. - Aguas
En el aguade bebida,tantodoradacomono tratada,no sedetectaronAcromonasspp.
mt

RESULTADOSY DISCUSION
móviles.Moyer (1987) adviertequeeJ aguano tratadaes un factor deriesgoimportanteen
relacióncon este grupo de microorganismos. La ausenciade Aeromonasspp.en el agua
doradaha sido justificaday demostradaporCattabiani(1986)y van der Kooij y Hijnen
(1988). Estos autoresno admitenqueAeromonassea resistenteal cloro, apesarde los
numerosos aislamientos efectuadosen aguasdoradas,(Hazeny col., 1978; Burke y col.,
1984c;Schubert,1991; Krovaceky col., 1992a),así comoen agua sabida deestuario
(Kapery col., 1981)e, incluso, enaguaembotellada(Síadey col., 1986).
El agua nopotablepresentéunaincidenciaelevadadeAeromonasspp.móviles (Tabla
IV.15. VéaseAnexo IVa, página157), que seaislaronen el 71,4%de las muestras,
resultadosque coincidencon los obtenidos por Huguety Ribas (1991)y Poffé y Op de
Beek (1991),confirmándoseasí la ampliadistribuciónde estegrupobacteriano enmedios
acuáticosy terrestres,registradapor Austin y Austin (1985)y Abeytay Wekell (1988).
Burke y col. (1984<,Krovaceky col. (1989)y Knóckely Jeppesen(1990)señalan
al aguade bebidacomoposible fuentede infección.La elevadafrecuenciadeaislamientos
en aguasde consumoha permitido que este grupode microorganismosseincluyaentrelos
tradicionalesindicadores higiénicosutilizadosparacontrastarla calidaddel agua(Rippeyy
Cabelil, 1980; Clark y col., 1982; Krovaceky col., 1989).Contrariamente,Nishikaway
Kishi (1988)consideranque las carnes,pescadosy vegetales,puedenserun vehículode
transmisiónmássignificativo queel agua.
IV.2.3.2. - Carnes
En catorcede lasveinte muestrasde carnede poíío, cerdo,corderoy ternera
analizadasseaislaronAeromonasspp. móviles (TablaIV.15. VéaseAnexo IVa, página
157),alcanzandoeste producto el segundopuestoen incidencia,con un 60%,despuésdel
aguano potable. Estosresultadoscoinciden con la marcadafrecuenciade aislamientosen
carnes, señaladaporFricker y Tompsett(1989), Majeedy col. (1989a,b)y Krovaceky col.
(1992b).La carnede pollo fue la quepresentóunamayorincidencia(100%).
Stemy col. (1987)informande la escasa detección deAeromonasspp.en lasheces
de los animales de abasto,lo que induce apensarque la elevadacontaminaciónde la carne
no se debe,únicamente,a contaminaciónfecal a nivel de matadero,sino tambiéna una
deficiente manipulacióny a la ampliadistribucióndeestosmicroorganismosen el entorno.
105

n
u
e’
es
u
u
RLS¿JLlADOS 1 L)ISCL’S/ÓN
IV.2.3.3. - Pescadosy mariscos
Hudsony De Lacy(1991)aíslanespeciesmóviles de Aerornonasen el 34% de las
muestrasde pescado que analizany Abeytay Wekell (1988)en el 70%de las de mansco.
En nuestro trabajo,la incidenciaen lasmuestrasutilizadasfue del 36% parael pescadoy45% parael marisco(TablaIV.15. VéaseAnexo IVa. página 157), llegandoal 80% en
algunaespeciede pescado (jurel)y al 60%en chirlasy mejillones.La importanciade estos
datos seincrementaal considerarque son productossusceptiblesdeconsumirsecrudoso
pocococinadosy quese hanestablecidorelacionessignificativasentre cepasprocedentesde
alimentosde origen marino(sobretodo moluscosbivalvos)y brotesdegastroenteritisen el
hombre(Altwegg y col., 1991).
IV.2.3.4. - Lechecruda y quesofresco
La presenciadel citado grupo seevidencióen el 20% de las muestraslácteas
analizadas (TablaIV.15. VéaseAnexo IVa, página157). Kielwein y col. (1969)y Palumboy col. (1985a) aíslanAeromonasspp.en lechecrudt Santosy col. (1991) y Córreay col.(1993)encuentranunaincidenciacercanaal 30%en quesofresco.La detecciónen quesofrescoelaboradocon leche pasterizada, posiblementeseadebidaa la recontaminación
posterioral tratamientotérmico,dada latermolabilidaddemostrada,al menosparaA.
hydrophfla,porPalumboy col. (1987)y Condóny col. (1992).
IV.2.3.5. - Ensaladas preparadas
En ninguna de las ensaladaspreparadasse detectaronespeciesmóviles de
Aeromonas.Lasensaladasestabanelaboradas condistintosvegetales(lechuga,tomate,
cebolla, zanahoria,escarola,col, pepino...)lavadasy listas parael consumo directo, lo cual
explicaría la ausenciade microorganismospatógenoso potencialmentepatógenos.Sólo un
tipo de ensaladallevabaen su composición mayonesa, cuyopH bajopodríaserun factor
limitante para el desarrolloy detecciónde Aeromonas. Fricker y Tompsett (1989)
encuentran una incidenciamuy bajaen este tipode ensaladas, mientrasKrovaceky col.
(1992b)no detectanAeronwnasen vegetales crudos.Contrariamente,Callister y Agger
(1987) informan de elevadastasas deaislamientosen vegetales,sobre todo tras su
almacenamientoen refrigeración,
106

RESLJLL4DOS Y D/SCL’S/ON
IV.2.4. - Distribución en especies de las cepas aisladas en los distintos
alimentos
Coincidiendocon Knóckel y Jeppesen(1990),como se observaen la Tabla IV.17
(VéaseAnexo IVa, página159),A. hydrophilafue la especie aisladaconmayorfrecuencia
(39,50%).Sólo seiscepasfueron identificadascomoA. caviae;en este sentido Krovacek y
col. (1992b)obtienen resultadossimilares al estudiarla incidenciade Aeromonasen
alimentossuizos.El 20,98%delas cepascorrespondíanal perfil bioquímicode A. sobriay
un 32,09%no se pudieron incluiren ningunade estastresespecies.
A. caviaese aislóprincipalmenteen mariscosy no sedetectóencarnesni pescado,A.
hydrophilaen carnesy pescadosy A. sobriaen carnes.En estesentido,Nishikaway Kishi
(1988)observanque mientrasA. hydrophita y A. sobriase aíslanmásfrecuentementeen
carnes,A. caviaeestápresente, sobretodo,en vegetales,pescadosy mariscos.
IV.3. - Resistenciaal cloro de seis cepasdiarreicasy seis alimentariasde
Atromonas spp. móviles
Laresistencia al clorodeestosmicroorganismosse comprobóefectuandorecuentosa
partirde suero fisiológico estéril,contaminadocon 6 cepas diarreicasy 6 alimentariasde
formaindependientey. posteriormente,dorandocon halazonahasta niveles de cloroactivo
de 1,2, 2,5 y 5 ppm.
Los controlesseefectuaroncon suerofisiológico contaminadoy no sometidoa
cloración.Los resultadosde los correspondientesrecuentos se recogen en la Tabla IV.18
(VéaseAnexo IVa, página160).
En los suerosfisiológicos, contaminados conlos mismos inóculos quelos sueros
controlesy, posteriormente.dorados,no sedetectóla presenciade lascepasdeAeromonas
spp. móviles,previamenteinoculadas,poniendode manifiesto lasensibilidadde estos
microorganismosa la cloraciónseñaladaporCattabiani(1986).En este sentido, Burkey
col. (1984c) tambiénindican que elnivel de cloro libre afectasignificativamentea la
presenciade acromonasen elaguay,Van der Kooij (1988)concluyequeel aislamiento de
estosmicroorganismosen aguadoradasedebea la insuficienteconcentraciónde cloroUbre
residualy al tratamientoincorrecto. Porotro lado, Yesim y Aykut (1994) señalanque
nivelesde hasta2,5 ppm de clorono afectana la supervivenciadeA. hydrophila.
Por lo tanto, se puedeconsiderarque estosmicroorganismos sonsensiblesa la
cloracióny quelos problemasen el aguadoradasedebena recontaminacionesposteriores.
107

0<
0<
u
e
0<
e
es
0<
0<
RES CEJADOSY L)/SCtS/OX
Por ello, es fundamentalhacerun seguimientode los nivelesde cloro residualque. por lo
general,debensercomomínimo superioresa 0,5 ppm, aunque estelímite se eleveen
determinadascircunstancias.
IV.4. Influencia de diferentesparámetrosexófenossobre el crecimientode seiscepasdiarreicasy seis alimentariasdc Atromonas spp. móviles
Se emplearonlas siguientescepas. De origen diarreico: A. hydroph¡la 6, A.
hydrop/z¡la 9, A. caviae 16, A. caviae 18, A. sobria 11 y A. sobria 13. De origen
alimentario:A. hydrophila55,A. hydrophila96,A. caviae54,A. caviae97, A. sobria34 yA. sobria92. Seconsiderécrecimientopositivocuandola lecturade absorbanciaen lospocillos inoculadosera 0.025unidadessuperiora la de cuatropocillos equivalentesno
inoculados. Dichaabsorbanciaeraaproximadamente0.080y secorrespondíacon unacarga
de 5.106 ufc/ml. Igualmente,se determinéque unaabsorbanciade 0,394 unidadessignificabaunatasabacterianade 10~ ufeiml.
IV.4.l. - Temperatura
Comoseobservaen la TablaIV.19 (VéaseAnexo IVa, página161),las temperaturasde incubación ensayadas correspondena 4, 10. 28, 37 y 450C. En este trabajo sedetecté
crecimientoa 40C en 7 cepas(2 diarreicasy 5 alimentarias),alcanzando4 de origen
alimentarioel nivel de io9 ufc/ml.
El grupodeAeromonasspp.móvilesha sido considerado de naturalezapsicrótrofa
pornumerososautores(Buchanany Palumbo,1985; Majeedy col., 1989b;Kirov y col.,
1990; Beuchat,1991; Kirov, 1993)e, incluso,con capacidadparaproducirexotoxinas,relacionadascon lavirulencia,atemperaturasde refrigeración(Majeedy col., 1990;Eley y
col., 1993).
Las cuatro cepasalimentariasque alcanzaronla tasa de io~ ufc/ml pueden
considerarse especialmente peligrosas desdeel punto de vistade la SaludPúblicaya que, de
acuerdocon Majeedy col. (1990).esnecesariollegar a esenivel de crecimientoparalaproduccióny detecciónde enterotoxinasy hemolisinas.Por otra parte.esprecisotener
presenteque esta tasa demicroorganismospotencialmentepatógenosrepresentaunadosis
infectivaquesealcanzaríaen unos9 díasa40C.El desarrolloelevadode las cepas aisladasen alimentosrefrigerados,en comparación
con las de origen diarreico,se podríaexplicarpor la mejory másrápida adaptación delas
los

RESULTADOS YDISCUSION
primerasa estastemperaturasbajas. Estehechoya ha sidosefialadopor Knoehel (199<)).Hudson(1992)y Buncicy Ave¡y (1995)queencuentranmenorcrecimientoa 50Cencepasclínicaso aisladasen ambientescálidosqueen las procedentesde alimentosrefrigeradosy
ambientesfríos.No obstante,en ningúncasodebedescartarse elcrecimientode cepasclínicasa 4”C.
Estaconclusiónno sóloestárespaldadapor los resultadosde estetrabajo (2 de 6), sino
tambiénpor los trabajosde Kirov y col. (1990) y Eley y col. (1993) en los que se afirmaque lamayoríade las cepasde origen clínico puedencrecera 50C y quelo hacenrápidamentea 100<2.
Cuandola temperatura deincubaciónseelevéa 100C, crecieronno sólo lascepasalimentariassino tambiénlas clínicasy no seencontrarondiferenciassignificativasentreellas. No obstanteesteincrementode temperaturasupuso que eltiempo de deteccióndecrecimientodisminuyese unascuatrovecesy que,el necesarioparaalcanzarla tasade 1o~ufc/ml, quedasereducidoa 4díasaproximadamente.
Los resultadosde la Tabla IV.19(VéaseAnexo IVa, página161), indicanqueelcrecimientomásefectivodeAeromonasspp. móviles seobtienea 280C, al igualque hanseñaladoPopoff (1984) y Vamam y Evans(1991)quefijaron estatemperaturacomo laóptima.
Cuandola incubaciónserealizóa370<2,unacepadeorigen alimentario nocreció.Enrelaciónconestehecho,Knochel (1990) apuntaqueciertascepasambientalesson incapaces
de crecera370C. Sólounacepade origendiarreicoalcanzóel nivel de i0~ ufc/ml a estatemperatura,lo que podríaexplicarunamenor producciónde factoresrelacionadosconlavirulencia a 370C. Mateosy col. (1993) indican que a 37~C las cepasambientales
disminuyenla producciónde proteasasy hemolisinasaunqueno de citotoxinas,mientrasque, encepasclínicas,sólo esmásbaja la producciónde proteasas,aumentandola decitotoxinasy hemolisinas.La dificultad paracrecerhastatasasmicrobianasimportantesa
37~C,de algunascepasdeAeromonas,podríaseruna de lascausas queinfluyeran en lapresentaciónde diferentesgradosde patogenicidaden el hombre.Así, las cepas concapacidadvirulentasólo causarían gastroenteritisen e] casode coincidir conotrosfactores
que estimularansu crecimientoy facilitaranla producciónde toxinasy enzimasa 370C.En estetrabajono sehadetectadocrecimiento,en ningúncaso,cuandola incubación
seefectuéa450<2. En la bibliografíaconsultada,Rouf y Rigney(1971) registrancepasquesedesarrollaninclusoa 550<2, mientrasqueparaVarnamy Evans(1991) la temperaturamáximade crecimientodeAeromonasspp. móvilessedade 38-41OC y paraPalumboy col.(1985b)de 420C. Estaheterogeneidadderesultados,posiblementese debaa lis diferencias
109

*
0<
0<
0<
0<
0<
0<
RI.SL’LIADO> 131StLSI(LS
de lascondicionesdecultivo y a la propiavariabilidad delas cepasensayadas.
IV.4.2. - Cloruro sódico
Como señalala Tabla IV.20 (Véase Anexo IVa,página 162), todas las cepas
alcanzaron unausabacterianade io~ ufc/ml en presenciade0,5 y 2,5%de CINa, aunque el
crecimientofue másrápidocon 0,5%de CINa. Knochel (1990)observaque enpresencia
de0,5%de CíNa, crecen el100% de las cepas,mientrasque conun 2% dc sal sólo se
desarrollanel 96%.Contrariamente,Paluniboy col. (1985b)no encontrarondiferencias
cuandolas concentracionesde CíNa erandel 0.5, 1.5 y 2.5%,detectandocepas capacesde
crecer hasta con 4.5%.Ennuestrocaso, se registré elcrecimientodetrescepas diarreicasy
tres alimentariasen presenciade 4.5% de CiNa, aunque ninguna alcanzóla tasade ío~
ufc/ml, por lo que estaconcentraciónde sal podría prevenir la actividadpatógenade
Aeromonas.Sin embargo, cuando elmedio de cultivo contenía6.5%deCíNa nosedetecté
crecimientoen ningún caso. En estesentido,Santosy col. (1994)apuntan que mientrasen
presenciade 4.5%de CíNa algunascepascrecen,un 5% de CINa esya suficientepara
inhibir el crecimiento.
IV.4.3. - pH
Como seobservaen la TablaIV.21 (Véase AnexoIVa, página163), tantolas cepasdiaffeicas comolas alimentariasalcanzaron tasassuperioresa ío~uit/ml a pH7, aunquelos
mejoresnivelesde crecimiento seobtuvierona pH8,5. Estehechojustifica la utilización de
aguade peptonaalcalina(pH 8,6)comomedio deenriquecimiento,previoal aislamiento.
Deacuerdoconnuestraexperienciatodaslas cepascrecierona pH 6, alcanzandoel
nivel de i09 ufc/ml cuatro diarreicasy cincoalimentarias.Cuandoel pH bajóa 4,5,sólo se
detectédesarrolloen doscepasprocedentesde alimentos,con tasasde crecimiento
menores.A pH 3,5, ya no se registró proliferaciónbacterianade ningún tipo. En este
sentido,Palumboy col. (1985b) señalan elcrecimientode cepasa pH 4,5 y Karem y col.
(1994) indicanque ciertascepasdeA. hydrophila,en medios ácidos(pH 3.5).sintetizan
proteínasprotectorasdela célulabacterianaparaadaptarsea estascondiciones.
Finalmente,se podríaconcluir que el pH óptimo de Aerotnonasspp. móviles se
encuentraalrededorde 8,5 y que, en condicionesexperimentalesfavorables,estos
microorganismosson bacterias relativamentetolerantesa la acidezo concapacidadpara
adaptarsea un mediode estas características.
110

RESULTADOS Y DISCUSION
IV.4.4. - Nitrito sádico
Como secompruebaen la TablaIV.22 (VéaseAnexo IVa,página164), apartirde
125 ~.tgJmlde nitritos secomenzóadetectarun efectonegativo sobreel crecimientodeAeromonasspp.móviles. Enpresenciade concentracionestanelevadascomo1000 I.Ig/ifllse desarrollaron seiscepasde alimentosy cuatrodiarreicas,aunqueprecisarontiempos de
incubación superiores.Otros autores, comoPalumboy col. (1992), compruebanque la
cepaK144 de A. hydroph¡la escapazde creceren presenciade 200 gg/ml de nitritos.En
consecuencia,sepuedeafirmar que los nitritos, por sí solos,a las concentracionespermitidasen los alimentos,prácticamenteno afectanal desarrollode Aeromonasspp.
móviles.
La influenciade la temperaturadeincubación,cloruro sódico, pHy nitritos sobreel
desarrollode Aeromonasspp. móvilesdependiómásdel origen(diarreicoo alimentario)que de las especies implicadas.No obstante,se puedematizarque las cepasde A.
hydrophilaresultaronserlasmásresistentesala accióndelos nitritos.
IV.4.5. - Efecto combinadode los parámetrosensayados
Los efectosindividualesdelos diversosparámetrosno se utilizanen la conservaci6n
de los alimentos, yaque raravez actúaun solo factor y, en cualquiercaso, laactividad
antunicrobianasemodificaal actuarconjuntamentevariosdeellos.A 4~C y 100C sepotencióel efectoinhibidor delpH, cloruro sódicoy nitrito sódico.
En la Tabla IV.23(VéaseAnexoIVa, páginas165y 166)seobservaqueunpH6 a40<2es
capazde impedir por completoel desarrollodeAeromonasspp.. Palumbo(1988) y
Palumboy Buchanan(1988)hanapuntado que lacombinaci6nde bajastemperaturasy pHsinferioresa 6 controla eficazmenteestos microorganismosen los alimentos.En este sentido,
Aytac y Ozbas (1994) han observado unainhibición completa delcrecimientoen yogura
40<2. Cuando el pH seelexaa 7 espreciso que la concentración decloruro sédicoseadel2,5% y que los nitritos seincrementenhasta 250-500xg/ml paraque la inhibición sea
completa.
De acuerdocon la Tabla IV.24(Véase AnexorVa,páginas167 y 168),a temperaturas
de 100<2 y pH 6 esnecesario adicionar125 ¡xg/ml de nitrito sódicoo bien 62 ¡sg/mlde este
antiniicrobianoy elevara 2.5%la concentraciónde nitrito sódicopara queel crecimientode
estos microorganismosno tenga lugar.El desarrollo de Aeromonasspp. a bajas
111

nr
RESULTADOSY I)ISCUSION
temperaturas esunade las razonesque justifican este trabajo,yaque se ha comprobadosu
presencia en carnesrefrigeradas(Jay,1967;bule y Murphy, 1978;Len, 1987),pescadoy
marisco(Smith y col., 1984; Gorczycay col., 1985;Gram y col., 1987). y especialmente
en platoscocinadosmantenidosen refrigeración(Toule y Murphy, 1978).El empleode
temperaturasde conservación de4oC~lOoC,combinadascon la adición denitrito sódico
(125 ~igIml),cloruro sódico (2.5%)y un gradode acidezmoderado (pH 6)essuficiente
paracontrolarla proliferaciónde estosmicroorganismosen los alimentos.Deacuerdocon
estos resultados,Palumboy col. (1985a),Palumboy Buchanan(1988),Knóchel (1990),
Beuchat (1991)y Palumboy col. (1991, 1992) señalanqueel desarrolloy supervivencia
deAeromonasen alimentos a bajas temperaturas estáinterrelacionadocon otrosfactores
(sal,pH, nitritos, entornoatmosféricoy flora competitiva).
A 40C y 100<2seobservarondiferenciassignificativas(p <0,05)entre cepas diarreicas
y alimentarias.Estos resultadosestánde acuerdocon los trabajosde Knóchel (1990) y
Hudson (1992)en los quesedemuestrala mayor adaptaciónde cepasalimentariaspara
crecer a bajas temperaturas, aligual quesu mayortoleranciaa la acidez,cloruro sódicoy
nitrito sódico.
En la Tabla IV.25 (Véase Anexo IVa, páginas169 y 170) secompruebaque la
temperaturaóptimade crecimiento,en funcióndel tiempode generación,fuede 280Ccomo
admitende formageneral diversosautores aunquey, al igual que observaPalumboy col.
(1991), la fasede latencia.en ocasiones,fue máscortaa 370<2. Las diferenciasentre las
distintascepasfueronescasas a estatemperatura,decanuindose,en todocaso, afavorde las
de origen diarreico, que fueronmásresistentes a laacidez.La influenciade los nitritos a
280<2estuvo fuertemente determinada porel pH, ya quelas cepasalimentarias fueronmás
resistentesa pH7 y las diarreicas cuando esteparámetro descendíaa 6. Estosresultados
podríanseñalarquelos pHs bajosjueganun papelimportanteen el control deAeromonas
spp. móvilesen alimentos, apoyadosporel hechoevidentede que,en todos los casos,el
pH óptimo de crecimientofue de 8,5.
A 370<2, comoseindicaen la Tabla IV.26(VéaseAnexo[Va, páginas171 y 172). las
cepas diarreicas mostraron mayor adaptación, resistiendo mejorlas condiciones
desfavorablesde pH, clorurosódicoy nitrito sódico. De modogeneral,se podríaconcluir
queel origen de las cepas(diarreicasy alimentarias)tiene ciertainfluenciaen laresistencia
frente alos diferentesagentesutilizadosen la conservaciónde los alimentos,de los que la
temperaturaes el factordemayorinterés.
112

RESULTADOS YDISCUSION
IV.5. Factoresde virulencia de Aeromonasspp. móviles
JV.5.1. - Productosexocelulares de cepas diarreicas y alimentarias
IV.5.1.1. - Hemolisinas
El ensayohemolítico, ademásde serpor símismo indicador devirulencia,ha sido
propuestopor Burkey col. (198la) como técnica sencillaparainvestigar, conun 5% de
error, la actividad enterotóxicade las cepasdeAeromonasspp., alconsideraral igual que
Majeedy col. (1989a),que en la granmayoríade lascepas,las actividadeshemolítica,citotóxica y enterotóxicaestánasociadas.En este sentido,Baloday col. (1995) han
propuestoel empleo de la reacciónen cadenade la polimerasaparaagilizarla detección de
cepashemolíticasdeAeronwnasspp.Sin embargo,Okitsuy col. (1985)y Krovaceky col.
(1994) no hanencontradounarelaciónsignificativa entrehemolisisy enterotoxicidad,yTodd y col. (1989) y Notermansy col. (1986) tampocodetectaroncorrelaciónentrecitotoxicidady hemolisis.No obstante,Asaoy col. (1984), Stelmay col. (1986),Bunningy col. (1986) y Burke y col. (1987) han purificadouna p-hemolisina citotóxicay
enterotóxica,apartir delsobrenadantedeun cultivo deA. hydrophila.
IV.5.1.1.1. - Hemólisis desarrolladapor las coloniasbacterianassobre agar sangre
De acuerdocon CaNil (1990) seconsideróa-hemolisiso hemolisisincompleta,cuandosobreagarsangrede caballoo corderoaparecíaun halo opaco alrededorde las
coloniasde aeromonas.Lascepasque originaronun halo transparente,al crecersobreagarsangrede caballoo cordero,seconsideraronf3-hemolíticas.
Comoseobservaen la TablaIV.27 (VéaseAnexo IVa, página173) sedetectóunmayor númerode cepashemolíticascuandoseempleóagarsangrede caballoquecuandose
utilizó agar sangrede cordero.Todaslas cepashemolíticas,sobreagarsangre de cordero,
lo fueron tambiénsobre agar sangrede caballo y un númeroimportantede cepas
a-hemolíticas,sobreagar sangrede cordero,originaronj3-hemolisissobre agarsangrede
caballo. Howardy Buckley (1986)advirtieronquela a-hemolisis, puede ser consecuencia
de la actividad residualde la 13-hemolisina,lo queexplicaríaquealgunascepasp-hemolíticas
sobre sangrede caballose comportencomo a-hemolíticassobresangrede cordero,sin que
ello implique laproducciónde las doshemolisinas(a y 3). Por tanto. sepodríadeducirque
113

tÉ
9
e
e
mv
e
nr
e
ÑLSLiJADOS VISÉ L SIUX
la sangrede caballo esmás inestableque la sangrede corderoo bien mássensiblea las
hemolisinas producidasporAeromonasspp.Dadoqueel agarsangrede caballoresultémás
eficaz para detectarcepas hemolíticasy actividad beta hemolíticasc consideraron,
fundamentalmente,los resultados obtenidosconestemedio.
No seapreciaron diferenciasimportantesen el númerode cepashemolíticaso en eltipo de hemolisis observadosobreagarsangre decaballo,al incubarlas placasa las
distintastemperaturasy tiemposespecificadasen la TablalV.27 (VéaseAnexoIVa, página
173).sin embargo,podría selialarse que al trabajarcon agarsangrede corderoa 370<2,en
las cepasde origen diarreicono se detectéj3-hemolisisy el númerode cepas¡3-hemolíticas
de A. sobriase redujoconsiderablemente.Contrariamente,Ljungh y col. (1981),aislaron
dos hemolisinas: la a-hemolisina,cuyo pico máximo de producción aparece,
aproximadamente,a las 50 horasde incubacióna 200C y no se detecta cuandola
temperatura es superior a300<2,y la p-hemolisina,cuyaproducción óptimaseconsiguea
370<2 en 18-20 horas.Caberesaltar que elensayohemolítico,basadoen laobservaciónde
los halos dehemolisis,que aparecenalrededorde las coloniassobreagarsangre,no
permite,lógicamente,detectarla producciónde a-hemolisinasen cepasproductorasdeambashemolisinas.
Los resultadosde la TablaIV.27 (VéaseAnexoIVa, página1>73) no permitenapreciardiferencias importantesen la actividadhemolíticade las cepasdiarreicas,alimentariaso de
colecciónsobre agarsangrede caballoy cordero,lo que coincide con los resultados
obtenidospor Mateosy col. (1993)al trabajarconcepasde origenacuáticoy humanosobreagarsangrede cordero.
Contrariamentea los resultadosobservados paralasdemásespecies, lascepasdeA.
cav¡ae, tanto diarreicascomoalimentarias,secomportaronpredominantementecomoa-hemolíticas.
IV.5.l.l.2. - Hemolisisdesarrolladaporel sobrenadante completolibre de célulassobre
eritrocitosen suspensión
El título hemolítico asignadoa cadasobrenadantefue la inversadela dilución más alta
queproducíael 100%de hemolisis(Mateosy col.. 1993).
Los resultadosmássatisfactoriosse obtuvieroncon eritrocitos de conejo(Tablas
114

RESULTADOSY DISCUSION
IV.28; IV.29; IV.30 y IV.31. VéaseAnexo IVa, páginas147 a 177). Así, el númerode
cepashemolíticas coneritrocitosde conejofuesiempresuperiory todasla cepasque Usaron
los eritrocitosdc cordero,hemolizarontambiénlos de conejo alcanzando, además,títulos
superiores.En este sentido,Ljungh (1987)ha señaladoquelos eritrocitosde roedores son
losmássensibles,por poseer unaglicoforina en la membranaqueactúa comoreceptorapara las hemolisinasde aeromonas.Todd y col. (1989) encontrarondiferenciasen losresultadosobtenidoscon eritrocitosde conejoy de hamster,siendoestos últimoslos más
sensibleso fácilesde lisar,e indican,al mismotiempo,que laactividad hemolítica dependetambiéndelmedio dondeseobtieneel sobrenadante.
El empleode ambostipos de eritrocitos en este ensayodio lugar a diferencias
estadísticanientesignificativasentrelos títulos hemolíticosregistradosa370<2 (p < 0.0005),
a 280<2(pc 0,0005) , a 20~<2 (ji <0,0005),y a 70<2 (ji <0,0005).
Dada lamayorlabilidad de los eritrocitosde conejoo su máselevadasensibilidad
frentea las hemolisinasde aeromonasen comparaciónconlos decordero,la discusión delos resultadossecentraen los datosobtenidoscon losprimeros.
Lossobrenadantesobtenidosapartir decultivosincubadosa‘70C-336h.(FablaIV.28.VéaseAnexoIVa, página174), dieron lugaratítuloshemoliticossobreeritrocitosde conejoinferioresa los obtenidosa 200C-SOh.,a 280C-24h.ya 370C-18h(TablasIV.29, JV.30,IV.31. VéaseAnexo IVa, páginas148 a 177). Las diferenciasson estadísticamentesignificativasen todoslos casos(ji <0,0005)y el númerodecepashemolíticas detectadas
a70<2 seredujonotablemente. Majeedy col. (1990)indicanque, la mayor partede lascepas
hemolíticasa 37~C,tambiénlo sona 50<2y señalan que laproduccióndeexotoxinasestáen
función dela poblaciónbacteriana,por lo quelas diferencias,queobservarona distintastemperaturas,fueron consideradas cuantitativasy dependientesde la tasabacterianaalcanzadaa esatemperatura.Complementandoestainformación, Mateosy col. (1993)
apuntanque la produccióndefactoresextracelularespuededepender tambiéndel efecto de
la temperaturasobresu síntesiso secreción.
El mayornúmero de cepashemolíticassedetectóa 280<2, sin embargo,los títulos
hemolíticos obtenidosa estatemperaturafueron inferioresalos registradosa 200<2 y a 370<2,
(ji < 0,025 y ji < 0,005, respectivamente).Este hechopone de manifiesto que la
temperaturaóptimade crecimientono coincidecon la temperaturaóptimadeproducciónde
factoresextracelulares.Al analizarel comportamientode las cepasde origen alimentario,elmayor númerode cepas hemolíticasse registraa 280<2, mientrasque el númerode las
detectadasa 20 y 370<2es bastantesimilar, indicandoqueexistencepashemolíticasa370<2
¡¡5

r
4,
r.
mv
w
u
PJiSUIiAIutSY DISÉL.StY\
queno lo son a 200C y sí a 280<?. Algo parecido ocurrecon algunascepashemolíticasa
200<2queno lo son a370<2 y si a28~C.Estoshechoshacenpensar queexisten temperaturas
óptimasde producciónde hemolisinasinferioreso superioresa 280C que daríanlugara
títulos hemolíticos,significativamentemásaltos,a20~Cy 370<?quea 280<?(p <0,025,en
ambos casos). En el caso delas cepas diarreicasno seapreciaron diferenciasen elnúmerode cepashemolíticasni en los títulos registradosa distintastemperaturas.Estamayorvariabilidaden las temperaturasde produccióndeexotoxinasde lascepasalimentarias frentealasdiarreicas hasidotambiénseñaladapor Kirov (1993).
Al compararlos títuloshemolíticoso el número decepashemolfticasdetectadascon
eritrocitos deconejo,a lasdistintastemperaturas,no seapreciaron diferenciasimportantes
asociadasal origen de las cepas (Tablas IV.28, IV.29, IV.30y JV.31. VéaseAnexo IVa,
páginas 147 a 177). Sin embargo,Knochel (1989a) ha señaladoun númerode cepas
hemolfticasnotablementemenora 37~C que a 1O~C, en aislamientosprocedentesde
ambientesfríos, concluyendoque muchascepasproductorasde toxinas a bajastemperaturasno tendríanun papelsignificativo in vivo. Contrariamente,Krovaceky col.(1994) no encontrarondiferenciasen el número de cepashemolíticasdiarreicasoprocedentesde sedimentosmarinos,detectadasa 37~C,aunqueestasúltimas mostrarontítulos más bajos. Igualmente, Figuray col. (1986) hallanactividadeshemolíticas,
citotóxicasy enterotóxicasbastantesimilaresentrecepasaisladasen pacientescon diarreay
encontrolessanos.
A 70<2 ningunacepadiarreicao alimentariadeA. caviaefue hemolítica,mientrasqueen las otrasespeciessedetectéun númeroconsiderabledecepashemolíticas.A 20, 28 y
370C las cepasdeA. caviaemostrarontítulos muchomásbajosqueel restode las cepas.A.
hydrophila fue la especieen la quesedetectéel mayornúmerode cepashemolíticasy los
títuloshemoliticos másaltos.De acuerdocon estosresultados,Majeedy col. (1989a,b)yCorreay col. (1993)apuntanque la producciónde hemolisinasestá asociadaa A.
hydrophulay A. sobria, en mayor medida queaA. caviae.Karunakarany Devi (1994) han
señaladoque la detecciónde hemolisinasen A. caviae dependede las condiciones
ambientales,principalmente,del hierro. Diversosestudiossobrela actividadhemolíticahan
puestode manifiestoque un 90% de cepasde A. hydrophilason positivasy existeuna
estrechacorrelaciónentrehemolisisy citotoxicidad(Callistery Agger, 1987;Okrendy col.
1987). Sin embargo,en un trabajo realizadoen Dinamarca,sóloel 37%de las cepasde A.
hydrophilafueron hemolíticas(Knochel y Jeppesen.1990).
116

RESULTADOS YDISCUSION
Por último, conviene señalarque la actividadhemolíticade lossobrenadantes,
obtenidosa28OC~24h. desapareciódespuésde someterlosa 560<2durante10 minutos,por
lo quelas hemolisinas detectadasse puedenconsiderartermolábiles.
IV.5.l.1.3. - Hemolisisdesarrolladapor el sobrenadantelibre de células, concentradoyfraccionadopordiafiltración, sobreeritrocitosde conejo
Se seleccionaron6 cepascuyos sobrenadantesmostraron títulos hemolíticossuperioresa 200 sobreeritrocitos de conejo, cuando laincubaciónserealizó a 280<? (A.
hydrophila 2 procedentede la ColecciónEspafiolade Cultivos Tipo,A. sobria11 de origen
diarreico y A. hydrophila 27, A. hydrophila 55, A. hydrophila 75 y A. hydrophila96 deorigenalimentario).A continuación,el sobrenadanteseconcentró20 vecesy se dividió en
cuatrofraccionespor diafiltración.
Comoseobservaen la Figura[‘1.1(VéaseAnexo iVa,página196), la fracciónqueconteníalos componentes delsobrenadantede pesomolecular superiora 50kDaalcanzó
títuloshemolíticossuperioresa 2000en todosloscepas y,además,era13-hemolíticasobreagar sangrede caballo.Estosresultadoscoincidencon los obtenidospor Ljungh y col.(1981)queaíslanuna¡3-hemolisinacitotóxicay no enterotóxicade 50kDa y porAsaoy col.
(1984), cuya13-hemolisinacitotóxicay enterotóxicamostróun peso molecularde 60 kDa.Stelma (1988) atribuye ladisparidadde resultadosde la enterotoxicidadal métodode
purificación,ya quemientras Ljunghsólo recupera el 0,6% de lasunidadeshemolíticas,
Asao recupera el 65%, por lo quesu ¡3-hemolisinaseríaindudablementemás activa. Burkey
col. (1987)y Stelmay col. (1986)handetectadouna¡3-hemolisinacitotóxicay enterotóxica
de 63 kDa, similar a lade Asao.Estosugiereque la actividadesenterotóxica,citotóxicay
hemolíticapodríanradicaren la mismamolécula. Contrariamente,Cahilí (1990)considera
que hemolisinas,enterotoxinasy citotoxinassonmoléculasdistintas,dadala existenciade
cepasquepresentanestas actividadesde formaaisladaeindependientey <2hakrabortyy col.
(1984) han señaladoque lasmoléculasresponsablesde estas actividades estarían
codificadasen genesdistintos.
Ljungh y col. (1981)describenunaa-hemolisinacitotónicay no enterotóxicade 63
kDa. Las técnicas empleadasen estatesisno hanpermitidoponerde manifiesto laposible
existenciade unaa-hemolisina,presenteen la fracciónconpesomolecularsuperiora 50
kDa,yaque hubierasido enmascaradapor la actividad¡3-hemolítica.
La fracciónde 30-50kDamostró títulos hemoliticosbajos(4-8)sobreeritrocitosde
conejoy no originó hemolisissobre agarsangrede caballo, lo quepodría debersea la
117

4,
1’•
u-
ev
e,
a
RuúSLIIIAI)úS Y I)LY(:tSIUV
diferente sensibilidadde los eritrocitosde una y otraespeciefrente a las hemolisinas de
acromonaso bien a la menordifusión del sobrenadanteen el agarsangrede caballo. La
actividaddetectadaen estafracción podríaconsiderarsedetipo residual,ya que la capacidad
de separaciónde la técnica empleadaesúnicamentedel90%.
En la fracción depesomolecularentre8-30kDa seregistrarontítulos hemoliticos
variablesen funcióndela cepa(entre64y 256)y permitióobservara-hemolisissobreagarsangrede caballo.Estafracción podríaconteneralgunasubunidadactivadelas hernolisinas
descritasporLjungh y col. (1981),Maoy col. (1984), Stelmay col. (1986)y Burke y col.
(1987) o algún factor extracelulara-hemolíticode peso molecularinferior al de la
a-hemolisinadetectadaporLjungh y col. (1981).En lafracciónque conteníalos componentesde pesomolecularinferioresa 8 k¿Dano
se detectóactividadhemolíticasobreeritrocitosdeconejo,ni sobreagarsangredecaballo.
Al considerarestos resultadosy parallegar a concretarla actividadde las fracciones
seríapreciso,previamente, purificary caracterizarcadaunade ellas.
IV.5. 1.1.4. - Evolución de la producciónde hemolisinas,activas sobre eritrocitos deconejo, en funcióndelcrecimientobacteriano
Seseleccionaronochocepas cuyostítulos hemoliticoseransuperioresa 100cuando la
incubación se realizó a280<2(A. sobria 13 y A. sobria14 deorigendiarreico;A. hydrophila
24, A. sobria34, A. hydrop/uita75,A. sobria92 A. hydrophila94 y A. hydrophila96 de
origenalimentario).
La producciónde hemolisinassobreeritrocitosde conejo(sin diferenciarportanto el
tipo de hemolisis)seempezóa detectaral final de la faselogarítmicao al principio de laestacionadade crecimiento(FiguraIV.2. VéaseAnexo IVa, página197). Ljungh y col.(1981)señalanquela ¡3-hemolisinaseproduceen la faselogarítmicay la a-hemolisinaen la
estacionana.En todaslas cepas laactividadhemolíticaseevidenciócuandola poblaciónbacteriana
alcanzó nivelespróximosa í09 ufe/mí. Enestesentido,Majeedy col. (1990) ya habían
indicadoque la tasa bacteriana mínimanecesariapara detectar laproducciónde factores
extracelulares. aunqueesespecíficaparacadacepa.siemprees superiora 108 ufc/ml.
118

RESULTADOSY DISCUSION
IV.5.l.2. - Enzimas(proteasasy lipasas)
IV.5.l.2.l. - Proteasas
IV.5. 1.2.1.1. - Caseinolisisy elastinolisisdesarrolladaporel sobrenadante completolibredecélulas
La mayoríade lascepas,tanto diarreicascomo alimentarias,hidrolizaronla caseína
(Tabla IV.32. VéaseAnexo IVa, página178). El nivel de producciónde proteasasde
ambosgrupos,expresadoen unidadesproteolfticas,fue similar. Eneste sentido,Krovaceky col. (1992b) observan quela mayoríade las cepasaisladasen alimentossuizos son
caseinolíticasy, en el trabajo de Mateosy col. (1993), no se apreciandiferencias
importantesen la actividadesproteolíticasde los sobrenadantesde cepasclínicas y
ambientales,obtenidosa28~>C.A. caviaeregistréel menorporcentajedecepas caseinolíticas,asícomola más baja
produccióndeproteasas.Igualmente,Leungy Stevenson (1988)observaronquetodaslascepasde A. hydrophila y la mayoríade las cepasde A. sobria produjeron proteasas,mientrasquesólotrescepasde A. caviae fueronproteolíticas.Krovaceky col. (1992b)
tambiénhandetectado actividadproteolítica tantoenA. hydrophilacomoenA. sobria.
La elastasaha sidoconsideradaun factor indicador devirulenciaen Pseudomonas
(Hsu y col., 1981). Como seobserva en la Tabla IV.32 (VéaseAnexoIVa, página178),
sólounacepano identificadaanivel deespeciey de origenalimentario,hidrolizóla elastina.Estosresultadosno concuerdancon los de Santosy col. (1988).queregistranun 42,4%delas cepasde Aeromonasconcapacidadde hidrolizarla elastina,al mismotiempoque,en el
casode A. hydrophila,estosautoresrelacionaronla virulenciaparapecesno sóloconla
elastinolisissino también con la estafilolisis. Por otro lado, Mateos y col. (1993)observaronque laactividad elastinolíticaes más frecuenteen cepasambientalesque en
clínicas.
IV.5. 1.2.1 .2. - Caseinolisis desarrollada por elsobrenadantelibre de células,concentradoy
fraccionadopor diafiltración
En todas lascepas,la mayoractividad proteoliticasedetectóen la fracción del
sobrenadante queconteníalos componentes depesomolecularsuperiora 50kDa(Figura
119

r
ev
a
a
o
e-
u-
u-
u-
u,
u-
RLSULlAVOS 1’ VISCUSION
IV.3. VéaseAnexo IVa, página198). No obstante,las tres cepas (unadiarreicay dos
alimentarias)que presentaronmáscapacidadde caseinolisis enestafracción, también
registraronunaactividadproteolíticaimportanteen la fracciónde pesomolecularentre8 y
30 kDa. En todaslas cepas,la fraccióninferiora 8 kDa hidrolizó la azocaseina, aunqueno
registré una actividad tan altacomo lasdosfraccionesyacitadas.En el tramocomprendido
entre 30y 50kDase puedeconsiderarquela escasaactividadproteolíticaregistrada(en tan
sólo doscepas)esde carácterresidual,ya que la eficaciade separaciónde la técnica
empleadaes de aproximadamenteel 90%. Sin embargo, Leungy Steveson(1988)
describen unametaiproteasatermoestablede 35 kDa y una serinoproteasatermolábil de
áSkDa, afirmandoquela secreciónde unao de las dos proteasasdependede la cepade
Aeronwnas.De acuerdocon esteautor,Rivero y col. (1990)hanconseguidodonaruna
proteasatermoestablede 38kDa.Porotro lado,Thunc y col. (1982)ponende manifiestola
existenciade una proteasatermoestableen el sobrenadantecrudo,que relacionancon la
acciónletal en peces.En la mismalínea, Kanai y Wakabayashi(1984) y Shieh(1987,1988) consideranque,en general,lasproteasas tendrían acciónletalsobrepeces.
Podría concluirse quela actividadproteolíticaseconcentraprincipalmenteen lasfracciones depesosuperiora 50 kDa y entre8 y 30 kDa, aunqueuna cepaalimentariamostróunaproteolisis importanteen la fracciónde pesoinferiora 8 kDa. Estosresultadosponende demanifiestola existenciade toda una variedadde proteasasexocelularessecretadasporAeromonasspp., asícomo de diferencias manifiestasentrelas distintas
cepas.En estesentido, Nietoy Ellis (1986) handescrito5 tipos deproteasasenAeromonas
spp.,al mismo tiempoqueregistrandiferenciasentrelasaisladasdedistintascepas.
IV.5.l.2.l.3. - Evolución de la producción de caseinasaen función del crecimiento
bacteriano
Seseleccionaronochocepasen las quesehabíadetectadouna actividadcaseinolítica
de 10 unidades, aproximadamente.
La producciónde proteasasscdetectóal final de la faselogarítmicadecrecimientoo al
principio de la estacionaria,cuandola poblaciónbacterianaalcanzóaproximadamentel0~j
ufc/ml (FiguraIV.4. Véase AnexoIVa, página199).
En lasFiguras IV.2y IV.4 (VéaseAnexoIVa. páginas19? y 199)secomprueba,tal
y como indica Cahilí (1990), que lamáximaproducciónde hemolisinascoincidecon el
aumentoen la elaboraciónde proteasas.En relacióncon este hecho, Howardy Buckley
(1985) apuntan que las proteasasno sólo activaríanlas hemolísinas que sesecretan
120

RESULTADOS Y DISCUSION
inactivas,sino queademássedancapacesde facilitar el acceso de otras enzimas asus
receptores.Contrariamentea esta acciónestimulante,Hardy y col. (1986) señalanque
inhibiendolas proteasasextracelularesse aumentadala estabilidadde las enterotoxinas.
Otro papelatribuidoa las proteasas, relacionadocon sucapacidadpatógenaparael hombre,hacereferenciaa lacapacidadde contrarrestar el efectobactericidadel suero,ademásde
facilitar la nutriciónde la célulabacteriana,apartir delos tejidosdelhospedador.
IV.5.1.2.2. - Lipasas
IV.5.1.2.2.l.- Lipolisis desaifollada porel sobrenadantecompletolibre de células
Las 100 cepas ensayadassecomportaroncomolipolifticas, y no sepudieron detectardiferencias significativasenel nivel de produccióndelipasasexocelulares(expresadocomo¡tg de ¡3-naftol liberados)en funciónde su origen(Tabla IV.33. Véase Anexo IVa, página
179). Wakabayashiy col. (1981)y Santosy col. (1988) estudianla produccióndeenzimas
en cepasambientales,observandoquela mayoríasonlipolíticas,aunqueen la bibliografíaconsultadano se¡tlacionala produccióndelipasasconvirulencia.
La cepasde A. hydrophlla, presentaron mayorcapacidad parahidrolizar elp-naftilcaprilato,mientrasquelas cepasdeA. sobria registraronmenoractividadlipoliltica.La fuerte producciónde proteasasy lipasasextracelularesen A. hydrophilaexplicalaimportanciade esta especieen la conservaciónde los alimentos.Algunos autores,comoSuhren (1989)y McKellar (1989), incluyen a las aeromonasdentro del grupo demicroorganismosalterantesde alimentosrefrigeradosyaque crecena bajastemperaturasyproducen unagranvariedadde enzimasexocelulares.
IV.5.I.2.2.2. - Lipolisis desarrollada porel sobrenadantelibre de células,concentradoyfraccionadopordiafiltración
Laactividadlipolítica seconcentróen la fracciónde pesomolecularsuperiora 50kDa. Comosepuedeobservaren la FiguraIV.5 (VéaseAnexo¡Va, página 200)la lipolisisregistradaen el restode las fraccionespuedeconsiderarseun artefactode la técnica.
Estudiosefectuadossobre lipasasen 1’. fragí, indican quepuedenexistir compuestoslipolíticos constituidospor subunidadesde 25 kDa que, por agregación,daríanlugar a
lipasasde 250 kDa(Lawrencey col.. 1967). Anguitay col. (1993),en uno de los escasostrabajosexistentessobrelas lipasasde Acromonas,hanpurificadouna lipasaextracelularde
121

r
u-
ev
u-
e
e
e
e>
e-
e-
u
0
a
0
e-
e-
RESULTADOS Y DISCUSION
67 kDa de A. hydrop/ziIa.
IV.5.1.2.2.3.- Evoluciónde la producciónde lipasasen funcióndelcrecimientobacteriano
Seseleccionaronochocepas cuyaactividad lipolíticadabalugar,aproximadamente,a
la liberaciónde 8 gg de¡3-naftol.
La actividad lipolítica comenzóa detectarseal final de la faselogarítmicade
crecimientoo principio de la estacionaria, aunqueen algunascepasno sepusode manifiesto
hastaalcanzarla faseestacionariaavanzada (FiguraIV.6. VéaseAnexo IVa, página201).
Estehecho explicarla la variedaden el comportamientode las cepas que, enalgunoscasos
(Priest, 1983),coincidecon la detección temprana obtenida enP.fluorescens,mientrasque
en otros (Mckellar, 1989>,la máxima producción delipasastiene lugaren la última fase de
crecimiento.
IV.5.1.3. - Citotoxinas
Comoya seafirmó al hablarde hemolisis,Burkey col. (1981a)hanseñaladoquelaactividadhemolítica,citotóxicay enterotóxicaestán correlacionadas, aunqueotrosautores,como Toddycol. (1989),no admitenestaasociaciónentrela producciónde enterotoxinasycitotoxinas.El estudio dela citopatogenicidad puedeampliarla informaciónobtenidaenlosensayoshemolíticos,si seconsideraqueLjungh y col. (1981)y Chakrabortyy col. (1984)han aislado unaenterotoxinano hemolíticacitotónica(quealterala morfología , pero nocausala muertecelular).La ¡3-hemolisinaobtenidaporMao y col. (1984)esenterotóxicaycitotóxica (determinamuertecelular). En cualquiercaso, ladetecciónde la enterotoxina
citotónicaesmuchomenos frecuente que lade la enterotoxina citotóxica(Seidíery col.,
1980; Johnsony Lior, 1981).Además,Burke y col. (1987) indican que,probablemente,la
enterotoxinacitotóxicatienemásimportanciacomo factorde virulencia que laenterotoxina
citotónicay Potomskiy col. (1987a)observan que laactividad citotóxicase asocia con
frecuenciaa cepasaisladasen heces diarreicas. En este sentido, Buningy col. (1986)
consideranqueel efectocitotónicopodríaserun efecto citotóxicoresidual,yaque,al diluirla enterotoxinacitotóxica hastadosis subletaleso calentarlaa 560C, el efecto sobrelas
célulaspasaa sercitotónico. Vadivelyy col. (1995)han detectado,en todaslas cepas
aisladasde pacientescon septicemia,actividadcitotóxicay hemolíticaperono enterotóxica.
122

RESULTADOS YDISCUSION
IV.5. 13.1. - Citotoxicidaddesarrolladapor elsobrenadantecompleto libre de célulassobre
líneascelularesestables
El título citotóxico se expresa como lainversade la dilución másalta queprovocael
50%de muertecelularsobrecélulasVero o el 100%sobrecélulasde mieloma.
La citotoxicidad sobrecélulasde mieloma se llevó a cabosobrelas 100 cepas.
Cuandoseemplearoncélulas Veroseensayaron28 cepasseleccionadaspor su actividad
hemolítica: 2procedentesde la ColecciónEspaifoladeCultivos Tipo(A. hydrophila 1 y A.
hydrophila2); 4 aisladasen hecesdiarreicashumanas(A. hydrophila8, A. sobria 11,.
sobria 13 y A. caviae18) y 22 aisladas enahimuno&Lagua& (A. sobria23,Aeromonasno
clasificada26, A. hydrophila 27, A. sobria 37, Aeromonasno clasificada40 , A.
hydrophila 43, A. caviae54, A. hydrophila 55, Aeromonasno clasificada62, A.
hydrophila63,A. sobria68, Aeromonasno clasificada70,Aeromonasno clasificada72,A. hydrophila75. Aeromonasno clasificada77,A. hydrophila78,A. caviae 87, A. sobria
91,Aeromonosno clasificada95,A. hydrophila96,A. caviae97 y A. hydrophila100)
La actividadcitotóxica deAeromonasspp. seha estudiadosobrenumerosaslíneascelulares:Vero, Int. 407, hígadode chimpancé, CHO, HeLa(Boulangery col., 1977;
Barer ycol., 1986; Kindschuchy col., 1987).Santosy col.(1988) incluso utilizanlíneascelulares poiquilotermas(de peces),comparándolascon los efectosobservadossobre
célulashomeotermas(HeLa, Vero y fibroblastos),sin apreciardiferencias clarasen latoxicidad.Sin embargo,enestetrabajo(TablaIV.34. Véase AnexoiVa, página180),las
células demielomaregistrarontítulos citotóxicossignificativamente(p<0.0005)más altos
que las célulasVero,a pesarde que,el título obtenidosobrecélulasde mielomasereferíaa
ladiluciónqueprovocabael 100%demuertecelulary, sobrelas Vero, a la queoriginabael
50%.Todaslas cepascitotóxicassobre células Verotambiénlo fueron sobre célulasde
mielomay sólo unacepacitotóxicasobrecélulasde mieloma, aunqueconun título muy
bajo (1,67),no lo fue sobrecélulasVero. Portanto,las célulasde mielomapodríansermásadecuadasparaestimarla actividadcitotóxicadeAeromonasspp.,principalmente,cuandoéstaesescasa.No obstante,dadala mayorproximidadfilogenéticade lascélulas Veroala
especiehumana,esposiblequela informaciónqueproporcionansobre lavirulenciade las
cepasestémásvinculadaa lapatogenicidadenel hombre.
El distinto comportamientode ambas líneas celularespodría debersea la mayor
sensibilidadde las célulasde mieloma frente alas citotoxinasde Aeromonas.En este
sentido, Bunnigy col. (1986)observanque las célulasde melanoma son unas100 veces
123

*
—,
e>
RESULTADOS Y DiSCUSION
más sensiblesquelas célulasCROmientrasque,Todd y col.(1989)Tegistranunaactividad
similar sobrecélulasHeLa(carcinomaepitelial de cervix humano),célulasYl (tumor de
cortezaadrenalen ratón)y célulasCRO, aunqueestasúltimas fueronligeramentemás
sensibles. También se podría pensarquelascélulastumorales.al crecermásrápidamentey
no alcanzaruna diferenciacióncelularcompleta, seríanmás inestables antecualquier
agresión,sin embargono existieron diferencias importantesen la toxicidad del
etilmetanosulfonato,utilizado como control positivo, sobre células Vero (303 ~.tgIml
provocaronel 50% demuerte celular)y célulasde mieloma (443¡tg/ml provocaronel 100%
de muerte celular).
Por otro lado, las lesionesmorfológicasindicadorasde toxicidad, tal y como se
observaen la FiguraIV.7 (VéaseAnexo[Va,página202), consistenfundamentalmenteenredondeamientoy desprendimiento,en el caso de las células Vero, pasandode unamonocapade células ligeramenteelongadasadheridaa la pareddel frasco a célulasredondeadasensuspensión.Lascélulasde mieloma lesionadassufrieronun cambiomenos
espectacular,perdieronsuturgenciay esfericidad,arrugándosey disminuyendode tamailo(FiguraIV.8. VéaseAnexo IVa, página203).
Veinte de las28 cepasensayadassobrecélulasVero (71,5%)y 75 de las 100 sobrecélulasdemielomasecomportaroncomocitotóxicas.Sin embargo,Frickery Tompsett(1989)observan citotoxicidad sobrecélulasHeLasólo en el 30%de las cepas.
Como semuestraen la Tabla IV.34 (VéaseAnexo IVa, página 180), lascepas
alimentariasregistraronlos títulosmásaltos,aunqueno sepudieronestablecerdiferencias
significativasentreéstosy los obtenidosde cepas diarreicas. Krovaceky col. (1994)
señalanque,aunquetodaslas cepasfueron citotóxicas sobre células CRO,las de origen
humanoalcanzarontítulos de citotoxicidadsuperioresa las aisladasen sedimentosmarinos,
mientrasque Mateosy col. (1993) no encuentrandiferenciasentrecepasclínicas yambientalescuandoutilizan célulasVero. Igualmente,Santosy col. (1988)observanque la
citopatogenicidadde cepas clínicas yambientaleses similar sobre varias lineas celulares
homeotermasy poiquilotermasy, contrariamente, Toranzo ycol. (1983a),Morgany col.
(1985)y Ciprianoy col. (1981)no encuentranasociación entrecitotoxicidady virulencia
parapeceso enteropatogenicidaden el hombre
Ninguna cepade A. caviae resultócitotóxicasobre células Veroy sólo cuatrolofueronsobre célulasde mieloma, registrandotítulos considerablementemásbajosque el
restode las cepas. Eleyy col. (1993),Correay col. (1993)y Majeedy MacRae (1994)
observan que la citotoxicidad se asocia,en escasasocasiones,a A. envine y.
¡24

RESULTADOS Y DISCUSION
frecuentemente,aA. hydrophila y A. sobria. Burke y col. (1984b)y Frickery Tompsett
(1989)no detectaron cepasde A. caviaecitotóxicasy señalanqueA. sobriaesla especieque, con mayorfrecuencia,muestra esta actividad.Yamamotoy col. (1994) empleanla
hibridación delADN paraclasificarlas cepasdeAeromonasy observanquela citotoxicidad
seasocia,preferentemente,aA. hydrophila,A. veronhibiotipo sobria y A. jandai (cuyo
fenotipo coincideconA sobria),y, conpocafrecuencia,aA. sobriabiotipo sobria y A.
caviae.Por tanto, el grupode cepasdenominadoA. sobria, segúnla clasificación
bioquímicade Popoff(1984), incluye cuatrogruposde hibridacióndistintos,destacandoA. sobriabiotiposobriadeescasa virulenciayA. veronil biotiposobriade mayor virulencia
e invasividad(Janday col., 1984b;Watsony col., 1985; Carelloy col., 1988; Grayy col.,
1990; Havelaary col., 1992).Namdariy Bottone (1990b)no estánde acuerdocon losautorescitados,señalandoqueA. caviaeescapazde producircitotoxinasy enterotoxinas en
funcióndel tiempoy lascondicionesde cultivo. Así, consideranqueestaespecienecesitaría16 horasde incubación,mientrasqueA. hydrophila y A. sobria produciríanexotoxinasa
partir de las 8 horasde incubaciónen caldo triptonasoja. Por otro ladopuntualizanque,
además,A. caviaenecesitaríacaldotriptonasojaadobleconcentracióny exentode glucosa,ya quela presenciadeesteazúcarpuedeinhibir la síntesisde estasmoléculas,al igual queescapazde reprimir la ruta delasácidostricarboxfficos,dandolugaral acúmulode ácidoacéticoresponsabledel fenómeno suicidaqueseasociapricipalmenteaA. caviae.Comofactoradicional,la mayorconcentraciónde salesdel mediopodríafavorecerla estabilidad
delas exotoxinas.
IV.5.l.3.2. - Citotoxicidaddesarrolladaporel sobrenadantelibre de células,concentradoyfraccionadopordiafiltración,sobrecélulasVero
Los mayorestítulos citotóxicos (Figura IV.9. Véase AnexoIVa. página204) se
registraronen la fraccióndepesomolecular superiora50 kDa.Lascepasconmásactividadcitotóxica,en estafracción, fueronlasmismasquemostraronmáspoderhemolítico(FiguraIV.1. VéaseAnexo [Va, página196). Estosresultadoscoinciden conlos obtenidos porLjunghy col. (1981),quedetectaron una¡3-hemolisinacitotóxica no enterotóxica de50-51kDa y por Asaoy col. (1984)que aislaronuna moléculade unos63 kDa con actividad¡3-hemolíticay citotóxica, quedifiere de la exotoxinade Ljungh enque, además,provoca
acúmulode fluidos al actuarsobreel asa intestinal ligadade conejo.
Igualmentese observócitotoxicidadcuandolas célulasVerose enfrentaroncon las
fraccionesdel sobrenadantede pesomolecular entre8 y 30 kDa. aunque lacorrespondiente
¡25

e>
e>
e
e>
e
e.
RESULTADOS YDISCUS,’ON
aA. sobria 11, que habíaobtenidouno de los títulos hemolíticosmáselevados.registróel
título citotóxico másbajo. A la vista de los resultados podríamos afirmar que, en esta
fracción, las actividades citotóxicasy hemolíticasno estaríanasociadas.A este respecto,
Ljungh y col. (1981)y Chopray col. (1994)describenunaenterotoxina citotónicay no
hemolíticade 15 kDa,y Potomskiy col. (1987b) detectanunamoléculasimilar, aunquede
pesomolecularsuperior(27 kDa). Estosautoresúnicamenteregistranactividadcitotónica
sobrecultivos celulares,pero como ya seha señaladocon anterioridad, la citotonicidad
puede debersea unaescasaconcentracióno a la parcialdesnaturalizaciónde la citotoxina,
comoconsecuenciadelmétodode purificaciónempleado.La diafiltración prácticamentenoafectaal estadonativodelas moléculas,aunqueal concentrarlas propiciaríala muertecelular
producidaporestafracción.Lasfraccionesdepesomolecularinferiora8 kDa y entre30y 50 kDa no afectarona
los cultivoscelulares.
IV.5.2. - Propiedadessuperficialesde cepasdiarreicasy alimentarias
IV.5.2.1. - Hemaglutinación
La hemaglutinación esunat&nica sencillaqueseutiliza paraobservarla capacidaddeuniónde los microorganismosacélulaseucariotas (Cahilí,1990). Singh y Sanyal(1993)handescritouna asociación relativaentrelacapacidadhemaglutinante,la resistencia al suero
y la actividad enterotóxicade cepasdeAeromonasspp.De acuerdo conBurke y col. (1984a),la aglutinaciónde los eritrocitos de conejoo
cordero,se considerófuertecuandola agregaciónerainmediatay completa(HGL++) y
débil si eraincompletao no instantánea(HGL+).
En la Tabla IV.35 (VéaseAnexo IVa, página181), seobservaquesólo43 cepasaglutinaronlos eritrocitosde conejo;de ellas, sólo 12 tambiénhemaglutinaronlos decordero. Estasdiferenciasfueronsignificativas(ji <0,001)eindicanquelos glóbulosrojosde conejo son mássensiblesa las hemaglutininasdeAeromonasspp. Eneste sentido,Parisy col. (1982) encuentrandiferentes gradosde hemaglutinación en función del tipo de
eritrocitos y Vadively y col. (1995) observaronque las cepasde Aeromonasno
hemaglutinabanlos eritrocitosde cordero,aunquesi los de hamster,conejoy ave. Santosy
col. (1988) observaronmayor númerodecepas hemaglutinantes coneritrocitoshumanosque con eritrocitos de trucha,no encontrando asociaciónentre lahemaglutinaciónde
126

RESULTADOS Y DISCUSION
eritrocitosde truchay la virulenciade la cepasparapeces,ni entrela hemaglutinaciónde
eritrocitoshumanosy la enterotoxicidaden ratón.Por otra parte,Atkinsony Trust(1980)
relacionanla aglutinaciónde eritrocitoshumanoscon laagregaciónacélulasdel epitelio
bucalaunque,en estecaso,el mecanismode adhesiónes diferente.
De acuerdo conlos resultadosobtenidos por Burkey col. (1984a),que noseñalan
diferenciasen la capacidad hemaglutinante entre cepasaisladasen pacientes con
gastroenteritis asociadaaAeromonasspp. y cepasacuáticas,en la TablaIV.35 (VéaseAnexo IVa, página181) seobservaque no existió unaasociaciónsignificativa entrelaaglutinaciónde eritrocitos de conejoo corderoy las cepas diarreicaso alimentarias.Coincidiendo conestosresultados,Singh y Sanyal (1993) no observanque el origenclínico o ambiental de las cepasse relacionecon unamayor o menor capacidadhemnaglutinante.Contrariamente,Atkinson y Trust (1980) detectanaglutinaciónfuertedeeritrocitoshumanosy adhesióna célulasdel epitelio bucal de cepasaisladasen hecesdiarreicashumanasy de cepasenterotoxigénicasy Caihill (1990)asociala hemaglutinacióncon unaproteínade la membranaexternaque posibilitala unión de las bacteriasa losreceptores antigénicosH deloseritrocitos humanos.EsteantígenoH tambiénaparece enlasuperficie delascélulasintestinales,por lo que podríaintervenir enla uniónaestascélulasantes dequetengalugar la liberación delasenterotoxinas.Burkey col. (1984a) sugieren
quelas hemaglutininasy, no sólo las enterotoxinas,deberían serconsideradasfactoresdevirulencia. Sin embargo,Jiwa (1983) y Santosy col. (1987) observaronque lashemaglutininasno son especificasdecepasaisladasenhecesdiarreicas humanas. Stewartycol. (1986)aislanunahemaglutininasoluble,apartir del sobrenandantelibre de célulasdeA. hydrophila, A.sobriay A. caviae,queno parecerelacionarseconla patogenicidad.
IV.5.2.2. - Inhibición dela hemaglutinaciónpor D-manosa, L-fucosay D-galactosa
La inhibiciónde la hemaglutinacióncon distintosazúcarespuedeaclarar, enparte, lanaturalezay especificidaddelas hemaglutininasdela superficiebacterianaqueinteraccionan
conlos receptores existentes enla superficiedeloseritrocitos (Trusty col., 1980).Dadalasensibilidad,especificidady probable naturaleza proteica,Cahilí (1990)recuerdaque lasmoléculasresponsablesde la hemaglutinación podríanser lectinas(que reconoceríanlosazúcaresterminales)y que secorresponderíancon aquellasque inhiben laspropiedades
aglutinantesde los receptoresde la superficiecelular.Hanney Finkelstein(1982) describen
cuatrohemaglutininasen y. cholerae con diferenteespecificidadfrentea eritrocitosde
¡27

e,
a
e>
e>
Rs
el
el
e>
e>
st
RESULTADOSY DISCUSION
distintasespecies,unade ellases inhibidapor fucosa,otra por manosa,la tercerano esinhibidapormanosani fucosay la cuarta esuna hemaglutininasolubleresistentetambiénaambosazúcares.
Seconsideróque hablaexistido inhibición de la hemaglutinación cuando lascepas
hemaglutinantesdejabandeserlo,o biencuandola hemaglutinaciónfuertese transformaba
endébil, en presenciade D-manosa,L-fucosao D-galactosa.
La capacidadhemaglutinantefrente a eritrocitosdecorderofue inhibidaporuno, doso
los tresazúcares. El23,3%de las cepashemaglutinantes coneritrocitosde conejopresentaronhemaglutinaciónresistentealos tresazúcaresy, en ningúncaso,sedetectóhemaglutinación
sensibleatodosellos(TablaIV.36. VéaseAnexo IVa, página182).Los patronesde inhibición con eritrocitosde cordero difierena los obtenidoscon
eritrocitosde conejo,por lo quepodríaconsiderarsela intervenciónde distintoselementossuperficialesen la adhesióna uno u otro tipo de eritrocito,o bien que la unión con los decorderoesmáslábil y fácil de inhibir. Morgany col. (1985)y Santosy col. (1988)observantambiéndistintos patrones deinhibición de la hemaglutinaciónen función del tipo de
eritrocitos.
El patrón de inhibición másfrecuentefue M+F-G-, tanto encepasdiarreicascomo
alimentariasy, la manosael azúcar quemásvecesinhibió la hemaglutinación(TablaIV.36.VéaseAnexo IVa,página182). El papelde la manosacomoinhibidorde la hemaglutinacióndeAeromonasspp. ha sidoseñaladopor Morgany col. (1985),al observarque todaslascepasmostrabanhemaglutinaciónsensibleamanosaconeritrocitosde alguna especie animal,
así comopor Atkinson y Trust (1980),Burke y col. (1984a)y Santosy col. (1988).Este
último autor,además,añadeque la hemaglutinaciónresistentea lamanosaesmásfrecuente
en cepasdiarreicasqueacuáticas.Santosy col. (1988) observaron que el patrónde inhibición más común,frente a
eritrocitoshumanosy de trucha,eraM+G-F+. Contrariamente, enla Tabla FV.36 (VéaseAnexo IVa, página 182) sepuede apreciarque sólo dos cepasde origen alimentario
mostraronestepatrón sobreeritrocitos de conejo.Burke y col. (1984a) indican quela
inhibición dela aglutinacióndeeritrocitoshumanospor manosa,fucosay no porgalactosaes
el patrónquemenosseasociaala diarrea,Por otro lado,Elbashir y Millership (1989) observaronque la hemaglutinación
resistentealos tres azúcares(M-G-F-) predominaen cepasaisladasen hecesdiarreicasy en
cepascitotéxicassobrecélulasVero.Contrariamente,los resultadosreflejadosen la Tabla
128

RESULTADOS Y DISCUSION
IV.36 (VéaseAnexo IVa. página182) indicanqueningunacepadiarreica mostréestepatrón.
Majeedy MacRae (1994)han señaladoquela hemaglutinaciónfueinhibida,enla mayoríadelos casos,poruno o másazúcares.
Sólo unacepadeA. caviaeerahemaglutinantecon eritrocitosde conejo,mostrandounpatrón de inhibición resistentea los tres azúcares,mientrasque Burkey col. (1984a),observanel patrón M+G-F+ paraestaespecie.
IV.5.2.3. - Autoaglutinación
Janday col. (1987)hanasociadola capacidadautoaglutinantedeAeromonosspp.condistintaspropiedadessuperficiales,proponiendoestecaráctercomomarcador de virulenciaal afirmar queel tipo SP-PAB+seasociaacepasaltamente virulentasaisladasde procesosinvasivos.En este sentido,Dooley y Trust (1988)observanquelas cepasPAB+ son másvirulentasparael ratón.
Lascepasautoaglutinantesmostraron precipitaciónespontAnea(SP)al crecerencaldoinfusióncerebrocorazónincubadoa300C-18h.y o precipitacióndespuésde mantenerestos
cultivos 1 horaa 1000C(PAR).El porcentajede precipitaciónespontáneasecalculécomparandola absorbanciaa
540nm. dela partesuperiordel cultivo, desarrolladoa 30<’C-18h.,con la de esemismocultivo, despuésde agitarlo.El porcentajede precipitaciónde las célulasbacterianas,mantenidasa 1000<2-lh,seobtuvo comparandola absorbanciaa 540nm.,de la partesuperiorde unafracción delcultivosometidaestetratamientotérmico,conla de la fracciónmantenidaatemperaturaambiente el mismotiempo.
El carácterautoaglutinante(Tabla IV.37. Véase AnexoIVa, página183) y losporcentajes deprecipitacióndelas cepasautoaglutinantes(TablaIV.38. VéaseAnexo IVa,página184)no pudieronasociarseconel origendiancicoo ajinientario.
En la TablaIV.37 (VéaseAnexo IVa, página183)sepuedeapreciarque5 cepasdelas 15 de origen diarreicoy 23 de las 81 alimentariasfueronautoaglutinantes,detectándose
los tres tipos de autoaglutinación(SP-PAiB+,SP+PAB+y SP+PAB-)en ambosgrupos.Doscepas diarreicasy 8 alimentariasmostraronel fenotipoSP-PAiB+. Coincidiendocon
estosresultados,Janday col. (1987)tampocoobservarondiferenciasentrecepasclínicasyambientales, registrandoun 30% de cepasautoaglutinantes,de las quemásde la mitad
fueron SP-PAB+.En estalínea, Santosy col. (1988) no apreciaron diferenciasen la
autoaglutinacidnde cepas virulentasy avirulentasparapeces.ni les fue posiblerelacionar
12<>

t
gr
e>
e>
e>
e>
e>
e>
e>
e>,
e>
e
gr
e>
RESULTADOS Y DISCUSION
autoaglutinacióny enterotoxicidad.
NingunacepadeA. caviaeeraautoaglutinante.Comosehaindicadoanteriormente,Janday col. (1987),asociaronla autoaglutinacióna cepasaisladasde procesos invasivos.
por lo que estosresultadosseríanaceptablesal considerarque A. caviae.rara vez, seaíslade bacteriemiasy estasson siemprepolimicrobianas(Janday col., 1984b).
IV.5.2.4. - Aglutinación con acriflavina
Comoseobservaen la Tabla IV.39(VéaseAnexo IVa,página185), el 53,3%de las
cepasdiarreicasy el 56,8%de lasalimentariasno seagregaron enpresenciadeacriflavina(Acri-). En estesentido,Santosy col. (1988) detectaronun 50% de cepasAcri-,
relacionandoestecarácterconla enterotoxicidadaunqueno conla virulenciapara peces.Janday col. (1987)hanasociadola incapacidadparaagzvgarseenpresenciadeacriflavinaconla letalidadparael ratón,aunquetambiénencuentranalgunascepasAcri+ letalesparael
ratón
IV.5.2.5. - Hidrofobicidad superficial
La adherencia bacteriana, esencialmente,esel resultadode dosmecanismosdiferentesde unión:uno decarácterespecíficoy otroinespecífico.El primeroimplicaríala interaccióndecomponentescomplementarios dela superficiebacterianay de la superficiecolonizable
con marcadaafinidad. La unión no específicaconlíevainteraccioneselectrostáticasohidrofóbicasde mucha menorafinidad (Sweet y col., 1987). ReferenteaAeronwnas,
Bartkovay Cizner(1994)hanseñaladoqueen cepasno fimbriadasla hidrofobicidadpuede
serel factor másimportanteresponsable dela adhesiónacélulasepiteliales.Las interaccioneshidrofóbicas jueganun papel importanteen la adherenciay
proliferación de los microorganismosa unaamplia variedadde superficiessólidas
(Marshall, 1976).La hidrofobicidadnaturaldela superficieexterna bacterianaintervieneenla distribuciónde los microorganismosen interfases(Marshall, 1976; Blanchardy Syzdek,
1978),en la adherenciabacterianaa superficies plásticasimpermeables(Dexter y col.,1975; Fletchery Loeb, 1979; Gersony Scheer,1980)y en la unión a células fagocitarias(van Oss,1978) y otrascélulasde mamíferos(Perers y col., 1977; Smythy col., 1978;
Beacheyy col., 1980; Rosenbergy col., 1981).Dillon y col. (1986) apuntanquelos resultadosobtenidosa travésde unasólatécnica
no reflejan la hidrofohicidadsuperficialreaJ.Por estemotivo, la hidrofobicidadseestimóen
130

RESULTADOS Y DISCUSION
base ala unión a filtros de nitrocelulosa(Lachicay Zink, 1984) y a la adsorcióna
hidrocarburoslíquidos: hexadecano,octanoy xileno (Rosembergy col., 1980; Sweetycol., 1987).Otrastécnicaspropuestaspara estudiar lahidrofobicidadson laagregacióncon
sal (Lindahí y col., 1981), la cromatograffade interacción hidrofóbica(Smyth y col..1978),la aglutinaciónde partículasde látex, la unión al cristal violeta (Paulay col., 1988)y
la adherenciaa superficiesde poliestireno(Rosenberg,1981).
IV.5.2.5.1. - Unión a filtrosde nitrocelulosa
Las 15 cepasde origen diarreico y las 81 de origen alimentariomostraronuna
capacidadde unión próximaal 75%,sin quesedetectasen diferenciassignificativasentreambosgrupos (TablaIV.40. VéaseAnexo IVa, página186). De acuerdocon Paulay col.(1988),la gran mayoríade lascepasdeAeromonas sepuedenconsiderar “fuertementehidrofóbicas”cuandoesta propiedadse estimaen función de la unión a filtros denitrocelulosa.EstosautoresconsideranfuertementehidrofóbicaaA. salmonicida449,que
presentaun porcentajedeunión del82%y relacionanestapropiedadcon laexistenciade lacapaA, al considerarquela mutante449-3,carentede capaA, muestraunacapacidaddeuniónde sóloel 20%.Porotro lado,la hidrofobicidadde la cepa449, estimadaen funciónde suadsorcióna hidrocarburos,esmoderadaconxileno y, prácticamente nula,conoctanoy hexadecano.
IV.5.5.2. - Adsorcióna hidrocarburos líquidos
La adsorciónahidrocarburoslíquidosy la unióna filtros de nitrocelulosapuedendarlugar a resultadosno concordantes,así Paulay col. (1988) indican queAcinetobacrer
calcoaceticusATCC 31012esfuertementehidrofóbicoen función desu adsorciónal xileno
(88%),al octano(90%)y al hexadecano(100%) y,moderadamentehidrofóbico, cuandosedeterminael porcentajede unióna filtros denitrocelulosa(46%).
IV.5.2.5.2.1. - Al hexadecano
Los resultadosrecogidos enla Tabla IV.40(VéaseAnexo IVa, página186)muestranque tanto las cepas diarreicas comolas alimentarias registraronun porcentajede adsorción
próximoal 15 %, porlo que la hidrofobicidadsepuedeconsiderarprácticamentenula. Por
lo general.los resultadosobtenidoscon el hexadecanofueron escasamente reproduciblesy.
131

e>
e>,
st
e>:
st
e>
gr
RESULTADOS Y DISCUSION
enocasiones,la absorbanciade la suspensiónbacterianatratadacon este hidrocarburo eraincluso superiora la delcontrol. En este sentido,Sweet y col. (1987) indican que la
fiabilidad de los ensayosde hidrofobicidad,en función de la adherenciaa hidrocarburos
líquidos, mejoracon la aireación finalde la suspensión bacterianaque eliminalos restosdel
hidrocarburo,exceptoen el casode que se empleehexadecano.Por tanto, aunque se
detectarondiferenciassignificativasen los porcentajesde adsorciónque presentaronlas
cepasdiarreicasy alimentarias,los resultados obtenidoscon el hexadecanono seconsideran
con el gradode fiabilidadpreciso.
IV.5.2.5.2.2.- Al octano
Lamayoríade las cepasmostraronhidrofobicidadmedia,en funciónde la capacidad
de adsorciónaestehidrocarburo,predominandolos valoresde adsorción próximosal 60%(TablaIV.40. VéaseAnexo[Va,página186). Al compararlosporcentajesdeadsorcióndecepasdiarreicasy alimentariasse observarondiferenciassignificativas (p < 0,005),resultandolas cepas aisladasen alimentosmás hidrofóbicas.Sin embargo,Janday col,(1987)observaronhidrofobicidadtanto en cepasvirulentascomoen avirulentas paraelratón.Contrariamente, Lachicay Zink (1984) asocianla hidrofobicidadsuperficialdeYersinia enterocolíticaa la existenciade un plásmidode 40-48 megadaltons, que
determinaríala colonizacióndel tractogastrointestinalen el ratón y cambiosen laspropiedadesffsico-quimicasde la superficie bacteriana.
IV.5.2.5.2.3. - Al xileno
Coincidiendo con Janday col. (1987) y Paulay col. (1988), elxileno fue el
hidrocarburocon elque seregistraronlos porcentajesmás altosde adhesión.Las cepasde
Aeromonasmostraronunahidrofobicidadmediacon unacapacidadde adhesiónpróximaal
62%, registrándosediferenciassignificativas (p <0,01)entrecepasdiarreicas (menoshidrofóbicas)y cepasalimentarias(máshidrofóbicas) (TablaIV.40. VéaseAnexo IVa,
página186).La superficiede las bacteriasgram-negativasesnormalmentehidrofiica, como
consecuenciade la presenciadelipopolisacáridoso cadenasde polisacáridos(LugtenbergyVan Alphen, 1983). El aumentode la hidrofobicidadsuperficial sueleasociarseamutacionesque conllevancambios de los lipopolisacáridoshacia formas rugosas(Lugtenbergy Van Alphen, 1983), a la producciónde adhesinas(Jones,1977: Smythy
i 32

RESULTADOSY DISCUSION
col., 1978; Lindahí y col., 1981) que incrementaríanla hidrofobicidadal facilitar la
adhesión(Jones,1977) oala presenciade unacapaextrasuperficialde naturalezaproteica,como la capaA de A. salmonicida(Trust y col., 1983). Sin embargo,la capa5 deAeromonasspp. móviles, similar por su naturalezay estructuraa la capaA de A.
salmonicida.no aumentala hidrofobicidadsuperficialde las células bacterianas(Dooley y
col., 1988).Tal y como sehaexpuestoanteriormente,la hidrofobicidad se relacionaría con
la capacidadde adhesióny. por tanto,con la virulencia.Sinembargo,Merino y col. (1995)
observanquelas cepasde A. hydrophila más virulentas para peces,productorasdesepticemias,poseen unacápsulade naturaleza polisacáriday, por tanto hidrofflica,que daríalugaraunadisminuciónde la bidrofobicidadsuperficialen estascepasaltamentevirulentas.
IV.5.2.6. - Aspectosmorfológicosde la superficiebacteriana
IV.5.2.6.1. - Capa5
Unaamplia variedadde bacteriaspresentancapassuperficiales proteicas (capas5) deestructuracristalina (Sletyr y Messner, 1983). Estasestructuras,normalmente,estánconstituidaspor monómerosproteicoso glicoproteicosensambladoscon simetríahexagonal,tetragonalo lineal(Sletyr y Messner,1983),queformanla capamásexterna dela célulay regulan,en granmedida,lasinteraccionesentrelos microorganismosy el medio
externo.Una de susfunciones esde simpleprotecciónfísica frenteaagenteslíticos, comoprotefnasdel suero(Munn y col., 1982) obacteriófagos(Ishiguroy col., 1981).La capaS,identificadapor Kay y col. (1981) en cepas autoaglutinantesde It. salmonicida,
(originalmentedenominadacapaA) esuno de los principalesfactoresdeterminantes de
virulenciapara peces,hastael puntodeque las cepas quecarecen decapaA seconsideranavirulentas(Evenbergy Lugtenberg,1982; Evenbcrgy col., 1982; Munny col., 1982;Parkery Munn 1984).
Posteriormente,seha observadola presenciade capa5 en cepasde A. hydrophila
GH-1 y A. veronil biotipo sobriaGH-8 (fenotípicamentesecorrespondecon A. sobria)
implicadasen distintasinfeccioneshumanasy animales(Dooley y Trust, 1988; Paulaycol., 1988; Kokka y col., 1991b).La capa5 de Aeromonasspp. móvilesúnicamente
aumentala resistenciaa la acción bactericidadelsuero(Murray y col., 1988)y no escapaz,comola de A. salmonicida,de facilitar la adhesióna macrófagos(Trust y col., 1983) o
presentarpuntos específicos de unión para porfirinas (Kay y col., 1985) e
inmunoglobulinas(Phippsy Kay, 1986).Dooleyy col. (1988)han indicadoquela capaS,
133

e>
1’
e>
gr
u
e
e>
st
gr
e,
RESULTADOS Y DISCUSION
constituidapor unidades proteicasde 52 kDa, esrelativamentehidrofóbica,aunquecuando
está unida a lacélulabacterianano le confieremayorhidrofobicidad.
Como seapreciaen la Tabla IV.4 1 (VéaseAnexoIVa, página187),la capa5 (Figura
IV.l0. VéaseAnexo IVa,página205) se detectóen 65 de las 100 cepas ensayadas
observándose,indistintamente,en cepasdiarreicasy alimentarias.Esteresultadopodría
suponerla presenciade cepaspotencialmentevirulentasen alimentos,si setieneen cuenta
que Dooley y col. (1986)y Murray y col. (1988)han apuntadoque las cepasde A.
hydrophila, altamentevirulentasparapeces,poseenun antígenosomáticocomún (0:11) y
unacapaproteicauniforme en la superficie externade la célula, cuyassubunidadessedenominanproteína5. Las unidadesproteicasde la capa5 no son idénticasa las proteínasde la capaA deA. salmoniciday, pareceserque, también sediferencianantigénicamente(Janday col. 1987). Por otro lado, Janday col. (1987) señalanqueno se ha podidorelacionarla existenciadela capa5 con elcomportamientoPAB+de cepasdeA. hydrophila
altamentevirulentas.Sin embargo,Paulay col. (1988)hacen referenciaa un nuevogrupodeIteromonasmesófiias patógenas,con capa5. implicadasen infeccionesgravesde peces
y sereshumanos,con un mismofenotipoautoaglutinante(PAB±)y un antígenosomático
común(0:11).
Porotro lado, lafrecuenciacon que se detecta la capa8 entrelascepasdeAeromonas
(TablaIV.41. VéaseAnexo[Va, página187) tanto diarreicascomo alimentarias, podría
ponerendudasu validezcomo indicadorde virulencia En estesentido,la opinión delos
investigadoresquehanestudiado este problemano escoincidente,asíCahill (1990) señalaquela capa5 proteicaesel mejor indicadorde virulencia, mientrasque, Merino y col.(1993)consideranmásfiables las pruebasinmunoenzimáticas paradetectarel antígeno
0:11.
1V.5.2.6.2. - Pu
Los pili de Aeromonasspp. móvilespueden clasificarse,en función de su
morfología,en dos tipos:pili cortosy rígidos, abundantesen la superficiecelular quesedetectanen cepasambientalesy pili largosy flexibles,en menornúmeroporcélula (1 a 60),queaparecentantoen cepaselínicáscomoambientales(Carrello y col., 1988;Hokamaycol., 1990; Ho y col., 1990; Kirov y col., 1993b). Carrello y col. (1988) apuntanque laadhesióna célulasHEp-2 estárelacionadacon la expresióndelos pilí flexibles,aunqueno
existe certezade que estalíneacelularestabletengalos mismosreceptoresde superficie
para bacteriasque las células intestinaleshumanasin vivo. Nishikawa y col. (1991>
‘34

RESULTADOS Y DISCUSION
demuestranque laadhesiónde las cepasdeAeromona,saestalíneacelularesparalelaa la
adhesiónacélulasintestinalesde embrión humanoy Rogersy col. (1990) yGreyy Kirov
(1993)afinnanquela adhesiónde las cepasdeAeromonasse correlaciona con la quetiene
lugar sobretejido ileal humanoformolizado.
Comose muestraen la FiguraIV. 11 (VéaseAnexo IVa. página206) sepudieron
observarlos dostipos depili en cepasdiarreicasy alimentarias.
En la TablaIV.41 (VéaseAnexo IVa, página187) seapreciaquesedetectaronpili
conosy rígidos en 22 cepas,y largos y flexibles en 5 cepas,sin que se apreciasendiferenciassignificativasentrecepasdiarreicasy alimentarias.Sehanpurificado los pilí
largos,a partir de doscepasde A. hydrophila(Hokaniay col., 1990; Ho y col., 1990) y detres cepasde A. sobria (Hokamae Iwanaga,1991, 1992; Iwanagay Hokama, 1992).Cuatrode lospili purificados(uno deA. hydrophilay tresdeA. sobria) secomportaron
comofactoresquefavoreceríanla colonización(Hokamay col., 1990;HokamaeIwanaga,1991, 1992;Iwanagay Hokama,1992),habiéndosecomprobadocapacidadhemaglutinante
en tresdeellos.También,Satoy col. (1989)aIslan un pilusen A. hydrophila,aunquenolograncorrelacionarlocon hidrofobicidadni hemaglutinación.Porotro lado,pareceserquelascepasenterotoxigénicasdeorigen fecal(A. veroniibiotiposobria)muestran menos¡>111
quelascepasambientalesde la mismaespecie(Kimv y col., 1986)y las bajastemperaturasdecrecimientofavorecenla expresiónde 1ospili, independientementedel origendela cepa
(Kirov y col., 1993a,b).
IV.5.3. - Característicasbioquímicas de cepasdiarreicasy alimentadas
Se hanpropuestocomo indicadoresde enteropatogenicidaddistintascaracterísticas
fenotípicas,tales como labemolisis, la descarboxilaciónde la lisina, la reacciónVoges
Proskauer, la fermentaciónde la arabinosa,de la salicinay del sorbitol (Burke y col.,
1984b;Abeytay col., 1986;Kirov y col., 1986; Callister y Agger, 1987).Sin embargo,Figuray col. (1986)y Kindschuchy col. (1987) han cuestionadola validezde algunosdeestos caracterescomo marcadoresde virulencia. Carnahany col. (1990)señalanque la
actividadpirazinamidasapuedeser indicadorde virulenciaen A. sobria.Por otro lado,Namdariy Bottone(1988)relacionan elcomportamientono suicida,al creceren presenciade 0,5% deglucosa,con laenteropatogenicidaddeA. sobriay A. hydrophila.Mateosy
col. (1993) consideraronque lautilización de la ramnosay el DL-lactatodependería delorigen.de modo que, ninguna cepa clínicaasimilaríala ramnosay ningunaambientalel
DL-lactato.
135

‘a
st
e,
e,
e,
e,
e,
e,
RESULTADOS Y DISCUSION
IV.5.3.1. - Voges Proskauer
Comose observaen la Tabla IV.42(VéaseAnexo IVa,página188) el 60% dc las
cepasdiarreicasy el 71,60%de las alimentadasson positivasparaestareacción.No se
apreciarondiferenciassignificativasen larespuestaa estareacciónen funcióndel origen,
mientras queCallistery Agger(1987) afirmanque secorrelacionacon lacitotoxicidad.Sin
embargo,Janday col. (1983b) encontraronque sólo el 42% de 12 cepascitotóxicasdeA.
hydrophila, aisladasen niñoscongastroenteritis,sonVogesProskauerpositivas aunque,deacuerdocon la clasificaciónde Popoff (1984),las cepasVoges Proskauernegativas
seríanA.caviaeo A. sobria. Abeyta y coL (1986), al igual que Burke y col. (1983b),observaronuna fuerte relación entre el carácter Voges Proskauerpositivo y laenterotoxigenicidadparael ratón.
Al aplicarel esquema de identificaciónbioquímicaempleadoen estetrabajo para
diferenciarlasespeciesningunacepadeA. caviaeresultáSVogesProskauerpositiva.
JN.3.3.2.- Descarboxilaciónde la lisina
Aproximadamente,el 67% de las cepasdiarreicasy el 50% de las alimentarias
descarboxilaronla lisina (TablaIV.42. VéaseAnexo IVa, página188). En consecuencia,estecarácterno pudoasociarseal origende las cepas.Cailistery Agger (1987)apuntanquela producciónde lisina descarboxilasaesun importanteindicadorde actividad citotóxica.Otros autores(Cumberbatchy col., 1979; Kaper y col., 1981; Burke y col., 1983b)observarontambiéncorrelaciónentreestecarácter fenotípicoy la producciónde toxinas.Sinembargo,Abeytay col. (1986)no handetectadoasociaciónalgunaentrela descarboxilación
de la lisina y la actividadcitotóxica.
IV.5.3.3. - Fennentaciónde la arabinosa
Un 60-70% de lascepas,tantodianeicascomoalimentarias,fermentaronesteazúcar,porlo queno sehanapreciado diferenciassignificativasenfunción del origen (Tabla IV.42.
VéaseAnexo IVa, página 188). Contrariamente,Abeytay col. (1986) y Burke y col.(1983b) relacionanla actividadenterotéxicacon laincapacidadparafermentarla arabinosay
la reacciónVP-positiva.Sin embargo, Mateesy col. (1993)no encuentrandiferenciasen la
fennentaciénde la arahinosa entre cepas clínicasy ambientales
136

RESULTADOS YDISCUSION
Despuésde estudiar los factores de virulencia, independientemente,en cepas
diarreicasy alimentarias, se hainvestigadosi las asociaciones entre estos factores,descritas
por otrosautorescomo indicadorasde virulencia,soncapacesdediferenciarambosgruposde cepas.En estesentido,Mittal y col. (1980) y Lallier y col. (1981) compruebanque lascepasPAiB+ y Acri- son altamentevirulentaspara peces,poseenun antígenoO comúnymayorresistenciaa la actividadbactericidadel suero.En la TablaIV.43 (Véase AnexoIVa,
página 189)seobservauna mayor incidenciade patronesPAiB+ Acri- entrelas cepasdiarreicas(20%)queentrelas alimentarias(6%), aunqueestaasociaciónno essignificativa(p c 0,1). Janday Brenden (1987),Paulay col. (1988) y Kokka y col. (1991a,b)
encuentranun grupo decepasautoaglutinantesconcapa5 y un antígenocomún(0:11)implicadasen infeccionesgravesde hombresy animales. LaTablaIV.43 (VéaseAnexoIVa, página189)muestraquelas cepasautoaglutinantesconcapa5 sedetectarontantoen
cepasde origen diarreicocomoalimentario.Burkey col. (1983b) y Abeytay col. (1986)señalanquela reacciónVoges Proskauerpositiva y la incapacidad parafermentarlaarabinosasecorrelacionan conla enterotoxicidad.De acuerdoconla Tabla IV.43(VéaseAnexo IVa, página 189), estecriterio no permitió diferenciar cepas diarreicasdealimentarias, enlas condiciones de nuestraexperiencia
IV.5.4. - Asociaciones detectadas entre los factores indicadoresde
virulencia
Contrariamentea lo indicadopor numerososautores(Boulangery col., 1977;Burkey col., 1984b;Tumbulíy col., 1984; Aggery col., 1985;Callistery Agger, 1987),aunque
coincidiendocon otros (Figura y col., 1986; Krovaceky col., 1992a), no se detectérelaciónentreactividadhemolíticay citotóxica.
En la Tabla IV.44 (Véase AnexoIVa, página 190) se apreciaque las cepashemaglutinantesmuestrantítuloshemolíticos significativamentemásaltos.Considerandoquela hemolisispuedeconstituirun indicador deenterotoxicidad(Burke y col., 1981a;Majeedy col.. 1989a),el empleoconjuntode estasdostécnicaspodría diferenciarlas cepas
másenterotóxicas.
Lascepascon pili muestran mayoractividad lipolítica(Tabla IV.45. Véase Anexo
IVa. página 190). La asociaciónde estosdos factorespodríaconstituir un indicadorde
¡17

r
y
e,
e,
e,
e,
e,
RESULTADOS Y DISCUSION
cepaspocovirulentasyaque. otrosmarcadoresde virulencia,comola autoaglutinación,la
descarboxilaciónde la lisina y la fermentaciónde la arabinosaindican quela cepaspotencialmentemáspatógenasseasociancon bajaactividadlipolítica y ausenciadepilí.
Lascepasque presentancapa5 mostraronmenor adhesiónal octano(Tabla IV.46.Véase AnexoIVa, página191), pero no sediferenciaron,en la capacidadde unión a losfiltros de nitrocelulosa,de lascepascarentesde capa 5. Por otro lado, las cepas
descarboxiladorasde la lisina presentan menorcapacidadde unión a los filtros denitrocelulosa(Tabla IV.47. Véase AnexoIVa, página 191), aunqueno seapreciarondiferenciasenla adhesiónal octano.Deacuerdoconnuestros resultados, asícomolos deDooley y col. (1988),la frecuenciadecapa5 enAeronwnasspp.móvilesrio aumentala
hidrofobicidad.Otros factores indicadoresde virulencia como la autoaglutinación,descarboxilaciónde la lisina y reacción VogesProskauerseasociaron enmayoro menorgradoala hidrofobicidad.
Las cepasno aglutinantesen presenciade acriflavinamuestran mayor actividadproteolítica(sobrela caseína)y capa5 (Tabla IV.48. VéaseAnexo IVa, página192).Aunquela bibliografíamencionaun grupode cepasaltamentevirulentasprovistasde capa
S, autoaglutinantesy conun antígenocomún(0:11), Janday col. (1987)no encuentranunacorrelaciónclara entreautoaglutinacióny presencia decapa5. La no aglutinaciónconacriflavinapodríaconstituirun indicadorútil paradiferenciar cepascon unacapaextrade
naturalezaproteicaen la superficiecelulary, porello, posiblementevirulentas.
La actividad proteolíticamarcada seha relacionadotambiéncon autoaglutinacióny
reacciónVogesProskauerpositiva. Por tanto, aunqueno estámuy claro el papelde estas
enzimasen la patogenicidad, las cepasmás virulentasparecen produciro secretarmásproteasas.
ParaJanday col. (1987) la autoaglutinaciónesun marcador decepasaltamentevirulentas,aisladasde procesosinvasivos. Larelaciónde la autoaglutinación con unaactividadproteolíticaelevaday unalipolítica baja(TablaIY.49. VéaseAnexo IVa, página193) indica queestasactividadesseasocianacepaspotencialmentemásvirulentas.Las
cepasautoaglutinantesfueron más hidrofóbicas,mientrasque otros indicadoresdevirulencia (descarboxilaciónde la Usina y presenciade capaS) se asociana escasa
hidrofocibidad,sin olvidarque. tal y como se ha expuestoanteriormente,las cepasdc
138

RESULTADOS Y DISCUSiON
origen diarreico mostraronmenor capacidadde adhesiónal octanoque las cepas
alimentarias.
El fracaso para fermentar la arabinosay, especialmente,la reacciónVogesProskauer
positivafueronlos factores asociados a altaactividad citotóxica(sobrecélulasde mieloma).
Además,las cepasno fermentadorasde la arabinosa se asociarona una bajaactividad
lipolítica, menorpresenciadepili, capacidadhemaglutinantey descarboxilaciónde la lisina
(TablaIV.50. VéaseAnexo IVa, página194), indicando que lano fermentaciónde laarabinosapodría seleccionarlas cepasconmás posibilidadesde servirulentas.
Lareacción VogesProskauerpositivaesla queseasociaal mayornúmerode factores
de virulencia(TablaIV.51. Véase AnexoIVa, página195). Además,esel indicadormásrelacionadoconlas actividadescitotóxicay hemolítica.Tambiénseasociaclaramentea la
actividadproteolitica,autoaglutinación,hemaglutinación.aglutinacióncon acriflavina,descarboxilaciónde la lisina e hidrofobicidad.Las cepasVoges Proskauer positivasmuestranmenorcapacidad deuniónalosfiltros de nitrocelulosay más adsorciónal octano,
indicandoque, aunquelas cepasdeAeromonassepuedenconsiderarhidrofóbicaspor
ambosmétodos,estosno diferencianlasquelo son enmayorgrado.La reacciónVogesProskauerpositiva, al ser empleadaen la identificación de
especies,excluyelascepasdeA. caviae.Comoyasehaindicado, estaespecieesdeficitariaparaun importantenúmero defactoresde virulenciay seconsideramenosvinilentaqueA.
hydrophllay A. sobria(Burke y col., 1984b;Callistery Agger, 1987;Correay col., 1993).Se puede deducir,de acuerdocon nuestros resultados,que la reacción Voges
Proskauerinforma, en cierta medida, sobre lapatogenicidadpotencialdeAeromonasspp.
móviles,comopreviamente ya han señaladoBurke y col. (1983b),Abeytay col. (1986)y
Callistery Agger(1987).
Seríanecesariodetenninarla implicación real decadauno de estosfactoresconla
virulenciapara escogerel indicadormásadecuado.Hay que teneren cuentaqueestosfactoresdevirulencia sehandetectadotanto en cepasdiarreicascomoalimentarias,por loque,estasúltimas podríanser,potencialmente,tanpeligrosascomolas diarreicas.
IV)

4,..
e,
e,.
e,
e,’
e,
e,

¡Va. ANEXO (Tablas y Figuras)

ty..
it
t

TablaIV.1. - Recuentode laflora naturalpresenteendiversosalimentossobrelos
mediosensayados
Mbncnlos
Aguadorada
ISA
Lechecruda
Carnevacuno
5,64
7,51
5,11 (90,60)
4,17 (55,52)
2,39 (42,37)
3,77 (50,19)
Carnepollo
Carnecerdo
Pescadilla
Trucha
Chirlas
Gambas
5,95
6,13
4,88
4,66
4,95
6,84
4,61 (77,47)
4,20 (68,51)
3,90 (79,91)
3,73 (80,04)
3,53 (71,31)
5,76 (84,21)
4,00 (67,22)
4,39 (71,61)
3,21 (65,77)
3,23 (69,31)
2,77 (55,95)
4,84 (70,76)
4,41 (74,11)
4,20 (68,51)
3,93 (80,53)
3,37 (72,31)
3,43 (69,29)
5,90 (86,25)
4,37 (75,94) 3,57 (61,64)
0,75 (10,76) 0,82 (10,85)
4,30 (74,54)
0,84 (11,29)
N.D. - Crecimiento nodetectado(a).- Expresadoen log. ufc/m¡. o«O. - PorcentajederecuperaciónTSA. - Agartriptonasoja
AAA. - AgaralmidónampicilinamA. - Medio deRyan (Unipath)AGAP. - Agarglutamato almidónpenicilina
X. - Mediaaritmética
D.S. - Desviación esténdar
g. de alimento
Aa
(a) (a) (b)
1
mA
(a) (b)
N.D.N.D.
AGAP
(a) (b)
N.D.
5,11
4,11
(90,60)
(54,72)
xD.S.
5,82
1
143

t
u,
e
u,
u,
Tablalvi- RecuentosobtenidosendiversosalimentosinoculadosconAeromírntzs
spp.móviles (cepas839,398,837y 838)sobrelos mediosensayados
Alimentos Nl MA mA AA~AP
(a) (a) (b) (a) (b) (a) (b)
Aguadorada 7,62 6,24 (81,88) 7,38 (96,85) 6,26 (82,21)
Lechecruda 7,48 5,94 (79,41) 7,02 (93,85) 6,01 (80,34>
Carnevacuno 7,47 5,80 (77,64) 6,95 (93,03) 5,86 (78,44)
Carne polio 7,2 5,85 (81,25) 6,66 (92,50) 5,70 (79,16)
Carnecerdo 7.02 5,63 (80,19) 6,64 (94,58) 5,68 (80,91)
Pescadilla 6,95 5,56 (80,00) 6,60 (94,96) 5,61 (80,71)
Trucha 6,93 5,58 (80,51) 6,34 (91,48) 5,52 (79,65)
Chirlas 7,03 5,58 (79,37) 6,63 (94,31) 5,60 (79,65)
Gambas 6,82 5,45 (79.91) 6,51 (95,45) 5,47 (80,20)
7,16 5,73 (80,01) 6,74 (94,11) 5,74 (80,14)
D.S. 0,28 0,24 (1,21) 0,31 (1,62) 0.25 (1,09)
Ni. - Nivel deinoculación(a). - ExpresadoenIog. ufc/ml. o g.dealimento(b). - PorcentajederecuperaciónAAA. - AgaralmidónampicilinamA. - Medio deRyan(Unipath)AGAP. - Agar glutamatoalmidón penicilina
X. - Media aritméticaD S - Desviaciónestándar
144

Tabla IV.3 - Recuentosobtenidosendiversos alimentos inoculadoscon
A. hydrophila CECT 839 sobrelos medios ensayados
Múnca Nl MA mA AGAE(a) (a> (b) (a) (b) (a) (b)
Aguadorada 6,93 6,76 (97,54) 6,59 (95,09) 6,66 (96,10)
Lechecruda 7,42 6,91 (93,12) 6,78 (91,37) 6,91 (93,12)
Carnevacuno 7,37 7,07 (95,92) 6,89 (93,48) 7,17 (97,28)
Carnepollo 7,3 6,90 (94,52) 6,70 (91,78) 6,85 (93,83)
Carne cerdo 7,31 7,00 (95,75) 6,72 (91,92) 7,04 (96,30)
Pescadilla 7,21 6,95 (96,39) 6,69 (92,78) 6,92 (95,97)
Trucha 7,19 7,07 (98,33) 6,61 (91,93) 6,79 (94,43)
Chirlas 7,2 6,82 (94,72) 6,64 (92,22) 6,82 (94,72)
Gambas 7,16 6,90 (96,36) 6,75 (94,27) 6,86 (95,81)
X 7,23 6,93 (95,85) 6,70 (92,76) 6,89 (95,28)
D.S. 0,14 0,10 (1,58) 0,09 (1,26) 0,14 (1,33)
N.I. - Nivel deinoculación(a). - Expresado en ¡og. uit/ml. o g. de alimento(b). - PorcentajederecuperaciónAAA. - Agar almidónampicilinamA. - MediodeRyan (Unipatb)AGAP. - Agarglutamato almidónpenicilina
X. - Media aritmética
D.S. - Desviaciónestándar
‘45

u,
Tabla JN.4 - Recuentos obtenidosen diversosalimentos Inoculadoscon
A. hydrophlla CEO? 398 sobrelos mediosensayados
Ni MA mA A=AE
(a) (a) (1,) (a) (b) (a) (b)
Aguadorada 7,94 7,65 (96,34) 7,67 (96,59> 7,69 (96,85)
Lechecruda 7,45 7,00 (93,95) 6,85 (91,94) 6,95 (93,28)
Carne vacuno 7,52 7,07 (94,01) 6,92 (92,02) 7,07 (94,01)
Carne poiio 7,06 6,66 (94,33> 6,50 (92,06) 6,57 (93,05)
Carnecerdo 6,99 6,80 (97,28) 6,82 (97,56) 6,83 (97,71)
Pescadilla 6,93 6,76 (97,54) 6,72 (96,96> 6,73 (97,11)
Trucha 6,96 6,83 (98,13) 6,34 (91,09) 6,90 (99,13)
Chirlas 7,81 7,32 (93,72) 7,23 (92,57) 7,25 (92,82)
Gambas 6,92 6,69 (96,67) 6,65 (96,09) 6,61 (95,52)
X 7,28 6,97 (95,77) 6,85 (94,09) 6,95 (95,49)
D.S. 0,4 0,32 (1,76) 0,39 (2,61) 0,34 (2,31)
N.I. - Nivel deinoculación(a). - Expresadoeníog. ufdnil. o g. dealimento(1». - PorcentajederecuperaciónAAA. - AgaralmidónampiciinamA. - Medio deRyan (tJnipatb)AGAP. - Agarglutamatoalmidónpenicilina
X. - Mediaaritmética
OS. - Desviaciónestándar
146

Tabla IV.5 - Recuentos obtenidosen diversos alimentosinoculadoscon A. caviae
CECT 838 sobrelos mediosensayados
Nl MA mA(a) (a) (b) (a) (b) (a) (b)
Aguaclarada 7,79 7,56 (97,04) 7,68 (98,58) 7,61 (97,68)
Lechecruda 7,43 6,92 (93,13) 7,04 (94,75) 6,91 (93,00)
Carnevacuno 7,48 6,85 (91,57) 6,50 (86,89) 6,89 (92,11)
Carnepollo 6,89 6,69 (97,09) 6,43 (93,32) 6,46 (93,75)
Carnecerdo 7,26 6,88 (94,76) 6,87 (94,62) 6,89 (94,90)
Pescadilla 7,24 6,83 (94,33) 6,93 (95,71) 6,87 (94,88)
Trucha ‘7,18 6,63 (92,33) 6,55 (91,22) 6,57 (91,50)
Chirlas 6,82 6,64 (97,36) 6,61 (96,92) 6,71 (98,38)
Gambas 6,88 6,46 (93,89) 6,60 (95,93) 6,55 (95,20)
X 7,21 6,82 (94,61) 6,80 (94,21) 6,82 (94,60)
D.S. 0,32 0,31 (2,14) 0,39 (3,45) 0,33 (2,32)
Ni. - Nivel deinoculación(a). - Expresadoen log. ufc/ml. og. dealimento(b). - PorcentajederecuperaciónMSA. - Agat almidónampicilinamA. - Medio deRyan(Unipath)AGAP. - AgarglutamawalmidónpenicilinaX. - Media aritmética
D.S. - Desviaciónestándar
147

e
u,
st
u,
u,
Tabal IV.6 - Recuentosobtenidos en diversosalimentos inoculadoscon A. sobria
CECT 837 sobrelos mediosensayados
AJhn~nUm Ni MA mA AGAf
(a) (a) (b) (a) (b) (a) (b)
Agua dorada 7,85 3,01 (38,34) 7,58 (96,56) 3,10 (39,49)
Lechecruda 7,65 2,93 (38,38) 7,44 (97,25) 3,27 (42,74)
Carnevacuno 7,52 2,23 (29,65) 7,49 (99,60) 2,33 (30,98)
Carnepoiío 7,55 3,16 (41,85) 7,04 (93,24) 2,94 (38,94)
Carnecerdo 6,55 1,87 (28,84) 6,17 (94,92) 1,98 (30,46)
Pescadilla 6,44 1,73 (26,94) 6,07 (94,25) 1,95 (30,27)
Trucha 6,39 1,79 (28,01) 5,86 (91,70) 1,84 (28,79)
Chirlas 6,3 1,56 (24,76) 6,04 (95,87) 1,64 (26,03)
Gambas 6,33 1,76 (27,80) 6,07 (95,98) 1,87 (29,54)
X 6,95 2,22 (31,61) 6,64 (95,48) 2,32 (33,02)
D.S. 0,66 0,63 (6,16) 0.72 (2.31) 0,61 (5,79)
N.I. - Nivel deinoculación(a). - Expresadoenlog. ufciml. o g. dealimento(b). - Porcentajede recuperaciónMSA. - Agar almidónampidiinamA. - MediodeRyan(Unipatb)AGAP. - AgarglutainaloalmidónpenicilinaX. - Media ariunéticaD.S. - Desviaciónestándar
148

en
—
~o2rnzooo~ 0’o~
oo~
00
CA
.úo
h~
0-
-o
~2.2~
~e
~o
ccn
SO
cn
o2
~.~
——
—en
—2
6-~
22
2‘O
S~
~g=
~>¿
9o
a~
o~
.-o
c~
8E
~~
zó
~cc~
o~
z
0%enQomt
mtmt
tutmt
•1<tt
PCUaU:
MCUU
Mi‘o‘o
‘oE0.
ut3..t4?
EocE-
(e)Cd
uunu¿U
C0OE-
o>4>u,caa.
UnE
—o
o.~o
o)o
56
>~—
S~
o~
cd
E~
•~
z1-,
EE-
‘4<.’

e
st
st
u,
e,
*
Tabla IV.8 - Recuentosde A. sobria CECT837 sobrelos mediosselectivos
con diferentes concentracionesde antibióticos
Medios Anlihi4licD
TSA
CQnrentad~n Rw1~
U9g~/mD
8,01
r~mpuadón
100
Ampicilina
Ampicilina
Ampicilina
Ampicilina
Ainpicilina
Anhpicilina
Penicilina
Piniaricina
Penicilina
Pimaricina
0 mg./1.
5 mgJl.
10 mg./1.
Orng./I.
5 mgJi.
lOmg./L
o U.Iil.o mg./1.
o u.ui.10 mg./1.
7,992,91
2,74
7,72
7,452,64
7,93
7,92
99,7536,32
34,2
96,3793
32,95
99
98,87
AGAP* Penicilina
Pimaricina
100.000U.L/1.
10 mgJl.
2,96 36,95
*.. Conceníraccionesdeantibióticosindicadas enlas fórmulas originalesdelos medios.TSA.- Agar ulptnnasojaAAA. - AgaralmidónampicilinamA. - Medio de Ryan (Unipath)
AC,AI’. - Agar glutamatoalmidónpenicilina
AAA
AAA
AAA*
mA
mA*
mA
AGAP
AGAP
150

Tabla IV.9 - Recuperaciónde A. sobria CECT 837sobre losmediosmA y AGAP
condistintas concentraccionesde penicilina G sódica
AadMúÉu Can~mntradñn En f~rccnU&dt
fi- —
TSA 8,38 100
mA Penicilina 0 7,85 93,67
mA Penicilina 25.000U.I./l. 3,9 46,53
mA Penicilina 50.000U.LI1. 3,2 38,18
mA Penicilina 75.000 UJJ1. 3.21 38,3
mA Penicilina 100.000U.L/L 3,17 37,82
AGAP Penicilina 0 7,84 93,55
AGAP Penicilina 25.000U.I.fl. 3,04 36,27
AGAP Penicilina 50.000U.I./1. 2,9 34,6
AGAP Penicilina 75X)00U.I.Il. 2,81 33,53
AGAP Penicilina 100.000UL/l. 2,93 34,96
TSA.- AgartriptonasojamA. - MediodeRyan (Unipath)AGAP. - Agarglutamatoalmidónpenicilina
151

st st st st st st e e u, e st
cgt, O 3 O E tJQ
1 1 15 5 2. 5’ 5
‘o u ~11
o-t
<~ o u-
.
~E.
a
33
oO
k3
1”
—O
wu
,
1> 39
OO
;>
uo
E
‘o—
——
u 3. Ml o’ 5 3? 1 o. o. o’ 5 a (t o ej m ej oc (ja -a
152

en-
-
oc‘n
cc
‘Cr
-
eN~r—
oc
00
00
00
EE
Eo
‘n
E-
E
IIji
E-
(e)mt1~u1uUMCCda=1<¿U‘oMi
¿UU4)
Ma
A3UE-
0’mO
Ce-
e-
OC
OC
en•en
ene-oc
00
00
EE
un
E•114(e 1iE¿U4?
1d
>,
~
153

e 5 * e u, st u, st u, u, u,
—o
.
~tJ
re ~E
.-‘
LA
iCA
CAO
aa
OQ
~Q
¿lo.;
~9 o’ E
a,
—M
u>u>
O-.
.>—
aO
U(7
0LA
LA¿l
~(7
0$
¡2. 5
’
Ml e E
M U o. o’
Cd a tu,
—ej m ej Qe cé
J-a
154

~e-e-N
Ce
-en
—r-4
cd
e¿
enen
enen
oe-
ocr4
r~e
4e
4
00
00
00
00
00
t~O
00
00
~0
0~
00.n
v~00
O«
fr~
Oc<
~O
0000
00
00
00
00
EE
EE
EE
Ocii[-4
e—
<¿
Ecu
~.E
~
‘~u
.
CC¡
HE
~
tu’14?M
a¿U‘oUMIMOa4?a.’a=U¿UTME4?1~.0=U.2U¿U=1<¿U‘e•0U8.’4?¿UU4?
Ma.5UUE-
JIjitAau•111u1¿Ue00E
-
Q
155

u, st. u,. st a a u, e
ej
EH
HO
~R
-a O&
cu‘<
n
Lie
~.u
~—
EC
ALA
CAO
E4
Ea
Ea
u
¿l0
(70
¿lo(JQ
¡u
~i~
tMl u E o.
e0
wu>
~u
00
(70
(70
00Cd
4
~2.
Ql
uO 1 9 0
Iv>.
wu
J—~
-JO
u>—
9 O ~1
O ¿U u‘o
0’o.C
AC
A-J
—R
stJ
tJ-
8r
__
__
__
__
__
__
__
__
__
__
__
__
156

00
—u~
0rl
0ci’
.n0
ci’—
—o
oo
o—
o—
e
00
N%O
N0
en
N0
00
‘erl
0—
O0
00
-4ej
-4e
00
0~
O0
O@
00
00
0<
n0
0N
O0
0O
00
en
~N
N~
00
tN0
r4
00
00
.3N
un
Cn
NS
(flt.40
WW
1t4
u—
‘-uu
-,
Os
rlO
rlci’
e—u~
ununu
nu
nufl
unun
un‘~
tfltfl
2un
unso
ent-.-4u—e’
-4ej
en
rlo’
-E
a4
>0
4>
un
-u.5aeu
e’.5
¾e
Miii
u‘aaEtui
Ea.1~ea
Mt
en
ue1uu
eMCgu.3aea‘uurPCuuu,‘uTMCuu-
‘a.30>1¡.3.1MI
14>u-4>u‘u
MtaO1!‘4u!
~1
CMCM0,uuaE§3..1>-Mc
1•1
4>PC
6
.5PC-ae•0OE-
M1.4
ca
St
u,
03
1EeE—
oQ
‘5-7

u,u,, st
st st u, u, u, st
e
u E
ej
rs,
4-
E
AIv.
>A
&
00‘A
00
00
OS
000%
0’
4fr
4C
h
1414
14
14
.AA
tu.>
~S
n
~1 A 00
-4 ate
ce
w~
O0
‘u.-
•O
——
——
——
——
—A
te—
14te
—14
CC
te0
0
O0
00
14
0—
0te
00
00
00
00
——
140
00
O0
00
—0
—te
U,
00
-.O
—14
te0
14
tu.>
‘4tu.
>0
0
O—
00
t~0
01
4—
——
teO
Os
40
144
14
00
15S
Ó o u e, tu’ e, e, u
0, E’c
E
CM
E Eu
E ‘-u
e, 1 e,A 0
0 Iv>
0
‘90
0
u -49
o’
a4’ a. 4’ ¡
——
—
¶1
a 2
A ‘o -u *
•1
44
A-’
-u
--u
$*
I u u
ca ‘a UF a u = 1 ¡
u
e1• 3 a’ a U -4 e o a
.u u a
’e
a‘a e u 20, CM
ue 1 a ‘a o
te u’
ti. te * -4 o’
‘-U -4 -4 le ‘o * u’ o’
ti, t E.
tu, a

00
rlU
en
rl0
en
ea—
0rIC
O0
)ci’
—0
—0
00
0
00
0ea
v-~
00
C0
00
00
40
—0
ci’
00
00
‘e
00
un
2rl~
—ci’t~
rI’o
N—
0e
n.-
4e
-4
O@
ti
oo
2r—
cn
~rlc
rlrl~u
ne
nrlr--’e
—ri’o
-4
u‘a
It~e’5
1u
~8I&
aE
~o
&a
2~
~<
~E
E~
o=
oS
<~
tQaE-~.>
CtD
.~”
0Q
~tQ
o>
4’
4’
159
uPC8EE‘Mc
o’0~
ea
Mt~1
ugt-.-.4
1
1MI12e‘ua4>u1uuOs
8.14>PC1u
.1‘11ta
-
‘McO
s0,
~1a‘ou‘usu2(5
E-

st
e’
st
e
st
it’
st
st’
e
st
st
st
a
Tabla IV.18 - Recuentos de los controles de suero fisiológico,
contaminadoscon seis cepasdiarreicasy seis alimentarias,no sometidosala cloración
Cepas de Aeromonasspp. Origen Recuento(ufc/ml)
A. hydrophila6A. hydrophila9
A. sobria 11A. sobria 13A. caviae16A. caviae18
A. hydrophila55A. hydrophila96
A. sobria34A. sobria 92A. caviae54A. cavíae97
HecesdiarreicasHecesdiarreicasHecesdiarreicasHecesdiarreicasHecesdiarreicasHecesdiarreicas
CarnedecerdoMejillones
Carne de corderoGambas
CarnedecerdoMejillones
160
2,26x 10~1,20x 10~4,30 x 1023,10 x8,54 x 1024,60 x 102
8,lOx 1022,47 x io~4,40x ío~3,02 x 10~1,17 x 1023,67 x 10~

O\Ifl
—~
e-c
ce
-rl—
‘nO
CO
60
O—
un
enCl
0’
‘1•unC
la
’
rlOC
e-e
-e
-so
tnn
ce
e-
rn~
.o
66
~
rl‘n
~~
so‘c
e-
.en
66
~
Ofl
‘05
0‘0
50
00
00
oce-e
-e
-—so
r-0%
50rl~e
nt
——
rl’nso
soso
so
50
W)
00
Ue
-cc
a%r-.
1*~‘c
cc
‘r,so
ci-
—cc
‘au
n
oo
C.C
o•C
.~s
•~s
so
Oc—
enrl
rna
’rl~
ene
-oc
11Voc
ccoc
6
oo
oo
e0
co
c•~
s•~
s~
~~
o>e
¼ococ
ocen
OO
Ce
-—
rle
n-Ú
00
04
uCd
QzoUQl
aai•14*M
a
10.’a4?00
o
1~
1>IDMaUi11a
Es
•1si
4?
11
‘II0<
.?1.’
=
Fi
UMa
*•~
uq
u~50.
uU
~
u,
tie-—•1III1‘o
tdLo>u.—
oaE
a
iiiUn
u-
St
sHe
o
4?¿U‘o
aUU—
‘
1-C
M>
42
o
E42E-
161

uit it w st
st.
st st’ u, st a 54 a
a’
4Iv
>O
CA
CA
CAC
A
u>
u>0
00
04
44 oc
gS
~s
oo
oo
4 00 o o
——
wtJ
:o~
oc
o Ml 20
0W
L>
ON
O’
O’O
s
Ma
—-4
...2
00
oc-J
o Ml z e
00
00
ON
O’
ON
O’
Ma
.JC
A
zz
so
e
MO
?P
P O’
-JO
’
u-O
-J0
0-J
u>
1C2
a o. a o,
E-o
”
u, rl’
u-
——
<tu za’
e<tu
iii
nR
~
<tu<tu
<tu
oE
’~ It a o. <tu~
—wo.
—<t
uo
—ctf
l a o
o 2 o Ml Ql o a W
C O Cd o Ml a.
jo’ La
-‘O
~ —9
Ma
o’
n.a
O ja d Co
.0
’0o,
Ma a 1< O

o
rl’~J’
co
so
66
—‘n
ce
e-0
0
<srl
soso
a’t-
e-c
cu
nso
66
66
co
00
00
~un
so5
0so
so
un
en
-c45
6—
—
rl‘c
e-
o’.
‘r-
rlrl—
——
00
Orl
soso
soso
so
so
enufl
en
—
——
ocso~
o~ce
o’
~
oo
t00
OC
enen
unen‘4
00
ocO
C‘1~
•t:~~
soe-
ti
0.4
UCd8ozua..aU0.
8a
dIQl
u,Ea-t4?MaU141Eu
!
¿UU
‘O41It
1-14?no
0~Ma
CCuo
tt‘o
E-E
a¿U‘14?‘a‘u0..¿U4>
¿U4
?’Q
CM
u,
su
s
—ie
IIII~11<iu~ou
-su
emi.,
Ou,
ej
tnOC
163

fr 54u, it st. u, 54 54 9* u 54 e 54 54 e u 54
u —-4
o”os
00
00
oo u>
r...>
—so
~.
oo —
—u
>tJ
LAIv
)Iv
)—
OCA
05 u>
O~
ii
e, ¿U o.
u o. o’
00
00
OC
OC
~.
oo o
0
(AL
A
o~
1.o
o
so
r5
00
0-J
CA
;>.¡
~..
bU
,O
CO
Cu
>
O’&
O’O
’O
NO
’O
NO
’O
NO
’O
NO
’2 Ma
u>
th
‘-4
?’>
CA
OS
—t
CA
O0
0
——
—u
>
u>
00
—-4
CA
t~J
tAo
.t~
.tu
.,
—-4
¿“‘u
>te
bu
>L
A
o MI z o
h>
tJW
OS
AO
NU
>O
NO
’O
NO
’2 ‘u
N>—
o—
oo
co
pp
pb
cbs
o~
oj
bo
g~
gO
LA
Oe
CA
‘oO
O~
LA~
~ag
Ou
>5
0
00
50
so—
09
05
OO
-J
09
00
00
‘o—
‘—aL
A
00
so$
csO
N>
00
00
00
00
164
2 e 2 o Qe
e e Ce
E, e> Ml a.
St
tu,o.
e cO
D ~ 85 9 ‘E j
O. 1’.U ~ E
n~
o.o
—
0~
50
si
o.<~ o. o
AM
¿U Qlu
-‘e ~ tu a u rl-
oQ
l
‘e¿U II ~ E, II 2
%
‘ao
’
u>
—
CA
CA

66
00
00
00
un
~
66
‘o
~11atu>
‘-u
‘0‘0
‘0‘0
‘0‘0
‘e‘e
‘O‘e
‘O‘0
‘e‘e
‘e‘e
‘e—
%.
——
——
——
——
—
OO
Oo
eo
wq
enea
00
0)
0)
0)
Mt
‘e
rlti’
e”
ci’—
Izo
.0ci’
‘tcT
h‘O
un
cr4
NN
‘0’0
’0~
’0u
wu
’O’e
’0’e
’e’o
’ec
——
——
—-..-
—‘u
.--%
-~-%
—~
-%‘u
.--‘u
-‘u—
OO
00
00
0)W
1C
t0tt’o
’eU
‘00
00
0r-flu
na
sti’
ZZ
ZZ
ZZ
6Z
ZZ
ZZ
6rl
‘nr
- nr-e
no
ó6
66
un<‘4rlrl
cc’0
ccc’e
wu
’ec’0
’e’e
’e’e
’e—
——
—‘u
-—
%•u
~‘u%
.—
—%
-—
‘u-.
—“u
-—
oo
oO
oo
ee
e.0
O—
—..e
<~
5<~
5
‘0‘0
‘0‘0
‘0‘0
‘e‘o
‘o‘e
‘e‘0
‘e‘e
‘e‘e
‘o‘u
-—
—“u
--.-
—-.
——
——
——
‘u—
——
‘u’-
oco
cco
elc
oe
co
en
en
en
en
en
rl
rICO
——
CO
flur5
o6
o¿
c’4
o6
600
——
rlti’
rlci’
ci’ci’
ci’
m0<
000
000
0000
un
un
un
un
q~
un
un
un
un
un
un
unun
unun
un
un
dS
oo
od
6S
SS
SS
dS
do
o
UQ
QU
WQ
QU
QU
Qu
.~u
-o
oo
oo
oo
oo
oo
oo
oo
oo
Mt
MtM
tM
tM
tM
t-r
nM
tM
tM
tM
tM
tM
tM
tM
tM
t
—ti
Ci’M
t‘ifl
‘000
0’
—C
iCi’
Mt
un‘0
tiII.1ufra
oc
¿UoUa2EocJ
1aUa4?1-
un1d
.Cd05a..
4?‘uu1eEiu!UtuID
a
‘VS-1¿U4>
rj.e
j0>
1-
—E
U’>
Ua41UQl
IDLbauCM
toME<Ib
4?PCaoaoCM)
U.u
500
~O
u>O
~g
o.
.‘o
.0L’.~‘~I*.iE00
~‘E
4~‘~
~
‘6‘~
‘~
~~M
II
1~
o—
o-
~
z~
á
a’‘fr~e—
o
3M
1it134
s3~1Ue
’
e’E
165

u, u, st
st
u, u, st u, st
u, Mu e
~jJ
~¿>
•~>
~j>
~jJ
<,4~
>1
41
41
41
41
41
41
41
41
41
4—
—0%
“A‘A
te1
4—
0‘0
00‘-4
OS
~A‘A
te14
05
00
0
‘A‘A
&‘A
‘A&
‘A&
‘A‘A
‘A‘A
‘A‘A
‘A&
&‘A
’A0
00
00
00
00
00
00
00
0
nfln
nn
fló
nn
nn
nn
nó
n
0000
0000
0000
00‘-4
~O
S0%
0%0%
0%0
’
‘st
ee
ee
tco
oO
OC
140
00
00
0-L
‘u—
—‘u
—‘u
‘u—
%‘u
-.‘u
.-‘u
-%
‘u%‘
u--
--‘u
----
-‘u
—%
‘u
o’
0%0%
0.
o’
0,0%
0%o
’Cm
0.
0%0%
0%0%
0%0%
0%0
.
ee
ee
ee
oo
o.e
eo
oo
oo
oe
—‘u
-—
‘u‘u
..-
‘u—
—‘u
——
‘u‘u
——
——
—‘u
—-‘u
‘u—
‘u—
‘u—
—
0.
0.
0.
o’
0.
0.
0%0%
040
.0
.0
,0%
0%0%
0%0
’0%
0.
14
14
14
14
14
14
teIv
)1
41
4
CA
OS
teO
(~0
%0
01
4O
kA
14 00
teA
tete
tete
OO
tete
&14
00
00
00
U,
—‘u
——
—‘u
—‘u
‘u—
’u-’u
-’u
%’u
’u~
“u-
——
-—
‘u—
‘u—
‘u—
0,
0.
0.
0.
0.
0.
0%0%
0.
0.
0.
0%0%
0%0%
0%0%
0%0
.
tete
te1
41
41
41
4te
—te
tete
tete
00
014
1414
00
00
oo
a—
‘u—
‘u—
-..
‘u-.
—‘u
-’--
—’—
’u’u
‘u—
%‘u
”’u
.~.-
’u’u
-.’u
~’u
‘u—
0%0
.o
’0
.0
.0%
0%O
S0%
0.
o’
0.
0%0%
OS
OS
0%0%
o’
—9
99
9—
4.
0%-<
400
Z~
tQn’
’.~’A
Z~
ZZ
~te
~N
CQ
C?
nO
tnN
)
ozz~
—4
166
u zn o
—
o.8
a’
~O
¿Ua ¿U
o.E
g’~1
<~
o
a-u
n u ‘a It”.o 2
u.-,
u
u’ E E u. u
0.0
—~
~~
IEe O
’
ee
—o tu’ o
.
o oe
,.
0% 0, o ~.
~M~ tu S
fte
<‘‘—
‘oe
oe u ~g.
o~ o.
—n
o‘o
‘e
o
o-’
-I —H
o—‘
2—u.
== o’
eo
o— o’
o. LE’.
o0.
e, Ot o.
2 Ma a a
r o 9 u CA a 4’
PC E u’ o o 4’ ‘a u
Ma
2 Ma E EM a o, a
WC

Osr-
ej50,0
00
~ji
ul
eC
Oouu-’
‘0‘0
‘O‘0
‘0‘0
‘O‘O
‘O‘0
‘O‘0
‘0‘0
‘0‘0
‘0-‘u
——
----—-—
-—-—
-‘u
-‘u—
rlrl
—0
00
‘00
)0
)C
l—
0‘0
‘0‘0
0)
‘0
‘0‘O
‘0‘0
‘0‘0
‘0‘0
‘0‘O
‘O‘O
‘0‘0
‘0‘0
‘0-‘u
‘u-
——
‘u—
-“u—
—-
—-
-‘u--
‘u.-
‘u——
——
—‘u
‘00
)04
00
0‘0
‘0‘0
b00
)‘0
‘0‘0
‘o‘0
~rn~
rn05rnIe
4I’e
LP
2
<n
os
~E
fl«~r—
—
oc5
0rn
vo
su
nv
O”
MtC
i’.Ó
CO
OC
O<
i’
Ud-r~
r~e-
i~<ev
n2
Q~
2to
~cj~
u~
¡~
~iajocícn
——
—~
——
——
—
1~
‘0‘0
‘0‘0
‘0‘0
W‘0
‘0‘0
‘O‘0
‘0‘0
‘0‘0
‘0-‘u
-—
‘u—
-‘u-
‘u.-
-‘u—
——
-‘u‘-‘u
‘u—
—‘u
—‘u
‘u-.
—-“u
~ftC
0OO
ffi~ftN
CO
’0’0’0’0’0
0%‘e
<“o
.
‘0‘O
‘0‘0
‘0‘0
‘0‘0
‘0‘0
%0
‘O‘0
‘0‘0
‘05
0-‘u
-‘u—
‘u‘u
-’—
‘u—
‘u—
‘u-—
-—-‘u
-‘u
‘0en
eo
oo
‘0‘0
‘0‘O
ene
‘0‘0
‘0‘0
‘0
Ei~r~i~ir~
LZLZIZJr-LZJ
‘0’0
’0’0
’0’0
’0’0
’0’0
’0’0
’0’0
’0’0
’0—
——
——
——
——
——
——
——
——
NrlrlN
rlN
NN
cr~
NC
irlN
~‘0’0
’0’0
’0’0
r’u-r-r-r-r--
unun
unun
unC
OC
OC
OC
OC
O
unun
unun
mr~un
uq
un
un
un
un
unun
unun
unun
oo
oo
oo
oo
oo
oo
oo
oo
oo
occo
oo
oo
oo
oo
occo
unr—
r——
—
0.4 ca
u1o‘a11
¡1¿UzEa14?64?4-U¿U4?6.4-1dIQ
lU
,
3.’.
‘<4?MaU1¡•1ULb
o—
¿U41>~u
u¡Q
l
Qn
-
rj~
—u-
¿U
’>E
-t
u141
•1U0<4?CM>9Uu0.
8oE8ID¿UO‘a¿UOCM)
n‘e00
5~
93n04e
u>93
930
:2o
b
~u>I
u>o
~u>
E-’-
ue
j.
-~o
’00
ti-
‘~1
.~0O
u-’
Iltia
u-~
u,~
‘eu
E-O
‘E
u>
’
y
040
1
‘>o’
u>‘u>
~1
‘~‘g
~L
Eu>
‘jt,ZC
~
E-o
o
caEj
U—
¡“1CM¿Ue
’
u’—
elEE-
167

Mu u, u,. u,, st’
st
st 9* st u, 54 54 e 54 54
CH
e- a R
~o
’—
?2.
o‘a
oo
a’
o-t
~ gfl
OH
t ~~
Á’~
~g~~
.g,
ni’
00
3
e,O
0.
o.
9.~
‘~
EE
-~o’
oo.
~e
.0‘u
’
oo e
o ~ti
-‘‘lo
6’~
E
~Z
o.
~ —o
’O
=3
3e
’
o~
, o
Raa o.
u.-
e, fl’~
tete
tete
tete
te14
14Iv
>14
1414
1414
1414
—‘A
te1
40
50
-40
’a’4
te1
40
50
00
——
——
——
——
——
——
——
00
00
00
00
00
00
90
00
3 o00
0000
0000
00-J
CO
--4
0%
0%
0%
14
14
14
14
14
14
14
14
14
14
14
14
14
14
14
14
14
14
14
——
——
-4
-L
-L
—-4
0%
05
0%
0%
0%
0%
0%
0%
0’0
%0
%0
%0
’OS
0’0
%0
%O
50
%
—rr
rrrr
~n
i2
~~
I~I’~
I0
’0%
OS
OS
CW
S0
%0
’05
00
00
0O
N0
%
‘u-
‘u—
‘u—
—‘u
——
‘u%‘u
~~•1
~~
‘—‘u
~’u
‘u—
—‘u
—‘u
—‘u
‘u—
0’0
%0
’0%
0’0
’0,0
,0%
OS
Om
0%0%
0’
0%0
’0
.
u>~r
ri-u
rrrr
i~
~Iz
Iww
~I~
14
—
Iv>
—>
te’u
-ItJ
--J
‘u
Li ‘M
o 2e
u
u o 9 u 2—
teO
S0%
0’
OSe
e14
‘AU
!te
00
00
C“
0.
-“u
-‘u
‘u-’
,‘u
,‘u
——
‘u—
%.
‘-‘u
—‘u
—-‘u
‘u—
——
‘u—
‘u—
‘u‘u
-’—
0.
0.
0’
0%0
’0%
0.
0.
0.
0.
0.
0.
0%0%
0%0%
0,
0.
0%
14
——
I¡J¡h
¡WC
ti 4h.
~
te—
LA
FN
~0
0
--4 ‘uÁ
LVtu
tu~
—m
uf
7W—
‘--
rW
~L
LW
áL
je, 4’ Mc u
0%O
S0
’0%
0%0%
te(f
i0%
0’
0%0%
00
00
Cm0%
0%—
‘u‘u
—‘u
—--
-‘u
-‘u
——
‘u‘u
—-—
-—-—
-—--
‘u-—
---
‘-“u
‘u—
—‘u
‘u-
0’
050%
0%OS
0’
0.
0%OS
0%0~
OSOS
0%0~
0%0
.0%
05
14tu
.)—
——
—
u>
0%
I0%
IIv>
I’0I
UL—
JL
JJ~
&t>
u>
——
—‘u
JLflb
>‘u
J0
0<
40
’‘u
-.)
&‘A
0’
0%0%
0’
—14
teU
,0
.(f
iO
OO
O‘A
te(f
i-“
u—
‘u-~
--.’-’u
-~%
’u~
‘u—
—.
‘u-’
—‘u
‘u-
—‘u
‘u-’
‘u—
—‘u
—‘u
‘u—
0.
0.
0%0
’0%
0’
o’
0.
0.
0.
0,
0.
0’
OS
0%0%
0.
0.
0.
3M e 4’ ‘a u
Wo
ze
fWF
¾p
99
ET
S1
~,~
~~
sp
no
,—
o, oe>
~o
a’
~E
,¿U
e>u
¿Uo.
Eg’ u—
,o
168
‘u u E. ‘u u EM O o, a u

ti’O
Cl
CO
OS
~C
OO
S’0#C
l&
S&
oce
0’r’uCO
¿UaOu
‘0‘0
‘0‘O
‘0‘0
50
‘0‘O
‘OO
‘O5
0‘0
50
‘0‘0
0)
Mt
Cl
—0
0‘0
‘0‘0
0)el
—5
05
05
05
05
0
—r—
ci’cn
to
50
ej
ej
~~
Mt
Mt
un¿
‘u-
ci’—
——
——
<‘4M
tC
iC
lC
lci’
——
——
Cl
Mt
‘0‘0
50‘0
‘050
‘050
50‘0
‘0‘0
‘0‘0
‘0‘0
‘0‘u
——
‘u—
‘-‘u
‘050
50U>
00
5050
‘0‘0
‘0‘O
‘0‘0
50‘0
‘0
U«
%C
OC
O50
O-4
<n
r-ON
Z‘a
N
<n
cn
oos
Mt’0
Or—
‘0C
Or—
Clu
nN
un
OS
O~
~v~
MtC
Or-r’u
CirIO
CO
N
O,O
Sr—
CO
0%
—N
O%
0u
nr—
Cl
‘OC
50
’O’0
50
50
50
’O’O
t’0%
O’0
’0’0
50
—-‘u
.-’.-‘u
‘u’-
——
-‘u—
-‘u—
‘u‘u
.-‘u
.-—
——
‘u—
‘u
tMtN
elo
O’0
’0tn
ten
ft50
’0’0
50
’0
——
——
——
——
——
—
5050
50‘O
‘0‘0
50‘0
‘0‘0
‘0‘O
‘0‘0
‘0‘0
50‘u
-’‘u—
—‘u
‘u-
—‘u
‘u—‘u—
‘u—‘u
--‘u
--‘u
—----’u
’u’u
’u--’u
‘05
05
0‘O
00
50
50
‘0‘0
50t
‘05
05
05
05
0
r-CO
t~
:un
CO
os-<
i’
~o
oo
oco
osZ
~o
oC
O0
’oso
s
~
0<C
OC
OC
OC
OC
O
unun
unun
unfl
unun
unun
unun
unun
unun
un
oo
oo
oO
OO
OO
OO
OO
OO
C,CC
O~
0000
CO
CO
CC
00
00
XO
CO
C0
0C
CO
CO
C<‘4
<‘4<‘4
cici
Cl
Cl
rlC
iC
iC
iti
Ci
Ci
<‘4
Ci
<‘4
20.. 1~1ut!N
uPM
¿UeU¿U:5E84?1-uU3¿UID
¡itOs
01a..IDeMaU1eEi•1UCM?
Cdo
4?
Lb
Lb
~
es
¿UejQl
el
—a..
c~
o.0
Lb
E-P
C
Ua¿Uu1MIUCd41Lb
.11uCdtouLb4?‘¿Uu¿Uo¿UOQ
53
93
u“u
‘93oa
¡C
CC
u>u
>—o‘8
u=
—ZAg
(5-
a.0
‘~5
Iii~ij1~
‘6’U’~
1‘g
eu>~
aU
~o
—u
U4>
las
O~
,~
~;
@~—
E~
o-E
‘
‘~‘.uil~
,u>
mu
’,t
o¼
C93
u>-
.~E-
~6
1~10)uPMu
¡4uuu’—elEE-
169

e, it u,, st st it. u,’ st st st a Vb e -4 e
tete
u>
tete
tete
14
14
14
14
14
14
tJ1
42,
4.te
14
—0
50
00
‘.40%
kA&
te1
40
50
00
e, E tmu
0000
0000
0000
00kA
kh
kA
kA
AA
——
——
——
14
——
——
—&
&tA
u>
’Ate
u>
C%
A&
U>
~14
’u.J
tA0
~‘~
~&
s&
U,0
’0’0
’ON
U,0
%0
%0
%0
%0
’00
0’0
50
.0%
—‘u
-‘u
--’.—
----
’u-
—-
-‘u
—-
‘u—
—‘u
O,
0.
0%0%
0%0
’0
.0%
0’
0’
0%OS
0%0%
0%0%
0%0
,0
’
14
Iv>
14
~—
te1
41’
——
~u
>u
>1
41
4kA
te1
40
00
00
’kA
14
50
’u4
CO
50
Z~
0%
u>
LA
MkA
‘AU
,O
S0’
Os0
’te
‘A‘A
(fi
0%0
’0
0—
u>‘A
Ch
0,
——
—‘u
—‘u
‘u—
“u-
‘u-
—“u
-.‘%
-—.—
——
‘u‘u
——
-%‘
u%
.—
0%0
,0’0
’0
%0
%0
,0
,0,0
,0%
0%
0%
0%
0,0
%0
,0,0
,
——
00
00
h.~
.00
OC
r,’
14
0
-osp
po
s-’J—
i~‘O
—-J
otA
’01
4te
.Cp
p‘A
pp
pp
po
p
‘u I 2 ‘~u
rl u’ MI u’ o ¡ 2 o u
00
0t.n
l.n’t’
uj
——
teN
>1
4-L
4.&
ZZ
kA
50
50
14
~O
S5
0Iv
>kA
Oti.
~tn
~¿
>&
&—
kAV
m1
40
0te
’0‘u’
41
4—
JO
s0
0-4
kA
0.
0,
0%0%
OS0’
0.O
S0’
0%0%
0%0
0—
0%0%
U,
0’
—‘u
—-‘“—
—“----“
—‘u
-‘u
‘u—
—0
.0
.0
’0%
OS
0’
0,O
S0
’0’
0’
0%0%
OSO
S0%
0’
0.
0’
14
14
14
k”
14
14
(Ji
O’
OS
OS
0’0
’0
ti0,
0,
0%0
’o
oe
u>ti
tiU
!—
‘u-‘u
-’--
-—--
--—
‘u—
—-
-“u
‘.-—
‘u’u
---’u
----
-—
‘u—
‘u—
‘u
0’0%
0%0’
0’0’
0’0
.0
.0
’0%
0%0%
0%0
.0’
0.0
.0
,
00
00
N>
00
00
bb
’oo
ll’k
Ae
’A5
0N
>o
oO
Su>
u’ O u CA e, 4’ PC E 1” e, 4’ ‘a u WC
00
00
00
00
00
00
00
Z—
t~7s
m~
e~
50
0C
b4
-4-I
D<J
u‘4
ZC
4.tI
50
0’
00
-a4
.0
’-‘4
4u
.u
o,
no
,
a:a
l~
E,
¿Ue>
~a
O’
u.—
O
o.O
H-, fl~
7o
—‘
‘—O
=e,
.03
E—
o
o -;~
‘~
3 ow
O’
o 3
a o.o
<1 oo o
-
a oo’
3o
.—
o
o~
o‘~
93
o
o-.
Oti o
=3
—o o’
30
O0
’o.
=o os
o —o
cf
Ma 1 E. a a E,
Ma 1 E E, o, a 1
ViO

00
00
00
00
00
00
~tC
OM
tr-~50N
N
50
00
05
<‘1C
OC
O
0~
0c.c
¿U
dcc
‘0‘0
50
‘05
0‘0
‘050
‘O‘O
‘0‘0
‘e‘e
‘O‘O
‘O‘u
-%‘u
—---
—-‘u
—-
—‘u
-‘u
00
00
00
0—
——
00
~fl
‘e‘e
‘een
3r-o
Cl
‘a—
—
‘00
00
<n
r-CO
t4r
4
‘O‘e
‘O‘O
‘0‘0
‘O‘0
‘0‘0
‘0‘0
‘O‘O
‘e‘O
‘O‘u—
-‘u-‘u
‘u-
‘u——
——
——
——
‘u.-
——
——
en0
)‘e
ej0
0U>
‘0‘0
‘0‘o
‘0sfl
un
uo
un0
)
U
ost
‘nr’-
c’S
oc
0’
Cle
nO
r-en
r’.~
‘0’00%
Cfl<fl
Ci’C
lC
l—
‘nM
ten
r’.en0
~
CO0
—,
,N’0
’nC
O<
nrlO
S—
—~
un
0%
:rlc~r—
c~
‘0’0
’05
05
05
0’0
%’0
’O’0
50
’O’O
’Ot’0
—-‘u
--‘u
..-‘u
-“u%
1~—
——
——
——
——
—O
O0
00
0—
—e
o0
0‘O
‘OO
——
——
——
—ti’
‘e‘0
‘0‘O
‘0‘0
‘O‘0
50
‘0‘0
‘0‘O
‘O‘0
‘O‘e
‘—-‘u
--’-‘-------------—
—-—
---‘u—
‘u—
—-
‘u-’
—‘u
‘Oó
‘O<n
00
‘0‘0
0‘0
‘0‘0
‘O‘e
‘O‘O
‘O
CO
r—te
n
——
—C
lO
sO
SC
O0%
00
0%CO
00
0.
——
44
40
04
44
44
40
00
00
00
00
00
00
0
uzu
50
50
50
‘05
050
3.-—r-
(‘u-
1’.-r’u
3.—Q
lCO
COCO
CO
un
~w
~u
nu
nu
nu
nu
n’n
un
~tu
nu
nu
nc’o
oc
od
SS
d&
d&
oo
oo
un
CO
un0~
oo
oo
oo
oo
oo
oo
oo
oo
oe—
r-—e—
r—e—
e—i’u-
e--e—
e--e--
e—e--
e—e--
e--e-
-en
enen
enC
<’uC
eC
i’fi’
Ci’
Ci’
Ci’
enC
i’en
enen
en
ne00
93‘93
‘90
33
o>u>
~U’>
É-.~
1ZA
‘~.~
.0“E~‘ti•j~E0
0‘—
(5
3u>
‘O
(5~;
‘~o-
u>u,-
a’.~
,~
u>u>
tu-.
ca
3Mu’
¿UoUa:5Ea01¿UID4-‘6U4?¿U1itdI.4U,a-
ID-‘MceM
aU1iUCM>ID
‘uMa
~
u•—C
d
it00
04
>
u•141
‘uuCM41<IbaMIUCd
8o>1.
ME‘mi
4?PC¿UOo
1~13PM•1Uo’—elUo>E-
¡71

e, 3.’ it * st st 54 54 st “u.
o.0
—~
o,‘-,~
~;‘
u>
wu
>te
u>
Wte
wta
au
>u
>o
’(tu
u>
tete
tete
ta>
MW
—u.
,
~a
fln
nn
nn
nn
nn
nn
nn
nn
n’n
c~
~->
—‘F
~—
—0
o -a!
=~
za o
30
00
00
00
00
00
000
Ql
o~
‘~--
“‘2
e>9
3.0
o.~
&oN
>6
~1
40
~cL
~N
>0
—t~
co
O~
.E
g’—
4u.
—
1~
o9
3o
o4
.4.&
4.4
.&A
&4
.4&
4.&
44
.&&
&<tu
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
~0
00
0
~u’ H
o
0.0
00
UiO
,uta
•3 e’
Cm
Cm
0,0
,0,0
,0,0
,0,0
,0,0
,0%
QsO
s0
,0,0
,o’
MI
14
>-”
.-’
‘““t-
J1
4u’
0g
8O
a‘-0
0N
>0
~U
o>M
aO
teC
ho
’o’0
,oM
’Aa
U,-
Lo
oo
I.a
ae
c
‘u“‘u
-—‘u
...
‘u‘u
..“u
.%
“..
‘u.
‘u.
‘u‘u
.“u
“u‘u
.
c~
0’0
,0,0
,0,0
,0%
0,0
,0,0
,0’0
%0
%0
’o’0
,0,0
%<tu
ou
~C
OC
Op
p>
~O
~j~
¼fl
oo’
“2.
o’1
~g
00
¡C
A1
4C
fl-J
kA&
~—
J‘0
00
t¿>
oo
’
3o.
—o
om
——
—,
——
05
01
4N
>N
>1
43
.J1
4>
u..,.4
4.4
.&
Um
’~1
4’—
5‘~
z‘~
.ib
ob
00
14
00
14
50
Wte
—te
Vi
3U
,(Dm
ChCh
U,Ch
‘AU
,Ch
(J,U
,(D
m0
00
14
ti—
U,
‘u-.
‘u-’
—‘u
’--’
‘u’-
‘u—
—-
‘—‘u
—-‘--
‘u‘u
—-‘u
-—-
—-
——
.—
‘-“u
eo
’0,0
.0,0
%0
,0,0
.0,0
,0.O
,0%
0%
Qso
’o’0
,0,
‘a 3Z
A-’
~1
0&
tM&
tMtM
Oe
’t.J
teu
.’.u
oo
om
..0
13
3-—
—.
—‘u
—.
-“u
—‘u
-—‘u
-’‘—
‘.—
~‘u
—--
—’u
’u—
‘u-‘u
-—
.‘u
—.—
o>
0’0
.0’O
sO
s0
’OS
Oso
’0’0
,OS
0’0
’OS
o’o
’o’0
’E
•a
3½
o,
Ea
~‘0
O0u
>14
—.-
’—
ao
--.
~W
CE
O~
~
30
09
9fu.
--
-V
i4
.V
i4-
V~
4.&
.>N
>V
4t~
4.te
~4
~te
tOte
teti—
~Z
C0
Ct4
.~5
0t>
172

-u~.
e-,
OO
~¿
O~
no~
Mt
Oen
COu
-’u
-u
-u
-—
Oe-
Oe-
enen
(5e
no
—o
A-~a
e-ci’
--
‘O&
oM
t9
u-—
—u
-st
4>—
enM
tM
t‘a
1e-
OO
‘ua
-(4
~.
00M
tO
en‘u
Nu
-u
-u
-PC
~“‘,
Mt
~U
d’t’
-u
3S
rO
O
1os
u-
cn
tM
e’
e.
e-<e)
e’
OunII
~e
nt9
~O
unen
ena
u-
e.
e’.
5.
4r
rl,O
,
4>u-O
e-
flu
-‘e
ee
nM
t
A-‘~
e”.e
.e
.3
enun
OO
en0*
~e-
~2
4.?00
u-
u-
—u
-1-
——
rlu
-e
-‘a
(5—
‘OM
te-
e’~C
Oe
uO
‘o•
o1
—O
rlu
-u
-u
-u
-50
OO
—rl
1::
7!!
6.e
’e
’.Os0,
u-
u-
-u
-rlrlu
-.
004
-50
Mte
--u
-u
—’
a..un
O—
o—
u-
u-
u-
u--
‘u00
‘aen
flO
0,Ue
.e
.0<
oo
qS
9a
4a
~u,.,-
u-
u-
e-u
-
eC
l~
ai’
‘url
e-<e~
5’
e’
2u
-C
iu
-
•—
u-
2—
uII
ID93D
i0,
oc
—1
~
a
e-’-
‘uE-”
‘a00
‘0U
t
¡73

stit it. it .4 st st, st u,. st 54
u
PC
ah
~—
o’
—Ci
io, CM o, (tu E
u>
tso
~
1’u
>—
-J-4
uu
> tu’>fix
o’
PC a&
000
’-4
u.o, CM o, 4’, <‘tu E
O~
FJ~
o o, ‘u,
~xi
tu.>
i~o
ou
>2
051
4=-
¡x n
CM <tu o <tu <‘tu E o o o CM <tu o <‘tu ¿U o o E o o
¡74
-~n
~M
o~
~E
-qg
zzfl-~
nc~
’ ~V
E
~.
E’3
’<b
t0
’o
e.=
~
tu,o.
o~
,CE’
Ocn
q~
E’~
~‘
EE’ 2
~E’
-cg
o E’*
-c ~E o
1’ o
CA
-4 o u> o u> OC
tu.>
-4 o tu.>
<b
u
Ma
tmM
F>
~
~ “E Ql~
’~
E,
29 U
a E, e> o A o .9 Ql
a o’
9 o, Cd u a e E PC a’
E, ¡ Ma a’
ev
¡
tTI a Ql e 8 o m a a’
E,
Ql
e 9 o
¿Di. so

oA..
ej1>
-licIDC
lC
lM
ao>
•M
cun
enC
do
o~
‘i~
e-
A8
~soQ
e-a
Ma•
1C
l—
rl
188—
MI¿Ua>
Mc
•P
C
~ors
.c—
~o
Mu
1-M
a0
4
cao
6.4-
‘1•n
o8
“‘E
o‘~
‘uLb
1<Iba>
—•
Mc
o’
ir>o
—‘
0Li
un
1cc
—ID
so‘
o>81
en—
rl
aM
Ee
j
-1’a>
E~
‘a
IBe
ja>
a>M
cPC
Ma
.Mc
—~
ECM<
U0
50
’’>U
Vr
~E
-—
~a>
—a-1.u
4)0
EQ
E-o
H~
E-
175

st mt st
‘u.. 9* e nr
o--
u 2< —~
o’a
—O
oQ
l(D
io~
—o
’E’
<‘jo
,a
h—
~~ rl,
IIo
O
0’.c
~—
at~I
‘u-]
4e>
así
‘II
~g
—~
‘>—
~—
4t’JL
A¡&
(A4
—•
Osu
btc
4a
-‘4O
CA
u>
E,
•1~
s-.;
~1
——
aO
OS
u>U
>Q
l
~uE
,o
e~ oE’
~A o
e> 0~
E—
O4
tu’>
&4
005
’CA
9
OQ
la
2.
—0
’a
ho
,¡
4”o
.a
<‘tuu a
lE
o,LA
4¿
D~
(A
C~
~ ao,
a E PL
Ql
al
so
oa
~—
2~
—M
lO
’U
>‘u-
J‘
~0
OM
a
x—
—~
,~
eva
.O 0
4”
0’
4”
4”
¡76

-ti’fl
‘ti’
rlC
l¡
unun
e-
ococsoe-
e——
en
unS
e-
6soO
C
SO‘It4
Cl
Un—
—oc
‘It‘ti,
oc
o’
unU
2~en
e-
Qen
en—
so
soso
ocso
IXti
un—
so—
enun—
so
*so
rl0
%
o’—4o
—e-5
ci’
‘ti.ci’’oc
‘o—
—ocu
CMbE
-OE-
r¡5rsUuoEa>
McuPCUMcoA-04
oej
‘u-
E4>M
coaua>Cm>a>
PCoLboEa>
McoE-0
4a>Cm>;4
>,
PC
a..ID-licID
‘auCd8AID
PC
a41Ea.3¿U41buao01
Ma6bu
PI
.2,oLb
‘EtIDo?1-aa
.
=2
ji‘a1?<U
—U
en
.o
E2
~
oa..4?.Ma1~84)
Ma
¡8o‘a~1
‘oOE~
~>
‘u‘~*
-m
¡‘1~o’u
fl~
As’.
AO
ni
~o
~o
‘o~
OO
—xj
U.E
u-~o
>
Q’>
OEa>M
crsUPCU.0EO
s
1•7•1

Wc it’ tu, 54 st st e m
r
mt u, u,’
54,
a u u,
o
a
E o,
PC ‘1 O
ah
~—
0-
—u
,~
‘ O’
PC -t O E
—o
4
o
OC
—4
OC
tu’>
1’>
LAt’J
CA
0’
su
--
o u>—
u>
u>
—4
——
CA
a’
0’
LA
XI
Y,,
—-4
u>
u>u>
LA
4O
’o’
sou
>•t
ou
>so
u>‘o
~ PC
~ah
~C
M 4’~ PC -‘5 o e>
—O
—o
~ ru>
-~u>
LAU
,-J
-J
z
1¡
xi p ce
III
178
>~-u
’-
o.g
EN
O‘e
>
E~
Ls
o—
—6
~~
p-a
.o <tu e
~ E<tu
~E
~<tu
O ~<tu E’
O PlO’
Ew 8’ -O -it O’ <tu O’ “5 <tu <tu
u E u 1.)
>~‘ a al
a al
‘a MI o 1 —‘u a’ 1~ o, .4 E, ¡ a: a Ql e, ¡ 2 ‘a u Ql
e ‘1 05 ‘a 7
o’ 9 1’. o, o’
Pl e, It a
O’
e> o <tu e E. CM o, CM e> e, PC -i o E o -4’ o CM o o e> -e ~4
~ e E. Os
ci Os
o e, PC -t O o’
-‘u o o, St

o1E‘1O‘u11‘u4u,o
~>
‘o•n
•~
‘(5
Ql
01tA-
1>-licu‘aUC
dIDLb
A4?‘aaU0.
1aou,
‘u—
‘aU‘a(Ib«ejU:5uE
-
179

w Pr
Sr 54,
nr — 9* Su.
tu 5k a, mt
utr
lz-
-.M
a~
-:u
e,o,
E,
MI~
MI-~
al 1
e’=
o-
e,‘u
¡-,
~z’
a,~
—O
oQ
l
—u
.
¡Ql
Stf
lO.
O.
cG
.C
e>M
I~o
O..
oO
’-‘
e>“u,)
O-e
E,
u,
4”so
oMM
a’
MI2
tJt
LAO
’4
Ce
—N
>O
e>isa
—~
—.0
<e
u>b
‘ocu~
E—
ocu
>Is>
3’3
o4
‘u-]
4u>
gl
ce
-ia,
~‘
so‘
O o
OW
a’
—3m
——
so
~a
wOs
u>
o~
~u>
~4
u>u
—.4
oe.,
<E
Oa
’E
~00
——
E.
~ta
•ON
•o’~
uE.
.<~
—~u>
—a—
J3
2E,
o—
MIe
‘u.9
8Q
lg
0’
0~ 4”
CMo
’O
<tuMI
oev
a,—
e>o
0o,
<tuN
iff
ls.
.~
‘os
—rm~
)--
_-
StLA O
o— It
o’E,
.e>
<5—
oE
——
—so
E Me
O’
so—a
u<tu
u>00
u>x
suu
’b
O”
—<5
00
—-2
“-2e,
O3 o
oe,’
.00
0000
‘‘u.]
—l
0-
¡4”
O’
4”—
S’
~=
u.-
•u.
¿U¡
-a
’—
e,-<
oIt
u>tu.
>u>
—~
—‘
PC—
uo
’a’
oso
4”?<
~<
Ita o
-tu>
tu>
-4.-
~-A
-- -JO
180

soCl
o’
u.—une-e
’
en‘ti:oc
<n
rlu.—
enCl
e’
+<e>
enen
——
u.—‘-u
Cl
e’
en
o.
<d
oc
unu.-,oc
e-
Un
——
oc
oeja>~uE e-
o.
en—
o.
sosc
e-O
Oen
Oso
u.—
‘-uen
Oso
99
unoc
e-ocu.’—oc
<e>o
c
o.
uno
’C
lC
lo
’—
rl
e’
~..
o-
unCl
—‘-u
‘u—
—-4
e’
o.
oe
-un
unC
le-un
nE-
E-
4>~0’-’
04
>
4>
4>
Effi
88
Oc’,
:11~‘E
a‘9
0)O>0
0”
-no>
—u
-
O‘O~
to~
++enenC
l
+(1,
o.
oCl
“-ue
ne-soso
‘-uo-4
++
ebu4?‘abu84?
x‘E841la.5uQl
e
e14)‘at4?¿U8Q
l01a.-lb4?
PCuQl
A41‘aau¿UuUEIDaMaU‘uu
;Lb
Un
1U:5UE-
u1obuCd
oz¿U4?WC‘Eo
o‘mlID‘ubuoCm)
181

Mu
r
u,
Su
0~
Sr
tu,
st
st
Sr
nr
mt
Tabla 1V.36- Inhibición de la hemaglutinación de las cepas de Atromonas spp, enpresenciade D-manosa,L-fucosa y D-galactosa
Cepashemaglutinantes(Eritrocitos deconejo)
M+ F+ G+
M+F+G-
M+ F- G+
M+ F Gm
M-F-G-
Mm F- G+
M-F+G-
M-F+G+
Patrones de
inhibición
Cepashemaglutinantes(Eritrocitosdecordero)
M+ F+ G+
M+F+Gm
M+ F- G+
M+F-G-
M-F-G-
M-F-G+
M-F+G-
M-F+G+
Patronesde
inhibición
Diarreico
7
o (0,0%)
0 (0,0%)
1(14,3%)
5 (71,4%)
0(0,0%)
1(14,3%)
0 (0,0%)
0(0,0%)
5
3 (60,0%)
0 (0,0%)
0 (0,0%)
2 (40,0%)
0 (0,0%)
0 (0,0%)
0 (0,0%)
0 (0,0%)
Origen
Alimentario
34
0 (0,0%)
2 (5,9%)
4(11,8%)
13 (38,2%)
9 (26,5%)
6 (17,6%)
0 (0,0%)
0(0,0%)
6
0(0,0%)
0(0,0%)
3 (50,0%)
1(16,7%)
0 (0,0%)
2 (33,3%)
0(0,0%)
0 (0,0%)
CECT - ColecciónEspañolade Cultivos Tipo
M: Manosa:F: ¡teosa:G: Galactosa
Se consideróque existíainhibición de lahemaglutinación,cuando¡as cepas perdieron sucapacidad
liemaglutinanteo cuandola hemaglutinaciónfuerte setransfonnóen débil, en presenciade manosa
(M+). fucos:i (F-4¾o galactosa(6+)
182
Total
43
0 (0,0%)
2 (4,7%)
5(11,6%)
19 (44,2)
10 (23,3%)
7(16,3%)
0 (0,0%)
0(0,0%)
CECT
2
0 (0,0%)
0(0,0%)
0 (0,0%)
1 (50,0%)1 (50,0%)
‘0(0,0%)
0(0,0%)
0 (0,0%)
1
0 (0,0%)
0(0,0%)
0 (0,0%)
1(100,0%)
0 (0,0%)
0(0,0%)
D (0,0%)
0 (0,0%)
12
3 (25,0%)
0(0,0%)
3 (25,0%)
4 (33,3%)
0 (0.0%)
2 (16,7%)
0(0,0%)
0(0,0%)

c’to
~oIr
Oc
u.—un
Cl
e-
oo~
o~o
’o
—u
-u
.—u
--
oco
O
e-4
so—
—C
l
enenu.—
<‘4
-‘u’
~u.o
o.
o—‘-u
o
e-o
.u.’—en
Cl
e-
u.—
e-
Un
——
QO
oE-
sososou—oene
n‘-u
Cl
1‘auYua41la~
¡4
E
0.4+m‘4<0.4
u,«0.4+0.4en+0.4+0.4u
,
dICd<fi<.~
t1e‘aUQl
A4?‘a¿U‘0tU¿U‘auU‘3uU:5UE
-
u>,.~
OC
O
oen
S
-~fl
~‘o‘~
u!~‘o
1141bu
¿U41WC‘Eo
80
~•~
~-)
1S3

Wc
nr nr st st nr’ ‘uf st tu mt
no
, o”
na
o,
E-~
U) 00
C.C
..
E—
~—
go,
8Dm
~~
CC
bOs
CM oQ
l
ceIb
~a
,o,
+M
au
‘a o
,a
.•
PS
-(J
AF
JJ
‘Og
OS
9L
A’a
‘6
o,
-Jtu.
>p
~--4
o,¡
41
u—,
ME
teo
’o E,
—te
—t
co
u...’
Ma
o’
tu’>
o’
LA
oo,
E’
u>
(A4
u>
o-J
00
04
.su
bs
mfi
su
E
u>
-~o
’u
>a
:o
u
1:—
1o
8g
+—
~h
m—
m.4
-~tu’
>Z
4”
+4— o,
cea o
’+
LA
O’
u.-’,
U>
~x
4—
—o
’It’>
u>
-J
——
——
jm
O’
~u
u>
LA
—L
Aca
e>—
tu.>
tu.)
—o
’‘
~‘
‘u’>‘-4
0.
o’
<fitu.
>1”
>5
00
S‘~
Ma
ti~
‘o O
’
tu.)
LA
It’>tu.
>a
’(J
i—
40
’u
>—5-’,
‘u-’>u
>0
’O
C‘o
0’
¡ __
18
4

en~
OOoc
u.M~
—-u
sc‘-‘
oc“ti’
un¡
—u.
te-
< tmisc~
tu.—
e-
mo
——
00o14?•<
e’
O.
Cl
Ou’-’
OC
l
qeE-
cm>
cm>1E-
ItLb¡uu<
UuU1¿U8u‘o•1uuCd
4?cm>
dIcli
01ulb.1<41MauQ
l8AeMaUuU1ID
Ma
1Xli¿UID¿U‘oUuu0%M
SaUE-
u1Ecli
.2:a4?WC-Eo
4>‘(5
uA><j00
II;
185

tu
st
tu
—
st
a
a
Sr
a
0
Su
mt
Tab¡a IV.40 - Hidrofobícidad superficial de las cepas de Aeromonas spp.
Origen
Diarreicas Alimentarias CECT Total
N de cepas probadas: 15 81 4 100
Unión a filtros de nitrocelulosa:
F II 50 2 63(63%)Cepas* M 4 31 1 36(36%)
1’ 0 0 1 1(1%)T 15(100%) 81 (100%) 4(100%) 100(100%>
R 78,68 74,35 64.30 74,59Porcentaje O. S. 13,77 14,59 24,47 14,96de unión Max. 94.67 95,67 - 83.59 95,67
Mix>. 51.33 40,28 32,24 32,24
Adsorciónahidrocarburos:
F O 0 0 0(0%)Cepas* M 1 3 0 4(5,19%)
1’ 6 66 1 73(94,80%)T 7(46,67%) 69(85,19%) 1(25%) 77(77%)
Hexa decanoX 15,67 15,87 4,06 15,70
Porcentaje D. S. 14,43 11,45 11,64de adsorción Max. 36.33 56,49 — 56,49Mm. 1,24 1,27 — 1,24
P 1 13 0 14(14%)Cepat M 13 61 4 84(84%)
P 1 1 0 2(2%)T 15 (100%) 81(100%) 4 (100%) 100 (100%)
OctanoX 52,02 60,57 56,70 59,14
Porcentaje O. S. 11,07 11,04 - 8,81 11.30de adsorción Max. 74,57 80,83 66,50 80.83
Mm. 27,96 9,75 45,58 9,75
F 0 17 1 18(18%)Cepas* M 15 64 3 82(82%)
P 0 0 0 0(0%)T 15 (100%) 81(100%) 4(100%) 100(100%)
XilenoX 57,90 63,52 62,73 62,65
Porcentaje O. S’ 5,57 8,55 7,47 8.32de adsorción Max. 65,90 83,95 70.08 83,95
Mm. 50,20 43,07 53,84 43,07
CECÍ - Colección EspañoladeCultivos Tipo
* - E: Cepasfuertemente hidrofébicas:Nl: Medianamentehidrofóhicas;P: Pocohidrofóbicas;T: Totales
Xu 118., Max.y Mm.; - Mediaaritmética.Ucs’iaciónestándar,Valor máximoy Valor mínimo, respectivamente,de lascepastotales 1>
186

64
o
u.—
en
unCl
ounu.-,
Cl
Coe
-e
-U
soQ
lsc
scU
sosO
.~-
u.,rl
ot
u.——
Sr>—
U‘uU‘oc‘.4Ql
‘rl—
—o
c
¿Uji
4?WC‘Eo
E-
u
un‘rlCl
tul
Cl
Cl
unso‘--u
unso
IDt‘E01U¿Ue•1u’oiu:5u;4?e.
u—
1St.
Qll>
~IDu
Ma
Mc—
54
1
80.
4?cm)
U0.
4)
£E-”u,
O‘aCm>‘(5Ou~u
Q
E-
187

Su It. st> nr
Eo,
o
a.
aE
,a
o,‘—
ah
—e,
E,
¡u
.a
a~
¡a
l~E
,u
a—
>P
CM
au
O a,z
‘aQ
lO
’o
<5‘a
al
nO
C“1
4—
~;
E 1“E
o ci,
Ma
~st
uue!~
-Ju
>L
Aso
oO
OC
e’
e’,’e
’
o, r¿U E, MI
-43
u.
u.—
u.—,
gE
lMI
gtu
t—
It’>~
‘>5
oO
.A
-’~
MI
o,!
u’z
ii
~‘a
¡u,
~E”
¡u.—
‘—-<
o,e
,—
O’ •
o,
.4 ~4
>E
,tu
~Q
l¡
aft~
41’>
a’
—e
’—m
ue
’o
,
u>
o. 0’
Ql
22
uE
,
EU
mt
mu
e
188
mt

oU—
‘a
¿U-‘-‘u
butS
UID
~1
ao2:¿U
—8
u
-mm
•<¿U‘oUa‘a¿UU-25
MdAm-b¿U‘o¿U“‘a¿U~8uU
0Cli
oU
CoUQl
ID‘uUml
UQl
IDu
iQl
41‘aiQ
l
41‘au
.—
¡Cd
4?Ma
eno
’en
sc~en
~‘
<‘5C
l
e-
4oC
l
scoc
soct’
soo
Cl
Clu
ji‘4)—
~¿U41
~
U‘1¡111uQl
a41¿Udua41‘aU1.5‘u4?laoMaU:5UE-
189

st
tu
a
a
mt
tu
Su
pu
w
st>
st
Tabla IVm44 - Asociaciónde factores de virulencia: hemaglutinadón y hemolisis
Cepas
Carácter
Actividadhemolítica
No
(Titulo hemolítico)*
Hemaglutinantes
No hemaglutinantes
p
41
55
174,93 (±154,86)
104,60(±164,37)
<0,025
* - Mediaaritmética(±Desviaciónestándar)
Título hemolítico - Inversadeladilución inésaltaenlaquese obseiva100%dehemólisis
TablaIV.45 - Asociación defactoresde virulencia: presencia depu ylIpollsis
* - Mediaaritmética(±Desviaciónestándar)
190
Cepas Actividad lipolítica
Carácter __________________________Q.tg. de B~naftol)*
con pu1 25 15,314 (±12,459)
sin puf 71 11,561(±7,417)
p <0,05

Tabla 1Vm46 - Asociaciónde factores de virulencia: Capa S ehidrofobicidad
64
HidrofobicidadCepas
NOCarácter(% de adsorciónal octano)*
con capaS 57,746(±12,029)
32sin capaS
p
* - Media artimética(±Desviaciónestándar)
62,22(±9,058)
<0,05
Tabla IV~47 - Asociaciónde factoresde virulencia: descarboxiladónde la usinae hidrofobicidad
Hidrofobicidad
NO
(% de unión a FNC)*
Descarboxiladoras 50 71,535(±15,17)
No descarboxiladoras
p
46
FNC - Fitros denitrocelulosa* - Media artlinética(±Desviaciónestándar)
Cepas
Carácter
78,795(±12,81)
<0,001
¡91

st st’ Pr’ stA re tu tu e tu Su e u a
*<tu
u
oO
Z
111
o,’
o¿
U.
aE
,MI
Ma
ci¿U
e,la
~
~‘;
e,E.
9-a,
al
ao
,Y
’a
l~
-o
,’a
~M
IA
¿U.
o8
o‘a
1-~
a‘a
=tJ
u>u>
e
<2~
u’
mt
‘au
.—,
aMI
o*
u8’
0. o,a,
al
e’
ev e, o,
al
Maa
E,
o,a,
e,U
)tb
u.-
’,
AM
ai
(AO
cM
a_
u1
b0
’1’>
e,o,
Eu
’o
’o
o,
U)
O’
e,<tu
u ‘au
ur
•1u,
__
-u
.-2.
192

4>1‘4-‘(5104>
PC£e0O
’4>
i4>
4>
cm>~
-ffi
~A*
—
¿U‘oU¿U¿U-a¿UUUa3‘E1U‘a:5o:541u,
¡405u,=o‘.4Q
lu
—U¿U4?
14)M
aE‘3LbUo’U:5
193

st Pr st Pr •1 “uf tu e tu tu mu tu u st- u-
z -~<tuo
.tE
”e>
u’
o E,
~r~
mO
-~
‘ag
~O
’MI
Mu
—O
a.~
a,-
’‘d
~O
’u
>—
~tu
.)
<tu-c
—~
ca
.
a,cm
Asctu
.m>
evQ
le
’e’
E.
—o
,o,
Ot
U—
.<tu
4—4
Ql~
uu
>’—
—e
vt
‘—u
1~
~~
—4
e,
.0u
e <tuu
—‘
Ou
>LA
~jn
ae-
‘oo
’— ¿U
s.a,
Au
>4
3.a
lD
.‘o
a’
o.
1;e
’e
5’5
O.
—OC
5’<tu
—a
tc05
9O
Cis
)¿
‘-u
u.*
24.
a~
‘m ti
Aa
lt1
•0—
-‘u
]¿
Uft
‘atI
]¿U
—
4E, a
¿U-
o,-
¡a
le
’ev .4
.
Au>
—0
0,
¡0
4”4
¿U
~%4
bb~
¡LA
LAu
>
II,‘.
Ma
al
¡ev
o,
rlA
al
9¿U
tiE.
LA—
E, E”
o,
194

Oa*
u,=O4>PCa‘O
aO
S~
O4>
u~
VO
4>-~
PCQ
fiPCO
.2PC
1‘e
Ot
tb
~£
O~
Ou
u‘u
0¿
tCb
~8
OC
en
<N‘e
—o
-.O
~0
U¡
-u
-’o
.¡
sAZ
y~
1‘1
~1)
ua,
‘OPC
~PC
uu
u.—e
~
uz
tu.
e.
‘o—
—5
CN
e.
u<
ne
n
~S
n
4>—
.2PC
OC
Nt
10
%fl*”4
‘~O
—~
Ci
EP
C‘o
itr—
tU
4<
t-
si
fitu
.a
u‘~
—o
’-
u!
-~tn
t~-—
‘~‘e
‘~~
J14•
r—0
~j
~•
a~-‘-.
§.—
-u
a,~
‘rrnS
PC~
OP
C’~
~‘—
‘y
iu
>u
-—
PC.2
OC
fl~
—‘o
—..~
te4
a,“
O4
>’O
‘~
~4<
‘-uo
~~
4>
0tu.
fi
¡~
gE
u•~
.,‘.,,&
—5
o.
cw
~PC
~‘
r—y,
OC
tP
C~
cfly
ju~
~~
y—
~m
du
PC
PC
Sa
‘~
‘Or~
u‘u
‘mc
uP
C
-!‘~
~¡
-e
u,O
r’uO
N¡
o
.28
—~1
—-o
u,4>4>
u,4
>4
>0
u..0
ou
<~
‘~Q
l~
t-~
4>utu
CuS
LC
OO
~O
’O~
”o
c~
t‘~
—~
Oti
OԦ
u4>
2et
~t
U0
CC
oC
U-
-J
195

r.
nr-
st’
‘st
a
re
mt
mt
tu
u
st>
e
pu
pu
Peso Molecular (kDa)
8-30
• A bydrophila 2 (CECT)• A sobria 11 (diarreica)
• A hydrophila27 (alinientariM
~ 4áY~igabUa55 (alimentaria)
E] A. hydropbila 75 (alimentaria)
E khy~lr~phila96(alimentaria)
-u
Peso Molecular(kDa)
Figura IV. 1 - Aclividau hemolítica detectadaen las fr iccionesdel sobrenadante.obtenidaspor diafútración,deseiscepasde Acromúnasspp.móvilestt—heínolisiso betaulisis incompletay ~1—lsernolisiso hemolisis completo
Título humo1 hico - loverso cte la dilución más altaen la que se observa 100%cíe bemol si
196
-<8 30-50 > 5010000
8400 -
6800 -
5200 -3600 -
2000 -
oLb-~1“4‘oE4?
o
400 -350
u
aa
a
300
250
200
150
100
50
o
/
/
> 50-cg 8-30 30-50

A. sobria 13 (diarreIca) A- sobria 14 (dIarreica>600—
400
200—o
o
cZ~~u u
10 20tiempo (h.)
11
9
7
5
30
A. hjdrqphila 75 (alimentaria)11
400
200
o0 10 20
tiempo (hm)
o
1 4>1
u E4>ae
— ou
o4>
91 1
U a7
yu
5 1~‘mli
600
400
200
o
11
<auuu
-7o
5
0 10 20 30tiempo (h-)
A.hydrophila 24 (alimentaria)11
600
400
200
o0 10 20
tiempo (h.)
91u
7
y5
A. sobria 34 (alimentaria)
600-
400-
200 -o u u
0 10 20tiempo (h.)
11
9
7
5
o
u1
Eu e>~ a
O‘31~
30
A. hydrophila 94 (alImentarla)11
600
400
200
o0 10 20
tiempo (h.)
91u
7o
5
A. sobria 92 (alimentaria)
600
400
200
o0 10 20
tiempo (h.)
o4>
§E4>a
eu
11
9
7
5
30
A- hydr9pkhila 96 (alimentaria)11
600
9400
7200
-5O,
o 10tiempo
20(h.)
1u
y
1u
o
30
Figura IV.2 Evolución de la producciónde hemolisinascon respectoa la población
bacterianade ochocepasdeAeronwnasspp.móviles. (a) Títulohemolítico.(e) Log ufc/mITítulo hemolítico - Inversa de la diluci’5n más alta enJaquese observa 100%dc heinolisis
e4>
oEtIaeu
o4>
ou41a
eulid
30 30
eu
oE41a
ou
E-
o4>
oE4>
o
u
E-
30
107

‘uf>
st
st
‘st
nr
st
sts’
st
st
st
re
nr.
re
st
12
10
8
6
4
2
o
• A. ~ydrqphda2 (CECT)
• ks~kna11 (diarreica)
• A- hydrophila27 (alimentaria)
O A. hydrophila55 (abinentaria)o Ahvdropbila75 (alimentaria)
E A. hydrophila96 (alimentaria)
> 50
1Pesomolecular (kDa)
Figura IV.3 - Actividad proteolíticadetectadaen las fracciones delsobrenadante.obtenidaspor diafiltración, deseis cepasde Aeromonasspp.móvilesUnidad proteolitica - Actividad enzirnática que produce un incrementoen laabsorbanciaa 440orn de 0,1. porml de sobrenadante
ULb
‘3IDobual
IDMaU
Ma-E 1
< 8 8-30 30-50
198

A. sobria 34 (alimentaria) A. sobria 13 (diarreica)
12
8
4
o0 10 20
tiempo (h.)
A. bydrophi¡a 94 (alimentaria)11
12
8
4
o0 10 20
tiempo (b.)
u,115>
9’ .¡2au uo’
e -— u,
‘u5 u
12 —8—
4-
o u • u
0 10 20
It
-9 iu
.7
y—5
30tiempo (h.)
u,u5>
viti —— §u 1-a.oca— u,
PC5 u
A. hydrophlla 96 (alimentaria)—11
9’
u~00
o
5
12
8
4
o
tiempo (h.)
A. sobria 11 (diarreica)ti
ti
1~
9! j-
t- au 1-7 oc a.
o— u‘u
5 ‘30 10 20 30
tiempo (h.)
A. bydrophila 24 (alimentaria)ti
12
9
87
4
5
10 20tiempo (h.)
30o
iuoco
u,U4>
oU>aa-a.
u,PCu
A. hydrophiia 6 (diarreica)
12
8
4
o0 ¡0 20
tiempo (h-)
A. hydropbila 9 (diarreica)
12 -
8~
4-
o0
u u
10 20tiempo (h.)
11
9,
u
o
5
—11
-9!
u-7 oco
-5
30
Figura IV.4 - Evolución de la producción deproteasasconrespectoa lapoblaciónbacterianadeocho cepasde Acrojaunasspp.móviles. (o) Unidadesproteolíticas.(e) Log ufc/mlUnidad proteolítica- Actividad enzimática que produceun incremento en la absorbanciaa 440 nni de 0,1 pormi de sobreTudante
u,0u
u. —
oU>
e1”a.
u,PCu
30
u,u<.1
aU>
au-a.
u,PC‘3
30 0 10 20 30
u,uu
§4>§a-a.
viPCu
12
8
4
o
u,04>
§U>
-4-§u-a.
u‘uu
30
19<)

mt
O:
O:
pu
mt
st
nr
9,
O
O
‘It
mt
• A hydrophila 2 (CECT)
• A. sobria It (diarreica)• A. hydropbila 27 (alimentaria)
El A. hydrophila 55 (alimentaria)E] A. hydrophita75 (alimentaria)
[II A. hydrophila 96 (alimentaria>
an-fl
<8
~rI-jfl
8-30
-a.-’
30-50 > 50
Peso molecular (kDa)
Figura IV.5 - Actividad lipolltica (expresada como~agde 13-naftolliberados)detectadaen lasfraccionesdel sobrenadante,obtenidaspor diafiltración,dc seiscepasdeAeronzonasspp.móviles
200
80
01o
MaUbu4?
Mc
4-•4~U¿U
41
MatÉ
6V
46
20-
o

A. sobria 34 <alimentaria) A. sobria 14 (diarreica)
-r r~~r0 10 20
tiempo (hj
11
9
7
5~1
aeno
ae
4>PCena
30
A. hydrophiia 75 (alimentaria)11
0 10 20tiempo (h)
9!
aeno
5
o<4~
ae4>
PC
en
A. hydrophila 9 (diarreica)
u u10 20tiempo (h)
‘4
30
A. hydrophila 5 (diarreica)
u ‘ u
0 10 20tiempo (b.)
— 11
—5
— oE ~
‘-‘u O4> aE-
een 4>e
ena
—11o
-9 E ‘(5~~1
4>u-
-7 4>enoen
—5
17 —13 —
9-
5-
1
í’t
9
7
5a u
0 10 20
tiempo (h.)30
A. hydrophila 24 (alimentaria)11
17
13 9
9 7
55
1’0 10 20 30
tiempo (h-)A- hydrophila 96 (alimentaria)
1117
13
9
5
10 10 20
tiempo (h.)
4>
a7 eno
5
A. hydrophila 94 (alimentaria)11
17
13
9
5
130 0 10 20
tiempo (It)
4>
a7 eno
5
Figura IV.6 - Evoluciónde la producción de lipasas con respectoa lapoblación bacterianadeochocepasdeAeromonasspp.móviles. (o) ~tgde 13-naftolliberados.(e) Log ufc/ml
ou-Oae4>
PCena
17 —
¡3 —
9-
5-t
1
‘7
13
o‘4-
‘(5ae4>
PCena
9
5
—‘u<a4-
eno
i—‘u4>
a
eno
130
oa-
ae
4>PCena
17 -
139-j
5-.
1o
17 —
13
o4-ae4>PCena
30
9—
5-
130
201

r.
mt
e
mt
mt
a
mt
e
a
e
e
mt
a
e
pu
a
pu
FiguraIV.7 - Efecto citotóxico de distintas diluciones de los sobrenadantes deAeromonasspp.sobre lamonocapadc célulasVero. (a y b): Diferentesgradosde citotoxicidad,en función de la dilución empleada.(e): Monocapano lesionada(control>
202
(a) (b)
(e>

(a)
(b)
Figura IV.8 - Efecto citotóxico de los sobrenadantesde Aeromonasspp. sobrecélulasde mieloma. (a): Célulaslesionadas.(10: Células nolesionadas (control)
203

pu
Su
st
pu
mt
u
nr
pu
pu
pu
e
a
mt
‘mu
Peso Molecular (kDa.)
-<8 8-30 30-50
• Ahydsuphda2 (CECT)
• A sobria II (dianeica)
• Ahydrophila2 (alimentaria)
~ A hydroph41x 55 (alimentaria)
E] A. hydrophila75 (alimentaria)
E A-_hydrophila96 (alimentaria)
> 50
-c 8 8-30 30-50 > 50
Peso molecular (kDa.)
Figura IV.9 - Actividad citotéxicasobrecélulasVerodetectadaen las fraccionesdel sobrenadante,obtenidaspor diafiltración,deseiscepasdeAeromonasspp’ móvilesTítulo citotóxico - Inversade la dilución más alta en la quese observa50% de muerte celular
204
4000
2800
e’
Oa-4)
u,
‘4)Lb
4)bu
Ou,
OLb
‘OO
e-)O
1600 -
400
300
200
100
o

(a)
(b)
Figura IV.1O - MicroscopíaelectrónicadeAerontonas70, provistadecapaS(a) y de Aeronionasó4desprovistade capaS (b)
205

nr
st
nr
st
mt
a
e
mu
mt
a
pu
mt
a
pu
U
e
(4
(a)
.4:
(b)
Figura IV. 11 - Microscopiaelectn5nicadeAeromonas32 con numerosospilí cortosy rígidos(a) ydeA. hydrophita25 conpili largosy flexibles (b)
206
*
‘~W ~.4, 1,“-4
‘4>
• “ti
--e
A
E

y. CONCLUSIONES

0’~
4’
‘Su
Su
st:
pu
9,
tu.
‘u.
9,
WC

CONCLUSIONES
1. - El medio selectivomásadecuadopara elaislamientodeAeromonasspp.móviles, a
partir de alimentos y aguas,es el medio de Ryan. La proporción de ampicilina y los
componentespresentesen el mediode Ryan permitenel crecimientode A. sobria CECT
837, a pesarde serla cepacon mayorsensibilidadfrentea esteantibiótico.
2. - Aeromonasspp.móviles estánampliamentedistribuidasen alimentos españoles(carnes
de avesy mamíferos, pescadosy mariscos,lechecruda,etc.).Su incidenciaes muy elevada
en aguas no potables de ríosy arroyos.
3. - AunqueAeromonasspp. móvilesson, en general,psicrótrofas, las cepas de origen
alimentariocrecenmejor a temperaturasde refrigeraciónque las diarreicas.Tantounas
como otras sonincapacesdeproliferara40C, si el pH esinferiora 6. El nitrito sádicoy el
cloruro sódico afectan, notablemente,al crecimientode estosmicroorganismosque,
además,son sensiblesal cloro.
4. - El mayor númerode cepashemolíticassedetectacuandola incubación se realiza a
280C,aunque la producción máxima dehemolisinassepuedeconseguirtantoatemperaturas
inferiorescomosuperiores.Los eritrocitosde conejoson más sensiblesa lashemolisinasy
hemaglutininasdeAeromonasspp.quelos de cordero.
Sm - La producciónde hemolisinas,proteasasy lipasasse detectaal final de la fase
logarítmicade crecimientoy principio de la estacionariadeAeromonasspp..
6. - Las actividades hemolítica,lipolítica, proteolítica y citotóxica se asocian,
principalmente,con los componentesdel sobrenadantede PM superiora 50 kDa y, en
ningúncaso,hanpermitido establecerdiferencias significativasentrecepasalimentariasy
diarreicas.
7. - Las célulasde mieloma son mássusceptiblesa las citotoxinas producidaspor
Aeromonosspp. quelas células Vero.
8. - Sehan detectado cepashemaglutinantes,autoaglutinantes,no aglutinantesen presencia
de acriflavina, hidrofóbicas,con capaS y con pUf, tantoenAeromonasspp. de origen
alimentario como diarreico,aunquela hidrofohicidadessignificativamentesuperioren las
2(X)

nr,
0
WC
mu
pu
e
0
mu
pu
pu
e
CONCLUSIONES
cepasde alimentos.
9. - Se hanseleccionadotrespruebas(VogesProskauerpositiva, no fermentaciónde laarabinosay no aglutinaciónen presenciadeacriflavina) que, consideradasconjuntamente,
permitensuasociacióncon latotalidadde los factoresde virulenciaestudiados. De ellas,la
reacción de Voges Proskauerpuedeconstituir una pruebasencillay de utilidad para
seleccionarcepaspotencialmentepat6genas,ya queserelaciona,significativamente,con la
hemolisis,proteolisis,citotoxicidad,hemaglutinación,autoaglutinacióny no aglutinaciónen
presenciadeacriflavina.
10. - La ampliadistribucióndeAeromonasspp.en alimentosy en los mediosacuáticosy
terrestres, el escaso número de brotesdeclaradosen los paísesdesarrollados,así como las
dificultadesparaconfirmarestaetiologíaen los mismos, nos inclinan apensarqueeste
grupo de microorganismosdebeserconsiderado,al menos, comopatógenopotencial para
el hombre.
210

VI. BIBLIOGRAFIA

nr
WC
u
a
u,
“4
mu
mu
e
pu
u
a

BIBUOCRAFIA
ABBOYI7. Si.; CLIFUNO, W.K.W.; KROSKE-BYSTROM, 5.; MALEKZADEH, T. Y JANDA, MJ.
(1992).Identification of Aeromonasstrainsto the genospecieslevel in thecinical
laboratory.J. Clin. Microbiol. 30. 1262-1266.ABBO’117. Si.; SERVE,H. Y JANDA, J.M. (1994).Caseof Aeromonasveronii (DNA group
10) bacteremia.1 Clin. Microbiol. 32, 3091-3093.
ABEYTA, C.JR.; KAYSNER, C.A.; WEKELL, M.M.; SULLIVAN, Ji. Y STELMA. G.N. (1986).
Recovery of Aeromonas hydrophilafrom oysers implicated in a outbreakoffoodborneillness.J Foad. Protect. 49, 643-646.
ABEYTA. C.JR. Y WEKELL, M.M. (1988). Potencial sourcesof Aeromonashydrophila.J.
FoodSafety.9, 1-22.
ABRUTYN, E. (1988).Hospital-associatedinfectionfrom leeches.Ann. Intern. Med.109,356-358.
AGBONLAI-IOR, DE., SHONEKAN. R.A.O.; KAZAK, W.H. Y COXER.A.O. (1982).Aeromonas
food poisoningin Nigeria: a casereport.CentAif. J. Med.28, 36-38.
AGOER, W.A. (1986). Diarrheaassociatedwith Aeromonashydrophila. Pediatr.Infect. Dis.
Suppl.5, S106-S108.
MiCER. W.A.; MCCORMICK, J.D. Y GURWITH, M.J. (1985).Clinical and microbiological
featuresof Aeromonas hydrophilaassociateddisease.J. Clin. Microbiol. 21,
909-913.
ALLEN, D.A.; AUSTIN, B. Y COLWELL, tR. (1983).Aeromonasmedia, a newspecies
isolatedfrom river watcr. fiat. 1 Syst. Bacteriol. 33,599-604.
ALTORFER, R.; ALTWEGG, M. Y ZOLLINGER, J. (1985). ítemandA. von Graevenitzgrowth
of Aeromonasspp. on cefsulodin-irgasan-novobiocinagarselectivefor Yersinia
enterocolitica.1 Clin. Microbiol. 22, 478-480.ALTWEGG. M. (1985).Aeromonascaviae:an entericparliogen?.Infection.13, 228-230.
ALTWEGC, M. Y GEISS,MI. (1989).Aeromonasas ahumanpathogen.CRC Cnt. Rey.
Microbiol. 16, 253-286.
ALTWEGO, M.; STEIGERWALT. A.G.; AiLTWEGG-BISSIG. R; LIJTHY-HO1TENSTEIN, 1 Y
BRENNER,Di. (1990).Biochemicalidentificationof Aeromonasgenospeciesiselated
from humans.¿ Clin. Microbio!. 27, 258-264.
ALTWEGG, M; MARTINE1TI LUCI-IINI, G.; LUTHY-HOYrENSTEIN, J. Y ROHRBACH, M.
(1991).Aeromonas-associatedgastroenteritis¿terconsumptionof contaminated
shrimp.Lur. J. Clin. Microbiol. Infect. Dis. 10, 44-45.
213

nr
nr
“4
“4
“4
mt
mu
e
mu
mu
“4
“4
mu
“4
nr;
BIBLIOGRAFL4
ALTWEGG. M. Y KERSTERS, K. (1994). CelluIar fatty acid composition as a
chemotaxonomicmarkerfor thedifferentiationof phenospeciesand hybridization
groups¡n the genusAeromonas.mr. J. Sysr.Bacceriol. 44, 651-658.ANGUITA, J.; RODRíGUEZAPARICIO, LB. Y NAVARRO, 0. (1993). Purif’¡cation genecloning
aminoacid sequenceanalysis, and expressionof an extracelularUpasefrom an
Aeromonashydrophilahumanisolate.Appl. Environ.Microbiol. 59, 2411-2417.AOKI, 17 Y HOLLAND, 81. (1985). The outer membraneproteifis of the fish pathogens
Aeromonashydrophila,Aeromonas salmonicidaandEdwardsiella tarda. FEMS
Microbiol. Len.27, 299-305.
ARDUINO, Mi.; HICKMAN-BRENNER, F?W. Y FARMER III 13. (1988).Phcnotypicanalysisof
132 Aeromonasstrainsrepresenting12 DNA hybridizationgroups.Ahstr.P-3. 2nd
mL WorkshopAeromonasPlesiomonas.
ASAO, T.; KINASKITZ, Y.; KOZAKI. S.; VEMURA, T. Y SACAGUCHI, 0. (1984). Purification
and somepropertiesof Aeromonashydrophila haemolysin. Infect. !mmun.46,
122-127.
ASAO, T.; KOZAKI, 5.; KATO, K.; KINOSHITA, Y.; OTSU, K.; VEMURA, T. Y SAKAGUCHI, 0.(1986).Purificationandcharacterizationofan Aeromonashydrophilahemolysin.¿
Clin. Microbio). 24, 228-232.
ASH. C.; MARTINEZ-MURCIA, A.J. Y COLLINS, M.D. (1993).Identification of Aeromonas
schubertii andAeromonasjandaeiby usinga polymerase chainreaction-probetest.
FEMSMicrobioLLea. 108, 151-155.ASHDOWN.L.R. Y KOEHLER, J.M. (1993).ThespectrumofAeromnonas-associateddiarrhea
in tropicalQueensland.Australia.SoutheastAsian¿ Trop. Mel Pubhc. Healrh.24,
347-353ATKINSON, H.M. Y TRUST, Ti. (1980).Hemagglutination properties and adherenceability
of Aeromonashydrophila.Infect Immun.27, 938-946.
ATKINSON, H.M.; ADAMS, O.; SAVVAS, R.S. Y TRUST, TI. (1987). Aeromonasadhesinantigens.Experienria.43, 372-374.
AUSTíN, B. Y AUSTíN, DA. (1985). Bacterial patihogensof fish. ¿ Appl. Bacreriot 58,
483-506.AUSTIN, 8. Y AUSTIN, DA. (1987).Bacterialfish pathogens:diseasein farmedand wild
fish. Bilis HorwoodLtd. ChichesterWestSussexEngland.
AYTAC, SA. Y OZBAS, Z.Y. 0994). Survey of 0w growth and survival of Yersinia
enterocoiltico andAcmmonoshydrophila in yogurt. Miichwissenscbaft.49. 322-325.
214

BIBLIOGRAFf4
BALOQA, S.B.; KROVACEK,K.; ERIKSSON,1.; LINNE, T. Y MANSSON, 1. (1995). Detectionofaerolysingenein Aeronionasstrains isolatedfrom drinking water, tish andfoodsbythc polymerasechain reaction.Comp.lmmunol.Microbio!. lnfect. DÍSu 18. 17-26.
BARER, M.R.; MILLERSI-{IP, SE. Y TABAQCHALI, S. (1986). Relationshipof toxinproductionte speciesin the genusAeromonas.1 Mal. Microbiol. 22. 303-309.
BARGHOUTHI, 5.; YOUNG, R.; OLSON, M.O.J.;ARCENEAUX, J.E.L,; CLEM, L.W. Y BYERS,
BR. (1989). Amonabactin,anovel tryptophan-or phenylalanine-containingphenotate
siderophorein Aeromonashydrophila.¿ Bacteriol. 171, 1811-1816.
BARTKOVA, O. Y CIZNIER, 1. (1994). Adherencepattern of non-piliatedAeromonas
hydrophilastrainsto tissuecultures.Microbios.77, 47-55.
BEACHEY, E.H.; SIMPSON, W.A.Y OFEK, 1. (1980). Interactionof surfacepolymersof
Streptococcuspyogeneswith animal cells. InMicrobial adbesionfo surfaces.eds.
Berkeley, R.C.W.; Lynch, J.M.; Melling, J. Rutter, P.R. y Vincent, B. PP.
389-405.London: Societyof Chemical Industry.BEEBE, ¿EL. (1986).Bloed-borne bacteria asreflectors and indicators of cancer.La!’. Man.
May, 45-50
BELLANO, R.J. Y TRUST, 1.1. (1985). Synthesis,expon, and assemblyof Aeromonas
salmonicida A-layer analyzedby transposonmutagenesis.¿ Bacteriol. 163,
877-881.
BELLAND, LI. Y TRUST, T.J. (1989).Aeromonas salmonicidaplasniids:plasmid-directed
synthesisof proteins ni vitroandin Escherichiacolí minicells. J. Gen. Microbiol.
135, 513-524.
BERNAOOZZI, M.; BIANUCCI, F.; SARRE,E. Y SACCHETrI, R. (1994). Assessmentof some
selectivemediafor therecoveryofAeronwnas hydrophllasurface waters.Zernralbk
Hyg. Umweltmed.195, 121-134.
BERNHEIMER, A.W. Y AVIGAD, L.S. (1974).Partialcharacterizationof aerolysin,a lyticexotoxinfrom Aeromonashydrophlla.Infecr. Immun.9, 1016-1021.
BERRANO, ME.; BRACKEYr, tE. Y BEUCHAT, L.R. (1989). Growthof Aeromonas
hydrophilaen freshvegetablesstored undera controlledatmosphere.Appl. Environ.
Microbiol. 55, 2167-2171.BEUCHAT. La. (1991).BehaviourofAeroinonasspeciesat refrigerationtemperatures.¡nl.
1 FoodMicrobio!. 13, 217-224.BEVERIDGE, 1.1 (1978).The responseof cdl walls of Bacillus subtiliste metalsami te
electron-microscopicstrains.Can. 1 Microbiol. 24, 89-104.
215

nr
“4.
“4
mu
mu
mu
fr,
“4
“uf
mu
mu
BII3L106RAFÍA
BEVERJOGE,Ti. (1979). Surfacearraysen the wall of Sporosarcinaureae.¿ Racteriol.
139. 1039-1048.
BEVERIOGE, TJ. Y MURRAY, RiLE. (1975). Surfacearraysen the cdl walI el’ Spirilummetamorphum.¿ Bacteria!. 124, 1529-1544.
BHAKDI, 5. Y TRANIJM-JENSEN,1 (1988). Damageto ccli membranesby pore-formingbacterialcytolisins. Prog.Allergy. 40, 1-43.
B[AMON, ElY ¡[AZEN, T.C. (1983).Survival anddistributionofAeromonashydrophik¿in
near-shorecoastalwatersof PuertoRico receivingrum distillery effluent.WaterRes.
17, 319-326.BICKEL, K.D.; LINEAWEAVER, W.C.; FOLLANSBEE, 5.; FEIBEL,R.; JACKSON, R.; BUNCKE,
H.J. (1994). Intestinalflora of the medicinal leechHirudinaria manillensis.1.
Reconstr. Microsurg.10, 83-85.BJORN, M.J.; SOKOL, P.A. Y IGLEWSKI, B.H. (1979). Influence of Iran on yields of’
extracellularproductsin Pseudomonasaeruginosacultures.¿ Bacteriol. 138.193-200.
BLACK, R.E. (1986).Pathogens thatcause¡ravelersdiarrheain Latin AmericaandMrica.
Rey.mf Dis. 8 (Supplj, 13 1-135.BLANCHARD, D.C. Y SYZDEK, L.D. (1978). Sevenproblemsin bubble andjet drop
researches.Limito!. Oceangr. 23, 389-400.BLAUSTEIN, A.R. Y WAKE, 0.0. (1995). Declive de las poblacionesde anfibios.
Investigacióny Ciencia.Junio, 8-13.
BLICKSTAND, E. YMOLIN, G. (1983).Carbondioxide as acontrollerof tic spoilage floraof
pork,with specialirferenceto texnperatureandsodiumchIoride.¿ FoadPro¡ect. 46,
756-763.BLOCH, S. Y MONTEIL, 11. (1989). Purification and characterizationof Aeromonas
hydrophilabeta-hemolisyn.Taxicon.27, 1279-1287.
BOULANGER, Y.; LAELIER, R. Y COUSINEAU. 6. (1977). Isolation of enterotoxigenic
Aeromonasfrom fish. Fish Can. J. Microbio!. 23, 1161-1164.BRENDEN, R.A. Y JM4DA, J.M. (1986).The interactionof complementcomponentswith
Aeromonasspecies.Can. ¿ Microbio!. 32, 1-3.
BUCHANAN, RL. Y PALUMBO, SA. (1985).AeromonoshydrophilaandAeromonas sobria
aspotentialfoad poisoningspecies:areview.¿ FoadSafety.7, 15-29.
BUNCIC, 5. Y AVERY, SM. (1995). Effcct of pre-incubation pH en the growth
characteristiesof Aeromonashydrophila at 50C. asassessedby two rnethods.Lea.
AppI. Microbio!. 20, 7-10.
216

BIBUOGRAFIA
BUNNING, V.K.; CRAWFORD, R.G.; STELMA, G.NJR.; KAYLOR, L.O. Y JOHNSON, C.H.
(1986).Melanogenesisin murineB16 celis exposed toAeromonas hydrophila
citotoxic enterotoxin.Can. ¿ Microb. 32. 814-819.
BURKE. V.; ROBINSON, 1.; ATRINSON, HM.; DIBLEY, M.; BERRY, R.J. Y GRACEY, M.
(198la). Exotoxins of Aeromonashydrophila.Aun. ¿ Exp. Biol. Mecí. Sci. 59,
753-761BURKE, y.; ROBINSON,J.; BERRY,RJ. Y GRACEY, M. (198lb). Detectionof enterotoxinsof
Aeromonashydrophila by suckling mousetcst.¿ MedMicrobio!. 14,401-408.BURKE, y.; GRACEY. M.; ROBINSON,3.; PECK,D.; BEAMAN, J. Y BUNDELL, C. (1983a).The
microbiology of childhoodgastroenteritis:Aeromonasspeciesand otherinfective
agents.1 Infect. Dis. 148, 68-74.
BURKE, y.; ROBINSON, J.; BEAMAN. 1; GRACEY, M.; LESMANA, M.; ROCKHILL, R.;
ECHEVERRIA, P. Y JANDA, J.M. (1983b). Correlation of enterotoxigenicity with
biotype in Aeromonasspp.1 Clin. Microbiol. 18, 1196-1200.BURKE, V.; COOPER,M.; ROBINSON, J.; GRACEY, M.; LESMANA, M.; ECHEVERRIA, P. Y
JANDA, J.M. (1984a).HemagglutinationpatternsofAeromonasspp. in relation to
biotype andsource.¿ Clin. Microbiol. 19, 3943.BURKE, y.; ROBINSON, 3.; COOPER. M.; BEAMAN, J.; PARTRJDGE, K.; PETERSON,D. Y
GRACEY, M. (1984b).Biotyping and virulence factorslii clinical andenvironmental
isolatesof Aeromonasspecies.Appl. Environ. Microbiol.47, 1146-1149.BUREE,V.; ROBINSON,1.; GRACEY. M.; PETERSON,D. Y PARTRIDGE,K. (1984c).Isolation
ofAeromonashydrophilafrom metropolitan watersupply: seasonalcorrelationwith
clinical isolates.Appl. Environ. Microbiol. 48, 361-366.
BURKE, y.; ROBINSON, 3. Y GRACEY, M. (1987). Enteroíoxinsof Aeromonasspecies.
Experientia.43, 368-369.
CAHILL, MM. (1990).Virulence factorsin motileAeromonasspecies.1 Appl. Bacteriol.
69, 1-16.
CALLISTER, SM. Y AGOER,W.A. (1987). Enumerationand characíerizationof Aeromonas
hydrophilaandcaviaeisolated fromgrocerystoreproduce.AppL Environ. Microbiol.
53, 249-253.
CAMPBELL, JO. Y HOUSTON,C.W. (1985). Effectofculturalconditionson the presenceof acholera-toxincross-reactivefactor in cultureflitratesof Aeromonashydrophilo. Cita.
Microbio!. 12, 10 [-¡06.
2l7

nr
nr
mu
“4
mu
e
“4
e
ArRUOC,RAFIA
CAREY, PE.; ELIIY, A. Y WWCOX, MU. (1994). Assessmentof a chemilurninnescent
umversalprebefor taxonomicalandepidemiologicalinvestigationsof Aeromonassp.
isolates.1 Clin. Pathol. 47, 642-646.
CARNAHAN, AM.; JOSEPH, 5W. Y JAlADA, J.M. (1989a). Species identification ofAeromonasstrainsbasedon carbonsubstrateoxidationprofiles. ¿ Clin. Microbiol.
27, 2128-2129.
CARNAl-LAN, AM.; MARII, MA.; FANNING, GR.; PASS, M.A. Y JOSEPH, 5W. (1989b).
Characterizationof Aeromonasschubertiistrains recently isolatedfrom traumatic
wourxd infections.J. Clin. Microbiol. 27. 1826-1830.
CARNAHAN, A.M.; HAMMONTREE, L.; BOURGEOIS, L. Y JOSEPH, S.W. (1990)
Pyrazinamidaseactivity as a phenotypicmarker for severalAeromonasspecies
isolatedfrom clinical specimens.¿ Clin. Microbio). 28, 391-394.
CARNAHAN, A.M.; BEHRAM. 5. Y JOSEPH,S.W. (1991a). Aerokey II: aflexible key for
identifying clinical Aeromonasspecies.¿ Clin. Microbio). 29, 2843-2849.
CARNAHAN, A.M.; CHAKRABORTY, T.; FANNING, G.R.; VERMA, O.; ALI, A.; JANDA, M. Y
JOSEPH,S.W. (1991b).Aeromonastrota sp. nov., an arnpicillin susceptible species
isolatedfrom clinical specimens.¿Clin. Microbio). 29, 1206-1210.
CARNAHAN, KM.; FANNING, G.R. Y JOSEPH,SW. (1991c).Aeromonasjando.ei(formerly
genospeciesDNA group9 Aeromonasobria)a newsucrose-negativespecies isolated
from clinical specimens.¿ Clin. Microbio). 29, 506-564.
CARNAHAN, A.M.; WATSKY, D. Y PEELER, R. (1992). A threeyearretrospectivestudy of
clinical AeromonasisolatedusingAerokeyII. Abstr.C-295.92ndGeneralMeetingof the American Societyfor Microbiology.
CARRELLO, A.; SILEURN, KA.; BUODEN, J.R. Y C}-IANG, B.J. (1988).Adhesienof clinical
and environmentalAeromonasisolated te HEp-2celís.1 Med. Microbiol. 26,
19-27.
CATrABIANI, E. (1986).Sensibilitadi disinfettaridi Aeromonashydrophilae di Vibri
fluvia)is.Arch. Vet. Ital. 37, 65-72.
CHAKRABORTY, T.; MONTENEGRO,MA.; SANYAL. St.; HELMUTH, R.; BULLING, E. Y
TIMMIS, K.N. (1984).Cloning of enterotoxingene fromAeromonashydrophila
providesof a cytotonicenterotoxin.JnfectInmun.46,435-441.
CHAKRABORTY, T.; HUHLE. B.; BERGBAUER, U. Y (jOEBEL, W. (1986).Cloningexpression
¿md mappingof theAcromonas/-zydrophila aerolysin genedeterminantin Escherichia
ccli KV. 1 Bacreriol. 167, 368-374.
218

BIBUOGRAFIA
CIIAKRABORTY, T.; HUHLE, B.; HOF, H.; BERGBAUER, H. Y GOEBEL, w. (1987). Marker
exchangemutagenesisof the aerolysindeterminantin Aeromonashydrophila
demonstratesthe roleof aerolysinin A. hydrophilaassociated systemicinfections.
Infec. ¡minien. 55. 2274-2280.
CHAMPSAUR, H.; ANDREMONT, A.; MATHIEU, O.; ROTTMAN, E. Y AUZEPY, P. (1982).
Cholera-lilceillness dueto Aeromonassobria.£ lnfect. Dis.145,248-254.
CHANG, B.J. Y BOLTON. SM. (1987).Plasmidsami resistanceto antimicrobial agentsin
Aeromonassobria andAeromonas hydrophilaclinical isolates.Anrimicrob.Agents
Chemoter.31, 1281-1282.
CHART, Mu Y TRUST, Ti. (1983). Acquisitionof iron by Aeromonassalmonicida.J.
Bacteriol. 156, 758-764.
CHOPRA, A.K.; HOUSTON, C.W.; GENAUX. C.T.; DIXON. J.D. Y KUROSKY. A. (1986).
Evidencefor productionof an enterotoxinami choleratoxincross-reactivefactorby
Aeromonashydroph¡la. ¿ Clin. Microbio). 24, 661-664.
CHOPRA, A.K.; HOUSTON, C.W.; PETERSON,J.W. Y JIN, G.F. (1993). Cloning, expression
andsequenceanalysisof a cytolyúc enterotoxingenefrom Aeromonashydrophila.
Can. ¿ Microbio). 39, 513-523.
CHOPRA, A.K.; PHAM, R. Y HOUSTON, C.w. (1994). Cloning andexpressionof putativecitotonic enterotoxin-encodinggenes fromAeromonashydrophila.Gene.139,
87-91-CIPRIANO, R.C; GRIFFIN. B.R. Y LIDGERDING, B.C. (1981). Aeromonas.valnwnicida:
relationshipbetweenextracellulargrowth productsandisolatevirulence. Can. 1
Fish. Aquat. Sci.38, 1322-1326.
CLARK, J.A.; BURGER, G.A. Y SABATINOS. LE. (1982). Characterizationof indicator
bacteriain municipal raw water,drinking waterand newmain water samples.Can. 1
Microbio!. 28, 1002-1013.
COLLINS, M.D.; MARTINEZ-MURCIA, A.J. Y CAl, 1. (1993).Aeromonasenteropelogenesand
Aeromonasichthiosm¡aare identicalto Aeromonastrota andAeromonasveronii,
respectively,asrevealedby small-subunit rRNAsequence analysis.fiat. J. Food
Microbiol. 43, 855-856.COLWELL, R.R.; MACDONELL, MT. Y DE LEY, j. (1986>. Proposalto recognizefamily
Aeromonadaceaefam. nov.¡nL ¿ Syst.Bacteriol. 36, 473-477.
CONDON, 5.; GARCIA, ML.; OTERO, A. Y SALA, Fi. (1992). Effect of culture age,
preincubationat low temperatureand pH en the thcrmalresistenceof Acromonas
hxdrophila. J. Appl. Bacteriol. 72. 322-326.
fil:

t
9,
“4
pu
pu
RIBLIOGRAFÍA
CORREA, A.; PAIVA, M.; MOREIRA, A. Y DELGADO. 1. (1993). Occurrence and
characterizationofAernmonasspeciesin pasteurizedmillc andwhite cheesein Rio de
JaneiroBrazil. ¿ Food Protect. 56, 62-65.
CROSA,1. (1989). Genetiesandmolecularbiology of siderophore-mediatediron transport
in bacteria.Microbio). Rey.53. 5 17-530.
CUMBERBATCH, N.; GURWITH, MI; LANGSTON, C.; SACK, RE. Y BRUNTON, j. (1979).
Cytotonic enterotoxinpreducedby Aeromonashydrophila: relationship oftexigenic
isolates te diarrhoeal disease.¡nfra. ¡minien. 23, 829-837.
DAILY, D.P.; JOSEPH,S.W.; COOLBAUGH, J.C.; WALKER, R.l.; MERRELL, E.R, ROLLINS,
DM.; SEIDLER, R.J.; COLWELL, R.R. Y LISSNER, C.R. (1981). Associationof
Aeromonassobriawith humaninfections.¿Clin. Microbio). 24,66 1-664.
DAVIS, W.A.; KANE, J.G. Y GARAGUSI, V.F.(1978).HumanAeromonasinfection: areview
of the literature and a case reportof endocarditis.Medicine.57,267-277.
DE LA MORENA, Mi.; VAN, R.; SINGH, K.; BRIAN, M.; MURRAY, ME.; PICKERING. L.K.
(1993).Diarrheaassociated withAeromonasspeciesin children in day catecenters.
.1. Infect. Dis.168, 215-218.
DESMOND, E. Y JANDA, J.M. (1986). Growthof Aeromonasspecies en entericagars.J.
Clin. Microbio). 23, 1065-1067.
DEXTER, SL.; SULLIVAN. J.D. IR.; WILLLAMS, 1. III Y WATSON. S.W. (1975). Influenceof
substrate wettabilityen the attachmentof marine bacteria te varioussurfaces.AppL
Microbio). 30, 298-308.
DICKSON, WA.; BOOTHMAN. P. Y liARE. K. (1984). An unusualsou’teof hospital wound
infection. Brir. Mal J. 289, 1727-1728.
OTiLLON. J.K.; FUERST,JA.; HAYWARD, A.C. Y DA VIS, 0.1-1.6.(1986).A comparison offive
methodsfor assaying bacterialhydrophobicity.1 Microbiot Merh.6, 13-19.
DOBRESCU,L. (1978). EnterotoxigenicAeromonashydrophila from a caseof piglet
diarrhea.ZentrabL Veteunaermed.(B) 25.713-718.
DOMAN, D.B.; GOLDING, M.l.; GOLDBERG, H.J. Y DOYLE, R.B. (1989). Aeromonas
hydrophila colitis presentingas medically refractoryinflammatorybewel disease.
Am.¿ Gastroenterol.84, 83-85.
DOOLEY. J.S.G.;LALLIER, R.; SHAW, 0.11. Y TRUST, TJ. (1985). Electrephoretic and
immunochemical analysisof the lipopolysaccharidesfrom various strains of
Aeromcnashydrophila.¿ Bacteriol. 164, 263-269.
0001EV, 1.5.6.: LAILIER, R. Y TRUST, T.J. (1986). Surfaceantigensof virulcnt strains of
Aerornonc¡s hvdrophilo. Vet. Imniunol. Immunopothól.12. 339-344.

BIBLIOGRAFÍA
DOOLEY, J.S.G.;MCCUBBIN, W.D.; KAY, C.M. Y TRUST, T.J. (1988). Isolation and
biochemical characterizationof the S-layerprotein from a pathogenicA eromonas
hydrophilastrain.J. Bacteriol.170, 2631-2638.
DOOLEY, J.S.G. Y TRUST, TI. (1988). Surfaceprotein compositionof Aeromonas
hydrophila strains virulentfor fish: identification of a surfacearray protein. J.
Bacteriol. 170, 499-506.
DORSCH, M.; ASHBOLT, N.J.; COX, P.T. Y GOOMAN, A.E. (1994). Rapid identificationof
Aeromonasspeciesusing 165 rDNA targeted oligonucleotide primers:amolecular
approachbasedon screeningof environmentalisolates.J. Appl. Bacteriol. 77,
722-726.
DUBEY. RS,; SANYAL, S.C. Y MALHORTA. D.P. (1980). Purification of Aeromonas
hydrophila enterotoxinand its modeof action in experimentalmodel. In Nature
Toxins.eds.Eaker,D. y Wadston,T. Oxford: PergamonPress.
ECHEVERRIA. P.; SACK, Rif Y BLACKLOW, N.R. (1984). Prophylacticdoxycycline for
traveler’s diarcheain Thailand:furthersupportiveevidenceof Aeromonas hydrophila
as anentexiepathegen.Am. ¿ Epidemiol.120, 912-921.
EDDY, B.P. (1960). Tbeuseand meaningof theterm“psychrophilic”. ¿ Appl. Bacteriol.
23, 189.
EDDY, BY. Y KJTCI{ELL. KG. (1959).Cold-tolerantfermentadve gram-negative organisms
from meatand other seurces.¿Appl. Bacteriol.22, 57-63.
ELBASMIR. KM. Y MILLERSHIP, S.E. (1989). Haemagglutinating activityofAeromonasspp.
from different saurces;attempteduseas atyp’mg system.Epidemiol. Infect. 102,
221-229.
ELEY, A.; GEARY, 1. Y WILCOX, M.H. (1993). Growthof Aeromonasspp. at 40C andrelated
toxin production.Lea. Appl. Microbio). 16, 36-39.
ELLISON, R.T. Y MOSTOW, S.R. (1984). Pyogenicmeningitismanifestationsduring therapy
forAeromonashydrophilasepsis.Arch. ¡titen Mecí. 144,2078-2079.
ENFORS,5.0.; MOUN, O. Y TERNSTROM,A. (1979).Effect of packagingundercarbon
dioxide, nitrogenor air en themicrobial flora of pork stored at40C. ¿ Appl.
Bacterio).47, 197-208.
ERICKSON, J.P. Y JENKINS, P. (1992). Behaviourof psychrotrophicpathogensListeria
monocytogenes,Yersiniaenterocolitica andAeromonas hydrophilain commercially
pasteurizedeggsheldat 2,6.7 and 12.80C.¿ FoodProtecr.55, 8-12.
221

mu
mu
mu
pu
t
mu
pu
a
y
BIBLIOGRAFíA
ESTEVE,C.; AMARO, C. Y TORANZO, A.E. (1994). O-serogroupingand surfacecomponents
of AeromonashydrophilaandAeromonasjandaeípathogenicfor ceIs. FEMS
Microbiol. Len. 117, 85-90.
ESTEVE,C.; AMARO, C.; GARAY, E.; SANTOS,Y. Y TORANZO, A.E. (1995). Pathogenicityof
live bacteriaand extracellularproductsof motile Aeromonasisolatedfrom cels.1
Appl. Bacreriol. 78, 555-562.
EVENBERG,D. Y LUGTENBERG, 8. (1982). Cdl surfaceof the fish pathogenic bacterium
Aeromonassalmonicida.1. Purificationandcharacterizationofa majorecli envelope
proteinrelatedte autoagglutination.adhesion,and virulence. Biochim. Biophys.
Acta. 684, 249-254.
EVENBERG,D.; VAN BOXTEL, R.; LUGTENBERG, B. Y SCHURER,E (1982). Ceil surfaceof
the fish pathogenicbacteriumAeromonassalmonicida.1. Relationship between
autoagglutinationand thepresenceof a major celí envelope protein.Biochim.
Biophys.Acta.684, 241-248.
FAINSTEIN, V.; WEAVER, 5. Y BODEY, G.P. (1982). In vitro susceptibility ofAeromonas
hydrophilaagainst newantibiotics.Antimicrob.AgentsChemoiher.22,513-514.
FANNINO, G.R.; HIKMAN-BRENNER, F¿W.; FARMER, JJ.llI. Y BRENNER,Di. (1985). DNA
relatednessandphenotypicanalysisof the genusAeromonas.Abstr. C- 116. Annual
Meetingof theAmericanSocietyfor Microbiology.
FARIS, A.; LINDAHL, M. Y WADSTROM, T. (1982). High surface hydrophobicity of
hemagglutinatingVibrio choleraeandetherVibrios. Curr. Microbio). 7,357-362.
FARMER, 3.1.111.;H¿IKMAN-BRENNER, F.W.; FANNING,G.R.; ARDIJINO, M.J. Y BRENNER,Di.
(1986).Analysis ofAeromonasandPlesiomonasby DNA-DNA hybridization and
phenotype.Abstr. P-1. lst lnt. WerkshopAeromonasPlesiomonas.
FARRAYE, F.A.; PEPPERCORN,MA.; CIANO, ¡‘.5. Y KAVESH, W.N. (1989). Segmental
colitis associatedwith Aeronwnashydrophila.Am.1 Gastroenterol.84, 436-438.
FASS,RJ. Y BARNISHAN, J. (1981). In vitro susceptibilityof Aeromonashydrophilato 32
antibioties.Antimicro!’. AgenrsChemorber.19, 357-358.FERREIRA,MA.S.S.Y LUNO B.M. (1987).Theinfluenceof pH andtemperatureen initiation
of growth of Sa)monellaspp.Len. Appl. Microbio). 5, 67-70.
FIGURA, N. Y GUGLIELMETfl, P. (1987). Differentiationof motile and mesophilic
Aeromonasstrainsinto speciesby testingfor CAMP-LIKE factor.J. Clin. Microbiol.
25, 1341 1342
991

BIBUOGRAFIA
FIGURA, N.; MARRI, L.; VERDIANI, 5.; CECCHERINI,C. Y BARBENI, A. (1986). Prevalence,
speciesdifferentiationand toxigenicity of Aeromonasstrainsin casesof childhood
gastroenteritisand in controls.1 Clin. Microbio). 23. 595-599.
FINLAY, BE. Y FALKOW, s.(1989).Commonthemesin microbialpathogenicity.Microbio!.
Rey.53, 210-230.
FLETCHER, M. Y LOEB, Gí. (1979). Influence of substratumcharacteristicson the
attachmentof a marinepseudomonadte solid surfaces.Appl. Environ. Microbio).
37, 67-72.
FRANCKI, K.T. Y CHANG, EJ. (1994). Variableexpressionof 0-antigenand therole of
lipopolysaccharideasan adhesinin Aeromonassobria. FEMSMicrobioL Lea. 122,97-100.
FREU, B.J. (1987). ExtraintestinalAeromonasvid Plesiomonasinfections of humans.
Experientia.43, 359-360.
FRICKER, C.R. (1987). Serotypingof mesophilic Aeromonasspp. en the basis of
Iipopolysaccharide antigens.Lea. Appl. Microbiol. 4, 113-116.
FRICKER, C.R. Y TOMPSETY, 5. (1989).Aeromonasspp. in foods: asignificantcauseof
feod poising?. Ini. ¿ FoodMicrobio). 9, 17-23.
GARCíA DE FERNANDO. GiD. Y FOX, RF. (1991). Extracellularpreteinasesfrom
MicrococcusOF: 1. Factorsaffectinggrowthandproduction.Lait. 71, 359-370.
GARCIA-LOPEZ. M.L.; OTERO, A.; GARCIA-FERNANDAZ, M.C. Y SANTOS J.A. (1993).
Incidencia,comportamientoy control deAeromonas hydrophilaen productos
cárnicosy lácteos.Microbiología.9,49-56.
GARCIA-RODRIGUEZ,J.A. (1987). Elementos facultativos: cápsula,glicoc~1ix, flagelosy
fimbrias. ElesporoEnMicrobiologíay ParasitologíaMédica.Pp.47-55.Barcelona:
Salvat.
GAEDNER,O.K (1965).Theaerobicflora ofstered meatwith panicularreferenceto the use
of selectivemedia.1 AppL BacterioL28, 252-264.
GARLAND, WJ. Y BUCKLEY, iT. (1988).Tic cytolytic toxin aerolysinmust aggregatete
disrupterythrocytes,and aggregationis stimulated byhumanglycephorin.Infect.
lmmun.56, 1249-1253.
GEORGE.WL.; NAKATA, MM.; THOMPSON,J. Y WHITE, ML. (1985).Aeromonas-related
diarcheain adults.Arch. Interia. Med.145. 2207-2211.
GEORGE, W.L.; iONES, Mi. Y NAKATA, MM. (1986). Phenetypiccharacteristicsof
Acm monasspecies isolatedfrom adult human J. Clin. Microbio?. 23, 1026-1029.
223

nr
mu
mu
“4
mu
mu
rL
mu
mu
u
mu-
BIBLIOGRAFÍA
GERSON, D.F. Y SO-lEER, 0. (1980). Ccii surfacecnergy, contact anglesand phase
partition. III. Adhesionof bacterialcelís tehydrephobic surfaces.Biochim. Biophys.
Acta.602, 506-510.
GHANEM. EH.; MUSSA, MuE. Y ERAL, HM. (1993).Aeromonasassociatedgastroenteritis
in Egypt. Zentralbí.Mikrobiol. 148, 441-447.
GILL, C.O. Y REICHEL, MP. (1989). Growth of 0w cold-tolerantpathogensYersinia
enterocolitica, AeromonashydrophilaandListerianwnocyrogeneson high-pH beef
packagedundervacuumor carbondioxide. FoodMicrobio). 6, 223-230.
GLUSKIN, 1.; BATASH, O.; SHOSEYOV,O.; MOR, A.; KAZAK, R.; AZUl, E. Y BOLDUR, 1.
(1992). A l5-year study of the role of Aeromonasspp. in gastroenteritisin
hospitalizedchildren.1 MaL Microbio). 37,315-318.
GOBAT, ¡‘.F. Y JEMMI, T. (1993).Distribution of mesophilicAeromonasspeciesin raw vid
readyte eatfish and meat productsin Switzerland.Inc ¿ FoadMicrobio!. 20,
117-120.
GOBAT,P.F. Y JBMMI, T. (1995>.Comparisonof sevenselectivemediafor the isoladonof
mesophilic Aeromonasspeciesin fish and meat. ¡ni. ¿ Food Microbio!. 24,375-384.
GOLO, W.L. Y SALIr, LE. (1993).Aeromonas hydrophilainfectionsof sUnvid seft tissue:
reponof 11 casesvid review. Clin. Infecr. Dis. 16, 69-74.
GOLDEN, D.A.; ENTES,M.J. Y BEUCHAT, L.R. (1989). Influenceof modified-atmosphere
storageen thegrowthof uninjuredvid heat-injuredAeromonashydrophilo.AppL
Enviran. Microbio!. 55, 3012-3015.
GOODWN, C.S.; HARPER,W.E.S.; STEWART, J.K.; GRACEY, M.; BURKE, y. y ROBINSON.1.(1983).EnterotoxigenicAeromonashydrophila¿mddiarrheain adults.Med.1 Aus:.
1, 25-26.GORCZYCA, E.; SUMNER, S.L.; COHEN. O. Y BRADY, pu (1985).Mesophilic fish spoilage.
Foad Techno).Aust.37, 24-26.GOSLING. Pi; TURNBULL, P.C.; LIGHTFOOT, N.F.; PETEER,1.V.; LEWIS, R.J. (1993).
Isolation and purification of Aeromonassobria cytotonic enterotoxinvid beta
haemolysin.¿ Med Microbio). 38, 227-234.
GRA¿BOW,W.O.K. Y DUPREEZ, M. (1979). Comparisonof ni-Ende LES, McConkey,and
Teepolmediafor membranefilter countingat total coliform bacteriain water.Appl?
Environ. Microbiol. 38, 35 1-358.
GRACEY, M.; BURKE, V. Y ROBLNSON, 1. (1982).Acromonassubspeciesin travefler’s
diarrhoca.&it. Mcd. J. 289. 658.
224

BIBLIOGRAFíA
GRACEY, M.: BIJRKE, Y. Y ROBINSON, J. (1984).Aeromonas-associatedgastroenteritis.
Lancer.ji, 1304-1306.
GRAM, L. (1991). Inhibition of mesophilic spoilageAeromonasspp. on fish by salt,
potasiumsorbate,liquid smoke, andchilling. ¿ Food Protect.54. 436-442.
GRAM, L.; TROLLE. G. Y HUSS, H.H. (1987). Detectionof specific spoilagebacteriafrom
fish storedat low (00C) vid high (20~<2) temperatures.mt. ¿ FoodMicrobiol. 4.
65-72.
GRAU, F.H. (1981). Role ofpH, lactatevid anaerobiosisin controlling thegrowthof some
fermentativegram-negativebacteriaon beef. AppL Environ. Microbio). 42,
1043-1050.
GRAY. RA. Y KIROV, S.M. (1993).Adherencete HEp-2 cellsvid enteropathegenicpotential
of Aeromonasspp. Epidemiol. Infecr. 110, 279-287.
GRAY, S.J. (1984). Aeromonas hydrophi)a in livestock: incidence, biochemical
characteristicsvid antibioticsusceptibility.¿Hyg.92, 365-375.
GRAY, S.J.; STICKER, D.J.Y BRYANT, T.N. (1990). Theincidenceof virulencefactorsin
mesophilicAeronwnasspecies isolatedfrom farm anima.lsand theirenvironment.
Epidemiof. Infeci. 105, 275-294.
GREENAWAY,c. (1988).Aeromonas:a futureconcernfor the dairyvid foed industries.In
PieDairy Technologisr.pp. 73-75.Victoria, Australia:Feod Research Institute.
GREY. RA. Y KIROV, 531. (1993).Adherence te HEp-2 celísvid enteropathogenicpotential
ofAeromonasspp.EpidemioLInfecí. 110,279-287.
HANES, D.E. Y CHANDLER, D.K. (1993). Therole of a 40-megadalton plasmidiii the
adherence andhemolytic propertiesofAeromonashydrophi)a.Micro!’. Pat/zog.15,
313-317.
HANNE, L.F. Y FINKELSTEIN, R.A. (1982). Characterizationand distribution of the
hemagglutininsproducedby Vibrio cholerae.lnfecr. ¡minien.36, 209-214.
HANNINEN, M.L. (1993).OccurrenceofAeromonasspp. in samplesof groundmealvid
chicken.¡nr. J. Foodmicrobio!. 18, 339-342.HANNINEN, ML. (1994). Phenotipiccharacteristicsof the threehybridation groupsof
Aeromonashydrophilacomplex isolatedfrom different sources.J. Appl. Bacteriol.
76, 455-462.
HARDY, J.C.; TODD, L.S. Y STRINGER,M.F. (1986). Toxin production by Aeromonas
hydrophilain bactercological mediaandfoods.Zentralbí. Bakteriol.Mikrobiol. Hyg.
L AbetellungSuppl. 15, 175-176.
‘OS

r
9
“4
U
“4
“4
U,
U
o
U,
“4
9,
u
B(BEJOGRAFÍA
HARRIS, RL.; FAINSmIN, V. Y ELTING, L. (1985). Bacteriemiacausedby Aeromonas
speciesin hospitalizedcancerpatients.Rey. ¡nfDis. 7. 314-320.
HAVELAAR, AH.; DURING, M. Y VERSTEEGH, 3P.M. (1987). Ampicillin-dextrin agar
mediumfor tic enumerarionof Aeromonasspeciesin water by mnembranefiltration.
1 AppL Bacteriol. 62, 279-287.
HAVELAAR, A.H. Y VONK, M. (1988). Thepreparatienof ampicillin dext¡-in agarfor tite
enumerationof Aeromonasin water.Len. Appl. Microbiol. 7. 169-171.
HAVELAAR, Adj.; SCHETS,F.M.; VAN SILFHOUT, A.; JANSEN,W.H.; ‘NIElEN, G. Y VAN DER
KOOJ. D. (1992).Typing ofAeromonasstrainsfrom patientswith diarrhea and frem
drinking water.£ AppL Bacteriol.72, 435-444.
HAZEN, Tt. (1983).A model for tite densityofAeromonashydrophi)ain Albemarle Sound
North Carolina.MicrobioL Ecol.9, 137-153.
HAZEN, T.C.; FLI¿ERMANS, C.B.; HIRSCII. Rl>. Y ESCH. G.W. (1978). Prevalenceand
distributionofAeromonas hydrophilain tite United States.AppL Enviren.Microbio).
36, 731-738.
HEIXIES, R.W.; SMITH, 1’. Y BRAZIL, 0. (1985). Resistanceplasmidsof aeromonads.J.
Gen.Microbio). 131, 2091-2095.
HEYWOOD. R. (1968).Aeromonasinfectionsiii snakes.Come))Va 58, 236-241.
HIKMAN-BRENNER, F.W.; MACDONALD, K.L.; STELNGERWALT, KG.; FANNING, GR.;
BRENNER, Di. Y FARMER, j.~.m. (1987>. Aeromonas veronii,a new ornititine
decarboxilase-positive speciesthat maycausediarrhea.J. C)in. Microbio). 25,
900-906.
1IIXIMAN-BRENNER, F.W.; PANNING, G.R.; ARDUINO,Mi.; BRENNER, [U. Y FARMER,Ji. III.
(1988).Acromonasschuberrii,a new mannilol-negativespeciesfound in human
clinie specimens.¿ Clin. Microbio). 26,1561-1564.
HO, A.S.Y.; MIETZNER,LA.; SMITH, AJ. Y SCHOOLNIK, O.K. (1990).The pu of Aeromonas
hydrophila: identificat.ionofan environmentallyregulated“mini pum”.¿ Exp. Mecí.
172, 795-806.
HOLMBERG, SD. Y FARMER, Ji. (1984). Aeromonashydrophi)a and Plesiomonas
shigelloidesascausesofintestinalinfections.Rey.¡nf Dis. 6, 633-639.
HOLMES, P. Y SARTORY, D.F.(1993).An evaluationof mediafor tite membranefiltration
enumerationofAeromonosfrom drinkingwater.Lea. Appl. Microbio). 17,58-60.i-IOKAMA. A.; IJUNMA. Y. Y NAKASONE, N. (199(J).PiIi of anAeronwnashydrophila strain
asa possiblecolenizationfactor.Microbio!. ¡mmuncl.34,901-915.
226

fiIBUOGRAFÍA
HOKAMA, A. Y IWANAGA. M. (1991).Purification andcharacterizationofAeromonassobria
pili, a possible colonization factor.Jnfect.Iminun.59, 3478-3483.I-IOKAMA, A. Y IWANAGA, M. (1992).Purification ¿mdcharacterizationofAeromonassobria
Ae24pili: a possible newcolonizationfactor.Micro!’. Pathog.13, 325-334.
I-IONMA, Y. Y NAKASONE, N. (1990). PiIi of Aeromonas hydrophila: purificatien,
characterization,andbiological role. Microbio). Immnunol.34, 83-98.
HOSTACKA. A.; CIZNAR, 3.; KORYCH, 8. Y KAROLCEH. 3. (1982). Toxie factors of
Aeromonashydrophilaand Plesiomonas shigelloides.Zentralbí. Bakteriol.Hyg.
252, 525-534.
HOUSTON, C.W.;CHOPRA, A.K.; ROSE,¿EM. Y KUROSKY, A. (1991). Reviewof Aeromonas
enterotoxins.Experientia. 47,424-426.
HOWARD, S.P. Y BUCKLEY, ¡ir. (1985). Activationof tite hole-forming toxinaerolysinby
extracellularprocessing.¿ BacíerioL 163, 336-340.HOWARD, S.P. YBUCKLEY, J.T. (1986). Molecular cloning¿md expressionin Escherichia
coli of tite structural gene for the hemolytic toxin aerolysin from Aeromonas
hydrophila.Mol. Gen.Ocian. 204, 289-295.
HSU, T.C.; WALTMAN, ‘N.B. Y SHOfl’S E.B. (1981). Correlationof extracellularenzymatic
activity and biochemicalcbaracteristicswith regardte virulence of Aeromonas
hydrophlla.Dep. Rbi Stand 49, 101-111.
HUDSON, J.A.(1992). Variationin growth kinetics andphenotypeof Aeromonasspp.fromclinical, meat processingand fleshfood sources.Ini. ¿ Food Microbio). 16,131-139.
HUDSON,¿LA. Y DE LACY. K.M. (1991).Incidence motileAeroi’nonadsin New Zeland retail
foods.1 Food Prorec. 54, 696-699.
HUDSON, LA. Y MOTT, S.J. (1993). Presencede Lysteria monocy:ogenes,motile
aeromonadsand Yersinia enterocolitica in environmental samplestakenfrom a
supermarketdelicatessen.¡ni. ¿ FoodMicrobiol. 18,333-337.HUDSON. J.A. Y AVERY, SM. (1994). Growthof Lisíeria monocyíogenes,Aeromonas
hydrophi)ay Yersiniaenierocoliricaon cooked musseltissueunderrefrigerationand
mild temperatureabuse.J. FoodSaIny. 14, 41-52.
HUGUET, J.M. Y RIBAS, F. (1991). SGAP-1OCagarfor the isolationand qualification of
Aeromonasfi-orn water.¿AppL Bacteriol.70, 81-88.
HUNTER. PR. Y SURGE,SAl. (1987).Isolation of Aerontonascaviae from ice-cream.Lert
AppI. Microbio!. 4, 45-46.
227

mu
pu
9
U,
U
U,:
9-
mu
BIBUOCRAFÍA
HUSSLEIN, y.; BERGBAUER. ¡-1. Y CI-IAKRAHORTY, T. (1991). Studieson aerolysinand
serme proteasefrom Aeromonastrata sp.nov. bxperientia.47,420-421.
HUSSLEIN, y.; CHAKRABORTY, T.: CARNAHAN, A. Y iOSEPH. SW. (1992). Molecular
studies entite aerolysingeneofAerornonasspecies¿mddiscoveryof species-specific
probeforAerornonasu-ctaspeciesnova. Clin. !nfecr. Dis. 14. 1061-1068.
HUYS, O.; VANCANNEYT, M.; COOPMAN, R.; JANSSEN, It; FALSEN, E.; ALTWEGG, M. Y
KERSTERS,K. (1994).CeIlular fatty acidcompositionas a chemotaxonomicmarker
for the differentiation of pitenospeciesand hybridization groupsin the genus
Aeromonas.mt. ¿ Sysr. Bacterio). 44, 651-658.
HUYS. G; KERSTERS,1.; VA¿NCANNEYT, M.; COOPMAN, R.; JANSSEN, It Y KERSTERS, K.
(1995). Diversityof Aeromonas sp. lii Flemishdrinking water productien plants as
determinedby gas-liquidchromatographicanalysisofcellular fatty acidmethyl esters
(FAMEs).¿ Appl. Bacteriol.78, 445-455.
IBRAHIM, A. Y MACRAE, f.C. (1991). Incideuceof Aeromonasvid Listeria spp. in redmeat¿mdmilk sampleslii Brisbane, Australia.¡nL ¿ FeodMicrobiol. 12, 263-270.
INGHAM, S.C. Y POlTER,N.N. (1988). GrowthofAeronwnashydrop/zilaami Pseudomonas
fragi en minee and surimis madefrom Atiantie pollock and storedunderair or
modified aymosphere.¿ FoodProtect. 51, 966-970.
INGHAM, s.C.(1990). Growthof Aeromonashydrophilavid Plesiomonasshigelloidesen
cooked crayfish tajís during caíd storageunder air, vacuum, and modified
atmosphere.J. Food Protect.53, 665-667.ISHIGURO, LE.; KAN, W.W.; AINSWORTH, T.; CHAMBERLAJN, J.B.; AUSTEN, RA.; BUCKLEY,
J.T. Y TRUST, T.J. (1981). Lossof virulenceduring cultureofAeromonas salmonicido
at high temperature.¿ Bacteriol. 148, 333-340.ISHIGURO, E.E.; AINSWORTH, ‘E; TRUST, Ti. Y KAY, W.W. (1985). Congo red agar, a
differential mediumfor Aeromonassa)monicida,detectste presenceof tite ceil
surfaceprotein arrayinvolved iii virulence.1 Bacteriol.164, 1233-1237.
IWANAGA, M. Y HOKAMA, A. (1992). CharacterizationofAeromonassobriaTAP 13 pili: a
possible new colonizationfactor.¿ Gen.Microbiol. 138, 1913-1919.JANDA, 3M. (1991).Recentadvancesin tite study of tite taxononiy,pathogenicityand
infectioussyrxdromesassociatedwith tite genusAeromonas.Clin Microbiol. Rey.4.
397-410.
JANDA, AM.; ¡3(YITONL, EJ. Y REITANO, M. (1983a). Aeromonasspeciesin clinical
microbiology: significance,epiderniology,ami speciation.Dio gn. Microbio!. Infecr.
Dis. 1, 221-228.

BIBLIOGRAFÍA
JANDA, J.M.; BOTT’ONE. EJ.: SKINNER, C.V. Y CALCATERRA, 1). (1983b). Phenotypicmarkersassociatedwith gastrointestinalAeromonas hydrophilaisolatesfrom
symptomatic children.¿ Clin. Microbiol. 17. 588-591.
JANDA, f.M.; BRENDEN, R. Y BOTTONE,Ef. (1984a).Differential susceptibility to human
serumby Aeromonasspp. Curr. Microbio). 11, 325-328.
JANDA, JkMu; REITANO, M. Y BOrIONE, EJ. (1984b). BiotypingofAeronwnasisolates as a
correlatete delineatinga species-a.ssociated diseasespectrum.¿ Clin. Microbiol. 19.44-47.
JANDA, J.M. Y MOTYL. M.R. (1985). Cephalothinsusceptibilityasa potentialmarkerforthe
Aeromonassobriagmup.¿ Clin. Microbiol. 22, 854-855.JANDA, J.M. Y BRENDEN, R. (1987). Importanceof Aeromonassobria in Aeromonas
bacteriemia.¿ Infect. Dis. 155, 589-591.
JANDA, J.M.; OSHIRO. L.S.; ABBO1T. S.L. Y DUFFEY. P.S. (1987). Virulence markersof
mesophiicaeromonads:associationof theautoagglutinationpitenomenonwith meuse
pathogenicityand thepresenceof a peripheral cell-associated layer.¡nfect. ¡minun.
55, 3070-3077.
JANDA, ¿EM. Y DUFFEY. P.S. (1988). Mesophilic aeromonadsin humandisease:current
taxonomy, laboratoryidentificationandinfectiousdiseasespectrum.Rey.Infect.Dis.
10, 980-997.
JANDA, J.M.; GUTHERTZ.L.S.; KOKKA, R.P. Y SHIMADA, T. (1994a).Aeromonasspeciesin
septicemia:laboratorycharacteristicsami clinical observations.Clin ¡rifen. Dis. 19,
77-83.JANDA, J.M.; KOKKA, R.P. Y GUTHERTZ,L.S. (1994b).Thesusceptibilityof S-layer-positive
vid S-layer-negativeAeromonasstrainsto complement-mediatedlysis. Microbiology.
140, 2899-2905.
JANOSSY, G. Y TARJAN, V. (1980).Enterotoxigenicityof Aeronwnasstrainsin sucklingmice. ActaMicrobioL Acad.Sci. Hung. 27, 63-69.
JAY, J.M. (1967). Natura, characteristics,vid proteolytic propertiesofbeefspoilage bacteria
at low vid high temperatures.AppL Environ. Microbio).15, 943-944.
JENSSEN,D. (1984). Aquantitativetestfor mutagenicityin V79 chinese hanistercelís.In
HandbookofMutagenicityTenProcedieres.eds.Kilbey, B.J.; Legator,M; Nichols,
W. y Rainel,C. pp.269-290.Amsterdam:ElsevierSelencePublishersBV.
22<>

“4
“4
mu
“4.
e-
“4
u
mu
a
a
U,
e
BIBLIOGRAFÍA
MANO, Z.D.; NELSON, A.C.; MATUEWSON. 1.3.; ERICSSON, C.D.Y DUPONT, 1-IL. (1991)
Intestinalsecretoryimmune responseto infection with Aeromonasspecies and
Plesiornonas shigelloidesamong studentsfrom dic UnitedStacesin Mexico.¿ Infect.
Vis. 164, 979-982.ITWA, S.F.H.(1983).Enterotoxigenicity. haernagglutinationandceil-surfacehydrophobicity
in Aeromonashydrophila,A. sobriaandA. salmonicida. Ver. Microbio). 8, 17-34.
JOHNSON, W.M, Y LIOR, H. (1981). Cito¡oxicity and suckling meuse reactivity of
Aeromonas hydrophila isolatedfrom human seurces,Can. J. Microbio). 27,
1019-1027.
IONES.G.W. (1977).Re attachmentof bacteria totite surfacesof animalcelís.ed. Reissig,
EL. In Receptors and recognirion, seriesE, vol. 3. Microbial Inreracrioris. pp.
139-176. London: Chapmany Hall.
JOSEPH,S.W.; COLWELL, R.R. Y MACDONELL, M.T. (1987). Acromonas taxonomy.
Experientia.43,349-350.
KALINA, Gr. (1977).Aeromonadsin feod productsand titeir possiblerole ascausative
agentsof feod poisoning.Gigiena¡ Sanirarila (Moskva).8, 97-100.
KAMPFER, P. Y ALTWEGG, M. (1992). Numerical classification and identificatien of
Aeromonasgenospecies.¿AppL Bacterio).72, 341-351.
KAMPFER, P.; BLASCZYK, K. Y AULING, 0. (1994).CharacterizationofAeromonasgenomie
speciesby usingquinone,polyamine,andfatty acid pattems.Can. J. Microbio). 40,844-850.
KANAI, K. Y WAKABAYASHI, H. (1984). Purificationami somepropertiesofproteasefrom
Aeromonashydrophila.BuIL Jap. Soc. Sé.Fish. 50, 1367-1374.
KAREM, 1<. U; FOSTER,5W, Y BEl, A.K. (1994).Adaptativeacid tolerante response (ATR)
in Aeromonashydmophila.Microbiology. 140, 1731-1736.
KAPER, 1.; SEIDLER, R.J.; LOCKMAN, H. Y COLWELL, R.R. (1979). Medium for the
presumptiveidentificationofAeromonashydrophilavid Enrerobacteriaceae.Appl.
Envir. Microbiol. 38, 1023-1026.
KAPER, LB.; LOCKMAN, H. Y COLWELL, R.R. (1981).AeronwnashydrophUa:Ecelogyvid
toxigenicityof isolatesfrom mi estuarie..1. AppL Bacteriot50, 359-377.
KARUNAKARAN, T. Y nEW, B.G. (1994). Characterizationof haemolytic activity from
Aeromonascaviae.EpidemioL¡nfect. 112, 291-298.
KAY, WW.; BUCKLFIY, J.T.: ISIIIGLRO, LE.; PHIPPS,BM.; MONEITE, J.P.L. Y TRtJS’I, T.i.
(1981). Purificationanddispotition of a surfaceprotein associaíedwith virulenceof
Aeromonos solmonicida.J. PaceÑo!.147, 1077-1084.
130

BIBLiOGRAFÍA
KAY, W.W.; PHIPPS, BM.; ISHIGURO, EF.; OLAFSON, R.W. Y TRUST, T.J. (1984). Surface
layer virulenceA-proteinsfrom Aeromonassalmonicidastrains.Can. ¿ Biochim.
Celí Biol. 62, 1064-1071.
KAY, W.W.; PHIPPS,BM.; ISHIGURO, Ef. Y TRUST, ‘EJ. (1985). Porphyrin bindingby tite
surfacearray virulenceprotein of Aeromonassalmonicida.J. Bacteriol. 164,
1332-1336.
KIELWE1N, G. (1969).Sn nahrbodenzur selektivenzuchtungvon PseudonwnadenundAeromonaden.Arch. Lebensmittelhyg.20, 131-133.
KIELWEIN, G.; GERLACH, R. Y JOHÁ”JE, H. (1969).PrevalenceofAeromonashydrophilain
rawnúlk. Arch. Lebensminelhyg.20,34-38.
KINDSCHUCH, M.; PICKERING, L.K.; CLEARY, T.G. Y RUIZ PALACIOS, G. (1987). Clinical
andbiochemicalsignificanceof toxin productionby Acromonashydrophlla.¿ Clin.
Microbiol. 25, 9 16-921.
KIROV, S.M. (1993).The public healthsignificanceof Aeromonasspp. in foeds.liar. 1
FoodMicrobiol. 20, 179-198.
KIROV, S.M.; REES, B.; WELLOCK, R.C.; GOLDSMID, ¿EM. Y VAN GALEN, A.D. (1986).
VirulencecharacteristicsofAero,nonasspp. in relationte sourcevid biotype.¿Clin.
Microbiol. 24, 827-834.
KIROV. S.M.; ANDERSON, Mi. Y MCMEEKIN, T.A. (1990). A note enAeromonasspecies
from chickensaspossible feod-bornepathogens.1 AppL Bacteriol.68, 327-334.
KIROV. S.M. Y BRODRIBB, F. (1993). Exotoxin productionby Aeromonasspp. in foods.
Lett AppL Microbio).17, 208-211.
KIROV, SM. Y HAY WARD, Li. (1993).Virulence traitsofAeromonasin relationto species
vid geographicregion.Aust. ¿ Mecí. Sci. 14, 54-58.
KIROV, SM.; ARDESTANI, E.K. Y HAYWARD, Li. (1993a).Tite growth andexpressionof
virulencefactorsat refrigeTationtemperaturesby Aeromonasstrains isolated from
foods. ¡nr. J. FoodMicrobiol. 20, 159-168.KIROV, S.M.; HUDSON, J.A.; I-IAYWARD, Li. Y MOTT, S.J. (1993b). Distribution of
Aeromonashydrophila hibridization groups and iheir virulence propertiesin
Australasiandinical andenvironmentalstrains.Lett AppL Microbio). 18,71-73.
KIROV, SM.; HUI, n~. y HAYWARD, Li. (1993c).MiIk as apotentialsourceofAeromonas
gastrointestinalinfection.1. FoodProtea. 56, 306-312.
KIROV, S.M.; HUI)SON, JA.; HAYWARD, L.J. Y MOTF, S.J. (1994). Distribution of
Acromonas hydrophila hybridizationgroups and titeir virulence propertiesin
Australasianclinical and environmentalstrains.Len. Appl. Microbio?. 18, 7 1-73.
231

“4
“4-
0
mu
mu
BIBLIOGRAFÍA
KLEEBERGER, A. (1975). Taxonomyof Aeromonadsfrom milk and other foods.
Milchwissenschaft.36. 602-603.
KNOCI-IEL, S. (1989a). Effect of temperatureen hemolysin production in Acromonas
speciesisolated from warm and coid environments.liar. 1 Food Microbio). 9,
225-235.
KNOCHEL, 5. (1989b).Thesuitability of feur mediafor enumeratingAeromonasspecies
frem environmentalsamples.Len. Appl. Microbiol. 9, 67-69.
KNOCHEL, 5. (1990). Growth characteristicsof motile Aeromonasspp. isolatedfrom
different environments.¡nr. ¿ FoodMicrobiol. 10, 235-244.
KNOCHEL, 5. Y JEPPESEN,C. (1990).Distributionandcharacteristicsof Aeromonasin food
and drinking waterin Denmark.¡nL 1 FoodMicrobiol. 10, 317-322.
KOBAYASHI, K. Y OHNAKA, 1. (1989).Foodpoisiningdueto newly recegnized pathogens.
Asian Mecí. J.32, 1-12.
KOKKA, Rl’.; VEDROS, N.A Y JANDA 3M. (1990). Electrophoreticanalysisof tite surface
componentsof autoagglutinating surface array protein-positivevid surfacearray
protein-negativeAeromonas hydrophilavid Aeromonassobria,£ C)in. Microbio).
28, 2240-2247.
KOKKA, Rl’.; JANDA J.M.; OSHIRO, L.S.; ALrHEGG, M.; SHIMADA, T.; SAKAZAKI, R. Y
BRENNER,Di. (1991a).Biochemicaland geneticcharacterizationofautoagglutinating
phenotypesof Áeronwnasassociatedwith invasiveand noninvasive disease.J.
Jnfecr. Dix. 161, 890-894.KOKKA, RS’.; VEDROS,N.A. Y JANDA 3M. (199ib). Characterizationof classicvid atypical
serogroup0:11Aeromonas:evidencethat thesurfacearrayin proteinis not directly
involved in meusepathogenici¡y.Microb. Pat/mg.10,71-79.
KOKKA, Rl’.; VEDROS, N.A. Y JANDA J.M. (1992). Immunochemicalanalysisvid possible
biological roleof vi Aeromonashydrophilasurfacearrayprotein in septicaemia.1
Gen. Microbio).138. 1229-1236.
KOZAKI, 5.; KATO, K.; ASAO, 1.; KAMATA, Y. Y SAKAGUCHI, G. (1987). Activities of
Aeromonashydrophilahemolysinsvid theirinteractionwith erythrocytemembranes.
¡nfect. ¡minien. 55, 1594-1599.
KOZAKI, 5.; ¡CATO, K.; KUROKAWA, A.; ¡CAMATA, Y.; ASAO, T. Y SAKAGUCHI, G. (1988).
Productionof monoclonalantibody againstAcromonas hydrophilahemolysin.J.
Metí. Microbiol. 25, 187-190.
232

BIBLIOGRAFÍA
KROVACEK, K, PETERZ, M, FARIS, A. Y MANSSON, 1. (1989). Enterotoxigenicityand
sensitivity of Aeromonashydrophila isolatedfrom well water in Sweden:a case
study. mr. 1 FoodMicrobiol.8, 149-154.
KROVACEK, K.; FARIS, A. Y MANSSON. í. (1991). Growth and toxin preduction by
A eromonashydrophila andAeromonassobria at low temperatures.¡nr. ¿ Food
Microbiol. 13, 165-175.KROVACEK, K.; FARIS, A.; BALODA, S.B.; LINDBERG, T.; PETERZ, M. Y MANSSON, 1.
(1992a).Isolatienvid virulenceprofilesofAeromonasspp. from differentmunicipal
drinkingwatersupplies inSweden.FoodMicrobio!. 9,215-222.
KROVACEK, K.; FARIS, A.; BALODA, S.B.; PETERZ, M.; LINDBERG, T. Y MANSSON, 1.
(1992b).PrevalenceandcharacterizationofAeronwnasspecies isolatedftom foodsiii
Uppsala Sweden.FoodMicrobio). 9, 29-36.
KROVACEK, K.; CONTE,M.; GALDERISI, P.; MOELLI, G; POSTIGLIONE.A. Y DUMONTET, 5.
(1993).FatalsepticaemiacausedbyAeromonashydrophilain a patientwith cirrhosis.
Comp.¡minunoL Microbio). ¡nfecr. Dis. 16, 267-272.
KROVACEK, K.; PASQUALE, y.; BALODA, 5.; SOPRANO, y.; CONTE, M. Y DUMONTET, 5.
(1994). Comparisonof putativevirulencefactors inAeromonas hydrophilastrains
isolated fromtite marine environmentandhuman diarrheal casesin SouthemItaly.
Appl. Environ. Microbio). 60, 1379-1382.
KUHN, 1.; LINDBERG, Y; OLSSEN, K. Y SThNSTROM, T.A. (1992). Biochemical
fingersprintingfor typing of Aeromonasstrainsfrom food and water.Lea.Appl.
Microbio). 15, 261-265.KIMJPER, EJ.; BOL, P.; PEETERS,M.F.; STEIGERWALT, A.G.; ZANEN, H.C. Y BRENNER,Di.
(1989a).Clinical and epidemiologicaspectsof membersof AeromonasDNA
hybridization groupsisolatedfroni humanfeces.¿ Clin. Microbio). 27, 1531-1537.
KUIJPER. EJ.; STEIGERWALT, A.G.; SCHOENMAKERS, B.S.C.I.M.; PEETERS, M.F.; ZANEN,
H.C. Y BRENNER, Di. (1989b).Phenotypiccharacterizationand DNA relatednessin
humanfecal isolatesof Aeromono.sspecies.J. Clin. Microbio?. 27, 132-138.
KULfPER, E.J.; VAN ALPHEN, L.; LEENDERS, E. Y ZANEN, H.C. (1989c). Typing of
Aeromonasstrainsby DNA restrictionendonucleaseanalysisandpolyacrylamidegel
electropitoresisof celí envelopes.¿ Clin. Microbiol. 27, 1280-1285.
KUIJPER, EJ.; VAN ALPEN, L.; PEETERS, MF. Y BRENNER Di. (1990). Human serum
antibodyresponseLo the presenceofAeromonasspp. in the intestinaltract. £ Clin.
Alicrobiol. 28, 584-590.
233

0
U
“4
“44
rL
e
44.
U,
RíA¡lOGRAFÍA
LACIIICA, R.V. Y ZINK, DL. (1984). Plasmid-associatedcelí surface charge and
hydrophobicityof Yersiniaenterocolirica. Infecr. Immun.44. 540-543.
LALLIER, R.; MITrAL, K.R.; LEBLANC. D.; LALONDE, G. Y OLIVIER, G. (1981). Rapid
methedsfor the differentiationof virulent and nonvirulentAeromonas hvdrophila
strains,p. 119-123.InternationalSymposiumon Fish Biologics.
LAWRENCE, R.C.; FRYER, TE. Y REITER, B. (1967).Theproduction¿mdcharactenzationof
lipasefrom a micrococusandpseudomonad.¿Gen.Microbio). 48, 401.
LAWSON. MA.; BURKE, Y. Y CHANG, B.J. (1985). Invasionof HEp-2celis by fecal isolates
ofAeromono,s hydrophila.Infect. Jmrnun.47, 680-683.
LE CHEVALLIER, M.W.; SEIDLER, R.J. Y EVANS, ‘EM. (1980). Enumeration and
characterizationof standar plate count bacteriain chlorinatedand rawwatersupplies.
AppL Environ. Microbiol. 40, 922-930.LE CHEVALLIER, M.W.; EVANS, T.M.; SEIDLER,RJ.;DAILY, O.P.;MEARELL, B.R.; ROLLINS,
D.M. Y JOSEPH,5W. (1982).Aeromonassobria in chiorinated drinkingwatcr
supplies.Microbiol. Ecol.8, 325-333.
LEN, P.P. (1987).Mesopitilie fish spoilageof marinefish: hay trout(Arripis trieno).breani
(Acanihopagnisburcheri> and millet (Alcírichetiaforsreri). Food Technol. Ausr.39,
277-282.
LEUNG, K.Y. Y STEVENSON,R.M.W. (1988).Characteristicsanddistribution ofextracellular
proteasesfrom Aeromonashydrophikt1 Gen.Microbio). 134, 151-160.
LEVEfl, P.N.YDANIEL,R.R. (1981). Adhesionofvitalesandaeromonadsto isolatedrabbit
brush borders.1 Gen. Microbio). 125, 167-172.
LINDAI-IL, M.; PARIS. A.; WADSTROM, T. Y HJERTEN. 5. (1981). A new tes¡ baseden
“salting out” te measurerelative surfacehydrophobicityof bacterialcelís.Biochim.
Biophys.Acta. 677, 471-476.
LJUNGH, A. (1987).Aeromonas-toxins¿md other virulencefactor. Experienria. 43,347-472.
LJUNGH, A.; WRETLLNG, B. Y MOLLBY. R. (1981). Separationand characterizationof
enterotoxinandtwo haemolysinsfrom Aeromonashydrophila.ActaParh. MicrobioL
Scand.89, 387-397.LJUNGH, A.; ENROTH, P. Y WADSTROM, T. (1982).Cytotonicenterotoxinfrom Aeromonas
hydrophila.Toxicon.20, 787-794.
LULWY. NG.; SANTER, LV.; WIECZOREK. M.; BLOOGETE, 1K.; iONES, 5W. Y CHERONIS,
J.C. (1993). Purificationandcharacterizationof a novel zinc-proteinasefrom cultures
of Aemunwnas hvdrophila.1. BioL Cheor 268. 9071-9078
234

BÍBUOGRAFÍA
LUGTENBERG, B Y VAN ALPHEN. L. (1983).Moleculararchitectureand functioningof the
outcr membraneof Escherichia coli and otiterGrani negative bacteria.Biochim.
Biophys.Acta.737, 51-115.
MACKENZJE. K.A. Y SKERMAN, V.B.D. (1982). Microbialspoilagein unpasteurizedliquid
whole egg.FoodTechnoLAun. 34. 524-528.MMEED, K.N.; EGAN, AY. Y MACRAE, f.C. (1989a).Enterotoxigenic aeromonadsen retail
lanib meatandoffal. 1 Appl. Bacteriol. 67, 165-170.
MAJEED, K.N.; EGAN, A.F. YMACRAL, f.c. (1989b).Incidenceof aeromenadsin samples
from an abattoirprocessinglanibs.J. AppL Bacteriol. 67, 597-604.
MAJEED, K.N.; EGAN, A.F. Y MACRAE, It. (1990).Productionof exotoxinsby Aeromonas
spp. at 50C.J. AppL Bacteriol.69, 332-337.
MAJEED, K.N. Y MACRAE. J.c. (1991). Experimentalevidencefor toxin productionbyAeromonashydrophilaandAeromoiaassobriain ameatextractal low tcmperatums.
Ini. 1 FoodMicrobiol. 12, 181-188.MAJEED,K.N. Y MACRAL, I.C. (1994). Cytotoxicandhaemagglutinatingactivitiesof motile
Aeromonasspecies.¿ Mecí. MicrobioL 40, 188-193.
MANO, S. Y GARCíA DEFERNANDO, G.D. (1995).Comunicaciónpersonal.
MARSHALL, ¡C.D. (1976). Interfacesin Microbiol Ecology.Canibridge,Mass: Harvard
UniversityPress.
MARTINEZ-MURCIA, A.!.; RSTEVE. C.; GARAY, E. Y COLLINS. M.D. (1992). Aeromoiaas
allosaccharophilasp. nov., a new mesophilicmemberof te genusAeromonas.
FEMS.Microbiot Len. 70, 199-205.
MASSAD, G.; ARCENEAUX. J.E.L. Y BYERS,B.R. (1991). Acquisition of iron from host
seurcesby mesophilicAeromonasspecies.J. Gen.Microbio). 137, 237-241.MASSAD, G.; ARCENEAUX, J.E.L.Y BYERS, B.R. (1994). Diversity of siderophoregenes
encoding boisynthesisof 2,3-dihydroxybenzoicacidin Aeromonasspp.Biometals.
7, 227-236.
MATEOS, D.; ANGUITA, J.; NAVARRO, G. Y PANIAGUA, C. (1993). Influence of growth
temperature entic productionofextracellularvirulencefactors andpathogenicityofenvironmentaland humanstrainsof Aeromonas hydrophila.1 AppL Bacteriol.74,
111-118.
MATTILA, L.; PELTOLA, H.; SUTONEN. A.; KYRONSEPPA, H.;SIMULA, 1. Y KATAYA, M.
(1993). Short-term treatmentof traveler’sdiarrheawith norfloxacin:a double-blind,
placebo-controlledstudyduring two scasons.Clin. !nfecr. Dis. 17. 779-782.
235

$4
9.
u
e
0
u
U
e
BIBLIOGRAFÍA
MCCOY, RU. Y PILO-tER, K.S. (1974). Peptonebeefextractglycogen agar. a selective¿md
differentialAeromonasmedium.¿ Fish. Res.BoarcíCon. 31, 1153-1155.
MCKELLAR, R.C. (1986). A rapid colorimetrie assayfor tite extracellularUpaseof
PseudomonasfluorescensB52 using ¡3-naphthylcaprylate.1 Dairy Res.57,
117-127.
MCKELLAR, R.C. (1989). Regulation and control of synhesys. In Enzymesof
psychrotrophsin raw food. cd. Robin C. Mackellar. PP. 153-172. Florida: CRC
Press,Inc.
MERCER.N.S.G.;BERRE,D.M.; BORNEMISZA, AJ. Y ThOMAS P. (1987). Medicalleeches as
seurcesof infection.Brit. Mal ¿ 294, 937.
MERINO, 5.; CAMPRUBI, 5. Y TOMAS, J.M. (1993). Detectionof Aeromonas hydrophilain
food wiih an enzime-linked immuneserbeníassay.¿ AppL Bacterio).74, 149-154.
MERINO. 5.; ALVAREZ, O.; HERNANDEZ-ALLES, 5. Y lUMAS, 131. (1994). Effectof grewthtemperature encomplement-mediatedkilling of mesophilicAeronionasspp.serotype
0:34. FEMSMicrobiol. Len. 118, 163-166.
MERINO, 5.; RUBIRES,X. Y TOMAS, J.M. (1995).The presenceof capsularpolysaccharide
in mesophilic Aeromonas hydrophila serotypes0:11 and 0:34. Abstr.
AAIR-PL920630.Mb Meetingof”Physiologyof FoodPoinsoningMicroorganisms“ -
MILLERSHIP, S.E.; CURNOW, S.R. Y CHA’fl?OPADHYAY, B. (1983).Faecalcarriagerate of
Aeromonashydrophila. ¿ Clin. Microbiol. 36, 920-923.
MILLERSHIP, S.L Y CHATrOPADHYAY, B. (1984>.Methods fortite isolationofAeromonas
hydrophlla¿mdPlesiomonasshigelloidesfrom faeces.1 Hyg.92, 145-152.
MILLERSHIP. S.E. Y WANT, S.V. (1989). Typing of Aeromonasspeciesby protein
fingerprinting:comparisonof radiolabe]ling¿mdsilve stainingfor labelling proteins.
¿ Med Microbio). 29, 29-32.
MILLERSHIP, S.L. Y WANT, 5V. (1993).Characterizationof strainsofAeromonasspp.by
phenotypeand whole-cellproticin fmgerprint.¿Mecí.Microbio,!. 39, 107-113.
MITI7AI, lCR.; LALONDE, G.; LEBLANC, D.; OLIVIER, O. Y LALLIER, It. (1980).Aeromonas
hydrophilain rainbowtrout: relation betweenvirulenceandsurfacecharacteristics.
Can. ¿ Microbiol. 26, 1501-1503.
MONTENEGRO,MA.; BI’ITER-SUERMANN. O.; TIMMIS, 3.K.; AGUERO,ME.; CABELLO, F.C.;
SANYAL, S.C. Y TIMMIS, RiN. (1985>. TraT genesequences,serurnresitanceand
pathogenici¡yrelated factorsin clinical isolatesof Escherichia coli and other
gram-negativebacteria.¿ Gen.Microbio!. 131, 1511-1521.
236

BIBLIOGRAFíA
MORGAN, D.R.; JOHNSON,P.C.; DUPONT, H.L.; SAYI’ERWHITE, T.K. Y WOOD, L.V. (1985).
Lack of correlationbetweenknown virulencepropertiesof Aeromonashydrophila
and enteropatogenicityfor humans.Infect Jinmun.50. 62-65.
MORGAN, DR. Y WOOD. L.V. (1985).IsAeromonassp. a foodbornepatbogen?Review of
tite clinical data..¿ Food Safery.9, 59-72.
MORITA, K.; WATANABE, N.; KURATA, S.Y KANAMORI, M. (1994).Beta-~lactamresistance
of motileAeronionasisolatesfrom clinical vid environmental seurces.Anrimicrob.
AgentsChemoter.38, 353-355.
MORSE, J.W.Y 1-LINO, D.W. (1984). Bacteria isolatedfrom Iymph nodesof California
slaughterswine.Am.¿ Ver. Res.45, 1648-1649.
MOSSEL, D.A.A.; BONATS-VAN LAARHOVEN, T.Mfl; LIGTEMBERG-MERCUS, A.M.TH. Y
WERDLER, M.E.B. (1983).Quality assuranceof selectiveculture mediafor bacteria,mouldsandyeast:aix attemptatstandarizationat tbeinternationallevel. .1. Bacterio).
54, 313-327,
MOYER, NS’. (1987).Clinical signifcanceofAero¡nonasspeciesisolated frompatientswith
diarrhea.£ Clin. MicrobioL 25, 2044-2048.
MOYER, N.P.; GEISS, H.K.; MARINESCO, M.; RIGBY, A.; ROBINSON. J. Y ALTWEGG, M.
(1991).Media ant! methodsfor isolation of aeromonadsfrom fecal specimens.A
multilaboratorystudy.F.xperientia.47, 409-412.MOLLA. R Y MILLERSHIP, 5. (1993).Typing ofAerotnonasspp. by numerica]analysisof
inmunoblotted SDS-PAGEgeis. ¿ Mecí. Microbiol. 39, 325-333.
MONN, C.B.; ISHIGURO. E.L.; KAY, W.W. YTRrJST,Ti. (1982). Roleofsurfacecomponentsin serumresistanceof virulent Aeromonas salmonícida. ¡nfect.¡mmun.36,1069-1075.
MUNOZ, It; FERNANDEZ-BACA, y.; PELAEZ,T.; SANCHEZ, R.; RODRIGUEZ-CREIXEMS,M. Y
BOOZA, E. (1994).Aeromonasperitonitis.Clin. Infect. Dix. 18, 32-37.MURRAY, R.GuL.; DOOLEY, J.S.G.;WHIPPEY, P.W. T TRUST, Ti. (1988). Structureof aix 8
layeren a patitogeniestrainofAeromonashydrophi)a.¿ Bacteriot 170, 2625-2630.
MYERS, B.R.; MARSHALL, R.T.; EDMONDSON, J.E. Y STRINGLR, W.C. (1982).Isolation of
pectinolyticAeromonashydrophilavid Versiniaeiarerocoliácafrom vacuum-packaged
pork. ¿ FoodProtect.45, 33-37.NAMDARI, H. Y BOTTONL, EJ. (1988). Correlationof the suicide phenomenonin
Aeromonasspecieswith virulenceand enteropathogenicity.¿ Clin Mit robiol 26,
26 15-2619.
237

U,
0.-
“4
u,
mu
a
mu
e
e’
“4
mu
BIBUOCRAFIA
NAMOARI, ¡1. Y BOYtONE, E.!. (1989). Suicide phenomenonin mesophilic aeromonadsasa
basisfor speciesidentiftcation.J. Clin, Microbio). 27, 788-789.
NAMDARI, ti. Y BOITONE, EJ. (1990a).Microbielegic ¿mdclinical evidence supportingtite
role of Aeromonascaviaeasa pedriaticentericpathogen.¿ Clin. Microbio). 28,
837-840.
NAMDARI, H. Y BOlTONE, EJ. (1990b). Cytotoxinand enterotoxinproductienas factors
delineating enteropathogenicityof Acromonascaviae.¿ Clin. Microbio). 28,
1796-1798.
NAMOARI, 11. Y BOTrONE, E.>. (1991). Aeromonascaviae:Ecologie adaptaúonin tite
intestinal tractof infantsceupledteadherenceandenterotoxinproductionasfactorsinenteropathogenicity.Experientia.47,434-436.
NkEVES, M.S; NONES, MP. Y MIIL1{OMEN, AM. (1994). Aeromenasspecies exhibit
aggregativeaditerenceto HEp-2cels.¿Clin. Microbio). 32, 1130-1131.NEWTON,J.A.~. Y KENNEDY, C.A. (1993).Woundinfection due Aeronwnas sobria.Clin.
¡nfect. Dis. 17, 1082-1083.
NIETO, Ti’. Y ELLIS, A.E. (1986). Characterizationof extracellular metallo- and
serine-proteasesof Aeromonashydrophi)astrain B51. ¿ Gen. Microbiol. 132,
1975-1979.
MSHIKAWA, Y. Y 1(15111.T. (1988).Isolation vid characterizationof mobile Aeronwnas
from humanfoodand environmentalspecimens.EpidemiotInfect.161, 213-223.NISHIKAWA. Y.; KJMURA, T. Y 1(15111, T. (1991). Mannose-resistantadhesionof motile
Aeromonaste 1NT407celisand tite differencesamong isolatesfrem humans,feod
¿mdwater.Epidemiol. ¡nfecr. 107, 17 1-179.
NISI-IIKAWA, Y.; OGAWASARA, 3. Y KIMURA, Y (1993).Heat¿mdacidsensitivityof motile
Aeromcnas:acomparisonwith otiter food poisoningbacteria,¡nr. J FoodMicrobio),
18, 271-278.
NISHIKAWA. Y.; BASE. A.; OGAWASARA, 1.; SCOTLAND, SM.; SMITh, H.R. Y ¡(¡MORA, 7,
(1994). Adhesion te andinvasion of human coloncarcinomaCaco-2celis by
Aeromonasstrains.¿ Mcd? Microbio).40, 55-61.
NOTLRMANS, 5.; HA VELAR, , JANSEN, W; KOZAKI. 5. Y GUINEE, 1’. (1986).Productionof “AsaoToxin” by Acromonasstrains isolatedfrom feces¿mddrinlcing water.1.
C)in. Microbio). 23, 1140-1142.
(JODEN, ID.; MILLAR, 1(3.; WATT, A.). Y WOOD, L. (1994). A comparison of three
identification kits for tihe confirmationof Arromonasspecies.Len.Appl. Microbio?,
18. 97-99.
235

BIBLIOGRAFíA
OKITSU, T.; ASAr. Y.; YASUDA. T. Y TAKIZAWA, K. (1985). Studiesof Aeromonasspp.
isolatedfrom patientswith sporadicdiarritoca.Kansenshogaku Zasshi.59, 977-983.
OKREND, AIG.; ROSE, BE. Y BENNETT, B. (1987). Incidence and toxigenicity of
Aeromonasspeciesin retail poultry beefandpork.¿ FoodFrotecr. 50, 509-513.
ORDONEZ. JA. (1979). Randomnumbersampling rnethodfor estimationof lactic acid
bacteria.J. AppL Bacterio). 46, 351-353.
(¡yERMAN. ‘EL.; KESSLER, LE. Y SEABOLT, iP. (1985).Comparisonof API 20E, API
Rapid E, and API Rapid NFT for identification of membersof the family
Vibrionaceae.J. Clin. Microbiol. 22.778-781.
PALOMBO, 5 A. (1986). Isrefrigerationenoughte restrain foodborne pathogens?¿ Food
Prorect. 49, 1003-1009.
PALUMBO, SA. (1988).The growthofAeromonashydrophilaK144 un greund pork al50C.
Ini. ¿ FoodMicrobiol. 7, 41-48.PALUMBO, 5 A.; MAXIMO, E.; WILLIANS, A.C.; BUCHANAN, RL. Y ThAYLR, D.W. (1985a).
Starchampicillin agarfor ihe quantitative detectionof Aeromonahydrophila.Appl.
Environ.. Microbio). 50, 1027-1030.
PALUMBO, 5 A.; MORGAN, D.R. Y BUCHANAN, R.L. (1985b). Influence of temperature,NaCí , ant! pH on tite growth of Aeromonashydrophila.¿ Food Protect. 50,
1417-1421.
PALUMBO, 8 A.; JENKINS, R.K.;BUCHANAN, RL. Y THAYER, D.W. (1986).DeterminationofirradiationD-valuesforAeromonashydrophila. ¿ FoodProtect.49, 189-191.
PALUMBO, 5 A.; WILLIANS, A.C.; BUCHANAN, Rl. Y PHILLIPS, 1.G. (1987). Thermal
resistanceofAeromonashydrophila.¿ FoodProtect. 50,761-764.
PALUMBO, 5 A. Y BUCHANAN, R.L. (1988). Factors affeccinggrowth or survival of
Aeronwnashydrophila in foods.1 FoodSafety.9, 37-51.
PALUMBO, 5 A.; BENCIVENGO, M.M.; DEL CORRAL, E.; WILLIANS, KC. Y BUCHANAN, Rl.
(1989). Characterizationof tihe Aeromonas hydrophilagroup isolatedfrom retailfoodsof animal origin.¿ Clin. Microbio!. 27, 854-859.
PALUMBO, 5 A.; WIILLIANS, A.C.; BUCHANAN, R.L. Y PHILLIPS, 1.0. (1991). Model fortite
aerobicgrowthofAeromonashydrophila K144.¿ FeodProzecí.54, 260-265.
PALUMBO, 5 A.; WILLIANS, A.C.; BUCHANAN, RL. Y PHILLIPS, 1.0. (1992). Model fer the
anaerebícgrowthof AeromonashydrophilaK144. ¿ Food Protect.55, 429-435.
PARRAS, F.: ~AZ, M.D.; REINA, 3.; MORENO, S.; GUERRERO,0. Y BOUZA, E. (1993).
Meningitis due Aeromonasspecies:casercport aud review. Clin. Infiel. fis. 17.
1058-1060.
239

A,
mu.
mu
“4
1~
mu
BÍBLIOGRAFÍA
PARKER, NO. Y MtJNN, CE. (1984). Increasedccii surfacehydrophobicityassociatedwith
possessionof an additional surfaceprotein by Aeromonassalmonicida.FEMS
Microbio,!. Leer. 21, 233-237.
PASQUALE, V.; BALODA, SR.; DUMONTET, 5. Y KROVACEK, K. (1994). Anoutbreakof
Aeromonas hydrophilainfectien in turtíes (Pseudemis seripta).Appl. Envirom.
Microbio). 60, 1678-1680.PAULA, S.J.; DUFFEY, PS.; AEBOrF, S.L.; KOKKA, R.P.; OSHIRO, LS.; JANDA, f.M.;
SHIMADA, T. Y SAKAZAKI, R. (1988). Surfacepropertiesof autoagglutinating
mesophilicaeromonads.Infect.Jmmun.56,2658-2665,
PAYNE, S.M. (1988). Iron ant! virulencein the family Enrerobacreriaceae.Cnt. Rey.
Microbiol. 16, 81-111.
PAZZAGLIA, 0.; ‘NINOTO. 1. Y JENNINGS,0. (1994). Oral challengewith Aeromonasinprotein-maineurishedmicej.Diarrhoeal Dix.. Res.12, 108-112.
PERERS,L.; ANDAKER, L. EDEBO, L.; STENDAHL, O. Y TAGESSON,C. (1977). Association
of sorneenterobacteriawith tite intestinalmucosaofmeusein telation to tiheir partitionin aquceuspolymertwo-phasesystems.ActaPathot Microbiol. Scand.Sect.13 85,
308-316..
PHIPPS,B.M. Y ¡CAY, W.W. (1986).Immunoglobulinbindingby the surface array virulence
proteinof Aeromonassalmonicida.Abstr. K225. Annual Meetingof tite American
Societyfor Microbiology.PLEiTO, 0.; LETENNEOR, 3. Y SCHWOERER, D.(1993). Infection osseusesaAeromonas
hydrophila- aproposof 2 casereporis.ActaOrthop. Belg. 59, 390-393.PITARXNGSI, C.; EC¡-IEVARRIA, It; WHITMIRE, It.; TIRAPAT, C.; FORMAL, 5.; DAMMITN, 0.1.
Y TINGTALAPONG, M, (1982). Enterepathogenicityof Aeromonas hydrophila¿md
Plesiomonas shigelloides:Prevalenceamongindividualswith ¿md without diarrhoea
in Thailand.Infecr. ¡mmun.35, 666-673.
POFFE,R. Y OP DE BEECK, E.(1991). Enumerationof Aeromonas hydrophi)afromdomestic waste water treatmentplants surface waters.J. Appl.. liacteriol. 71.
366-370.
POIRIER, T.; LAURENS, E.; yIAND, J.V.; POUJOL, E. Y LORRE, G. (1993). Nosocomial
Aeromonashydrophila pneumoniacomplicatingtoxic coma.Ania. Fr. Aneseh
Reanim.12, 72-74.pQ¡)QJZp Ni. (1984).Acrornonas.En BergeVs Manualof Systemaricflacteriology. cd.
Krieg N.R. Y Holt, J.G. Pp. 545-548. Vol.1. Baltimore: WiIliams and Wilkins.
24<

BÍBLÍOGRAFíA
POPOFF, M.;COYNAULT, C.; KIREDJIAN, M. Y LEMELIN. M. (1981). Polynucleotide
sequencerelatednessamong motileAeromonasspecies..Curr. Microbio). 5, 109-114.
POPOFF, M. Y yERON, M. (1976). A taxonomic study of the Aeromonas
hydrophila-Aeromonaspuiactatagroup.¿ Gen.Microbiol. 94. 11-22.
POTOMSKI, J.; BURKE, y.; WATSON, 1. Y GRACEY, M. (1987a).Purification of citotoxie
enterotoxin of Aeromonassobria by use of monoclonal antibodies.J. Med.
Microbio). 23, 171-177.
POTOMSKI, J.; BURKE. y.; ROBINSON, 1.; FUMAROLA, D. Y MIRAGLIOTA, 0. (1987b).
Aeromonascitotonicenterotoxincross reactivewith choleratoxin. 1 MaL Microbio).
23, 179-186.PRIEST, F.G. (1983). Enzyme synthesys: regulation and process of secretionby
microorganisms.In Microbial Enzymesand Biotechnology.ed. Fogarty,W.M.London:Applied Science.
PUMAROLA, A. (1987a). Factoresdeterminantesde la acciónpatógena:capacidadlesional.
Modelos de infección.. En Microbiologíay Parasitología Médica. PP.. 178-188.
Barcelona: Salvat.POMAROLA, A. (1987b)..Factoresdeterminantesde la acción patógena:penetración,
multiplicacióneinvasión. En Microbiología yParasitologíaMédica.pp. 171-177.Barcelona: Salvat.
PUMAROLA, A. (1987c). Infección.Poderpatógenoy virulencia.Factores determinantesdela acciónpatógena:colonización.En Microbiología y Parasftología Médica.pp.
161-170.Barcelona:Salvat.
QADRI, S.M.H.; GORDON, LS’..; WENDL, R.D. Y WILLIAMS. R.C.. (1976). Meningitis due teAeromonashydrophila.1 Clin. Microbio). 3, 102-104.
QUINN. D..M.; ATKINSON, H..M.; BRETAG, A.M.; TESTER. M.; TRUST, TJ..; ‘NONO, C.Y.F. Y
FLOWER, R.L.P.. (1994). Carbohydrate-reactivepore-forming outer membrane
proteinsofAeromonashydrophila.¡nfect. Immun.62, 4054-4058.
RAHIM, Z.; SANYAL, SC.; AZIZ, K.M.S.; HUQ, Mi. Y CHOWDHURY, A.A. (1984). Isolation
of enterotoxigenic, hemofilic,¿mdantibioúc-resistantAerornonashydrophila.strains
from infectedfish in Bangladesh.AppL Environ.Microbio). 48, 865-867.
REINA, .¡. (1992). Situación taxonómicaactual de las especiesincluidas en el grupo
Aeromonasspp. aisladasen sereshumanos.Rey.Esp. Microbio). Clin. Octubre.
37 1-377.
RILAMON. A.F.M.S. Y WILLOUGHBY, ¡MT. (1980). Dysentery-likesyndrome associated
with ,4eronwnashvdrophilo. Brit.. Med. J. 281, 976.
241

w
0
mu.
mu
E
BIBUOGRAFL4
RIBAS, E.; ARAUJO, It.; ERIAS, 3.; HUGUET, J.M.; RIBAS, FR. Y LUCENA, F. (1991)
Comparisonof differentmediaforthe identification ¿mdquantificationofAerotnonas
specicsin water.Antonie van Leeuwehvek.59, 225-228.
RIPPEY, SR. Y CABELLI, y.>. (1979). Membranefilter procedurefor enumerationof
Aeromonashydrophila in freshwaters.AppL Environ. Microbio). 38, 108-113.
RJPPEY,SR. Y CA.BELLI, VJ. (1985). Growth charactcristicsof Acromonashydrophila in
limnetic waterof varying trophicstate.Arch. Microbiol. 104, 311-319.
RIVERO, O.; ANGUITA, 1.; PANIAGUA, C. Y NAHARRO, 0. (1990). Molecular cloning and
characterizationof an extracellularpreteasegenefrom Aeromonashydrophi)a.J.
Bacteriol. 172, 3905-3908..
ROBERTS,hM.; PARENTI,D.M. Y ALBERT, M.B. (1987).Acromonas hydrophila-asseciated
colitis in a malehomosexual.Arch. miera. Mcd. 147, 1502-1503.
ROGLRS, 5.; MATHIAS. 0>4.; WILLIAMS, R.M.. Y PARK, R.W.A. (1990). Tite attachmentof
feod ¿mdclinical Acromonasisolateste tite suifaceof formalised humanintestinal
epithelium.Abstr.mt. Symp.Aeromonas¿mdPlesiemonas.
ROGOL, M.; SECHTER,1.; GRINBERG,L. Y GERICHTER,C.D. (1979). Pril-xylosa-ampicillinagar,a newselective mediumfor tite isolationof Acromonashydrophila. 1 Mcd
Microbiol. 12, 229-231.
ROLAN!?), F.P.. (1977).SaIt-starch xyloselysine deoxycholateagar.A singlemedium fortiteisolationof sodiumandnon-sodiumdependententericgrani-negativebacilli. Mcd
Microbio). ¡mmunol. 163, 241-249.ROSE, J.M.; HOUSTON, C.W.; COPPE1&HAVER, Dli.; DIXON. J.D. Y KUBOSKY, A. (1989a).
Purification¿mdchemicalcharacterizationof a cholera toxin-cross-reactive cytolyúc
enteretoxin produc&d bya hum¿m isolateofAcromonashydrophila. lnfect. Immun.
57, 1165-1169.
ROSE,J.M.; HOUSTON, CM’. Y KUROSKY, A. (198%). Bioactivity ant! immunological
characterizationof a cholera toxin-cross-reactivecytolytic enterotoxinfrom
Acromonashydrophila. ¡nfect ¡mimen.57, 1170-1176.
ROSENBERG,M. (1981).Bacteria]aditerencete polystyrene:a replica metitodfor screening
tihe bacteñalhydrophobicity.Appl. Environ. Microbio). 42, 375-377.ROSENBERG, M.; GUTMCK, D. Y ROSENBERG,E. (1980). Adherenceof bacteriato
hydrocarbons:a simple metitodfor measuringceil-surfacehydropitobicity. FEMS
Microbio!. Leí!. 9, 29-33.
242

BÍBUOGRAFÍA
ROSENBERG,M..; PERRY, A.; BAYER, E,A,; GUTNICK. DL.: ROSENBERG. E. Y OFEK. 1.
(1981).Adherenceof Acinerobacterca)coacedcusRAG-1 te humanepititelial celis
¿md te hexadecane.!nfect lmmun.33, 29-33.
ROUF, M.A. Y RIGNEY, M.M. (1971). Growth temperaturesand temperaturecharacteristics
ofAeromonas.AppL Microbiol. 22, 503-506.
RUIMY, It.; BREITTMAYER, V.; ELBAZE, P.Q.; LAFAY, B..; BOUSSEMART,O.; GAUTHLER, M.
Y CHRiSTEN, R. (1994). Phylogeneticvialysis ant! assessmentof ihe generaVibrio,
Phorobacterium, Aeromonas¿mdPlesiomonasdeducedfrom small-subunitrRNA
sequences.¡ni. ¿ FoodMicrobio). 44, 416-426.
SACK, R.B.; LM4ATA, C. Y KAY, B.A. (1987). EpidemiologicalstudiesofAeromonas-reíateddiarrhealdiseases.Experientia.43, 364-365.
SAKAI, D.K. (1985). Lossof virulencein a protease-deficientmutantof Aeromonas
salmonicéda.Iuxfen. Immun..48, 146-152.
SAN JOAQUíN, V.H. Y PICKETT, DA. (1988).Aeromonas-associatedgastroenteritisin
children.Pediatr. Infecz.Dis. 1 7, 53-57.
SANTOS,1.; TORANZO, A.E.; DOPAZO, C.P.; NIETO, T.P. Y BARJA, JI. (1987). Relationships
amongvirulencefor fish, enterotoxigenieity,vid phenotypiccharacteristicsof motile
Aeromonas.Aquaculture. 67, 29-39..
SANTOS. 1.; TORANZO, A.L..; BARiA. 1.L.; METO, LP. Y VILLA, T.G. (1988). Virulence
propertiesand toxin productionof Aeromonasstrains isolatedfrom fish. Infecí.
Inmun.56, 3285-3293.
SANTOS. ¿LA.; SANZ, J.; LOPEZ. T.M..; GARCíA. M.C. Y OTERO. A. (1991). AislamientoycaracterizacióndeAeromonasmóvilesobtenidasde distintosalimentes.Res. XII
CongresoNacionalde MicrobiologíaEspañol.
SANTOS, 1.; LOPEZ, T.M.; GARCíA, ML.; GARCíA, M..C. Y OTERO, A. (1994). Minimum
water activity for tite growthof Aeromonas hydroph¡la as affectcdby strain,temperature¿mdhumectant.Uit Appl. Microbiol. 19, 76-78.
SATO, M.; ARITE, M.; HONDA, T.. Y MIWATANI, T. (1989). Characterizationof a pilusproducedby Aeromonashydrophila. FEMSMicrobio,!. Lea. 59, 325-330.
SCHLFFER, 1.; KONIG, W.; BRAUN, V.. Y GOLBEL, W. (1988). Comparison of fourhaemolysin-producingorganisms(Escherichiacoli, Serradamarcescens,Aeromonas
hydrophilaasidListeria monocytogenes)for releaseof inflanxmatorymediatorsfrom
varicuscells.£ Clin.. Microbiol. 26, 544-551.
SCHIEMANN, DA. (1979).Synthesisof a selective agarmediumfor Yersiniaenrerocolirica.
Can. £ Microbio!. 25, 1298-1304.
243

mu
mu
Mr
rL
U,
0
BÍBI.IOGRAFíA
SCHUBERT,R.1I.W. (1967).Dasvockommender acromonadenin oberirdischenGewasse¡n..
Arch. Hyg.150, 688-708.SCHUBERT, R.H.W. (1977). Veberden Nachweisvon PlesionwnasshigelloidesHabsand
Schubert,1962. und ein elektivmedium,deninositel-brillant grum-gallesolz-agar.E.
Rodenwald:.Arch. 4. 97-103.
SCHUBERT, R.H.W.(1991).Aeromonadsand titeir significanceas potential pathogensin
water. JAppl. Bacreriol.76 (Supplj, 131S-135S.
SCHUBERT, R..H.W.. Y HEGAZI, M. (1988). Aeroínonas eucrenophilaspeciesnava
Aeromonascaviaea later¿mdilegitimatesynonymofAeromonaspunctata.Zenrralbl.
Bakreriolo.Ryg. Abr. 1 origin Reihe A.268, 34-39.
SCHUBERT, R..H.W. Y HEGAZI, M. Y WAHLING, W. (1990a).Aeromonasenreropelogenes
speciesnova.HygieneMedizin.15, 471-472.SCHUBERT.R.H.’N.. Y HEGAZI, M. Y WA¿HLING, W. (1990!».Aeromonasichrhiosmiaspecies
nova. HygieneMedizin.15, 477-479.
SCHULTZ, A.]. Y MCCARDELL, B.A.. (1988). DNA homology ant! immunological
cross-reactivitybetweenAeromonashydrophilacytotonic toxin asidcholeratoxin. ¿
Clin. Microbio). 26, 57-61.SEIDLER, RJ.; ALLEN, D..A.; LOCKMAN, U..; COLWELL, R..R..; JOSEPH, SM’. Y DAILY, O.P..
(1980)..Isolation, enumerationant! characterizationof Aeromonasfrom polluted
watersencounteredun diving operations.AppL Environ. Microbio).39, 1010-1018.
SHERAGO. M (1936). Aix epizootic septicemiaof young guinea pigs causedby
Pseudomonas caviaenov. sp.1 Bacreriol. 31, 83.
SHIEH, H.S. (1987). Toxicity of Aeromonashydrophila proteasete Atlantic salmon.
Microbios Len. 36, 17-21.
SHIEH, H..S. (1988). Lethal toxicityof Aeromonassobria proteaseto Atlantic salmon.
MicrobiosLen.37. 65-68.
SHOrtS,E..B.JR.Y RIMLER, R. (1973). Mediumfor theisolationof Aeromonashydrophila.
AppL Microbio). 26, 550-553.
SUREAD, P.;DONOVAN, ‘EJ. Y LEE,J.V. (1981). A surveyof tite incidenceofAeromonaslii
human faeces.Soc. Gen.Microbio). Q, 8, 184.
SIMARO. RE..; ZEE,1. Y L’HEIJREUX, L.. (1984)..Microbial growth in carcassesant! boxed
beefduring storage.£ FoodProtect.47, 773-777.
SINGIL, DV.. Y SA=4YAL,SC. (1992a).Euiwrotoxicity of clirxical ¿mdenvironmentalisolates
of Acromonasspp.£ Med. Microbio!. 36, 269-272.
244

BIBLIOGRAFÍA
SINGH, D.V. Y SANYAL, SC. (1992!». Haemolysin and enterotexinproduction by
Aeromonascaviae isolatedfrom diarrhoealpatients, fishandenvironmental..J.
Diarrhoeal Dix. Res. 10. 16-20.SINGLE, D.V.; DUBEY, RS.Y SANYAL, S.C. (1993). Adherenceof haemagglutinatingand
non-haemagglutinatingclinical and environmentaliselatesof Aeromonas.J.
Diarrhoea)Dis. Res.11, 157-160
SINGLE, D.V. Y SANYAL, SC. (1993). Haemagglutinatingactivity serumsensivity and
enterotoxigenicityof Aeromonasspp.1 Med. Microbio).38, 49-53.SLADE. P.J..;FALAIl, M.A. Y AL-GRADY, A.M.R. (1986). Isolationof Aeromonashydrophila
from botiledwaters and domesticwatersuppliesin SaudiArabia. 1 Food Protect.
49. 471-476.
SLEYTR, u..B. (1978).Regulararraysof macromoleculeson bacteñalcelí walls: structure,cbemistry,assembly,asidfunction. ¡nL Rey.Cyrol. 53, 1-64.
SLEYRT, U..B. Y MESSNER,P. (1983).Crystallinesurface layers enbacteria.Annu. Rey.
Microbio). 37, 311-319.SLOTNICK, i.J. (1970). Aeromonasspeciesisolates.Ann. N.Y AcadSc).. 174, 503-510.
SMIT, it; GRANO, D..A.; GLAESER, R.M. Y AGABIAN, N.. (1981). Periodicsurfacearray in
Cau)obacter crescentus:Fine structureasid chemical analysis.1 Bacteriol. 146,
1135-1150.SMITH. H. (1990). Pathogenicityand Ihe microbe in vivo.. ¿ Gen. Microbio). 136,
377-383.SMITH, R.; NICKELSON, R.; MARTIN, R. Y FINNE, 0.. (1984). Bacteriology of indole
productionin shrimp homogenatesheld at different temperatures.¿Food Protect.
47. 86 1-864.SMYTH, C.J.; JONSSON, P.; OLSSON, E..; SODERLIND, O..; ROSENGREN, 1.; HIERTEN, 5. Y
WADSTROM, T. (1978).Differencesin hydrophobicsurfacecharacteristicsofporcine
enteropathogenicEscherichia col) with or without K88 antigenas revealedbyhydrophobicinteractionchromatography..Infect. ¡mmun.22, 462-472..
SNO’NER, D.P.; RUEF, C.; ¡CURIrZA. A.P. Y EDBERG, S.C. (1989). Aeromonashydrophila
infection associatedwith the useof tite medicinal leeches.J. C)in. Microbiol. 27,
1421-1422.
STLLMA. G.N.JR. (1988).Virulence factorsassociatedwitb pathogenicityof Aeromonas
iselated.¿ EoodSafery.9, 1-9.
STET,MA. G.NJR. (1989).Acromonashydrophila.In FoodborneBacterial Pathogens..ed.
DoYle. MP. PP. 1-20..New York: Marce! Dekker..
245

U,
Mt,
u
e’
u
—
0
a
a
BIBLIOGRAFlA
STELMA, G.NJR..: JONLESON. CH. Y SPAtJLD[NG, P. (1986). Evidence lcr the direct
involvementof f3-hemelysinin Aeromonas hydrophilaenteropathogenicity.Curr.
Microbio). 14. 71-77.
STEPHENSON,J..R.; MILLERSHIP, SE. Y TABAQCHALI, S.. (1987). Typingof Aeromonas
speciesby polyacrylamide gelelectrophoresisof radiolabelledcelí proteins.¿ Med.
Microbio), 208, 980-984.
STERN, Nl.; ORAZEK, ES. Y JOSEPH,S.W. (1987).. Low incidenceof Aerontonassp. in
livestok feces.J. Foad Prorecr..50, 66-69.
STEWART, G..A.; BONDELL, C.S.Y BURME, y. (1986). Partialcharacterizationof a soluble
haemaggiutininfrom human diarrhoealisolatesof Aeromonas..¿ Med? Microbio)..
21, 319-324.
S’rE’NART. M. Y BEVERIDGE, Ti. (1980).. Structureof tibe regularsurface layer of
Sporosarcinaureae.¿ Bacterio).. 142,302-309.STEWART, M. Y MURRAY, R.G.E. (1982). Structureof tihe regularsurfacelayer of
AquaspirillumserpensMW5.1 Bacteriol.. 150, 348-357.
SUGITA, 1-1..; NAKAMURA, T..; TANAKA, 1<. Y DEGUCHI, Y. (1994). Identificationof
Aeromonasspecies isolatedfrom freshwaterfish with tbenijeroplatehybridization
Method.AppL Environ. Microbio).60, 3036-3038.
SUHREN, 6. (1989).Producermicroorganisms.lix Enzymesofpsychrotrophsin rawfood..
ed. RobinC. Mackellar.PP.3-34.Florida: CRC Press,Inc.
SWEET, SS’..; MACFAR¿LANE, 1W. Y SAMARANAYAKE, L.P.. (1987). Determinationof tibeceil surfacehydwphobicity of oralbacteriausinga modified hydrocarbonaditerencemethod.FEMSMicrobio). Len. 4S, 159-163.
TELFORD, J.R.; LEARY, JA.; TUNSTAD, L.M.G.; BYERS, BR. Y RAYMOND, K.N.. (1994).Amonabactin:characterizationof a seriesof siderophoresfrom Aeromonas
hydrophila.¿ Am.Chem.Soc.116, 4499-4500.
THOMAS, L.y..; GROSS, Rl.; CHEASTY, 1.. Y ROWE, B. (1990). Extended serogrouping
sehemefor motilemesophilicAerornonas.J. Clin. Microbiol. 208, 980-984.
THOMAS, S..R. Y TRUST, T.J.. (1995). Tyrosinasephosphorylationof tite tetragonal para
crystailine array of Aeromonas hydrophi)a: molecularcloning ant! high-level
expressionof theS-layerproteingene.¿Mo).Biol. 245,568-581.
IHORNE, K..3.I.; OLIVER, R.C.. Y GLAUERT.. AM. (1976). Synthesis andturnoverof the
regulary arrangedsurface protein of Acinetobactersp. relative tic tihe ether
componentsof tihe ceil envelope.¿Bacteriol.. 127, 440-450.
24fi

BIBLIOGRAFíA
THUNE, R.L.; GRAHAM, T.E..; RIDDLE, L.M. Y AMBORSKI, RL. (1982). Extracellular
proteasesfrom Aeromonashydrophila: Partialpuriticationand effectsen age-Ochannel catfish.Trans. Am. Fish. Soc.3, 404-408.
TODD, L.S.; HAROY, J.C.; STRINGER, MF? Y BARTHOLOMEW, A.. (1989). Toxin production
by strainsof Aeroínona hydrophilagrownin laboratorymedia¿md prawn puree.Inc
¿ FoodMicrobiol. 9, 145-156.
TORANZO, A.E.; BARIA, J.L.; COLWELL, R.R.. Y HETRICK, EM. (1983a)..Characterizationof
plasmidsin bacterialfxsh pathogens.Infect. ¡mimen.39, 184-192.
TORANZO, A..E.; BARJA, J.L..; COLWELL, R.R.; HETRICK, F.M.. Y CROSA. ¿[LE. (1983b)..Haemagglutinatinghaemolyticasidrelatedvibrios isolatedfrom stripedbasson the
atlanticcoast.FEMSMicrobiol Len. 18, 257-262.TORANZO, A.E.; SANTOS. Y..; NIETO, T..P. Y BORJA, J.L. (1986). Evaluation of different
assay systems foridentificationof environmentalAeromonasstrains.AppL Environ.
MicrobioL 51, 652-656.
TOULE, J. Y MIJRPHY, 0. (1978). A study of bacteriacontaminatingrefrigeratedcooked
chicken; their spoilagepotential and possible origin.1 Hyg. Cambridge.81,
161-169.
TRUST, Ti.; HOWARD, P..S.; CHAMBERLAIN, 1.B.; ISHIGURO, E.E.Y BUCICLLY, J.T. (1980).
Additional surfaceprotein in autoaggregatingstrainsof atypical Aeromonas
salmonicida.FEMSMicrobio)Len. 9, 35-38.
TRUST, T.J..; KAY, W.W. Y ISHIGURO, E.E. (1983>. Celí surfacehydrophobicity asidmacrophageassociationofAeromonassalmonicida.Ciar. Microbio). 9, 315-318.
TSUBOI, A.; TSUKAGOSLEI, N. Y UDAKA, 5.. (1982). Reassemblyiii vitro of hexagonal
surfacearraysin a protein-producing bacterium,Baci)lusbrevis 47. J. Bacíeriol..
151, 1485-1497.
TURNBIJLL. P..C.B.; LEE, J..V.; MILIOTIS, M.D.; VAN DER WALLL, 5.; KOORNHOF, H.J.;
JEFFREY,L. Y BRYANT, ‘EN. (1984).Enterotoxinproductionin relationto taxonomicgreupingand sourceof isolation of Aeromonasspecies.J. Clin. Microbiol. 19,
175-180..
VADIVLLY, J.; PT.JTHOCHEARY, S.D.; PHIPPS, M. Y CEfEE, Y.W. (1995).. Possiblevirulence
factors involved in bacteraemiacausedby Aeromonas hydrophila.¿ Med.
Microbio,!. 42, 171-174.
VAN DER GOOT, F.G.; AUSIO, .1.; WONG, K.R.; PATFUS, E. Y BUCKLEY, J.T.. (1993).
Dimerizationstabilizesthe pre-formingtexin aerolysinin solution..¿ Rio!? Chem.
268. 18272-18279.
24?

“4-.
pv
“4-
“4
“4-
“4
BÍBLIOGRAFÍA
VAN DER GOOT, EG.; PAYI’US, E; PARKER, M. Y BUCKLEY, J.T. (1994a).Tite cytolytic
toxin aerolysin: from thesolublefon to tite transrnembranechannel.Toxicology.
87, 19-28.
VAN DER GOOT, F.G.; HARCHE, K..R..; PARKER, M. Y BUCKLEY, IT. (1994b). The
C-terminalpeptide producedupon proteolytic activationof the cytolytic toxin
aerolysinis not involvedin channelformation. 1 Rial. Chem.269, 30496-30501.
VAN DER KOOU, 0. (1988).Properties ofaeromonadsvid theiroccurrencevid hygienic
significancein drinking water.ZenrralbL Bacrerio!?Hyg.. B 187, 1-17.
VAN DER ¡COQId, D. y HIJNEN, W.A.M. (1988).Nutritional versatilityvid growth kineticsof
Aeromonashydrophilastrainisolatedftom drinkingwater.AppL. Enviran.MicrobioL
54, 2842-2851..
VAN OSS.C.J. (1978).Phagocytosisasa surface pitenomenon.Annu. Rey. Microbio).32,19-39.
VARNAM, A..LE. Y EVANS, M.G. (1991).Aeronionas.In FoodbornePathogens:An illustrated
tefl. Pp. 185-200.London:Wolfe publishingLtd.
VILLUENDAS, M.C.; ESTEBAN. A.; LOPEZ-GOMEZ, C.; FORTONO, B.; SERRANO, M. Y
MOLES, B. (1991).Aeromonasen infecciones extraintestinales.Rey..Esp. Microbiol.
Clin. 9, 4446-4450..VON GRAEVENITZ, A. (1985). AcromonasasidP)esiomonas.Itt Manual of C)inical
Microbio)ogy. ed. Lennette,E.H; Balows, A.; Hausler,W..J.JR.y Shadomy,H.J.PP..278-281.. WashingtonDC: AmericanSocietyfor Microbiology.
VONGRAEVENITZ. A. (1987).Research enAeromonasvid Plesiomonas.Experientia..43,
348-349.
VON GRAEVENITZ, A. Y MENSCH, A.H. (1968). Tite genusAeromonas in itumanbacteriology,reportof 30 casesvid review of tite literature.N. EngL J.. Med.278,
245-249.
VON GRAEVENflZ, A. YZINTERLEOFER,L. (1970).ThedetectionofAeromonashydrophi)a
in stoolsspecimens.HealthLab. Sci.7, 124-126.
VON GRAEVENITZ, A. Y BUCHER,C. (1983).Evaluationofdifferential and selective media
for iselatienof Aeromonasvid Plesiomonasspp. from ituman feces..1 Clin.
Microbiol. 17, 16-21.
WADSTROM, T. Y LJUNGLE, A.. (1991). Aeromonasy Plesiomonasas food- and
watcrborne-pathogens.mt. ¿ Food Microbio!. 12, 303-312.
24$

BíBLÍOGRAFIA
WAKABAYASIII, ti.; KANAI, K.; ¡-ISU, T.C. Y EGUSA. s. (1981). Pathcgenic activitiesof
Aeromonashydrophilabiovar. hydrophi)a (Chester)Popoff and Veten, 1976 te
fishes.Fish Parhol.. 15, 3 19-325.
WALKER, S.J. Y STRINGER, M.F. (1987). Growtit of Listeria monocytogenesand
Aeromonashydrophila at chilí temperatures..TechnicalMemerandumN0 462.
CampdenFoodPreservationResearchAssociation,Englasid.
WANDERSMAN, C. (1989).Secretion,precessing,¿md activationof bacterialextracellular
proteases.Mo!? Microbio). 3, 1825-1831.
WARBORTON, D..W.; MCCORMICK, J.K. Y BOWEN, B.. (1994). Survival and recoveryofAeromonashydrophila in water: developmentof methodolegyfor testingbottled
waterin Canada.Can.. J. Microbiol. 40, 145-148.
WATSON, I.M.; ROBINSON, J.O.; BURKL, y. y GRACEY, M. (1985). Invasivenessof
Acromonasspp.unrelatentebiotype,virulencefactors, amiclinical features.J. Clin.
Microbio). 22, 48-51.
WILCOX, M..H.; COOK, A.M.; ELEY, A. Y SPENCLR, R.C.(1992). Aeromonasspp. as a
potencialcause ofdiarritoeain children..J. Clin. PajIzo). 45, 959-963.WILCOX, M.LE.; COOK, A.M.; GEARY, 1. Y ELEY, A. (1994).Toxin production, adherenceasid
proteinexpressionby clinical Aeromonasspp. isolatesin broth asid human pooled
ileostomyfluid. Epidemio!?¡nfecr. 113, 235-245.
WILKEN, G.B. Y A¿PPLETON,C.C. (1993). Bacteriologicalinvestigationof the ocunenceasid
antibiotic sensitivitiesto tibe gut-floraofthepotentialsouthemMrican medicalleech
Asiañcobdelía buntonensis(Hirudin)dae).1 Hosp..Jnfect.23, 223-228.
WILLIAMS, L.A. Y LAROCHE, P.A. (1985). Temporal occurrenceof Vibrio speciesand
Aeromonashydrophila in estuarine sediments..Appl. Environ, Microbio). 50,
1490-1495..
WILLOOGHBY, ¡MT.; RALEMAN, A..F.M.S.. Y GREGORY,M.M. (1989). Chronic colitisaher
Aeromonasinfection. 0w. 30, 686-690..
WOLELEGEMUTLE, K.; PIERCE,RL. Y KIRKBRIDGE, C.A. (1972).Bevineabortiensasseciated
with Aeromonashydrophila.J. Am. Ver.. Med? Assoc.160, 1001-1002.
WOLFF, R.L..; WISEMAN, S.L. Y KITCHENS, S.C.(1980)..Aeromonas hydrophilabacteremiain ambulatoryinimunocempromisedhosts.Am.1 Med.68. 238-242..
YAMADA, H..; TSUKAGOSHI, N.Y tJDAKP., s. (1981).Morphologicalalterationsof ccli wallconcomitantwith proteinreleasein a protein-producingbacterium,Bacillus brevis47.
J Bacteriol. 148, 322-332.
24>

“4*
Mt
u
“4
e
u
u
e
$4
BÍBLiOGRAU4
YAMAMOTO, K.; NAGAMINE, 5.; KAMIYA. T.; MURAKAMI, R.; KIKUCHI, J. Y SATo, Ii.
(1994).Identificationte tite genospecies levelofAerotnonasstrainsisolatedfrom raw
meat and carp.and evaluationof titeir biological activities. f EcodJ-fyg. Soc. Jap.
35, 187-194.
YESIM OZBAS, Z. Y AYKUT AYTAC, S. (1994).Effectof chloride en grewtit and survivalof
Aeromonashydrophila¿md Yersinia enteroco)itica in water.Chem. Mikrobiol.
Technol. Lebensmitrel.16. 146-150,
ZYWNO, 5.; BAILEY, C.; ARCENEAUX, 1. Y BYERS, R. (1989). Siderophore distributionin
isolatesof tite Aeromonasspecies.Abstr. D91.. Annual Meetingof tite American
Societyfor Microbiology.






![Clinical and Therapeutic Implications of Aeromonas Bacteremia: … · 2016-12-28 · Aeromonas bacteremia are malignancy and hepatobiliary dis-eases [5]. Aeromonas spp. tend to produce](https://img.pdfslide.net/doc/110x75/5ec79db8c2bd727c0b32cc58/clinical-and-therapeutic-implications-of-aeromonas-bacteremia-2016-12-28-aeromonas.jpg)