Embed Size (px)
Citation preview

vol. 137, No. 3, 1986 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
June 30. 1986 Pages 1153-1158
ALTERATIONS IN DEOXYNUCLEOSIDE TRIPHOSPBATE METABOLISM IN DNA DAMAGED CELLS: IDENTIFICATION AND CONSEQUENCES OF POLY(ADP-RIBOSE)
POLYMERASE DEPENDENT AND INDEPENDENT PROCESSES
Shishir K. Das and Nathan A. Berger
Hematology/Oncology Division, Departments of Medicine and Biochemistry &se West& Reserve University School of Medicine and
R. Livingston Ireland Cancer Center Cleveland, Ohio 44106
Received May 12, 1986
SUMMARY: Treatment of L1210 cells with increasing concentrations of MNNG produces heterogeneous perturbations of cellular deoxynucleoside triphos- phate pools, with the magnitude and direction of the shift depending on the deoxynucleotide and on the concentration and time of exposure of the DNA damaging agent. 5 uM MNNG stimulated an increase in dATP, dCTP and dTTP but dGTP pools remained constant. These increases were not affected by 3-aminobenzamide, indicating that the pool size increases were produced by poly(ADP-ribose) polymerase independent reactions. 30 nM MNNG caused a time dependent decrease in dATP, dGTP, dTTP and dCTP. The dGTP pool was most drastically affected, becoming totally depleted within 3 hours. The fall in all 4 dNTP pools was substantially prevented by 3-aminobenzamide, suggesting that the decrease in dNTPs following DNA damage is mediated by a poly(ADP-ribose) polymerase dependent reaction. Severe depression of dGTP pools consequent to NAD and ATP depletion may provide a metabolic pathway for rapidly stopping DNA synthesis as a consequence of DNA damage and the activation of poly(ADP-ribose) polymerase. @ 1986 Academic Press, Inc.
Deoxynucleostde triphosphates (dNTPs) are the ultimate precursors for
DNA replication (1). Their synthesis is finely regulated and their relative
cellular pool sizes have important consequences on the rate and fidelity of
DNA replication and repair (2). One of us has previously shown in Chinese
hamster V79 cells, that several DNA damaging agents can perturb cellular
dNTP levels, with increases in dATP and dTTP pools occurring following
treatment with MNNG or W-irradiation (3.4). In other studies, with L1210
leukemia cells and normal human lymphocytes, we have shown that DNA damaging
agents can activate the chromosomal enzyme poly(ADP-ribose) polymerase to
consume cellular NAD pools and consequently deplete ATP (5). In contrast to
the increase in dNTP pools noted above, this depletion of ATP is expected to
ABBREVIATIONS: ADP-ribose, Adenosine diphosphoribose; 3-AB, 3-aminobenza- mide; dNTP, deoxynucleoside triphosphate; MNNG, N-methyl-N'nitro-N-nitroso- guanidine.

Vol. 137, No. 3, 1986 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
diminish and possibly deplete cellular dNTP pools. The apparently divergent
studies alluded to above were conducted in separate laboratories, with
different cell types using different concentrations of DNA damaging agents.
The present study was therefore conducted to investigate the alteration in
dNTP pool sizes induced by a wide concentration range of DNA damaging agents
and to examine the role of poly(ADP-ribose) polymerase in these processes
using a single cell type, the L1210 lymphocytic leukemia cell.
Poly(ADP-ribose) synthesis can be involved in altering nucleotide pools
by several mechanisms. First, the enzyme uses NAD as a substrate, leading
to pool shifts of this dinucleotide by direct consumption (5). Second, it
is possible that poly(ADP-ribosylation) could produce allosteric modifica-
tions of nucleotide metabolizing enzymes, resulting in alteration of their
synthesis or degradation rate. Since the enzyme can be inhibited by nicoti- ,
namide analogs such as 3-aminobenzamide, this agent can be used to determine
whether poly(ADP-ribose) polymerase is involved in a par 'icular process (6).
4 While 3-aminobenzamide has other effects, we have previo , ly shown that it
is highly effective as an inhibitor of poly(ADP-ribose) pplymerase and that
it can block the NAD and ATP depletion that occurs when the enzyme is activa-
ted by high levels of DNA damage (6,7,8). Thus, these studies were carried
out in the presence and absence of 3-aminobenzamide to determine whether the
DNA damage induced alterations of dNTP pool sizes were dependent or
independent of the activation of poly(ADP-ribose) polymerase.
MATERIALS AND METHODS: Cell culture and drug treatment - L1210 cells were grown at 37°C
in a-modified Eagle's medium buffered with 25 mM Iiepes, pH 7.2, and supplemented with 10% heat-inactivated fetal calf serum, 50 units/ml penicillin, and 50 rig/ml streptomycin. Logarithmically growing L1210 cells were diluted to 1 x 105/m1 and treated with MNNG that was freshly dissolved in dimethylsulfoxide and diluted with 0.9% NaCl.
Measurement of dNTP levels - For the measurement of dNTPs, L1210 cells were pelleted by centrifugation at 2OOOg for ten minutes. The medium was aspirated carefully and the pellets were extracted with 0.5N HC104. These suspensions were cleared by centrifugation and the extracts were neutralized with 5N KOH. After centrifuging again to remove the KC104 precipitate, the clear supernatants were collected and used for dNTP assays. dNTP levels were quantified by using the E. coli DNA polymerase I assay with either poly (dA-dT) or poly(dG-dC) as primer-template as previously described (3,4). Multiple volumes of extracts were used to establish the linearity of the assay for the calculation of the amounts of dNTPs.
RESULTS: Figure 1 shows the effects of treating exponentially growing L1210
cells with different concentrations of M&NG for 12 hours. Upon treatment
with 5 ClM MNNG, there was a substantial increase in the levels of MTP,
dTTP, and dCTP, whereas the dGTP levels were not significantly altered. At
10 uM MNNG, the response was heterogeneous; dATP and dTTP pools became
elevated above control levels, the dCTP pool fell slightly below control
1154

Vol. 137, No. 3, 1986
dATP 500 -
I---- 0 IO 20 30
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
1 dTTP
L.'.' 0 IO 20 30
t
dGTP 500 t
dCTP
dependence of MNNG-induced alterations in FIG. 1: Concentration L1210 cell dNTP poolsizes. L1210 cells in the log phase of growth were treated with the indica- ted concentration of MNNG and incubated at 37OC in the presence (0) or absence (0) of 2mn 3AB. After 12 hours incubation, the cells were collected and HC104 extracts prepared for dNTP analysis as indicated in Methods. The dGTP level in cells treated with 30 ti MNNG was beyond the limit of detection.
(u Ml MNNG
level and the dGTP pool fell significantly. When the cells were treated
with 30 VM MNNG, there were drastic decreases in all four dNTPs; the
greatest effect occurred in the dGTP pool, which was totally depleted by
this treatment.
When these incubations were carried out in the presence of 2 mM 3-AR,
there was no interference with the MNNG induced increases in dATP, dTTP or
dCTP, suggesting that these processes are independent of poly(ADP-ribose)
polymerase activation. In contrast, the decrease of the dNTPs that occurred
at 30 uM MNNG was substantially retarded by 3-AB. Most strikingly, the
total depletion of dGTP pools was completely prevented by 3-AB.
To determine the rapidity of these pool changes, we examined the time
courses of their alterations upon treatment of the cells with 5 or 30 m
MNNG. Figure 2 shows that after treatment with 5 pM MNNG, the increases
in dATP, dTTP and dCTP are already apparent within 3 hours. The dTTP pool
remains at a relatively constantly elevated level during the next nine hours
whereas the dCTP pool continues to undergo a small increase, and the dATP
pool continues to undergo a more marked increase. At 5 nM MNNG, the dGTP
pool remained essentially constant during the entire incubation period.
None of the responses to 5 pM MNNG were inhibited by 3-AB, confirming
that this increase in dNTP pools is independent of poly(ADP-ribose)
polymerase activation.
1155
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS Vol. 137, No. 3, 1986
dATP
500 t t 5uM MNNG \ fA 00 5uMMNNG
-K “0 t3AB
. 50 . 30uMMNNG
+ ?A9
1 30 uM MNNG
I- O 6 12
dTTP
t 5~ M MNNG
1 30 UM MNNG
Lt.1 0 6 12
dGTP
I 5uMMNNG
I
0 6 I2
dCTP
t 5uMMNNG
E 301.1 M MNNG
L- 0 6 12
HOURS
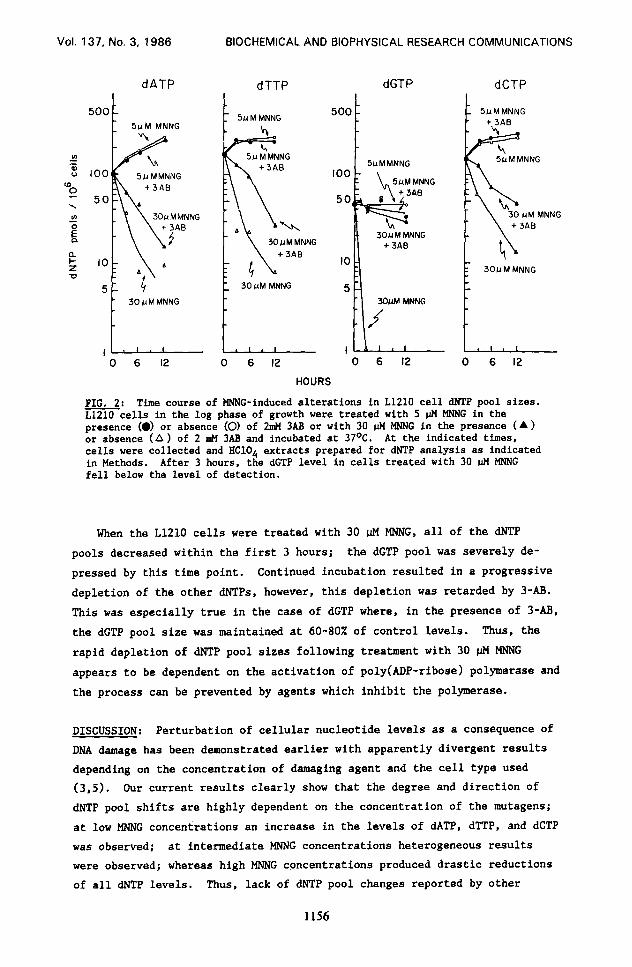
FIG. 2: Time course of MNNG-induced alterations in L1210 call dNTP pool sizes. L1210 cells in the log phase of growth were treated with 5 pM MNNG in the presence (0) or absence (0) of 2mM 3AB or with 30 IJM MNNG in the presence (A) or absence (A) of 2 mM 3AB and incubated at 37OC. At the indicated times, cells were collected and HC104 extracts prepared for dNTP analysis as indicated in Methods. After 3 hours, the dGTP level in cells treated with 30 fl MNNG fell below the level of detection.
When the LX10 cells were treated with 30 nM MNNG, all of the dNTP
pools decreased within the first 3 hours; the dGTP pool was severely de-
pressed by this time point. Continued incubation resulted in a progressive
depletion of the other dNTPs, however, this depletion was retarded by 3-AB.
This was especially true in the case of dGTP where, in the presence of 3-m,
the dGTP pool size was maintained at 60-807; of control levels. Thus, the
rapid depletion of dNTP pool sizes following treatment with 30 $4 MNNG
appears to be dependent on the activation of poly(ADP-ribose) polymerase and
the process can be prevented by agents which inhibit the polymerase.
DISCUSSION: Perturbation of cellular nucleotide levels as a consequence of
DNA damage has been demonstrated earlier with apparently divergent results
depending on the concentration of damaging agent and the cell type used
(3,5). Our current results clearly show that the degree and direction of
dNTP pool shifts are highly dependent on the concentration of the mutagens;
at low MNNG concentrations an increase in the levels of MTP. dTTP, and dCTP
was observed; at intermediate MNNG concentrations heterogeneous results
were observed; whereas high MNNG concentrations produced drastic reductions
of all dNTP levels. Thus, lack of dNTP pool changes reported by other
1156

Vol. 137, No. 3, 1986 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
workers could be due to the dose of mutagen tested (9,lO); alternatively,
the dNTP pool response to mutagen treatment may be a,variable phenomenon
influenced by the cell type used in the study. However, preliminary studies
in our lab indicate that qualitatively similar responses occur at high and
low doses of DNA damaging agents in Chinese hamster, V79 cells.
Poly(ADP-ribose) is synthesized in eukaryotic nuclei by the chromatin-
bound enzyme poly(ADP-ribose) polymerase (11). The enzyme cleaves NAD at
the N-glycosylic bond between the nicotinamide and ribose rings and trans-
fers the ADP-ribose moieties to covalent linkages with various acceptor mole-
cules. The enzyme adds successive residues by ribose to ribose 0-glycosidic
linkages to form homopolymers of repeating ADP-ribose units. Poly(ADP-
ribose) polymerase activity is stimulated by treating cells with a variety
of DNA damaging agents; the common lesion which stimulates enzyme activity
is DNA strand breaks. We have previously suggested that when cells sustain
high levels of DNA damage and DNA strand breaks, the consequent activation
of poly(ADP-ribose)polymerase initiates a suicide response or final common
pathway leading to cell death (8). Thus, activation of poly(ADP-ribose)
polymerase by DNA strand breaks can lead to consumption of its substrate NAD
and subsequent depletion of ATP (5,8). This process of energy depletion
causes drastic alterations in intermediary metabolism (12) and in DNA, RNA
and protein synthesis, leading to rapid cell death (5,8).
We have previously shown that MNNG, in the concentration range of
30 IJM will rapidly deplete the NAD and ATP levels of L1210 cells (5,12).
Our present results indicate that this loss of NAD and ATP has generalized
effects on cellular metabolites resulting in a decrease in all four dNTPs.
The role of poly(ADP-ribose) polymerase in this process is demonstrated by
the fact that 3AB substantially prevents depletion of these dNTP pools. It
is interesting to note that in earlier studies of poly(ADP-ribose)
polymerase, it was suggested that polymer synthesis was involved in stopping
DNA synthesis in response to DNA damage (13). However, in subsequent
studies, no specific structural or regulatory effect has been identified by
which the enzyme or polymer could suppress DNA synthesis. Our present
studies demonstrate that DNA damage can cause the rapid depression of dGTP
pools in a poly(ADP-ribose) dependent reaction. Since replicative DNA
synthesis is dependent on the presence of all four dNTPs (I), the rapid
depression of dGTP consequent to NAD and ATP depletion may provide a
metabolic pathway for acutely halting DNA synthesis as a consequence of DNA
damage and the activation of poly(ADP-ribose) polymerase. Following
treatment of cells with 10 nM MNNG, the dTTP and dATP pools were clearly
elevated, whereas the dGTP pool was significantly reduced. These elevated
dATP and dTTP pools might contribute to an increased error rate in DNA
1157

Vol. 137, No. 3, 1986 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
synthesis (3,4), however, the lowering of the dGTP pool through a
poly(ADP-ribose) polymerase dependent process may provide the cells with a
mechanism to turn off DNA synthesis and avoid this mutagenic potential.
ACKNOWLEDGEMENTS: This research was supported by National Institutes of Health grants CA-35983, GM32647 and T32RI.07147. Some of the studies were performed in facilities supported by the Sohio Foundation.
REFERENCES
1. Kornberg, A. (1980) DNA Replication. San Francisco, Freeman. 2. Thelander, L. and Reichard, P. (1979) Ann. Rev. Biochem. 48, 133-158. 3. Das, S.K., Benditt, E.P., and Loeb, L.A. (1983) Biochem. Biophys. Res.
comm., 114, 458-464. 4. Das, S.K., and Loeb, L.A. (1984) Mutat. Res., 131, 97-100. 5. Sims, J.L., Berger, S.J., and Berger, N.A. (1983) Biochemistry, 22,
5188-5194. 6. Sims, J.L., Sikorski, G.W., Catino, D.M., Berger, S.J. and Berger, N.A.
(1982) Biochemistry, 21, 1813-1821. 7. Milam, K.M. and Cleaver, J.E. (1984) Science 223, 589-591. 8. Berger, N.A. (1985) Radiat. Res., 101, 4-15. 9. Hyodo, M., Ito, N., and Suzuki, K. (1984) Biochem. Biophys. Res. Comm.,
122, 1160-1165. 10. Trukla, D., Otsuka, II. and Plunkett, W. (1985) Biochem. Biophys. Res.
comnl., 133, 1064-1071. 11. Iiayaishi, O., and Ueda, K. (1977) Ann. Rev. Biochem., 46, 95-116. 12. Berger, S.J., Sudar, D.C. and Berger, N.A. (1986) Biochem. Biophys.
Res. Comm., 134, 227-232. 13. Burzio, L. and Koide S.S. (1970) Biochem. Biophys. Res. Comm. 40.
1013-1020.
1158