Embed Size (px)
Citation preview

Proc. Natl. Acad. Sci. USAVol. 80, pp. 7556-7560, December 1983Genetics
ampC (8-lactamase hyperproduction in Escherichia coli: Naturalampicillin resistance generated by horizontal chromosomalDNA transfer from Shigella
(DNA sequence analysis/antibiotic-resistance evolution)
OLOF OLSSON, SVEN BERGSTROM, FREDERIK P. LINDBERG, AND STAFFAN NORMARKDepartment of Microbiology, University of UmeA, S-901 87 Ume&, Sweden
Communicated by Robert L. Sinsheimer, July 25, 1983
ABSTRACT Six ampicillin-resistant clinical isolates of Ewch-erichia coli that hyperproduced the chromosomal ampC (-lacta-mase were studied. By DNA sequence analysis, we found that fiveof them were identical over an entire 449-base-pair sequence andcarried a novel strong ampC promoter [Olsson, O., Bergstrom, S.& Normark, S. (1982) EMBOJ. 1, 1411-1416]. Except for one basepair this sequence was identical to that of a low 13-lactamase-pro-ducing clinical isolate of Shigella sonnei. Spontaneous one-stepmutants of S. sonnei that overproduced the ampC 1-lactamase by45-fold were characterized and found to be mutated at the singlebase that distinguishes S. sonnei from the five E. coli hyperproduc-ers. The most likely explanation for this result is that chromosomalDNA was transferred in vivo from ShigeUa to E. coli across thespecies barrier.
Escherichia coli contains a chromosomal gene, ampC, that codesfor the production of a class C f3-lactamase (1). By using ampCDNA as a probe in DNA-DNA hybridization experiments it hasbeen possible to detect DNA homologous to this gene, not onlyin clinical isolates of E. coli but also in other enterobacterialspecies (2). An especially strong signal was obtained when hy-bridizing to DNA from Shigella sonnei and Shigella flexneri.
Expression from the E. coli K-12 ampC gene is very low, andampC wild-type strains are resistant to ampicillin at about 1 jug/ml. The low production of ampC 13-lactamase in E. coli K-12is due to a relatively weak promoter and to the presence of atranscriptional attenuator in the ampC leader (3). Transcrip-tional termination from the ampC attenuator decreases with in-creasing growth rate, since the expression from ampC is undergrowth rate control (4).
By selection for ampicillin resistance in batch cultures of E.coli K-12, different classes of ampC f3-lactamase hyperproduc-ing mutants have been isolated. Up-promoter mutations, in eachcase increasing the fitness to the consensus sequence of E. colipromoters (5, 6), occurred at an incidence of between 10-10 and10-11 and increased ampC expression 7- to 21-fold (3, 7). Otherone-step genetic events known to increase E. coli K-12 ampCexpression are (i) integration of insertion sequence IS2(II) intothe -10 hexamer of the ampC promoter, which increasedexpression 20-fold (incidence 101 to 10-1") (8); (ii) ampC at-tenuator mutations, which increased expression 4-fold duringfast growth (incidence 10-8 to 10-9) (3), and (iii) gene ampli-fication, which may increase expression 2- to 20-fold (incidence10-5 to 10-10) (9).
It has not been possible to increase expression from the E.coli K-12 ampC gene more than 21-fold by a single genetic event.To achieve an even higher level of expression from the E. coli
ampC gene, two or more mutations have to be combined.In view of these findings in E. coli K-12, we found it intrigu-
ing to elucidate how naturally occurring ampC hyperproduc-tion may have evolved. Earlier we characterized six clinical E.coli isolates that produced 24- to 48-fold-increased levels of theampC enzyme (10). By determining the sequence of the ampCregulatory regions and the surrounding DNA sequences of these,/3lactam-resistant E. coli isolates and comparing them to wild-type E. coli and S. sonnei, we conclude that in five of the sixE. coli isolates /3-lactamase hyperproduction must have evolvedby a single mutation from a Shigella ampC control region. Thisgenerates a novel, strong ampC promoter that differs from, butoverlaps, the ampC promoter defined in E. coli K-12.
MATERIALS AND METHODSBacteria. The E. coli K-12 strain LA5 and the clinical isolates
of E. coli and S. sonnei used in this study, the relative amountsof 1-lactamase produced by these strains in rich media, andtheir LD50 value on rich ampicillin media are listed in Table 1.The E. coli strains C11, C13, C14, C15, C16, and C17 wereobtained by screening 109 f3-lactam resistant isolates from pa-tients with urinary tract infections as previously described (10).E. coli strains C115 and C123 and S. sonnei strain OS10 werealso isolated from clinical specimens and were considered aswild-type ampC strains on the basis of their low (3-lactamaseproduction. E. coli K-12 strain JM103 was used as a host forM13 phage growth (13).
Media. In most experiments bacteria were grown in minimalmedium E (14) or in LB medium of Bertani (15). The carbonsources used are indicated in the legend to Fig. 1. Phage M13was propagated on strain JM103 in YT medium (16).DNA Techniques and Cloning of the ampC Gene. Isolation
of chromosomal DNA was as described (2). Plasmid pBEU28carrying kanamycin resistance and a temperature-sensitive rep-lication control (17) was kindly provided by B.-E. Uhlin (thislaboratory). Plasmid pNU78 has been described (2). Chromo-somal DNAs from the E. coli strains C123, C115, Cli, C13,C14, C15, and C17 and from the S. sonnei strains OS106, OS107,OS112, and OS113 (Table 1) were isolated, cleaved with EcoRI,ligated with EcoRI-digested plasmid pBEU28, and used totransform the P-lactamase-negative strain SNO3. Transformantscarrying the ampC gene were selected on ampicillin plates. TheampC gene of strain C16 was inserted into plasmid pNU78 asdescribed (18). Hybrid plasmids obtained were characterizedby physical mapping.The ampC region from S. sonnei strain OS10 was isolated
from a A phage library as described (19) and inserted into plas-mid pNU78.
Abbreviation: bp, base pair(s).
7556
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertise-ment" in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Proc. Natl. Acad. Sci. USA 80 (1983) 7557
Table 1. Bacterial strainsRelative Ampicillin
,-lactamase resistance,t SourceStrain production* pg/ml Comments or ref.
E. coli K-12LA5 1 2.3 Laboratory strain 11
E. coliCli 51 100 Clinical isolate 10C13 25 38 Clinical isolate 10C14 46 160 Clinical isolate 10C15 50 170 Clinical isolate 10C16 44 90 Clinical isolate 10C17 50 100 Clinical isolate 10C115 1.4 2.1 Clinical isolate This
paperC123 0.9 2.6 Clinical isolate This
paperS. sonneiOslo 1.1 1.5 Clinical isolate 2OS106 4.5 14 Mutant from OS10 2OS107 7.5 21 Mutant from OS10 2OS112 41 90 Mutant from OS10 20S113 37 80 Mutant from OS10 2
*Determined by rocket immunoelectrophoresis according to Laurell (12).The cells were grown in rich medium (LB).
t Ampicillin concentration permitting 50% of single cells to form col-onies.
Plasmids carrying the ampC gene on an EcoRI fragment (Fig.2) were prepared as described (18). Restriction enzyme frag-ments were prepared from polyacrylamide gels (20), and a 931-base-pair (bp) Sal I/Xho I restriction fragment was inserted intothe M13 cloning vector Gori 1 (21). Plaques of phage carryingthe ampC gene were identified by plaque hybridization (22).The M13 vector mp8 carrying the E. coli K-12 Sal I/Xho I frag-ment of the ampC control region (Fig. 2) inserted in either ori-entation was labeled by synthesizing the M13 complementarystrand from a probe primer in the presence of [a-32P]dCTP (23)and were used directly as probes for plaque hybrdization. PhageDNA was prepared from positive plaques, and its sequence wasdetermined by the dideoxy method (24). Both strands of theDNA were analyzed, using two oligodeoxynucleotides as strand-specific primers.
Other Methods. Ampicillin resistance was determined forsingle cells as described (10). To quantitate ampC 3-lactamaseexpression, the immunoelectrophoresis method of Laurell wasused (12).
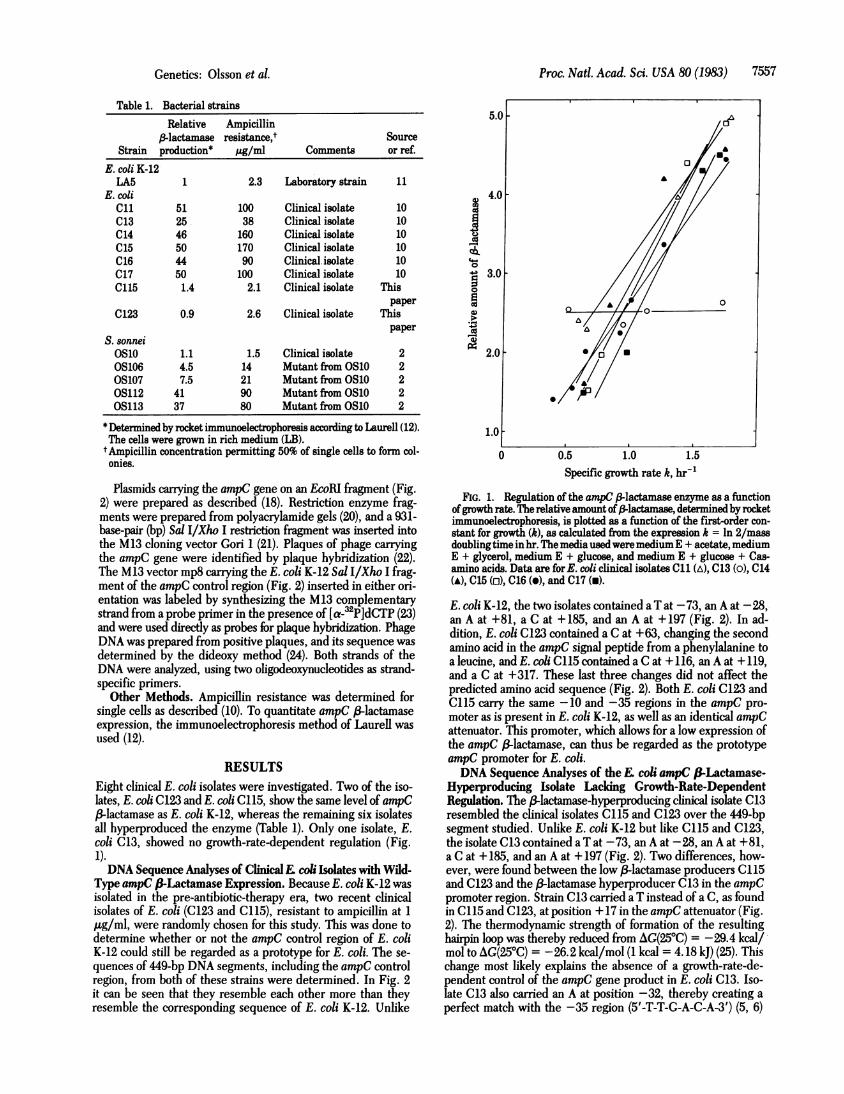
RESULTSEight clinical E. coli isolates were investigated. Two of the iso-lates, E. coli C123 and E. coli C115, show the same level of ampC3-lactamase as E. coli K-12, whereas the remaining six isolates
all hyperproduced the enzyme (Table 1). Only one isolate, E.coli C13, showed no growth-rate-dependent regulation (Fig.1).DNA Sequence Analyses of ClinicalE coli Isolates with Wild-
Type ampC &-Lactamase Expression. Because E. coli K-12 wasisolated in the pre-antibiotic-therapy era, two recent clinicalisolates of E. coli (C123 and C115), resistant to ampicillin at 1pug/ml, were randomly chosen for this study. This was done todetermine whether or not the ampC control region of E. coliK-12 could still be regarded as a prototype for E. coli. The se-quences of 449-bp DNA segments, including the ampC controlregion, from both of these strains were determined. In Fig. 2it can be seen that they resemble each other more than theyresemble the corresponding sequence of E. coli K-12. Unlike
a2)
C6.40
'Ua1)P.-4
v;
4.0
3.0
2.0
1.0
0 0.5 1.0 1.5Specific growth rate k, hr-'
FIG. 1. Regulation of the ampC 13lactamase enzyme as a functionofgrowth rate. The relative amount of -lactamase, determined by rocketimmunoelectrophoresis, is plotted as a function of the first-order con-stant for growth (k), as calculated from the expression k = ln 2/massdoublingtime in hr. Themedia usedweremediumE + acetate, mediumE + glycerol, medium E + glucose, and medium E + glucose + Cas-amino acids. Data are forE. coli clinical isolates C11 (A), C13 (0), C14(A), C15 (ci), C16 (A), and C17 (a).
E. coli K-12, the two isolates contained a T at -73, an A at -28,-an A at +81, a C at +185, and an A at +197 (Fig. 2). In ad-dition, E. coli C123 contained a C at +63, changing the secondamino acid in the ampC signal peptide from a phenylalanine toa leucine, and E. coli C115 contained a C at +116, an A at +119,and a C at +317. These last three changes did not affect thepredicted amino acid sequence (Fig. 2). Both E. coli C123 andC115 carry the same -10 and -35 regions in the ampC pro-moter as is present in E. coli K-12, as well as an identical ampCattenuator. This promoter, which allows for a low expression ofthe ampC ,-lactamase, can thus be regarded as the prototypeampC promoter for E. coli.DNA Sequence Analyses of the coli ampC &fiLactamase-
Hyperproducing Isolate Lacldng Growth-Rate-DependentRegulation. The /3-lactamase-hyperproducing clinical isolate C13resembled the clinical isolates C115 and C123 over the 449-bpsegment studied. Unlike E. coli K-12 but like C115 and C123,the isolate C13 contained a T at -73, an A at -28, an A at +81,a C at + 185, and an A at + 197 (Fig. 2). Two differences, how-ever, were found between the low 3-lactamase producers C115and C123 and the (3-lactamase hyperproducer C13 in the ampCpromoter region. Strain C13 carried a T instead of a C, as foundin C115 and C123, at position + 17 in the ampC attenuator (Fig.2). The thermodynamic strength of formation of the resultinghairpin loop was thereby reduced from AG(250C) = -29.4 kcal/mol to AG(250C) = -26.2 kcal/mol (1 kcal = 4.18 kJ) (25). Thischange most likely explains the absence of a growth-rate-de-pendent control of the ampC gene product in E. coli C13. Iso-late C13 also carried an A at position -32, thereby creating a
perfect match with the -35 region (5'-T-T-G-A-C-A-3') (5, 6)
A
0
.
D -
Genetics: Olsson et al.
5.(
-

7558 Genetics: Olsson et al.
-iCO)0
0o = _c c cox CO) CO)1 * *
Proc. Natl. Acad. Sci. USA 80 (1983)
00
I I kb
E. coli K12
E. coli C123E. .coli C115E. coli C13E. coli Cll,C14-C17Sh. sonnei OS10Sh. sonnei OS106Sh. sonnei OS107Sh. sonnei OS112,113
22
449 basesequenced fi
13 4 5 6
Ppairsfragment
5-ArgMetHisHisAlaMetHisAspLeuLyslIeHisValProAlaGlyLysTrpValPheTyrGlyLeuAlaAlalI1iGTATGCACCACGCGATGCACGATCTGAAAATCCACGTACCTGCGGGCAAATGGGTTTTCTACGGTCTGGCTGCTAT-120 -100 -80 -60
-------_-_- --------------------T-----------------------------------------------_ ---_------------T------ -----------------------------------------_--------------T------------------------------T-------____ --------T-----G--------------------------------------------------__ _ - T-__ G-----------------------------__ ------_- T- G--G ---- -----------_------------------
--------------------------------T----G---------- _----------------------- ------------------T-----G--------------------------------------
eLeuThrValValThrLeulleGlyValValThrIleOch +1 MetOch TerminationE. coli K12 CCTACAGTTGTCACGCTGATTGTGTCGTTACATCTACGCATCGCCATGTAAATCCGGCCCGCCTATGGCGGGCCGTTTTGTATGGAACCAGACCCT
-40 -35 -20 -10 0 ' 40
E. coli C123E. coli C115E. coli C13E. coli C11,C14-C17Sh. sonnei OSloSh. sonnei OS106Sh. sonnei OS107Sh. sonnei OS112,113
-------_--A--_-----------
------------------------A---------------T---------------------------------------------------------T--------------------------A----------------T------------------T---------------------------A----------------T-----------------W-------------------------------------T--------------------------A-----4 --------T---------------------------------------------------------T-
------------------A---------------T---------------------------------------------------------T-
MetPheLysThrThrLeuCysAlaLeuLeuI1eThrAlaSerCysSerThrPheAlaAlaPro GInLyslieProGlyMetAl aVal AlaVal I1eTyrGE. coli K12 ATGTTCAAAACGACGCTCTGGCCTTATTATTACCGCCTJTTGCTCCACATTTGCTGCCsCT//CAAAAITCCCGGGTATGGCGGTGGC6GTATTTATCto 80 100 120 180 200
E. coli C123E. coli C115E. coli C13E. coli Ci1.C14-C17Sh. sonnei OS1OSh. sonnei OS106Sh. sonnei OS107Sh. sonnei OS112,113
---C ----------------A----------------------1-------------------/ -----------C-----------A----------------------------------A----------------------------------C--A---//-----------C-----------A----------------------------------A-----------------------------------------//-----------C-----------A-------------
-G---------------------------------------------------------//-----------C----------------G---------------------------------------------------------//-----------C------------------------------G-------------------------------------------------G--------//----C-------C------------------------------G-----------------------------------------C---------------//-----------C---------------------------G---------------------------------------------------------//-----------C-------------------------
InGlyLysPro ThrLeuPheGluLeuGtySerValSerLysThrPheThrGlyValLeuGly 3'E. coli K12 AGGGTAAACCT//AgGTTGTTTGAGTTAGGTTC6GTCAGCAAAACATTTACTGG GTGCTTGGT
220 280 300 320
E. coli C123E. coli C115E. coli C13E. coli C11, C14-C17Sh. sonnei OSlOSh. sonnei OS106Sh. sonnei OS107Sh. sonnel OS112,113
//-C------------//--------------------------------------G-----------------------//--------------------------------------G-----------------------//--------------------------------------G-----------------------//--------------------------------------G-----------------------//--------------------------------------G-----------------------//--------------------------------------G------------
FIG. 2. Physical and genetic structure of the frd and ampC operons and a 449-bp nucleotide sequence including the frdD-ampC' overlap, theampC control region, and the 90 NH2-terminal amino acids of the ampC gene. The E. coli K-12 DNA sequence is written in capital letters. Thethree-letter abbreviations for the amino acids appear directly over their three-base codons (Och, ochre termination); the start of transcription ismarked by +1; and every 20th base pair in either direction from +1 is marked by a dot with the corresponding number written below each dot.The -35 and -10 promoter regions are underlined, and the stem and loop structure of the ampC attenuator is marked by horizontal arrows. Thenucleotide sequence of the clinical isolates ofE. coli and S. sonnei described in the text are written as broken lines below the E. coli K-12 sequence.Capital letters indicate base pair differences between the E. coli K-12 sequence and the clinical isolates. Encircled capital letters indicate base pairdifferences that directly influence expression of the ampC 3-lactamase; a capital letter with a A indicates an insertion of a base pair; and an un-derlining below the broken line indicates the -35 and -10 promoter regions. The nucleotide sequence between the slashes was determined but notwritten out in the figure. The nucleotide sequence from +210 to +329 was not determined in the E. coli isolate C123.
of the E. coli consensus promoter. A mutation at this positionin E. coli K-12 is known to increase transcription initiation bya factor of about 20 (7). It is therefore likely that ampC hy-perproduction in E. coli C13 is a combined effect of these twodiscrepancies from the prototype ampC promoter and atten-uator.
DNA Sequence Analyses of the E. coli ampC f-LactamaseHyperproducers with a Retained Metabolic Control. The fiveindependently isolated ampC ,B-lactamase hyperproducers (Cll,C14, C15, C16, and C17) that had retained growth-rate-de-pendent control were found to be identical over the entire 449-bp DNA segment. This sequence differed by 9 bp from the cor-
00wi
I frdA I frdB I fr-dCtgdD+4=i ApC

Proc. Natl. Acad. Sci. USA 80 (1983) 7559
responding sequence of E. coli K-12. With the E. coli ampCprototype transcriptional start base as a reference point, thesedifferences occurred at positions -88, -82, -42, -18, -1,+58, +65, + 185, and +317 (Fig. 2). When aligned with C115and C123, 13 and 12 differences were found, respectively (Fig.2). Thus, the five identical l3-lactamase hyperproducers (Cli,C14, C15, C16, and C17) had a sequence quite different fromthe other E. coli sequences. We had previously demonstratedthat for E. coli C16, the T at position -42 and the A at -18create a strong promoter distinct from, but overlapping, thatdefined as the ampC prototype promoter (18). In E. coli C16,transcription initiation starts four and five bases upstream fromthe start in E. coli K-12 (18). Since E. coli Cii, C14, C15, andC17 are identical toE. coli C16 in the ampC region, in the levelof (3-lactamase production, and in the transcriptional initiationsite (data not shown), they must all use the strong ampC pro-
moter as defined by E. coli C16.DNA Sequence Analyses of the ampC Region in S. sornws.
Because DNADNA hybridization has not shown any detect-able differences between the ampC regions from E. coli and S.sonnei (2) and because their respective ampC gene products are
immunologically identical (2), the ampC region of the 3lactam-sensitive clinical isolate S. sonnei OS10 was cloned (19). TheDNA sequence of the entire 449-bp segment defined in thispaper, carrying the ampC control region, was, apart for 1 bp,identical to the sequence found in the five different E. coli ampChyperproducing isolates (Cli, C14, C15, C16, and C17) (Fig.2). The difference was a C-G base pair in the OS10 sequence
instead of a TA base pair, as found in the E. coli hyperproduc-ers, at position -42 relative to the E. coli K-12 transcriptionstart (Fig. 2). This difference is within the hexameric sequence
that we previously have defined as the -35 region of the ampCpromoter in E. coli C16 (18). This sequence reads 5'-C-T-G-A-C-A-3' in S. sonnei OS10 and 5'-T-T-G-A-C-A-3' in the resis-tant E. coli isolates.DNA Sequence Analyses of Spontaneous S. soniw ampC
Lactamase-Hyperproducing Mutants. S. sonnei OS10 is resis-tant to ampicillin at 1 kg/ml and its production of ampC ,B-lac-tamase is comparable to that of E. coli K-12 wild type (strainLA5) and the E. coli clinical isolates C123 and C115. As in theseE. coli strains, the production of ampC ,&lactamase of S. sonneiOS10 increases with growth rate (2). Spontaneous ampicillin-resistant mutants from S. sonnei OSlO were selected with am-
picillin at 5 ,ug/ml and were obtained at an incidence of 10-10.Out of 20 mutants tested in rich medium, 18 were resistant toampicillin at 5-10 ug/ml and two were resistant to about 90Ag/ml. The ampC regions from the two representatives of themore resistant group, S. sonnei OS112 and OS113, as well as
two mutants resistant to ampicillin at about 10 ,ug/ml, S. sonneiOS106 and OS107, were cloned and their DNA sequences were
determined. The ampC regions of OS112 and OS113 were
identical and differed from the parental strain OS10 only by a
C-G to TA transition at position -42 relative to the E. coli ampCprototype transcription start base (Fig. 2). Consequently, theampC regions of S. sonnei OS112 and OS113 are identical to
those found in the E. coli clinical isolates Cli, C14, C15, C16,and C17. In vitro transcription confirmed that these two am-
picillin-resistant S. sonnei mutants initiated transcription fourand five bases upstream from the E. coli K-12 initiation site (datanot shown), as earlier shown for E. coli C16 (18). Thus S. sonneiOS112 and OS113 utilize the ampC promoter defined in E. coliC16 (18). However, the parental S. sonnei OS10 uses the same
transcription initiation site as E. coli K-12 and the two clinicalE. coli ampC prototypes, since in vitro transcription gives the41-nucleotide attenuated leadertranscript (datanot shown). Thus,
the novel strong promoter is generated solely by the base sub-stitution at -42.
In S. sonnei OS107 a COG to TEA transition at position -11had changed the -10 region in the ampC prototype promoterfrom 5'-T-A-C-A-A-T-3' to 5'-T-A-T-A-A-T-3' (Fig. 2). Anidentical base pair substitution in E. coli K-12 is known to in-crease 3lactamase production about 7-fold (7). Finally, S. son-nei OS106 was mutated in the ampC attenuator in that it carriedan insertion of aGC base pair between positions + 18 and + 19(Fig. 2). This alteration will decrease the thermodynamic sta-bility of the hairpin structure by changing the free energy offormation from -29.4 kcal/mol to -21.4 kcal/mol (25). Thisis in agreement with the finding that this isolate has lost its growthrate control of 3lactamase expression (2).
DISCUSSIONThe purpose of this study was to deduce how increased expres-sion from the E. coli chromosomal 3-lactamase gene, ampC, canevolve under natural conditions. We therefore characterized sixclinical isolates of E. coli. These isolates produced very highlevels of ampC 1&lactamase, higher than we have been able toselect for as a single mutational event in E. coli K-12 under lab-oratory conditions.
Here we demonstrate that five of these six clinical E. coliisolates have obtained their ampC (-lactamase-hyperproducingability as a consequence of a natural gene exchange with Shi-gella. Although independently isolated, they are all identical inthe 449-bp DNA sequence investigated. They differ markedlyfrom the corresponding sequences of E. coli K-12 and the otherclinical E. coli isolates. Furthermore, the five isolates con-tained, relative to the prototype ampC promoter, the two basepair substitutions that generate the novel 45-fold stronger ampCpromoter (18). S. sonnei OS10, which we assume carries a Shi-gella ampC prototype promoter, was identical in the ampC con-trol region to these five clinical 3-lactamase hyperproducersexcept for position -42 (Fig. 2). The expression from the S.sonnei OS10 ampC promoter is very low (Table 1), and in vitrotranscription revealed that the same start base is used in thisstrain as in the E. coli K-12 prototype promoter (data not shown).However, the Shigella promoter has an A at position -18.Therefore, a single mutational event (C to T at -42) is sufficientto shift transcription initiation from the prototype promoter tothe novel strong one. This unique property distinguishes theShigella promoter from that of the E. coli prototype and givesShigella the potential to mutate to a very high ampicillin re-sistance level.
It can be argued that the ampC control region of E. coli K-12 is not typical of E. coli strains present today. The five E. coliampC hyperproducers could have evolved from a recent E. coli.However, the sequenced DNA of hyperproducers differedmarkedly from that of two recent initial E. coli isolates with low3-lactamase expression (C115 and C123). These latter isolatesused the ampC prototype promoter and showed only one basepair difference in the promoter region relative to E. coli K-12(position -28).
It can also be argued that a transfer has occurred to S. sonneiof the high ampicillin resistance sequence present in E. coli Cli,C14, C15, C16, and C17. This sequence could subsequentlyhave mutated and generated the S. sonnei OS10 sequence, whichthen would be an atypical Shigella sequence. We cannot ex-clude this possibility, since it should require DNA sequenceanalysis of a large number of E. coli and Shigella isolates. How-ever, despite attempts with many different isolates, we havenever been able to isolate one-step mutants in E. coli with more
Genetics: Olsson et al.

Proc. Natl. Acad. Sci. USA 80 (1983)
than a 21-fold increased ampicillin-resistance level. This sug-gests that E. coli isolates normally lack the A residue at position-18 that is required to generate the novel strong ampC pro-moter. On the other hand, different Shigella isolates are easilymutated to high ampicillin resistance, favoring the idea that theShigella OS10 ampC promoter sequence represents the pro-totype for Shigella.The sixth isolate, E. coli C13, however, most likely owes its
25-fold increased ampC production to one promoter and oneattenuator mutation in an E. coli prototype sequence. E. coliC13 is also mutated in the ampC signal peptide at position +81,which may negatively affect the secretion or stability of the ,3lactamase enzyme.
The ampC promoter overlaps the distal genefrdD, in thefrdoperon (26). The protein encoded byfrdD is thought to act asa membrane anchoring protein for the fumarate reductase com-plex (27). It is possible that evolution of ampC hyperproductioncan only proceed in such a way that it will not adversely affectthe function of the frdD gene product. The -42 mutation inthe Shigella sequence, generating the strong novel ampC pro-moter, will not affect thefrdD gene product, whereas all knownup-promoter mutations in the E. coli ampC prototype promoterwill do so.
In our hands transfer of ampC-mediated ampicillin resis-tance between S. sonnei and clinical isolates of E. coli could eas-ily be achieved in batch cultures either by phage P1 transduc-tion or R-factor-mediated mobilization (data not shown). Wefavor the idea that a similar transfer has occurred in areas whereshigellosis is endemic. We believe that such transfer has gen-erated a small E. coli subpopulation containing the Shigella ampCprototype region. Strains carrying the novel ampC promoterare then selected from this subpopulation in the intestine by asingle mutational event. Alternatively, 3-lactam resistance mayhave evolved already in Shigella and then transferred to E. coli.
In this study we have only compared 449-bp DNA segments.Therefore we do not know the sizes of the Shigella sequencesthat are present in the five resistant E. coli isolates. If the Shi-gella E. coli hybrids have been generated by conjugation, theseisolates may represent very large natural intergeneric hybrids.It is conceivable that such hybrids are quite common in nature,putting into question the traditional classification system of E.coli and Shigella.
In conclusion, naturally occurring chromosomal ,B-lactam re-sistance in E. coli may have evolved by horizontal transfer ofampC DNA from Shigella species. A single base pair substi-tution in the Shigella prototype ampC sequence, while not af-fecting the protein of the overlapping frD gene, appears to give
rise to a level of resistance impossible to achieve by the E. coliprototype ampC sequence through a single mutational event.We thank Stina Olofsson for excellent technical assistance. We are
indebted to Richard Goldstein (Harvard) for valuable discussions andto Christopher Korch for carefully reading the manuscript. This workwas supported by grants from the Swedish Natural Science ResearchCouncil (Dnr 3373), the Swedish Medical Research Council (Dnr 5428),and the Board for Technological Development (Dnr 81-3384).
1. Jaurin, B. & Grundstrom, T. (1981) Proc. Natl. Acad. Sci. USA 78,4897-4901.
2. Bergstrom, S., Olsson, 0. & Normark, S. (1982)J. Bacteriol. 150,528-534.
3. Jaurin, B., Grundstrom, T., Edlund, T. & Normark, S. (1981) Na-ture (London) 290, 221-225.
4. Jaurin, B. & Normark, S. (1979) J. Bacteriol. 138, 896-902.5. Rosenberg, M. & Court, 0. (1979) Annu. Rev. Genet. 13, 256-275.6. Siebenlist, U., Simpson, R. B. & Gilbert, W. (1980) Cell 20, 269-
281.7. Jaurin, B., Grundstr6m, T. & Normark, S. (1982) EMBOJ 1, 875-
881.8. Jaurin, B. & Normark, S. (1983) Cell 32, 809-816.9. Edlund, T., Grundstrom, T. & Normark, S. (1979) Mol. Gen. Ge-
net. 173, 115-125.10. Bergstrom, S. & Normark, S. (1979) Antimicrob. Agents Chemo-
ther. 16, 427-433.11. Normark, S., Edlund, T., Grundstr6m, T., Bergstrom, S. & Wolf-
Watz, H. (1977)J. Bacteriol. 132, 912r922.12. Laurell, C. B. (1966) Anal. Biochem. 15, 45-52.13. Messing, J., Crea, R. & Seeburg, P. H. (1981) Nucleic Acids Res.
9, 309-321.14. Vogel, H. J. & Bonner, D. M. (1956) J.:Biol. Chem. 218, 97-106.15. Bertani, G. (1951) J. Bacteriol. 62, 293-300.16. Miller, J. H. (1972) Experiments in Molecular Genetics (Cold Spring
Harbor Laboratory, Cold.Spring Harbor, NY).17. Uhlin, B. E., Schweikaetz, V. & Clark, A. J. (1983) Gene 22, 255-
265.18. Olsson, O., Bergstr6m, S. & Normark, S. (1982) EMBOJ 1, 1411-
1416.19. Bergstrom, S., Lindberg, F., Olsson, 0. & Normark, S. (1983)J.
Bacteriol. 155, 1297-1305.20. Akusjarvi, G. & Pettersson, U. (1978) Virology 91, 477-480.21. Hines, C. J. & Ray, D. S. (1980) Gene 11, 207-218.22. Benton, W. D. & Davies, R. W. (1977) Science 196, 180-182.23. Hu, N. & Messing, J. (1982) Gene 17, 271-277.24. Sanger, F., Nicklen, S. & Coulson, A. R. (1977) Proc. Natl. Acad.
Sci. USA 74, 5463-5467.25. Tinoco, I., Jr., Borer, P. N., Dengler, B., Levine, M. D., Uhlen-
beck, O. C., Crothers, D. M. & Gralla, J. (1973) Nature (London)New Biol. 246, 40-41.
26. Grundstrom, T. & Jaurin, J. B. (1982) Proc. Natl. Acad. Sci. USA79, 1111-1115.
27. Lemire, B. D., Robinson, J. J. & Weiner, J. H. (1983)J. Bacteriol.152, 1126-1131.
7560 Genetics: Olsson et al.





























