Embed Size (px)
Citation preview

Angiogenesis and its control in the femalereproductive system
Hamish M Fraser and Stephen F LunnMRC Human Reproductive Sciences Unit, Centre for Reproductive Biology, Edinburgh, UK
The rapid, controlled and cyclical nature of angiogenesis in the femalereproductive tract suggests that interference with this process should provide anovel approach to manipulation of reproductive function. Many factors involvedin the regulation of angiogenesis have been identified, and the possibility ofstimulating or inhibiting these paracrine control mechanisms is being addressedusing current advances in the development of angiogenic and anti-angiogeniccompounds. Studies with animal models indicate that the normal processes offolliculogenesis, ovulation and corpus luteum function in the ovary, and thecontrol of menstruation and implantation in the endometrium could beprofoundly influenced by manipulation of angiogenesis. Novel therapeuticagents targeted to the angiogenic pathway may also have a wide range ofapplications in pathological processes in the reproductive tract such as cancer,endometriosis, fibroid growth, and ovarian hyperstimulation syndrome.
Correspondence toDr Hamish M Fraser, MRC
Human ReproductiveSciences Unit, Centre forReproductive Biology, 37
Chalmers StreetEdinburgh EH3 9ET, UK
Angiogenesis is the development of new blood vessels by endothelial cellproliferation and outgrowth from pre-existing vessels. It is regulated by acomplex series of growth factor interactions, including stimulatory,modulatory and inhibitory regulators and is associated with changes inthe extracellular matrix required to allow migration of the newly-formingvessels. Apart from during tumour growth and wound healing, the adultvascular endothelium is generally quiescent. The notable exceptions arewithin the female reproductive system; the ovaries, uterus and placenta1"6.These tissues undergo cyclic changes in angiogenesis necessary to supplythe nutrients and hormone precursors essential for the establishment andmaintenance of pregnancy. During the menstrual cycle, this physiologicalangiogenesis in the corpus luteum and endometrium is distinctive in that,in the absence of conception, it is followed by tissue regression and re-initiation of cyclicity. This tight physiological control contrasts to thesituation in tumours, and raises the possibility that endogenous inhibitorsof angiogenesis play an integral role in the reproductive tract. Thus, thesetissues provide an outstanding system in which to study the normalphysiology of angiogenesis. Understanding the mechanisms of control ofangiogenesis in the reproductive system and the harnessing of major
British Medical Bulletin 2000, 56 (No 3) 787-797 O The British Council 2000
Downloaded from https://academic.oup.com/bmb/article-abstract/56/3/787/351359by gueston 17 February 2018

Human reproduction: pharmaceutical and technical advances
advances in the development of angiogenesis inhibitors and stimulatorsmay reveal new approaches to regulation of reproductive function andtreatment of pathological conditions of the female reproductive tract. Inaddition, insight should be gained into the changes which precipitate therelatively uncontrolled angiogenesis responsible for the growth of solidtumours and how this may be inhibited.
This chapter describes briefly the potential angiogenic control mechan-isms within the female reproductive tract and how these might be targeted,reviews recent data investigating the consequences of angiogenesisinhibition in experimental models, and examines the projected clinicalapplications. Since mechanisms involved in regulation of cyclic ovarianand uterine angiogenesis have much in common, progress with respect tothe ovary is highlighted and the reader referred to recent reviews for theuterus5"7.
Angiogenic control mechanisms
Folhcular angiogenesis commences at, or shortly before, the developmentof a recognisable thecal layer and increases in follicles undergoing matur-ation to the antral stage. Blood vessels increase in number and size as thefollicle develops, but do not penetrate the granulosa cell layer while thebasement membrane surrounding it remains intact3-8. The extent to whichangiogenesis governs follicular atresia on the one hand, and selection of thedominant follicle on the other, is of great interest; in some species, arelationship has been demonstrated3, but differences are not obvious in thehuman ovary8.
During the process of ovulation, the basement membrane loses its integrityand the thecal blood vessels access the granulosa cell layer. Subsequently,formation of the corpus luteum is associated with intense angiogenesis (Fig.I)9. By the mid-luteal phase, when the corpus luteum (CL) is fully functional,blood flow is among the greatest of any tissue in the body. Proliferation ofendothelial cells continues at a moderate level, decreasing towards the end ofthe cycle. By the mid-luteal phase, the microvascular tree is established. Itappears that the rescue of the CL in early pregnancy is not associated with afurther spurt of intense angiogenesis since chorionic gonadotrophin (hCG)does not influence endothelial cell proliferation (Fig. I)9.
Angiogenesis is regulated in the reproductive tract and elsewhere by atleast 20 angiogenic growth factors and inhibitors identified to date. A keyplayer is vascular endothelial growth factor (VEGF) also known asvascular permeability factor10. VEGF is a member of the platelet-derivedgrowth factor family, bemg a dimeric protein existing as 5 different homo-dimeric isoforms generated by alternative splicing from a single gene. Thesecreted forms are composed of 121 and 165 amino acids and act upon
788 British Medical Bulletin 2000,56 (No 3)
Downloaded from https://academic.oup.com/bmb/article-abstract/56/3/787/351359by gueston 17 February 2018

Angiogenesis
300
glow-
- . a )
100-
0 J
150-
1 loo-'s
E 50-z
0 J
b)
Early Mid Late Rescued
Fig. 1 (a) Number of endothelial cells and (b) proliferating endothehal cells per field inthe human corpus luteum during the early, mid and late luteal phase and after rescue byhCG administration. Data are means ± SEM; *shows significant differences betweengroups Note the intense proliferation during the early luteal phase and the resultantincrease in endothelial cell number. From Rodger etaP with permission.
vascular endothelial cells in which its receptor tyrosine kinases (Flt-1and KDR) are expressed selectively. VEGF is expressed in the ovary in aregulated manner11'12 and its importance in ovarian angiogenesis hasbeen established following studies in which its action has been inhibitedduring the ovulatory cycle (see below).
Particular attention is being given to the inter-relationship of VEGFwith a novel family of angiogenic regulators, the angiopoietins, whichconsist of about 500 ammo acids and act via another receptor tyrosinekinase, Tie-2, on the endothelial cells. Angiopoietins are relatively highlyexpressed in the ovary, uterus and placenta12. Since they appear to playa major role in stabilisation of blood vessels on the one hand andendothelial cell death on the other, the angiopoietins are attractivecandidates as regulators of these divergent processes as they occur in thefemale reproductive tract. When active angiogenesis is occurring in thepresence of VEGF, Ang-2 acts in concert to enhance angiogenesis, whileAng-1 is involved in the process of maturation and stabilisation of newblood vessels12. Paradoxically, in circumstances of reduced VEGFexpression, Ang-2 may act to destabilise blood vessels and inducevascular regression by competitive inhibition of Ang-112'13.
The temporal changes in the pattern of expression of VEGF andangiopoietin mRNA and in the rat ovary have shown that VEGF is
British Medical Bulletin 2000, 56 (No 3) 789
Downloaded from https://academic.oup.com/bmb/article-abstract/56/3/787/351359by gueston 17 February 2018

Human reproduction- pharmaceutical and technical advances
expressed in the pre-ovulatory follicle and in the hormone-producingcells of the corpus luteum12. Ang-2 is present in theca of the pre-ovulatory follicle and in the endothelial cell sprouts of the developingcorpus luteum, suggesting an angiogenic role in concert with VEGF atthis time. Intriguingly, Ang-2 is highly expressed in regressing corporalutea, when VEGF is declining, implying its involvement in the luteolyticprocess via destabilisation of blood vessels. Similarly, quantitative PCRin the bovine revealed the Ang-2 :Ang-l mRNA ratio increases duringluteolysis14. While luteolysis in the normal cycle is associated withendothelial cell loss, stabilisation and maturation of luteal blood vesselsmay be crucial to the maintenance of the corpus luteum in earlypregnancy4. The mechanisms involved in these divergent processes arenot understood. If luteolysis is associated with an increase in Ang-2expression and blood vessel destabilisation perhaps 'rescue' of thecorpus luteum is associated with an increased expression of Ang-1resulting in recruitment of pericyte cells which stabilise the bloodvessels. Control of luteal function differs markedly between species andit is not yet known whether similar inter-relationships between positiveand negative angiogenic mechanisms exist in the primate corpus luteum.
Two further natural inhibitors of angiogenesis have been discoveredrecently, both being fragments of larger molecules; angiostatin is 98%identical to an internal fragment of plasminogen,15 while endostatincorresponds to the C-terminal fragment of collagen XVIH16. Both proteinsinhibit tumour growth, but their role in reproductive physiology isunknown. Since it appears that an 'angiogenic switch' regulates thedivergent states of follicular development, luteal rescue and luteolysis, andendometrial function, it is of importance to localise and quantify thetemporal changes in the angiostatin/endostatin system in these situationsto explore the hypothesis that production of these proteins is increased inthe regressing state and suppressed in association with rescue.
Changes in the extracellular matrix (ECM) are an essential part of theangiogenic process and are regulated by metalloproteinases and theirinhibitors. Cell adhesion to the ECM is primarily through integrins, a familyof heterodimeric transmembrane proteins17. These mediate cell adhesion tothe ECM leading to intracellular signalling events that regulate cell survival,proliferation and migration18. During angiogenesis, it is thought that anumber of integrins expressed on the surface of activated endothelial cellsregulate critical adhesive interactions with a variety of ECM proteins.
A complete understanding of these complex regulatory mechanismswithin the ovary and uterus will allow systematic targeting in pathologieswhich have a vascular component. While basic studies strive to achievethis goal, targeting of putative key regulators has been conducted both inthe laboratory and clinic during the last decade, particularly in the area ofsolid tumour biology.
790 British Medical Bulletin 2000,56 (No 3)
Downloaded from https://academic.oup.com/bmb/article-abstract/56/3/787/351359by gueston 17 February 2018

Angiogenesis
Manipulation of angiogenesis
Development of anti-angiogenic agents
Over 300 endogenous, natural or synthetic anti-angiogenic agents havebeen described19. The initial development involved screening for suchactivity in compounds developed for other indications. A second grouprequired the design of specific antagonists to known positive angiogenicfactors and their receptors, while a third class have been identified fromtumour sources as having negative effects on angiogenesis (Table 1).
Numerous studies have shown that inhibition both by broad-spectrumor systematic targeting of the angiogenic pathway can have profoundsuppressive effects upon tumour growth in animal models16-20"22. Theseinclude the effects of neutralisation of VEGF in an ovarian cancermodel23. Most of the clinical trials on angiogenesis inhibitors haveemployed first generation compounds19 such as the fumagillin analogue,TNP-470, but in common with many of these compounds its mechanismof action is multi-factorial and is still under investigation.
Specific angiogenic peptide or receptor antagonists are being developedfor clinical application. A humanised monoclonal antibody to VEGF iscurrently on clinical trial and the VEGF receptor is also being targetedusing a soluble truncated Flt-1 receptor or tyrosine kinase inhibitors, thelatter having the advantage of oral activity. Combination treatments such
Table 1 Examples of anti-angiogenic agents
First generationSuraminFumagillin derivative (TNP-470)ThahdomideMedroxyprogesteroneAnti-oestrogens
Inhibition of positive regulatorsVEGF inhibitors
VEGF immunoneutrahsing antibodiesTruncated soluble receptorTyrosine kinase receptor antagonists, eg Sugen 5416VEGF receptor immunoneutrahsing antibodiesVEGF diphtheria toxinRibozymes
Angiopoietin/Tle receptor antagonistsMetalloproteinase inhibitors (e g Marimastat)Integrin inhibitors (e.g Vitaxin)
Administration of negative regulatorsAngiogemnAngiostatinThrombospondinEndostatin
British Medical Bulletin 2000, 56 (No 3) 791
Downloaded from https://academic.oup.com/bmb/article-abstract/56/3/787/351359by gueston 17 February 2018

Human reproduction: pharmaceutical and technical advances
as VEGF inhibitor followed by Ang-2 may prove to be exceptionallyeffective in preventing angiogenesis13.
The involvement of the ECM in angiogenesis has led to development ofmetalloproteinase inhibitors and neutralising antibodies capable of inhibit-ing specific integrins. The therapeutic application of such approaches iscurrently under investigation in the clinic.
Initial studies in experimental tumour models using negative regulatorsof angiogenesis, such as recombinant angiostatin, were encouraging inthat they demonstrated that tumours may be treated successfully withthese agents without generatmg resistance15'16-22, hi addition, prolongedtumour dormancy was observed post-treatment. The challenge ofextending these results to man is eagerly awaited.
As increasing numbers of specific antagonists are developed, it shouldbecome possible to target systematically key steps in the angiogenicpathway at selected periods of the reproductive cycle using suitableanimal models and their effects upon angiogenesis in the ovary, uterusand placenta determined. This potential for inhibition of physiologicalangiogenesis is only just beginning to be explored and three of the firstexperiments are reviewed below.
Rodent studies
In the reproductive system, the pioneering study underlining the importanceof angiogenesis was carried out in the mouse using TNP-47024. Femaleswere treated at various times after mating and it was shown that TNP-470prevents pregnancy by interfering with the process of deciduahsanon andplacental and yolk sac formation. In mice with regular oestrous cyclestreated for 16 days, mating was abolished, there was a marked decrease inendometrial stromal and glandular cell proliferation, uterine blood vesseldevelopment was severely restricted, and the ovaries contained fewer andsmaller corpora lutea than cyclic controls.
The importance of angiogenesis in the immature rat ovary was shownin studies in which VEGF was blocked using a truncated Fit receptor25.Follicular development was induced by gonadotrophin treatment andresulted in multiple ovulations and elevated serum progesterone incontrol rats, whereas those treated with antagonist starting 4-6 h priorto gonadotrophin administration showed small antral follicles and onlya few corpora lutea. The latter were relatively avascular and oftendemonstrated central ischaemic necrosis. While the uteri of the gonado-trophin-treated rats demonstrated morphological evidence of steroidstimulation, such changes were largely blocked in antagonist-treatedanimals, presumably secondary to the comparative lack of steroidstimulation, but also in part as a result of inhibition of uterine VEGF.
792 British Medical Bulletin 2000,56 (No 3)
Downloaded from https://academic.oup.com/bmb/article-abstract/56/3/787/351359by gueston 17 February 2018

Angiogenesis
Primate studies
The mechanisms which regulate luteal function in rodents and primates aremarkedly different, hi the non-fertile cycle of the rodent, the corpus luteumis active for less than a day, while that of primates is functional for 2 weeksprior to its regression. In addition, in many non-primates, regression of thecorpus luteum is mediated by a specific luteolysin, a phenomenon notapparent in the human. In the rat model, since VEGF antagonist treatmentwas initiated prior to the onset of follicular hyperstimulation,25 it was notpossible to dissociate the luteal inhibition from the consequences ofinhibition of angiogenesis in the developing follicle. Experiments outlinedbelow targeted the corpus luteum specifically by treating animals duringthe normal cycle at the time of ovulation.
When macaques or marmosets were treated with TNP-470, there wereno apparent effects upon angiogenesis or luteal function26 in contrast tothe results in the mouse. The reason for this difference is unclear, butcould be related to dose or route of administration of the compound orreal species differences. The effects of 'first generation' compounds maybe comparatively weak relative to the more specific antagonists beingdeveloped. These primate results emphasise the need for caution inextrapolating results from the rodent to the human.
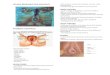
Luteal angiogenesis is suppressed by neutralisation of VEGF27.Marmosets were treated with a monoclonal antibody to human VEGFstarting at the time of ovulation and continuing for 3 days (early lutealphase group) or 10 days (mid-luteal group). Treatment effects weredetermined by measuring plasma progesterone concentrations, endo-thelial cell proliferation index using bromodeoxyuridine (BrdU)incorporation, and endothelial cell number using the specific cell marker,factor Vin. Neutralisation of VEGF during the early luteal phase inhibitedthe intense angiogenesis associated with luteal formation (Fig. 2a,b). Thisresulted in blockade of development of the normally extensive capillarybed since, in the mid-luteal phase treated animals, the endothelial cellnumbers were reduced (Fig. 2c,d). Luteal function, as judged by secretionof progesterone, was markedly compromised by the treatment, beingreduced by 60% in comparison with controls and demonstrating theprinciple of manipulation of ovarian function by this approach. Furtherstudies using this model should allow the identification of the essentialcomponents of the angiogenic pathway and provide a vehicle fortranslating these findings into safe and effective clinical application.
Information on stimulation of angiogenesis within the reproductivesystem, e.g. by recombinant protein or by gene transfer, is currently limited.However, the feasibility of this approach has been demonstrated in clinicaltrials in patients with critical limb ischaemia and myocardial ischaemia19.
British Medical Bulletin 2000; 56 (No 3) 793
Downloaded from https://academic.oup.com/bmb/article-abstract/56/3/787/351359by gueston 17 February 2018

Human reproduction: pharmaceutical and technical advances
Fig. 2 Photomicrographs of early luteal phase marmoset corpora lutea showing(a) proliferating endothehal cells in microvessels and capillaries (dark staining nuclei) usingBrdU incorporation as a marker, (b) reduced proliferation following anti-VEGF treatment;(c) localisation of endothehal cells (dark staining) using Factor VIII as a marker in the mid-luteal corpus luteum; and (d) reduction in the vascular tree following anti-VEGF treatment.(Scale bar = 200 |im). SE Dickson and HM Fraser unpublished.
Clinical studies and possible applications
Anti-angiogenic compounds have been used primarily for treatment ofcancer, but are also envisaged for non-malignant pathologies which have aneovascular component (e.g. rheumatoid arthritis, psoriasis and retinalneovascularisation)20*22. Results from the use of 31 agents which haveentered clinical trials at over 140 centres have been reviewed19. In additionto treatment efficacy, such studies allow evaluation of side-effects and thesehave turned out to be more wide-spread than anticipated. This may suggestthat, in tissues in which there appears to be no angiogenesis, angiogenicregulators have a more general 'housekeeping' role. The introduction ofdrugs having a selective site of action may dissociate specific from non-specific adverse effects on the vasculature or overcome them altogether.Paradoxically, such observations could shed light on novel physiologicalsites and/or pathways of vascular regulation. In any event, it is importantthat future trials incorporate a multidisciplinary team approach19 whichincludes a reproductive component. Ultimately, as with any other drug, arisk-benefit assessment will have to be undertaken prior to commencementof treatment.
794 British Medical Bulletin 2000,56 (No 3)
Downloaded from https://academic.oup.com/bmb/article-abstract/56/3/787/351359by gueston 17 February 2018

Angiogenesis
To date, there have been no reports on the effects of regulators ofangiogenesis on the reproductive system in women, although it is acknow-ledged that it offers numerous potential targets both for manipulation offertility and treatment of pathologies involving a vascular com-ponent1^'24'25. Manipulation of angiogenesis indicates a potentially power-ful approach to either promoting or inhibiting the normal processes offolliculogenesis, ovulation and corpus luteum function in the ovary, andto the control of menstruation and implantation in the endometrium. It isunlikely that for fertility control, one would aim to suppress angiogenesisfor prolonged periods as the risk of disruptive side-effects would beunacceptable. Targeting anti-angiogenic treatment to the folhcular phasewould probably compromise follicular development by inhibition of thethecal vasculature. Treatment during the early luteal phase wouldsuppress progesterone production and probably inhibit implantation.Despite the fact that the functioning corpus luteum is essential for theestablishment of pregnancy, there is no agent currently in use which canprevent the supporting effect of hCG during the conceptual cycle. Thus,the use of angiogenesis inhibitors during the postovulatory period couldhave potential for development.
One clinical condition in which inhibition of follicular angiogenesis wouldbe indicated is ovarian hyperstimulation syndrome (OHSS) which occurs ina proportion of patients undergoing ovulation induction. The patho-physiology of OHSS is complex, but increased ovarian angiogenesis andpermeability are major factors28. An effective treatment for this potentiallyfatal condition has proved elusive and it is likely that the introduction of anti-angiogenic approaches will offer a major clinical advantage.
Angiogenesis is implicated to varying degrees in numerous pathologicalprocesses of the endometrium including dysfunctional uterine bleeding,endometrial response to exogenous hormonal treatments, bleedingassociated with intra-uterine devices, uterine leiomyomata, endometriosisand endometrial carcinoma7. The pathophysiology of the vasculature inthese conditions is currently the focus of active research. For example,VEGF has been implicated in endometriosis5. Inhibition of angiogenesisshould have potential in the treatment of these conditions, possibly as anadjunct to existing therapies operating via a separate pathway.
Administration of angiogenic agents to increase blood vessel develop-ment could be of use in stimulation of follicular development, for thetreatment of inadequate luteal function, early pregnancy failure, pre-eclampsia and intra-uterine growth retardation.
Studies in the female reproductive tract are gathering momentum asthe potential for control of angiogenesis in normal and pathologicalconditions come under scrutiny. This is an exciting field in whichadvances are likely in the near future and this review has given someindication as to the prospects for further developments.
British Medical Bulletin 2000, 56 (No 3) 795
Downloaded from https://academic.oup.com/bmb/article-abstract/56/3/787/351359by gueston 17 February 2018

Human reproduction: pharmaceutical and technical advances
References
1 Folkman J. Angiogenesis in female reproductive organs. In: Alexander NJ, D'Arcangues C (Eds)Steroid Hormones and Uterine Bleeding. Washington DC: AAAS Press, 1992; 143-58
2 Reynolds LP, Kilhlea SD, Redmer DA. Angiogenesis in the female reproductive system. FASEBJ 1992; 6: 886-92
3 Redmer DA, Reynolds LP. Angiogenesis in the ovary. Rev Reprod 1996; 1: 182-924 Augustin HG. Development of the vascular system of the corpus luteum. In: Risau W, Rubanyi
G (Eds) Morphogenesis of Endotbeltum Reading, UK Harwood, 2000; 237-2545 Smith SK. Angiogenesis, vascular endothelial growth factor and the endometnum. Hum
Reprod Update 1998; 4: 509-196 Rees M, Hague S, Oehler MK, Bicknell R. Regulation of endometnal angiogenesis. Climateric
1999; 2: 52-87 Abulafia O, Sherer DM Angiogenesis of the endometnum. Obstet Gynecol 1999; 94: 148-538 Suzuki T, Sasano H, TaKaya R, Fukaya T, Yajima A, Nagura H. Cyclic changes of vasculature
and vascular phenotypes in normal human ovaries. Hum Reprod 1998; 13: 953-99 Rodger FE, Young FM, Fraser HM, Dhngworth PJ. Endothelial cell proliferation follows the
mid-cycle luteimzing hormone surge, but not human chononic gonadotrophin rescue, in thehuman corpus luteum Hum Reprod 1997; 12: 1723-9
10 Ferrara N, Davis-Smyth T The biology of vascular endothelial growth factor Endocrine Rev1997; 18: 4-25
11 Gordon JD, Mesiano S, Zaloudek CJ, Jaffe RB. Vascular endothelial growth factor localizationin human ovary and fallopian tubes: possible role in reproductive function and ovanan cystformation. / Clm Endocrinol Metab 1996; 81 353-9
12 Maisonpierre PC, Sun,C, Jones PF et al. Angiopoietin-2, a natural antagonist for Tie2 thatdisrupts in vtvo angiogenesis. Science 1997, 277: 55-60
13 Hanahan D. Signaling vascular morphogenesis and maintenance. Science 1997, 277: 48-5014 Goede V, Schmidt T, Kimmina S, Kozian D, Augustin HG. Analysis of blood vessel maturation
processes during cyclic ovarian angiogenesis. Lab Invest 1998; 78: 1385-9415 O'Reilly MS, Holmgren L, Chen C, Folkman J. Angiostatin induces and sustains dormancy of
human primary tumors in mice. Nat Med 1996; 2: 689-9216 O'Reilly MS, Boehm T, Shing Y et al. Endostatin: an endogenous inhibitor of angiogenesis and
tumor growth Cell 1997; 88: 277-8517 Bazzoni G, Dejana E, Lampugnani G. Endothelial adhesion molecules in the development of
the vascular tree: the garden of forking paths. Curr Opm Cell Biol 1999; 11: 573-8118 Ehcein BP, Cheresh DA. The role of av integnns during angiogenesis: insights into potential
mechanisms of action and clinical development./ Clm Invest 1999, 103: 1227-3019 Thompson WL), Li WW, Maragoudakis M. The clinical manipulation of angiogenesis:
pathology, side-effects, surprises, and opportunities with novel human therapies. / Pathol 1999;187: 503-10
20 Auerbach W, Auerbach R. Angiogenesis inhibition: a review. Pharm Ther 1994; 63: 265-31121 Skobe M, Rockwell P, Goldstein N, Vosseler S, Fusenig NE. Halting angiogenesis suppresses
carcinoma cell invasion. Nat Med 1997; 3: 1222-722 Jones A, Bicknell R Angiogenesis and invasion. In- Souhami RL, Tannock I, Hohenberger P,
Honot JC (Eds) Oxford Textbook of Oncology. Oxford. Oxford University Press, 2000; Inpress
23 Mesiano S, Ferrara N, Jaffe RB Role of vascular endothelial growth factor in ovanan cancer:inhibition of ascites formation by lmmunoneutralization. Am J Pathol 1998; 153: 1249-56
24 Klauber N, Rohan RM, Flynn E, D'Amato RJ. Cntical components of the female reproductivepathway are suppressed by the angiogenesis inhibitor AGM-1470. Nat Med 1997; 3: 443-6
796 British Medical Bulletin 2000;56 (No 3)
Downloaded from https://academic.oup.com/bmb/article-abstract/56/3/787/351359by gueston 17 February 2018

Angiogenesis
25 Ferrara N, Chen H, Davis-Smyth T et al. Vascular endothehal growth factor is essential forcorpus luteum angiogenesis Nat Med 1998; 4: 336-40
26 Fraser HM, Dickson SE, Morris KD, Erickson GF, Lunn SF. The effects of the angiogenesisinhibitor TNP-470 on luteal establishment and function in the primate. Hum Reprod 1999; 14:2054-60
27 Fraser HM, Dickson SE, Morris KD, Lunn SF, Carroll V, Bicknell R. Suppression of lutealangiogenesis by neutralization of vascular endothehal growth factor in the primate.Endocrinology 2000, 141: 995-1000
28 Schenker JG. Clinical aspects of ovarian hyperstimulation syndrome. Eur J Obstet Gynecol1999; 85 13-20
British Medical Bulletin 2000, 56 (No 3) 797
Downloaded from https://academic.oup.com/bmb/article-abstract/56/3/787/351359by gueston 17 February 2018